Результат интеллектуальной деятельности: СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ СЕРУСОДЕРЖАЩИХ АМИНОКИСЛОТ
Вид РИД
Изобретение
Настоящее изобретение относится к способу ферментативного получения серусодержащих аминокислот, выбранных из группы, включающей L-метионин, L-цистеин, L-цистин, L-гомоцистеин и L-гомоцистин.
Серусодержащие аминокислоты имеют важное экономическое значение. L-цистеин применяют в качестве пищевой добавки, в качестве сырья для фармакологически активных соединений (например, N-ацетилцистеина) и для косметических средств. Аминокислота L-метионин имеет очень важное значение для питания животных. Она относится к незаменимым аминокислотам, которые не могут образовываться путем биосинтеза в процессе метаболизма у позвоночных животных. Следовательно, при выкармливании животных необходимо гарантировать потребление в достаточных количествах метионина с кормом. Однако, поскольку L-метионин часто присутствует в традиционных кормовых растениях (таких как соя или зерновые культуры) в количествах, слишком низких для того, чтобы гарантировать оптимальное питание животных, прежде всего свиней и домашней птицы, то целесообразно примешивать метионин в качестве добавки к корму для животных. В организме позвоночных животных D-метионин может превращаться в биологически активный L-метионин. Поэтому в корм для животных, как правило, добавляют рацемат D- и L-метионина. В организме животных L-гомоцистеин может превращаться путем трансметилирования в L-метионин и поэтому может служить его заменой.
Из существующего уровня техники известно получение аминокислот, таких как метионин, путем химического синтеза. При таком получении сначала подвергают взаимодействию акролеин и метилмеркаптан с получением 3-метилтиопропиональдегида, из которого путем взаимодействия с цианидом, аммиаком и монооксидом углерода получают гидантоин. На конечной стадии его гидролизуют с получением рацемата, эквимолярной смеси обоих стереоизомеров, т.е. D- и L-метионина. Поскольку обладающая биологической активностью форма молекулы представляет собой исключительно L-форму, то D-форма, входящая в корм, сначала должна превратиться в процессе метаболизма путем де- и трансаминирования в активную L-форму.
В отличие от метионина большинство других встречающихся в естественных условиях протеиногенных аминокислот получают главным образом путем ферментации с использованием микроорганизмов. Этот метод основан на том факте, что для микроорганизмов характерны соответствующие пути биосинтеза, обеспечивающие синтез встречающихся в естественных условиях аминокислот. Кроме того, многие процессы ферментации позволяют осуществлять очень выгодное с точки зрения стоимости производство с выделением недорогих эдуктов, таких как глюкоза и минеральные соли, и кроме того, позволяют получать биологически активную L-форму конкретной аминокислоты.
Однако в штаммах дикого типа пути биосинтеза аминокислот находятся под строгим метаболическим контролем, это гарантирует, что аминокислоты продуцируются только для обеспечения собственных потребностей клетки. Таким образом, важной предпосылкой для эффективных процессов производства является создание соответствующих микроорганизмов, которые в отличие от организмов дикого типа обладают резко увеличенной способностью продуцировать требуемую аминокислоту.
Указанные микроорганизмы, отличающиеся сверхпроизводством аминокислот, можно получать общепринятыми методами осуществления мутации/селекции и/или с использованием современных целенаправленных методов рекомбинации (метаболическая инженерия). В последнем случае прежде всего идентифицируют гены или аллели, которые влияют на сверхпроизводство аминокислот в результате их модификации, активации или инактивации. Затем эти гены/аллели интродуцируют в штамм микроорганизма или инактивируют с использованием методов молекулярной биологии таким образом, чтобы достигать оптимального сверхпроизводства. Однако часто только комбинация нескольких различных путей приводит к действительно эффективному производству.
В E. coli и C. glutamicum L-цистеин образуется в результате биохимических процессов из L-серина. L-серин активируется с образованием O-ацетилсерина с помощью серинацетилтрансферазы CysE. Затем O-ацетилсерин-(тиол)лиаза переносит восстановленную серу в форме сульфида в результате чего образуется L-цистеин. В отличие от C. glutamicum у E. coli присутствуют две различные O-ацетилсерин-(тиол)лиазы, в том числе O-ацетилсерин-(тиол)лиаза В («CysM»), которая тоже обладает способностью переносить тиосульфат на 0-ацетилсерин. В результате образуется S-сульфоцистеин, который затем расщепляется на L-цистеин и сульфит или сульфат с помощью пока не изученного механизма (Kredich N.M. в: «Escherichia coli and Salmonella», под ред. Neidhardt F.C. и др., 2-ое изд., 1996, сс. 514-527).
L-метионин наряду с лизином и треонином образуется из аспартата. Серу интродуцируют в L-метионин в форме L-цистеина (через цистатионин в качестве промежуточного продукта) путем транссульфурирования (в C. glutamicum и E. coli; у E. coli этот путь является единственным). Параллельно этому, например, у C. glutamicum, присутствует путь прямого сульфурирования, при котором сульфид ассимилируется в форме L-гомоцистеина (B-J Hwang, H-J Yeom, Y. Kirn, H-S Lee J Bacteriol., 184(5), 2002, сс. 1277-1286). С 1-группа L-метионина образуется в результате С 1-метаболизма и переносится на L-гомоцистеин с помощью метионинсинтаз MetE и MetH (обзор: Greene R.C. в: «Escherichia coli and Salmonella», под ред. Neidhardt F.C. и др., 2-ое изд., 1996, сс. 542-560). Биосинтез L-метионина в C. glutamicum описан у Ruckert с соавторами (Ruckert С., PUhler A., Kalinowski J., J Biotechnol., 104(1-3), 2003, сс. 213-228). Штаммы и методы ферментативного получения L-метионина описаны, например, для E. coli (WO 2006/001616, WO 2009/043803) и C. glutamicum (WO 2009/144270).
Использование тиосульфата в качестве источника серы позволяет значительно увеличивать теоретический выход производства серусодержащих аминокислот (по сравнению с сульфатом) (Krömer J.O., Wittmann С., Schroder H., Heinzle Е., Metab Eng., 8 (4), 2006, сс. 353-369; и WO 2007/020295). Преимущество тиосульфата объясняется следующим образом: в сульфате (S04) присутствует атом серы, имеющий степень окисления +6. Для ассимиляции степень окисления должна быть снижена до -2 (сульфид, S "). Для восстановления сульфата до сульфида клетки должны использовать две молекулы АТФ и 4 НАДФ-Н (т.е. в общей сложности 8 электронов). В тиосульфате центральный атом серы имеет степень окисления +5, а концевой атом серы имеет степень окисления -1 (средняя формальная степень окисления двух атомов серы: +2). Для восстановления обоих атомов серы тиосульфата требуется только 8 электронов по сравнению с 16 электронами, которые требуются для восстановления двух сульфатов.
Применение тиосульфата в качестве источника S для производства L-цистеина с использованием E. coli описано, например, в DE 102007007333 и WO 2001/27307.
В WO 2007/077041 описано, что при применении тиосульфата вместо сульфата можно повышать также в значительной степени производство L-метионина в E. coli.
К настоящему времени пока не продемонстрировано экспериментально, что применение тиосульфата приводит также к повышению производства L-метионина в C. glutamicum. В WO 2007/020295 (с. 240) указано, что при применении Na-тиосульфата в качестве источника серы C. glutamicum образует больше биомассы. Это определяли, используя в качестве показателя снижение уровня потребления АТФ и НАДФ-Н. Производство L-метионина с использованием тиосульфата на практике не изучено.
У E. coli тиосульфат поглощается с помощью транспортера сульфата/тиосульфата CysPUWA-Sbp (Kredich N.M., в: «Escherichia coli and Salmonella», под ред. Neidhardt F.C. и др., 2-ое изд., 1996, сс. 514-527). CysP и Sbp представляют собой два различных периплазматических связывающих белка. CysP обладает повышенной аффинностью к тиосульфату, а Sbp обладает повышенной аффинностью к сульфату. CysA образует АТФ-связывающий компонент.CysU и CysW представляют собой трансмембранные компоненты. В WO 2009/043803 описано усиление экспрессии оперона cysPUWAMb E. coli и достижение тем самым повышения производства L-метионина. Сведения о поглощении тиосульфата у C. glutamicum отсутствуют.«Knock ом/»-мутанты предполагаемой системы поглощения сульфата CysZ (cg3112) все еще могут сохранять способность к росту в присутствии тиосульфата в качестве источника S (Riickert С. и др., ВМС Genomics., т. 6 (121), 2005). Таким образом, CysZ не является (единственным) элементом, ответственным за транспорт тиосульфата.
O-ацетилсерин-(тиол)лиаза В (т.е. CysM, КФ 2.5.1.47) обеспечивает способность E. coli использовать тиосульфат в качестве источника S для синтеза L-цистеина и L-метионина. Указанный фермент катализирует образование S-сульфоцистеина (R-S-SO3) и ацетата из O-ацетилсерина и тиосульфата. Затем S-сульфоцистеин расщепляется на L-цистеин и сульфит или сульфат с помощью пока не известного пути (Kredich N.M. в: «Escherichia coli and Salmonella», под ред. Neidhardt F.C. и др., 2-ое изд., 1996, сс. 514-527). Сульфат восстанавливается до сульфита (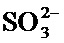 ) в результате трехстадийного процесса с использованием АТФ-сульфурилазы (CysDN), APS-киназы (CysC) и PAPS-сульфотрансферазы (CysH). Затем сульфит восстанавливается с помощью НАДФ⋅Н-сульфитредуктазы с образованием сульфида (S2-), который O-ацетилсерин(тиол)лиаза A (CysK) и O-ацетилсерин(тиол)лиаза В (CysM) могут использовать для синтеза второй молекулы L-цистеина.
) в результате трехстадийного процесса с использованием АТФ-сульфурилазы (CysDN), APS-киназы (CysC) и PAPS-сульфотрансферазы (CysH). Затем сульфит восстанавливается с помощью НАДФ⋅Н-сульфитредуктазы с образованием сульфида (S2-), который O-ацетилсерин(тиол)лиаза A (CysK) и O-ацетилсерин(тиол)лиаза В (CysM) могут использовать для синтеза второй молекулы L-цистеина.
C. glutamicum может расти в минимальной среде с тиосульфатом в качестве источника серы (Rflckert С. и др., ВМС Genomics., т.6(121), 2005). Однако у C. glutamicum отсутствует O-ацетилсерин(тиол)лиаза В («CysM») (Ruckert и Kalinowski в: «Corynebacteria: Genomics and Molecular Biology», под ред. А. Burkovski, изд-во Caister Academic Press, 2008). Таким образом, тиосульфат может ассимилироваться иным образом.
В основу настоящего изобретения была положена задача разработать способ и штаммы микроорганизмов, позволяющие увеличивать сверхпроизводство серусодержащих аминокислот, в частности L-метионина.
Указанная задача решается с помощью способа ферментативного получения серусодержащих аминокислот, выбранных из группы, включающей L-метионин, L-цистеин, L-цистин, L-гомоцистеин и L-гомоцистин, заключающегося в том, что осуществляют стадии, на которых:
а) получают микроорганизм семейства Enterobacteriaceae или микроорганизм семейства Corynebacteriaceae, который обладает повышенной тиосульфат-сульфотрансферазной активностью по сравнению с конкретным исходным штаммом;
б) ферментируют микроорганизм, полученный на стадии а), в среде, которая содержит соль дитиосерной кислоты или смесь соли дитиосерной кислоты и соли серной кислоты в качестве неорганического источника серы, получая ферментационный бульон, и
в) концентрируют серусодержащую аминокислоту в ферментационном бульоне, полученном на стадии б).
Изобретение относится также к способу ферментативного получения серусодержащих аминокислот, выбранных из группы, включающей L-метионин, L-цистеин, L-цистин, L-гомоцистеин и L-гомоцистин, заключающемуся в том, что осуществляют стадии, на которых:
а) получают микроорганизм семейства Enterobacteriaceae или микроорганизм семейства Corynebacteriaceae, в котором имеет место сверхэкспрессия гена, кодирующего полипептид с активностью тиосульфат-сульфотрансферазы;
б) ферментируют микроорганизм, полученный на стадии а), в среде, которая содержит соль дитиосерной кислоты или смесь соли дитиосерной кислоты и соли серной кислоты в качестве неорганического источника серы, получая ферментационный бульон, и
в) концентрируют серусодержащую аминокислоту в ферментационном бульоне, полученном на стадии б).
Тиосульфат-сульфотрансферазы, которые обозначают также как «роданазы» (КФ 2.8.1.1), представляют собой ферменты, которые переносят восстановленный атом серы с тиосульфата на цианид (CN-).
Некоторые тиосульфат-сульфотрансферазы могут переносить также восстановленный атом серы на альтернативные субстраты, например, на дигидролипоат (Alexander К., Volini M, J. Biol. Chem., 262, 1987, сс. 6595-6604). В базе данных «Clusters of Orthologous Groups of proteins» (кластеры ортологичных групп белков) (COG) в категории COG0607 «Rhodanese-related sulphurtransferase» в настоящее время присутствуют 168 гомологов сульфотрансфераз во всех трех доменах жизни (Tatusov R.L., Fedorova N.D., Jackson J.D., Jacobs A.R., Kiryutin В., Koonin E.V., Krylov D.M., Mazumder R., Mekhedov S.L., Nikolskaya A.N., Rao B.S., Smirnov S., Sverdlov A.V., Vasudevan S., Wolf Y.I., Yin J.J., Natale D.A., BMC Bioinformatics, 4, 2003, с. 41; http://www.ncbi.nlm.nih.gov/COG/). Родоназы имеют один или несколько характерных роданазаподобных доменов (база данных PFAMe PF00581, http://pfam.sanger.ac.uk/: Gliubich F., Gazerro M., Zanotti G., Delbono S., Bombieri G., Berni R., J Biol Chem., 271 (35), 1996, сс. 21054210-61).
Ген RDL2 (rhodanese-like protein) (роданазаподобный белок) из Saccharomyces cerevisiae S288c кодирует состоящий из 149 аминокислот белок Rdl2p.Он содержит роданазаподобный домен (база данных PFAM PF00581, http://pfam.sanger.ac.uk/). Экспериментально установлено, что Rdl2p представляет собой тиосульфат-сульфотрансферазу (роданаза, КФ 2.8.1.1; Foster M.W., Forrester M.T., Stamler J.S., Proc Nati Acad Sci USA, 106(45), 2009, сс. 18948-18953). В SEQ ID NO: 1 представлена ДНК-последовательность гена RDL2. В SEQ ID NO: 2 представлена аминокислотная последовательность белка Rdl2p, кодируемого RDL2.
E. coli несет помимо хорошо охарактеризованных роданаз GlpE и PspE (Adams H., Teertstra W., Koster M., Tommassen J., FEBS Lett. 518, 2002, сс. 173-176) семь дополнительных паралогичных ферментов с роданазаподобными доменами (Cheng H., Donahue J.L., Battle S.E., Ray W.K., Larson T.J., Open Microbiol J., 2, 2008, сс. 18-28):
Фермент YgaP содержит N-концевой роданазаподобный домен и два трансмембранных домена. Для него обнаружена in vitro роданазная активность (Ahmed F., 2003, диссертация).
Фермент YbbB представляет собой тРНК-2-селенуридинсинтазу и имеет N-концевой роданазаподобный домен (Wolfe M.D., Ahmed F., Lacourciere G.M., Lauhon C.T., Stadtman T.C., Larson T.J., J Biol Chem., 279(3), 2004, сс. 1801-1809).
Фермент ThiI необходим для синтеза тиамина и играет роль в превращении уридина в тиоуридин в положении 8 в тРНК (Palenchar P.M., Buck C.J., Cheng H., Larson T.J., Mueller E.G., J Biol Chem., 275(12), 2000, сс. 8283-8286).
Фермент SseA представляет собой 3-меркаптопуриватщианид-сульфотрансферазу, которая главным образом переносит 3-меркаптопуриват вместо тиосульфата на цианид (Colnaghi R., Cassinelli G., Drummond M., Forlani F., Pagani S., FEBS Lett., 500(3), 2001, сс. 153-156). Фермент имеет два роданазаподобных домена.
OPC ynjE кодирует предполагаемую сульфотрансферазу с тремя роданазаподобными доменами (Hanzelmann P., Dahl J.U., Kuper J., Urban A., Muller-Theissen U., Leimkuhler S., Schindelin H., Protein Sci., 18 (12), 2009, сс. 2480-2491).
OPC yibN кодирует предполагаемую сульфотрансферазу с одним C-концевым роданазаподобным доменом.
OPC yceA кодирует предполагаемую сульфотрансферазу с одним роданазаподобным доменом.
У Corynebacterium glutamicum по меньшей мере 7 OPC кодируют предполагаемые сульфотрансферазы:
OPC thtR (т.е. cg0803, NCg10671) кодирует предполагаемую сульфотрансферазу с двумя роданазаподобными доменами, состоящую из 301 аминокислоты.
OPC sseA2 (т.е. eg 1613, NCg11369) кодирует предполагаемую сульфотрансферазу с двумя роданазаподобными доменами, состоящую из 289 аминокислот.
OPC cg3000 (т.е. NCg12616) кодирует предполагаемую сульфотрансферазу с одним роданазаподобным доменом, состоящую из 96 аминокислот.
OPC cg0073 (т.е. NCg10053) кодирует предполагаемую сульфотрансферазу с одним роданазаподобным доменом, состоящую из 97 аминокислот.
ORF cg0074 (т.е. NCg10054) кодирует предполагаемую сульфотрансферазу с одним роданазаподобным доменом, состоящую из 197 аминокислот.
OPC sseAl (т.е. cg3073, NCg12678) кодирует предполагаемую сульфотрансферазу с двумя роданазаподобными доменами, состоящую из 274 аминокислот.
OPCF cg3319 (т.е. NCg12890) кодирует предполагаемую сульфотрансферазу с одним роданазаподобным доменом, состоящую из 312 аминокислот.
Тиосульфат-сульфотрансфераза из быков (Bos taurus) имеет два роданазаподобных домена и хорошо охарактеризована (Cannella С., Costa М., Pensa В., Ricci G., Pecci L., Cavallini D., Eur J Biochem., 119 (3), 1981, сс. 491-495).
В предпочтительном варианте способа у микроорганизма происходит сверхэкспрессия одного гена или нескольких генов, кодирующего(их) полипептид с активностью тиосульфат-сульфотрансферазы, при этом полипептид с активностью тиосульфат-сульфотрансферазы выбирают из следующих полипептидов, указанных в подпунктах а)- г):
а) полипептид, состоящий из или содержащий полипептиды Rdl2p, GlpE, PspE, YgaP, ThiI, YbbB, SseA, YnjE, YceA, YibN, NCg10671, NCg11369, NCg12616, NCg10053, NCg10054, NCg12678, NCg12890; тиосульфат-сульфотрансферазу млекопитающих, например, тиосульфат-сульфотрансферазу из бычьей печени (Bos taurus); предпочтительно Rdl2p, GlpE, PspE и наиболее предпочтительно Rdl2p;
б) полипептид, состоящий из или содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2;
в) полипептид, аминокислотная последовательность которого идентична на 70% или более аминокислотной последовательности, указанной в подпунктах а) или б), где полипептид обладает тиосульфат-сульфотрансферазной активностью;
г) полипептид, имеющий аминокислотную последовательность, которая имеет делецию, замену, инсерцию и/или добавление от 1 до 45 аминокислотных остатков относительно аминокислотной последовательности, представленной в SEQ ID NO: 2, где полипептид обладает тиосульфат-сульфотрансферазной активностью.
Как отмечалось выше, полипептиды с активностью тиосульфат-сульфотрансферазы включают также варианты ферментов, указанных в подпунктах а) или б), аминокислотная последовательность которых идентична на 70% или более аминокислотной последовательности последовательностей, указанных в подпунктах а) или б), где вариант обладает тиосульфат-сульфотрансферазной активностью. Предпочтительными вариантами осуществления изобретения являются варианты, которые идентичны по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% описанным выше аминокислотным последовательностям, т.е. в которых по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% аминокислотных положений идентичны положениям в указанных выше аминокислотных последовательностях. Процент идентичности предпочтительно рассчитывают для всей длины аминокислотной или нуклеотидной области. Специалистам в данной области доступен ряд программ для сравнительного анализа последовательностей на основе большого количества алгоритмов. В этом плане наиболее надежные результаты дают алгоритмы Needleman и Wunsch или Smith и Waterman. Для сравнительного анализа первичной структуры последовательностей можно использовать программа PileUp (J. Mol. Evolution., 25, 1987, сс. 351-360, Higgins и др., CABIOS, 5, 1989, сс. 151-153) или программы Gap и BestFit [Needleman и Wunsch (J. Mol. Biol. 48, 1970, сс. 443-453) и Smith и Waterman (Adv. Appl. Math. 2, 1981, сс. 482-489)], которые входят в пакет программ GCG [Genetics Computer Group, 575 Science Drive, Мэдисон, шт.Висконсин, США 53711, 1991]. Указанные выше величины процентов, характеризующие идентичность последовательностей, для всей области последовательности предпочтительно рассчитывают с помощью программы GAP.
Кроме того, полипептиды с активностью тиосульфат-сульфотрансферазы включают также фрагменты ферментов, указанных в подпунктах а) или б). Эти фрагменты обладают описанной выше активностью. Фрагмент полипептида с активностью тиосульфат-сульфотрансферазы в контексте настоящего описания предпочтительно содержит по меньшей мере 30, по меньшей мере 50 или по меньшей мере 80 последовательно расположенных аминокислотных остатков одной из указанных выше аминокислотных последовательностей.
Как отмечалось выше, полипептиды с активностью тиосульфат-сульфотрансферазы включают также варианты ферментов, указанных в подпунктах а) или б), имеющие аминокислотную последовательность, которая имеет делецию, замену, инсерцию и/или добавление от 1 до 45 аминокислотных остатков относительно аминокислотной последовательности, представленной в SEQ ID NO: 2, где полипептид обладает тиосульфат-сульфотрансферазной активностью. В предпочтительных вариантах осуществления изобретения аминокислотная последовательность имеет делецию, замену, инсерцию и/или добавление от 1 до 40, более предпочтительно от 1 до 30, еще более предпочтительно от 1 до 20, предпочтительно от 1 до 15, еще более предпочтительно от 1 до 10 и наиболее предпочтительно от 1 до 5 аминокислотных остатков относительно аминокислотной последовательности, представленной в SEQ ID NO: 2.
В другом предпочтительном способе полипептид кодируется геном, который включает нуклеотидную последовательность SEQ ID NO: I, соответствующую последовательности дикого типа гена RDL2 из штамма Saccharomyces cerevisiae S288c.
Предпочтительно также полипептид может кодироваться геном, который включает нуклеотидную последовательность SEQ ID NO: 3. В этом случае наиболее часто встречающийся кодон в определенной степени адаптируют для E. coli.
В другом предпочтительном способе полипептид кодируется геном, который включает нуклеотидную последовательность SEQ ID NO: 4. В этом случае наиболее часто встречающийся кодон адаптируют для E. coli в существенно большей степени.
В другом предпочтительном способе полипептид кодируется геном, который включает нуклеотидную последовательность SEQ ID NO: 5. В этом случае наиболее часто встречающийся кодон полностью адаптируют для E. coli.
При осуществлении способа, предлагаемого в изобретении, экспрессию гена, кодирующего полипептид с активностью тиосульфат-сульфотрансферазы, предпочтительно повышают с помощью одного или нескольких следующих путей:
а) Экспрессию гена осуществляют под контролем промотора, который в применяемом для осуществления способа микроорганизме обеспечивает усиленную экспрессию по сравнению с исходным штаммом. Для этой цели можно применять конститутивный промотор GAPDH гена gapA Escherichia coli или индуцибельный промотор lac, tac, trc, lambda, ara или tet (обзор: Makrides SC. Micro-biol Rev. 60(3), сентябрь 1996 г., сс. 512-538).
б) Увеличивают количество копий гена, кодирующего полипептид с активностью тиосульфат-сульфотрансферазы, по сравнению с исходным штаммом. Для достижения этого можно, например, встраивать ген в плазмиды с увеличенным количеством копий, или интегрировать ген в несколько копий хромосомы микроорганизма (Baneyx F., Curr. Opin. Biotechnol. 10, 1999, сс. 411-421).
в) Экспрессию гена осуществляют с использованием сайта связывания рибосом, что приводит к усиленной трансляции в применяемом для осуществления способа микроорганизме по сравнению с исходным штаммом (Makrides S.C. Microbiol Rev., 60 (3), сентябрь 1996 г., сс. 512-538).
г) Усиливают экспрессию гена путем оптимизации наиболее часто встречающегося кодона гена применительно к используемому для осуществления способа микроорганизму (Welch, Villalobos, Gustafsson и Minshull, J R Soc Interface, 6, 2009, сс. 467-476). Например, индекс адаптации кодонов (CAI), можно применять в качестве меры адаптации наиболее часто встречающего кодона гена к организму (Sharp P.M., Li W.H., Nucleic Acids Res., 15(3), 1987, сс. 1281-1295).
д) Усиливают экспрессию гена путем снижения уровня вторичных структур мРНК в мРНК, транскрибируемой с гена (Kudia G., Murray A.W., Tollervey D., Plotkin J.B., Science, 10, 324(5924), апрель 2009 г., сс. 25525-8; Welch, Villalobos, Gustafsson и Minshull, J R Soc Interface, 6, 2009, сс. 467-476).
е) Усиливают экспрессию гена путем элиминации терминаторов для РНК-полимеразы в мРНК, транскрибируемой с гена (Welch, Villalobos, Gustafsson иа Mmshull, J R Soc Interface, 6, 2009, сс. 467-476).
ж) Осуществляют экспрессию гена с использованием стабилизирующих мРНК последовательностей в мРНК, транскрибируемой с гена (Carrier T.A., Keasling J.D., Biotechnol Prog., 13(6), ноябрь-декабрь 1997 г, сс. 699-708). В качестве примера последовательности, стабилизирующей мРНК гена metF E. coli, можно упомянуть последовательность ARNmstl7 (WO 2009/043803).
В другом варианте способа, предлагаемого в изобретении, предпочтительным является, чтобы соль дитиосерной кислоты представляла собой соль, выбранную из группы, включающей соль щелочного металла, соль щелочноземельного металла, соль аммония и их смеси, предпочтительно аммониевую соль.
Предпочтительно также, чтобы соль серной кислоты представляла собой соль, выбранную из группы, включающей соль щелочного металла, соль щелочноземельного металла, соль аммония (и их смеси), предпочтительно аммониевую соль.
Предпочтительно концентрация соли дитиосерной кислоты в среде или ферментативном бульоне составляет от 0,05 до 100 г/кг, предпочтительно от 0,1 до 20 г/кг и наиболее предпочтительно от 0,2 до 12 г/кг.
В предпочтительном варианте способа концентрацию соли дитиосерной кислоты в среде или в ферментационном бульоне в процессе ферментации поддерживают на уровне, составляющем по меньшей мере от 0,05 до 100 г/кг, предпочтительно от 0,1 до 20 г/кг и наиболее предпочтительно от 0,2 до 12 г/кг.
В другом предпочтительном варианте способа в процессе ферментации содержание соли дитиосерной кислоты в пересчете на общее содержание неорганической серы в среде и в ферментационном бульоне поддерживают на уровне, составляющем по меньшей мере 5 мол.%.
Способ, предлагаемый в изобретении, можно осуществлять в виде периодического процесса, периодического процесса с подпиткой, периодического процесса с повторной подпиткой и непрерывного процесса.
Кроме того, серусодержащую аминокислоту можно получать из ферментационного бульона в виде твердого продукта или в растворенной форме в виде жидкого продукта.
В предпочтительном способе, предлагаемом в настоящем изобретении, получаемая серусодержащая аминокислота представляет собой L-метионин.
Предпочтительно также, чтобы микроорганизм принадлежал к роду Escherichia, наиболее предпочтительно виду Escherichia coli.
В альтернативном способе микроорганизм принадлежит к роду Corynebacterium, наиболее предпочтительно виду Corynebacterium glutamicum.
В наиболее предпочтительном способе, предлагаемом в настоящем изобретении, микроорганизм выбирают из
- Corynebacterium glutamicum с повышенной активностью и/или уровнем экспрессии аспартаткиназы и с ослабленным по сравнению с исходным штаммом или с удаленным путем делеции регуляторным белком McbR;
- Escherichia coli с повышенной активностью и/или уровнем экспрессии аспартаткиназы и с ослабленным по сравнению с исходным штаммом или с удаленным путем делеции регуляторным белком MetJ.
Изобретение относится также к микроорганизму, выбранному из
- Corynebacterium glutamicum с повышенной активностью и/или уровнем экспрессии аспартаткиназы и с ослабленным по сравнению с исходным штаммом или с удаленным путем делеции регуляторным белком McbR;
- Escherichia coli с повышенной активностью и/или уровнем экспрессии аспартаткиназы и с ослабленным по сравнению с исходным штаммом или с удаленным путем делеции регуляторным белком MetJ;
где микроорганизм секретирует или продуцирует L-метионин и где микроорганизм обладает повышенной тиосульфат-сульфотрансферазной активностью по сравнению с исходным штаммом.
Изобретение относится также к микроорганизму, выбранному из
- Corynebacterium glutamicum с повышенной активностью и/или уровнем экспрессии аспартаткиназы и с ослабленным по сравнению с исходным штаммом или с удаленным путем делеции регуляторным белком McbR;
- Escherichia coli с повышенной активностью и/или уровнем экспрессии аспартаткиназы и с ослабленным по сравнению с исходным штаммом или с удаленным путем делеции регуляторным белком MetJ;
где микроорганизм секретирует или продуцирует L-метионин и где микроорганизм сверхэкспрессирует ген, кодирующий полипептид с активностью тиосульфат-сульфотрансферазы.
Штамм C. glutamicum, который секретирует или продуцирует L-метионин, предпочтительно обладает повышенной ферментативной активностью аспартаткиназы (КФ 2.7.2.4), при этом предпочтительными являются устойчивые к регуляции по типу обратной связи аллели. У представителей Corynebacterium указанная аспартаткиназа кодируется геном lysC. Из-за ослабления или делеции регуляторного белка McbR (который кодируется геном mcbR), имеет место также дополнительная утилизация серы. McbR является репрессором полного каскада утилизации серы у C. glutamicum (Rey D.A., Nentwich S.S., Koch D.J., Riickert C., Puhler A., Tauch A., Kalinowski J., Mol Microbiol., 56 (4), 2005, сс. 871-887).
Штамм E. coli, который секретирует или продуцирует L-метионин, предпочтительно обладает повышенной ферментативной активностью аспартаткиназы (КФ 2.7.2.4), при этом предпочтительными являются устойчивые к регуляции по типу обратной связи аллели. У E. coli присутствуют три различные аспартаткиназы, которые кодируются генами thrA, metL или lysC. Из-за ослабления или делеции регуляторного белка MetJ, который кодируется геном metJ, имеет место дополнительный биосинтез L-метионина. MetJ является основным репрессором биосинтеза L-метионина у E. coli.
Предпочтительным является также, если для указанного микроорганизма характерна сверхэкспрессия одного или нескольких гена(ов), кодирующего(их) полипептид с активностью тиосульфат-сульфотрансферазы, где полипептид с активностью тиосульфат-сульфотрансферазы можно выбирать из полипептидов, указанных в следующих подпунктах а)-г):
а) полипептид, состоящий из или содержащий полипептиды Rdl2p, GlpE, PspE, YgaP, ThiI, YbbB, SseA, YnjE, YceA, YibN, NCg10671, NCg11369, NCg12616, NCg10053, NCg10054, NCg12678, NCg12890; тиосульфат-сульфотрансферазу млекопитающих, например, тиосульфат-сульфотрансферазу из бычьей печени (Bos taurus); предпочтительно Rdl2p, GlpE, PspE и наиболее предпочтительно Rdl2p;
б) полипептид, состоящий из или содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2;
в) полипептид, аминокислотная последовательность которого идентична на 70% или более аминокислотной последовательности, указанной в подпунктах а) или б), где полипептид обладает тиосульфат-сульфотрансферазной активностью;
г) полипептид, имеющий аминокислотную последовательность, которая несет делецию, замену инсерцию и/или добавление от 1 до 45 аминокислотных остатков относительно аминокислотной последовательности, представленной в SEQ ID NO: 2, где полипептид обладает тиосульфат-сульфотрансферазной активностью.
Предпочтительно также исходный штамм микроорганизма выводят из группы, включающей Escherichia coli MG1655, Escherichia coli W3110, Escherichia coli DH5a, Escherichia coli DH10B, Escherichia coli BW2952, Escherichia coli REL606, Corynebacterium glutamicum ATCC13032, Corynebacterium glutamicum R, Corynebacterium glutamicum DSM20411 (прежнее название Brevibacterium flavum), Corynebacterium glutamicum DSM20412 (прежнее название Brevibacterium lactofermentum), Corynebacterium glutamicum DSM1412 (прежнее название Brevibacterium lactofermentum), Corynebacterium efficiens YS-314T (=DSM44549), Corynebacterium glutamicum ATCC21608, Corynebacterium glutamicum DSM17322.
Другим штаммом, который секретирует или продуцирует L-метионин, является, например, продуктивный штамм E. coli MG1655 AmetJ Ptrc-metH Ptrc-metF PtrcF-cysPUWAM PtrcF-cysJIH Ptrc09-gcvTHP, содержащий продуктивную плазмиду pME101-thrA*1-cysE-Pgap-metA*11 (WO 2009/043803).
Клонирование продуктивного штамма E. coli MG1655AmetJ Ptrc-metH Ptrc-metF PtrcF-cysPUWAM PtrcF-cysJIH Ptrc09-gcvTHP описано в заявке на патент WO 2009/043803. Основой указанного штамма является штамм дикого типа E. coli K12 MG1655. В геном указанного штамма были интродуцированы следующие модификации:
- Удаляли путем делеции ген репрессора биосинтеза L-метионина metJ.
- Против хода транскрипции относительно гена metH (кодирует кобаламинзависимую метионинсинтазу), встраивали сильный промотор trc.
- Против хода транскрипции относительно гена metF (кодирует 5,10-метилентетрагидрофолатредуктазу), встраивали сильный промотор trc.
- Против хода транскрипции относительно оперона cysPUWAM встраивали сильный промотор trcF. cysPUWA кодирует транспортер поглощения сульфата/тиосульфата. cysM кодирует цистеинсинтазу B.
- Против хода транскрипции относительно оперона cysJIH встраивали сильный промотор trcF. cysJI кодирует сульфитредуктазу, a cysH кодирует 3'-фосфоаденилил-сульфатредуктазу.
- Против хода транскрипции относительно оперона gcvTHP встраивали сильный промотор trcO. gcvT, gcvH и gcvP кодируют три компонента системы расщепления глицина.
Клонирование в E. coli продуктивной плазмиды pME101-thrA*1-cysE-Pgap-metA*11 описано в заявке на патент WO 2007/077041. Основой этой плазмиды с малым количеством копий (малокопийная плазмида) являлся вектор pCL1920 (Lerner C.G. и Inouye M., Nucl. Acids Res. 18, 1990, с.4631 [PMID: 2201955]). Незагруженная (пустая) плазмида pMe101 несет ген lacIq, который кодирует высокоэкспрессируемый аллель репрессора lac. Ген thra*1 клонировали по ходу транскрипции относительно сильного промотора trc, который может подавляться репрессором Lac. Он кодирует устойчивый к регуляции по типу обратной связи вариант аспартаткиназы /гомосериндегидрогеназы ThrA из E. coli. В той же ориентации за ним расположен ген cysE вместе с его встречающимся в естественных условиях промотором. Он кодирует серинацетилтрансферазу из E. coli. По ходу транскрипции относительно cysE расположен сильный промотор gapA из E. coli, который контролирует экспрессию гена metA*11. Ген metA*11 кодирует устойчивый к регуляции по типу обратной связи вариант гомосерин-O-сукцинилтрансферазы из E. coli.
В качестве примеров других микроорганизмов, которые секретируют или продуцируют L-метионин, можно упомянуть следующие штаммы:
- C. glutamicum Ml 179 (DSM17322) (WO 2007/011939)
- E. coli TF4076BJF metA#10 + metYX (Lm) (WO 2008/127240; с 46);
- E. coli W3110ΔJ/pKP451 (EP 1445 31 OBI, с.7, пример 4);
- E. coli WΔthrBCΔmetJmetK32 pMWPthrmetA4Δ569 (Yoshihiro Usuda и Osamu Kurahashi, Applied and Environmental Microbiology, т.71, №6, 2005, сс. 3228-3234);
- W3HO/pHC34 (WO 01/27307 с.13, пример 3).
Дополнительные примеры различных пригодных микроорганизмов описаны у Gomes и др.. Enzyme and Microbial Technology 37, 2005, сс. 3-18.
В других предпочтительных вариантах осуществления бактерии, которые продуцируют L-метионин, обладают одним или несколькими признаками, выбранными из группы:
1) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов транспортной системы тиосульфата/сульфата CysPUWA (КФ 3.6.3.25),
2) способность к сверхэкспрессии полинуклеотида, который кодирует 3'-фосфоаденозин-5'фосфосульфатредуктазу CysH (КФ 1.8.4.8),
3) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов сульфитредуктазы CysJI (КФ 1.8.1.2),
4) способность к сверхэкспрессии полинуклеотида, который кодирует цистеинсинтазу A CysK (КФ 2.5.1.47),
5) способность к сверхэкспрессии полинуклеотида, который кодирует цистеинсинтазу В CysM (КФ 2.5.1.47),
6) способность к сверхэкспрессии полинуклеотида, который кодирует серинацетилтрансферазу CysE (КФ 2.3.1.30),
7) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов системы расщепления глицина GcvTHP-Lpd (КФ 2.1.2.10, КФ 1.4.4.2, КФ 1.8.1.4),
8) способность к сверхэкспрессии полинуклеотида, который кодирует липоилсинтазу LipA (КФС 2.8.1.8),
9) способность к сверхэкспрессии полинуклеотида, который кодирует липоилпротеинлигазу LipB (КФ 2.3.1.181),
10) способность к сверхэкспрессии полинуклеотида, который кодирует фосфоглицератдегидрогеназу SerA (КФ 1.1.1.95),
11) способность к сверхэкспрессии полинуклеотида, который кодирует 3-фосфосеринфосфатазу SerB (КФС 3.1.3.3),
12) способность к сверхэкспрессии полинуклеотида, который кодирует 3-фосфосерин/фосфогидрокситреонин-аминотрансферазу SerC (КФ 2.6.1.52),
13) способность к сверхэкспрессии полинуклеотида, который кодирует серингидроксиметилтрансферазу GlyA (EC 2.1.2.1),
14) способность к сверхэкспрессии полинуклеотида, который кодирует аспартокиназу I и гомосериндегидрогеназу I ThrA (КФ 2.7.2.4, КФ 1.1.1.3),
15) способность к сверхэкспрессии полинуклеотида, который кодирует аспартаткиназу LysC (КФ 2.7.2.4),
16) способность к сверхэкспрессии полинуклеотида, который кодирует гомосериндегидрогеназу Нот (КФ 1.1.1.3),
17) способность к сверхэкспрессии полинуклеотида, который кодирует гомосерин-0-ацетилтрансферазу MetX (КФ 2.3.1.31),
18) способность к сверхэкспрессии полинуклеотида, который кодирует гомосерин-0-сукцинилтрансферазу MetA (КФ 2.3.1.46),
19) способность к сверхэкспрессии полинуклеотида, который кодирует цистатионин-гамма-синтазу MetB (КФ 2.5.1.48),
20) способность к сверхэкспрессии полинуклеотида, который кодирует β-С-S-лиазу AecD (КФ 4.4.1.8, обозначают также как бета-лиаза),
21) способность к сверхэкспрессии полинуклеотида, который кодирует цистатионин-бета-лиазу MetC (КФ 4.4.1.8),
22) способность к сверхэкспрессии полинуклеотида, который кодирует В12-независимую гомоцистеин-3-метилтрансферазу MetE (КФ 2.1.1.14),
23) способность к сверхэкспрессии полинуклеотида, который кодирует В12-зависимую гомоцистеин-8-метилтрансферазу MetH (КФ 2.1.1.13),
24) способность к сверхэкспрессии полинуклеотида, который кодирует метилентетрагидрофолатредуктазуМе1Р (КФ 1.5.1.20),
25) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов экспортера L-метионина BrnFE из Corynebacterium glutamicum,
26) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов экспортера валина YgaZH из Escherichia coli (b2682, b2683),
27) способность к сверхэкспрессии полинуклеотида, который кодирует предполагаемый транспортер YjeH из Escherichia coli (b4141),
28) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов пиридин-нуклеотид-трансгидрогеназы PntAB (КФ 1.6.1.2),
29) способность к сверхэкспрессии полинуклеотида, который кодирует 0-сукцинилгомосеринсульфгидрилазу MetZ (КФ 2.5.1.48),
30) способность к сверхэкспрессии полинуклеотида, который кодирует фосфоенолпируваткарбоксилазу Рус (КФ 4.1.1.31).
В контексте настоящего описания предпочтительными признаками является(ются) один или несколько признаков, выбранных из группы:
1) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов транспортной системы тиосульфата/сульфата CysPUWA (КФ 3.6.3.25),
2) способность к сверхэкспрессии полинуклеотида, который кодирует 3'-фосфоаденозин-5'-фосфосульфатредуктазу CysH (КФ 1.8.4.8),
3) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов сульфитредуктазы CysJI (КФ 1.8.1.2),
4) способность к сверхэкспрессии полинуклеотида, который кодирует цистеинсинтазу A CysK (КФ 2.5.1.47),
5) способность к сверхэкспрессии полинуклеотида, который кодирует цистеинсинтазу В CysM (КФ 2.5.1.47),
6) способность к сверхэкспрессии полинуклеотида, который кодирует серинацетилтрансферазу CysE (КФ 2.3.1.30),
7) способность к сверхэкспрессии полинуклеотида, который кодирует один или несколько компонентов системы расщепления глицина GcvTHP-Lpd (КФ 2.1.2.10, КФ 1.4.4.2, КФ 1.8.1.4),
8) способность к сверхэкспрессии полинуклеотида, который кодирует липоилсинтазу LipA (КФС 2.8.1.8),
9) способность к сверхэкспрессии полинуклеотида, который кодирует липоилпротеинлигазу LipB (КФ 2.3.1.181),
10) способность к сверхэкспрессии полинуклеотида, который кодирует фосфоглицератдегидрогеназу SerA (КФ 1.1.1.95),
11) способность к сверхэкспрессии полинуклеотида, который кодирует 3-фосфосеринфосфатазу SerB (КФС 3.1.3.3),
12) способность к сверхэкспрессии полинуклеотида, который кодирует 3-фосфосерин/фосфогидрокситреонин-аминотрансферазу SerC (КФ 2.6.1.52),
13) способность к сверхэкспрессии полинуклеотида, который кодирует серингидроксиметилтрансферазу GlyA (EC 2.1.2.1),
14) способность к сверхэкспрессии полинуклеотида, который кодирует аспартокиназу I и гомосериндегидрогеназу I ThrA (КФ 2.7.2.4, КФ 1.1.1.3),
15) способность к сверхэкспрессии полинуклеотида, который кодирует аспартаткиназу LysC (КФ 2.7.2.4),
16) способность к сверхэкспрессии полинуклеотида, который кодирует гомосериндегидрогеназу Нот (КФ 1.1.1.3),
17) способность к сверхэкспрессии полинуклеотида, который кодирует гомосерин-O-ацетилтрансферазуМе1Х (КФ 2.3.1.31),
18) способность к сверхэкспрессии полинуклеотида, который кодирует гомосерин-O-сукцинилтрансферазу MetA (КФ 2.3.1.46),
19) способность к сверхэкспрессии полинуклеотида, который кодирует цистатионин-гамма-синтазу MetB (КФ 2.5.1.48),
20) способность к сверхэкспрессии полинуклеотида, который кодирует β-С-S-лиазу AecD (КФ 4.4.1.8, обозначают также как бета-лиаза),
21) способность к сверхэкспрессии полинуклеотида, который кодирует цистатионин-бета-лиазу MetC (КФ 4.4.1.8),
22) способность к сверхэкспрессии полинуклеотида, который кодирует B12-независимую гомоцистеин-S-метилтрансферазу MetE (КФ 2.1.1.14),
23) способность к сверхэкспрессии полинуклеотида, который кодирует B12-зависимую гомоцистеин-3-метилтрансферазу MetH (КФ 2.1.1.13),
24) способность к сверхэкспрессии полинуклеотида, который кодирует метилентетрагидрофолатредуктазу MetF (КФ 1.5.1.20).
В контексте настоящего описания наиболее предпочтительными признаками являются признаки, выбранные из группы:
1) способность к сверхэкспрессии полинуклеотида, который кодирует аспартокиназу I и гомосериндегидрогеназу I ThrA (КФ 2.7.2.4, КФ 1.1.1.3),
2) способность к сверхэкспрессии полинуклеотида, который кодирует аспартаткиназу LysC (КФ 2.7.2.4),
3) способность к сверхэкспрессии полинуклеотида, который кодирует гомосериндегидрогеназу Нот (КФ 1.1.1.3),
4) способность к сверхэкспрессии полинуклеотида, который кодирует гомосерин-O-ацетилтрансферазу MetX (КФ 2.3.1.31),
5) способность к сверхэкспрессии полинуклеотида, который кодирует гомосерин-O-сукцинилтрансферазу MetA (КФ 2.3.1.46),
6) способность к сверхэкспрессии полинуклеотида, который кодирует цистатионин-гамма-синтазу MetB (КФ 2.5.1.48),
7) способность к сверхэкспрессии полинуклеотида, который кодирует β-С-S-лиазу AecD (КФ 4.4.1.8, обозначают также как бета-лиаза),
8) способность к сверхэкспрессии полинуклеотида, который кодирует цистатионин-бета-лиазу MetC (КФ 4.4.1.8),
9) способность к сверхэкспрессии полинуклеотида, который кодирует B12-независимую гомоцистеин-8-метилтрансферазу MetE (КФ 2.1.1.14),
10) способность к сверхэкспрессии полинуклеотида, который кодирует B12-зависимую гомоцистеин-3-метилтрансферазу MetH (КФ 2.1.1.13),
11) способность к сверхэкспрессии полинуклеотида, который кодирует метилентетрагидрофолатредуктазу MetF (КФ 1.5.1.20).
Для повышения производства L-метионина в C. glutamicum может оказаться целесообразным ослаблять один или несколько генов, выбранных из группы:
а) ген pgi, кодирующий глюкозо-6-фосфатизомеразу (Pgi, КФ 5.3.1.9),
б) ген thrB, кодирующий гомосеринкиназу (ThrB, КФ 2.7.1.39),
в) ген, metK, кодирующий S-аденозилметионинсинтазу (MetK, КФ 2.5.1.6),
г) ген dapA, кодирующий дигидродипиколинатсинтазу (DapA, КФ 4.2.1.52),
д) ген pck, кодирующий фосфоенолпируваткарбоксикиназу (Pck, КФ
4.1.1.49),
е) ген cg3086, кодирующий цистатионин-γ-лиазу (Cg3086, КФ 4.4.1.1),
ж) ген cg2344, кодирующий цистатионин-β-синтазу (Cg2344, КФ 4.2.1.22),
з) ген cg3031, кодирующий регуляторный белок Cg3031,
и) ген mcbR, кодирующий регулятор транскрипции при биосинтезе L-метионина (McbR),
к) ген metQ, кодирующий субъединицу транспортера L-метионина (MetQNI),
л) ген metN, кодирующий субъединицу транспортера L-метионина (MetQNI),
м) ген metl, кодирующий субъединицу транспортера L-метионина (MetQNI),
н) ген metP, кодирующий транспортер L-метионина (MetP).
Для повышения производства L-метионина в E. coli может оказаться целесообразным ослаблять один или несколько генов, выбранных из группы:
а) ген metJ (b3938, ECK3930), кодирующий регулятор транскрипции при биосинтезе L-метионина (MetJ),
б) ген pgi (b4025, ECK4017), кодирующий глюкозо-6-фосфатизомеразу (Pgi, КФ. 5.3.1.9),
в) ген thrB (b0003, ECK0003), кодирующий гомосеринкиназу (ThrB, КФ
2.7.1.39),
г) ген metK (b2942, ECK2937), кодирующий S-аденозилметионинсинтазу (MetK, КФ. 2.5.1.6),
д) ген dapA (b2478, ECK2474), кодирующий дигидродипиколинатсинтазу (DapA, КФ 4.2.1.52),
е) ген pck (b3403, ECK3390), кодирующий фосфоенолпируваткарбоксикиназу (Pck, КФ. 4.1.1.49),
ж) ген purU (b1232, ECK1227), кодирующий формилтетрагидрофолатгидролазу (PurU, КФ 3.5.1.10),
з) ген pykA (b1854, ECK1855), кодирующий пируваткиназу II (PykA, КФ 2.7.1.40),
и) ген pykF (bl676, ECK1672), кодирующий пируваткиназу I (PykF, КФ 2.7.1.40),
к) ген metQ (b0197, ECK0197), кодирующий субъединицу транспортера L-метионина (MetQNI),
л) ген metI (b0198, ECK0198), кодирующий субъединицу транспортера L-метионина (MetQNI),
м) ген metN (b0199, ECK0199), кодирующий субъединицу транспортера L-метионина (MetQNI),
н) ген dcd (b2065, ECK2059), кодирующий дезоксицитидин-5'-трифосфатдеаминазу (Dcd, КФ. 3.5.4.13),
о) ген yncA (b1448, ECK1442), кодирующий предполагаемую N-ацилтрансферазу (YncA, Metabolic Explorer (метаболический зонд) WO 2010/020681),
п) ген fnrS (b4699, ECK4511) кодирующий регуляторную тРНК FnrS.
Подробное описание изобретения
Изобретение относится к способу ферментативного получения серусодержащих аминокислот, выбранных из группы, включающей L-метионин, L-цистеин, L-цистин, L-гомоцистеин и L-гомоцистин. Способ осуществляют с использованием микроорганизма из семейства Enterobacteriaceae или микроорганизма из семейства Corynebacteriaceae, в котором происходит сверхэкспрессия гена, кодирующего полипептид, который обладает ферментативной активностью тиосульфат-сульфотрансферазы.
В контексте настоящего описания понятие «ген» относится к сегменту дезоксирибонуклеиновой кислоты (ДНК), который содержит информацию, необходимую для производства (транскрипции) прежде всего рибонуклеиновой кислоты (РНК), и она содержит информацию, необходимую для производства (трансляции) белка (полипептида), а именно, в контексте настоящего описания полипептида с активностью тиосульфат-сульфотрансферазы. Тот факт, что ген или полинуклеотид содержит информацию, необходимую для производства белка, называют также кодированием белка или полипептида геном или РНК. Под эндогенными генами или полинуклеотидами подразумеваются открытые рамки считывания (ОРС), гены или аллели или полинуклеотиды, присутствующие в популяции вида. Понятие «ген» и «ОРС» (открытая рамка считывания) в настоящем изобретении используют в качестве синонимов.
Понятие «полинуклеотид» в целом относится к полирибонуклеотидам и полидезоксирибонуклеотидам, которые могут представлять собой немодифицированную РНК или ДНК или модифицированную РНК или ДНК.
Понятие «полипептид» относится к пептидам или белкам, которые содержат две или большее количество аминокислот, сцепленных помощью пептидных связей. Понятие полипептид и белок используют в качестве синонимов. Белки принадлежат к основным единицам всех клеток. Они не только обеспечивают структуру клетки, но представляют собой молекулярные «устройства», которые транспортируют субстанции, катализируют химические реакции и распознают сигнальные субстанции.
Под «протеиногенными аминокислотами» подразумеваются аминокислоты, которые встречаются в природных белках, т.е. в белках микроорганизмов, растений, животных и человека. Они включают, в частности, L-аминокислоты, выбранные из группы, включающей L-аспарагиновую кислоту, L-аспарагин, L-треонин, L-серин, L-глутаминовую кислоту, L-глутамин, глицин, L-аланин, L-цистеин, L-валин, L-метионин, L-изолейцин, L-лейцин, L-тирозин, L-фенилаланин, L-гистидин, L-лизин, L-триптофан, L-пролин и L-аргинин, а также селоноцистеин. В этом контексте протеиногенные аминокислоты всегда представляют собой α-аминокислоты. За исключением аминокислоты глицина, у всех протеиногенных аминокислот α-углеродный атом является асимметричным (т.е. молекулы являются хиральными): существует два энантиомера каждой из этих аминокислот. В этом контексте только один из двух энантиомеров является протеиногенным, и он фактически представляет собой L-аминокислоту: аппарат, необходимый для построения белков - рибосома, тРНК, аминоацил-тРНК-синтетаза (которая загружает тРНК аминокислотами) и другие компоненты сами также обладают хиральностью и могут распознавать только L-варианты.
Понятие «генная экспрессия» (для краткости «экспрессия») в целом относится к экспрессии генетической информации с образованием фенотипа. В узком смысле генная экспрессия обозначает транскрипцию гена с образованием РНК и последующую трансляцию РНК с образованием полипептида, который может обладать ферментативной активностью.
Понятие «сверхэкспрессия» в целом относится к увеличению внутриклеточной концентрации или активности рибонуклеиновой кислоты, белка или фермента по сравнению с исходным штаммом (родительский штамм) или штаммом дикого типа. В контексте настоящего изобретения имеет место сверхэкспрессия генов или полинуклеотидов тиосульфат-сульфотрансферазы, которые кодируют полипептид с тиосульфат-сульфотрансферазной активностью.
Под «исходным штаммом» (родительским штаммом» подразумевается штамм микроорганизма, на который оказывают воздействия, обеспечивающие увеличение продуктивности одной/одного или нескольких аминокислот, пептидов или белков, или оказывают воздействия, обеспечивающие активность одного или нескольких ферментов (например, воздействия, приводящие к сверхэкспрессии). Исходный штамм может представлять собой штамм дикого типа, но также штамм, который уже ранее был предварительно модифицирован (например, продуктивный штамм (штамм-продуцент)).
«Дикий тип» клетки предпочтительно относится к клетке, геном которой находится в том состоянии, в котором он сформировался в природе в процессе эволюции. Понятие применяют как к целой клетке, так и к индивидуальным генам. Таким образом, те конкретные клетки или те гены, генные последовательности которых были по меньшей мере частично модифицированы человеком с использованием методов рекомбинации, не подпадают под понятие «дикий тип».
Мутанты, полученные согласно настоящему изобретению, отличаются повышенной секрецией или производством требуемой аминокислоты в процессе ферментации по сравнению с применяемым исходным штаммом или родительским штаммом. В этом контексте аминокислоты высвобождаются в среду, окружающую их или накапливаются внутри клетки (аккумуляция).
Понятие «усиление» или «повышенная активность» в этом контексте относится к повышению внутриклеточной ферментативной активности одного или нескольких ферментов в микроорганизме, которые кодируются соответствующей ДНК.
В принципе, повышения ферментативной активности можно достигать, например, путем увеличения количества копий генной последовательности или генных последовательностей, которая(ые) кодирует(ют) фермент, путем использования сильного промотора или использования гена или аллеля, который кодирует соответствующий фермент, обладающий повышенной активностью, и необязательно, объединяя эти пути. Клетки, генетически модифицированные согласно изобретению, получают, например, путем трансформации, трансдукции, конъюгации или объединения этих методов, с использованием вектора, который содержит требуемый ген, аллель этого гена или его часть, и вектора, который обеспечивает возможность экспрессии гена. Для достижения гетерологичной экспрессии, в частности, интегрируют ген или аллели в хромосому клетки или в реплицирующийся вне хромосомы вектор.
Обзор возможностей повышения ферментативной активности в клетках, например в случае пируваткарбоксилазы, представлен в DE-A-10031999, который включен в настоящее описание в качестве ссылки, и содержание которого в части, касающейся возможностей повышения ферментативной активности в клетках, включено в качестве составной части в описание настоящего изобретения.
Повышения ферментативной активности можно достигать, например, путем увеличения количества копий соответствующих полинуклеотидов внутри хромосомы или вне хромосомы по меньшей мере на одну копию.
Широко применяемый метод увеличения количества копий заключается в том, что встраивают соответствующий полинуклеотид в вектор, предпочтительно плазмиду, которая реплицируется в бактерии.
Приемлемыми плазмидными векторами для Enterobacteriaceae являются, например, клонирующие векторы, выведенные из pACYC184 (Bartolome и др., Gene 102, 1991, сс. 75-78), pTrc99A (Amann и др.. Gene 69, 1988, сс. 301-315), или производные pSC101 (Vocke и Bastia; Proceedings of the National Academy of Sciences USA 80(21), 1983, сс. 6557-6561). Предпочтительными являются также плазмиды, выведенные из pCL1920 (Lerner C.G. и Inouye M., Nucl. Acids Res., 18, 1990, с.4631 [PMID: 2201955]). Для увеличения количества копий соответствующих полинуклеотидов в E. coli. можно применять также плазмидные векторы, выведенные из бактериальных искусственных хромосом (ВАС), такие, например, как pCC1BAC (фирма EPICENTRE Biotechnologies, Мэдисон, США).
Приемлемыми плазмидными векторами для C. glutamicum являются, например, pZ1 (Menkel и др.. Applied and Environmental Microbiology 64, 1989, сс. 549-554), pEKExl (Eikmanns и др.. Gene 107, 1991, сс. 69-74) или pHS2-1 (Sonnen и др.. Gene 107, 1991, сс. 69-74). Их основой являются криптические плазмиды рНМ1519, pBL1 или pGA1. Аналогично этому можно применять другие плазмидные векторы, такие, например, как векторы, основой которых являются pCG4 (US 4489160) или pNG2 (Serwold-Davis и др., FEMS Microbiology Letters 66, 1990, сс. 119-124) или pAGI (US 5158891). В качестве обзорной статьи, в которой описаны плазмиды для C. glutamicum, см. Tauch и др., Journal of Biotechnology 104, 2003, сс. 27-40).
В качестве векторов можно применять также транспозоны, инсерционные элементы (IS-элементы) или фаги. Указанные генетические системы представлены, например, в описаниях патентов US 4822738, US 5804414 и US 5804414. Аналогично этому можно применять IS-элемент ISaBI, описанный в WO 92/02627, или транспозон Tn 45 плазмиды pXZ10142 (в качестве ссылки см. «Handbook of Corynebacterium glutamicum», под ред. L. Eggeling и M. Bott).
Другим широко применяемым методом достижения сверхэкспрессии является метод амплификации генов в хромосомах. Согласно этому методу по меньшей мере одну дополнительную копию представляющего интерес полинуклеотида встраивают в хромосому бактерии. Указанные методы амплификации описаны, например, в WO 03/014330 или WO 03/040373.
Еще один метод достижения сверхэкспрессии предусматривает функциональное соединение (осуществление функциональной связи) соответствующего гена или аллеля с промотором или кассетой экспрессии. Приемлемые промоторы для C. glutamicum представлены, например, на фиг.1 обзорной статьи Patek и др., Journal of Biotechnology 104 (1-3), 2003, сс. 311-323. Аналогично этому можно применять варианты промотора dapA, например, промотор А25, описанный у Vasicova и др.. Journal of Bacteriology 181, 1999, сс. 6188-6191. Можно применять также промотор gap C. glutamicum (EP 06007373).
Для Е. colt известны, например, промоторы Т3, Т7, SP6, М13, lac, tac и trc, которые описаны у Amann и др., Gene 69 (2), 1988, сс. 301-315 и у Amann и Brosius, Gene 40 (2-3), 1985, сс. 183-190, некоторые из которых можно применять также для C. glutamicum. Указанный промотор можно встраивать, например, против хода транскрипции относительно рассматриваемого гена, как правило, на расстоянии примерно 1-500 нуклеотидных оснований от стартового кодона. В US 5939307 описано, что для повышения экспрессии можно применять включение кассет экспрессии или промоторов, таких, например, как промотор tac, промотор trp, промотор lpp или промотор PL и промотор PR фага X, например, против хода транскрипции относительно хромосомного оперона треонина. Аналогично этому можно применять промоторы фага Т7, промотор gear-box или промотор nar. Указанные кассеты экспрессии или промоторы можно применять также для сверхэкспрессии связанных с плазмидами генов согласно методу, описанному в EP 0593792. С использованием аллеля lacIQ, в свою очередь, можно контролировать экспрессию связанных с плазмидами генов (Glascock и Weickert, Gene 223, 1998, сс. 221-231). Можно также для повышения активности промоторов применять модификацию их последовательности посредством одного или нескольких изменений нуклеотидов с помощью инсерции(ий) и/или делеции(ий).
С помощью путей, обеспечивающих сверхэкспрессию, активность или концентрацию соответствующего полипептида, как правило, повышают по меньшей мере на 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500%, максимум вплоть до 1000% или 2000% относительно активности или концентрации полипептида в соответствующем исходном штамме (микроорганизм дикого типа или исходный) до воздействия на него путей, приводящих к сверхэкспрессии.
Методы определения ферментативной активности тиосульфат-сульфотрансфераз описаны, например, у Cheng H., Donahue J.L., Battle S.E., Ray W.K., Larson T.J., Open Microbiol J., 2, 2008, сс. 18-28 и у Alexander К., Volini M., J. Biol. Chem., 262, 1987, сс. 6595-6604.
Экспрессию вышеуказанных ферментов или генов можно выявлять с помощью 1-й 2-мерного разделения белков в геле и последующего определения оптическими методами концентрации белка в геле с использованием соответствующей предназначенной для оценки программы. Если повышение ферментативной активности базируется только на повышении уровня экспрессии соответствующего гена, то повышение ферментативной активности можно оценивать количественно просто путем сравнения результатов 1- или 2-мерного разделения белков в клетке дикого типа и генномодифицированной клетки. Общепринятый метод получения гелей для разделения белков в случае бактерий и идентификации белков представляет собой метод, описанный у Hermann и др., Electrophoresis, 22, 2001, сс. 1712-1723. Концентрацию белка можно определять также путем анализа методом вестерн-блоттинга, осуществляя гибридизацию с антителом, специфическим в отношении подлежащего определению белка (Sambrook и др.. Molecular Cloning: a laboratory manual, 2-ое изд., изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY USA, 1989), и последующего определения концентрации оптическими методами с использованием соответствующей предназначенной для оценки концентрации программы (Lohaus и Meyer, Biospektrum, 5, 1989, сс. 32-39; Lottspeich, Angewandte Chemie 111, 1999, cc. 2630-2647). Активность ДНК-связывающих белков можно измерять путем анализа сдвига полос ДНК (который называют также методом гель-ретардации; Wilson и др.. Journal of Bacteriology, 183, 2001, cc. 2151-2155). Воздействие ДНК-связывающих белков на экспрессию других генов можно определять с помощью различных подробно описанных методов анализа с использованием репортерного гена (Sambrook и др.. Molecular Cloning:
a laboratory manual, 2-е изд., изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY USA, 1989). Внутриклеточную ферментативную активность можно определять различными известными из литературы методами (Donahue и др., Journal of Bacteriology 182 (19), 2000, cc. 5624-5627; Ray и др., Journal of Bacteriology 182 (8), 2000, cc. 2277-2284; Freedberg и др., Journal of Bacteriology 115 (3), 1973, cc. 816-823). Если ниже не указаны конкретные методы определения активности конкретного фермента, то определение повышения ферментативной активности, а также определение снижения ферментативной активности предпочтительно осуществляют с помощью методов, описанных у Hermann и др., Electophoresis, 22, 2001, cc. 1712-1723 (2001), Lohaus и др., Biospektrum 5, 1998, cc. 32-39, Lottspeich, Angewandte Chemie 111, 1999, cc. 2630-2647 и Wilson и др.. Journal of Bacteriology 183, 2001, cc. 2151-2155).
Если повышение ферментативной активности осуществляют посредством мутации эндогенного гена, то указанные мутации можно создавать либо случайным образом с использованием общепринятых методов, таких, например, как УФ-облучение, или с использованием индуцирующих мутации химических соединений, либо целенаправленно с помощью методов генной инженерии, таких как делеция(и), инсерция(и) и/или нуклеотидная(ые) замена(ы). Путем осуществления таких мутаций получают генетически модифицированные клетки. Конкретными предпочтительными мутантами ферментов являются, в частности, ферменты, которые более не могут ингибироваться по типу обратной связи или могут ингибироваться по типу обратной связи по крайней мере в меньшей степени, чем фермент дикого типа.
Если повышение ферментативной активности осуществляют путем повышения уровня экспрессии фермента, то для этого увеличивают количество копий, например, соответствующих генов, или изменяют посредством мутации промотор и регуляторную область или сайт связывания рибосом против хода транскрипции относительно структурного гена. Кассеты экспрессии, которые встраивают против хода транскрипции относительно структурного гена, действуют аналогичным образом. С помощью индуцибельного промотора можно дополнительно повышать уровень экспрессии в любой требуемый момент времени. Кроме того, можно создавать также так называемые энхансеры для гена фермента в качестве регуляторных последовательностей, которые также могут обусловливать повышение уровня экспрессии гена путем улучшения взаимодействия между РНК-полимеразой и ДНК. Аналогично этому уровень экспрессии можно повышать путем пролонгирования жизни мРНК. Кроме того, ферментативную активность можно повышать также, предупреждая расщепление белка фермента. В этом контексте гены или генные конструкции либо присутствуют в плазмидах с другим количеством копий, либо их интегрируют в хромосому и амплифицируют.Альтернативно этому, для достижения сверхэкспрессии рассматриваемых генов можно модифицировать также состав сред и процедуру культивирования. Инструкции по осуществлению модификации специалист в данной области может среди прочего почерпнуть у Martin и др,. Bio/Technology 5, 1987, сс. 137-146, Guerrero и др., Gene 138, 1994, сс. 35-41, Tsuchiya и Morinaga, Bio/Technology 6, 1988, сс. 428-430, Eikmanns и др.. Gene 102, 1991, сс. 93-98, в ЕР-А-0472869, в US 4601893, у Schwarzer и Puhler, Bio/Technology 9, 1991, сс. 84-87, Reinscheid и др., Applied and Environmental Microbiology 60, 1994, сс. 126-132, LaBarre и др., Journal of Bacteriology 175, 1993, сс. 1001-1007, в WO-A-96/15246, у Malumbres и др.. Gene 134, 1993, сс. 15-24, в JP-А-10-229891, у Jensen и Hammer, Biotechnology and Bioengineering 58, 1998, сс. 191-195 и в известных руководствах по генетике и молекулярной биологии. Описанные выше подходы, также как и мутации, приводят к получению генетически модифицированных клеток.
Можно применять также те плазмидные векторы, с помощью которых можно осуществлять метод генной амплификации путем интеграции в хромосому, что описано, например, у Reinscheid и др.. Applied and Environmental Microbiology 60, 1994, сс. 126-132), для дупликации или амплификации оперона hom-thrB. Согласно этому методу полный ген клонируют в плазмидном векторе, который может реплицироваться в хозяине (как правило, в Escherichia coli), но не в Corynebacterium glutamicum. Приемлемые векторы представляют собой, например, pSUP301 (Simon и др., Bio/Technology 1, 1983, сс. 784-791), pK18mob или pK19mob (Schäfer и др.. Gene 145, 1994, сс. 69-73), pGEM-T (фирма Promega Corporation, Мэдисон, шт.Висконсин, США, pCR2.1-TOPO (Shuman, Journal of Biological Chemistry 269, 1994, сс. 32678-32684), pCR-BluntII-TOPO (фирма Invitrogen, Гронинген, Нидерланды), pEM1 (SchrumpiH др., Journal of Bacteriology 173, сс. 4510-4516) или pBGS8 (Spratt и др., Gene 41, 1986, сс. 337-342). Затем плазмидный вектор, который содержит подлежащий амплификации ген, вносят в требуемый штамм Corynebacterium glutamicum путем конъюгации или трансформации. Метод конъюгации описан, например, у Schafer и др., Applied and Environmental Microbiology 60, 1994, сс. 756-759. Методы трансформации описаны, например, у Thierbach и др.. Applied Microbiology and Biotechnology 29, 1988, сс. 356-362, Dunican и Shivnan, Bio/Technology 7, 1989, сс. 1067-1070 и Tauch и др., FEMS Microbiology Letters 123, 1994, сс. 343-347. После гомологичной рекомбинации в результате случая кроссовера образовавшийся штамм содержит по меньшей мере две копии требуемого гена. Аналогичный метод для E. coli описан, например, у Link, A.J., Phillips, D. и Church G.M., J. Bacteriology 179, 1997, сс. 6228-6237.
Для инсерции или делеции ДНК в хромосоме можно применять также методы, основанные на использовании рекомбиназы, такие, как методы, описанные, например у Datsenko K.A., Wanner B.L.,, Ргос Nati Acad Sci USA., 97 (12), 2000, сс. 6640-6645.
Применяемую выше и далее фразу «активность фермента, повышенная по сравнению с активностью штамма дикого типа или исходного штамма» предпочтительно всегда следует понимать как активность конкретного фермента, которая увеличена по меньшей мере в 2 раза, в частности, предпочтительно по меньшей мере в 10 раз, более предпочтительно по меньшей мере в 100 раз, еще более предпочтительно по меньшей мере в 1000 раз и наиболее предпочтительно по меньшей мере в 10000. К клетке, предлагаемой в изобретении, которая обладает «активностью фермента, повышенной по сравнению с активностью штамма дикого типа или исходного штамма» относится также такая клетка, штамм которой дикого типа или исходный штамм не обладает вообще или по меньшей мере не обладает выявляемой активностью указанного фермента, и которая обладает выявляемой активностью указанного фермента только после повышения ферментативной активности, например, посредством сверхэкспрессии. В этом контексте понятие «сверхэкспрессия» или фраза «повышение экспрессии», которые используются далее, включают также случай, при которой в исходной клетке, например, клетке дикого типа, экспрессия вообще не обнаружена или по меньшей мере не обнаружена выявляемая экспрессия фермента, и выявляемая экспрессия фермента индуцируется только с помощью методов рекомбинации.
Выделенные бактерии, полученные с помощью путей, предлагаемых в изобретении, отличаются повышенной секрецией или повышенным производством требуемой аминокислоты в процессе ферментации по сравнению с применяемым исходным штаммом или родительским штаммом.
Под выделенными бактериями следует понимать мутантные и рекомбинантные бактерии, предлагаемые в изобретении, в частности бактерии семейства Enterobacteriaceae или Corynebacteriaceae, которые выделены или получены и которые обладают повышенной активностью тиосульфат-сульфотрансферазы по сравнению с исходным штаммом.
Выход выделенных бактерий и процесса ферментации с их использованием касательно одного или нескольких параметров, выбранных из группы, включающей концентрацию продукта (продукт на единицу объема), выход продукта (продукт, образовавшийся на единицу поглощенного источника углерода) и образование продукта (продукт, образовавшийся на единицу объема и в единицу времени), или также другие параметры процесса и их комбинации, повышаются по меньшей мере на 0,5%, по меньшей мере на 1%, по меньшей мере на 1,5% или по меньшей мере на 2% относительно исходного штамма или родительского штамма или процесса ферментации с их использованием.
Согласно способу, предлагаемому в изобретении, бактерии можно культивировать непрерывно, что описано, например в PCT/EP 2004/008882, или с перерывами с помощью периодического процесса (культивирование партий) или периодического процесса с подпиткой (процесс с подпиткой), или периодического процесса с повторной подпиткой (процесс с повторной подпиткой), для получения L-аминокислот.Обзор общих принципов, на которых основаны известные методы культивирования, можно найти в учебнике Chmiel, Bioprozesstecknik [Bioprocess Technology], 1. Einfuhrung in die Bioverfahrenstechnik [Introduction to Bioprocess Engineering], изд-во Gustav Fischer Verlag, Stuttgart, 1991, или в учебнике Storhas, Bioreaktoren und periphere Einrichtungen [Bioreactors and Peripheral Equipment], изд-во Vieweg Verlag, Braunschweig/Wiesbaden, Germany, 1994.
Предназначенная для применения культуральная среда или ферментационная среда должна соответствующим образом удовлетворять требованиям, зависящим от конкретных штаммов. В руководстве «Manual of Methods for General Bacteriology» Американского общества бактериологов (Washington D.C., USA, 1981) приведено описание культуральных сред для различных микроорганизмов. Понятия «культуральная среда» и «ферментационная среда» или среда используются взаимозаменяемо.
В качестве источника углерода можно использовать сахара и углеводы, такие, например, как глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, содержащие сахарозу растворы, полученные в результате обработки сахарной свеклы или сахарного тростника, крахмал, гидролизат крахмала и целлюлозу, масла и жиры, такие, например, как соевое масло, подсолнечное масло, арахисовое масло и кокосовый жир, жирные кислоты, такие, например, как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие, например, как глицерин, метанол и этанол, и органические кислоты, такие, например, как уксусная кислота. Эти субстанции можно применять индивидуально или в виде смеси.
В качестве источника азота можно использовать содержащие азот органические соединения, такие как пептоны, дрожжевой экстракт, мясной экстракт, солодовый экстракт, кукурузный экстракт, соевая мука и мочевина, или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Источники азота можно применять индивидуально или в виде смеси.
В качестве источника фосфора можно использовать фосфорную кислоту, первичный кислый фосфат калия или вторичный кислый фосфат калия или соответствующие содержащие натрий соли.
Согласно изобретению применяемым источником серы является соль дитиосерной кислоты (тиосульфат), необязательно в сочетании с другими источниками серы, такими, например, как сульфат, сульфит или дитионит.
Кроме того, культуральная среда может содержать соли, например, в форме хлоридов металлов, таких, например, как натрий, калий, магний, кальций и железо, сульфатов, таких, например, как сульфат магния или сульфат железа, которые необходимы для роста. И, наконец, помимо указанных выше субстанций можно применять незаменимые факторы роста, такие как аминокислоты, например, гомосерин, и витамины, например, тиамин, биотин или пантотеновую кислоту.
В культуральную среду можно добавлять также приемлемые предшественники конкретной аминокислоты.
Указанные исходные продукты можно добавлять в культуру в виде одной партии или осуществлять соответствующим образом подпитку в процессе культивирования.
Значение рН культуры можно контролировать соответствующим образом путем применения оснований, таких как гидроксид натрия, гидроксид калия, аммиак или водный раствор аммиака, или кислот, таких как фосфорная кислота или серная кислота. Как правило, pH доводят до значения, составляющего от 6,0 до 9,0, предпочтительно от 6,5 до 8. Для контроля пенообразования можно применять противовспенивающие средства, такие, например, как эфиры жирной кислоты и полигликоля. Для поддержания стабильности плазмид можно добавлять в среду пригодные обладающие избирательным действием субстанции, такие, например, как антибиотики. Для поддержания аэробных условий в культуру вводят кислород или кислородсодержащие газовые смеси, такие, например, как воздух. Можно применять также жидкости, обогащенные пероксидом водорода. Ферментацию необязательно осуществляют при избыточном давлении, например, при избыточном давлении, составляющем от 0,03 до 0,2 МПа. Температура культуры обычно составляет от 20 до 45°C и предпочтительно от 25 до 40°C. В случае периодических процессов культивирование продолжают вплоть до образования максимального количества требуемой аминокислоты. Как правило, для достижения этой цели требуется от 10 до 160 ч. В случае непрерывных процессов может потребоваться осуществлять культивирование в течение более длительных промежутков времени.
Примеры пригодных ферментационных сред описаны, среди прочего, в US 6221636, US 5840551, US 5770409, US 5605818, US 5275940 и US 4224409.
Методы определения L-аминокислот известны из существующего уровня техники. Анализ можно осуществлять, например, согласно методу, описанному у Spackman и др.. Analytical Chemistry, 30, 1958, с. 1190), с помощью ионообменной хроматографии с последующей дериватизацией нингидрином, или его можно осуществлять с помощью ЖХВР с обращенной фазой согласно методу, описанному у Lindroth и др.. Analytical Chemistry, 1979, 51, сс. 1167-1174).
Полученный таким образом ферментационный бульон затем дополнительно перерабатывают с получением твердого или жидкого продукта.
Под ферментационным бульоном подразумевают ферментационную среду, в которой осуществляли культивирование микроорганизма в течение определенного периода времени и при определенной температуре. Ферментационная среда или среды, применяемая/применяемые при ферментации, содержит/содержат все субстанции или компоненты, обеспечивающие размножение и жизнеспособность микроорганизма и образование требуемой аминокислоты.
После завершения ферментации полученный ферментационный бульон содержит соответственно:
а) бактериальную биомассу (клеточную массу), образовавшуюся в результате размножения микроорганизма,
б) требуемую аминокислоту, образовавшуюся в результате ферментации,
в) органические побочные продукты, образовавшиеся в процессе ферментации, и
г) компоненты применявшейся ферментационной среды или исходных продуктов, такие, например, как витамины, например, биотин, аминокислоты, такие как гомосерин, или соли, такие как сульфат магния, которые не были поглощены в процессе ферментации.
К органическим побочным продуктам относятся субстанции, которые могут производиться и могут секретироваться бактериями, применяемыми для ферментации, помимо конкретной требуемой L-аминокислоты. Они включают L-аминокислоты, содержание которых составляет менее 30%, 20% или 10% по сравнению с содержанием требуемой аминокислоты. К ним относятся также органические кислоты, которые несут одну-три карбоксильных группы, такие, например, как уксусная кислота, молочная кислота, лимонная кислота, яблочная кислота или фумаровая кислота. И, наконец, к ним относятся также сахара, такие, например, как трегалоза.
Как правило, в ферментационных бульонах, которые можно применять для промышленных процессов, содержание аминокислоты составляет от 40 до 180 или от 50 до 150 г/кг. Содержание биомассы (в виде высушенной биомассы), как правило, составляет от 20 до 50 г/кг.
Для иллюстрации заявленного изобретения приведена фиг.1. На ней содержится плазмида рМА-RDL2, содержащая AmpR: ген, обусловливающий устойчивость к ампициллину; кодирует бета-лактамазу;
upstream: SEQ ID NO: 6; содержит последовательности, распознаваемые рестриктазами PacI и FseI, а также сайт связывания рибосом;
RDL2: последовательность гена RDL2 из штамма Saccharomyces cerevisiae S288c;
downstream: SEQ ID NO: 7; содержит второй стоп-кодон ТАА, за которым расположен терминатор Т1 гена rnpB из штамма E. coli MG1655. За ним расположены последовательности, распознаваемые рестриктазами PmeI und SbH
Примеры:
Пример 1: Синтез и клонирование гена RDL2
Для экспрессии OPC RDL2 (SEQ ID NO: 1) из S. cerevisiae синтезировали de novo нуклеотидную последовательность (фирма GENEART AG; Регенсбург, Германия), которая содержит расположенную против хода транскрипции последовательность («upstream», SEQ ID NO: 6), последовательность OPC RDL2 («RDL2», SEQ ID NO: 1) и расположенную по ходу транскрипции последовательность («downstream», SEQ ID NO: 7). Расположенная против хода транскрипции последовательность содержит последовательности, распознаваемые рестриктазами PacI и FseI, и сайт связывания рибосом. Расположенная по ходу транскрипция последовательность содержит второй стоп-кодон ТАА, за которым следует терминатор Т1 гена rnpB из штамма E. coli MG1655. За ним расположены последовательности, распознаваемые рестриктазами PmeI и SbfI. После синтеза нуклеотидную последовательность расщепляли Sad и KpnI и клонировали в плазмиде рМА (ampR), также расщепленной с помощью Sad и KpnI. Полученную плазмиду обозначали как «pMA-RDL2» (SEQ ID NO: 8, фиг. 1).
Используя в качестве исходной аминокислотную последовательность белка Rdl2p, кодируемую OPC RDL2 (последовательность «RDL2p», SEQ ID NO: 2), создавали три различные ДНК-последовательности, которые кодируют белки Rdl2p с аминокислотной последовательностью дикого типа. Последовательности обозначали как «RDL2a» (SEQ ID NO: 3), «RDL2b» (SEQ ID NO: 4) и «RDL2c» (SEQ ID NO: 5). Каждую из этих трех последовательностей синтезировали de novo вместе с описанными выше расположенными против и по ходу транскрипции последовательностями и клонировали в описанной выше плазмиде рМА (фирма GENEART AG; Регенсбург, Германия). Полученные плазмиды обозначали как pMA-RDL2a, pMA-RDL2b и pMA-RDL2c. Указанные выше варианты различались оптимизацией наиболее часто встречающихся кодонов в зависимости от применяемого для осуществления способа микроорганизма. Так, использовали следующие индексы адаптации наиболее часто встречающихся кодонов:
RDL2: отсутствует адаптация наиболее часто встречающихся кодонов (индекс адаптации кодонов CAI=0,27),
RDL2a: невысокая степень адаптации наиболее часто встречающихся кодонов к E. coli (CAI=0,38),
RDL2b: более высокая степень адаптации наиболее часто встречающихся кодонов к E. coli (CAI=0,72),
RDL2c: полная адаптация наиболее часто встречающихся кодонов к E. coli (CAI=1).
Пример 2: Клонирование OPC RDL2. RDL2a. RDL2b и RDL2c в плазмиде pME101-thrA*1-cysE-Pgap-metA*11.
Для экспрессии в E. coli каждый из четырех вариантов генов, кодирующих белок RDL2, клонировали в применяемой для E. coli продуктивной плазмиде pME101-thrA*1-cysE-Pgap-metA*11 (WO 2007/077041), расположенной по ходу транскрипции относительно metA*11.
Для этой цели каждый из генов амплифицировали с использованием праймеров RDL2-t4-f(SEQ ID NO: 11) и RDL2-r (SEQ ID NO: 12), применяя полимеразную цепную реакцию (ПЦР) с ДНК-полимеразой Phusion (фирма Finnzymes Oy, Эспоо, Финляндия). Плазмиды pMA-RDL2, pMA-RDL2a, рМА-RDL2b и pMA-RDL2c, описанные в примере 1, служили в качестве матриц.
RDL2-t4-f(SEQIDNO: 11):
5' aggacagtcgacggtaccgcaagcttcggcttcgcACTGGAAAGCGGGCAGTGAG 3' RDL2-r (SEQ ID NO: 12):
5' AGCGCGACGTAATACGACTC 3'
Состоящий из 804 пар оснований ПЦР-продукт и плазмиду pME101-thrA*1-cysE-PgapA-metA*11 расщепляли рестриктазами SaiI и SbfI, лигировали и трансформировали ими штамм E. coli DH5α. Несущие плазмиду клетки отбирали путем культивирования на LB-агаре с 50 мкг/мл стрептомицина. После выделения плазмидной ДНК успешное клонирование подтверждали путем контрольного расщепления рестриктазой AgeI. И, наконец, клонированные ДНК-фрагменты секвенировали с использованием следующих праймеров:
pME-RDL2-Seqf(SEQ ID NO: 13):
5' ATGTGGAAGCCGGACTAGAC 3'
pME-RDL2-Seqr (SEQ ID NO: 14):
5' TCGGATTATCCCGTGACAGG 3'.
Полученные таким образом плазмиды обозначали как pME-RDL2, pME-RDL2a, pME-RDL2b и pME-RDL2c.
Пример 3: Трансформация E. coli с использованием MG1655ΔmetJPtrc-metH Ptrc-metF PtrcV-cvsPUWAM PtrcV-cvsJIH VtrcW-gcvTHP
Штамм E. coli MG1655ΔwetJ metA*11 Ptrc-metHPtrc-metFPtrcF-cysPUWAM PtrcF-cysJIH (ниже обозначен как DM2219), который продуцирует L-метионин, описан в заявке WO 2009/043803 A2.
Указанный штамм трансформировали по отдельности каждой из плазмид pME-RDL2, pME-RDL2a, pME-RDL2b и pME-RDL2c и плазмидой pME101-thrA*1-cysE-VgapA-metA*11 (WO 2009/043803 A2) и несущие плазмиду клетки отбирали на LB-агаре с 50 мкг/мл стрептомицина. Трансформантов пересевали в каждом случае в 10 мл жидкой LB-среды, дополненной 1% глюкозы и 50 мкг/мл стрептомицина, и культивировали при 37°C в течение 16 ч. Добавляли глицерин до конечной концентрации 10% и культуры замораживали при -70°C.
Пример 4: Оценка продуцирующих L-метионин штаммов E. coli
Конечную производительность продуцирующих L-метионин штаммов E. coli оценивали с помощью экспериментов по оценке продуктивности в 100-миллилитровых колбах Эрленмейера. В качестве предварительных культур в каждом случае по 10 мл среды для предварительных культур (10% LB-среды, дополненной 2,5 г/л глюкозы, и 90% минимальной среды РС1) инокулировали 100 мкл клеточной культуры и культуры выращивали при 37°C в течение 10 ч. Затем в каждом случае инокулировали ими по 10 мл минимальной среды PC1 (таблица 1) до достижения величины ОП при 600 нм, составляющей 0,2 (фотометр Eppendorf Bio; фирма Eppendorf AG, Гамбург, Германия), и культивировали при 37С в течение 24 ч. Определяли концентрацию внеклеточного L-метионина с использованием аминокислотного анализатора (фирма Sykam GmbH, Эрезинг, Германия) с помощью ионообменной хроматографии и постколоночной дериватизации нингидрином. Внеклеточную концентрацию глюкозы определяли с помощью анализатора YSI 2700 Select Glucose (фирма YSI Life Sciences, Йеллоу-Спрингс, шт. Огайо, США). Данные демонстрируют, что экспрессия OPC RDL2, RDL2a, RDL2b или RDL2c приводит к существенному увеличению концентрации L-метионина (таблица 2).
|
|
Пример 5: Клонирование OPC RDL2. RDL2a. RDL2b и RDL2c в плазмиде pEC-ХТ99А
В качестве основного вектора для экспрессии OPC RDL2, RDL2a, RDL2b и RDL2c в C. glutamicum применяли бифункциональную экспрессионную плазмиду для E. coli-C. glutamicum рЕС-ХТ99А (ЕР 1085094 В1). Каждую из плазмид pMA-RDL2, pMA-RDL2a, pMA-RDL2b и pMA-RDL2c, описанных в примере 1, расщепляли рестриктазами StuI и Nael и разделяли в агарозном геле 0,8%-ной крепости. Вырезали из геля ДНК-фрагменты длиной 638 пар оснований и экстрагировали ДНК (набор для экстракции из геля QIAquick, фирма QIAGEN, Хильден, Германия). Экспрессионную плазмиду рЕС-ХТ99А расщепляли рестриктазой Eel 13 611. Затем в каждом случае осуществляли встраивание путем лигирования ДНК-фрагментов длиной 638 пар оснований с помощью набора для лигирования Ready-To-Go (фирма Amersham GE Healthcare Europe GmbH, Фрайбург, Германия). Штамм E. coli DH5a (фирма Invitrogen GmbH; Дармштадт, Германия) трансформировали полученными в результате лигирования партиями и отбирали несущие плазмиду клетки на LB-агаре с 5 мкг/мл тетрациклина.
После выделения плазмидной ДНК успешное клонирование подтверждали путем контрольного расщепления рестриктазами Pmel и SacII. И, наконец, плазмиды секвенировали с использованием следующих праймеров:
pECf(SEQIDNO:9):
5' TACTGCCGCCAGGCAAATTC 3'
pECr (SEQ ID NO: 10):
5' TTTGCGCCGACATCATAACG 3'.
Плазмидные конструкции, в которых собственный промотор trc плазмиды и ОРС, кодирующие RDL2, находились в одинаковой ориентации, обозначали как:
pEC-RDL2, pEC-RDL2a, pEC-RDL2b и pEC-RDL2c.
Пример 6: Трансформация C. glutamicum M1179
Штамм Corynebacterium glutamicum M1179 представляет собой устойчивый к этионину штамм-продуцент L-метионина (WO 2007/011939). Он депонирован в соответствии с Будапештским договором в Немецкой коллекции микроорганизмов и клеточных культур (Deutsche Sammlung fur Mikroorganismen und Zeilkulturen) (DSMZ, Брауншвейг, Германия) 18.5.2005 г. под номером DSM17322.
Плазмиды pEC-RDL2, pEC-RDL2a, pEC-RDL2b, pEC-RDL2c и рЕС-ХТ99А встраивали путем электропорации в штамм Ml 179 с использованием условий электропорации, описанных у Tauch и др., FEMS Microbiological Letters, 123, 1994, сс. 343-347.
Отбор несущих плазмиду клеток осуществляли на LB-агаре с 5 мкг/мл тетрациклина. Выделяли плазмидную ДНК трансформантов (Peters-Wendisch и др., Microbiology 144, 1998, сс. 915-927) и анализировали путем расщепления рестриктазами и гель-электрофореза. Полученные таким образом штаммы обозначали как Ml 179/pEC-RDL2, Ml 179/pEC-RDL2a, Ml 179/pEC-RDL2b, M1179/pEC-RDL2c и М1179/рЕС-ХТ99А.
Пример 7: Оценка продуцирующих L-метионин штаммов С.slutamicum
Полученные штаммы C. glutamicum Mil 79/pEC-RDL2, Ml 179/pEC-RDL2a, Ml 179/pEC-RDL2b, Ml 179/pEC-RDL2c и Ml 179/pEC-XT99A культивировали в питательной среде, пригодной для производства L-метионина, и определяли содержание L-метионина в супернатанте культур.
Для этой цели штаммы сначала высевали штрихами на агаровые пластины (агар с сердечно-мозговой вытяжкой с канамицином (25 мг/л)) и инкубировали при 33°C в течение 24 ч. Клетки из культур, выращенных на указанных агаровых пластинах, пересевали в каждом случае в 10 мл BH-среды (бульон с сердечно-мозговой вытяжкой, фирма Merck, Дармштадт, Германия) с 5 мг/л тетрациклина и культуры встряхивали в 100-миллилитровых колбах Эрленмейера при 33°C и 240 об/мин в течение 24 ч. Затем в каждом случае ими инокулировали 10 мл PM-среды (таблица 3) до достижения величины ОП при 600 нм, составляющей 0,1 (устройство Genios, фирма Tecan Deutschland GmbH, Крайльсхайм, Германия), и культивировали при 33°C в 100-миллилитровых колбах Эрленмейера с дефлекторами в течение 24 ч.
|
Через 24 ч определяли ОП при 660 нм (устройство Genios, фирма Tecan Deutschland GmbH, Крайльсхайм, Германия) и затем определяли концентрацию образовавшегося L-метионина. L-метионин определяли с использованием аминокислотного анализатора (фирма Sykam GmbH, Эрезинг, Германия) с помощью ионообменной хроматографии и постколоночной дериватизации нингидрином. В таблице 4 представлены оптические плотности и концентрации L-метионина в культурах. Установлено, что экспрессия OPC RDL2, RDL2a, RDL2b или RDL2c приводит к существенному увеличению концентрации L-метионина.
|
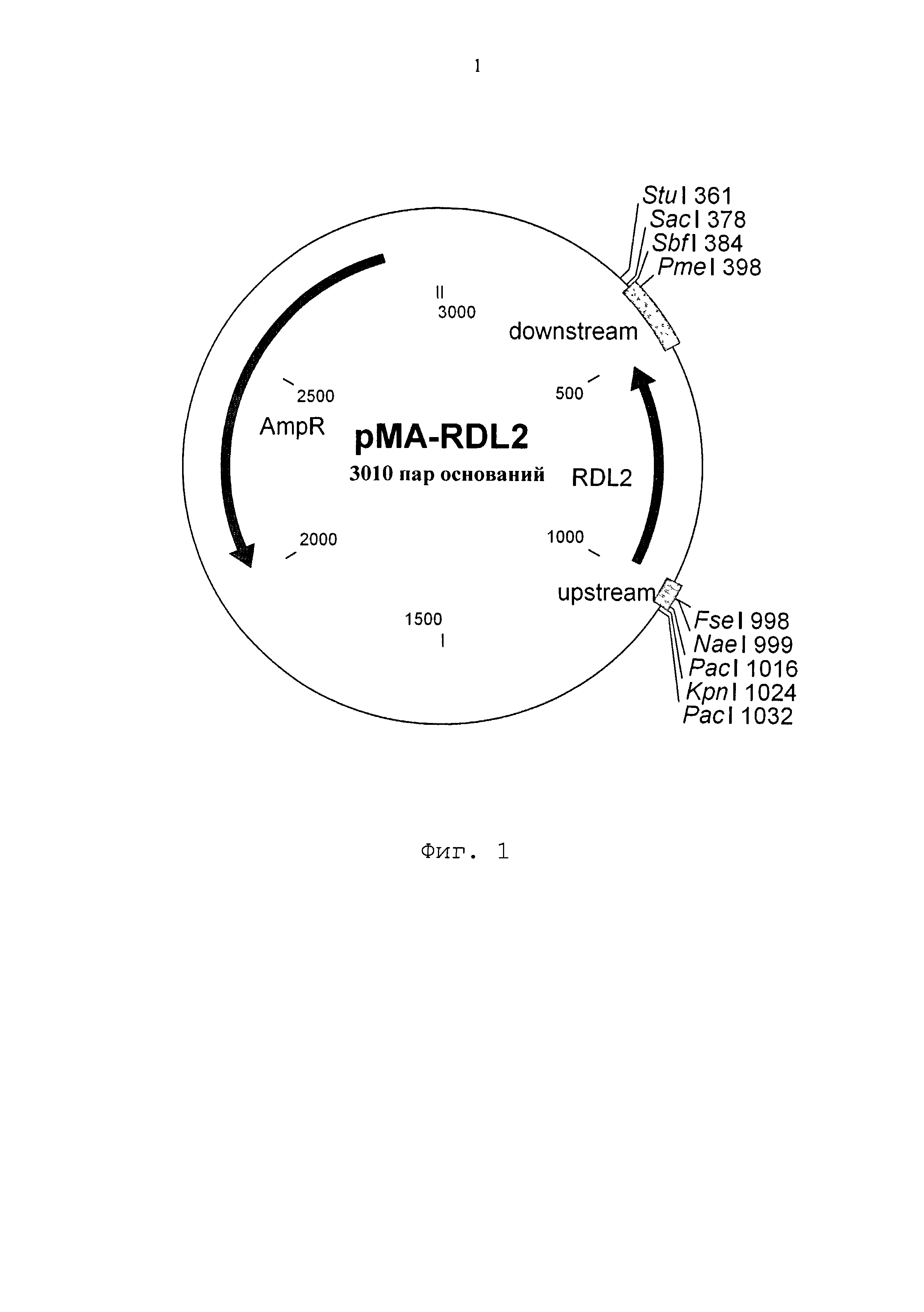







Способ получения метионина из гомосерина
Клей на основе привитых сополимеров сложных полиэфиров и поли(мет)акрилата
Смешанные оксидные катализаторы для каталитического окисления в газовой фазе
Кетометионинкетали и их производные
Способ регенерации катализатора, используемого при дегидратации глицерина
Способ непрерывного получения метилмеркаптана из углерод- и водородсодержащих соединений
Содержащие вольфрамовые соединения катализаторы и способ дегидратации глицерина
Волокнистые композитные материалы и их применение в системах вакуумной изоляции
Применение полиамидной формовочной массы для облицовки трубопроводов
Способ получения метилмеркаптана из диалкилсульфидов и диалкилполисульфидов
Привод для педального транспортного средства, предназначенного, прежде всего, для детей
Мобильная дробильная установка
Низковибрационная щековая дробилка
Способ ферментативного получения l-аминокислот с использованием улучшенных штаммов семейства enterobakteriaceae
Игрушечная машина-экскаватор
Трубчатая пружина для транспортных средств и способ изготовления трубчатой пружины
Трубчатая пружина для транспортных средств и способ изготовления трубчатой пружины

