Результат интеллектуальной деятельности: АНТИТЕЛА ПРОТИВ GDF8 ЧЕЛОВЕКА
Вид РИД
Изобретение
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, которые являются специфическими в отношении фактора 8 роста и дифференциации (GDF8).
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Фактор 8 роста и дифференциации (GDF8), также известный как миостатин, является членом суперсемейства факторов роста TGF-β. GDF8 является отрицательным регулятором массы скелетных мышц, высокой степени экспрессируемым в ходе развития, и скелетных мышц взрослых.
GDF8 является высоко консервативным для видов, и аминокислотные последовательности GDF8 мыши и человека являются идентичными (нуклеотидная последовательность и аминокислотная последовательность GDF8 человека представлены в SEQ ID NO:338-339) (McPherron et al. 1977 Nature 387:83-90).
Ряд заболеваний человека, например, мышечная дистрофия, мышечная атрофия, синдром истощения мышечной массы, саркопения и кахексия, сопровождаются уменьшением или повреждением мышечной ткани, и ингибиторы GDF8 могут использоваться для лечения этих заболеваний или нарушений.
Антитела против GDF8 и способы лечения описаны, например, в патентах США № 6096506, 7320789 и 7807159, в заявках WO 2007/047112 и WO 2005/094446, в заявке на патент США № 2007/0087000, в патенте США № 7261893 и в заявке WO 2010/070094.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам человека или гуманизированным антителам и антигенсвязывающим фрагментам антител человека или гуманизированных антител, которые специфически связываются с фактором 8 роста и дифференциации (GDF8). Эти антитела характеризуются связыванием с GDF8 с высокой аффинностью и способностью к нейтрализации активности GDF8. Антитела могут быть полноразмерными (например, антителом IgG1 или IgG4) или могут содержать только антигенсвязывающую часть (например, Fab-, F(ab')2- или scFv-фрагмент), и могут быть модифицированы для оказания воздействия на выполняемые функции, например, для исключения остаточных эффекторных функций (Reddy et al. (2000) J. Immunol. 164: 1925-1933).
В одном из вариантов осуществления антитело по настоящему изобретению содержит аминокислотную последовательность вариабельной области тяжелой цепи (HCVR), выбранную из группы, состоящей из SEQ ID NO:2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 360 и 376, или в значительной степени идентичную ей последовательность.
В одном из вариантов осуществления антитело по настоящему изобретению содержит аминокислотную последовательность вариабельной области легкой цепи (LCVR), выбранную из группы, состоящей из SEQ ID NO:10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 322, 368 и 384, или в значительной степени идентичную ей последовательность.
В одном из вариантов осуществления антитело по настоящему изобретению содержит аминокислотную последовательность HCVR и аминокислотную последовательность LCVR, причем последовательности пар HCVR/LCVR выбраны из группы, состоящей из SEQ ID NO:2/10, 18/26, 34/42, 50/58, 66/74, 82/90, 98/106, 114/122, 130/138, 146/154, 162/170, 178/186, 194/202, 210/218, 226/234, 242/250, 258/266, 274/282, 290/298, 306/314, 114/322, 360/368 и 376/384.
Отличительным признаком настоящего изобретения служит также антитело человека или гуманизированное антитело или антигенсвязывающий фрагмент антитела, содержащее(ий) аминокислотную последовательность определяющего комплементарность участка 3 тяжелой цепи (HCDR3) и аминокислотную последовательность CDR3 легкой цепи (LCDR3), причем аминокислотная последовательность HCDR3 выбрана из группы, состоящей из SEQ ID NO:8, 24, 40, 56, 72, 88, 104, 120, 136, 152, 168, 184, 200, 216, 232, 248, 264, 280, 296, 312, 366 и 382, или в значительной степени идентичную ей последовательность, а аминокислотная последовательность LCDR3 выбрана из группы, состоящей из SEQ ID NO:16; 32, 48, 64, 80, 96, 112, 128, 144, 160, 176, 192, 208, 224, 240, 256, 272, 288, 304, 320, 328, 374 и 390, или содержащее(ий) в значительной степени идентичную ей последовательность. В другом варианте осуществления антитело или его фрагмент содержит пару аминокислотных последовательностей HCDR3/LCDR3, выбранную из группы, состоящей из SEQ ID NO:8/16, 24/32, 40/48, 56/64, 72/80, 88/96, 104/112, 120/128, 136/144, 152/160, 168/176, 184/192, 200/208, 216/224, 232/240, 248/256, 264/272, 280/288, 296/304, 312/320, 120/328, 366/374 и 382/390.
В соответствующем варианте осуществления антитело или его фрагмент, кроме того, содержит аминокислотные последовательности CDR1 тяжелой цепи (HCDR1) и CDR2 тяжелой цепи (HCDR2) и аминокислотные последовательности CDR1 легкой цепи (LCDR1) и CDR2 легкой цепи (LCDR2), причем аминокислотная последовательность HCDR1 выбрана из группы, состоящей из SEQ ID NO:4, 20, 36, 52, 68, 84, 100, 116, 132, 148, 164, 180, 196, 212, 228, 244, 260, 276, 292, 308, 362 и 378, или в значительной степени идентичную ей последовательность; аминокислотная последовательность HCDR2 выбрана из группы, состоящей из SEQ ID NO:6, 22, 38, 54, 70, 86, 102, 118, 134, 150, 166, 182, 198, 214, 230, 246, 262, 278, 294, 310, 364 и 380, или в значительной степени идентичную ей последовательность; аминокислотная последовательность LCDR1 выбрана из группы, состоящей из SEQ ID NO:12, 28, 44, 60, 76, 92, 108, 124, 140, 156, 172, 188, 204, 220, 236, 252, 268, 284, 300, 316, 324, 370 и 386, или в значительной степени идентичную ей последовательность; а аминокислотная последовательность LCDR2 выбрана из группы, состоящей из SEQ ID NO:14, 30, 46, 62, 78, 94, 110, 126, 142, 158, 174, 190, 206, 222, 238, 254, 270, 286, 302, 318, 326, 372 и 388, или в значительной степени идентичную ей последовательность. В другом варианте осуществления HCDR1, HCDR2 и HCDR3 выбраны из группы, состоящей из SEQ ID NO:36/38/40, 116/118/120, 228/230/232, 362/364/366 и 378/380/382; а LCDR1, LCDR2 и LCDR3 выбраны из группы, состоящей из SEQ ID NO:44/46/48, 124/126/128, 236/238/240, 370/372/374 и 386/388/390. Тем не менее, в другом варианте осуществления CDR тяжелой и легкой цепей выбрана из группы, состоящей из SEQ ID NO:36/38/40/44/46/48 (например, 21-E5), 116/118/120/124/126/128 (например, 8D12), 228/230/232/236/238/240 (например, 1A2), 362/364/366/370/372/374 (например, H4H1657N2) и 378/380/382/386/388/390 (например, H4H1669P).
В соответствующем варианте осуществления настоящее изобретение содержит антитело или антигенсвязывающий фрагмент антитела, которое(ый) специфически связывается с GDF8, причем антитело или фрагмент содержит CDR-домены тяжелой и легкой цепей, содержащиеся внутри последовательностей вариабельных доменов тяжелой и легкой цепей, выбранных из группы, состоящей из SEQ ID NO:2/10, 18/26, 34/42, 50/58, 66/74, 82/90, 98/106, 114/122, 130/138, 146/154, 162/170, 178/186, 194/202, 210/218, 226/234, 242/250, 258/266, 274/282, 290/298, 306/314, 114/322, 360/368 и 376/384. Способы и методы идентификации CDR внутри аминокислотных последовательностей HCVR и LCVR хорошо известны в данной области и могут использоваться для идентификации CDR внутри указанных аминокислотных последовательностей HCVR и/или LCVR, раскрытых в настоящем описании. Приводимые в качестве примера правила, которые могут использоваться для определения границ CDR, содержат, например, определение по Kabat, определение по Chothia, и определение в случае AbM. Вообще говоря, определение по Kabat основывается на вариабельности последовательностей, определение по Chothia основывается на местонахождении структурных петлевых участков, а определение в случае AbM является компромиссом между подходами Kabat и Chothia. См., например, Kabat, "Sequences of Proteins of Immunological Interest," National Institutes of Health, Bethesda, Md. (1991); AI-Lazikani et al., J. Mol. Biol. 273: 927-948 (1997); и Martin et al., Proc. Natl. Acad. Sci. USA 86: 9268-9272 (1989). В распоряжении также имеются общедоступные базы данных для идентификации последовательностей CDR внутри антитела.
Настоящее изобретение также относится к молекулам нуклеиновых кислот, кодирующим антитела или антигенсвязывающие фрагменты по настоящему изобретению. Настоящее изобретение также относится к рекомбинантным экспрессионным векторам, несущим кодирующие антитела нуклеиновые кислоты по настоящему изобретению, и клетки-хозяева, в которые такие векторы были введены, равно как и способы получения антител по настоящему изобретению путем культивирования клеток-хозяев по настоящему изобретению.
В одном из вариантов осуществления антитело по настоящему изобретению содержит HCVR, кодируемую нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:1, 17, 33, 49, 65, 81, 97, 113, 129, 145, 161, 177, 193, 209, 225, 241, 257, 273, 289, 305, 359 и 375, или в значительной степени сходной с ней последовательностью, гомологичной ей по крайней мере на 95%.
В одном из вариантов осуществления антитело по настоящему изобретению содержит LCVR, кодируемую нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:9, 25, 41, 57, 73, 89, 105, 121, 137, 153, 169, 185, 201, 217, 233, 249, 265, 281, 297, 313, 321, 367 и 383, или в значительной степени сходной с ней последовательностью, гомологичной ей по крайней мере на 95%.
В одном из вариантов осуществления антитело по настоящему изобретению содержит аминокислотную последовательность HCVR и аминокислотную последовательность LCVR, причем последовательности пар HCV/LCVR кодируются парами молекул нуклеиновых кислот, выбранными из группы, состоящей из SEQ ID NO:1/9, 17/25, 33/41, 49/57, 65/73, 81/89, 97/105, 113/121, 129/137, 145/153, 161/169, 177/185, 193/201, 209/217, 225/233, 241/249, 257/265, 273/281, 289/297, 305/313, 113/321, 359/367 и 375/383.
Отличительным признаком настоящего изобретения служит также антитело человека или гуманизированное антитело или фрагмент антитела, содержащее(ий) HCDR3, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:7, 23, 39, 55, 71, 87, 103, 119, 135, 151, 167, 183, 199, 215, 231, 247, 263, 279, 295, 311, 365 и 381, или в значительной степени сходной с ней последовательностью, гомологичной ей на по крайней мере 95%, и LCDR3, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:15, 31, 47, 63, 79, 95, 111, 127, 143, 159, 175, 191, 207, 223, 239, 255, 271, 287, 303, 319, 327, 373 и 389, или в значительной степени сходной с ней последовательностью, гомологичной ей на по крайней мере 95%. В одном из вариантов осуществления совокупность HCDR3/LCDR3 кодируется парой нуклеотидных последовательностей, выбранной из группы, состоящей из SEQ ID NO:7/15, 23/31, 39/47, 55/63, 71/79, 87/95, 103/111, 119/127, 135/143, 151/159, 167/175, 183/191, 199/207, 215/223, 231/239, 247/255, 263/271, 279/287, 295/303, 311/319, 119/327, 365/373 и 381/389.
В соответствующем варианте осуществления антитело или фрагмент антитела, кроме того, содержит HCDR1 и HCDR2, и LCDR1 и LCDR2, причем HCDR1 кодируется нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:3, 19, 35, 51, 67, 83, 99, 115, 131, 147, 163, 179, 195, 211, 227, 243, 259, 275, 291, 307, 361 или 377, или в значительной степени сходной с ней последовательностью, гомологичной ей на по крайней мере 95%, HCDR2 кодируется нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:5, 21, 37, 53, 69, 85, 101, 117, 133, 149, 165, 181, 197, 213, 229, 245, 261, 277, 293, 309, 363 и 379, или в значительной степени сходной с ней последовательностью, гомологичной ей на по крайней мере 95%, LCDR1 кодируется нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:11, 27, 43, 59, 75, 91, 107, 123, 139, 155, 171, 187, 203, 219, 235, 251, 267, 283, 299, 315, 323, 369 и 385, или в значительной степени сходной с ней последовательностью, гомологичной ей по крайней мере на 95%, а LCDR2 кодируется нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:13, 29, 45, 61, 77, 93, 109, 125, 141, 157, 173, 189, 205, 221, 237, 253, 269, 285, 301, 317, 325, 371 и 387, или в значительной степени сходной с ней последовательностью, гомологичной ей на по крайней мере 95%. В одном из вариантов осуществления антитело или фрагмент антитела содержит CDR тяжелой и легкой цепей, кодируемые совокупностями последовательностей нуклеиновых кислот SEQ ID NO:35/37/39/43/45/47, 115/117/119/123/125/127, 227/229/231/235/237/239, 361/363/365/369/371/373 или 377/379/381/385/387/389.
Отличительным признаком настоящего изобретения служит также выделенное антитело или фрагмент антитела, которое(ый) специфически связывается с GDF8, содержащее(ий) CDR тяжелой и легкой цепи, выбранные из группы, состоящей из (a) HCDR1, содержащего аминокислотную последовательность формулы X1-X2-X3-X4-X5-X6-X7-X8 (SEQ ID NO:329), где X1 представляет собой Gly; X2 представляет собой Phe; X3 представляет собой Thr; X4 представляет собой Phe; X5 представляет собой Ser; X6 представляет собой Ala или Ser; X7 представляет собой Phe или Tyr; X8 представляет собой Gly или Ala; (b) HCDR2, содержащего аминокислотную последовательность формулы X1-X2-X3-X4-X5-X6-X7-X8 (SEQ ID NO:330), где X1 представляет собой Ile; X2 представляет собой Gly или Ser; X3 представляет собой Tyr или Gly; X4 представляет собой Ser или Asp; X5 представляет собой Gly; X6 представляет собой Gly; X7 представляет собой Ser или Asn; а X8 представляет собой Ala или Glu; (c) HCDR3, содержащего аминокислотную последовательность формулы X1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12-X13-X14 (SEQ ID NO:331), где X1 представляет собой Ser или Ala; X2 представляет собой Thr или Lys; X3 представляет собой Asp или Ile; X4 представляет собой Gly или Ser; X5 представляет собой Ala или His; X6 представляет собой Trp или Tyr; X7 представляет собой Lys или Asp; X8 представляет собой Met или Ile; X9 представляет собой Ser или Leu; X10 представляет собой Gly или Ser; X11 представляет собой Leu или Gly; X12 представляет собой Asp или Met; X13 представляет собой Val или Asp; X14 представляет собой Val или отсутствует; и (d) LCDR1, содержащего аминокислотную последовательность формулы X1-X2-X3-X4-X5-X6 (SEQ ID NO:332), где X1 представляет собой Gln; X2 представляет собой Asp или Gly; X3 представляет собой Ile; X4 представляет собой Ser; X5 представляет собой Asp или Asn; а X6 представляет собой Tyr или Trp; (e) LCDR2, содержащего аминокислотную последовательность формулы X1-X2-X3 (SEQ ID NO:333), где X1 представляет собой Thr или Ala; X2 представляет собой Thr или Ala; а X3 представляет собой Ser; и (f) LCDR3-участка, содержащего аминокислотную последовательность формулы X1-X2-X3-X4-X5-X6-X7-X8-X9 (SEQ ID NO:334), где X1 представляет собой Gln; X2 представляет собой Lys или Gln; X3 представляет собой Ala или Tyr; X4 представляет собой Asp или Asn; X5 представляет собой Ser; X6 представляет собой Ala или Phe; X7 представляет собой Pro; X8 представляет собой Leu; и X9 представляет собой Thr.
Методика определения вышеуказанных консенсусных последовательностей (SEQ ID NO:329-334) показана на фиг. 4A и 4B.
Отличительным признаком настоящего изобретения служит также полностью антитело человека или гуманизированное антитело или фрагмент антитела, которое связывается с CDF8 с аффинностью (выражаемой в виде константы диссоциации, «KD»), составляющей приблизительно 1 нМ или менее, как определено с помощью анализа с использованием поверхностного плазмонного резонанса (например, BIACORETM). В некоторых вариантах осуществления антитело по настоящему изобретению демонстрирует KD, составляющую приблизительно 700 пМ или менее; приблизительно 500 пМ или менее; приблизительно 320 пМ или менее; приблизительно 160 пМ или менее; приблизительно 100 пМ или менее; приблизительно 50 пМ или менее; приблизительно 10 пМ или менее, или приблизительно 5 пМ или менее.
В одном из вариантов осуществления настоящее изобретение относится к полностью антителу человека или гуманизированному моноклональному антителу (mAb), которое специфически связывается с GDF8 человека и ингибирует его, и демонстрирует IC50, составляющую менее или равную приблизительно 10 нМ; приблизительно 5 нМ или менее; приблизительно 3 нМ или менее; приблизительно 2 нМ или менее; приблизительно 1 нМ или менее; приблизительно 500 пМ или менее, или приблизительно 200 пМ, или менее, как определено с помощью исследования индуцируемой GDF8 люциферазной активности. Как показано в экспериментальной части ниже, некоторые из антител против GDF8 по настоящему изобретению блокируют активность близкородственных белков, таких как GDF11, с намного большей IC50, чем GDF8 в биологическом исследовании люциферазной активности. В одном из вариантов осуществления настоящее изобретение относится к антителу или антигенсвязывающему фрагменту антитела, которое демонстрирует IC50 в случае блокирования активности GDF11, превышающую в по крайней мере приблизительно 10 раз, по крайней мере приблизительно 50 раз, по крайней мере приблизительно 100 раз, по крайней мере приблизительно 200 раз, по крайней мере приблизительно 500 раз, по крайней мере приблизительно 1000 раз или по крайней мере приблизительно 1500 раз таковую в случае блокирования активности GDF8.
Настоящее изобретение относится к антителу против GDF8, характеризующемуся измененным профилем гликозилирования. В вариантах использования может быть эффективной модификация для удаления нежелательных сайтов гликозилирования, или антитело, в котором отсутствует составляющая в виде фукозы, присутствующая в олигосахаридной цепи, например, для усиления функции антителозависимой клеточноопосредованной цитотоксичности (ADCC) (см. Shield et al. (2002) JBC 277: 26733). В случае других вариантов использования можно изменить галактозилирование для изменения комплементзависимой цитотоксичности (CDC).
Настоящее изобретение относится к антителам против GDF8, которые связываются со специфическими эпитопами GDF8 и способны к блокировке биологической активности GDF8. В первом варианте осуществления антитело по настоящему изобретению связывается с эпитопом зрелого белка GDF8 (SEQ ID NO:340) в пределах аминокислот с приблизительно 1 до приблизительно 109; с приблизительно 1 до приблизительно 54; с приблизительно 1 до приблизительно 44; с приблизительно 1 до приблизительно 34; с приблизительно 1 до приблизительно 24 и с приблизительно 1 до приблизительно 14. Во втором варианте осуществления антитело по настоящему изобретению связывается с одним или более эпитопов зрелого белка GDF8 (SEQ ID NO:340) в пределах аминокислот с приблизительно 35 до приблизительно 109; с приблизительно 45 до приблизительно 109; с приблизительно 55 до приблизительно 109; с приблизительно 65 до приблизительно 109; с приблизительно 75 до приблизительно 109; с приблизительно 85 до приблизительно 109; с приблизительно 92 до приблизительно 109 или с приблизительно 95 до приблизительно 109. В третьем варианте осуществления антитело или антигенсвязывающий фрагмент антитела связывается в эпитопе зрелого белка GDF8 человека с приблизительно аминокислотного остатка 48 до приблизительно 72; с приблизительно 48 до приблизительно 69; с приблизительно 48 до приблизительно 65; с приблизительно 52 до приблизительно 72; с приблизительно 52 до приблизительно 65 или с приблизительно 56 до приблизительно 65.
В соответствующем варианте осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое(ый) конкурирует за специфическое связывание с GDF8 с другим антителом, содержащим комбинацию аминокислотных последовательностей HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3 - SEQ ID NO:36/38/40/44/46/48, 116/118/120/124/126/128, 228/230/232/236/238/240, 362/364/366/370/372/374 или 378/380/382/386/388/390. В одном из вариантов осуществления антитело или антигенсвязывающий фрагмент по настоящему изобретению конкурирует за специфическое связывание с GDF8 с другим антителом, содержащим пару аминокислотных последовательностей HCVR/LCVR - SEQ ID NO:2/10, 18/26, 34/42, 50/58, 66/74, 82/90, 98/106, 114/122, 130/138, 146/154, 162/170, 178/186, 94/202, 210/218, 226/234, 242/250, 258/266, 274/282, 290/298, 306/314, 114/322, 360/368 или 376/384. Тем не менее, в другом связанном с этим варианте осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое(ый) распознает эпитоп GDF8, который распознается другим антителом, содержащим комбинацию аминокислотных последовательностей HCDR/LCDR - SEQ ID NO:36/38/40/44/46/48, 116/118/120/124/126/128, 228/230/232/236/238/240, 362/364/366/370/372/374 или 378/380/382/386/388/390. В одном из вариантов осуществления антитело или антигенсвязывающий фрагмент по настоящему изобретению распознает эпитоп GDF8, который распознается другим антителом, содержащим пару аминокислотных последовательностей HCVR/LCVR - SEQ ID NO:2/10, 18/26, 34/42, 50/58, 66/74, 82/90, 98/106, 114/122, 130/138, 146/154, 162/170, 178/186, 194/202, 210/218, 226/234, 242/250, 258/266, 274/282, 290/298, 306/314, 114/322, 360/368 или 376/384.
Настоящее изобретение также относится к композиции, содержащей рекомбинантное антитело человека или гуманизированное антитело против GDF8 человека и приемлемый носитель. Настоящее изобретение, кроме того, относится к векторам и клеткам-хозяина, содержащим векторы, которые содержат молекулы нуклеиновых кислот, кодирующие антитело человека против GDF8 по настоящему изобретению, а также к способам получения этих новых антител, включающим выращивание клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело против GDF8 по настоящему изобретению или фрагмент антитела, в условиях, дающих возможность получить белок и выделить полученный таким образом белок.
Настоящее изобретение также относится к способам ингибирования активности GDF8 с использованием антитела или его антигенсвязывающей части по настоящему изобретению. В одном из вариантов осуществления способ включает введение антитела или фрагмента антитела по настоящему изобретению человеку, страдающему нарушением, которое ослабляется в результате ингибирования активности GDF8. В предпочтительных вариантах осуществления человек, получающий лечение антителом или фрагментом антитела по настоящему изобретению, нуждается в улучшение гомеостаза глюкозы, уменьшении жировой массы, увеличении чувствительности к инсулину, улучшению функционирования почек и/или уменьшении жирового отложения. Антитело или фрагмент антитела по настоящему изобретению можно использовать для лечения, профилактики или подавления заболевания или состояния, характеризующегося уменьшением костной массы, содержащего остеопороз, остеопению, остеоартрит и переломы кости, лечения метаболического синдрома, противодействия истощению мышечной массы в результате длительного введения глюкокортикоида или стероидного гормона, или уменьшению мышечной массы, связанному с мышечной дистрофией, мышечной атрофией, синдромом истощения мышечной массы, саркопенией и кахексией.
Другие варианты и преимущества станут очевидными в результате ознакомления со следующим подробным описанием.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1. Иммуноблот ограниченного протеолиза GDF8 человека с помощью протеиназы K. Гели представляли собой 18% SDS-ПААГ для электрофореза в невосстанавливающих условиях с загрузкой 0,2 мкг GDF8 на каждую дорожку и использованием 2 мкг/мл любого из антител - контроля I (А), 1А2 (В) или 21-Е9 (С). Дорожка 1: время расщепления - 10 мин, 1 мкг GDF8, 0 мкг протеиназы К; Дорожка 2: время расщепления - 10 мин, 1 мкг GDF8, 1 мкг протеиназы К; Дорожка 3: время расщепления - 10 мин, 1 мкг GDF8, 6 мкг протеиназы К; Дорожка 4: время расщепления - 45 мин, 1 мкг GDF8, 0 мкг протеиназы К; Дорожка 5: время расщепления - 4 5 мин, 1 мкг GDF8, 1 мкг протеиназы К; Дорожка 6: время расщепления -4 5 мин, 1 мкг GDF8, 6 мкг протеиназы К.
Фиг. 2. Иммуноблот ограниченного протеолиза GDF8 человека с использованием больших доз протеиназы К. Гели представляли собой 18% SDS-ПААГ для электрофореза в невосстанавливающих условиях с загрузкой 0 или 4 мкг GDF8 на каждую дорожку и использованием 2 мкг/мл любого из антител - контроля I (А) или 1А2 (В). Дорожка 1: время расщепления - 16 ч, 0 мкг GDF8, 9 6 мкг протеиназы К; Дорожка 2: время расщепления - 16 ч, 4 мкг GDF8, 0 мкг протеиназы К; Дорожка 3: время расщепления - 16 ч, 4 мкг GDF8, 24 мкг протеиназы К; Дорожка 4: время расщепления - 16 ч, 4 мкг GDF8, 96 мкг протеиназы К; Дорожка 5: время расщепления - 1 ч, 4 мкг GDF8, 2 4 мкг протеиназы К; Дорожка 6: время расщепления - 1 ч, 4 мкг GDF8, 9 6 мкг протеиназы К; Дорожка 7: время расщепления 10 мин, 4 мкг GDF8, 0 мкг протеиназы К; Дорожка 8: время расщепления - 10 мин, 4 мкг GDF8, 24 мкг протеиназы К.
Фиг. 3А и 3В. Графики, демонстрирующие процент от исходных уровней глюкозы в динамике по времени у мышей, подвергнутых проверке на чувствительность к инсулину до (фиг .3А) и после (фиг. 3В) лечения антителом.
Фиг. 4А и 4В. Совмещение аминокислотных последовательностей
CDR тяжелой цепи (фиг. 4A) и CDR легкой цепи (фиг. 4B) приводимых в качестве примера антител против GDF8 H4H1657N2 и H4H1669P, демонстрирующее консенсусные последовательности, общие для этих последовательностей.
ПОДРОБНОЕ ОПИСАНИЕ ПО НАСТОЯЩЕМУ ИЗОБРЕТЕНИЮ
Перед описанием способов по настоящему изобретению следует отметить, что настоящее изобретение не ограничивается описываемыми конкретными способами и экспериментальными условиями, и такие способы и условия могут изменяться. Также следует понимать, что используемая в настоящем описании терминология служит в целях описания конкретных вариантов осуществления и не предназначена для ограничения, поскольку объем настоящего изобретения ограничен лишь прилагаемой формулой изобретения.
Используемые в этом описании изобретения и прилагаемой формуле изобретения формы единственного числа включают указания на множественное число, если только контекст явно не подразумевает иное. Таким образом, например, указание на “способ” включает один или более способов, и/или операций описываемого в настоящем описании типа, и/или которые станут очевидны квалифицированным в данной области специалистам после прочтения настоящего описания изобретения.
Если не указано иного, все технические и научные термины, используемые в настоящем описании, имеют значение, одинаковое с тем, в котором они обычно подразумеваются средним специалистом в области, к которой относится настоящее изобретение. Используемый термин «приблизительно», при использовании в отношении конкретного изложенного числового значения, означает, что это значение может отличаться от изложенного значения на более чем 1%. Например, используемое в настоящем описании выражение «приблизительно 100» включает 99 и 101, и все значения между ними (например, 99,1, 99,2, 99,3, 99,4 и т.д.).
Хотя любые способы и материалы, аналогичные с описываемыми в настоящем описании способами или материалами или эквивалентные им, могут использоваться при осуществлении на практике или осуществлении настоящего изобретения, ниже описаны предпочтительные способы и материалы.
Определения
«Фактор-8 роста и дифференциации человека», «GDF8» и «миостатин» используются взаимозаменяемо для обозначения белка, кодируемого последовательностью нуклеиновой кислоты SEQ ID NO:338, и белка с аминокислотной последовательностью SEQ ID NO:339 (пропептид) и 340 (зрелый белок).
Используемый в настоящем описании термин «антитело», как предполагается, относится к молекулам иммуноглобулинов, содержащим четыре полипептидных цепи, две тяжелые (H) цепи и две легкие (L) цепи, связанные с помощью дисульфидных связей, а также их мультимерам (например, IgM). Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (в настоящем описании сокращенно HCVR или VH) и константную область тяжелой цепи. Константная область тяжелой цепи содержит три домена, CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельную область легкой цепи (в настоящем описании сокращенно LCVR или VL) и константную область легкой цепи. Константная область легкой цепи содержит один домен (CL1). VH- и VL-области можно, кроме того, подразделить на участки гипервариабельности, называемые определяющими комплементарность участками (CDR), чередующиеся с областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, с расположением от амино-конца к карбоксильному концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В различных вариантах осуществления настоящего изобретения FR антитела против GDF8 (или его антигенсвязывающей части) могут быть идентичны последовательностям зародышевой линии человека, или они могут природно или искусственно модифицированными. Аминокислотную консенсусную последовательность можно определить на основе одновременного анализа двух или более CDR.
Используемый в настоящем описании термин «антитело» также содержит антигенсвязывающие фрагменты полноразмерных молекул антитела. Используемые в настоящем описании термины «антигенсвязывающая часть» антитела, «антигенсвязывающий фрагмент» антитела и т.п. содержат встречающийся в природе, ферментативно получаемый, синтетический или полученный с помощью методов генетической инженерии полипептид или гликопротеин, который специфически связывается с антигеном с образованием комплекса. Антигенсвязывающие фрагменты антитела можно получить, например, из полноразмерных молекул антител, используя любые подходящие стандартные способы, такие как протеолитическое расщепление или способы генетической инженерии для создания рекомбинантных молекул, содержащие манипулирование ДНК, кодирующей вариабельные и необязательно константные домены, и ее экспрессию. Такая ДНК известна и/или легкодоступна, например, из коммерческих источников, библиотек ДНК (содержащих, например, библиотеки антител в фагах), или ее можно синтезировать. ДНК можно подвергнуть секвенированию и химическому манипулированию или манипулированию с использованием способов молекулярной биологии, например, с целью приведения одного или более вариабельных и/или константных доменов в подходящую конфигурацию или с целью введения кодонов, создания остатков цистеина, модифицирования, добавления или делетирования аминокислот и т.д.
Неограничивающие примеры антигенсвязывающих фрагментов включают: (i) Fab-фрагменты; (ii) F(ab')2-фрагменты; (iii) Fd- фрагменты; (iv) Fv-фрагменты; (v) одноцепочечные Fv (scFv) молекулы; (vi) dAb-фрагменты и (vii) минимальные рекогниционные единицы, состоящие из аминокислотных остатков, которые имитируют гипервариабельный участок антитела (например, выделенный определяющий комплементарность участок (CDR)). В используемое в настоящем описании выражение «антигенсвязывающий фрагмент» также включены другие сконструированные молекулы, такие как диатела, триатела, тетратела и минитела.
Антигенсвязывающий фрагмент антитела, как правило, содержит по крайней мере один вариабельный домен. Вариабельный домен может быть любого размера или с любым составом аминокислот и будет, как правило, содержать по крайней мере один CDR, который расположен рядом или в рамке считывания с одной или более последовательностей каркасных областей. В антигенсвязывающих фрагментах, содержащих VH-домен, связанный с VL-доменом, VH- и VL-домены могут располагаться относительно друг друга в любом подходящем порядке. Например, вариабельная область может быть димерной и содержать димеры VH-VH, VH-VL или VL-VL. Альтернативно, антигенсвязывающий фрагмент антитела может содержать мономерный VH- или VL-домен.
В некоторых вариантах осуществления антигенсвязывающий фрагмент антитела может содержать по крайней мере один вариабельный домен, ковалентно связанный с по крайней мере одним константным доменом. Неограничивающие, приводимые в качестве примера конфигурации вариабельных и константных доменов, которые можно обнаружить в антигенсвязывающем фрагменте антитела по настоящему изобретению, включают: (i) VH-CH1; (ii) VH-CH2; (iii) VH-CH3; (iv) VH-CH1-CH2; (v) VH-CH1-CH2-CH3; (vi) VH-CH2-CH3; (vii) VH-CL; (viii) VL-CH1; (ix) VL-CH2; (x) VL-CH3; (xi) VL-CH1-CH2; (xii) VL-CH1-CH2-CH3; (xiii) VL-CH2-CH3 и (xiv) VL-CL. В случае любой конфигурации вариабельных и константных доменов, в том числе любой из приводимых в качестве примера конфигураций, перечисленных выше, вариабельные и константные домены могут быть связаны друг с другом либо непосредственно, либо с помощью полноразмерной или частичной шарнирной или линкерной области. Шарнирная область может состоять из по крайней мере 2 (например, 5, 10, 15, 20, 40, 60 или более) аминокислот, которые приводят к гибкому или полужесткому соединению между соседними вариабельными и/или константными доменами в одной полипептидной молекуле. Сверх того, антигенсвязывающий фрагмент антитела по настоящему изобретению может включать гомодимер или гетеродимер (или другой мультимер) любой из перечисленных выше конфигураций вариабельных и константных доменов в нековалентном соединении с одним другим мономерным VH- или VL-доменом и/или с одним или более мономерных VH- или VL-доменов (например, с помощью дисульфидной связи(ей)).
Как и в случае полноразмерных молекул антител, антигенсвязывающие фрагменты могут быть моноспецифическими или полиспецифическими (например, биспецифическими). Полиспецифический антигенсвязывающий фрагмент антитела, как правило, содержит по крайней мере два различных вариабельных домена, причем каждый вариабельный домен способен к специфическому связыванию с отдельным антигеном или с отличным эпитопом одного и того же антигена. Любой формат полиспецифического антитела, в том числе приводимые в качестве примера форматы биспецифического антитела, описываемые в настоящем описании, можно приспособить для применения в ситуации антигенсвязывающего фрагмента антитела по настоящему изобретению, используя стандартные способы, имеющиеся в данной области.
Антитела по настоящему изобретению могут функционировать в ходе комплементзависимой цитотоксичности (CDC) или антителозависимой клеточноопосредованной цитотоксичности (ADCC). «Комплементзависимая цитотоксичность» (CDC) относится к лизису экспрессирующих антиген клеток под действием антитела по настоящему изобретению в присутствии комплемента. «Антителозависимая клеточноопосредованная цитотоксичность» (ADCC) относится к клеточноопосредованной реакции, в ходе которой неспецифические цитотоксические клетки, экспрессирующие рецепторы Fc (FcR), (например, являющиеся природными киллерами (NK) клетки, нейтрофилы и макрофаги) распознают связавшееся антитело на поверхности клетки-мишени и в связи с этим приводят к лизису клетки-мишени. CDC и ADCC можно определить, используя исследования, которые хорошо известны и имеются в данной области. (См., например, патенты США № 5500362 и 5821337 и Clynes et al., Proc. Natl. Acad. Sci. (USA) 95: 652-656 (1998)).
Термин «специфически связывается», или тому подобный, означает, что антитело или его антигенсвязывающий фрагмент образует комплекс с антигеном, который является относительно стабильным в физиологических условиях. Специфическое связывание может характеризоваться константой диссоциации, составляющей 1×10-6 M или менее. Способы определения того, связываются ли две молекулы специфически, хорошо известны в данной области и содержат, например, равновесный диализ, поверхностный плазмонный резонанс и т.п. Например, антитело, которое «специфически связывается» с GDF8 человека, как используется в рамках по настоящему изобретению, содержит антитела, которые связываются с GDF8 человека или его частью (например, пептидом, содержащим по крайней мере 6 следующих друг за другом аминокислот SEQ ID NO: 340), с KD, составляющей менее чем приблизительно 1000 нМ, менее чем приблизительно 500 нМ, менее чем приблизительно 300 нМ, менее чем приблизительно 200 нМ, менее чем приблизительно 100 нМ, менее чем приблизительно 90 нМ, менее чем приблизительно 80 нМ, менее чем приблизительно 70 нМ, менее чем приблизительно 60 нМ, менее чем приблизительно 50 нМ, менее чем приблизительно 40 нМ, менее чем приблизительно 30 нМ, менее чем приблизительно 20 нМ, менее чем приблизительно 10 нМ, менее чем приблизительно 5 нМ, менее чем приблизительно 4 нМ, менее чем приблизительно 3 нМ, менее чем приблизительно 2 нМ, менее чем приблизительно 1 нМ или менее чем приблизительно 0,5 нМ, как определено в исследовании с использованием поверхностного плазмонного резонанса. (См., например, пример 3, в настоящем описании). Выделенное антитело, которое специфически связывается с GDF8 человека, может, однако, обладать перекрестной реактивностью с другими антигенами, такими как молекулы GDF8 других видов.
Термин антитело с «высокой аффинностью» относится к таким антителам, которые способны к связыванию с GDF8 с константной диссоциации (KD), составляющей приблизительно 10-8 M или менее, приблизительно 10-9 M или менее, приблизительно 10-10 M или менее, приблизительно 10-11 M или менее или приблизительно 10-12 M или менее, как определено с помощью поверхностного плазмонного резонанса, например, BIACORETM, или ELISA для определения аффинности в растворе.
Под термином «медленная скорость диссоциации» или «Koff» подразумевается, что антитело подвергается диссоциации от GDF8 с константой скорости, составляющей 1×10-3 сек-1 или менее, предпочтительно, 1×10-4 сек-1 или менее, как определено с помощью поверхностного плазмонного резонанса, например, BIACORETM.
«Нейтрализующее» или «блокирующее» антитело, как предполагается, относится к антителу, связывание которого с GDF8 приводит к ингибированию биологической активности GDF8. Это ингибирование биологической активности GDF8 можно оценить посредством определения одного или более индикаторов биологической активности GDF8. Оценку этих индикаторов биологической активности GDF8 можно осуществить с помощью одного или более из нескольких стандартных in vitro или in vivo исследований, известных в данной области (смотрите примеры ниже).
Полностью антитела человека против GDF8, описываемые в настоящем описании, могут содержать одну или более замен, вставок и/или делеций аминокислот в каркасных областях и/или CDR-участках вариабельных доменов тяжелой и легкой цепей по сравнению с соответствующими последовательностями зародышевой линии. Такие мутации можно легко определить посредством сравнения аминокислотных последовательностей, раскрытых в настоящем описании, с последовательностями зародышевой линии, доступными, например, из общедоступных баз данных, касающихся последовательностей антител. Настоящее изобретение включает антитела, и их антигенсвязывающие фрагменты, которые получают, исходя из любой из описанных в настоящем описании аминокислотных последовательностей, причем одну или более аминокислот в одной или более каркасных областей и/или CDR-участков подвергают обратной мутации к соответствующему остатку(ам) последовательности зародышевой линии или к консервативной аминокислотной замене (природной или неприродной) соответствующего остатка(ов) последовательности зародышевой линии (такие изменения последовательности называют в настоящем описании «обратными мутациями к последовательности зародышевой линии»). Средний специалист в данной области, исходя из последовательностей вариабельных областей тяжелой и легкой цепей, раскрытых в настоящем описании, может легко создать многочисленные антитела и антигенсвязывающие фрагменты, которые содержат одну или более отдельных обратных мутаций к последовательности зародышевой линии или их комбинации. В некоторых вариантах осуществления все остатки каркасных областей и/или CDR в VH и/или VL-доменах подвергают обратным мутациям к последовательности зародышевой линии. В других вариантах осуществления лишь некоторые остатки подвергают обратным мутациям к последовательности зародышевой линии, например, лишь мутированные остатки, обнаруживаемые в пределах первых 8 аминокислот FR1 или в пределах последних 8 аминокислот FR4, или лишь мутированные остатки, обнаруживаемые внутри CDR1, CDR2 или CDR3. Кроме того, антитела по настоящему изобретению могут содержать любую комбинацию из двух или более обратных мутаций к последовательности зародышевой линии внутри каркасных областей и/или CDR-участков, т.е., в которых некоторые отдельные остатки подвергнуты обратным мутациям к последовательности зародышевой линии, в то время как некоторые другие остатки, которые отличаются от последовательности зародышевой линии, сохранены. После получения антитела и антигенсвязывающие фрагменты, которые содержат одну или более обратных мутаций к последовательности зародышевой линии, можно легко проверить на предмет наличия одного или более желаемых свойств, таких как увеличенная специфичность связывания, увеличенная аффинность связывания, улучшенные или усиленные антагонистические или агонистические биологические свойства (исходя из конкретной ситуации), уменьшенная иммуногенность и т.д. Антитела и антигенсвязывающие фрагменты, полученные этим общим способом, включены в настоящее изобретение.
Настоящее изобретение также относится к антителам против GDF8, содержащим варианты любой из раскрытых в настоящем описании аминокислотных последовательностей HCVR, LCVR и/или CDR, содержащих одну или более консервативных замен. Например, настоящее изобретение относится к антителам против GDF8, которые содержат аминокислотные последовательности HCVR, LCVR и/или CDR, например, с 10 или менее, 8 или менее, 6 или менее, 4 или менее и т.д. консервативными аминокислотными заменами относительно любой из раскрытых в настоящем описании аминокислотных последовательностей HCVR, LCVR и/или CDR. В одном из вариантов осуществления антитело содержит HCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:360 и 376, с 8 или менее консервативных аминокислотных замен. В другом варианте осуществления антитело содержит HCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:360 и 376, с 6 или менее консервативных аминокислотных замен. В другом варианте осуществления антитело содержит HCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:360 и 376, с 4 или менее консервативных аминокислотных замен. В другом варианте осуществления антитело содержит HCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:360 и 376, с 2 или менее консервативных аминокислотных замен. В одном из вариантов осуществления антитело содержит LCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:368 и 384, с 8 или менее консервативных аминокислотных замен. В другом варианте осуществления антитело содержит LCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:368 и 384, с 6 или менее консервативных аминокислотных замен. В другом варианте осуществления антитело содержит LCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:368 и 384, с 4 или менее консервативных аминокислотных замен. В другом варианте осуществления антитело содержит LCVR, имеющую аминокислотную последовательность, выбранную из SEQ ID NO:368 и 384, с 2 или менее консервативных аминокислотных замен.
В некоторых вариантах осуществления антитело или фрагмент антитела по настоящему изобретению может быть конъюгировано с терапевтической составляющей, («иммуноконъюгат»), такой как цитотоксин, химиотерапевтическое лекарственное средство и иммунодепрессант или радиоизотоп.
«Выделенное антитело», как используется в настоящем описании, означает антитело, которое было идентифицировано и выделено, и/или отделено от по крайней мере одного компонента его природного окружения. Например, антитело, которое было выделено или отделено от по крайней мере одного компонента организма, ткани или клетки, в котором(ой) антитело существует в природе или продуцируется в природе, является «выделенным антителом» в рамках настоящего изобретения. Выделенное антитело также содержит антитело in situ в рекомбинантной клетке, а также антитело, которое было подвергнуто по крайней мере одной стадии очистки или выделения. В соответствии с некоторыми вариантами осуществления выделенное антитело может по существу не содержать другой клеточный материал и/или другие химические вещества.
Используемый в настоящем описании термин «поверхностный плазмонный резонанс» относится к оптическому явлению, которое позволяет исследовать биоспецифические взаимодействия в реальном времени посредством выявления изменений концентраций белка в матрице биосенсора, используя, например, систему BIACORETM (Pharmacia Biosensor AB, Uppsala, Швеция и Piscataway, N.J.).
Используемый в настоящем описании термин «KD», как предполагается, относится к равновесной константе диссоциации конкретного взаимодействия антитело-антиген.
Термин «эпитоп» включает любую детерминанту, предпочтительно, полипептидную детерминанту, способную к специфическому связыванию с иммуноглобулином или T-клеточным рецептором. В некоторых вариантах осуществления эпитопы содержат химически активные поверхностные группировки молекул, такие как аминокислоты, сахарные боковые цепи, фосфорильные группы или сульфонильные группы, и в некоторых вариантах осуществления могут обладать специфическими характеристиками трехмерной структуры и/или специфическими зарядными характеристиками. Эпитоп представляет собой район антигена, который связывается антителом. В некоторых вариантах осуществления говорят, что антитело специфически связывается с антигеном, если оно, предпочтительно, распознает являющийся его мишенью антиген в сложной смеси белков и/или макромолекул. Например, говорят, что антитело специфически связывается с антигеном, если KD составляет менее или равна 10-8 M, составляет менее или равна 10-9 M или составляет менее или равна 10-10 M.
Белок или полипептид является «по существу чистым», «по существу гомогенным» или «в значительной степени очищенным», когда по крайней мере приблизительно 60-75% образца демонстрирует один вид полипептида. Полипептид или белок может быть мономерным или мультимерным. По существу чистый полипептид или белок, как правило, содержит приблизительно 50%, 60%, 70%, 80% или 90% в весовом отношении образца белка, обычно приблизительно 95%, и, предпочтительно, характеризоваться составляющей свыше 99% степенью чистоты. О чистоте белка или его гомогенности может свидетельствовать ряд способов, хорошо известных в данной области, таких как электрофорез в полиакриламидном геле образца белка с последующей визуализацией единственной полипептидной полосы после окрашивания геля красящим веществом, хорошо известным в данной области. Для некоторых целей можно обеспечить большую разрешающую способность, используя HPLC или другие способы, хорошо известные в данной области для очистки.
Используемый в настоящем описании термин «аналог или вариант полипептида» относится к полипептиду, который состоит из сегмента по крайней мере из 25 аминокислот, который по существу идентичен части аминокислотной последовательности, и который обладает по крайней мере одним из следующих свойств: (1) специфическим связыванием с GDF8 в подходящих условиях связывания или (2) способностью к блокированию биологической активности GDF8. Как правило, аналоги или варианты полипептида содержат консервативную аминокислотную замену (или вставку, или делецию) относительно встречающейся в природе последовательности. Характерная длина аналогов составляет по крайней мере 20 аминокислот, по крайней мере 50, 60, 70, 80, 90, 100, 150 или 200 аминокислот или более, и они часто могут быть размером с полноразмерный встречающийся в природе полипептид.
Предпочтительными аминокислотными заменами являются замены, которые: (1) уменьшают подверженность протеолизу, (2) уменьшают подверженность окислению, (3) изменяют аффинность связывания в отношении образования белковых комплексов, (4) изменяют аффинности связывания и (4) придают таким аналогам другие физико-химические или функциональные свойства или изменяют их. Аналоги могут включать различные мутации последовательности, отличные от встречающейся в природе пептидной последовательности. Например, одну или множество аминокислотных замен (предпочтительно, консервативных аминокислотных замен) можно осуществить во встречающейся в природе последовательности (предпочтительно, в части полипептида, находящейся вне домена(ов), образующего межмолекулярные контакты. Консервативная аминокислотная замена не должна в значительной степени изменять структурные характеристики исходной последовательности (например, аминокислота замены не должна проявлять тенденцию к разрушению спирали, которая встречается в исходной последовательности, или нарушению других типов вторичной структуры, которые характеризуют исходную последовательность). Примеры принятых в данной области вторичных и третичных структур полипептидов описаны в Proteins, Structures and Molecular Principles (Creighton 1984 W. H. Freeman and Company, New York; Introduction to Protein Structure (Branden & Tooze, eds., 1991, Garland Publishing, NY); и Thornton et al. 1991 Nature 354: 105.
Непептидные аналоги обычно используются в фармацевтической промышленности в качестве лекарственных средств, обладающих свойствами, аналогичными таковым матричного пептида. Эти типы непептидного соединения называют «миметиками пептидов» или «пептидомиметиками» (см., например, Fauchere (1986) J. Adv. Drug Res. 15: 29; и Evans et al. (1987) J. Med. Chem. 30: 1229). Методичное замещение одной или более аминокислот консенсусной последовательности D-аминокислотой того же типа (например, D-лизином вместо L-лизина) может также использоваться для создания более стабильных пептидов. Кроме того, пептиды с ограничениями, содержащие консенсусную последовательность или вариант, по существу идентичный консенсусной последовательности, можно создать с помощью известных в данной области способов (Rizo et al. (1992) Ann. Rev. Biochem. 61: 387), например, посредством добавления внутренних остатков цистеина, способных образовывать внутримолекулярные дисульфидные мостики, которые приводят к циклизации пептида.
Термин «процент идентичности последовательностей» применительно к последовательностям нуклеиновых кислот относится к остаткам в двух последовательностях, которые являются одинаковыми после совмещения для максимального соответствия. Участок сравнения для определения идентичности последовательностей может быть больше чем участок из по крайней мере приблизительно девяти нуклеотидов или более, обычно по крайней мере приблизительно 18 нуклеотидов, обычнее по крайней мере приблизительно 24 нуклеотида, как правило, по крайней мере приблизительно 28 нуклеотидов, как правило, по крайней мере приблизительно 32 нуклеотидов и, предпочтительно, по крайней мере приблизительно 36, 48 или более нуклеотидов. В данной области известен ряд различных алгоритмов, которые могут использоваться для определения идентичности нуклеотидных последовательностей. Например, полинуклеотидные последовательности можно сравнить, используя FASTA, Gap или Bestfit, которые являются программами в пакете Wisconsin версии 10.0, Genetics Computer Group (GCG), Madison, Wis. FASTA, который содержит, например, программы FASTA2 и FASTA3, обеспечивает совмещения и процент идентичности последовательностей участков наилучшего совмещения между последовательностями запроса и поиска (Pearson (1990) Methods Enzymol. 183: 63-98 and (2000) Methods Mol. Biol. 132: 185-219). Кроме особо оговоренных случаев, используют параметры по умолчанию для конкретной программы или алгоритма. Например, процент идентичности между последовательностями нуклеиновых кислот можно определить, используя FASTA вместе с параметрами по умолчанию для нее (размер слова - 6 и опция NOPAM для матрицы замен) или используя Gap вместе с параметрами по умолчанию для нее, предоставленной в GCG версии 6.1.
Ссылка на последовательность нуклеиновой кислоты содержит ее комплемент, кроме особо оговоренных случаев. Таким образом, ссылка на молекулу нуклеиновой кислоты, имеющую конкретную последовательность, как следует истолковывать, содержит комплементарную ей цепь, с комплементарной ей последовательностью. Как правило, термины «процент идентичности последовательностей», «процент подобия последовательностей» и «процент гомологии последовательностей» используются в данной области взаимозаменяемо. В настоящей заявке эти термины имеют такое же значение по отношению к последовательностям нуклеиновых кислот.
Термин «значительное подобие» или «значительное подобие последовательностей», при ссылке на нуклеиновую кислоту или ее фрагмент, означает, что, после оптимального совмещения с использованием соответствующих вставок или делеций нуклеотидов с другой нуклеиновой кислотой (или комплементарной ей цепью), в по крайней мере приблизительно 90%, по крайней мере приблизительно 95%, по крайней мере приблизительно 96%, по крайней мере приблизительно 97%, по крайней мере приблизительно 98% или по крайней мере приблизительно 99% оснований нуклеотидов существует идентичность нуклеотидных последовательностей, определяемая с помощью любого широко известного алгоритма идентичности последовательностей, такого как FASTA, BLAST или Gap, обсуждавшихся выше.
Применительно к полипептидам, термин «значительная идентичность» или «в значительной степени идентичные» означает, что две пептидные последовательности, после оптимального совмещения, например, с помощью программ GAP или BESTFIT с использованием значений для пропусков по умолчанию, имеют составляющую по крайней мере приблизительно 80% идентичность последовательностей, составляющую по крайней мере приблизительно 90%, по крайней мере приблизительно 95%, по крайней мере приблизительно 98% или по крайней мере приблизительно 99% идентичность последовательностей. Предпочтительно, когда отличиями положений остатков, которые не являются идентичными, являются консервативные аминокислотные замены. «Консервативной аминокислотной заменой» является замена, в случае которой аминокислотный остаток заменен другим аминокислотным остатком, имеющим боковую цепь (группу R), обладающую схожими химическими свойствами (например, зарядом или гидрофобностью). Обычно консервативная аминокислотная замена не будет в значительной степени изменять функциональные свойства белка. В тех случаях, когда отличиями друг от друга двух или более аминокислотных последовательностей являются консервативные замены, процент идентичности последовательностей или степень подобия может быть увеличен для внесения поправки на консервативную природу замены. Способы осуществления этой корректировки хорошо известны квалифицированным в данной области специалистам. Смотрите, например, Pearson (1994) Methods Mol. Biol. 24: 307-331. Примеры групп аминокислот, которые имеют боковые цепи со схожими химическими свойствами, содержат 1) алифатические боковые цепи: глицин, аланин, валин, лейцин и изолейцин; 2) алифатические гидроксильные боковые цепи: серин и треонин; 3) амидсодержащие боковые цепи: аспарагин и глютамин; 4) ароматические боковые цепи: фенилаланин, тирозин и триптофан; 5) основные боковые цепи: лизин, аргинин и гистидин; и 6) серосодержащие боковые цепи: цистеин и метионин. Предпочтительными группами консервативных аминокислотных замен являются: валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, глютамат-аспартат и аспарагин-глютамин. Альтернативно, консервативной заменой является любая замена с положительным значением в матрице логарифмического правдоподобия PAM250, раскрытой в Gonnet et al. (1992) Science 256: 1443-45. «Умеренно консервативной» заменой является любая замена с неотрицательным значением в матрице логарифмического правдоподобия PAM250.
Подобие последовательностей полипептидов, которую также называют идентичностью последовательностей, как правило, определяют, используя программное обеспечение для анализа последовательностей. Программное обеспечение для анализа белков сопоставляет схожие последовательности, используя показатели подобия, заданные для различных замен, делеций и других модификаций, в том числе консервативных аминокислотных замен. Например, GCG содержит такие программы, как «Gap» и «Bestfit», которые могут использоваться вместе с параметрами по умолчанию для определения гомологии последовательностей или идентичности последовательностей между близкородственными полипептидами, такими как гомологичные полипептиды различных видов организмов, или между белком дикого типа и его мутеином. См., например, GCG версии 6.1. Полипептидные последовательности можно также сравнить, используя FASTA с использованием параметров по умолчанию или рекомендуемых параметров, программу в GCG версии 6.1. FASTA (например, FASTA2 и FASTA3) обеспечивает совмещения и процент идентичности последовательностей участков наилучшего совмещения между последовательностями запроса и поиска (Pearson (2000), выше). Другим предпочтительным алгоритмом при сравнении последовательности по настоящему изобретению с базой данной, содержащей большое количество последовательностей различных организмов, является компьютерная программа BLAST, особенно blastp или tblastn, с использованием параметров по умолчанию. См., например, Altschul et al. (1990) J. Mol. Biol. 215: 403-410 и Altschul et al. (1997) Nucleic Acids Res. 25: 3389 402.
Размер полипептидных последовательностей, сравниваемых на предмет гомологии, как правило, составляет по крайней мере приблизительно 16 аминокислотных остатков, по крайней мере приблизительно 20 остатков, по крайней мере приблизительно 24 остатков, по крайней мере приблизительно 28 остатков или по крайней мере приблизительно 35 остатков. При переборе базы данных, содержащей последовательности большого количества различных организмов, предпочтительным является сравнение аминокислотных последовательностей.
Термин «эффективное количество» означает концентрацию или количество антитела или антигенсвязывающего фрагмента антитела, которая приводит к достижению конкретной заданной цели. «Эффективное количество» антитела против GDF8 или его антигенсвязывающего фрагмента можно определить эмпирически. Кроме того, «терапевтически эффективное количество» представляет собой концентрацию или количество антитела против GDF8 или его антигенсвязывающего фрагмента, которое является эффективным для достижения заданного терапевтического эффекта. Это количество можно также определить эмпирически.
Получение антител человека
Способы создания моноклональных антител, в том числе полностью человеческих моноклональных антител, хорошо известны в данной области. Любые такие известные способы могут использоваться в рамках настоящего изобретения для получения антител человека, которые специфически связываются с GDF8.
Используя технологию VELOCIMMUNETM или любой другой известный способ создания моноклональных антител, в начале выделяют химерные антитело против GDF8, обладающие высокой аффинностью, содержащие человеческую вариабельную область и мышиную константную область. Как и в экспериментальной части ниже, получают характеристики антител, и осуществляют отбор антител в отношении желательных характеристик, содержащих аффинность, селективность, эпитоп и т.д. Мышиные константные области заменяют желаемой человеческой константной областью для создания полностью антитела человека по настоящему изобретению, например IgG1 или IgG4 дикого типа или модифицированного IgG1 или IgG4. Тогда как выбранная константная область может изменяться в соответствии с конкретным применением, характеристики, относящиеся к высокоаффинному связыванию с антигеном и специфичности в отношении мишени, присущи вариабельной области.
Обычно антитела по настоящему изобретению обладают очень высокой аффинностью, как правило, обладая KD, составляющей от приблизительно 10-12 по приблизительно 10-9 M, при определении посредством связывания с антигеном, который либо иммобилизован на твердой фазе, либо находится в жидкой фазе. Мышиные константные области заменяют желаемыми человеческими константными областями для создания полностью человеческих антител по настоящему изобретению, например, IgG1 дикого типа (SEQ ID NO:335) или IgG4 дикого типа (SEQ ID NO:336), или модифицированного IgG1 или IgG4 (например, SEQ ID NO:337). Тогда как выбранная константная область может изменяться в соответствии с конкретным применением, характеристики, относящиеся к высокоаффинному связыванию с антигеном и специфичности в отношении мишени, присущи вариабельной области.
Биоэквиваленты
Антитела против GDF8 и фрагменты антител по настоящему изобретению содержат белки с аминокислотными последовательностями, которые отличаются от таковых описанных антител, но которые сохраняют способность к связыванию с GDF8 человека. Такие варианты антител и фрагментов антител содержат одно или более добавлений, делеций или замен аминокислот по сравнению с исходной последовательностью, но проявляют биологическую активность, которая по существу эквивалента таковой описанных антител. Точно так же, кодирующие антитела против GDF8 последовательности ДНК по настоящему изобретению включают последовательности, которые содержат одно или более добавлений, делеций или замен нуклеотидов по сравнению с раскрытой последовательностью, но которые кодируют антитело против GDF8 или фрагмент антитела, которое(ый) по существу биоэквивалентно антителу против GDF8 или фрагменту антитела по настоящему изобретению. Примеры таких вариантов аминокислотных и ДНК последовательностей обсуждаются выше.
Два антигенсвязывающих белка, или антитела, считаются биоэквивалентными, если, например, они являются фармацевтическими эквивалентами или фармацевтическим вариантами, скорость и степень абсорбции которых не отличаются значительно после введения в одинаковой молярной дозе, либо однократной дозе, либо в многократной дозе, в схожих экспериментальных условиях. Некоторые антитела будут считаться эквивалентами или фармацевтическими вариантами, если они являются эквивалентными по степени их абсорбции, но не по скорости их абсорбции, и все еще могут считаться биоэквивалентными, поскольку такие различия в скорости абсорбции являются умышленными и находят отражение при отнесении к определенной категории, не являются существенными для достижения эффективных концентраций лекарственного средства в организме, например, при постоянном применении, и считаются терапевтически незначимыми для конкретного исследуемого лекарственного продукта.
В одном из вариантов осуществления два антигенсвязывающих белка являются биоэквивалентными, если нет клинически значимых различий в их безопасности, степени чистоты и эффективности.
В одном из вариантов осуществления два антигенсвязывающих белка являются биоэквивалентными, если пациента можно перевести один или более раз с контрольного продукта на биологический продукт без ожидаемого увеличения риска неблагоприятных эффектов, содержащих клинически значимое изменение иммуногенности, или уменьшения эффективности, по сравнению с продолжающейся терапией без такого перевода.
В одном из вариантов осуществления два антигенсвязывающих белка являются биоэквивалентными, если они оба функционируют по общему механизму или механизмам действия для используемого состояния или состояний, в случае, если такие механизмы известны.
Биоэквивалентность может быть показана способами in vivo и in vitro. Показатели биоэквивалентности включают, например, (a) in vivo исследование на людях или других млекопитающих, в ходе которого определяют концентрацию антитела или его метаболитов в крови, плазме, сыворотке или другой биологической жидкости в зависимости от времени; (b) in vitro исследование, которое было увязано с данными, касающимися in vivo биодоступности у людей, и корректно предсказывает их; (c) in vivo исследование на людях или других млекопитающих, в ходе которого определяют соответствующий кратковременный фармакологический эффект антитела (или его мишени) в зависимости от времени; и (d) полностью контролируемое клиническое испытание, в ходе которого устанавливается безопасность, эффективность, или биодоступность или биоэквивалентность антитела.
Биоэквивалентные варианты антител против GDF8 по настоящему изобретению можно сконструировать с помощью, например, осуществления различных замен остатков или последовательностей, или делетирования концевых или внутренних остатков, или последовательностей, которые не требуются для биологической активности. Например, остатка цистеина, не являющиеся необходимыми для биологической активности, можно делетировать или заменить другими аминокислотами для предотвращения образования ненужных или неправильных внутримолекулярных дисульфидных мостиков при ренатурации. В других ситуациях биоэквивалентные антитела могут содержать варианты антител против GDF8, содержащие аминокислотные замены, которые изменяют особенности гликозилирования антител, например, мутации, которые исключают или устраняют гликозилирование.
Эпитопное картирование и родственные технологии
Для скрининга на предмет нахождения антител, которые связываются с конкретным эпитопом (например, те, которые блокируют связывание IgE со своим высокоаффинным рецептором), может быть выполнен обычный перекрестный конкурентный анализ, такой как тот, который описан в Harlow and Lane (1990), выше. Другие способы содержат сканирующий аланином мутагенез, блоты пептидов (Reineke (2004) Methods Mol Biol 248: 443-463) или анализ расщепления пептидов. Кроме того, могут использоваться такие способы, как исключение эпитопа, удаление эпитопа и химическая модификация антигенов (Tomer (2000) Protein Science 9: 487-496).
Термин «эпитоп» относится к участку антигена, на который реагируют B- и/или T-клетки. B-клеточные эпитопы могут быть образованы из расположенных рядом аминокислот или из не расположенных рядом аминокислот, которые стали соседними в результате свертывания белка в третичную структуру. Эпитопы, образованные из расположенных рядом аминокислот, обычно сохраняются при подверганию воздействию денатурирующих растворителей, тогда как эпитопы, образованные в результате свертывания в третичную структуру, обычно утрачиваются при обработке денатурирующими растворителями. Эпитоп обычно содержит по крайней мере 3, обычно по крайней мере 5 или 8-10 аминокислот в уникальной пространственной конформации.
Выполняемый с помощью модификаций анализ профиля (MAP), также известный как анализ профиля антител на основе структуры антигена (ASAP), является способом распределения по категориям больших количеств моноклональных антител (mAb), направленных против одного и того же антигена, в соответствии со сходствами профиля связывания каждого антитела с химически или ферментативно модифицированными поверхностями антигена (US 2004/0101920). Каждая категория может отражать уникальный эпитоп, либо явно отличный от эпитопа, представленного другой категорией, либо частично перекрывающийся с ним. Эта технология позволяет быстро отфильтровать генетически идентичные антитела, так что снятие характеристик может быть сосредоточено на генетичных отличных антителах. В случае использования для скрининга гибридом MAP может способствовать идентификации редких гибридомных клонов, которые продуцируют mAb с желаемыми характеристиками. MAP может использоваться для распределения антител против GDF8 по настоящему изобретению по группам антител, связывающихся с различными эпитопами.
Настоящее изобретение относится к антителам против GDF8 и антигенсвязывающим фрагментам антител, которые связываются со специфическими эпитопами GDF8 человека (SEQ ID NO:340), и способны к блокированию биологической активности GDF8. В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент связывается в эпитопе, содержащем аминокислотные остатки 1-109; 1-54; 1-44; 1-34; 1-24 и 1-14. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент связывается в эпитопе, содержащем аминокислотные остатки 65-72; 35-109; 45-109; 55-109; 65-109; 75-109; 85-109; 92-109 или 95-109. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент связывается в эпитопе, содержащем аминокислотные остатки 48-72; 48-69; 48-65; 52-72; 52-65 или 56-65. В особых вариантах осуществления антитело или его антигенсвязывающий фрагмент может связываться в 2 или более эпитопах.
Настоящее изобретение также относится к антителам и их антигенсвязывающим фрагментам, которые связываются со зрелым белком GDF8 дикого типа (SEQ ID NO:340), но не связываются с выделенными пептидами, имеющими менее чем полноразмерную аминокислотную последовательность SEQ ID NO:340. Например, настоящее изобретение относится к антителам против GDF8, которые связываются со зрелым белком GDF8 дикого типа (SEQ ID NO:340), но не связываются с выделенными пептидами, состоящими из 10-40 следующих друг за другом аминокислот SEQ ID NO:340. Настоящее изобретение также относится к антителам против GDF8, которые не связываются с какими-либо линейными эпитопами зрелого белка GDF8 дикого типа. В некоторых вариантах осуществления по настоящему изобретению антитела против GDF8 связываются со зрелым белком GDF8 дикого типа, содержащим SEQ ID NO:340, но не связываются с одним или более выделенных пептидов GDF8, имеющих аминокислотную последовательность, выбранную из группы, состоящей из 1-14, 1-18, 17-42, 48-65, 48-69, 48-72, 52-65, 52-72, 56-65, 56-72, 65-72, 73-90, 75-105 и 91-105 SEQ ID NO:340. В некоторых вариантах осуществления антитела против GDF8 не связываются с любым из вышеупомянутых пептидов GDF8. Способы определения того, способно ли это антитело к связыванию с конкретным пептидом GDF8, известны средним специалистам в данной области. В примере 7 показан один пример способа, в котором пептиды GDF8 присоединяют к микросферическим частицам, к микросферическим частицам с конъюгированными пептидами добавляют антитела, и после стадий промывки выявляют микросферические частицы со связанными антителами. Отсутствие связанных антител означает, что антитела не связываются с конкретными исследуемыми пептидами.
Настоящее изобретение также относится к выделенным антителам человека или к их антигенсвязывающим фрагментам, которые специфически связываются со зрелым белком GDF8 человека дикого типа (например, белком или полипептидом, содержащим SEQ ID NO:340), но не связываются с химерной конструкцией CDF8, в которой некоторые аминокислоты GDF8 замены соответствующей аминокислотной последовательностью(ями) из неидентичного, но родственного белка, такого как TGFβ-1. В одном из примеров химерной конструкцией является химера GDF8/TGFβ-1, в которой аминокислоты 48-72 зрелого белка GDF8 замены соответствующей аминокислотной последовательностью TGFβ-1 (например, аминокислотами 49-76 TGFβ-1). SEQ ID NO:352 представляет собой пример одной такой химеры (см. примеры 4 и 6 в настоящем описании). Таким образом, в некоторых вариантах осуществления антитела по настоящему изобретению специфически связываются со зрелым белком GDF8 человека дикого типа (SEQ ID NO:340), но не связываются с химерной конструкцией GDF8/TGFβ-1 с SEQ ID NO:352, что указывает на то, что эпитоп, с которым связываются такие антитела, включает или содержит аминокислоты, локализованные в пределах остатков 48-72 SEQ ID NO:340. Биоанализы блокирования, такие как анализ, изложенный в примере 4 в настоящем описании, могут также использоваться для непрямого определения, связывается ли антитело со зрелым белком GDF8 человека дикого типа (SEQ ID NO:340), и не связывается ли оно с химерной конструкцией GDF8/TGFβ-1, например, конструкцией с SEQ ID NO:352. Например, антитело, которое блокирует биологическую активность зрелого белка GDF8 человека дикого типа, но не блокирует биологическую активность химерной конструкцией GDF8/TGFβ-1, как считают, связывается с частью GDF8, которая заменена соответствующей последовательностью TGFβ-1 в химерной конструкции.
Точно так же, настоящее изобретение также относится к выделенным антителам человека или к их антигенсвязывающим фрагментам, которые блокируют опосредованную зрелым белком GDF8 дикого типа активность в биоанализе, но не блокируют активность химерной конструкции GDF8 (например, химеры GDF8/TGFβ-1, в которой аминокислоты 48-72 зрелого белка GDF8 замены соответствующей аминокислотной последовательностью TGFβ-1 (например, SEQ ID NO:352)). Приводимым в качестве примера биоанализом GDF8, который может использоваться в рамках этого аспекта настоящего изобретения, является исследование индуцируемой GDF8 люциферазной активности, изложенное в примере 4 в настоящем описании, хотя в настоящем описании также предусматриваются другие схожие биоанализы, с помощью которых можно определить клеточную активность GDF8.
Настоящее изобретение относится к антителам против GDF8, которые связываются с одним и тем же эпитопом, как и любые из конкретных, приводимых в качестве примера антител, описываемых в настоящем описании. Точно так же, настоящее изобретение также относится к антителам против GDF8, которые перекрестно конкурируют за связывание с GDF8 или фрагментом GDF8 с любым из конкретных, приводимых в качестве примера антител, описываемых в настоящем описании.
Можно легко определить, связывается ли антитело с тем же эпитопом, что и контрольное антитело против GDF8, или конкурирует ли антитело за связывание с контрольным антителом, используя обычные способы, известные в данной области. Например, для определения того, связывается ли исследуемое антитело с тем же эпитопом, что и контрольное антитело против GDF8 по настоящему изобретению, допускают связывание контрольного антитела с белком или пептидом GDF8 в условиях насыщения. Затем оценивают способность исследуемого антитела к связыванию с молекулой GDF8. Если исследуемое антитело способно к связыванию с GDF8 после связывания в условиях насыщения с контрольным антителом против GDF8, можно сделать вывод, что исследуемое антитело связывается с эпитопом, отличным от такового для контрольного антитела против GDF8. С другой стороны, если исследуемое антитело не способно к связыванию с молекулой GDF8 после связывания в условиях насыщения с контрольным антителом против GDF8, то исследуемое антитело может связываться с эпитопом, одинаковым с эпитопом, с которым связывается контрольное антитело против GDF8 по настоящему изобретению. Можно также провести дополнительное рутинное экспериментирование (например, мутационный анализ пептидов и анализ связывания пептидов) для подтверждения, действительно ли отмеченное отсутствие связывания исследуемого антитела обусловлено связыванием с эпитопом, одинаковым с таковым для контрольного антитела, или ответственно ли за отсутствие наблюдаемого связывания стерическое блокирование (или другое явление). Эксперименты такого типа можно выполнить, используя ELISA, RIA, Biacore, проточную цитометрию или любые другие количественные или качественные анализы связывания антител, имеющиеся в данной области. В соответствии с некоторыми вариантами осуществления по настоящему изобретению два антитела связываются с одним и тем же (или перекрывающимся) эпитопом, если, например, 1-, 5-, 10-, 20- или 100-кратное избыточное количество одного антитела ингибирует связывание другого антитела на по крайней мере 50%, но, предпочтительно, на 75%, 90% или даже 99%, что определено в конкурентном анализе связывания (см., например, Junghans et al., Cancer Res. 1990:50:1495-1502). Альтернативно, считают, что два антитела связываются с одним и тем же эпитопом, если по существу все мутации аминокислот в антигене, которые уменьшают или исключают связывание одного антитела, уменьшают или исключают связывание другого антитела. Считают, что антитела связываются с «перекрывающимися эпитопами», если лишь подмножество мутаций аминокислот, которые уменьшают или исключают связывание одного антитела, уменьшают или исключают связывание другого антитела.
Для определения того, конкурирует ли антитело за связывание с контрольным антителом против GDF8, описанную выше методику связывания выполняют в двух установках. В первой установке допускают связывание контрольного антитела с молекулой GDF8 в условиях насыщения с последующей оценкой связывания исследуемого антитела с молекулой GDF8. Во второй установке допускают связывание исследуемого антитела с молекулой GDF8 в условиях насыщения с последующей оценкой связывания контрольного антитела с молекулой GDF8. Если, в обеих установках, лишь первое (используемое для насыщения) антитело способно к связыванию с молекулой GDF8, то делают вывод, что исследуемое антитело и контрольное антитело конкурируют за связыванием с GDF8. Как это будут понятно среднему специалисту в данной области, антитело, которое конкурирует за связывание с контрольным антителом, может необязательно связываться с тем же эпитопом, что и контрольное антитело, но может стерически блокировать связывание контрольного антитела в результате связывания с перекрывающимся или расположенным рядом эпитопом.
Видовая селективность и межвидовая перекрестная реактивность
В соответствии с некоторыми вариантами осуществления по настоящему изобретению антитела против GDF8 связываются с GDF8 человека, но не с GDF8 других видов. Альтернативно, антитела против GDF8 по настоящему изобретению, в некоторых вариантах осуществления, связываются с GDF8 человека и с GDF8 одного или более не являющихся людьми видов. Например, антитела против GDF8 по настоящему изобретению могут связываться с GDF8 человека и могут связываться или могут не связываться, в зависимости от конкретного случая, с одним или более GDF8 мыши, крысы, морской свинки, хомячка, песчанки, свиньи, кошки, собаки, кролика, козы, овцы, коровы, лошади, верблюда, яванского макака, мартышки, макака-резус или шимпанзе.
Иммуноконъюгаты
Настоящее изобретение относится к антителу человека или гуманизированному моноклональному антителу против GDF8, конъюгированному с терапевтической составляющей («иммуноконъюгат»), такой как цитотоксин, химиотерапевтическое лекарственное средство, иммунодепрессант или радиоизотоп. Цитотоксические агенты включают любой агент, который является вредным для клеток. Примеры подходящих цитотоксических агентов и химиотерапевтических средств для образования иммуноконъюгатов известны в данной области, см., например, WO 05/103081.
Полиспецифические антитела
Антитела по настоящему изобретению могут быть моноспецифическими, биспецифическими или полиспецифическими. Полиспецифические антитела могут быть специфическими для различных эпитопов одного полипептида-мишени или могут содержать антигенсвязывающие домены, специфические для более чем одного полипептида-мишени. См., например, Tutt et al., 1991, J. Immunol. 147: 60-69; Kufer et al., 2004, Trends Biotechnol. 22: 238-244. Антитела против GDF8 по настоящему изобретению можно связать или коэкспрессировать с другой функциональной молекулой, например, другим пептидом или белком. Например, антитело или его фрагмент можно функционально связать (например, с помощью химической связи, генетического слияния, нековалентной связи или иного) с одной или более других молекулярных структур, таких как другое антитело или фрагмент антитела, с целью получения биспецифического или полиспецифического антитела со второй специфичность связывания. Например, настоящее изобретение относится к биспецифическим антителам, при этом одно плечо иммуноглобулина является специфическим для GDF8 человека или его фрагмента, а другое плечо иммуноглобулина является специфическим для второй терапевтической мишени или конъюгировано с терапевтической составляющей.
Приводимый в качестве примера формат биспецифического антитела, который можно использовать в рамках настоящего изобретения, включает использование СН3-домена первого иммуноглобулина (Ig) и СН3-домена второго Ig, причем СН3-домены первого и второго Ig отличаются друг от друга по крайней мере одной аминокислотой, и причем различие в по крайней мере одной аминокислоте уменьшает связывание биспецифического антитела с белком A по сравнению с биспецифическим антителом, в котором отсутствует различие в аминокислоте. В одном из вариантов осуществления СН3-домен первого Ig связывается с белком A, а СН3-домен второго Ig содержит мутацию, которая уменьшает или аннулирует связывание с белком A, такую как модификации H95R (в соответствии с нумерацией экзонов IMGT; H435R в соответствии с нумерацией EU). Второй СН3 может, кроме того, содержать модификацию Y96F (в соответствии с IMGT; Y436F в соответствии с EU). Дополнительные модификации, которые можно обнаружить во втором CH3, включают: D16E, L18M, N44S, K52N, V57M и V82I (в соответствии с IMGT; D356E, L358M, N384S, K392N, V397M и V422I в соответствии с ЕС) в случае антител IgG1; N44S, K52N и V82I (IMGT; N384S, K392N и V422I в соответствии с ЕС) в случае антител IgG2; и Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (в соответствии с IMGT; Q355R, N384S, K392N, V397M, R409K, E419Q и V422I в соответствии с ЕС) в случае антител IgG4. Вариации в формате биспецифического антитела, описанные выше, предполагаются находящимися в объеме настоящего изобретения.
Терапевтическое введение и препараты
Настоящее изобретение относится к терапевтическим композициям, содержащим антитела или их антигенсвязывающие фрагменты по настоящему изобретению. Введение терапевтических композиций в соответствии с настоящим изобретением осуществляется вместе с подходящими носителями, наполнителями и другими агентами, которые содержатся в препараты для обеспечения улучшенного переноса, доставки, переносимости и т.п. Множество соответствующих препаратов можно найти в фармакологическом справочнике, известном всем фармакохимикам: Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, PA. Эти препараты включают, например, порошки, пасты, мази, гели, воски, масла, липиды, содержащие липиды (катионные или анионные) везикулы (такие как LIPOFECTINTM), ДНК-конъюгаты, безводные пасты с абсорбционными свойствами, эмульсии типа «масло-в-воде» или «вода-в-масле», карбовакс (полиэтиленгликоли с различными молекулярными массами), полутвердые гели и полутвердые смеси, содержащие карбовакс. Любая из вышеупомянутых смесей может быть подходящей для лечений и терапий в соответствии с настоящим изобретением при условии, что препарат не инактивирует активный ингредиент в препарате, и препарат является физиологически совместимым и переносимым при использовании пути введения. См. также Powell et al. "Compendium of excipients for parenteral formulations" PDA (1998) J Pharm Sci Technol 52: 238-311.
Доза может изменяться в зависимости от возраста и массы тела индивида, которому ее вводят, болезни-мишени, состояний, пути введения и т.п. Если антитело по настоящему изобретению используют для лечения различных состояний и заболеваний, связанных с GDF8, например, мышечной дистрофии, мышечной атрофии, синдрома истощения мышечной массы, саркопении и кахексии, у взрослого пациента, то преимущественным является внутривенное введение антитела по настоящему изобретению, которое обычно проводят в однократной дозе, составляющей от приблизительно 0,01 до приблизительно 20 мг/кг веса тела, от приблизительно 0,1 до приблизительно 10 мг/кг веса тела или от приблизительно 0,1 до приблизительно 5 мг/кг веса тела. Частоту введения и продолжительность лечения можно регулировать в зависимости от тяжести состояния. В случае другого парентерального введения и перорального введения антитело можно вводить в дозе, соответствующей приведенной выше дозе. Когда состояние является особенно тяжелым, дозу можно увеличить в соответствии с состоянием вплоть до количества, которое вызывает значительные побочные эффекты, если это имеет место.
Для введения фармацевтической композиции по настоящему изобретению различные системы доставки известны и могут использоваться, например, инкапсуляция в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать мутантные вирусы, рецептор-опосредованный эндоцитоз (см., например, Wu et al. (1987) J. Biol. Chem. 262: 4429-4432). Способы введения включают, но без ограничения, внутрикожный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, интраназальный, эпидуральный и пероральный пути. Композицию можно вводить любым удобным путем, например, посредством инфузии или болюсной инъекции, посредством абсорбции через эпителиальные или кожно-слизистые выстилки (например, слизистую оболочку ротовой полости, слизистую оболочку прямой кишки и кишечника и т.д.), и ее можно вводить вместе с другими биологически активными агентами. Введение может быть системным или местным.
Фармацевтическую композицию можно также доставить в везикуле, в частности, липосоме (см. Langer (1990) Science 249: 1527-1533).
Фармацевтическую композицию по настоящему изобретению можно доставить подкожно или внутривенно с помощью стандартной иглы и шприца. Кроме того, что касается подкожной доставки, устройство для доставки в виде ручки охотно используют для доставки фармацевтической композиции по настоящему изобретению. Такое устройство для доставки в виде ручки может быть многократного или одноразового использования. В устройстве для доставки в виде ручки многократного использования, как правило, используется заменяемый картридж, который содержит фармацевтическую композицию. После того, как вся фармацевтическая композиция, находящаяся в картридже, введена, и картридж является пустым, пустой картридж можно без труда удалить и заменить новым картриджем, который содержит фармацевтическую композицию. Устройство для доставки в виде ручки можно затем снова использовать. В случае одноразового устройства для доставки в виде ручки заменяемого картриджа нет. Точнее, одноразовое устройство для доставки в виде ручки поставляется предварительно наполненным фармацевтической композицией, удерживаемой в резервуаре внутри устройства. Коль скоро резервуар не содержит фармацевтическую композицию, пустое устройство выбрасывают.
Многочисленные устройства для доставки в виде ручек и автоинжекторов многократного использования применяют для подкожной доставки фармацевтической композиции по настоящему изобретению. Примеры включают, но без ограничения, AUTOPENTM (Owen Mumford, Inc., Woodstock, UK), ручку DISETRONICTM (Disetronic Medical Systems, Bergdorf, Швейцария), ручку HUMALOG MIX 75/25TM, ручку HUMALOGTM, ручку HUMALIN 70/30TM (Eli Lilly and Co., Indianapolis, IN), NOVOPENTM I, II и III (Novo Nordisk, Copenhagen, Дания), NOVOPEN JUNIORTM (Novo Nordisk, Copenhagen, Дания), ручку BDTM (Becton Dickinson, Franklin Lakes, NJ), OPTIPENTM, OPTIPEN PROTM, OPTIPEN STARLETTM и OPTICLIKTM (Sanofi-Aventis, Frankfurt, Германия), не говоря уже у других. Примеры одноразовых устройств для доставки в виде ручек, применяемых для подкожной доставки фармацевтической композиции по настоящему изобретению, включают, но без ограничения, ручку SOLOSTARTM (Sanofi-Aventis), FLEXPENTM (Novo Nordisk) и KWIKPENTM (Eli Lilly), автоинжектор SURECLICKTM (Amgen, Thousand Oaks, CA), PENLETTM (Haselmeier, Stuttgart, Германия), EPIPEN (Dey, L.P.) и ручку HUMIRATM (Abbott Labs, Abbott Park IL), не говоря уже у других.
В некоторых ситуациях фармацевтическая композиция может доставляться в системе контролируемого высвобождения, например, с использованием насоса или полимерных материалов. В другом варианте осуществления систему контролируемого высвобождения можно поместить вблизи мишени композиции, тем самым приводя к необходимости лишь части системной дозы.
Примеры композиции для перорального введения включают твердые или жидкие лекарственные формы, особенно таблетки (в том числе драже и покрытые оболочкой таблетки), пилюли, гранулы, порошкообразные препараты, капсулы (в том числе мягкие капсулы), сироп, эмульсии, суспензии и т.д. Такую композицию получают с помощью общеизвестных способов, и она содержит носитель, разбавитель или наполнитель, обычно используемый в области фармацевтических препаратов. Примерами носителя или наполнителя для таблеток являются лактоза, крахмал, сахароза, стеарат магния и т.п.
Инъецируемые препараты могут включать лекарственные формы для внутривенных, подкожных, внутрикожных и внутримышечных инъекций, капельных инфузий и т.д. Эти инъецируемые препараты можно получить с помощью общеизвестных способов. Например, инъецируемые препараты можно получить, например, посредством растворения, суспендирования или превращения в эмульсию антитела или его солей, описанных выше, в стерильной водной среде или масляной среде, обычно используемой для инъекций. В качестве водной среды для инъекций существуют, например, физиологический солевой раствор, изотонический раствор, содержащий глюкозу и другие вспомогательные средства, и т.д., которые могут использоваться в сочетании с соответствующим солюбилизатором, таким как спирт (например, этиловый спирт), полиспирт (например, пропиленгликоль, полиэтиленгликоль), неионное поверхностно-активное вещество [например, полисорбат 80, HCO-50 (продукт присоединения полиоксиэтилена (50 моль) к гидрогенизированному касторовому маслу], и т.д. В качестве масляной среды используют, например, кунжутное масло, соевое масло и т.д., которые могут использоваться в сочетании с соответствующим солюбилизатором, таким как бензилбензоат, бензиловый спирт и т.д. Инъекционным раствором, полученным таким образом, предпочтительно, наполняют подходящую ампулу.
Преимущественно, фармацевтические композиции для перорального или парентерального применения, описанные выше, получают в лекарственных формах в стандартной дозе, соответствующей дозе активных ингредиентов. Такие лекарственные формы в стандартной дозе включают, например, таблетки, пилюли, капсулы, инъекционные растворы (ампулы), суппозитории и т.д. Количество ранее указанного содержащегося антитела составляет, как правило, приблизительно 5-500 мг в каждой лекарственной форме в стандартной дозе, особенно в форме инъекционного раствора; предпочтительно, когда ранее указанное антитело содержится в количестве, составляющем приблизительно 5-100 мг и приблизительно 10-250 мг, в случае других лекарственных форм.
Терапевтические применения антител
Антитела по настоящему изобретению можно использовать, кроме прочего, для лечения, профилактики и/или ослабления любого заболевания или нарушения, связанного с активностью GDF8. Конкретнее, антитела по настоящему изобретению могут использоваться для лечения любого состояния или болезни, которое можно улучшить посредством увеличения мышечной силы и/или мышечной массы, и/или функционирования мышц у индивидуума, или посредством положительного изменения метаболизма (углеводного, липидного и процессирования белков) в результате блокирования активности GDF8. Приводимые в качестве примера заболевания, нарушения и состояния, лечение которых можно проводить антителами против GDF8 по настоящему изобретению, включают, но ими не ограничиваются, саркопению, кахексию (или идиопатическую, или на фоне других состояний, например, рака, хронической почечной недостаточности или хронического обструктивного заболевания легких), травму мышцы, истощение мышечной массы и мышечную атрофию, например, мышечную атрофию или истощение мышечной массы, вызванную недогрузкой, обездвижением, постельным режимом, травмой, терапевтическим лечением или хирургическим вмешательством (например, переломом тазобедренного сустава, замещением тазобедренного сустава, замещением коленного сустава и т.д.) или необходимостью механической вентиляции, или связанную с ними. Антитела против GDF8 по настоящему изобретению могут также использоваться для лечения, профилактики и/или ослабления таких заболеваний, как рак, ожирение, диабет, артрит, рассеянный склероз, мышечная дистрофия, боковой амиотрофический склероз, болезнь Паркинсона, остеопороз, остеоартрит, остеопения, метаболические синдромы (содержащие, но без ограничения, диабет, ожирение, нарушения питания, атрофию органов, хроническое обструктивное заболевание легких и анорексию).
Настоящее изобретение относится к терапевтическим схемам введения, которые включают введение антитела против GDF8 по настоящему изобретению в сочетании с по крайней мере одним дополнительным терапевтически активным компонентом. Неограничивающие примеры таких дополнительных терапевтически активных компонентов включают другие антагонисты GDF8 (например, ингибиторы GDF8 в виде небольших молекул или другие антитела против GDF8 или связывающиеся с GDF8 молекулы), ингибиторы факторов роста, иммунодепрессанты, противовоспалительные средства, ингибиторы метаболизма, ингибиторы ферментов и цитотоксические/цитостатические агенты. Дополнительный терапевтически активный компонент(ы) можно вводить до введения антитела против GDF8 по настоящему изобретению, одновременно с ним или после такого введения.
Применения антител для диагностирования
Антитела против GDF8 по настоящему изобретению могут также использоваться для выявления и/или измерения GDF8 в образце, например, с диагностической целью. Например, антитело против GDF8, или его фрагмент, можно использоваться для диагностирования состояния или заболевания, характеризующегося аберрантной экспрессией (например, сверхэкспрессией, недостаточной экспрессией, отсутствием экспрессии и т.д.) GDF8. Приводимые в качестве примера диагностические исследования GDF8 могут включать, например, приведение образца, полученного от пациента, в контакт с антителом против GDF8 по настоящему изобретению, при этом антитело против GDF8 помечено выявляемой меткой или молекулой репортера. Альтернативно, в случае применений для диагностирования немеченое антитело против GDF8 может использоваться в сочетании со вторым антителом, которое само является выявляемым образом меченным. Выявляемой меткой или молекулой репортера может быть радиоизотоп, такой как 3H, 14C, 32P, 35S или 125I; флуоресцентная или хемилюминесцентная составляющая, такая как флуоресцеин изотиоцианат или родамин; или фермент, такой как щелочная фосфатаза, β-галактозидаза, пероксидаза хрена или люцифераза. Конкретные, приводимые в качестве примера исследования, которые могут использоваться для выявления или измерения GDF8 в образце, содержат иммуноферментный твердофазный анализ (ELISA), радиоиммуноанализ (RIA) и клеточный сортинг с активацией флуоресценции (FACS).
Образцы, которые могут использоваться в диагностических исследованиях GDF8 в соответствии с настоящим изобретением, содержат любой образец ткани или жидкости, который можно получить от пациента, который содержит выявляемые количества белка GDF8, или его фрагментов, в нормальных или патологических условиях. Как правило, будут определяться уровни GDF8 в конкретном образце, полученном от здорового пациента (например, пациента, не пораженного заболеванием или состоянием, связанным с анормальными уровнями или активностью GDF8), для установления в начале исходного, или стандартного, уровня GDF8. Этот исходный уровень GDF8 можно затем сопоставить с уровнями GDF8, определенными в образцах, полученных от индивидуумов с подозрением на наличие связанного GDF8 заболевания или состояния.
ПРИМЕРЫ
Пример 1. Создание антител человека против GDF8 человека
Мышей можно подвергнуть иммунизации любым способом, известным в данной области (см., например, Harlow и Lane, выше). В одном из вариантов осуществления антиген GDF8 вводят непосредственно, вместе с адъювантом для стимуляции иммунного ответа, мыши VELOCIMMUNE®, содержащей ДНК, кодирующую вариабельные области тяжелой цепи и легкой цепи каппа Ig человека. Подходящие адъюванты содержат полный и неполный адъювант Фрейнда, адъювантную систему MPL+TDM (Sigma) или RIBI (мурамилдипептиды) (смотрите O'Hagan 2000 Vaccine Adjuvant, by Human Press, Totawa, N·J). Гуморальный ответ отслеживают с помощью стандартного специфического в отношении антигена иммуноанализа. После достижения желаемого иммунного ответа, в одном из вариантов осуществления, экспрессирующие антитела B-клетки получают и сливают с клетками миеломы мыши для сохранения их жизнеспособности и образования линий гибридомных клеток. Линии гибридомных клеток подвергают скринингу и отбору для идентификации линий клеток, которые получают специфические в отношении антигена антитела.
Альтернативно, специфические в отношении антигена гибридомные клетки можно выделить с помощью проточной цитометрии. Коротко, после слияния с клетками миеломы объединенные гибридомные клетки выращивают в течение 10 дней в среде HAT. Затем клетки собирают и подвергают окрашиванию меченным биотином GDF8 в концентрации 2 мкг/мл в течение одного часа, с последующим добавлением фикоэритрина-стрептавидина. Флуоресцентно меченные клетки сортируют с помощью проточной цитометрии (одну клетку в каждую лунку 96-луночного планшета, содержащую среду для выращивания гибридомы), культивируют в течение 8-10 дней, и кондиционированные среды скринируют на предмет присутствия функционально желательных моноклональных антител.
В другом варианте осуществления антитела против GDF8 получают через прямое выделение спленоцитов. Специфические в отношении антигена антитела выделяют непосредственно из антиген-иммунизированных B-клеток без слияния с клетками миеломы, как описано в заявке на патент США № 2007/0280945A1. Стабильные линии рекомбинантных, экспрессирующих антитела клеток CHO устанавливают, исходя из выделенных специфических рекомбинантов.
Пример 2. Анализ использования генов
С целью анализа структуры продуцируемых антител были клонированы и секвенированы нуклеиновые кислоты, кодирующие вариабельные области антител. Исходя из последовательности нуклеиновой кислоты и предсказанной аминокислотной последовательности антител, было определено использование генов для каждой цепи антитела. В таблице 1 изложено использование генов для выбранных антител в соответствии с настоящим изобретением. Идентификаторы антител (HCVR/LCVR): 21-E5 (SEQ ID NO:34/42); 21-B9 (SEQ ID NO:18/26); 21-E9 (SEQ ID NO:98/106); 21-A2 (SEQ ID NO:2/10); 22-D3 (SEQ ID NO:50/58); 22-E6 (SEQ ID NO:66/74); 22-G10 (SEQ ID NO:82/90); 1A2 (SEQ ID NO:226/234); 20B12 (SEQ ID NO:274/282); 58C8 (SEQ ID NO:242/250); 19F2 (SEQ ID NO:258/266); 8D12-1 (SEQ ID NO: 114/122); 4E3-7 (SEQ ID NO:194/202); 9B11-12 (SEQ ID NO:162/170); 4B9 (SEQ ID NO:226/234); 1H4-5 (SEQ ID NO:210/218); 9B4-3 (SEQ ID NO:178/186); 3E2-1 (SEQ ID NO:290/298); 4G3-25 (SEQ ID NO:306/314); 4B6-6 (SEQ ID NO:130/138); H4H1657N2 (SEQ ID NO:360/368); H4H1669P (SEQ ID NO:376/384).
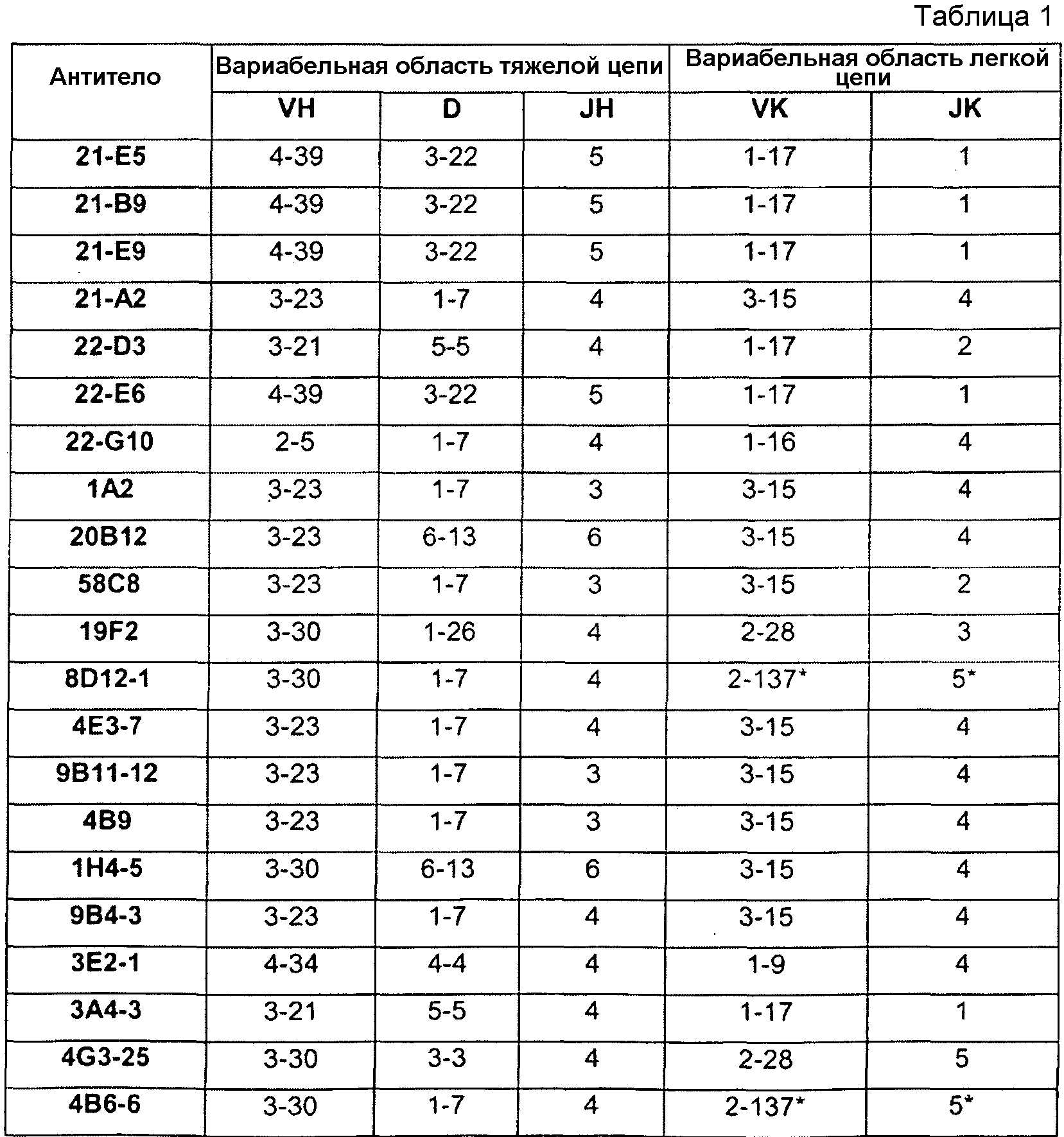
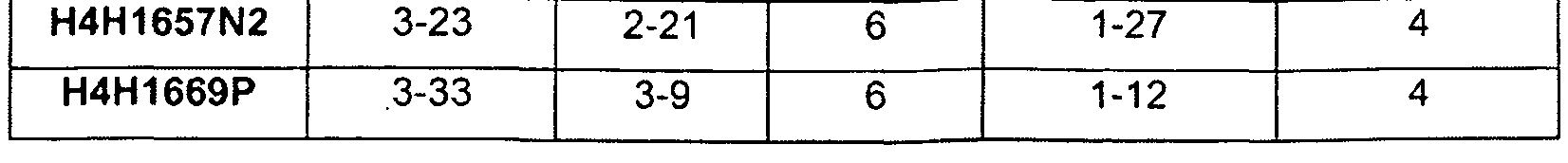
Контрольные конструкции, используемые в следующих примерах
С целью сравнения в следующие эксперименты были включены различные контрольные конструкции (антитела против GDF8 и другие антагонисты GDF8). Разработаны следующие контрольные конструкции: контроль I: антитело человека против GDF8, содержащее вариабельные области тяжелой и легкой цепей, имеющие аминокислотные последовательности соответствующих доменов «Myo29» (т.е. SEQ ID NO:16 и 18), изложенное в патенте США № 7261893; контроль II: антитело человека против GDF8, содержащее вариабельные области тяжелой и легкой цепей, имеющие аминокислотные последовательности соответствующих доменов «2_112_K» (т.е. SEQ ID NO:118 и 120), изложенное в заявке на патент США № 2006/0263354; контроль III: слитая конструкция ActRIIB-Fc, имеющая аминокислотную последовательность SEQ ID NO: 391; и контроль IV: слитая конструкция вариант ActRIIB-Fc, идентичная контролю III за исключение того, что аланин в положении 64 SEQ ID NO:391 [A64] заменен аргинином [R64]. (Не все контрольные конструкции использовались в каждом примере).
Пример 3. Определение аффинности связывания с антигеном.
Равновесные константы диссоциации (значения KD) для связывания антигена с выбранными антителами были определены согласно кинетике поверхностных явлений, используя анализ с помощью поверхностного плазмонного резонанса в режиме реального времени с использованием биосенсора (BIACORETM 2000). Каждое выбранное антитело фиксировали либо на поверхности с козьими поликлональными антителами против IgG мыши, либо на поверхности с козьими поликлональными антителами против Fc человека (hFc) (Jackson Immuno Research Lab), созданной благодаря прямой химической связи с чипом BIACORETM для образования поверхности с фиксированными антителами. Гомодимер GDF8 человека, гомодимер hGDF11 или гомодимер hGDF5 в концентрации 25 нМ подавали с использованием инъекции по поверхностям с фиксированными антителами, и следили за связыванием антигена с антителом и их диссоциацией в режиме реального времени при комнатной температуре. Выполняли кинетический анализ для расчета KD, констант скоростей диссоциации (kd), констант скоростей ассоциации (ka) и полупериода диссоциации комплекса антиген/антитело (таблица 2).
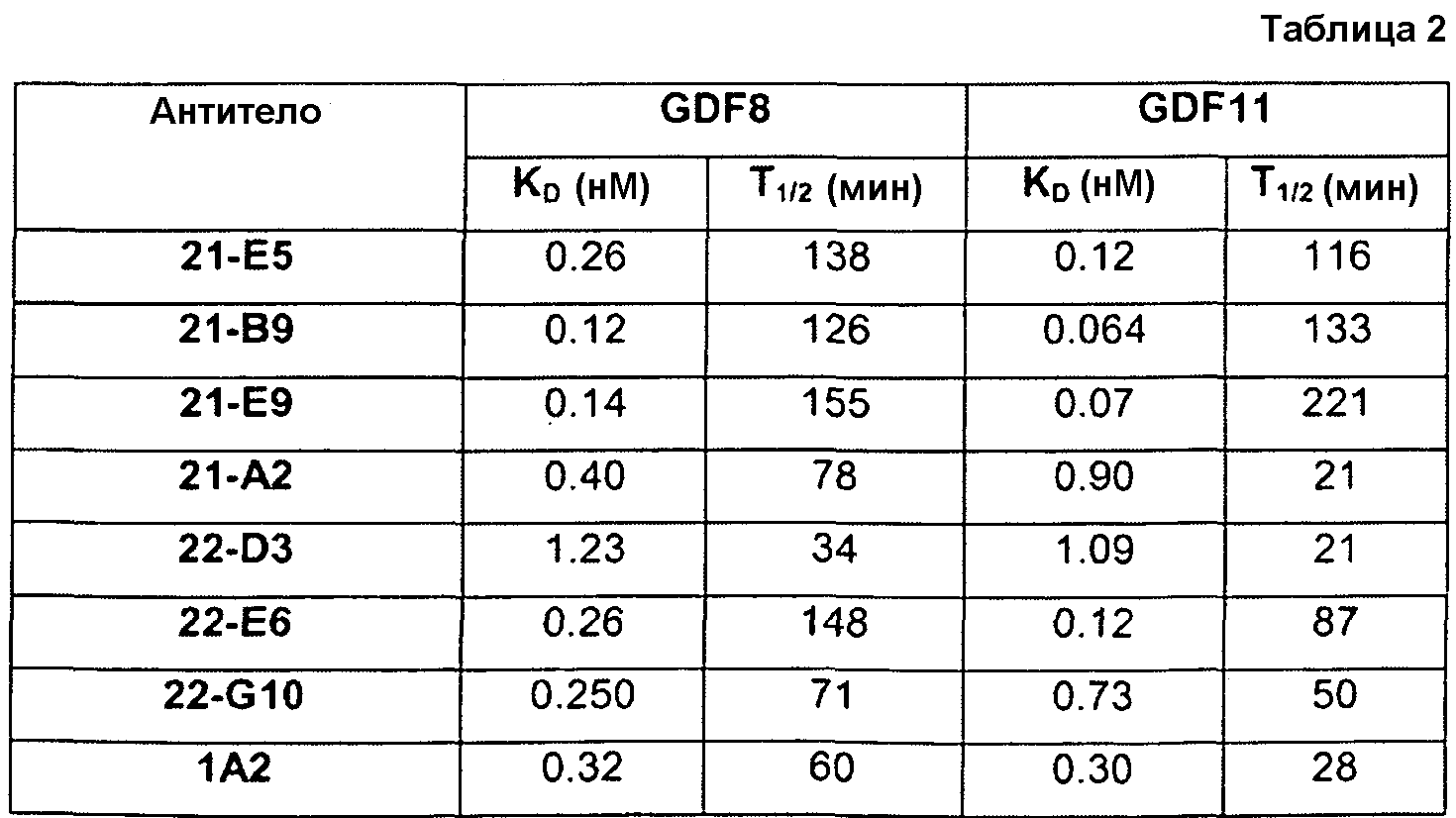
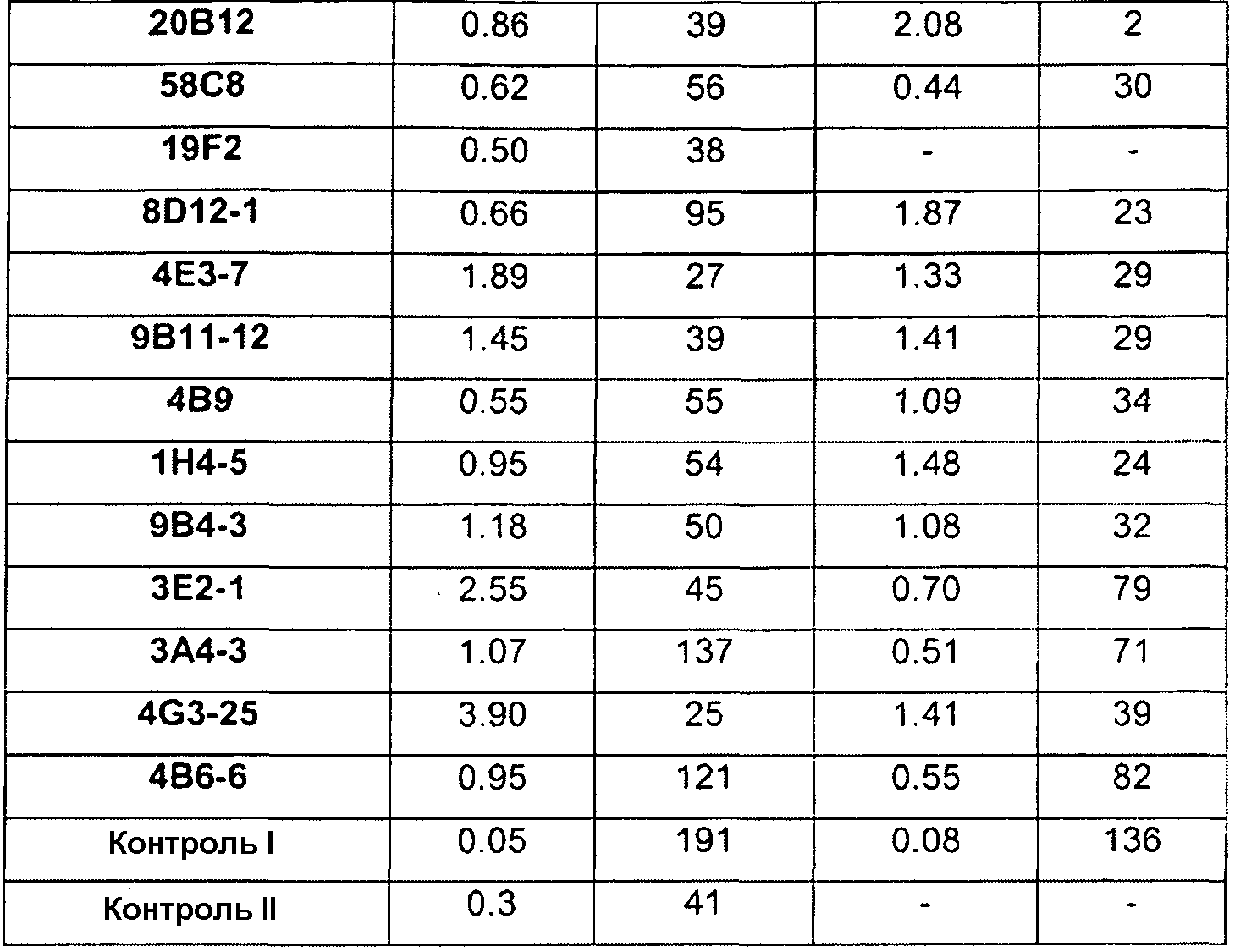
Вышеизложенный эксперимент также проводили с GDF8, наносимым по поверхности с фиксированными антителами - антителами-кандидатами H4H1669P или H4H1657N2. Предварительные данные выявили очень медленную скорость диссоциации для обоих антител, что означает KD, составляющую 1-2 пМ или менее.
Значения KD для связывания антигена с выбранными антителами также определяли, как описано выше, с использованием модифицированного подвижного буфера, который не содержит BSA.
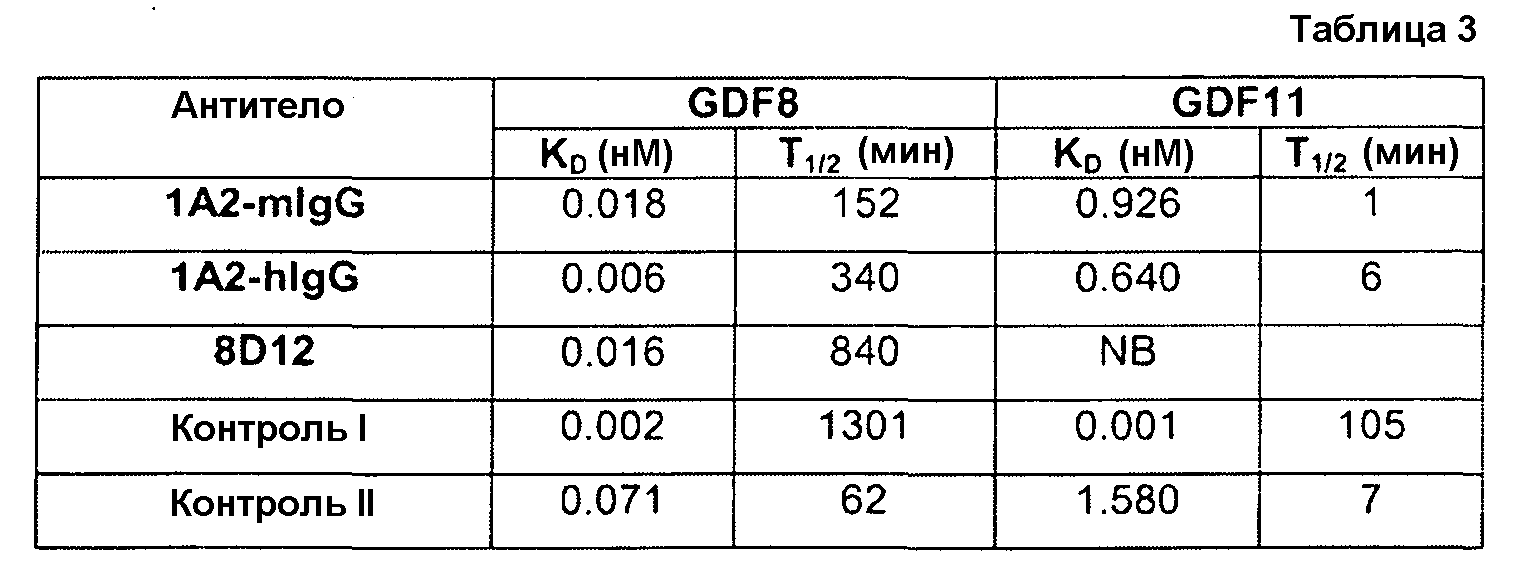
Были проведены дополнительные эксперименты по связыванию антигена, в которых GDF8 и GDF11 наносили по поверхности выбранных антител против GDF8 и контрольных антител при 25ºC и 37ºC. Равновесные константы диссоциации (значения KD) для связывания антигена с выбранными антителами были определены согласно кинетике поверхностных явлений, используя анализ с помощью поверхностного плазмонного резонанса в режиме реального времени с использованием биосенсора (BIACORETM T100). Каждое выбранное антитело и контроль фиксировали на поверхности с козьими поликлональными антителами против hFc (Jackson Immuno Research Lab, каталожный # 109-005-098), созданной благодаря прямой химической связи с чипом биосенсора BIACORETM CM5 для образования поверхности с фиксированными антителами. Различные концентрации (2,5-0,625 нМ, 2-кратные разведения) гомодимера hGDF8 или гомодимера hGDF11 (или в некоторых экспериментах, активина A) подавали с использованием инъекции по поверхностям с фиксированными антителами, и следили за связыванием антигена с антителом и их диссоциацией в режиме реального времени. Выполняли кинетический анализ для расчета KD, констант скоростей диссоциации (kd), констант скоростей ассоциации (ka) и полупериода диссоциации комплекса антиген/антитело. Результаты суммированы в таблице 4. (NB - не отмечаемое связывание).
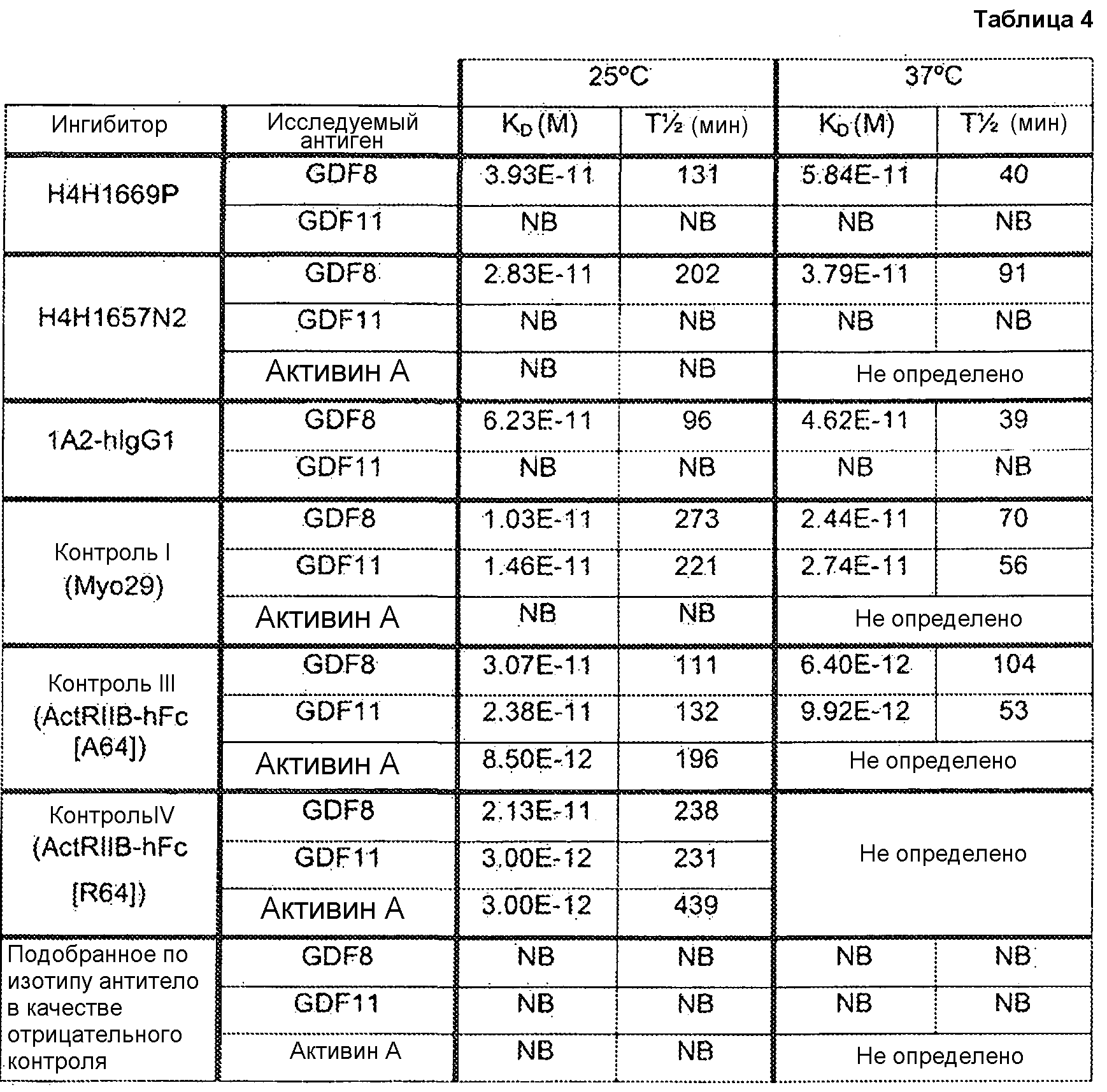
Как продемонстрировано выше, все антитела H4H1669P, H4H1657N2 и 1A2-hIgG1 по настоящему изобретению проявляли сильное связывание с GDF8, но не с GDF11. Напротив, контрольные молекулы продемонстрировали связывании и с GDF8, и с GDF11.
Пример 4. Блокирование антителами ответной реакции Smad2/люцифераза
Анализ GDF8-индуцируемой люциферазной активности: Был разработан биоанализ для определения способности выбранных антител против GDF8 к нейтрализации GDF8-опосредованной или GDF11-опосредованной клеточной функции in vitro, используя созданную линию клеток A204 (клеток рабдомиосаркомы человека, ATCC), которая содержит реагирующий на GDF8 или GDF11 промотор, запускающий экспрессию люциферазы. Ингибирование GDF8- или GDF11-индуцируемой люциферазной активности определяли следующим образом. Клетки засевали в 96-луночный планшет в количестве 2×104 клеток/лунку в средах и инкубировали в течение ночи при 37ºC, 5% CO2. Белок антитела (в последовательных разведениях, начиная с 25 нМ в средах для клеток) добавляли в лунки с клетками A204/Smad2 в трех повторах в двух планшетах; в каждую лунку добавляли GDF8 или GDF11 (0,8 нМ). Планшеты инкубировали при 37ºC, 5% CO2 в течение 6 часов. Люциферазную активность определяли посредством добавления субстрата BRIGHT-GLO® (Promega), и определяли значения IC50 (таблица 5).
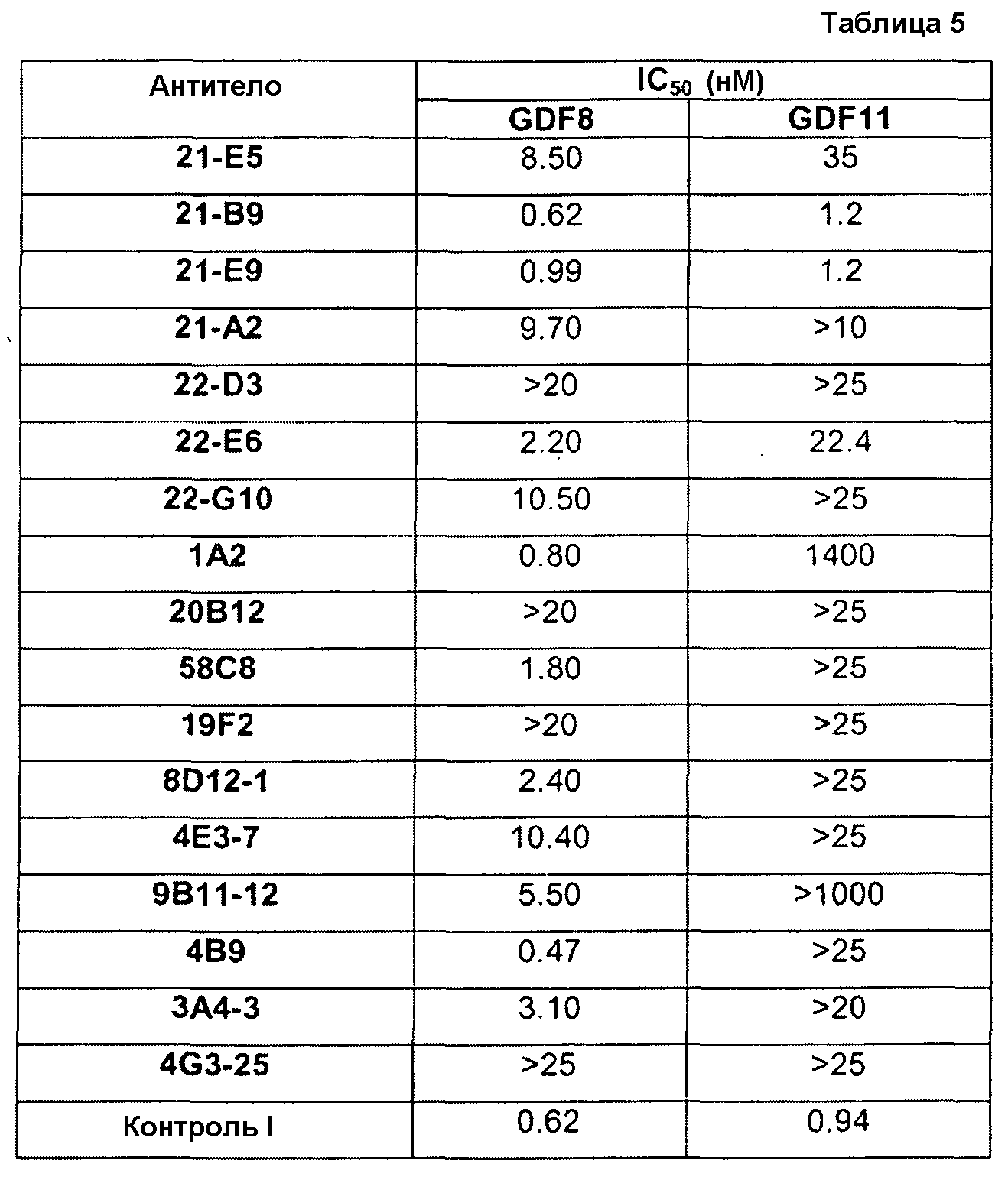
Способность выбранных антител против GDF8 к нейтрализации GDF8-опосредованной или GDF11-опредованной клеточной функции, кроме того, исследовали, как описано выше, с использованием различных концентраций GDF8 или GDF11 (таблица 6). (nd = не определено; NB = нет связывания).
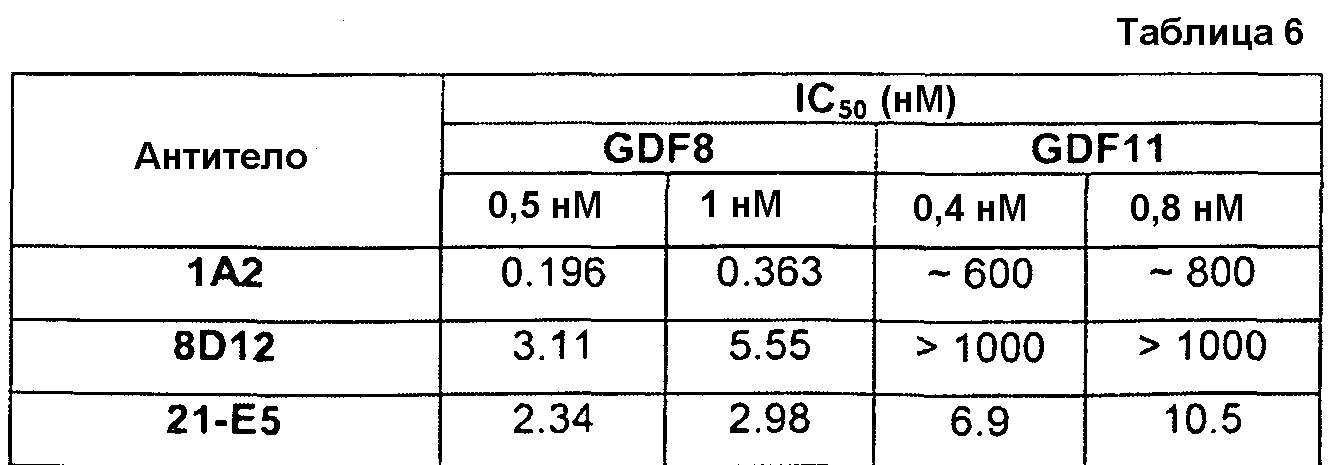

Описанный выше биоанализ повторяли, используя химерную конструкцию GDF8/TGFβ-1 в качестве активирующего пептида. В частности, в этом эксперименте была использована химера, состоящая из зрелого белка GDF8, в котором аминокислоты 48-72 были заменены соответствующими аминокислотами TGFβ-1 (SEQ ID NO:352, также называемая в настоящем описании «GDF8/TGFβ[48-72]»). Эта конструкция было создана, исходя из экспрессионной конструкции, кодирующей полноразмерный предшественник GDF8 человека с последовательностью TGFβ-1 человека, замещающей соответствующую последовательность GDF8. Биоактивность исследовали в кондиционированной среде, полученной в результате транзиторной трансфекции конструкции GDF8/TGFβ[48-72] в клетки CHO. Экспрессию и процессирование оценивали с помощью Вестерн-блоттинга. Кондиционированную среду концентрировали в 20 раз, нагревали до 80ºC в течение 5 минут для инактивации связанного пропептида GDF8, и активность в ней оценивали при последовательном разведении в биоанализе. Хотя точная концентрация химерного белка не была определена в этих экспериментах, характерная концентрация находилась в диапазоне 1-10 мкг/мл до концентрирования. Как описано выше, клетки, содержащие реагирующий на GDF8 промотор, запускающий экспрессию люциферазы, засевали в 96-луночный планшет. Выбранные антитела (в последовательных разведениях, начиная с 100 нМ, в средах для клеток) добавляли к количеству кондиционированной среды с химерным белком GDF8/TGFβ[48-72], которое, как установлено, дает максимальную ответную реакцию. Эту смесь предварительно инкубировали в течение 45 минут и добавляли к клеткам-репортерам. Определяли люциферазную активность, и рассчитывали приблизительные значения IC50, представленные в таблице 7 ниже. (NB = нет блокирования).
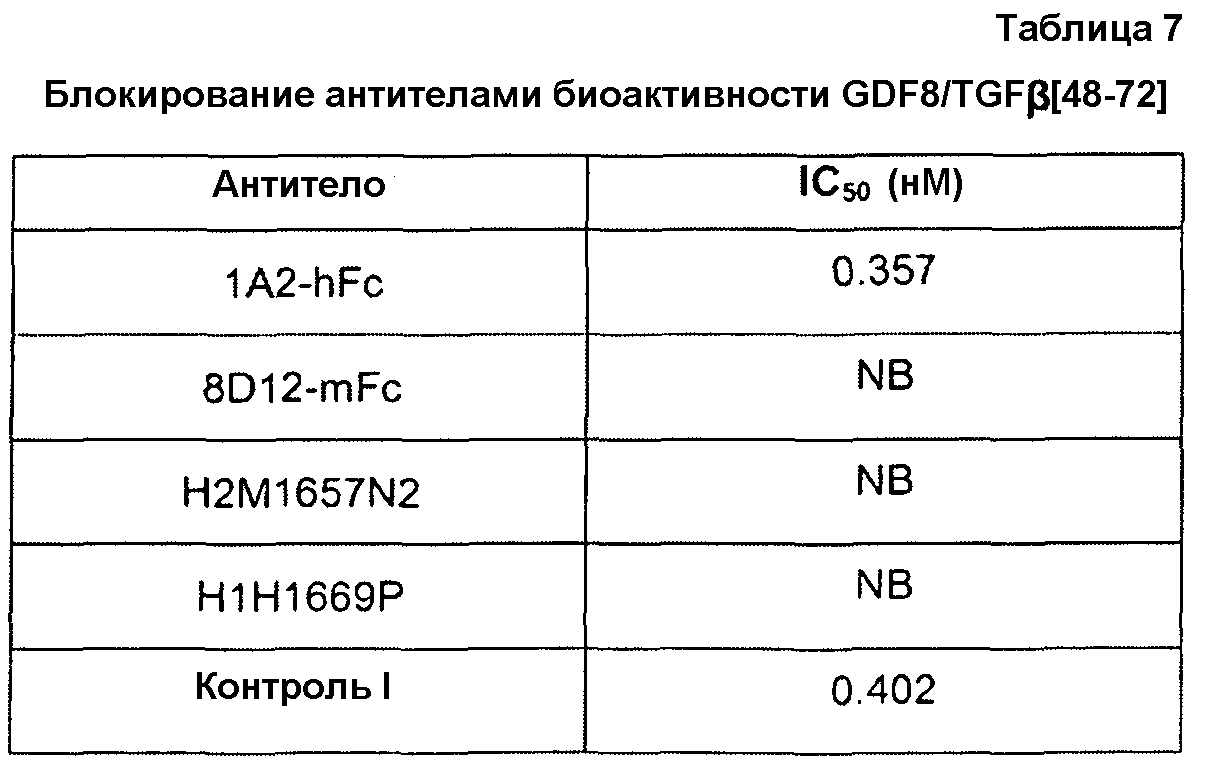
Химерная конструкция GDF8/TGFβ[48-72] была способна активировать экспрессию люциферазы в этом анализе, а 1A2-hFc и контроль I были способны блокировать биоактивность этой конструкции. Однако антитела H2M1657N2 и H1H1669P не блокировали биоактивность химерной конструкции GDF8/TGFβ[48-72]. Поскольку было установлено, что эти два антитела блокируют биоактивность GDF8 дикого типа в этой системе анализа (см. таблицу 6), можно сделать вывод, что H2M1657N2 и H1H1669P по всей вероятности проявляют свои биологические эффекты в результате взаимодействия с эпитопом, находящимся в пределах аминокислот 48-72 GDF8.
Пример 5. Иммуноблоттинг фрагментов hGDF8, образованных в результате расщепления протеиназой K.
Анализ с помощью Вестерн-блоттинга использовали для определения иммунореактивности исследуемых и контрольных mAb с GDF8 человека, подвергнутого протеолитическому расщеплению протеиназой K. Ферментативные реакции, содержащие 1 мкг GDF8 человека и 0, 1 или 6 мкг протеиназы K, инкубировали в течение либо 10, либо 45 мин в буфере для расщепления. Равные аликвоты, содержащие 20% от количества GDF8, присутствующего в реакционной смеси, (200 нг) вносили в 3 отдельных 18% SDS-ПААГ для электрофореза в неденатурирующих условиях и подвергали переносу под действием электрического тока на мембраны PVDF. Каждую мембрану инкубировали с первым антителом в концентрации 2 мкг/мл, а затем с соответствующим вторым антителом, конъюгированным с HRP. Результирующая иммунореактивность для mAb в виде контроля I, mAb 1A2 и mAb 21-E9, соответственно, представлена на фиг. 1А-С. Результаты указывают на утрату реактивности с GDF8 в случае контрольного mAb (A) на дорожках 3 и 6. В очевидном отличии от этого, антитело 1A2 сохраняет иммунореактивность с более маленьким фрагментом GDF8 с молекулярной массой, составляющей приблизительно 17-19 кДа (фиг. 1B).
Эксперимент повторяли с использованием периодов времени расщепления вплоть до 10 мин, 1 ч или 16 ч, в присутствии 0 или 4 мкг GDF8 и 0, 24 или 96 мкг протеиназы K. Результаты (фиг. 2A-B) показывают, что в отсутствие протеиназы K как контрольное mAb, так и mAb 1A2 обладали иммунореактивностью с полноразмерным зрелым (нерасщепленным) белком GDF8 (смотрите фиг. 2A-B, дорожки 2 и 7). В присутствии протеиназы K иммунореактивность утрачивается в случае mAb в виде контроля I во все моменты времени. В очевидном отличии от этого, mAb 1A2 оставалось иммунореактивным с фрагментом подвергнутого расщеплению GDF8 человека (фиг. 2B, дорожки 3-6 и 8). Эти результаты указывают на то, что mAb 1A2 обладает иммунореактивностью с более маленьким фрагментом GDF8, который остается интактным в присутствии протеиназы K, тогда как контрольное mAb утрачивает иммунореактивность с более маленьким фрагментом (17-19 кДа).
Модифицированный анализ с помощью Вестерн-блоттинга использовали, чтобы далее определить эпитоп hGDF8 для выбранных антител против hGDF8. Модификация, состоящая в том, что до инкубации специфического для hGDF8 первого антитела с мембраной каждое антитело против hGDF8 предварительно инкубировали с 1000-кратным или 50-кратным молярным избытком пептидных фрагментов hGDF8, состоящих из аминокислот 1-14, 17-42, 48-72 или 75-105. Результаты указывают на то, что предварительная инкубация пептидного фрагмента из аминокислот 48-72 в 50-кратном молярном избыточном количестве была способна блокировать связывание антитела 8D12 с hGDF8.
Пример 6. Связывание антител с химерами GDF8
Было создано двенадцать химерных пробелков GDF8. В таблице 8 представлены структуры зрелых химерных белков GDF8. Химерные белки GDF8 включали две совокупности: одна совокупность, имеющая различные замены последовательностей GDF8 последовательностями BMP2, другая совокупность, имеющая различные замены последовательностей GDF8 последовательностями TGFβ1. Эти химерные белки использовали для проверки связывания с антителами и определения места такого связывания.

Различные химерные пробелки GDF8 транзиторно трансфицировали в созданную стабильную линию клеток CHO.hFurin. Анализ с помощью Вестерн-блоттинга, схожий с описанным выше анализом, использовали для выявления связывания различных антител против hGDF8 с каждым из химерных белков GDF8. Вкратце, 10 мкг супернатанта культуры клеток CHO вносили на каждую дорожку SDS-ПААГ (для электрофореза в неденатурирующих или денатурирующих условиях) и подвергали переносу под действием электрического тока на мембрану PVDF. Затем мембрану инкубировали с антителом против GDF8 в концентрации 2 мкг/мл с последующим подверганием воздействию соответствующего второго антитела, конъюгированного с HRP. Как показано в таблице 9, антитело 8D12 было не способно связываться ни с B48, ни с T48. Этот результат означал, что аминокислоты 48-72 зрелого GDF8 участвуют в связывании антитела 8D12 с GDF8.
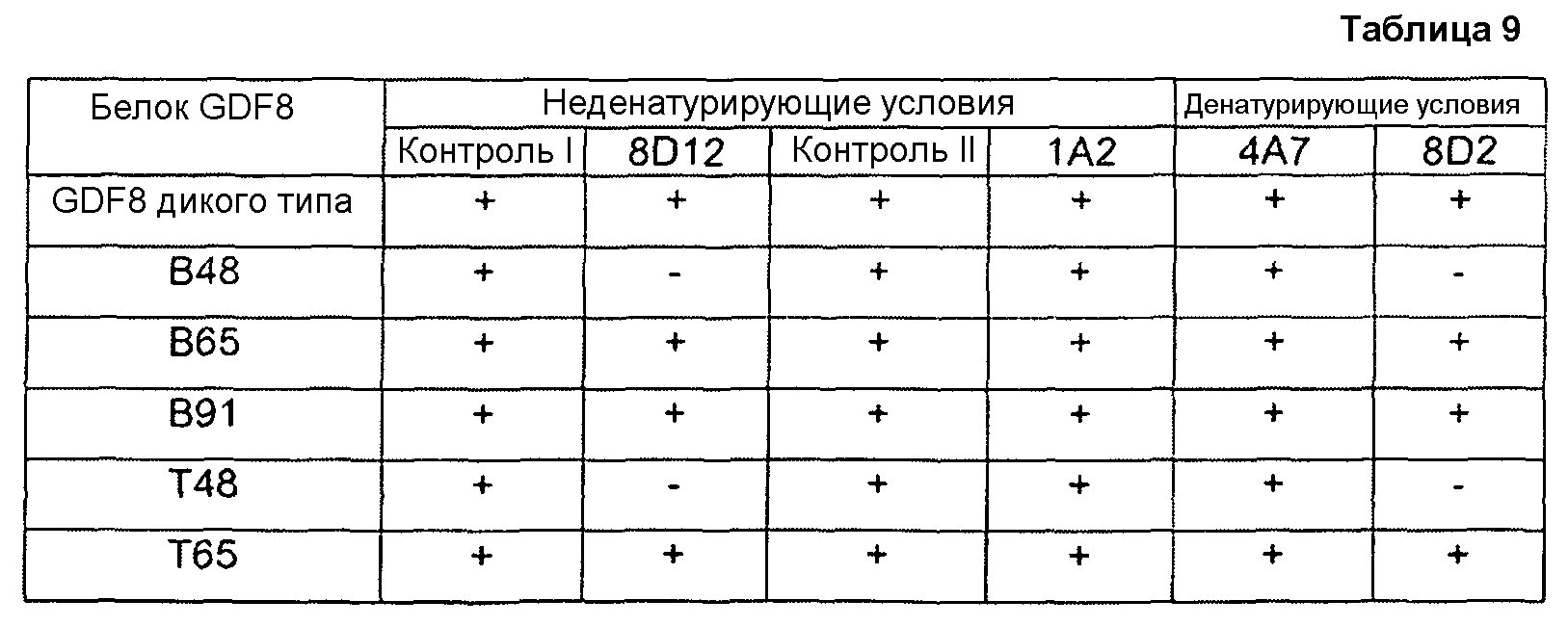
Пример 7. Связывание антител с пептидами hGDF8
Четырнадцать пептидов (таблица 10) было образовано из зрелого белка hGDF8 (SEQ ID NO: 340). Немодифицированные пептиды, биотилированные с N-конца пептиды (N-term) или биотилированные с C-конца пептиды (C-term) были использованы для проверки связывания с антителами и определения места такого связывания. Каждый из полноразмерного hGDF8, hGDF11 и немодифицированных пептидов по отдельности связывали через аминогруппу с микросферическими частицами (или сферами) xMAP® Multi-Analyte COOH. Каждый из биотилированных пептидов связывали с микросферическими частицами xMAP® Multi-Analyte LumAvidin. Суспензию связанных с пептидами сфер затем смешивали с равным объемом буфера для блокирования (PBS, 1% BSA, 0,05% Tween 20, 0,05% азида натрия), а затем распределяли по 96-луночному планшету с фильтрами (Millipore, MULTISCREEN® BV). К суспензии связанных с пептидами сфер затем добавляли контрольные и исследуемые антитела против hGDF8, в концентрации 2,5 мкг/мл, и допускали их связывание со сферами при комнатной температуре в течение ночи. Подвергнутые инкубации с антителами сферы затем отмывали дважды PBST (PBS + 0,05% Tween 20) и инкубировали либо с конъюгированным с фикоэритрином (PE) антителом против hFC, либо с PE-конъюгированным антителом против mFC при комнатной температуре в течение 45 мин. Сферы снова отмывали, и сигналы, указывающие на связывание антител с различными пептидами, выявляли с использованием инструментальных средств LUMINEX® 100TM или 200TM. Как показано в таблице 10, антитело против hGDF8 - 8D12 способно к связыванию с пептидами 4, 5, 6, 7, 8 и 9. Напротив, антитело против hGDF8 - H4H1657N2 не связывалось с каким-либо из пептидов (не представленные данные).

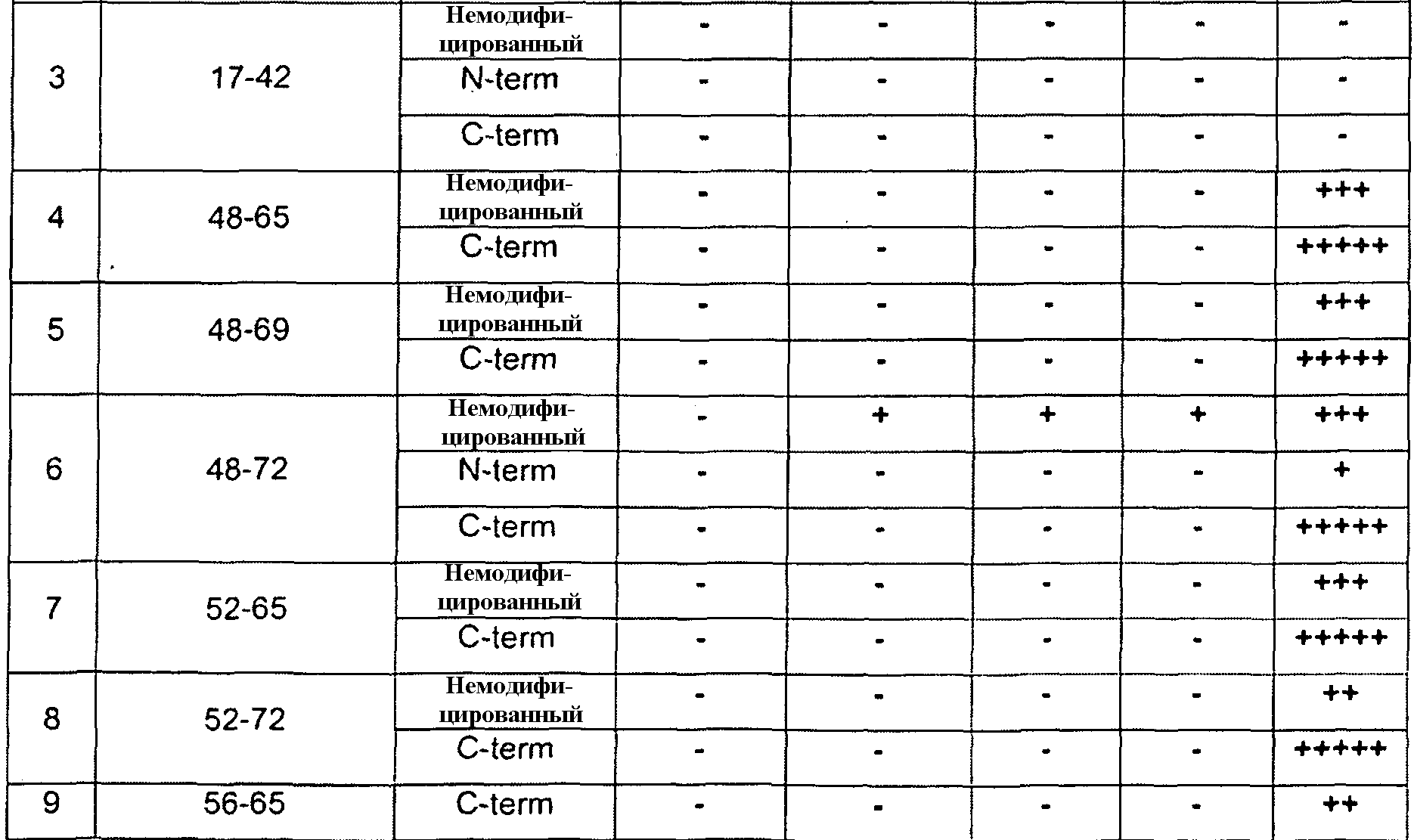
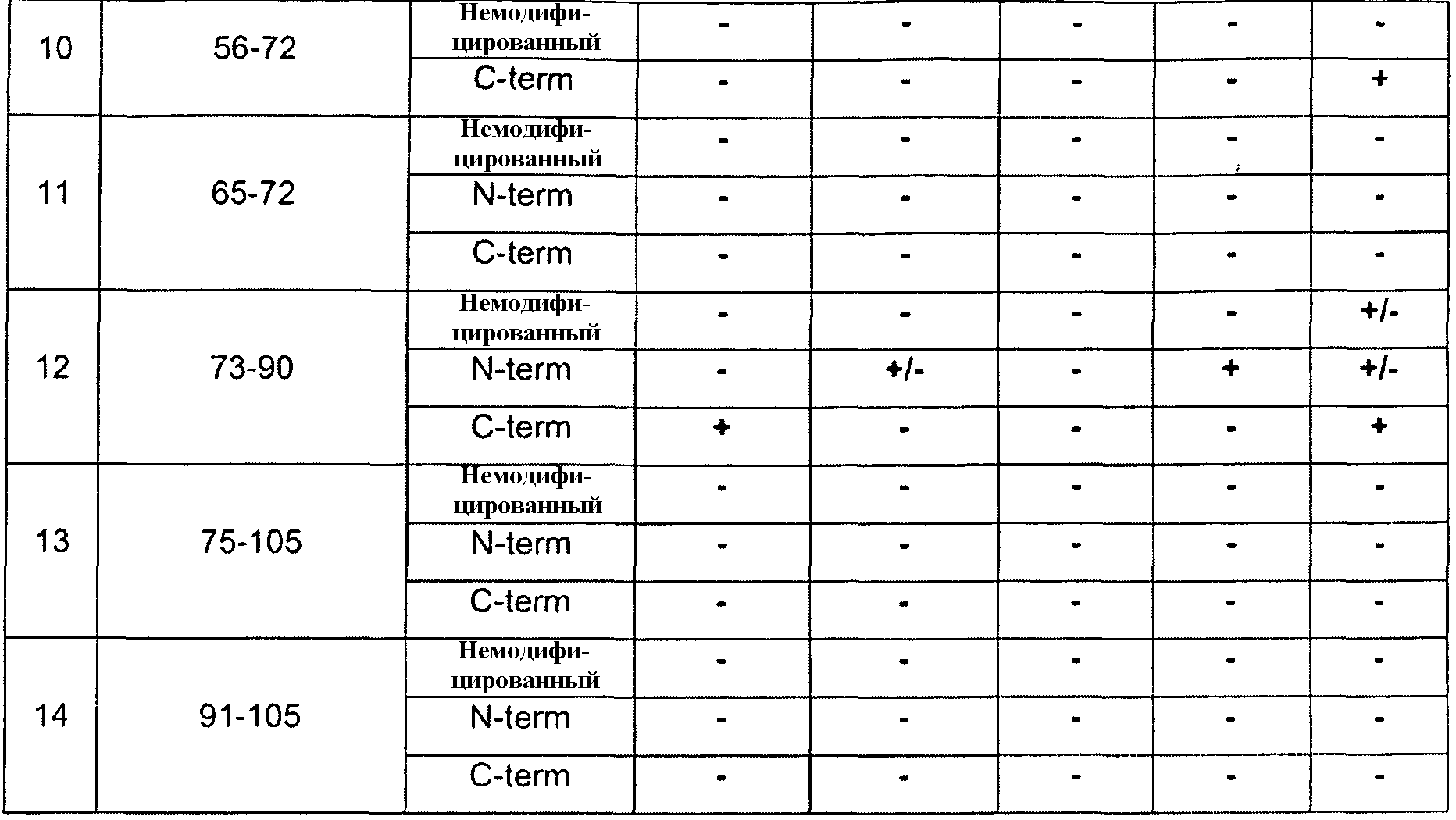
Пример 8. Эффект антител человека против GDF8 на связывание GDF8 с активином RIIB
Сначала зрелый белок hGDF8 связывали через аминогруппу со сферами Luminex®. Затем покрытые hGDF8 сферы Luminex® инкубировали с различными антителами против hGDF8, в концентрации 1,25 мкг/мл, в течение 2 ч при комнатной температуре. Затем к смеси сферы-антитело добавляли активин человека RIIB-mFc, и осуществляли инкубацию в течение дополнительных 2 ч при комнатной температуре. Затем сферы отмывали и окрашивали конъюгированным с R-фикоэритрином (R-PE) поликлональным антителом против mFc, и определяли среднюю интенсивность флуоресценции (MFI). Как показано в таблице 11, хотя и контрольное mAb I, и антитело 21-E5 были способны к блокированию связывания между hGDF8 и активином человека RIIB, контрольное mAb II было способно лишь к частичному блокированию связывания. Антитело 1A2 не было способно к блокированию связывания hGDF8 со своим рецептором, активином RIIB (таблица 11, n=3).

Пример 9. Варианты антитела 8D12
Варианты антитела 8D12 с модифицированной LCVR были созданы посредством модифицирования одной или более из следующих аминокислот LCVR 8D12: A7S или T, A8P, 9PL, S18P, V19A, M21I, K27Q, F41Y, V42L, R44K, R55L или T, M56G или L, N58Y, L59R, A75D, R79K, A105G, L109V и L111I.
Аффинность связывания (KD) вариантов антител по отношению к hGDF8 определяли, используя анализ с помощью поверхностного плазмонного резонанса в режиме реального времени с использованием биосенсора (BIACORETM 3000), как описано выше, в модифицированном подвижном буфере, который не содержит BSA (таблица 12).

Также проверяли связывание между вариантами антитела и пептидами hGDF8, как описано выше в примере 7. Варианты антитела 8D12-v2 и 8D12-v3 продемонстрировали сильное связывание с биотинилированными с C-конца пептидами 4, 5, 7 и 10.
Пример 10. Эффект антител против GDF8 на массу скелетной мускулатуры
Эффективность выбранных mAb против GDF8 в отношении вызова гипертрофии скелетной мускулатуры была определена in vivo. Коротко, 20 самцов мышей CB17 SCID, возрастом приблизительно 9 недель, делили поровну в соответствии с весом тела на 4 группы. Выбранное mAb (контроль I, 1A2, 21-E5, 8D12, 1A2-hIgG или контроль II) вводили в трех увеличивающихся дозах, составляющих 2,5 мг/кг/дозу, 5 мг/кг/дозу и 10 мг/кг/дозу. В качестве отрицательного контроля использовали Fc-фрагмент IgG человека. Антитела вводили внутрибрюшинно дважды в течение первой недели и один раз в неделю в течение последующих трех недель. В день 28 мышей подвергали эвтаназии и взвешивали, и передние большеберцовые (TA) мышцы, икроножные (GA) мышцы и четырехглавые (Quad) мышцы, сердце, селезенку и почку иссекали и взвешивали. Ткани приводили к исходному весу тела, и рассчитывали изменение веса в процентах по сравнению с отрицательным контролем. Были повторены шесть отдельных экспериментов с использованием антител: контроля I, 1A2, 21-E5, 8D12, 1A2-hIgG и контроля II (таблицы 13-18). Кроме того, был также повторен эксперимент с использованием более высоких увеличивающихся доз антитела в виде контроля I, составляющих 10 мг/кг/дозу, 30 мг/кг/дозу и 50 мг/кг/дозу (таблица 19). Результаты представляли в виде увеличения в процентах по сравнению с отрицательным контролем ± среднеквадратическое отклонение.
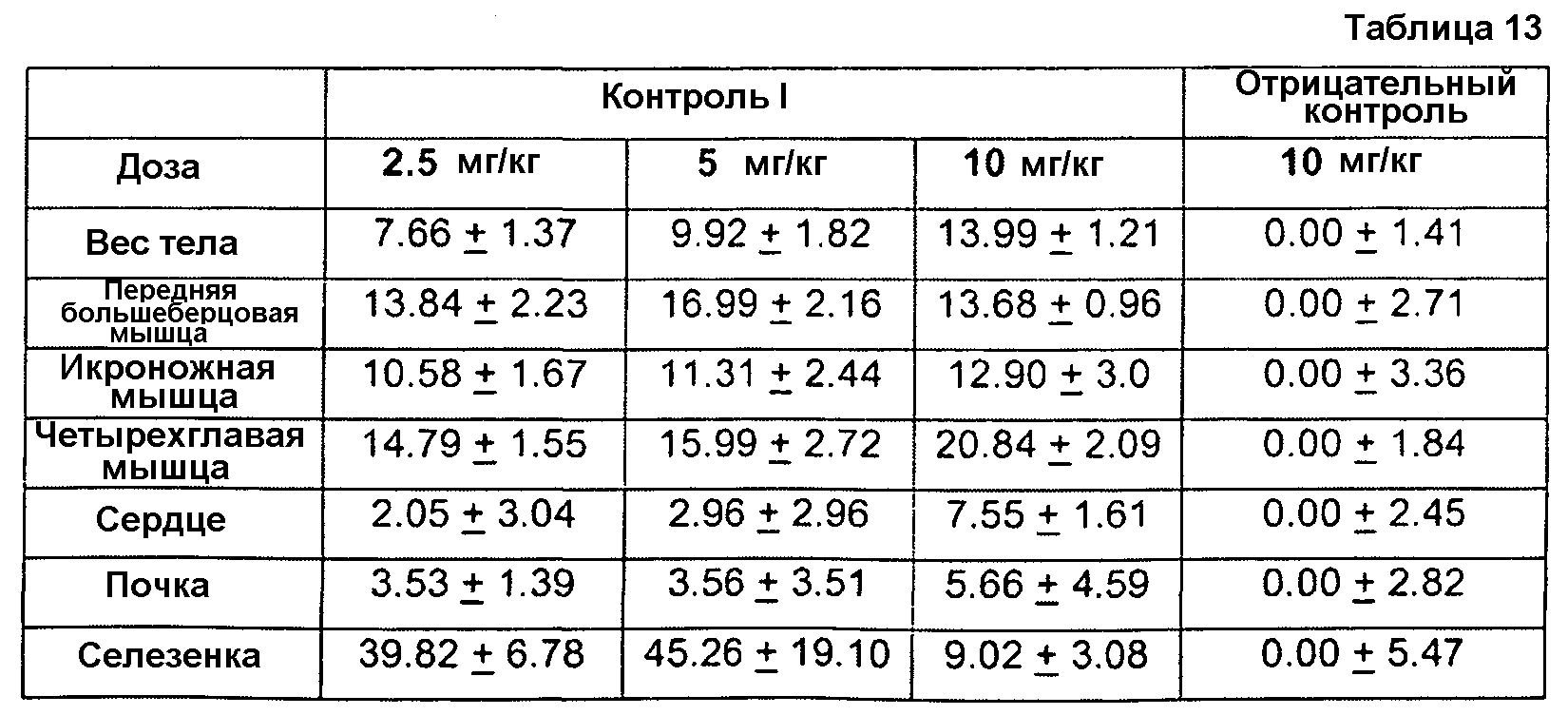
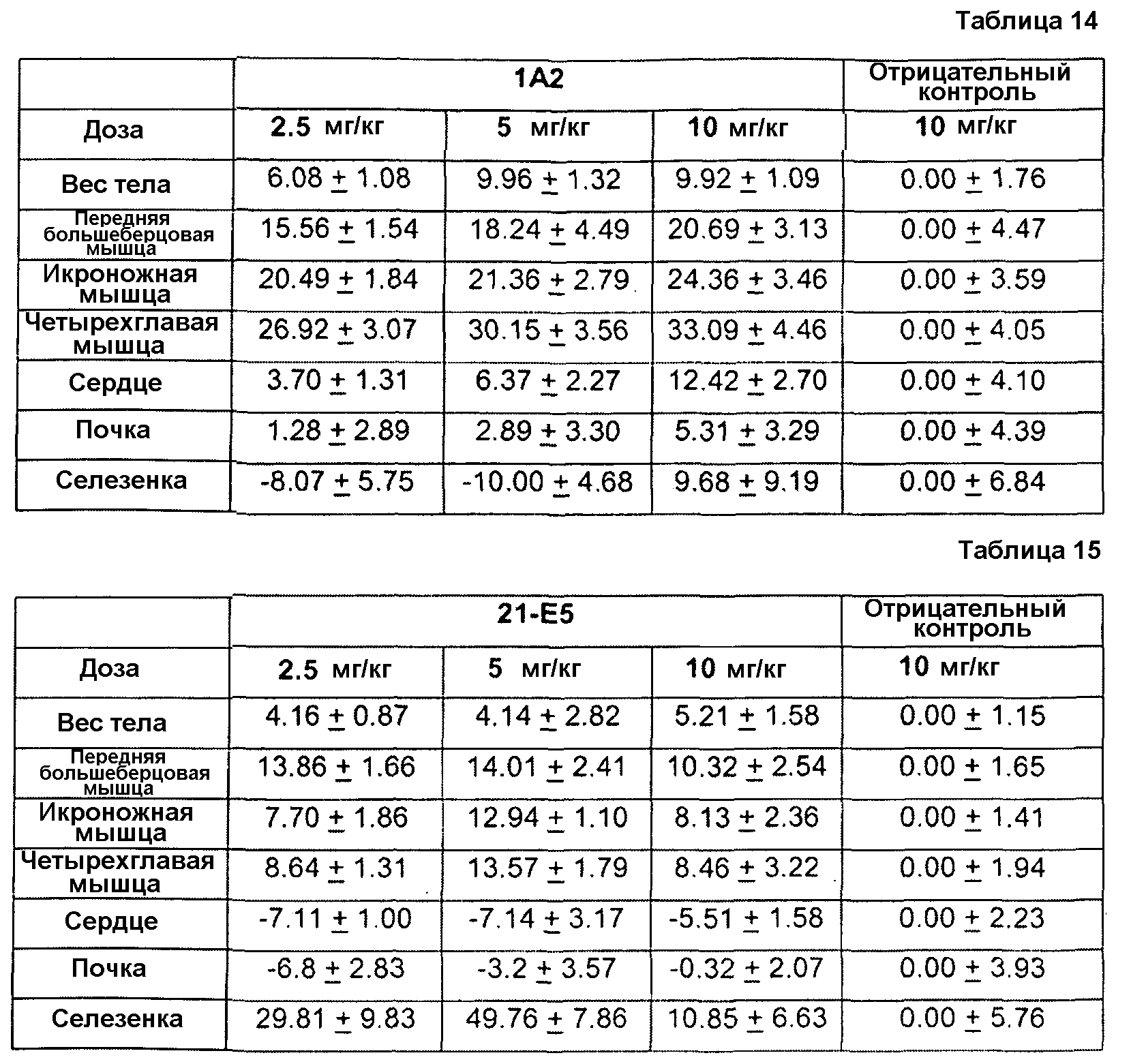
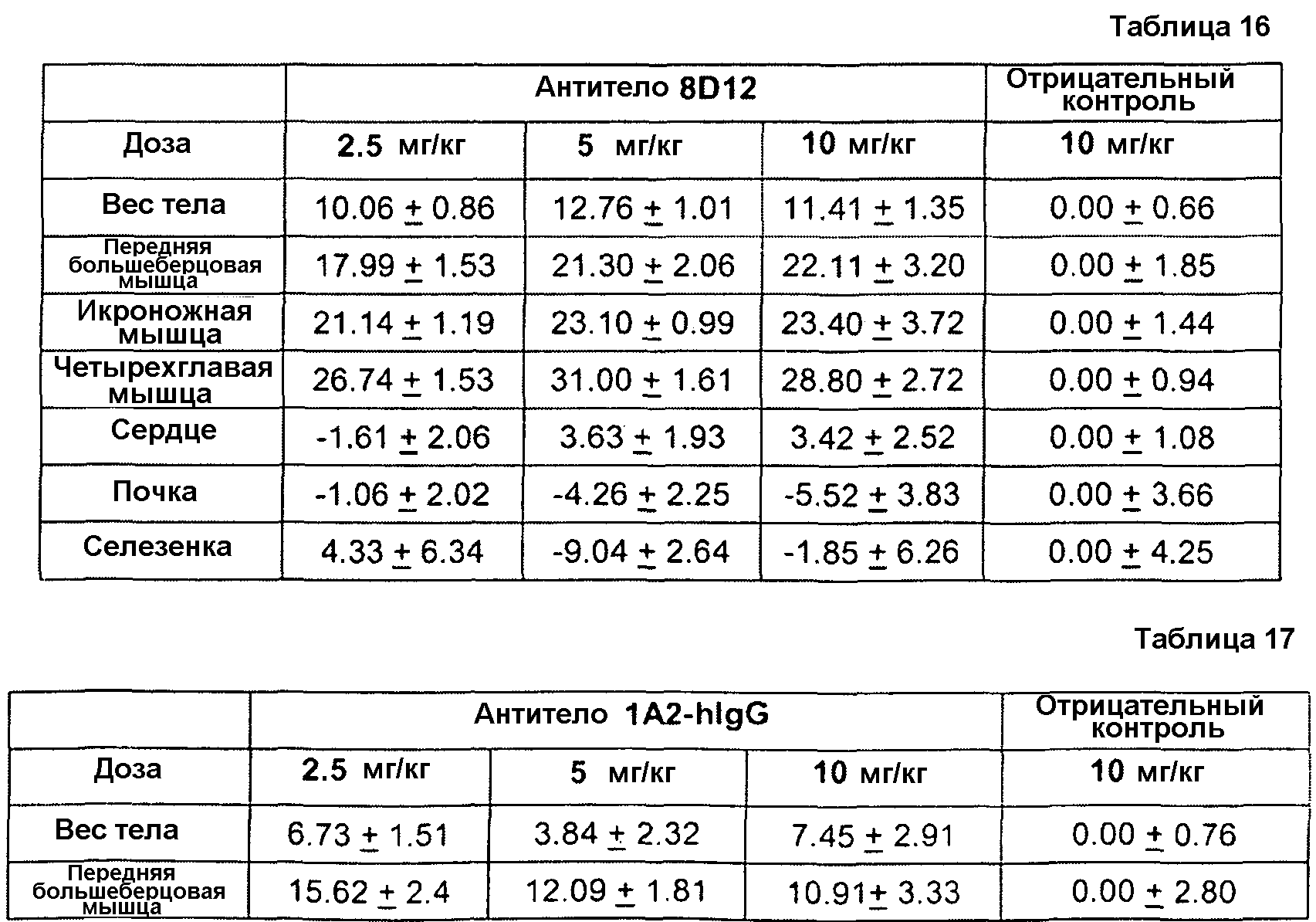
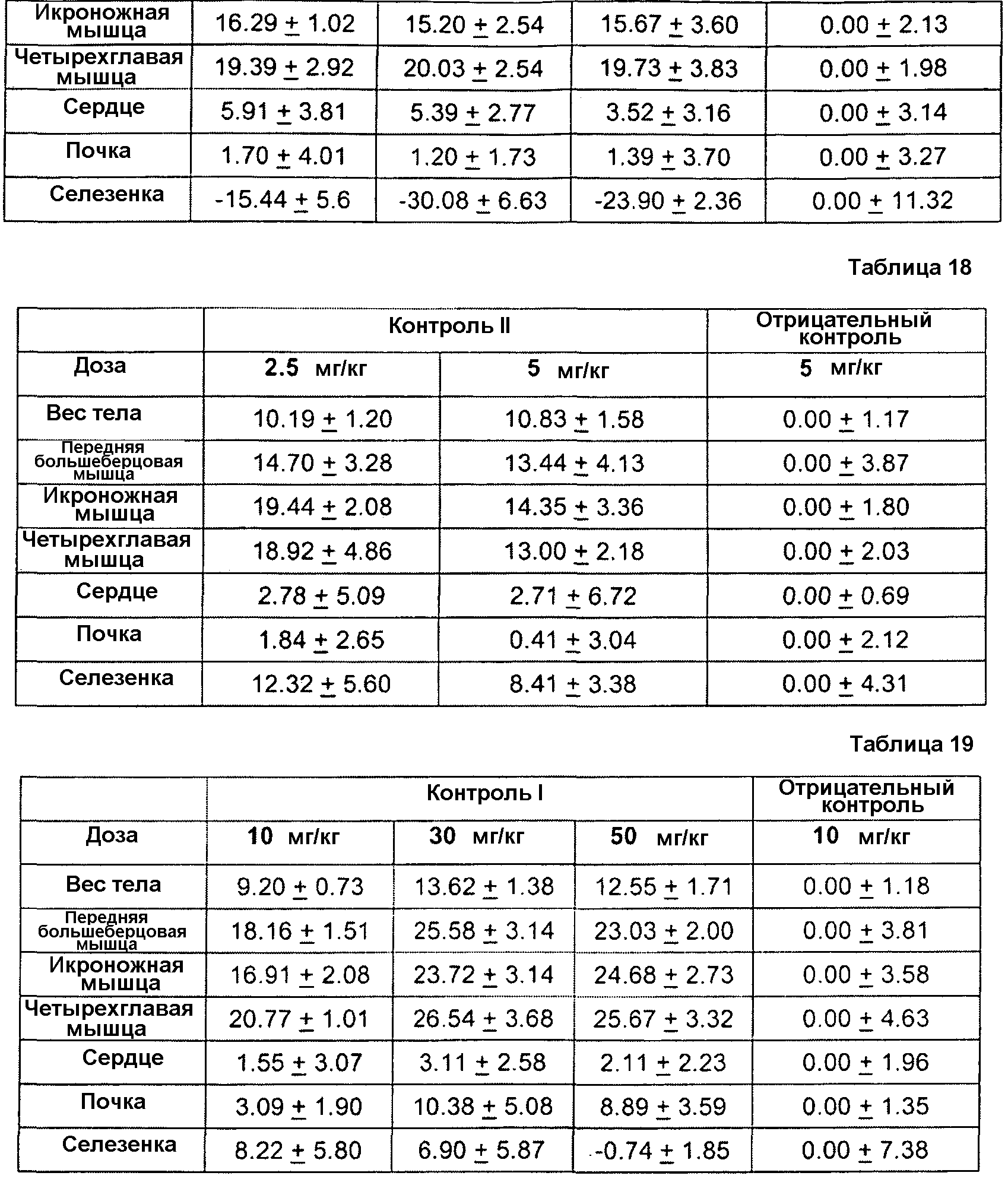
Схожий эксперимент был проведен, используя антитела H4H1657N2 и H4H1669P и контроли, вводимые мышам SCID. В частности, самцам мышей SCID в возрасте 10 недель вводили подкожно антитело в дозе 10 мг/кг в соответствии со следующей схемой введения доз: 2 раза на недели 1 и 1 раз в неделю на неделях 2 и 3. Общий период времени лечения составлял 28 дней. В случае этого эксперимента 5 мышам вводили подобранное по изотипу антитело в качестве отрицательного контроля; 5 мышам вводили контроль I (Myo29); 6 мышам вводили H4H657N2; и 6 мышам вводили H4H1669P. Результаты суммированы в таблице 20 и представлены в виде увеличения в процентах по сравнению с отрицательным контролем ± среднеквадратическое отклонение.
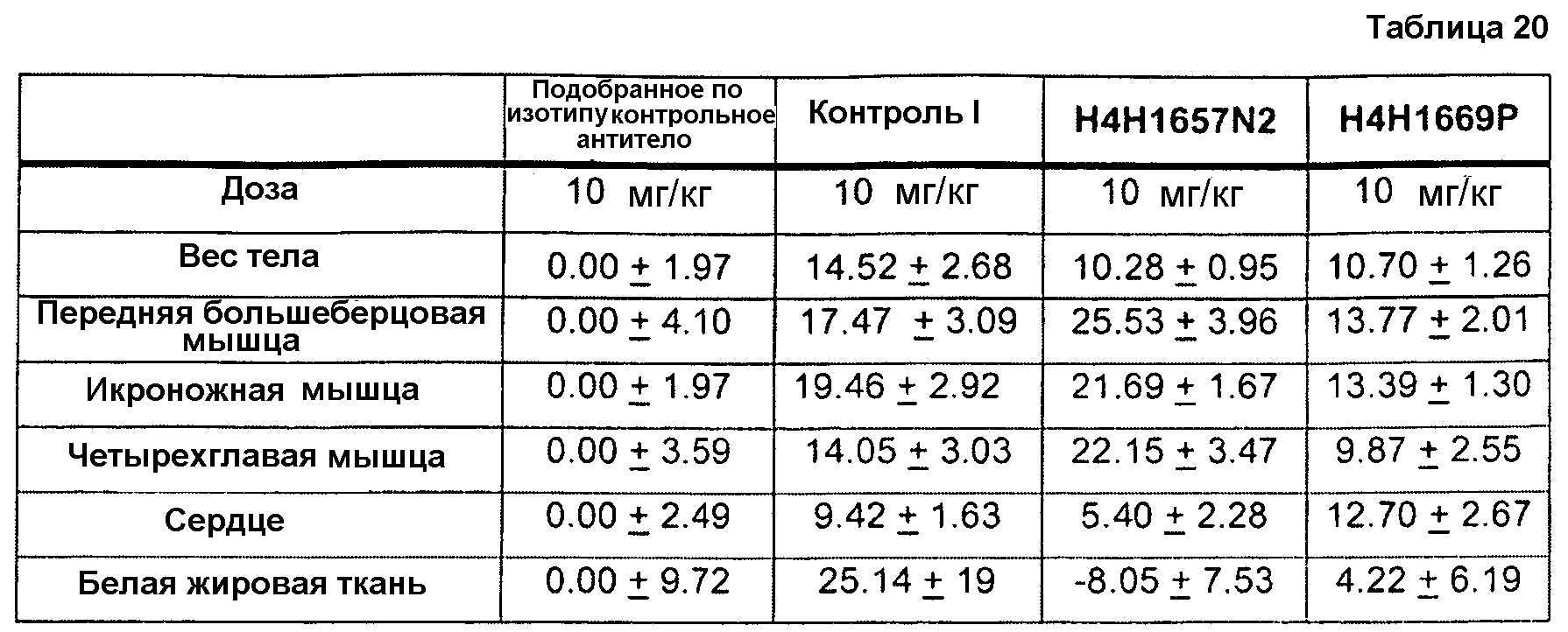
Схожий эксперимент был проведен, используя антитела H4H1657N2 и H4H1669P и контроли, вводимые мышам C57. В частности, самцам мышей C57 в возрасте 10 недель вводили подкожно антитело в дозе 10 мг/кг в соответствии со следующей схемой введения доз: 2 раза в неделю в течение двух недель. В случае этого эксперимента 5 мышам вводили подобранное по изотипу антитело в качестве отрицательного контроля; 5 мышам вводили контроль I (Myo29); 5 мышам вводили H4H1657N2; и 6 мышам вводили H4H1669P. Результаты суммированы в таблице 21 и представлены в виде увеличения в процентах по сравнению с отрицательным контролем ± среднеквадратическое отклонение.
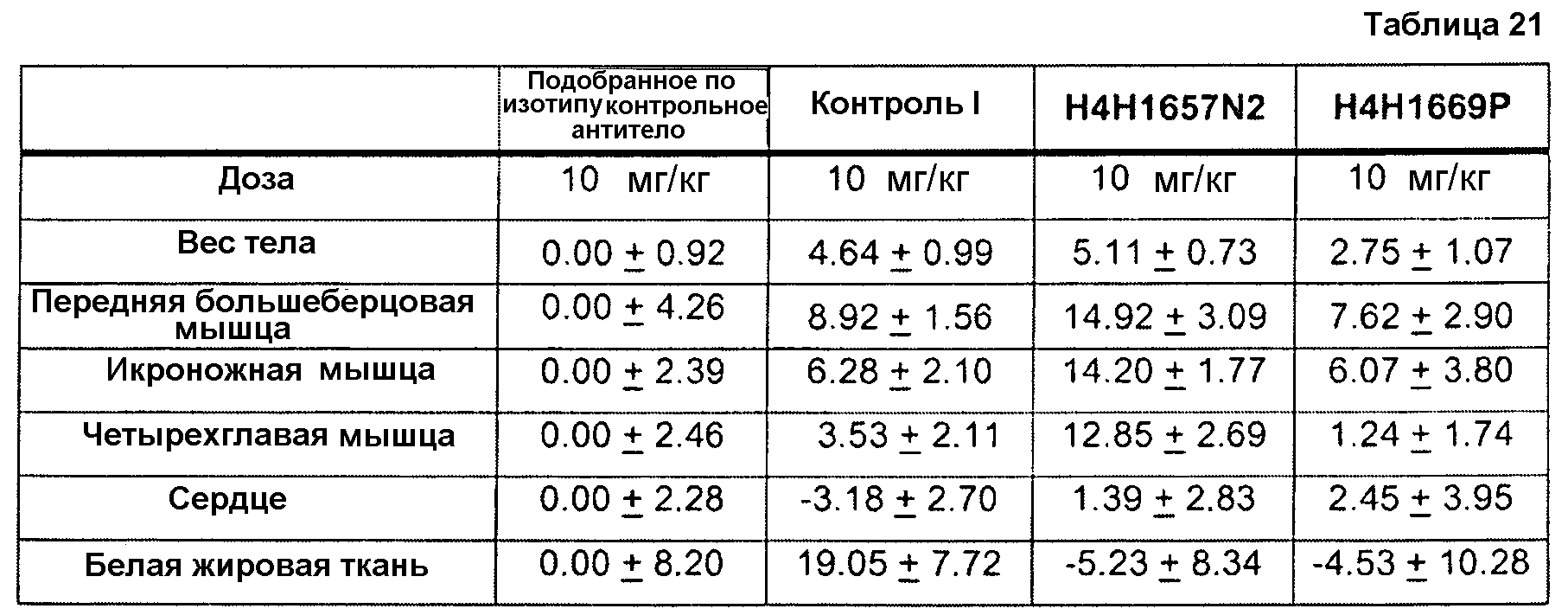
Затем были проведены эксперименты для выявления зависимости ответной реакции от величины дозы, используя антитела H4H1657N2 и H4H1669P, на мышах SCID. В частности, самцам мышей SCID в возрасте 10 недель вводили подкожно контрольные антитела в дозе 30 мг/кг и H4H1657N2 или H4H1669P в дозе, составляющей 2,5, 10 или 30 мг/кг, в соответствии со следующей схемой введения доз: 2 раза в неделю на недели 1 и 1 раз в неделю после этого. Результаты суммированы в таблицах 22 и 23 и представлены в виде увеличения в процентах по сравнению с отрицательным контролем ± среднеквадратическое отклонение.
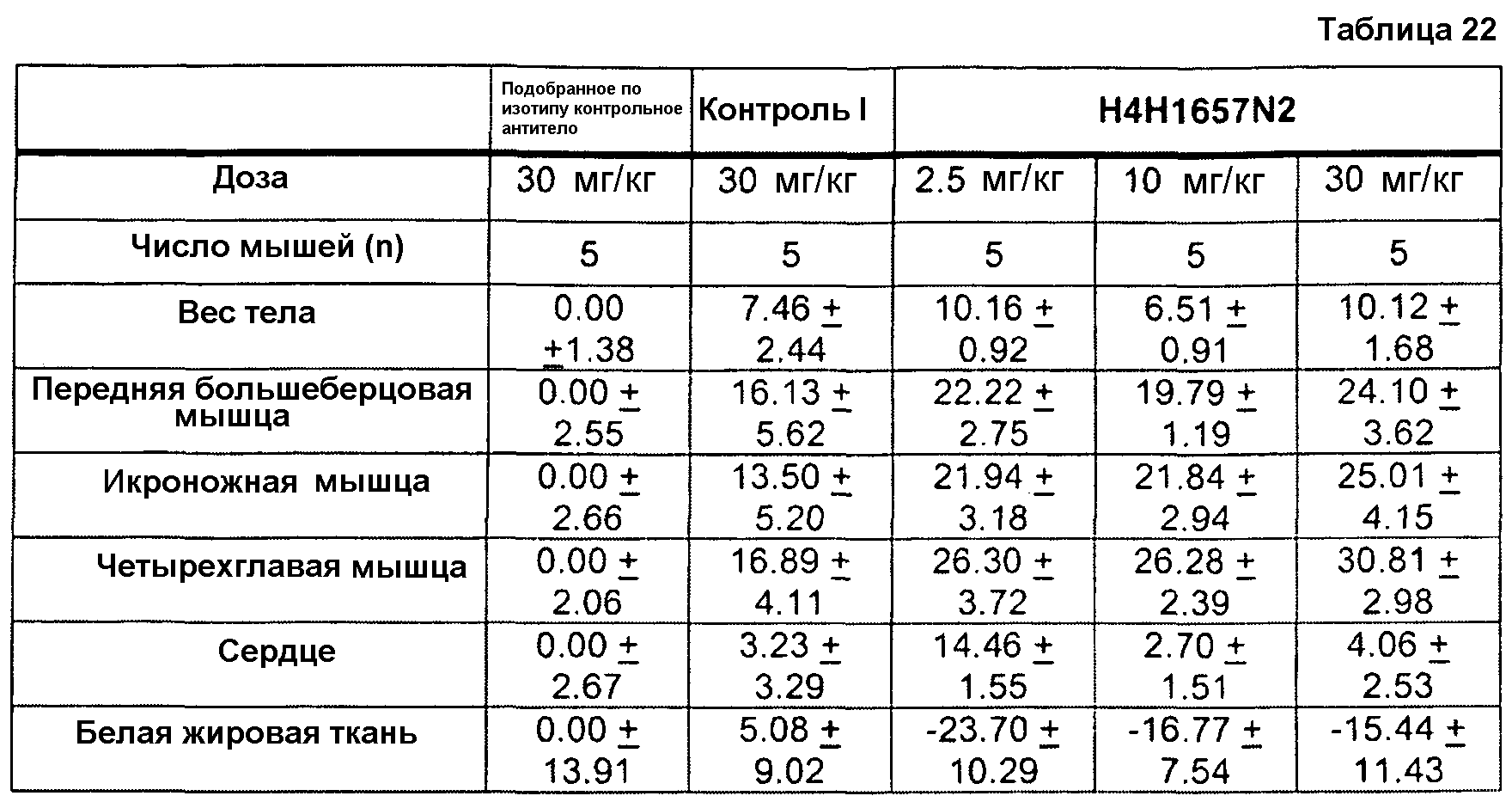
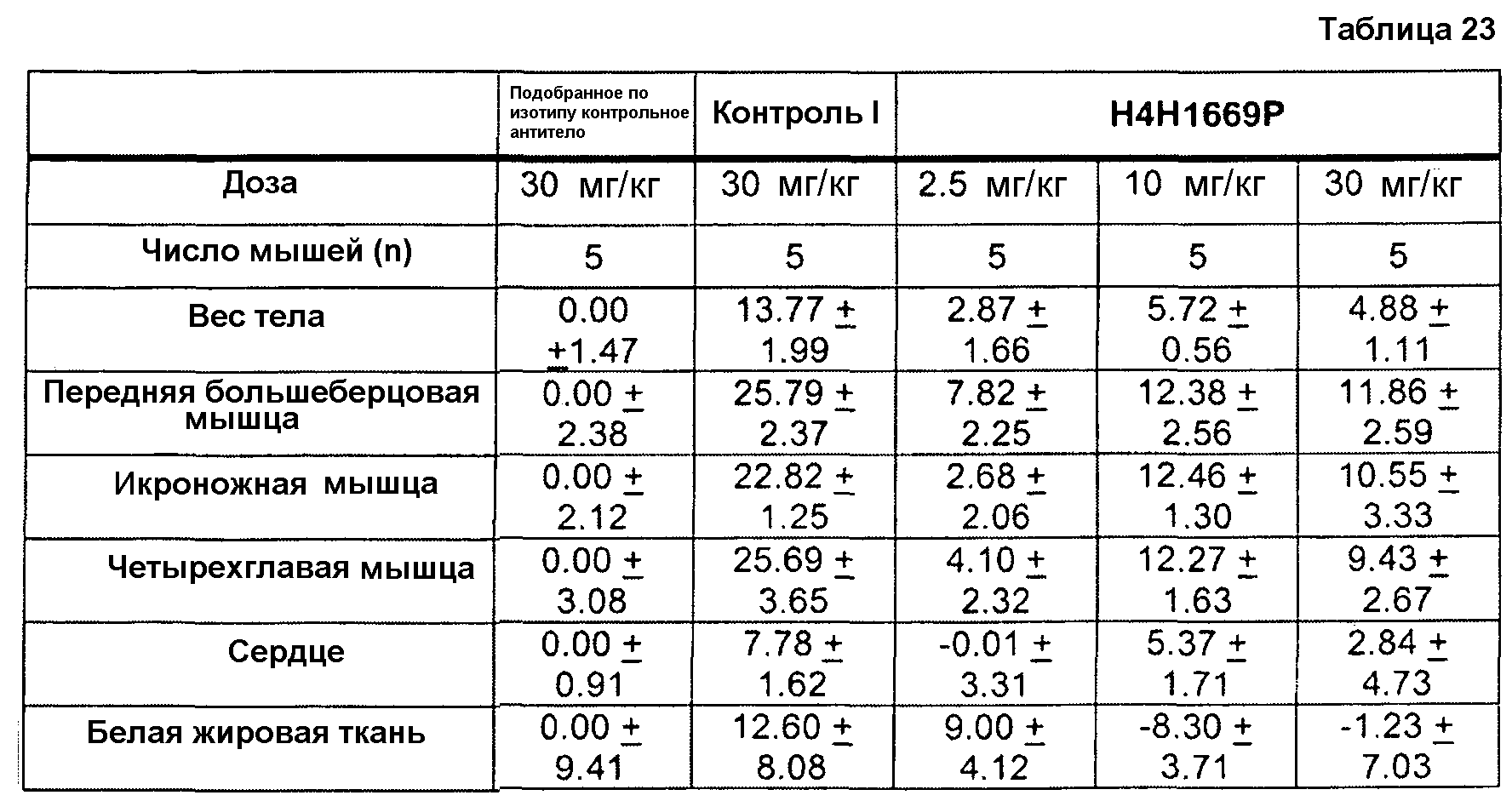
В отдельном эксперименте 5 группам из 6 самцов мышей SCID в возрасте 10 недель вводили подкожно подобранное по изотипу антитело в качестве отрицательного контроля в дозе, составляющей 10 мг/кг, и H4H1657N2 в дозе, составляющей 0,1, 0,75, 2,5 или 10 мг/кг, в соответствии со следующей схемой введения доз: 2 раза в неделю на недели 1 и 1 раз в неделю после этого в течение всего 28 дней лечения. Результаты суммированы в таблице 24 и представлены в виде увеличения в процентах по сравнению с отрицательным контролем ± среднеквадратическое отклонение.
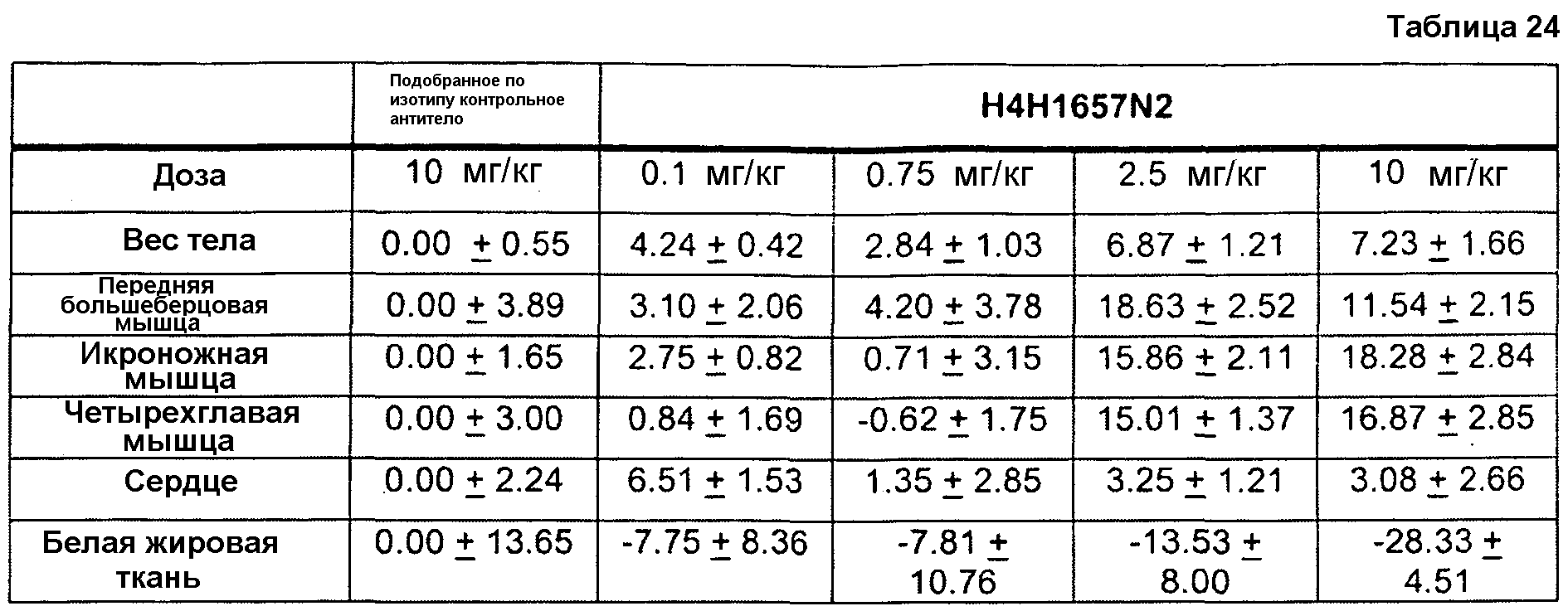
Как продемонстрировано в этом эксперименте, антитела H4H1657N2 и H4H1669P вызывали значительные увеличения мышечной массы при введении мышам на всем протяжении диапазона доз.
Пример 11. Эффект антител против GDF8 на гомеостаз глюкозы
Были проверены эффекты антител против GDF8 по настоящему изобретению на гомеостаз глюкозы и чувствительность к инсулину в модели на мыши вызванного диетой ожирения (DIO). В этом эксперименте у мышей добивались DIO посредством кормления мышей C57BL6 пищей с высоким содержанием жира (45 ккал% жира) в течение 7 недель, начиная с 9-недельного возраста. Начиная с недели 8, вводили антитела в дозе, составляющей 30 мг/кг, дважды в неделю в течение еще двух недель, и исследование прекращали спустя неделю после этого (21 день после начала лечения). Используемыми в этом эксперименте антителами были H4H1669P и H4H1657N2, а также подобранное по изотипу антитело в качестве отрицательного контроля и контроль I (антитело против GDF8, соответствующее Myo29). Тесты на чувствительность к инсулину проводили до и после лечения. Инсулин (2 МЕ/кг) вводили с помощью внутрибрюшинной инъекции после 4 часов голодания, и определяли уровни глюкозы. Результаты продемонстрированы на фиг. 3A (до лечения антителом) и 3B (после лечения антителом).
Как продемонстрировано в этом эксперименте, ингибирование миостатина в результате введения антитела против GDF8 улучшало гомеостаз глюкозы у мышей с DIO. Значимое различие в снижении уровня глюкозы в ответ на болюсную инъекцию инсулина было отмечено между группами лечения антителом против GDF8 (H4H1669P, H4H1657N2 или контролем I) и группой лечения подобранным по изотипу контрольным антителом.
Пример 12. In vivo блокирование мышечной атрофии посредством введения H4H1657N2
С целью определения in vivo способностей H4H657N2 к профилактики мышечной атрофии, вызванной иммобилизацией/обездвижением и введением дексаметазона, были использованы мыши С57.
В этом примере иммобилизации три группы из 8 мышей C57 подвергали анестезии, и голеностопный сустав фиксировали под углом в 90º с использованием иммобилизирующего материала на 14 дней. Четвертую группу оставляли не потревоженной и использовали в качестве контроля - не подвергнутой иммобилизации группы. В течение 14 дней иммобилизации мышам вводили подобранное по изотипу антитело в качестве отрицательного контроля; контроль I (Myo29) или H4H1657N2. Трем группам инъецировали подкожно антитело в дозе, составляющей 30 мг/кг, дважды в неделю в течение двух недель, начиная с момента времени иммобилизации. В таблице 25 суммированы результаты, представленные в виде изменения в процентах по сравнению с отрицательным контролем ± среднеквадратическое отклонение. Результаты указывали на то, что через 14 дней иммобилизации группа, которую лечили антителом H4H1657N2, продемонстрировала значимое уменьшение потери массы скелетной мускулатуры в сравнении с группой лечения подобранным по изотипу контрольным антителом.

В примере с использованием дексаметазона две группы из 5 мышей C57 подвергали анестезии, и им имплантировали подкожно осмотические насосы, которые доставляли 1 мкг/г/день дексаметазона. Двум дополнительным группам имплантировали осмотические насосы, которые доставляли солевой раствор, и их использовали в качестве контролей. В течение 14 дней лечения двум группам мышей (одной группе введения солевого раствора и одной группе введения дексаметазона) вводили подобранное по изотипу антитело в качестве отрицательного контроля; и двум группам мышей (одной группе введения солевого раствора и одной группе введения дексаметазона) вводили антитело H4H1657N2. Антитела вводили подкожно в дозе, составляющей 30 мг/кг, дважды в неделю в течение двух недель. Лечение антителом начинали в момент времени имплантации насоса. В таблице 26 суммированы результаты, представленные в виде изменение в процентах по сравнению с отрицательным контролем ± среднеквадратическое отклонение. Сравнение группы введения дексаметазона, которая получала антитело H4H1657N2, с группой, которая получала подобранное по изотипу контрольное антитело, указывает на то, что лечение антителом H4H1657N2 предотвращает потерю мышечной массы, вызванную введением дексаметазона.

Пример 13. Специфичность H4H1657N2 in vivo
С целью исследования специфичности H4H1657N2 in vivo мышам C57BL6 инъецировали лекарственное средство, и сыворотку, полученную от повергнутых лечению мышей, подвергали экспериментальному исследованию «улавливания» лиганда на основе масс-спектрометрии. Вкратце, лекарственное средство (4H1657N2, контроль IV или подобранное по изотипу антитело в качестве отрицательного контроля) инъецировали многократно (в день 0, день 3, день 7, день 10) в течение 14-дневного периода времени мышам C57BL6. Животных умерщвляли в день 14, и сыворотку инкубировали со сферами с антителами против Fc человека. Сферы отмывали, и связанный материал подвергали элюированию с использованием буфера для загрузки при электрофорезе в SDS-ПААГ. Подвергнутый элюированию материал подвергали электрофорезу в SDS-ПААГ, и вырезали части геля, соответствующие диапазону молекулярных масс 5-20 кДа. Для масс-спектрометрии образцы подвергали обработке, используя стандартные условия восстановления, алкилирования и трипсинизации. Гидролизаты разделяли на наноколонке C18 и автоматически наносили в виде пятен на содержащие «якоря» кружки от Bruker. MALDI-S (MS/MS) анализ (Bruker Ultraflextreme) выполняли в режиме автоматизированной работы, используя LC-WARP (Bruker Daltonics). Масс-спектры исследовали, используя MASCOT (Matrix Science), и оценку результатов осуществляли относительно подобранного по изотипу контрольного антитела. Элюированные белки перечислены в таблице 27.
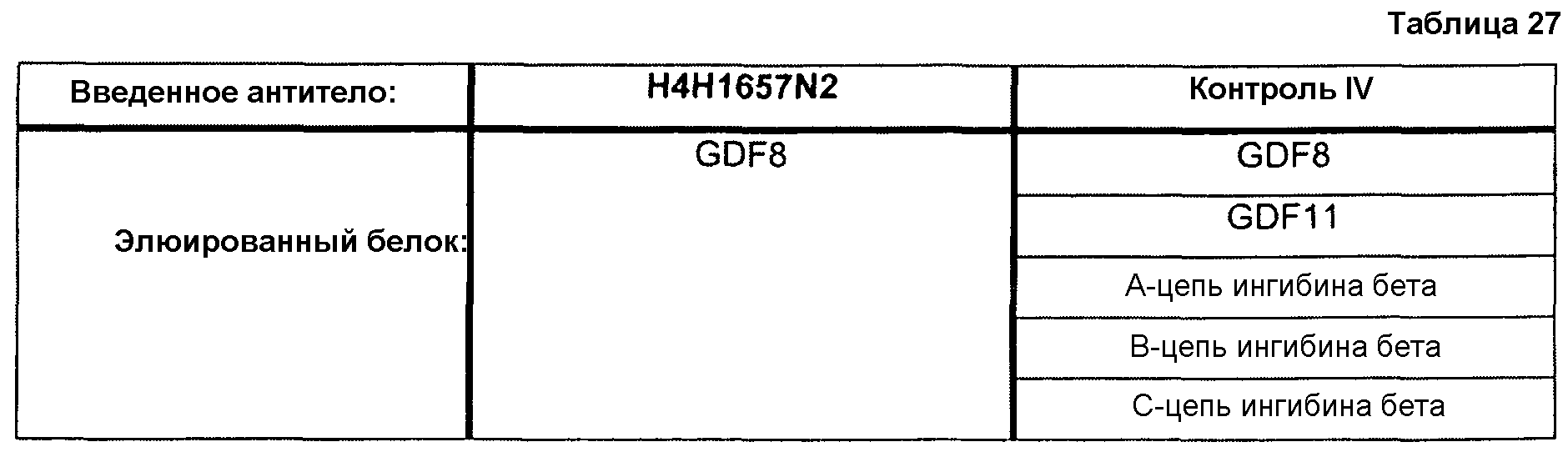
Как продемонстрировано в таблице 27, лишь GDF8 мыши был идентифицирован в качестве партнера по связыванию для H4H1657N2 с помощью этого эксперимента. (Последовательности GDF8 мыши и человека являются идентичными.) В отличие от этого, контроль IV связывался с несколькими другими членами семейства лигандов TGF бета, кроме GDF8, содержащими, в числе прочих, GDF11. Этот эксперимент подтверждает специфичность H4H1657N2 в отношении GDF8 в in vivo ситуации.
Объем настоящего изобретения не ограничивается конкретными вариантами осуществления, описанными в настоящем описании. На самом деле, различные модификации настоящего изобретения, помимо описанных в настоящем описании модификаций, станут очевидными квалифицированным в данной области специалистам благодаря вышеприведенному описанию и сопроводительным фигурам. Предполагается, что такие модификации попадают в объем прилагаемой формулы изобретения.












































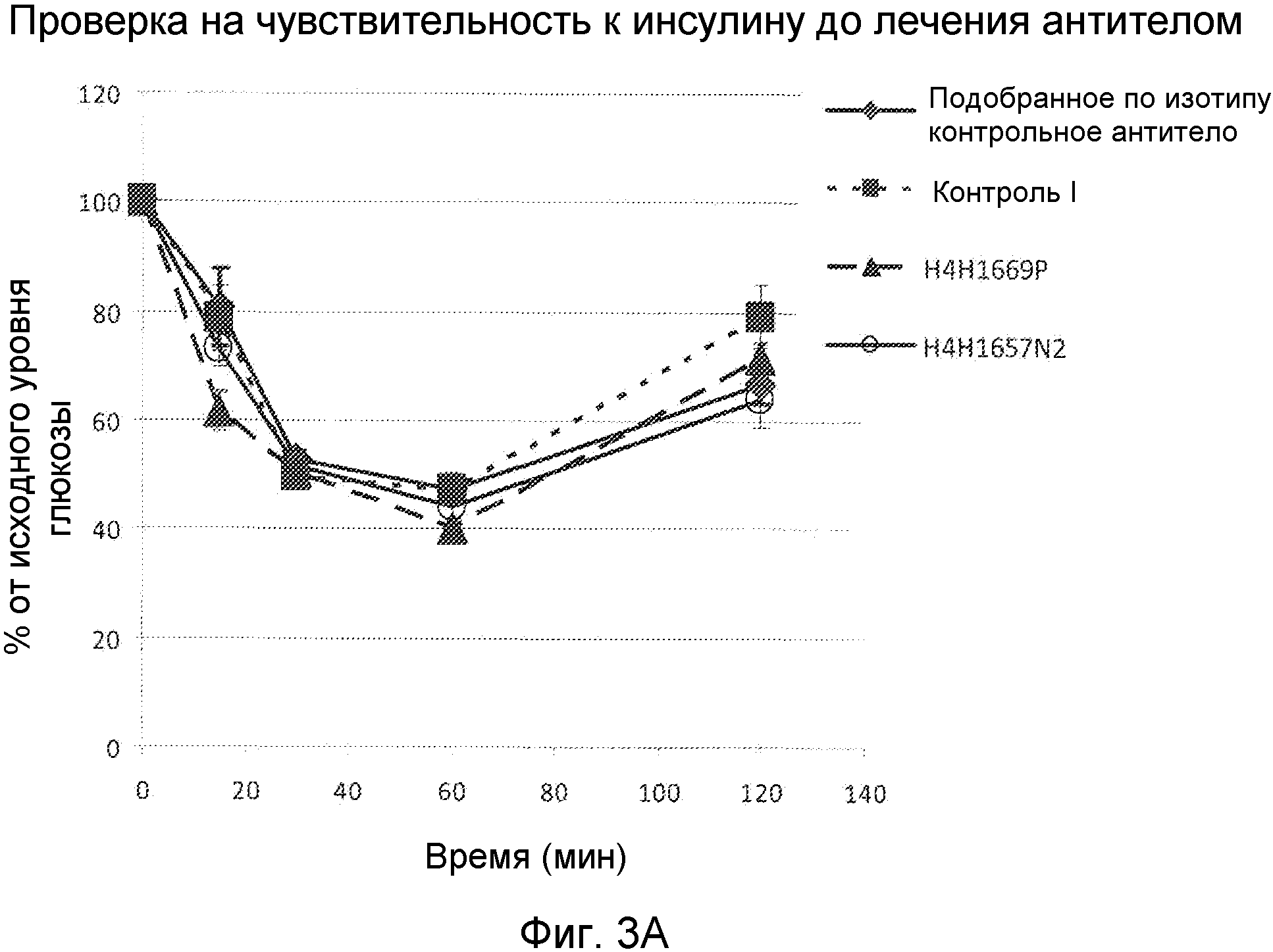






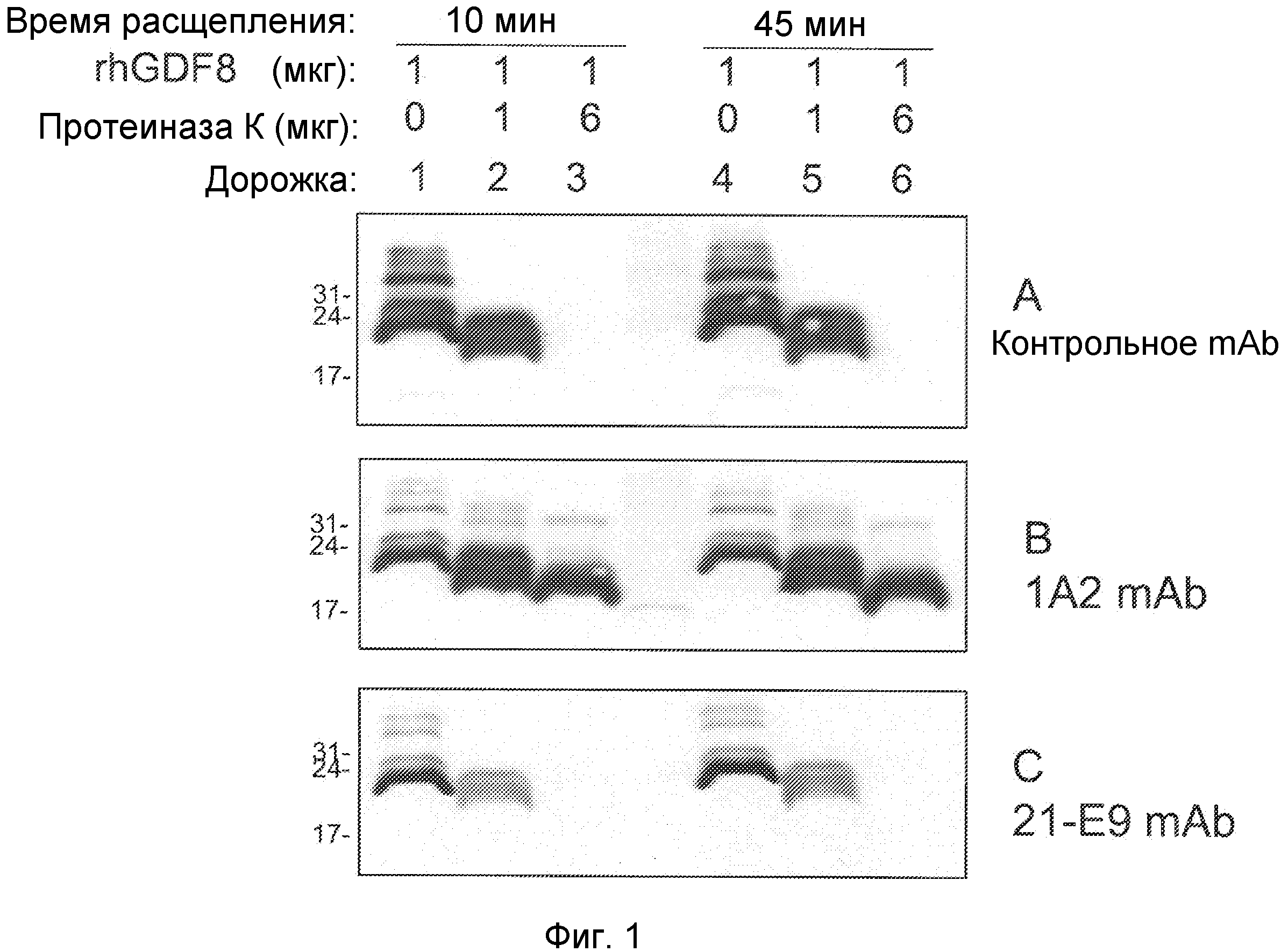
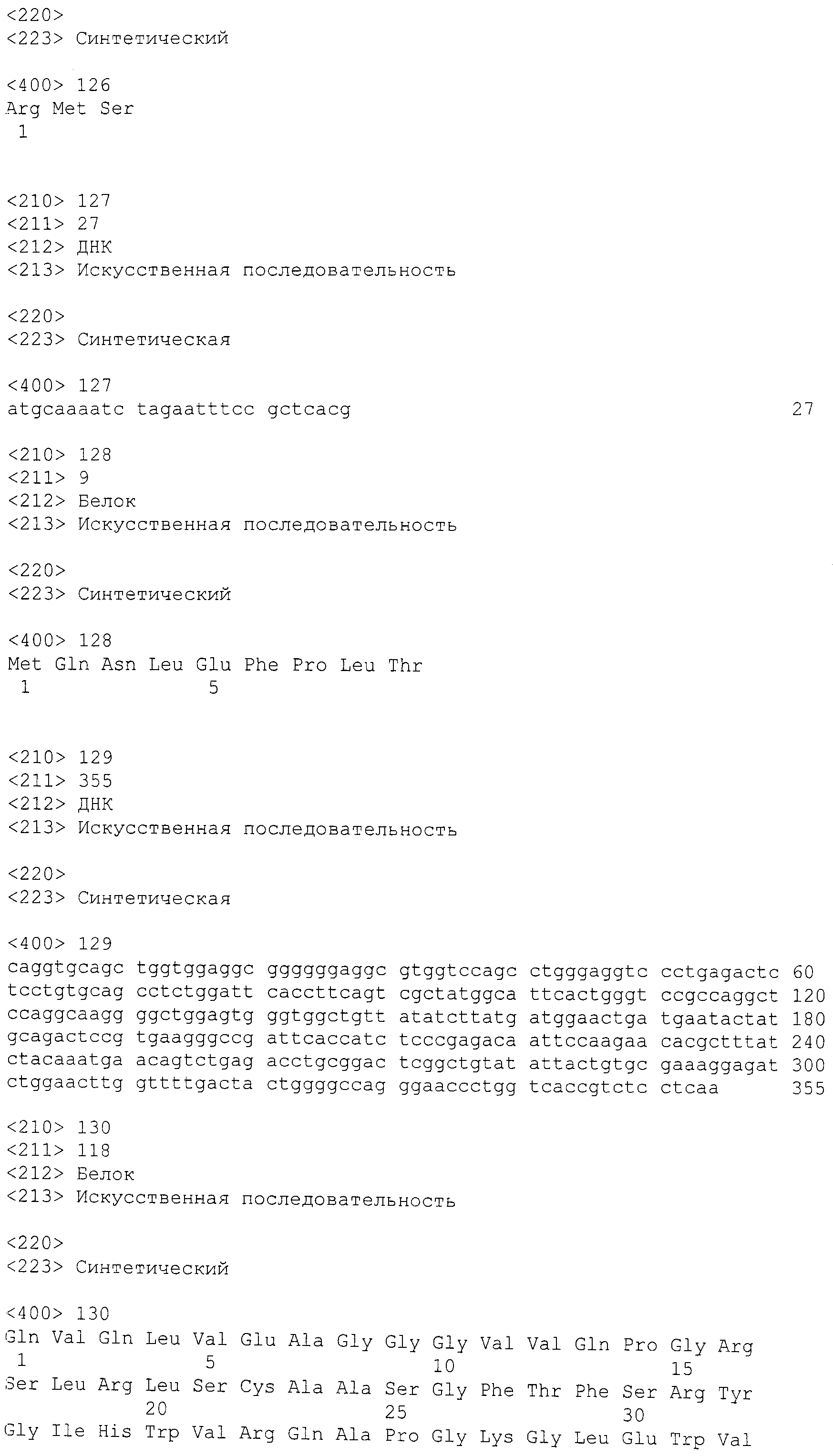
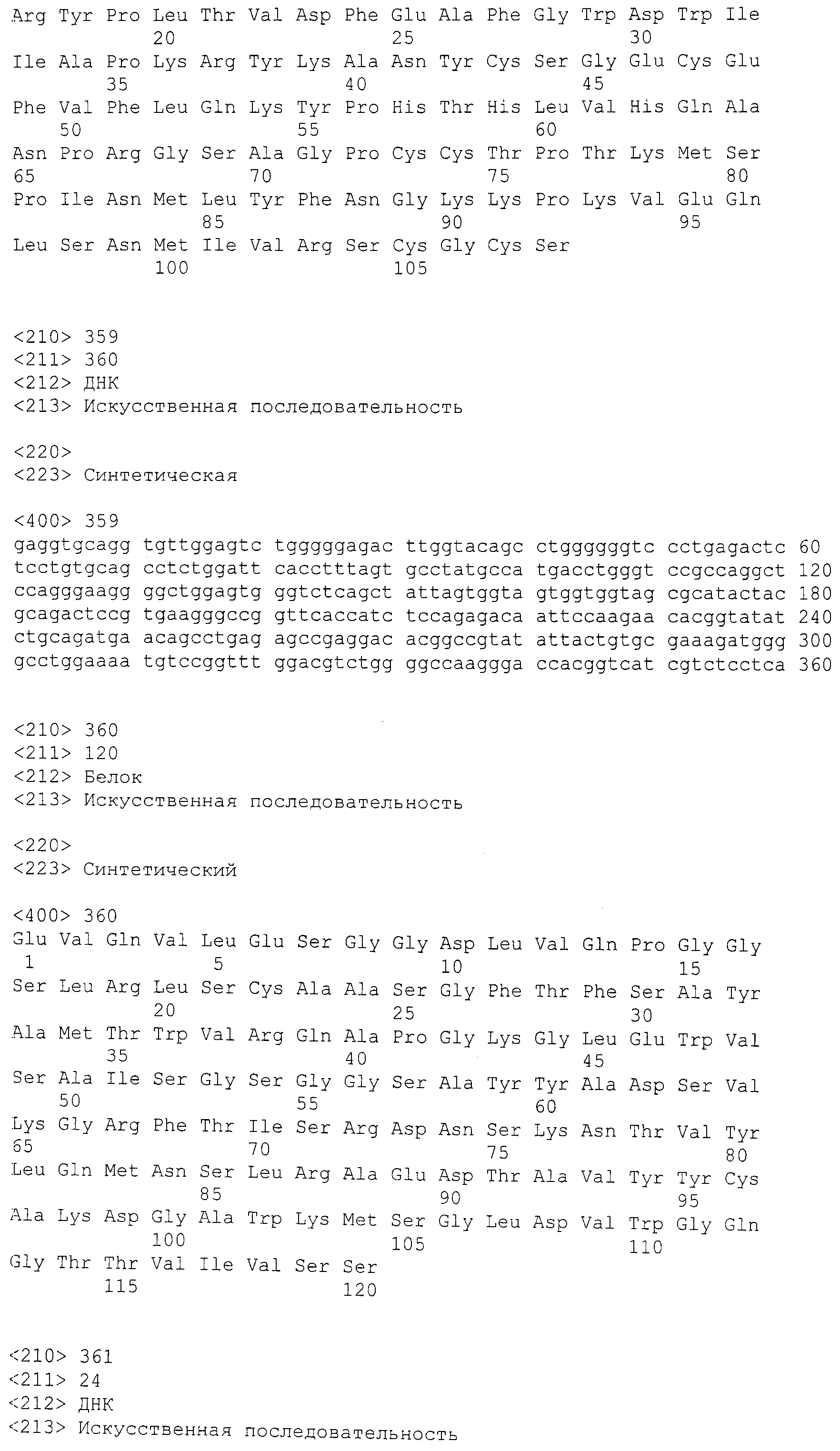





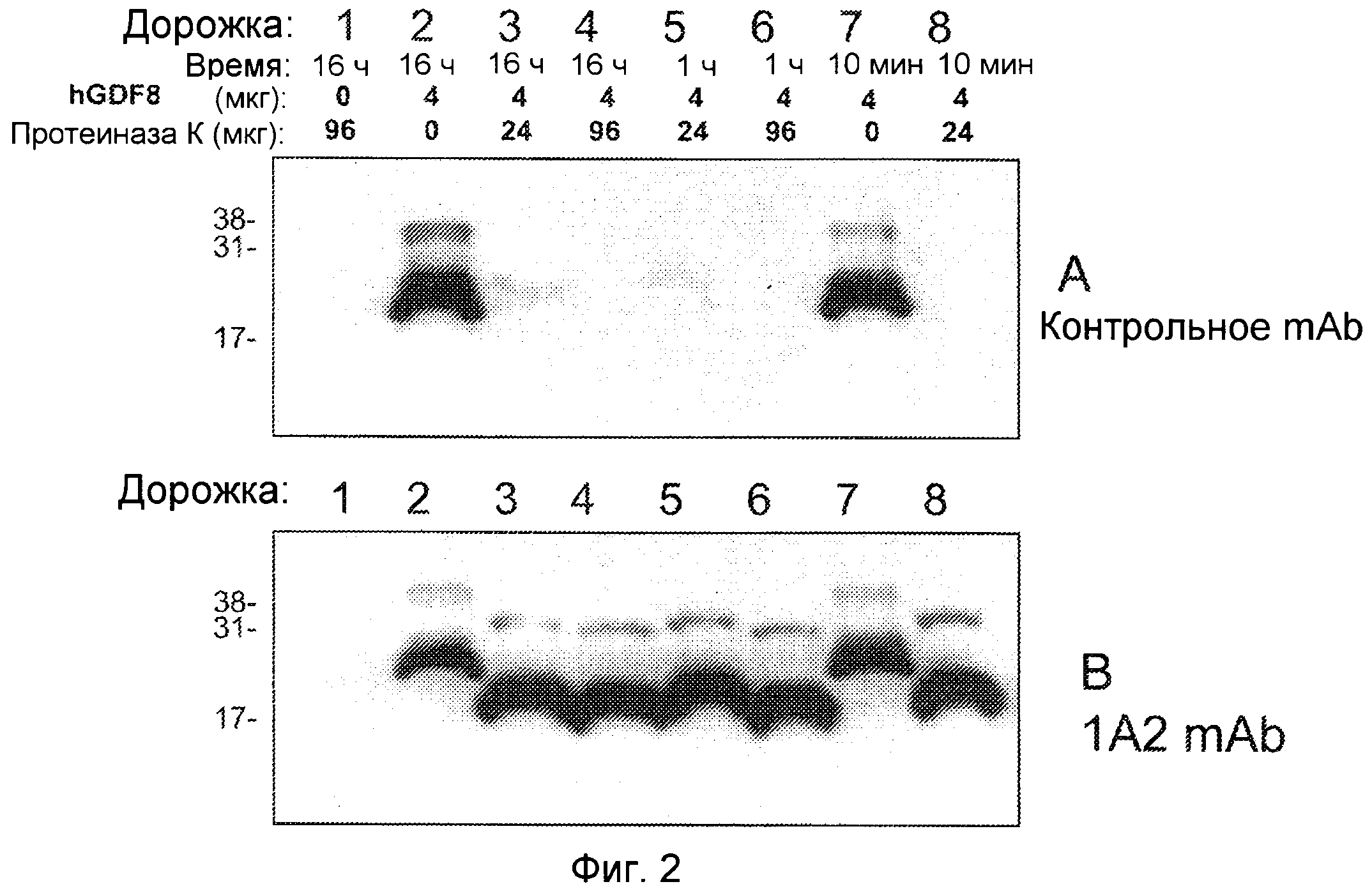
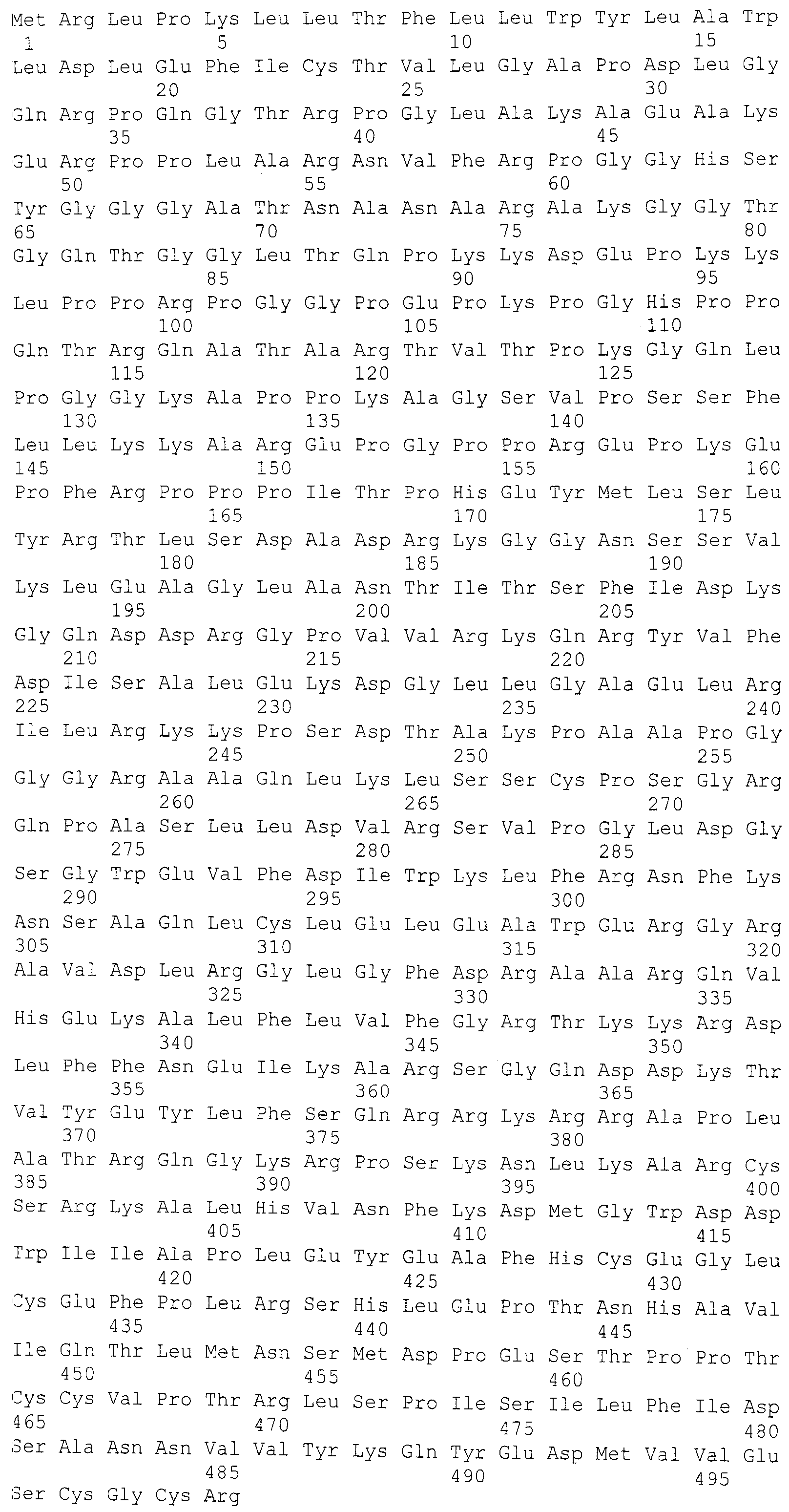













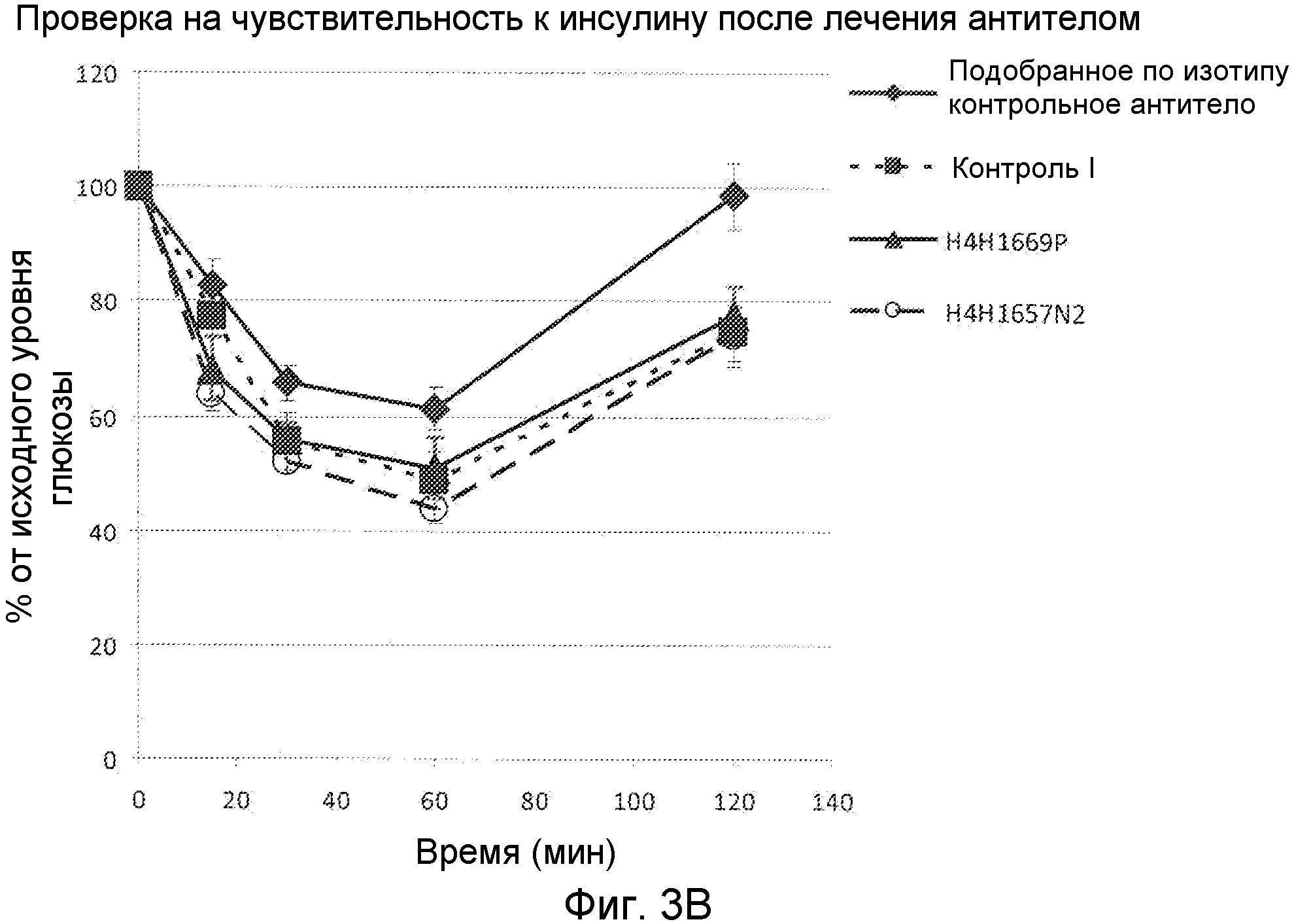
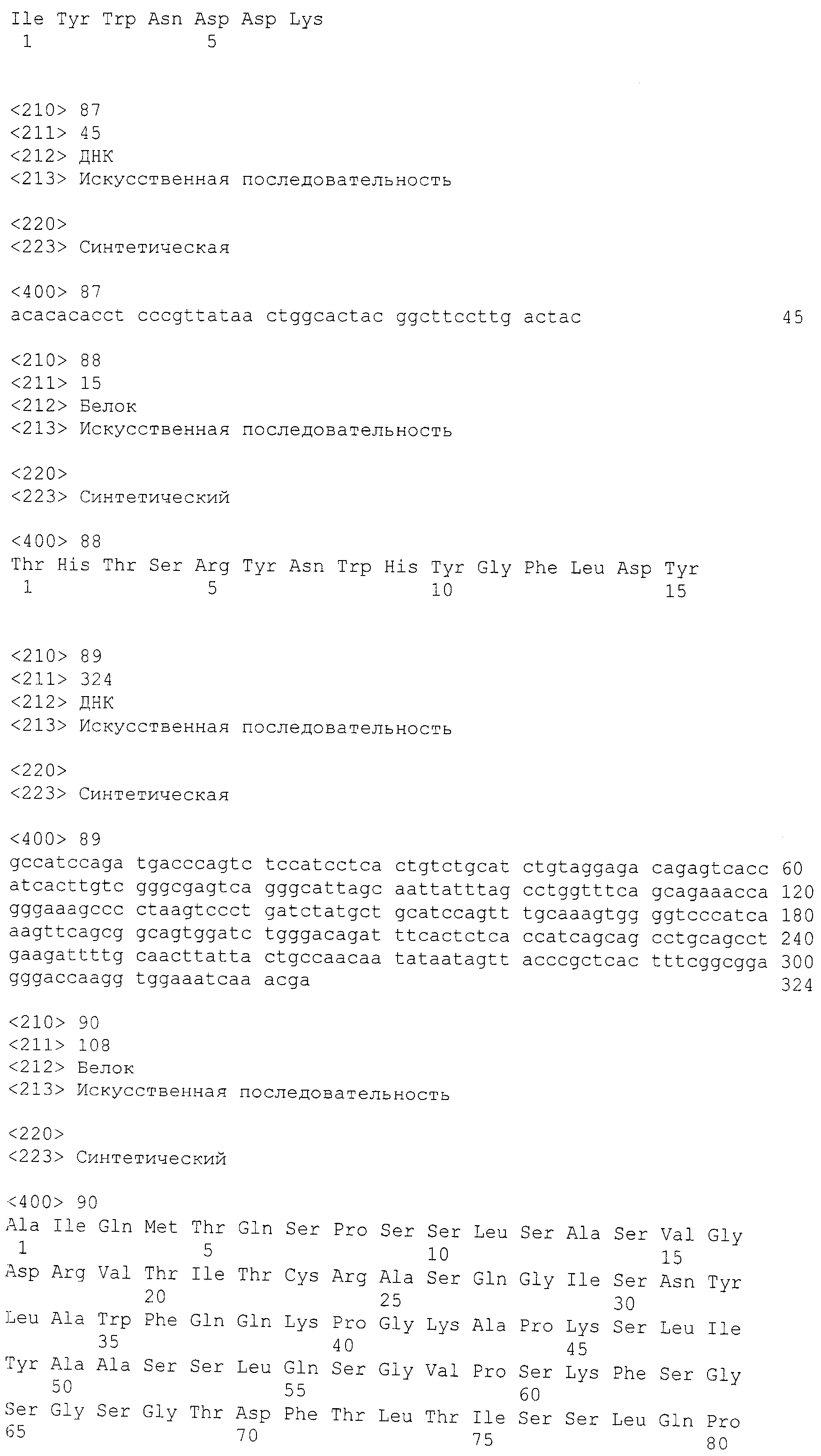






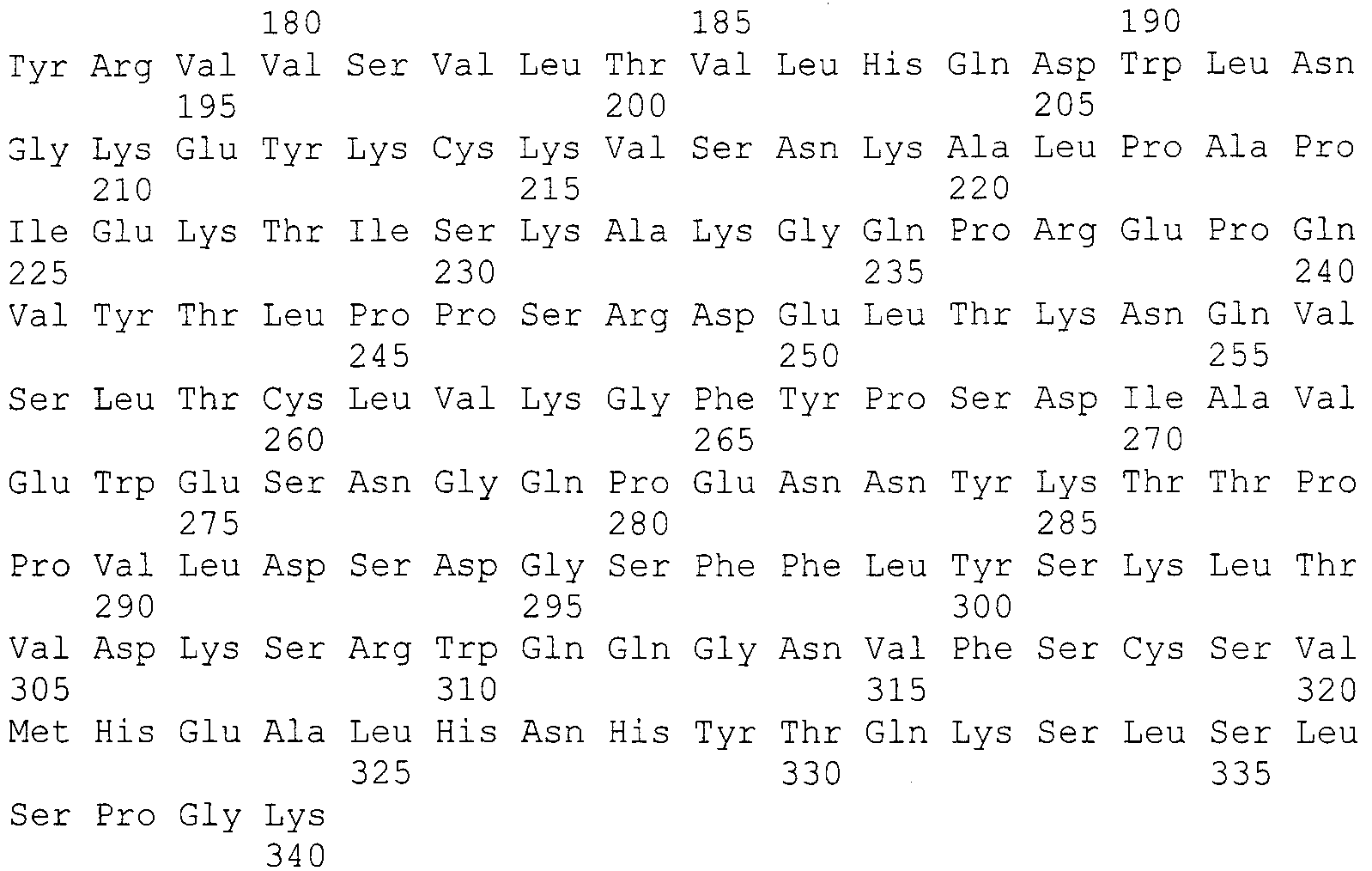





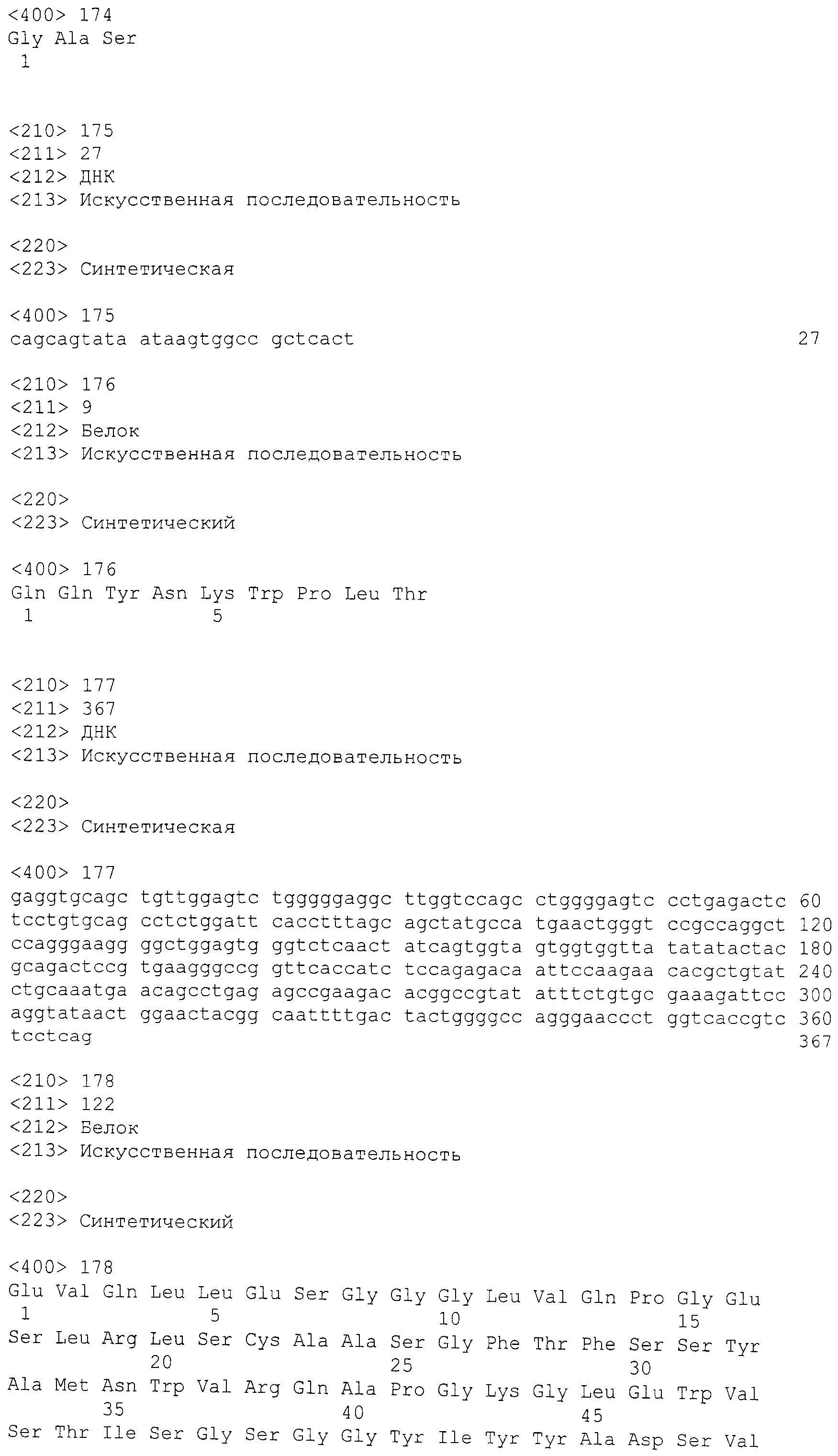












Антитела человека с высокой аффинностью к фактору роста нервов человека
Антитела человека к cd20 человека и способ их использования
Легковыделяемые биспецифические антитела с природным иммуноглобулиновым форматом
Человеческие антитела с высокой аффинностью к рецепторам il-4 человека
Высокоаффинные человеческие антитела к человеческому ангиопоэтину-2
Гуманизированные fcγr мыши
Высокоаффинные человеческие антитела к pcsk9
Стабилизированные составы, содержащие антитела к рецептору интерлейкина 6 (il-6r)
Легковыделяемые биспецифические антитела с природным иммуноглобулиновым форматом
Мышь с общей легкой цепью
Антитела человека с высокой аффинностью к фактору роста нервов человека
Антитела человека к cd20 человека и способ их использования
Легковыделяемые биспецифические антитела с природным иммуноглобулиновым форматом
Человеческие антитела с высокой аффинностью к рецепторам il-4 человека
Высокоаффинные человеческие антитела к человеческому ангиопоэтину-2
Гуманизированные fcγr мыши
Высокоаффинные человеческие антитела к pcsk9
Стабилизированные составы, содержащие антитела к рецептору интерлейкина 6 (il-6r)
Легковыделяемые биспецифические антитела с природным иммуноглобулиновым форматом
Мышь с общей легкой цепью

