Результат интеллектуальной деятельности: ИНСЕКТИЦИДНЫЕ БЕЛКИ
Вид РИД
Изобретение
УРОВЕНЬ ТЕХНИКИ
Настоящее изобретение относится к отраслям белковой инженерии, молекулярной биологии растений и борьбы с вредителями. Более конкретно, изобретение относится к новым сконструированным гибридным белкам, обладающим инсектицидной активностью, к нуклеиновым кислотам, результатом экспрессии которых являются эти инсектицидные белки, а также к способам получения и способам применения этих инсектицидных белков и соответствующих нуклеиновых кислот в борьбе с насекомыми.
Насекомые-вредители являются главной причиной потерь урожая. Только в США ежегодные потери, вызванные инвазией различных видов насекомых, составляют миллиарды долларов. Кроме того, что насекомые-вредители приводят к потерям урожая полевых культур, они также причиняют неприятности производителям овощей и фруктов, досаждают садоводам и домовладельцам.
Самыми вредоносными кукурузными вредителями считаются разные виды кукурузного корневого жука. Наиболее значительными его видами в США являются Diabrotica virgifera virgifera, западный кукурузный корневой жук, D.longicornis barberi, северный кукурузный корневой жук и D. undecimpunctata howardi, южный кукурузный корневой жук. Основными кукурузными вредителями в Кукурузном поясе США считаются только западный и северный кукурузные корневые жуки. Важным видом кукурузного корневого вредителя в южной части США является мексиканский кукурузный корневой жук, Diabrotica virgifera zeae. Наиболее существенный вред растению наносят личинки кукурузного корневого жука, поскольку они питаются исключительно корнями кукурузы. Этот вред выражается в увеличении полегания растений, снижении урожая зерна и растительной массы, а также в изменении содержания питательных веществ в зерне. Осуществление питания личинками оказывает также косвенное воздействие на кукурузу тем, что в его результате в корнях открываются ходы для инвазии бактерий и грибков, что приводит к болезням, выражающимся в гниении корня и стебля. Взрослые особи кукурузного корневого жука проявляют свою активность на кукурузных полях поздним летом, когда они питаются початками, «шелком», а также пыльцой, что препятствует нормальному опылению.
Борьбу с кукурузными корневыми жуками ведут главным образом путем интенсивного применения химических пестицидов, активность которых проявляется через препятствование росту насекомых, предотвращение их питания или размножения, или же гибель насекомых. Таким путем может быть достигнут хороший результат в борьбе против кукурузного корневого жука, однако иногда эти химикаты могут воздействовать и на другие, полезные организмы. Другой проблемой, проявляющейся в результате широкого применения химических пестицидов, является появление видов насекомых, устойчивых к ним. Еще одна проблема вызвана тем, что личинки кукурузного корневого жука питаются под землей, что затрудняет их контакт с вносимыми инсектицидами. Поэтому, в большинстве случаев применение инсектицидов осуществляется профилактически, во время посева. Результатом этой практики является большой вред для окружающей среды. Частично этот вред удалось снизить за счет применения различных способов ведения фермерского хозяйства, однако потребность в альтернативных механизмах борьбы с вредителями постоянно растет.
Биологические агенты борьбы с вредителями, такие как штаммы Bacillus thuringiensis (Bt) экспрессирующие пестицидные токсины, такие как δ-эндотоксины (дельта-эндотоксины, которые также называются кристаллическими токсинами или Cry-белками), также применялись для хлебных злаков и показали удовлетворительные результаты главным образом в борьбе с чешуекрылыми насекомыми-вредителями. Эти δ-эндотоксины представляют собой белки, заключенные в кристаллическую матрицу, известные своей инсектицидной активностью, проявляющейся при проглатывании их определенными видами насекомых. Классификация различных δ-эндотоксинов была выполнена на основе их спектра активности и гомологии последовательностей. До 1990 г. основные классы определялись по их спектру активности, причем белки Cry1 обладают активностью против чешуекрылых (молей и бабочек), белки Cry2 обладают активностью как против чешуекрылых, так и против двукрылых (мух и комаров), белки Cry3 обладают активностью против жесткокрылых (жуков), а белки Cry4 обладают активностью в борьбе против двукрылых (Hofte и Whitely, 1989, Microbiol. Rev. 53:242-255). В 1998 г. была разработана новая номенклатура, которая дала систематическую классификацию Cry-белков на основе гомологии последовательности аминокислот, а не на активности по отношению к определенным видам насекомых (Crickmore и др. 1998, Microbiol. Molec. Biol. Rev. 62:807-813).
Спектр инсектицидной активности отдельного δ-эндотоксина из Bt является довольно узким, то есть, данный δ-эндотоксин проявляет активность только против небольшого числа видов в данном отряде насекомых. Например, известно, что токсин Cry3A обладает очень высокой токсичностью по отношению к колорадскому картофельному жуку, Leptinotarsa decemlineata, однако обладает очень слабой или нулевой токсичностью по отношению к родственным жукам из рода Diabrotica (Johnson и др., 1993, J. Econ. Entomol. 86:330-333). Согласно работе Slaney и др. (1992, Insect Biochem. Molec. Biol. 22:9-18) токсичность токсина Cry3A, по меньшей мере, в 2000 раз слабее по отношению к личинкам южного кукурузного корневого жука, чем по отношению к колорадскому картофельному жуку. Известно также, что токсичность Cry3A по отношению к западному кукурузному корневому жуку или к северному кукурузному корневому жуку очень слабая или совсем отсутствует.
Специфичность δ-эндотоксинов является результатом эффективности различных шагов, предпринятых в процессе производства активного токсичного белка, и его последующего взаимодействия с эпителиальными клетками в средней кишке насекомого. Для того чтобы обладать инсектицидными свойствами, большинство из известных δ-эндотоксинов должны попасть в организм насекомого путем проглатывания и протеолитически активироваться для образования активного токсина. Активация инсектицидных кристаллических (Cry) белков представляет собой многоэтапный процесс. После проглатывания насекомым кристаллы должны в первую очередь раствориться в пищеварительном канале насекомого. После растворения δ-эндотоксины активируются путем специфичного протеолитического расщепления. Протеазы в пищеварительном канале насекомого могут играть важную роль в специфичности путем определения места, где перерабатывается δ-эндотоксин. После того, как δ-эндотоксин растворился и переработался, он привязывается к определенным рецепторам на поверхности эпителия средней кишки насекомого, а впоследствии интегрируется в липидный бислой мембраны щеточной каемки. Затем формируются ионные каналы, нарушающие нормальное функционирование средней кишки, в конечном итоге приводя к гибели насекомого.
У чешуекрылых, уровень pH в пищеварительных каналах которых является щелочным, протеазы кишечника перерабатывают 5-эндотоксины, например, Cry1Aa, Cry1Ab, Cry1Ac, Cry1B и Cry1F, из протоксинов размером 130-140 кДа в токсичные белки размером приблизительно 60-70 кДа. Процесс переработки протоксина в токсин, согласно опубликованным данным, осуществляется путем перемещения как N-концевых, так и C-концевых аминокислот, а точное местоположение процесса переработки зависит от конкретного δ-эндотоксина, и участие в нем принимают специфичные текучие вещества пищеварительного канала насекомого (Ogiwara и др., 1992, J.Invert. Pathol. 60:121-126). Таким образом, процесс активации требует отщепления всего C-концевого хвостового участка протоксина. Эта протеолитическая активация δ-эндотоксина может играть значительную роль в определении его специфичности.
Жесткокрылые насекомые имеют пищеварительные каналы с уровнем кислотности от более нейтрального до кислотного, и δ-эндотоксины, специфичные для жесткокрылых, подобны по своему размеру активированным токсинам, специфичным для чешуекрылых. Поэтому, прежде считалось, что переработка δ-эндотоксинов, специфичных для жесткокрылых, не является необходимой для их токсичности. Однако, исходя из полученных данных, можно предположить, что δ-эндотоксины, активные против жесткокрылых, растворяются и протеолитически перерабатываются в токсичные полипептиды меньшего размера. Белковый δ-эндотоксин Cry3A размером 73 кДа, вырабатываемый бактериями В.thuringiensis var. tenebrionis, легко поддается переработке в бактерии на N-конце, теряя 49-57 остатков в процессе формирования кристалла или после него, образуя в итоге обычную изолированную форму размером 67 кДа (Carroll и др., 1989, Biochem. J. 261:99-105). В своей работе McPherson и др., (1988, Biotechnology 6:61-66) также продемонстрировали, что нативная кодирующая последовательность cry3A содержит два функциональных кодона, инициирующих трансляцию, в одной и той же рамке считывания, один - для кодирования белка размером 73 кДа, а другой - для кодирования белка размером 67 кДа, начиная соответственно с Met-1 и Met-48 расшифрованной последовательности аминокислот. Поэтому оба белка могут считаться полноразмерными белками Cry3A природного происхождения.
По мере расширения знаний о том, как функционируют δ-эндотоксины, увеличилось число попыток разработать δ-эндотоксины, обладающие новыми видами активности. Разработка δ-эндотоксинов стала более возможна в результате определения трехмерной структуры Cry3A в 1991 г.(Li и др., 1991, Nature 353:815-821). Li и др. определили, что белок Cry3A имеет три структурных домена: N-концевой домен I, из остатков 58-290, состоящий из семи α-спиралей, домен II, из остатков 291-500, состоящий из трех β-листов в упаковке так называемым греческим ключом, а также C-концевой домен III, из остатков 501-644, представляющий собой β-сэндвич в так называемой упаковке рулетом. Также была определена трехмерная структура активного против чешуекрылых токсина Cry1Aa (Grochulski и др., 1995, J. Mol. Biol. 254:447-464). Токсин Cry1Aa имеет три домена: N-концевой домен I, из остатков 33-253, домен II из остатков 265-461, а также домен III из остатков 463-609 с дополнительной внешней цепочкой в одном из β-листов из остатков 254-264. Если структуры Cry3A и Cry1Aa спроектировать на другие последовательности Cry1, то к домену I относятся аминокислотные остатки от 28 до 260, к домену II - приблизительно от 260 до 460, а к домену III - приблизительно от 460 до 600. См., работы Nakamura и др., Agric. Biol. Chem. 54(3): 715-724 (1990); Li и др., Nature 353: 815-821 (1991); Ge и др., J. Biol. Chem. 266(27): 17954-17958 (1991); и Honee и др., Mol. Microbiol. 5(11): 2799-2806 (1991); каждая из которых включена в настоящую заявку путем ссылки. Таким образом, в настоящее время известно, что на основе гомологии последовательности аминокислот известные Bt δ-эндотоксины имеют подобную трехмерную структуру, состоящую из трех доменов.
Токсинная часть Bt Cry-белков также характеризуется наличием пяти консервативных блоков по их последовательности аминокислота, имеющих номера от СВ1 до СВ5, расположенных в таком порядке на протяжении от N-конца до С-конца (Hofte и Whiteley, см. выше). Консервативный блок 1 (СВ1) содержит приблизительно 29 аминокислот, консервативный блок 2 (СВ2) содержит приблизительно 67 аминокислот, консервативный блок 3 (СВ3) содержит приблизительно 48 аминокислот, консервативный блок 4 (СВ4) содержит приблизительно 10 аминокислот, а консервативный блок 5 (СВ5) содержит приблизительно 12 аминокислот. Последовательности, расположенные перед этими пятью консервативными блоками и после них, являются в высокой степени вариабельными, а поэтому называются «вариабельными участками» V1-V6. В типичном случае домен I Bt δ-эндотоксина состоит из вариабельного участка 1, консервативного блока 1, вариабельного участка 2 и 52-х N-концевых аминокислот консервативного блока 2. Домен II в типичном случае состоит из C-концевых аминокислот (приблизительно 15) консервативного блока 2, вариабельного участка 3 и N-концевых аминокислот (приблизительно 10) консервативного блока 3. Домен III в типичном случае состоит из C-концевых аминокислот (приблизительно 38) консервативного блока 3, вариабельного участка 4, консервативного блока 4, вариабельного участка 5 и консервативного блока 5. Активные против чешуекрылых токсины Cry1, как и другие дельта-токсины, имеют вариабельный участок 6, состоящий приблизительно из 1-3 аминокислот, лежащий в пределах домена III.
Многие Bt-штаммы и δ-эндотоксины являются активными по отношению к различным видам насекомых и нематод. Однако относительно небольшое число этих штаммов и токсинов обладают активностью против жесткокрылых насекомых. Кроме того, большинство активных по отношению к жесткокрылым δ-эндотоксинов, известных в настоящее время, например, Cry3A, Cry3B, Cry2C, Cry7A, Cry8A, Cry8B, и Cry8C, обладают недостаточной оральной токсичностью против кукурузного корневого жука, что не позволяет обеспечить адекватные меры борьбы с ним в случае подачи их через микробы и трансгенные растения. Поэтому, существует необходимость разработки других подходов для производства новых токсинов, активных против кукурузного корневого жука.
Были разработаны активные против чешуекрылых 5-эндотоксины в попытках улучшения специфической активности или расширения спектра инсектицидной активности. Например, домен специфичности для тутового шелкопряда (Bombyx топ) из белка CrylAa переместили в белок Cry 1 Ас, таким образом наделив новой инсектицидной активностью полученный в результате гибридный Bt-белок (Ge и др. 1989, PNAS 86: 4037-4041). Кроме того, Bosch и др. 1998 (Патент США 5,736,131, включенный в настоящую заявку путем ссылки) описывает гибридные токсины Bacillus thuringiensis, содержащие на своем С-конце домен III первого Cry-белка, а на своем N-конце - домены I и II второго Cry-белка. Такие гибридные токсины продемонстрировали изменившуюся инсектицидную специфичность против чешуекрылых насекомых. Например, гибридный токсин Н04, описанный также в источнике De Maagd и др., Appl. Environ. Microbiol. 62(5): 1537-1543 (1996), содержит на своем N-конце домены I и II белка Cry1Ab, а на своем С-конце - домен III белка Cry1C. Согласно полученным данным, Н04 обладает высокой токсичностью по отношению к чешуекрылому насекомому Spodoptera exigua (свекловичная совка) по сравнению с родительским токсином Cry1Ab, a также значительно большей токсичностью по сравнению с родительским токсином Cry1C. Также было показано, что заменой домена III в токсинах, не являющихся активными против свекловичной совки, таких как Cry1E и Cry1Ab, на домен III из Cry1C, являющегося активным против свекловичной совки, можно получить гибридные токсины, обладающие активностью против этого насекомого. Во всех гибридах, описанных в работе Bosch и др., используются домены из Cry-белков, активных против чешуекрылых, для получения новых токсинов с активностью против чешуекрылых. Полученные результаты дают основание предполагать, что домен III белка Cry1C является важным определителем специфичности относительно свекловичной совки. См. также источники Bosch и др., FEMS Microbiology Letters 118:
129-134 (1994); Bosch и др., Bio/Technology 12: 915-918 (1994); De Maagd и др., Appl. Environ. Microbiol. 62(8): 2753-2757 (1996); и De Maagd и др., Mol. Microbiol. 31(2): 463-471 (1999); каждый из которых включен сюда путем ссылки.
Имеются данные о нескольких попытках разработки δ-эндотоксинов, активных против жесткокрылых. Chen и Stacy (Патент США 7,030,295, включенный сюда путем ссылки) успешно создали токсин, активный против кукурузного корневого жука путем введения не встречающегося в природе сайта узнавания протеазы в домен I, в домен III, или как в домен I, так и в домен III белка Cry2A. Один из полученных в результате модифицированных Cry3A-белков, обозначенный Cry3A055, в домен I которого ввели сайт узнавания протеазы, оказался активным по отношению к различным видам Diabrotica. Van Rie и др., 1997, (Патент США №. 5,659,123) разрабатывали Cry3A путем произвольной замены аминокислот, считающихся важными в вопросе доступности растворителю, в домене II на аминокислоту аланин. Некоторые из этих произвольных замен в домене II, согласно полученным данным, проявились в повышении активности против западного кукурузного корневого жука. Однако другие исследователи показали, что некоторые замены аланином в домене II белка Cry3A дали в результате разрушение связи с рецептором или нестабильность структуры (Wu и Dean, 1996, J. Mol. Biol. 255: 628-640). Согласно данным English и др., 1999, (Номер публикации международной патентной заявки WO 99/31248) в результате замены аминокислот в Cry3 Bb повысилась токсичность против южного и западного кукурузного корневого жука. Однако из полученных данных по 35 мутантам Cry3 Bb видно, что только три из них, с мутациями в основном в домене II и на стыке домен I-домен II, оказались активными против западного кукурузного корневого жука. Кроме того, вариации токсичности исходного Cry3 Bb против западного кукурузного корневого жука в одних и тех же пробах превышали различия между подвергшимися мутациям токсинами Cry3Bb и исходными токсинами Cry3 Bb. Shadenkov и др. (1993, Mol. Biol. 27:586-591) получили гибридный белок путем слияния аминокислот 48-565 белка Cry3A с аминокислотами 526-725 белка Cry1Aa. Таким образом, скрещивание последовательностей Cry3A и Cry1Aa произошло в консервативном блоке 4, расположенном в домене III. Cry3A является очень активным против колорадского картофельного жука (Leptinotarsa decemlineatd). Однако гибридный белок, описанный Shadenkov и др., не был активным против колорадского картофельного жука, несмотря на то, что гибридный белок состоял из последовательности Cry3A более чем на 75%. Таким образом, добавление только 25% последовательности Cry1Aa уничтожило активность против жесткокрылых насекомых, которой обладал родительский белок Cry3A. Это позволяет предположить, что гибридные белки, полученные путем слияния части Cry-белка, активного против жесткокрылых, например, белка Cry3A, с активным против чешуекрылых Cry-белком, например, Cry1A, не будут обладать активностью против жесткокрылых насекомых, в частности, против жесткокрылого насекомого, не имеющего такой природной восприимчивости к Cry3A, как у кукурузного корневого жука.
Исходя из вышеописанного, очевидно, что остается потребность разработки новых эффективных средств борьбы с вредителями, которые были бы экономически выгодными фермерам и приемлемыми для окружающей среды. В особенности, нужны такие белки, обладающие токсичностью против видов Diabrotica, главного вредителя кукурузы, способ действия которых отличался бы от существующих средств борьбы с вредителями в плане ослабления развития сопротивляемости. Кроме того, желательно, чтобы подача таких средств борьбы осуществлялась через продукты, минимизирующие вред для окружающей среды, например, через трансгенные растения.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В свете этих потребностей целью настоящего изобретения является предоставление новых сконструированных гибридных инсектицидных белков (сГИБов). Такие новые сГИБы получают путем слияния уникальных комбинаций вариабельных участков и консервативных блоков, по меньшей мере, двух различных Cry-белков и, по выбору, включения протоксинного хвостового участка из Bt Cry-белка в С-конец, или N-концевого пептидильного фрагмента, или того и другого. Не ограничивающим примером является комбинирование полных или неполных вариабельных участков и консервативных блоков из первого Cry-белка, обладающего активностью против жесткокрылых, с полными или неполными вариабельными участками и консервативными блоками второго Cry-белка, обладающего активностью против чешуекрылых и отличающегося от первого Cry-белка, и, по выбору, включение протоксинной хвостовой области из активного против чешуекрылых Bt Cry-белка или N-концевого пептидильного фрагмента, или обоих, в результате чего получаются новые сконструированные гибридные инсектицидные белки, обладающие активностью против спектра насекомых, отличающегося от спектра первого или второго родительский Cry-белков или того и другого. Такие новые сГИБы могут содержать полные или неполные вариабельные участки, консервативные блоки или домены из модифицированного белка Cry3A, а также из Cry-белка, отличающегося от модифицированного белка Cry3A. Пептидильный фрагмент может наделять сГИБ инсектицидной активностью, или может делать инсектицидную активность сГИБа более высокой, чем у сГИБа, не имеющего пептидильного фрагмента, или может делать сГИБ более стабильным, чем сГИБ, не имеющий пептидильного фрагмента. Токсичность сГИБа по настоящему изобретению против кукурузного корневого жука (Diabrotica sp.) неожиданно оказалась удивительно высокой. Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим данный сГИБ или комплементарным к гибридизирующемся в жестких условиях рекомбинантным гибридным нуклеиновым кислотам по настоящему изобретению.
Кроме того, в изобретение включены векторы, содержащие такие рекомбинантные (или комплементарные к ним) нуклеиновые кислоты, растение или микроорганизм, включающие в себя такие нуклеиновые кислоты и дающие возможность их экспрессии, например, трансгенная кукуруза; потомство таких растений, в котором содержатся нуклеиновые кислоты, устойчиво включенные в него и наследуемые согласно Менделю, а также/или семена таких растений и такого потомства.
Изобретение также включает в себя композиции и формулировки, содержащие сГИБы, обладающие возможностью ингибировать способность насекомых выживать, расти и размножаться, или позволяющие ограничить причиняемый насекомыми ущерб урожаю зерновых, например, путем применения этих сГИБов или композиций или формулировок в местах инвазии насекомых, или профилактической обработкой восприимчивых к насекомым площадей или растений для защиты их от насекомых-вредителей.
Кроме того, изобретение относится к способу получения сГИБов и к способам использования нуклеиновых кислот, например, в микроорганизмах, для борьбы с насекомыми или в трансгенных растениях для придания защиты от насекомых.
Описанные здесь новые сГИБы обладают высокой активностью против насекомых. Например, сГИБы по настоящему изобретению можно использовать для борьбы с серьезными вредителями-насекомыми, такими как западный кукурузный корневой жук (Diabrotica virgifera virgifera), северный кукурузный корневой жук (Л longicornis barberi) и мексиканский кукурузный корневой жук (D. virgifera zeae). Некоторые сГИБы можно также применять для борьбы с европейским мотыльком кукурузным (Ostrinia nubilalis) и другими чешуекрылыми насекомьми. СГИБы можно применять отдельно или в комбинации с другими стратегиями борьбы с насекомыми для получения максимальной эффективности борьбы с вредителями при минимальном вреде для окружающей среды.
Другие аспекты и преимущества настоящего изобретения станут понятными специалистам в данной отрасли в процессе изучения следующего описания изобретения с не ограничивающими его примерами.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Поданные ниже рисунки являются частью настоящего описания и включены с целью продемонстрировать некоторые аспекты настоящего изобретения. Понимание изобретения может быть облегчено путем обращения к одному или нескольким из этих фигур в комбинации с подробным описанием конкретных исполнений изобретения,
представленным здесь.
Фиг.1А-1Е показывает сопоставление последовательности в некоторых исполнениях сГИБ с родительским Cry-белками или модифицированными белками Cry3A, применявшимися для конструирования этих сГИБов, включая Cry3A, Cry1Ab, и Cry3A055, а также указывает процент идентичности. Подчеркнуты N-концевые пептидильные фрагменты. Пять консервативных блоков обозначены ярлычками СВ1-СВ5. Местоположения соединений между доменами I, II и III обозначены вертикальной пунктирной линией. Последовательность AAPF узнавания протеазы катепсин G показана жирным.
На Фиг.2А-2Е показан набор выполнений сГИБ, активных, по меньшей мере, против западного кукурузного корневого жука, и указан процент идентичности по сравнению со сГИБ 8AF. N-концевые пептидильные фрагменты подчеркнуты одной линией. С-концевые протоксинные хвостовые участки подчеркнуты двойной линией. Пять консервативных блоков обозначены ярлычками СВ1-СВ5. Местоположения соединений между доменами I, II и III показаны значком "↓" и имеют соответствующие ярлычки. Местоположения позиций кроссинговера обозначены значком "◆", Последовательность AAPF узнавания протеазы катепсин G показана жирным.
На Фиг.3 показана карта рекомбинантного вектора 12207, используемого для трансформации кукурузы и включающего экспрессионную кассету с промотором убиквитина кукурузы, операбельно связанным с кодирующей последовательностью FRCG, операбельно связанной с NOS-терминатором.
На Фиг.4 показана карта рекомбинантного вектора 12161, используемого для трансформации кукурузы и включающего в себя экспрессионную кассету с промотором убиквитина кукурузы, операбельно связанным с кодирующей последовательностью FR8a, операбельно связанной с NOS-терминатором.
На Фиг.5 показана карта рекомбинантного вектора 12208, используемого для трансформации кукурузы и включающего в себя экспрессионную кассету с промотором, которым является вирус скручивания желтых листьев цеструма (cmp),
операбельно связанный с кодирующей последовательностью FRCG, операбельно связанной с NOS-терминатором.
На Фиг.6 показана карта рекомбинантного вектора 12274, используемого для трансформации кукурузы и включающего в себя экспрессионную кассету с промотором, которым является вирус скручивания желтых листьев цеструма (cmp), операбельно связанный с кодирующей последовательностью FR8a, операбельно связанной с NOS-терминатором.
На Фиг.7 показана карта рекомбинантного вектора 12473, используемого для трансформации кукурузы и включающего в себя экспрессионную кассету с промотором убиквитин кукурузы (ubi), операбельно связанный с кодирующей последовательностью FRD3, операбельно связанной с NOS-терминатором.
На Фиг.8 показана карта рекомбинантного вектора 12474, используемого для трансформации кукурузы и включающего в себя экспрессионную кассету с промотором, которым является вирус скручивания желтых листьев цеструма (cmp), операбельно связанный с кодирующей последовательностью FRD3, операбельно связанной с NOS-терминатором.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, СОДЕРЖАЩИХСЯ В ПЕРЕЧНЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 - нуклеотидная последовательность 20L-8a.
SEQ ID NO: 2 - последовательность 20L-8a, кодированная SEQ ID NO: 1.
SEQ ID NO: 3 - нуклеотидная последовательность FR8a.
SEQ ID NO: 4 - последовательность FR8a, кодированная SEQ ID NO: 3.
SEQ ID NO: 5 - нуклеотидная последовательность FRCG.
SEQ ID NO: 6 - последовательность FRCG, кодированная SEQ ID NO: 5,
SEQ ID NO: 7 - нуклеотидная последовательность FR8a-9F.
SEQ ID NO: 8 - последовательность FR8a-9F, кодированная SEQ ID NO: 7.
SEQ ID NO: 9 - нуклеотидная последовательность FR-9F-catg.
SEQ ID NO: 10 - последовательность FR-9F-catg, кодированная SEQ ID NO: 9.
SEQ ID NO: 11 - нуклеотидная последовательность FR8a-12AA. SEQ ID NO: 12 - последовательность FR8a-12AA, кодированная SEQ ID NO: 11.
SEQ ID NO: 13 - нуклеотидная последовательность WR-9mut.
SEQ ID NO: 14 - последовательность WR-9mut, кодированная SEQ ID NO: 13.
SEQ ID NO: 15 - нуклеотидная последовательность FRD3.
SEQ ID NO: 16 - последовательность FRD3 кодированная SEQ ID NO: 15.
SEQ ID NO: 17 - нуклеотидная последовательность FR-12-cg-dm3.
SEQ ID NO: 18 - последовательность FR-12-cg-dm3, кодированная SEQ ID NO: 17.
SEQ ID NO: 19 - нуклеотидная последовательность 9F-cg-del6.
SEQ ID NO: 20 - последовательность 9F-cg-del6, кодированная SEQ ID NO: 19.
SEQ ID NO: 21 - нуклеотидная последовательность FR-cg-dm3.
SEQ ID NO: 22 - последовательность FR-cg-dm3, кодированная SEQ ID NO: 21.
SEQ ID NO: 23 - нуклеотидная последовательность 9F-cg-dm3.
SEQ ID NO: 24 - последовательность 9F-cg-dm3, кодированная SEQ ID NO: 23.
SEQ ID NO: 25 - нуклеотидная последовательность В8а.
SEQ ID NO: 26 - последовательность В8а, кодированная SEQ ID NO: 25.
SEQ ID NO; 27 - нуклеотидная последовательность 5*В8а.
SEQ ID NO: 28 - последовательность 5*B8a, кодированная SEQ ID NO: 27.
SEQ ID NO: 29- нуклеотидная последовательность V3A.
SEQ ID NO: 30 - последовательность V3A, кодированная SEQ ID NO: 29.
SEQ ID NO: 31 - нуклеотидная последовательность V4F.
SEQ ID NO: 32 - последовательность V4F кодированная SEQ ID NO: 31.
SEQ ID NO: 33 - нуклеотидная последовательность 5*V4F.
SEQ ID NO: 34 - последовательность 5*V4F, кодированная SEQ ID NO: 33.
SEQ ID NO: 35 - нуклеотидная последовательность 2OL-7.
SEQ ID NO: 36 - последовательность 20L-7, кодированная SEQ ID NO: 35.
SEQ ID NO: 37 - нуклеотидная последовательность Т7-2OL-7.
SEQ ID NO: 38 - последовательность T7-20L-7, кодированная SEQ ID NO: 37.
SEQ ID NO: 39 - нуклеотидная последовательность 5*2OL-7.
SEQ ID NO: 40 - последовательность 5*20L-7, кодированная SEQ ID NO: 39.
SEQ ID NO: 41 - нуклеотидная последовательность 2OL-10.
SEQ ID NO: 42 - последовательность 20L-10, кодированная SEQ ID NO: 41.
SEQ ID NO: 43 - нуклеотидная последовательность 5*2OL-10.
SEQ ID NO: 44 - последовательность 5*2OL-10 кодированная SEQ ID NO: 43.
SEQ ID NO: 45 - нуклеотидная последовательность 2OL-12A.
SEQ ID NO: 46 - последовательность 20L-12A, кодированная SEQ ID NO: 45.
SEQ ID NO: 47 - нуклеотидная последовательность 2OL-13.
SEQ ID NO: 48 - последовательность 201-13, кодированная SEQ ID NO: 47.
SEQ ID NO: 49 - нуклеотидная последовательность V5 и 6.
SEQ ID NO: 50 - последовательность У5и6, кодированная SEQ ID NO: 49.
SEQ ID NO: 51 - нуклеотидная последовательность 5*V5 и 6.
SEQ ID NO: 52 - последовательность 5*У5и6, кодированная SEQ ID NO: 51.
SEQ ID NO: 53 - нуклеотидная последовательность 88A-dm3.
SEQ ID NO: 54 - последовательность 88A-dm3, кодированная SEQ ID NO: 53.
SEQ ID NO: 55 - нуклеотидная последовательность FR(1Fa).
SEQ ID NO: 56 - последовательность FR(1Fa), кодированная SEQ ID NO: 55.
SEQ ID NO: 57 - нуклеотидная последовательность FR(1Ac).
SEQ ID NO: 58 - последовательность FR(1Ac), кодированная SEQ ID NO: 57.
SEQ ID NO: 59 - нуклеотидная последовательность FR(1Ia).
SEQ ID NO: 60 - последовательность FR(1Ia), кодированная SEQ ID NO: 59.
SEQ ID NO: 61 - нуклеотидная последовательность DM2 ЗА.
SEQ ID NO: 62 - последовательность DM23A, кодированная SEQ ID NO: 61.
SEQ ID NO: 63 - нуклеотидная последовательность 8AF.
SEQ ID NO: 64- последовательность 8AF, кодированная SEQ ID NO: 63.
SEQ ID NO: 65 - нуклеотидная последовательность 5*cry3A055.
SEQ ID NO: 66 - последовательность 5*Cry3A055, кодированная SEQ ID NO: 65.
SEQ ID NO: 67 - оптимизированная для кукурузы нуклеотидная последовательность cry3A.
SEQ ID NO: 68 - последовательность Cry2A, кодированная SEQ ID NO: 67.
SEQ ID NO: 69 - нуклеотидная последовательность cry3A055.
SEQ ID NO: 70 - последовательность Cry3A055, кодированная SEQ ID NO: 69.
SEQ ID NO: 71 - оптимизированная для кукурузы нуклеотидная последовательность cry1Ab.
SEQ ID NO: 72 - последовательность Cry1Ab, кодированная SEQ ID NO: 71.
SEQ ID NO: 73 - оптимизированная для кукурузы нуклеотидная последовательность cry1Ba.
SEQ ID NO: 74 - последовательность Cry1Ba, кодированная SEQ ID NO: 73.
SEQ ID NO: 75 - оптимизированная для кукурузы нуклеотидная последовательность cry1Fa.
SEQ ID NO: 76 - последовательность Cry1Fa, кодированная SEQ ID NO: 75.
SEQ ID NO: 77 - нуклеотидная последовательность cry8Aa.
SEQ ID NO: 78 - последовательность Cry8Aa, кодированная SEQ ID NO: 77.
SEQ ID NO: 79 - нуклеотидная последовательность cry1Ac.
SEQ ID NO: 80 - последовательность Cry1Ac кодированная SEQ ID NO: 79.
SEQ ID NO: 81 - нуклеотидная последовательность cry1Ia.
SEQ ID NO: 82 - последовательность Cry На, кодированная SEQ ID NO: 81.
SEQ ID NO: 83-125 - праймерные последовательности, применяемые в изобретении.
SEQ ID NO: 126-134 - N-концевые пептидильные фрагменты.
SEQ ID NO: 135 - полноразмерный белок Cry3A.
SEQ ID NO: 136-143 - праймерные последовательности, применяемые в изобретении.
SEQ ID NO: 144 - кодирующая последовательность T7-8AF.
SEQ ID NO: 145 - последовательность T7-8AF, кодированная ASEQ ID NO: 144.
SEQ ID NO: 146 - кодирующая последовательность -catGSAF.
SEQ ID NO: 147 - последовательность -CatG8AF, кодированная SEQ ID NO: 146.
SEQ ID NO: 148 - кодирующая последовательность 8AFdm3.
SEQ ID NO: 149 - последовательность 8AFdm3, кодированная SEQ ID NO: 148.
SEQ ID NO: 150 - кодирующая последовательность 8AFlongdm3.
SEQ ID NO: 151 - последовательность 8AFlongdm3, кодированная SEQ ID NO: 150.
SEQ ID NO: 152 - кодирующая последовательность cap8AFdm3.
SEQ ID NO: 153 - последовательность cap8AFdm3, кодированная SEQ ID NO: 152.
SEQ ID NO: 154 - кодирующая последовательность 8AFdm3T.
SEQ ID NO: 155 - последовательность 8AFdm3T, кодированная SEQ ID NO: 154.
SEQ ID NO: 156 - кодирующая последовательность 8AFlongdm3T.
SEQ ID NO: 157 - последовательность 8AFlongdm3T, кодированная SEQ ID NO: 156.
SEQ ID NO: 158 - кодирующая последовательность cap8AFdm3T.
SEQ ID NO: 159 - последовательность cap8AFdm3T, кодированная SEQ ID NO: 158.
SEQ ID NO: 160 -FR8a+34 сГИБ.
ОПРЕДЕЛЕНИЯ
Для внесения ясности ниже поданы определения некоторых терминов, используемых в настоящем описании:
«Активность» сГИБов по настоящему изобретению означает, что данные сГИБы функционируют как орально активные средства борьбы с насекомыми, обладают токсическим эффектом и способны нарушить или затруднить питание насекомого, что может приводить или не приводить к гибели насекомого. Если сГИБ по настоящему изобретению доставлен в организм насекомого, то результатом в типичном случае является гибель насекомого или невозможность насекомого питаться источником, из которого сГИБ попадает в его организм.
Термин «ассоциированные с / операбельно связанные» относится к двум нуклеиновым кислотам, связанным друг с другом физически или функционально. Например, говорят, что промотор или регуляторная последовательность ДНК «ассоциированы с» последовательностью ДНК, кодирующей РНК или белком, в том случае, если эти две последовательности связаны оперативно или расположены так, что регуляторная последовательность ДНК будет воздействовать на уровень экспрессии кодирующей или структурной последовательности ДНК.
В контексте настоящего изобретения «химерный инсектицидный белок» (ХИБ) представляет собой инсектицидный белок, содержащий пептидильный фрагмент, который ввели путем слияния в N-концевую часть сГИБа. Этот пептидильный фрагмент может наделять данный сГИБ инсектицидной активностью, может делать активность сГИБа выше, чем у сГИБа, не имеющего такого пептидильного фрагмента, или же может сделать сГИБ более стабильным, чем сГИБ без пептидильного фрагмента, в частности против, по меньшей мере, западного кукурузного корневого жука. Пептидильный фрагмент представляет собой последовательность аминокислот, которая в типичном случае гетерологична Bt Cr-белку (не получена из него), но может быть и полученной из Bt Cry-белка. Такие пептидильные фрагменты вытянуты за N-концевую часть инсектицидного белка и в природе не встречаются в N-концевой части Bt Cry-белков. Одним из примеров N-концевого пептидильного фрагмента является последовательность аминокислот MTSNGRQCAGIRP (SEQ ID NO: 129), которая не получена из Bt Cry-белка.
«Кодирующая последовательность» - это нуклеиново-кислотная последовательность, транскрибированная в РНК, такую как мРНК, рРНК, тРНК, мяРНК, смысловая РНК или антисмысловая РНК. Желательно, чтобы затем эта РНК транслировалась в организме для производства белка.
В контексте настоящего изобретения «связывание» нуклеиновых кислот означает, что две или несколько нуклеиновых кислот соединяют вместе при помощи любых способов, известных в данной отрасли. Для примера, не являющегося ограничивающим, скажем, что нуклеиновые кислоты можно лигировать вместе при помощи, например, ДНК-лигазы или соединять отжигом при помощи ПЦР, Нуклеиновые кислоты также можно соединять путем химического синтеза нуклеиновой кислоты, используя последовательность из двух или нескольких отдельных нуклеиновых кислот.
«Борьба» с насекомыми означает препятствование посредством токсического воздействия способности насекомых-вредителей выживать, расти, питаться и/или размножаться или ограничение ущерба или потери урожая злаков, Вызываемых насекомыми. «Борьба» с насекомыми может означать или не означать уничтожение их, хотя в предпочтительном варианте она означает уничтожение насекомых.
В контексте настоящего изобретения термин «соответствует» означает, что когда последовательность аминокислот определенных белков (например, Bt Cry-белков или модифицированных Cry3A-белков) совместить друг с другом, то аминокислоты, которые совпадают с определенными пронумерованными позициями, например, в токсине Cry3A (либо SEQ ID NO: 68, либо SEQ ID NO: 134); в токсине Cry3A055 (SEQ ID NO: 70); или в токсине Cry1Ab (SEQ ID NO: 72), хотя не обязательно они должны находиться точно в этих пронумерованных позициях относительно ссылочной последовательности аминокислот, особенно по отношению к идентификации доменов I, II and III, а также/или консервативных блоков и вариабельных участков, то позиции этих аминокислот «соответствуют» друг другу. Например, в схеме домена I гибридного белка аминокислоты 11-244 белка Cry3A055 (SEQ ID NO: 70) соответствуют аминокислотам 58-290 нативного белка Cry3A (SEQ ID NO: 135) или аминокислотам 11-243 нативного белка Cry3A (SEQ ID NO: 68) или аминокислотам 33-254 нативного белка Cry1Ab.
В контексте настоящего изобретения слова «Cry-белок» могут использоваться как взаимозаменяющие со словами «дельта-эндотоксин» или «δ-эндотоксин».
В контексте настоящего изобретения «сконструированный гибридный инсектицидный белок» (сГИБ) представляет собой инсектицидный белок, созданный путем слияния уникальных комбинаций вариабельных участков и консервативных блоков из, по меньшей мере, двух различных Cry-белков. Такие вновь созданные сГИБы могут содержать полные или неполные вариабельные участки, консервативные блоки или домены из модифицированного белка Cry3A и из Cry-белка, отличающегося от модифицированного белка Cry3A. СГИБы по настоящему изобретению могут, по выбору, включать в себя протоксинный хвостовой участок из Bt Cry-белка, или N-концевой пептидильный фрагмент, или и то и другое. В не ограничивающем примере сГИБ создан путем комбинирования (в направлении от N-конца до С-конца) аминокислотами 1-468 белка Cry3A055 (SEQ ID NO: 70), что включает вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислотами 477-648 белка Cry1Ab (SEQ ID NO: 72), что включает в себя C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6, а также участком из 38 аминокислот хвостового участка протоксина Cry1Ab. Сконструированный ГИБ, содержащий N-концевой пептидильный фрагмент можно также называть «химерным инсектицидным белком» (ХИБ).
«Доставить» сГИБ означает, что данный сГИБ вступает в контакт с насекомым, результатом чего является токсичное воздействие и борьба с насекомым. Доставка сГИБ может осуществляться различными путями, например, путем экспрессии сГИБ трансгенным растением, с помощью составленной белковой композиции (композиций), распыляемыми белковыми композициями (композицией), с помощью приманочной матрицы или с помощью любых других, известных в отрасли систем доставки токсинов.
«Количество, эффективное для борьбы с насекомыми» означает такую концентрацию сГИБа, которая путем токсичного воздействия препятствует способности насекомых выживать, расти, питаться и/или размножаться, или ограничивает причиняемые насекомыми ущерб или потерю урожая злаков. «Количество, эффективное для борьбы с насекомыми» может означать или не означать уничтожение насекомых, хотя предпочтительно, чтобы оно означало уничтожение насекомых.
Термин «экспрессионная кассета» здесь используется в значении последовательности нуклеиновых кислот, способной направлять экспрессию конкретной нуклеотидной последовательности в соответствующую клетку-хозяин, содержащей промотор, операбельно связанный со значимой нуклеотидной последовательностью, операбельно связанной с сигналами терминации. В типичном случае она также содержит последовательности, требуемые для надлежащей трансляции значимой нуклеотидной последовательности. По меньшей мере, один из компонентов экспрессионной кассеты, содержащей значимую нуклеотидную последовательность, может быть гетерологичным по отношению к, по меньшей мере, одному из других ее компонентов. Эта экспрессионная кассета может быть такой, которая встречается в природе, но была получена в рекомбинантной форме, полезной для гетерологичной экспрессии. Однако в типичном случае экспрессионная кассета является гетерологичной по отношению к хозяину, т.е., конкретная последовательность нуклеиновых кислот этой экспрессионной кассеты не встречается в природе в клетке-хозяине, и должна вводиться в клетку-хозяин или в предшественник клетки-хозяина путем трансформации. Экспрессия нуклеотидной последовательности может находиться под управлением конституитивного промотора или индуцируемого промотора, который инициирует транскрипцию только тогда, когда клетка-хозяин открыта для определенного внешнего стимула. В случае многоклеточного организма, например, растения, промотор может также обладать специфичностью к конкретной ткани, или органу, или к стадии развития.
«Ген» - это определенная область, расположенная внутри генома, которая, кроме вышеупомянутой кодирующей последовательности нуклеиновых кислот, содержит также другие, главным образом регуляторные, нуклеиновые кислоты, ответственные за управление экспрессией, то есть, так сказать за транскрипцию и трансляцию кодирующей части. Ген может также содержать другие 5' и 3' нетранслируемые последовательности и терминирующие последовательности. Другими присутствующими элементами могут быть, например, интроны. Регуляторная последовательность нуклеиновых кислот в гене может не быть нормально оперативно связанной с ассоциированной последовательностью нуклеиновых кислот, как это встречается в природе, и в таком случае этот ген будет химерным геном.
Термин «значимый ген» относится к любому гену, который при перемещении в растение наделяет это растение желаемыми характеристиками, такими как устойчивость к антибиотикам, сопротивляемость вирусам, сопротивляемость насекомым, сопротивляемость болезням или сопротивляемость другим вредителям, толерантность к гербицидам, повышение питательного значения, улучшение промышленно значимых качеств или измененная репродуктивная способность, «Значимым геном» может также быть ген, который пересаживают в растения для выработки коммерчески ценных энзимов и метаболитов в таком растении.
«Гетерологичная» нуклеиново-кислотная последовательность представляет собой нуклеиново-кислотную последовательность, не являющуюся натурально ассоциированной с клеткой-хозяином, в которую ее ввели, включая искусственные множественные копии природной нуклеиново-кислотной последовательности. Гетерологичная последовательность аминокислот - это такая, которая не является натурально ассоциированной с нативной последовательностью аминокислот, например, с последовательностью аминокислот Cry-белка.
«Гомологичная» нуклеиново-кислотная последовательность - это нуклеиново-кислотная последовательность, натурально ассоциированная с клеткой-хозяином, в которую ее ввели.
«Гомологичная рекомбинация» - это взаимный обмен фрагментами нуклеиновых кислот между гомологичными молекулами нуклеиновых кислот.
Термин «идентичность» или «процент идентичности» относится к степени подобия между двумя нуклеиново-кислотными или белковыми последовательностями. Для сравнения последовательностей в типичном случае одна последовательность служит ссылочной (эталонной), с которой сравниваются тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестовая и ссылочная последовательности вводятся в компьютер, при необходимости задаются координаты подпоследовательности, а также задаются параметры программы алгоритма. Затем алгоритм сравнения последовательностей вычисляет процент идентичности тестовой последовательности (последовательностей) по отношению к ссылочной последовательности на основе заданных параметров программы.
Оптимальное совмещение последовательностей для сравнения может производиться, например, с помощью алгоритма локальной гомологии, представленного в работе Smith и Waterman, Adv. Appl. Math. 2: 482 (1981), алгоритма совмещения гомологии, представленного в работе Needleman и Wunsch, J. Mol. Biol. 48: 443 (1970), способом поиска подобности, представленным в работе Pearson и Lipman, Proc. Natl. Acad. Sci. USA 85: 2444 (1988), компьютерным воплощением этих алгоритмов (GAP, BESTFIT, FASTA, и TFASTA в пакете программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI), или путем визуальной проверки (см. в целом, Ausubel и др., ниже).
Одним из примеров алгоритма, пригодного для определения процента идентичности последовательности и подобности последовательностей является алгоритм BLAST, описанный в работе Altschul и др., J. Mol. Biol. 215: 403-410 (1990). Программное обеспечение для выполнения анализов по BLAST можно получить через национальный центр информации по биотехнологии (http://www.ncbi.nlm.nih.gov/). Этот алгоритм включает в себя, прежде всего нахождение пар с наиболее высокой степенью идентичности (ПВИ) путем идентификации коротких слов длиной W в тестируемой последовательности, которые либо полностью совпадают либо удовлетворяют некоторому пороговому положительному значению Т при совмещении со словом такой же длины из последовательности, полученной в базе. Т - это пороговое значение близости слова (Altschul и др., 1990). Эти первоначальные нахождения близости слов (совпадений) служат затравкой для инициации поиска более длинных ПВИ, содержащих эти слова. Затем эти совпадения слов расширяются в обоих направлениях вдоль каждой последовательности настолько далеко, насколько может увеличиваться совокупное значение баллов за совпадения. Совокупные значения вычисляются при помощи (для нуклеотидных последовательностей) параметров М (премиальный балл, начисляемый за пару совпадающих остатков; он всегда > 0) и N (штрафной бал за несовпадение остатков; он всегда < 0). Для вычисления совокупного значения по последовательностям аминокислот применяется матрица начисления баллов. Расширение совпадений слов в каждом направлении останавливается тогда, когда совокупное значение баллов за совпадения падает от максимального достигнутого значения на величину X, когда совокупное значение счета падает до нуля или ниже нуля вследствие накопления одного или нескольких отрицательных результатов совпадения, или же при достижении конца любой из последовательностей. Параметры W, Т, и Х алгоритма BLAST определяют чувствительность и скорость совмещения. В программе BLASTN (для нуклеотидных последовательностей) по умолчанию длина слова (W) принимается равной 11, ожидаемое значение (Е) равным 10, падение (отсечка) равным 100, М=5, N=-4, и сравнение выполняется по обеим цепочкам. Для последовательностей аминокислот программа BLASTP по умолчанию принимает длину слова (W) равной 3, ожидаемое значение (Е) равным 10, а также использует матрицу начисления баллов BLOSUM62 (см. НешкоПи Henikoff, Proc. Nati. Acad. Sci. USA 89: 10915 (1989)).
Кроме вычисления процента идентичности последовательности алгоритм BLAST также выполняет статистический анализ подобности между двумя последовательностями (см. напр., Karlin и Altschul, Proc. Natl. Acad. Sci. USA 90: 5873-5787 (1993)). Одной из величин определения подобности, предоставляемой алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), показывающая вероятность, с которой совпадение между двумя нуклеотидными или аминокислотными последовательностями может произойти случайно. Например, тестируемая последовательность нуклеиновых кислот считается подобной ссылочной последовательности, если наименьшая суммарная вероятность при сравнении тестовой последовательности нуклеиновых кислот со ссылочной последовательностью нуклеиновых кислот меньше 0,1, более предпочтительно меньше чем 0,01, а наиболее предпочтительно меньше, чем 0,001.
Другой широко используемой и общепринятой компьютерной программой сравнения последовательностей является CLUSTALW v1.6 (Thompson, и др. Nuc. Acids Res., 22: 4673-4680, 1994). Число совпадающих оснований или аминокислот делится на общее число оснований или аминокислот и умножается на 100 для получения процента идентичности. Например, если у двух последовательностей из 580 пар оснований совпадают 145 оснований, то эти последовательности будут идентичными на 25 процентов. Если две сравниваемые последовательности имеют разную длину, то число совпадений делится на меньшее из двух значений длины. Например, если при сравнении белков из 200 и 400 аминокислот совпали 100 аминокислот, то идентичность их составляет 50 процентов по отношению к более короткой последовательности. Если более короткая последовательность содержит меньше, чем 150 оснований или 50 аминокислот, то число совпадений делится на 150 (для оснований нуклеиновых кислот) или на 50 (для аминокислот) и умножается на 100 для получения процента идентичности.
Другим показателем того, что две нуклеиновых кислоты являются в целом идентичными, служит то, что эти две молекулы гибридизуются друг к другу в жестких условиях. Фраза «гибридизуется специфично к» относится к связыванию, дуплексированию или гибридизации молекулы только к конкретной нуклеотидной последовательности в жестких условиях, когда эта последовательность присутствует в сложной смеси (напр., полностью клеточной) ДНК или РНК. Фраза «связывается в целом» относится к комплементарной гибридизации между нуклеиновой кислотой-зондом и нуклеиновой кислотой-мишенью и охватывает мелкие несовпадения, с которыми можно справиться путем снижения жесткости среды гибридизации для того, чтобы достичь требуемого обнаружения нуклеиново-кислотной последовательности-мишени.
«Жесткие условия гибридизации» и «жесткие условия гибридизационной отмывки» в контексте экспериментов по гибридизации нуклеиновых кислот, таких как Южная и Северная гибридизация, являются зависимыми от последовательности и отличаются при различных параметрах окружающей среды. Длинные последовательности гибридизуются специфично при повышенных температурах. Обширная информация по гибридизации нуклеиновых кислот содержится в работе Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes часть I глава 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays" Elsevier, Нью Йорк. В целом, температура для гибридизации и отмывки в условиях высокой жесткости выбирается приблизительно на 5°C ниже температуры плавления (Tпл) для конкретной последовательности при определенных ионной силе и уровне pH. В типичном случае, при «жестких условиях» зонд будет гибридизироваться к своей последовательности-мишени, но ни к каким другим последовательностям.
Температура Tпл представляет собой температуру (при определенных значениях ионной силы и pH) при которой 50% последовательности-мишени гибридизируется к идеально совпадающему зонду. Для очень жестких условий температура выбирается равной Tпл для конкретного зонда. Пример жестких условий гибридизации для гибридизации комплементарных нуклеиновых кислот, содержащих более 100 комплементарных остатков, на фильтре в Южному или северному блоттинге представляет собой: 50% формамид с 1 мг гепарина при 42°C, гибридизация выполняется в течение ночи. Пример отмывки в условиях высокой жесткости: 0,1 5М NaCl при 72°C приблизительно в течение 15 минут. Пример жестких условий отмывки: двукратная отмывка в стандартном растворе SSC при 65°C в течение 15 мин. (описание буфера SSC см., Sambrook, ниже). Часто отмывке в условиях высокой жесткости предшествует отмывка в условиях низкой жесткости для удаления исходного зондового сигнала. Пример отмывки в условиях средней жесткости для дуплекса, содержащего, например, более 100 нуклеотидов: 1х SSC при 45°C в течение 15 минут. Пример отмывки в условиях низкой жесткости для дуплекса, содержащего, например, более 100 нуклеотидов: 4-6х SSC при 40°C в течение 15 минут. Для коротких зондов (напр. приблизительно от 10 до 50 нуклеотидов) жесткие условия в типичном случае означают, что концентрации соли составляют менее 1,0 М ионов Na, в типичном случае приблизительно от 0,01 до 1,0 М ионов Na (или других солей) при уровне pH от 7,0 до 8,3, и типичном значении температуры, по меньшей мере, приблизительно 30°C. Жесткие условия также могут быть достигнуты путем добавления дестабилизирующих агентов, например, формамида. В общем случае, отношение сигнала к шуму, превышающее в 2 (или большее число) раза значение, полученное для неопределенного зонда в данном конкретном образце для гибридизации, свидетельствует об обнаружении специфической гибридизации. Нуклеиновые кислоты, не гибридизирующиеся друг к другу в жестких условиях, тем не менее являются в целом идентичными, если в целом идентичны белки, кодирующиеся ими. Такое происходит, например, если копия нуклеиновой кислоты создается с использованием максимальной дегенерации кодонов, допустимой данным генетическим кодом.
Ниже приведены примеры наборов условий гибридизации/отмывки, которые могут использоваться для клонирования гомологичных нуклеотидных последовательностей, являющихся в целом идентичными ссылочными нуклеотидными последовательностями по настоящему изобретению: ссылочная нуклеотидная последовательность в оптимальном варианте гибридизируется к ссылочной нуклеотидной последовательности в 7% додецил сульфата натрия (ДСН), 0,5 М NaPO4, 1 мМ ЭДТА при 50°C с отмывкой: 2Х SSC, 0,1% ДСН при 50°C, более предпочтительно в 7% додецил сульфата натрия (ДСН), 0,5 М NaPO4, 1 мМ ЭДТА при 50°C с отмывкой: IX SSC, 0,1% ДСН при 50°C, еще более предпочтительно в 7% додецил сульфата натрия (ДСН), 0,5 М NaPO4, 1 мМ ЭДТА при 50°С с отмывкой: 0,5Х SSC, 0,1% ДСН при 50°C, предпочтительно в 7% додецил сульфата натрия (ДСН), 0,5 М NaPO4, 1 мМ ЭДТА при 50°C с отмывкой: 0,1Х SSC, 0,1% ДСН при 50°C, еще более предпочтительно в7% додецил сульфата натрия (ДСН), 0,5 М NaPO4, 1 мМ ЭДТА при 50°C с отмывкой: 0,1Х SSC, 0,1% ДСН при 65°C.
Дальнейшим свидетельством того, что две нуклеиновые кислоты или два белка являются существенно идентичными, служит то, что белок, кодированный первой нуклеиновой кислотой, является иммунологически кросс-реактивным с белком, кодированным второй нуклеиновой кислотой, или специфически связывается с ним. Таким образом, в типичном случае один белок является существенно идентичным другому белку, если эти два белка отличаются только консервативными заменами.
Термин «инсектицидный» означает токсичную биологическую активность, способную осуществлять борьбу с насекомыми, предпочтительно уничтожая их.
Последовательность нуклеиновых кислот является «изокодированной с» ссылочной последовательностью в том случае, если эта последовательность нуклеиновых кислот кодирует полипептид, имеющий такую же последовательность аминокислот, как и полипептид, кодированный ссылочной последовательностью нуклеиновых кислот.
«Изолированная» молекула нуклеиновой кислоты или изолированный токсин представляют собой молекулу нуклеиновой кислоты или токсин, которые вследствие действий человека существуют отдельно от своей естественной среды, а поэтому не являются продуктом природы. Изолированная молекула нуклеиновой кислоты или изолированный токсин могут существовать в очищенной форме или могут существовать в не естественной среде, например (что не означает ограничений) такой, как рекомбинантная микробная клетка, растительная клетка, растительная ткань или растение.
«Модифицированный токсин Cry3A» или «Cry3A055» по настоящему
изобретению относится к токсину, полученному из Cry3A и имеющему, по меньшей мере один дополнительный сайт узнавания протеазы, который распознается протеазой кишечника насекомого-мишени и не встречается в природе в токсине Cry3A, как описано в патенте США 7,030,295, включенном сюда путем ссылки.
«Модифицированная кодирующая последовательность Cry3A» согласно настоящему изобретению может быть получена из нативной кодирующей последовательности cry3A или синтетической последовательности cry3A и содержит кодирующую последовательность, по меньшей мере, одного дополнительного сайта узнавания протеазы, которая не встречается натурально в немодифицированном гене cry3A.
«Молекула нуклеиновой кислоты» или «нуклеиново-кислотная последовательность» представляет собой сегмент одно- или двухцепочечной ДНК или РНК, который может быть изолирован из любого источника. В контексте настоящего изобретения молекула нуклеиновой кислоты в типичном случае является сегментом ДНК.
Термин «растение» относится к любому растению на любой стадии развития, в особенности, к семенному растению.
«Растительная клетка» представляет собой структурную и физиологическую единицу растения, состоящую из протопласта и клеточной стенки. Растительная клетка может быть в форме изолированной единичной клетки или культивированной клетки, либо являться частью более высокоорганизованной единицы, например, растительной ткани, органа растения или целого растения.
«Культура растительной клетки» означает культуры структурных единиц растения, таких как, например, протопласты, клетки клеточных культур, клетки в растительных тканях, пыльца, пыльцевые трубки, яйцеклетки, зародышевые мешки, зиготы и зародыши на различных стадиях развития.
«Растительный материал» относится к листьям, стеблям, корням, цветам или частям цветов, фруктам, пыльце, яйцевым клеткам, зиготам, семенам, срезам, клеточным культура или культурам ткани или к любой части или продукту растения.
«Растительный орган» - это различимая и визуально структурированная и дифференцированная часть растения, такая как корень, стебель, лист, бутон цветка или зародыш.
Термин «растительная ткань» здесь означает группу растительных клеток, организованную в структурную и функциональную единицу. Сюда включается любая ткань растения in planta или в культуре. Этот термин включает в себя (не ограничивая) целые растения, органы растений, семена растений, культуры ткани и любые группы растительных клеток, организованные в структурные и/или функциональные единицы. Использование этого термина в совокупности с (или в отсутствии) каким-либо конкретным типом растительной ткани, перечисленной выше или иным образом подпадающей под это определение, не имеет своей целью исключение любого другого типа растительной ткани.
«Промотор» - нетранслированная последовательность ДНК, находящаяся до кодирующей области и содержащая место для связывания РНК-полимеразы, а также инициирующая транскрипцию ДНК. Область промотора может также включать в себя другие элементы, работающие регуляторами экспрессии генов.
Термин «регуляторные элементы» относится к последовательностям, включенным в управление экспрессией нуклеотидной последовательности. Регуляторные элементы включают в себя промотор, операбельно связанный со значимой нуклеотидной последовательностью, и терминирующие сигналы. В типичном случае они также охватывают последовательности, требуемые для надлежащей трансляции нуклеотидной последовательности.
«Трансформация» - это процесс для введения гетерологичной нуклеиновой кислоты в клетку-хозяин или в организм. В частности, «трансформация» означает устойчивую интеграцию молекулы ДНК в геном интересующего организма.
«Трансформированный/трансгенный/рекомбинантный» относится к организму-хозяину, такому как бактерия или растение, в который ввели гетерологичную молекулу нуклеиновой кислоты. Эта молекула нуклеиновой кислоты может быть устойчиво интегрирована в геном хозяина, или же эта молекула нуклеиновой кислоты также может присутствовать как внехромосомная молекула. Такая внехромосомная молекула может быть способна к саморепликации. Следует понимать, что к трансформированным клеткам, тканям или растениям относятся не только конечные продукты процесса трансформации, но также и их трансгенное потомство. Термины «не трансформированный», «не трансгенный» или «не рекомбинантный» относятся к природному организму-хозяину, например, к бактерии или растению, который не содержит гетерологичной молекулы нуклеиновой кислоты.
Нуклеотиды обозначаются по их основаниям следующими стандартными сокращенными обозначениями: аденин (А), цитозин (С), тимин (Т) и гуанин (G). Аналогичным образом аминокислоты обозначаются следующими стандартными сокращенными обозначениями: аланин (А1а; А), аргинин (Arg; R), аспарагин (Asn; N), аспарагиновая кислота (Asp; D), цистеин (Cys; С), глутамин (Gin; Q), глутаминовая кислота (Glu; Е), глицин (Gly; G), гистидин (His; Н), изолейцин (Не; 1), лейцин (Leu; L), лизин (Lys; К), метионин (Met; М), фенилаланин (Phe; F), пролин (Pro; Р), серии (Ser;
S), треонин (Thr; Т), триптофан (Trp; W), тирозин (Туг; Y), и валин (Val; V).
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новьм сконструированным гибридным инсектицидным белкам (сГИБам), созданными для того, чтобы обладать активностью, по меньшей мере, против западного кукурузного корневого жука, а также могут быть активными против северного кукурузного корневого жука, мексиканского кукурузного корневого жука и/или колорадского картофельного жука. Некоторые сГИБы обладают активностью против чешуекрылых вредителей, европейского кукурузного мотылька. Такие новые сГИБы производятся путем слияния уникальных комбинаций полных или неполных вариабельных участков и консервативных блоков, по меньшей мере, двух разных Cry-белков и, по выбору, включают в себя протоксинную хвостовую область из Bt Cry-белка на C-концевой части, или N-концевой пептидильный фрагмент, или и то и другое. Неограничивающим примером может служить комбинирование полных или неполных вариабельных участков и консервативных блоков из первого Cry-белка, обладающего активностью против жесткокрылых, с полными или неполными вариабельными участками и консервативными блоками из второго Cry-белка, обладающего активностью против чешуекрылых и отличающегося от первого Bt Cry-белка, а также включение, по выбору, протоксинного хвостового участка из Bt Cry-белка на C-концевой части, или N-концевого пептидильного фрагмента, или того и другого, в результате чего создается новый сконструированный гибридный инсектицидный белок, обладающий активностью против целого спектра насекомых и отличающийся от первого или второго родительских Cry-белков, или от обоих. Такие новые сГИБы могут также содержать полные или неполные вариабельные участки, консервативные блоки или домены из модифицированного Cry3A-белка и Cry-белка, отличающегося от модифицированного Cry3A-белка. N-концевой пептидильный фрагмент или протоксинный хвостовой участок могут наделять сГИБ инсектицидной активностью, могут делать инсектицидную активность сГИБа более высокой, чем у сГИБа, не содержащего пептидильного фрагмента или протоксинного хвостового участка, и/или может делать сГИБ более стабильным, чем сГИБ, не содержащий пептидильного фрагмента или протоксинного хвостового участка, в особенности против, по меньшей мере, западного кукурузного корневого жука. Последовательность аминокислот пептидильного фрагмента в типичном случае является гетерологической к Bt Cry-белку (т.е., не полученной из него). Однако на основе изложенного здесь материала квалифицированный специалист увидит, что N-концевой пептидильный фрагмент может быть создан с применением последовательности аминокислот, полученной из Bt Cry-белка. Оказалось, что сГИБы по настоящему изобретению обладают неожиданно удивительной токсичностью по отношению к кукурузному корневому жуку, особенно к западному, северному и мексиканскому кукурузным жукам. Настоящее изобретение также относится к нуклеиновым кислотам, результатом экспрессии которых являются сГИБы, а также к производству и применению сГИБов для борьбы с насекомыми-вредителями. Результатом экспрессии этих нуклеиновых кислот являются сГИБы, которые можно применять для борьбы с жесткокрылыми насекомыми, такими как западный, северный или мексиканский кукурузные корневые жуки, либо применять для борьбы с чешуекрылыми насекомыми, такими как европейский кукурузный мотылек, особенно, когда они экспрессированы в трансгенное растение, такое как трансгенная кукуруза.
В одном исполнении настоящее изобретение охватывает сконструированный гибридный инсектицидный белок, содержащий последовательность аминокислот из первого Bacillus thuringiensis (Bt) Cry-белка, включающую полные или неполные вариабельные участки и консервативные блоки первого Cry-белка, слитую с последовательностью аминокислот из второго Bt Cry-белка, отличающегося от первого Bt Cry-белка, включающей полные или неполные вариабельные участки и консервативные блоки второго Cry-белка, и, по выбору, содержащий: (а) протоксинный хвостовой участок Bt Cry-белка, расположенный на С-конце; или (б) N-концевой пептидильный фрагмент, или как (а) так и (б), причем этот сГИБ обладает активностью против, по меньшей мере, западного кукурузного корневого жука.
В другом исполнении настоящее изобретение охватывает сГИБ, содержащий N-концевую часть первого Bt Cry-белка, слитую с С-концевой областью второго Bt Cry-белка, отличающегося от первого Bt Cry-белка, причем, по меньшей мере, одна позиция кроссинговера между первым и вторым Bt Cry-белками расположена в консервативном блоке 2, консервативном блоке 3, вариабельном участке 4 или консервативном блоке 4, а также, по выбору, содержащий (а) протоксинный хвостовой участок Bt Cry-белка, расположенный на С-конце; или (б) N-концевой пептидильный фрагмент, или как (а) так и (б), причем этот сГИБ обладает активностью против, по меньшей мере, западного кукурузного корневого жука.
В другом исполнении сГИБ по изобретению содержит (в направлении от N-конца до С-конца) вариабельный участок 1 или C-концевую часть вариабельного участка 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевую часть консервативного блока 3 первого Bt Cry-белка, срощенные с C-концевой частью консервативного блока 3, вариабельным участком 4, консервативным блоком 4, вариабельным участком 5, консервативным блоком 5 и вариабельным участком 6 второго Bt Cry-белка.
В другом исполнении сГИБ по изобретению содержит, по меньшей мере, две позиция кроссинговера между последовательностью аминокислот первого Bt Cry-белка и последовательностью аминокислот второго Bt Cry-белка. В одном исполнении первая позиция кроссинговера расположена в консервативном блоке 2, а вторая позиция кроссинговера расположена в консервативном блоке 3. В другом исполнении первое соединение кроссинговера расположено в консервативном блоке 3, а вторая позиция кроссинговера расположена в консервативном блоке 4.
В другом исполнении сГИБ по изобретению содержит в С-концевой части хвостовой участок Bt Cry-белка. Протоксинный хвостовой участок может наделять сГИБ инсектицидной активностью (это означает, что такой сГИБ без протоксинного хвостового участка не был бы активным), может делать активность сГИБа более высокой, чем у сГИБа без протоксинного хвостового участка, или может делать сГИБ более стабильным, чем сГИБ без протоксинного хвостового участка. В одном исполнении изобретения протоксинный хвостовой участок берется из Bt Cry-белка, обладающего активностью против чешуекрылых. В другом исполнении протоксинный хвостовой участок берется из Cry1A-белка. Еще в одном исполнении протоксинный хвостовой участок берется из белка Cry1Aa или Cry1Ab. Протоксинный хвостовой участок по настоящему изобретению может включать в себя целый протоксинный хвост Bt Cry-белка или любой его фрагмент. В одном аспекте этого исполнения протоксинный хвостовой участок сГИБа содержит, по меньшей мере, 38 аминокислот из N-конца протоксинного хвоста Cry1Ab-белка. В другом аспекте этого исполнения протоксинный хвостовой участок содержит последовательность аминокислот, соответствующую аминокислотам 611-648 последовательности SEQ ID NO: 72. Еще в одном аспекте этого исполнения протоксинный хвостовой участок содержит аминокислоты 611-648 последовательности SEQ ID NO: 72.
Еще в одном исполнении сГИБ содержит N-концевой пептидильный фрагмент. Этот N-концевой пептидильный фрагмент может наделять данный сГИБ инсектицидной активностью (это значит, что без N-концевого пептидильного фрагмента этот белок не будет обладать инсектицидной активностью), или же N-концевой пептидильный фрагмент может делать инсектицидную активность сГИБа более высокой, чем у сГИБа без N-концевого пептидильного фрагмента, или же N-концевой пептидильный фрагмент может делать сГИБ более стабильным, чем сГИБ без N-концевого пептидильного фрагмента. В одном аспекте этого исполнения пептидильный фрагмент содержит последовательность аминокислот, гетерологичную Bt Cry-белку (т.е., не полученную из него). В другом аспекте этого исполнения N-концевой пептидильный фрагмент содержит, по крайней мере, 9 аминокислот. В другом аспекте этого исполнения пептидильный фрагмент содержит, по меньшей мере, 10, 11, 12, 13, 14, 15, 16, 17,18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 70, 80, 90 или 100 аминокислот. В другом аспекте этого исполнения пептидильный фрагмент содержит более 100 аминокислот. Еще в другом аспекте этого исполнения N-концевой пептидильный фрагмент содержит последовательность аминокислот YDGRQQHRG (SEQ ID NO: 133) или TSNGRQCAGIRP (SEQ ID NO: 134). Еще в другом аспекте этого исполнения N-концевой пептидильный фрагмент содержит последовательность аминокислот, выбранную из группы, состоящей из последовательностей SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131 и SEQ ID NO: 132.
Еще в одном исполнении вариабельные участки и консервативные блоки первого Cry-белка, активного против жесткокрылых насекомых, для получения сГИБа по изобретению используются в комбинации с вариабельными участками и консервативными блоками второго Cry-белка, активного против чешуекрылого насекомого. Перечень Cry-белков, активных против жесткокрылых насекомых, включает в себя (не ограничивая) белки Cry3, Cry7, Cry8 и Cry34/Cry35. Перечень Cry-белков, активных против чешуекрылых насекомых, включает в себя (не ограничивая) белки Cry1 and Cry9. В одном аспекте этого исполнения первый Cry-белок представляет собой белок Cry3A, а второй Cry-белок - белок Cry1A. В другом аспекте белок Cry3A может быть заменен модифицированным Cry3A, например, белком Cry3A055, описанным в патенте США 5,659,123, включенном сюда путем ссылки. Еще в одном аспекте этого исполнения белком Cry3A является белок Cry3Aa, а белком Cry1A - белок Cry1Aa или Cry1Ab. В другом аспекте этого исполнения белок Cry3Aa выбирается из следующей группы и имеет указанный номер доступа в базе данных GenBank: Cry3Aa1 (М22472), Cry3Aa2 (J02978), Cry3Aa3 (Y00420), Cry3Aa4 (M30503), Cry3Aa5 (M37207), Cry3Aa6 (U10985), Cry3Aa7 (AJ237900), Cry3Aa8 (AAS79487), Cry3Aa9 (AAW05659), Cry3Aa10 (AAU29411) и Cry3Aa11 (AY882576). В другом аспекте этого исполнения белок CrylAa выбирается из следующей группы и имеет указанный номер доступа в базе данных GenBank: Cry1Aa1 (Ml 1250), CrylAa2 (M10917), Cry1Aa3 (D00348), Cry1Aa4 (X13535), Cry1Aa5 (D17518), Cry1Aa6 (U43605), Cry1Aa7 (AF081790), Cry1Aa8 (126149), Cry1Aa9 (AB026261), Cry1Aa10 (AF154676), CrylAal 1 (Y09663), Cry1Aal2 (AF384211), Cry1Aa13 (AF510713), Cry1Aa14(AY197341) и Cry1Aa15 (DQ062690). Еще в одном аспекте этого исполнения белок Cry1Ab выбирается из следующей группы и имеет указанный номер доступа в базе данных GenBank: Cry1Ab1 (M13898), CrylAb2 (M12661), Cry1Ab3 (M15271), Cry1Ab4 (D00117), Cry1Ab5 (X04698), Cry1Ab6 (M37263), Cry1Ab7 (X13233), Cry1Ab8 (M16463), Cry1Ab9 (Х54939), Cry1Ab10 (A29125), Cry1Ab11 (112419), Cry1Abl2 (AF059670), Cry1Ab13 (AF254640), Cry1Ab14 (U94191), Cry1Ab15 (AF358861), Cry1Ab16 (AF37560), Cry1Ab17 (AAT46415), Cry1Ab18 (AAQ88259), Cry1Ab19 (AY847289), Cry1Ab20 (DQ241675), Cry1Ab21 (EF683163) и Cry1Ab22 (ABW87320). Еще в одном аспекте этого исполнения первый Cry-белок содержит последовательность аминокислот, выбранную из группы в составе: SEQ ID NO: 68, SEQ ID NO: 70 и SEQ ID NO: 135, а второй Cry-белок содержит последовательность аминокислот, записанную в SEQ ID NO: 72.
В одном исполнении настоящее изобретение охватывает сГИБ, в котором имеется, по меньшей мере, одна позиция кроссинговера между N-концевым участком первого Cry-белка и C-концевым участком второго Cry-белка, расположенная в консервативном блоке 3, вариабельном участке 4 или в консервативном блоке 4. В одном аспекте этого исполнения эта позиция кроссинговера в консервативном блоке 3 расположена сразу же за аминокислотой, соответствующей Ser451, Phe454, или Leu468 последовательности SEQ ID NO: 70. В другом аспекте этого исполнения эта позиция кроссинговера расположена в консервативном блоке 3 сразу же после Ser451, Phe454 или Leu468 последовательности SEQ ID: 70, либо после Ser450, Phe453, или Leu467 последовательности SEQ ID NO: 68; либо Ser497, Phe 100, Leull4 последовательности SEQ ID NO: 135. Позиции кроссинговера в некоторых исполнениях сГИБов с Cry3A/Cry1Ab или с модифицированным Cry3A/Cry1Ab исполнения сГИБов по настоящему изобретению показаны на Фигуре 2, указывающей процент идентичности.
В другом исполнении сГИБ по настоящему изобретению содержит, по меньшей мере, две позиции кроссинговера между последовательностью аминокислот из первого Bt Cry-белка и последовательностью аминокислот из второго Bt Cry-белка. В одном аспекте этого исполнения одна позиция кроссинговера между Cry3A или модифицированным Cry3A и Cry1Ab или Cry1Aa расположена в консервативном блоке 2 сразу же после аминокислоты, соответствующей Asp232 последовательности SEQ ID N0: 70, а вторая позиция кроссинговера между Cry1Ab и Cry3A или модифицированным Cry3A расположена в консервативном блоке 3 сразу же после аминокислоты, соответствующей Leu476 последовательности SEQ ID NO: 72. В другом аспекте этого исполнения позиция кроссинговера между Cry3A или модифицированным Cry3A и Cry1Ab или Cry1Aa расположена в консервативном блоке 2 сразу же после аминокислоты, соответствующей Asp232 последовательности SEQ ID N0: 70, или Asp231 последовательности SEQ ID NO: 68, или Asp278 последовательности SEQ ID NO: 135, а вторая позиция кроссинговера между Cry1Ab и Cry3A или модифицированным Cry3A расположена в консервативном блоке 3 сразу же после аминокислоты, соответствующей Leu476 последовательности SEQ ID NO: 72.
Еще в одном аспекте этого исполнения первая позиция кроссинговера между белками Cry3A или модифицированным Cry3A и Cry1Ab расположена в консервативном блоке 3 сразу же за аминокислотой, соответствующей Leu468 последовательности SEQ ID NO: 70, а вторая позиция кроссинговера между CrylAb и Cry3A или модифицированным Cry3A расположена в консервативном блоке 4 сразу же после аминокислоты, соответствующей IIe527 последовательности SEQ ID NO: 72. В другом аспекте этого исполнения первая позиция кроссинговера между белками Cry3A или модифицированным Cry3A и Cry1Ab расположена в консервативном блоке 3 сразу же за аминокислотой, соответствующей Leu468 последовательности SEQ ID NO: 70 или Leu467 последовательности SEQ ID NO: 68, или Leu114 последовательности SEQ ID NO: 135, а вторая позиция кроссинговера между Cry1Ab и Cry3A или модифицированным Cry3A расположена в консервативном блоке 4 сразу же после аминокислоты, соответствующей I1e527 последовательности SEQ ID NO: 72. Еще в одном аспекте этого исполнения сГИБ содержит последовательность аминокислот SEQ ID NO: 28 или SEQ ID NO: 34.
В одном исполнении настоящее изобретение охватывает сГИБ, в котором первый Cry-белок представлен белком Cry3A или модифицированным Cry3A, а второй Cry-белок представлен белком Cry1Aa или Cry1Ab, причем этот сГИБ содержит последовательность аминокислот, по меньшей мере, на 80% идентичную SEQ ID NO:
64. В другом исполнении сГИБ содержит аминокислотную последовательнсть, по меньшей мере, на 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентичную SEQ ID NO: 64. Совмещение некоторых исполнений настоящего изобретения с последовательностью SEQ ID NO: 64 представлено на Фигуре 2, которая показывает процент идентичности.
В другом исполнении настоящее изобретение охватывает сГИБ, в котором первый Cry-белок представлен белком Cry3A или модифицированным Cry3A, а второй Cry-белок представлен белком Cry1Aa или Cry1Ab, причем этот сГИБ содержит последовательность аминокислот, по меньшей мере, на 75% идентичную SEQ ID NO:
70. В другом исполнении сГИБ содержит аминокислотную последовательнсть, по меньшей мере, на 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93. 94, 95, 96, 97, 98 или 99% идентичную SEQ ID NO: 70. Совмещение некоторых исполнений настоящего изобретения с последовательностью SEQ ID NO: 70 представлено на Фигуре 1, которая показывает процент идентичности.
В другом исполнении настоящее изобретение охватывает сГИБ, в котором имеется первая позиция кроссинговера между Cry3A или модифицированным Cry3A и Cry1Aa или Cry1Ab в консервативном блоке 2 и вторая позиция кроссинговера между Cry1Aa или Cry1Ab и Cry3A или модифицированным Cry3A в консервативном блоке 3, и который содержит последовательность аминокислот, по меньшей мере, на 56% идентичную SEQ ID NO: 64. В одном аспекте этого исполнения сГИБ имеет, по меньшей мере, 60, 70 или 80% идентичности с последовательностью SEQ ID NO: 64. В другом аспекте этого исполнения сГИБ имеет, по меньшей мере, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97. 98 или 99% идентичности с SEQ ID NO: 64.
Еще в другом исполнении настоящее изобретение охватывает сГИБ, содержащий последовательность аминокислот, выбранную из группы в составе: SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 30, SEQ ID NO: 34, SEQ ID NO: 62; SEQ ID NO: 64. SEQ ID NO: 147, SEQ ID NO: 153, SEQ ID NO: 155, SEQ ID NO: 159 и SEQ ID NO: 160.
В одном исполнении сГИБ по настоящему изобретению обладает активностью против других насекомых-вредителей, включая (но не ограничиваясь) северного кукурузного корневого жука, мексиканского кукурузного корневого жука, колорадского картофельного жука и/или европейского кукурузного мотылька.
В другом исполнении настоящее изобретение охватывает молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует сГИБ по настоящему изобретению. В одном аспекте этого исполнения эта молекула нуклеиновой кислоты содержит нуклеотидную последовательность, выбранную из группы в составе: SEQ ID NO: I, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 21, SEQ ID NO: 23, SEQ ID NO: 29, SEQ ID NO: 33, SEQ ID NO: 61; SEQ ID NO: 63, SEQ ID NO: 146, SEQ ID NO: 152, SEQ ID NO: 154 и SEQ ID NO: 158. Разъяснение на конкретных примерах способов создания молекулы нуклеиновой кислоты, которая кодирует сГИБ, можно найти в Примерах 1-41. Специалисты в данной отрасли увидят, какие модификации можно внести в приведенные в примерах способы для создания сГИБов по настоящему изобретению.
Настоящее изобретение также охватывает экспрессионные кассеты, содержащие молекулы нуклеиновой кислоты, а также рекомбинантные векторы и трансгенные клетки-хозяева, не из организма человека, такие как бактериальные клетки или растительные клетки, в которые помещаются экспрессионные кассеты по изобретению.
Настоящее изобретение также охватывает рекомбинантные векторы, содержащие нуклеиновые кислоты по настоящему изобретению. В таких векторах нуклеиновые кислоты в оптимальном варианте содержатся в экспрессионных кассетах, включающих в себя регуляторные элементы для экспрессии нуклеотидных последовательностей в клетке-хозяине, пригодной для экспрессии таких нуклеотидных последовательностей. Такие регуляторные элементы обычно включают в себя промотор и сигналы терминации, а в оптимальном варианте также содержат элементы, позволяющие выполнять эффективную трансляцию полипептидов, кодированных нуклеиновыми кислотами настоящего изобретения. Векторы, содержащие эти нуклеиновые кислоты, могут быть способны к репликации в особых клетках-хозяевах, предпочтительно как внехромосомные молекулы, и таким образом используются для увеличения нуклеиновых кислот по настоящему изобретению в клетках-хозяевах. В одном исполнении клетками-хозяевами для таких векторов являются микроорганизмы, такие как бактерии, в частности. Bacillus thuringiensis или Е.coll. В другом исполнении клетками-хозяевами для таких рекомбинантных векторов являются эндофиты или эпифиты. Еще в одном исполнении такие векторы являются вирусными векторами и используются для репликации нуклеотидных последовательностей в конкретных клетках-хозяевах, например, в клетках насекомых или растительных клетках. Рекомбинантные векторы также используются для трансформации нуклеотидных последовательностей по настоящему изобретению в клетки-хозяева, посредством чего эти нуклеотидные последовательности устойчиво интегрируются в ДНК трансгенного хозяина. В одном исполнении таким трансгенным хозяином является растение, например, кукуруза.
Испытания сГИБов по настоящему изобретению в биопробах показали наличие у них активности против насекомых. В одном исполнении сГИБы по изобретению активны против жесткокрылых насекомых, или против чешуекрылых насекомых, или против и тех и других. В одном аспекте этого исполнения сГИБы по изобретению активны против западного кукурузного корневого жука, северного кукурузного корневого жука, мексиканского кукурузного корневого жука и/или колорадского картофельного жука. В другом аспекте этого исполнения сГИБы по изобретению активны против европейского кукурузного мотылька. Свойства сГИБов по борьбе с насекомыми проиллюстрированы далее в Примерах 43, 45 и 46.
Настоящее изобретение также охватывает композиции, содержащие сГИБ в количестве, эффективном для борьбы с насекомыми.
В другом исполнении изобретение охватывает способ производства сГИБа, активного против насекомых, состоящий в том, что: (а) получают клетку-хозяин, содержащую ген, который сам содержит гетерологичную последовательность-промотор, оперативно связанную с молекулой нуклеиновой кислоты по изобретению; и (б) выращивают трансгенную клетку-хозяин таким образом, чтобы экспрессировать сГИБ, активный против насекомых.
Еще в одном исполнении изобретение охватывает способ производства трансгенного растения, устойчивого против насекомых, состоящий во введении молекулы нуклеиновой кислоты по изобретению в трансгенное растение, причем молекула нуклеиновой кислоты вызывает экспрессию сГИБа в этом трансгенном растении в количестве, эффективном для борьбы с насекомыми. В одном аспекте этого исполнения такими насекомыми являются жесткокрылые или чешуекрылые насекомые. В другом аспекте этого исполнения такими жескокрылыми насекомыми являются западный кукурузный корневой жук, северный кукурузный корневой жук, мексиканский кукурузный корневой жук и/или колорадский картофельный жук. Еще в одном аспекте этого исполнения таким чешуекрылым насекомым является европейский кукурузный мотылек.
Еще в одном исполнении изобретение охватывает способ борьбы с насекомыми, включающий в себя подачу насекомым эффективного количества сГИБа по изобретению. В одном аспекте этого исполнения такими насекомыми являются жесткокрылые или чешуекрылые насекомые. В другом аспекте этого исполнения такими жескокрылыми насекомыми являются западный кукурузный корневой жук, северный кукурузный корневой жук, мексиканский кукурузный корневой жук и/или колорадский картофельный жук. Еще в одном аспекте этого исполнения таким чешуекрылым насекомым является европейский кукурузный мотылек. В типичном случае подача сГИБа насекомым осуществляется орально. В одном аспекте сГИБ подается орально через трансгенное растение, содержащее последовательность аминокислот, экспрессирующую сГИБ по настоящему изобретению.
Настоящее изобретение также охватывает способ борьбы с насекомыми, в котором трансгенное растение также содержит вторую молекулу нуклеиновой кислоты или группы молекул нуклеиновой кислоты, кодирующие второй пестицидный элемент. Примерами такой второй нуклеиновой кислоты могут быть нуклеиновые кислоты, кодирующие Bt Cry-белок, а также кодирующие вегетативный инсектицидный белок, описанный в патентах США 5,849,870 и 5,877,012, включенных сюда путем ссылки, или же те, которые кодируют канал для производства небелковых инсектицидных элементов.
Настоящее изобретение также охватывает способ получения сконструированного гибридного инсектицидного белка (сГИБа), состоящий в том, что: (а) получают первый Bt Cry-белок или модифицированный Bt Cry-белок; (б) получают второй Bt Cry-белок, отличающийся от первого Bt Cry-белка или модифицированного Bt Cry-белка; (в) комбинируют полные или неполные вариабельные участки и консервативные блоки первого Bt Cry-белка или модифицированного Bt Cry-белка с полными или неполными вариабельными участками и консервативными блоками второго Bt Cry-белка для образования сГИБа, обладающего активностью против, по меньшей мере, западного кукурузного корневого жука; и, по выбору, (г) вводят пептидильный фрагмент в N-концевую часть сГИБа, или протоксинный хвостовой участок Bt Cry-белка в С-концевую часть сГИБа, или обоих, причем эти N-концевой пептидильный фрагмент, или С-концевой протоксинный участок, или оба придают сГИБу инсектицидную активность, или повышают инсектицидную активность сГИБа, или делают сГИБ более стабильным, чем сГИБ, не имеющий пептидильного фрагмента, или протоксинного хвостового участка, или обоих.
В другом исполнении настоящее изобретение охватывает способ изготовления сконструированного гибридного инсектицидного белка (сГИБа), состоящий в том, что: (а) получают первую нуклеиновую кислоту, кодирующую первый Bt Cry-белок или модифицированный Bt Cry-белок, и вторую нуклеиновую кислоту, кодирующую второй Сгу-белок, отличающийся от первого Cry-белка или модифицированного Bt Cry-белка; (б) выделяют из первой и второй нуклеиновых кислот нуклеотидную последовательность, кодирующую полные или неполные вариабельные участки и консервативные блоки первого Bt Cry-белка или модифицированного Bt Cry-белка и второго Bt Cry-белка; (в) соединяют полученные на этапе (б) нуклеиновые кислоты таким образом, чтобы получить новую гибридную нуклеиновую кислоту, кодирующую белок, а также, по выбору, выполняют гибридизацию нуклеиновой кислоты, кодирующей пептидильный фрагмент, с 5' концом вышеупомянутой гибридной нуклеиновой кислоты, в результате чего получается удлинение 5' конца, или выполняют гибридизацию нуклеиновой кислоты, кодирующей протоксинный хвостовой участок Bt Cry-белка с 3' концом вышеупомянутой гибридной нуклеиновой кислоты, в результате чего получается удлинение 3' конца, или то и другое; (г) в экспрессионную кассету вводят гибридную нуклеиновую кислоту, имеющую или не имеющую одно или оба удлинения концов 5' или 3'; (д) трансформируют эту экспрессионную кассету в клетку-хозяин, в результате чего вышеупомянутая клетка-хозяин производит сГИБ; и (е) выполняют биоиспытание сГИБа против, по меньшей мере, западного кукурузного корневого жука, результатом чего является инсектицидная активность против западного кукурузного корневого жука.
В следующих исполнениях способов по изобретению первый Bt Cry-белок или модифицированный Bt Cry-белок представлен белком Cry3A или модифицированным Cry3A, а второй Bt Cry-белок представлен белком A Cry1Aa или Cry1Ab.
В другом исполнении способов по изобретению N-концевой пептидильный фрагмент содержит 9 аминокислот.В одном аспекте этого исполнения N-концевой пептидильный фрагмент содержит последовательность аминокислот YDGRQQHRG (SEQ ID NO: 132) или последовательность аминокислот TSNGRQCAGIRP (SEQ ID NO: 133). В другом аспекте этого исполнения N-концевой пептидильный фрагмент выбирается из группы последовательностей в составе: SEQ ID NO: 126, SEQ ID NO: 127, SEQ ID NO: 128, SEQ ID NO: 129, SEQ ID NO: 130, SEQ ID NO: 131 и SEQ ID NO: 132.
Еще в одном исполнении способов изобретения протоксинный хвостовой участок берется из Cry1Aa или Cry1Ab. В одном аспекте этого исполнения этот протоксинный участок содержит, по меньшей мере, 38 аминокислот. В другом аспекте этого исполнения протоксинный хвостовой участок содержит последовательность аминокислот, соответствующую аминокислотам 611-648 последовательности SEQ ID N0: 72. Еще в другом аспекте этого исполнения протоксинный хвостовой участок содержит аминокислоты 611-648 последовательности SEQ ID NO: 72.
Разъяснение на конкретных примерах способов создания гибридной нуклеиновой кислоты и сГИБов можно найти в Примерах 1-41.
В последующих исполнениях нуклеотидные последовательности по изобретению, в особенности, последовательность, кодирующая пептидильный фрагмент, протоксинный хвост и/или консервативные блоки 2, 3 и 4, может быть еще модифицирована путем включения случайных мутаций по технологии, известной как in vitro рекомбинация или перетасовка ДНК. Эта технология описана в работе Stemmer и др.. Nature 370:389-391 (1994) и в патенте США 5,605,793, включенным сюда путем ссылки. Миллионы мутантных копий нуклеотидной последовательности производятся на основе оригинальной нуклеотидной последовательности по настоящему изобретению, и отбираются те варианты, которые имеют улучшенные свойства, такие как повышенная инсектицидная активность, повышенная стабильность или разные специфичность или диапазон мишеней-вредителей. Способ включает в себя формирование мутагенизированного двухцепочечного полинуклеотида из шаблонного двухцепочечного полинуклеотида, содержащего нуклеотидную последовательность по настоящему изобретению, причем шаблонный двухцепочечный полинуклеотид был расщеплен на случайные двухцепочечные фрагменты требуемой длины, кроме того, способ включает в себя этапы: добавления к полученной популяции двухцепочечных случайных фрагментов одного или нескольких одно- или двухцепочечных олигонуклеотидов, причем, вышеупомянутые олигонуклеотиды содержат область идентичности и область гетерологии к шаблонному двухцепочечному полинуклеотиду; денатурации полученной смеси двухцепочечных случайных фрагментов и олигонуклеотидов в одноцепочечные фрагменты; инкубацию полученной популяции одноцепочечных фрагментов с полимеразой в условиях, результатом которых является отжиг вышеупомянутых одноцепочечных фрагментов в вышеуказанных областях идентичности для формирования пар отожженных фрагментов, причем эти области идентичности достаточны для того, чтобы из одного члена пары выполнить точную репликацию второго, таким образом формируя мутагенизированный двухцепочечный полинуклеотид; а также, по меньшей мере, два цикла повтора второго и третьего этапов, причем полученная смесь на втором этапе следующего цикла включает в себя мутагенизированный двухцепочечный полинуклеотид из третьего этапа предыдущего цикла, а следующий цикл формирует следующий мутагенизированный двухцепочечный полинуклеотид. В предпочтительном варианте исполнения изобретения концентрация одноцепочечных видов двухцепочечных случайных фрагментов в популяции двухцепочечных случайных фрагментов составляет менее 1% от веса всей ДНК. В еще более предпочтительном варианте исполнения шаблонный двухцепочечный полинуклеотид содержит, по меньшей мере, около 100 видов полинуклеотидов. В другом предпочтительном исполнении размер двухцепочечных случайных фрагментов составляет приблизительно от 5 п.о. до 5 кб. В другом предпочтительном исполнении четвертый этап способа содержит повторении второго и третьего этапов по крайней мере для 10 циклов.
В качестве биологических инсектицидных контролирующих средств сГИБы производятся путем экспрессии нуклеиновых кислот в гетерологичных клетках-хозяевах, пригодных для экспрессии этих нуклеиновых кислот. В одном исполнении получаются клетки В. thuringiensis, содержащие модификации нуклеиновой кислоты по настоящему изобретению. Такие модификации охватывают мутации или устранение существующих регуляторных элементов, приводящие к измененной экспрессии данной нуклеиновой кислоты, или включение новых регуляторных элементов, контролирующих экспрессию этой нуклеиновой кислоты. В другом исполнении к клеткам Bacillus thuringiensis добавляются копии одной или нескольких нуклеиновых кислот путем либо инсерции в хромосому, либо введением содержащих нуклеиновые кислоты молекул, которые реплицируются внехромосомно.
В другом исполнении, по меньшей мере, одна из нуклеиновых кислот по изобретению вводится в приемлемую экспрессионную кассету, содержащую промотор и сигнал терминации. Экспрессия нуклеиновой кислоты является конституитивной, или же используется индуцируемый промотор, реагирующий на различные типы стимулов для инициации транскрипции. В другом исполнении клетка, в которой экспрессируется сГИБ, представляет собой микроорганизм, например, вирус, бактерию или грибок. Еще в одном исполнении вирус, например, бакуловирус содержит нуклеиновую кислоту по изобретению в своем геноме и экспрессирует большие количества соответствующего инсектицидного белка после инфицирования соответствующих эукариотических клеток, пригодных для репликации вируса и экспрессии нуклеиновой кислоты. Полученный таким образом белок используется в качестве инсектицидного агента. В альтернативном варианте сконструированные бакуловирусы, содержащие нуклеиновую кислоту, используются для инфицирования насекомых in vivo и убивают их либо путем экспрессии инсектицидного токсина, либо комбинацией вирусной инфекции и экспрессии инсектицидного токсина.
Клетки бактерий также являются клетками-хозяевами для экспрессии нуклеиновых кислот по изобретению. В одном исполнении применяются непатогенные симбиозные бактерии, способные жить и размножаться внутри растительной ткани, так называемые эндофиты, или непатогенные симбиозные бактерии, способные колонизировать филлосферу или ризосферу, так называемые эпифиты. К таким бактериям относятся бактерии видов Agrobacterium, Alcaligenes, Azospirillum, Azotobacter, Bacillus, Clavibacter, Enterobacter, Erwinia, Flavobacter, Klebsiella, Pseudomonas, Rhizobium, Serratia, Streptomyces и Xanthomonas. Симбиотические грибки, такие как Trichoderma Gliocladium также способны быть хозяевами для экспрессии изобретенных нуклеиновых кислот с той же самой целью.
Технологии этих генетических манипуляций являются специфичными для различных имеющихся хозяев, и хорошо известны в отрасли. Например, экспрессирующие векторы рКК223-3 и рКК223-2 могут применяться для экспрессии гетерологичных генов в Е.coli либо транскрипционным, либо трансляционным слиянием за tac-промотором или trc-промотором. Для экспрессии оперонов, кодирующих множественные белки ORF простейшая процедура представляет собой введение оперона в вектор, такой как рКК223-3, трансляционным слиянием, что позволяет использовать место связывания родственной рибосомы гетерологичных генов. Технологии сверхэкспрессии в грам-положительных видах, таких как Bacillus, также известны в отрасли и могут использоваться в контексте настоящего изобретения (Quax и др. В: Industrial Microorganisms: Basic and Applied Molecular Genetics, авторы Baltz и др., American Society for Microbiology, Washington (1993)). Альтернативные системы для сверхэкспрессии основываются, например, на дрожжевых векторах и включают использование Pichia, Saccharomyces и Kluyveromyces (Sreekrishna, В: Industrial microorganisms: basic and applied molecular genetics, Baltz, Hegeman, и Skatrud авторы, American Society for Microbiology, Washington (1993); Dequin и Barre, Biotechnology L2: 173-177 (1994); van den Berg и др.. Biotechnology 8:135-139 (1990)).
В одном исполнении, по меньшей мере, один сГИБ по изобретению экспрессирован в высшем организме, таком как растение. В этом случае трансгенные растения, экспрессирующие эффективные количества сГИБа, защищают сами себя от насекомых-вредителей. Когда насекомое начинает есть такое трансгенное растение, оно также поглощает экспрессированный сГИБ. Это удержит насекомое от дальнейшего вгрызания в это растение, а может даже привести к повреждению или гибели насекомого. Нуклеиновая кислота по настоящему изобретению вводится в экспрессионную кассету, которая затем может быть устойчиво интегрирована в геном растения. В другом исполнении такая нуклеиновая кислота включается в непатогенный самореплицирующий вирус. Растения, трансформированные по настоящему изобретению, могут быть однодольными или двухдольными и включают, но не ограничиваясь ими, кукурузу, пшеницу, ячмень, рожь, сладкий картофель, фасоль, горох, цикорий, салат-латук, капусту, цветную капусту, брокколи, турнепс, редис, шпинат, спаржу, лук, чеснок, перец, сельдерей, кабачки, тыкву, коноплю, цукини, яблоко, грушу, айву, дыню, сливу, черешню, персик, нектарин, абрикос, клубнику, виноград, малину, черную смородину, ананас, авокадо, папайю, манго, банан, сою, помидор, сорго, сахарный тростник, сахарную свеклу, подсолнечник, рапс, клевер, табак, морковь, люцерну, рис, картофель, баклажан, огурец, резушку Таля, а также хвойные и лиственные деревья.
После перемещения требуемой нуклеиновой кислоты в конкретный вид растения она может с помощью традиционных технологий разведения распространяться в этом виде или переходить в другие сорта этого же вида, в особенности, включая коммерчески важные сорта.
Предпочтительно, чтобы экспрессия нуклеиновой кислоты по настоящему изобретению осуществлялась в трансгенных растениях, вызывая таким образом биосинтез соответствующего сГИБа в этих трансгенных растениях. Таким путем получаются трансгенные растения с повышенной сопротивляемостью к насекомым, в частности, кукурузному корневому жуку. Для экспрессии в трансгенных растениях нуклеиновым кислотам по изобретению могут потребоваться другие модификации и оптимизация. Хотя во многих случаях гены из микробных организмов могут экспрессироваться в растениях на высоких уровнях и без модификации, но в трансгенных растениях может происходить и слабая экспрессия микробных нуклеиновых кислот, имеющих кодоны, не являющиеся предпочтительными в растениях. В данной отрасли известно, что у всех организмов есть специфические преференции по использованию кодонов, и кодоны нуклеиновых кислот, описанных в настоящем изобретении, могут быть изменены для соответствия преференциям растений, в то же время, поддерживая кодируемые ими аминокислоты. Более того, высокая экспрессия в растениях достигается из кодирующих последовательностей, GC-содержание в которых составляет, по меньшей мере, 35%, предпочтительно более 45%, еще более предпочтительно свыше 50%, а наиболее предпочтительно свыше 60%. Микробные нуклеиновые кислоты с низким GC-содержанием могут слабо экспрессироваться в растениях вследствие существования мотивов АТТТА, которые могут дестабилизировать сообщения, и мотивов ААТААА, которые могут вызвать ненадлежащее полиаденилирование. Хотя предпочтительные генные последовательности могут быть адекватно экспрессированы как в однодольных так и в двудольных видах растений, последовательности могут быть и модифицированы с учетом специфичных преференций по кодонам и GC-содержанию в однодольных и двудольных, поскольку эти преференции отличаются (Murray и др. Nucl. Acids Res. 17:477-498 (1989)). Кроме того, осуществляется скрининг нуклеиновых кислот на наличие нестандартных мест сплайсинга, которые могут вызвать усечение сообщений. Все изменения, которые требуется выполнить внутри нуклеиновых кислот, такие как описаны выше, выполняются с применением хорошо известных технологий сайт-направленного мутагенеза, ПЦР и конструирования синтетических генов с применением способов, описанных в опубликованных патентных заявках ЕР 0385962, ЕР 0359472, и WO 93/07278.
В одном исполнении изобретения кодирующая последовательность сГИБа и/или кодирующая последовательность родительского Bt Cry-белка получается/получаются в соответствии с процедурой, описанной в патенте США 5,625,136, включенном сюда путем ссылки. В этой процедуре используются кодоны, являющиеся предпочтительными для кукурузы, т.е., единственный кодон, наиболее часто кодирующий аминокислоту в кукурузе. Предпочтительный для кукурузы кодон для конкретной аминокислоты может быть получен, например, из известной генной последовательности для кукурузы. Применение кодона кукурузы для 28 генов из растений кукурузы встречается в работе Murray и др.. Nucleic Acids Research 17:477-498 (1989), описание которой включено сюда путем ссылки.
Таким способом нуклеотидная последовательность может быть оптимизирована для экспрессии в любом растении. Признано, что вся генная последовательность или любая ее часть могут быть оптимизированными или синтетическими. То есть, могут также использоваться синтетические или частично оптимизированные последовательности.
Для эффективной инициации трансляции последовательностям, примыкающим к инициирующему метионину, может потребоваться модификация. Например, они могут быть модифицированы путем включения последовательностей, известных своей эффективностью в растениях. Joshi предложил соответствующую типичную последовательность для растений (NAR 15:6643-6653 (1987)), a Clonetech предлагает дальнейший общий инициатор трансляции (1993/1994 каталог, страница 210). Эти консенсусы пригодны для использования с нуклеиновыми кислотами по настоящему изобретению. Эти последовательности включены в конструкции, содержащие нуклеиновые кислоты до ATG включительно (оставляя немодифицированной вторую аминокислоту) или в альтернативном варианте - до и включая GTC, следующую за ATG (с возможностью модифицирования второй аминокислоты данного трансгена).
Экспрессию нуклеиновых кислот в трансгенных растениях ведут промоторы, функционирующие в растениях. Выбор промотора будет варьировать в зависимости от пространственно-временных требований к экспрессии, а также в зависимости от видов-мишеней, Поэтому предпочтительной является экспрессия нуклеиновых кислот по настоящему изобретению в листьях, стеблях или стволах, в початках, в соцветиях (например, в шипах, метелках, косточках и т.п.), в корнях и/или саженцах. Однако во многих случаях ищут защиту от более чем одного типа насекомых-вредителей, а поэтому желательна экспрессия во многих тканях. Хотя многие промоторы из двудольных продемонстрировали свою действенность в однодольных и наоборот, в идеальном случае для экспрессии в двудольных выбираются промоторы двудольных, а промоторы однодольных - для экспрессии в однодольных. Однако не существует никаких ограничений относительно происхождения выбранных промоторов; достаточно, чтобы они эффективно вели экспрессию нуклеиновых кислот в требуемой клетке.
В одном исполнении применяются промоторы с конститутивной экспрессией, включая актин, или убиквитин, или промоторы cmp, или промоторы CaMV 35S и198. Нуклеиновые кислоты по настоящему изобретению также могут экспрессироваться под управлением промоторов с химической регуляцией. Это дает возможность сГИБам синтезироваться только тогда, когда семенные растения обрабатываются индуцирующими химикатами. Оптимальная технология химической индукции генной экспрессии подробно описана в опубликованной заявке ЕР 0 332 104 (Ciba-Geigy) и в патенте США 5,614,395. Оптимальным промотором для химической индукции является табачный PR-1a.
В другом исполнении может применяться категория промоторов, индуцируемых повреждением. Описано много промоторов, экспрессирующихся в местах повреждений или в местах инфицирования фитопатогенами. В идеальном случае такой промотор должен активироваться локально в местах инфекции, и таким путем сГИБ аккумулируется только в тех клетках, которым требуется синтезировать сГИБ, чтобы убить вторгшегося насекомого-вредителя. К оптимальным промоторам этого вида относятся описанные в работах Stanford и др. Mol. Gen. Genet. 215:200-208 (1989), Xu и др. Plant Molec. Biol. 22:573-588 (1993), Logemann и др. Plant Cell 1:151-158 (1989), Rohrmeier и Lehle, Plant Molec. Biol. 22:783-792 (1993), Firek и др. Plant Molec. Biol. 22:129-142 (1993), и Wamer и др. Plant J. 3:191-201 (1993).
Предпочтительными для ткани или преферентными для ткани промоторами, применимыми для экспрессии генов, кодирующих сГИБы в растениях, в особенности, в кукурузе, являются промоторы, выполняющие прямую экспрессию в корне, сердцевине, листе или пыльце, особенно в корне. Такими промоторами являются, например, выделенные из РЕРС или trpA. описанные в патенте США №. 5,625,136, или из MTL, описанные в патенте США №5,466,785. Оба эти патента включены сюда полностью путем ссылки.
Следующими исполнениями являются трансгенные растения, экспрессирующие нуклеиновые кислоты способом индукции повреждением или патогенной инфекцией.
Для процесса разработки гибридной нуклеиновой кислоты с использованием генов сГИБ по настоящему изобретению кроме промоторов применяется еще ряд терминаторов транскрипции, имеющихся в наличии. Терминаторы транскрипции отвечают за прекращение транскрипции за пределами трансгена и за правильное полиаденилирование. Приемлемыми терминаторами транскрипции являются терминаторы, известные своим функционированием в растениях, включая терминатор CaMV 35S, терминатор нопалин синтазы (NOS), гороховый терминатор rbcS E9 и другие, известные в отрасли. Они могут применяться как в однодольных, так и в двудольных. В контексте настоящего изобретения может применяться любой имеющийся терминатор, известный своим функционированием в растениях.
В экспрессионную кассету, описанную в настоящем изобретении, могут быть включены многочисленные другие последовательности. К ним относятся последовательности, показавшие способность повышать экспрессию, такие как интронные последовательности (напр. из Adhl и bronzel) и ведущие вирусные последовательности (напр. из TMV, MCMV и AMV).
Желательно было бы направлять экспрессию нуклеиновых кислот по настоящему изобретению к различным клеточным локализациям в растении. В некоторых случаях желательной может быть локализация в цитозоле, а в других случаях может быть предпочтительной локализация в некоторой субклеточной органелле. Субклеточная локализация трансгенно-кодированных энзимов осуществляется с помощью хорошо известных в отрасли технологий. В типичном случае ДНК, кодирующая пептид-мишень из известного генного продукта, направленного на органеллу, направляется и гибридизируется перед нуклеиновой кислотой. Известно много таких последовательностей-мишеней для хлоропласта, и их функционирование в гетерологичных конструкциях было показано. Экспрессия нуклеиновых кислот по изобретению также направлена на эндоплазматический ретикулум или на вакуоли клеток-хозяев. Технологии достижения этого также хорошо известны в отрасли.
Векторы, пригодные для трансформации растений, описаны в разных местах настоящего описания. Для переноса посредством агробактерий пригодны бинарные векторы, или векторы, несущие, по меньшей мере, одну граничную последовательность Т-ДНК, в то время как для прямого генного переноса подходит любой вектор, и предпочтительной может быть линейная ДНК, включающая только значимую конструкцию. В случае прямого переноса генов может применяться трансформация с единственным видом ДНК или ко-трансформация (Schocher и др. Biotechnology 4:1093-1096 (1986)). Как для прямого переноса генов, так и для агробактериального переноса трансформация обычно (но не обязательно) осуществляется с выбираемым маркером, который может обеспечить устойчивость к антибиотикам (канамицину, гигромицину или метотрексату) или к гербицидам (basta). Векторы трансформации растений, содержащие гены сГИБ по настоящему изобретению, могут также содержать гены (напр. изомеразу фосфоманнозы; PMI), обеспечивающие положительную селекцию трансгенных растений, как описано в патентах США 5,767,378 и 5,994,629, включенных сюда путем ссылки. Однако выбор такого маркера не является критичным для настоящего изобретения.
В другом исполнении нуклеиновая кислота по настоящему изобретению прямо трансформируется в геном пластид. Большим преимуществом пластидной трансформации является то, что пластиды в общем случае способны к экспрессию бактериальных генов без существенной оптимизации кодонов, и пластиды способны к экспрессии множественных открытых рамок чтения под управлением единственного промотора. Технология пластидной трансформации широко описана в патентах США №№5,451,513, 5,545,817, и 5,545,818, в заявке РСТ №WO 95/16783 и в работе McBride и др. (1994) Proc. Nati. Acad. Sci. USA 91, 7301-7305. Базовая технология хлоропластной трансформации включает в себя введение участков клонированной пластидной ДНК, ограничивающих выбираемый маркер, совместно со значимым геном в соответствующую ткань-мишень, напр., с помощью биолистики или протопластной трансформации (напр., трансформации посредством хлорида кальция или PEG). Ограничивающие участки размером от 1 до 1,5 кб, именуемые последовательностями-мишенями, осуществляют гомологичную рекомбинацию с геномом пластид и таким образом дают возможность замены или модификации специфичных участков пластома. Изначально точечные мутации в генах хлоропластов 16S рРНК и rps12, придающие сопротивляемость спектиномицину и/или стрептомицину, используются в качестве выбираемых маркерова для трансформации (Svab, Z., Hajdukiewicz, P., и Maliga, P. (1990) Proc. Nati. Acad. Sci. USA 87, 8526-8530; Staub, J. M., и Maliga, P. (1992) Plant Cell 4, 39-45). Результатом этого являются устойчивые гомоплазменные трансформанты с частотой приблизительно один на 100 бомбардировок листьев-мишеней. Наличие сайтов клонирования между этими маркерами дало возможность создания пластид-направленного вектора для введения чужеродных генов (Staub, J.M., и Maliga, Р. (1993) ЕМВО J. 12, 601-606). Существенные повышения частоты трансформации получены путем замены рецессивных генов сопротивляемости антибиотикам рРНК или p-белка на доминантный выбираемый маркер, бактериальный ген aadA, кодирующий энзим аминогликозид-3'-аденилтрансферазу, обезвреживающий спектиномицин (Svab, Z., и Maliga, Р. (1993) Proc. Nati. Acad. Sci. USA 90, 913-917). Ранее этот маркер успешно применялся для высокочастотной трансформации генома пластид зеленой водоросли Chlamydomonas reinhardtii (Goldschmidt- Clermont, M. (1991) Nucl. Acids Res. 19:4083-4089). Другие выбираемые маркеры, полезные для пластидной трансформации также известны в отрасли и охватываются рамками настоящего изобретения. В типичном случае для достижения гомопластидного состояния требуется приблизительно 15-20 циклов клеточного деления после трансформации. Пластидная экспрессия, в которой гены вводятся путем гомологичной рекомбинации во все несколько тысяч копий кольцевого генома пластид, присутствующих в каждой клетке, использует преимущество от огромного числа копий над ядерной экспрессией генов, позволяя получать уровни экспрессии, легко превышающие 10% от всего растворимого растительного белка. В предпочтительном исполнении нуклеиновая кислота по настоящему изобретению вводится в пластид-направленный вектор и трансформируется в геном пластид желаемого растения-хозяина. Получаются гомопластические растения с геномами пластид, содержащими нуклеиновую кислоту по настоящему изобретению, предпочтительно способные к высокому уровню экспрессии этой нуклеиновой кислоты..
СГИБы по изобретению могут применяться в комбинации с другими пестицидными элементами (напр., с Bt Cry-белками) с целью расширения диапазона вредителей, на которых они направлены. Более того, применение сГИБов по изобретению в комбинации с модифицированными Cry3A-токсинами, Bt Cry-белками или другими элементами, активными против КЮК (кукурузного корневого жука), такими как РНКи, обладающими другим видом активности или направленными на другие рецепторы в пищеварительном канале насекомого, особенно полезно для предотвращения возникновению и/или регулирования сопротивляемости у кукурузного корневого жука. Перечень других инсектицидных элементов включает, не ограничиваясь ими, лектины, α-амилазу, пероксидазу и оксидазу холестерина. Также полезными в комбинации со сГИБами по настоящему изобретению являются Vip-гены, описанные в патенте США №5,889,174, включенном сюда путем ссылки.
Эта коэкспрессия более чем одного инсектицидного элемента в одном трансгенном растении может быть достигнута путем приготовления одного рекомбинантного вектора, содержащего кодирующие последовательности более чем одного инсектицидного элемента в так называемом молекулярном стеке и генетического конструирования растения, которое бы содержало и экспрессировало все эти инсектицидные элементы. Такие молекулярные стеки могут быть также получены применением мини-хромосом, как описано, например, в патенте США 7,235,716. В альтернативном варианте трансгенное растение, содержащее одну нуклеиновую кислоту, кодирующую первый инсектицидный элемент, может быть ретрансформировано другой нуклеиновой кислотой, кодирующей другой инсектицидный элемент, и т.д. В альтернативном варианте растение, Родитель 1, может быть генетически разработано для экспрессии генов по настоящему изобретению. Второе растение, Родитель 2, может быть генетически разработано для экспрессии дополнительного элемента борьбы с насекомыми. Скрещивание Родителя 1 с Родителем 2 дает потомство растений, экспрессирующих все гены, введенные в Родителей 1 и 2.
Трансгенные семена по настоящему изобретению также можно обрабатывать инсектицидным покрытием, как описано в патентах США №№5,849,320 и 5,876,739, включенных сюда путем ссылки. Если и инсектицидное покрытие семян, и трансгенные семена по изобретению являются активными против одного и того же насекомого, то такая комбинация полезна (i) в способе повышения активности сГИБа по изобретению против такого насекомого и (ii) в способе предотвращения развития сопротивляемости сГИБу по изобретению за счет наличия второго механизма действия против насекомого-мишени. Таким образом, изобретение предоставляет способ повышения активности против насекомого-мишени или предотвращения развития сопротивляемости у такого насекомого, например, у кукурузного корневого жука, состоящий в нанесении инсектицидного покрытия на трансгенные семена, содержащие один или несколько сГИБов по изобретению.
Даже если инсектицидное покрытие активно против другого насекомого, это инсектицидное покрытие полезно для расширения диапазона насекомых, против которых ведется борьба, например, путем нанесения инсектицидного покрытия, активного против чешуекрылых насекомых, на трансгенные семена по изобретению, активные против жесткокрылых насекомых, в результате чего полученные трансгенные семена с покрытием будут активны как против чешуекрылых, так и против жесткокрылых насекомых-вредителей.
ПРИМЕРЫ
Далее описание изобретения подано на следующих детально представленных примерах. Эти примеры являются только иллюстративными и не предназначены для какого-либо ограничения, если оно не указывается. Используемые здесь стандартные технологии рекомбинантной ДНК или клонирования молекул хорошо известны в отрасли и описаны в работах J. Sambrook, и др.. Molecular Cloning: A Laboratory Manual, третье издание, Cold Spring Harbor, Нью Йорк: Cold Spring Harbor Laboratory Press (2001); T.J.Silhavy, M.L.Berman, и L.W.Enquist, Experiments -with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, Нью Йорк (1984) и Ausubel, F.M. и др.. Current Protocols in Molecular Biology, Нью Йорк, John Wiley and Sons Inc., (1988), Reiter, и др.. Methods in Arabidopsis Research, World Scientific Press (1992), а также Schultz и др.. Plant Molecular Biology Manual, Kluwer Academic Publishers (1998).
Пример 1. Родительские кодирующие последовательности
Оптимизированные для кукурузы кодирующие последовательности cry3Aa, crylAb, cry1Ba, и cry1Fa, обозначенные здесь соответственно как mocry3Aa, mocry1Ab, mocry1Ba и mocry1Fa, были получены в соответствии с процедурой, описанной в патенте США 5,625,136, включенном сюда полностью путем ссылки.
Последовательность cry3A055 (SEQ ID NO: 67), кодирующая белок Cry3A055 (SEQ ID NO: 68) была получена путем модификации кодирующей последовательности mocry3A введением нуклеотидной последовательности, кодирующей сайт узнавания протеазы катепсин G в домен I согласно патенту США 7,030,295, включенному сюда полностью путем ссылки.
Последовательности mocrY3Aa (SEQ ID NO: 67), кодирующая белок, представленный в SEQ ID NO: 68, Cry3A055 (SEQ ID NO: 69), кодирующая белок, представленный в SEQ ID NO: 70, mocry1Ab (SEQ ID NO: 71), кодирующая белок, представленный в SEQ ID NO: 72, mocry1Ba (SEQ ID NO: 73), кодирующая белок, представленный в SEQ ID NO: 74, mocry1Fa (SEQ ID NO: 75), кодирующая белок, представленный в SEQ ID NO: 76, cry8Aa (SEQ ID NO: 77). кодирующая белок, представленный в SEQ ID NO: 78, cry1Ac (SEQ ID NO: 79), кодирующая белок, представленный в SEQ ID NO: 80, и crylla (SEQ ID NO: 81), кодирующая белок, представленный в SEQ ID NO: 82, были использованы в конструировании кодируемых ими гибридных нуклеиновых кислот и белков, как описано в следующих примерах.
Пример 2. Применение ПЦР-праймеров для конструирования гибридных нуклеиновых кислот
Полимеразная цепная реакция (ПЦР) представляет собой повторяющийся, ферментный, инициируемый праймером синтез последовательности нуклеиновых кислот. Эта процедура хорошо известна и широко применяется специалистами в данной отрасли (См. Mullis, патенты США №№4,683,195, 4,683,202 и 4,800,159; Saiki, Randall K., Stephen Scharf, Fred Faloona, Kary B. Mullis, Glenn T. Horn, Henry A. Eriich, Norman Amheim [1985] "Enzymatic Amplification of.beta.-Globin Genomic Sequences and Restriction Site Analysis for Diagnosis of Sickle Cell Anemia," Science 230:1350-1354). ПЦР основана на ферментном увеличении значимого фрагмента ДНК, по краям которого находятся два олигонуклеотидных праймера, гибридизирующиеся к противоположным цепочкам последовательности-мишени. Эти праймеры сориентированы так, что их 3' концы направлены друг на друга. Повторяющиеся циклы денатурации нагревом, отжиг праймеров к их комплементарным последовательностям и удлинение отожженных праймеров с помощью ДНК-полимеразы дают в результате увеличение сегмента, определенного 5' концами этих ПЦР-праймеров. Поскольку продукт удлинения каждого праймера может служить шаблоном для другого праймера, каждый цикл практически удваивает количество фрагментов ДНК, полученное в предыдущем цикле. Результатом этого является экспоненциальное накопление конкретного фрагмента-мишени в несколько миллионов раз за несколько часов. Используя термостабильную ДНК-полимеразу, например, Taq-полимеразу, выделенную из термофильной бактерии Thermus aquaticus, процесс увеличения можно сделать полностью автоматическим, Химерные кодирующие последовательности, описанные в следующих примерах, были разработаны с применением различных комбинаций примеров праймеров, показанных в Таблице 1. Смеси для реакций ПЦР и протоколы температурных циклов ПЦР, применявшиеся в экспериментах, представлены в Таблицах 2 и 3 соответственно. В каждом из последующих примеров праймеры ПЦР обозначаются наименованием и "SEQ ID NO:", а смеси для реакций ПЦР и протоколы температурных циклов ПЦР обозначены их соответствующими номерами. Квалифицированный специалист увидит, что для конструирования химерных кодирующих последовательностей по настоящему изобретению могут применяться другие праймеры ПЦР, а также другие условия ПЦР, и, что перечень примеров праймеров и условий ПЦР, используемых в настоящем изобретении, ни в коем случае не является ограничивающим.
|
|
|
|
|
|
В Таблице 4 показано взаимное соответствие между тремя доменами белков Cry3A055, Cry1Ab и Cry3A с их соответствующими вариабельными участками и консервативными блоками. Для каждого белка показано, какие аминокислоты содержатся в доменах, консервативных блоках и вариабельных участках.
Таблица 4
|
Пример 3. Разработка 20L-8a
Фрагмент первой нуклеиновой кислоты, кодирующий N-концевую часть белка Cry2A055 (SEQ ID NO: 70) увеличили способом ПЦР из плазмиды, содержащей cry3A055 (SEQ ID NO: 69), с применением праймеров 5'3A-1-bam (SEQ ID NO: 83) и С3-3А-б (SEQ ID NO: 84), а также Смеси 1 и температурного цикла Профиля 1. Эта реакция ПЦР ввела точечную мутацию путем удаления нуклеотида 28 последовательности SEQ ID NO: 69 (cry3A055), что привело к сдвигу рамки считывания сгу2А055, и удалила сайт BamHI и последовательность Козака (Kozak, М., 1986. Cell 44:283-92) на 5' конце полученного ампликона.
Фрагмент второй нуклеиновой кислоты, кодирующий C-концевую часть белка Cry1Ab (SEQ ID NO: 72) увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71), с применением праймеров С3-1Ab-3 (SEQ ID NO: 85) и 1Ab-6-Sac (SEQ ID NO: 86), а также Смеси 1 и температурного цикла Профиля 1.
Описанные выше первая и вторая нуклеиновые кислоты были соединены путем использования их в качестве шаблонов в расширенной (overlap) ПЦР реакции (Horton и др., 1989. Gene 77: 61-68) с праймерами 5'3A-l-bam (SEQ ID NO: 83) и 1Ab-6-Sac (SEQ ID NO: 86), а также Смеси 2 и температурного цикла Профиля 1, с тем отличием, что в качестве температуры отжига использовали градиент 45-65°С.
Полученный в результате ампликон лидировали в качестве фрагмента с затупленным концом к вектору pCR2.1-TOPO (Invitrogen, Carlsbad, CA), разрезанному Smal, для образования плазмиды p20L8a/CR2.1. Затем фрагмент BamHI-SacI из p20L8a/CR2.1 лигировали к рЕТ21а (EMD Biosciences, Inc., San Diego, CA), который был разрезан с помощью BamHI-SacI, и трансформировали в Е. coli. Фрагмент BamHI-SacI из p20L8a/CR2.1 содержал 40 нуклеотидов, полученных из вектора pCR2.1-TOPO, примыкающих к внерамочному ампликону из первой ПЦР реакции. Лигирование этого фрагмента BamHI-SacI к рЕТ21а создало открытую рамку считывания, начинающуюся стартовым кодоном (ATG) метки Т7 и заканчивающуюся сайтом Sad введенной ДНК. Эта открытая рамка считывания обозначена 20L-8a (SEQ ID NO: 1) и кодирует химерный инсектицидный белок 20L-8a (SEQ ID NO: 2). Таким образом, в химерном инсектицидном белке 20L-8a от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MASMTGGQQMGRGSTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 126), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70), составляющие вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), куда входят С-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6; а также 38 аминокислот протоксинного хвостового участка белка Cry1Ab.
Нуклеотиды, кодирующие аминокислоты 1-14 пептидильного фрагмента, получены из метки Т7 и сайта расщепления BamHI вектора рЕТ21. Нуклеотиды, кодирующие аминокислоты 15-26 пептидильного фрагмента, получены из вектора pCR2.1-TOPO. А нуклеотиды, кодирующие аминокислоты 27-35 пептидильного фрагмента, получены из фрагмента cry3A055, находящегося за рамками остальной части кодирующей последовательности cry3A055.
Пример 4. Разработка FR8a
Кодирующая последовательность FR8a была разработана путем размещения последовательности Козака (АСС) и стартового кодона (ATG) сразу же после N-концевого сайта BamHI в 20L-8a (См. Пример 3). Кроме того, сайт EcoRI в 20L-8a был разрушен для помощи будущей векторизации FR8a. Все эти изменения были сделаны в ходе одной ПЦР с применением 20L-8a в качестве шаблона, а в качестве праймеров: 8a-atg-delRI (SEQ ID NO: 87) и С2-ЗА-4 (SEQ ID NO: 88), Смеси 2 и температурного цикла Профиля 2.
Полученный в результате ампликон лигировали к вектору pCR2.1-TOPO (Invitrogen). Фрагмент BamHI-PpuMI из клонированного продукта ПЦР затем лигировали к фрагменту PpuMI-NcoI из 20L8a/pCR2.1 (См. Пример 3) и к фрагменту NcoI-BamHI из 20L8a/pCR2.1 для создания FR8a (SEQ ID NO: 3), которая кодирует химерный инсектицидный белок FR8a (SEQ ID NO: 4). Таким образом, в химерном инсектицидном белке FR8a от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70), составляющие вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6; а также 38 аминокислот протоксинного хвостового участка белка Cry1Ab.
Этот сГИБ FR8a имеет очень высокую активность против западного кукурузного корневого жука, как показано в Таблице 5. Следовательно, удаление последовательности аминокислоты Т7 из N-концевого пептидильного фрагмента сГИБа 20L-8a не сказалось отрицательно на инсектицидной активности.
Добавлением дополнительных 34 аминокислот к N-концу FR8a создали сГИБ, получивший обозначение FR8a+34 (SEQ ID NO: 160), имеющий N-концевой пептидильный фрагмент из 56 аминокислот (SEQ ID NO: 131). Этот N-концевой пептидильный фрагмент из 56 аминокислот не оказал отрицательного воздействия на активность FR8a против западного кукурузного корневого жука (См. Табл.5). Пример 5. Разработка FRCG.
Для того чтобы определить, является ли сайт узнавания протеазы катепсин G необходимым для инсектицидной активности гибридного белка, содержащего N-концевой фрагмент от Cry3A055, была изготовлена модель, устраняющая сайт катепсина G из гибридного белка FR8a (Пример 4). Первый фрагмент нуклеиновой кислоты MluI-PpuMI из плазмиды, содержащей FR8a (SEQ ID NO: 3), и второй фрагмент нуклеиновой кислоты PpuMI/MluI из плазмиды, содержащей mocry3Aa (SEQ ID NO: 67) дотировали с помощью стандартной в молекулярной биологии технологии для создания FRCG (также обозначаемой FR8a-catg) (SEQ ID NO: 5), которая кодирует гибридный белок FRCG (SEQ ID NO: 6). Таким образом, в химерном инсектицидном белке FRCG от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-467 белка Cry3A (SEQ ID NO: 68), составляющие вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6; а также 38 аминокислот протоксинного хвостового участка белка Cry1Ab.
Белок FRCG показал такую же активность против западного кукурузного корневого жука, как и белок FR8a (см. Таблицу 5), из чего можно предположить, что сайт протеазы катепсина G не является необходимым для инсектицидной активности сГИБа.
Пример 6. Разработка FR8a-9F
Первый фрагмент нуклеиновой кислоты размером приблизительно 323 бп увеличили способом ПЦР из плазмиды, содержащей FR8a (SEQ ID NO: 3), с применением праймеров reverse (SEQ ID NO: 89) и FR8a-OL-l (SEQ ID NO: 90), a также Смеси 2 и температурного цикла Профиля 2. Второй фрагмент нуклеиновой кислоты размером приблизительно 470 бп увеличили способом ПЦР из плазмиды, содержащей FR8a, при помощи праймеров FR8a-OL-2 (SEQ ID NO: 91) и С1-3А-2 (SEQ ID NO: 92), а также Смеси 2 и температурного цикла Профиля 2. Полученные в результате два ампликона соединили путем использования их в качестве шаблонов в расширенной ПЦР реакции с применением праймеров 5'FR8a (SEQ ID NO: 93) и С1-ЗА-2 (SEQ ID NO: 92), используя для ПЦР Смесь 2 и температурный цикл Профиля 2, для увеличения 5' конца последовательности FR8a-9F. Продукт расширенной ПЦР клонировали в вектор pCR2.1-TOPO (Invitrogen), обозначенный 5'FR-9F/pCR2.1. Фрагмент BamHI/PpuMI последовательности 5'FR-9F/pCR2.1 затем дотировали к фрагменту PpuMI/BamHI последовательности FR8a для создания последовательности FR8a-9F (SEQ ID NO: 7), кодирующей химерный белок FR8a-9F (SEQ ID NO: 8). Таким образом, в химерном инсектицидном белке FR8a-9F от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRP (SEQ ID NO; 129), аминокислоты 1-468 белка Cry3A055 (SEQ ID NO: 70), куда входят вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), куда входят С-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6; а также 38 аминокислот протоксинного хвостового участка белка Cry1Ab.
Активность сГИБа FR8a-9F против западного кукурузного корневого жука оказалась немного ниже, чем у сГИБа FR8a (См. Таблицу 5), из чего можно предположить, что C-концевые 9 аминокислот пептидильного фрагмента SEQ ID NO: 127 играют роль в наделении белка FR8a полной инсектицидной активностью.
Пример 7. Разработка FR-9F-catg
Кодирующая последовательность FR-9F-catg была создана для того, чтобы вернуть полученные из cry3A055 внерамочные нуклеотиды FR8a обратно в рамку и устранить сайт узнавания протеазы катепсин G. Фрагмент BamHI/PpuMI из 5TR-9F/pCR2.1 (См. Пример 6) лидировали с фрагментом PpuMI/BamHI последовательности FRCG (См. Пример 5) для создания кодирующей последовательности FR-9F-catg (SEQ ID NO: 9), которая кодирует химерный белок FR-9F-catg (SEQ ID NO: 10). Таким образом, в химерном белке FR-9F-catg от N-конца до С-конца располагаются: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRP (SEQ ID NO: 129), аминокислоты 1-467 белка Cry3A (SEQ ID NO: 68), куда входят вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6; а также 38 аминокислот протоксинного хвостового участка белка Cry1Ab.
Уровень активности сГИБа FR8a-9F-catg против западного кукурузного корневого жука оказался таким же, как у FR8a (См. Таблицу 5), это подтверждает то, что сГИБ может быть получен из последовательности либо модифицированного Cry3A, либо нативного Cry3.
Пример 8. Разработка FR8a-12аа
Нуклеотиды, кодирующие аминокислоты 2-13 пептидильного фрагмента, содержащегося в FR8a (SEQ ID NO: 4) извлекли при помощи ПЦР. Фрагмент увеличили способом ПЦР из плазмиды, содержащей FR8a (SEQ ID NO: 3), при помощи праймеров 5'FR8a-12aa (SEQ ID NO: 94) и С1-3А-2 (SEQ ID NO: 90), a также Смеси 1 и температурного цикла Профиля 1. Полученный в результате ампликон клонировали в pCR2.1-TOPO (Invitrogen). Фрагмент BamHI-PpuMI из клона pCR2.1-TOPO затем лигировали с фрагментом PpuMI-BamHI из плазмиды, содержащей FR8a для создания последовательности FR8a-12аа (SEQ ID NO; 11), которая кодирует химерный инсектицидный белок FR8a-12aa (SEQ ID NO: 12). Таким образом, в химерном инсектицидном белке FR8a-12aa от N-конца до С-конца располагаются: пептидильный фрагмент, содержащий последовательность аминокислот MYDGRQQHRG (SEQ ID NO: 128), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70), куда входят вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6; а также 38 аминокислот протоксинного хвостового участка белка Cry1Ab.
Уровень активности сГИБа FR8a-12aa против западного кукурузного корневого жука оказался таким же, как у FR8a (См. Таблицу 5), на основании чего можно предположить, что для полной инсектицидной активности FR8a не требуются N-концевые 12 аминокислот пептидильного фрагмента последовательности SEQ ID NO: 127.
Пример 9. Разработка Wr-9mut
Фрагмент нуклеиновой кислоты увеличили способом ПЦР из FR8a/pCR2.1 (Пример 2) при помощи праймеров 5'FR8a-12aa (SEQ ID NO: 94) и С1-3А-2 (SEQ ID NO: 92), а также Смеси 1 и температурного Профиля 2. Полученный в результате ампликон клонировали в pCR2.1-TOPO (Invitrogen). Затем фрагмент BamHI/PpuMI дотировали к фрагменту PpuMI/BamHI последовательности FR8a (SEQ ID NO: 3) для создания последовательности Wr-9mut (SEQ ID NO: 13), которая кодирует белок WR-9mut (SEQ ID NO: 14), в котором от N-конца до С-конца располагаются пептидильный фрагмент, содержащий последовательность аминокислот MYDGRQQHRG (SEQ ID NO: 128) и аминокислоты 10-598 белка Cry3A055 (SEQ ID NO: 70). Таким образом, белок WR-9mut представляет собой Cry3A055 с N-концевым пептидильным фрагментом по изобретению.
Белок WR-9mut не обладает активностью против западного кукурузного корневого жука. Следовательно, добавление N-концевого пептидильного фрагмента к негибридному модифицированному белку Cry3a уничтожило инсектицидную активность. Из этого можно сделать вывод, что, возможна некоторая взаимосвязь между C-концевой частью Cry1Ab гибридного белка FR8a и N-концевым пептидильным фрагментом, придающая инсектицидную активность белку FR8a.
Пример 10. Разработка FRD3
3' конец этой кодирующей последовательности создали путем увеличения способом ПЦР фрагмента из плазмиды, содержащей FR8a (SEQ ID NO: 3), с применением праймеров С2-3А-3 (SEQ ID NO: 95) и 3'1Ab-dm3 (SEQ ID NO: 96), а также Смеси 2 и температурного цикла Профиля 2. Полученный в результате ампликон клонировали в pCR2.1-TOPO (Invitrogen). Фрагмент ApaI/SacI клонированного ампликона размером 364 п.о., обозначенный 3'FRD3/pCR2.1, лигировали с фрагментом SacI/Apal последовательности FR8a для создания FRD3 (SEQ ID NO: 15), которая кодирует химерный белок FRD3 (SEQ ID NO: 16). В этом химерном белке FRD3 от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70), куда входят вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-610 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6. Таким образом, химерный инсектицидный белок FRD3 представляет собой вариант химерного инсектицидного белка FR8a, у которого удален участок из 38 аминокислот, представляющий собой протоксинный хвост Cry1Ab.
СГИБ FRD3 показал такой же уровень активности против западного кукурузного корневого жука, как и у FR8a (См. Таблицу 5), это дает основания предполагать, что хвостовой протоксинный участок из 38 аминокислот белка FR8a не является необходимым для полной инсектицидной активности.
Пример 11. Разработка FR-12-cg-dm3
Фрагмент SacI/PpuMI размером 3082 п.о. из плазмиды, содержащей FR8a-12 (См. Пример 8), фрагмент PpuMI/MluI размером 721 п.о. от последовательности FRCG (См. Пример 5) и фрагмент MluI/SacI размером 923 п.о. от последовательности FRD3 (См. Пример 10) скомбинировали для создания кодирующей последовательности FR-12-cg-dm3 (SEQ ID NO: 17), которая кодирует химерный белок FR-12-cg-dm3 (SEQ ID NO: 18). В этом химерном белке FR-12-cg-dm3 от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MYDGRQQHRG (SEQ ID NO: 129), аминокислоты 10-467 белка Cry3Aa (SEQ ID NO: 70), куда входят полный вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-610 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6. Таким образом, химерный белок FR-12-cg-dm3 представляет собой вариант FR8a, у которого удалены 12 N-концевых аминокислот пептидильного фрагмента, сайт узнавания протеазы катепсин G, а также 38 аминокислот протоксинного хвостового участка Cry1Ab.
Этот сГИБ FR-12-cg-dm3 не настолько активен против западного кукурузного корневого жука, как FR8a (См. Таблицу 5), из чего можно сделать вывод, что для полной инсектицидной активности требуется некоторое взаимодействие между C-концевой частью FR8a и N-концевым пептидильным фрагментом.
Пример 12. Разработка 9F-cg-de16
5'конец этой кодирующей последовательности получили путем увеличения способом ПЦР фрагмента из плазмиды, содержащей FR-9F-catg (См. Пример 7), с применением праймеров 5'FR-del6 (SEQ ID NO: 97) и С1-3А-2 (SEQ ID NO: 92), a также Смеси З и температурного цикла Профиля 3. Полученный в результате ампликон клонировали в pCR2.1-TOPO. Затем фрагмент BamHI/PpuMI размером 215 п.о. дотировали с фрагментом PpuMI/BamHI размером 4668 п.о. от последовательности FR-9F-catg для создания FR-9F-cg-del6 (SEQ ID NO: 19), которая кодирует химерный белок FR-9F-cg-del6 (SEQ ID NO: 20). В этом химерном белке FR-9F-cg-del6 от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MCAGIRP (SEQ ID NO: 130), аминокислоты 1-467 белка Cry3A (SEQ ID NO: 68), куда входят вариабельные участки 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6, а также 38 аминокислот протоксинного хвостового участка Cry1Ab. Таким образом, химерный белок FR-9F-cg-del6 представляет собой вариант FR8a-9F-catg, у которого удалены аминокислоты 2-7 пептидильного фрагмента.
Этот FR-9F-cg-del6 не обладает активностью против западного кукурузного корневого жука, из чего можно сделать вывод, что для активности против западного кукурузного корневого жука N-концевому пептидильному фрагменту нужны, по меньшей мере, 7 аминокислот из 9 C-концевых аминокислот последовательности SEQ ID NO: 127.
Пример 13. Разработка FR-cg-dm3
Фрагмент MluI/SacI размером 3839 п.о. от FRCG (Пример 5) и фрагмент MluI/SacI размером 923 п.о. от FRD3 (Пример 10) лидировали для создания FR-cg-dm3 (SEQ ID NO: 21), которая кодирует белок FR-cg-dm3 (SEQ ID NO: 22). В этом химерном инсектицидном белке FR-cg-dm3 от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-467 белка Cry3A (SEQ ID NO: 68), куда входят вариабельные участки 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-610 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6.
Одинаковый уровень активности у сГИБа FRD3 и у FR8a против западного кукурузного корневого жука (См. Таблицу 5) подтверждает, что сайт катепсина G и протоксинный хвостовой участок белка FR8a не требуются для полной инсектицидной активности против западного кукурузного корневого жука.
Пример 14. Разработка 9F-cg-dm3
Фрагмент MluI/SacI плазмиды, содержащей FR-9F-cg (См. Пример 7) датировали с фрагментом MluI/SacI размером 923 п.о. из плазмиды, содержащей FRD3 (См. Пример 10) для создания 9F-cg-dm3 (SEQ ID NO: 23), которая кодирует химерный белок 9F-cg-dm3 (SEQ ID NO: 24). В этом химерном белке 9F-cg-dm3 от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRP (SEQ ID NO: 129), аминокислоты 1-467 белка Cry3A (SEQ ID NO: 68), куда входят вариабельные участки 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-610 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6.
Уровень активности сГИБа 9F-cg-dm3 против западного кукурузного корневого жука оказался таким же (См. Таблицу 5), это подтверждает, что C-концевые 9 аминокислот пептидильного фрагмента могут придавать активность в том случае, если домен сГИБа состоит либо из вариабельных участков и консервативных блоков модифицированного Cry3A (Cry3A055), либо из вариабельных участков и консервативных блоков Cry3A.
Пример 15. Разработка В8а
Фрагмент нуклеиновой кислоты, кодирующий N-концевую часть белка Cry3A055 (SEQ ID NO: 70), увеличили способом ПЦР из плазмиды, содержащей cry3A055 (SEQ ID NO: 69), с применением праймеров 5'3А-1-bam (SEQ ID NO: 83) и С3-ЗА-8 (SEQ ID NO: 99), а также Смеси 1 и температурного цикла Профиля 1. Фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry1Ab (SEQ ID NO: 72), содержащий вариабельные участки 4-6 увеличили из плазмиды, содержащей mocrylAb (SEQ ID NO: 71), с применением праймеров С3-ЗА-7 (SEQ ID N0: 100) и 1Ab-6-sac (SEQ ID NO: 86), а также Смеси 1 и температурного цикла Профиля 1. Полученный в результате ампликон обозначили 20L-8b.
Фрагмент нуклеиновой кислоты, кодирующий N-концевую часть белка Cry3A055 (SEQ ID NO: 70), увеличили способом ПЦР из плазмиды, содержащей cry3A055 (SEQ ID NO: 69), с применением праймеров 5'3А-1-bam (SEQ ID NO: 83) и C2-3A-4 (SEQ ID NO: 88), а также Смеси 1 и температурного цикла Профиля 1.
Фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry1Ba (SEQ ID NO: 74) увеличили способом ПЦР из плазмиды, содержащей mocry1Ba (SEQ ID NO: 73), с применением праймеров 1В-5 (SEQ ID NO: 101) и 1В-10 (SEQ ID NO: 102), а также Смеси 1 и температурного цикла Профиля 1, с тем отличием, что применялась температура отжига 60°C.
Потом два описанных выше продукта ПЦР использовали в качестве шаблонов в расширенной ПЦР реакции с праймерами 5'3А-1-bam (SEQ ID NO; 83) и 1В-10 (SEQ ID NO: 102) и с применением Смеси 1 и температурного цикла Профиля 2. Полученный в результате ампликон обозначили В10.
Далее фрагмент нуклеиновой кислоты cry3A055 (SEQ ID NO: 69) увеличили способом ПЦР, используя 20L-8b (см. выше) в качестве шаблона, с праймерами 5'3А-1-bam (SEQ ID NO: 83) и 3А-22 (SEQ ID NO: 103) при следующих условиях ПЦР: Смесь 1, профиль температурного цикла: 94°C-45 секунд, градиент 50°C-70°C-45 секунд, 72°C-90 секунд, 30 циклов. Фрагмент другой нуклеиновой кислоты увеличили способом ПЦР, используя в качестве шаблона В 10 (см. выше), праймеры 75-7 (SEQ ID NO: 104) и 1 В-10 (SEQ ID NO: 102), с применением Смеси 1 раекции ПЦР и температурного цикла Профиля 2, с тем отличием, что применяли температуру отжига 60°C. Полученные в результате ПЦР два продукта затем использовали в качестве шаблонов в расширенной реакции ПЦР с праймерами 5'3А-1-bam (SEQ ID NO: 83) и 1 В-10 (SEQ ID NO: 102). с использованием следующих условий ПЦР: Смесь 2, профиль температурного цикла: 94°C-30 секунд, градиент 40°C-60°C-30 секунд, 72°C-60 секунд, 30 циклов.
Полученный в результате ПЦР продукт лидировали к вектору pCR2.1-TOPO (Invitrogen) и обозначили B10/pCR2.1. Фрагмент BamHi-SacI из B8a/pCR2.1 затем дотировали к рЕТ21а (Novagen), который был разрезан с помощью BamHi/SacI, для создания кодирующей последовательности В8а (SEQ ID NO: 25), которая кодирует гибридный белок В8а (SEQ ID NO: 26). Этот гибридный белок В8а содержит от N-конца до С-конца аминокислоты 1-468 белка Cry3A055 (SEQ ID NO: 70), а также аминокислоты 505-656 белка CrylBa (SEQ ID NO: 74).
Пример 16. Разработка 5*В8а
Фрагмент BamHI-XbaI из плазмиды, содержащей 20L-8a (См. Пример 3) и фрагмент XbaI-SacI из плазмиды содержащей В8а (См. Пример 15) лигировали для создания 5*В8а (SEQ ID NO: 27), кодирующей химерный белок 5*В8а (SEQ ID NO: 28). В этом белке 5*В8а от N-конца до С-конца располагаются: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-467 белка Cry3A055 (SEQ ID NO: 70) и аминокислоты 505-656 белка Cry1Ba (SEQ ID NO: 74). Таким образом, химерный белок 5*В8а представляет собой гибридный белок В8а, к которому добавили N-концевой пептидильный фрагмент.
Пример 17. Разработка V3A
Этот ген увеличили способом ПЦР, используя 3 фрагмента вместе в качестве шаблонов: первый шаблон увеличили из плазмиды, содержащей Cry3A055 (SEQ ID N0: 69) с применением праймеров 5'3A-l-bam (SEQ ID NO: 83) и С2-3А-4 (SEQ ID NO: 88), а также Смеси 1 и температурного цикла Профиля 1; второй фрагмент увеличили из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71), с применением праймеров С2-ЗА-3 (SEQ ID NO: 95) и С3-1Ab-2 (SEQ ID NO: 105), а также Смеси 1 и температурного цикла Профиля 1; а третий фрагмент увеличили из плазмиды, содержащей Cry3A055 (SEQ ID NO: 69), с применением праймеров С3-3А-5 (SEQ ID N0: 106) и 3А-12-sac (SEQ ID NO: 107), а также Смеси 1 и температурного цикла Профиля 1. Эти три продукта ПЦР затем использовали в качестве шаблонов в расширенной ПЦР реакции с праймерами 5'3A-l-bam (SEQ ID NO: 83) и 3А-12-sac (SEQ ID NO: 107), а также с применением Смеси 1 и Профиля 1 для получения кодирующей последовательности v3A (SEQ ID NO: 29), которая кодирует гибридный белок V3A (SEQ ID NO: 30). В гибридном белке V3A от N-конца до С-конца расположены: аминокислоты 1-226 белка Cry3A055 (SEQ ID NO: 70), куда входят вариабельный участок 1, консервативный блок 1, вариабельный участок 2 и N-концевые 34 аминокислоты консервативного блока 2, аминокислоты 237-474 белка Cry1Ab (SEQ ID NO: 72), куда входят C-концевые 33 аминокислоты консервативного блока 2, вариабельный участок 3 и N-концевые 20 аминокислот консервативного блока 3, а также аминокислоты 467-598 белка Cry3A055 (SEQ ID NO: 70), куда входят C-концевые 28 аминокислот консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5 и консервативный блок 5.
В сГИБе V3A содержатся две позиции кроссинговера. Первое скрещивание между Cry3A055 и Cry1Ab, расположено в консервативном блоке 2, а второе скрещивание между Cry1Ab и Cry3A055 расположено в консервативном блоке 3. Таким образом, V3A является вариантом Cry3A055, в котором весь вариабельный участок 3 заменили вариабельным участком 3 белка Cry1Ab. Этот сГИБ V3A оказался не столь активным против западного кукурузного корневого жука, как FR8a, на основании чего можно предположить, что наличие последовательности Cry1Ab в консервативном блоке 3, вариабельном участке 4, консервативном блоке 4, вариабельном участке 5, консервативном блоке 5 и/или вариабельном участке 6 является важным для полной инсектицидной активности белка FR8a.
Кодирующую последовательность v3A лигировали к вектору pCR2.1-TOPO, а затем субклонировали в рЕТ21а с помощью фрагмента BamHI/SacI. Белок V3A, экспрессированный вектором рЕТ21а, имеет метку Т7 на N-конце. Этот белок обозначили T7-V3A.
Пример 18. Разработка V4F
Первый фрагмент нуклеиновой кислоты, кодирующий участки 1-3 белка Cry3A055 увеличили способом ПЦР из плазмиды, содержащей Cry3A055 (SEQ ID NO: 69), с применением праймеров 5'3A-l-bam (SEQ ID NO: 83) и С3-3А-6 (SEQ ID NO: 84), а также Смеси 1 и температурного цикла Профиля 1.
Второй фрагмент нуклеиновой кислоты, кодирующий вариабельный участок 4 белка Cry1Ab, ПЦР увеличили из плазмиды, содержащей mocrylAb (SEQ ID NO: 71), с применением праймеров С3-1Ab-3 (SEQ ID NO: 85) и С4-3А-10 (SEQ ID NO: 108) а также Смеси 1 и температурного цикла Профиля 1.
Третий фрагмент нуклеиновой кислоты, кодирующий вариабельные участки 5-6 белка Cry3A055 увеличили способом ПЦР из плазмиды, содержащей Cry3A055 (SEQ ID NO: 69), с применением праймеров С4-ЗА-9 (SEQ ID NO: 109) и 3А-12-sac (SEQ ID NO: 107), а также Смеси 1 и температурного цикла Профиля 1.
Все три ампликона ПЦР скомбинировали и использовали в качестве шаблона в расширенной ПЦР с праймерами 53А-1-barn (SEQ ID NO: 83) и 3А-12-sac (SEQ ID NO: 107) с применением следующих условий ПЦР: Смесь 1 и профиль термоцикла: 94°C-30 секунд, градиент 50°C-70°C-30 секунд,, 72°C-30 секунд, 20 циклов. Полученный в результате ампликон v4F -последовательность (SEQ ID NO: 31), кодирующая гибридный токсин V4F (SEQ ID NO: 32), клонировали в вектор pCR2.1-TOPO и обозначили v4F/pCR2.1. Гибридный белок V4F содержит от N-конца до С-конца аминокислоты 1-468 белка Cry3A055 (SEQ ID NO: 70), аминокислоты 477-520, составляющие вариабельный участок 4 белка Cry1Ab (SEQ ID NO: 72), а также аминокислоты 512-598 белка Cry3A055 (SEQ ID NO: 70).
Белок V4F имеет две позиции кроссинговера. Первая позиция кроссинговера между Cry3A055 и Cry1Ab находится в консервативном блоке 3, а второе скрещивание между Cry1Ab и Cry3A055 - в консервативном блоке 4. Следовательно, белок V4F является вариантом Cry3A055, в котором весь вариабельный участок 4 заменен вариабельным участком 4 белка Cry1Ab. Гибридный белок V4F не активен против западного кукурузного корневого жука, это дает основание предполагать, что последовательность из Cry1Ab в C-концевой части FR8a вносит вклад в инсектицидную активность FR8a.
Фрагмент BamHI-SacI из v4F/pCR2.1 субклонировали в рЕТ21. Белок, экспрессированный результирующей плазмидой, обозначили T7-V4F.
Пример 19. Разработка 5*V4F
Фрагмент BamHI-XbaI из плазмиды, содержащей FR8a (См. Пример 4) и фрагмент XbaI-SacI из V4F/pCR2.1 (См Пример 18) лигировали к рЕТ21, разрезанному с помощью BamHI-SacI, в результате получили 5*V4F/pET21. Кодирующая последовательность 5*V4F(SEQ ID NO: 33) кодирует химерный белок 5*V4F (SEQ ID NO: 34). Этот химерный инсектицидный белок 5*V4F содержит от N-конца до С-конца пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-491 белка Cry3A055 (SEQ ID NO: 70), аминокислоты 501-520, составляющие вариабельный участок 4 белка CrylAb (SEQ ID NO: 72), а также аминокислоты 512-598 белка Cry3A055 (SEQ ID NO: 70).
СГИБ 5*V4F представляет собой гибридный белок V4F с добавленным N-концевым пептидильным фрагментом (SEQ ID NO: 127). Активностью против западного кукурузного жука сГИБ 5*V4F обладает, хотя и не на том же уровне, что FR8a. Следовательно, N-концевая часть придала инсектицидную активность белку V4F, подтверждая, что возможно наличие вносящего вклад взаимодействия между C-концевой частью и N-концевым пептидильным фрагментом FR8a.
Белок, экспрессированный плазмидой 5*V4F/pET21, обозначается T7-5*V4F и имеет метку Т7 N-концевую для пептидильного фрагмента 5*V4F.
Пример 20. Разработка 20L-7
Фрагмент нуклеиновой кислоты, кодирующий вариабельный участок 1 белка Cry3A055, увеличили способом ПЦР из плазмиды, содержащей cry3A 055 (SEQ ID NO: 69), с применением праймеров 5'3A-l-bam (SEQ ID NO: 83) и С1-ЗА-2 (SEQ ID NO: 92), а также Смеси 1 и температурного цикла Профиля 1.
Фрагмент нуклеиновой кислоты, кодирующий вариабельные участки 2-6 белка Cry1Ab увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71) с применением праймеров C1-1Ab-1 (SEQ ID NO: 110) и 1Ab-6-sac (SEQ ID NO: 86), а также Смеси 1 и температурного цикла Профиля 1.
Полученные в результате два ампликона использовали в качестве шаблонов для расширенной ПЦР реакции с праймерами 5'3A-1-bam (SEQ ID NO: 83) и 1Ab-6-sac (SEQ ID NO: 86) с применением Смеси 2 и температурного цикла Профиля 1 для создания кодирующей последовательности 20L-7 (SEQ ID NO: 35), которая кодирует гибридный белок 20L-7 (SEQ ID NO: 36). В этом гибридном белке 20L-7 от N-конца до С-конца расположены аминокислоты 1-156 белка Cry3A055 (SEQ ID N0: 70), составляющие вариабельный участок 1 и N-концевые 14 аминокислот консервативного блока 1, а также аминокислоты 167-648 белка Cry1Ab (SEQ ID NO: 72), составляющие C-концевые 15 аминокислот консервативного блока 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3, консервативный блок 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6, а также 38 аминокислот протоксинного хвостового участка Cry1Ab. Таким образом, 20L-7 является вариантом белка Cry1Ab, в котором вариабельный участок 1 заменен вариабельным участком 1 белка Cry3A055.
Кодирующую последовательность 20L-7 клонировали в pCR2.1-TOPO (Invitrogen), а затем переместили в рЕТ21 с помощью BamHI/SacI и обозначили 20L-7/pET21a. Кодирующая последовательность в 20L-7/pET21 была обозначена T7-20L-7 (SEQ ID NO: 37). Белок, экспрессируемый вектором 20L-7/pET21a, был обозначен T7-20L-7 (SEQ ID NO: 38).
Пример 21. Разработка 5*20L-7
Фрагмент BamHI/Xbal последовательности FR8a {См. Пример 4), фрагмент PpuMI/SacI последовательности 20L-7 (См. Пример 20) и фрагмент BamHI/SacI от рЕТ21а лидировали для получения 5*20L-7/pET21a. Кодирующая последовательность 5*20L-7 (SEQ ID NO: 39) кодирует химерный белок 5*20L-7 (SEQ ID NO: 40). Белок 5*20L-7 от N-конца до С-конца содержит пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-156 белка Cry3A055 (SEQ ID NO: 70), а также аминокислоты 167-643 белка Cry1Ab (SEQ ID NO: 72). Таким образом, гибридный белок 5*20L-7 представляет собой гибридный белок 20L-7, к которому прибавлен N-концевой пептидильный фрагмент.
Пример 22. Разработка 20L-10
Фрагмент нуклеиновой кислоты, кодирующий N-концевую часть белка Cry3A055, увеличили способом ПЦР из плазмиды, содержащей cry3A055 (SEQ ID NO: 69), с применением праймеров 53А-1-bam (SEQ ID NO: 83) и С2-3А-4 (SEQ ID NO: 88), а также Смеси 1 и температурного цикла Профиля 1. Фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry1Ab, увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71), с применением праймеров С2-3А-3 (SEQ ID NO: 95) и 1Ab-6-sac (SEQ ID NO: 86), a также Смеси 1 и температурного цикла Профиля 1. Эти два продукта ПЦР затем использовали в качестве шаблонов в расширенной ПЦР реакции с праймерами 5'3А-1-bam (SEQ ID NO: 83) и 1Ab-6-sac (SEQ ID NO: 86) при следующих условиях ПЦР: Смесь 2, профиль термоцикла: 94°C-30 секунд, градиент 45°C-65°C-30 секунд, 72°C-30 секунд, 20 циклов, в результате чего получили кодирующую последовательность 20L-10 (SEQ ID NO: 41), которая кодирует гибридный белок 20L-10 (SEQ ID NO: 42). Гибридный белок 20L-10 содержит от N-конца до С-конца аминокислоты 1-232 белка Cry3A055 (SEQ ID NO: 70) и аминокислоты 243-648 белка Cry1Ab (SEQ ID NO: 72). Таким образом, гибридный белок 20L-10 представляет собой по существу домен I белка Cry3A055 и домены II и III белка Cry1Ab.
Кодирующую последовательность 20L-10 клонировали в вектор pCR2.1-ТОРО (Invitrogen), который затем переместили в рЕТ21а с помощью BamHI/SacI. Белок, экспрессируемый 20L-10/pET21a получил обозначение T7-20L-10.
Пример 23. Разработка 5*20L-10
Фрагмент BamHI-Xbal из плазмиды, содержащей FR8a {См, Пример 4) и фрагмент XbaI-SacI из 20L-10/pCR2.1 (См. Пример 22) лигировали к рЕТ21, разрезанному с помощью BamHI-SacI, чтобы получить 5*20L-10/pET21. Кодирующая последовательность 5*20L-10 (SEQ ID NO: 43) кодирует химерный белок 5*20L-10 (SEQ ID NO: 44). Белок 5*20L-10 содержит от N-конца до С-конца пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-232 белка Cry3A055 (SEQ ID NO: 70) и аминокислоты 243-648 белка Cry1Ab (SEQ ID NO: 72). Таким образом, химерный белок 5*20L-10 представляет собой гибридный белок 20L-10 с прибавленным к нему N-концевым пептидильным фрагментом.
Пример 24.Разработка 20L-12A
Первый фрагмент нуклеиновой кислоты, кодирующий N-концевую часть белка Cry1Ab, увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71), с применением праймеров 5'1Ab-bam (SEQ ID NO: 98) и С3-1Ab-2 (SEQ ID NO: 105), а также Смеси 1 и температурного цикла Профиля 1.
Второй фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry3A055, увеличили способом ПЦР из плазмиды, содержащей Cry3A055 (SEQ ID NO: 69), с применением праймеров С3-3А-5 (SEQ ID NO: 106) и 3А-12-sac (SEQ ID NO: 107), а также Смеси 1 и температурного цикла Профиля 1.
Описанные выше первый и второй фрагменты нуклеиновой кислоты соединили путем использования их в качестве шаблонов в расширенной ПЦР реакции с праймерами 5'1Ab-bam (SEQ ID NO: 98) и 3А-12-sac (SEQ ID NO: 107). с применением Смеси 1 и температурного цикла Профиля 1, для создания кодирующей последовательности 20L-12A (SEQ ID NO: 45), которая кодирует сГИБ 20L-12A (SEQ ID NO: 46). Белок 20L-12A включает в себя от N-конца до С-конца аминокислоты 1-476 белка CrylAb (SEQ 1D NO: 72) и аминокислоты 469-598 белка Cry3A055 (SEQ ID NO: 70).
У сГИБа 20L-12A отсутствует активность против западного кукурузного корневого жука, но он активен против европейского кукурузного мотылька (см. Таблицу 6). Это демонстрирует, что сГИБ может быть разработан из белков, активных против чешуекрылых, и белков, активных против жесткокрылых, не теряя активности против чешуекрылых насекомых.
Кодирующую последовательность 20L-12A клонировали в pCR2.1-TOPO (Invitrogen), а затем переместили в рЕТ21а при помощи BamHI/SacI. Белок, экспрессируемый вектором 20L-12A/pET21a, обозначили T7-20L-12A.
Пример 25. Разработка 20L-13
Четыре нуклеиново-кислотных фрагмента получили следующим образом: фрагмент 1 увеличили способом ПЦР из плазмиды, содержащей Cry3A055 (SEQ ID N0: 69), с применением праймеров 5'3A-1-bam (SEQ ID NO: 83) и С1-3А-2 (SEQ ID NO: 92), а также Смеси 1 и температурного цикла Профиля 1; фрагмент 2 увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71), с применением праймеров С2-3А-3 (SEQ ID NO: 95) и С3-1Ab-2 (SEQ ID NO: 105). a также Смеси 1 и температурного цикла Профиля 1; фрагмент 3 увеличили способом ПЦР из плазмиды, содержащей mocrylAb (SEQ ID NO: 71) С3-1Ab-3, с применением праймеров (SEQ ID NO: 85) и С4-3А-10 (SEQ ID NO: 108), а также Смеси 1 и температурного цикла Профиля 1; а фрагмент 4 увеличили способом ПЦР из плазмиды, содержащей cry 3А 055 (SEQ ID NO: 69), с применением праймеров С4-3А-9 (SEQ ID NO: 109) и ЗА-12-sac (SEQ ID NO: 107), а также Смеси 1 и температурного цикла Профиля 1.
Затем все четыре фрагмента использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров 5'3А-bam (SEQ ID NO: 83) и ЗА-12-sac (SEQ ID NO: 107), а также Смеси 1 и температурного цикла Профиля 1, для создания кодирующей последовательности 20L-13 (SEQ ID NO: 47), которая кодирует гибридный токсин 20L-13 (SEQ ID NO: 48). Белок 20L-13 содержит от N-конца до С-конца аминокислоты 1-159 белка Cry3A055 (SEQ ID NO; 70), аминокислоты 170-522 белка Cry1Ab (SEQ ID NO: 72), а также аминокислоты 515-598 белка Cry3A055 (SEQ ID NO: 70). Таким образом, гибридный токсин 20L-13 содержит VI и N-концевую часть СВ1 из белка Cry3A055; c-концевую часть блока СВ1, V2, СВ2, V3, СВЗ и V4 из белка CrylAb; а также СВ4, V5 и СВ5 из белка Cry3A055.
Кодирующую последовательность 20L-13 клонировали в pCR2.1-TOPO (Invitrogen), а затем переместили в рЕТ21 с помощью BamHI/SacI. Белок, экспрессируемый вектором 20L-13/pET21a, был обозначен T7-20L-13.
Пример 26. Разработка 20L-20
Фрагмент BamHI/NspI из плазмиды, содержащей mocry3A (SEQ ID NO: 67), фрагмент NspI/HindIII из плазмиды, содержащей 20L-8A (SEQ ID NO: 1), и фрагмент HmdIII/BamHI из рЕТ21 а дотировали для получения 20L-20/pET21a.
Пример 27. Разработка У5и6
Фрагмент нуклеиновой кислоты, кодирующий N-концевую часть белка Cry3A055, увеличили способом ПЦР из плазмиды, содержащей Cry3A055 (SEQ ID N0: 69), с применением праймеров 5 '3А-1-bam (SEQ ID NO: 83) и С4-3А-10 (SEQ ID NO: 108), а также Смеси 1 и температурного цикла Профиля 1.
Фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry1Ab увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71), с применением праймеров С4-3А-9 (SEQ ID NO: 109) и 1Ab-6-sac (SEQ ID NO: 86), а также Смеси 1 и температурного цикла Профиля 1.
Эти два продукта ПЦР затем использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров 5'3А-1-bam (SEQ ID NO: 83) и 1Ab-6-sac (SEQ ID NO: 86), а также Смеси 1 и температурного цикла Профиля 2 для создания кодирующей последовательности V5 и 6 (SEQ ID NO: 49), которая кодирует гибридный токсин V5 и 6 (SEQ ID NO: 50). Белок V5 и 6 содержит от N-конца до С-конца аминокислоты 1-524 белка Cry3A055 (SEQ ID NO: 70), составляющие VI, СВ1, V2, СВ2, V3, СВ3, V4 и СВ4, а также аминокислоты 533-648 белка Cry1Ab (SEQ ID NO: 72), составляющие V5, СВ5 и V6, а также 38 аминокислот протоксинного хвостового участка Cry1Ab.
Кодирующую последовательность V5u6 клонировали в pCR2.1-TOPO, a затем переместили в рЕТ21 с помощью BamHI/SacI. Белок, экспрессируемый У5и6/рЕТ21а, обозначили Т7-V5 и б.
Пример 28. Разработка 5*V5 и 6
Фрагмент BamHI/XbaI последовательности FR8a (См. Пример 4), фрагмент XbaI/SacI последовательности V5u6 (См. Пример 27) и фрагмент BamHI/SacI из рЕТ21 а лигировали для получения 5*У5и6/рЕТ21, Кодирующая последовательность 5*V5 и 6 (SEQ ID NO: 51) кодирует химерный белок 5*V5 и 6 (SEQ ID NO: 52). В химерном инсектицидном белке 5*V5 и 6 от N-конца до С-конца расположены:
пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-524 белка Cry3A055 (SEQ ID NO: 70), составляющие VI, CB1, V2, CB2, V3, CB3, V4 и СВ4, аминокислоты 533-648 белка Cry1Ab (SEQ ID NO: 72), составляющие V5, CB5 и V6, а также 38 аминокислот протоксинного хвостового участка Cry1Ab. Таким образом, химерный инсектицидный белок 5*V5 и 6 представляет собой гибридный белок У5и6, к которому прибавлен N-концевой пептидильный фрагмент.
Пример 29. Разработка 88A-dm3
Фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry8Aa (SEQ ID NO: 78) увеличили способом ПЦР из плазмиды, содержащей cry8Aa (SEQ ID NO: 77), с применением праймеров 5'8Aa-dm3 (SEQ ID NO: 111) и 3'8Aa-dm3 (SEQ ID NO: 112), а также Смеси 2 и температурного цикла Профиля 2. Полученный в результате ампликон клонировали в pCR2.1-TOPO (Invitrogen) и обозначили 88A-dm3/pCR2.1.
Фрагмент MluI/SacI из 88A-dm3/pCR2.1 и фрагмент SacI/MluI из плазмиды, содержащей FR8a (См. Пример 4) лигировали для создания кодирующей последовательности 88A-dm3 (SEQ ID NO: 53), которая кодирует гибридный белок 88A-dm3 (SEQ ID NO: 54). В этот белок 88A-dm3 от N-конца до С-конца входят: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70) и аминокислоты 532-664 белка Cry8Aa (SEQ ID NO: 78).
Кодирующую последовательность 88A-dm3 также трансформировали в рЕТ21а при помощи рестрикции BamHI/SacI и лидирования. Белок, экспрессированный 88A-dm3/pET21, обозначили T7-88A-dm3.
Пример 30. Разработка FR(1Fa)
Фрагмент нуклеиновой кислоты, кодирующий N-концевую часть FR8a (См. Пример 3) увеличили способом ПЦР из плазмиды, содержащей FR8a (SEQ ID NO: 1), с применением праймеров С2-3А-3 (SEQ ID NO: 95) и tant-OL-2 (SEQ ID NO: 113), а также Смеси 3 и температурного цикла Профиля 3.
Фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry1Fa (SEQ ID NO: 76) увеличили способом ПЦР из плазмиды, содержащей mocry1Fa (SEQ ID NO: 75), с применением праймеров tant-OL-1 (SEQ ID NO: 114) и tant-3'sac (SEQ ID NO: 115), а также Смеси 3 и Температурного цикла Профиля 3.
Эти два продукта ПЦР затем использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров С2-3А-3 (SEQ ID NO: 95) и tant-3'sac (SEQ ID NO: 115), а также Смеси 3 и температурного профиля Профиля 3. Полученный в результате ПЦР продукт клонировали в pCR2.1-TOPO (Invitrogen). Фрагмент BamHI/MluI из плазмиды, содержащей FR8a, фрагмент MluVSacI из продукта расширенной ПЦР в pCR2.1, а также фрагмент BamHI/SacI из рЕТ21а затем лигировали для создания FR(lFa)/pET21a. Кодирующая последовательность FR(lFa) (SEQ ID NO: 55) кодирует химерный белок FR(lFa) (SEQ ID NO: 56). В белке FR(lFa) от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70) и аминокислоты 470-649 белка Cry1Fa (SEQ ID NO: 76).
Пример 31. Разработка FR(1Ac)
Домены I и II белка FR8a увеличили способом ПЦР из плазмиды, содержащей FR8a (SEQ ID NO: 1) с применением праймеров С2-3А-3 (SEQ ID NO: 95) и 1Ac-OL-2 (SEQ ID NO: 116), а также Смеси 3 и температурного цикла Профиля 3. Домен III белка Cry1Ac (SEQ ID NO: 80) увеличили способом ПЦР из плазмиды, содержащей cry1Ac (SEQ ID NO: 79), с применением праймеров 1Ac-OL-7 (SEQ ID NO: 117) и 1Ac-3'sac (SEQ ID NO: 118), а также Смеси 3 и температурного цикла Профиля 3.
Эти два продукта ПЦР использовали в качестве шаблонов в расширенной ПЦР с применением праймеров С2-3А-3 (SEQ ID NO: 95) и 1Ac-3'sac (SEQ ID NO: 118) и следующих условий реакции: Смесь 3 и профиль температурного цикла: 94°C-30 секунд, 68°C-30 секунд, 68°C-30 секунд, 20 циклов. Продукт расширенной реакции ПЦР клонировали в pCR2.1-TOPO (Invitrogen). Фрагмент BamHI/MluI из плазмиды, содержащей FR8a, фрагмент MluI/SacI из продукта расширенной ПЦР в pCR2.1, а также фрагмент BamHI/SacI из рЕТ21а дотировали для создания FR(lAc)/pET21a. В белке FR(lAc) от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO: 127), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70) и аминокислоты 477-608 белка CrylAc (SEQ ID NO: 80).
Пример 32. Разработка FR(1Ia)
Нуклеотидный фрагмент, кодирующий домены I и II белка FR8a, увеличили способом ПЦР из плазмиды, содержащей FR8a (SEQ ID NO: 3), с применением праймеров С2-3А-3 (SEQ ID NO: 95) 1Ia-OL-2 (SEQ ID NO: 119), а также Смеси 3 и температурного цикла Профиля 3. Второй нуклеотидный фрагмент, кодирующий домен III белка Cry1Ia (SEQ ID NO: 82), увеличили способом ПЦР из плазмиды, содержащей cry1Ia (SEQ ID NO: 81), с применением праймеров 1Ia-OL-1 (SEQ ID NO: 120) и 1Ia-3'sac (SEQ ID NO: 121), а также Смеси 3 и температурного цикла Профиля 3. Эти два продукта ПЦР использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров С2-3А-3 (SEQ ID NO: 95) и 1Ia-3'sac (SEQ ID NO: 121), а также Смеси 3 и профиля температурного цикла: 94°C-30 секунд, 68°C-45 секунд, 20 циклов. Продукт расширенной ПЦР клонировали в pCR2.1-TOPO (Invitrogen). Фрагмент BamHI/MluI из плазмиды, содержащей FR8a, фрагмент MluI/SacI из продукта расширенной ПЦР в pCR2.1, а также фрагмент BamHI/SacI из рЕТ21а дотировали для создания FR(1Ia)/pET21a. В белке FR(1Ia) от N-конца до С-конца расположены: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO; 127), аминокислоты 10-468 белка Cry3A055 (SEQ ID NO: 70) и аминокислоты 513-719 белка Cry1la (SEQ ID NO: 82).
Пример 33. Разработка Dm2-3A
Часть 5' конца этой кодирующей последовательности увеличили способом ПЦР из плазмиды, содержащей cry3A055 (SEQ ID NO: 69), с применением праймеров С2-3А-3 (SEQ ID NO: 95) и FR-1Ab-2 (SEQ ID NO: 122), а также Смеси 3 и температурного цикла Профиля 2. Нуклеотидный фрагмент, кодирующий домен III белка Cry1Ab, увеличили способом ПЦР из плазмиды, содержащей mocrylAb (SEQ ID NO: 71), с применением праймеров FR1Ab-1 (SEQ ID NO: 123) и 1Ab-6-sac (SEQ ID NO: 86), а также Смеси 3 и температурного цикла Профиля 2. Эти два продукта ПЦР использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров С2-ЗА-3 (SEQ ID NO: 95) и lAb-6-sac (SEQ ID NO: 86), a также Смеси 3 и температурного цикла Профиля 2. Полученный в результате ампликон клонировали в pCR2.1-TOPO (Invitrogen). Фрагмент BamHI/MluI из FR8a и вышеупомянутый продукт ПЦР в pCR2.1-TOPO AflIII, FR8a AflIII/SacI лидировали в BamHI/SacI pET21. Затем всю эту кодирующую последовательность (BamHI/SacI) переместили в 1454. В химерном инсектицидном белке DM2-3A от N-конца до С-конца располагаются: пептидильный фрагмент, содержащий последовательность аминокислот MTSNGRQCAGIRPYDGRQQHRG (SEQ ID NO:
127), аминокислоты 10-451 белка Cry3A055 (SEQ ID NO: 70), составляющие вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 7 аминокислот консервативного блока 3, а также аминокислоты 460-648 белка CrylAb (SEQ ID NO: 72), составляющие 41 С-концевую аминокислоту консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6. Таким образом, сГИБ DM2-3A имеет позицию кроссинговера между Cry3A055 и Cry1Ab, расположенную в консервативном блоке 3 сразу же после Ser451, что находится до соединения домена II с доменом III. DM2-3A обладает активностью против западного кукурузного корневого жука, но его активность меньше, чем у сГИБов SAF и FR8a как показано в Таблице 5.
Пример 34. Разработка T7-8AF
Фрагмент нуклеиновой кислоты, кодирующий N-концевую часть белка Cry3A055 (SEQ ID NO: 70), увеличили способом ПЦР из плазмиды, содержащей cry3A055 (SEQ ID NO: 69), с применением праймеров 5'3A-1-bam (SEQ ID NO: 83) и С3-3А-6 (SEQ ID NO: 84), а также смеси 1 и температурного цикла Профиля 1.
Фрагмент нуклеиновой кислоты, кодирующий C-концевую часть белка Cry1Ab (SEQ ID NO: 72), увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71) с применением праймеров С3-1Ab-3 (SEQ ID NO: 85) и 1Ab-6-Sac (SEQ ID NO: 86), а также Смеси 1 и температурного цикла Профиля 1.
Затем эти два вышеописанных продукта ПЦР использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров 5'3А-1-bam (SEQ ID NO: 83) и 1Ab-6-Sac (SEQ ID NO: 86), а также Смеси 2 и температурного цикла Профиля 1.
Полученный в результате ампликон лигировали как тупоконечный фрагмент к вектору pCR2.1-TOPO (Invitrogen, Carlsbad, СА), разрезанному с помощью Smal, для образования плазмиды p8AF/CR2.I. Затем фрагмент BamHI-SacI из p8AF/CR2.1 лигировали к рЕТ21а (EMD Biosciences, Inc., San Diego, СА), разрезанному с помощью BamHI-SacI, и трансформировали в Е.coli. Эта открытая рамка чтения обозначается T7-8AF (SEQ ID NO: 144) и кодирует гибридный белок T7-8AF (SEQ ID NO: 145). В гибридном белке T7-8AF от N-конца до С-конца располагаются: пептидильный фрагмент, содержащий последовательность аминокислот MASMTGGQQMGRGS (аминокислоты 1-14 из SEQ ID NO: 126), аминокислоты 1-468 белка Cry3A055 (SEQ ID NO: 70), составляющие вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), составляющие С-концевые 24 аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6, а также 38 аминокислот протоксинного хвостового участка белка Cry1Ab. Гибридный белок T7-8AF имеет очень слабую или нулевую инсектицидную активность против западного кукурузного корневого жука.
Пример 35. Разработка 8AF
Фрагмент BamHI-SacI из плазмиды p8AF/CR2.1 (См. Пример 34) дотировали к плазмиде, содержащей конститутивный промотор Cry 1 Ас, который был модифицирован из описанного в работе Schnepfy др. (1985. J. Biol. Chem. 260:6264-6272) с целью коррекции внутреннего стартового кодона ATG, который существует в кодоне по SchnepfM др., к кодону АТС, который был разрезан с помощью BamHI-SacI и трансформирован в Е. coli. Эта рамка чтения обозначается 8AF (SEQ ID NO: 63) и кодирует сГИБ 8AF (SEQ ID NO: 64). Этот сГИБ 8AF подобен сГИБу FR8a, но не содержит N-концевого пептидильного фрагмента. В сГИБе 8AF от N-конца до С-конца расположены: аминокислоты 1-468 белка Cry3A055 (SEQ ID NO: 70), составляющие вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 24 аминокислоты консервативного блока 3, а также аминокислоты 477-648 белка Cry1Ab (SEQ ID NO: 72), составляющие 24 C-концевые аминокислоты консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6, а также протоксинный хвостовой участок Cry1Ab из 38 аминокислот. Таким образом, сГИБ 8AF имеет позицию кроссинговера между Cry3A055 и Cry1Ab, расположенную в консервативном блоке 3 сразу же после Leu468 последовательности SEQ ID NO: 70, что находится после соединения домена II и домена III. Этот сГИБ 8АРобладает высокой активностью против западного кукурузного корневого жука.
Пример 36. Разработка - catG8AF
Была изготовлена модель без сайта катепсина G (Cat G) для того, чтобы определить, является ли наличие сайта Cat G в домене I сГИБа 8AF необходимым для активности против кукурузного корневого жука. Фрагмент BamHI/Sa1I размером 1359 п.о. из плазмиды, содержащей moCry3A (SEQ ID NO: 67) и фрагмент BamHI/SalI размером 3483 п.о. из плазмиды, содержащей 20L-8a (SEQ ID NO: 1) дотировали для создания -catG8AF (SEQ ID NO: 146), которая кодирует сГИБ -catG8AF (SEQ ID NO: 147).
Этот сГИБ -catG8AF очень активен против западного кукурузного корневого жука, это демонстрирует, что сайт узнавания протеазы катепсина G в сГИБе 8AF не является необходимым для инсектицидной активности.
Пример 37. Разработка 8AFdm3
Описанный в Примере 35 сГИБ 8AF имеет точку кроссинговера между Cry3A055 и Cry1Ab, расположенную в СВ3 после соединения доменов II/III, в результате чего домен III сГИБа SAP имеет маленький N-концевой участок домена III белка Cry3A055, а остальную часть домена III составляет последовательность домена III белка Cry1Ab. Для того, чтобы определить, требуется ли маленький N-концевой участок домена III белка Cry3A055 для инсектицидной активности 8AF, была изготовлена другая модель, имеющая скрещивание между Cry3A055 и Cry1Ab, расположенное в СВЗ точно на соединении доменаП с доменом III.
Фрагмент нуклеиновой кислоты, кодирующий часть домена I и домена II белка Cry3A055 увеличили способом ПЦР из плазмиды, содержащей FR8a (SEQ ID NO; 3), с применением праймеров CMS96 (SEQ ID NO: 138) и CMS97 (SEQ ID NO: 139), а также Смеси 5 и температурного цикла Профиля 5.
Фрагмент нуклеиновой кислоты, кодирующий домен III белка moCry1Ab увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71) с применением праймеров CMS98 (SEQ ID NO: 140) и CMS99 (SEQ ID NO: 141), a также Смеси 5 и температурного цикла Профиля 5.
Полученные в результате два ампликона использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров CMS96 (SEQ ID NO: 138) и CMS98 (SEQ ID NO: 140), а также Смеси 5 и температурного цикла Профиля 6. Полученный в результате ампликон клонировали в pCR4 Blunt (Invitrogen, Carlsbad, CA). Фрагмент StuI/SacI клонированного ампликона размером 1633 п.о., обозначенный pCR4Blunt-OLWrdm3 и фрагмент StuI/SacI размером приблизительно 3089 п.о. из плазмиды, содержащей Cry3A055 (SEQ ID NO: 69), скомбинировали для создания 8AFdm3 (SEQ ID NO: 148), кодирующей гибридный белок 8AFdm3 (SEQ ID NO: 149).
В гибридном белке 8AFdm3 от N-конца до С-конца располагаются: аминокислоты 1-454 белка Cry3A055 (SEQ ID NO: 70), составляющие домены I и II, включающие вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 10 аминокислот консервативного блока 3, а также аминокислоты 463-610 белка Cry1Ab (SEQ ID NO: 72), составляющие весь домен III, включающий C-концевые 38 аминокислот консервативного блока 3, вариабельный участок 4, консервативный участок 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6.
Таким образом, белок 8AFdm3 имеет позицию кроссинговера между Cry3A055 и Cry1Ab сразу же после Phe454 последовательности SEQ ID NO: 70, что находится на соединении домен II - домен III. Белок 8AFdm3 не обладает активностью против западного кукурузного корневого жука. Это дает основания предположить, что 24 аминокислоты N-концевого участка СВЗ белка Cry3A055 или Cry3A (поскольку они оба имеют одну и ту же последовательность на этом участке) являются необходимыми для активности сГИБа 8AF.
Пример 38. Разработка 8AFlongdm3
Для того чтобы определить, является ли критичной позиция кроссинговера в СВЗ между Cry3A или Cry3A005 и Cry1Ab для активности против кукурузного корневого жука, была изготовлена модель, в которой позиция кроссинговера расположена в СВ4 сразу же после аминокислоты 519 белка Cry3A055.
Фрагмент нуклеиновой кислоты, кодирующий часть домена I и весь домен II, а также часть домена III белка Cry3A055, увеличили способом ПЦР из плазмиды, содержащей Cry3A055 (SEQ ID NO: 69), с применением праймеров CMS96 (SEQ ID N0: 138) и CMS101 (SEQ ID NO: 143), а также Смеси 5 и температурного цикла Профиля 5.
Фрагмент нуклеиновой кислоты, кодирующий часть домена III белка Cry1Ab, увеличили способом ПЦР из плазмиды, содержащей mocry1Ab (SEQ ID NO: 71), с применением праймеров CMS98 (SEQ ID NO: 140) и CMS100 (SEQ ID NO: 142), а также Смеси 5 и температурного цикла Профиля 5,
Полученные в результате два ампликона использовали в качестве шаблонов в расширенной ПЦР реакции с применением праймеров CMS96 (SEQ ID NO: 138) и CMS98 (SEQ ID NO: 140), а также Смеси 5 и температурного цикла Профиля 6. Полученный в результате ампликон клонировали в pCR4 Blunt (Invitrogen, Carlsbad, CA). Фрагмент SalI/SacI клонированного ампликона размером 460 бп, обозначенный pCR4Blunt-OL8AFlongdm3, и фрагмент SalI/SacI размером 4265 бп из плазмиды, содержащей 8AFdm3 (SEQ ID NO: 147) скомбинировали для создания 8AFlongdm3 (SEQ ID NO: 150), которая кодирует гибридный белок 8AFlongdm3 (SEQ ID NO: 151).
В гибридном белке 8AFlongdm3 от N-конца до С-конца располагаются аминокислоты 1-519 белка Cry3A055 (SEQ ID NO: 70), составляющие домены I и II, куда входят вариабельный участок 1, консервативный блок 1, вариабельный блок 2, консервативный блок 2, вариабельный участок 3, консервативный блок 3, вариабельный участок 4 и N-концевые 6 аминокислот консервативного блока 4, а также аминокислоты 528-610 белка Cry1Ab (SEQ ID NO: 72), составляющие C-концевой участок домена III, включающий C-концевые 4 аминокислоты консервативного блока 4, вариабельный участок 5, консервативный блок 5 и вариабельный участок 6.
Таким образом, белок 8AFlongdm3 имеет позицию кроссинговера между Cry3A055 и Cry1Ab в консервативном блоке 4 сразу же после Ile519 of SEQ ID NO: 70. Гибридный Cry-белок 8AFlongdm3 не обладает активностью против западного корневого жука. Это дает основания предположить, что участок, критический для активности сГИБа Cry3A-CrylA против кукурузного корневого жука лежит в области между аминокислотами, соответственно от аминокислоты 6 из СВЗ до аминокислоты 7 из СВ4.
Пример 39. Разработка cap8AFdm3
Фрагмент BamHI/SalI размером приблизительно 1363 п.о. из плазмиды, содержащей 8AFdm3 (SEQ ID NO: 148), и фрагмент BamHI/SalI размером приблизительно 3362 п.о. из плазмиды, содержащей FR8a (SEQ ID NO: 3) дотировали для создания cap8AFdm3 (SEQ ID NO: 152), которая кодирует сГИБ cap8AFdm3 (SEQ ID NO: 153).
Белок cap8AFdm3 обладает некоторой активностью против западного кукурузного корневого жука, как показано в Таблице 5. Единственное отличие между гибридным белком 8AFdm3, не являющимся инсектицидным, и сГИБом cap8AFdm3 состоит в наличии N-концевого пептидильного фрагмента (SEQ ID NO: 127). Таким образом, прибавление пептидильного фрагмента к неактивному гибридному Cry-белку дало активный против кукурузного корневого жука гибридный инсектицидный белок.
Пример 40. Разработка 8APdm3T
Фрагмент PmlI/SacI размером приблизительно 4654 и.о. из плазмиды, содержащей 8AFdm3 (SEQ ID NO: 148), и фрагмент PmlI/SacI размером приблизительно 190 п.о. из плазмиды, содержащей FR8a (SEQ ID NO: 3), лигировали для создания 8AFdm3T (SEQ ID NO: 154), которая кодирует сГИБ 8AFdm3T (SEQ ID NO: 155). В сГИБе 8AFdm3T от N-конца до С-конца располагаются аминокислоты 1-454 белка Cry3A055 (SEQ ID NO: 70), составляющие домены I и II, куда входят вариабельный участок 1, консервативный блок 1, вариабельный участок 2, консервативный блок 2, вариабельный участок 3 и N-концевые 10 аминокислот консервативного блока 3, а также аминокислоты 463-610 белка Cry1Ab (SEQ ID NO: 72), составляющие весь домен III, куда входят C-концевые 38 аминокислот консервативного блока 3, вариабельный участок 4, консервативный блок 4, вариабельный участок 5, консервативный блок 5, вариабельный участок 6, а также протоксинный хвостовой участок Cry1Ab из 38 аминокислот.
Единственное отличие между гибридным белком 8AFdm3 и сГИБом 8AFdm3T состоит в добавке 38 аминокислот протоксинного хвостового участка Cry1Ab, отсюда можно сделать вывод, что прибавление протоксинного хвостового участка может изменить неактивный гибридный Cry-белок в активный сГИБ.
Пример 41. Разработка 8AFlongdm3T
Фрагмент PmlI/SacI размером приблизительно 4693 п.о. из плазмиды, содержащей 8AFlongdm3 (SEQ ID NO: 150) и фрагмент PmlI/SacI размером приблизительно 190 п.о. из плазмиды, содержащей FR8a (SEQ ID NO: 3), лигировали для создания 8AFlongdm3T (SEQ ID NO: 156), которая кодирует гибридный Cry-белок 8AFlongdmT (SEQ ID NO: 157).
Единственное отличие между гибридным Cry-белком 8AFlongdm3 и гибридным Cry-белком 8AFlongdm3T, не обладающим активностью против западного кукурузного корневого жука, состоит в добалении 38 аминокислот протоксинного хвостового участка Cry1Ab, это свидетельствует о том, что добавление протоксинного участка само по себе не достаточно для придания инсектицидной активности гибридному Cry-белку 8AFlongdm3. Это указывает на то, что для создания некоторых сГИБов может быть необходимой комбинация вариабельных участков и консервативных блоков в качестве добавки к протоксинному хвостовому участку и/или N-концевому пептидильному фрагменту.
Пример 42. Разработка cap8AFdm3 Т
Фрагмент PmlI/SacI размером приблизительно 4693 п.о. из плазмиды, содержащей cap8AFdm3 (SEQ ID NO: 152), и фрагмент PmlI/SacI размером приблизительно 190 п.о. из плазмиды, содержащей FR8a (SEQ ID NO: 3) дотировали для создания cap8AFdm3T (SEQ ID NO: 158), которая кодирует сГИБ cap8AFdm3T (SEQ ID NO: 159).
Белок cap8AFdm3T обладает более высокой активностью против западного кукурузного корневого жука, чем сГИБ cap8AFdm3, как показано в Таблице 5. Единственное различие между сГИБом cap8AFdm3, обладающим определенной активностью против западного кукурузного корневого жука, и сГИБом cap8AFdm3T является наличие 38 аминокислот протоксинного хвостового участка Cry1Ab. Следовательно, некоторые гибридные Cry-белки можно сделать активными путем прибавления N-концевого пептидильного фрагмента и протоксинного хвостового участка.
Пример 43. Испытание гибридных белков на инсектицидную активность. Западный кукурузный корневой жук
Гибридные белки, созданные в описанных выше примерах, испытали на инсектицидную активность против западного кукурузного корневого жука в лабораторных биопробах. Биопробы выполняли способом включения в пищу. Клоны Е.coli, экспрессирующие один из белков, выращивали в течение ночи. 500 мкл выращенной за ночь культуры обработали ультразвуком, и определили количество белка для испытания. Затем белковый раствор смешали с 500 мкл расплавленного искусственного питания, подобного описанному в работе Man-one и др. (1985, J. of Economic Entomology 78:290-293). После затвердения питания его разложили в чашку Петри, и на это питание поместили 20 новорожденных кукурузных корневых жуков. Температура чашек Петри поддерживалась приблизительно на уровне 30°C. Смертность записали через 6 дней.
Результаты биопроб показаны в Таблице 5. В столбце 1 указаны наименования гибридных Cry-белков, сконструированных гибридных инсектицидных белков и химерных инсектицидных белков. В столбце 2 указаны относительные уровни активности против западного кукурузного корневого жука ("-"=<40% смертности; "+"=40-49% смертности; "++"=50- 59% смертности; «+++"=60-80% смертности; и "++++"=>80% смертности). В столбце 3 указаны относительные уровни соответствующего белка, определенные по вестерн-блоттингу. Столбец 4 указывает наличие пептидильного фрагмента ("-"=нет пептидильного фрагмента; #1=SEQ ID NO: 126; #2=SEQ ID NO: 127; #3=SEQ ID NO: 128; #4=SEQ ID NO: 129; #5=-- SEQ ID NO: 130; #6=SEQ ID NO: 131; #7=SEQ ID NO: 132). Столбцы 5-7 показывают комбинации и расположение вариабельных участков (V1-V6), консервативных блоков (С1-С5) и соответствующих доменов (домены I-III) из первого Bt Cry-белка или модифицированного Cry-белка и второго Bt Cry-белка, отличающегося от первого Cry-белка или модифицированного Cry-белка, составляющих сердцевинный гибридный белок и не являющихся активными против западного кукурузного корневого жука, а также сГИБов, обладающих активностью против западного кукурузного корневого жука. В столбце 8 указано число аминокислот в протоксинном хвостовом участке, если он имеется, а также Cry-белок, из которого взяли этот хвостовой участок ("1Ab-38"=38 аминокислот в протоксинном хвосте из Cry1Ab; "1Ba-18"=18 аминокислот в протоксинном хвосте из Cry1Ba).
|
|
Химерные инсектицидные белки 20L-8a и FR8a, а также сГИБ 20L-12A испытали против различных видов насекомых для определения спектра активности. Среди насекомых, принимавших участие в испытаниях, были западный кукурузный корневой жук (ЗКЖ), северный кукурузный корневой жук (СКЖ), южный кукурузный корневой жук (ЮЮК), колорадский жук (КЖ) и европейский кукурузный мотылек (ЕКМ). Результаты биопроб представлены в Таблице 6. Знак «+» указывает на наличие инсектицидной активности. Знак «-» указывает на отсутствие инсектицидной активности. СГИБы 20L-8a и FR8a оказались активными против ЗКЖ, СКЖ и КЖ. СГИБ 20L-12A оказался удивительно активным против ЕКМ.
|
Пример 44. Введение генов, кодирующих сГИБы, в растения
Для трансформации в кукурузные растения были выбраны три гена, кодирующие химерные инсектицидные белки FR8a, FRCG и FRD3. Экспрессионную кассету, содержащую кодирующую последовательность FR8a, или FRCG, или FRD3, встроили в соответствующий вектор для агробактериальной трансформации в кукурузу. Для этого примера в экспериментах по трансформации применялись следующие векторы: 12207 (Фиг.3), 12161 (Фиг.4), 12208 (Фиг.5), 12274 (Фиг.6). 12473 (Фиг.7) и 12474 (Фиг.8).
Трансформация незрелых кукурузных зародышей выполнялась в целом так, как описано в работе Negrotto и др., 2000, Plant Cell Reports 19: 798-803. Для этого эксперимента все составляющие среды были в основном такими же, как описано в указанной выше работе Negrotto и др.. Однако можно использовать и другие составляющие, известные в отрасли.
Применяющиеся для трансформации гены клонировали в вектор, пригодный для трансформации кукурузы. Применяющиеся в этом примере векторы содержат ген изомеразы фосфоманнозы (ФМИ) для селекции трансгенных линий (Negrotto и др., выше).
Вкратце, штамм Agrobacterium LBA4404 (pSBl), содержащий плазмиду трансформации растения, выращивали на твердой среде YEP (дрожжевой экстракт (5 г/л), пептон (10 г/л), NaCl (5 г/л), агар (15 г/л), pH 6.8) в течение 2-4 дней при 28°C. Приблизительно 0.8Х 109 Agrobacterium суспендировали в среде LS-inf, дополненной 100 мкМ As (Negrotto и др., выше). Предварительная индукция бактерий в этой среде происходила в течение 30-60 мин.
Незрелые зародыши из А 18 8 или другого пригодного генотипа извлекают из початков возрастом 8-12 дней и помещают в жидкую среду LS-inf+100 µM As. Зародыши один раз споласкивают свежей инфекционной средой. Затем добавляют раствор Agrobacterium, и подвергают встряхиванию в течение 30 секунд, после чего дают возможность осесть с бактериями на 5 минут. Затем зародыши переносят заднеспинкой вверх в среду LSAs и культивируют в темноте в течение двух или трех дней. Затем от 20 до 25 зародышей на каждую чашку Петри переносят в среду LSDc с добавлением цефотаксима (250 мг/л) и нитрата серебра (1, 6 мг/л) и культивируют в темноте при 28°C в течение 10 дней.
Незрелые эмбрионы, образующие эмбриогенный каллус, перенесли в среду LSD1M0.5S. Селекция этих культур на этой среде осуществлялась в течение около 6 недель с этапом субкультивирования около 3 недель. Сохранившиеся каллусы перенесли в среду Regl с добавлением маннозы. После культивирования на свету (16 часов света/8 часов темноты) зеленые ткани перенесли в среду Reg2 без регуляторов роста и инкубировали 1-2 недели. Всходы перенесли в коробки Magenta GA-7 (Magenta Corp, Chicago 111.), содержащие среду Reg3, и выращивали на свету. Спустя 2-3 недели растения тестировали способом ПЦР на присутствие гена pmi, а также генов FR8a или FRCG. Растения, показавшие положительные результаты тестирования ПЦР, перенесли в теплицу и испытывали на сопротивляемость кукурузному корневому жуку.
Пример 45, Анализ растений трансгенной кукурузы на эффективность против кукурузного корневого жука: биопроба с извлечением корня
В типичном случае образцы растений кукурузы берут при их пересаживании из коробок Magenta GA-7 в почву. Это позволяет получить образцы корней из стерильной среды по сравнению с условиями почвы. Образцы берут путем отрезания небольшого кусочка корня (длиной ок. 2-4 см) и помещения его в небольшую чашку Петри с обогащенным фитагаром (фитагар,12 г, сахароза, 9 г, MS-соли, 3 мл, MS-витамины, 3 мл, нистатин (25 мг/мл), 3 мл, цефотаксим (50 мг/мл), 7 мл, ауреомицин (50 мг/мл), 7 мл, стрептомицин (50 мг/мл), 7 мл dH2O, 600 мл). Отрицательными контрольными образцами являются либо трансгенные растения, получившие отрицательный результат при тестировании ПЦР на присутствие гена FR8a или FRCG, из этого же эксперимента по трансформации, либо не трансгенные растения (такого же размера, как и тестовые растения), которые выращивались в фитотроне.
Образцы корней берут также после выращивания растений в почве. Если образцы корней берут из почвы, то эти кусочки корней промывают водой для удаления остатков почвы, окунают в раствор нистатина (5 мг/л), вынимают из раствора, высушивают бумажным полотенцем и помещают в чашку с фитагаром, как описано выше.
На образцы корней осуществили инокуляцию западного кукурузного корневого жука, поместив приблизительно 10 личинок первого возраста на внутреннюю поверхность крышки каждой из чашек с фитагаром, а затем вновь плотно закрыв крышки над образцом корня. Личинки переносили тонкой кисточкой. После инокуляции всех чашек лоток с чашками поместили в темноту с комнатной температурой до сбора данных.
Данные собрали приблизительно через 2-4 дня после инокуляции корней. Вычисляли процент смертности личинок, а также визуально оценивали степень повреждения корня. Степень повреждения определяли, исходя из числа обнаруженных прогрызенных личинками кукурузного корневого жука отверстий (ПО) в образце корня, и оценивали ее как высокую, среднюю, низкую или нулевую с присвоением категории, соответственно, 3, 2, или 1 (категория 1 присваивается при низкой степени повреждения корня или при отсутствии повреждений). В типичном случае, в растениях категории 1 имелось от 0 до 2 ПО, в растениях категории 2 - от 3 до 4 ПО, а в растениях категории 3 - более 5 ПО. Считали, что растения категории 1 по повреждениям - отличный результат, категория 2 -средний результат, категория 3 - слабый результат. Растения категории 1 отобрали для дальнейшего испытания в теплице и на поле.
Результаты в Таблице 7 показывают, что растения, экспрессирующие сГИБы FR8a и FRCG защищают корни от повреждений западным кукурузным корневым жуком. В большинстве случаев экспрессирующие химерный инсектицидный белок растения были отнесены к категории 1, в то время, как контрольные растения, не экспрессирующие химерный инсектицидный белок, относились к категории 3. Растения, экспрессирующие сГИБ FRD3, продемонстрировали сравнимые уровни борьбы против западного кукурузного корневого жука.
|
Пример 46. Анализ эффективности растений трансгенной кукурузы против кукурузного корневого жука в полевых условиях
Часть растений, получивших положительные оценки в описанных выше биопробах с извлечением корня, оценивали на поле. По восемнадцать растений из каждого опыта извлекли из полевых участков и оценивали степень повреждения их корней. Степень повреждения корней оценивалась на основе принятой в штате Айова линейной шкалы повреждения корней от 0 до 3 (Oleson, J.D. и др., 2005. J. Econ Entomol. 98(1): 1-8), где 0,00 = отсутствие повреждений (самый низкий возможный уровень); 1,00 = один узел (круг корней) или эквивалент целого узла, выеденный приблизительно до 11/2 дюйма стебля (линия почвы на 7-ом узле); 2,00 = два полных выеденных узла; 3,00=три или большее количество выеденных узлов (самый высокий возможный уровень); а повреждение, выражающееся в не целом числе узлов, отмечается как доля отсутствующего узла, т.е., 1,50=1 1/2 узла выедено, 0,25=1/4 одного узла выедено и т.п.
Результаты полевых испытаний против западного и северного кукурузного корневого жука показаны в Таблице 8, а против мексиканского кукурузного корневого жука - в Таблице 9. Вся трансгенная кукуруза, экспрессирующая химерный инсектицидный белок FR8a, показала результаты, лучшие, чем стандартный коммерческий химический инсектицид против западного, северного и мексиканского кукурузного корневого жука.
|
|










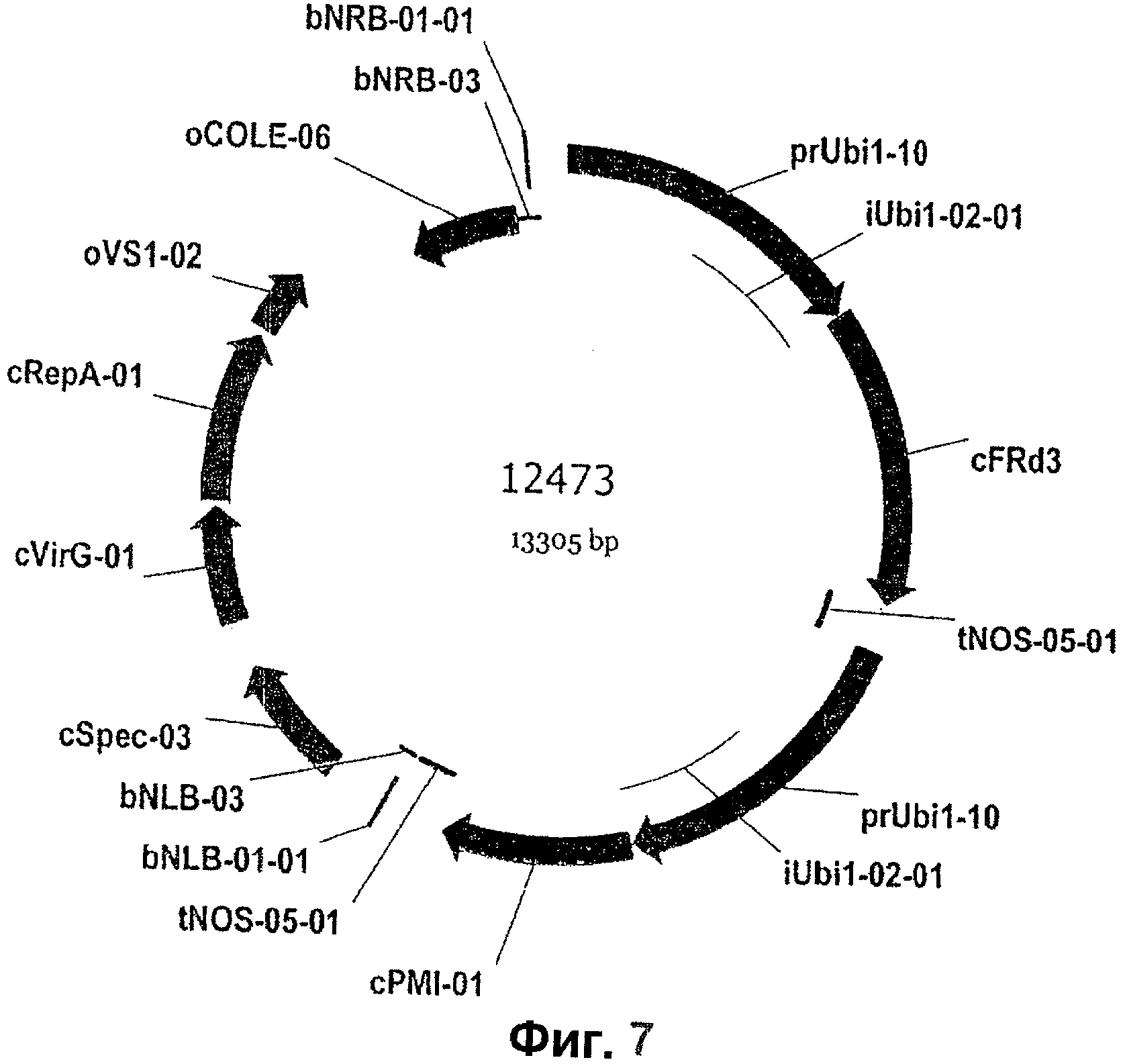


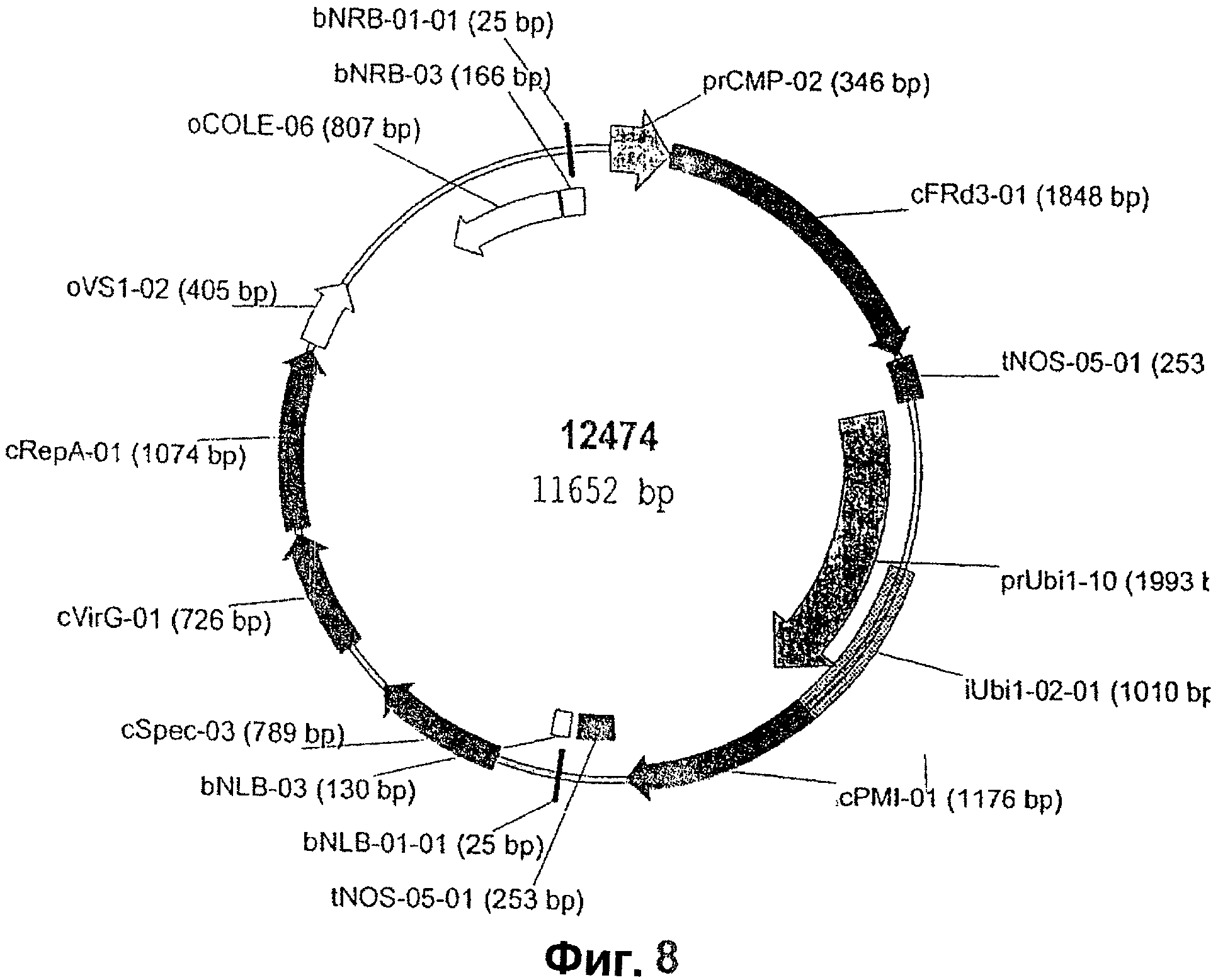































































































































































































































































































Производные иминопиридина и их применение в качестве микробиоцидов
Гербицидные композиции и их применение для борьбы с сорняками
Гербицидные композиции, способ борьбы с сорняками и способ борьбы с сорняками в сельскохозяйственной культуре
Инсектицидные соединения
Гибридный инсектицидный белок, молекула нуклеиновой кислоты, кодирующая такой белок, трансгенные растения и их семена, содержащие такой белок, способ получения белка и его применение
Композиция для контроля развития растений
Растение, устойчивое к насекомым
Производные иминопиридина и их применение в качестве микробиоцидов
Способ повышения и создания совместимой концентрации смеси обладающих электролитическими свойствами пестицидов в водном растворе и совместимая водная препаративная форма пестицидов
Растения кукурузы, отличающиеся локусам количественных признаков qtl
Производные иминопиридина и их применение в качестве микробиоцидов
Гербицидные композиции и их применение для борьбы с сорняками
Гербицидные композиции, способ борьбы с сорняками и способ борьбы с сорняками в сельскохозяйственной культуре
Инсектицидные соединения
Гибридный инсектицидный белок, молекула нуклеиновой кислоты, кодирующая такой белок, трансгенные растения и их семена, содержащие такой белок, способ получения белка и его применение
Композиция для контроля развития растений
Бисамидные производные и их применение в качестве инсектицидных соединений
Растение, устойчивое к насекомым
Производные иминопиридина и их применение в качестве микробиоцидов
Способ повышения и создания совместимой концентрации смеси обладающих электролитическими свойствами пестицидов в водном растворе и совместимая водная препаративная форма пестицидов

