Результат интеллектуальной деятельности: НОВЫЕ СЕЛЕКТИВНЫЕ ВЕКТОРЫ И СПОСОБЫ СЕЛЕКЦИИ ЭУКАРИОТИЧЕСКИХ КЛЕТОК-ХОЗЯЕВ
Вид РИД
Изобретение
Область техники изобретения
Настоящее изобретение относится к новой системе селекции, основанной на использовании мутантного фолатного рецептора в качестве селективного маркера, для селекции клеток-хозяев, в частности клеток-хозяев млекопитающих, экспрессирующих интересующий полипептид. Изобретение относится к соответствующим экспрессионным векторам, клеткам-хозяевам и способам селекции клеток-хозяев, экспрессирующих рекомбинантный интересующий полипептид с высоким выходом. Кроме того, настоящее изобретение относится к способу эффективного получения рекомбинантных полипептидов с высоким выходом.
Уровень техники изобретения
Возможность клонировать и экспрессировать интересующие продукты, такие как рекомбинантные пептиды и белки, в больших количествах становится все более важной. Возможность очищать большие количества белков, используемых человеком, важна в фармацевтической и биотехнологической области, например, для производства белковых фармацевтических препаратов, а также в научно-исследовательской работе, например, для кристаллизации белков, позволяющей определять их трехмерную структуру. Белки, которые иным образом трудно получать в больших количествах, могут быть избыточно экспрессированы в клетке-хозяине, а затем выделены и очищены.
Выбор системы экспрессии для продуцирования рекомбинантных белков зависит от многих факторов, в том числе ростовых характеристик клеток, уровней экспрессии, внутриклеточной и внеклеточной экспрессии, посттрансляционных модификаций и биологической активности интересующего белка, а также нормативных вопросов и экономических соображений при производстве терапевтических белков. Основными преимуществами клеток млекопитающих по сравнению с другими системами экспрессии, такими как бактерии или дрожжи, являются способность осуществлять правильное сворачивание белков, комплексное N-связанное гликозилирование и аутентичное O-связанное гликозилирование, а также широкий спектр других посттрансляционных модификаций. Вследствие описанных преимуществ эукариотические клетки, и в частности, клетки млекопитающих, являются предпочтительной системой экспрессии для производства сложных терапевтических белков, таких как моноклональные антитела.
Наиболее распространенным подходом для получения экспрессирующих на высоком уровне клеток-хозяев (так называемых суперпродуцентов) является, в качестве первого этапа, создание подходящего экспрессионного вектора для экспрессии интересующего полипептида. Экспрессионный вектор приводит в действие экспрессию полинуклеотида, кодирующего интересующий полипептид, в клетке-хозяине и предоставляет по меньшей мере один селективный маркер для получения рекомбинантной линии клеток.
Одним из общепризнанных методов получения продуцирующих на высоком уровне линий клеток, экспрессирующих интересующий полипептид с высоким выходом, является стабильная трансфекция клеток-хозяев. Затем интересующий полипептид секретируется в культуральную среду и может быть получен из нее в больших количествах. Однако стабильная интеграция в геном является редким событием и только небольшое подмножество стабильно трансфицированных клеток являются суперпродуцентами.
Селективные маркеры и системы селекции широко используют для получения клеток-хозяев, экспрессирующих интересующий полипептид с высоким выходом. Соответствующие системы также полезны для создания и идентификации стабильно трансфицированных клонов. Основной целью использования соответствующих селективных маркеров и систем селекции является введение селективного гена, который под воздействием селективных условий роста позволяет идентифицировать клетки, способные на высоком уровне продуцировать интересующие рекомбинантные продукты. Общепризнанные селективные маркеры включают, например, дигидрофолатредуктазу (DHFR) или глутамин-синтетазу (GS).
Другая система селекции основана на использовании селекции с помощью переносчика восстановленных фолатов. Переносчик восстановленных фолатов (RFC) представляет собой повсеместно экспрессируемый мембранный гликопротеин, который служит основным переносчиком при поглощении восстановленных фолатов, таких как 5-метил-THF и 5-формил-THF. Однако RFC обладает очень слабой аффинностью для окисленного фолата, фолиевой кислоты. Таким образом, клетки, в которых отсутствует экспрессия RFC или у которых был делетирован геномный локус RFC, могут служить в качестве реципиентов для трансфекции гена селективного маркера RFC в условиях, в которых восстановленные фолаты, такие как 5-формил-THF, постепенно выводят из ростовой среды, тем самым заставляя клетки экспрессировать повышенные количества этого переносчика фолатов. У системы селекции с помощью RFC существует ряд недостатков: a) необходимо использовать RFC-нулевые клетки-реципиенты, в которых эндогенный локус RFC был подвергнут нокауту или инактивирован за счет направленного нокаута или мутаций потери функции, b) RFC обладает чрезвычайно низкой способностью к переносу фолиевой кислоты и, следовательно, этот окисленный фолат нельзя использовать для селекции, c) в отличие от системы, основанной на фолатном рецепторе (смотри ниже), которая представляет собой однонаправленную систему поглощения фолатов, RFC является двунаправленным переносчиком фолатов, в равной степени эффективный для введения и выведения фолатов. Это означает, что в условиях депривации фолатов, избыточная экспрессия RFC может быть губительной для реципиентных клеток, из которых дополнительно выводятся фолаты через избыточно экспрессированный RFC.
Другая система селекции, предложенная недавно, основана на использовании фолатного рецептора, такого как фолатный рецептор альфа, в качестве селективного маркера. Эта система описана в WO 2009/080759. Данная система имеет ряд преимуществ в том, что для селекции нет необходимости в токсичных веществах и, более того, нет необходимости в нокауте эндогенного фолатного рецептора клетки-хозяина. Другая система селекции, основанная на использовании фолатного рецептора в качестве селективного маркера, описана в WO 2010/097240.
Фолатные рецепторы и их мутанты описаны, например, в публикации Shen et al. «Identification of amino acid residues that determine the differential ligand specificities of folate receptors alpha and beta» (Biochemistry 1997, 36, 6157-6163). Мутации в фолатном рецепторе альфа, ассоциированные с медицинскими нарушениями, также описаны. Анализ положений аминокислот в фолатных рецепторах приведен в публикации Ramamoorthy et al. «In silico analysis of functionally important residues in folate receptors» (Bioinformation 2 (4): 157-162 (2007)).
Строгость системы селекции имеет решающее значение для селекции и, следовательно, обогащения продуцирующих на высоком уровне клеток из трансфицированной популяции. Чем строже система селекции, тем меньше число продуцирующих на низком уровне клеток после процесса селекции и тем выше шанс обнаружить очень редкие клоны-суперпродуценты. Кроме того, существует большая потребность в создании системы селекции, позволяющей получать продуцирующие на высоком уровне клоны быстрее, чем способами предшествующего уровня техники.
Целью настоящего изобретения является предложение строгой системы селекции для отбора клеток-хозяев, продуцирующих интересующий полипептид с высоким выходом, а также соответствующих экспрессионных векторов и клеток-хозяев. В частности, целью настоящего изобретения является предложение новой системы селекции, имеющей определенные преимущества по сравнению с вышеупомянутыми системами селекции.
Сущность изобретения
Настоящее изобретение относится к системе селекции, подходящей для отбора клеток-хозяев, экспрессирующих интересующий полипептид с высоким выходом. Указанная система селекции основана на использовании мутантного функционального мембраносвязанного фолатного рецептора в качестве селективного маркера. В частности, указанная система селекции позволяет проводить более строгую и быструю селекцию суперпродуцентов, чем система селекции, в которой используют соответствующий функциональный мембраносвязанный фолатный рецептор дикого типа в качестве селективного маркера.
Первый аспект настоящего изобретения относится к экспрессионному вектору или сочетанию по меньшей мере двух экспрессионных векторов, содержащим:
a) полинуклеотид, кодирующий мутантный фолатный рецептор в качестве селективного маркера, при этом мутантный фолатный рецептор имеет сниженную аффинность связывания фолата по сравнению с фолатным рецептором дикого типа, и
b) по меньшей мере один полинуклеотид, кодирующий интересующий полипептид,
при этом когда указанный экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов вводят в клетку-хозяина, интересующий полипептид секретируется из указанной клетки-хозяина.
Второй аспект настоящего изобретения относится к клетке-хозяину, жизнеспособность которой зависит от поглощения фолатов, содержащей
a) введенный полинуклеотид, кодирующий мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата по сравнению с фолатным рецептором дикого типа, в качестве селективного маркера,
и
b) по меньшей мере один введенный полинуклеотид, кодирующий интересующий полипептид,
при этом указанный интересующий полипептид секретируется из указанной клетки-хозяина.
Третий аспект настоящего изобретения относится к способу получения клетки-хозяина в соответствии со вторым аспектом настоящего изобретения, включающему этап введения в клетку-хозяина, жизнеспособность которой зависит от поглощения фолатов, по меньшей мере
a) полинуклеотида, кодирующего мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата по сравнению с фолатным рецептором дикого типа, в качестве селективного маркера,
и
b) по меньшей мере одного полинуклеотида, кодирующего интересующий полипептид, при этом интересующий полипептид секретируется из указанной клетки-хозяина.
Четвертый аспект настоящего изобретения относится к способу селекции по меньшей мере одной клетки-хозяина, способной экспрессировать интересующий полипептид, включающему
a) получение множества клеток-хозяев в соответствии со вторым аспектом;
b) культивирование указанного множества клеток-хозяев в селективной культуральной среде, содержащей фолат в ограниченной концентрации;
и
c) получение по меньшей мере одной клетки-хозяина, экспрессирующей интересующий полипептид.
Пятый аспект настоящего изобретения относится к способу получения интересующего полипептида, включающему
a) культивирование клетки-хозяина в соответствии со вторым аспектом и/или a клетки-хозяина, отобранной в соответствии с четвертым аспектом, в условиях, допускающих экспрессию и секрецию интересующего полипептида;
b) выделение интересующего полипептида из среды для культивирования клеток и
c) необязательно, дальнейшую обработку выделенного интересующего полипептида.
Шестой аспект настоящего изобретения относится к применению полинуклеотида, кодирующего
a) мутантный фолатный рецептор, содержащий следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9),
где Xaa не является аланином, и при этом аффинность связывания фолата у мутантного фолатного рецептора снижена по сравнению с соответствующим фолатным рецептором дикого типа, где Xaa является аланином (SEQ ID NO 1),
или
b) мутантный фолатный рецептор, содержащий аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9, при этом Xaa не является аланином в указанном мутантном фолатном рецепторе и при этом аффинность связывания фолата у указанного мутантного фолатного рецептора снижена по сравнению с последовательностью зрелого человеческого фолатного рецептора альфа дикого типа, где Xaa является аланином (смотри SEQ ID NO 1),
в качестве селективного маркера для селекции клеток, жизнеспособность которых зависит от поглощения фолатов.
Седьмой аспект настоящего изобретения относится к применению полинуклеотида, кодирующего
a) мутантный фолатный рецептор, содержащий следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9)
где Xaa является лейцином;
или
b) мутантный фолатный рецептор, содержащий аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9, и при этом Xaa является лейцином в указанном мутантном фолатном рецепторе по пункту b),
в качестве селективного маркера для отбора клеток, жизнеспособность которых зависит от поглощения фолатов. Как показано в примерах, соответствующий мутантный фолатный рецептор является очень эффективным и строгим селективным маркером, который также позволяет отбирать клетки-хозяева, экспрессирующие соответствующий селективный маркер, более быстро, чем фолатный рецептор дикого типа.
Другие цели, признаки, преимущества и аспекты настоящей заявки станут очевидными для специалистов в данной области из следующего далее описания и прилагаемой формулы изобретения. Следует понимать, однако, что следующее далее описание, прилагаемая формула изобретения и конкретные примеры, хотя и указывают на предпочтительные варианты осуществления изобретения, приведены исключительно для иллюстрации.
Краткое описание чертежей
Фигуры 1-5 демонстрируют продуктивность при производстве антител у отдельных клеточных клонов, полученных путем серийных разведений из поликлональных пулов клеток, которые ранее были трансфицированы различными экспрессионными векторами и получены с использованием различных условий селекции. Таким образом, показана продуктивность клонов, полученных после селекции. Для одноклеточного клонирования клетки культивировали либо в полной среде (таким образом, не поддерживая давление отбора после селекции), либо в селективной среде (таким образом, поддерживая давление отбора после селекции).
Фигура 1: Одноклеточное клонирование трансфектантов с V-DHFRреф после селекции (125 нМ MTX).
Фигура 2: Одноклеточное клонирование трансфектантов с V-DHFRреф после селекции (250 нМ MTX).
Фигура 3: Одноклеточное клонирование трансфектантов с V-дтFRальфа после селекции (15 нМ FA).
Фигура 4: Одноклеточное клонирование трансфектантов с V-мутFRальфа (5 нМ). Как можно видеть, экспрессирующие на более высоком уровне клеточные клоны были получены при использовании мутантного фолатного рецептора в качестве селективного маркера, по сравнению с использованием фолатного рецептора дикого типа в качестве селективного маркера. Более того, степень экспрессии была выше, чем та, которую наблюдали при селекции с использованием DHFR в качестве селективного маркера.
Фигура 5: Одноклеточное клонирование совместно трансфицированной V-мутFRальфа/V-DHFRреф популяции (50 нМ фолиевая кислота (FA)/50 нМ MTX). Как можно видеть, значительно больше клеточных клонов, экпрессирующих на более высоком уровне, было получено при использовании такой стратегии совместной селекции.
Подробное описание изобретения
Неожиданно было обнаружено, что система селекции, основанная на использовании фолатного рецептора в качестве селективного маркера, может быть значительно усовершенствована за счет использования мутантной формы фолатного рецептора. Мутантный селективный маркер можно использовать в качестве доминантного селективного маркера для селекции эукариотических клеток, таких как клетки млекопитающих. Указанный мутант имеет модулированную аффинность связывания фолата по сравнению с соответствующим фолатным рецептором дикого типа. Было обнаружено, что мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата по сравнению с соответствующим фолатным рецептором дикого типа, обладает важными преимуществами в качестве селективного маркера.
Новую систему можно использовать для ускоренной селекции, скрининга и создания клонов эукариотических клеток, в частности клеток млекопитающих, стабильно экспрессирующих и секретирующих рекомбинантные полипептиды с высоким выходом. Селекцию можно проводить с использованием культуральной среды, содержащей ограниченную концентрацию фолата, в частности, ограниченную концентрацию фолиевой кислоты. Новая система селекции, помимо преимуществ общего характера, связанных с использованием фолатного рецептора в качестве селективного маркера, имеет ряд важных преимуществ по сравнению с системами селекции, доступными на предшествующем уровне техники, а также в сравнении с использованием фолатного рецептора дикого типа в качестве селективного маркера, как будет объяснено ниже.
1. Улучшенная скорость и характеристики роста. Как показано в примерах, использование мутантного фолатного рецептора в качестве селективного маркера позволяет значительно ускорять селекцию по сравнению со стандартными системами селекции, которые основаны, например, на использовании DHFR в качестве селективного маркера. Кроме того, система селекции по настоящему изобретению требует гораздо меньше времени, чем система селекции, основанная на использовании фолатного рецептора дикого типа в качестве селективного маркера. В частности, в сравнении с использованием фолатного рецептора дикого типа в качестве селективного маркера, клетки, содержащие мутантный фолатный рецептор по настоящему изобретению в качестве селективного маркера, делятся и восстанавливаются быстрее при культивировании их в селективной культуральной среде, содержащей очень низкие концентрации фолиевой кислоты. Эта достигнутая скорость является значительным преимуществом, уменьшающим продолжительность селекционного цикла. Соответствующее преимущество в росте наблюдается даже при использовании очень строгих условий селекции и, соответственно, крайне ограниченных концентраций фолиевой кислоты в селективной культуральной среде, что даже приводит к нарушению роста клеток, трансфицированных фолатным рецептором дикого типа в качестве селективного маркера. Таким образом, можно применять более строгие условия селекции при использовании мутантного фолатного рецептора по изобретению в качестве селективного маркера. Это преимущество мутантного фолатного рецептора по настоящему изобретению перед фолатным рецептором дикого типа было совершенно неожиданным. Фолат, предпочтительно такой, как фолиевая кислота, должен присутствовать в культуральной среде и должен эффективно включаться в клетки-хозяева для поддержания роста клеток, биосинтеза пуриновых и пиримидиновых нуклеотидов, репликации ДНК и, следовательно, пролиферации клеток. С учетом этого ожидалось, что клетки, трансфицированные мутантным фолатным рецептором, имеющим сниженную аффинность связывания фолата, не будут иметь преимущество в росте по сравнению с клетками, которые трансфицированы фолатным рецептором дикого типа. Даже предполагалось, что клетки, трансфицированные таким мутантным фолатным рецептором в качестве селективного маркера, могут не иметь преимуществ в росте по сравнению с нетрансфицированными клетками, которые эндогенно экспрессируют фолатный рецептор дикого типа, имеющий полную аффинность связывания фолата. Это особенно, поскольку известно, что экспрессия эндогенного фолатного рецептора возрастает, если нетрансфицированные клетки культивировать в культуральной среде, содержащей ограниченную концентрацию фолата (смотри Zhu et al., Journal of Cellular Biochemistry 81: 205-219 (2001)). Таким образом, было совершенно неожиданно, когда авторы изобретения обнаружили, что мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата, является эффективным селективным маркером, даже превосходящим фолатный рецептор дикого типа.
2. Повышенная строгость и продуктивность. Клетки, содержащие мутантный фолатный рецептор по настоящему изобретению в качестве селективного маркера, как ни удивительно, переносят более низкие концентрации фолата в селективной культуральной среде, чем клетки, содержащие фолатный рецептор дикого типа в качестве селективного маркера. Это позволяет использовать более строгие условия селекции. Таким образом, клетки с более высокой продуктивностью можно получать быстрее при использовании нового селективного маркера, описанного в настоящем документе. Это было совершенно неожиданно, учитывая тот факт, что аффинность связывания фолата у мутантного фолатного рецептора по настоящему изобретению снижена по сравнению с рецептором дикого типа.
3. Повышенная надежность. При использовании мутантного фолатного рецептора по настоящему изобретению в качестве селективного маркера наблюдали линейную зависимость от концентрации фолата в культуральной среде. Чем ниже концентрация фолата в селективной среде, тем выше достигнутая в результате производительность отобранных клеток. Соответствующую зависимость не наблюдали в аналогичной ситуации при использовании фолатного рецептора дикого типа в качестве селективного маркера. Эта линейная зависимость от концентрации способствует более надежному контролю и оптимизации условий селекции. Этот результат также был совершенно неожиданным.
Таким образом, новый способ селекции при помощи фолата, описанный в настоящем документе, который основан на использовании мутантного фолатного рецептора в качестве селективного маркера, который по сравнению с соответствующим фолатным рецептором дикого типа имеет сниженную аффинность связывания фолата, является отличной стратегией, которая хорошо подходит для ускоренной селекции стабильных клеток, экспрессирующий рекомбинантный интересующий полипептид с высоким выходом. Полезные результаты, описанные в настоящем документе, могут быть достигнуты при низких концентрациях фолата в среде для культивирования клеток и даже без селекции при помощи цитотоксического лекарственного средства, обычно используемой в различных других системам селекции.
Экспрессионный вектор и сочетание экспрессионных векторов
Первый аспект настоящего изобретения относится к экспрессионному вектору или сочетанию по меньшей мере двух экспрессионных векторов, содержащим:
a) полинуклеотид, кодирующий мутантный фолатный рецептор в качестве селективного маркера, при этом мутантный фолатный рецептор имеет сниженную аффинность связывания фолата по сравнению с фолатным рецептором дикого типа, и
b) по меньшей мере один полинуклеотид, кодирующий интересующий полипептид,
при этом когда указанный экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов вводят в клетку-хозяина, интересующий полипептид секретируется из указанной клетки-хозяина.
«Вектор» в соответствии с настоящим изобретением, в частности, означает полинуклеотид, способный нести по меньшей мере один полинуклеотидный фрагмент. Вектор действует как молекулярный носитель, доставляющий полинуклеотиды в клетку-хозяина. Экспрессионный вектор может содержать по меньшей мере одну экспрессионную кассету, содержащую регуляторные последовательности для правильной экспрессии включенного в нее полинуклеотида. Полинуклеотиды (например, кодирующие интересующий полипептид или селективный маркер), которые предстоит вводить в клетку, можно встраивать в экспрессионную кассету(ы) вектора для экспрессии из нее. При введении в клетку-хозяина экспрессионная кассета, в частности, способна управлять аппаратом клетки для транскрипции встроенного полинуклеотида, кодирующего интересующий полипептид, в РНК, которая затем, как правило, дополнительно процессируется и в конечном итоге транслируется в интересующий полипептид. Вектор может находиться в кольцевой или линейной (линеаризованной) форме. Термин «вектор» также включает искусственные хромосомы, вирусные векторы или аналогичные соответствующие полинуклеотиды, позволяющие переносить фрагменты чужеродной нуклеиновой кислоты.
Термин «полинуклеотид» означает полимер из нуклеотидов, которые, как правило, имеют связи между одной дезоксирибозой или рибозой и другой, и относится к ДНК, а также РНК, в зависимости от контекста. Термин «полинуклеотид» не предполагает никаких ограничений по размеру.
Далее авторы изобретения описывают варианты осуществления экспрессионного вектора и сочетания по меньшей мере двух экспрессионных векторов по настоящему изобретению. Полинуклеотид, кодирующий мутантный фолатный рецептор, и полинуклеотид, кодирующий интересующий полипептид, могут быть расположены на одном и том же экспрессионном векторе или на отдельных экспрессионных векторах, если используют сочетание по меньшей мере двух экспрессионных векторов. Если используют сочетание по меньшей мере двух экспрессионных векторов, из которых один экспрессионный вектор содержит полинуклеотид, кодирующий интересующий полипептид, и другой экспрессионный вектор содержит полинуклеотид, кодирующий мутантный фолатный рецептор, указанное сочетание совместно трансфицируют в одни и те же клетки-хозяева, чтобы можно было проводить селекцию. Соответствующие стратегии совместной трансфекции хорошо известны специалистам и также описаны в примерах. Далее авторы изобретения описывают конкретные варианты осуществления и преимущества, главным образом связанные с вариантом осуществления, в котором оба полинуклеотида расположены на одном и том же экспрессионном векторе. Однако указанное описание с соответствующими изменениями применимо и к варианту осуществления, в котором используют сочетание по меньшей мере двух экспрессионных векторов, совместно трансфицируемых в клетки. Когда это целесообразно, авторы изобретения описывают преимущества, связанные с экспрессионным вектором или сочетанием по меньшей мере двух экспрессионных векторов, при использовании указанного экспрессионного вектора(ов) для селекции клеток-хозяев, экспрессирующих интересующий полипептид с высоким выходом.
Мутантный фолатный рецептор
Используемый в настоящем документе термин «фолатный рецептор» относится к рецептору, который является функциональным и, следовательно, способным вводить или привносить фолат или его производное в эукариотическую клетку, в частности, клетку млекопитающего. Предпочтительно, фолатный рецептор способен к однонаправленному введению или привнесению фолата или его производного в эукариотическую клетку-хозяина, в частности, клетку млекопитающего. Кроме того, описанный в настоящем документе фолатный рецептор является мембраносвязанным. Таким образом, фолатные рецепторы, описанные в настоящем документе, являются функциональными мембраносвязанными фолатными рецепторами. Это относится как к мутантному фолатному рецептору, так и к фолатному рецептору дикого типа. Заякоривание в мембране может достигаться за счет, например, трансмембранного якоря или гликозилфосфатидилинозитольного (GPI) якоря. GPI якорь является предпочтительным, поскольку он соответствует естественному состоянию фолатного рецептора. Фолатные рецепторы (FR) представляют собой высокоаффинные фолат-связывающие гликопротеины. Они кодируются тремя разными генами: FR альфа, FR бета и FR гамма. FR альфа также известен как фолат-связывающий белок взрослых, или FDP, как фолатный рецептор 1, или FOLR (у мышей folbp1), и как антиген, ассоциированный с раком яичника. FR бета также известен как FOLR2 (эмбриональный) и как FBP/PL-1 (плацента). FR гамма также известен как FOLR3 и как FR-G (обзор в публикации M.D. Salazar and M. Ratnam, Cancer Metastasis Rev. 2007 26(1), pp.141-152). Зрелые рецепторы FR, которые хорошо охарактеризованы, представляют собой гомологичные белки с ~70-80% аминокислотной идентичности и содержат 229-236 аминокислот, а также два-три сайта N-гликозилирования. FR альфа и FR бета являются мембраносвязанными белками. FR альфа и FR бета представляют собой GPI-заякоренные гликопротеины клеточной поверхности, в то время как FR гамма лишен GPI якоря и является секретируемым белком. Однако он может быть генетически изменен для включения трансмембранного домена или GPI якоря. Такая измененная форма FR гамма, содержащая мембранный якорь, также считается фолатным рецептором дикого типа, если она способна вводить или привносить фолат или его производное в эукариотическую клетку, как описано выше. FR альфа и FR бета проявляют высокую аффинность в отношении фолиевой кислоты (Kd = 0,1-1 нМ), 5,10-дидеазатетрагидрофолиевой кислоты (DDATHF; лометрексол; Ki = 0,4-1,3 нМ при использовании [3H]фолиевой кислоты в качестве субстрата) и BGC945 (который представляет собой ингибитор тимидилатсинтазы на основе циклопента[g]хиназолина, специфически переносимый исключительно через FR альфа и не через переносчик восстановленных фолатов) (Kd = 1 нМ), но намного меньшую аффинность в отношении MTX (Kd >100 нМ). FR-зависимое поглощение фолатов и антифолатов происходит по классическому механизму рецептор-опосредованного эндоцитоза.
«Мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата по сравнению с фолатным рецептором дикого типа» или аналогичные выражения, используемые в настоящем документе, в частности, относятся к мутантному фолатному рецептору, который по сравнению с соответствующим фолатным рецептором дикого типа имеет сниженную аффинность связывания для по меньшей мере одного фолата, выбранного из группы восстановленных фолатов и окисленных фолатов. Указанный термин, в частности, относится к мутантным фолатным рецепторам, которые по сравнению с соответствующим фолатным рецептором дикого типа имеют сниженную аффинность связывания фолата для конкретного фолата. Аффинность связывания фолата для других фолатов, то есть, фолатов, отличных от указанного конкретного фолата, может быть неизменной. В одном варианте осуществления мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата, имеет по меньшей мере одну мутацию, которая, по сравнению с соответствующим фолатным рецептором дикого типа, приводит к снижению аффинности связывания для по меньшей мере одного фолата, выбранного из группы восстановленных фолатов и окисленных фолатов. В одном варианте осуществления мутантный фолатный рецептор имеет, по сравнению с соответствующим фолатным рецептором дикого типа, сниженную аффинность связывания для восстановленного фолата. В одном варианте осуществления мутантный фолатный рецептор имеет, по сравнению с соответствующим фолатным рецептором дикого типа, сниженную аффинность связывания для 6S диастереоизомера 5-метилтетрагидрофолата. В одном варианте осуществления мутантный фолатный рецептор имеет величину IC50 для восстановленного фолата, предпочтительно для 6S диастереоизомера 5-метилтетрагидрофолата, которая по меньшей мере в 20 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз, по меньшей мере в 50 раз или по меньшей мере в 55 раз выше, чем величина IC50 соответствующего фолатного рецептора дикого типа. Вследствие значительно более высокого значения IC50 он имеет существенно сниженную аффинность связывания для указанного восстановленного фолата по сравнению с фолатным рецептором дикого типа. В одном варианте осуществления мутантный фолатный рецептор демонстрирует ослабленное связывание с фолиевой кислотой.
По меньшей мере одна мутация, которая приводит к сниженной аффинности связывания фолата, может представлять собой, например, замену, делецию или вставку аминокислоты. В одном варианте осуществления по меньшей мере одна мутация присутствует в предполагаемом кармане связывания фолата. В одном варианте осуществления указанная мутация представляет собой замену в предполагаемом кармане связывания фолата.
Мутантный фолатный рецептор, используемый по настоящему изобретению в качестве селективного маркера, имеет сниженную аффинность связывания фолата по сравнению с соответствующим фолатным рецептором дикого типа. Как описано выше и как показано в примерах, выгодно использовать мутантный фолатный рецептор, который, по сравнению с соответствующим фолатным рецептором дикого типа, имеет сниженную аффинность связывания для по меньшей мере 6S диастереоизомера 5-метилтетрагидрофолата. Снижение аффинности связывания фолата может быть достигнуто путем введения одной или более мутаций в последовательность дикого типа. Соответствующие примеры описаны ниже. Без привязки к теории, считается, что вследствие сниженной аффинности связывания фолата клетки, трансфицированные экспрессионным вектором(ами) по настоящему изобретению, должны экспрессировать больше мутантного фолатного рецептора для достижения степени поглощения фолатов, достаточной для выживания в селективных условиях депривации фолата. Таким образом, интересующий полипептид также экспрессируется на более высоком уровне популяцией выживших клеток. Как показано в примерах, при использовании мутантного фолатного рецептора, описанного в настоящем документе, в качестве селективного маркера продуктивность возрастает, если концентрация фолата в селективной культуральной среде снижается. Соответствующую корреляцию не наблюдают в аналогичной ситуации при использовании фолатного рецептора дикого типа в качестве селективного маркера. Более того, при использовании мутантного фолатного рецептора в качестве селективного маркера можно даже дополнительно снижать концентрацию фолата в селективной культуральной среде и, таким образом, дополнительно увеличивать давление отбора на трансфицированные клетки. За счет этого создается очень строгая и быстрая система селекции, превосходящая систему селекции, в которой используют фолатный рецептор дикого типа в качестве селективного маркера. Это было неожиданно и очень удивительно, с учетом того факта, что аффинность связывания фолата у мутантного фолатного рецептора по настоящему изобретению снижена по сравнению с фолатным рецептором дикого типа. Кроме того, неожиданно было обнаружено, что клетки, которые были трансфицированы мутантным переносчиком фолатов, демонстрировали превосходные характеристики, и в частности, раньше восстанавливались из селекционных условий, даже в случае использования очень строгих условий селекции.
Предпочтительно, мутантный фолатный рецептор, используемый в качестве селективного маркера, имеет по меньшей мере одну мутацию в кармане связывания фолата, при этом указанная мутация оказывает тот эффект, что аффинность связывания фолата снижается по сравнению с соответствующим фолатным рецептором дикого типа. Надлежащие мутации описаны далее. Введение мутации в карман связывания фолата является очень эффективным подходом для снижения аффинности связывания фолата. Только клетки, экспрессирующие в большом избытке введенный мутантный фолатный рецептор, способны поглощать достаточные количества фолата из культуральной среды для поддержания клеточного роста, репликации ДНК и, следовательно, клеточной пролиферации. Удивительно, но даже если клетки содержали мутантный фолатный рецептор, имеющий сниженную аффинность для фолата, в качестве селективного маркера, трансфицированные клетки демонстрировали значительно ускоренный рост по сравнению с клетками, трансфицированными фолатным рецептором дикого типа, или по сравнению с клетками, трансфицированными общепринятым селективным маркером, таким как DHFR. Этот ускоренный рост является существенным преимуществом, поскольку это сокращает время, необходимое для проведения селекции.
Мутантный фолатный рецептор, используемый в соответствии с настоящим изобретением, можно получать из фолатного рецептора любого вида при условии, что он будет функциональным в рамках настоящего изобретения, то есть, совместимым с используемой клеткой-хозяином и при экспрессии в трансфицированной клетке-хозяине будет вводить фолат, в частности фолиевую кислоту, из культуральной среды в клетку-хозяина.
Как правило, мутантный фолатный рецептор, который вводят в эукариотическую клетку-хозяина и используют в качестве селективного маркера, может быть гомологичным или гетерологичным для эндогенного фолатного рецептора клетки-хозяина (если эндогенный фолатный рецептор имеется, что предпочтительно). Если он является гомологичным, он будет происходить из того же вида, что и клетка-хозяин. Если он является гетерологичным, он будет происходить из иных видов, чем клетка-хозяин. Например, человеческий фолатный рецептор можно использовать в качестве селективного маркера для клетки-хозяина грызунов, например, клетки CHO. Предпочтительно, используют фолатный рецептор, полученный от каких-либо видов млекопитающих, например, полученный от грызунов, таких как мышь, крыса или хомяк, либо, более предпочтительно, полученный от человека. В одном варианте осуществления в качестве селективного маркера используют мутантный фолатный рецептор, полученный из человеческого фолатного рецептора альфа.
Мутантный фолатный рецептор можно выбирать из группы, состоящей из фолатного рецептора альфа, фолатного рецептора бета и фолатного рецептора гамма. Мутантный фолатный рецептор можно получать из фолатного рецептора дикого типа, содержащего аминокислотную последовательность, приведенную в SEQ ID NO 1, 3, 4, 6, 7 и 8, ниже, при этом, однако, указанный мутантный фолатный рецептор имеет по меньшей мере одну мутацию, которая приводит к снижению аффинности связывания фолата по сравнению с соответствующим фолатным рецептором дикого типа. Предпочтительно, мутантный фолатный рецептор получают из фолатного рецептора альфа, в частности, человеческого фолатного рецептора альфа.
Зрелый человеческий фолатный рецептор альфа дикого типа содержит следующую аминокислотную последовательность (SEQ ID NO 1, 1-буквенный код, показано в направлении от N-конца к C-концу):
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEAHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA
Фолатный рецептор альфа естественным образом заякорен в клеточной мембране с помощью GPI якоря. Сигнальная последовательность для GPI якоря не приведена в SEQ ID NO 1. В одном варианте осуществления мутантный фолатный рецептор альфа, полученный из SEQ ID NO 1, содержит сигнал GPI якоря на C-конце. Можно использовать любой подходящий сигнал GPI якоря. Природная сигнальная последовательность GPI якоря человеческого фолатного рецептора альфа является следующей (SEQ ID NO 2, 1-буквенный код, показано в направлении от N-конца к C-концу):
AAMSGAGPWAAWPFLLSLALMLLWLLS
Альтернативно, заякоривание в мембране можно осуществлять с использованием мембранного якоря, например, трансмембранного якоря. В таком варианте осуществления мутантный фолатный рецептор содержит мембранный якорь на своем C-конце. Подходящие якоря известны в данной области техники.
Мутантный фолатный рецептор альфа, полученный из SEQ ID NO 1, может содержать лидерную последовательность на N-конце. Можно использовать любую подходящую лидерную последовательность, которая обеспечивает функциональную экспрессию мутантного фолатного рецептора.
Полная аминокислотная последовательность, включая природную лидерную последовательность (на N-конце, подчеркнута) и природную сигнальную последовательность GPI якоря (на C-конце, подчеркнута) человеческого фолатного рецептора альфа дикого типа является следующей (SEQ ID NO 3, 1-буквенный код, показано в направлении от N-конца к C-концу):
MAQRMTTQLLLLLVWVAVVGEAQTRIAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEAHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYAAAMSGAGPWAAWPFLLSLALMLLWLLS
Последовательность зрелого человеческого фолатного рецептора бета дикого типа представляет собой следующую аминокислотную последовательность (SEQ ID NO 4, 1-буквенный код, показано в направлении от N-конца к C-концу):
QDRTDLLNVCMDAKHHKTKPGPEDKLHDQCSPWKKNACCTASTSQELHKDTSRLYNFNWDHCGKMEPACKRHFIQDTCLYECSPNLGPWIQQVNQTWRKERFLDVPLCKEDCQRWWEDCHTSHTCKSNWHRGWDWTSGVNKCPAGALCRTFESYFPTPAALCEGLWSHSYKVSNYSRGSGRCIQMWFDSAQGNPNEEVARFYA
Фолатный рецептор бета естественным образом заякорен в клеточной мембране с помощью GPI якоря. Сигнальная последовательность для GPI якоря не приведена в SEQ ID NO 4. В одном варианте осуществления мутантный фолатный рецептор бета, полученный из SEQ ID NO 4, содержит сигнал GPI якоря на C-конце. Можно использовать любой подходящий сигнал GPI якоря. Природная сигнальная последовательность GPI якоря человеческого фолатного рецептора бета является следующей (SEQ ID NO 5, 1-буквенный код, показано в направлении от N-конца к C-концу):
AAMHVNAGEMLHGTGGLLLSLALMLQLWLLG
Заякоривание в мембране также можно осуществлять с использованием мембранного якоря, например, трансмембранного якоря. В таком варианте осуществления мутантный фолатный рецептор содержит мембранный якорь на своем C-конце. Подходящие якоря известны в данной области техники.
Мутантный фолатный рецептор бета, полученный из SEQ ID NO 4, может содержать лидерную последовательность на N-конце. Можно использовать любую подходящую лидерную последовательность, которая обеспечивает функциональную экспрессию мутантного фолатного рецептора.
Полная аминокислотная последовательность, включая лидерную последовательность (на N-конце, подчеркнута) и природную сигнальную последовательность GPI якоря (на C-конце, подчеркнута) человеческого фолатного рецептора бета дикого типа является следующей (SEQ ID NO 6, 1-буквенный код, показано в направлении от N-конца к C-концу):
MVWKWMPLLLLLVCVATMCSAQDRTDLLNVCMDAKHHKTKPGPEDKLHDQCSPWKKNACCTASTSQELHKDTSRLYNFNWDHCGKMEPACKRHFIQDTCLYECSPNLGPWIQQVNQTWRKERFLDVPLCKEDCQRWWEDCHTSHTCKSNWHRGWDWTSGVNKCPAGALCRTFESYFPTPAALCEGLWSHSYKVSNYSRGSGRCIQMWFDSAQGNPNEEVARFYAAAMHVNAGEMLHGTGGLLLSLALMLQLWLLG
Кроме того, можно использовать фолатный рецептор, который естественным образом не связан с мембраной. Такой не связанный с мембраной фолатный рецептор можно изменять, делая его мембраносвязанным. Например, можно добавлять мембранный якорь и указанный фолатный рецептор можно экспрессировать в виде слитого белка, содержащего фолатный рецептор и мембранный якорь из другого полипептида. Кроме того, последовательность можно модифицировать для включения сигнальной последовательности GPI якоря. Соответствующие сигнальные последовательности GPI якоря описаны выше и также известны в данной области. Таким образом, фолатный рецептор можно заякоривать в клеточной мембране при помощи GPI якоря. Аналогично, можно использовать другие варианты, которые легко доступны для специалиста в данной области. Предпочтительным примером в этом отношении может быть мутантный фолатный рецептор на основе фолатного рецептора гамма, предпочтительно человеческого фолатного рецептора гамма, который был генетически изменен для включения мембранного якоря. В настоящем описании последовательность фолатного рецептора гамма мутирована в соответствии с идеями настоящего изобретения для демонстрации снижения аффинности связывания фолата.
Человеческий растворимый фолатный рецептор гамма дикого типа имеет следующую аминокислотную последовательность (SEQ ID NO 7, 1-буквенный код, показано в направлении от N-конца к C-концу):
qprsarartdllnvcmnakhhktqpspedelygqcspwkknacctastsqelhkdtsrlynfnwdhcgkmeptckrhfiqdsclyecspnlgpwirqvnqswrkerilnvplckedcerwwedcrtsytcksnwhkgwnwtsginecpagalcstfesyfptpaalceglwshsfkvsnysrgsgrciqmwfdsaqgnpneevakfyaaamnagapsrgiids
Кроме того, мутантный фолатный рецептор гамма, полученный из SEQ ID NO 7, может содержать лидерную последовательность на N-конце. Можно использовать любую подходящую лидерную последовательность, которая обеспечивает функциональную экспрессию мутантного фолатного рецептора.
Полная аминокислотная последовательность, включая лидерную последовательность человеческого фолатного рецептора гамма дикого типа (подчеркнута), является следующей (SEQ ID NO 8, 1-буквенный код, показано в направлении от N-конца к C-концу):
mdmawqmmqllllalvtaagsaqprsarartdllnvcmnakhhktqpspedelygqcspwkknacctastsqelhkdtsrlynfnwdhcgkmeptckrhfiqdsclyecspnlgpwirqvnqswrkerilnvplckedcerwwedcrtsytcksnwhkgwnwtsginecpagalcstfesyfptpaalceglwshsfkvsnysrgsgrciqmwfdsaqgnpneevakfyaaamnagapsrgiids
Мутантный фолатный рецептор по настоящему изобретению на основе фолатного рецептора гамма имеет по меньшей мере одну мутацию в соответствующей последовательности для получения мутантного фолатного рецептора, имеющего сниженную аффинность связывания фолата. Предпочтительно, мутация имеет место в кармане связывания фолата.
В одном варианте осуществления мутантный фолатный рецептор получен из фолатного рецептора альфа или бета. В одном варианте осуществления мутантный фолатный рецептор получен путем создания химерной аминокислотной последовательности, происходящей из фолатного рецептора альфа и бета. В фолатном рецепторе альфа и бета важными аминокислотными положениями, участвующими в связывании лиганда, являются, при соотнесении с соответствующей аминокислотной последовательностью зрелого фолатного рецептора (смотри, например, SEQ ID NO 1 и 4), положения 49, 104 и 166 (смотри также Ramamoorthy et al., 2007). В одном варианте осуществления мутантный фолатный рецептор имеет по меньшей мере одну замену в аминокислотном положении, соответствующем структурно или по гомологии аминокислотных последовательностей аминокислотному положению, выбранному из положений 49, 104 и 166 соответствующей последовательности дикого типа. Кроме того, более чем одна аминокислота может быть заменена в соответствующих положениях в мутантном фолатном рецепторе. Замена в одном или более из этих аминокислотных положений оказывает сильное влияние на аффинность связывания фолата. Предпочтительно, замена приводит к снижению аффинности связывания фолата у мутантного фолатного рецептора по сравнению с соответствующим фолатным рецептором дикого типа. В одном варианте осуществления полученный мутантный фолатный рецептор имеет, по сравнению с соответствующим фолатным рецептором дикого типа, сниженную аффинность связывания для 6S диастереоизомера 5-метилтетрагидрофолата. В одном варианте осуществления полученный мутантный фолатный рецептор имеет более слабое связывание с фолиевой кислотой. В одном варианте осуществления аминокислота, естественным образом присутствующая в соответствующей последовательности дикого типа, заменена за счет неконсервативной аминокислотной замены, при этом указанная замена снижает аффинность связывания фолата у мутантного фолатного рецептора. В одном варианте осуществления аминокислота, естественным образом присутствующая в соответствующей последовательности дикого типа, заменена за счет консервативной аминокислотной замены. При консервативной замене аминокислоту заменяют другой аминокислотой в пределах группы с аналогичными свойствами. Примерами соответствующих групп являются:
- Аминокислоты, имеющие неполярные боковые цепи: A, G, V, L, I, P, F, W, M
- Незаряженные аминокислоты, имеющие полярные боковые цепи: S, T, G, C, Y, N, Q
- Аминокислоты, имеющие ароматические боковые цепи: F, Y, W
- Положительно заряженные аминокислоты: K, R, H
- Отрицательно заряженные аминокислоты: D, E
- Аминокислоты с аналогичным размером или молекулярной массой, при этом молекулярная масса заменяющих аминокислот отличается максимально на +/- 25% (или +/- 20%, +/- 15%, +/- 10%) от молекулярной массы исходной аминокислоты.
Само собой разумеется, что группы также включают модифицированные аминокислоты и неприродные аминокислоты с соответствующим профилем боковых цепей, такие как, например, гомоаргинин в случае группы с положительно заряженными боковыми цепями. В одном варианте осуществления аминокислота, естественным образом присутствующая в последовательности дикого типа, заменена природной L-аминокислотой для получения мутантного фолатного рецептора.
Предпочтительно, мутантный фолатный рецептор представляет собой фолатный рецептор альфа. Он может быть получен из рецептора грызунов, таких как мышь, крыса или хомяк, или может быть получен из человеческого фолатного рецептора альфа. Предпочтительно, мутантный фолатный рецептор получен из человеческого фолатного рецептора альфа. В одном варианте осуществления мутантный фолатный рецептор по настоящему изобретению получен из человеческого фолатного рецептора альфа дикого типа, имеющего SEQ ID NO 1 или SEQ ID NO 3, приведенные выше, однако при этом указанный мутантный фолатный рецептор альфа имеет по меньшей мере одну мутацию, которая приводит к снижению аффинности связывания фолата по сравнению с фолатным рецептором дикого типа. В одном варианте осуществления полученный мутантный фолатный рецептор имеет, по сравнению с соответствующим фолатным рецептором дикого типа, сниженную аффинность связывания для 6S диастереоизомера 5-метилтетрагидрофолата. В одном варианте осуществления полученный мутантный фолатный рецептор альтернативно или дополнительно имеет сниженную аффинность связывания для фолиевой кислоты.
Предпочтительно, мутантный фолатный рецептор по настоящему изобретению имеет замену в аминокислотном положении, которое соответствует структурно или по гомологии аминокислотных последовательностей аминокислоте 49 в последовательности зрелого человеческого фолатного рецептора альфа дикого типа, приведенной в SEQ ID NO 1. Мутация в положении 49 зрелой последовательности фолатного рецептора альфа дикого типа соответствует мутации в кармане связывания фолата и, таким образом, оказывает сильное влияние на аффинность связывания фолата. Этот аланин в положении 49 последовательности дикого типа присутствует в последовательности человеческого, а также в последовательности соответствующего мышиного фолатного рецептора альфа дикого типа. Разумеется, мутантный фолатный рецептор по настоящему изобретению может содержать дополнительные мутации в других положениях, при условии, что мутантный фолатный рецептор является функциональным. В одном варианте осуществления по меньшей мере одна мутация, приводящая к снижению аффинности связывания фолата по сравнению с фолатным рецептором дикого типа, представляет собой замену аланина, находящегося в положении 49 последовательности зрелого фолатного рецептора альфа дикого типа, аминокислотой, выбранной из группы, состоящей из лейцина, глицина, валина, изолейцина, гистидина и аспарагиновой кислоты. Предпочтительно, аланин заменен лейцином. Авторы изобретения неожиданно обнаружили, что замена A49L в последовательности фолатного рецептора альфа приводит к созданию мутантного фолатного рецептора альфа, имеющего превосходящие характеристики в качестве селективного маркера по сравнению с соответствующим фолатным рецептором альфа дикого типа. Мутантный фолатный рецептор альфа, содержащий соответствующую замену A49L, имеет, по сравнению с соответствующим фолатным рецептором альфа дикого типа, сниженную аффинность связывания для фолата, а именно, 6S диастереоизомера 5-метилтетрагидрофолата. Кроме того, как показано в примерах, мутант A49L человеческого фолатного рецептора альфа имеет существенные преимущества при использовании в качестве селективного маркера для идентификации и селекции успешно трансфицированных клеток-хозяев млекопитающих. Таким образом, его предпочтительно используют в качестве селективного маркера для идентификации клеток-хозяев, экспрессирующих рекомбинантный интересующий полипептид с высоким выходом.
В одном варианте осуществления зрелый мутантный фолатный рецептор содержит аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97% или по меньшей мере 98%, или по меньшей мере 99% со зрелой последовательностью дикого типа человеческого фолатного рецептора альфа (SEQ ID NO 1), однако при этом аминокислотная последовательность зрелого мутантного фолатного рецептора имеет по меньшей мере одну мутацию, которая приводит к снижению аффинности связывания фолата по сравнению с человеческим фолатным рецептором альфа дикого типа. Как обсуждалось выше, по меньшей мере одна мутация, приводящая к снижению аффинности связывания фолата по сравнению с фолатным рецептором дикого типа, предпочтительно представляет собой замену аланина, находящегося в положении 49 последовательности зрелого фолатного рецептора альфа дикого типа (смотри SEQ ID NO. 1), аминокислотой, выбранной из группы, состоящей из лейцина, глицина, валина, изолейцина, гистидина и аспарагиновой кислоты. Предпочтительно, аланин в положении 49 заменен лейцином. Такой мутантный фолатный рецептор имеет, по сравнению с соответствующим фолатным рецептором дикого типа, сниженную аффинность связывания для 6S диастереоизомера 5-метилтетрагидрофолата и улучшенные характеристики в качестве селективного маркера.
В одном варианте осуществления первый полинуклеотид кодирует мутантный фолатный рецептор, при этом указанный мутантный фолатный рецептор имеет следующие характеристики:
a) зрелый мутантный фолатный рецептор содержит следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9)
где Xaa не является аланином и где, предпочтительно, Xaa является аминокислотой, выбранной из лейцина, глицина, валина, изолейцина, гистидина и аспарагиновой кислоты, и при этом более предпочтительно, Xaa является лейцином;
или
b) зрелый мутантный фолатный рецептор содержит аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, или 98%, или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9, и при этом Xaa не является аланином в указанном мутантном фолатном рецепторе и предпочтительно Xaa является аминокислотой, выбранной из лейцина, глицина, валина, изолейцина, гистидина и аспарагиновой кислоты, и более предпочтительно, Xaa является лейцином, и при этом аффинность связывания фолата у указанного мутантного фолатного рецептора снижена по сравнению с последовательностью зрелого человеческого фолатного рецептора альфа дикого типа, в которой Xaa является аланином (смотри SEQ ID NO 1). В одном варианте осуществления указанный мутантный фолатный рецептор имеет, по сравнению с соответствующим фолатным рецептором дикого типа, сниженную аффинность связывания для 6S диастереоизомера 5-метилтетрагидрофолата. В одном варианте осуществления полученный мутантный фолатный рецептор дополнительно или альтернативно имеет сниженную аффинность связывания для фолиевой кислоты. Мутантный фолатный рецептор по пункту b) может рассматриваться как функциональный вариант рецептора по пункту a) и может иметь одну или более дополнительных аминокислотных мутаций по сравнению с мутантным фолатным рецептором по пункту a). Например, он может иметь одну или более дополнительных замен, делеций и/или добавлений одной или более аминокислот при условии, что его функция как фолатного рецептора не исчезает. Сюда также относятся слитые белки, содержащие последовательность соответствующего мутантного фолатного рецептора.
Как обсуждалось выше, предпочтительно, если Xaa является лейцином. Как показано в примерах, мутация остатка аланина, находящегося в положении 49 последовательности дикого типа фолатного рецептора альфа, на остаток лейцина приводит к созданию мутантного фолатного рецептора, который, по сравнению с соответствующей последовательностью дикого типа, обладает превосходными характеристиками в качестве селективного маркера. Как показано в примерах, клетки, содержащие в качестве селективного маркера мутантный фолатный рецептор, несущий мутацию в положении, соответствующем положению 49 зрелой последовательности дикого типа фолатного рецептора альфа, демонстрируют после селекции высокую продуктивность для интересующего полипептида, которая зачастую даже значительно выше, чем продуктивность, которая достигается при использовании соответствующего фолатного рецептора дикого типа в качестве селективного маркера, и которая также выше, чем продуктивность, которая достигается при использовании других мутантных форм рецептора. Более того, клетки быстрее восстанавливаются после селекции. Эти важные преимущества делают A49L мутантный фолатный рецептор особенно подходящим в качестве селективного маркера. Указанный мутантный фолатный рецептор альфа описан и охарактеризован в публикации Shen et al., 1997. В этой публикации продемонстрировано, что указанный мутантный вариант имеет сниженную аффинность связывания для 6S диастереоизомера 5-метилтетрагидрофолата, о чем можно судить по величине IC50 (нМ), которая возрастает по сравнению с фолатным рецептором альфа дикого типа (2,9) почти в 60 раз (до 179,0).
Мутантный фолатный рецептор является мембраносвязанным и может содержать, например, GPI якорь или трансмембранный якорь. Как описано выше, фолатные рецепторы альфа и бета естественным образом заякорены при помощи GPI якоря в клеточной мембране. При использовании GPI якоря для заякоривания в мембране кодирующий полинуклеотид должен иметь соответствующую сигнальную последовательность для присоединения GPI якоря. Подходящие сигнальные последовательности для GPI якоря известны в данной области и также описаны выше. Как указано выше, соответствующие сигнальные последовательности GPI якоря находятся на C-конце и могут быть использованы в сочетании с настоящим изобретением.
В одном варианте осуществления незрелый мутантный фолатный рецептор содержит лидерную последовательность функционального человеческого фолатного рецептора альфа дикого типа, приведенную далее (SEQ ID NO 10, 1-буквенный код, показано в направлении от N-конца к C-концу):
MAQRMTTQLLLLLVWVAVVGEAQTR
Лидерные последовательности человеческих фолатных рецепторов бета и гамма дикого типа приведены далее (SEQ ID NO 11 и 12, 1-буквенный код, показано в направлении от N-конца к C-концу):
MVWKWMPLLLLLVCVATMCSA (SEQ ID NO 11)
mdmawqmmqllllalvtaagsa (SEQ ID NO 12)
В одном варианте осуществления первый полинуклеотид кодирует мутантный фолатный рецептор, при этом указанный мутантный фолатный рецептор имеет следующие характеристики:
a) мутантный фолатный рецептор содержит следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYAAAMSGAGPWAAWPFLLSLALMLLWLLS (SEQ ID NO 13)
где Xaa является лейцином;
или
b) мутантный фолатный рецептор содержит аминокислотную последовательность, которая имеет идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 13, где Xaa является лейцином в указанном мутантном фолатном рецепторе по пункту b), и при этом аффинность связывания указанного мутантного фолатного рецептора для 6S диастереоизомера 5-метилтетрагидрофолата снижена по сравнению с последовательностью зрелого человеческого фолатного рецептора альфа дикого типа, где Xaa является аланином (смотри SEQ ID NO 1).
Интересующий полипептид
Экспрессионный вектор или сочетание экспрессионных векторов содержат по меньшей мере один полинуклеотид, кодирующий интересующий полипептид. Когда указанный экспрессионный вектор или сочетание векторов вводят в зависимую от фолата клетку-хозяина, такую как, например, клетка млекопитающего, описанная в настоящем документе, интересующий полипептид секретируется из указанной клетки-хозяина. Таким образом, интересующий полипептид является секретируемым полипептидом. Полинуклеотид может кодировать полипептид, который естественным образом секретируется, или он может быть изменен, чтобы стать секретируемым, за счет соответствующей секреторной лидерной последовательности. Большинство секретируемых полипептидов имеют аминоконцевой лидерный пептид (также называемый лидерной последовательностью или сигнальным пептидом), который отщепляется от образующегося полипептида-предшественника во время биосинтеза. Секреторные лидерные пептиды, как правило, имеют длину от 5 до 60 аминокислот. Эта последовательность является необходимой и достаточной для секреции. Многочисленные примеры секреторных лидерных последовательностей хорошо известны в данной области и, таким образом, нет необходимости в подробном описании их в настоящем документе. Анализ большого числа этих секреторных лидерных пептидов выявил общий структурный мотив, который существует в отсутствие значительной гомологии аминокислотных последовательностей [Von Heijne, 1981; Perlman et al., 1983, Bird et al., 1990]. Как правило, секреторная лидерная последовательность состоит из положительно заряженного амино-конца (n), гидрофобного ядра (h) и более полярного карбокси-конца (c), который определяет сайт расщепления сигнальной пептидазой. Нарушение области h за счет делеции или за счет замены гидрофобных остатков гидрофильными или заряженными аминокислотами приводит к утрате сигнальной функции, в то время как изменения в области «n» оказывают небольшое влияние. Карбокси-конец, или сайт расщепления, как правило, имеет длину примерно 6 аминокислот. Эта область участвует в узнавании и расщеплении сигнальной пептидазой, что обычно необходимо для окончательного сворачивания и секреции белка.
Интересующий полипептид может представлять собой фармацевтически или терапевтически активное соединение, или исследовательский инструмент, используемый в анализах, и тому подобное. Интересующий полипептид может быть полипептидом любого типа. Термин «полипептид» означает молекулу, представляющую собой полимер из аминокислот, связанных вместе пептидной связью(ми). Полипептиды включают полипептиды любой длины, в том числе белки (например, содержащие более 50 аминокислот) и пептиды (например, содержащие 2-49 аминокислот). Полипептиды включают белки и/или пептиды с любой активностью, функцией или размером, и могут включать, например, ферменты (например, протеазы, киназы, фосфатазы), рецепторы, переносчики, бактерицидные и/или эндотоксин-связывающие белки, структурные полипептиды, гликопротеины, глобулярные белки, иммунные полипептиды, токсины, антибиотики, гормоны, факторы роста, факторы крови, вакцины или тому подобное. Полипептид может быть выбран из группы, состоящей из пептидных гормонов, интерлейкинов, тканевых активаторов плазминогена, цитокинов, иммуноглобулинов, в частности, антител или функциональных фрагментов антител, или их вариантов и Fc-слитых белков. Интересующий полипептид, экспрессируемый в соответствии с идеями, изложенными в настоящем документе, также может представлять собой субъединицу или домен полипептида, такой как, например, тяжелая цепь или легкая цепь антитела, или их функциональный фрагмент или производное. Термин «интересующий полипептид» может относиться к такой отдельной субъединице или домену, или конечному белку, состоящему из соответствующих субъединиц или доменов, в зависимости от контекста. В предпочтительном варианте осуществления интересующий полипептид представляет собой молекулу иммуноглобулина, более предпочтительно, антитело или его субъединицу, или домен, такой как, например, тяжелая или легкая цепь антитела. Используемый в настоящем документе термин «антитело», в частности, относится к белку, содержащему по меньшей мере две тяжелые цепи и две легкие цепи, связанные дисульфидными связями. Термин «антитело» охватывает природные антитела, а также рекомбинантные формы антител, например, гуманизированные антитела, полностью человеческие антитела и химерные антитела. Каждая тяжелая цепь, как правило, состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи (CH). Каждая легкая цепь, как правило, состоит из вариабельной области легкой цепи (VL) и константной области легкой цепи (CL). Однако термин «антитело» также включает другие типы антител, такие как однодоменные антитела, антитела из тяжелых цепей, то есть, антитела, состоящие только из одной или более, в частности, двух тяжелых цепей, и нанотела, то есть, антитела, состоящие только из одного мономерного вариабельного домена. Как обсуждалось выше, полинуклеотид, кодирующий интересующий полипептид, также может кодировать одну или более субъединиц или доменов антитела, например, тяжелую или легкую цепь, или их функциональный фрагмент, или производное, в качестве интересующего полипептида. Указанные субъединицы или домены могут экспрессироваться либо с одной и той же, либо с разных экспрессионных кассет. Термин «функциональный фрагмент или производное» антитела, в частности, относится к полипептиду, который получен из антитела и способен связывать тот же антиген, в частности, тот же эпитоп, что и антитело. Было показано, что антигенсвязывающая функция антитела может осуществляться фрагментами полноразмерного антитела или его производными. Примеры фрагментов или производных антитела включают (i) Fab фрагменты, одновалентные фрагменты, состоящие из вариабельной области и первого константного домена каждой из тяжелых и легких цепей; (ii) F(ab)2 фрагменты, двухвалентные фрагменты, содержащие два Fab фрагмента, связанные дисульфидной связью в шарнирной области; (iii) Fd фрагменты, состоящие из вариабельной области и первого константного домена CH1 тяжелой цепи; (iv) Fv фрагменты, состоящие из вариабельной области тяжелой цепи и легкой цепи одного плеча антитела; (v) scFv фрагменты, Fv фрагменты, состоящие из одной полипептидной цепи; (vi) (Fv)2 фрагменты, состоящие из двух Fv фрагментов, ковалентно связанных вместе; (vii) вариабельный домен тяжелой цепи и (viii) мультитела, состоящие из вариабельной области тяжелой цепи и вариабельной области легкой цепи, ковалентно связанных вместе таким образом, что ассоциация вариабельных областей тяжелой цепи и легкой цепи может быть только межмолекулярной, но не внутримолекулярной.
Дополнительные селективные маркеры
В одном варианте осуществления экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов по настоящему изобретению дополнительно содержит(содержат) один или более полинуклеотидов, кодирующих дополнительный селективный маркер. Селективный маркер позволяет в соответствующих селективных условиях культивирования отбирать клетки-хозяева, экспрессирующие указанный селективный маркер. Селективный маркер обеспечивает носителя указанного маркера в селективных условиях преимуществом для выживания и/или роста. Как правило, ген селективного маркера придает устойчивость к селективному средству, такому как лекарственное средство, например, антибиотик или другое токсическое средство, или компенсирует метаболический или катаболический дефект клетки-хозяина. Маркер может быть маркером для положительной или отрицательной селекции. Для селекции успешно трансфицированных клеток-хозяев при культивировании клеток-хозяев можно использовать культуральную среду, содержащую селективное средство, позволяющее отбирать на используемый селективный маркер. В других вариантах осуществления селективный маркер позволяет клетке-хозяину выживать и пролиферировать при отсутствии или при недостатке соединения, которое необходимо для выживания и/или пролиферации клеток-хозяев, лишенных селективного маркера. В одном варианте осуществления селективный маркер представляет собой маркер устойчивости к лекарственному средству, кодирующий белок, придающий устойчивость к селективным условиям с применением указанного лекарственного средства. Различные гены селективных маркеров хорошо известны специалистам и описаны в литературе (смотри, например, WO 92/08796, WO 94/28143, WO 2004/081167, WO 2009/080759, WO 2010/097240). В одном варианте осуществления селективный маркер может представлять собой амплифицируемый селективный маркер. Гены селективных маркеров, обычно используемые с клетками млекопитающих, включают гены аминогликозид фосфотрансферазы (APH), гигромицин фосфотрансферазы (hyg), дигидрофолатредуктазы (DHFR), тимидинкиназы (tk), глутамин-синтетазы, аспарагин-синтетазы, а также гены, кодирующие устойчивость к неомицину (G418), пуромицину, гигромицину и зеоцину. Такие селективные маркеры можно использовать в дополнение к мутантному фолатному рецептору.
В одном варианте осуществления экспрессионный вектор или сочетание экспрессионных векторов содержат дополнительный полинуклеотид, кодирующий селективный маркер, участвующий в метаболизме фолата, и при этом на активность указанного селективного маркера, по меньшей мере частично, оказывает влияние активность мутантного фолатного рецептора. Характерная особенность, заключающаяся в том, что на активность дополнительного селективного маркера, по меньшей мере частично, оказывает влияние активность мутантного фолатного рецептора, в частности, означает, что активность указанного дополнительного селективного маркера находится под влиянием и/или зависит, по меньшей мере в какой-то степени, прямо или косвенно от активности или функции мутантного фолатного рецептора. Эту зависимость/взаимодействие мутантного фолатного рецептора и дополнительного селективного маркера можно использовать для значительного увеличения давления отбора на клетки-хозяева при селективных условиях культивирования.
В одном варианте осуществления дополнительный селективный маркер представляет собой фермент, осуществляющий процессинг субстрата, который представляет собой фолат, производное фолата и/или продукт, который может быть получен в результате процессинга фолата, например, DHF или THF, или функциональный вариант или производное вышеперечисленных. Соответствующие субстраты важны для производства нуклеиновых кислот. Предпочтительно, дополнительный селективный маркер представляет собой дигидрофолатредуктазу (DHFR) или фермент, действующий после или в сочетании с DHFR, такой как тимидилатсинтаза (TS) и серин-гидроксиметилтрансфераза (SHMT). Предпочтительно, дополнительный селективный маркер представляет собой DHFR. DHFR также может экспрессироваться в виде части слитого белка.
Использование соответствующего сочетания селективных маркеров, то есть, мутантного фолатного рецептора по настоящему изобретению и дополнительного селективного маркера, участвующего в метаболизме фолата, который описан выше, предпочтительно DHFR, обеспечивает очень строгую систему селекции для получения и обогащения продуцирующих на высоком уровне клеток из популяции трансфицированных клеток-хозяев. Данная концепция использования фолатного рецептора в качестве селективного маркера в сочетании с дополнительным селективным маркером, участвующим в метаболизме фолата, таким как, предпочтительно, DHFR, и связанные с этим преимущества раскрыты в WO 2010/097240, который включен в настоящий документ посредством ссылки. Как показано в примерах, высокая строгость системы селекции по данному варианту осуществления значительно снижает количество слабых продуцентов в популяции, полученной после селекции, и тем самым повышает шансы обнаружить очень редкие клоны-суперпродуценты. Кроме того, после селекции получают более гомогенную популяцию продуцирующих на высоком уровне клеток, что уменьшает усилия, затрачиваемые на скрининг. Это упрощает одноклеточное клонирование продуцирующих на высоком уровне клеток. Как показано в примерах, использование мутантного фолатного рецептора, описанного в настоящем документе, в сочетании с дополнительным селективным маркером, участвующим в метаболизме фолата, описанным выше, предпочтительно DHFR, приводит к улучшенным результатам по сравнению с использованием фолатного рецептора дикого типа в сочетании с таким селективным маркером. Таким образом, при использовании сочетания соответствующих селективных маркеров настоящее изобретение также обеспечивает значительные преимущества за счет использования мутантного фолатного рецептора.
Как обсуждалось выше, дополнительный селективный маркер предпочтительно представляет собой фермент DHFR. В данной области известно несколько подходящих ферментов DHFR и соответствующих генов, которые можно использовать в качестве селективного маркера в сочетании с настоящим изобретением. Термины «дигидрофолатредуктаза» или «DHFR» означают DHFR дикого типа, а также ферменты DHFR, имеющие одно или более изменений аминокислотной последовательности (например, делеции, замены или добавления) относительно аминокислотной последовательности соответствующего фермента DHFR дикого типа, слитые белки, содержащие фермент DHFR, и ферменты DHFR, которые были модифицированы для создания дополнительной структуры и/или функции, а также функциональные фрагменты вышеперечисленных, которые все-еще сохраняют по меньшей мере одну функцию фермента DHFR. Такие варианты осуществления хорошо известны в данной области и, таким образом, нет необходимости в их подробном описании. Например, в качестве селективного маркера можно использовать фермент DHFR, который более или менее чувствителен к антифолатам, таким как MTX, чем фермент DHFR дикого типа и/или фермент DHFR, эндогенно экспрессируемый клеткой-хозяином, в случае его экспрессии. Соответствующие ферменты DHFR хорошо известны в данной области и описаны, например, в EP 0 246 049 и других документах. Фермент DHFR можно получать из любых видов при условии, что он будет функциональным в рамках настоящего изобретения, то есть, совместимым с используемой клеткой-хозяином млекопитающего. Например, мутантный мышиный фермент DHFR с основной устойчивостью к MTX широко используется в качестве доминантного селективного маркера в клетках млекопитающих. В качестве селективного маркера можно использовать фермент DHFR, который менее чувствителен к ингибитору DHFR, такому как MTX, чем фермент DHFR, эндогенно экспрессируемый в DHFR+ (плюс) клетке-хозяине, и, следовательно, клетке-хозяине, содержащей функциональный эндогенный ген DHFR. В одном варианте осуществления интрон или его фрагмент помещают на 3’-конце открытой рамки считывания гена DHFR. Интрон, используемый в экспрессионной кассете DHFR, приводит к меньшему по размеру, нефункциональному варианту гена DHFR (Grillari et al., 2001, J. Biotechnol. 87, 59-65). Таким образом, уровень экспрессии гена DHFR снижается, что дополнительно повышает строгость селекции. Альтернативные способы использования интрона для снижения уровня экспрессии гена DHFR описаны в EP0 724 639 и также могут быть использованы.
Полинуклеотид, кодирующий дополнительный селективный маркер, может быть расположен на том же экспрессионном векторе, что и полинуклеотид, кодирующий мутантный фолатный рецептор и/или по меньшей мере один полинуклеотид, кодирующий интересующий полипептид, или он может быть расположен на отдельном экспрессионном векторе, если используют сочетание экспрессионных векторов. В этом случае сочетание экспрессионных векторов, содержащих все полинуклеотиды (кодирующие мутантный фолатный рецептор, интересующий полипептид и дополнительный селективный маркер), будут совместно трансфицированы в клетки-хозяева для обеспечения селекции.
В предпочтительном варианте осуществления экспрессионный вектор или сочетание экспрессионных векторов содержат
- полинуклеотид, кодирующий мутантный фолатный рецептор, имеющий по меньшей мере одну мутацию, соответствующую структурно или по аминокислотному положению аминокислоте 49 в зрелой последовательности дикого типа человеческого фолатного рецептора альфа (смотри SEQ ID NO.1), при этом указанная мутация приводит к снижению аффинности связывания фолата по сравнению с фолатным рецептором альфа дикого типа, при этом, предпочтительно, аланин, находящийся в последовательности дикого типа в указанном положении, заменен лейцином, и
- полинуклеотид, кодирующий фермент DHFR, который менее чувствителен к MTX, чем фермент DHFR дикого типа и/или фермент DHFR, эндогенно экспрессируемый клеткой-хозяином, в качестве дополнительного селективного маркера. Указанный фермент DHFR предпочтительно также включает интрон, описанный выше. Соответствующее сочетание маркеров является особенно предпочтительным, если в качестве клеток-хозяев используют DHFR+ (плюс) клетки. DHFR+ (плюс) клетки экспрессируют эндогенный DHFR. Как показано в примерах, продуцирующие на очень высоком уровне клеточные клоны можно эффективно отбирать при использовании соответствующего вектора или сочетания экспрессионных векторов.
Экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов по настоящему изобретению могут дополнительно содержать один или более дополнительных полинуклеотидов, кодирующих селективный маркер. Такой дополнительный селективный маркер может присутствовать помимо мутантного фолатного рецептора и дополнительного селективного маркера, участвующего в метаболизме фолата, который предпочтительно представляет собой DHFR.
Помимо дополнительных эукариотических селективных маркеров, позволяющих проводить селекцию эукариотических клеток-хозяев, в экспрессионном векторе или сочетании экспрессионных векторов также могут присутствовать и прокариотические селективные маркеры. Это, например, позволяет амплифицировать вектор(ы) в прокариотах. «Прокариотический селективный маркер» представляет собой селективный маркер, позволяющий проводить селекцию в прокариотических клетках-хозяевах при подходящих условиях селекции. Примерами соответствующих прокариотических селективных маркеров являются маркеры, которые обеспечивают устойчивость к антибиотикам, таким как, например, ампициллин, канамицин, тетрациклин и/или хлорамфеникол.
Дополнительные элементы вектора и варианты осуществления экспрессионного вектора(ов)
Экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов могут, кроме того, содержать дополнительные элементы вектора. Например, может содержаться по меньшей мере один дополнительный полинуклеотид, кодирующий дополнительный интересующий полипептид. Как описано выше и как станет ясно из описанных примеров полипептидов, которые можно экспрессировать, конечный полипептид, который будет продуцироваться и секретироваться клеткой-хозяином, также может представлять собой белок, состоящий из нескольких отдельных субъединиц или доменов. Предпочтительным примером соответствующего белка является молекула иммуноглобулина, в частности, антитело, которое содержит, например, тяжелую и легкую цепи. Существует несколько вариантов продуцирования соответствующего белка, состоящего из различных отдельных субъединиц или доменов, и конструкции соответствующих векторов известны в данной области. В одном варианте осуществления две или более субъединиц или доменов указанного белка экспрессируются с одной экспрессионной кассеты. В этом варианте осуществления получают один длинный транскрипт с соответствующей экспрессионной кассеты, содержащий кодирующие области отдельных субъединиц или доменов белка. В одном варианте осуществления по меньшей мере один элемент IRES (внутренний сайт связывания рибосомы) функционально расположен между кодирующими областями отдельных субъединиц или доменов, и каждой кодирующей области предшествует секреторная лидерная последовательность. Тем самым гарантируется, что отдельные продукты трансляции будут получены с указанного транскрипта и что конечный белок может быть корректно собран и секретирован. Соответствующие технологии известны в данной области и, следовательно, нет необходимости в их подробном описании в настоящем документе.
Однако в объем настоящего изобретения также входит, и для некоторых вариантов осуществления, таких как экспрессия антител, даже является предпочтительной, экспрессия отдельных субъединиц или доменов с разных экспрессионных кассет. В одном варианте осуществления экспрессионная кассета, используемая для экспрессии интересующего полипептида, представляет собой моноцистронную экспрессионную кассету. Предпочтительно, все экспрессионные кассеты, содержащиеся в экспрессионном векторе или сочетании экспрессионных векторов, являются моноцистронными. В одном варианте осуществления, соответственно, каждая экспрессионная кассета содержит полинуклеотид, кодирующий одну субъединицу или домен белка, экспрессируемого в качестве интересующего полипептида. Например, в случае антител одна экспрессионная кассета кодирует легкую цепь антитела, а другая экспрессионная кассета кодирует тяжелую цепь антитела. После экспрессии отдельных субъединиц/доменов с отдельных экспрессионных кассет конечный белок, такой как антитело, собирается из указанных субъединиц или доменов и секретируется из клетки-хозяина. Этот вариант осуществления особенно подходит для экспрессии молекул иммуноглобулинов, таких как антитела. В этом случае первый полинуклеотид, кодирующий интересующий полипептид, кодирует, например, тяжелую или легкую цепь молекулы иммуноглобулина и второй полинуклеотид, кодирующий интересующий полипептид, кодирует другую цепь молекулы иммуноглобулина. В одном варианте осуществления экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов содержат по меньшей мере одну экспрессионную кассету, содержащую полинуклеотид, кодирующий тяжелую цепь молекулы иммуноглобулина или ее функциональный фрагмент, и по меньшей мере одна экспрессионную кассету, содержащую полинуклеотид, кодирующий легкую цепь молекулы иммуноглобулина или ее функциональный фрагмент. Указанные полинуклеотиды могут быть расположены на одном и том же или на разных экспрессионных векторах в случае использования сочетания по меньшей мере двух экспрессионных векторов. При экспрессии указанных полинуклеотидов в трансфицированной клетке-хозяине образуется функциональная молекула иммуноглобулина и секретируется из клетки-хозяина.
Экспрессионные векторы, используемые для экспрессии рекомбинантных интересующих продуктов, как правило, содержат в качестве элементов экспрессионной кассеты элементы контроля транскрипции, подходящие для управления транскрипцией, такие как, например, промоторы, энхансеры, сигналы полиаденилирования, сигналы временной остановки или терминации транскрипции. Предпочтительно, включены соответствующие элементы контроля трансляции, такие как, например, 5' нетранслируемые области, ведущие к структурам 5'-кэпа, предназначенным для рекрутинга рибосом, и стоп-кодоны для терминации процесса трансляции. Полученные транскрипты гена(ов) селективного маркера и гена(ов) интересующего полипептида содержат функциональные трансляционные элементы, способствующие экспрессии белка (то есть, трансляции) на значительных уровнях и надлежащей терминации трансляции. Функциональную единицу экспрессии, способную надлежащим образом управлять экспрессией встроенного полинуклеотида, в настоящем документе также называют «экспрессионной кассетой». Полинуклеотид(ы), кодирующий интересующий секретируемый полипептид, и полинуклеотиды, кодирующие селективные маркеры, описанные в настоящем документе, предпочтительно содержатся в экспрессионной кассете. Подходящими являются несколько вариантов осуществления, например, каждый из указанных полинуклеотидов может содержаться в отдельной экспрессионной кассете. Однако по меньшей мере два из соответствующих полинуклеотидов могут также содержаться в одной экспрессионной кассете. В одном варианте осуществления по меньшей мере один элемент внутреннего сайта связывания рибосомы (IRES) функционально расположен между полинуклеотидами, которые экспрессируются с одной и той же экспрессионной кассеты. Тем самым гарантируется, что с указанного транскрипта будут получены отдельные продукты трансляции. Соответствующие технологии экспрессии на базе IRES и другие би- и полицистронные системы хорошо известны и, следовательно, не нуждаются в дополнительном описании в настоящем документе.
Как описано, экспрессионный вектор или сочетание экспрессионных векторов по настоящему изобретению могут содержать по меньшей мере один элемент промотора и/или промотора/энхансера в качестве элемента экспрессионной кассеты. Промоторы можно подразделять на два класса, те, которые действуют конститутивно, и те, которые регулируются индукцией или дерепрессией. Оба вида подходят для использования в соответствии с идеями настоящего изобретения. Промоторы, используемые для продуцирования белков на высоком уровне в клетках млекопитающих, должны быть сильными и предпочтительно активными в широком диапазоне типов клеток. Сильные конститутивные промоторы, управляющие экспрессией в клетках многих типов, включают, но не ограничиваются ими, основной поздний промотор аденовируса, предранний промотор цитомегаловируса человека, промотор SV40 и вируса саркомы Рауса, а также промотор мышиной 3-фосфоглицераткиназы, EF1a. В одном варианте осуществления промотор и/или энхансер получен из CMV и/или из SV40. Промоторы транскрипции можно выбирать из группы, состоящей из промотора SV40, промотора CMV, промотора EF1альфа, промотора RSV, промотора BROAD3, промотора rosa 26 мыши, промотора pCEFL и промотора β-актина.
В одном варианте осуществления по меньшей мере один полинуклеотид, кодирующий интересующий полипептид, полинуклеотид, кодирующий мутантный фолатный рецептор, и/или полинуклеотид, кодирующий второй селективный маркер, находятся под контролем разных промоторов транскрипции. Отдельные промоторы транскрипции, управляющие экспрессией полинуклеотидов, могут быть одинаковыми или разными.
В одном варианте осуществления используют более сильный промотор и/или энхансер для управления экспрессией по меньшей мере одного полинуклеотида, кодирующего интересующий полипептид, чем для управления экспрессией полинуклеотида, кодирующего мутантный фолатный рецептор и/или один или более дополнительных селективных маркеров. Такая схема имеет тот эффект, что больше транскриптов создается для интересующего полипептида, чем для селективных маркеров. Предпочтительно, если продуцирование интересующего полипептида доминирует над продуцированием селективных маркеров, поскольку способность отдельной клетки продуцировать гетерологичные продукты не безгранична и, таким образом, следует сосредоточиться на интересующем полипептиде. Кроме того, процесс селекции имеет место только на начальных стадиях создания линии экспрессирующих клеток, которые затем постоянно продуцируют интересующий полипептид. Таким образом, предпочтительно сосредотачивать ресурсы клеток на экспрессии/продуцировании интересующего полипептида. Кроме того, если менее сильный промотор используют для экспрессии селективного маркера, чем для экспрессии интересующего полипептида, дополнительно возрастает давление отбора на трансфицированные клетки-хозяева.
В одном варианте осуществления промотор, управляющий экспрессией полинуклеотида(ов), кодирующего интересующий полипептид, представляет собой промотор CMV, а промотор, управляющий экспрессией полинуклеотида, кодирующего мутантный фолатный рецептор, представляет собой промотор SV40. Как известно, промотор CMV является одним из самых сильных промоторов, используемых для экспрессии в клетках млекопитающих, и позволяет достигать хорошего уровня экспрессии. Считается, что он обеспечивает больше транскриптов, чем промотор SV40. Однако также можно использовать и другие промоторы.
В следующем варианте осуществления по меньшей мере один полинуклеотид, кодирующий интересующий полипептид, и полинуклеотид, кодирующий мутантный фолатный рецептор, и/или полинуклеотид, кодирующий селективный маркер, при наличии, находятся под контролем одного и того же промотора транскрипции. Подходящие промоторы описаны выше. В этом варианте осуществления получают один длинный транскрипт с соответствующей экспрессионной кассеты, которая находится под контролем указанного промотора транскрипции. В одном варианте осуществления по меньшей мере один элемент IRES функционально расположен между полинуклеотидами, которые экспрессируются с одной и той же экспрессионной кассеты.
Экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов могут содержать соответствующий сайт терминации транскрипции в качестве элемента экспрессионной кассеты. Это явление, когда продолжающаяся транскрипция с расположенного выше промотора через вторую единицу транскрипции может ингибировать функцию расположенного ниже промотора, известно как окклюзия промотора или транскрипционная интерференция. Сайты терминации транскрипции хорошо охарактеризованы и, как показано, их включение в экспрессионные векторы имеет множество полезных эффектов на экспрессию гена.
Экспрессионные кассеты могут содержать сайт полиаденилирования. Существует несколько эффективных сигналов полиA, которые можно использовать в экспрессионных векторах млекопитающих, включая те, которые получены из бычьего гормона роста (bgh), мышиного бета-глобина, ранней транскрипционной единицы SV40 и гена тимидинкиназы вируса простого герпеса. Однако известны также синтетические сайты полиаденилирования (смотри, например, экспрессионный вектор pCI-neo от компании Promega на основе публикации Levitt el al., 1989, Genes Dev. 3, (7): 1019-1025). Сайт полиаденилирования можно выбирать из группы, состоящей из сайта полиA SV40, такого как поздний и ранний сайт поли-A SV40 (смотри, например, плазмиду pSV2-DHFR, описанную в публикации Subramani et al., 1981, Mol.Cell. Biol. 854-864), синтетического сайта полиA (смотри, например, экспрессионный вектор pCI-neo от компании Promega на основе публикации Levitt el al., 1989, Genes Dev. 3, (7): 1019-1025) и сайта полиA bgh (бычьего гормона роста).
Кроме того, экспрессионная кассета может содержать по меньшей мере один интрон. Как правило, интроны помещают на 5’-конце открытой рамки считывания, однако их также можно помещать на 3’-конце. Соответственно, интрон может присутствовать в экспрессионной кассете(ах) для повышения степени экспрессии. Указанный интрон может быть расположен между элементом(ами) промотора и/или промотора/энхансера и 5'-концом открытой рамки считывания экспрессируемого полинуклеотида. В данной области известно несколько подходящих интронов, которые можно использовать в сочетании с настоящим изобретением. В одном варианте осуществления интрон, используемый в экспрессионных кассетах для экспрессии интересующего полипептида, представляет собой синтетический интрон, такой как интрон SIS или RK. Интрон RK состоит из донорского сайта сплайсинга интрона промотора CMV и акцепторного сайта сплайсинга вариабельной области тяжелой цепи IgG мыши (смотри, например, Eaton et al., 1986, Biochemistry 25, 8343-8347, Neuberger et al., 1983, EMBO J. 2(8), 1373-1378; его можно получать из вектора pRK-5 (BD PharMingen)), и его предпочтительно помещают перед инициирующим кодоном ATG интересующего гена.
Экспрессионный вектор или сочетание векторов по настоящему изобретению можно трансфицировать в клетку-хозяина в кольцевой форме или в линеаризованной форме. Линеаризация экспрессионного вектора перед трансфекцией часто повышает эффективность стабильной трансфекции. Это также позволяет контролировать точку линеаризации, если экспрессионный вектор линеаризуют перед трансфекцией. Соответствующие конструкции для указанного сайта линеаризации описаны, например, в WO 2009/080720. Экспрессионный вектор(ы) также может содержать прокариотическую точку начала репликации.
Экспрессионный вектор или сочетание экспрессионных векторов по настоящему изобретению могут содержать дополнительные элементы, позволяющие комбинировать способ селекции по настоящему изобретению, основанный на использовании мутантного фолатного рецептора, с другими системами селекции, известными в данной области.
Один общепризнанный способ селекции, известный в данной области, основан на использовании проточной цитометрии, в частности, активированной флуоресценцией сортировки клеток (FACS), для отбора экспрессирующих на высоком уровне клеток-хозяев. Способы селекции с использованием проточной цитометрии имеют то преимущество, что большое число клеток можно подвергать быстрому скринингу на желаемый выход характерной экспрессии. В одном способе селекции, который особенно полезен для идентификации продуцирующих на высоком уровне клеточных клонов, часть интересующего полипептида, например, антитела, экспрессируют в виде мембраносвязанного слитого полипептида. Таким образом, часть продукта экспонируется в виде слитого полипептида на клеточной поверхности. Поскольку количество продуцируемого слитого полипептида коррелирует с общим уровнем экспрессии, клетки-хозяева можно отбирать методом проточной цитометрии на основании количества слитого полипептида, экспонированного на клеточной поверхности. Это позволяет быстро отбирать продуцирующие на высоком уровне клетки-хозяева. Систему селекции по настоящему изобретению можно успешно комбинировать с соответствующими способами селекции, основанными на использовании проточной цитометрии. Для эффективной селекции с использованием FACS предпочтительно использовать специальную экспрессионную кассету для экспрессии интересующего полипептида. Таким образом, в одном варианте осуществления полинуклеотид, кодирующий интересующий полипептид, находится в экспрессионной кассете, которая сконструирована так, что часть экспрессируемого интересующего полипептида содержит трансмембранный якорь. Есть несколько путей для достижения этого результата.
В одном варианте осуществления указанная экспрессионная кассета для экспрессии интересующего полипептида содержит по меньшей мере
(i) полинуклеотид, кодирующий интересующий полипептид,
(ii) по меньшей мере один стоп-кодон ниже полинуклеотида, кодирующего интересующий полипептид, и
(iii) другой полинуклеотид ниже стоп-кодона, кодирующий мембранный якорь и/или сигнал для мембранного якоря.
Транскрипция полинуклеотида, кодирующего интересующий полипептид, находящегося в вышеописанной экспрессионной кассете, приводит к получению транскрипта, содержащего в последовательном порядке по меньшей мере
(i) полинуклеотид, трансляция которого приводит к интересующему полипептиду;
(ii) по меньшей мере один стоп-кодон ниже указанного полинуклеотида;
(iii) полинуклеотид ниже указанного стоп-кодона, кодирующий мембранный якорь и/или сигнал для мембранного якоря.
Часть транскрипта транслируется в слитый полипептид, содержащий интересующий полипептид и мембранный якорь, за счет сквозной трансляции через по меньшей мере один стоп-кодон. Такая конструкция экспрессионной кассеты имеет тот эффект, что в процессе сквозной трансляции (стоп-кодон «дает утечку») часть интересующего полипептида продуцируется в виде слитого полипептида, содержащего мембранный якорь. Остальная часть экспрессируется в виде секретируемого интересующего полипептида. Слитый полипептид экспонируется на клеточной поверхности, и клетки, экспонирующие заякоренный в мембране слитый полипептид в больших количествах, могут быть отобраны методом проточной цитометрии, предпочтительно методом FACS, например, с использованием соответствующих методик окрашивания клеточной поверхности. Таким образом, отбираются клетки-хозяева, имеющие высокий уровень экспрессии. Подробности предпочтительных вариантов осуществления этой технологии на основе стоп-кодонов описаны в WO 2005/073375 и WO 2010/022961. Смотри данное описание.
В одном варианте осуществления экспрессионная кассета дополнительно содержит (iv) полинуклеотид, кодирующий репортер, такой как, например, GFP. Указанный полинуклеотид, кодирующий репортер, расположен ниже стоп-кодона. При сквозной трансляции через стоп-кодон получают слитый полипептид, содержащий репортер, что позволяет проводить селекцию методом проточной цитометрии, основанным на характеристиках экспрессируемого репортера, таких как, например, его флуоресценция. Предпочтительно, полинуклеотид, кодирующий репортер, расположен ниже полинуклеотида, кодирующего мембранный якорь.
В альтернативном варианте осуществления указанная экспрессионная кассета содержит по меньшей мере:
(i) полинуклеотид, кодирующий интересующий полипептид,
(ii) интрон, содержащий 5' донорский сайт сплайсинга и 3' акцепторный сайт сплайсинга и содержащий в рамке считывания стоп-кодон трансляции и сигнал полиаденилирования, и
(iii) полинуклеотид ниже указанного интрона, кодирующий мембранный якорь и/или сигнал для мембранного якоря.
Такая конструкция экспрессионной кассеты имеет тот эффект, что в процессе транскрипции и процессинга транскрипта с экспрессионной кассеты получают по меньшей мере две разные зрелые мРНК (мРНК-POI) и (мРНК-POI-ЯКОРЬ). Трансляция мРНК-POI приводит к получению интересующего полипептида. Трансляция мРНК-POI-ЯКОРЬ приводит к получению слитого полипептида, содержащего интересующий полипептид (POI) и мембранный якорь. В результате этот слитый полипептид, опять-таки, экспонируется на клеточной поверхности, и клетки, экспонирующие заякоренный в мембране слитый полипептид в больших количествах, могут быть отобраны методом проточной цитометрии, предпочтительно методом FACS. Таким образом, отбираются клетки-хозяева, имеющие высокий уровень экспрессии. Подробности и предпочтительные варианты осуществления данной технологии на основе интронов описаны в WO 2007/131774. Смотри данное описание. В одном варианте осуществления экспрессионная кассета дополнительно содержит (iv) полинуклеотид, кодирующий репортер, такой как, например, GFP. Указанный полинуклеотид, кодирующий репортер, расположен ниже интрона. Таким образом, получают слитый полипептид, содержащий репортер, что позволяет проводить селекцию методом проточной цитометрии, основанным на характеристиках экспрессируемого репортера, таких как, например, его флуоресценция. Предпочтительно, полинуклеотид, кодирующий репортер, расположен ниже полинуклеотида, кодирующего мембранный якорь. Таким образом, репортер находится внутри клетки-хозяина.
В одном варианте осуществления экспрессионная кассета сконструирована так, что образуется примерно ≤50%, ≤25%, ≤15%, ≤10%, ≤5%, ≤2,5%, ≤1,5%, ≤1% или менее ≤0,5% слитого полипептида. Оставшаяся часть продуцируется в форме секретируемого полипептида, не содержащего мембранный якорь. Мембранный якорь может быть любого типа при условии, что он обеспечивает заякоривание интересующего полипептида в клеточной мембране и, таким образом, делает возможным экспонирование слитого полипептида на клеточной поверхности. Подходящие варианты осуществления включают, но не ограничиваются ими, GPI якорь или трансмембранный якорь. Трансмембранный якорь является предпочтительным для гарантированного прочного связывания слитого полипептида с клеточной поверхностью и избегания отделения слитого белка. Особенно предпочтительно, в частности, при экспрессии антител в качестве интересующего полипептида, использовать трансмембранный якорь иммуноглобулинов. Другие мембранные якоря и предпочтительные варианты осуществления трансмембранного якоря иммуноглобулинов описаны в WO 2007/131774, WO 2005/073375 и WO 2010/022961.
В одном варианте осуществления интересующий полипептид представляет собой молекулу иммуноглобулина, такого как антитело. Полинуклеотид, кодирующий тяжелую цепь молекулы иммуноглобулина, и полинуклеотид, кодирующий легкую цепь молекулы иммуноглобулина, могут содержаться в одной и той же экспрессионной кассете или, предпочтительно, содержатся в отдельных экспрессионных кассетах, как описано выше. При использовании описанной выше конструкции экспрессионной кассеты, в которой часть интересующего полипептида продуцируется в виде заякоренного в мембране слитого полипептида вследствие сквозной трансляции или альтернативного сплайсинга, такую конструкцию экспрессионной кассеты используют для экспрессии тяжелой цепи антитела.
Клетки-хозяева
Второй аспект настоящего изобретения относится к клетке-хозяину, жизнеспособность которой зависит от поглощения фолатов, содержащей по меньшей мере
a) введенный полинуклеотид, кодирующий мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата по сравнению с фолатным рецептором дикого типа, в качестве селективного маркера
и
b) введенный полинуклеотид, кодирующий интересующий полипептид,
при этом указанный интересующий полипептид секретируется из указанной клетки-хозяина.
«Введенный полинуклеотид» означает полинуклеотидную последовательность, которая была введена в клетку-хозяина, например, с использованием рекомбинантных методов, таких как трансфекция. Клетка-хозяин может содержать или не содержать эндогенный полинуклеотид, функционально соответствующий или идентичный введенному полинуклеотиду. Предпочтительно, введение осуществляют при помощи экспрессионного вектора, который содержит экспрессионную кассету, содержащую полинуклеотид, который должен быть введен, например, кодирующий интересующий полипептид или кодирующий мутантный фолатный рецептор. Предпочтительные примеры экспрессионных векторов и сочетания экспрессионных векторов по настоящему изобретению описаны выше в связи с первым аспектом настоящего изобретения. Введение можно осуществлять, например, путем трансфекции соответствующего экспрессионного вектора, который может интегрироваться в геном клетки-хозяина (стабильная трансфекция). Если полинуклеотид не встраивается в геном, он может быть потерян на более поздней стадии, например, когда клетки претерпевают митоз (временная трансфекция). Соответствующие векторы также могут сохраняться в клетке-хозяине без интегрирования в геном, например, за счет эписомальной репликации. Стабильная трансфекция является предпочтительной для создания экспрессирующих на высоком уровне клеточных клонов, которые подходят для продуцирования интересующего полипептида в промышленных масштабах. В данной области известно несколько подходящих методов введения полинуклеотида, такого как экспрессионный вектор, в эукариотические клетки-хозяева. Соответствующие методы включают, но не ограничиваются ими, трансфекцию с помощью фосфата кальция, электропорацию, нуклеофекцию, липофекцию, биолистический и опосредованный полимерами перенос генов и тому подобное. Помимо традиционных методов, основанных на случайной интеграции, для переноса полинуклеотида, который должен быть введен в геном клетки-хозяина, также можно использовать подходы на основе рекомбинации. Поскольку соответствующие методы хорошо известны в данной области, нет необходимости в их подробном описании в настоящем документе. Однако в данной области известны также и другие методы введения полинуклеотида в клетку-хозяина, которые более подробно описаны ниже.
В одном варианте осуществления клетка-хозяин содержит экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов по первому аспекту, подробно описанные выше и в формуле изобретения. Авторы изобретения ссылаются на указанное описание, которое здесь также применимо. Предпочтительно, указанный экспрессионный вектор или сочетание экспрессионных векторов стабильно интегрированы в геном.
Чтобы была возможна селекция с помощью системы по настоящему изобретению, жизнеспособность клетки-хозяина должна зависеть от поглощения фолатов, предпочтительно от поглощения фолиевой кислоты. Подходящие эукариотические клетки можно выбирать из группы, состоящей из клеток млекопитающих, клеток насекомых, клеток дрожжей, клеток растений и клеток грибов. Клетки грибов и клетки растений могут быть прототрофными в отношении фолатов (то есть, такие клетки могут автономно синтезировать собственные фолаты, необходимые для их клеточной жизнеспособности, то есть, клеточного роста и пролиферации). Настоящее изобретение включает такие клетки грибов и растений, которые являются или становятся ауксотрофными в отношении фолатов. Это может происходить, например, вследствие генетической манипуляции, то есть, клетки впоследствии становятся неспособными синтезировать достаточное количество фолатов, необходимых для жизнеспособности клеток. Предпочтительно, клетка-хозяин представляет собой клетку млекопитающего. Все клетки млекопитающих зависят от поглощения фолатов и, соответственно, могут быть использованы для системы селекции, описанной в настоящем документе. В одном варианте осуществления клетку млекопитающего выбирают из группы, состоящей из клетки грызуна, клетки человека и клетки обезьяны. Особенно предпочтительной является клетка грызуна, которую можно выбирать из группы, состоящей из клетки CHO, клетки BHK, клетки NS0, клетки мышиных фибробластов 3T3 и клетки SP2/0. Особенно предпочтительной клеткой грызуна является клетка CHO. Человеческие клетки также можно использовать и можно выбирать из группы, состоящей из клетки HEK293, клетки MCF-7, клетки PerC6, клетки CAP и клетки HeLa. Клетки обезьяны можно выбирать из группы, состоящей клетки COS-1, клетки COS-7 и клетки Vero.
В одном варианте осуществления у клетки-хозяина отсутствует полная активность по меньшей мере одного эндогенного фолатного рецептора. Соответствующие линии клеток можно получать путем селекции/скрининга или методами генетической инженерии, например, для создания нокаутных линий клеток. Таким образом, предложена также клетка-хозяин, в которой у эндогенной однонаправленной функциональной системы транспорта фолата, например, содержащей по меньшей мере один эндогенный фолатный рецептор, отсутствует полная активность, то есть, активность ослаблена. Такое ослабление можно создавать, например, путем любого мутагенеза соответствующей эндогенной системы транспорта фолатов, например, эндогенного фолатного рецептора, например, за счет точечной мутации, разрушения гена и тому подобного. Ослабление может быть частичным или полным. В этом случае клетка-хозяин по настоящему изобретению не содержит эндогенную однонаправленную функциональную систему транспорта фолатов, например, эндогенный фолатный рецептор.
Однако в предпочтительном варианте осуществления клетка-хозяин по настоящему изобретению содержит по меньшей мере одну эндогенную однонаправленную функциональную систему транспорта фолатов в дополнение к мутантному фолатному рецептору, который введен в указанную клетку-хозяина, например, при помощи экспрессионного вектора или сочетания экспрессионных векторов, описанных выше, в частности один или более эндогенных фолатных рецепторов. Таким образом, генетически неизмененные клетки можно использовать для трансфекции экспрессионным вектором или сочетанием экспрессионных векторов по настоящему изобретению. Преимуществом настоящего изобретения является то, что систему селекции, описанную в настоящем документе, можно использовать даже в присутствии такой эндогенной однонаправленной функциональной системы транспорта фолатов, то есть, в тех случаях, когда такая система сохранена. Это является преимуществом, поскольку использование соответствующих клеток-хозяев для последующего продуцирования интересующего полипептида, которое происходит в не селективных условиях относительно фолата, проще осуществлять, если эндогенная система сохранена и, следовательно, функциональна. Как описано выше, предпочтительными являются клетки-хозяева млекопитающих.
Соответственно, также предложена клетка-хозяин, содержащая по меньшей мере одну эндогенную однонаправленную функциональную систему транспорта фолатов, при этом такая эндогенная однонаправленная функциональная система транспорта фолатов предпочтительно содержит по меньшей мере один эндогенный фолатный рецептор. В предпочтительном варианте осуществления эндогенный фолатный рецептор выбирают из группы, состоящей из фолатного рецептора альфа и фолатного рецептора бета.
В одном варианте осуществления клетка-хозяин дополнительно содержит введенный полинуклеотид, кодирующий дополнительный селективный маркер, который участвует в метаболизме фолата. Варианты осуществления описаны выше в связи с первым аспектом изобретения, смотри приведенное выше описание. Предпочтительно, указанный дополнительный селективный маркер представляет собой DHFR. В соответствии с данным вариантом осуществления, например, клетки-хозяева (например, клетки CHO), лишенные гена DHFR (например, в результате направленной делеции в геноме, также называемые клетки-хозяева DHFR- (минус)), можно использовать в качестве реципиентов для совместной трансфекции гена DHFR в качестве селективного маркера. Однако также возможно и предпочтительно использовать клетки-хозяева, эндогенно экспрессирующие DHFR (клетки-хозяева DHFR+ (плюс)), при проведении селекции с использованием DHFR. В этом случае в качестве селективного маркера предпочтительно используют фермент DHFR, который менее чувствителен к MTX, чем эндогенный фермент DHFR, экспрессируемый клеткой-хозяином DHFR+ (плюс).
В одном варианте осуществления метаболизм эндогенных фолатов или аппарат клетки-хозяина генетически не изменяют перед введением полинуклеотидов путем трансфекции.
По меньшей мере один полинуклеотид, кодирующий интересующий полипептид, полинуклеотид, кодирующий мутантный фолатный рецептор, и, необязательно, полинуклеотид, кодирующий дополнительный селективный маркер, участвующий в метаболизме фолата (который предпочтительно представляет собой DHFR), а также, необязательно, другие полинуклеотиды, описанные выше в связи с первым аспектом изобретения, можно стабильно вводить в указанную клетку-хозяина. Стабильное введение, соответственно, трансфекция, имеет преимущества для создания экспрессирующей линии клеток, и в частности, для крупномасштабного производства секретируемого интересующего полипептида, такого как антитело.
Способ получения рекомбинантных клеток-хозяев
Третий аспект относится к способу получения клетки-хозяина в соответствии со вторым аспектом изобретения, включающему введение в клетку-хозяина, жизнеспособность которой зависит от поглощения фолатов, по меньшей мере
a) полинуклеотида, кодирующего мутантный фолатный рецептор, имеющий сниженную аффинность связывания фолата по сравнению с фолатным рецептором дикого типа, в качестве селективного маркера
и
b) по меньшей мере одного полинуклеотида, кодирующего интересующий полипептид, при этом интересующий полипептид секретируется из указанной клетки-хозяина.
Несколько подходящих методов известно в данной области для введения полинуклеотидов и экспрессионных векторов в клетки-хозяева, в том числе, эукариотические клетки-хозяева, такие как клетки-хозяева млекопитающих. Соответствующие методы хорошо известны в данной области и также описаны выше. Помимо традиционных методов, основанных на случайной интеграции, также можно использовать подходы на основе рекомбинации для переноса полинуклеотида, кодирующего интересующий полипептид, полинуклеотида, кодирующего мутантный фолатный рецептор, и, необязательно, полинуклеотида, кодирующего дополнительный селективный маркер (и/или другие полинуклеотиды), в геном клетки-хозяина. Такие методы рекомбинации могут включать использование сайт-специфических рекомбиназ, таких как Cre, Flp или ΦC31 (смотри, например, Oumard et al., Cytotechnology (2006) 50: 93-108), которые могут опосредовать прямую вставку трансгенов. Альтернативно, можно использовать механизм гомологичной рекомбинации для вставки указанных полинуклеотидов (обзор в Sorrell et al., Biotechnology Advances 23 (2005) 431-469). Опосредованная рекомбинацией вставка генов позволяет сводить к минимуму число элементов, включаемых в гетерологичную нуклеиновую кислоту, которую переносят/вводят в клетку-хозяина. Например, можно использовать локус вставки, который уже имеет промотор и сайт поли-A (экзогенный или эндогенный), так что только оставшиеся элементы необходимо переносить/трансфицировать в клетку-хозяина. Подробности относительно интересующего полипептида, мутантного фолатного рецептора и одного или более селективных маркеров (при использовании), а также их сочетаний приведены выше; авторы изобретения ссылаются на приведенное выше описание. В одном варианте осуществления экспрессионный вектор или сочетание экспрессионных векторов по первому аспекту изобретения вводят в клетку-хозяина. Экспрессионный вектор и сочетание экспрессионных векторов подробно описаны выше и в формуле изобретения. Смотри соответствующее описание. Кроме того, соответствующие примеры клеток-хозяев, жизнеспособность которых зависит от поглощения фолатов, также приведены выше; смотри соответствующее описание.
Способ селекции
Четвертый аспект настоящего изобретения относится к способу селекции по меньшей мере одной клетки-хозяина, способной экспрессировать рекомбинантный интересующий полипептид с высоким выходом, включающему
a) получение множества клеток-хозяев в соответствии со вторым аспектом настоящего изобретении;
b) культивирование указанного множества клеток-хозяев в селективной культуральной среде, содержащей ограниченную концентрацию фолата;
и
c) получение по меньшей мере одной клетки-хозяина, экспрессирующей интересующий полипептид.
Используемый в настоящем документе термин «отбор» или «селекция», в частности, означает процесс использования селективного маркера и селективных условий культивирования для отбора и, соответственно, получения клеток-хозяев, в которые включены полинуклеотиды, предназначенные для введения, такие как экспрессионный вектор или сочетание векторов по настоящему изобретению. Успешно трансфицированные клетки-хозяева можно получать, например, путем выделения и/или обогащения из популяции трансфицированных клеток-хозяев. Успешно трансфицированные клетки-хозяева способны выживать в селективных условиях и экспрессировать интересующий полипептид. Способ селекции является ex vivo способом.
Используемый в настоящем документе термин «ограниченная концентрация фолата», в частности, означает концентрацию фолата(ов) в селективной культуральной среде, которая создает давление отбора на клетку-хозяина. Соответственно, фолаты не содержатся в селективной культуральной среде в достатке, и это ограничение фолата(ов) в культуральной среде создает давление отбора на клетки-хозяева. В таких селективных условиях в основном только те клетки-хозяева растут и/или пролиферируют, в которые включен фолатный рецептор в качестве селективного маркера. Клетки-хозяева, в которые не были успешно включены полинуклеотиды, предназначенные для введения, такие как экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов, и, следовательно, которые не экспрессируют мутантный фолатный рецептор в качестве селективного маркера или в которых экспрессия осуществляется на низком уровне, не способны пролиферировать, расти и/или погибают в селективных условиях культивирования с ограниченной концентрацией фолата. Напротив, клетки-хозяева, в которые был успешно включен экспрессионный вектор или сочетание векторов по настоящему изобретению, и которые экспрессируют мутантный фолатный рецептор в качестве селективного маркера (и соответственно экспрессируют совместно введенный интересующий полипептид) с достаточным выходом, являются устойчивыми или в меньшей степени подвержены давлению отбора и, следовательно, могут в процессе селекции перерастать клетки-хозяева, которые не были успешно трансфицированы или в которых сайт интеграции в геном клетки не благоприятен в случае стабильной трансфекции.
Фолат, содержащийся в селективной культуральной среде в ограниченной концентрации, может поглощаться и процессироваться клеткой-хозяином, в частности, клетками-хозяевами, в которые был введен мутантный фолатный рецептор, используемый качестве селективного маркера. Фолаты и, в частности, производные фолатов, которые не будут или не могут быть процессированы клеткой-хозяином, не вносят вклад в давление отбора, которое оказывается с целью селекции клеток-хозяев, в которые включен фолатный рецептор в качестве селективного маркера, и соответственно, не вносят вклад в ограниченную концентрацию фолата. Однако соответствующие фолаты, такие как, например, антифолаты, могут присутствовать и даже предпочтительно присутствуют, если, например, проводят комбинированную селекцию с DHFR в качестве дополнительного селективного маркера, как описано в настоящем документе. Фолат, присутствующий в селективной культуральной среде в ограниченной концентрации, может, например, быть окисленным фолатом или восстановленным фолатом, либо их производным. Окисленные фолаты, такие как фолиевая кислота, а также восстановленные производные фолиевой кислоты, известные как восстановленные фолаты или тетрагидрофолаты (THF), представляют собой группу витаминов B9, которые являются необходимыми кофакторами и/или коферментами для биосинтеза пуринов, тимидилата и некоторых аминокислот в клетках млекопитающих. Примеры восстановленных фолатов включают 5-метилтетрагидрофолиевую кислоту, 5-формилтетрагидрофолиевую кислоту, 10-формилтетрагидрофолиевую кислоту и 5,10-метилентетрагидрофолиевую кислоту. Как правило, фолат является полезным при условии, что такой фолат будет способен поглощаться и процессироваться клеткой-хозяином для поддержания роста и пролиферации. Предпочтительно, фолат, который содержится в ограниченной концентрации в селективной культуральной среде, представляет собой фолиевую кислоту. Подходящие диапазоны концентраций для создания ограниченной концентрации фолата, описаны ниже.
В процессе селекции клетки-хозяева, в которые был успешно включен экспрессионный вектор(ы) по настоящему изобретению, можно обогащать в виде пула из популяции трансфицированных клеток-хозяев. Затем такой пул можно, например, анализировать для идентификации присутствия клеток-хозяев, которые экспрессируют интересующий полипептид и, например, имеют особенно хорошие уровни экспрессии, ростовые характеристики и/или стабильность. Также отдельные клетки-хозяева можно выделять в виде отдельных клонов из популяции трансфицированных и отобранных клеток-хозяев (например, путем клональной селекции или селекции с помощью FACS). Соответствующие варианты осуществления процедур селекции для получения успешно трансфицированных одиночных клонов из популяции выживающих клеток-хозяев, полученной после селекции (например, методом FACS сортировки или методом серийных разведений), хорошо известны в данной области и, соответственно, не нуждаются в подробном описании.
Подходящие и предпочтительные варианты осуществления клеток-хозяев, мутантного селективного маркера, дополнительных селективных маркеров и сочетаний маркеров, экспрессионных векторов и сочетаний векторов подробно описаны выше, смотри вышеприведенное описание.
Как описано, способ селекции по настоящему изобретению основан на ограниченной доступности фолата, предпочтительно фолиевой кислоты, в среде для культивирования клеток. Система имеет широкое применение, и в частности, может быть использована для селекции эукариотических клеток, жизнеспособность которых зависит от поглощения фолатов, в частности, фолиевой кислоты, таких как, в частности, клетки млекопитающих. Примеры клеток млекопитающих приведены выше. Эта селекция на основе фолата в сочетании с использованием мутантного фолатного рецептора в качестве селективного маркера является отличной стратегией, которая хорошо подходит для ускоренной, стабильной и сверхизбыточной экспрессии полипептидов в культивируемых клетках млекопитающих. Как показано в примерах, способ по настоящему изобретению, в котором мутантный фолатный рецептор используют в качестве селективного маркера, способствует ускоренной селекции, скринингу и созданию клеток-хозяев, в частности, клеток-хозяев млекопитающих, которые избыточно экспрессируют большие количества рекомбинантных продуктов, таких как антитела. Полученные результаты превосходят результаты использования фолатного рецептора дикого типа в качестве селективного маркера.
Система селекции по настоящему изобретению, как описано выше, не требует делеции в геноме или ослабления гена(ов) эндогенного фолатного рецептора перед трансфекцией и, таким образом, может быть применена к любой реципиентной клетке, даже при наличии экспрессии гена эндогенного фолатного рецептора. Это ключевое преимущество основано на том факте, что после трансфекции мутантного фолатного рецептора в качестве селективного маркера, клетки могут быть подвергнуты резкой и сильной депривации фолатов (например, фолиевой кислоты) из ростовой среды. В настоящем изобретении при использовании мутантного фолатного рецептора, имеющего сниженную аффинность связывания фолата, даже более низкие концентрации фолата можно использовать в селективной культуральной среде по сравнению с системой селекции, в которой используют фолатный рецептор дикого типа. Только клетки-трансфектанты, экспрессирующие значительные количества мутантного фолатного рецептора в качестве селективного маркера, способны переносить достаточно фолата в клетку-хозяина для поддержания репликации ДНК и клеточной пролиферации. Это происходит даже в отсутствие какого-либо значительного повышения экспрессии гена эндогенного фолатного рецептора альфа во время цикла селекции. Более того, система селекции по настоящему изобретению, судя по всему, не страдает от потери строгости селекции вследствие ослабления давления отбора из-за повышенной экспрессии альтернативных путей поглощения фолатов, включая повышенную экспрессию эндогенных RFC. Это важное преимущество является результатом того факта, что в то время как фолатный рецептор альфа имеет выдающуюся аффинность для фолиевой кислоты (Kd = 0,1 нМ), RFC демонстрирует чрезвычайно низкую аффинность для фолиевой кислоты (Km = 0,2-0,4 мМ).
Клетки, полученные в результате строгой процедуры скрининга/селекции по настоящему изобретению, можно выделять и обогащать из неотобранных клеток исходной клеточной популяции. Их можно выделять и культивировать как отдельные клетки или пулы клеток. Полученные клетки-хозяева также можно использовать в одном или более дополнительных раундах селекции, необязательно, для дополнительного качественного или количественного анализа, или можно использовать, например, в создании клональной линии клеток для производства белка. В одном варианте осуществления обогащенную популяцию продуцирующих клеток-хозяев, выбранных, как описано выше, непосредственно используют в качестве популяции для продуцирования интересующего полипептида с хорошим выходом.
Предпочтительно, выбирают клетку-хозяина, которая стабильно экспрессирует и, таким образом, секретирует интересующий полипептид. Преимущества стабильной трансфекции/экспрессии подробно описаны выше. Авторы изобретения ссылаются на приведенное выше описание. Предпочтительно, клональную линию клеток создают из выбранной клетки-хозяина, экспрессирующей интересующий белок с желаемым высоким выходом.
Селективная культуральная среда, которую используют в по меньшей мере одном этапе селекции b), может содержать один или более видов фолатов. Фолат, содержащийся в селективной культуральной среде в ограниченной концентрации, может поглощаться и процессироваться трансфицированными клетками-хозяевами, позволяя выживать и, предпочтительно, позволяя поддерживать рост и пролиферацию клеток. Селективная культуральная среда, используемая на этапе b), может иметь одну или более из следующих характеристик:
(a) она содержит ограниченную концентрацию фолата, при этом указанный фолат предпочтительно представляет собой фолиевую кислоту, концентрацию выбирают из концентрации примерно 2000 нМ или менее, примерно 1750 нМ или менее, примерно 1500 нМ или менее, примерно 1000 нМ или менее, примерно 500 нМ или менее, примерно 350 нМ или менее, примерно 300 нМ или менее, примерно 250 нМ или менее, примерно 150 нМ или менее, примерно 100 нМ или менее, примерно 75 нМ или менее, примерно 50 нМ или менее, примерно 40 нМ или менее, примерно 35 нМ или менее, примерно 30 нМ или менее, примерно 25 нМ или менее, примерно 20 нМ или менее, примерно 15 нМ или менее, примерно 10 нМ или менее, примерно 5 нМ или менее и примерно 2,5 нМ или менее; и/или
(b) она содержит фолиевую кислоту в концентрации, выбранной из концентрации примерно 2000 нМ или менее, примерно 1750 нМ или менее, примерно 1500 нМ или менее, примерно 1000 нМ или менее, примерно 500 нМ или менее, примерно 100 нМ или менее, примерно 75 нМ или менее, примерно 50 нМ или менее, примерно 40 нМ или менее, примерно 35 нМ или менее, примерно 30 нМ или менее, примерно 25 нМ или менее, примерно 20 нМ или менее, примерно 15 нМ или менее, примерно 10 нМ или менее, примерно 5 нМ или менее и примерно 2,5 нМ или менее.
Предпочтительные концентрации фолата, и в частности, фолиевой кислоты, в селективной культуральной среде можно выбирать из:
(a) примерно 2000 нМ - 0,1 нМ;
(b) примерно 1750 нМ - 0,1 нМ;
(c) примерно 1500 нМ - 0,1 нМ;
(d) примерно 1250 нМ - 0,1 нМ;
(e) примерно 1000 нМ- 0,1 нМ;
(f) примерно 750 нМ - 0,1 нМ;
(г) примерно 500 нМ - 0,1 нМ;
(h) примерно 250 нМ - 0,1 нМ; предпочтительно примерно 250 нМ - 1 нМ или примерно 250 нМ - 2,5 нМ;
(i) примерно 150 нМ - 0,1 нМ; предпочтительно примерно 150 нМ - 1 нМ или примерно 150 нМ - 2,5 нМ;
(j) примерно 100 нМ - 0,5 нМ; предпочтительно примерно 100 нМ - 1 нМ или примерно 100 нМ - 2,5 нМ;
(k) примерно 75 нМ - 0,5 нМ, предпочтительно примерно 75 нМ - 1 нМ или примерно 75 нМ - 2,5 нМ;
(l) примерно 50 нМ - 1 нМ; предпочтительно примерно 50 нМ - 2,5 нМ или примерно 50 нМ - 5 нМ;
(m) примерно 35 нМ - 0,5 нМ; и
(n) примерно 25 нМ - 1 нМ или примерно 25 нМ - 2,5 нМ, примерно 20 нМ - 3 нМ, примерно 15 нМ - 4 нМ или 10 нМ - 5 нМ.
В одном варианте осуществления фолиевая кислота является единственным фолатом, содержащимся в селективной культуральной среде, который вносит вклад в ограниченную концентрацию фолата.
Концентрации и диапазоны концентраций, описанные выше, особенно подходят для быстрорастущих суспензионных клеток, таких как клетки CHO, которые являются предпочтительным фенотипом для коммерческих линий клеток-продуцентов. Фолат, содержащийся в селективной культуральной среде, предпочтительно представляет собой фолиевую кислоту. Однако разные линии клеток могут иметь разные способности поглощения фолиевой кислоты. Впрочем, подходящие концентрации специалист может с легкостью определить экспериментально. Как показано в примерах, использование мутантного фолатного рецептора в качестве селективного маркера позволяет использовать более низкие концентрации фолата в селективной культуральной среде.
В одном варианте осуществления клетки-хозяева предварительно культивируют в не содержащей фолат культуральной среде или в культуральной среде, содержащей ограниченную концентрацию фолата, перед этапом b) трансфекции и/или селекции. Тем самым клетки вынуждают использовать все их внутренние запасы фолата. Соответствующие ограниченные концентрации фолата описаны выше. Предпочтительно, указанная культуральная среда для предварительного культивирования клеток-хозяев содержит фолат, в частности, фолиевую кислоту, в концентрации 100 нМ или менее, 75 нМ или менее, 50 нМ или менее, предпочтительно 25 нМ или менее, более предпочтительно 15 нМ или менее, наиболее предпочтительно 10 нМ или менее, или даже может совсем не содержат фолат. В одном варианте осуществления банк клеток, например, маточный банк клеток или рабочий банк клеток, создают из таких клеток-хозяев, предварительно культивированных при ограниченных концентрациях фолата, например, фолиевой кислоты. Преимуществом этого является более короткое время подготовки для трансфекции и создания линии клеток.
В предпочтительном варианте осуществления мутантный фолатный рецептор, используемый в качестве селективного маркера в соответствии с идеями настоящего изобретения, используют в сочетании с дополнительным селективным маркером, описанным выше. Как обсуждалось выше, указанный дополнительный селективный маркер предпочтительно участвует в метаболизме фолата и предпочтительно представляет собой DHFR. В одном варианте осуществления, в котором клетки дополнительно трансфицированы другим селективным маркером, селективная культуральная среда, которую используют на этапе b), содержит по меньшей мере один соответствующий ингибитор для указанного дополнительного селективного маркера. Используемая концентрация указанного ингибитора в селективной культуральной среде (которую также можно повышать постепенно), вносит свой вклад в строгость условий селекции. Кроме того, для поддержания давления отбора культуральная среда не должна содержать достаточные количества метаболитов, которые позволили бы обходить активность дополнительного селективного маркера. Например, если DHFR используют в качестве дополнительного селективного маркера, участвующего в метаболизме фолата, предпочтительно, если селективная культуральная среда не будет содержать соответствующие нуклеотиды. Как правило, метаболиты или другие добавки, препятствующие выбранной стратегии селекции, должны находиться под контролем, например, отсутствовать в селективной среде.
Селективные условия для мутантного фолатного рецептора (ограниченная концентрация фолата) и для дополнительного селективного маркера (например, ингибитор DHFR, если DHFR используют в качестве селективного маркера) можно применять одновременно на этапе b), используя соответствующую селективную культуральную среду. Это повышает давление отбора и способствует более эффективной процедуре селекции, тем самым сокращая время для получения подходящей линии клеток, экспрессирующей интересующий полипептид с высоким выходом. Для сочетания селективных маркеров мутантного фолатного рецептора/DHFR на этапе b) предпочтительно используют селективную культуральную среду, содержащую ограниченную концентрацию фолата (соответствующие концентрации и примеры фолата описаны выше) и, кроме того, содержащую ингибитор DHFR, такой как антифолат. Ингибитор DHFR, в частности, представляет собой соединение, которое ингибирует активность дигидрофолатредуктазы (DHFR). Соответствующий ингибитор может, например, конкурировать с субстратом DHFR за связывание с DHFR. Подходящими ингибиторами DHFR являются, например, антифолаты, такие как метотрексат (MTX). Дополнительные примеры включают, но не ограничиваются ими, триметрексат глюкуронат (нейтрексин), триметоприм, пириметамин и пеметрексед. Таким образом, в одном варианте осуществления селективная культуральная среда, используемая на этапе b), дополнительно содержит по меньшей мере один ингибитор DHFR, предпочтительно такой как антифолат, например, MTX.
Таким образом, в одном варианте осуществления клетки-хозяева, получаемые на этапе a), дополнительно содержат введенный полинуклеотид, кодирующий селективный маркер, который представляет собой DHFR, и на этапе b) используют селективную культуральную среду, содержащую антифолат в концентрации 1500 нМ или менее, 1250 нМ или менее, 1000 нМ или менее, 750 нМ или менее, 500 нМ или менее, 250 нМ или менее, 200 нМ или менее, 150 нМ или менее, 125 нМ или менее, 100 нМ или менее, или 75 нМ или менее. В одном варианте осуществления селективная культуральная среда содержит MTX в качестве антифолата. Предпочтительно, селективная культуральная среда содержит MTX в концентрации примерно 350 нМ или менее, 200 нМ или менее, предпочтительно примерно 150 нМ или менее, 125 нМ или менее, 100 нМ или менее, 75 нМ или менее, или 50 нМ или менее. Как показано в примерах, особенным преимуществом является то, что в способе по настоящему изобретению можно использовать очень низкие концентрации MTX. Предпочтительные концентрации антифолата, и в частности MTX, можно выбирать из:
(a) примерно 500 нМ - 1 нМ;
(b) примерно 350 нМ - 2,5 нМ;
(c) примерно 200 нМ - 5 нМ;
(d) примерно 150 нМ - 7,5 нМ;
(e) примерно 100 нМ - 10 нМ; и
(f) примерно 75 нМ - 10 нМ.
Предпочтительные концентрации и диапазоны концентраций для фолата и антифолата, описанные выше, можно использовать в сочетании друг с другом. В одном варианте осуществления концентрацию фолата примерно 0,1 нМ - 100 нМ, предпочтительно 1 нМ - 75 нМ, более предпочтительно 5 нМ - 50 нМ используют в сочетании с концентрацией антифолата 2,5 нМ - 150 нМ, предпочтительно 5 нМ - 125 нМ, более предпочтительно 7,5 нМ - 100 нМ, более предпочтительно 10 нМ - 50 нМ в селективной культуральной среде. Как описано, предпочтительно используют фолиевую кислоту в качестве фолата и MTX в качестве антифолата.
Кроме того, также установлено, что используемые концентрации фолата и антифолата могут влиять друг на друга. Таким образом, помимо абсолютной концентрации фолатов и антифолатов, также и их соотношение может быть фактором для создания подходящих селективных условий. Концентрация антифолатов (предпочтительно MTX) может вплоть до примерно 20 раз превышать концентрацию фолата (предпочтительно фолиевой кислоты). Концентрация антифолата (предпочтительно MTX) может примерно в 10 раз превышать концентрацию фолата (предпочтительно фолиевой кислоты). Предпочтительно, селективная культуральная среда содержит фолат и антифолат в соотношении концентраций от 1:10 до 10:1, предпочтительно в соотношении концентраций от 1:5 до 5:1. Очень хорошие результаты получают при использовании примерно эквимолярных концентраций фолата и антифолата. Как показано в примерах, эти соотношения обеспечивают очень подходящие условия культивирования для получения продуцирующих на высоком уровне клеток-хозяев при использовании желаемого сочетания селективных маркеров.
Такой вариант осуществления настоящего изобретения, в котором мутантный фолатный рецептор используют в сочетании с селективным маркером, участвующим в метаболизме фолата, предпочтительно DHFR, для селекции имеет то преимущество, что продуктивность популяции клеток, переживших селекцию, заметно возрастает. В частности, средняя продуктивность заметно возрастает, как показано в примерах, если этот принцип использовать в сочетании с мутантным фолатным рецептором по настоящему изобретению. В примерах показано, что клетки-хозяева, полученные после применения способа селекции, продуцируют интересующий полипептид с особенно высоким выходом. Таким образом, повышаются шансы обнаружить клоны-суперпродуценты, затрачивая меньше усилий на скрининг. Таким образом, система селекции по настоящему изобретению превосходит системы селекции предшествующего уровня техники.
Более того, установлено, что степень продуктивности можно еще больше увеличивать, если этап b) селекции выполнять по меньшей мере дважды и при этом между всеми этапами b) селекции трансфицированные клетки культивировать в культуральной среде, содержащей не ограниченные или по меньшей мере менее ограниченные концентрации фолата и, предпочтительно, не содержащей ингибитор DHFR, и, следовательно, например, без антифолата. Таким образом, между всеми этапами b) селекции предпочтительно культивировать клетки в не селективных условиях. Установлено, что соответствующая повторяющаяся селекция, при которой клеткам дают возможность восстанавливаться между этапами селекции или циклами селекции, позволяет получать клетки-хозяева, экспрессирующие интересующий белок с особенно высоким выходом, и, более того, число суперпродуцентов значительно возрастает.
Как описано выше, также можно использовать один или более дополнительных селективных маркеров в дополнение к мутантному фолатному рецептору и в дополнение к селективному маркеру, участвующему в метаболизме фолата. Селективные условия для такого дополнительного селективного маркера можно применять до (например, на этапе предварительной селекции, которую проводят между этапами a) и b)) или одновременно с применением на этапе b) селективных условий для мутантного фолатного рецептора и, необязательно, селективного маркера, участвующего в метаболизме фолата. Например, в случае, если ген неомицин фосфотрансферазы (neo) используют в качестве дополнительного селективного маркера, клетки можно сначала выращивать в среде, например, содержащей G418, для предварительной селекции клеток, содержащих экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов по настоящему изобретению. Затем экспрессирующие на высоком уровне клетки отбирают из указанной предварительно отобранной клеточной популяции с использованием селекции на основе мутантного фолатного рецептора, в предпочтительном варианте осуществления в сочетании с селекцией на основе DHFR.
Кроме того, как описано выше, способ селекции по настоящему изобретению можно комбинировать со способами селекции на основе проточной цитометрии, известными в данной области. Таким образом, в одном варианте осуществления этап селекции методом проточной цитометрии выполняют после того, как клетки-хозяева были отобраны в соответствии со способом по настоящему изобретению и, следовательно, после этапа c). Это можно осуществлять для селекции из выжившей популяции клеток-хозяев, экспрессирующих интересующий полипептид с высоким выходом. Такой подход делает этапы клонирования вручную (например, серийные разведения) устаревшими. С этой целью предпочтительно по меньшей мере часть интересующего полипептида экспрессируют в виде заякоренного в мембране слитого полипептида, который экспонируется на клеточной поверхности клетки-хозяина. На основании количества экспонированного слитого полипептида можно отбирать клетки-хозяева с использованием проточной цитометрии, предпочтительно с использованием FACS, которые экспрессируют интересующий полипептид с высоким выходом. Подходящие экспрессионные кассеты для экспрессии полинуклеотида, кодирующего интересующий полипептид, которые делают возможной соответствующую селекцию, описаны выше. Смотри соответствующее описание. Для селекции клетки-хозяева культивируют, чтобы дать возможность экспрессироваться интересующему полипептиду таким образом, что по меньшей мере часть интересующего полипептида экспрессируется в виде слитого полипептида, содержащего мембранный якорь, при этом указанный слитый полипептид экспонируется на поверхности указанной клетки-хозяина и при этом по меньшей мере одну клетку-хозяина отбирают на основании количества слитого полипептида, экспонированного на клеточной поверхности. В этой ситуации можно использовать меченое детектируемое соединение, которое связывается с внеклеточной частью слитого белка. Например, можно использовать флуоресцентно меченое детектируемое соединение. Альтернативно, слитый белок может дополнительно содержать репортер, такой как GFP, который маркирует клетку, тем самым позволяя проводить прямую селекцию на основе характеристик репортера. Предпочтительно, репортер расположен ниже по ходу транскрипции от трансмембранного якоря, и таким образом, находится внутри клетки. Как обсуждалось выше, клетки-хозяева можно отбирать на основании выхода экспрессии, используя метод проточной цитометрии, в частности, FACS.
Способ получения интересующего полипептида
Третий аспект изобретения относится к способу получения рекомбинантного интересующего полипептида, включающему этап культивирования клетки-хозяина по настоящему изобретению и/или клетки-хозяина, выбранной в соответствии с идеями настоящего изобретения, в условиях, допускающих экспрессию и секрецию интересующего полипептида. Использование клеток-хозяев по настоящему изобретению для получения интересующего полипептида имеет то преимущество, что интересующий полипептид может быть получен с высоким выходом. Это, в частности, происходит при применении способа селекции по настоящему изобретению для отбора соответствующих клеток-хозяев для экспрессии. Таким образом, в настоящем изобретении предложен усовершенствованный способ получения интересующего полипептида. Подходящие клетки-хозяева описаны выше; авторы изобретения ссылаются на приведенное выше описание.
Полипептид секретируется в культуральную среду и может быть получен из нее. С этой целью в интересующем полипептиде присутствует соответствующий секреторный лидерный пептид. Примеры приведены выше. Таким образом, рекомбинантные полипептиды могут быть продуцированы и получены/выделены эффективно с высоким выходом. В одном варианте осуществления указанные клетки-хозяева культивируют в бессывороточных условиях.
Способ получения интересующего полипептида может включать по меньшей мере один из следующих этапов:
- выделения интересующего полипептида из указанной клеточной культуральной среды; и/или
- обработки выделенного интересующего полипептида.
Интересующий полипептид, полученный в соответствии с настоящим изобретением, также можно подвергать дополнительным этапам обработки, таким как, например, этапы очистки и/или модификации, для получения интересующего полипептида желаемого качества. Например, продукт можно извлекать из питательной среды общепринятыми методами, включая, но не ограничиваясь ими, центрифугирование, фильтрацию, ультрафильтрацию, экстракцию или осаждение. Очистку можно осуществлять различными методами, известными в данной области, включая, но не ограничиваясь ими, хроматографию (например, ионообменную, аффинную, гидрофобную, хроматофокусирование и эксклюзионную), электрофоретические методы (например, препаративное изоэлектрофокусирование), использование разной растворимости (например, осаждение сульфатом аммония) или экстракцию. Выделенный интересующий полипептид можно формулировать в виде фармацевтической композиции.
Примеры интересующего полипептида приведены выше в связи с первым аспектом изобретения, смотри соответствующее описание. Клетка млекопитающего может содержать или не содержать эндогенный полинуклеотид, соответствующий или идентичный полинуклеотиду, кодирующему интересующий полипептид. В одном варианте осуществления клетка млекопитающего не содержит эндогенный ген, соответствующий интересующему полипептиду. Также предложен полипептид, получаемый способом по настоящему изобретению, описанным выше и формуле изобретения. Указанный полипептид может, в частности, представлять собой молекулу иммуноглобулина или ее функциональный фрагмент.
Варианты применения
Шестой аспект настоящего изобретения относится к применению полинуклеотида, кодирующего
a) мутантный фолатный рецептор, имеющий или содержащий следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9)
где Xaa не является аланином, и при этом аффинность связывания фолата у мутантного фолатного рецептора снижена по сравнению с соответствующим фолатным рецептором дикого типа, где Xaa является аланином (SEQ ID NO 1),
или
b) мутантный фолатный рецептор, содержащий аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9, при этом Xaa не является аланином в указанном мутантном фолатном рецепторе и при этом аффинность связывания фолата у указанного мутантного фолатного рецептора снижена по сравнению с последовательностью фолатного рецептора альфа дикого типа, где Xaa является аланином (смотри SEQ ID NO 1),
в качестве селективного маркера. Указанный селективный маркер можно использовать для селекции успешно трансфицированных клеток-хозяев, жизнеспособность которых зависит от поглощения фолатов, таких как, в частности, клетки млекопитающих. В частности, его можно использовать в качестве селективного маркера для идентификации клеток-хозяев, экспрессирующих рекомбинантный интересующий полипептид с высоким выходом. Предпочтительно, указанный мутантный фолатный рецептор содержится в экспрессионном векторе. Подробности, сочетания и преимущества использования соответствующего мутантного фолатного рецептора в качестве селективного маркера и подходящие экспрессионные векторы описаны выше, смотри приведенное выше описание. Особенно предпочтительным является применение способов по настоящему изобретению. Как описано выше, Xaa предпочтительно представляет собой аминокислоту, выбранную из лейцина, глицина, валина, изолейцина, гистидина и аспарагиновой кислоты. Наиболее предпочтительно, Xaa является лейцином. Предпочтительно, мутантный фолатный рецептор заякорен с помощью GPI. В одном варианте осуществления указанный селективный маркер используют в сочетании с DHFR в качестве дополнительного селективного маркера. Подробности данного варианта осуществления и соответствующие селективные условия описаны выше, смотри приведенное выше описание.
Седьмой аспект настоящего изобретения относится к применению полинуклеотида, кодирующего
a) мутантный фолатный рецептор, содержащий следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9)
или
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYAAAMSGAGPWAAWPFLLSLALMLLWLLS (SEQ ID NO 13)
где Xaa является лейцином;
или
b) мутантный фолатный рецептор, содержащий аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9 или SEQ ID NO 13, и при этом Xaa является лейцином в указанном мутантном фолатном рецепторе по пункту b),
в качестве селективного маркера. Указанный селективный маркер можно использовать для селекции клеток, жизнеспособность которых зависит от поглощения фолатов, таких как, в частности, клетки млекопитающих. В частности, его можно использовать в качестве селективного маркера для идентификации клеток-хозяев, экспрессирующих рекомбинантный интересующий полипептид с высоким выходом. Предпочтительно, указанный мутантный фолатный рецептор, используемый в качестве селективного маркера, содержится в экспрессионном векторе. Подробности, сочетания и преимущества использования соответствующего мутантного фолатного рецептора (мутант A49L) в качестве селективного маркера и подходящие и предпочтительные экспрессионные векторы описаны выше и также описаны в примерах. Смотри соответствующее описание. Особенно предпочтительным является применение способов по настоящему изобретению. Предпочтительно, мутантный фолатный рецептор заякорен с помощью GPI. В одном варианте осуществления указанный селективный маркер используют в сочетании с DHFR в качестве дополнительного селективного маркера. Подробности данного варианта осуществления и соответствующие селективные условия описаны выше, смотри приведенное выше описание. Предпочтительные варианты осуществления этого седьмого аспекта, кроме того, описаны далее.
В одном варианте осуществления седьмого аспекта используют экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов, содержащие:
a) полинуклеотид, кодирующий мутантный фолатный рецептор в качестве селективного маркера, при этом
(i) указанный мутантный фолатный рецептор содержит следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9)
или
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYAAAMSGAGPWAAWPFLLSLALMLLWLLS (SEQ ID NO 13)
где Xaa является лейцином;
или
(ii) указанный мутантный фолатный рецептор содержит аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9 или SEQ ID NO 13, и при этом Xaa является лейцином в указанном мутантном фолатном рецепторе по пункту ii)
b) по меньшей мере один полинуклеотид, кодирующий интересующий полипептид.
Предпочтительно, полинуклеотид, кодирующий мутантный фолатный рецептор, и полинуклеотид, кодирующий интересующий полипептид, содержатся в отдельных экспрессионных кассетах. Подробности подходящих и предпочтительных вариантов осуществления экспрессионных кассет и экспрессионных векторов описаны выше, смотри приведенное выше описание. Предпочтительно, интересующий полипептид является секретируемым полипептидом. Подробности приведены выше в связи с первым аспектом изобретения. В одном варианте осуществления экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов дополнительно содержат полинуклеотид, кодирующий селективный маркер, участвующий в метаболизме фолата, предпочтительно дигидрофолатредуктазу. Подходящие и предпочтительные варианты осуществления описаны выше, смотри приведенное выше описание. Указанный селективный маркер, который предпочтительно представляет собой DHFR, предпочтительно содержится в отдельной экспрессионной кассете.
В одном варианте осуществления данного аспекта также предложена клетка-хозяин, жизнеспособность которой зависит от поглощения фолатов, содержащая
a) введенный полинуклеотид, кодирующий мутантный фолатный рецептор, при этом
i) указанный мутантный фолатный рецептор содержит следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9)
или
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYAAAMSGAGPWAAWPFLLSLALMLLWLLS (SEQ ID NO 13)
где Xaa является лейцином;
или
i) указанный мутантный фолатный рецептор содержит аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9 или SEQ ID NO 13, и при этом Xaa является лейцином в указанном мутантном фолатном рецепторе по пункту ii) и
b) по меньшей мере один введенный полинуклеотид, кодирующий интересующий полипептид, при этом интересующий полипептид секретируется из указанной клетки-хозяина.
Предпочтительно, указанная клетка-хозяин содержит экспрессионный вектор или сочетание по меньшей мере двух экспрессионных векторов, как описано выше. В одном варианте осуществления клетка-хозяин представляет собой клетку млекопитающего. Предпочтительно, она представляет собой клетку грызуна, более предпочтительно клетку CHO. В одном варианте осуществления клетка-хозяин млекопитающего экспрессирует эндогенный фолатный рецептор. В одном варианте осуществления клетка-хозяин млекопитающего содержит введенный полинуклеотид, кодирующий селективный маркер, участвующий в метаболизме фолата, который предпочтительно представляет собой дигидрофолатредуктазу. Подходящие и предпочтительные варианты осуществления подробно описаны выше, как и способы получения соответствующей клетки-хозяина. Смотри соответствующее описание.
В одном варианте осуществления данного аспекта также предложен способ селекции по меньшей мере одной клетки-хозяина, способной экспрессировать рекомбинантный интересующий полипептид с желаемым выходом, включающий
a) получение множества клеток-хозяев, жизнеспособность которых зависит от поглощения фолатов, содержащих
aa) введенный полинуклеотид, кодирующий мутантный фолатный рецептор, при этом
i) указанный мутантный фолатный рецептор содержит следующую последовательность
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYA (SEQ ID NO 9)
или
IAWARTELLNVCMNAKHHKEKPGPEDKLHEQCRPWRKNACCSTNTSQEXaaHKDVSYLYRFNWNHCGEMAPACKRHFIQDTCLYECSPNLGPWIQQVDQSWRKERVLNVPLCKEDCEQWWEDCRTSYTCKSNWHKGWNWTSGFNKCAVGAACQPFHFYFPTPTVLCNEIWTHSYKVSNYSRGSGRCIQMWFDPAQGNPNEEVARFYAAAMSGAGPWAAWPFLLSLALMLLWLLS (SEQ ID NO 13)
где Xaa является лейцином;
или
ii) указанный мутантный фолатный рецептор содержит аминокислотную последовательность, имеющую идентичность последовательности по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% с последовательностью, приведенной в SEQ ID NO 9 или SEQ ID NO 13, и при этом Xaa является лейцином в указанном мутантном фолатном рецепторе по пункту ii) и
bb) по меньшей мере один введенный полинуклеотид, кодирующий интересующий полипептид;
b) культивирование указанного множества клеток-хозяев в селективной культуральной среде, содержащей фолат в ограниченной концентрации;
и
c) получение по меньшей мере одной клетки-хозяина, экспрессирующей интересующий полипептид.
Селективная культуральная среда, используемая на этапе b), содержит ограниченную концентрацию фолата, при этом указанный фолат предпочтительно представляет собой фолиевую кислоту; концентрацию выбирают из концентрации примерно 2000 нМ или менее, примерно 1750 нМ или менее, примерно 1500 нМ или менее, примерно 1000 нМ или менее, примерно 500 нМ или менее, примерно 350 нМ или менее, примерно 300 нМ или менее, примерно 250 нМ или менее, примерно 150 нМ или менее, примерно 100 нМ или менее, примерно 75 нМ или менее, примерно 50 нМ или менее, примерно 40 нМ или менее, примерно 35 нМ или менее, примерно 30 нМ или менее, примерно 25 нМ или менее, примерно 20 нМ или менее, примерно 15 нМ или менее, примерно 10 нМ или менее, примерно 7,5 нМ или менее, примерно 5 нМ или менее и примерно 2,5 нМ или менее. Предпочтительно, в качестве фолата используют фолиевую кислоту. Клетка-хозяин предпочтительно представляет собой клетку млекопитающего. В одном варианте осуществления клетка-хозяин дополнительно содержит введенный полинуклеотид, кодирующий селективный маркер, который представляет собой дигидрофолатредуктазу. В этом варианте осуществления селективная культуральная среда, используемая на этапе b), дополнительно содержит, в соответствии с одним вариантом осуществления, антифолат в концентрации, выбранной из концентрации 1500 нМ или менее, 1000 нМ или менее, 750 нМ или менее, 500 нМ или менее, 200 нМ или менее, 150 нМ или менее, 125 нМ или менее, 100 нМ или менее, 75 нМ или менее, 50 нМ или менее, 25 нМ или менее, 20 нМ или менее, 15 нМ или менее, 12 mM или менее и 10 нМ или менее. В одном варианте осуществления после этапа c) клетки культивируют в культуральной среде, содержащей не ограниченную концентрацию фолата, а затем вновь культивируют в соответствии с этапом b) и получают в соответствии с этапом c). Дополнительные подробности относительно предпочтительных и подходящих селективных культуральных сред и вариантов осуществления способа селекции описаны выше в связи с четвертым аспектом изобретения, смотри соответствующее описание.
В следующем варианте осуществления данного аспекта также предложен способ получения интересующего полипептида, включающий
a) культивирование клетки-хозяина, описанной в предшествующих параграфах данного аспекта, и/или клетки-хозяина, выбранной способом, описанным в предшествующих параграфах данного аспекта, в условиях, допускающих экспрессию и секрецию интересующего полипептида;
b) выделение интересующего полипептида из среды для культивирования клеток и
c) необязательно, обработку выделенного интересующего полипептида.
Подробности, относящиеся к соответствующему способу получения, а также подходящие и предпочтительные варианты осуществления интересующего полипептида также описаны выше, смотри приведенное выше описание. Предпочтительно, интересующий полипептид представляет собой терапевтически активный полипептид, такой как антитело.
Данное изобретение не ограничено иллюстративными способами и материалами, раскрытыми в настоящем документе. Числовые диапазоны, описанные в настоящем документе, включают все числа, определяющие диапазон. Заголовки, приведенные в настоящем документе, не ограничивают различные аспекты или варианты осуществления данного изобретения, которые можно понять, обращаясь к описанию в целом. В одном варианте осуществления объект, описанный в настоящем документе как содержащий определенные элементы, также означает объект, состоящий из соответствующих элементов. В частности, полинуклеотиды, описанные в настоящем документе как содержащие определенные последовательности, также могут состоять из соответствующих последовательностей. Предпочтительно выбирать и комбинировать предпочтительные варианты осуществления, описанные в настоящем документе, и конкретный объект, возникающий в результате соответствующей комбинации предпочтительных вариантов осуществления, также входит в объем настоящего изобретения.
Следующие далее примеры служат для иллюстрации настоящего изобретения, никоим образом не ограничивая его объем. В частности, примеры относятся к предпочтительным вариантам осуществления настоящего изобретения.
ПРИМЕРЫ
В описанных далее экспериментах были использованы следующие векторы:
Референсный вектор «V-DHFRреф» содержал следующие основные экспрессионные кассеты: экспрессионную кассету, содержащую полинуклеотид, кодирующий DHFR в качестве селективного маркера; экспрессионную кассету, содержащую полинуклеотид, кодирующий легкую цепь антитела; экспрессионную кассету, содержащую полинуклеотид, кодирующий тяжелую цепь антитела, и экспрессионную кассету, содержащую полинуклеотид, кодирующий неомицин фосфотрансферазу. Все экспрессионные кассеты были ориентированы в одном и том же направлении. С указанного референсного вектора экспрессировалось полное антитело. Конструкция соответствующего вектора также описана в WO 2009/080720.
Векторы, содержащие фолатный рецептор в качестве селективного маркера, были сконструированы на основе референсного вектора путем замены полинуклеотида, кодирующего DHFR в качестве селективного маркера, на полинуклеотид, кодирующий фолатный рецептор в качестве селективного маркера. В остальном экспрессионные кассеты оставались такими же. Вектор «V-дтFRальфа» содержал человеческий фолатный рецептор альфа дикого типа в качестве селективного маркера. Вектор «V-мутFRальфа» содержал мутантный человеческий фолатный рецептор альфа с мутацией A49L.
Пример 1: Одиночные трансфекции
Для одиночных трансфекций человеческий рецептор фолиевой кислоты альфа дикого типа (вектор: V-дтFRальфа) или мутантный человеческий фолатный рецептор альфа (вектор: V-мутFRальфа) были введены в качестве селективных маркеров в клетки CHO. Целью данного эксперимента был анализ функции системы селекции с использованием фолатного рецептора, которая в отличие от системы с использованием DHFR/MTX не основана на токсическом ингибировании роста клеток, а основана на ингибировании роста вследствие депривации фолиевой кислоты в культуральной среде. Фолиевая кислота является окисленной формой витамина B9. Фолиевая кислота является биологически активной в ее восстановленной форме, тетрагидрофолата (THF). Фолиевая кислота восстанавливается в клетке в ее биологически активную форму тетрагидрофолат (THF) через дигидрофолат (DHF) в НАДФН-зависимой реакции, катализируемой ферментом дигидрофолатредуктазой (DHFR). Поглощение фолиевой кислоты необходимо клеткам млекопитающих для поддержания клеточного роста и клеточной пролиферации.
Только клетки, которые интегрируют трансфицированный вектор в геном и экспрессируют либо фолатный рецептор альфа дикого типа (V-дтFRальфа), либо мутантный фолатный рецептор альфа (V-мутFRальфа) с высокой эффективностью, могут выживать в селективных условиях, основанных на ограничении концентрации фолата в культуральной среде. Эти клетки способны включать достаточное количество фолиевой кислоты из культуральной среды в клетки для поддержания пролиферации и роста клеток, даже если культуральная среда содержит ограниченную концентрацию фолиевой кислоты. Поскольку экспрессионные векторы также вводят в клетки полинуклеотиды, кодирующие интересующий белок (в данных экспериментах тяжелую и легкую цепь антитела), можно отбирать стабильные, продуцирующие на высоком уровне линии клеток с использованием технологии селекции, основанной на использовании фолатного рецептора.
Для определения влияния концентрации фолиевой кислоты на рост клеток, тестировали различные селективные культуральные среды. Стандартная культуральная среда содержит 11,7 мкМ фолиевой кислоты (полная среда). Установлено, что наиболее строгие условия селекции достигались при использовании ≤50 нМ фолиевой кислоты в культуральной среде. Кроме того, при использовании различных концентраций фолиевой кислоты наблюдали различия в скоростях роста клеток. Чем меньше было фолиевой кислоты в культуральной среде, тем более медленным был рост клеток.
Во-первых, внутренние запасы фолиевой кислоты в клетках CHO были снижены и сопутствующий перенос фолиевой кислоты из стандартной среды для культивирования (полной среды) в селективную среду был предотвращен. Для этого перед трансфекцией клетки, предназначенные для селекции в условиях ограниченной концентрации фолиевой кислоты, промывали три раза PBS и инокулировали в не содержащую фолиевую кислоту среду. Референсный контроль (вектор V-DHFRреф) пересевали при той же плотности клеток в полной среде. Рост клеток анализировали перед трансфекцией и установили, что культура, растущая в полной среде (5×106 LZ/мл), росла примерно в 2 раза лучше, чем культура, которая росла в селективной среде (2,5×106 LZ/мл).
Векторы трансфицировали в клетки методом нуклеофекции. 5×106 жизнеспособных клеток (LZ/мл) были трансфицированы 3 мкг векторной ДНК. Клетки CHO, содержащие векторы V-дтFRальфа, V-мутFRальфа и отрицательные контроли (V-DHFRреф и без ДНК), были переведены в селективную среду; трансфицированные референсными векторами клетки были переведены в полную среду. Сводные данные по проведенным одиночным трансфекциям приведены в таблице 1. Селекцию начинали через 48 часов после трансфекции, тем самым давая возможность клеткам восстанавливаться после нуклеофекции и начинать экспрессию введенных экспрессионных векторов. Клетки, содержащие селективные маркеры, предназначенные для тестирования, подвергали воздействию ограниченных концентраций фолиевой кислоты. Параллельно, клетки с селективным маркером DHFR (V-DHFRреф) подвергали воздействию различных содержащих MTX культуральных сред, а также различных содержащих фолиевую кислоту (FA) сред. Культура без дополнительной ДНК служила отрицательным контролем.
Таблица 1
Сводные данные по проведенным одиночным трансфекциям
|
Эффективность трансфекции определяли через 48 часов путем контролирования GFP. В следующей далее таблице 2 приведены сводные данные по достигнутой плотности жизнеспособных клеток в день 12 селекции на основе фолиевой кислоты.
Таблица 2
Сводные данные по плотности клеток (LZ/мл) в день 12 селекции на основе фолиевой кислоты (FA)
|
В таблице 2 приведена плотность клеток [LZ/мл] пулов клеток, трансфицированных контролями (V-DHFRреф; без ДНК), V-дтFRальфа и V-мутFRальфа и отобранных с использованием различных концентраций фолиевой кислоты. Как можно видеть, рост уменьшается при снижении концентрации фолиевой кислоты в селективной среде. Пулы клеток, трансфицированных мутантным фолатным рецептором альфа в качестве селективного маркера, в селективной среде, содержащей ограниченные концентрации фолиевой кислоты, демонстрировали клеточный рост, который превышал примерно в два раза или даже более, клеточный рост, наблюдаемый в других пулах клеток. В день 12 клетки, трансфицированные фолатным рецептором альфа дикого типа, не имели ростового преимущества. Они росли примерно так же хорошо, как популяции, содержащие V-DHFRреф, или отрицательные контроли. Однако ростовое преимущество становилось заметно в случае фолатного рецептора альфа дикого типа на более поздней стадии - в зависимости от используемой концентрации фолиевой кислоты - примерно в день 16-20. Таким образом, фолатные рецепторы обоих типов (дикого типа и мутант) подходят для селекции клеток, основанной на ограниченной концентрации фолиевой кислоты в культуральной среде. Однако использование мутантного фолатного рецептора альфа в качестве селективного маркера в соответствии с идеями настоящего изобретения более выгодно, поскольку ростовые преимущества успешно трансфицированных клеток видны раньше, чем в случае фолатного рецептора альфа дикого типа. Более того, мутантный фолатный рецептор делал возможной селекцию при более низких концентрациях фолиевой кислоты, таких как 15 нМ и даже 5 нМ. Таким образом, можно использовать более строгие условия селекции в случае применения мутантного фолатного рецептора по настоящему изобретению в качестве селективного маркера.
При анализе жизнеспособности пулов клеток в день 12 селекции наблюдаемая жизнеспособность была примерно одинаковой при концентрациях фолиевой кислоты 35-50 нМ (жизнеспособность пулов клеток >90%). Вследствие высокой жизнеспособности было возможно пересевать популяции клеток в этот день. Однако при более низких концентрациях фолиевой кислоты жизнеспособность снижалась. В селективных средах с самым низким содержанием фолиевой кислоты только трансфицированная V-мутFRальфа популяция демонстрировала относительно высокую жизнеспособность 76%.
Кроме того, анализировали время, необходимое для селекции. Общее время, необходимое для проведения селекции, важно при разработке нового селективного маркера. При культивировании клеток CHO один этап селекции с использованием DHFR-MTX может быть завершен в зависимости от используемых условий селекции в пределах 15-16 дней. Однако многоэтапная амплификация генов, как правило, занимает гораздо больше времени. На протяжении этого периода времени клетки должны восстанавливаться после кризиса, вызванного давлением отбора. В таблице 3 приведено количество дней селекции до следующего пересева, то есть, период времени, необходимый для достижения клетками жизнеспособности более 90% и для того, чтобы клетки можно было использовать для скрининга титров антитела.
Таблица 3
Продолжительность селекции в днях
|
В таблице 3 показано количество дней селекции до следующего пересева, то есть, период времени, необходимый клеткам для переживания селекционного кризиса и достижения жизнеспособности более 90%. Как можно видеть, вначале все клеточные пулы восстанавливаются при культивировании в селективной среде, содержащей 50, 45 или 35 нМ фолиевой кислоты. Однако более низкие концентрации фолиевой кислоты оказывают высокое давление отбора на клетки так, что только использование V-мутFRальфа допускает хорошее восстановление и, следовательно, жизнеспособность после 16 дней в этих очень строгих условиях. В этой ситуации популяция клеток, трансфицированных V-мутFRальфа, восстанавливалась и демонстрировала жизнеспособность более 90% в селективной среде, содержащей всего лишь от 5 нМ до 25 нМ фолиевой кислоты. Клеткам, трансфицированным V-дтFRальфа, требовалось больше времени и они не могли восстанавливаться при очень низких концентрациях фолиевой кислоты.
Пример 2: Определение продуктивности в отношении антител
Для анализа успешности трансфекции и селекции на основании экспрессии интересующего гена (в данном случае референсного антитела), клетки, полученные, то есть, отобранные в соответствии с примером 1, культивировали как периодические культуры во встряхиваемых колбах в течение 13 дней для определения продуктивности клеток после селекции. Клетки имели жизнеспособность более 90%. В день 13 концентрацию антитела в культуральном супернатанте определяли с использованием аффинной хроматографии на белке A [мг/л]. Результаты приведены в таблице 4.
Таблица 4
Концентрация антитела (мАт) в культуральном супернатанте периодической культуры (мг/л)
|
Антитело, экспрессируемое с введенных экспрессионных векторов, можно было обнаружить во всех клеточных популяциях. Поскольку низкие количества антитела были обнаружены также в пулах клеток, не трансфицированных ДНК, только значения, превышающие 9 мг/л, считали значимыми. Клетки референсной популяции в 125 нМ MTX (стандартная система селекции с DHFR/MTX) продуцировали 28 мг/л. Результаты для четырех популяций клеток, трансфицированных V-дтFRальфа, при высоких концентрациях фолиевой кислоты были в пределах фона. Однако при снижении концентрации фолиевой кислоты в культуральной среде до 15 нМ фолиевой кислоты, экспрессия антитела была примерно такой же высокой (24 мг/л), как в случае референсного контроля V-DHFRреф в 125 нМ MTX (28 мг/л). Это подтверждает предыдущий вывод, что фолатный рецептор дикого типа может служить в качестве селективного маркера и при этом достигается эффективность, сопоставимая с эффективностью общепризнанной системы DHFR/MTX, хотя никакие токсические средства не были использованы для селекции. Пулы клеток, трансфицированных V-мутFRальфа, демонстрировали линейное возрастание титра антитела, которое зависело от концентрации фолиевой кислоты в культуральной среде. Чем ниже была концентрация фолиевой кислоты в культуральной среде, тем выше был достигнутый уровень экспрессии антитела. Таким образом, использование мутантного фолатного рецептора по настоящему изобретению в качестве селективного маркера имело преимущества перед использованием фолатного рецептора дикого типа в качестве селективного маркера.
Поскольку число интегрированных трансгенов оказывает важное влияние на уровень экспрессии, число копий наиболее важных элементов, а именно, легкой и тяжелой цепей (LC, HC) экспрессируемого антитела, а также число копий фолатного рецептора (мутантного или дикого типа) определяли с использованием количественной ПЦР на основе геномной ДНК пулов клеток.
В отношении измеренного титра антитела, число копий может косвенно обеспечить понимание того, является ли место интегрирования в геном причиной сильной или слабой экспрессии. Количественный ПЦР-анализ, проведенный в данном случае, позволяет определять среднее значение числа копий трансгена, поскольку анализируют не одиночные клеточные клоны, а популяцию различных клеток, переживших селекцию.
Таблица 5
Определение числа копий методом количественной ПЦР
|
В таблице 5 приведены результаты количественного ПЦР-анализа V-дтFRальфа и V-мутFRальфа трансфектантов после селекции. Из контрольных пулов анализировали пул клеток, которые выжили при максимальной строгости селекции (V-DHFRреф: 125 нМ MTX, 35 нМ фолиевой кислоты, без ДНК: 25 нМ фолиевой кислоты). Кроме того, не трансфицированные клетки CHO, культивируемые без давления селекции, анализировали в качестве отрицательного контроля. В таблице 5 приведено число копий для легкой цепи и тяжелой цепи, а также число копий для фолатного рецептора, используемого в качестве селективного маркера. Величины относятся к теоретическому размеру генома клеток CHO. В таблице 5 показано, что в не трансфицированных клетках CHO, как и ожидалось, никаких последовательностей антитела невозможно было определить. Более того, были определены только копии эндогенного фолатного рецептора альфа дикого типа. В случае референсного контроля V-DHFRреф при отборе с 125 нМ MTX обнаруживали в среднем двукратное интегрирование трансгенов антитела. При рассмотрении популяции клеток с V-DHFRреф, отобранных с использованием 35 нМ фолиевой кислоты, можно видеть гораздо большую степень интегрирования легкой и тяжелой цепей антитела. Было обнаружено 5,5 копий тяжелой цепи и 6,8 копий легкой цепи. Число копий генов FR альфа было сопоставимо с родительскими клетками CHO и объяснялось эндогенными аллелями.
Пулы клеток, трансфицированных V-дтFRальфа, по сравнению с контролями V-DHFRреф имели лишь несколько копий HC и LC. Никаких различий в зависимости от концентрации не наблюдалось. Число копий для фолатного рецептора возрастало при изменении от 50 нМ до 35 нМ фолиевой кислоты до 2,8 копий, однако затем снижалось при концентрации 25 нМ. Кроме того, в пуле клеток, отобранных с использованием 15 нМ фолиевой кислоты, было включено в среднем 2,6 копий для цепей антитела. Напротив, в пуле клеток, трансфицированных V-мутFRальфа, наблюдали почти линейное возрастание числа копий генов цепей антитела, которое шло параллельно с уменьшением содержания фолиевой кислоты в селективной среде. Таким образом, чем ниже была концентрация фолиевой кислоты в селективной среде, тем большее число копий генов LC и HC обнаруживали в отобранных клетках. Число копий гена мутантного фолатного рецептора альфа демонстрировало сопоставимую тенденцию. Пул клеток, отобранных с использованием 5 нМ фолиевой кислоты, отличался наличием примерно в три раза большего числа копий интегрированных генов антитела, чем в клетках референсного контроля с V-DHFRреф, отобранных с использованием 125 нМ MTX.
Пример 3: Одноклеточное клонирование
Для создания линии клеток, стабильно продуцирующих интересующий ген с высоким выходом, необходимо отбирать из полученной популяции различных продуцирующих клеток, которые переживают селекцию в соответствии с примером 1, лучшие клеточные клоны, которые демонстрируют как высокий уровень экспрессии антитела, так и хороший рост клеток. Для этой цели получали линии клеток из одиночных клеток методом серийных разведений. Серийные разведения позволяют получать моноклональную клеточную популяцию, начиная с поликлональной массы клеток, переживших селекцию в соответствии с примером 1. Это достигается путем проведения серии возрастающих разведений родительской (поликлональной) культуры клеток. Получают суспензию родительских клеток. Затем выполняют соответствующие разведения, в зависимости от количества клеток в исходной популяции. После выполнения последних разведений, одиночную клетку помещают в лунку планшета для культивирования клеток и из нее получают клон. Создание популяции моноклональных клеток гарантирует стабильную экспрессию антитела в течение длительного периода времени. Выбранные популяции клеток с V-дтFRальфа (15 нМ фолиевой кислоты), V-мутFRальфа (5 нМ фолиевой кислоты), V-DHFRреф (125 нМ MTX) и V-DHFRреф (250 нМ MTX - от другой трансфекции) были соответственно клонированы. Клонирование проводили в полной среде и, кроме того, в соответствующей селективной среде, которую использовали ранее для селекции. Таким образом, в последнем случае давление отбора поддерживали во время одноклеточного клонирования. После успешного роста клоны при конфлюэнтности более 70% переносили в 24-луночные планшеты и тестировали в периодическом культивировании (продолжительность 10 дней) на продуцирование ими антитела. В процессе периодического культивирования вновь использовали либо полную среду (без давления отбора), либо селективную среду (поддержание давления отбора). Клоны располагали (в зависимости от среды) в порядке от наивысшего до самого низкого уровня экспрессии. Результаты показаны на фигурах 1-4.
Показаны результаты клонирования, достигнутые либо в полной среде (без поддержания давления отбора после селекции), либо в селективной среде (давление отбора поддерживали после селекции). В данном случае имело место типичное выполнение процедуры клонирования вручную. Было обнаружено 1-5 продуцирующих на высоком уровне клонов клеток, а затем кривая быстро опускалась вниз до экспрессирующих на низком уровне или совсем не экспрессирующих клеточных клонов. Кроме того, среди продуцирующих на низком уровне клеточных клонов наблюдали широкий спектр продуктивности клеток, в котором, однако, большинство находилось ниже порога 9 мг/л.
Клонирование клеток, трансфицированных V-DHFRреф (отобранных с использованием 125 нМ MTX), в полной среде и селективной среде привело к получению 86 клонов (53 в полной среде, 33 в селективной среде) из 6 96-луночных планшетов. Максимально продуцирующий клон культивировали в селективной среде и его продуктивность составила в 24-луночной партии 28,4 мг/л. В 250-мл встряхиваемой колбе (всего 50 мл) исходный поликлональный пул клеток также достигал титра 28 мг/л. Клонирование клеток, трансфицированных V-DHFRреф (отобранных с использованием 250 нМ MTX), в полной среде и селективной среде позволило выделить 76 клонов (41 в полной среде и 35 в селективной среде). Три лучших клона были выделены из клеток, культивируемых в полной среде после селекции, в остальном, клетки, выращенные в селективной среде, демонстрировали более высокую общую продуктивность, чем клоны в полной среде. В начальном поликлональном пуле титр составлял 27 мг/л, у максимально продуцирующего клона он достигал 42 мг/л в 24-луночном планшете. Оба референсных контроля показали, что необязательно наивысшая концентрация MTX, используемая в процессе селекции, приводит к наивысшему титру. Чтобы выделить наилучший клон, необходимо анализировать большое число клеточных клонов.
Клетки, трансфицированные вектором V-дтFRальфа, содержащим фолатный рецептор альфа дикого типа в качестве селективного маркера (отобранные с использованием 15 нМ фолиевой кислоты), также клонировали либо в полной среде (не поддерживая давление отбора после селекции), либо в селективной среде (тем самым поддерживая давление отбора во время клонирования). Два максимально продуцирующих клона были выделены в селективной среде. У наилучшего клона титр достигал 53 мг/л в 24-луночном формате. Примечательно, что только 7 клонов выжили в этой селективной среде. В полной среде выжили 49 клонов. У исходного пула в 250-мл встряхиваемой колбе концентрация антитела составляла примерно 24 мг/л.
Клетки, трансфицированные вектором V-мутFRальфа, содержащим мутантный фолатный рецептор альфа в качестве селективного маркера (отобранные с использованием 5 нМ фолиевой кислоты), также клонировали либо в полной среде (не поддерживая давление отбора после селекции), либо в селективной среде (тем самым поддерживая давление отбора во время клонирования). Два максимально продуцирующих клона были выделены в полной среде, а также в селективной среде. У максимально продуцирующего клона титр составлял 42 мг/л в супернатанте. В общей сложности, сто клонов могли быть перенесены в 24-луночные планшеты, в том числе 52 в полной среде и 48 в селективной среде. В этом случае аналогичные результаты были достигнуты в селективной и в полной среде. В таблице 6 приведены сводные данные по продуктивности наилучших клонов-продуцентов.
Таблица 6
Сводные данные по максимально продуцирующим клонам (мАт [мг/л]) после конечного разведения
|
Из таблицы 6 видно, что селекция с двумя селективными маркерами, фолатным рецептором альфа дикого типа (V-дтFRальфа) и мутантным фолатным рецептором альфа (V-мутFRальфа), позволяла получать после одноклеточного клонирования клеточные клоны, которые достигали в проведенном эксперименте клонирования по меньшей мере таких же хороших результатов, как и при использовании в качестве референсного селективного маркера DHFR. Дальнейшие эксперименты (смотри ниже) показали, что более высокие общие уровни продуктивности могут быть достигнуты при использовании мутантного фолатного рецептора по настоящему изобретению в качестве селективного маркера.
Пример 4: Эксперименты по совместной трансфекции
Для анализа строгости селекции и эффективности давления двойного отбора в виде депривации фолиевой кислоты в селективной среде и добавления MTX клетки были совместно трансфицированы V-DHFRреф и V-мутFRальфа. Все трансфицированные векторы содержали одинаковые гены антитела в качестве интересующего белка. Два отдельных экспрессионных вектора были совместно трансфицированы, при этом каждый вектор содержал экспрессионные кассеты для экспрессии легкой и тяжелой цепи антитела. Перед трансфекцией клетки CHO (за исключением клеток, которые использовали для референсного контроля с DHFR) промывали три раза PBS для удаления фолиевой кислоты, перенесенной из полной среды, и пересевали в селективную среду для трансфекции. Пересев клеток CHO, использованных для трансфекции референсного контроля V-DHFRреф, проводили в полной среде.
Векторы трансфицировали в клетки методом нуклеофекции. В отличие от трансфекций одиночными векторами, для выполнения совместной трансфекции использовали двойное количество клеток (1×107 LZ/мл) и двойное количество ДНК (3 мкг на вектор). Однако в расчете на клетку количество трансфицированной ДНК было тем же самым. Клетки CHO, трансфицированные V-DHFRреф/V-мутFRальфа, и контроли переводили в селективную среду; референсные трансфицированные клетки переводили в полную среду. Селекцию начинали через 48 часов после трансфекции. Клетки, полученные после трех трансфекций на каждые тестируемые условия, объединяли через 48 часов, центрифугировали, ресуспендировали в 9 мл селективной среды или полной среды и разделяли на порции триплетами в три селективные среды. Три партии пулов совместно трансфицированных клеток и контролей подвергали воздействию трех разных концентраций фолиевой кислоты/MTX для селекции. Параллельно с ними выполняли референсный контроль с вектором V-DHFRреф, используя селекцию при помощи G418/MTX. В этом случае клетки культивировали в полной среде, содержащей фолиевую кислоту в достатке. Затем для начала цикла селекции добавляли селективные средства, создавая давление отбора. В качестве отрицательного контроля проводили трансфекцию без добавления ДНК. Данные по проведенным трансфекциям и использованным культуральным средам приведены в следующей далее таблице 7.
Таблица 7
Сводные данные по проведенным совместным трансфекциям
|
После завершения селекции из отобранных клеток готовили периодические культуры для определения уровня экспрессии интегрированных генов антитела. Во время периодического культивирования клеточные популяции культивировали в полной среде. Во всех клеточных популяциях определяли концентрации антитела через 13 дней периодического культивирования для совместной и референсной трансфекции, используя аффинную хроматографию с белком A для определения экспрессии антитела. Результаты приведены в таблице 8, в которой периодические культуры обозначены в соответствии с их происхождением в селективной среде, то есть, они обозначены в соответствии с проведенной селекцией (50/50, 50/100, 12,5/50 [нМ FA/нМ MTX] или V-DHFRреф-G418-MTX - выполнено в трех повторах). Пулы клеток, которые не выжили, не показаны. Опять-таки, вследствие использованного метода измерения только значения выше 9 мг/л считали значимыми.
Таблица 8
Концентрация антитела (мАт) в культуральном супернатанте периодической культуры в [мг/л]
|
Как можно видеть из таблицы 8, в пулах клеток, полученных референсным способом с DHFR после трех этапов селекции (0,8 мг/мл G418 - 500 нМ MTX - 1 мкм MTX), титр антитела составлял до 58 мг/л. После совместной трансфекции с использованием V-мутFRальфа/V-DHFRреф почти все пулы клеток выживали, когда клетки были перенесены после селекции в полную среду. Пулы клеток с V-мутFRальфа/V-DHFRреф, полученные из селективной среды 50/50, продуцировали до 25 мг/л, в пулах клеток 50/100 и 12,5/50 клетки продуцировали с титрами до 36 мг/л.
Использовали три последовательных цикла селекции в референсном способе с DHFR, поскольку за селекцией с G418 следовали два цикла селекции с MTX (500 нМ и 1 мкМ MTX). Таким образом, было также протестировано, можно ли повышать уровни экспрессии при проведении двух циклов селекции, используя ограниченную концентрацию фолиевой кислоты и MTX в среде для культивирования клеток. Вследствие этого, после проведения первого цикла селекции с использованием ограниченной концентрации фолиевой кислоты и MTX (относительно использованных концентраций смотри выше), клетки переводили в полную среду для восстановления. Затем клетки вновь подвергали воздействию во втором цикле селекции в той же самой селективной среде и, следовательно, такому же давлению отбора. Результаты приведены в таблице 9.
Таблица 9
Концентрация антитела (мАт) в культуральном супернатанте периодической культуры в [мг/л] после проведения двух циклов селекции
|
Из таблицы 9 видно, что, когда клетки, трансфицированные V-мутFRальфа/V-DHFRреф, вновь подвергали воздействию давления отбора через 35-38 дней, клетки демонстрировали очень значительное увеличение продуктивности от 21 до 670 мг/л. Это соответствует тридцатикратному возрастанию титра антитела. Другая популяция в селективной среде 50/50 также продуцировала в условиях повторного давления отбора в 20 раз больше, чем культура, культивируемая в полной среде. Кроме того, полученные результаты были значительно лучше, чем при использовании референсного способа с DHFR (смотри таблицу 8), в котором выполняли три цикла селекции. Таким образом, данный принцип селекции, когда два цикла селекции проводят с использованием селективной среды, содержащей ограниченную концентрацию фолата и антифолат, с промежуточным этапом культивирования в не селективной среде, приводит к исключительно высоким титрам экспрессии.
Пример 5: Одноклеточное клонирование
Для получения клеточных линий со стабильной экспрессией вектора популяции клеток, полученные в соответствии с примером 4, были клонированы после завершения селекции. После селекции поликлональный пул трансфицированных клеток 50/50 клонировали методом серийных разведений в полной среде (тем самым не поддерживая давление отбора в процессе клонирования) и селективной среде (тем самым поддерживая давление отбора в процессе клонирования) в 6 96-луночных планшетах. После удачного роста клонов клоны переводили при конфлюэнтности более 70% в 24-луночные планшеты и тестировали после 10 дней периодического культивирования на их продуктивность в отношении антитела. Результаты, достигнутые при клонировании популяции клеток, совместно трансфицированных V-мутFRальфа/V-DHFRреф и отобранных, приведены на фигуре 5. Как показано на фигуре 5, клонирование клеток с V-мутFRальфа/V-DHFRреф в полной среде и селективной среде привело к получению 65 клонов (55 в полной среде, 10 в селективной среде, содержащей 50 нМ FA/50 нМ MTX) из 6 96-луночных планшетов. Клоны располагали (в зависимости от среды) в порядке от наивысшего до самого низкого уровня экспрессии. Максимально продуцирующий клон был выделен в результате клонирования в полной среде и продуцировал в 24-луночной партии 450 мг/л. В 250-мл встряхиваемой колбе (всего 50 мл) исходный пул достигал титра 670 мг/л.
Для сравнения проводили клонирование методом серийных разведений содержащих DHFR клеток с вектором V-DHFRреф после предшествующей селекции с использованием G418-MTX. Соответствующие результаты были получены из предыдущего эксперимента и были получены в аналогичных условиях. Клонирование проводили в селективной среде, тем самым поддерживая давление отбора во время клонирования. Результаты приведены в таблице 10.
Таблица 10
Клонирование референсного пула клеток, трансфицированных V-DHFRреф
|
В данном случае проводили постепенную селекцию с G-418, 500 нМ MTX, 1 мкМ MTX. В таблице 10 приведены результаты конечного разведения популяции клеток в селективной среде (1 мкМ MTX). Клоны располагали в порядке от наивысшего до самого низкого уровня экспрессии. Максимально продуцирующий клон в данной референсной популяции достигал титра 51 мг/л в 20-луночной партии. Был получен широкий спектр клеточной продуктивности, который, однако, находился в основном ниже порогового значения 9 мг/л.
Результаты, показанные на Фиг.5 и в таблице 10, свидетельствуют, что совместная трансфекция V-мутFRальфа/V-DHFRреф приводила к значительно более высоким показателям продуктивности, и более того, число выделенных продуцирующих на высоком уровне клеточных клонов значительно возрастало. Более 50% клонов, выделенных из отобранной поликлональной популяции, достигали титра, превышающего 300 мг/л. Совместная трансфекция при наивысшей строгости селекции позволяла достигать девятикратного возрастания концентрации антитела среди максимально продуцирующих клеточных клонов с V-мутFRальфа, V-DHFRреф (другие способы селекции) и V-мутFRальфа/V-DHFRреф. Результаты приведены в таблице 11.
Таблица 11
Сводные данные по максимально продуцирующим клонам
|
В таблице 11 приведена концентрация антитела (мАт [мг/л]) у суперпродуцентов, полученных в результате проведенной селекции, после клонирования. Как можно видеть, совместная трансфекция V-мутFRальфа/V-DHFRреф и селекция в селективной среде, содержащей ограниченную концентрацию фолиевой кислоты, и дополнительно содержащей антифолат, приводила к наилучшим результатам.
Приведенные выше результаты свидетельствуют о том, что селекция, основанная на использовании мутантного фолатного рецептора в качестве селективного маркера, позволяла выживать успешно трансфицированным клеткам при использовании селективной среды, содержащей ограниченное количество фолата, в данном случае, фолиевой кислоты. Селекция с использованием мутантного фолатного рецептора альфа в качестве селективного маркера проходила гораздо быстрее, чем при использовании фолатного рецептора альфа дикого типа в качестве селективного маркера. Поскольку фолатный рецептор альфа дикого типа связывает с высокой аффинностью фолиевую кислоту (KD = 0,1 нМ), то давление отбора намного ниже переносимого порога концентрации фолиевой кислоты, ниже которого клетки, трансфицированные без ДНК, также могли бы выживать. Это также находит отражение в определяемой концентрации антитела. Клетки, которые были трансфицированы мутантным фолатным рецептором, могли выживать во всех протестированных селективных средах. Более того, наблюдали относительно однородный рост клеток. В трех наиболее концентрированных культуральных средах клетки можно было пересевать через 12 дней; в трех средах с наиболее низкой концентрацией фолиевой кислоты восстановление было достигнуто ко дню 16. В этих случаях давление отбора на клетки по сравнению с использованием фолатного рецептора дикого типа еще больше возрастало. Поскольку избыточная экспрессия мутантного фолатного рецептора коррелирует с экспрессией интересующего белка, определяемый титр антитела обратно пропорционален концентрации фолиевой кислоты в селективной среде.
Пример 6: Трансфекция векторов с dhfr, FolR дикого типа и FolR A49L в качестве селективных маркеров
В данном примере тестировали и сравнивали различные селективные условия. Клетки CHO трансфицировали векторами V-DHFRреф, V-дтFRальфа и V-мутFRальфа (мутант A49L). Для создания давления отбора на клетки-хозяева использовали ограниченную концентрацию фолиевой кислоты в селективной среде, что в настоящем документе также называют депривацией фолиевой кислоты.
Культивирование клеток, трансфекцию и скрининг проводили во встряхиваемых колбах с использованием растущих в суспензии клеток CHO в культуральной среде с определенным химическим составом. Клетки трансфицировали методом электропорации (нуклеофекции). Для селекции, основанной на депривации фолиевой кислоты, клетки пересевали в не содержащую фолиевую кислоту среду за 3 дня до трансфекции и трансфицировали в не содержащей фолиевую кислоту среде для уменьшения внутренних запасов фолиевой кислоты. В зависимости от жизнеспособности клеток селекцию начинали через 24-48 часов после трансфекции путем добавления селективной среды к клеткам. Трансфицированные V-дтFRальфа и V-мутFRальфа клетки отбирали с использованием 6 разных концентраций фолиевой кислоты (11700, 150, 50, 5, 0,5 и 0 нМ), в то время как в случае трансфицированных V-DHFRреф клеток для сравнения тестировали 6 разных концентраций MTX (2000, 1000, 500, 250, 125 и 0 нМ).
После восстановления клеток до жизнеспособности выше 80% после селекции анализировали продуктивность популяции выживших клеток. Продуктивность отобранных клеточных популяций анализировали после селекции с помощью переросших периодических культур во встряхиваемых колбах в полной среде, содержащей 11,7 мкм фолиевой кислоты. Периодические культуры высевали во встряхиваемые колбы (125) с рабочим объемом 50 мл и культивировали на шейкере в боксе (без увлажнения) в условиях 150 об/мин и 10% CO2. Жизнеспособность клеток должна была быть >90% в начале анализа. Плотность посева клеток составляла 2×105 клеток/мл. Определение титра проводили в день 13. Титры антитела в супернатанте клеточной культуры определяли методом ВЭЖХ с белком A через 13 дней после начала культивирования.
Результаты данного эксперимента описаны далее. Для оценки строгости селекции при использовании обоих вариантов фолатного рецептора в условиях ограниченных концентраций фолиевой кислоты тестировали различные концентрации фолиевой кислоты в диапазоне от 11700 нМ (референсная среда, полная среда) до 0 нМ для отбора избыточно экспрессирующих антитело клеток. Параллельно тестировали разные концентрации MTX с референсным вектором DHFR для сравнения эффективности. Все трансфицированные клеточные популяции смогли восстановиться. Популяции в случае 0 нМ фолиевой кислоты предположительно содержали следовые количества фолиевой кислоты, перенесенные из среды для предварительного культивирования. Этих остаточных количеств фолиевой кислоты, судя по всему, было достаточно для стимулирования выживания некоторой части клеток. Однако дальнейшая подкормка средой, содержащей фолиевую кислоту, была необходима для восстановления этих популяций. Продуктивность оценивали, как описано выше. В таблице 12 приведены сводные данные по продуктивности.
Таблица 12
Продуктивность клеточных популяций после селекции
|
В таблице 12 приведены результаты для трансфицированных клеток, выбранных при различных концентрациях фолиевой кислоты или MTX, которые анализировали в выращиваемых во встряхиваемых колбах периодических культурах. В день 13 культивирования образцы культуральной среды отбирали и анализировали на содержание антитела методом ВЭЖХ с белком A. Было обнаружено, что все клеточные популяции продуцировали антитело. В случае V-дтFRальфа максимальная продуктивность была достигнута в условиях селекции с 5 нМ фолиевой кислоты. Эта концентрация фолиевой кислоты ниже, чем концентрация, наблюдаемая в описанных выше экспериментах, и, вероятно, объясняется тем фактом, что некоторое количество фолиевой кислоты было перенесено из исходной культуральной среды в данном примере. Это также должно объяснять уровни восстановления и продуцирования, достигнутые при 0 нМ фолиевой кислоты. Дальнейшее уменьшение содержания фолиевой кислоты не приводило к более высокой продуктивности при использовании фолатного рецептора дикого типа. Напротив, в случае V-мутFRальфа была достигнута более высокая продуктивность при самой низкой концентрации фолиевой кислоты во время селекции. Продуктивность, достигнутая в случае мутанта, значительно превышала продуктивность, достигнутую при использовании фолатного рецептора дикого типа, и также значительно превышала продуктивность в случае DHFR/MTX.
Кроме того, при анализе восстановления клеток во время селекции было обнаружено, что клетки, трансфицированные V-мутFRальфа, восстанавливались значительно быстрее при очень низких концентрациях фолиевой кислоты, в частности, <25 нМ. Таким образом, более быстрое восстановление, описанное выше, было также подтверждено в данном эксперименте.
Вышеописанные примеры 1-6 демонстрируют преимущества, которые достигаются благодаря идеям настоящего изобретения, заключающимся в том, что для селекции используют мутантный фолатный рецептор. Например, в вышеописанных примерах секреция антитела в референсной популяции клеток (DHFR) составляла 28 мг/л, при максимальной для выживания строгости селекции в случае трансфицированных V-дтFRальфа клеток был получен результат 24 мг/л и в случае трансфицированных V-мутFRальфа клеток был получен результат 26,6 мг/л. Таким образом, показатели продуктивности, определенные в соответствующем эксперименте, были в сходном диапазоне, что указывало на то, что фолатный рецептор альфа дикого типа, а также мутантный фолатный рецептор альфа в качестве селективных маркеров позволяют достигать результатов, сопоставимых с результатами общепринятой системы селекции DHFR/MTX, которую можно считать «золотым стандартом». Более того, при сравнении периодов времени, необходимых для проведения селекции, видно, что селекцию можно проводить гораздо быстрее, используя систему селекции, основанную на мутантном фолатном рецепторе, предложенном по настоящему изобретению. Даже клетки, трансфицированные вектором V-дтFRальфа, которые в условиях 15 нМ фолиевой кислоты и 25 нМ фолиевой кислоты нуждались в более длительной фазе восстановления, чем клетки, трансфицированные V-мутFRальфа, при той же концентрации (16 дней), продемонстрировали через 20 дней очевидное преимущество перед клетками референсного контроля (DHFR), которые к этому времени все еще переживали кризис. Таким образом, использование мутантного фолатного рецептора по настоящему изобретению, по сравнению с системой DHFR/MTX, позволяет сохранять время во время фазы селекции при создании клеточной линии и также ускоряет процесс по сравнению с системой селекции на основе рецептора дикого типа. Данные результаты также свидетельствуют о том, что использование мутантного фолатного рецептора обеспечивает клеткам преимущество по сравнению с использованием фолатного рецептора дикого типа в тестируемых селективных средах, поскольку клетки выживают в более широком диапазоне концентраций фолиевой кислоты, и в частности, способны выживать при более низких концентрациях фолиевой кислоты, что позволяет создавать более строгие условия селекции. Кроме того, клетки выходят из кризиса, вызванного селекцией, гораздо раньше в случае мутантного фолатного рецептора, чем в случае, когда клетки трансфицированы фолатным рецептором дикого типа. Результаты показывают, что использование мутантного фолатного рецептора, описанного в настоящем документе, имеет важные преимущества.
Кроме того, также проводили эксперименты, в которых осуществляли двойную селекцию, используя фолатный рецептор и DHFR в качестве селективных маркеров, в условиях селективной среды, содержащей фолат в ограниченной концентрации и дополнительно содержащей антифолат, и они показали четкие преимущества мутантного фолатного рецептора. Мутация в фолатном рецепторе, очевидно, оказывала положительное влияние на рост клеток при указанном двойном давлении отбора. Без привязки к теории, возможно, что аффинность в отношении антифолатов, таких как MTX, снижена у мутанта, так что меньше MTX включается в клетки. Кроме того, установлено, что выгодно повторять селекцию и переводить клетки в полную среду в промежутке между двумя циклами селекции, чтобы позволить клеткам восстанавливаться после первого раунда селекции. После повторного перевода клеток в селективную среду было возможно обогащать клетки, которые имели оба интегрированных вектора в их геноме, и таким образом, были способны выживать при давлении двойного отбора. Было обнаружено, что продуктивность возрастала до двадцати-тридцати раз по сравнению с ситуацией в полной среде. Это является значительным преимуществом. По сравнению с референсным контролем после стандартной многоэтапной селекции с использованием G418/MTX все еще наблюдали возрастание продуктивности от шести- до тринадцати раз. Таким образом, система селекции по настоящему изобретению, в которой используют мутантный фолатный рецептор в сочетании с DHFR, имеет очевидные преимущества перед системой селекции предшествующего уровня техники. Более того, при использовании данной стратегии двойной селекции можно выделять более 50% продуцирующих на высоком уровне клонов с титром свыше 300 мг/л. Таким образом, поиск клонов-суперпродуцентов был менее трудоемким, что является существенным усовершенствованием по сравнению с существующими технологиями скрининга, в частности, для целей промышленного производства белков.
Мутантный фолатный рецептор, который по настоящему изобретению используют в качестве селективного маркера, является очень полезным, поскольку трансфицированные популяции клеток переживают кризис роста быстрее, чем в референсных системах селекции. Более того, популяции клеток, трансфицированных мутантным фолатным рецептором, демонстрировали после селекции в различных селективных средах экспрессию рецептора и антитела, которая была обратно пропорциональна концентрации фолиевой кислоты. Эту корреляцию можно продемонстрировать с использованием молекулярно-биологического анализа геномной ДНК (число копий), а также на уровне РНК. Кроме того, установлено, что использование мутантного фолатного рецептора, описанного в настоящем документе, является очень выгодным при выполнении совместной селекции с DHFR/MTX. Используемые контрольные клетки (после одиночных трансфекций FRдт, FRмут и DHFR) не могли пережить летальный эффект строгих условий сочетания депривации фолиевой кислоты и MTX. Хотя высокая строгость данной системы селекции также имела тот эффект, что были отобраны некоторые из совместно трансфицированных популяций. Однако за счет добавления фолиевой кислоты в качестве промежуточного этапа и проведения второго раунда селекции были достигнуты очень хорошие результаты с принципом совместной трансфекции при использовании принципа двойной селекции с FRмут/DHFR. Показано, что данный способ позволяет быстрее и с большей легкостью проводить скрининг на лучшие (первоклассные) клоны-продуценты. Одноклеточное клонирование максимально продуцирующей клеточной популяции (670 мг/л в 50 мл объема культуры) приводило к примерно 50% восстановлению продуцирующих на высоком уровне клеточных клонов, которые продуцировали более чем 300 мг/л. По сравнению с результатами одиночной и референсной трансфекции клонирование совместно трансфицированной популяции позволяло достигать средней продуктивности 240 мг/л, что является сорокакратным увеличением продуктивности. Для максимально продуцирующего клеточного клона были достигнуты результаты 450 мг/л в 24-луночной партии. Эти результаты подтверждают, что настоящее изобретение, основанное на использовании мутантного фолатного рецептора в качестве селективного маркера, вносит значительный вклад в существующие системы селекции.
Пример 7: Трансфекция экспрессионного вектора, содержащего два селективных маркера
В данном примере клетки CHO трансфицировали (нуклеофекцией) экспрессионным вектором, который содержал экспрессионную кассету, содержащую полинуклеотид, кодирующий мутантный человеческий фолатный рецептор альфа (мутант A49L - мутFRальфа, смотри выше), и экспрессионную кассету, содержащую полинуклеотид, кодирующий DHFR (V-мутFRальфа/DHFRреф). Таким образом, оба селективных маркера, мутFRальфа и DHFR, были на одном и том же экспрессионном векторе. Кроме того, экспрессионный вектор содержал экспрессионную кассету, содержащую полинуклеотид, кодирующий легкую цепь антитела, и экспрессионную кассету, содержащую полинуклеотид, кодирующий тяжелую цепь антитела. В данном эксперименте было экспрессировано иное антитело, чем в предыдущих примерах. Тестировали пять различных условий селекции с использованием 50 нМ фолиевой кислоты (FA) и разных концентраций MTX. Сводные данные по селективным средам приведены в следующей далее таблице 13. После селекции отобранные пулы клеток переводили в полную среду и выращивали в виде периодических культур во встряхиваемых колбах. В день 13 культивирования образцы культуральной среды отбирали и анализировали на содержание антитела методом ВЭЖХ с белком A. Результаты также приведены в таблице 13.
Таблица 13
Продуктивность пулов, полученная с экспрессионным вектором V-мутFRальфа/DHFRреф при использовании различных условий селекции
|
Как можно видеть, даже такая низкая концентрация MTX, как 5 нМ, обеспечивает значительное селекционное преимущество при использовании экспрессионного вектора V-мутFRальфа/DHFRреф, содержащего мутантный фолатный рецептор и DHFR в качестве селективного маркера. Это подтверждает преимущества использования мутантного фолатного рецептора в сочетании с DHFR для селекции, которые также продемонстрированы в других примерах. Продуктивность в отношении антитела значительно возрастала, и более того, более низкие концентрации MTX можно было использовать в процессе селекции, что является существенным преимуществом, учитывая, что MTX является токсичным средством.
Пример 8: Трансфекция с упрощенной предварительной обработкой родительских клеток CHO
Для проверки того, возможно ли избежать этапов центрифугирования/промывания клеток в данном способе, родительские клетки CHO брали для культивирования с использованием культуральной среды, содержащей ограниченную концентрацию 50 нМ фолиевой кислоты, из клеток, криоконсервированных либо в полной среде, либо в среде с 50 нМ фолиевой кислоты. После нескольких пересевов в этой среде клетки трансфицировали с использованием метода нуклеофекции и экспрессионного вектора V-мутFRальфа/DHFRреф, кодирующего моноклональные антитело. Эту трансфекцию и последующее культивирование осуществляли, используя ту же среду с 50 нМ фолиевой кислоты. Затем, через 48 часов после трансфекции давление отбора увеличивали за счет добавления 10 нМ MTX в культуру. Продуктивность культур, восстановленных после селекции, оценивали в периодических культурах, выращиваемых во встряхиваемых колбах, используя полную среду. Результаты приведены в таблице 14. Как показано в таблице 14, такие упрощенные протоколы процедур трансфекции и селекции приводили к достижению продуктивности, сопоставимой с продуктивностью в случае процедур, описанных в предыдущих примерах (например, таблица 13).
Таблица 14
Продуктивность пулов, полученная с экспрессионным вектором V-мутFRальфа/DHFRреф при использовании различных условий селекции
|
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Novartis AG
<120> Новые селективные векторы и способы селекции эукариотических клеток-хозяев
<130> PAT055303-WO-PCT
<150> US 61/860,439
<151> 2013-07-31
<160> 13
<170> PatentIn version 3.5
<210> 1
<211> 205
<212> PRT
<213> Homo sapiens
<400> 1
Ile Ala Trp Ala Arg Thr Glu Leu Leu Asn Val Cys Met Asn Ala Lys
1 5 10 15
His His Lys Glu Lys Pro Gly Pro Glu Asp Lys Leu His Glu Gln Cys
20 25 30
Arg Pro Trp Arg Lys Asn Ala Cys Cys Ser Thr Asn Thr Ser Gln Glu
35 40 45
Ala His Lys Asp Val Ser Tyr Leu Tyr Arg Phe Asn Trp Asn His Cys
50 55 60
Gly Glu Met Ala Pro Ala Cys Lys Arg His Phe Ile Gln Asp Thr Cys
65 70 75 80
Leu Tyr Glu Cys Ser Pro Asn Leu Gly Pro Trp Ile Gln Gln Val Asp
85 90 95
Gln Ser Trp Arg Lys Glu Arg Val Leu Asn Val Pro Leu Cys Lys Glu
100 105 110
Asp Cys Glu Gln Trp Trp Glu Asp Cys Arg Thr Ser Tyr Thr Cys Lys
115 120 125
Ser Asn Trp His Lys Gly Trp Asn Trp Thr Ser Gly Phe Asn Lys Cys
130 135 140
Ala Val Gly Ala Ala Cys Gln Pro Phe His Phe Tyr Phe Pro Thr Pro
145 150 155 160
Thr Val Leu Cys Asn Glu Ile Trp Thr His Ser Tyr Lys Val Ser Asn
165 170 175
Tyr Ser Arg Gly Ser Gly Arg Cys Ile Gln Met Trp Phe Asp Pro Ala
180 185 190
Gln Gly Asn Pro Asn Glu Glu Val Ala Arg Phe Tyr Ala
195 200 205
<210> 2
<211> 27
<212> PRT
<213> Homo sapiens
<400> 2
Ala Ala Met Ser Gly Ala Gly Pro Trp Ala Ala Trp Pro Phe Leu Leu
1 5 10 15
Ser Leu Ala Leu Met Leu Leu Trp Leu Leu Ser
20 25
<210> 3
<211> 257
<212> PRT
<213> Homo sapiens
<400> 3
Met Ala Gln Arg Met Thr Thr Gln Leu Leu Leu Leu Leu Val Trp Val
1 5 10 15
Ala Val Val Gly Glu Ala Gln Thr Arg Ile Ala Trp Ala Arg Thr Glu
20 25 30
Leu Leu Asn Val Cys Met Asn Ala Lys His His Lys Glu Lys Pro Gly
35 40 45
Pro Glu Asp Lys Leu His Glu Gln Cys Arg Pro Trp Arg Lys Asn Ala
50 55 60
Cys Cys Ser Thr Asn Thr Ser Gln Glu Ala His Lys Asp Val Ser Tyr
65 70 75 80
Leu Tyr Arg Phe Asn Trp Asn His Cys Gly Glu Met Ala Pro Ala Cys
85 90 95
Lys Arg His Phe Ile Gln Asp Thr Cys Leu Tyr Glu Cys Ser Pro Asn
100 105 110
Leu Gly Pro Trp Ile Gln Gln Val Asp Gln Ser Trp Arg Lys Glu Arg
115 120 125
Val Leu Asn Val Pro Leu Cys Lys Glu Asp Cys Glu Gln Trp Trp Glu
130 135 140
Asp Cys Arg Thr Ser Tyr Thr Cys Lys Ser Asn Trp His Lys Gly Trp
145 150 155 160
Asn Trp Thr Ser Gly Phe Asn Lys Cys Ala Val Gly Ala Ala Cys Gln
165 170 175
Pro Phe His Phe Tyr Phe Pro Thr Pro Thr Val Leu Cys Asn Glu Ile
180 185 190
Trp Thr His Ser Tyr Lys Val Ser Asn Tyr Ser Arg Gly Ser Gly Arg
195 200 205
Cys Ile Gln Met Trp Phe Asp Pro Ala Gln Gly Asn Pro Asn Glu Glu
210 215 220
Val Ala Arg Phe Tyr Ala Ala Ala Met Ser Gly Ala Gly Pro Trp Ala
225 230 235 240
Ala Trp Pro Phe Leu Leu Ser Leu Ala Leu Met Leu Leu Trp Leu Leu
245 250 255
Ser
<210> 4
<211> 203
<212> PRT
<213> Homo sapiens
<400> 4
Gln Asp Arg Thr Asp Leu Leu Asn Val Cys Met Asp Ala Lys His His
1 5 10 15
Lys Thr Lys Pro Gly Pro Glu Asp Lys Leu His Asp Gln Cys Ser Pro
20 25 30
Trp Lys Lys Asn Ala Cys Cys Thr Ala Ser Thr Ser Gln Glu Leu His
35 40 45
Lys Asp Thr Ser Arg Leu Tyr Asn Phe Asn Trp Asp His Cys Gly Lys
50 55 60
Met Glu Pro Ala Cys Lys Arg His Phe Ile Gln Asp Thr Cys Leu Tyr
65 70 75 80
Glu Cys Ser Pro Asn Leu Gly Pro Trp Ile Gln Gln Val Asn Gln Thr
85 90 95
Trp Arg Lys Glu Arg Phe Leu Asp Val Pro Leu Cys Lys Glu Asp Cys
100 105 110
Gln Arg Trp Trp Glu Asp Cys His Thr Ser His Thr Cys Lys Ser Asn
115 120 125
Trp His Arg Gly Trp Asp Trp Thr Ser Gly Val Asn Lys Cys Pro Ala
130 135 140
Gly Ala Leu Cys Arg Thr Phe Glu Ser Tyr Phe Pro Thr Pro Ala Ala
145 150 155 160
Leu Cys Glu Gly Leu Trp Ser His Ser Tyr Lys Val Ser Asn Tyr Ser
165 170 175
Arg Gly Ser Gly Arg Cys Ile Gln Met Trp Phe Asp Ser Ala Gln Gly
180 185 190
Asn Pro Asn Glu Glu Val Ala Arg Phe Tyr Ala
195 200
<210> 5
<211> 31
<212> PRT
<213> Homo sapiens
<400> 5
Ala Ala Met His Val Asn Ala Gly Glu Met Leu His Gly Thr Gly Gly
1 5 10 15
Leu Leu Leu Ser Leu Ala Leu Met Leu Gln Leu Trp Leu Leu Gly
20 25 30
<210> 6
<211> 255
<212> PRT
<213> Homo sapiens
<400> 6
Met Val Trp Lys Trp Met Pro Leu Leu Leu Leu Leu Val Cys Val Ala
1 5 10 15
Thr Met Cys Ser Ala Gln Asp Arg Thr Asp Leu Leu Asn Val Cys Met
20 25 30
Asp Ala Lys His His Lys Thr Lys Pro Gly Pro Glu Asp Lys Leu His
35 40 45
Asp Gln Cys Ser Pro Trp Lys Lys Asn Ala Cys Cys Thr Ala Ser Thr
50 55 60
Ser Gln Glu Leu His Lys Asp Thr Ser Arg Leu Tyr Asn Phe Asn Trp
65 70 75 80
Asp His Cys Gly Lys Met Glu Pro Ala Cys Lys Arg His Phe Ile Gln
85 90 95
Asp Thr Cys Leu Tyr Glu Cys Ser Pro Asn Leu Gly Pro Trp Ile Gln
100 105 110
Gln Val Asn Gln Thr Trp Arg Lys Glu Arg Phe Leu Asp Val Pro Leu
115 120 125
Cys Lys Glu Asp Cys Gln Arg Trp Trp Glu Asp Cys His Thr Ser His
130 135 140
Thr Cys Lys Ser Asn Trp His Arg Gly Trp Asp Trp Thr Ser Gly Val
145 150 155 160
Asn Lys Cys Pro Ala Gly Ala Leu Cys Arg Thr Phe Glu Ser Tyr Phe
165 170 175
Pro Thr Pro Ala Ala Leu Cys Glu Gly Leu Trp Ser His Ser Tyr Lys
180 185 190
Val Ser Asn Tyr Ser Arg Gly Ser Gly Arg Cys Ile Gln Met Trp Phe
195 200 205
Asp Ser Ala Gln Gly Asn Pro Asn Glu Glu Val Ala Arg Phe Tyr Ala
210 215 220
Ala Ala Met His Val Asn Ala Gly Glu Met Leu His Gly Thr Gly Gly
225 230 235 240
Leu Leu Leu Ser Leu Ala Leu Met Leu Gln Leu Trp Leu Leu Gly
245 250 255
<210> 7
<211> 223
<212> PRT
<213> Homo sapiens
<400> 7
Gln Pro Arg Ser Ala Arg Ala Arg Thr Asp Leu Leu Asn Val Cys Met
1 5 10 15
Asn Ala Lys His His Lys Thr Gln Pro Ser Pro Glu Asp Glu Leu Tyr
20 25 30
Gly Gln Cys Ser Pro Trp Lys Lys Asn Ala Cys Cys Thr Ala Ser Thr
35 40 45
Ser Gln Glu Leu His Lys Asp Thr Ser Arg Leu Tyr Asn Phe Asn Trp
50 55 60
Asp His Cys Gly Lys Met Glu Pro Thr Cys Lys Arg His Phe Ile Gln
65 70 75 80
Asp Ser Cys Leu Tyr Glu Cys Ser Pro Asn Leu Gly Pro Trp Ile Arg
85 90 95
Gln Val Asn Gln Ser Trp Arg Lys Glu Arg Ile Leu Asn Val Pro Leu
100 105 110
Cys Lys Glu Asp Cys Glu Arg Trp Trp Glu Asp Cys Arg Thr Ser Tyr
115 120 125
Thr Cys Lys Ser Asn Trp His Lys Gly Trp Asn Trp Thr Ser Gly Ile
130 135 140
Asn Glu Cys Pro Ala Gly Ala Leu Cys Ser Thr Phe Glu Ser Tyr Phe
145 150 155 160
Pro Thr Pro Ala Ala Leu Cys Glu Gly Leu Trp Ser His Ser Phe Lys
165 170 175
Val Ser Asn Tyr Ser Arg Gly Ser Gly Arg Cys Ile Gln Met Trp Phe
180 185 190
Asp Ser Ala Gln Gly Asn Pro Asn Glu Glu Val Ala Lys Phe Tyr Ala
195 200 205
Ala Ala Met Asn Ala Gly Ala Pro Ser Arg Gly Ile Ile Asp Ser
210 215 220
<210> 8
<211> 245
<212> PRT
<213> Homo sapiens
<400> 8
Met Asp Met Ala Trp Gln Met Met Gln Leu Leu Leu Leu Ala Leu Val
1 5 10 15
Thr Ala Ala Gly Ser Ala Gln Pro Arg Ser Ala Arg Ala Arg Thr Asp
20 25 30
Leu Leu Asn Val Cys Met Asn Ala Lys His His Lys Thr Gln Pro Ser
35 40 45
Pro Glu Asp Glu Leu Tyr Gly Gln Cys Ser Pro Trp Lys Lys Asn Ala
50 55 60
Cys Cys Thr Ala Ser Thr Ser Gln Glu Leu His Lys Asp Thr Ser Arg
65 70 75 80
Leu Tyr Asn Phe Asn Trp Asp His Cys Gly Lys Met Glu Pro Thr Cys
85 90 95
Lys Arg His Phe Ile Gln Asp Ser Cys Leu Tyr Glu Cys Ser Pro Asn
100 105 110
Leu Gly Pro Trp Ile Arg Gln Val Asn Gln Ser Trp Arg Lys Glu Arg
115 120 125
Ile Leu Asn Val Pro Leu Cys Lys Glu Asp Cys Glu Arg Trp Trp Glu
130 135 140
Asp Cys Arg Thr Ser Tyr Thr Cys Lys Ser Asn Trp His Lys Gly Trp
145 150 155 160
Asn Trp Thr Ser Gly Ile Asn Glu Cys Pro Ala Gly Ala Leu Cys Ser
165 170 175
Thr Phe Glu Ser Tyr Phe Pro Thr Pro Ala Ala Leu Cys Glu Gly Leu
180 185 190
Trp Ser His Ser Phe Lys Val Ser Asn Tyr Ser Arg Gly Ser Gly Arg
195 200 205
Cys Ile Gln Met Trp Phe Asp Ser Ala Gln Gly Asn Pro Asn Glu Glu
210 215 220
Val Ala Lys Phe Tyr Ala Ala Ala Met Asn Ala Gly Ala Pro Ser Arg
225 230 235 240
Gly Ile Ile Asp Ser
245
<210> 9
<211> 205
<212> PRT
<213> Искусственная
<220>
<223> Зрелый мутантный фолатный рецептор
<220>
<221> MISC_FEATURE
<222> (49)..(49)
<223> Xaa является любой аминокислотой, за исключением Ala
<400> 9
Ile Ala Trp Ala Arg Thr Glu Leu Leu Asn Val Cys Met Asn Ala Lys
1 5 10 15
His His Lys Glu Lys Pro Gly Pro Glu Asp Lys Leu His Glu Gln Cys
20 25 30
Arg Pro Trp Arg Lys Asn Ala Cys Cys Ser Thr Asn Thr Ser Gln Glu
35 40 45
Xaa His Lys Asp Val Ser Tyr Leu Tyr Arg Phe Asn Trp Asn His Cys
50 55 60
Gly Glu Met Ala Pro Ala Cys Lys Arg His Phe Ile Gln Asp Thr Cys
65 70 75 80
Leu Tyr Glu Cys Ser Pro Asn Leu Gly Pro Trp Ile Gln Gln Val Asp
85 90 95
Gln Ser Trp Arg Lys Glu Arg Val Leu Asn Val Pro Leu Cys Lys Glu
100 105 110
Asp Cys Glu Gln Trp Trp Glu Asp Cys Arg Thr Ser Tyr Thr Cys Lys
115 120 125
Ser Asn Trp His Lys Gly Trp Asn Trp Thr Ser Gly Phe Asn Lys Cys
130 135 140
Ala Val Gly Ala Ala Cys Gln Pro Phe His Phe Tyr Phe Pro Thr Pro
145 150 155 160
Thr Val Leu Cys Asn Glu Ile Trp Thr His Ser Tyr Lys Val Ser Asn
165 170 175
Tyr Ser Arg Gly Ser Gly Arg Cys Ile Gln Met Trp Phe Asp Pro Ala
180 185 190
Gln Gly Asn Pro Asn Glu Glu Val Ala Arg Phe Tyr Ala
195 200 205
<210> 10
<211> 25
<212> PRT
<213> Homo sapiens
<400> 10
Met Ala Gln Arg Met Thr Thr Gln Leu Leu Leu Leu Leu Val Trp Val
1 5 10 15
Ala Val Val Gly Glu Ala Gln Thr Arg
20 25
<210> 11
<211> 21
<212> PRT
<213> Homo sapiens
<400> 11
Met Val Trp Lys Trp Met Pro Leu Leu Leu Leu Leu Val Cys Val Ala
1 5 10 15
Thr Met Cys Ser Ala
20
<210> 12
<211> 22
<212> PRT
<213> Homo sapiens
<400> 12
Met Asp Met Ala Trp Gln Met Met Gln Leu Leu Leu Leu Ala Leu Val
1 5 10 15
Thr Ala Ala Gly Ser Ala
20
<210> 13
<211> 232
<212> PRT
<213> Искусственная
<220>
<223> Мутантный фолатный рецептор
<400> 13
Ile Ala Trp Ala Arg Thr Glu Leu Leu Asn Val Cys Met Asn Ala Lys
1 5 10 15
His His Lys Glu Lys Pro Gly Pro Glu Asp Lys Leu His Glu Gln Cys
20 25 30
Arg Pro Trp Arg Lys Asn Ala Cys Cys Ser Thr Asn Thr Ser Gln Glu
35 40 45
Leu His Lys Asp Val Ser Tyr Leu Tyr Arg Phe Asn Trp Asn His Cys
50 55 60
Gly Glu Met Ala Pro Ala Cys Lys Arg His Phe Ile Gln Asp Thr Cys
65 70 75 80
Leu Tyr Glu Cys Ser Pro Asn Leu Gly Pro Trp Ile Gln Gln Val Asp
85 90 95
Gln Ser Trp Arg Lys Glu Arg Val Leu Asn Val Pro Leu Cys Lys Glu
100 105 110
Asp Cys Glu Gln Trp Trp Glu Asp Cys Arg Thr Ser Tyr Thr Cys Lys
115 120 125
Ser Asn Trp His Lys Gly Trp Asn Trp Thr Ser Gly Phe Asn Lys Cys
130 135 140
Ala Val Gly Ala Ala Cys Gln Pro Phe His Phe Tyr Phe Pro Thr Pro
145 150 155 160
Thr Val Leu Cys Asn Glu Ile Trp Thr His Ser Tyr Lys Val Ser Asn
165 170 175
Tyr Ser Arg Gly Ser Gly Arg Cys Ile Gln Met Trp Phe Asp Pro Ala
180 185 190
Gln Gly Asn Pro Asn Glu Glu Val Ala Arg Phe Tyr Ala Ala Ala Met
195 200 205
Ser Gly Ala Gly Pro Trp Ala Ala Trp Pro Phe Leu Leu Ser Leu Ala
210 215 220
Leu Met Leu Leu Trp Leu Leu Ser
225 230
<---
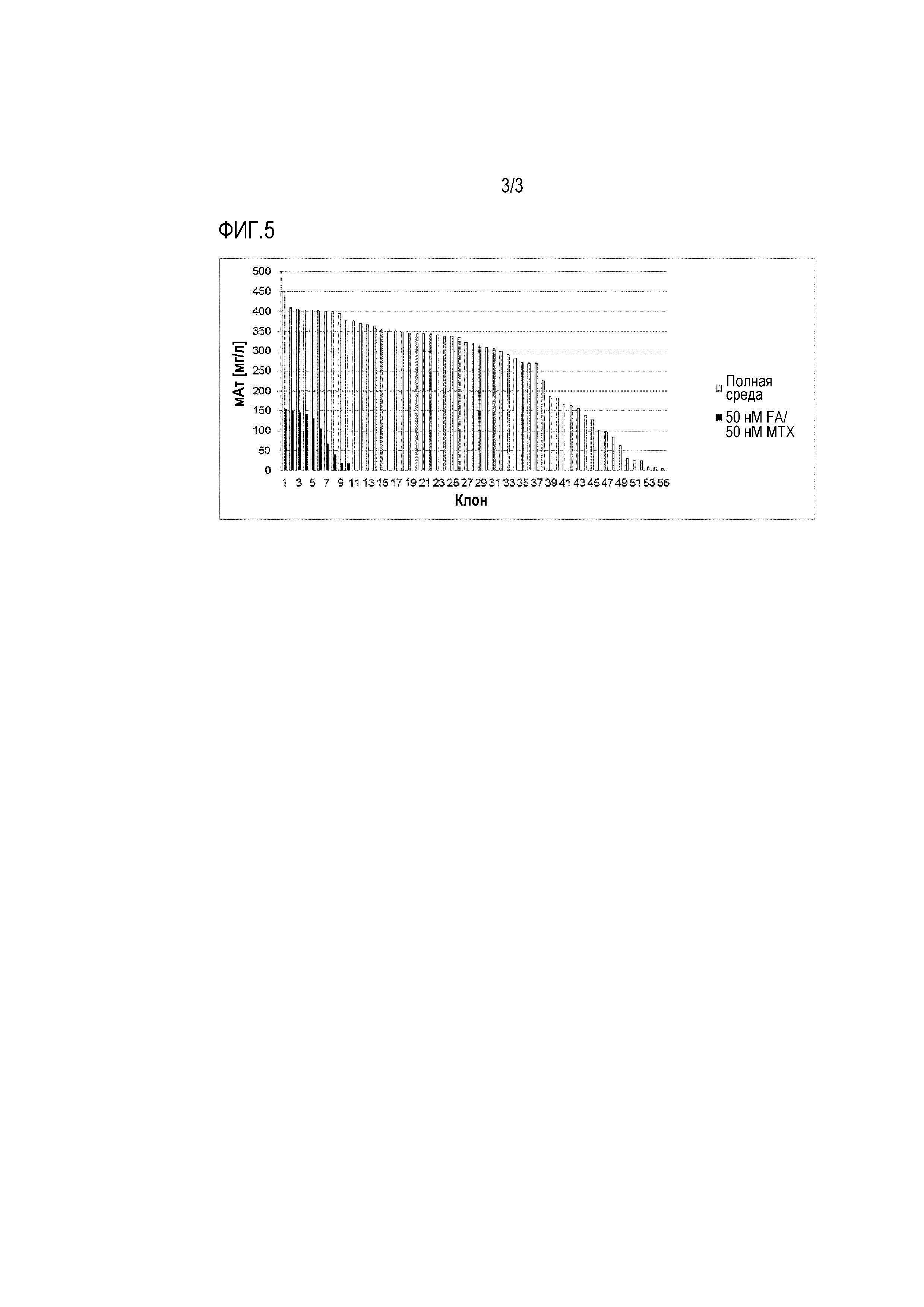
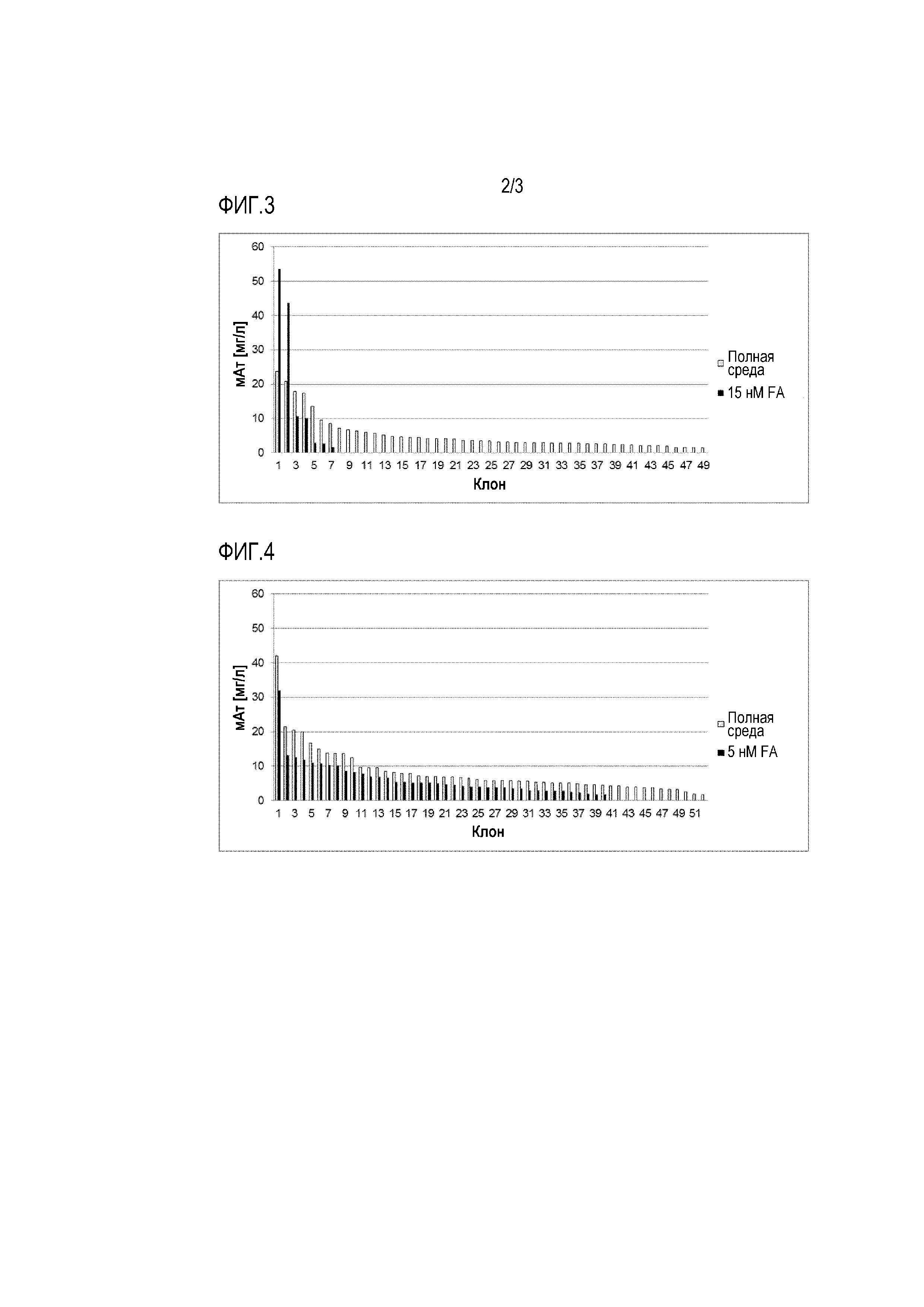
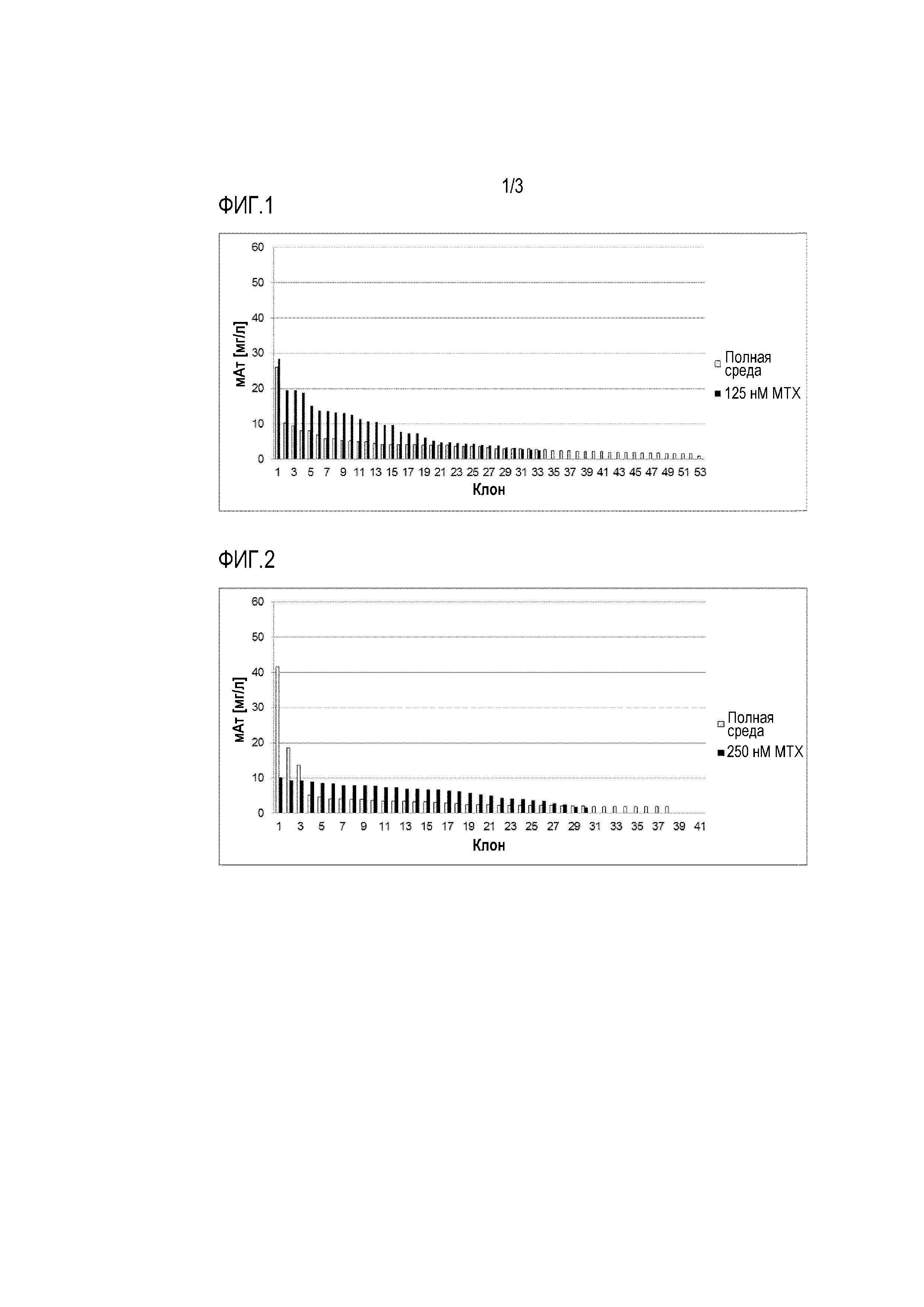
Мутантные антигены gas57 и антитела против gas57
[(1н-индол-5-ил)-гетероарилокси]-1-(азабицикло[3.3.1]нонаны, как холинергические лиганды n-achr, предназначенные для лечения психотических и нейродегенеративных нарушений
Лечение туберозного склероза
Органические соединения
Способ получения фармацевтической композиции
Предназначенная для перорального применения фармацевтическая композиция
Менингококковые полипептиды fhbp
Стабилизированные полипептиды инсулиноподобного фактора роста
Органические соединения
Курс лечения с использованием агониста рецептора s1p
Новые эукариотические клетки и способы их получения для рекомбинантной экспрессии представляющего интерес продукта
Новые эукариотические клетки и способы их получения для рекомбинантной экспрессии представляющего интерес продукта

