Результат интеллектуальной деятельности: СИСТЕМА ДЛЯ СТАБИЛЬНОЙ ЭКСПРЕССИИ ОПУХОЛЬ-АССОЦИИРОВАННЫХ АНТИГЕНОВ НА ОСНОВЕ ЛЕНТИВИРУСНОГО ВЕКТОРА
Вид РИД
Изобретение
Изобретение относится к области биотехнологии. Может быть использовано для экспрессии опухоль-ассоциированных антигенов с их последующим включением в эндосому через систему комплекса гистосовместимости МНС II.
Ежегодно в мире от злокачественных новообразований умирает около 7 млн. человек. По данным ВОЗ онкологические заболевания по частоте встречаемости стоят на 2 месте, а в некоторых промышленно развитых регионах - на первом месте. Традиционно до недавнего времени основными методами лечения считались: хирургический, цитотоксическая химиотерапия и лучевая терапия.
Достижения иммунологической науки за последние годы привели к созданию принципиально новых подходов и методов терапии онкологических заболеваний.
Так, уже более 40 лет тому назад было показано, что многие опухоли иммуногенны. Однако цитотоксические Т-лимфоциты не активируются при появлении раковых клеток и их специфических антигенов и не способны секретировать цитокины, которые они секретируют при встрече с чужеродными антигенами. Поэтому одна из стратегий борьбы с опухолями заключается в стимулировании иммунной системы к узнаванию опухолевых клеток.
В частности, были предприняты попытки создания индивидуальных противоопухолевых вакцин (А.Ю. Барышников, Принципы и практика вакцинотерапии рака, Бюллетень СО РАМН, №2(112). 2004, стр. 59-63).
В настоящее время значительное внимание уделяется применению дендритных клеток и препаратов (вакцин) на их основе.
Дендритные клетки являются профессиональными антиген-презентирующими клетками (АПК) и играют ключевую роль в формировании первичного иммунного ответа. Важной особенностью дендритных клеток является их способность захватывать из окружающей среды различные антигены при помощи пино- и эндоцитоза. После поглощения антигена дендритные клетки мигрируют в лимфоидные органы, где презентируют антигенные детерминанты в контексте главного комплекса гистосовместимости (МНС) различным популяциям Т-лимфоцитов. Существует два типа рецепторов главного комплекса гистосовместимости. Рецепторы МНС I экспрессируются во всех ядерных клетках организма, осуществляя механизм распознавания «свой-чужой» и презентируют эндогенные антигены, такие как вирусные или раковые антигены. В данном случае связывание рецептора и антигена происходит в эндоплазматическом ретикулуме. Рецепторы МНС II экспрессируются антигенпрезентирующими клетками и другими типами клеток крови, например В-клетками, и презентируют экзогенные антигены, поглощенные в результате эндоцитоза. В отличие от МНС I, связывание МНС II и антигена происходит в области эндосомы. В случае МНС I презентация чужеродного антигена, например, вирусного пептида, вызывает активацию CD8+ цитотоксических Т-лимфоцитов и приводит к немедленному иммунному ответу. В результате происходит уничтожение зараженной клетки. Презентация чужеродного экзогенного пептида молекулами комплекса МНС II вызывает активацию как CD8+ цитотоксических Т-лимфоцитов, так и активацию CD4+ хэлперных Т-лимфоцитов, что приводит к возникновению механизма «иммунной памяти». Такой механизм характеризуется возникновением популяции В-лимфоцитов, продуцирующих специфические антитела и популяции более специфичных цитотоксических Т-лимфоцитов, нацеленных на чужеродный антиген.
Известен способ индукции высокого уровня CD4+ и CD8+ Т-клеточного ответа и специфических типов цитокинов с целью достижения высокого уровня защиты и терапевтической активности за счет иммуногенных HER2-специфичных конструкций полиэпитопа, содержащих CTL и/или эпитопы Th и оптимизированные последовательности спейсера. Конструкции могут использоваться для продуцирования антигенпредставляющих клеток, например, дендритных клеток (DC), для представления желаемых эпитопов лимфоцитам (заявка ЕА 201290813 (А1), опубл. 2013-04-30).
Известен способ экспрессии полиэпитопных опухоль-ассоциированных антигенов в дендритных клетках, способных стимулировать специфические цитотоксические клетки, основанный на использовании рекомбинантной плазмидной ДНК pCI-UB-POLYEPI, которая содержит 11 эпитопов опухоль-ассоциированных антигенов колоректального рака, имеет размер 6355 п.н. (патент РФ 2507265, опубл. 20.02.2014). Данный источник может быть указан в качестве ближайшего аналога-прототипа.
Задачей изобретения является создание системы, способной включать специфические антигенные детерминанты через систему комплекса МНС II, что позволяет значительно расширить возможности применения дендритных клеток пациента, и тем самым увеличить эффективность противораковой терапии.
Задача решается ДНК конструкцией на основе лентивирусого вектора для экспрессии опухоль-ассоциированных антигенов с последующим их транспортом в эндосому через систему комплекса МНС П.
Предлагаемые плазмидные лентивирусные конструкции включают последовательность инвариантного домена CD74/Ii комплекса МНС II, в качестве лидирующей сигнальной области, присоединенной к целевым полипептидам (антигенам). CD74/Ii выступает в роли сигнальной последовательности, направляющей транспортировку комплекса МНС II в эндосомальный компартмент.
Техническим результатом изобретения является создание универсальной лентивирусной системы, позволяющей транспортировать широкий спектр слитых белков-антигенов в эндосому, где они будут расщеплены и презентированы молекулами комплекса МНС II.
В отличие от случая с комплексом МНС I, МНС II препятствует присоединению эндогенных пептидных молекул в области шероховатого ЭПР (эндоплазматического ретикулума). Также CD74/Ii выступает в роли сигнальной последовательности, направляющей транспортировку комплекса МНС II в эндосомальный компартмент. В эндосоме МНС II претерпевает протеолитическое расщепление, в результате которого происходит отщепление Ii домена и присоединение уже протеолизированных экзогенных пептидов (антигенов). Таким образом, использование последовательности CD74/Ii в качестве лидирующей сигнальной области, присоединенной к целевым полипептидам (антигенам), экспрессированным с плазмидных ДНК или из генома дендритной клетки, позволит транспортировать такие слитые белки в эндосому, где они будут расщеплены и презентированы молекулами комплекса МНС II.
Согласно изобретению экспрессии может подлежать широкий спектр опухоль-ассоциированных антигенов, например: СБА, TGF-PRII (колоректальная карцинома), TAG-72 (карцинома простаты), HPV Е6, Е7 (цервикальная карцинома), BING-4, SSX-2, TRP-1/-2 ( меланома), Cyclin-B1, EphA3, Her2/neu (мульти рак), Ер-САМ (рак груди), простато-специфический антиген (рак простаты), BRCA1/2 (карцинома груди и яичников), MUC1 ( дуктальная карцинома), Ig, ТСЯ(лимфома) и т.п.
Для доставки последовательностей антигенов, слитых с сигнальными последовательностями CD74/Ii цепей, в антигенпрезентирующие дендритные клетки использована методика лентивирусного переноса целевых последовательностей.
Использование лентивирусных векторов имеет несколько значительных преимуществ по сравнению с такими подходами, как трансфекционный перенос плазмидных ДНК или электропорирование клеток ДНК конструкциями:
- лентивирусные векторы способны быстро интегрироваться в геном клетки-мишени, обеспечивая стабильную экспрессию трансгена;
- лентивирусные векторы позволяют клонировать достаточно длинные последовательности ДНК до 10 т.п.о, при этом сохраняют высокий уровень упаковки вирусных частиц и свою трансдукционную способность;
- лентивирусные векторы обладают широким тропизмом за счет внедрения типирующих белков в оболочку вирусных частиц. Данное свойство позволяет значительно увеличить трансдукционную способность вируса для определенного типа клеток, что крайне важно в работе с труднотрансфецируемыми первичными культурами клеток крови;
- лентивирусные векторы позволяют доставить ДНК даже в труднотрансфецируемые, неделящиеся клетки, в том числе, первичные дифференцированные клетки, такие как дендритные клетки и лимфоциты.
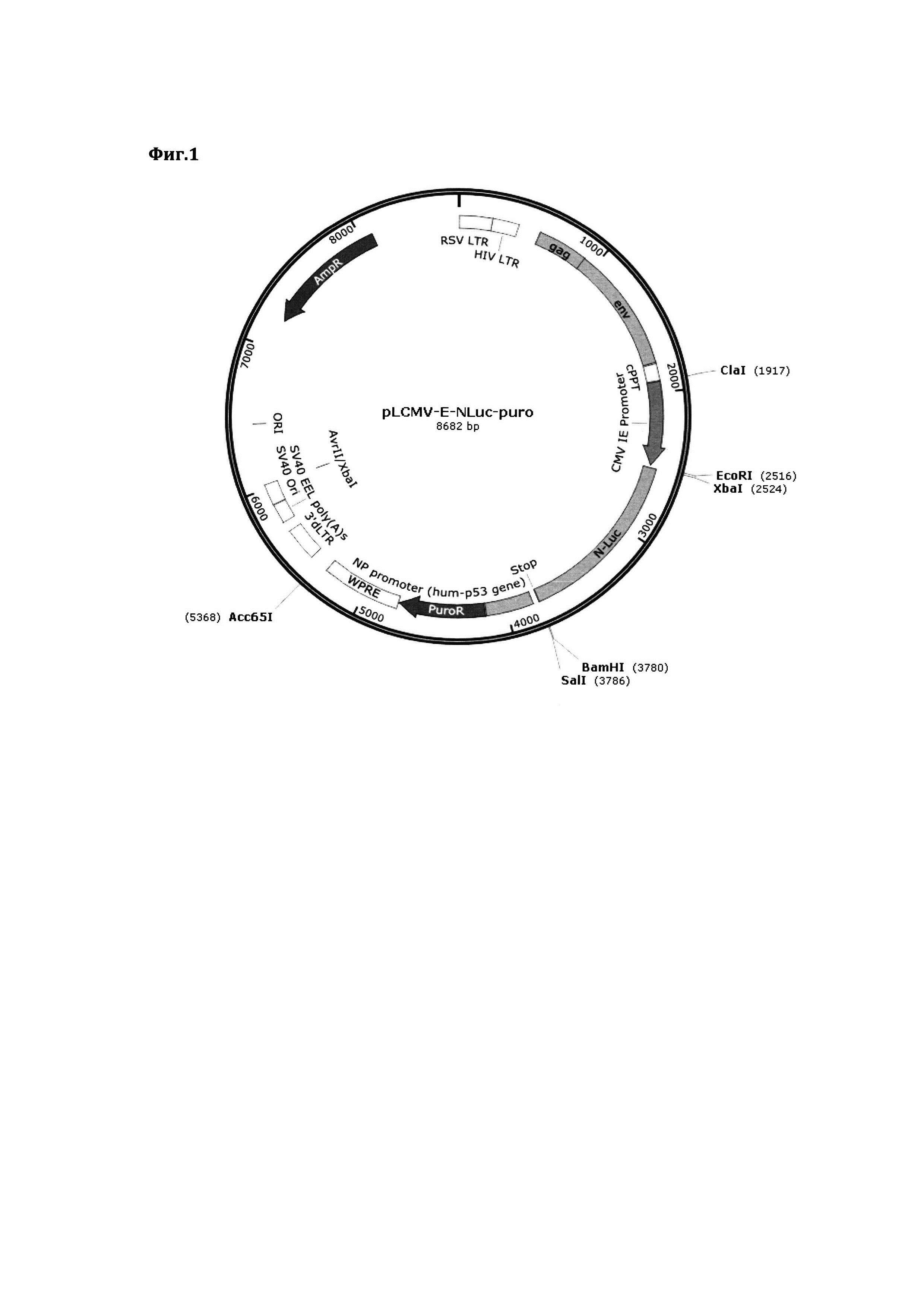
В качестве основного генетического каркаса, формирующего лентивирусную векторную систему использовалась ранее сконструированная нами универсальная плазмидная лентивирусная ДНК конструкция (Фиг. 1). Изображение иллюстрирует схему плазмидной лентивирусной ДНК-конструкции pLA-CMV-ELuc-puro.
В состав данной плазмиды входит ряд ключевых генетических элементов:
- AmpR - последовательность устойчивости к ампициллину, необходимая для амплификации плазмидной ДНК в клетках бактериального штамма E. coli;
- HIV 5' и 3' LTR - длинные концевые повторы ретровирусов (в частности, вируса иммунодефицита человека), обеспечивают встраивание последовательности генома вируса в геном клетки хозяина;
- сРРТ (central polypurine tract) - центральный полипуриновый тракт, способствует встраиванию ДНК вируса в геном митотически неактивных клеток;
- CMV - промотерный регион, обеспечивает стабильный и высокий уровень экспрессии 3' нижележащей последовательности ДНК;
- NLuc - участок N-концевой области люциферазы, на 5' и 3' концах содержит ряд консенсусных участков, распознаваемых ферментами рестрикции-модификации;
- WPRE (Woodchuck hepatitis virus Posttranscriptional Regulatory Element) - энхансерный регион, способствует повышению уровня экспрессии целевых белков.
Презентация опухоль-ассоциированных антигенов, опосредованная молекулами главного комплекса гистосовместимости II, напрямую зависит от эффективного переноса новоэксперссированного антигена в область эндосомального компартмента. В связи с тем, что одним из основных регуляторов транспортировки МНС II в эндосому является белок CD74/Ii, который используют в качестве сигнальной последовательности, направляющей антиген в эндосому. Однако, большая часть последовательности полипептида служит для формирования особой пространственной структуры белка, необходимой для его взаимодействия с молекулой МНС II. За саму транспортировку CD74 отвечает лишь небольшая функциональная область, которая может использоваться отдельно.
Более того, в данный N-концевой домен может входить несколько вариантов еще более коротких последовательностей, отвечающих за перенос CD74/Ii-MHC II. Для определения сигнальной последовательности, приводящей к наилучшей загрузке в МНС II было протестировано несколько вариантов сигнальных пептидов CD74/Ii-рецептора.
Для амплификации была использована серия праймеров (табл). Изображение иллюстрирует последовательности праймеров для амплификации различных сигнальных участков N-концевого домена инвариантной цепи.
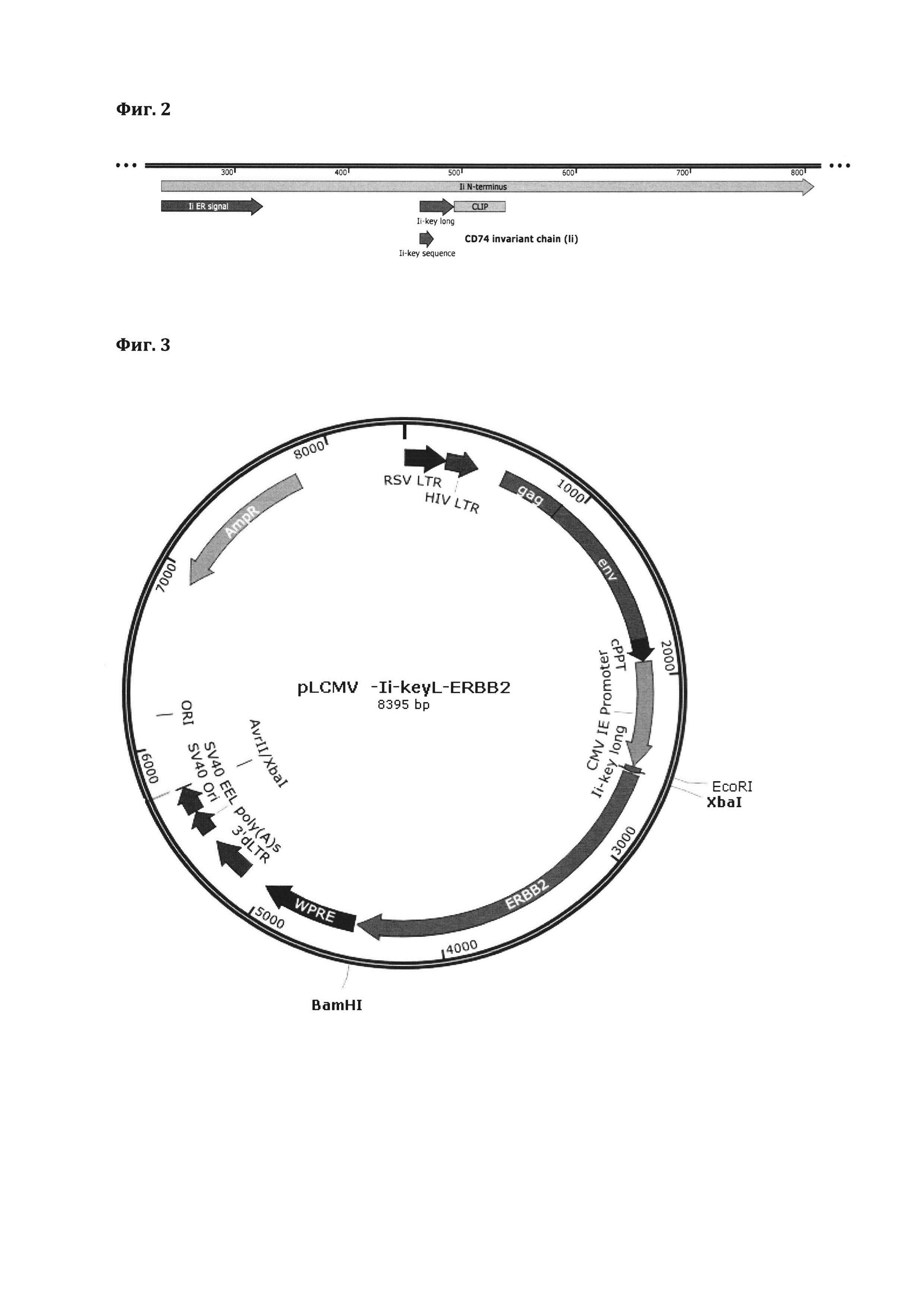
Схема последовательности белка CD74 с отмеченными участками сигнальных пептидов эндосомальной локализации представлена на фиг. 2
В качестве наиболее оптимального варианта отобран удлиненный вариант сигнального участка Ii key (далее в тексте обозначается как "Ii key long"). В качестве опухолевого антигена используют белок ERBB2 (HER2/neu). Рецептор ERBB2 является одним из высокоспецифичных онкологических маркеров, за счет того, что повышенная амплификация и экспрессия данного рецептора характерна для ряда агрессивных типов рака молочной железы и некоторых других видов злокачественных опухолеобразований. Рецептор также ERBB2 является одним из самых больших специфичных опухолевых антигенов, поэтому успешная презентация антигенных детерминант такой молекулы позволит судить об успешности загрузки остальных опухоль-ассоциированных антигенов.
Таким образом, получена лентивирусная ДНК-конструкция, несущая в своем составе последовательность опухоль-ассоциированного антигена ERBB2/HER2 и последовательность сигнального участка гена CD74, обеспечивающего транспорт данного антигена в эндосому (фиг. 3). Изображение иллюстрирует схему лентивирусной плазмидной ДНК конструкции pLA-CMV-Ii-key-long-ERBB2, содержащей последовательность сигнального участка Ii key long. Данная конструкция является одним из предпочтительных, но не ограничивающих изобретение вариантов.
Изобретение иллюстрировано следующим графическим материалом:
Фиг. 1 - Схема плазмидной лентивирусной ДНК-конструкции pLA-CMV-ELuc-puro
Фиг. 2 - Схема последовательности белка CD74 с отмеченными участками сигнальных пептидов эндосомальной локализации
Фиг. 3 - Схема лентивирусной плазмидной ДНК конструкции pLA-CMV-Ii-key-long-ERBB2, содержащей последовательность сигнального участка Ii key long.
Табл. - Последовательности праймеров для амплификации различных сигнальных участков N-концевого домена инвариантной цепи.
SEQ ID№1 CD74 N-концевой участок
Нуклеотидная последовательность
ATGGATGACCAGCGCGACCTTATCTCCAACAATGAGCAACTGCCCATGCTGGGCCGGCGCCCTGGGGCCCCGGAGAGCAAGTGCAGCCGCGGAGCCCTGTACACAGGCTTTTCCATCCTGGTGACTCTGCTCCTCGCTGGCCAGGCCACCACCGCCTACTTCCTGTACCAGCAGCAGGGCCGGCTGGACAAACTGACAGTCACCTCCCAGAACCTGCAGCTGGAGAACCTGCGCATGAAGCTTCCCAAGCCTCCCAAGCCTGTGAGCAAGATGCGCATGGCCACCCCGCTGCTGATGCAGGCGCTGCCCATGGGAGCCCTGCCCCAGGGGCCCATGCAGAATGCCACCAAGTATGGCAACATGACAGAGGACCATGTGATGCACCTGCTCCAGAATGCTGACCCCCTGAAGGTGTACCCGCCACTGAAGGGGAGCTTCCCGGAGAACCTGAGACACCTTAAGAACACCATGGAGACCATAGACTGGAAGGTCTTTGAGAGCTGGATGCACCATTGGCTCCTGTTTGAAATGAGCAGGCACTCCTTGGAGCAAAAGCCCACTGACGCTCCACCG
Аминокислотная последовательность
MDDQRDLISNNEQLPMLGRRPGAPESKCSRGALYTGFSILVTLLLAGQATTAYFLYQQQGRLDKLTVTSQNLQLENLRMKLPKPPKPVSKMRMATPLLMQALPMGALPQGPMQNATKYGNMTEDHVMHLLQNADPLKVYPPLKGSFPENLRHLKNTMETIDWKVFESWMHHWLLFEMSRHSLEQKPTDAPP
SEQ ID №2 Ii-ER сигнальная последовательность
Нуклеотидная последовательность
ATGGATGACCAGCGCGACCTTATCTCCAACAATGAGCAACTGCCCATGCTGGGCCGGCGCCCTGGGGCCCCGGAGAGCAAGTGCAGCCGC
Аминокислотная последовательность
MDDQRDLISNNEQLPMLGRRPGAPESKCSR
SEQ ID №3 Ii key сигнальная последовательность
Нуклеотидная последовательность
CTGCGCATGAAG
Аминокислотная последовательность
LRMK
SEQ ID №4
Ii key long (удлиненная сигнальная последовательность)
Нуклеотидная последовательность
CTGCGCATGAAGCTTCCCAAGCCTCCCAAG
Аминокислотная последовательность
LRMKLPKPPK
SEQ ID №5 CLIP сигнальная последовательность
Нуклеотидная последовательность
CCTGTGAGCAAGATGCGCATGGCCACCCCGCTGCTGATGCAGGCG
Аминокислотная последовательность
PVSKMRMATPLLMQA

Полипептид из морской анемоны heteractis crispa, обладающий анальгетическим действием
Лигнан, обладающий анальгетическим действием
Способ включения квантовых точек методом соосаждения в пористые частицы карбоната кальция
Флуоресцентный зонд и тест-система для определения активности фосфолипазы а2
Способ определения неспецифической устойчивости патогенных микроогранизмов к антибиотикам на основании измерения каталитической активности фосфодиэстераз, расщепляющих циклический дигуанозинмонофосфат
Способ длительного хранения in vitro растений осины
Рекомбинантные плазмидные днк, кодирующие гибридные полипептиды со свойствами красного флуоресцентного белка mcherry, для продуцирования гибридных флуоресцентных белков в escherichia coli
Способ оценки чувствительности клеток рака легкого к доксорубицину на основании уровней экспрессии маркерных генов и набор для его осуществления
Способ формирования многофункциональных микросистем
Способ выявления белков в разных типах клеток млекопитающих и человека с помощью флуоресцеин-5-изотиоцианата на микроскопическом уровне
Способ диагностики рака молочной железы
Аттенуированный штамм вируса сендай
Способ получения системы направленной доставки белковых молекул (онколитических белков) в опухолевые ткани на основе активированных лимфоцитов
Способ определения огнестойкости кирпичных столбов с растворной обоймой
Набор синтетических олигонуклеотидов для определения уровней экспрессии гена pdlim4
Экспрессионный плазмидный лентивирусный вектор для гетерологичной экспрессии рекомбинантного человеческого белка cd47
Репортерная система на основе лентивирусных репортерных конструкций для изучения белок-белковых взаимодействий
Аптамеры, специфичные к внеклеточному гликозилированному домену человеческого рецептора cd47
Экспрессионный плазмидный лентивирусный вектор для гетерологической экспрессии рекомбинантного человеческого белка cd44
Селективный бифункциональный препарат на основе фрагментов одноцепочечных антител верблюжьих, нацеленный на опухолевые рецепторы cd47/erbb2, предназначенный для терапии злокачественных новообразований

