Результат интеллектуальной деятельности: КОМПОЗИЦИЯ И СПОСОБ ЛЕЧЕНИЯ ОПУХОЛЕЙ
Вид РИД
Изобретение
Область изобретения
Опухолью называют аномальное разрастание ткани, которое может носить злокачественный или доброкачественный характер. Доброкачественные опухоли характеризуются только локальным ростом, тогда как злокачественная опухоль распространяется на другие ткани.
Рак относится к генетическим заболеваниям. Способность противостоять апоптозу является отличительной чертой раковых клеток. Известно, что многие генетические нарушения обеспечивают устойчивость раковых клеток к программируемой гибели клеток (апоптозу). Ферменты, называемые каспазами, осуществляют процесс разрушения клетки в ходе апоптоза. Инициаторные каспазы расщепляют неактивные формы предшественников каспаз-эффекторов, тем самым активируя их. Затем активированные каспазы-эффекторы начинают разрушать белки внутри клетки. В обычной ткани скорость апоптоза уравновешивается скоростью клеточного роста в ткани, и это регулирует рост ткани. В случае нарушения процесса апоптоза в клетке она может продолжать делиться, что приводит к неконтролируемому росту тканей.
Лиганд, индуцирующий апоптоз, связанный с фактором некроза опухоли (TRAIL), является перспективным терапевтическим реагентом для лечения опухолей. TRAIL входит в суперсемейство факторов некроза опухоли (TNF), включающее мощные индукторы апоптоза TNF и лиганд Fas (FasL). TRAIL представляет особый интерес не только потому, что он индуцирует быстрый апоптоз в самых разнообразных опухолях человека и линиях раковых клеток, но и потому, что он проявляет незначительную токсичность для обычных клеток и тканей.
Экспрессия TRAIL обычно осуществляется многими тканями организма, однако его можно вводить и экзогенно в ходе лечения опухоли. Введение растворимой активной формы TRAIL мышам с солидными опухолями индуцировало апоптоз, подавляло распространение опухоли и улучшало выживаемость. Имеются также свидетельства того, что комбинированное лечение с использованием TRAIL и различных химиотерапевтических композиций может еще более усилить регрессию опухоли.
TRAIL связывается с рецепторами смерти DR4 и DR5 на клетках опухоли и линиях раковых клеток и быстро и специфично вызывает апоптоз. В зависимости от типа клеток связывание TRAIL с рецептором смерти индуцирует либо и внешний, и внутренний каналы апоптоза, либо только один из этих каналов. В обоих каналах при связывании TRAIL с его рецептором формируется сайт связывания для адапторного белка FADD (Fas-связанный белок с доменом смерти), который захватывает и активирует инициаторную каспазу 8. В некоторых клетках каспаза 8 активирует внешний канал апоптоза, напрямую вызывая активирование каспаз-эффекторов 3, 6, и 7, что достаточно для индуцирования апоптоза. TRAIL может также индуцировать апоптоз по внутреннему каналу, где активация каспазы 8 инициирует апоптоз с участием митохондрий и образованием апоптосомы. Каспаза 8 расщепляет ингибирующий Bcl-2 белок, который содержит домен BH3 (Bid), вызывая транслокацию укороченного Bid из цитоплазмы во внешнюю мембрану митохондрии и индуцируя олигомеризацию Bax и Bak. Поры, образованные Bax (Bcl-2-связанный X белок) и Bak (Bcl-2-антагонист/киллер 1), приводят к потере поляризации митохондриальной мембраны и выбросу цитохрома c, вызывая метаболический отказ клеточных митохондрий. Затем цитохром с в цитоплазме взаимодействует с каспазой 9, с тем чтобы последовательно активировать каспазы-эффекторы 3, 6 и 7.
В то же время лечение с использованием соединений из суперсемейства TNF имеет определенные ограничения, связанные с тем, что различные клетки опухоли устойчивы по отношению к апоптозу, вызванному соединениями суперсемейства TNF, в том числе TRAIL. В настоящем изобретении рассматриваются такие ограничения.
Опубликованные результаты
Известно, что определенные молекулы, присутствующие в организме, могут подавлять активность соединений из суперсемейства TNF (такие вещества в дальнейшем называются супрессорами TNF). Например, сообщалось о том, что клеточный FLICE-ингибирующий белок (c-FLIPs), Bax ингибитор 1 и Bcl-XL подавляют апоптотическую активность TRAIL. Burns, T.F. and W.S. El-Deiry, J. Biol. Chem., 276: 37879-37886 (2001). Сообщалось также, что гены AKT и MIRSA тоже подавляют апоптотическую активность TRAIL. См., например, Aza-Blanc et al., Molecular Cell, 12: 627-637 (2003).
Кроме того, известно, что биологические молекулы, ингибирующие активность супрессора TNF, служат сенсибилизаторами клетки таким образом, чтобы повысить апоптотическую активность TRAIL. Например, показано, что введение миРНК, ориентированной на гены AKT и MIRSA в клетке, сенсибилизирует клетку по отношению к апоптотической активности TRAIL. См., например, Aza-Blanc et al., Molecular Cell, 12: 627-637 (2003).
Настоящее изобретение относится к созданию средств, функция которых также заключается в повышении апоптотической активности TRAIL.
Краткое описание изобретения
Настоящее изобретение относится к композиции, включающей: (A) ингибитор, который ингибирует активность белка комплекса III, например Core 1, и (B) носитель.
Кроме того, изобретение относится к способу получения композиции, включающей ингибитор, ингибирующий активность белка комплекса III, например Core 1, и носитель, включающий смешивание указанного ингибитора с указанным носителем.
В другом аспекте изобретение относится к способу получения композиции, в которую входят: ингибитор, ингибирующий активность белка комплекса III, например Core 1, носитель, а также член суперсемейства фактора некроза опухоли, включающий смешивание указанного ингибитора, указанного носителя и указанного члена суперсемейства.
Еще один аспект настоящего изобретения относится к способу проведения анализа с целью определить, является ли то или иное соединение ингибитором TNF-подавляющей активности белка комплекса III, например Core 1, включающий сравнение жизнеспособности клеток, экспрессирующих белок комплекса III, которые выращивали в присутствии члена суперсемейства TNF и приводили в контакт с указанным соединением, с жизнеспособностью клеток той же клеточной линии (также экспрессирующих указанный белок комплекса III), которые выращивали в отсутствие члена суперсемейства TNF и приводили в контакт с указанным соединением.
Следующий аспект настоящего изобретения относится к способу проведения анализа с целью определить, является ли то или иное соединение ингибитором TNF-подавляющей активности белка комплекса III, например Core 1, включающий приведение вещества в контакт с клетками, экспрессирующими указанный белок и которые выращивали в присутствии члена суперсемейства TNF, в течение заранее заданного промежутка времени; приведение вещества в контакт с клетками той же самой клеточной линии, которые экспрессируют белок и которые выращивали в отсутствие члена суперсемейства TNF, в течение такого же заранее заданного промежутка времени; и сравнение жизнеспособности этих клеток.
Еще один аспект настоящего изобретения относится к способу проведения анализа с целью определить, является ли то или иное соединение ингибитором активности белка комплекса III, например Core 1, включающий выращивание клеток, экспрессирующих белок из клеточной линии в присутствии члена суперсемейства TNF; выращивание клеток, экспрессирующих белок из той же клеточной линии в отсутствие члена суперсемейства TNF; приведение вещества в контакт с этими клетками; сравнение жизнеспособности клеток после заранее заданного промежутка времени.
Еще один дополнительный аспект настоящего изобретения относится к способу проведения анализа с целью определить, является ли то или иное соединение ингибитором TNF-подавляющей активности белка комплекса III, например Core 1, включающий сравнение жизнеспособности клеток, экспрессирующих указанный белок и которые приводили в контакт с членом суперсемейства TNF и этого соединения, с жизнеспособностью клеток той же клеточной линии, экспрессирующих указанный белок комплекса III и приводили в контакт с этим соединением, но в отсутствие члена суперсемейства TNF.
Другой дополнительный аспект настоящего изобретения относится к способу проведения анализа с целью определить, является ли то или иное соединение ингибитором TNF-подавляющей активности белка комплекса III, например Core 1, включающий приведение экспрессирующих белок клеток из клеточной линии в контакт с членом суперсемейства TNF и указанным соединением; приведение экспрессирующих белок клеток из той же клеточной линии в контакт с указанным соединением, но в отсутствие члена суперсемейства TNF; и сравнение жизнеспособности клеток.
Описание фигур
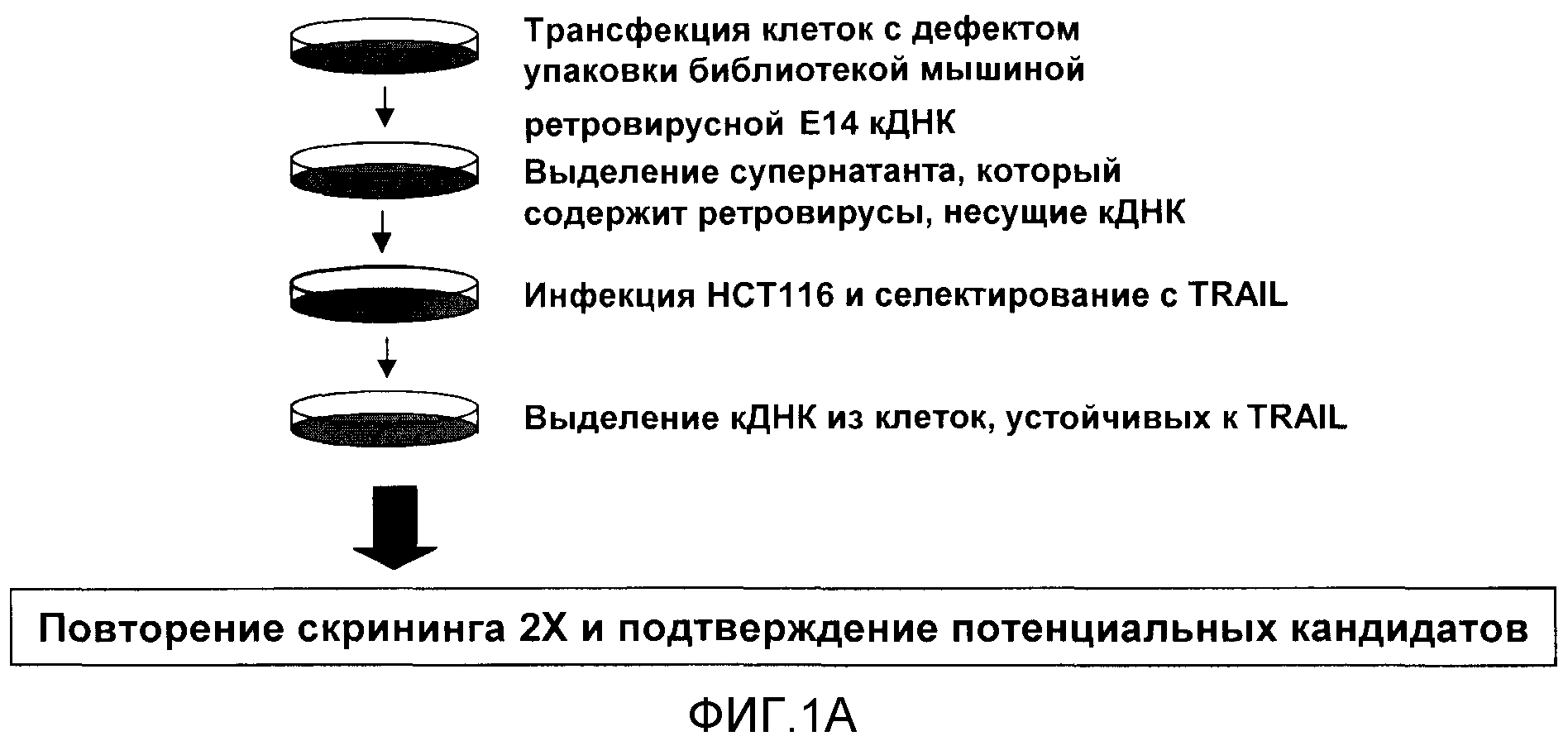
На фигуре 1A приводится схема, описывающая метод скрининга библиотеки кДНК для выявления генов, которые определяют резистентность к апоптозу, индуцированному TRAIL. На схематической диаграмме приводится генетический скрининг библиотеки мышиной кДНК E14 для выявления резистентности к апоптозу, индуцированному TRAIL. кДНК из клеток, устойчивых к TRAIL, были выделены, секвенированы и проверены.
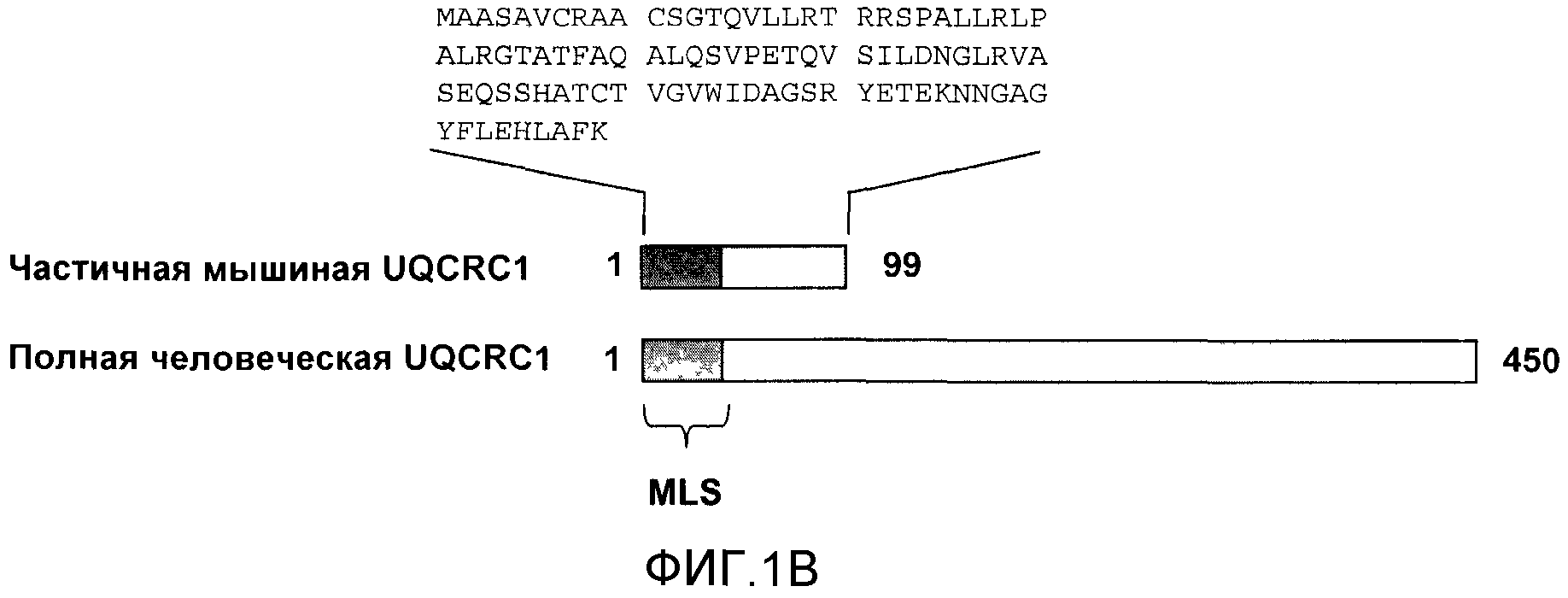
На фигуре 1B приводится частичная аминокислотная последовательность мышиной кДНК UQCRC1 (Core 1) в сравнении с полной последовательностью человеческой кДНК UQCRC1. Обе содержат последовательность митохондриальной локализации (MLS).
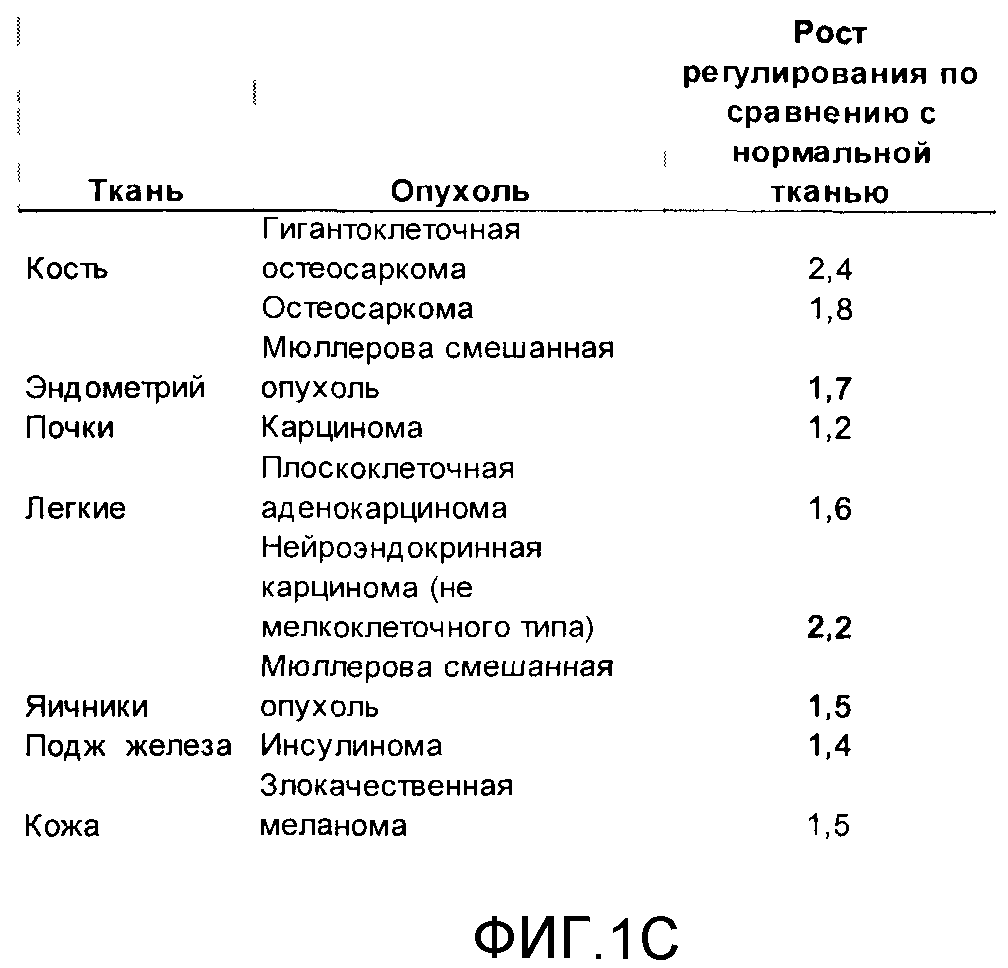
На фигуре 1C приводится график, демонстрирующий уровень сверхэкспрессии Core 1 в различных опухолевых тканях (из базы данных Ascenta).
Фигура 2A представляет собой гистограмму, отражающую жизнеспособность (что определяется процентом клеток, являющихся Annexin V-отрицательными) клеток HCT116, экспрессирующих доминантный отрицательный FADD (dnFADD), зеленый флуоресцентный белок (GFP) или частичный Core 1; эти клетки были обработаны TRAIL в концентрации 10 нг/мл или 50 нг/мл либо не приводились в контакт с TRAIL (0 нг/мл).
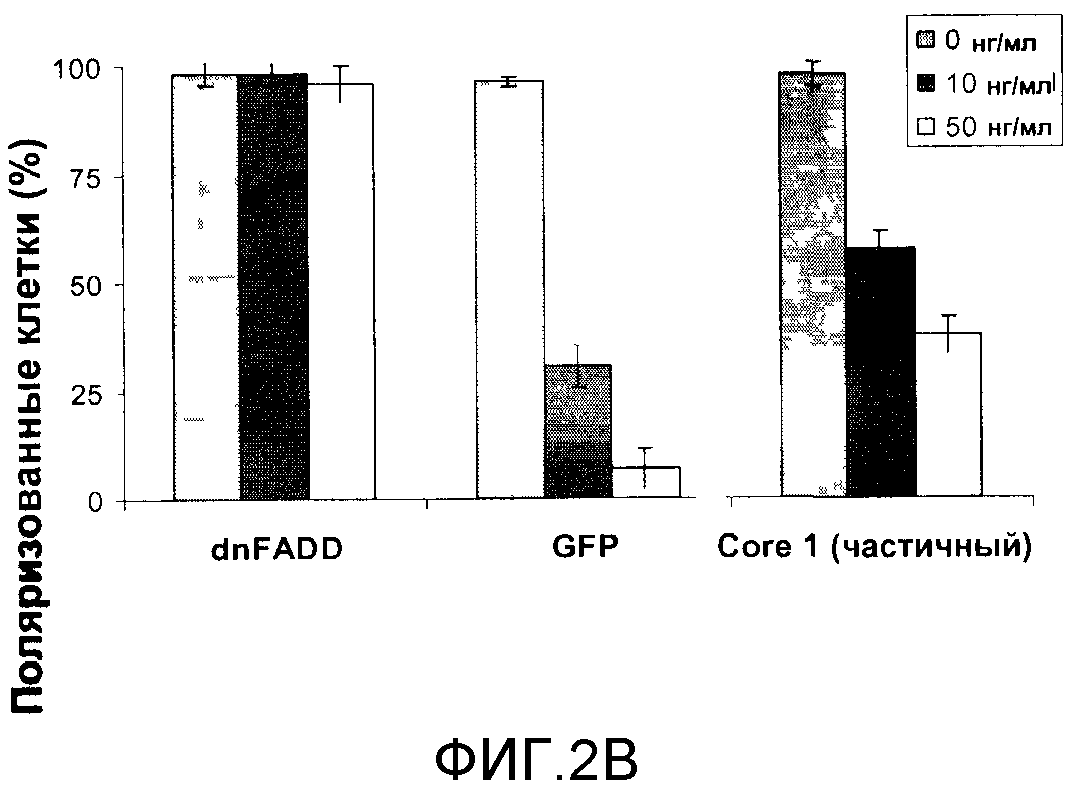
Фигура 2B представляет собой гистограмму, отражающую жизнеспособность (что определяется процентом поляризованных митохондрий в клетках) клеток HCT116, экспрессирующих доминантный отрицательный FADD (dnFADD), зеленый флуоресцентный белок (GFP) или частичный Core 1, которые были обработаны TRAIL в концентрации 10 нг/мл или 50 нг/мл либо не приводились в контакт с TRAIL (0 нг/мл).
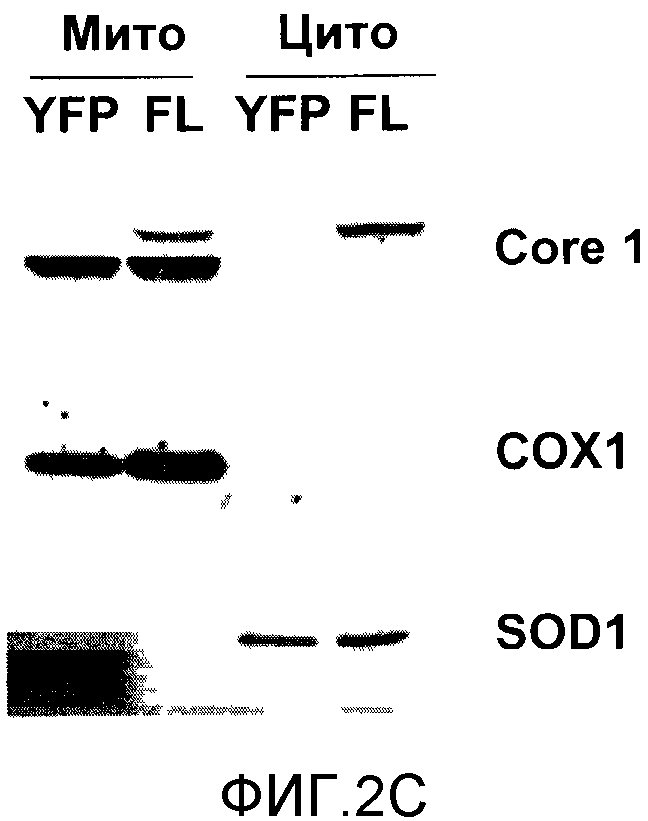
На фигуре 2C представлен вестерн-блоттинг, проведенный на цитоплазматических и митохондриальных фракциях, которые были приготовлены из стабильных клеточных линий, экспрессирующих желтый флуоресцентный белок (YFP) или полную последовательность человеческого Core 1 (FL), с использованием антител к Core 1, COX 1 (митохондриальный маркер) и SOD1 (цитоплазматический маркер).
На фигуре 2D представлена гистограмма, отражающая жизнеспособность (что определяется процентом клеток, являющихся Annexin V-отрицательными) клеток HCT116, экспрессирующих желтый флуоресцентный белок (YFP) или полную последовательность человеческого Core 1, которые были обработаны TRAIL в концентрации 10 нг/мл или 50 нг/мл либо не приводились в контакт с TRAIL (0 нг/мл).
На фигуре 3A представлен вестерн-блоттинг, проведенный на фракциях из стабильных клеточных линий HCT116, экспрессирующих желтый флуоресцентный белок (YFP) или полную последовательность человеческого Core 1 (FL), которые либо были обработаны 200 нг/мг TRAIL в течение 24 часов, либо не приводились в контакт с TRAIL, с использованием антител к каспазе 8, каспазе 9, каспазе 3 и тубулину (пятна снимались и повторно анализировались с использованием антител к тубулину в качестве контроля загрузки).
Фигура 3B представляет собой гистограмму, отражающую количество цитохрома с, присутствующего в клетках, экспрессирующих либо желтый флуоресцентный белок (YFP), либо Core 1, которые были обработаны 100 нг/мл TRAIL или не приводились в контакт с TRAIL (0 нг/мл). Количество цитохрома с в лизатах определялось иммуноферментным анализом ELISA.
Фигура 3С представляет собой гистограмму, отражающую жизнеспособность клеток, экспрессирующих желтый флуоресцентный белок (YFP) или Core 1, которые либо не подвергались обработке, либо были обработаны TRAIL (100 нг/мл) или FasL (16 нг/мл). Жизнеспособность клеток определялась по титру Cell Titer Blue.
Фигура 3D представляет собой гистограмму, отражающую жизнеспособность клеток, экспрессирующих желтый флуоресцентный белок (YFP) или полную последовательность человеческого Core 1, которые либо не подвергались обработке, либо были обработаны 200 нМ камптотецина, 10 нМ таксотера или DMSO. Жизнеспособность клеток определялась по титру Cell Titer Blue.
На фигуре 4A представлен вестерн-блоттинг, проведенный на фракциях из клеток HCT116 дикого типа, которые инкубировали только с липофектамином (контрольные клетки), 3,75 нМ Null миРНК или Core 1 миРНК в течение 72 часов, с использованием антител к Core 1 и тубулину (пятна снимали и повторно анализировали с использованием антител к тубулину в качестве контроля загрузки).
Фигура 4B представляет собой гистограмму, отражающую уровень экспрессии Core 1 в каждой из клеток, которые были описаны выше, на фигуре 4A.
На фигуре 4C приведена гистограмма, описывающая жизнеспособность клеток, которые ранее были обработаны только липофектамином, Null миРНК или Core 1 миРНК с последующей обработкой возрастающими концентрациями TRAIL.
На фигуре 5A приведена нуклеотидная последовательность человеческой кДНК UQCRC1 (SEQ ID NO: 1).
На фигуре 5B приведена аминокислотная последовательность человеческого Core 1 (SEQ ID NO: 2).
Подробное описание изобретения
Настоящее изобретение описывает применение ингибитора, который ингибирует TNF-подавляющую активность белка, относящегося к митохондриальному дыхательному комплексу III. Для простоты такой белок в настоящем документе называется белком комплекса III. Изобретение применимо к белку в той форме, в которой он связан с комплексом III, а также к белку, находящемуся в той форме, в которой он не связан с комплексом III, например, в выделенной форме. В одном из вариантов осуществления настоящего изобретения белком комплекса III является Core 1.
В состав настоящего изобретения входит фармацевтически эффективное количество ингибитора, который ингибирует TNF-подавляющую активность белка комплекса III, например Core 1. По сути, любое соединение, способное ингибировать TNF-подавляющую активность белка комплекса III, например Core 1, может использоваться для практического применения настоящего изобретения. Примеры соединений, которые могут использоваться для практического применения настоящего изобретения, включают: антисмысловую нуклеиновую кислоту, которая ингибирует экспрессию гена, кодирующего белок комплекса III, например Core 1; нуклеиновую кислоту, кодирующую антисмысловую нуклеиновую кислоту, которая ингибирует экспрессию гена белка комплекса III, например Core 1; миРНК, которая ингибирует экспрессию гена, кодирующего белок комплекса III, например Core 1; нуклеиновую кислоту, которая кодирует такую миРНК; антитело, которое способно связываться с белком комплекса III, например Core 1, и снижать его активность; нуклеиновую кислоту, кодирующую антитело, которое способно связываться с белком комплекса III, например, Core 1, и снижать его TNF-подавляющую активность; рибозим, который способен снижать уровень экспрессии белка комплекса III, например Core 1; нуклеиновую кислоту, кодирующую рибозим, который способен снижать уровень экспрессии белка комплекса III, например Core 1; соединение с малыми молекулами, которое способно снижать TNF-подавляющую активность белка комплекса III, например Core 1.
Ингибитор присутствует в композиции в концентрации, достаточной для снижения TNF-подавляющей активности белка комплекса III, например Core 1. Она может определяться рядовыми специалистами в данной области с учетом возможных колебаний эффективных концентраций в зависимости от конкретного используемого ингибитора, а также количеств и природы других компонентов композиции. В рамках общих рекомендаций предполагается, что в большинстве приложений будет использоваться ингибитор в количествах примерно от 0,01% до 15% от веса композиции. В некоторых вариантах осуществления ингибитор присутствует в количествах приблизительно от 0,01% до 10% от веса композиции, либо примерно от 0,1% до 5% от веса композиции.
В вариантах осуществления, где в качестве ингибитора используется антисмысловая нуклеиновая кислота, она гибридизуется по крайней мере с частью дезоксирибонуклеиновой кислоты (ДНК), кодирующей белок комплекса III, например Core 1, с тем чтобы за счет этого ингибировать экспрессию белка. Антисмысловой нуклеиновой кислотой может быть также кислота, которая снижает экспрессию гена, кодирующего белок комплекса III, например Core 1, за счет ингибирования сплайсинга его первичного транскрипта. Антисмысловая нуклеиновая кислота может содержать ДНК или рибонуклеиновую кислоту (РНК), либо обе кислоты. Антисмысловая последовательность любой длины достаточна для практического применения настоящего изобретения, при условии что она способна ингибировать экспрессию белка комплекса III, например Core 1. Предпочтительная антисмысловая последовательность имеет длину по меньшей мере 20 нуклеотидов. В предпочтительных вариантах осуществления антисмысловой нуклеиновой кислотой является олигонуклеотид.
Антисмысловая нуклеиновая кислота может быть получена синтетическим путем, например посредством экспрессии всей или части нуклеиновой кислоты, кодирующей белок комплекса III, например Core 1, в противоположной ориентации, как описано в European Patent № 140308. Последовательность нуклеиновой кислоты, кодирующей Core 1, приводится на фигуре 5A. Приготовление и использование синтетических антисмысловых нуклеиновых кислот в общей форме описано в International Application Publication WO 92/15680.
Антисмысловая нуклеиновая кислота может быть химически модифицирована, к примеру для улучшения стабильности и/или селективности. Например, одна из таких модификаций предусматривает замену свободного кислорода в фосфодиэфирной связи на сульфогруппу с образованием фосфоротиоатной связующей цепочки. Фосфоротиоатные антисмысловые нуклеиновые кислоты растворимы в воде, являются полианионами и устойчивы по отношению к эндогенным нуклеазам. Кроме того, можно синтезировать антисмысловые нуклеиновые кислоты с фосфорамидитными и полиамидными (пептидными) связями. Такие нуклеиновые кислоты обычно очень устойчивы к распаду в присутствии нуклеазы. Кроме того, химические группы могут добавляться к 2'-углероду сахара и 5-углероду (C-5) пиримидинов для увеличения стабильности антисмысловой нуклеиновой кислоты и способствования связыванию антисмысловой нуклеиновой кислоты с целевым сайтом. Модификации могут включать 2'-деокси, O-пентокси, O-пропокси, O-метокси, фтор, метоксиэтоксифосфоротиоаты, модифицированные основания, а также другие модификации, известные рядовым специалистам в данной области.
Антисмысловая нуклеиновая кислота присутствует в составе композиции в концентрации, достаточной для ингибирования экспрессии нуклеиновой кислоты, кодирующей белок комплекса III, например Core 1.
В тех вариантах осуществления, где нуклеиновая кислота, кодирующая антисмысловую нуклеиновую кислоту, используется в качестве ингибитора, такая нуклеиновая кислота относится к кислотам, кодирующим антисмысловую нуклеиновую кислоту описанного выше типа. Эта нуклеиновая кислота присутствует в составе композиции в концентрации, достаточной для экспрессии такого количества антисмысловой нуклеиновой кислоты, которое будет обеспечивать эффективное ингибирование экспрессии белка комплекса III, например Core 1.
В тех вариантах осуществления, где миРНК (малая интерферирующая РНК) используется в качестве ингибитора, такая миРНК гибридизуется с частью мРНК, кодирующей белок комплекса III, например Core 1, и способна включаться в индуцированный РНК сайленсинг-комплекс (RISC), который идентифицирует мРНК и ингибирует ее экспрессию. миРНК присутствует в составе композиции в концентрации, достаточной для ингибирования экспрессии нуклеиновой кислоты, кодирующей белок.
В еще одном варианте осуществления настоящего изобретения нуклеиновая кислота, кодирующая вышеуказанную миРНК, может использоваться в качестве ингибитора.
В тех вариантах осуществления, где в качестве ингибитора используется антитело, такое антитело связывается с Core 1 и ингибирует его TNF-подавляющую активность. Это антитело может быть, например, моноклональным или поликлональным, химерным, фрагментом Fab, фрагментом Fv, либо продуктом библиотеки экспрессии Fab или Fv.
Поликлональные антитела могут быть получены из антигенного фрагмента белка комплекса III, например Core 1, или полного белка. Моноклональные антитела могут быть получены, например, с помощью метода, изложенного в Mishell, B. B., et al., Selected Methods in Cellular Immunology, (W.H. Freeman, ed.) San Francisco (1980). Моноклональные антитела по настоящему изобретению могут быть «гуманизированы», чтобы исключить формирование иммунного ответа реципиента на антитела. «Гуманизированным антителом» называют такое антитело, в котором участки, определяющие комплементарность (CDR) и/или другие части каркасных доменов легкой и/или тяжелой вариабельной цепи взяты из не принадлежащего человеку иммуноглобулина, а остальные части молекулы взяты из одного или нескольких иммуноглобулинов человека. К гуманизированным также относятся антитела, для которых характерна гуманизированная тяжелая цепь, связанная с немодифицированной легкой цепью донора или акцептора либо с химерной легкой цепью, или наоборот. Гуманизация антител может проводиться с помощью известных специалистам методов (см., например, G.E. Mark and E.A. Padlan, "Chapter 4. Humanization of Monoclonal Antibodies", The Handbook of Experimental Pharmacology Vol. 113, Springer-Verlag, New York, 1994). Для экспрессии гуманизированных антител могут использоваться трансгенные животные.
Антитело может быть одноцепочечным. Известные специалистам методики получения одноцепочечных антител могут быть адаптированы для получения одноцепочечных антител к белку комплекса III, например Core 1.
Антитело присутствует в составе композиции в концентрации, достаточной для ингибирования активности белка комплекса III, например Core 1.
В тех вариантах осуществления, где нуклеиновая кислота, кодирующая антитело, используется в качестве ингибитора, такая нуклеиновая кислота относится к кислотам, кодирующим антитело, которое связывается с белком комплекса III, например Core 1, и ингибирующим его активность. Нуклеиновая кислота присутствует в составе композиции в концентрации, достаточной для экспрессии такого количества антитела, которое оказывается эффективным для ингибирования активности белка.
В тех вариантах осуществления, где в качестве ингибитора используется рибозим (энзим рибонуклеиновой кислоты), таким рибозимом является тот рибозим, который содержит каталитический домен и домен, связывающий субстрат. Домен, связывающий субстрат, гибридизуется по крайней мере с частью мРНК, которая кодирует белок комплекса III, например Core 1. Каталитическим доменом называют домен, который способен расщеплять мРНК.
В предпочтительных вариантах осуществления настоящего изобретения образующийся рибозим имеет молоточковый мотив. Другие формы имеют шпилечный мотив, мотивы вируса гепатита-дельта, интрона группы I или РНК РНКазы Р (в связи со вспомогательной последовательностью РНК) либо мотив РНК Neurospora VS. Молоточковые мотивы описаны в работе Rossi et al., Aids Research and Human Retroviruses, 8: 183 (1992). Шпилечные мотивы описаны в работе Hampel and Tritz Biochemistry, 28: 4929 (1989), и Hampel et al., Nucleic Acids Res., 18: 299 (1990). Мотив вируса гепатита-дельта описан в работе Perrotta and Been, Biochemistry, 31: 16 (1992); мотив РНКазы Р описан в работе Guerrier-Takada et al., Cell, 35: 849 (1983); мотив рибозима РНК Neurospora VS описан в работе Collins (Saville and Collins, Cell, 61: 685-696 (1990); Saville and Collins, Proc. Natl. Acad. Sci. USA, 88: 8826-8830 (1991); Collins and Olive, Biochemistry, 32, 2795-2799 (1993); мотив интрона группы I описан в Cech et al., US Patent № 4987071.
Одним из примеров метода получения рибозима является химический синтез олигодезоксирибонуклеотида с каталитическим доменом рибозима (-20 нуклеотидов) с боковыми последовательностями, которые после транскрипции гибридизуются с целевой мРНК Core 1. Амплификация олигодезоксирибонуклеотида проводится с использованием связывающих субстрат последовательностей в качестве праймеров. Амплифицированный продукт клонируется в эукариотический вектор экспрессии. Рибозим по настоящему изобретению может экспрессироваться в клетках эукариотов из соответствующего вектора ДНК. При желании активность рибозима может усиливаться посредством его выделения из первичного транскрипта вторым рибозимом (Ohkawa et al., Nucleic Acids Symp. Ser., 27: 15-6 (1992); Taira et al., Nucleic Acids Res., 19: 5125-30 (1991); Ventura et al., Nucleic Acids Res., 21, 3249-55 (1993).
Рибозим присутствует в составе композиции в концентрации, достаточной для ингибирования экспрессии нуклеиновой кислоты, кодирующей белок комплекса III, например Core 1.
В вариантах осуществления, в которых нуклеиновая кислота, кодирующая рибозим, используется в качестве ингибитора, такая нуклеиновая кислота относится к кислотам, кодирующим рибозим, который способен гибридизоваться с мРНК, кодирующей белок комплекса III, например Core 1, и расщеплять ее. Нуклеиновая кислота присутствует в составе композиции в концентрации, достаточной для получения рибозима в количестве, ингибирующем экспрессию белка. Количество может определяться рядовыми специалистами в данной области.
В вариантах осуществления, где используется соединение с малыми молекулами, ингибирующее активность белка комплекса III, например Core 1, таким соединением является соединение, которое сенсибилизирует клетки по отношению к апоптозу, индуцированному TRAIL, посредством ингибирования функции митохондриального комплекса III, например за счет ингибирования активности вышеуказанного белка. Соединение с малыми молекулами присутствует в композиции в концентрации, достаточной для ингибирования активности белка комплекса III, например Core 1, или для усиления эффекта воздействия TRAIL на клетки опухоли независимо от активности белка комплекса III в этих клетках.
В вариантах осуществления, в которых используется нуклеиновая кислота, кодирующая соединения с малыми молекулами, такая нуклеиновая кислота относится к тем кислотам, которые кодируют соединение с малыми молекулами, ингибирующим TNF-подавляющую активность белка комплекса III, например Core 1. Нуклеиновая кислота присутствует в составе композиции в концентрации, достаточной для получения вышеуказанного соединения в количестве, ингибирующем активность белка. Это может определяться рядовыми специалистами в данной области.
В вышеприведенных вариантах осуществления, которые предусматривают использование нуклеиновой кислоты, кодирующей антисмысловую нуклеиновую кислоту или белок, например антитело, рибозим или соединение с малыми молекулами, применяемую для ингибирования TNF-подавляющей активности белка комплекса III, например Core 1, такая нуклеиновая кислота может также содержать одну или несколько регуляторных областей, которые регулируют экспрессию кодирующей части нуклеиновой кислоты. Выбор соответствующей регуляторной области или областей представляет собой стандартную процедуру и не превышает уровня знаний рядовых специалистов в данной области. Регуляторные области включают промоторы, энхансеры, супрессоры и т.д.
К промоторам, которые могут использоваться в настоящем изобретении, относятся конститутивные промоторы и регулятивные (индуцируемые) промоторы. Примеры промоторов, применяемых на практике в настоящем изобретении: убиквитарные промоторы (например HPRT, виментин, актин, тубулин); промоторы промежуточного филамента (например десмин, нейрофиламенты, кератин, GFAP); терапевтические генные промоторы (например, MDR типа, CFTR, фактор VIII), а также тканеспецифические промоторы (например актиновый промотор в клетках гладкой мускулатуры или промоторы Flt и Flk, активные в эндотелиальных клетках).
К тканеспецифическим промоторам относятся транскрипционные контрольные области, которые проявляют специфичность по отношению к тканям, например: контрольная область гена эластазы I, проявляющая активность в ацинарных клетках поджелудочной железы (Swift et al., 1984, Cell 38:639-646; Ornitz et al., 1986, Cold Spring Harbor Symp. Quant. Biol., 50:399-409; MacDonald, 1987, Hepatology 7:425-515); контрольная область гена инсулина, проявляющая активность в бета-клетках поджелудочной железы (Hanahan, 1985, Nature 315:115-122); контрольная область гена иммуноглобулина, которая проявляет активность в лимфоидных клетках (Grosschedl et al., 1984, Cell 38:647-658; Adames et al., 1985, Nature 318:533-538; Alexander et al., 1987, Mol. Cell. Biol., 7:1436-1444); контрольная область опухоли молочной железы мышей, проявляющая активность в тестикулярных клетках, клетках грудной железы, лимфоидных и тучных клетках (Leder et al., 1986, Cell 45:485-495); контрольная область гена альбумина, которая проявляет активность в печени (Pinkert et al., 1987, Genes and Devel. 1:268-276); контрольная область гена альфа-фетопротеина, проявляющая активность в печени (Krumlauf et al., 1985, Mol. Cell. Biol., 5:1639-1648; Hammer et al., 1987, Science 235:53-58); контрольная область гена альфа-1-антитрипсина, которая проявляет активность в печени (Kelsey et al., 1987, Genes and Devel., 1:161-171); контрольная область гена бета-глобина, которая проявляет активность в миелоидных клетках (Mogram et al., 1985, Nature 315:338-340; Kollias et al., 1986, Cell 46:89-94); контрольная область гена основного белка миелина, которая проявляет активность в клетках олигодендроцитов мозга (Readhead et al., 1987, Cell 48:703-712); контрольная область гена легкой цепи миозина-2, проявляющая активность в скелетной мускулатуре (Sani, 1985, Nature 314:283-286), а также контрольная область гена гонадотропин-высвобождающего гормона, которая проявляет активность в гипоталамусе (Mason et al., 1986, Science 234:1372-1378).
К другим промоторам, которые могут использоваться в практических применениях по настоящему изобретению, относятся, например, промоторы, которые преимущественно активируются в делящихся клетках, промоторы, реагирующие на стимул (например рецептор стероидных гормонов, рецептор ретиноевой кислоты), тетрациклин-регулируемые транскрипционные модуляторы, предранний промотор цитомегаловируса, промоторы ретровирусного LTR, металлотионеина, SV-40, E1a и MLP. Тетрациклин-регулируемые транскрипционные модуляторы и промоторы CMV описаны в International Application Publication № WO 96/01313 и US Patent № 5168062 и 5385839.
В одном из вариантов осуществления настоящего изобретения в состав композиции входит вектор, который, в свою очередь, включает описанную выше нуклеиновую кислоту. Вектор представляет собой средство для переноса нуклеиновой кислоты в клетку. Термин «вектор» включает как вирусные, так и невирусные средства введения нуклеиновой кислоты в клетку. К невирусным векторам относятся, например, плазмиды, липосомы, электрически заряженные липиды (цитофектины), комплексы ДНК-белок и биополимеры. К вирусным векторам относятся, например, ретровирус, аденосателлитный вирус, векторы чумы, бакуловируса, осповакцины, симплекса герпеса, Эпштейна-Барра и аденовируса. Кроме нуклеиновой кислоты вектор может также содержать один или несколько селектируемых маркеров, которые применяются для выбора, измерения и мониторинга результатов переноса нуклеиновой кислоты (ткани, в которые осуществляется перенос, продолжительность экспрессии и т.д.).
Что касается использования вирусного вектора, он может быть дефектным по отношению к репликации, то есть неспособным к автономной репликации в целевых клетках. В общем случае в геноме дефектных по отношению к репликации вирусных векторов отсутствует по крайней мере одна область, которая необходима для репликации вируса в инфицированной клетке. Эти области можно либо удалить (полностью или частично), либо сделать нефункциональными с помощью любой методики, известной специалистам в данной области. К таким методикам относятся полное удаление, замена (другими последовательностями, в частности за счет введенной нуклеиновой кислоты), частичное удаление либо добавление одного или нескольких оснований к важной (для репликации) области. Такие методики могут осуществляться in vitro (на изолированной ДНК), или in situ, с использованием методик манипуляций с генами или за счет обработки мутагенными агентами. Предпочтительно, вирус, дефектный по отношению к репликации, сохраняет последовательности своего генома, которые необходимы для инкапсулирования вирусных частиц.
Что касается использования ретровирусов, то они интегрируют вирусы, которые инфицируют делящиеся клетки. Геном ретровируса включает две LTR, последовательность инкапсуляции и три кодирующие области (gag, pol и env). Структура рекомбинантных ретровирусных векторов описана, например, в European Patent № 453242 и 178220 и в Bernstein et al., Genet. Eng., 7: 235 (1985) and McCormick, BioTechnology, 3: 689 (1985). В рекомбинантных ретровирусных векторах гены gag, pol и env обычно удаляются, полностью или частично, и заменяются представляющей интерес последовательностью гетерологичной нуклеиновой кислоты. Такие векторы могут конструироваться из различных видов ретровируса, например, MoMuLV («вирус мышиного лейкоза Молони»), MSV («вирус мышиной саркомы Молони»), HaSV («вирус саркомы Харви»), SNV («вирус некроза селезенки»); RSV («вирус саркомы Рауса») и вирус Фрейнда. Системы векторов лентивируса могут также использоваться в практическом применении настоящего изобретения. Лентивирусный геном представляет собой полиаденилированную РНК с позитивным геномом с 9000-10000 пар оснований, содержащую три структурных гена с организацией от 5' до 3' (gag, pol, env), характерной для всех ретровирусов. Подробный обзор лентивирусных систем см. в Fields Virology, Second Edition, Volume 2, Chapter 55, "Lentiviruses," pp. 1571-1589, Raven Press, New York, 1990.
Как правило, для того чтобы построить рекомбинантные ретровирусы, содержащие нуклеиновую кислоту по настоящему изобретению, конструируется плазмида, которая содержит LTR, инкапсулирующую и кодирующую последовательности. Данная конструкция используется для трансфицирования линии клеток с дефектом упаковки, которая способна передавать ретровирусные функции, недостающие в плазмиде. Соответственно, линии клеток с дефектом упаковки обычно могут экспрессировать гены gag, pol и env. Такие линии клеток с дефектом упаковки описаны ранее, в частности, линия клеток PA317 (US 4861719); линия клеток PsiCRIP (International Application Publication № WO 90/02806) и линия клеток GP+envAm-12 (International Application Publication № WO 89/07150). Кроме того, рекомбинантные ретровирусные векторы могут содержать модификации внутри LTR для подавления транскрипционной активности, а также экстенсивных инкапсулирующих последовательностей, которые могут включать часть гена gag (Bender et al., J. Virol. 61 (1987) 1639). Рекомбинантные ретровирусные векторы очищают с помощью стандартных методик, известных рядовым специалистам в данной области.
Что касается использования аденоассоциированных вирусов (AAV), то они представляют собой ДНК-вирусы сравнительно небольшого размера, способные стабильным и сайт-специфичным образом интегрироваться в геном клеток, которые они инфицируют. Они могут инфицировать широкий спектр клеток, не индуцируя никаких эффектов клеточного роста, морфологии или дифференциации, и, по-видимому, не связаны с патологиями человека. Геном AAV был клонирован, секвенирован и охарактеризован. Он включает примерно 4700 оснований и содержит инвертированную терминальную повторяющуюся область (ITR) примерно из 145 оснований на обоих концах, которая служит в качестве исходной для репликации вируса. Остальная часть генома делится на две важные области, выполняющие функции инкапсуляции: левая часть генома, которая содержит ген rep, задействованный в репликации вируса и экспрессии генов вируса, и правая часть генома, которая содержит ген сар, кодирующий капсидные протеины вируса.
Использование векторов, полученных из AAV для передачи генов in vitro и in vivo, было описано ранее (см. International Application Publication № WO 91/18088 и WO 93/09239; US Patent №№ 4797368 и 5139941, и European Patent № 488528). В этих публикациях описаны различные конструкции из AAV, в которых гены rep и (или) cap удалены и заменены представляющим интерес геном, а также использование этих конструкций для передачи представляющих интерес генов in vitro (в культивированные клетки) или in vivo (непосредственно в организм). Используемый в настоящем изобретении дефектный по отношению к репликации рекомбинантный AAV может готовиться посредством котрансфицирования плазмиды, которая содержит представляющую интерес последовательность нуклеиновой кислоты, ограниченную двумя инвертированными терминальными повторяющимися областями (ITR) AAV, а также плазмиды, несущей гены инкапсуляции AAV (гены rep и cap), в линию клеток, инфицированных человеческим вирусом-помощником (например, аденовирусом). Полученные AAV рекомбинанты затем очищаются с помощью стандартных методик.
В предпочтительном варианте осуществления используемым вектором настоящего изобретения является аденовирусный вектор. Аденовирусы являются эукариотическими ДНК-вирусами, которые могут быть модифицированы для эффективной доставки нуклеиновой кислоты в клетки различного типа.
Существуют различные серотипы аденовируса. К предпочтительным серотипам для практического применения в настоящем изобретении относятся аденовирусы человека типа 2 или 5 (Ad 2 или Ad 5) либо аденовирусы животного происхождения (см. International Application Publication № WO 94/26914). К аденовирусам животного происхождения относятся, например, аденовирусы собаки, коровы, мыши (пример: Mav1, Beard et al., Virology 75: 81 (1990)), овцы, свиньи, птицы и обезьяны (пример: SAV). Предпочтительным аденовирусом животного происхождения является аденовирус собаки, более предпочтительным - аденовирус CAV2 (например, штамм Manhattan или A26/61 (ATCC VR-800)).
Предпочтительно аденовирусные векторы, дефектные по отношению к репликации, включают ITR, инкапсулирующую последовательность и представляющую интерес нуклеиновую кислоту. Еще более предпочтительно, чтобы по крайней мере область Е1 аденовирусного вектора была нефункционирующей. Делеция области Е1 предпочтительно распространяется на нуклеотиды с 455 по 3329 в последовательности аденовируса Ad5. Другие области также могут быть модифицированы, в частности: область E3 (см. International Application Publication № WO 95/02697); область E2 (см. International Application Publication № WO 94/28938); область E4 (см. International Application Publication № WO 94/28152, WO 94/12649 и WO 95/02697) или в любом из поздних генов L1-L5. Дефектные ретровирусные векторы рассматриваются в International Application Publication № WO 95/02697.
В предпочтительном варианте осуществления аденовирусный вектор имеет делеции в областях E1 и E4. В другом предпочтительном варианте осуществления аденовирусный вектор имеет делеции в области E1, в которую вставляются область E4 и последовательность, кодирующая представляющую интерес нуклеиновую кислоту (см. French Publication № 9413355).
Любая удобная методика может использоваться для приготовления рекомбинантных аденовирусов, дефектных по отношению к репликации. Примеры методик описаны в работах Levrero et al., Gene 101:195 (1991); European Patent № 185573; Graham, EMBO J., 3: 2917 (1984). В частности, аденовирусы могут быть получены посредством гомологичной рекомбинации между аденовирусом и плазмидой, которая, помимо всего прочего, включает представляющую интерес нуклеиновую кислоту. Гомологичная рекомбинация проводится после котрансфицирования указанного аденовируса и плазмиды в соответствующую линию клеток. Используемая линия клеток предпочтительно должна (i) быть трансформируемой под действием указанных элементов и (ii) содержать последовательности, которые способны дополнять часть генома аденовируса, дефектного по отношению к репликации, предпочтительно в интегральной форме, с тем чтобы избежать рисков рекомбинации. Примерами линий клеток, которые могут использоваться, являются линия клеток эмбриональной почки человека 293 (см. Graham et al., J. Gen. Virol., 36: 59 (1977)), содержащая левую часть генома аденовируса Ad5 (12%), интегрированную в его геном, а также линии клеток, которые способны дополнять функции E1 и E4, как описано в International Application Publication № WO 94/26914 и WO 95/02697. Рекомбинантные аденовирусы выделяются и очищаются при помощи стандартных методик молекулярной биологии, которые хорошо известны рядовым специалистам в данной области.
Для этих процедур использовались некоторые невирусные системы, которые способны упростить введение нуклеиновой кислоты в клетку.
Нуклеиновую кислоту можно ввести в клетку посредством липофекции, например с использованием липосомы. Применение катионных липидов может способствовать инкапсуляции отрицательно заряженных нуклеиновых кислот, а также слиянию с отрицательно заряженными клеточными мембранами (см. Felgner and Ringold, Nature 337:387-388 (1989)). Особенно удобные липидные соединения и составы для переноса нуклеиновых кислот описаны в International Application Publication № WO 95/18863 и WO 96/17823, а также в US Patent № 5459127. Применение липофекции для введения экзогенных генов в конкретные органы in vivo имеет определенные практические преимущества. К одной группе преимуществ относится молекулярное направленное воздействие липосом на конкретные клетки. Очевидно, что направление трансфицирования на конкретные виды клеток будет иметь особые преимущества для тканей с клеточной гетерогенностью, например поджелудочной железы, печени, почек и мозга. Для направленного воздействия липиды могут быть химически связаны с другими молекулами. Пептиды-мишени, например гормоны или нейротрансмиттеры, а также белки, например антитела, или непептидные молекулы, могут химически связываться с липосомами.
Для облегчения трансфицирования нуклеиновой кислоты in vivo также могут применяться другие молекулы, например катионный олигопептид (см., например, International Patent Application Publication № WO 95/21931), пептиды, полученные из ДНК-связывающих белков (см., например, International Patent Application Publication № WO 96/25508), и катионный полимер (см., например, International Patent Application Publication № WO 95/21931).
Существует также возможность введения нуклеиновой кислоты в клетку в форме чистого вектора нуклеиновой кислоты (см. US Patent № 5693622; 5589466 и 5580859). Чистые векторы нуклеиновых кислот для генной терапии могут вводиться в нужные клетки хозяина с применением методов, известных специалистам в данной области; к таким методам относятся, например, трансфицирование, электропорация, микроинъекция, трансдукция, слияние клеток, декстран DEAE, осаждение фосфатом кальция, использование генной пушки или применение транспортера ДНК вектора (см., например, Wilson et al., J. Biol. Chem. 267:963-967 (1992); Wu and Wu, J. Biol. Chem. 263:14621-14624 (1988); Hartmut et al., Canadian Patent № 2,012,311; Williams et al., Proc. Natl. Acad. Sci. USA 88:2726-2730 (1991)). Могут также использоваться рецепторные подходы к доставке ДНК (Curiel et al., Hum. Gene Ther. 3:147-154 (1992); Wu and Wu, J. Biol. Chem. 262:4429-4432 (1987)).
В настоящем изобретении предлагаются составы, которые включают ингибитор, описанный в настоящем изобретении, и носитель. Носитель представляет собой среду, в которой ингибитор сохраняет свою активную форму, к примеру форму, способную проявлять биологическую активность. Например, нуклеиновая кислота будет способна к репликации, передаче сообщения или гибридизации в комплементарную нуклеиновую кислоту; вектор будет способен к трансфицированию целевой клетки; антитело будет связывать Core 1. Как правило, таким носителем будет водный буфер, например, буфер Tris, фосфатный или HEPES, содержащий ионы солей. Как правило, концентрация ионов солей будет соответствовать физиологическим уровням. В рамках общих рекомендаций предполагается, что в большинстве приложений будет использоваться носитель в количествах примерно от 40% до примерно 98% от веса композиции. В некоторых вариантах осуществления носитель присутствует в количествах примерно от 50 примерно до 98% от веса композиции.
Композиция также может включать стабилизирующие агенты, консерванты и другие наполнители.
Состав композиции по настоящему изобретению может создаваться для перорального и парентерального введения (например местного, интраназального, подкожного и внутриглазного применения). Подразумевается, что парентеральное введение включает внутривенную инъекцию, внутримышечную инъекцию, внутриартериальную инъекцию или инфузионные методики. Композиция может вводиться парентерально в единицах дозы, содержащей стандартные, хорошо известные нетоксичные физиологически приемлемые носители, адъюванты и среды - в зависимости от необходимости.
Предпочтительным стерильным инъекционным составом может быть раствор или суспензия ингибитора в нетоксичном, приемлемым для парентерального введения носителе. Примерами фармацевтически приемлемых носителей являются физиологический раствор, буферный физиологический раствор, изотонический физиологический раствор (например, мононатрий или динатрийфосфат, хлорид натрия, калия, кальция или магния либо смесь таких солей), раствор Рингера, декстроза, вода, стерильная вода, глицерин, этанол и их сочетание. 1,3-бутандиол и стерильные жирные масла удобно использовать в качестве растворителей или суспендирующих сред. Может применяться любая смесь жирных масел, в том числе синтетические моно- или диглицериды. В композициях для инъекций применение находят также жирные кислоты, такие как олеиновая кислота.
Носителем также может быть гидрогель, который получается из любого биосовместимого или нецитотоксичного (гомо- или гетеро-) полимера, например, гидрофильный полимер полиакриловой кислоты, который может выступать в качестве губки, на поверхности которой адсорбируется ингибитор. Подобные полимеры были описаны, например, в International Application Publication № WO 93/08845. Гидрогель может наноситься непосредственно на поверхность обрабатываемой ткани.
Другой предпочтительный вариант осуществления настоящего изобретения включает фармацевтическую композицию, содержащую в качестве носителя полоксамер, который пропитывается дефектным по отношению к репликации рекомбинантным вирусом, включающим, в соответствии с настоящим изобретением, нуклеиновую кислоту. Предпочтительным полоксамером является Poloxamer 407, который производится серийно (BASF, Parsippany, NJ) и представляет собой нетоксичный биосовместимый полиол. Полоксамеры обладают, по существу, теми же преимуществами, что и гидрогели, при более низкой вязкости.
Настоящее изобретение обеспечивает способ лечения, предусматривающий введение человеку или другому животному эффективного количества композиции по настоящему изобретению. Эффективные количества композиции могут меняться в зависимости от возраста, характера и остроты заболевания, которое подлежит лечению, массы тела, желательной продолжительности лечения, метода введения и других параметров. Эффективные количества определяются врачом или другим квалифицированным медицинским работником.
Настоящее изобретение также обеспечивает способ анализа соединений, позволяющий определить, способны ли они действовать как ингибиторы TNF-подавляющей активности белка комплекса III, например, Core 1. В ходе анализа жизнеспособность клеток, которые экспрессируют белок комплекса III (например Core 1), проявляют природную устойчивость к TNF-индуцированному апоптозу и приводятся в контакт с членом суперсемейства TNF (в дальнейшем TNF) и указанного соединения, сопоставляется с жизнеспособностью контрольных клеток из той же линии клеток, которые также экспрессируют белок и приводятся в контакт с указанным соединением, но в отсутствие TNF. Жизнеспособность может определяться с использованием титра Cell Titer Blue Assay, который измеряет метаболический потенциал (и тем самым жизнеспособность) клеток по их способности восстанавливать краситель резазурин до его высокофлуоресцентной формы резоруфина, которая может детектироваться с помощью спектрофотометра.
Если соединение значительно снижает жизнеспособность клеток, которые приводились в контакт с TNF, и контрольных клеток, то считается, что активность соединения не связана с TNF, и соединение не относят к представляющим интерес. Если соединение существенно снижает жизнеспособность клеток, которые контактировали с TNF, но не жизнеспособность контрольных клеток, то считается, что оно сенсибилизирует клетки по отношению к апоптотической активности TNF, поскольку было известно, что данная клеточная линия ранее проявляла устойчивость по отношению к TNF. Поскольку клетка экспрессирует белок комплекса III, делается вывод о том, что анализируемое соединение действует посредством ингибирования TNF-подавляющей активности белка комплекса III.
Считается, что соединение значительно снижает жизнеспособность клетки, если жизнеспособность клеток, которые приводились в контакт с TRAIL, и анализируемого соединения оказывается по крайней мере на 30% ниже по сравнению с жизнеспособностью контрольных клеток.
В одном из вариантов осуществления настоящего изобретения клетки, экспрессирующие белок комплекса III, например Core 1, и которые относятся к линии клеток, известных своей природной устойчивостью по отношению к TNF-индуцированному апоптозу, культивируются в присутствии TNF, например, в течение 24 часов, а затем приводятся в контакт с анализируемым соединением. Контрольные клетки из той же линии клеток культивируют в отсутствие TNF в течение того же периода времени, а затем приводятся в контакт с тем же соединением. Затем измеряется и сопоставляется жизнеспособность клеток.
Для проведения такого анализа может использоваться, по существу, любая линия клеток, которая известна своей природной устойчивостью к TRAIL-индуцированному апоптозу и экспрессирует представляющий интерес белок комплекса III либо модифицирована таким образом, чтобы экспрессировать этот белок, например, посредством трансфицирования трансгена, экспрессирующего белок. Примером такой клеточной линии является линия клеток опухоли толстой кишки HCT116, которые были модифицированы для стабильной экспрессии Core 1. Трансген Core 1 может вводиться, например, посредством инфицирования клеток HCT116 с использованием применяемых в данной области стандартных лентивирусных, ретровирусных или аденовирусных векторов экспрессии. В настоящем изобретении также предполагается, что клеткам нет необходимости экспрессировать весь белок, если они экспрессируют ту часть белка, которая придает способность подавлять индуцирующую апоптоз активность TNF. Соответственно, клетка может содержать или может быть модифицирована таким образом, чтобы содержать только нуклеиновую кислоту, которая кодирует такую часть белка комплекса III.
Клетки могут приводиться в контакт с анализируемым соединением в диапазоне концентраций от наномолярного до микромолярного в период от 24 до 96 часов при 37°C, и жизнеспособность клеток измеряется с использованием серийно выпускаемых наборов для анализа, например WST-1, MTT и анализа каспазы-3. Эффект воздействия соединений на развитие клеточного цикла может измеряться с помощью анализа FACS или другими стандартными методами анализа изображений, используемыми в данной области. Все описанные эксперименты с соединениями могут также проводиться в условиях гипоксии (1-2% кислорода), чтобы установить, в какой мере гипоксические условия усиливают, или индуцируют, гибель клеток в присутствии TRAIL.
Клетки могут приводиться в контакт с TRAIL посредством прямого добавления TRAIL в растворе соответствующего носителя (например, фосфатный буферный физиологический раствор) в приемник, содержащий клетки в концентрации примерно от 50 до 200 нг/мл, и культивирования клеток в его присутствии в течение примерно 96 часов.
Примеры
Пример 1
В настоящем примере описывается открытие того факта, что Core 1 связан с устойчивостью опосредованного TRAIL апоптоза.
Проводили функциональный генетический скрининг библиотеки мышиной кДНК E14 для линии карциномы толстой кишки g HCT116, линии клеток, известной своей чувствительностью к TRAIL-индуцированному апоптозу [фигура 1A]. В ходе скрининга была идентифицирована UQCRC1, кодирующая Core 1, как определяющая устойчивость к опосредованному TRAIL апоптозу.
Клетки HCT116 (American Type Culture Collection, Manassas, Virginia) и линия клеток с дефектом упаковки 293 EBNA (Invitrogen) культивировали раздельно в модифицированной по Дульбекко среде Игла (DMEM) в присутствии 10% термоинактивированной фетальной бычьей сыворотки (FBS) и 1% пенициллин-стрептомицина (Gibco) при 37°C и 5% CO2.
Библиотеку ретровирусной кДНК 14-дневных мышиных эмбрионов (George Daley, MIT, Cambridge, Massachusetts) клонировали в ретровирусный вектор pEYK и трансфицировали в указанную выше линию клеток с дефектом упаковки 293 EBNA с использованием липофектамина (Invitrogen). Супернатант собирали в период времени от 48 до 72 часов после трансфицирования. Супернатант смешивали с полибеном концентрации 10 мг/мл и использовали для инфицирования указанных выше линий клеток HCT116, которые высеивали на планшеты за 48 часов до инфицирования. Через 48 часов после инфицирования 1,0×106 клеток высеивали в 15-см чашки и обрабатывали 200 нг/мл рекомбинантным человеческим TRAIL (BioMol, Plymouth Meeting, Pennsylvania) в течение 10 дней, причем свежая среда и TRAIL добавляли каждые 48 часов. Собирали индивидуальные клоны, устойчивые по отношению к индуцированному TRAIL апоптозу.
Геномную ДНК выделяли из устойчивого клона посредством повторного суспендирования клеток в питательном буфере (100 нМ NaCl, 10 мМ Tris pH 8, 25 мМ EDTA, 0,5% SDS) с последующей обработкой протеиназой K с конечной концентрацией 100 мкг/мл при 50°C в течение 12 ч. Образцы экстрагировали фенолом-хлороформом с осаждением этанолом. Потенциальные кДНК затем амплифицировали с использованием специфичных для библиотеки праймеров или же выделенную ретровирусную плазмиду использовали непосредственно для проведения очередного цикла скрининга. Одной из интересных потенциальных кДНК, которая, по-видимому, придавала устойчивость по отношению к индуцированному TRAIL апоптозу, была кДНК основной субъединицы 1 убихинол-цитохром С редуктазы (UQCRC1), которая кодирует Core 1 [фигура 1B].
Пример 2
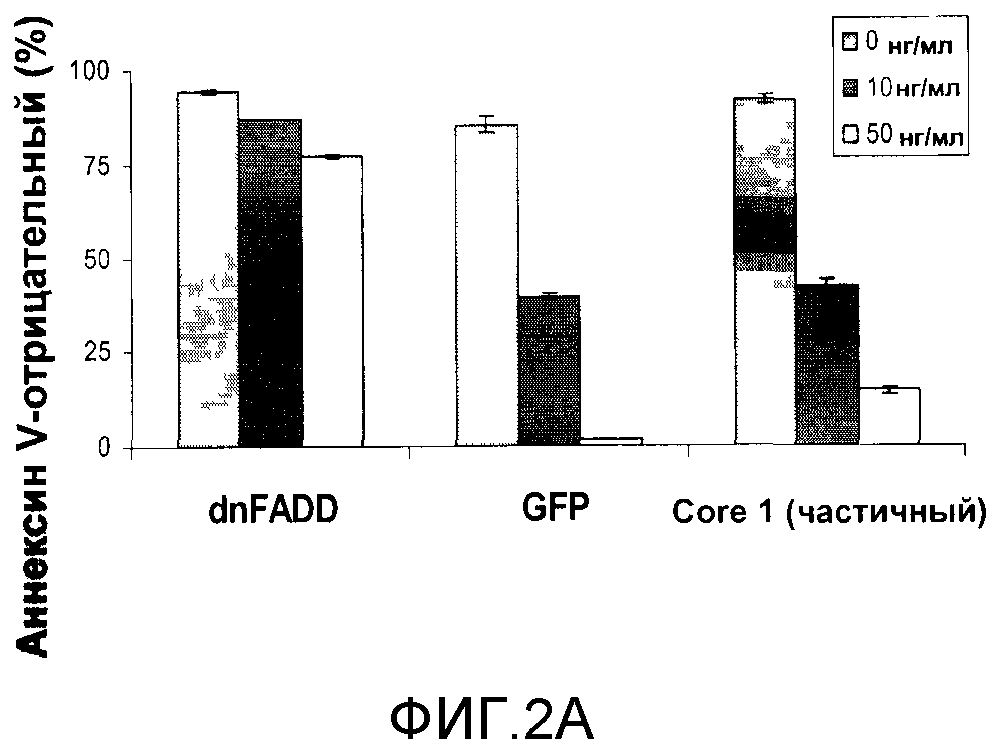
Для дополнительной характеристики способности Core 1 защищать от индуцированного TRAIL апоптоза выделенный при скрининге вектор pEYK, содержащий частичную кДНК UQCRC1, использовали для прямого инфицирования клеток HCT116. В качестве контроля использовали клетки, экспрессирующие доминантно-негативный FADD (dnFADD), который ингибирует сигнал рецепторов TRAIL. В течение 48 часов клетки обрабатывали 0, 10 или 50 нг/мл TRAIL и анализировали содержание аннексина V, раннего маркера апоптоза [фигура 2A]. Экспрессия dnFADD ингибировала индуцированный TRAIL апоптоз, причем 78% клеток были аннексин V-отрицательными или не проявляющими апоптоз при концентрации TRAIL 50 нг/мл. Среди клеток, экспрессирующих зеленый флуоресцентный белок (GFP), только 2% были аннексин V-отрицательными при 50 нг/мл по сравнению с 14,5% в случае клеток, экспрессирующих Core 1, что соответствует 7-кратному увеличению уровня защиты от TRAIL-индуцированного апоптоза. Этот результат показывает, что экспрессия частичного белка Core 1 обеспечивает определенную защиту, но не полностью ингибирует TRAIL-индуцированный апоптоз.
Пример 3
Поскольку Core 1 представляет собой субъединицу митохондриального дыхательного комплекса III, проводили исследования для установления, защищает ли экспрессия Core 1 от митохондриальной деполяризации, критически важной стадии митохондриально-опосредованного апоптоза. Изменения потенциала митохондриальной мембраны в ходе обработки TRAIL исследовали с помощью набора JC-1 Assay Kit (Molecular Probes). JC-1 (5, 5',6,6',-тетрахлор-1,1',3,3'-тетраэтилбензимидазолилкарбоцианин) является катионным красителем, который обладает диффузной зеленой флуоресценцией в цитоплазме. Накопление JC-1 в митохондриях, которое зависит от численности интактных поляризованных митохондрий, приводит к образованию красных флуоресцентных агрегатов. Отношение зеленой и красной флуоресценции измеряли для клеток, описанных выше в примере 2, с помощью анализа методом сортировки активированных флуоресценцией клеток (FACS). FACS представляет собой метод сортировки клеток, использующий специфичные маркеры клеточной поверхности (например красная или зеленая флуоресценция). Указанное выше соотношение откладывали на графике как процент поляризованных митохондрий.
Экспрессия dnFADD полностью защищала от митохондриальной деполяризации при всех концентрациях TRAIL, в то время как клетки, экспрессирующие GFP, демонстрировали всего 7% поляризованных митохондрий, которые сохранялись при 50 нг/мл TRAIL. Экспрессия частичного Core 1 демонстрировала 38% поляризованных митохондрий, обеспечивая 5-кратную защиту от митохондриальной деполяризации по сравнению с GFP [фигура 2B]. Таким образом, Core 1 также защищает от митохондриальной деполяризации.
Пример 4
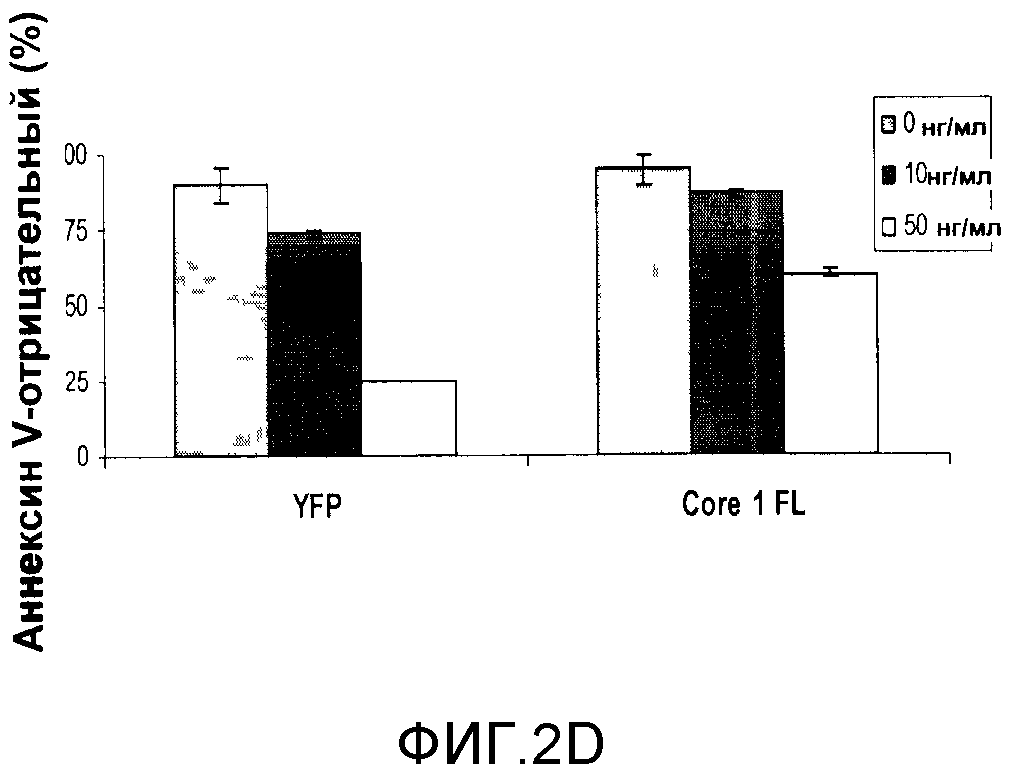
Для подтверждения того, что защита, наблюдаемая в случае частичного белка Core 1, свидетельствует о его функции, полную последовательность человеческого белка Core 1 клонировали в pLenti-вектор (содержащий HA-тэг) и использовали для инфицирования HCT116, с тем чтобы получить стабильную линию клеток, экспрессирующих Core 1. Аналогичным образом готовили контрольную линию клеток для экспрессии желтого флуоресцентного белка (YFP). Чтобы подтвердить экспрессию полного белка Core 1, клетки гомогенизировали с целью приготовления митохондриального и цитоплазматического лизата, а затем проводили вестерн-блоттинг с антителами к Core 1 [фигура 2C]. Эндогенный Core 1 детектировали в митохондриальной фракции клеток, экспрессирующих YFP и Core 1. Эндогенно экспрессированный Core 1 связывается с тэгом HA и обнаруживается как более активно движущаяся полоса в митохондриальной фракции, а также в цитоплазматической фракции. Такой характер экспрессии не является результатом загрязнения цитоплазматической фракции митохондриальными белками, поскольку COX1 (субъединица I комплекса IV) обнаруживается только в митохондриальной фракции, тогда как SOD1 (супероксиддисмутаза 1), цитоплазматический маркер, не обнаруживает цитоплазматических белков в митохондриальной фракции. Поэтому Core 1 экспрессируется в этих стабильных клеточных линиях HCT116. Чтобы установить, защищает ли также полный Core 1 от TRAIL-индуцированного апоптоза, клетки обрабатывали TRAIL и анализировали на предмет апоптоза с помощью анализа FACS для клеток, которые были аннексин V-положительными или отрицательными. Экспрессирующие Core 1 клетки демонстрировали 2,5-кратную защиту от TRAIL-индуцированного апоптоза при концентрации 50 нг/мл [фигура 2D]. Эти данные показывают, что экспрессия Core 1 в HCT116 защищает от TRAIL-индуцированного апоптоза и митохондриальной деполяризации.
Пример 5
В приведенном примере продемонстрировано ингибирующее действие Core 1 на активность каспаз 8, 9 и 3.
Клетки, стабильно экспрессирующие YFP или полный человеческий Core 1, обрабатывали в течение 24 часов 200 нг/мл TRAIL, лизировали, а затем проводили вестерн-блоттинг с антителами к нескольким каспазам. Активацию каспазы детектировали по распаду полной прокаспазы на более мелкие активные формы. Клетки HCT116, экспрессирующие полный человеческий Core 1, демонстрировали меньшую активность каспазы 8 при стимуляции TRAIL, чем клетки, экспрессирующие YFP [фигура 3A]. Кроме того, экспрессия Core 1 аналогичным образом приводила к пониженной активации каспазы 9 и каспазы 3. Таким образом, экспрессия Core 1 ингибирует полную активацию каспаз 8, 9 и 3 в условиях стимуляции TRAIL.
Пример 6
Настоящий пример демонстрирует эффект воздействия Core 1 на высвобождение цитохрома С из митохондрий.
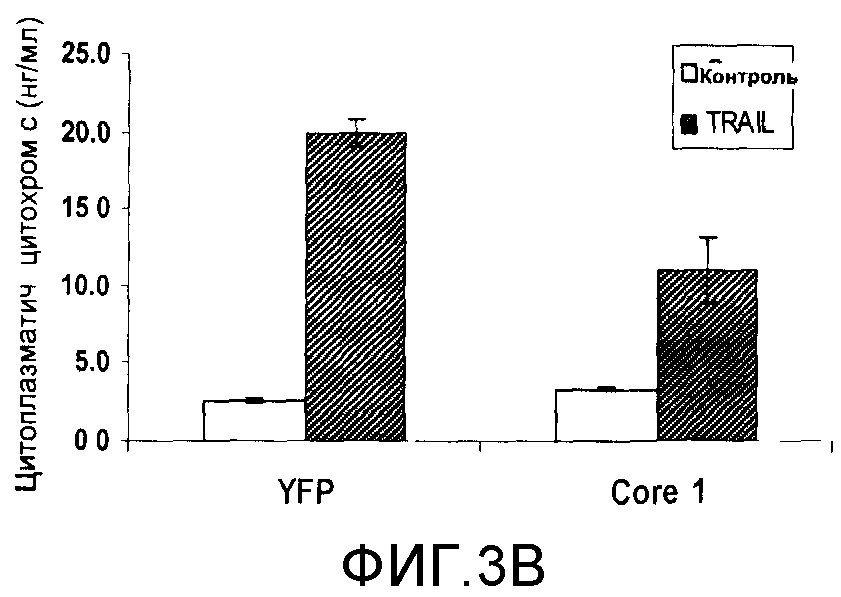
Клетки HCT116, стабильно экспрессирующие YFP или полный человеческий Core 1, в течение 24 часов обрабатывали 100 нг/мл TRAIL. Контрольные клетки, экспрессирующие YFP или полный человеческий Core 1, не обрабатывали TRAIL. Затем собирали культуры клеток. Среды, содержащие плавающие клетки, вначале переносили в новые пробирки. Оставшиеся прилипшие клетки отделяли скребком для клеток в ледяном PBS и добавляли к средам, содержащим плавающие клетки. Клетки таблетировали центрифугированием при 1000 g в течение 10 минут при 4°C и повторно суспендировали в изоосмотическом буфере (0,3 M сахарозы, 10 мМ Tris pH 7,5, 1 мМ EDTA, таблетка свободной от EDTA протеазы). После пяти минут инкубации на льду клетки лизировали с помощью гомогенизаторов Даунса до лизирования примерно 90% плазматических мембран. Эту процедуру проводили с целью выделения цитоплазматической фракции. Степень лизиса клеток контролировали смешиванием 10 мкл лизата с трипановым синим и последующим контролем на гемоцитометре. Супернатант переносили в чистые пробирки и центрифугировали при 8000 g в течение 10 минут при 4°C для таблетирования митохондрий. Концентрацию лизатов определяли с использованием набора для анализа белка BioRad DC (BioRad) или по методу BCA (Pierce, Rockford, Illinois). Для количественного определения уровня цитохрома С лизаты анализировали на содержание цитохрома С с использованием набора для иммуноферментного анализа Function ELISA (Active Motif, Carlsbad, California) [фигура 3B]. В цитоплазме необработанных клеток обнаруживались низкие базовые уровни цитохрома С. После обработки TRAIL клетки, экспрессирующие YFP, демонстрировали 10-кратное увеличение количества цитохрома С, выделяемого в цитоплазматическую фракцию, тогда как клетки, экспрессирующие Core 1, выделяли лишь половину этого количества цитохрома С [фигура 3B]. Этот эксперимент показывает, что экспрессия Core 1 также уменьшает количество цитохрома С, выделяемого из митохондрий в цитозоль при обработке TRAIL.
Пример 7
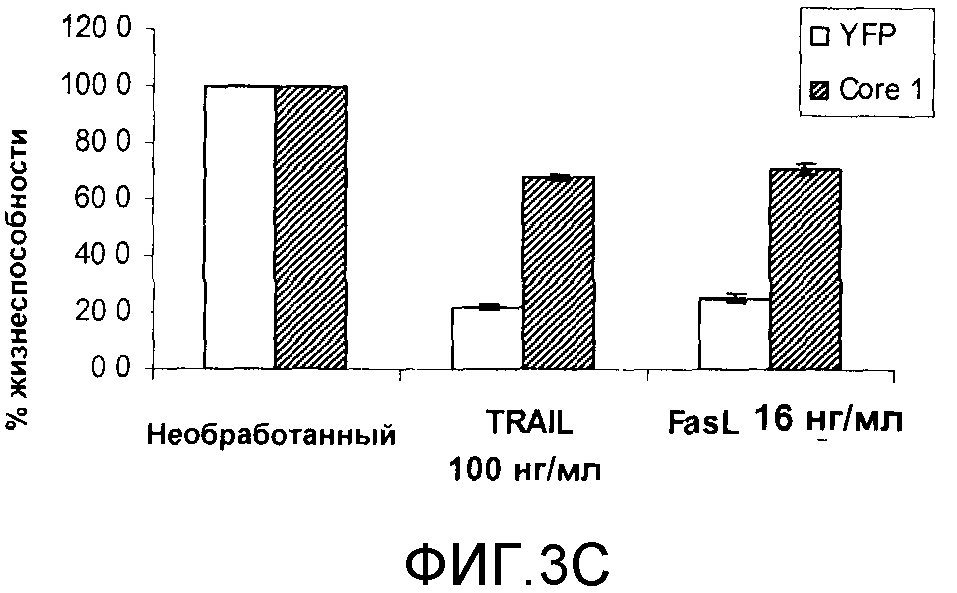
Чтобы выяснить, защищает ли Core 1 от апоптоза, индуцированного TRAIL или по другим каналам, клетки обрабатывали несколькими различными индукторами апоптоза и анализировали на предмет жизнеспособности. Лиганд Fas (FasL), также относящийся к семейству TNFα, который передает сигнал через рецепторы смерти, использует один и тот же белок-адаптер FADD для индукции активации каспаз и апоптоза. Клетки, стабильно экспрессирующие либо желтый флуоресцентный белок (YFP), либо полный человеческий Core 1, обрабатывали 100 нг/мл TRAIL или 16 нг/мл FasL в течение 24 часов. Жизнеспособность клеток анализировали с помощью набора Cell-Titer Blue, в котором превращение индикаторного красителя ресазурина применяли для измерения метаболического потенциала клеток, что является показателем жизнеспособности клеток. Экспрессия Core 1, подобно TRAIL, обеспечивала трехкратную защиту от FasL-индуцированного апоптоза [фигура 3C]. Это означает, что Core 1 может обеспечивать защиту от апоптоза, вызванного сигналами рецепторов гибели клетки.
Затем клетки обрабатывали двумя соединениями, которые индуцируют апоптоз через каналы, не связанные с рецепторами гибели. Камптотецин ингибирует ДНК-топоизомеразу I, что в конечном счете приводит к повреждению ДНК и гибели, тогда как таксотер является ингибитором деполимеризации микротрубочек, что препятствует клеточному делению и вызывает митотическую смерть. При обработке 200 нМ раствором камптотецина в диметилсульфоксиде или 10 нМ раствором таксотера в диметилсульфоксиде не отмечалось никаких различий в жизнеспособности клеток, экспрессирующих YFP или полный человеческий Core 1 [фигура 3D]. Эти результаты показывают, что Core 1 не защищает от других форм апоптоза, не опосредованных рецепторами смерти, вызванных повреждением ДНК или ингибированием деполимеризации микротрубочек. Полученные данные убедительно показывают, что Core 1 играет определенную роль в регулировании TRAIL-индуцированного апоптоза, и согласуются с выводом о том, что Core 1 обеспечивает частичную защиту от TRAIL-индуцированного апоптоза.
Пример 8
В настоящем примере описано использование миРНК для ингибирования экспрессии Core 1.
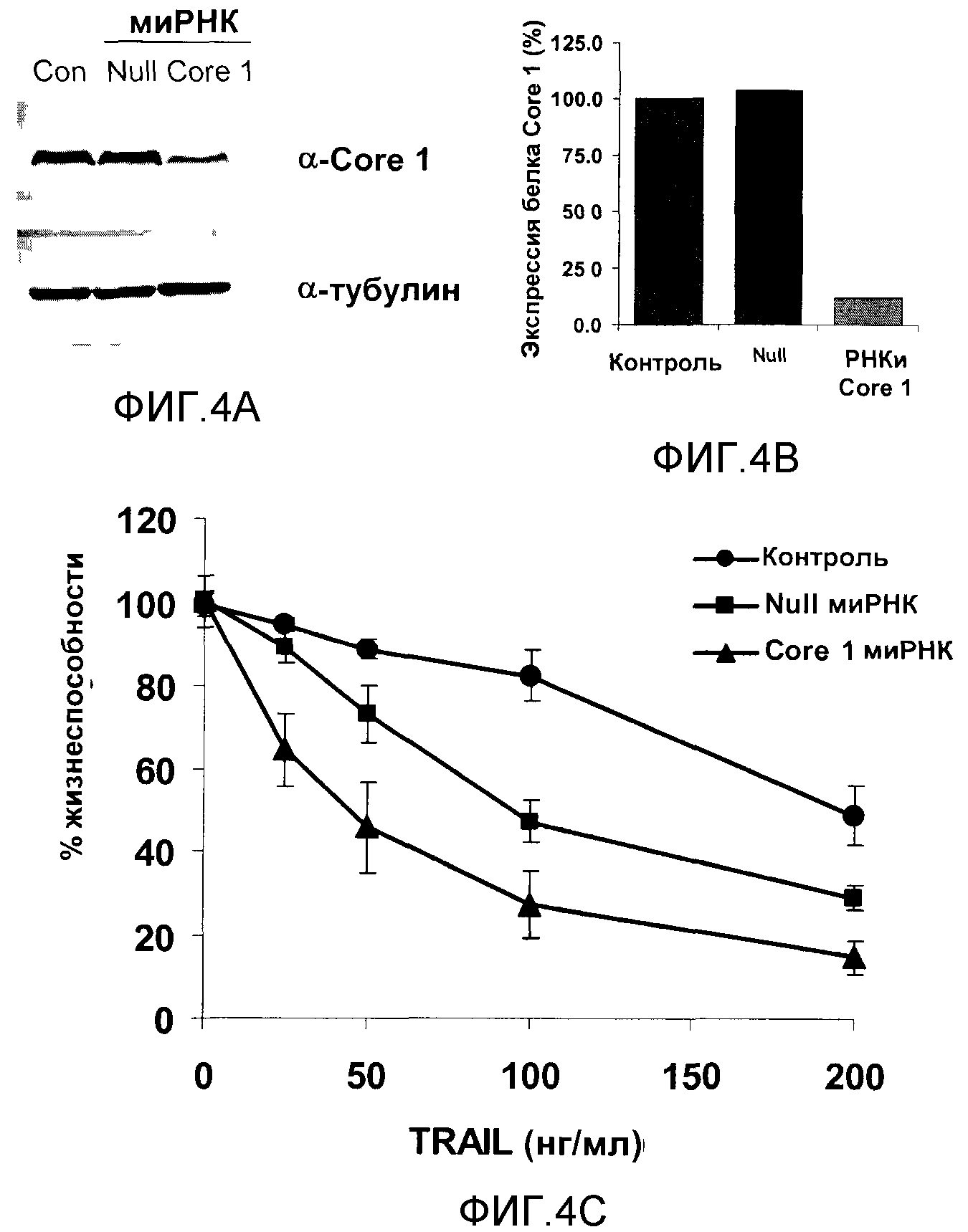
Перед трансфицированием клетки HCT116, содержащие эндогенный Core 1 (и не содержащие экзогенного Core 1), высеивали по 1×105 клеток на 6-ячеечную планшету на 24 часа в среде, не содержащей пенициллина/стрептомицина. В соответствии с протоколами изготовителя клетки трансфицировали липофектамином 2000 (Invitrogen). Клетки инкубировали в течение 72 часов с трансфицированной смесью, содержащей только липофектамин (контрольные клетки), 3,75 нМ миРНК UQCRC1 (SMARTpool) или неориентированную миРНК (Null) (Dharmacon, Lafayette, CO). Концентрацию белка определяли посредством приготовления лизатов и вестерн-блоттингом с использованием антител к Core 1 [фигура 4A]. Пятна повторно снимали и анализировали с применением антител к антитубулину в качестве контроля загрузки. Трансфицирование Core 1 приводило к 90% снижению уровня белка Core 1, тогда как трансфицирование только липофектамином или Null миРНК не влияло на уровни белка Core 1 [фигура 4B].
Каждую из перечисленных выше групп клеток затем обрабатывали различными концентрациями TRAIL в течение дополнительных 24 часов, и жизнеспособность клеток определяли с помощью набора для анализа Cell Titer Blue (Promega, Madison, Wisconsin). В ходе этого анализа определяется метаболический потенциал клетки (а значит, ее жизнеспособность) по ее способности восстанавливать краситель ресазурин до высокофлуоресцентного резоруфина. Количество добавляемого реагента Cell Titer Blue (буферный раствор, содержащий высокоочищенный ресазурин) составляло 20% от объема среды. Клетки инкубировали при 37°C в течение 2-3 часов и анализировали с использованием флуоресцентного ридера со дна планшет при длине волны возбуждения 560 нм и длине волны испускания 590 нм. Посев клеток проводили в трипликатах. Клетки затем дважды промывали в холодном PBS (Gibco) и лизировали в 100 мкл ледяного буфера лизирования (50 нМ HEPES pH 7,4, 250 нМ NaCl, 2 нМ EDTA, 1% Triton X-100, свободные от EDTA ингибиторы протеазы (Roche)). Лизаты центрифугировали в течение 10 минут при 3500 об/мин и температуре 4°C. Супернатант смешивали с 4-кратным объемом аналитического буфера LDS (Invitrogen) и нагревали при 90°C в течение 10 минут, а затем хранили при -20°C. Результаты анализа приводятся на фигуре 4C. Приведенные результаты представляют собой усреднение 4 отдельных экспериментов. Клетки с нокдауном Core 1 проявляли лишь 60% жизнеспособности при 25 нг/мл TRAIL по сравнению с 90-95% при той же концентрации в контрольных клетках или клетках, трансфицированных Null [фигура 4C]. Клетки с нокдауном Core 1 проявляли менее 50% жизнеспособности при 50 нг/мл, в то время как для индукции 50% жизнеспособности в клетках, трансфицированных Null, необходимо в два раза большее количество TRAIL. Несмотря на то что трансфицирование только Null миРНК, по-видимому, оказывает воздействие на чувствительность к TRAIL по сравнению с одним только липофектамином, эти данные однозначно показывают, что снижение уровня белка Core 1 сенсибилизирует клетки по отношению к TRAIL-индуцированному апоптозу, особенно при более низких концентрациях. Полученные результаты убедительно показывают, что Core 1 играет определенную роль в регулировании TRAIL-индуцированного апоптоза.
Пример 9
В приведенном примере описывается анализ, призванный установить, ингибирует ли то или иное соединение активность Core 1.
Анализируемое соединение добавляли в концентрации 10 мкМ в подходящем носителе (например, диметилсульфоксид, этанол, буферный физиологический раствор) к клеткам HCT116 (American Type Culture Collection, Manassas, VA), которые были преинфицированы ДНК Core 1. Затем клетки культивировали в модифицированной Дульбекко среде Игла (DMEM) с 10% FBS в присутствии необходимых концентраций до 200 нг/мл TRAIL и выдерживали в течение 24 часов. Соединение добавляли также в концентрации 10 мкМ к клеткам HCT116, которые были предварительно инфицированы ДНК Core 1. Затем клетки культивировали в DMEM с 10% FBS в отсутствие TRAIL (контрольные клетки) и выдерживали в течение 24 часов.
Затем клетки промывали в PBS и их жизнеспособность определяли с помощью анализа WST-1. В анализе WST-1 жизнеспособность клеток определяется посредством измерения восстановления химического субстрата WST в растворимую соль формазана, которая детектируется с помощью спектрофотометра. Анализ титра Cell Titer Blue может использоваться в качестве альтернативы методу WST-1.
Если соединение значительно снижает жизнеспособность клеток, которые культивировали в присутствии TRAIL, и контрольных клеток, которые культивировали в отсутствие TRAIL, считается, что активность соединения не связана с TRAIL, и соединение не относят к представляющим интерес соединениям. Если соединение существенно снижает жизнеспособность клеток, которые культивировали в присутствии TRAIL, но не жизнеспособность контрольных клеток, тогда считается, что оно сенсибилизирует клетки по отношению к апоптотической активности TRAIL, поскольку было известно, что клеточная линия HCT116, экспрессирующая Core 1, ранее проявляла устойчивость по отношению к TRAIL. Поскольку ранее было известно, что устойчивость клеток по отношению к TRAIL-индуцированному апоптозу вызвана Core 1, делается вывод о том, что анализируемое соединение действует, ингибируя TRAIL-подавляющую активность Core 1.




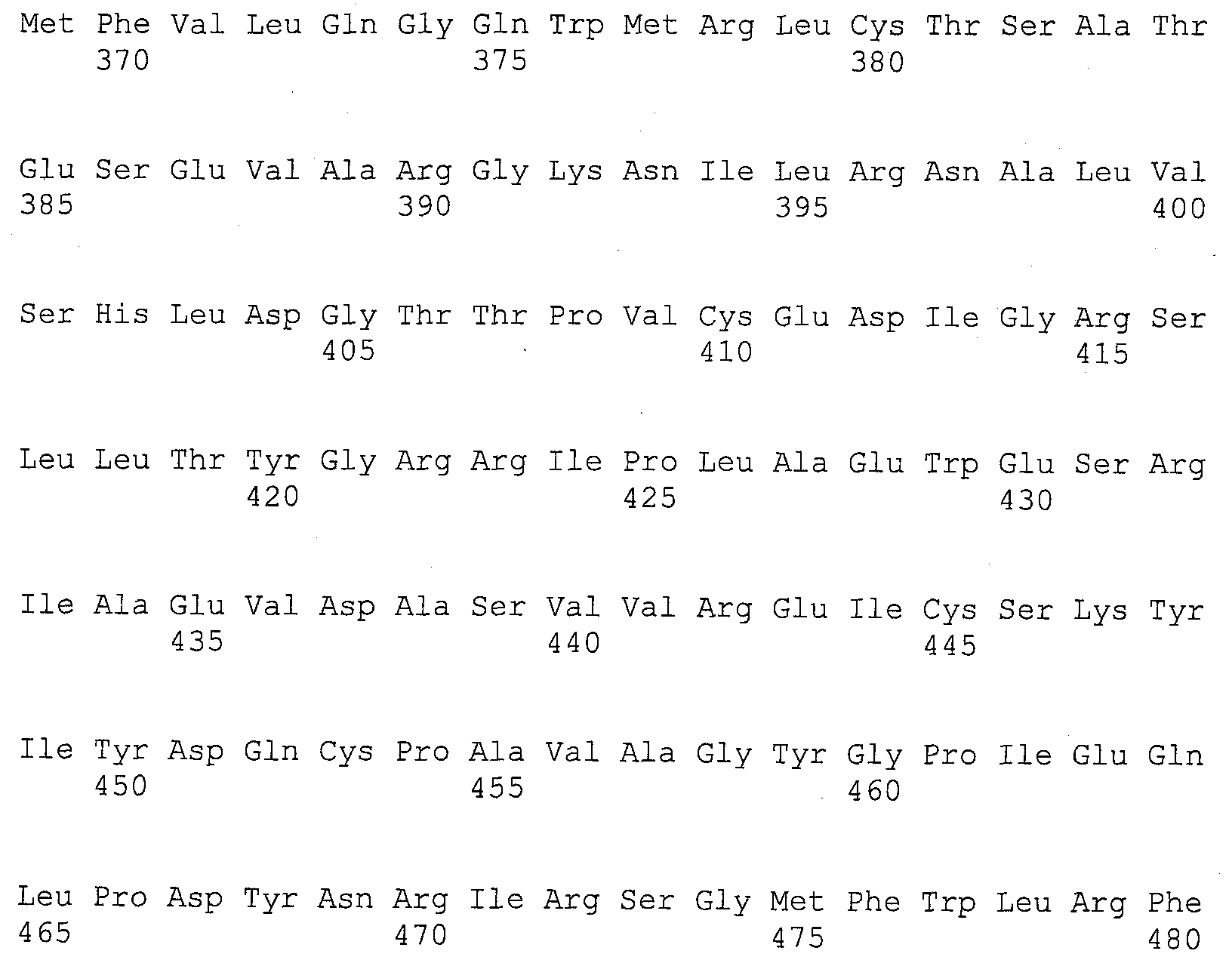
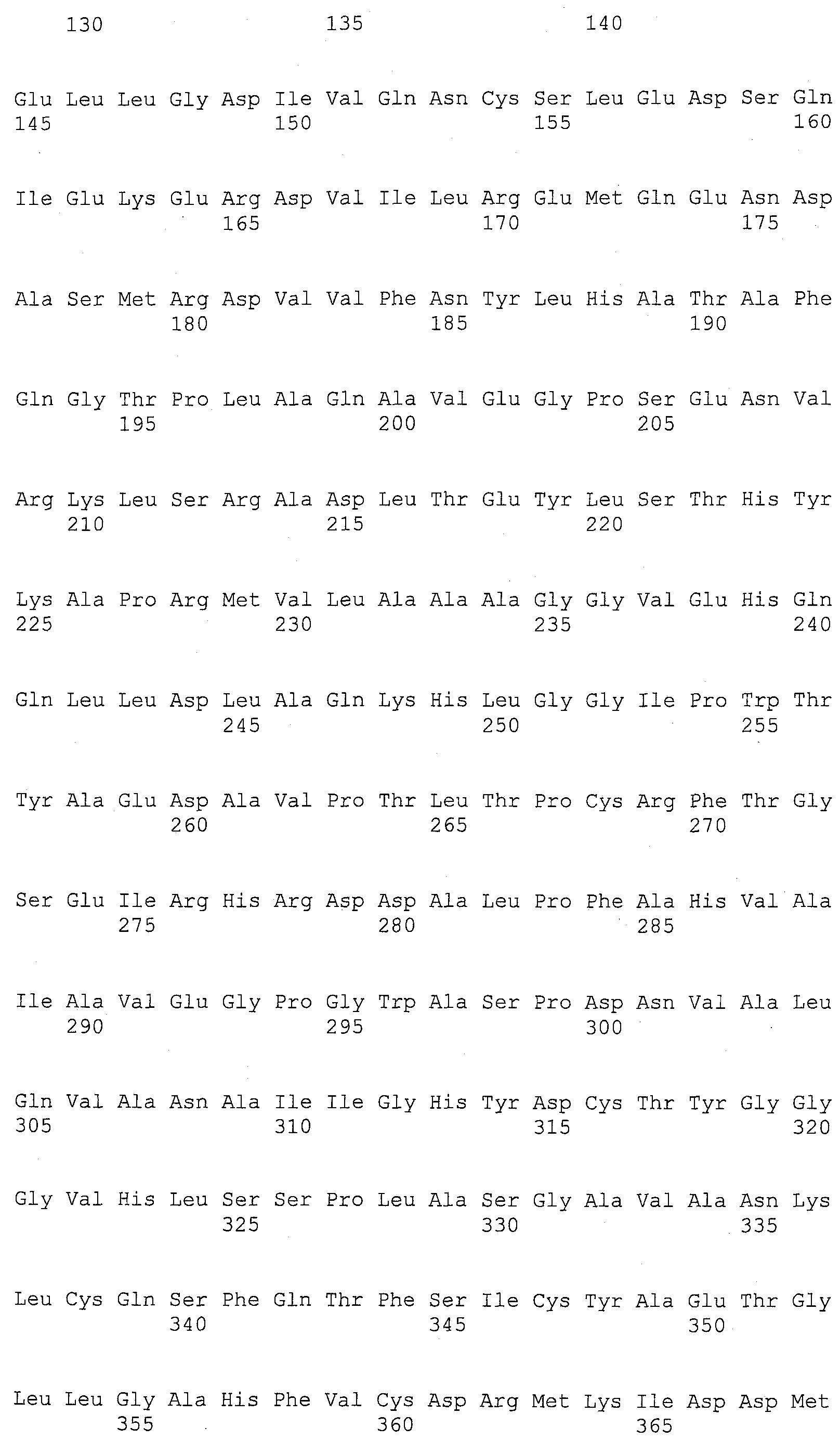


Идентификация молекул, модулирующих белок-белковое взаимодействие
Замещенные бензоиламиноиндан-2-карбоновые кислоты и родственные соединения
Замещенные n-фенилбипирролидинкарбоксамиды и их применение в лечебных целях
Спироциклические нитрилы в качестве ингибиторов протеазы
Циклические индол-3-карбоксамиды, их получение и их применение в качестве лекарственных препаратов
Способ идентификации агента на основе высокопроизводительного скрининга
Циклические азаиндол-3-карбоксамиды, их получение и применение в качестве лекарственных препаратов
Антитела, связывающие il-4 и/или il-13, и их применение
Триазоло[1,5-a]хинолины в качестве лигандов рецептора аденозина а3
Пептиды с большим числом мостиковых связей, выделяемые из actinomadura namibiensis
Идентификация молекул, модулирующих белок-белковое взаимодействие
Замещенные бензоиламиноиндан-2-карбоновые кислоты и родственные соединения
Замещенные n-фенилбипирролидинкарбоксамиды и их терапевтическое применение
Замещенные n-фенилбипирролидинкарбоксамиды и их применение в лечебных целях
Замещенные n-фенилпирролидинилметилпирролидинамиды и их терапевтическое применение в качестве модуляторов рецептора н3 гистамина
Замещенные n-фенил-бипирролидинмочевины и их терапевтическое применение
Спироциклические нитрилы в качестве ингибиторов протеазы
Применение расбуриказы для лечения или профилактики расстройств или косвенных осложнений на сердце, вызванных приступами ишемии или реперфузией
Циклические индол-3-карбоксамиды, их получение и их применение в качестве лекарственных препаратов
Способ идентификации агента на основе высокопроизводительного скрининга

