Результат интеллектуальной деятельности: СПОСОБ КОНСЕРВАЦИИ И РАСКОНСЕРВАЦИИ ЖИВЫХ КЛЕТОК
Вид РИД
Изобретение
Изобретение относится к областям удовлетворения жизненных потребностей человека и химии, а именно - к биотехнологии консервации живых клеток, может быть использовано в медицине, ветеринарии, сельском хозяйстве и промышленных биотехнологиях.
Более детально - предполагаемое изобретение относится к созданию способа подготовки живых клеток к консервации за счет доставки сахара трегалозы в консервируемые живые клетки и последующего удаления трегалозы с использованием белка TRET-1 насекомого Polypedilum vanderplanki. Разрабатываемый способ предназначен для использования при консервации живых клеток с целью их (живых клеток) хранения и транспортировки, например - при консервации ооцитов.
В области хранения биоматериалов известные технологии консервации живых клеток непригодны для некоторых чувствительных клеток, например - ооцитов некоторых животных, в том числе свиней. Это связано с тем, что используемые технические приемы и материалы при консервации и хранении не обеспечивают защиту чувствительных клеток на достаточном для предотвращения их гибели уровне. Заявляемое техническое решение нацелено на создание способа подготовки живых клеток к консервации путем доставки в защищаемые клетки защитного агента - трегалозы - с использованием белка TRET-1. Известно, что белок TRET-1 является эффективным белком-транспортером трегалозы, свойственным насекомому P. vanderplanki [1].
Заявляемый способ предполагает увеличение эффективности доставки трегалозы по сравнению с известными способами консервации живых клеток, и удаление трегалозы из живых клеток по мере необходимости. Основой предлагаемого подхода является предшествующее консервации введение белка TRET-1 в живые клетки, доставка трегалозы в живые клетки путем добавления ее (трегалозы) в состав окружающей клетки среды (среды культивирования, среды консервации и пр.), и последующее удаление трегалозы из клеток путем замены окружающей клетки среды на среду, не содержащую трегалозу.
Общеизвестно, что наиболее крупным и сложно устроенным живым организмом, способным восстанавливать жизнедеятельность после полного высыхания (обезвоживания на воздухе до влажности менее 2%) является личинка насекомого P. vanderplanki [2]. Такая способность личинки P. vanderplanki и составляющих ее клеток обусловлена сложным сочетанием молекулярных факторов, защищающих внутриклеточные компоненты при обезвоживании. В обезвоженном состоянии личинка P. vanderplanki характеризуется высокой устойчивостью к неблагоприятным воздействиям окружающей среды, таким как действие высокой температуры, радиация и других [2]. В обезвоженном состоянии личинки P. vanderplanki могут длительно (10 лет и более), храниться с сохранением жизнеспособности [3].
Одним из ключевых механизмов, позволяющих личинкам P. vanderplanki сохранять жизнеспособность клеток организма во время обезвоживания и иных стрессов, является способность данных личинок накапливать внутри клеток сахар трегалозу [4]. Трегалоза предотвращает денатурацию внутриклеточных белков, разрушение клеточных мембран, действие активных форм кислорода [5]. Трегалоза оказывает защитный эффект на живые клетки многих организмов при различных видах стресса, в том числе - при их высушивании или заморозке [6-8].
Показано, что защитное действие трегалозы существенно усиливается при ее (трегалозы) доставке внутрь живой клетки [9]. Однако оболочки клеток (окружающие живые клетки мембраны) непроницаемы для трегалозы. Кроме того, клетки млекопитающих и подавляющего большинства иных организмов не способны к синтезу трегалозы или ее эффективному транспорту через клеточную мембрану.
Единственным механизмом транспорта трегалозы у клеток большинства организмов является протекающий с низкой скоростью неспецифический процесс эндоцитоза, непригодный для создания внутри клеток концентраций трегалозы, достаточных для обеспечения защитного эффекта [10]. Доставка трегалозы внутрь живых клеток за счет химических агентов сопряжена с побочным действием (химических агентов) на живые клетки [8]. Кроме того, доставка трегалозы в живые клетки с помощью известных способов не позволяет вывести трегалозу из живых клеток после их расконсервации (вывода клеток из законсервированного состояния). Необходимость искусственного удаления трегалозы из клеток обусловлена тем, что большинство живых клеток неспособны расщеплять трегалозу ввиду естественного для них отсутствия в них фермента трегалазы [11]. Это (длительное наличие трегалозы после расконсервации) вызывает нарушения естественного метаболизма клеток ввиду осмотического стресса и невозможности повторной активации поврежденных (например - денатурированных) белков в присутствии трегалозы [12].
С учетом вышесказанного, разработка эффективного способа доставки трегалозы в живые клетки является основой для создания нового способа их (живых клеток) консервации, хранения и транспортировки. Основные требования к разрабатываемым способам доставки трегалозы в живые клетки - обеспечение доставки трегалозы в живые клетки, причем - с обеспечением возможности последующего удаления трегалозы из клеток после расконсервации.
Известно техническое решение [13] под названием «Cryopreservation of human adult and fetal pancreatic cells and human platelets» (Криохранение клеток печени человека (у взрослых и плода) и тромбоцитов человека). Сущность [13] заключается в использовании комбинации трегалозы и вещества диметилсульфоксид (далее по тексту - ДМСО) в качестве криопротектора (защитного компонента при криохранении, т.е. консервации путем замораживания). Решение [13] предлагается для криохранения островковых клеток человека, представляющих собой специфический тип клеток печени, и их аналогов, в том числе - человеческих тромбоцитов. По [13], доставка трегалозы в живые клетки осуществляется за счет ее (трегалозы) абсорбции клетками из культуральной среды при охлаждении защищаемых клеток со скоростью 1°С в минуту от температуры культивирования до обеспечивающей фазовый переход мембран температуры (плюс 15-20°С). Указанный фазовый переход представляет собой переход материала мембран клеток из жидкокристаллического состояния в состояние геля при понижении температуры. При фазовом переходе нарушается целостность мембран и они (мембраны клеток) становятся проницаемыми для трегалозы. Ввиду низкой скорости охлаждения, фазовый переход длится некоторое время, достаточное для доставки трегалозы в живые клетки. В [13] добавление ДМСО не способствует доставке трегалозы в клетки в необходимом количестве, поскольку данное вещество (ДМСО) вносится в культуральную среду непосредственно перед замораживанием клеток, и до консервации путем замораживания действует весьма кратковременно.
Недостатком [13] является отсутствие возможности вывести трегалозу из живых клеток после их (клеток) размораживания (расконсервации). Сохраняющаяся длительное время в клетках трегалоза оказывает токсический эффект (отравляющее воздействие) на клетки ввиду осмотического стресса (для клеток) и невозможности повторной активации поврежденных, например - денатурированных, белков в присутствии трегалозы [12]. Следствием недостатка является невозможность использования [13] для оптимального использования трегалозы в качестве защитного компонента при консервации и хранении разнообразных типов живых клеток для их последующего использования.
Известно техническое решение [14] под названием «А method of cryopreserving cells and tissues by liposomal delivery of sugars to enhance post-thaw viability» (Метод криохранения клеток и тканей с липосомальной доставкой Сахаров для увеличения жизнеспособности после размораживания). Сущность [14] выражается в заключении сахаров-криопротекторов (трегалозы, сукрозы, рафинозы и стахиозы) в липосомы (пузырьки жидкости, окруженные липидной мембраной подобно живым клеткам) и их последующей доставке в клетки. Доставка трегалозы и прочих указанных выше Сахаров осуществляется за счет естественного физико-химического взаимодействия липосом с мембраной живых клеток. Решение [14] описывает также различные режимы охлаждения и заморозки клеток после внесения липосом, различные варианты состава среды, используемой для заморозки и различные типы защищаемых клеток.
Недостатком [14] является отсутствие возможности вывести трегалозу из живых клеток после их (клеток) расконсервирования. Наличие трегалозы после расконсервации вызывает нарушения естественного метаболизма клеток [12]. Недостаток [14] существенно ограничивает область его применения, например -для консервации клеток с целью получения биотехнологических продуктов.
Известно техническое решение [15] под названием «А freezing medium composition for cryopreserving amniotic fluid-derived stem cells and a method for cryopreserving the same» (среда для криохранения стволовых клеток из амниотической жидкости и метод их криохранения) по совокупности совпадающих признаков наиболее близкое к заявляемому техническому решению и принятое в качестве прототипа. Сущность [15] заключается в использовании для консервации состава среды, включающего трегалозу, каталазу, ДМСО и вещество zVAD-fmk, предотвращающее апоптоз (смерть клеток). Прототип [15] предусматривает использование ДМСО в качестве увеличивающего проницаемость мембран криопротектора, обеспечивающего транспорт трегалозы внутрь клеток. В отличие от [14], прототип предусматривает длительное воздействие ДМСО, достаточное для обеспечения транспорта трегалозы. Прототип [15] содержит описание метода криохранения клеток с использованием среды указанного выше состава.
Недостатком прототипа [15] является отсутствие возможности вывести трегалозу из живых клеток после их (клеток) расконсервации (размораживания). Наличие трегалозы после расконсервации вызывает нарушения естественного метаболизма клеток ввиду осмотического стресса и невозможности повторной активации поврежденных (например - денатурированных) белков в присутствии трегалозы [12]. Кроме того, применение ДМСО в качестве обеспечивающего проницаемость мембран для трегалозы агента сопровождается токсическим эффектом (результатом действия ДМСО) на клетки, что отмечается авторами [15]. Недостатки делают невозможным использование прототипа [15] для применения трегалозы в качестве оптимального защитного компонента при консервации и хранении живых клеток.
Целью изобретения является разработка способа консервации и расконсервации живых клеток с сохранением жизнеспособности клеток, расширение возможностей консервации, хранения живых клеток для последующего использования, расширение перечня способов сохранения и области применения живых клеток.
Цели достигают тем, что в живые клетки вводят белок TRET-1 путем экспрессии содержащей ген TRET-1 насекомого Polypedilum vanderplanki генетической конструкции, после введения белка TRET-1 в живые клетки при необходимости осуществляют проверку наличия белка в живых клетках непосредственно после чего осуществляют доставку трегалозы в указанные живые клетки путем добавления трегалозы в окружающую клетки среду, а при расконсервации осуществляют удаление трегалозы из живых клеток путем замены окружающей клетки среды с трегалозой на среду без трегалозы. Расконсервированные клетки используют по назначению.
Заявляемое техническое решение иллюстрируется материалами, оформленными в соответствии со стандартами ВОИС.
Фиг. 1 - Аминокислотная последовательность белка TRET-1.
Фиг. 2 - Нуклеотидные последовательности праймеров (коротких фрагментов ДНК) для получения кодирующего белок TRET-1 гена.
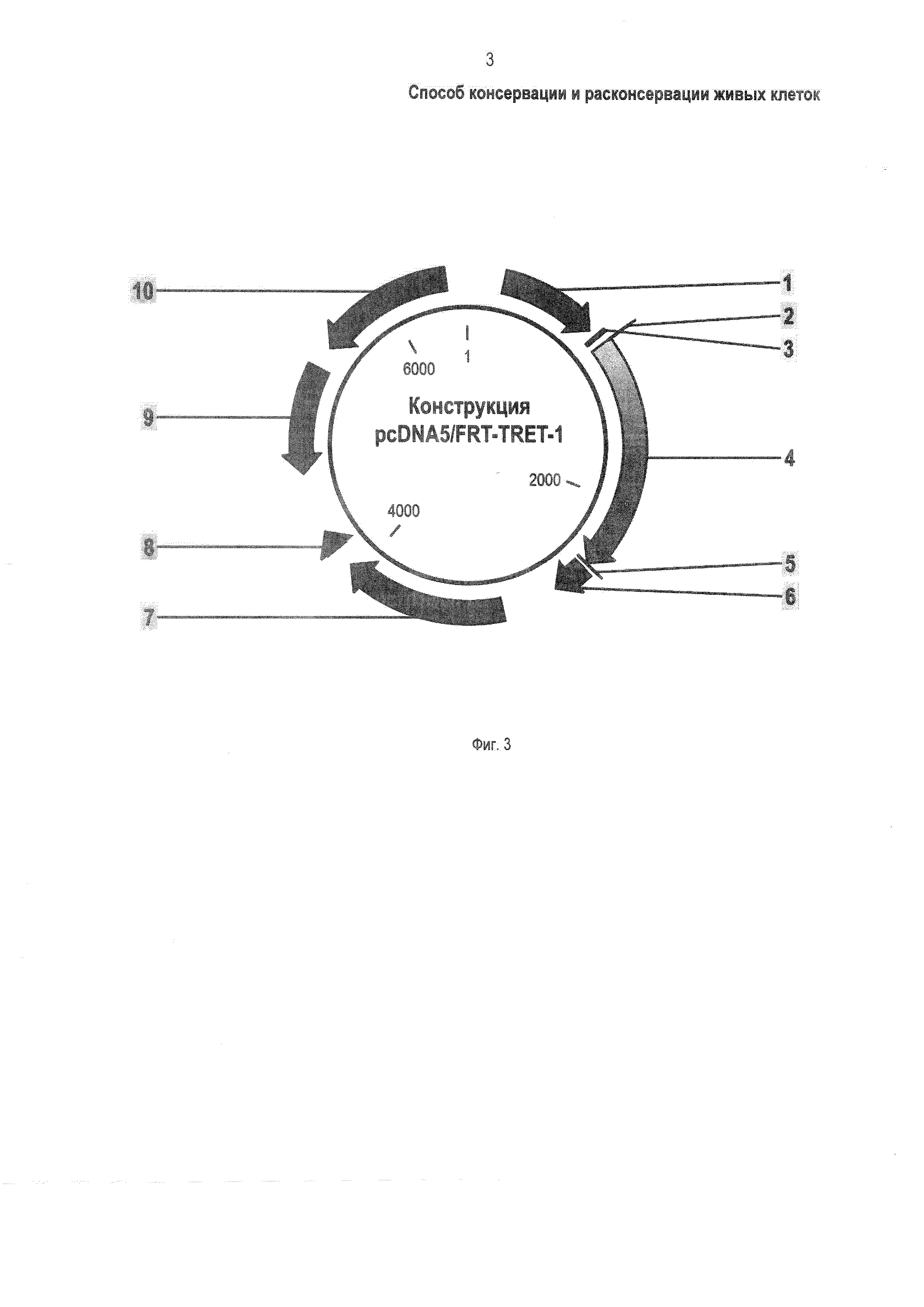
Фиг. 3 - Физическая карта генетической конструкции pcDNA5/FRT-TRET-1, где: 1 - промотор CMV; 2 - промотор Т7; 3 - распознаваемый ферментом BamHI участок; 4 - ген TRET-1; 5 - распознаваемый ферментом Xhol участок,; 6 - участок полиаденилирования; 7 - ген устойчивости к антибиотику гигромицин; 8 - участок полиаденилирования; 9 - точка начала репликации PUC; 10 - ген устойчивости к антибиотику ампициллин.
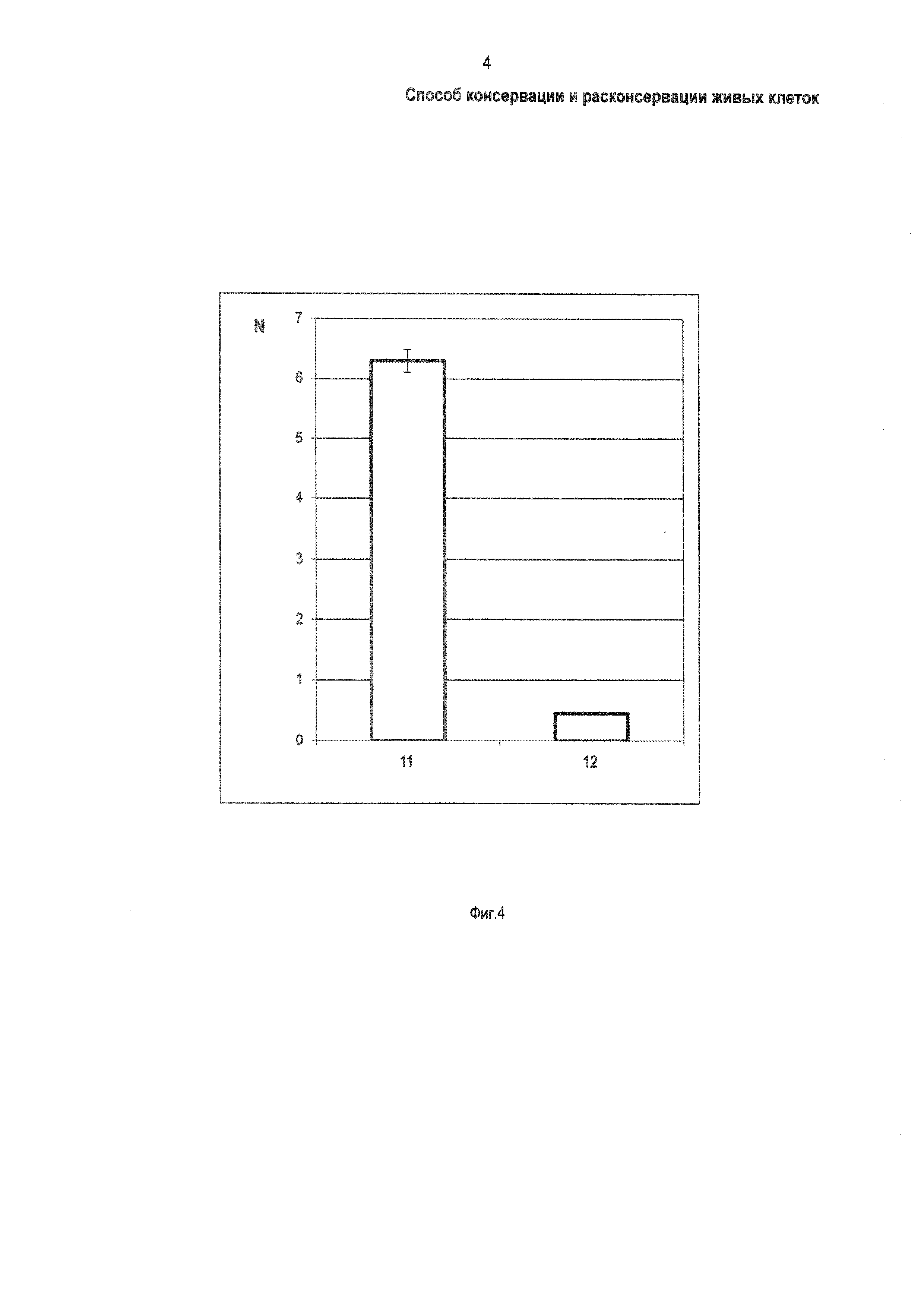
Фиг. 4 - Экспериментально полученная диаграмма содержания трегалозы в клетках СНО (Chinese hamster ovarian cells, клетки яичников китайского хомяка) после доставки трегалозы с использованием белка TRET-1 в соответствии с заявляемым способом, в сравнении с контролем, где: 11 - содержание трегалозы в содержащих белок TRET-1 клетках СНО; 12 - содержание трегалозы в клетках СНО без белка TRET-1; N - содержание трегалозы, 10-9 моль в расчете на 1×105 клеток.
Осуществление заявляемого технического решения показывает ПРИМЕР, демонстрирующий доставку трегалозы в живые клетки, например - клетки яичников китайского хомяка, за счет использования белка TRET-1 и последующее удаление трегалозы из клеток.
ПРИМЕР
Опытным путем выбрано средство доставки (транспорта) трегалозы. Белок TRET-1 выбран на основе экспериментальных данных, как обеспечивающий наиболее эффективный транспорт трегалозы. Белок TRET-1 осуществляет транспорт трегалозы в живые клетки или из живых клеток, действуя без затрат энергии, за счет разницы концентраций трегалозы по разные стороны клеточной мембраны. Ввиду способности белка TRET-1 к транспорту трегалозы в обе стороны (в живые клетки и из них) этот белок пригоден для удаления трегалозы из живых клеток после их (живых клеток) расконсервации.
Вводят в живые клетки белок TRET-1 с аминокислотной последовательностью по Фиг. 1, например, способом, описанным ниже. Получают экземпляры насекомого Polypedilum vanderplanki, например - путем культивирования в лабораторных условиях. Из P. vanderplanki выделяют общую ДНК, например - с использованием коммерческого набора Nucleospin tissue kit (производства фирмы Macherey Nagel, Германия).
Специфический фрагмент ДНК с геном, кодирующим белок TRET-1 (геном TRET-1) получают, например, в полимеразной цепной реакции (ПЦР) с использованием общей ДНК P. vanderplanki в качестве матрицы. Для ПЦР (здесь и далее в тексте) используют высокоточную полимеразу, например - полимеразу Q5 производства фирмы New England Biolabs, США, и праймеры (короткие фрагменты ДНК), например - с последовательностью по Фиг. 2 (I, II). Фрагмент ДНК с геном TRET-1 встраивают в генетическую конструкцию, обеспечивающую экспрессию генов в соответствующих живых клетках. Например, фрагмент ДНК с геном TRET-1 встраивают в генетическую конструкцию pcDNA5/FRT (производства фирмы Invitrogen, США) с использованием ферментов Bam HI, Xhol и Т4 лигазы (производства фирмы New England Biolabs, США) в стандартных условиях в соответствии с рекомендациями производителей. По завершении встраивания получают генетическую конструкцию pcDNA5/FRT-TRET-1, Фиг. 3. Полученную генетическую конструкцию с геном TRET-1, например - конструкцию pcDNA5/FRT-TRET-1 (производства фирмы Thermo Fisher scientific, США), вводят в живые клетки, например - культуру клеток китайского хомячка СНО (Chinese Hamster Ovarian, производства фирмы Invitrogen, США). Генетическую конструкцию вводят в клетки СНО (проводят трансфекцию клеток). Трансфекцию осуществляют, например, с использованием реагента FuGENE (производства фирмы Promega, США) в соответствии с рекомендациями производителя.
В контрольном варианте трансфекцию культуры клеток СНО осуществляют с использованием стандартной генетической конструкции pcDNA5/FRT - вместо используемой в заявляемом техническом решении конструкции pcDNA5/FRT-TRET-1. Конструкция pcDNA5/FRT содержит все элементы генетической конструкции pcDNA5/FRT-TRET-1, за исключением гена TRET-1.
После введения белка TRET-1 в живые клетки, при необходимости осуществляют проверку наличия белка в живых клетках, например - способом, описанным ниже. После проведенной в соответствии с рекомендациями (в настоящем примере - рекомендациями по применению реагента FUGENE) трансфекции клетки СНО культивируют в питательной среде RPMI в стандартных условиях в соответствии с рекомендациями фирмы-производителя клеток. Наличие экспрессии гена TRET-1 в клетках СНО проверяют через 48 часов после трансфекции. Для этого проводят окрашивание фиксированного препарата клеток СНО на предметном стекле первичными антителами, специфически связывающимися с белком TRET-1, например - производства фирмы Invitrogen (США) и вторичными антителами, содержащими краситель AlexaFluor (например, производства фирмы Invitrogen, США) в соответствии с рекомендациями производителей антител. Связывание первичных антител с клетками СНО, определяемое по флюоресцентному окрашиванию клеток СНО свидетельствует о наличии в них (клетках) белка TRET-1. Наличие флюоресценции подтверждает факт состоявшейся экспрессии белка TRET-1. В контрольном варианте (клеток СНО после трансфекции конструкцией pcDNA5/FRT) флюоресценция не наблюдается, что свидетельствует об отсутствии белка TRET-1 в контрольных клетках. То есть экспериментально доказано появление и наличие белка TRET-1 в клетках СНО после их обработки вышеописанным путем.
Далее осуществляют доставку трегалозы в живые клетки, содержащие белок TRET-1, путем добавления трегалозы в окружающую клетки среду, например - среду культивирования. В приводимом примере - осуществляют доставку трегалозы в клетки СНО. Определяют количество клеток СНО известным способом, например - с использованием камеры Горяева [16]. Отбирают по 1×105 клеток СНО после трансфекции генетическими конструкциями pcDNA5/FRT-TRET-1 и pcDNA5/FRT. Известным способом проводят замену стандартной питательной среды RPMI на среду RPMI с добавлением трегалозы в концентрации 0,1 моль/дм3. После замены среды клетки СНО культивируют в стандартных условиях в соответствии с рекомендациями фирмы-производителя (клеток) в течение 3 час. После культивирования клетки СНО трижды промывают стандартным буферным раствором DPBS (производства фирмы Gibco, США) при температуре 0°С. Проводят повторное определение количества клеток СНО известным способом, например - с использованием камеры Горяева. Известным способом определяют концентрацию трегалозы в клетках, например - методом высокоэффективной жидкостной хроматографии (далее по тексту - ВЭЖХ) в расчете на 1×105 клеток. На Фиг. 4 приведена диаграмма содержания трегалозы в клетках СНО при доставке трегалозы в соответствии с заявляемым способом (Фиг. 4, позиция 11) в сравнении с контролем (поз. 12). Клетки СНО с белком TRET-1 (после их трансфекции генетической конструкцией pcDNA5/FRT-TRET-1) содержат 6,3×10-9 моль трегалозы в расчете на 1×105 клеток. В контроле, клетки СНО без белка TRET-1 (после трансфекции конструкцией pcDNA5/FRT) содержат не более 0,45×10-9 моль трегалозы в расчете на 1×105 клеток. Так, экспрессирующие белок TRET-1 клетки СНО содержат в 14 раз (6,3×10-9/0,45×10-9=14) больше трегалозы, по сравнению с клетками СНО, не экспрессирующими TRET-1. 14-кратное увеличение содержания трегалозы в экспрессирующих белок TRET-1 клетках СНО подтверждает факт доставки трегалозы в живые клетки белком TRET-1. Таким путем завершают доставку трегалозы в живые клетки белком TRET-1 и подготовку клеток к консервации. Последующую (после подготовки живых клеток) консервацию выполняют избранным путем, например - с использованием способа криохранения. Способы консервации не ограничиваются криохранением и включают любые известные способы консервации, причем оптимальным применительно к живым клеткам является, например - высушивание или охлаждение или замораживание.
На Фиг. 3 основным элементом конструкции pcDNA5/FRT-TRET-1 является ген TRET-1 (Фиг. 3, поз. 4), обеспечивающий экспрессию (синтез) белка TRET-1 в живых клетках (в приведенном ПРИМЕРЕ - клетках СНО). Конструкция pcDNA5/FRT-TRET-1 (Фиг. 3) содержит также известные из существующего уровня техники вспомогательные, не являющиеся обязательными элементы: позиции 1,2- обеспечивающие синтез встроенного в конструкцию гена TRET-1 промоторы CMV и Т7; поз. 3, 5 - участки, распознаваемые ферментами BamHI и Xhol при встраивании гена TRET-1; поз. 6, 8 - участки полиаденилирования, необходимые для синтеза встроенного в конструкцию гена TRET-1; поз. 7, 10 - гены устойчивости к антибиотикам гигромицину и ампициллину, которые обеспечивают селекцию несущих генетическую конструкцию клеток известным способом [17].
По потребности, законсервированные клетки расконсервируют, удаляют из них трегалозу путем замены окружающей клетки среды (например, среды культивирования) на среду без трегалозы, и применяют клетки по назначению.
В приводимом примере трегалозу удаляют из клеток СНО. Для этого известным способом проводят замену питательной среды RPMI с добавлением трегалозы на стандартную среду RPMI без трегалозы. После замены питательной среды клетки культивируют (инкубируют) в стандартных условиях в соответствии с рекомендациями фирмы-производителя клеток в течение 3 ч. После завершения инкубации определяют концентрацию трегалозы в клетках СНО методом ВЭЖХ в соответствии с описанием выше. Через три часа инкубации в не содержащей трегалозу среде RPMI, клетки СНО не содержат детектируемых количеств трегалозы, то есть клетки СНО освобождаются (избавляются) от ранее введенной для консервации трегалозы.
Отсутствие трегалозы в ранее содержавших трегалозу клетках СНО подтверждает факт удаления трегалозы из клеток СНО за счет действия белка TRET-1.
Состоявшееся и экспериментально установленное повышение и последующее снижение концентрации трегалозы в экспрессирующих белок TRET-1 клетках СНО подтверждает достижение цели заявляемого изобретения - использование белка TRET-1 в соответствии с описанием предполагаемого изобретения обеспечивает доставку трегалозы в живые клетки и последующее удаление трегалозы по мере потребности.
В контрольном варианте экспериментов, выполняемых с использованием вышеприведенных реактивов, расходных материалов и оборудования, доставку трегалозы в культуру клеток СНО осуществляют с использованием прототипа [15]. Для этого в культуральную среду RPMI добавляют трегалозу в концентрации 0,1 моль/дм3 и ДМСО в концентрации 5% [15]. Клетки СНО инкубируют при температуре плюс 4°С в течение 4 мин в соответствии с приведенным описанием [15]. Охлаждают клетки СНО до 0°С, помещая их на лед, и незамедлительно определяют концентрацию трегалозы методом ВЭЖХ в соответствии с описанием выше. Затем обеспечивают условия для самопроизвольного выхода трегалозы из клеток СНО, заменяя среду RPMI с добавкой трегалозы и ДМСО на новую среду RPMI без добавок. Повторно инкубируют клетки в стандартных условиях в течение 3-х час аналогично заявляемому способу. После инкубации определяют концентрацию трегалозы в клетках СНО методом ВЭЖХ в соответствии с описанием выше.
Использование прототипа [15] позволяет достичь содержания трегалозы в клетках СНО на уровне 2,1×10-9 моль трегалозы в расчете на 1×105 клеток. После инкубации в питательной среде RPMI. без добавок в течение 3 ч, содержание трегалозы снижается до 1,4×10-9 моль трегалозы в расчете на 1×105 клеток. Таким образом, с использованием прототипа [15] не достижимо и невозможно обеспечиваемое заявляемым способом внутриклеточное содержание трегалозы 6,3×10-9 моль в расчете на 1×105 клеток, и последующее за 3 ч полное освобождение клеток СНО от трегалозы.
Таким образом, сравнение результатов осуществления заявляемого способа и контрольного варианта показывает и доказывает, что успешно обеспечиваемые заявляемым техническим решением доставка трегалозы в живые клетки и последующее удаление трегалозы за счет белка TRET-1, не осуществимы по прототипу [15].
Приведенный пример показывает осуществимость предполагаемого изобретения с использованием стандартного оборудования и материалов, полезность его применения в биотехнологиях, например - для консервации живых клеток. Применимость заявляемого технического решения не ограничивается культурой клеток китайского хомячка СНО, использованной в Примере для показа возможности осуществления заявляемого технического решения - живые клетки иных организмов также могут быть подготовлены к консервации заявляемым способом и законсервированы, в том числе - в составе тканей и органов. Заявляемое техническое решение относится к способу подготовки живых клеток к консервации путем доставки сахара трегалозы в консервируемые живые клетки и последующего удаления трегалозы с использованием белка TRET-1 насекомого Polypedilum vanderplanki. Способ предназначен для использования при консервации живых клеток, например - ооцитов, с целью их (живых клеток) хранения и транспортировки, с сохранением возможности последующего восстановления жизнедеятельности законсервированных клеток и их использования, например - для воспроизводства потомства.
В заявленном техническом решении заявителем реализована поставленная цель: осуществлена доставка трегалозы в живые клетки с использованием белка TRET-1; доказательством достижения заявленной цели является Фиг. 4, где продемонстрировано повышение содержания трегалозы в живых клетках СНО, обусловленное использованием белка TRET-1 за счет применения генетической конструкции. Именно доставка трегалозы в живые клетки за счет использования белка TRET-1 позволяет осуществить их (клеток) последующую консервацию и хранение. После завершения хранения последующее, по мере потребности, удаление трегалозы из живых клеток за счет использования белка TRET-1 и замены среды, окружающей клетки, позволяет избежать токсических эффектов на живые клетки. Применение заявляемого способа обеспечивает существенное расширение возможностей консервации, хранения и транспортировки живых клеток в весьма неблагоприятных для нормальной жизнедеятельности условиях, с сохранением жизненных функций живого организма, например, при длительном пребывании в космическом пространстве.
Заявляемое техническое решение удовлетворяет критерию «новизна», предъявляемому к изобретениям, так как при определении уровня техники не выявлены средства, которым присущи признаки, идентичные (то есть совпадающие по исполняемой ими функции и форме выполнения этих признаков) всем признакам, перечисленным в формуле предполагаемого изобретения, включая характеристику назначения.
Заявляемое техническое решение удовлетворяет критерию «изобретательский уровень», т.к. не является очевидным для специалиста в данной области техники и из исследованного уровня техники заявителем не выявлены технические решения, совпадающие по технической сущности с предлагаемым решением, и не установлена известность влияния отличительных признаков на полученный технический результат.
Заявляемое техническое решение можно реализовать для консервации клеток в медицине, ветеринарии, сельском хозяйстве и биотехнологии, в том числе - производстве биотехнологических продуктов посредством использования известных стандартных технических устройств, оборудования и материалов. Это соответствует критерию «промышленная применимость», предъявляемому к изобретениям.
ИСПОЛЬЗОВАННЫЕ ИСТОЧНИКИ
1. Kikawada Т. et al. Trehalose transporter 1, a facilitated and high-capacity trehalose transporter, allows exogenous trehalose uptake into cells. // Proc. Natl. Acad. Sci. U. S. A. 2007. Vol. 104, №28. P. 11585-11590.
2. Gusev O. et al. Comparative genome sequencing reveals genomic signature of extreme desiccation tolerance in the anhydrobiotic midge // Nat. Commun. Nature Publishing Group, 2014. Vol. 5. P. 4784.
3. Cornette R., Kikawada T. The induction of anhydrobiosis in the sleeping chironomid: current status of our knowledge // IUBMB Life. 2011. Vol. 63, №6. P. 419-429.
4. Watanabe M. et al. Mechanism allowing an insect to survive complete dehydration and extreme temperatures //J. Exp. Biol. 2002. Vol. 205. P. 2799-2802.
5. Crowe J.H. Trehalose as a "Chemical chaperone": fact and fantasy // Advances in experimental medicine and biology / ed. Csermely P., Vigh L. New York, NY: Springer New York, 2007. Vol. 594. P. 143-158.
6.  J. et al. Favorable effects of trehalose on the development of UVB-mediated antioxidant/pro-oxidant imbalance in the corneal epithelium, proinflammatory cytokine and matrix metalloproteinase induction, and heat shock protein 70 expression //
J. et al. Favorable effects of trehalose on the development of UVB-mediated antioxidant/pro-oxidant imbalance in the corneal epithelium, proinflammatory cytokine and matrix metalloproteinase induction, and heat shock protein 70 expression //  Arch. Clin. Exp. Ophthalmol. 2011. Vol. 249, №8. P. 1185-1194.
Arch. Clin. Exp. Ophthalmol. 2011. Vol. 249, №8. P. 1185-1194.
7. Guo N. et al. Trehalose expression confers desiccation tolerance on human cells // Nat. Biotechnol. Nature America Inc., 2000. Vol. 18, №2. P. 168-171.
8. Holovati J.L., Gyongyossy-lssa M.I.C., Acker J.P. Effects of trehalose-loaded liposomes on red blood cell response to freezing and post-thaw membrane quality. // Cryobiology. 2009. Vol. 58, №1. P. 75-83.
9. Eroglu A. et al. Intracellular trehalose improves the survival of cryopreserved mammalian cells. // Nat. Biotechnol. Nature America Inc., 2000. Vol. 18, №2. P. 163-167.
10. Oliver A.E. et al. Loading Human Mesenchymal Stem Cells with Trehalose by Fluid-Phase Endocytosis // Cell Preserv. Technol. Mary Ann Liebert, Inc., 2004. Vol. 2, №1. P. 35-49.
11. The Human Protein Atlas portal. Trehalase protein expression in human tissues and organs [Electronic resource]. 2016. URL: http://www.proteinatlas.org/ENSG00000118094-TREH/tissue(accessed: 04.10.2016).
12. Singer M.A., Lindquist S. Multiple effects of trehalose on protein folding in vitro and in vivo. // Mol. Cell. 1998. Vol. 1, №5. P. 639-648.
13. Gillian M. Beattie, John H. Crowe, Fern Tablin A.H. Cryopreservation of human adult and fetal pancreatic cells and human platelets: pat. US 5827741 A USA. USA, 1996. P. 16.
14. Acker J.P., Gyongyossy-lssa M. A method of cryopreserving cells and tissues by liposomal delivery of sugars to enhance post-thaw viability. Google Patents, 2005. P. 19.
15. Yun-Hee SHON, Jang-Soo SUH J.J.Y. A freezing medium composition for cryopreserving amniotic fluid-derived stem cells and a method for cryopreserving the same: pat. WO 2012091206 A1 USA. 2011. P. 9.
16. Stone L.R. et al. Accuracy and precision comparison of the hemocytometer and automated cell counting methods // FASEB J. 2009. Vol.23, №(Meeting abstracts). P. 827.
17. Sambrook J., W Russell D. Molecular Cloning: A Laboratory Manual // Cold Spring Harb. Lab. Press. Cold Spring Harb. NY. 2001. P. 999.
Способ консервации и расконсервации живых клеток, предусматривающий использование трегалозы и белка TRET-1, заключающийся в том, что в живые клетки вводят белок TRET-1 путем экспрессии содержащей ген TRET-1 насекомого Polypedilum vanderplanki генетической конструкции, после введения белка TRET-1 в живые клетки при необходимости осуществляют проверку наличия белка в живых клетках непосредственно, после чего осуществляют доставку трегалозы в указанные живые клетки путем добавления трегалозы в окружающую клетки среду, а при расконсервации осуществляют удаление трегалозы из живых клеток путем замены окружающей клетки среды с трегалозой на среду без трегалозы.

Штамм бактерий bacillus pumilus мк-10 с низкой протеолитической активностью и его применение
Штамм бактерий bacillus pumilus 2a-5 с низкой протеолитической активностью, повышенной активностью фосфатазы, способ его получения и применения
Генетическая конструкция для экспрессии генов в клетках насекомого polypedilum vanderplanki
Способ подготовки шламов
Способ обезвреживания углеводородсодержащих шламов

