Результат интеллектуальной деятельности: Штамм Escherichia coli - продуцент L-треонина
Вид РИД
Изобретение
Настоящее изобретение относится к микробиологической промышленности, в частности, к микробиологическому синтезу L-треонина с использованием бактерии вида Escherichia coli.
Традиционно L-аминокислоты в промышленном масштабе получают методом ферментации с использованием штаммов микроорганизмов, выделенных из природных источников, или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано множество методов увеличения продукции L-аминокислот, например, путем трансформации микроорганизма рекомбинантной ДНК (US 4278765). Указанные методы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот и/или уменьшении чувствительности целевого фермента к обратному ингибированию продуцируемой L-аминокислотой (WO 9516042, US 5661012, US 6040160).
Известны различные штаммы, использующиеся для производства L-треонина методом ферментации. Это штаммы с увеличенными активностями ферментов, вовлеченных в биосинтез L-треонина (US 5175107; US 5661012; US 5705371; US 5939307; ЕР 219027), штаммы, устойчивые к некоторым химических реагентам, таким как L-треонин и его аналоги (WO 0114525, ЕР 301572, US 5376538), штаммы с инактивированными ферментами системы деградации L-треонина (US 5939307 и US 6297031), штаммы, в которых устранена чувствительность целевого фермента к ингибированию продуцируемой аминокислотой или ее побочными продуктами по типу обратной связи (US 5175107 и US 5661012).
В настоящее время известны штаммы-продуценты L-треонина с различным уровнем продукции аминокислоты. Штамм Escherichia coli ВКПМ В-11820 (RU 2546237) за 24 часа ферментации способен синтезировать до 7,1 г/л L-треонина, штамм Escherichia coli B-3996/pMrelA (RU 20041177) - 12,2 г/л за 24 часа ферментации, Escherichia coli ВКПМ В-13240 (RU 2697499) - 16,5 г/л за 24 часа культивирования, штамм Escherichia coli B-3996/pMW118-ptsG-mut-43 (RU 2335536) продуцирует до 12,0 г/л за 48 часов ферментации, при более продолжительном культивировании в течение 65 часов штамм Escherichia coli B-3996-ΔgcvB (RU 2395567) синтезирует L-треонин до 31,0 г/л. Различие в уровне продукции L-треонина связано как с биотехнологическими показателями конкретного штамма, так и с условиями его культивирования: количество субстрата, продолжительность ферментации, способ рН-статирования, условия аэрации и др.
Однако, все вышеприведенные штаммы получены на основе штамма Escherichia coli ВКПМ В-3996, который сам был получен на основе дикого штамма Escherichia coli ВКПМ В-7. в том числе с использованием ненаправленного мутагенеза, что предопределило наличие в нем мутаций, отрицательно влияющих на биотехнологические показатели.
В соответствии с этим, важную роль при конструировании продуцента L-треонина играет направленное введение модификаций в геном дикого штамма Escherichia coli, не подвергавшегося мутагенезу и селекционным работам, что позволяет получить конечный штамм продуцент с известной нуклеотидной последовательностью.
Преимущество такого подхода заключается в том, что влияние на продукцию L-треонина каждой из вводимой генетической модификации возможно оценить независимо от других, в то время, как при использовании штамма, ранее подвергшегося ненаправленному мутагенезу, накопленные мутации, могут искажать эффект от вновь вводимой модификации.
Технической задачей, на решение которой направлено настоящее изобретение является расширение арсенала штаммов Escherichia coli, способных к продукции L-треонина.
Поставленная задача решена тем, что на основе дикого штамма Escherichia coli MG1655 ВКПМ В-13106 в несколько этапов, путем введения направленных модификаций, получен штамм бактерии Escherichia coli ВКПМ В - 13427.
Штамм Escherichia coli MG1655 ВКПМ В-13106 взят за основу благодаря своей изученности. Он является одним из популярных штаммов Е. coli K-12 дикого типа, используемых в метаболической инженерии. Именно для этого штамма была впервые определена последовательность генома Е. coli.
В штамм Escherichia coli MG1655 ВКПМ В-13106 введены следующие модификации:
- С целью улучшения ростовых свойств в штамме ВКПМ В-13106 исправлена природная мутация rph-1 в гене rph.
- Для повышения продукции L-треонина в штамме оверэкспрессированы гены пути биосинтеза треонина thrABC под конторолем протомора PH207 и терминатора транскрипции TrrnB. Для устранения ретро-ингибирования с продукта гена thrA, в последовательность гена введена десенсибилизирующая мутация Gly433Arg
- Биодеградация L-треонина в клетке снижена за счет удаления генов tdcB, kbl-tdh.
- Экспорт L-треонина в культуральную жидкость усилен путем экспрессии гена rhtA, кодирующего траспортер L-треонина, под контролем конститутивного промотора P1077PR
- Во избежание обратного транспорта аминокислоты внутрь клетки удалены гены sstT, tdcC.
- С целью расширения арсенала используемых селективных маркеров, которые в последствии могут облегчить конструирование продуцента, введен ряд мутаций rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG
- Увеличение продукции L-треонина достигнуто также путем снижения ppGpp-гидролазной активности в клетках за счет введения мутаций T252ins(CATGAT) 253А, G520T, C1585T в ген spoT
Большая часть указанных модификаций описана и успешно применена при конструировании штаммов-продуцентов аминокислот на основе Escherichia coli (Бирюкова И.В. и др. Генетика. - 2010. - Т. 46. - №. 3. - С. 349-355; Lee K.Н. et al, Molecular systems biology. - 2007. - Т. 3. - №. 1; Livshits V.A. et al., Research in microbiology. - 2003. - Т. 154. - №. 2. - C. 123-135; Bubnov D.M. et al., Journal of microbiological methods. - 2018. - T. 151. - C. 48-56; Sarubbi E. et al., Journal of Biological Chemistry. - 1989. - T. 264. - №. 25. - C. 15074-15082; RU 2212448; RU 2288265; RU 2697219, EP 2628792 A1).
Сочетания модификаций, использованного в заявляемом изобретении, в источниках информации нами не обнаружено.
Изобретение проиллюстрировано следующими фигурами графического изображения:
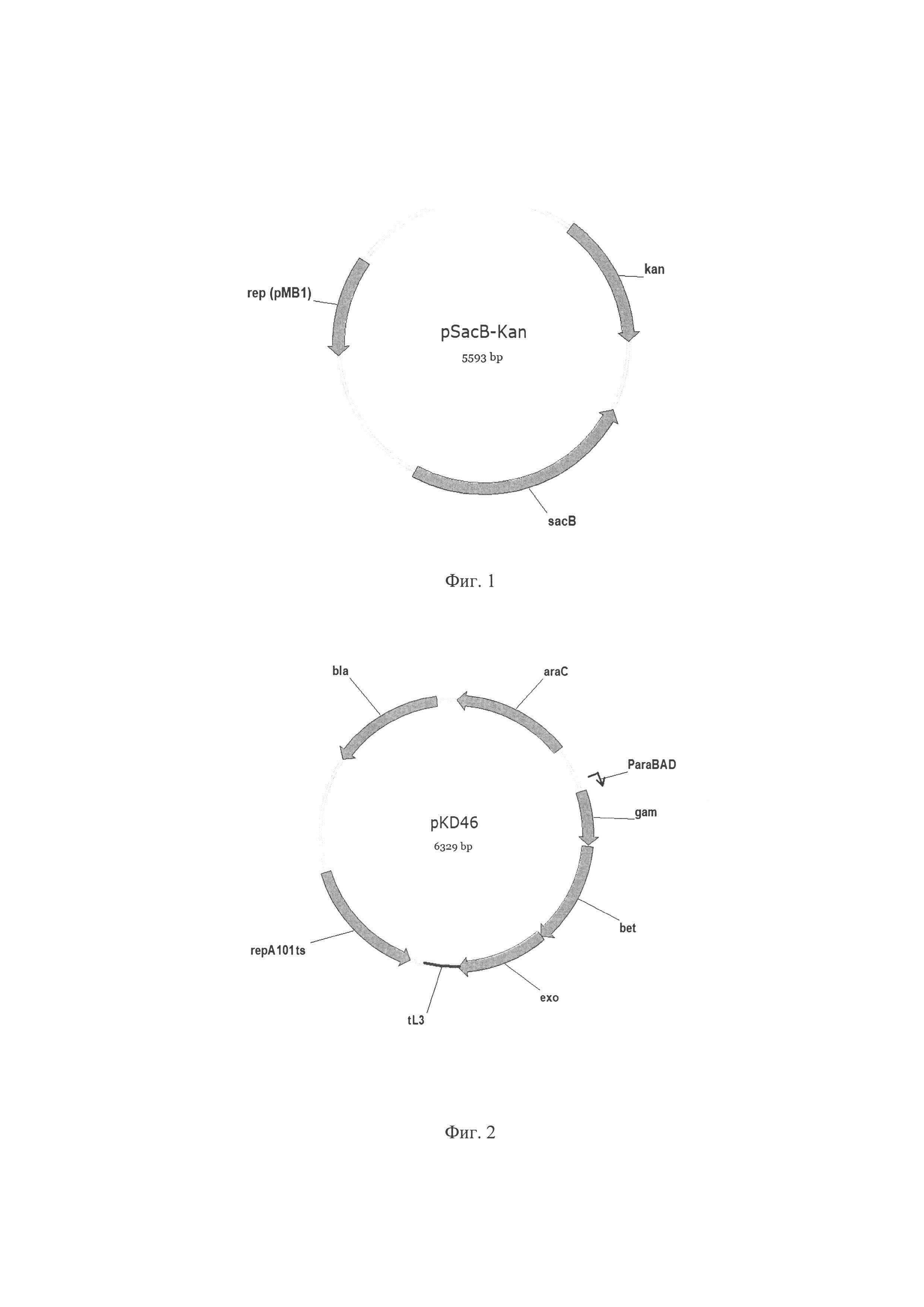
Фиг. 1. Схема плазмиды pSacB-Kan, где kan - ген kan (из Tn903), кодирует аминогликозид-3'-фосфотрансферазу, обуславливает устойчивость к канамицину; sacB - ген sacB, кодирует левансукразу Bacillus subtilis; rep (pMB1) - репликон плазмиды pMB1.
Фиг. 2. Схема плазмиды pKD46, где araC - ген araC регулятор транскрипции генов катаболизма L-арабинозы; ParaBAD - промотор оперона araBAD, индуцируется L-арабинозой; gam, bet, exo - гены gam, bet, exo Red системы гомологичной рекомбинации фага λ, repA101ts - температурочувствительный репликон; tL3 - терминатор транскрипции tL3; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину.
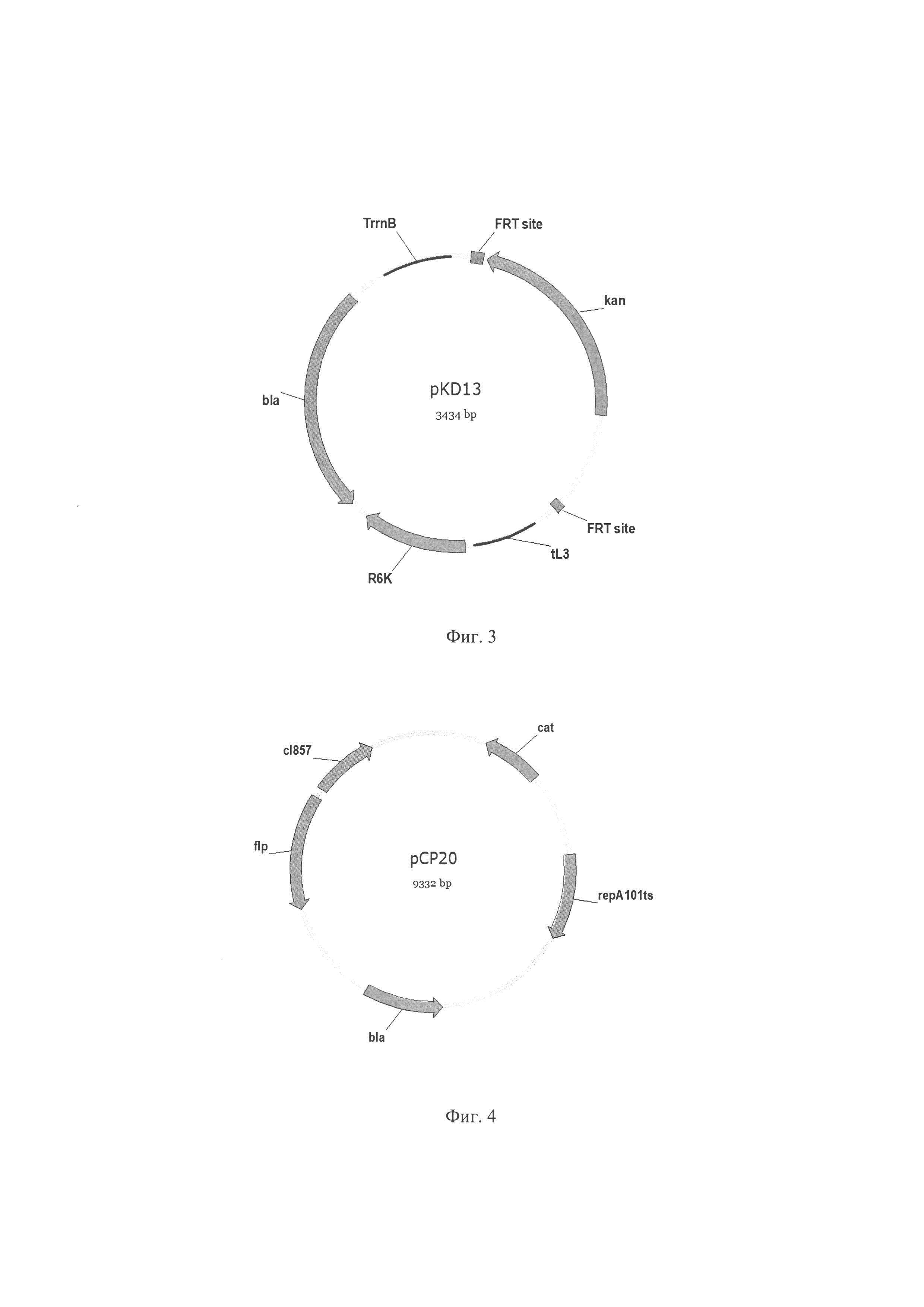
Фиг. 3. Схема плазмиды pKD13, где kan - ген kan (из Tn5) кодирует неомицинфосфотрансферазу, обуславливающую устойчивость к канамицину; FRT site - сайты FLP-FRT системы сайт-специфической рекомбинации S. cerevisiae; tL3 - терминатор транскрипции tL3; R6K - репликон плазмиды pR6K; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину; TrrnB - терминатор транскрипции.
Фиг. 4. Схема плазмиды рСР20, где flp - ген flp, кодирует FLP рекомбиназу сайт-специфической системы рекомбинации S. cerevisiae; cI857 - cI термочувствительный репрессор фага λ, cat - ген cat, кодирует хлорамфениколацетилтрансферазу, обуславливает устойчивость к хлорамфениколу; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину; repA101ts - температурочувствительный репликон.
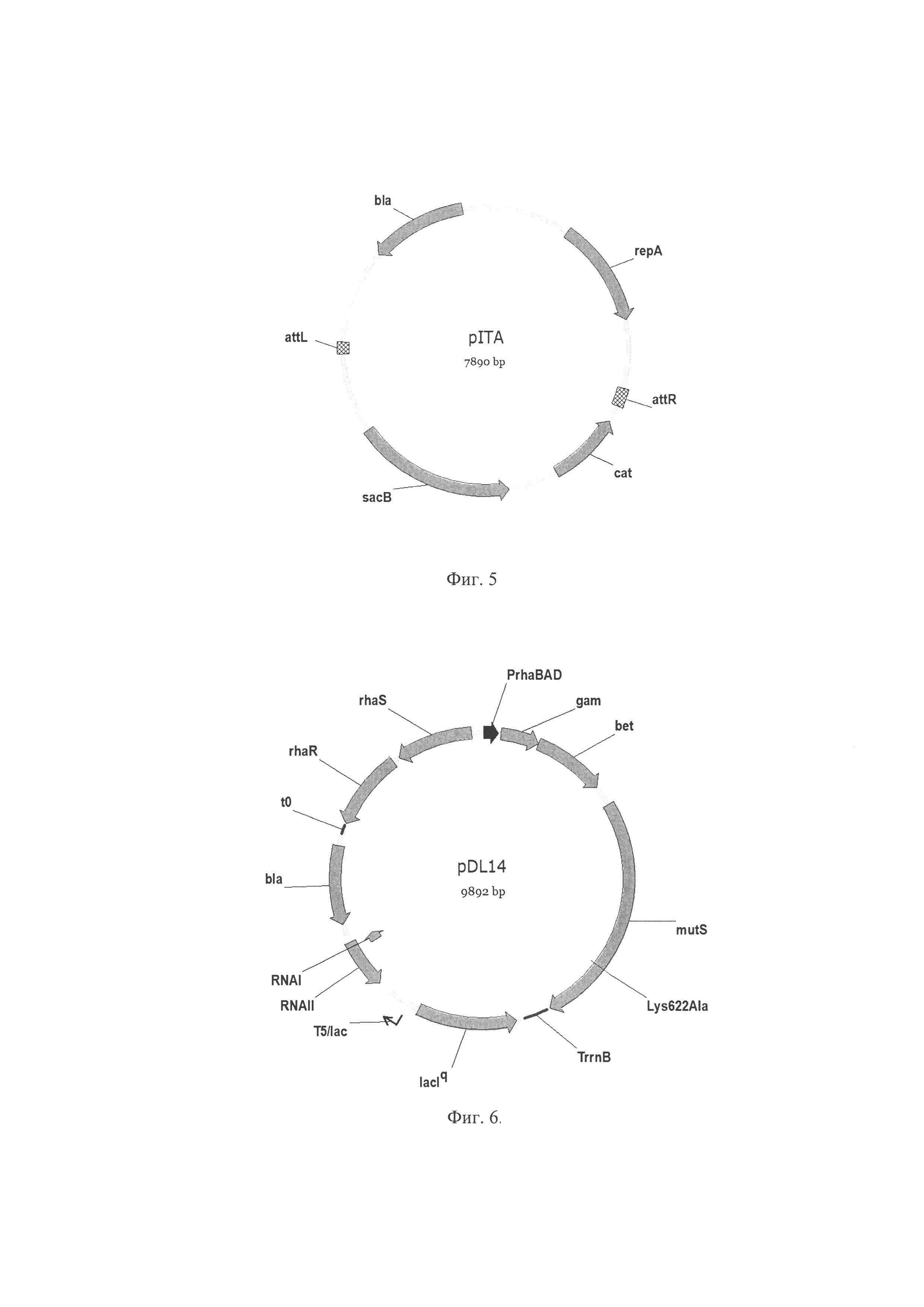
Фиг. 5. Схема плазмиды pITA, где attL, attR - attL (левый) и attR (правый) сайты унавания сайт-специфической рекомбинации фага λ; sacB - ген sacB, кодирует левансукразу Bacillus subtilis; cat - ген cat, кодирует хлорамфениколацетилтрансферазу, обуславливает устойчивость к хлорамфениколу, repA - репликон; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину.
Фиг. 6. Схема плазмиды pDL14 где rhaS, rhaR - гены rhaS, rhaR кодируют активаторы транскрипции оперона rhaBAD; t0 - терминатор транскрипции; PrhaBAD - промотор оперона rhaBAD, индуцируется L-рамнозой; gam, bet, exo - гены gam, bet, exo Red системы гомологичной рекомбинации фага λ; mutS - ген mutS Salmonella enterica, кодирует основной фермент системы репарации неканонических пар нуклеотидов, несет замену Lys622Ala; TrrnB - терминатор транскрипции; lacIq - мутантный аллель репрессора LacI, Т5/lac - сильный промотор Т5 с lac оператором; RNAI, RNAII - короткие транскрипты RNAI, RNAII (pBR322) контролирующие число копии плазмиды плазмиды; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину.
Фиг. 7. Схема плазмиды pKK-up-SC-dw-thrABC-433 где Ptac - промотор Ptac; thrA - ген thrA треонинового опрерона, кодирует аспартаткиназу / гомосериндегидрогеназу, несет мутацию Gly433Arg(G1297A); thrB - ген thrB треонинового опрерона, кодирует гомосеринкиназу; thrC - ген thrC треонинового опрерона, кодирует треонинсинтазу; TrrnB - терминатор транскрипции; up / down-thr-homology - фрагменты ДНК, гомологичные послетовательностям на хромосоме Е. coli выше и ниже треонинового оперона; rep (pMB1) - репликон плазмиды pMB1; attL, attR - attL (левый) и attR (правый) сайты унавания сайт-специфической рекомбинации фага λ; sacB - ген sacB, кодирует левансукразу Bacillus subtilis; cat - ген cat, кодирует хлорамфениколацетилтрансферазу, обуславливает устойчивость к хлорамфениколу, repA - репликон; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину.
Фиг. 8. Схема плазмиды pDL17, где rhaS, rhaR - гены rhaS, rhaR кодируют активаторы транскрипции оперона rhaBAD; t0 - терминатор транскрипции; PrhaBAD - промотор оперона rhaBAD, индуцируется L-рамнозой; gam, bet, exo - гены gam, bet, exo Red системы гомологичной рекомбинации фага λ; tL3 - терминатор транскрипции; TrrnB - терминатор транскрипции; lacIq - мутантный аллель репрессора LacI, Т5/lac - сильный промотор Т5 с lac оператором; RNAI, RNAII - короткие транскрипты RNAI, RNAII (pBR322) контролирующие число копии плазмиды плазмиды; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину.
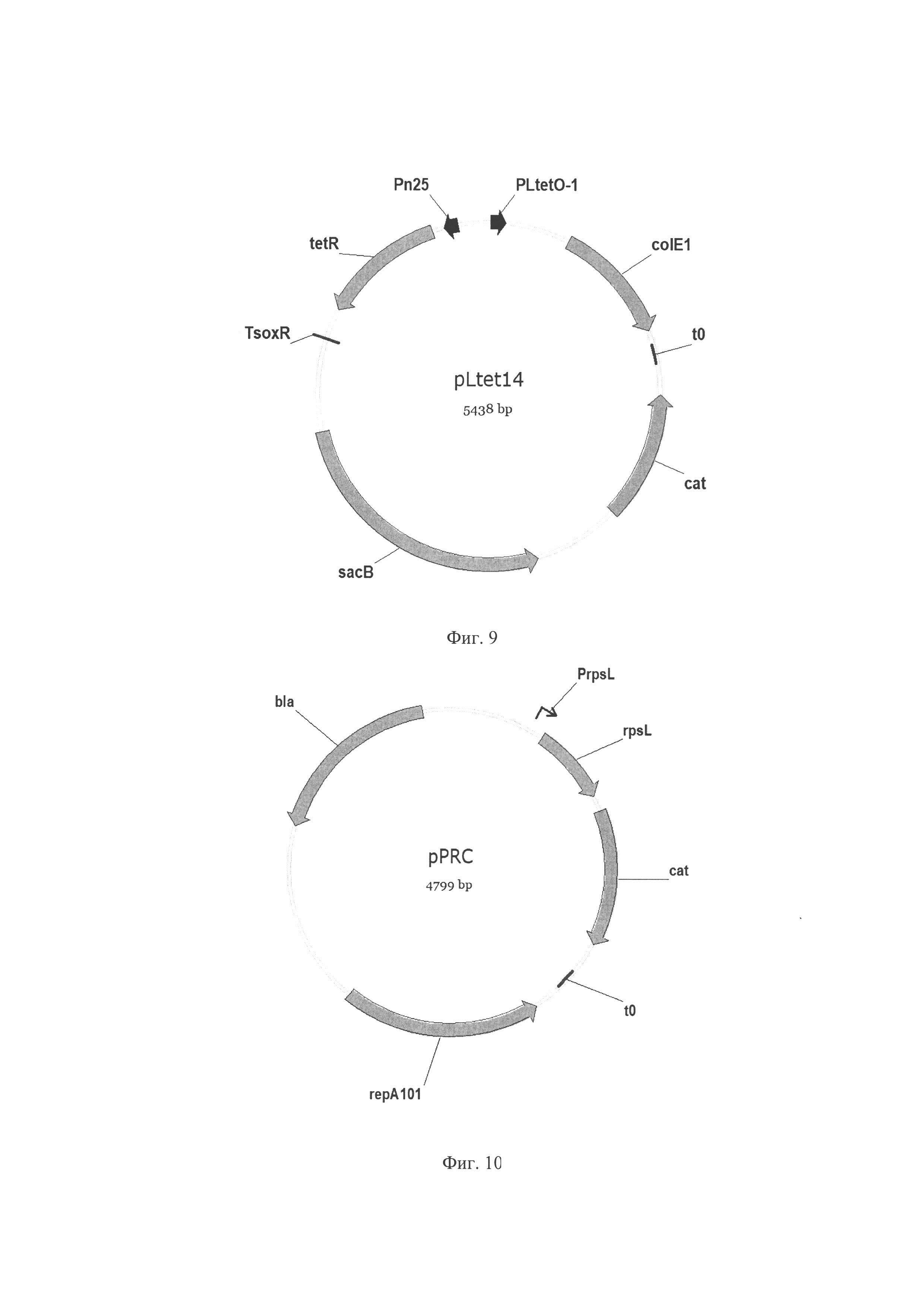
Фиг. 9. Схема плазмиды pLtet14 где PLtetO-1 - регулируемый промотор, индуцируется ангидротерациклином; PN25 - промотор; terR - репрессор; TsoxR - терминатор транскрипции; colE1 - репликон colE1; t0 - терминатор транскрипции; sacB - ген sacB, кодирует левансукразу Bacillus subtilis; cat - ген cat, кодирует хлорамфениколацетилтрансферазу, обуславливает устойчивость к хлорамфениколу.
Фиг. 10. Схема плазмиды pPRC, где PrpsL - промотор гена rpsL; rpsL - ген rpsL кодирует S12 белок рибосомной субчастицы 30S, мутация в гене rpsL приводит к рецессивному фенотипу устойчивости к стрептомицину; cat - ген cat, кодирует хлорамфениколацетилтрансферазу, обуславливает устойчивость к хлорамфениколу; t0 - терминатор транскрипции; repA101ts - репликон; bla - ген bla, кодирует бета-лактамазу, обуславливающую устойчивость к ампициллину.
Изобретение подтверждено следующими примерами.
Пример 1. Конструирование заявляемого штамма
1.1 Удаление мутации rph-1 в гене rph
Имеющуюся в штамме Escherichia coli MG1655 ВКПМ В-13106 мутацию rph-1 ранее рассматривали как несущественную. Однако впоследствии выяснилось, что она негативно влияет на рост бактерий в определенных условиях. Удаление мутации rph-1 в гене rph (номер последовательности по базе данных GenBank NC_000913.3 U00096.3: 3815863…3816549) в штамме MG1655 выполняют в два этапа.
На первом этапе заменяют последовательность мутантного гена на последовательность, несущую два маркера прямой и обратной селекции, ген устойчивости к канамицину и левансукразу, кодируемую sacB. На втором - после подтверждения генотипа замещают указанную последовательность диким геном rph.
Конструкцию для введения в локус rph гена устойчивости к канамицину и маркера негативной селекции sacB получают методом ПЦР, в качестве матрицы используют плазмиду pSacB-Kan (RU 2546237), представленную на фиг. 1. Амплификацию проводят по праймерам:
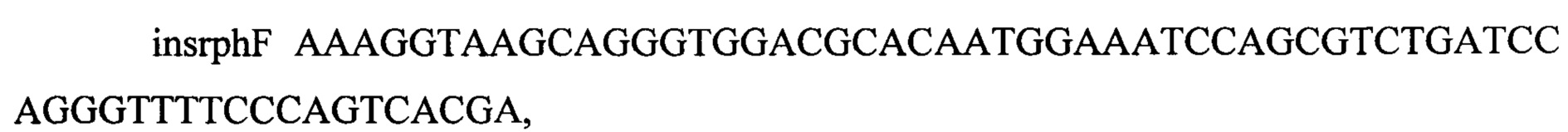
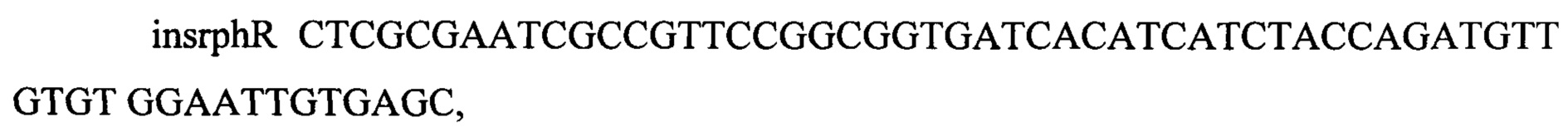
содержащим последовательности, гомологичные участкам в локусе rph длиной 45 и 43 нуклеотида соответственно, и последовательности, гомологичные участкам плазмиды, фланкирующим гены kan и sacB длиной 20 и 21 нуклеотид соответственно.
Введение амлифицированной кассеты в мутантный ген rph штамма MG1655 осуществляют по методике основанной на Red-зависимой рекомбинации фага λ (Datsenko K.A., Wanner В.L., Proceedings of the National Academy of Sciences. - 2000. - T. 97. - №. 12. - C. 6640-6645.). Согласно методике для интеграции фрагмента ДНК в хромосому штамм MG1655 трансформируют плазмидой pKD46 (фиг. 2). Плазмида pKD46 необходима для интеграции продукта ПЦР в хромосому штамма MG1655. Электрокомпетентные клетки штамма MG1655, несущего плазмиду pKD46, наращивают при 30°С в среде SOB (% мас, объем: триптон - 2; дрожжевой экстракт - 0,5; NaCl - 0,0585; MgSO4⋅7H2O - 0,246) с добавлением 1 мМ L-арабинозы. Клетки трасформируют методом электропорации. Трасформанты отбирают на чашках с L-агаром (% мас, объем: триптон - 1; дрожжевой экстракт - 0,5; NaCl - 1, агар-агар - 2, вода - остальное) по устойчивости к канамицину (100 мкг/мл). Генотип полученных трансформантов подтверждают методом ПЦР с использованием синтетических олигонуклеотидов
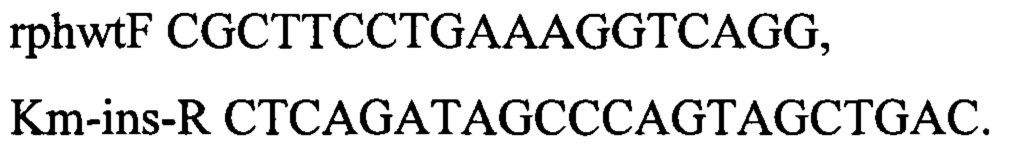
С хромосомы штамма Е. coli DH10B амплифицируют последовательность дикого гена rph, используя синтетические олигонуклеотиды
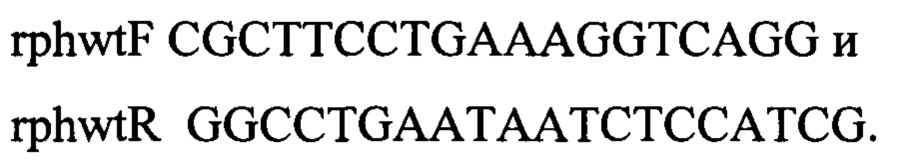
Полученным ПЦР-продуктом длиной 1063 п.н. трасформируют компетентные клетки штамма MG1655 Δrph::KmR-sacB, несущего плазмиду pKD46 (фиг. 2). Трасформанты отбирают на чашках со средой SuLA (% мас, объем: триптон - 1; дрожжевой экстракт - 0,5; сахароза - 6; агар-агар - 2; вода - остальное). Трансформанты проверяют методом ПЦР на корректность встройки. Один из полученных клонов с подтвержденным генотипом обозначают, как MG1655thpwt.
1.2 Деления генов tdcBCDE
Гены tdcBCDE (номер последовательности по базе данных GenBank NC_000913.3 U00096.3: 3266028…3260124) входят в состав оперона, отвечающего за транспорт L-треонина внутрь клетки, а также за деградацию L-треонина в анаэробных условиях. Инактивацию гена выполняют по коротким гомологичным участкам с использованием системы, основанной на Red-зависимой рекомбинации фага λ. Селективный маркер, ген устойчивости к канамицину, фланкированный FRT-сайтами, амплифицируют методом ПЦР, в качестве матрицы для синтеза используют плазмиду pKD13 (Datsenko K., et al, Proc natl Acad Sci, 2000), представленную на фиг. 2. Для специфической амплификации используют олигонуклеотиды:
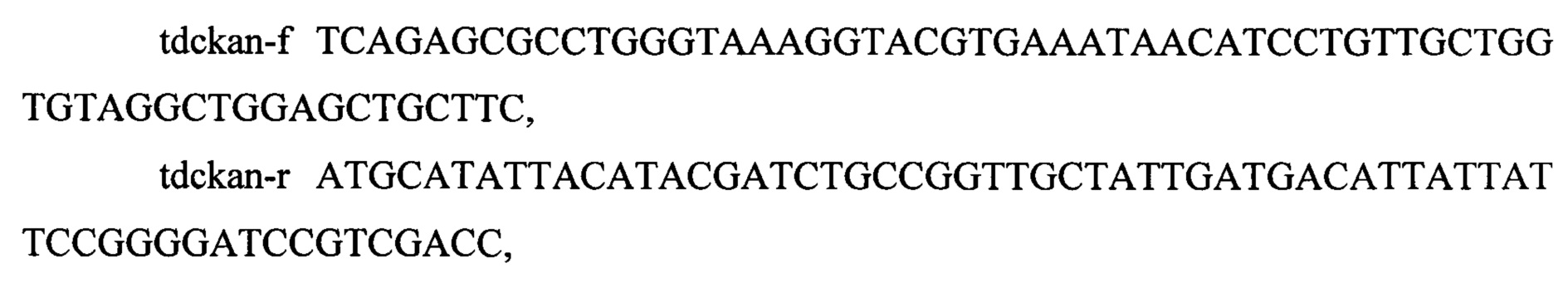
содержащие участки по 40 н.о, гомологичные участкам, расположенным выше и ниже генов tdcB и tdcE. Полученный ПЦР продукт размером 1394 п.н. содержит ген устойчивости к канамицину, фланкированный сайтами FRT рекомбиназы и гомологичными участками длинной 40 п. о выше и ниже генов tdcB и tdcE.
Штамм MG1655rphwt, трансфомируют плазмидой pKD46 (фиг. 2). Трасформанты отбирают на чашках с L-агаром с добавленим ампициллина 250 мкг/мл. Из одного трасформанта наращивают компетентные клетки при 30°С в среде SOB с добавкой 15 мМ арабинозы. Компетентные клетки трасформируют методом элетропорации очищенным ПЦР продуктом для инактивации генов tdcBCDE. Трансформанты отбирают на чашках с L-агаром с добавлением канамицина (100 мкг/мл) и проверяют на наличие мутации по генам tdcBCDE методом ПЦР с использованием праймеров
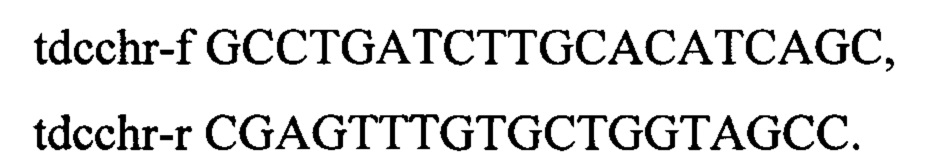
В результате получен штамм с генотипом rphwt, ΔtdcBCDE::FRT-KmR-FRT.
Маркер устойчивости к канамицину удаляют путем применения сайт-специфической рекомбинации с использованием рекомбиназы Flp. Для этой цели штамм с генотипом rphwt, ΔtdcBCDE:: FRT-KmR-FRT трансформируют плазмидой рСР20 (Cherepanov P.P., et. Al., Gene, 1995), схема которой предствлена на фиг. 4. Трансформанты отбирают при 30°С на чашках с L-агаром сдобавлением 250 мкг/мл ампициллина. Для элиминации плазмиды рСР20, которая несет термочувствительный репликон штамм культивируют при температуре 37°С. Отсутствие маркера подтверждают по отсутствию устойчивости к канамицину и по наличию ПЦР продукта размером 819 п.о. по праймерам
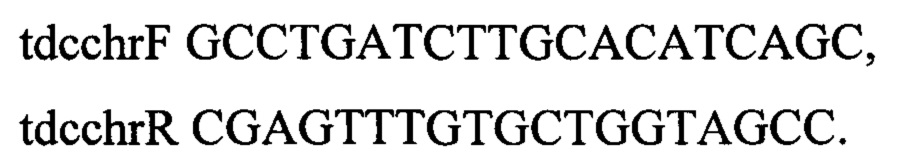
В результате отбирают один из клонов с подтвержденным генотипом rphwt, ΔtdcBCDE, штамм обозначают как MG-THR1.2
1.3 Деления генов kbl-tdh
Инактивацию генов kbl-tdh, находящихся в хромосоме Е. coli в одном опероне (номер последовательности по базе данных GenBank NC_000913.3 U00096.3:3790320…3792551) проводят путем замены нуклеотидной последовательности генов на кассету attR-cat-sacB-attL, содержащую селективные маркеры устойчивости к хлорамфениколу и левансукразу. Кассету амплифицируют методом ПЦР с плазмиды, pMW-attL-SacB-Cm-attR (RU 2546237) (далее pITA, фиг. 5) по праймерам
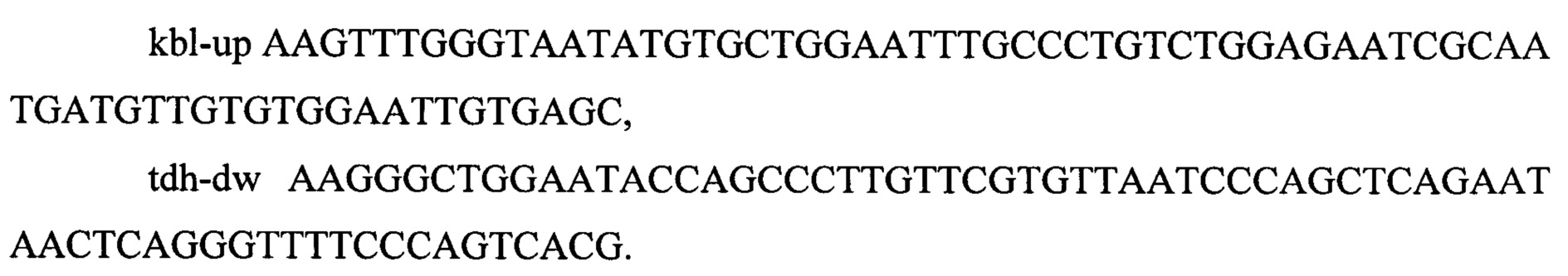
ПЦР продукт размером размером 3944 п.о. очищают методом экстракции из агарозного геля и далее используют для трансформации штамма MG-THR1.2, который предварительно трансформируют плазмидой pKD46 (фиг. 2). Отбор трансформантов, несущих делецию в генов kbl-tdh проводят на чашках с L-агаром с добавлением хлорамфеникола (25 мкг/мл). Отобранные клоны далее проверяют методом ПЦР на наличие инсерции в локусе kbl-tdh, используя олигонуклеотиды
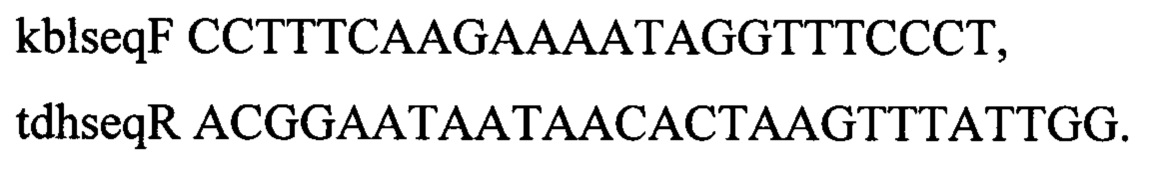
Для вырезания маркеров из локуса kbl-tdh хромосомы штамма MG1655 амплифицируют два фрагмента размером 243 п.о. и 245 п.о. по двум парам праймеров kblupF и kblupR, а также tdhdwR и tdhdwF:

Оба полученных ПЦР-продукта используют в качестве матрицы в следующей ПЦР по праймерам kblupF и tdhdwR. Полученным продуктом 447 п.о. трансформируют штамм с делецией Δkbl-tdh::attR-cat-sacB-attL, содержащий плазмиду pKD46. Трансформанты отбирают на среде с SuLA при 37°С. Вырезание маркера подтверждают методом ПЦР по праймерам  и kblupF. Потерю плазмиду pKD46 регистрируют по отсутствию роста на L-агаре с ампициллином 250 мкг/мл. В результате отобран штамм MG-THR1.3 с rphwt, ΔtdcBCDE, Δkbl-tdh.
и kblupF. Потерю плазмиду pKD46 регистрируют по отсутствию роста на L-агаре с ампициллином 250 мкг/мл. В результате отобран штамм MG-THR1.3 с rphwt, ΔtdcBCDE, Δkbl-tdh.
1.4. Деления гена sstT
Делецию гена sstT (номер последовательности по базе данных GenBank NC_000913.3 U00096.3: 3266028…326) проводят путем замены нуклеотидной последовательности гена на кассету attR-cat-sacB-attL, содержащую селективные маркеры устойчивости к хлорамфениколу и левансукразу. Кассету размером 3946 п.о. амплифицируют методом ПЦР с плазмиды pITA (фиг. 5) по праймерам:
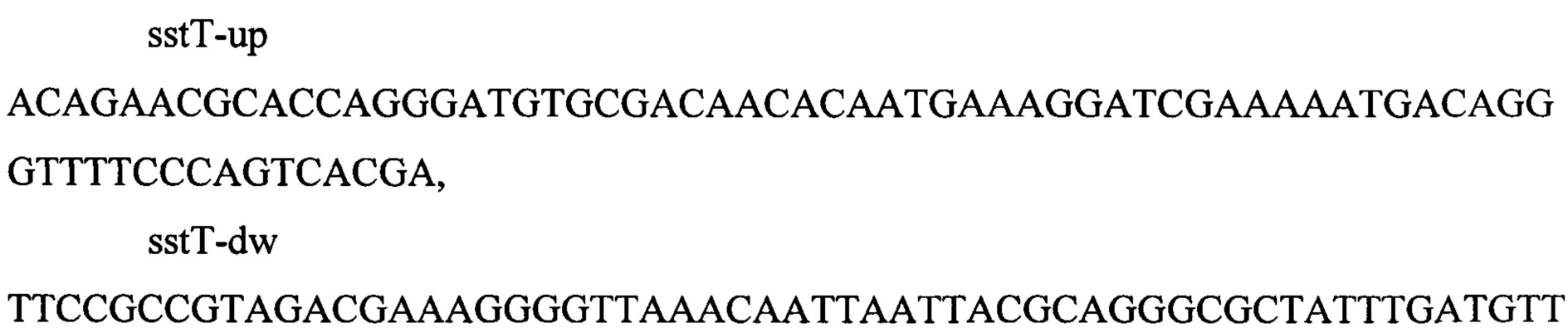
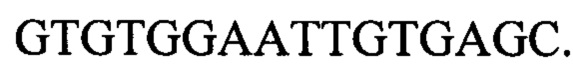 Штамм MG-THR1.3 трансформируют плазмидой pKD46 (фиг. 2). Из полученных трансформантов наращивают компетентные клетки при 30°С с индукцией арабинозой. Компетентные клетки трансформируют кассетой для инактивации гена sstT, трансформанты отбирают на чашках с L-агаром с добавлением хлорамфеникола (25 мкг/мл) и проверяют методом ПЦР на наличие целевой вставки. Для амплификации фрагмента используют следующие олигонуклеотиды
Штамм MG-THR1.3 трансформируют плазмидой pKD46 (фиг. 2). Из полученных трансформантов наращивают компетентные клетки при 30°С с индукцией арабинозой. Компетентные клетки трансформируют кассетой для инактивации гена sstT, трансформанты отбирают на чашках с L-агаром с добавлением хлорамфеникола (25 мкг/мл) и проверяют методом ПЦР на наличие целевой вставки. Для амплификации фрагмента используют следующие олигонуклеотиды
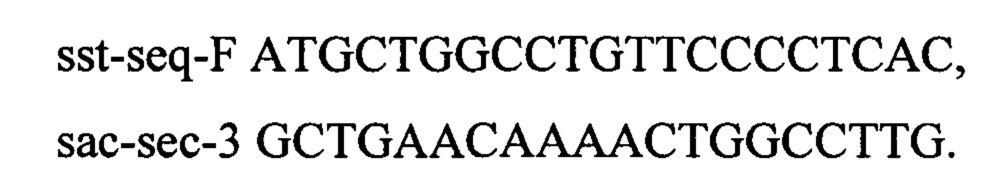
В результате получают штамм с генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT::attR-cat-sacB-attL.
Для вырезания маркеров из локуса sstT полученный штамм с делецией ΔsstT::attR-cat-sacB-attL, несущий плазмиду pKD46 (фиг. 2) трансформируют ПЦР продуктом sstT-up-down размером 522 п.о. Для получения такого продукта, используя хромосому штамма MG1655, амплифицируют два фрагмента размерами 245 п.о. и 243 п.о. по двум парам праймеров sstTupR, sstTupF и sstTdwF, sstTdwR, соответственно.
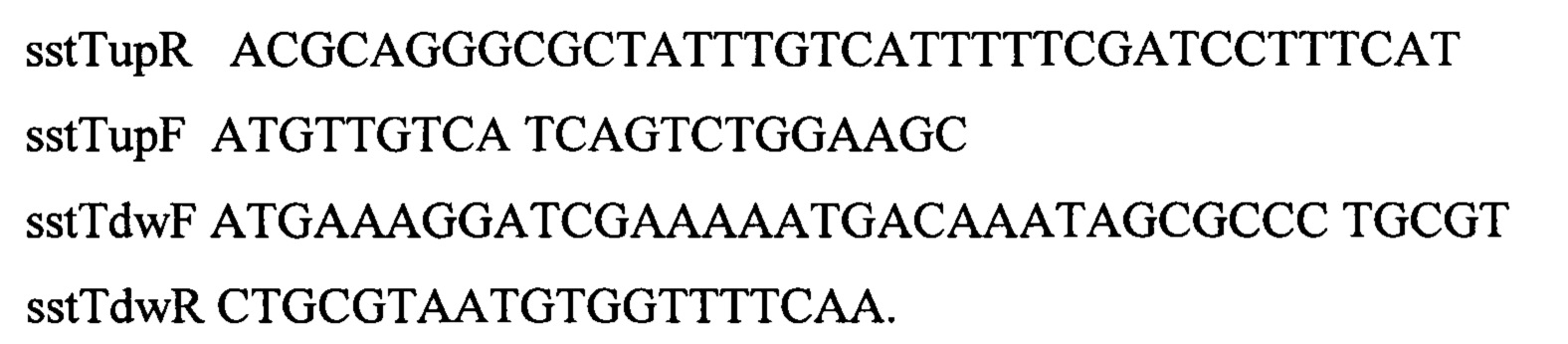
Оба полученных ПЦР-продукта используют в качестве матрицы в следующей ПЦР по праймерам sstTupF и sstTdwR для получения продукта sstT-up-down. После трансформации клетки переносят в пробирку объемом 20 мл с 2 мл рабочего раствора среды SOB. Пробирки инкубируют на качалке при 37°С со скоростью перемешивания 200 об/мин в течение 24 часов. После инкубирования различные разведения культуральной жидкости высевают на чашки с L-агаром. Чашки инкубируют в течение 20 часов при 37°С. Полученные клоны рассевают на чашки с L-агаром, содержащим хлорамфеникол (25 мкг/мл), с добавлением ампициллина (250 мкг/мл) или без добавления антибиотиков. После инкубирования в течение 20 часов при 37°С отбирают клоны с отсутствием устойчивости к ампициллину и хлорамфениколу. Отобранные клоны далее проверяют методом ПЦР на наличие делеции по гену sstT. Для амплификации фрагмента используют следующие олигонуклеотиды sst-seq-F и sstTseqR. Отбирают один клон с подтвержденным генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT. Штамм обозначают как MG-THR1.4.
1.5 Получение мутаций rpsLK43R, galKTyr38TAG. araBTyr57TAG. rhaBTyr163TAG
Полученный штамм MG-THR1.4, трансформируют плазмидой pDL14 (Bubnov D.М. et al, Journal of microbiological methods. - 2018. - T. 151. - C. 48-56.), схема которой представлена на Фиг. 6. Трансформантов отбирают на L-агаре по устойчивости к ампициллину (250 мкг/мл). Из одного трансформанта наращивают компетентные клетки, которые трансформируют смесью олигонуклеотидов
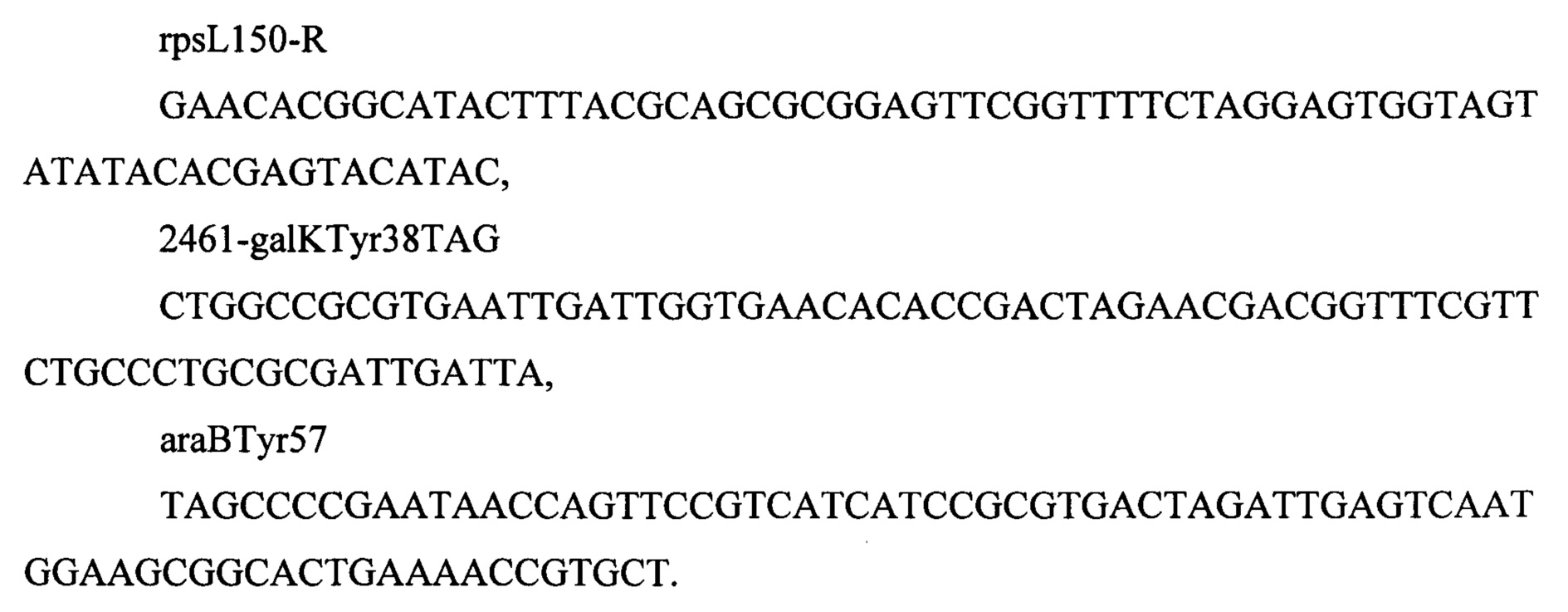
Трансформанты отбирают на среде Эндо (производства Himedia, Индия, кат. номер М029) с добавлением 2 г/л галактозы, 2 г/л арабинозы и 250 мкг/мл стрептомицина. Рекомбинанты отбирают по соответствующей окраске. Наличие мутаций во всех трех генах подтверждают секвинированием. Плазмиду pDL14 удаляют из штамма путем двойного рассева до отдельной колонии на чашках с L-агром с добавлением 1 мМ ИПТГ (Изопропил-β-D-1-тиогалактопиранозид). В результате отобран штамм с генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG.
Затем в полученный штамм вводят мутацию rhaBY163Am. Для этого его предварительно трансформируют плазмидой pDL14 (фиг. 6). Электрокомпетентные клетки, несущие плазмиду DL14, трасформируют олигонуклеотидом
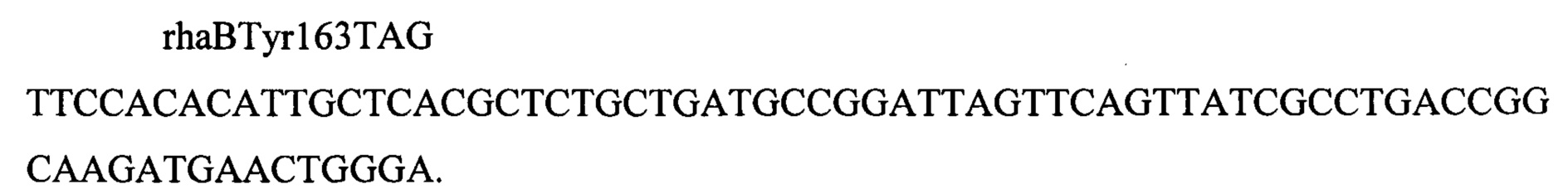
Трансформанты отбирают на среде Эндо (производства Himedia, Индия, кат. номер М029) с добавлением 2 г/л рамнозы. Наличие мутации подтверждают секвинированием. Плазмиду pDL14 удаляют рассевом на L-агаре с 1 мМ ИПТГ. Полученный штамм с генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG обозначают как MG-THR1.5.
1.6 Экспрессия оперона thrABC с десенсибилизирующей мутацией в гене thrA под регуляцией промотора PH-207
Экспрессию генов треонинового оперона с десенсибилизирующей мутацией в штамме MG-THR1.5 проводят в два этапа. На первом этапе в хромосому в локус thrABC (номер последовательности по базе данных GenBank NC_000913.3 U00096.3: 377…5020) интегрируют конструкцию, которая заменяет нативный промотор оперона thrABC на фаговый промотор Ptac, удаляет последовательность регуляторного пептида ThrL и вводит точечную мутацию, вызывающую замену Gly433Arg в аминокислотной последовательности ThrA, природный терминатор оперона заменяют терминатором TrrnB. На втором этапе вырезают селективные маркеры из хромосомы и заменяют промотор Ptac на промотор PH-207.
Интегративную конструкцию получают путем рестрикции по Eco72I плазмиды pKK-up-SC-dw-thrABC-433 (RU 2546237), схема которой представлена на фиг. 7. Полученный после рестрикции фрагмент несет в своем составе модифицированный треониновый оперон, селективные маркеры cat и sacB, а также области гомологии к хромосоме выше и ниже thrABC. Полученным фрагментом ДНК размером 9882 п.о. трансформируют компетентные клетки штамма MG-THR1.5, несущего плазмиду pDL17 (Bubnov D.М. et al, Journal of microbiological methods. - 2018. - T. 151. - C. 48-56.) (фиг. 8). Трансформанты отбирают на чашках с L-агаром в присутствии хлорамфеникола (25 мкг/мл). Плазмиду pDL17 удаляют путем культивирования штамма на чашках с L-агаром, содержащим 1 мМ ИПТГ. Наличие модификации в нужном локусе подтверждают по наличию ПЦР-продукта размером 1096 п.о. по праймерам
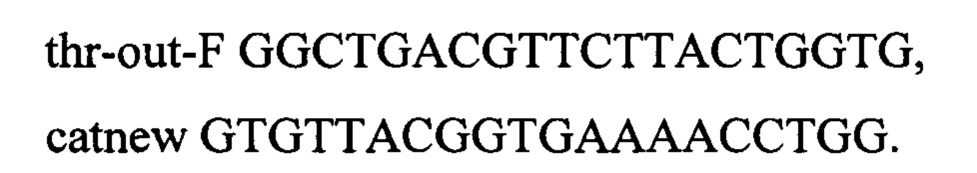
Один из полученных таким образом штаммов имеет генотип rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG, attSC-Ptac-thrA433BC-TrrnB.
Для того чтобы удалить селективные маркеры, отобранный штамм повторно трансформируют плазмидой pDL17, как описано выше. Затем наращивают компетентные клетки и трансформируют фрагментом 718 п.о., полученным после ПЦР по праймерам
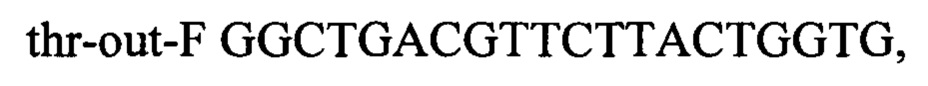
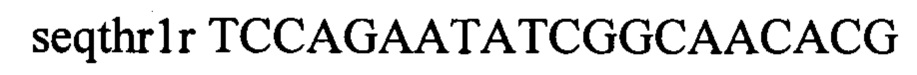
В качестве матрицы ПЦР используют хромосому штамма Escherichia coli ВКПМ В-13207 (RU 2697499), несущего промотор PH207 перед треониновым опероном. Плазмиду pDL17 удаляют путем культивирования на L-агаре с 1 мМ ИПТГ. Один из полученных таким образом штаммов с генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG, PH207-thrA433BC-TrrnB обозначен как MG-THR1.6.
1.7 Экспрессия гена rhtA под регуляцией промотора P1107PR
Экспрессию гена rhtA (номер последовательности по базе данных GenBank NC_000913.3 U00096.3: 849210…850097) под промотором P1107PR проводят в два этапа: 1) заменяют нативный промотор PrhtA кассетой, несущей маркеры cat, sacB, репрессор tetR и промотор PLtetO-1; 2) вырезают маркеры из хромосомы, заменяя регулируемый промотор PLtetO-1 на конститутивный P1107PR.
Компетентные клетки Штамма MG-THR1.6, трансформируют плазмидой pDL17 (фиг. 8). Трансформанты отбирают на L-агаре с ампициллином 250 мкг/мл, и трансформируют ПЦР продуктом 4531 п.о. полученным амплификацией с плазмиды pLtet14, представленной на фиг. 9 (Выборная Т.В. и др., Биотехнология. - 2019. - Т. 35. - №. 4. - С. 42-54.) по праймерам
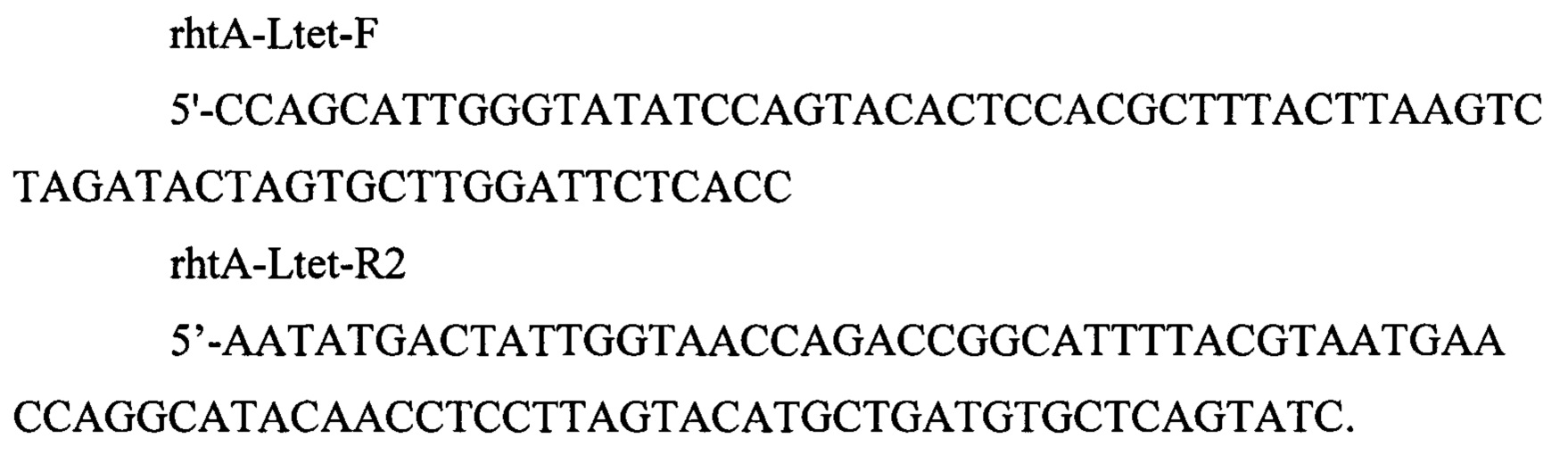
Трансформанты отбирают на L-агаре с добавлением хлорамфеникола (25 мкг/мл). Плазмиду pDL17 удаляют путем культивирования на L-агаре с 1 мМ ИПТГ. Наличие модификации в нужном локусе подтверждают по наличию ПЦР-продукта размером 839 п.о. по праймерам:
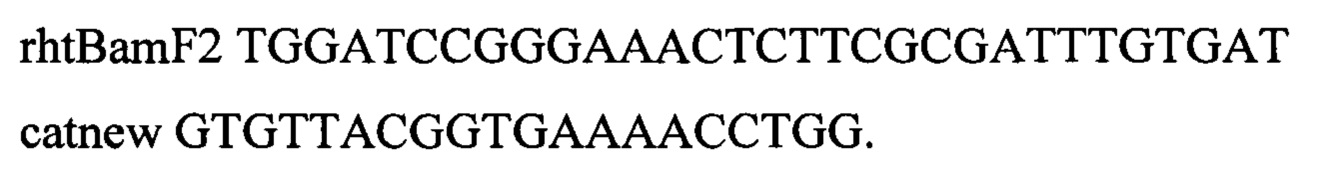
Последовательности гена rhtA и промотора PLtetO-1 были подтверждают полностью методом секвенирования. Один из клонов с подтверженным генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG, PH207-thrA433BC-TrrnB, SC-tetR-PLtetO-1-rhtA используют для дальнейшей работы.
Отобранный штамм повторно трасформируют плазмидой pDL17 (фиг. 8), затем компентеные клетки, несущие pDL17, трасформируют фрагментом 803 п.о., полученным после ПЦР по праймерам

В качестве матрицы ПЦР используют хромосому штамма Escherichia coli ВКПМ В-13207 (RU 2697499), несущего промотор P1107PR перед геном rhtA. Трансформантов отбирают на чашках со средой SuLA и проверяют методом секвенирования на наличие последовательности промотора P1107PR перед геном rhtA. Один из отобранных трансформантов обозначен MG-THR1.7, он имеет генотип rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG, PH207-thrA433BC-TrrnB, P1077PR-rhtA
1.8 Получение мутаций spoTT252ins(CATGAT)253A; G520T; G1585T
Штамм продуцент L-треонина E.coli ВКПМ B-3996 несет в геноме мутантный ген spoT (номер последовательности по базе данных GenBank NC_000913.3 U00096.3: 3822400…3824508): инсерция CATGAT с. 252_253, замена Cys176Gly, замена Phe531Leu. Известно, что указанные мутации увеличивают продукцию аминокислот в штаммах Е. coli. Перенос мутаций из ВКПМ В-3996 в разрабатываемый штамм продуцент осуществляют в несколько стадий: внесение маркеров прямой и обратной селекции выше гена spoT в штамме ВКПМ В-3996; амлификация кассеты с полученного штамма, включая маркеры и ген мутантный ген spoT; трансформация полученной кассетой штамма MG-THR1.7 с последующим удалением маркеров.
Штамм ВКПМ В-3996 трансформируют плазмидой pDL17 (фиг. 8), отбирают трансформанты на L-агаре с добавлением ампициллина (250 мкг/мл.) Компетентные клетки штамма ВКПМ В-3996, несущего pDL17 трансформируют ПЦР продуктом, полученным в результате амплификации по праймерам
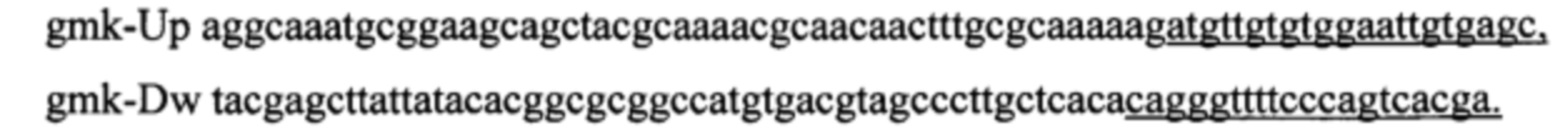
В качестве матрицы используют плазмиду pPRC (фиг. 10). В своем составе плазмида несет два маркера прямой и обратной селекции: ген cat, обуславливающий устойчивость к хлорамфениколу, и ген rpsL, мутация в котором обуславливает устойчивость к стрептомицину. В результате трансформации отбирают клоны, устойчивые к хлорамфениколу. Один из клонов с подтвержденным генотипом rpsL-cat-gmK-spoTmut испольуют в качетстве матрицы при амплификации продукта по праймерам:
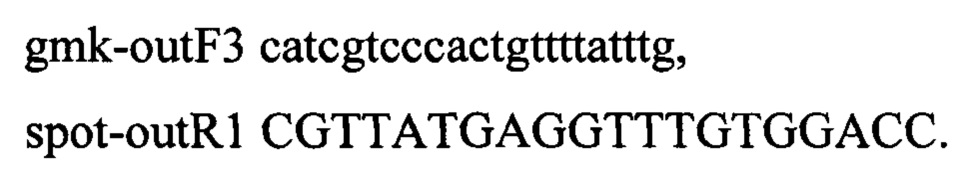
Амплифирированным продуктом размером 5134 п.о. трансформируют компетентные клетки штамма MG-THR1.7, несущие плазмиду pDL17. Трансформанты отбирают по устойчивости к хлорамфениколу. Наличие целевой вставки подтверждают амплификацией по специфическим праймерам, наличие мутаций подтверждают методом секвенирования. В результате отбирают один клон с подтвержденным генотипом. Не теряя плазмиду pDL17, культуру клеток трансформируют ПЦР продуктом для вырезания селективных маркеров. Для удаления маркеров амплифицируют фрагмент ДНК с хромосомы MG1655 по праймерам

Трансформанты отбирают на чашках с L-агаром с добавлением стрептомицина (250 мкг/мл). Элиминацию плазмиды pDL17 проводят пересевом отобранных трансформантов на среде с добавлением 1 мМ ИПТГ. В результате работы отбирают 8 клонов с генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG, PH207-thrA433BC-TrrnB, P1077PR-rhtA, spoTT252ins(CATGAT) 253A; G520T; C1585T.
Пример 2. Синтез L-треонина заявляемым штаммом в пробирках
Для оценки биотехнологического потенциала клонов, полученных на этапе 1.7, проводят ферментацию в пробирках. В качестве контрольного штамма используют штамм MG-THR1.6, полученный на этапе 1.6 и базовый штамм MG1655.
Штаммы выращивают на чашках с L-агаром в течение 24 часов при 37°С. Для приготовления инокулята используют посевную среду следующего состава (мас. %)
|
В пробирки объемом 50 мл с рабочим объемом 5 мл посевной среды вносят биомассу клеток до стартового значения оптической плотности равной ОП660нм 0,1 ед. Пробирки инкубируют на роторной качалке в течение 5 часов при 37°С и скорости перемешивания 220 об/мин. Для основного процесса ферментации используют среду следующего состава (мас. %):
|
Растворы глюкозы, сульфата марганца и сульфата магния стерилизуют отдельно автоклавированием при 121°С в течение 40 мин. Навески СаСО3 по 40 мг стерилизуют в стеклянных пробирках автоклавированием при 121°С в течение 20 мин. рН доводят до значения 7,0. Раствор сульфата железа стерилизуют фильтрованием через мембрану диаметром пор 22 мкм.
Полученный инокулят вносят в пробирки объемом 50 мл с рабочим объемом 2 мл ферментационной среды до стартовой оптической плотности ОП660 нм 0,1 ед. Клетки культивируют в течение 24 часов при 37°С на роторной качалке - 220 об/мин.
После выращивания количество накопленного в среде L-треонина определяют с методом ВЭЖХ (Waters 2695, Alliance). Результаты ферментации восьми клонов приведены в табл. 1.
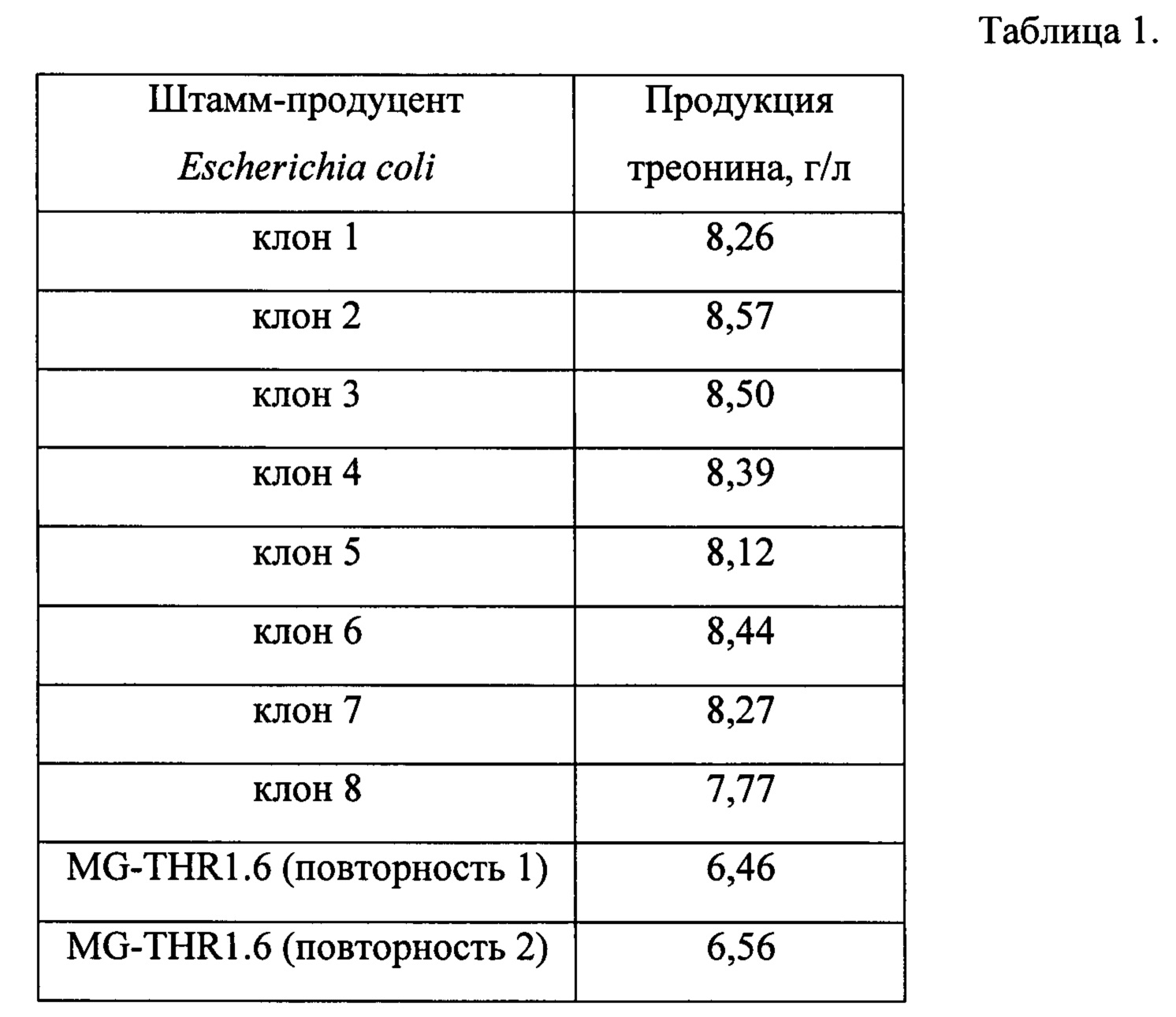

Как видно из табл. 1, клон 2, накапливает большее количество L-треонина по сравнению с родительским штаммом MG-THR1.6 и базовым штаммом MG1655. Отобранный клон 2 депонирован в Биоресурсном центре Всероссийская Коллекция Промышленных Микроорганизмов (БРЦ ВКПМ) НИЦ «Курчатовский институт» - ГосНИИгенетика как Escherichia coli ВКПМ В-13427.
Штамм Escherichia coli ВКПМ В-13427 характеризуется следующими признаками:
Культурально-морфологические характеристики заявляемого штамма
Грамм-отрицательная бактерия. Суточная культура в жидкой LB представлена слабо подвижными клетками округлой формы 1 мкм в диаметре. При культивировании на L-агаре в течение 18-24 часов при 37°С образует круглые, беловатый, полупрозрачные на свет колонии колонии 1 - 2 мм, поверхность колонии гладкая, края ровные или слегка волнистые, центр колоний приподнят, структура однородная, консистенция пастообразная, легко эмульгируется. При культивировании на агаризованной среде Эндо при 37°С образуются колонии, круглой формы с ровным четко очерченным краем малиново-красного цвета с металлическим блеском. Диаметр колоний 0,5-1,5 мм. При культивировании на тетразолиевом агаре с добавлением одного из сахаров: арабинозы, галактозы, рамнозы образуются красно-вишневые колонии.
Физиолого-биохимические характеристики
Факультативный анаэроб. Сахара не сбраживает. Ассимилирует: D-глюкозу, D-маннитол, D-ксилозу. Не ассимилирует: галактозу, лактозу, арабинозу и рамнозу. Отсутствует способность к гидролизу крахмала. Заявляемый штамм продуцирует L-треонин. Оптимальное значение рН для роста 7,2-7,4. Не растет при температурах свыше 45°С. Оптимальная температура роста 37°С. Штамм не патогенен.
Таким образом, сконструирован штамм Escherichia coli ВКПМ В-13427, обладающий следующим генотипом rphwt, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsLK43R, galKTyr38TAG, araBTyr57TAG, rhaBTyr163TAG, PH207-thrA433BC-TrrnB, P1077PR-rhtA, spoTT252ins(CATGAT) 253A; G520T; C1585TAG, способный синтезировать до 8,6 г/л L-треонина при культивировании в пробирках. Штамм Escherichia coli ВКПМ В-13427 получен из штамма дикого типа MG1655 путем введения направленных модификаций и не несет неизвестных мутаций. Штамм Escherichia coli ВКПМ В-13427 характеризуется высокой скоростью роста и скоростью синтеза L-треонина.
Штамм Escherichia coli ВКПМ В-13427 с генотипом rph, ΔtdcBCDE, Δkbl-tdh, ΔsstT, rpsL, galK, araB,rhaB, P-thrABC-T P-rhtA, spoT - продуцент L-треонина.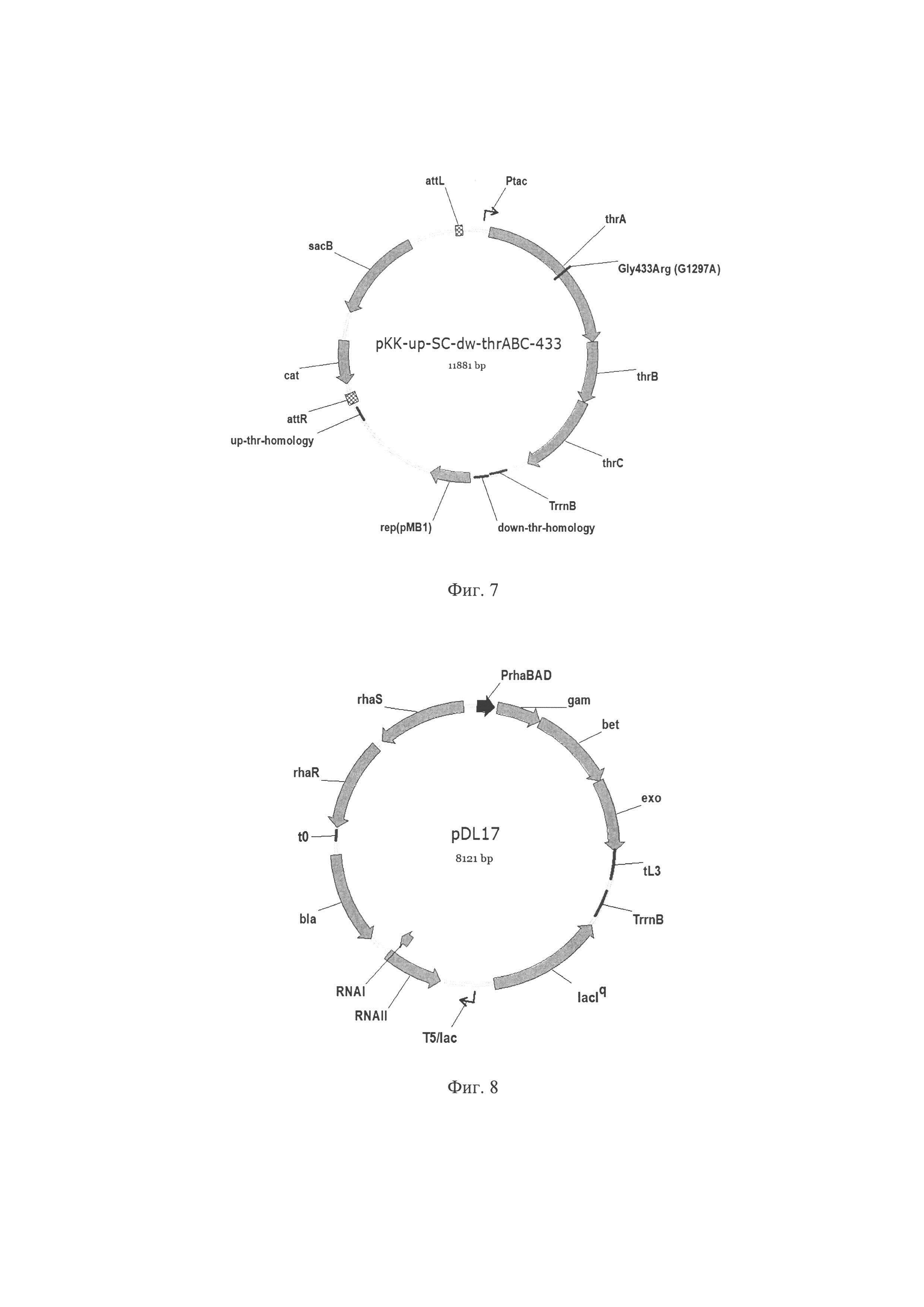
Применение полипептида, полученного на основе глюкагоноподобного пептида 1 человека, в качестве средства для контроля за весом тела
Штамм бактерий paenibacillus species - продуцент ксиланазы
Штамм бактерий paenibacillus species - продуцент β-глюканазы
Рекомбинантный штамм бактерии escherichia coli - продуцент l-треонина
Бактерия вида escherichia coli - продуцент l-треонина, способ микробиологического синтеза l-треонина с ее использованием.
Рекомбинантный штамм дрожжей pichia pastoris - продуцент ксиланазы
Штамм спорообразующих бактерий bacillus amyloliquefaciens, обладающий фунгицидным действием против фитопатогенных грибов, вызывающих заболевания овощных растений, биологический препарат на его основе
Рекомбинантный штамм дрожжей pichia pastoris - продуцент фитазы
Штамм дрожжей pichia pastoris - продуцент ксиланазы
Рекомбинантный штамм дрожжей komagataella kurtzmanii - продуцент бета-глюканазы
Рекомбинантный штамм escherichia coli-продуцент l-треонина
Трансформант дрожжей schizosaccharomyces pombe, продуцирующий молочную кислоту (варианты), способ его получения (варианты), способ микробиологического синтеза молочной кислоты с использованием такого трансформанта
Бесплазмидный рекомбинантный штамм escherichia coli, обладающий конститутивной аспартазной активностью и способ синтеза l-аспарагиновой кислоты с использованием этого штамма в качестве биокатализатора
Применение штамма мицелиального гриба aspergillus awamori в качестве реципиента для конструирования продуцентов целевых ферментов
Способ спектрофотометрического определения молочной кислоты
Штамм дрожжей yarrowia lipolytica - продуцент янтарной кислоты (варианты)
Штамм schizosaccharomyces pombe - продуцент молочной кислоты
Мутантная рекомбинантная гепариназа i с повышенной удельной активностью из pedobacter heparinus, фрагмент днк, кодирующий указанную гепариназу
Способ модификации дрожжей schizosaccharomyces pombe с помощью cre-lox системы бактериофага р1, трансформант, полученный таким способом
Способ изготовления интегральных элементов микросхем на эпитаксиальных структурах арсенида галлия

