Результат интеллектуальной деятельности: ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ФЛУОРЕСЦЕНТНЫЙ БИОСЕНСОР ДЛЯ ДЕТЕКЦИИ ПЕРОКСИДА ВОДОРОДА, КАССЕТА ЭКСПРЕССИИ, КЛЕТКА, ПРОДУЦИРУЮЩАЯ БИОСЕНСОР, ВЫДЕЛЕННЫЙ ФЛУОРЕСЦЕНТНЫЙ БИОСЕНСОР ДЛЯ ДЕТЕКЦИИ ПЕРОКСИДА ВОДОРОДА, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ФЛУОРЕСЦЕНТНЫЙ БИОСЕНСОР, ОПЕРАТИВНО СЛИТАЯ С НУКЛЕИНОВОЙ КИСЛОТОЙ, КОДИРУЮЩЕЙ СИГНАЛ ВНУТРИКЛЕТОЧНОЙ ЛОКАЛИЗАЦИИ
Вид РИД
Изобретение
Предлагаемое изобретение относится в основном к области биологии и химии. В частности, изобретение направлено на биосенсоры для детекции пероксида водорода, сконструированные на основе флуоресцентных белков.
Флуоресцентные белки семейства GFP (Green Fluorescent Protein, GFP), включая собственно GFP из медузы Aequorea victoria (avGFP), его мутанты и гомологи, сегодня широко известны благодаря их интенсивному использованию в качестве флуоресцентных маркеров in vivo в биомедицинских исследованиях, что детально рассмотрено Lippincott-Schwartz и Patterson в Science (2003) 300(5616):87-91 и Chudakov et al. (Physiol Rev 2010 Jul; 90(3):1103-63).
Флуоресцентные белки способны к флуоресценции при облучением светом подходящей длины волны. Флуоресцентные свойства этих белков обусловлены взаимодействием двух или более аминокислотных остатков, формирующих хромофор, а не флуоресценцией какого-либо одного аминокислотного остатка.
GFP гидромедузы Aequorea aequorea (синоним A. victoria), был описан Johnson et al. в J Cell Comp Physiol.(1962), 60:85-104, как часть биолюминесцентной системы медузы, где GFP играет роль вторичного эммитера, преобразовывающего синий свет от фотобелка экворина в зеленый свет, а
кДНК, кодирующая A. victoria GFP была клонирована Prasher et al. (Gene (1992), 111(2):229-33). Оказалось, что этот ген может быть гетерологично экспрессирован в практически любом организме благодаря уникальной способности GFP автокаталитически образовывать хромофор (Chalfie et al., Science 263 (1994), 802-805). Эти сведения открыли широкие перспективы для использования GFP в клеточной биологи, в качестве генетически кодируемой флуоресцирующей метки.
GFP был использован в широком спектре приложений, включая исследование экспрессии генов и локализацию белков (Chalfie et al.. Science 263 (1994), 802-805, and Heim et al. in Proc. Nat. Acad. Sci. (1994), 91: 12501-12504), как инструмент для визуализации внутриклеточного распределения органелл (Rizzuto et al., Curr. Biology (1995), 5: 635-642), для визуализации транспорта белков по секреторному пути (Kaether and Gerdes, FEBS Letters (1995), 369: 267-271).
Были проведены многочисленные, исследования для улучшения свойств avGFP (Aequorea victoria GFP) и для получения вариантов GFP, пригодных и оптимизированных для различных исследовательских целей. Была проведена оптимизация генетического кода avGFP (codon usage) для повышения уровня экспрессии в клетках млекопитающих ("гуманизированный" GFP, Haas, et al.. Current Biology (1996), 6: 315-324; Yang, et al., Nucleic Acids Research (1996), 24: 4592-4593). Были получены различные мутанты GFP, в том числе "усиленный зеленый флуоресцентный белок" (EGFP), имеющий две аминокислотные замены: F64L и S65T (Heim et al., Nature 373 (1995), 663-664). Другие мутанты являются синим, голубым и желто-зеленым спектральными вариантами avGFP и содержат замены аминокислотных остатков, формирующих хромофор, и/или остатков, формирующих окружение хромофора.
В 1999 г., гомологи GFP были клонированы из небиолюминесцентных видов Anthozoa (Matz et al., Nature Biotechnol. (1999), 17: 969-973). Это открытие продемонстрировало, что эти белки не являются обязательно компонентом биолюминесцентной системы.СРР-подобные белки из Anthozoa обладали большим спектральным разнообразием и включали циановые, зеленые, желтые, красные флуоресцентные белки и фиолетово-синие не-флуоресцентные хромопротеины (CPs) (Matz et al., Bioessays (2002), 24(10):953-959). В дальнейшем кДНК GFP-подобных белков были клонированы из ряда гидроидных медуз и из копепод (Shagin et al., Mol Biol Evol. (2004), 21(5):841-850). Сегодня семейство GFP-подобных белков включает сотни флуоресцентных и окрашенных гомологов GFP. Сходство этих белков с GFP варьирует от 80-90% до менее, чем 25% идентичности аминокислотной последовательности.
Были получены кристаллические структура avGFP дикого типа, GFP S65T мутанта и ряда гомологов GFP (Ormo et al. Science (1996) 273: 1392-1395; Wall et al. Nat Stmct Biol (2000), 7: 1133-1138; Yarbrough et al. Proc Nati Acad Sci U S A (2001) 98: 462-467; Prescott et al. Structure (Camb) (2003), 11: 275-284; Petersen et al. J Biol Chem (2003), 278: 44626-44631; Wilmann et al. J Biol Chem (2005), 280: 2401-2404; Remington et al. Biochemistry (2005), 44, 202-212; Quillin et al. Biochemistry (2005), 44: 5774-5787). Было постулировано, что все члены семейства обладают общей 3D структурой (GFP-подобным доменом), представляющей собой так называемый "бочонок" из 11 бета-слоев, образующих компактную встречно-параллельную структуру, внутри которой располагается альфа-спираль, содержащая хромофор. Хромофор формируется внутри GFP-подобного домена путем окислительной циклизации трех консервативных аминокислотных остатков в центральном регионе альфа-спирали (Cody et al., Biochemistry (1993) 32, 1212-1218). Положения аминокислотных остатков, формирующих хромофор, соответствует Ser65-Tyr66-Gly67 региону avGFP. Эти аминокислотные остатки легко могут быть идентифицированы у любого GFP-подобного белка путем выравнивания его последовательности с последовательностью avGFP.
Флуоресцентные белки представляют собой уникальное семейство структурно родственных белков, которые способны формировать хромофор автокаталитически без привлечения внешних субстратов или кофакторов. Под действием индуцирующего света, хромофор производит флуоресценцию, легко детектируемую с помощью современного лабораторного оборудования (спектрофлуориметр, флуоресцентный микроскоп, флуорес-центно-активируемый клеточный сортер, планшетный флуориметр).
Процесс автокаталитического формирования хромофора белков с различными спектральными свойствами подробно описан в ряде статей и включает несколько химических реакций (Heim et al. Proc Nati Acad Sci USA. 1994; 91: 12501-12504; Ormo et al. Science. 1996;273:1392-1395; Yang et al. Nat Biotechnol. 1996; 14:1246-1251; Brejc et al. J. Proc Nati Acad Sci USA. 1997; 94: 2306-2311; Palm et al. Nat Struct Biol. 1997;4:361-365; Gurskaya et al., BMC Biochem. 2001; 2:6; Gross et al. Proc Nati Acad Sci USA. 2000; 97:11990-11995; Wall et al. Nat Struct Biol. 2000; 7:1133-1138; Yarbrough et al., J. Proc Nati Acad Sci USA. 2001;98:462-467; Pakhomov, A. A. and Mar-tynov, V. I. Chem. Biol. 2008, 15, 755- 764; Quillinet al. (2005) Biochemistry 44, 5774-5787; Yampolsky et al. (2005) Biochemistry 44, 5788- 5793; Shu et al. (2006) Biochemistry 45, 9639- 9647; Kikuchi et al. (2008) Biochemistry 47, 11573-11580; Yampolsky et al., Biochemistry, 2009, 48 (33), pp 8077-8082).
GFP-подобные белки широко используют для создания генетически-кодируемых биосенсоров. Исследование внутриклеточных процессов с помощью таких генетически кодируемых биосенсоров становятся все более популярным, так как только такие сенсоры могут дать информацию об изменении исследуемого параметра.непосредственно в живой системе. Такие биосенсоры востребованы как в фундаментальных исследованиях сигнальных путей организма, так и при тестировании токсических и лекарственных препаратов на модельных клеточных линиях или организмах. В сравнении с химическими и физическими методами регистрации биологически активных субстанций биосенсорами, требующими экзогенно добавляемых красителей, субстратов или кофакторов, генетически кодируемые нанобиосенсоры относятся к классу;, безреагентных и многоразовых сенсоров. Использование флуоресцентных белков в качестве передатчиков информации стало наиболее эффективным подходом к разработке генетически-кодируемых сенсоров.
Биосенсоры на основе флуоресцентных белков представляют собой химерные белки, в состав которых входит сенсорный домен - белковый домен, чувствительный к изменению определенного параметра клетки, например, изменению концентрации какого-либо иона или молекулы (ионов кальция, перекиси водорода, ионов водорода и т.д.). В качестве сигнальной части биосенсора используют GFP-подобные белки или их варианты, подвергнутые круговой пермутации (Suslova and Chudakov, Trends Biotechnol. 23(12) (2005), 605-13; Griesbeck, Curr Opin NeurobioL, 2004, v.14(5), pp.636-641; Bunt and Wouters, Int Rev CytoL, 2004, v.237, pp.205-277).
Создание пермутированных GFP-подобных белков необходимо для увеличения подвижности хромофорного окружения и, следовательно, для большей лабильности спектральных свойств белка. Круговая пермутация флуоресцентных белков описана Topell S. и Glockshuber R. (Methods in Molecular Biology. 183, 31-48; 2002). Например, для круговой пермутации avGFP в его первичную структуру вносится разрыв в область между 144 и 149 аминокислотами, нативные N- и С - концы совмещаются при помощи полипептидного линкера. Новые N- и С - концы находятся в непосредственной близости от хромофора и могут влиять на его микроокружение. Круговая пермутация производится.на уровне нуклеиновой кислоты путем оперативного сшивания 3'- и 5'- концов нуклеотидной последовательности, кодирующей флуоресцентный белок, и внесения разрыва в последовательность между кодонами, кодирующими новые N и С - концевые аминокислоты. Методы для получения таких конструкций хорошо известны специалистам в данной области.
В результате круговой пермутации кпфб приобретает способность реагировать на конформационные перестройки в области новых N и С - концов изменением спектра флуоресценции (сенсоры с использованием обычных флуоресцентных белков оказались малочувствительными). Эффективность биосенсоров на основе кпфб, полученных из avGFP была продемонстрирована на примере сенсора на ионы кальция (Nagai et al, Proc Nati Acad Sci USA. 98(6) (2001), 197-202). кпфб был так же использован для изготовления сенсора на пероксид водорода HyPer (Belousov et al., Nature Method.4, 281-286; 2006).
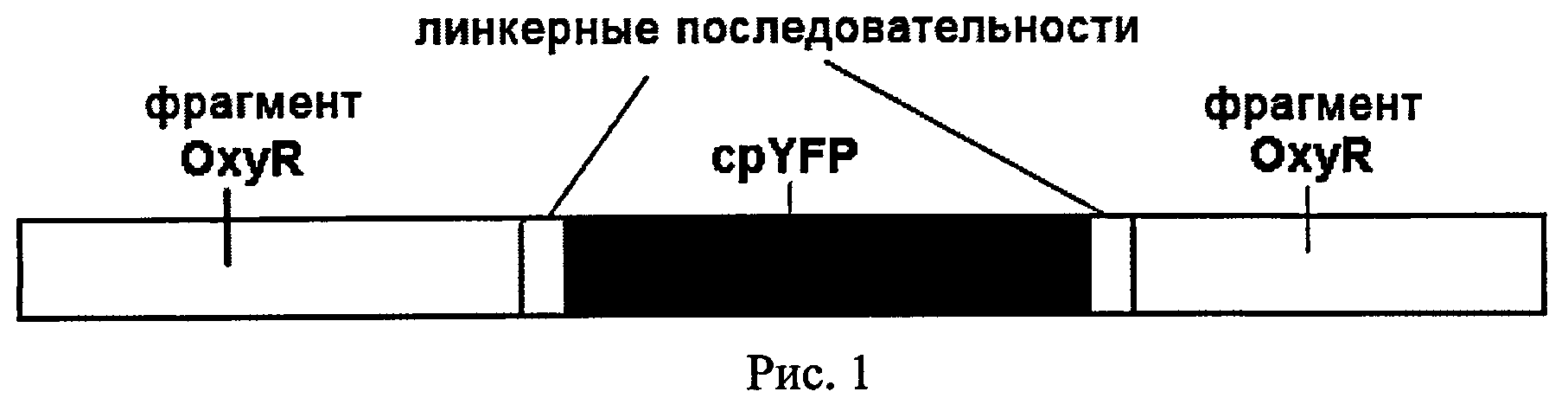
HyPer представляет собой химерный белок, состоящий из регуляторного домена белка OxyR E.coli, реагирующего с пероксидом водорода и интегрированного в пептидную цепь OxyR пермутированного желтого флуорес-центного белка cpYFP (рис.1). HyPer представляет собой конструкцию, в которой cpYFP соединен с двумя: фрагментами чувствительного к H2O2 домена OxyR посредством коротких пептидных линкеров. Нуклеотидные последовательности, кодирующие фрагменты белка OxyR и cpYFP оперативно сшиты между собой с использованием линкерных последовательностей. Линкерные последовательности - нуклеотидные последовательности, кодирующие одну или несколько аминокислот, используются для повышения подвижности частей химерного белка относительно друг друга и способствуют его лучшему фолдированию (формированию правильной 3D-структуры).
Транскрипционный фактор Е.coli OxyR активирует экспрессию ряда генов в ответ на увеличение концентрации H2O2. OxyR состоит из двух доменов, различающихся функционально. N-концевой домен (первые 80 аминокислот) образует ДНК-связывающий участок, в то время как С-концевой домен (81 - 305 аминокислот) выполняет регуляторную функцию и способствует олигомеризации белка (Choi et al., Cell. 105, 103 - 113;2001). В присутствии Нг02 происходит образование дисульфидной связи между цис-теиновыми остатками Cys-199 и Cys-208 OxyR белка, что приводит к кон-формационным изменениям всего регуляторного домена OxyR (Belousov et al., Nature Method.4, 281 - 286; 2006).
HyPer является уникальным инструментом, позволяющим анализировать изменение концентрации перекиси водорода внутри живых клеток. Перекись водорода (H2O2) - важная сигнальная молекуля, ответственная за включение ряда внутриклеточных каскадов (Yoshizumi et al, J Biol Chem. 2000, V. 275(16), pp.11706-11712; Abe et al, J Biol Chem. 1996, V. 271(28), pp.16586-16590; Schreck et al, EMBO J. 1991, V. 10(8), pp.2247-2258; Meyer et al, EMBO J. 1993, V. 12(5), pp.2005-2015). Многие внешние стимулы вызывают внутриклеточную продукцию H2O2, которая в свою очередь приводит к активации белковых каскадов. При появлении перекиси водорода в окружающей HyPer среде, происходит окисление домена OxyR и изменение его конформации, которое в свою очередь вызывает изменение спектральных характеристик флуоресцентного белка. Эти изменения могут быть зарегистрированы с помощью визуального скрининга, спектрофото-метрии, спектрофлуориметрии, флуоресцентной микроскопии, с помощью FACS или другим общепринятым способом для регистрации спектральных характеристик флуоресценции. Через некоторое время происходит восстановление биосенсора за счет работы клеточных системам, восстанавливающих дисульфидные связи в домене OxyR. Восстановление OxyR in vitro глуторедоксином-1 и тиоредоксином было показано Aslund et al. (Biochemistry. 96, 6161 - 6165;1999).
Был разработан вариант HyPer, названный HyPer2 с увеличенным динамическим диапазоном (Markvicheva et al., Bioorg Med Chem. 2011; 19(3):1079-84). HyPer2 был получен внесением; мутации A406V в последовательность HyPer. HyPer2 имеет улучшенный примерно в 2-раза динамический диапазон, нежели HyPer.
HyPer и HyPer2 позволяют отслеживать динамические изменения концентрации Нз02 в режиме реального времени, однако вследствие относительно высоких скоростей восстановления OxyR домена, невозможно выявлять и продолжительное время отслеживать клетки, которые прореагировали на стимуляцию производством пероксида водорода. Иными словами, биосенсоры HyPer и HyPer2 работают как «динамические» сенсоры, но не могут быть использованы как «накопительные». Это ограничивает область применения HyPer и HyPer2 для мониторинга изменений концентрации НзОз, например, в системах in vivo, или при сортировке клеток активированной флюоресценцией (FACS). То есть, когда нужно, чтобы сигнал биосенсора сохранялся в течение времени, необходимого для обработки биологических образцов (например, когда необходимо фиксировать ткани и изготовлять срезы тканей для дальнейшего'изучения или когда нужно время для нанесения образцов на FACS-машину).
Настоящее изобретение обеспечивает выделенные молекулы нуклеиновых кислот, кодирующих флуоресцентный биосенсор для детекции пероксида водорода, обладающий уменьшенной скоростью восстановления после реакции с пероксидом водорода по сравнению с HyPer и HyPer2. В некоторых воплощениях, нуклеиновая кислота настоящего изобретения кодирует флуоресцентный биосенсор для детекции пероксида водорода, аминокислотная последовательность которого отличается от последовательности HyPer2 (SEQ ID NO:1) по крайней мере, одной аминокислотной заменой. В некоторых воплощениях, это замена в регуляторном домене OxyR. В некоторых воплощениях, это замена по положению 34 в аминокислотной последовательности, представленной в (SEQ ID NO:1). В некоторых воплощениях, это замена H34Y.
В некоторых воплощениях, нуклеиновая кислота настоящего изобретения кодирует флуоресцентный биосенсор для детекции пероксида водорода, имеющий аминокислотную последовательность, показанную в SEQ ID NO:4. В некоторых воплощениях, нуклеиновая кислота настоящего изобретения имеет нук-леотидную последовательность, показанную в SEQ ID NO:03. В некоторых воплощениях, нуклеиновая кислота настоящего изобретения кодирует флуоресцентный биосенсор для детекции пероксида водорода, слитый с сигналом внутриклеточной локализации.
В некоторых воплощениях, нуклеиновая кислота настоящего изобретения кодирует флуоресцентный биосенсор для детекции пероксида водорода, который обладает меньшей скоростью восстановления после окисления, происходящего в процессе взаимодействия сенсора с пероксидом водорода, чем с HyPer и HyPer2.
Молекулы нуклеиновых кислот, которые отличаются от представленных нуклеотидных последовательностей вследствие вырожденности генетического кода так же входят в рамки настоящего изобретения. В других воплощениях также обеспечиваются векторы, включающие нуклеиновую кислоту настоящего изобретения. Кроме того, настоящее изобретение обеспечивает кассеты экспрессии, включающие нуклеиновую кислоту настоящего изобретения и регуляторные элементы, необходимые для экспрессии нуклеиновой кислоты в выбранной клетке-хозяине. Кроме того, также обеспечиваются клетки, стабильные клеточные линии, трансгенные животные и трансгенные растения, включающие нуклеиновые кислоты, векторы или экспрессионные кассеты настоящего изобретения. В других воплощениях обеспечиваются функциональные флуоресцентные биосенсоры настоящего изобретения, которые кодируются нуклеиновыми кислотами указанными выше. Кроме того, обеспечиваются набор, содержащий нуклеиновые кислоты и/или векторы и/или экспрессионные кассеты, включающие указанные нуклеиновые кислоты настоящего изобретения.
Краткое описание рисунков
Рисунок 1 илюстрирует принципиальную схему строения биосенсора для детекции пероксида водорода.
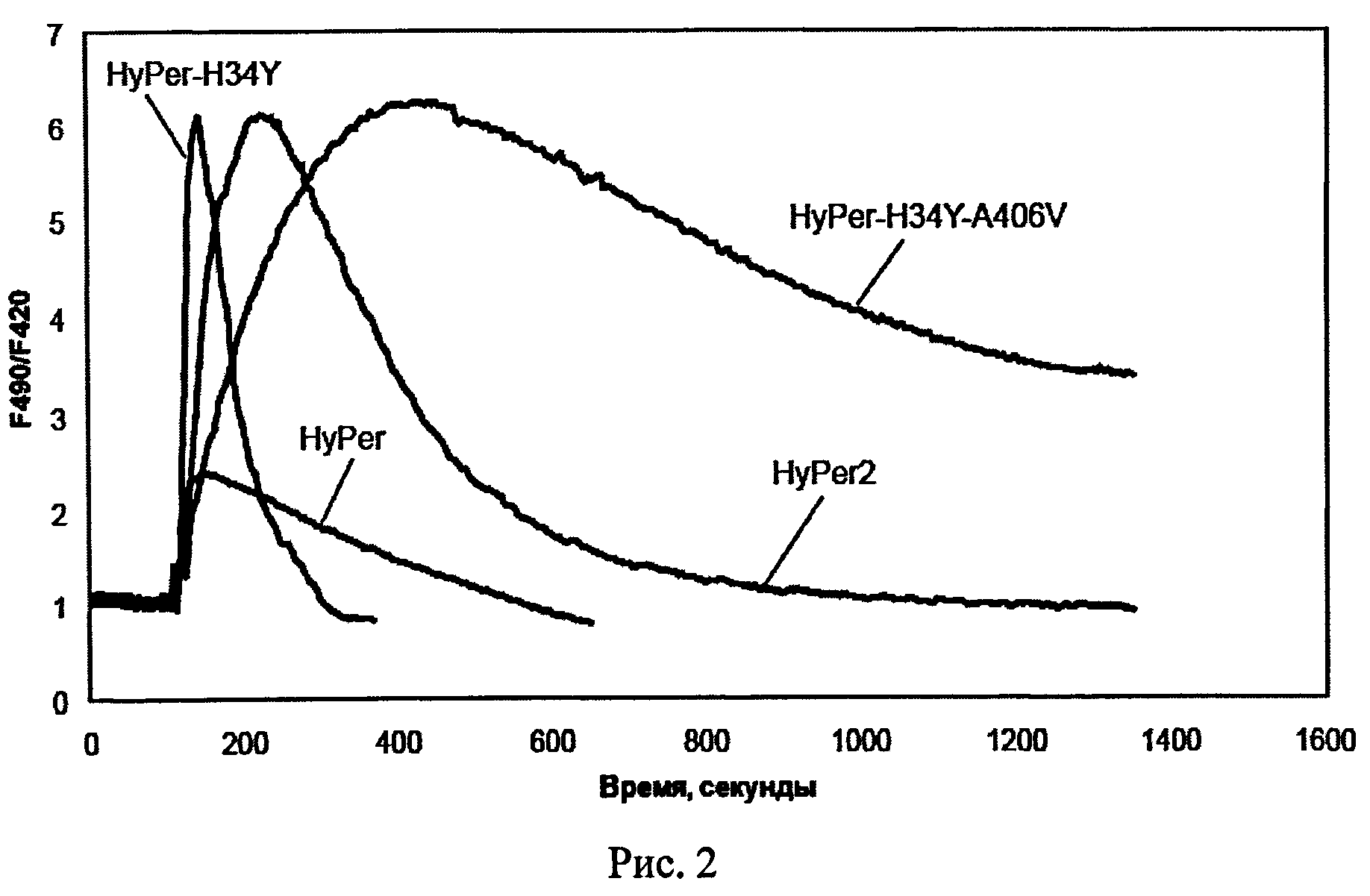
Рисунок 2 иллюстрирует динамику изменения сигнала вариантов биосенсора HyPer во времени после добавления пероксида водорода.
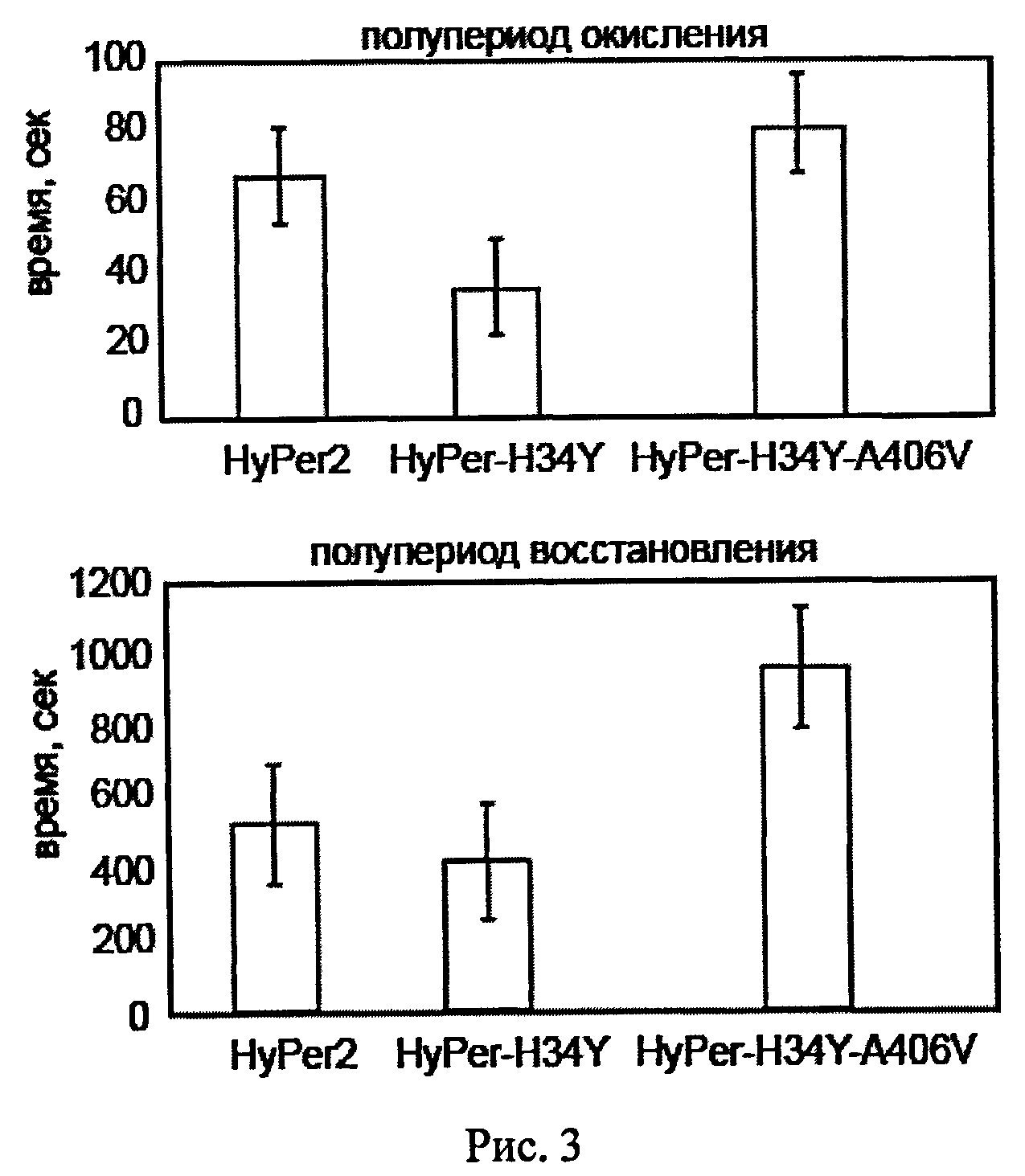
Рисунок 3 показывает сравнение среднего значения для полупериода окисления и восстановления вариантов биосенсора HyPer (n=20, Р<0.05).
Осуществление изобретения
Для более полного раскрытия вышеперечисленных характеристик настоящего изобретения ниже предлагается детальное описание изобретения, кратко сформулированного выше, в виде ссылок на воплощения, некоторые из которых проиллюстрированы дополнительными фигурами. При этом следует отметить, что прилагаемые фигуры иллюстрируют лишь типичные воплощения настоящего изобретения и, следовательно, не должны быть восприняты в качестве ограничения объема изобретения, которое может допускать другие в равной степени эффективные воплощения.
Как указано выше, настоящее изобретение направлено на молекулы нуклеиновых кислот, которые кодируют мутантный вариант биосенсора HyPer, обладающие уменьшенной скоростью восстановления после реакции с пероксидом водорода. Нуклеиновые кислоты настоящего изобретения получены с помощью рекомбинантных технологий. В предпочтительных воплощениях, нуклеиновые кислоты настоящего изобретения кодируют белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO:4. В некоторых воплощениях, нуклеиновые кислоты, кодируют белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO:4, слитый с сигналом определенной внутриклеточной локализации. Также обеспечиваются векторы и кассеты экспрессии, включающие нуклеиновую кислоту настоящего изобретения. Кроме того, обеспечиваются клетки, стабильные клеточные линии, трансгенные животные и трансгенные растения, включающие нуклеиновые кислоты, векторы или экспрессионные кассеты настоящего изобретения.
Указанные нуклеиновые кислоты применяются во многих различных приложениях и методах, в частности, для мониторинга изменений концентрации пероксида водорода внутри живых клеток и выявления клеток, в которых произошла активация продукции переоксида водорода. Наконец, обеспечиваются наборы для их использования в таких методах и приложениях.
Определения
Различные термины, относящиеся к биологическим молекулам настоящего изобретения, используются выше и также в описании и в формуле изобретения.
Как здесь используется, термин "флуоресцентный белок" означает белок, относящийся к семейству GFP-подобных белков, который обладает способностью к флуоресценции; например, он может проявлять низкую, среднюю или интенсивную флуоресценцию при облучении светом с подходящей для возбуждения длиной волны. Флуоресцентное свойство флуоресцентного белка представляет собой такое свойство, которое является результатом работы хромофора, образующегося путем автокаталитической циклизации трех или более аминокислотных остатков в полипептидной цепи. Как таковые флуоресцентные белки настоящего изобретения не включают белки, которые обладают флуоресценцией за счет отдельных флуоресцирующих остатков, таких как триптофан, тирозин и фенилаланин. Термин «флуоресцентный белок» относится так же к флуоресцентным белкам GFP-семейства, подвергнутым круговой пермутации.
Как здесь используется, термин "avGFP" относится к зеленому флуоресцентному белку из медузы Aequorea victoria, включая варианты avGFP, известные из уровня техники, сконструированные для обеспечения большей интенсивности флуоресценции или флуоресценции в других цветовых областях. Последовательность дикого типа avGFP была раскрыта в Prasher et al. (1992, Gene 111: 229-33).
Как здесь используется, термин "флуоресцентный белок, подвергнутый круговой пермутации" или "кпфб" относится к белку, полученному из флуоресцентного белка (например из avGFP) с помощью генно-инженерной модификации нуклеиновой кислоты, в результате которой С- и N- концы исходного флуоресцентного белка оказываются оперативно слиты, а новые С- и N- концы формируются вблизи хромофора. Круговая пермутация не влияет на формирование «бочонка» GFP-подобного домена и формирование активного (способного к флуоресценции) хромофора. Круговая пермутация приводит к тому, что кпФБ приобретает способность менять спектральные характеристики при конформационных изменениях белковых доменов или полипептидов, оперативно слитых с его С- и N-концами.
Термин "гуманизированный" относится к изменению нуклеотидной последовательности флуоресцентного белка, сделанной для оптимизации генетического кода кодонов для экспрессии в клетках млекопитающих (Yang et al„ 1996, Nucleic Acids Research 24:4592-4593).
Как здесь используется, термин "выделенный" означает молекулу или клетку, которые находятся в среде, отличной от среды, в которой молекула или клетка находятся в естественных условиях.
Как здесь используется, термин "мутант" или "производное" относятся к белку, раскрытому в настоящем изобретении, в котором одна или более аминокислот добавлены и/или замещены и/или удалены (делегированы) и/или вставлены (инсертированы) в N-конец и/или С-конец, и/или в пределах нативных аминокислотных последовательностей белков настоящего изобретения. Как здесь используется, термин "мутант" относится к молекуле нуклеиновой кислоты, которая кодирует мутантный белок. Кроме того, термин "мутант" здесь относится к любому варианту, который короче или длиннее белка или нуклеиновой кислоты.
Как здесь используется, "гомология" - это термин, использующийся для описания взаимосвязи последовательностей нуклеотидов или аминокислот с другими последовательностями нуклеотидов или аминокислот, которая определена степенью идентичности и/или сходства между указанными сравниваемыми последовательностями.
Как здесь используется, аминокислотная или нуклеотидная последовательности "по существу сходны" или "по существу такие же как референсная последовательность, если аминокислотная или нуклеотидная последовательности имеют по крайней мере 85% идентичности с указанной последовательностью внутри выбранного для сравнения региона. Таким образом, по существу сходные последовательности включают те, которые имеют, например, по крайней мере, 85% идентичности, по крайней мере, 90% идентичности, по крайней мере, 95% идентичности или по крайней мере, 96%б 97%б 98% или 99% идентичности. Две последовательности, которые идентичны одна другой, так же по существу сходны.
Процент идентичности последовательностей определяется на основании референсной последовательности. Алгоритмы для анализа последовательности известны в данной области, такие как BLAST, описанный в Altschul et al., J. Mol. BioL, 215, pp.403-10 (1990). Для целей настоящего изобретения сравнение нуклеотидных и аминокислотных последовательностей производимое с помощью пакета программ Blast, предоставляемого National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/blast) с использованием содержащего разрывы выравнивания со стандартными параметрами, может быть использовано для определения уровня идентичности и сходства между нуклеотидными последовательностями и аминокислотными последовательностями.
Как здесь используется, термин "подобные белки" или "по существу сходные белки" относится к белкам, которые имеют аминокислотные последовательности, идентичные по крайней мере на 85%, как правило идентичные на 90% или более, чаще всего идентичные по крайней мере на 95% или более (например на 96% и более, 97% и более, 98% и более, 99% и более, 100%). Длина гомологичных аминокислотных последовательностей у «подобных белков» при этом может составлять, по крайней мере, 100 аминокислотных остатков, чаще, по крайней мере, 200 аминокислотных остатков, или 300 аминокислотных остатков.
В некоторых воплощениях, термин "подобные белки" или "по существу сходные белки" относится к белкам, которые имеют аминокислотные последовательности целого белка идентичные по крайней мере на 85%, как правило идентичные на 90% или более, чаще всего идентичные по крайней мере на 95% или более (например на 96% и более, 97% и более, 98% и более, 99% и более, 100%).
Как здесь используется, термин "функциональный" означает, что нуклеотидная или аминокислотная последовательность может функционировать для указанного испытания или задачи. Термин "функциональный", используемый для описания биосенсора для детекции пероксида водорода настоящего изобретения, означает, что он меняет спектральные характеристики при увеличении концентрации пероксида водорода в среде.
Как здесь используется, термин «среда» по отношению к биосенсору для детекции пероксида водорода означает любую среду, в которой этот биосенсор может функционировать. Для выделенного белка это может быть любой буферный раствор, в котором этот сенсор сохраняет функциональность. Для белка, экспрессированного в клетке, это цитоплазма или внутриклеточный компартмент, в котором биосенсор локализован.
Как здесь используется, "биохимические свойства" относятся к белковому фолдингу (сворачиванию) и скорости созревания, полупериоду окисления, полупериоду восстановления после реакции с пероксидом водорода, времени полужизни, способности к агрегации, способности к олигомеризации, рН и температурной стабильности, и другим подобным свойствам.
Как здесь используется, "флуоресцентные свойства" или "спектральные свойства" относятся к коэффициенту молярной экстинкции при подходящей длине волны, к квантовому выходу флуоресцентции, форме спектра возбуждения флуоресценции или спектра испускания, длине волны, соответствующей максимуму возбуждения флуоресценции, и длине волны, соответствующей максимуму испускания, отношению амплитуды возбуждения флуоресценции при двух разных длинах волн, отношению амплитуды испускания при двух разных длинах волн, времени жизни возбужденного состояния, и анизотропии оптических свойств. Измеряемая разница может быть определена как количество любого количественного флуоресцентного свойства, например, интенсивность флуоресцентции при определенной длине волны, или интегральная флуоресценция на всем спектре испускания.
Как здесь используется, "агрегация" относится к склонности или способности экспрессированного белка формировать нерастворимый осадок (агрегаты). "Агрегация" должна быть отличаема от "олигомеризации". В частности, мутанты с уменьшенной способностью к агрегации, например, с увеличенной растворимостью, не обязательно имеют уменьшенную способность к олигомеризации.
Как здесь используется, "олигомеризация" относится к склонности или способности экпрессированного белка формировать комплексы (олигоме-ры) в результате специфического взаимодействия двух или более полипептидов. Указанное специфическое взаимодействие наблюдается в специальных условиях, например, в физиологических условиях, и относительно стабильно в этих условиях. Ссылка на "способность" белков олигомеризоваться означает, что белки могут формировать димеры, триммеры, тетрамеры или подобные комплексы в специальных условиях. Как правило, флуоресцентные белки обладают способностью к олигомеризации в физиологических условиях. Флуоресцентные белки могут также олигомеризоваться при других, например, рН, нежели рН при физиологических условиях. Условия, при которых флуоресцентные белки формируют олигомеры или проявляют склонность к олигомеризации могут быть определены с помощью хорошо известных методов, таких как гель-фильтрация или иным способом известным в данной области.
Ссылка на нуклеотидную последовательность "кодирующую" полипептид означает, что с нуклеотидной последовательности в ходе трансляции и транскрипции мРНК продуцируется этот полипептид. При этом может быть указана как кодирующая цепь, идентичная мРНК и обычно используемая в списке последовательностей, так и комплементарная цепь, которая используется как матрица при транскрипции. Как очевидно для любого специалиста в данной области техники, термин так же включает любые вырожденные нуклеотидные последовательности кодирующие одинаковую аминокислотную последовательность. Нуклеотидная последовательности кодирующие полипептид включают последовательности, содержащие интроны.
Термин "оперативно связанный", «оперативно слитый» или ему подобный при описании структуры биосенсора или химерных белков на его основе относится к полипептидным последовательностям, которые находятся в физической и функциональной связи одна с другой. В наиболее предпочтительных воплощениях, основные функции полипептидных компонентов химерной молекулы не изменены по сравнению с функциональными свойствами выделенных полипептидных компонентов. Например, OxyR домен, входящий в состав биосенсора настоящего изобретения, сохраняет способность связывать пероксид водорода, а кпБФ - способность к флуоресценции. В случае, когда биосенсор настоящего изобретения сшит с представляющим интерес сигналом внутриклеточной локализации, химерный белок сохраняет способность реагировать на появление пероксида водорода, но локализуется в определенном клеточном компартменте. Как очевидно для любого специалиста в данной области техники, нуклеотидные последовательности, кодирующие химерный белок, включающий "оперативно связанные" компоненты (белки, полипептиды, линкерные последовательности, белковые домены и т.д.), состоят из фрагментов, кодирующих указанные компоненты, где эти фрагменты ковалентно связаны таким образом, что в ходе трансляции и транскрипции нуклеотидной последовательности продуцируется полноразмерный химерный белок. Иными словами, фрагменты соединены таким образом, что в местах их соединения отсутствуют 'сбойки' рамки считывания и стоп-кодоны.
Как здесь используется, термины «перекись водорода» и «пероксид водорода» относятся к молекуле, имеющей формулу H2O2.
Как здесь используется, термин «сигнал биосенсора» означает детектируемое изменение спектральной характеристики биосенсора в ответ на перок-сид водорода.
Как здесь используется, термин «динамический диапазон» означает отношение максимального уровня сигнала биосенсора (в полностью окисленном состоянии) к уровню исходного сигнала (в полностью восстановленном состоянии).
Как здесь используется, термин «скорость ответа» или «скорость окисления» в отношении биосенсора настоящего изобретения относится к скорости, с которой молекула биосенсора реагирует на увеличение концентрации или появление перекиси водорода в среде. Для нужд настоящего изобретения скорость ответа может быть охарактеризована «полупериодом окисления» (Т1/20Х) биосенсора - половиной времени, за который биосенсор достигает максимального значения сигнала, считая от момента добавления пероксида водорода в среду.
Как здесь используется, термин «скорость восстановления» в отношении биосенсора настоящего изобретения, относится к скорости, с которой молекула окисленного биосенсора восстанавливает способность реагировать на появление в среде пероксида водорода за счет взаимодействия с клеточными системами, восстанавливающими дисульфидные связи. Для нужд настоящего изобретения скорость восстановления может быть охарактеризована «полу периодом восстановления» (Т 1/2 RED) - половина времени, за которое биосенсор достигает минимального (первоначального, характекр-ного для восстановленной формы) значения сигнала, считая от момента достижения максимума сигнала при окислении.
Молекулы нуклеиновых кислот
Настоящее изобретение обеспечивает молекулы нуклеиновых кислот кодирующие флуоресцентный биосенсор для детекции пероксида водорода, имеющий аминокислотную последовательность SEQ ID NO:4.
Как здесь используется, молекула нуклеиновой кислоты это молекула ДИК, такая как геномная ДИК или кДНК молекула, или молекула РНК, такая как молекула мРНК. Как здесь используется, термин "кДНК" относится к нуклеиновым кислотам, которые обладают размещением элементов последовательности найденным в нативных зрелых видах мРНК, где элементы последовательности - это экзоныи 5' и 3' некодирующие области. Структура биосенсора схематически изображена на рисунке 1. Методы получения таких слитых белков хорошо известны в данной области. Например, части нуклеиновой кислоты, кодирующие различные элементы, могут быть встроены в полилинкер вектора, таким образом, что эти части окажутся оперативно связаны и между ними не будет стоп-кодонов в рамке считывания и не будет сбоек рамки считывания. Альтернативно, желательная нуклеотидная последовательность может быть собрана из фрагментов с помощью ДНК-лигазы или ПЦР с праймерами, содержащими части, комплементарные концевым последовательностям соединяемых фрагментов.
Молекула нуклеиновой кислоты, кодирующая флуоресцентный биосенсор, может быть синтезирована из подходящих нуклеозидтрифосфатов или выделена из биологических источников. Оба метода основаны на хорошо известных в данной области протоколах. Например, доступность информации о последовательности аминокислот или информации о нуклеотидной последовательности дает возможность изготовить выделенные молекулы нуклеиновых кислот настоящего изобретении с помощью олигонуклео-тидного синтеза. В случае информации о последовательности аминокислот может быть синтезировано несколько нуклеиновых кислот отличающихся друг от друга вследствие вырожденности генетического кода. Методы выбора вариантов кодонов для требуемого хозяина хорошо известны в данной области. Синтетические олигонуклеотиды могут быть приготовлены с помощью фосфорамидитного метода, и полученные конструкты могут быть очищены с помощью методов хорошо известных в данной области, таких как высокоэффективная жидкостная хроматография (ВЭЖХ) или других методов как описано, например, в Sambrook et al.. Molecular Cloning: A Laboratory Manual, 2nd Ed., (1989) Cold Spring Harbor Press, Cold Spring Harbor, NY, и по инструкции, описанной в, например. United States Dept. of HHS, National Institute of Health (NIH) Guidelines for Recombinant DNA Research. Длинные двухцепочечные молекулы ДНК настоящего изобретения могут быть синтезированы за следующие стадии: несколько меньших фрагментов с необходимой комплементарностью, которые содержат подходящие концы способные к когезии с соседним фрагментом, могут быть. Соседние фрагменты могут быть сшиты с помощью ДНК-лигазы или метода, основанного на ПЦР. Нуклеиновая кислота, кодирующая биосенсор или его фрагмент, может быть выделена любым из многих известных методов.
Кроме того, также обеспечиваются вырожденные варианты нуклеиновых кислот, которые кодируют биосенсор настоящего изобретения. Вырожденные варианты нуклеиновых кислот включают замены кодонов нуклеиновой кислоты на другие кодоны, кодирующие те же самые аминокислоты. В частности вырожденные варианты нуклеиновых кислот создаются, чтобы увеличить экспрессию в клетке-хозяине. В этом воплощении, кодоны нуклеиновой кислоты, которые не являются предпочтительными или являются менее предпочтительными в генах клетки-хозяина, заменены кодонами, которые обильно представлены в кодирующих последовательностях генов в клетке-хозяине, где указанные замененные кодоны кодируют ту же самую аминокислоту.
Особенный интерес представляют гуманизированные версии нуклеиновых кислот настоящего изобретения. Как здесь используется, термин "гумани-зированный" относится к заменам, сделанным в последовательности нуклеиновой кислоты для оптимизации кодонов для экспрессии белка в клетках млекопитающих (человека) (Yang et al., Nucleic Acids Research (1996) 24: 4592-4593). См. также Патент США №5795737, который описывает гуманизацию белков, раскрытие которого здесь включено ссылкой. В некоторых воплощениях, молекула нуклеиновой кислоты настоящего изобретения - это ДНК (или кДНК) молекула, содержащая открытую рамку считывания, которая кодирует биосенсор настоящего изобретения и способна в подходящих условиях (например, физиологические внутриклеточные условия) быть использована для экспрессии белка в клетке-хозяине. Настоящее изобретение так же охватывает нуклеиновые кислоты, которые гомологичны, по существу сходны, идентичны, или получены из нуклеиновых кислот, кодирующих белки настоящего изобретения. Указанные нуклеиновые кислоты находятся в среде, отличной от среды, в которой они находятся в естественных условиях, например, они выделены, представлены в увеличенном количестве, находятся или экспрессированы в системах in vitro или в клетках или организмах, отличных от тех, в которых они находятся в естественных условиях.
Заявленные нуклеиновые кислоты могут быть выделены и получены по существу в очищенной форме. По, существу очищенная форма означает, что нуклеиновые кислоты являются, по меньшей мере, на 50% чистыми, обычно, по меньшей мере, на 90% чистыми и обычно являются "рекомбинантными", то есть, фланкированы одним или более нуклеотидов, - с которыми она обычно не связана в хромосоме, встречающейся в природе в ее естественном организме-хозяине.
Изменения или различия в нуклеотидной последовательности между высоко сходными нуклеотидными последовательностями могут представлять нуклеотидные замены в последовательности, которые возникают в процессе нормальной репликации или дупликации. Другие замены могут быть специально рассчитаны и вставлены в последовательность для определенных целей таких, как изменение кодонов определенных аминокислот или нуклеотидной последовательности регуляторного региона. Такие специальные замены могут быть произведены in vitro с помощью различных технологий мутагенеза или получены в организмах-хозяевах, находящихся в специфических селекционных условиях, которые индуцируют или отбирают эти изменения. Такие специально полученные варианты последовательности могут быть названы "мутантами" или "производными" исходной последовательности.
Мутантные или производные нуклеиновые кислоты могут быть получены на матричной нуклеиновой кислоте, выбранной из вышеописанных нуклеиновых кислот, путем модификации, делеции или добавления одного или более нуклеотидов в матричной последовательности или их комбинации, для получения варианта матричной нуклеиновой кислоты. Модификации, добавления или делеции могут быть выполнены любым способом, известным в данной области (см. например Gustin et al., Biotechniques (1993) 14: 22; Barany, Gene (1985) 37: 111-123; и Colicelli et al., Mol. Gen. Genet. (1985) 199:537-539, Sambrook et al., Molecular Cloning: A Laboratory Manual, (1989), CSH Press, pp.15.3-15.108), включая подверженный ошибкам ПЦР (error-prone PCR), shuffling, олигонуклеотид-направленный мута-генез, ПЦР со сборкой, парный ПЦР мутагенез, мутагенез in vivo, кассетный мутагенез, рекурсивный множественный мутагенез, экспоненциальный множественный мутагенез, сайт-специфический мутагенез, случайный мутагенез, генная реассемблирование (gene reassembly), генный сайт-насыщающий мутагенез (GSSM), искусственное перестройку с лигирова-нием (SLR) или их комбинации. Модификации, добавления или делеции могут быть также выполнены методом, включающим рекомбинацию, рекурсивную рекомбинацию последовательностей, фосфотиоат-модифицированный мутагенез ДНК, мутагенез на урацил-содержащей матрице, мутагенез с двойным пропуском, точечный восстановительный по рассогласованию мутагенез, мутагенез штамма, дефицитного по восстановлениям, химический мутагенез, радиоактивный мутагенез, делетацион-ный мутагенез, рестрикционно-избирательный мутагенез, рестрикционный мутагенез с очисткой, синтез искусственных генов, множественный мутагенез, создание химерных множественных нуклеиновых кислот и их комбинации.
Также обеспечиваются нуклеиновые кислоты, кодирующие химерные белки, состоящие из биосенсора настоящего изобретения и сигнала определенной внутриклеточной локализации. Такие химерные белки могут быть получены путем оперативного соединения нуклеиновой кислоты настоящего изобретения, кодирующей биосенсор, и нуклеиновой кислоты, кодирующей сигнал внутриклеточной локализации. Методы получения таких нуклеиновых кислот хорошо известны специалистам в данной области.
Также обеспечиваются вектор и другие конструкции нуклеиновой кислоты, содержащие заявленные нуклеиновые кислоты. Подходящие векторы включают вирусные и невирусные векторы, плазмиды, космиды, фаги и т.д., предпочтительно плазмиды, и используются для клонирования, амплификации, экспрессии, переноса и т.д., последовательности нуклеиновой кислоты настоящего изобретения в подходящего хозяина. Выбор подходящего вектора является понятным для квалифицированного специалиста в данной области, и известно много таких доступны коммерчески векторов. Для приготовления конструкции, полноразмерная нуклеиновая кислота или ее часть обычно встраивается в вектор посредством прикрепления ДНК-лигазой к расщепленному ферментами рестрикции сайту в векторе. Альтернативно, желательная нуклеотидная последовательность может быть вставлена гомологичной рекомбинацией in vivo, обычно, присоединением гомологичных участков к вектору на флангах желательной нуклео-тидной последовательности. Гомологичные участки добавляют лигирова-нием олигонуклеотидов или полимеразной цепной реакцией, с использованием праймеров, включающих, например, как гомологичные участки, так и часть желательной нуклеотидной последовательности.
Также обеспечиваются кассеты экспрессии или системы, использованные inter alia для получения заявленных^ бйосенсоров или химерных белков на их основе или для репликации заявленных молекул нуклеиновой кислоты.
Кассета экспрессии может существовать как внехромосомный элемент или может быть включена в геном клетки в результате введения указанной кассеты экспрессии в клетку.
В экспрессионном векторе или кассете экспрессии указанная нуклеиновая кислота является функционально связанной с регуляторной последовательностью, которая может включать промоторы, энхансеры, терминаторы, операторы, репрессоры и индукторы и обеспечивает инициацию считывания РНК (транскрипции) в клетке-хозяине. В экспрессионном векторе или кассете экспрессии нуклеиновая кислота настоящего изобретения может быть также связана с сигналами терминации транскрипции, функциональным в клетке-хозяине. Методы изготовления кассет экспрессии или систем для экспрессии желаемого продукта известны специалистом, квалифицированным в данной области.
Вышеописанные системы экспрессии могут использоваться в прокариоти-ческих или эукариотических хозяевах. Клеточные линии, которые устойчиво экспрессируют белки настоящего изобретения, могут быть выбраны способами, известными в данной области (например, котрансфекция с селектируемым маркером, таким как dhfr, gpt, неомицин, гигромицин, что делает возможным выявление и выделение транфецированных клеток, которые содержат ген, включенный в геном).
Белковый продукт, кодируемый нуклеиновой кислотой изобретения, может быть получен путем экспрессии в любой удобной системе экспрессии, включая, например, бактериальные системы, дрожжевые системы, клетки насекомых, земноводных или клетки млекопитающих. Например, для получения белка могут использоваться клетки-хозяева, такие как Е.coli, В.subtilis, S.cerevisiae, клетки насекомого в комбинации с бакуловирусными векторами, или клетки высшего организма, такого как позвоночные, например, COS 7 клетки, НЕК 293, СНО, ооциты Xenopus и т.д.
Если используется любая вышеупомянутая клетка-хозяин или другие подходящие клетки-хозяева или организмы для репликации и/или экспрессии нуклеиновых кислот изобретения, то полученная реплицированная нуклеиновая кислота, экспрессированный белок или полипептид находятся в рамках притязания изобретения как продукт клетки-хозяина или организма. Продукт может быть выделен подходящим способом, известным в данной области.
Белки Нуклеиновые кислоты по настоящему изобретению кодируют флуоресцентный биосенсор для детекции пероксида водорода. Аминокислотная последовательность биосенсора настоящего изобретения отличается от аминокислотной последовательности HyPer (SEQ ID NO:2) наличием аминокислотных замен A406V и H34Y. Аминокислотная последовательность биосенсора настоящего изобретения отличается от аминокислотной последовательности HyPer2 (SEQ ID NO:1) наличием аминокислотной замены H34Y. Специфические биосенсоры, представляющие интерес, включают биосенсор, имеющий аминокислотную последовательность SEQ ID No:04.
Заявленный биосенсор обладает способностью к детектируемой флуоресценции, которая может быть зарегистрирована с помощью визуального скрининга, спектрофотометрии, спектрофлуориметрии, флуоресцентной микроскопии, с помощью FACS или другим общепринятым способом для регистрации флуоресценции. В отсутствие пероксида водорода, заявленный биосенсор имеет максимум (пик) эмиссии в диапазоне от 500 нм до 550 нм, например, 520 нм, и два пика возбуждения флуоресценции; первый в диапазоне 400-450 нм (например, 420 нм) и второй в диапазоне 470-510 нм (например, 500 нм).
При появлении пероксида водорода заявленный биосенсор переходит в окисленное состояние. При окислении биосенсора меняется интенсивность флуоресценции при возбуждении светом с длиной волны 500 нм (F500), к интенсивности флуоресценции при возбуждении светом с длиной волны 420 нм (F420). Для нужд настоящего изобретения соотношение «F500/F420» является сигналом биосенсора. Заявленный биосенсор обладает широким динамическим диапазоном. Заявленный биосенсор имеет динамический диапазон от 3 до 10, чаще от 5 до 8.
Заявленный биосенсор обладает полупериодом окисления (Т1/20Х) находящимся в пределах от 15 до 80 сек, чаще от 40 до 75 сек, как правило, от 50 до 70 сек.
Заявленный биосенсор обладает скоростью восстановления, которая ниже, чем скорость восстановления биосенсоров HyPer и HyPer2. Это означает, что указанный биосенсор демонстрирует больший полупериод восстановления (T1/2RED), чем HyPer и HyPer2. Полупериод восстановления биосенсора по настоящему изобретению более 11 мин, чаще более 13 мин и находится в пределах от 11 до 25 мин, чаще от 13 до 21 мин.
Для определения Т1/20Х и T1/2RED для биосенсоров на пероксид водорода можно использовать биосенсор, экспрессированный в гетерологической системе экспрессии. Например, можно использовать клетки линии HeLa
Kyoto. Для определения Т1/20Х и T1/2RED после трансфекции клеток конструкцией для экспрессии биосенсора в клетках-хозяевах (вектором, кассетой экспрессии и т.д.) и культивацией в подходящих условиях в течение времени, необходимого для наращивания клеток и наработки в них биосенсора в среду культивации добавляют пероксид водорода в определенной концентрации и фиксируют зависимость интенсивности флуоресценции при возбуждении светом с длиной волны 420 нм и 500 нм от времени. На основании полученных данных определяют Т1/20Х и T1/2RED. Также обеспечиваются функциональные биосенсоры, которые по существу сходны с указанным выше биосенсором, где по существу сходны означает, что эти белки имеют аминокислотную последовательность, идентичную последовательности SEQ ID No:04, по крайней мере, на 85% идентичности, обычно, по крайней мере, 90% и чаще, по крайней мере, 95%, (например 95% и выше; 96% и выше, 97% и выше; 98% и выше: 99% и выше или 100% идентичности последовательности).
Например, мутанты биосенсора могут быть получены с использованием стандартных методов молекулярной биологии, как подробно описано в разделе "молекулы нуклеиновых кислот" выше. Примеры обеспечивают общие приемы, и использование стандартных способов, так что специалисты, квалифицированные в данной области, могут легко получить ряд дополнительных мутантов и проверить, было ли изменено биологическое (например, биохимическое, спектральное, и т.д.) свойство. Например, интенсивность флуоресценции может быть измерена с использованием спектрофлуориметра при различных длинах волн возбуждения. Биосенсоры настоящего изобретения присутствуют в среде, отличной от их естественной среды; например, они рекомбинантны. Белки настоящего изобретения могут находиться в выделенном состоянии, что означает, что белки по существу свободны от других белков и других биологических молекул, присутствующих в естественной среде, таких как олигосахариды,
нуклеиновые кислоты и их фрагменты и т.п., где термин "по существу свободны" в этом случае означает, что меньше чем 70%, обычно меньше чем 60% и чаще меньше чем 50% композиции, содержащей выделенный белок, представляет собой некоторые другие биологические молекулы, чем встречающиеся в природе. В некоторых воплощениях, белки присутствуют в по существу очищенной форме, где "по существу очищенная форма" означает очищенная, по меньшей мере, на 95%, обычно, по меньшей мере, на 97% и чаще, по меньшей мере, на 99%.
Заявленные биосенсоры могут быть получены искусственным путем, например, экспрессией рекомбинантной нуклеиновой кислоты, кодирующей последовательность, белка, представляющего интерес, в соответствующем хозяине, как описано выше. Для очистки белка могут применяться любые обычные методики, где подходящие методы очистки белка описаны в Guide to Protein Purification, (Deuthser ed., Academic Press, 1990). Например, лизат может быть приготовлен из исходного источника и очищен с использованием ВЭЖХ, вытеснительной хроматографии, гель-электрофореза, аффинной хроматографии и т.п.
Также обеспечиваются белки слияния, включающие биосенсор настоящего изобретения, слитые с последовательностью внутриклеточной локализации (например, с сигналом локализации в ядре, в пероксисомах, аппарате Гольджи, митохондрии и т.д.). Полипептид, обеспечивающий определенную внутриклеточную локализацию, может быть оперативно присоединен к N-концу и/или С-концу биосенсора. Сигналы внутриклеточной локализации хорошо известны специалистам в данной области и описаны, например, Nakai К. (Advances in Protein Chemistry, Vol.54,2000, p.277-344).
Трансформанты
Нуклеиновые кислоты настоящего изобретения используют для получения трансформантов, включая трансгенных организмов или сайт-специфичных генных изменений в клеточных линиях. Трансгенные клетки, заявленные в изобретении, содержат одну или более нуклеиновых кислот, заявленных по настоящему изобретению, в качестве трансгена. Для целей изобретения любая приемлемая клетка-хозяин может быть использована, включая про-кариотические (например, Escherichia coli, Streptomyces sp., Bacillus subtilis, Lactobacillus acidophilus, и т.д.) или эукариотические клетки-хозяева. Трансгенный организм, заявленный по изобретению, может быть прокариотическим или эукариотическим организмом, включая бактерии, цианобактирии, грибы, растения и животные, в которых одна или больше клеток организма содержат гетерогенную нуклеиновую кислоту, заявленную по изобретению, введенную посредством вмешательства человека, такими способами как технологии трансгеноза, которые известны в данной области.
Выделенная нуклеиновая кислота настоящего изобретения может быть введена в хозяина способами, известными в данной области, например инфицированием, трансфекцией, трансформацией или трансконъюгацией. Заменяющий лист
Способы переноса молекулы нуклеиновой кислоты (то есть. ДНК) в такие организмы широко известны и обеспечивается в ссылках, таких как Sambrook et al. (Molecular Cloning: A Laboratory Manual, 3nd Ed., (2001) Cold Spring Harbor Press, Cold Spring Harbor, NY).
В одном воплощении, трансгенный организм может быть прокариотическим организмом. Способы трансформации прокариотических хозяев хорошо описаны в данной области (например, см. Sambrook et al. Molecular Cloning: A Laboratory Manual, 2nd edition (1989) Cold Spring Harbor Laboratory Press и Ausubel et al.. Current Protocols in Molecular Biology (1995) John Wiley & Sons, Inc).
В другом воплощении, трансгенными организмами могут быть грибы, например дрожжами. Дрожжи широко используются как носители для экспрессии гетерогенного гена (например, см. Goodey et al Yeast biotechnology, D R Berry et al, eds, (1987) Alien and Unwin, London, pp 401-429) и King et al Molecular and Cell Biology of Yeasts, E F Walton and G Т Yarronton, eds, Blackie, Glasgow (1989) pp 107-133). Несколько типов дрожжевых векторов доступны, включая интегративные векторы, которые требуют рекомбинации с геномом хозяина для их поддержки, и автономно реплицирующиеся плазмидные векторы.
В другом воплощении, трансгенными организмами могут быть животные. Трансгенные животные могут быть получены трансгенными способами, известными в данной области и обеспечиваются в ссылках, таких как Pinkert, Transgenic Animal Technology: a Laboratory Handbook, 2nd edition (2203) San Diego: Academic Press; Gersenstein and Vintersten, Manipulating the Mouse Embryo: A Laboratory Manual, 3rd ed, (2002) Nagy A. (Ed), Cold Spring Harbor Laboratory; Blau et al., Laboratory Animal Medicine, 2nd Ed., (2002) Fox J.G., Anderson L.C., Loew F.M., Quimby F.W. (Eds), American Medical Association, American Psychological Association; Gene Targeting: A Practical Approach by Alexandra L. Joyner (Ed.) Oxford University Press; 2nd edition (2000). Например, трансгенные животные могут быть получены гомологичной рекомбинацией, где изменяется эндогенный локус. Альтернативно, конструкция нуклеиновой кислоты включается случайным образом в геном. Векторы для устойчивого включения включают плазмиды, ретро-вирусы и другие животные вирусы, YAC, и т.п. Нуклеиновые кислоты могут быть введены в клетку непосредственно или опосредованно, введением в прекурсор клетки, путем намеренной генетической манипуляции, такой как микроинъекция или инфицирование рекомбинантным вирусом или рекомбинантным вирусным вектором и т.п. Термин «генетическая манипуляция» не включает классическое скрещивание или оплодотворение in vitro, а предпочтительно является направленным на введение рекомбинантных молекул нуклеиновых кислот. Эти молекулы нуклеиновой кислоты могут быть включены в хромосому или они являться внехромосомными реплицирующими ДНК. Конструкции ДНК для гомологичной рекомбинации будут содержать, по меньшей мере, часть нуклеиновой кислоты настоящего изобретения, где ген имеет желательную генетическую модификацию (ции) и включает области гомологии с целевым локусом. Конструкциям ДНК для произвольного включения не обязательно содержать область гомологии с медиатором рекомбинации. Легко могут быть включены маркеры для положительной и отрицательной селекции. Способы получения клеток, имеющих целевые генные модификации, через гомологическую комбинацию известны в данной области. Для различных способов трансфекции клеток млекопитающих, см. Keown et al., Meth. Enzymol. (1990) 185:527-537.
Трансгенные животные могут быть любыми животными, не относящимися к человеку, включая млекопитающее, не относящееся к человеку, (например мышь, крыса), птица или амфибия и т.д., и использованы в функциональном исследовании, скрининге лекарственного средства и т.п."
Также могут быть получены трансгенные растения. Способы получения трансгенных растительных клеток и растений описаны в патентах США №№5767367; 5750870; 5739409; 5689049; 5689045; 5674731; 5656466; 5633155; 5629470; 5595896; 5576198; 5538879; 5484956; раскрытия которых включены сюда ссылкой. Способы получения трансгенных растений также рассмотрены в Plant Biochemistry and Molecular Biology (eds. Lea and Leegood, John Wiley & Sons) (1993) pp.275-295 и в Plant Biotechnology and Transgenic Plants (eds. Oksman-Caldentey and Barz), (2002) 719 p.Например, эмбриогенные эксплантаты, содержащие соматические клетки, могут использоваться для получения трансгенного хозяина. После сбора клеток или тканей, экзогенная ДНК, представляющая интерес, вводится в растительные клетки, при этом известен для такого введения ряд различных способов. При наличии выделенных протопластов, возникает возможность для введения ДНК через опосредованные протоколы передачи гена, включая инкубацию протопластов с очищенной ДНК, такой как плазмида, содержащая целевую экзогенную последовательность, в присутствии поливалентных катионов (например, PEG или PLO); или электропорацию протопластов в присутствии выделенной ДНК, включающей целевую экзогенную последовательность. Протопласты, которые успешно включили экзогенную ДНК, затем отбирают, выращивают в каллус, и, в конечном счете, в трансгенное растение при контакте с подходящими количествами стимулирующих факторов, таких как ауксины и цитокины. Могут использоваться другие подходящие способы получения растения, которые доступны для квалифицированных специалистов в, данной области, такие, как применение "генной пушки", или Agrobacterium-опосредованная трансформация.
Способы применения Биосенсоры настоящего изобретения являются генетически кодируемыми флуоресцентными белками, меняющими спектральные свойства в присутствии перекиси водорода. Они могут быть использованы для мониторинга продукции перекиси водорода внутри живых клеток при различных биологических процессах. Благодаря увеличенному периоду восстановления биосенсоры настоящего изобретения сохраняются в окисленной форме продолжительное время, что позволяет выявлять и отслеживать клетки, подвергшиеся стимулированию продукции перекиси водорода. Для осуществления мониторинга должна быть получена нуклеиновая кислота, кодирующая биосенсор. Получение таких конструкций очевидно для любого специалиста в данной области. Полученная конструкция должна быть встроена в кассету экспрессии (или вектор), обеспечивающую временную или постоянную экспрессию этой нуклеиновой кислоты в клетках-хозяевах. Вектор или кассета экспрессии может содержать элементы, обеспечивающие адресную доставку конструкции в интересующие клетки, или находится в составе частиц, обеспечивающих адресную доставку. После трансфекции клеток вектором и по истечении времени, необходимого для наработки в клетках продукта экспрессии, может быть осуществлен мониторинг продукции перекиси водорода в данных клетках.
Биосенсоры настоящего изобретения находят использование в скрининге препаратов, вызывающих активацию определенных белковых каскадов, например, активацию тирозинкиназ в клеточных линиях. Примером применения биосенсоров могут служить автоматизированные скрининги множества клеток, описанные, например, в патенте США №5989835.
Заявленные биосенсоры также находят использование в применении при сортировке клеток, активированных флюоресценцией, (FACS). В таких применениях заявленный биосенсор, используется для мечения популяции клеток, стимулированных каким-либо фактором к продукции перекиси водорода, и полученная меченая популяция клеток затем подвергается сортировке в устройстве для сортировки клеток, активированных флуоресценцией, как известно в данной области. Способы FACS описаны в патентах США №№5968738 и 5804387.
Заявленные биосенсоры также находят применение как метки продукции перекиси водорода in vivo для трансгенных животных. Например, экспрессия заявленного белка может сопровождаться тканеспецифичными промоторами, где такие способы находят применение в исследованиях для генной терапии, таких как тестирование эффективности лекарственных препаратов на животных моделях. Типичное применение флуоресцентных биосенсоров у трансгенных животных, которое поясняет такие применения, описано в WO 00/02997.
Наборы
Также обеспечиваются в соответствии с настоящим изобретением наборы для использования при осуществлении одного или более вышеописанных применений. В предпочтительных воплощениях наборы обычно включают нуклеиновую кислоту, кодирующую биосенсор, предпочтительно с элементами для экспрессии биосенсора в клетке-хозяине. Например, наборы могут включать вектор, включающий нуклеиновую кислоту, кодирующую заявленный белок. Компоненты наборов обычно присутствуют в соответствующей среде для хранения, такой как водный или буферный раствор, обычно в соответствующей емкости. В отдельных воплощениях набор включает множество различных векторов, каждый их которых кодирует заявленный белок, где векторы предназначены для экспрессии в различных средах и/или при различных условиях. Например, для конститутивной экспрессии, набор включает вектор, имеющий сильный промотор для экспрессии в клетках млекопитающих, и вектор с ослабленным промотором. Могут быть так же включены векторы с множественными клонированными сайтами для выборочной вставши промотора и/или векторы для экспрессии в различных организмах, и т.д.
В дополнение к описанным выше компонентам, заявленные наборы будут дополнительно включать инструкции для осуществления заявленных способов. Эти инструкции могут присутствовать в заявленных наборах в различных формах (например, в печатном варианте или на электроном носителе в виде текстового и/или графического файла) в количестве, одна или более.
Следующие примеры предлагается в качестве иллюстративных, но не ограничивающих.
Примеры
Пример 1
Получение мутантных вариантов биосенсора Ну Per для детекции пероксида водорода.
Для направленного мутагенеза использовали нуклеиновую кислоту, кодирующую HyPer, из коммерчески доступного вектора pHyPer-cyto (Евроген, Россия). Для получения конструкций с содержанием нужных мутаций проводили сайт-направленный мутагенез с использованием "overlap extention" продуктов ПЦР как описано Wurch et all, Methods in Molecular Biology. 12 (9), 653 - 657 (2004). Для ПЦР использовали праймеры, структуры которых показаны в SEQ ID NOs: 5-10, и Tersus полимеразу (Евроген). Использовали следующий режим амплификации в 22 циклах ПЦР: денатурация 95°С - 12 сек; отжиг 55°С - 20 сек; элонгация 72°С - 1 мин. Амплификацию проводили на приборе РТС-100 Thermal Cycler (MJ Reserch). Каждая реакционная проба так же содержала праймеры (0,5 мкМ), эквимолярную смесь dNTP (0,5 мкМ), матричную ДНК (10-100 нг). Для доведения-смеси до нужного объема использовали воду степени очистки milliQ.
После амплификации полученные фрагменты очищали при помощи элек-тофореза в 1% агарозном геле и дальнейшей экстракцией. Выделенные из геля фрагменты ДНК, содержащие требуемую мутацию, объединяли в целую конструкцию, содержащую точечную замену методом «overlap extention» ПЦР (денатурация 95°С - 12 сек; отжиг 55°С - 2 мин; элонгация 72°С - 1 мин, 8 циклов ПЦР). Этот метод подразумевает, что в качестве матрицы и затравки применяют присутствующие в реакционной смеси фрагменты с комплементарными друг другу участками.
Далее амплификацию всей конструкции, полученной на предыдущем этапе, производили методом стандартной ПЦР с добавлением концевых праймеров, структуры которых показаны в SEQ ID NOs: 11, 12. Использовали следующий режим амплификации: 14 циклов ПЦР; денатурация 95°С - 12 сек; отжиг 55°С - 20 сек; элонгация 72°С - 1 мин. Полученные нуклеиновые кислоты клонировали в вектор pQE30 по стандартному протоколу, соответствие структуры вставки проверяли секвени-рованием с использованием стандартных плазмидных праймеров. Были получены следующие новые варианты HyPer, содержащие следующие мутации: H34Y и A406V (HyPer-H34Y-A406V), H34Y (HyPer-H34Y), I32D (HyPer-I32D), F392A (HyPer-F392A) и L46D (HyPer-L46D). Все указанные мутации находятся в области регуляторного домена OxyR, ответственного за димеризацию.
Пример 2
Измерение динамического диапазона и полупериодов окисления и восстановления полученных мутантных вариантов HyPer.
Векторы на основе pQE-30 для экспрессии мутантных вариантов HyPer в Е.coli были получены, как описано в примере 1. Для осуществления пере-клонирования полученных конструктов из вектора pQE30 в вектор pCleGFP (Клонтех, США) фрагменты нуклеиновых кислот, содержащие кодирующую область биосенсора, амплифицировали с использованием праймеров (SEQ ID NO:13, 14), специфических к концам фрагментов и содержащих сайты рестрикции для переклонирования, и полимеразу Tersus (Евроген). Проводили 22 цикла ПЦР в режиме: денатурация 95°С - 12 сек; отжиг 55°С - 20 сек; элонгация 72°С - 1 мин. Полученные фрагменты очищали с помощью фенольно-хлороформной экстракции и лигировали в pCleGFP, предварительно подвергнутый рестрикции по аналогичным сай-там, на место EGFP.
Клетки HeLa Kyoto рассаживали в слайды (p-Slide VI, Ibidi) в среде DMEM, содержащей 10% инактивированной эмбриональной сыворотки (РАА Laboratories) из расчета 5-7 тыс.клеток на одну дорожку, трансфецировали соответствующим вектором и культивировали по стандартному протоколу.
Перед микроскопией среду во всех слайдах заменяли на раствор Хэнкса без бикарбоната натрия. Для визуализации флуоресценции трансфецированных клеток использовали флуоресцентный микроскоп Leica DMI 6000 В. Микроскопию клеток проводили при температуре 37°С, детекцию флуоресценции проводили в двух каналах, соответствующих двум пикам возбуждения хромофора cpYFP (420 нм и 500 нм), эмиссия 510 нм. Для канала соответствующего протонированной форме хромофора cpYFP (420 нм) использовали фильтры EFW Excitation: 427/10 (CFP), Emission: BP 542/27 (YFP). Для депротонированной формы хромофора cpYFP(500 нм) - фильтры Excitation: BP 504/12 (YFP); Emission: BP 542/27 (YFP). Использовали объектив НСХ Р2 ApoLambda blue 63*1,4 Oil. Интервалы съемки составляли 5 секунд. К клеткам добавляли пероксид водорода до конечной концентрации 100 мкМ. Регистрировали сигнал биосенсора и рассчитывали полупериоды окисления и восстановления.
Были получены следующие результаты: сенсоры с мутациями 13 2D, L46D, F392A демонстрировали низкий динамический диапазон по сравнению с исходным HyPer. Амплитуды ответа HyPer-H34Y-A406V и HyPer-H34Y на добавление Н2О2 значительно превосходили таковую для HyPer, не уступая HyPer-2.
HyPer-H34Y демонстрировал более высокие скорости окисления и восстановления по сравнению с HyPer и HyPer-2, тогда как HyPer-H34Y-A406V имел более длительные полупериоды окисления и восстановления, чем HyPer и HyPer-2.
Результаты эксперимента суммированы в Таблице 1. Графики, демонстрирующие динамику сигнала для вариантов биосенсора, показаны на рис.2. Сравнение средних полупериодов окисления и восстановления вариантов биосенсора показаны на рис.3.
Вариант HyPer-H34Y-A406V (SEQ ID NOs:3, 4) был отобран для дальнейших исследований как обладающий наибольшим полупериодом восстановления и широким динамическим диапазоном.
Таблица 1.
Основные характеристики вариантов биосенсора.
|
Пример 3
Тестирование варианта биосенсора HyPer-H34Y-A406V в стабильно трансфецированных клеточных линиях
Линия клеток HeLa Kyoto, стабильно экспрессирующая белок HyPer-H34Y-A406V, была получена с помощью лентивирусной трансдукции. Для этого открытая рамка считывания HyPer-H34Y-A406V, полученная как описано в примере 1 была клонирована в лентивирусный вектор pRRLSIN.EFl.WPRE. Трансдукция была проведена согласно стандартным протоколам. Популяция клеток с наибольшей интенсивностью флуоресценции была отобрана с помощью проточного флуоресцентного клеточного сортера MoFlo (Dako). Полученная линия стабильно экспрессировала HyPer-H34Y-A406V. Добавление в среду H2O2 в различных концентрациях вызывало ожидаемое изменение спектральных характеристик биосенсора, подтверждая его функциональность. Тестирование проводили, как описано в примере 2. Окисленная форма биосенсора детектировалась в клетках по истечении 20-30 мин после добавления пероксида, тогда как в клеточной линии, экспрессирующей HyPer и приготовленной по тому же протоколу, окисленную форму биосенсора не удавалось обнаружить позднее, чем через 5 мин после добавления пероксида водорода.
Пример 4
Получение и тестирование биосенсора HyPer-H34Y-A406V с различной плокализацией в клетках млекопитающих.
Нуклеиновую кислоту, кодирующую HyPer-H34Y-A406V, полученную как описано в примере 1, оперативно встраивали в следующие коммерчески доступные векторы на место флуоресцентных маркеров TagRFP и mKate2:
1. pTagRFP-Golgi (Евроген), содержащий последовательность из бета-1,4-галактозилтрансферазы человека, обеспечивающую локализацию белка в аппарате Гольджи;
2. pTagRFP-mito (Евроген), содержащий последовательность из VIII субъединицы цитохром-С-оксидазы (Rizzuto et al., J Biol Chem. 1989;264(18):10595-600.; Rizzuto et al., Curr Biol. 1995;5(6):635-42), обеспечивающую локализацию белка в митохондриях;
3. pmKate2-peroxi (Евроген), содержащий трипептид SKL, обеспечивающий локализацию белка в пероксисомах. Полученные векторы использовали для трансфекции почечных эпителиальных клеток человека 293 Т. Трансфекция приводила к возникновению флуоресцентного сигнала в ожидаемых компартментах клетки. Тестирование экспрессированных слитых белков на способность реагировать на добавление перекиси водорода было подтверждено, как описано в примере 2.
Все публикации и патентные заявки, цитируемые в настоящем описании, вводятся в настоящее описание посредством ссылки, как если бы каждая отдельная публикация или патентная заявка была конкретно и отдельно введена посредством ссылки. Цитирование любой публикации приводится в соответствии с контекстом и интерпретацией по настоящему изобретению и не должно истолковываться как признание любой такой публикации прототипом данного изобретения.





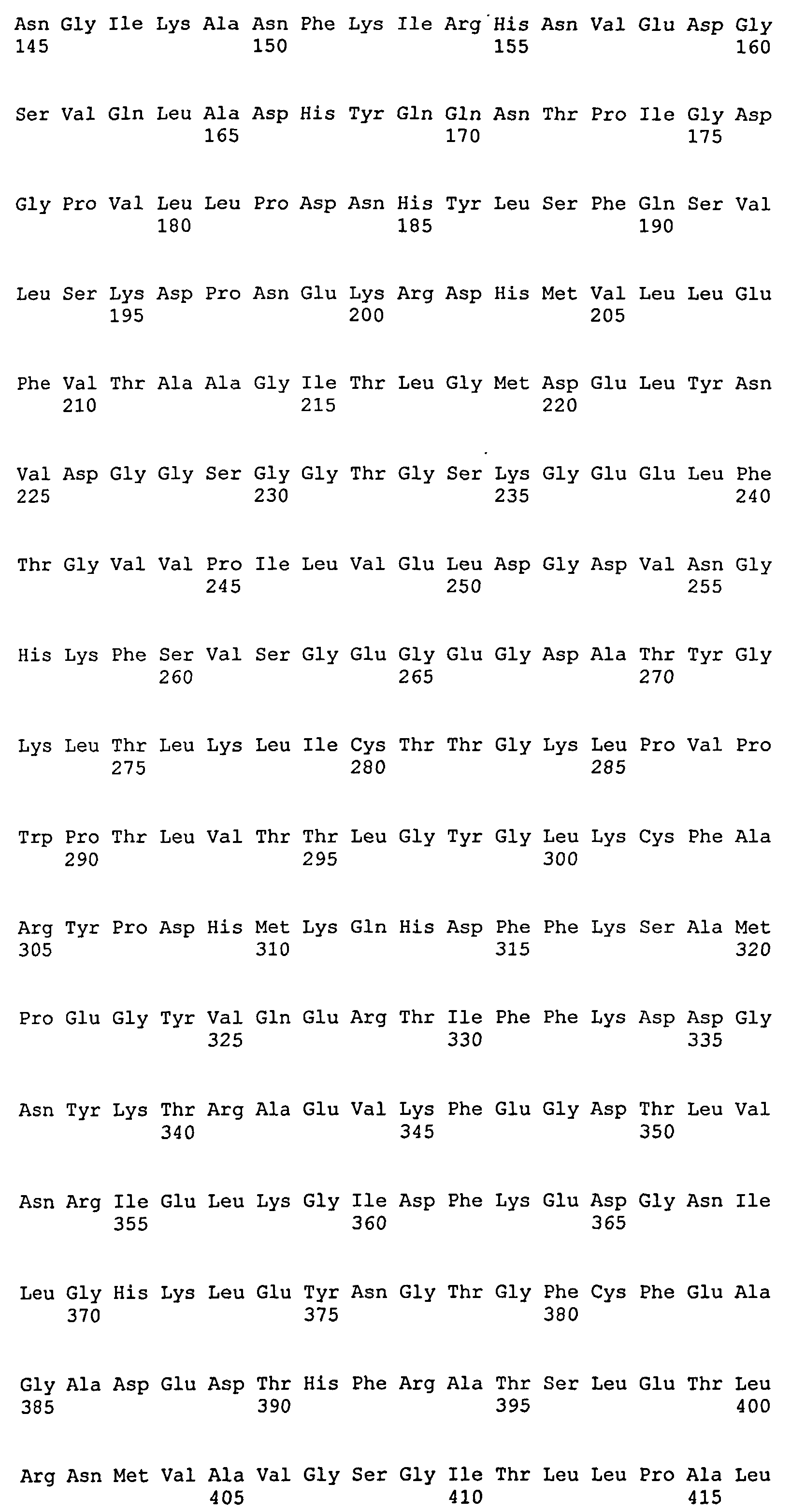
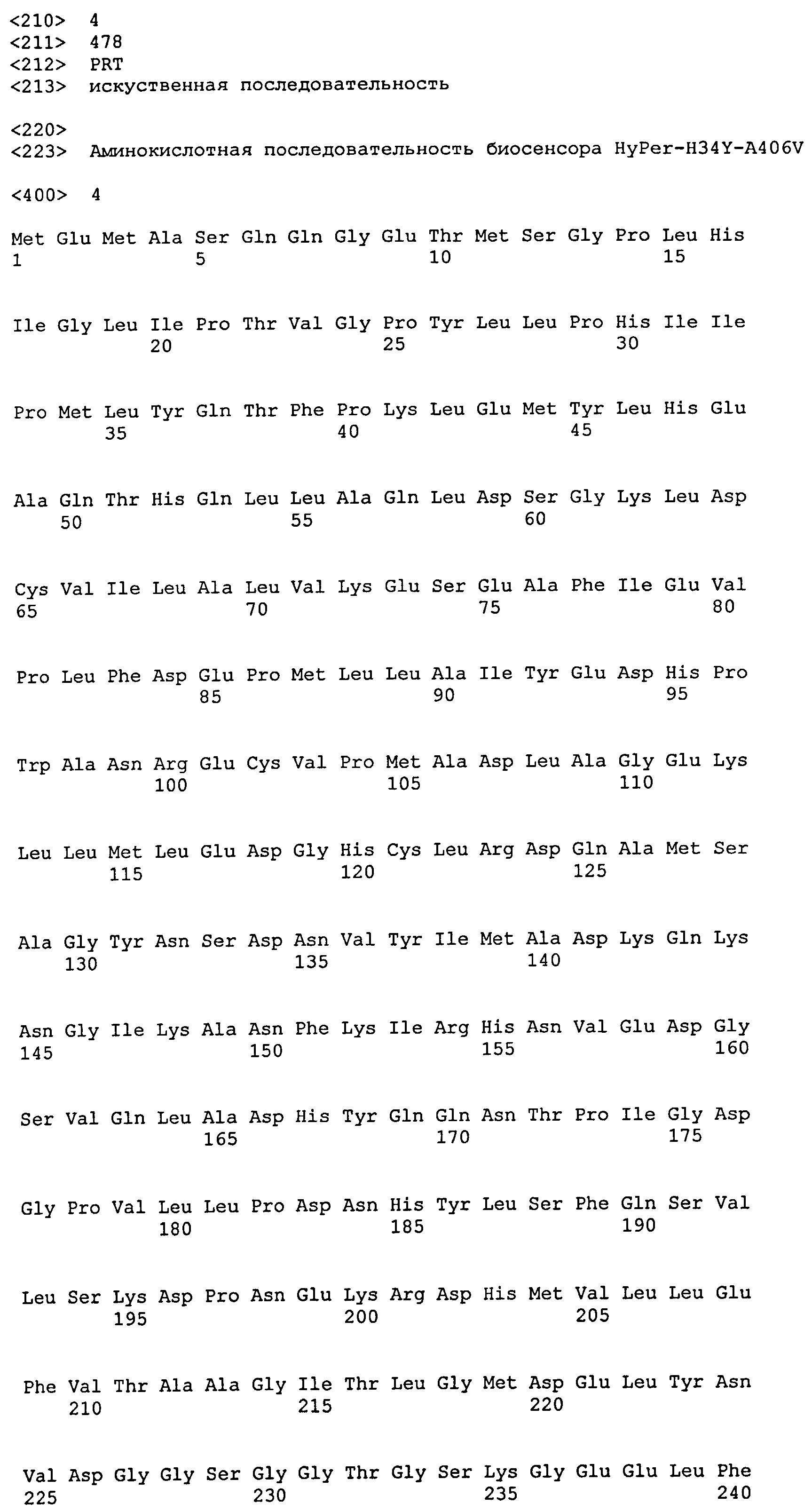
Выделенная нуклеиновая кислота, кодирующая оперативно слитый интрамолекулярный димер белка killerred, кассета экспрессии, клетка, продуцирующая химерный белок и содержащая кассету экспрессии, выделенный химерный белок
Способ оценки содержания пероксида водорода в опухолевых клетках при воздействии на них противоопухолевого препарата
Химерный белок, являющийся флуоресцентным биосенсором для одновременной детекции пероксида водорода и фосфатидилинозитол-3,4,5-трифосфата, нуклеиновая кислота, кодирующая такой белок, кассета экспрессии и эукариотическая клетка-хозяин
Способ холодной непрерывной прокатки тонких стальных полос
Выделенная нуклеиновая кислота, кодирующая флуоресцентный биосенсор, кассета экспрессии, клетка продуцирующая флуоресцентный биосенсор, выделенный флуоресцентный биосенсор
Способ фотодинамической терапии опухолей
Красный флуоресцентный биосенсор для детекции пероксида водорода в живых клетках
Способ регистрации внутриклеточного ph опухолевых клеток
Способ обработки тяжелых переувлажненных почв под яровые зерновые культуры
Модифицированный генетически кодируемый фотосенсибилизатор
Способ прогнозирования жизнеугрожающих желудочковых аритмий у больных сахарным диабетом 2 типа, страдающих хронической сердечной недостаточностью
Способ проведения наркоза в эксперименте
Способ диагностики декомпенсации сахарного диабета 2 типа
Способ стимуляции сперматогенеза в эксперименте
Способ диагностики вибрационной болезни
Выделенная нуклеиновая кислота, кодирующая оперативно слитый интрамолекулярный димер белка killerred, кассета экспрессии, клетка, продуцирующая химерный белок и содержащая кассету экспрессии, выделенный химерный белок
Способ профилактики раневых осложнений после пластики грыжи с использованием сетчатого эндопротеза
Способ лечения хронического неспецифического эндометрита
Способ оценки содержания пероксида водорода в опухолевых клетках при воздействии на них противоопухолевого препарата
Химерный белок, являющийся флуоресцентным биосенсором для одновременной детекции пероксида водорода и фосфатидилинозитол-3,4,5-трифосфата, нуклеиновая кислота, кодирующая такой белок, кассета экспрессии и эукариотическая клетка-хозяин

