Результат интеллектуальной деятельности: НОВЫЙ ГЕН ФЛАВОНОИД 3,5-ГИДРОКСИЛАЗЫ КОЛОКОЛЬЧИКА И ЕГО ПРИМЕНЕНИЕ
Вид РИД
Изобретение
Область техники, к которой относится изобретение
Настоящее изобретение относится к методу генетической инженерии, в котором экзогенный ген вводится в растительные клетки. Более конкретно, изобретение относится к новому гену флавоноид 3',5'-гидроксилазы колокольчика, к плазмиде, содержащей ген под контролем 35S промотора вируса мозаики цветной капусты, и к способу получения растения садовой розы со значительно повышенным содержанием дельфинидина в лепестках, путем трансформации с использованием плазмиды способа с Agrobacterium.
Предшествующий уровень техники
Способы кроссбридинга растений включают (i) гибридизацию путем скрещивания тычинки и пестика, (ii) естественную или искусственную мутацию, и (iii) генную рекомбинацию. Из этих способов только технология генной рекомбинации может быть использована для придания новых признаков растениям путем экспрессии полезных генов в растениях-мишенях, без стеснения генетическими ограничениями видов. Широкий спектр растений с рекомбинантными генами, созданных таким образом, уже выращиваются по всему миру.
Компоненты окраски цветка включают антоцианины, каротеноиды и беталаины. Антоцианины являются представителями вторичных метаболитов общей группы соединений, названных флавоноидами, и они синтезируются из фенил ал анина под действием многочисленных ферментов. Цвет антоцианина зависит от его структуры. В частности, цвет становится более синим при увеличении количества гидроксильных групп на кольце анотицианидина В, которое является хромофорной группой антоцианина. В ряду основных антоцианидинов, состоящем из дельфинидина, цианидина и пеларгонидина, количество гидроксильных групп на В-кольце растет в указанном порядке. Антоцианины, полученные из дельфинидина, накапливаются в большинстве голубых цветов. Также известно, что при росте количества ароматических ацильных групп (кумарароильных групп, кафеоильных групп и т.п.), модифицирующих антоцианины, происходит усиление синего цвета антоцианинов (при котором максимум абсорбции смещается к длинноволновому концу), что соответствует повышенной стабильности антоцианинов. В частности, антоцианины, к которым присоединено множество ароматических ацильных групп, известные как полиацилированные антоцианины, присутствуют в синих лепестках горечавки, клитории и т.п.(см. Plant J. 54, 737-749, 2008). Ферменты, вовлеченные в биосинтез антоцианинов, и гены, кодирующие их, были выделены из множества растений (см. Plant J. 54, 737-749, 2008). Цвета антоцианинов зависят от локального рН вакуолей, и от присутствия других флавоноидов, металлических ионов и т.п. В частности, антоцианины демонстрируют красную окраску при низком рН вакуолей, и синюю окраску при нейтральном значении. Флавоны и флавонолы относятся к копигментам, и они обладают способностью делать антоцианины синими, когда они присутствуют вместе с антоцианинами. Также известно, что ионы железа и алюминия, координированные гидроксильными группами В-кольца антоцианина, делают антоцианин синим.
Цветки растений имеют большой спектр различных окрасок, но не существует разновидности, в которой были бы цветки всех цветочных окрасок. Это происходит из-за того, что пигменты, которые могут быть синтезированы разновидностью, определены генетически. Например, розы, гвоздики, хризантемы, лилии, герберы и т.п. не имеют естественных синих или фиолетовых разновидностей. Основная причина этого заключается в том, что эти растения не имеют гена для флавоноид 3',5'-гидроксилазы (далее «F3'5'H»), который необходим для синтеза дельфинидина. В нескольких сообщениях зафиксированы попытки экспрессировать ген F3'5'H в лепестках для выработки дельфинидина и получения синей цветочной окраски. Такое исследование основано на важности выбора соответствующего промотора и соответствующего гена F3'5'H для каждого разновидности, как описано ниже, в целях повышения содержания дельфинидина в лепестках и смещения цветочной окраски к синей.
При экспрессии гена F3'5'H в петуниях, которые не имеют F3'5'H, повышается уровень антоцианинов, полученных из дельфинидина (Nature, 1993, 366, 276-279, FEBS Lett. 1999, 461, 241-245). Также, экспрессия гена F3'5'H в табаке (Nicotiana tabacum), который накапливает цианидин, приводит к синтезу дельфинидина и придает слабосинюю окраску цветкам (FEBS Lett. 1999,461, 241-245).
В NPL 1 сообщается, что когда гены F3'5'H, полученные из колокольчика, гиацинтоидеса неописанного и петунии, экспрессировали в табаке под контролем 35S промотора вируса мозаики цветной капусты, наиболее эффективная выработка дельфинидина была получена с геном F3'5'H колокольчика (см. реферат публикации).
Также в PTL 1 описывается введение полученного из петунии гена F3'5'H в розу под контролем 35S промотора вируса мозаики цветной капусты (см. Пример 9), но там же ген F3'5'H, полученный из колокольчика, вводили просто в табак, при этом ничего не упоминая относительно выработки дельфинидина и изменения окраски растения розы или лепестков розы.
В NPL 2 также описывается экспрессия генов F3'5'H клитории и вербены в вербене, при этом отмечается, что экспрессия гена, полученного из клитории, приводит к большей выработке дельфинидина и четко измененной окраске цветов.
В PTL 2 также утверждается, что при экспрессии в розе генов F3'5'H, полученных из петунии, горечавки, цинерарии, гиацинтоидеса, анютиных глазок и лаванды, экспрессия гена F3'5'H, полученного из анютиных глазок, под контролем 35S промотора вируса мозаики цветной капусты, или промотора гена халькон синтазы, полученного из розы, приводит к выработке дельфинидина, составляющей несколько десятков процентов от общего содержания антоцианидинов, при этом полученный из анютиных глазок ген F3'5'H удовлетворительно функционирует в розе под контролем 35S промотора вируса мозаики цветной капусты или промотора гена халькон синтазы, полученного из розы.
В NPL 3 сообщается о том, что дополненный энхансером 35S промотор вируса мозаики цветной капусты дает более эффективную экспрессию экзогенных генов, чем 35S промотор вируса мозаики цветной капусты. Роза, обладающая геном F3'5'H анютиных глазок, введенного под контролем E12 35S промотора, вырабатывает дельфинидин и имеет измененную окраску цветков. В зависимости от хозяйской разновидности возможно получить розу, содержащую дельфинидин с 95% или большим содержанием от общего количества антоцианидинов, и имеющую окраску цветков измененную до фиолетовой, и одна такая разновидность уже продается (учетный номер разновидности No. 20992).
Более того, в PTL 3 сообщается, что в розе, в которую введена кассета, которая переносит ген F3'5'H анютиных глазок под контролем промотора Е12 35S, ген дигидрофлавонол 4-редуктазы (далее «DFR») под контролем промотора Е12 35S, и двухцепочечную РНК DFR розы, содержание дельфинидина составляет около 100%, а окраска цветка изменилась в интродуцированных разновидностях розы, вне зависимости от разновидности.
Для изменения окраски цветка розы, имеющей товарную ценность, естественно желательно повысить содержание дельфинидина в лепестках, но также важно не оказывать нежелательного воздействия на рост розы. Также существенно проведение перекрестного скрещивания с использованием в качестве родителя розы, рекомбинантной по гену высокой экспрессии дельфинидина. Другими словами, для разновидности жизнеспособной садовой розы необходима трансформация плазмидой содержащей специфическую комбинацию специфического высокоэкспрессирующего промотора, позволяющего осуществить высокую экспрессию F3'5'H, и F3'5'H, имеющего специфическую последовательность, которые позволяют максимизировать содержание дельфинидина в лепестках.
Также в PTL 4 описывается перенос гена F3'5'H, полученного из анютиных глазок, в разновидности хризантемы Improved Reagan и Dark Splendid Reagan с использованием промотора халькон синтазы розы, на основании чего была получена рекомбинантная хризантема, в которой запасалось 50% и более дельфинидина, и имеющая окраску цветков, измененную на синий.
В PTL 5, при экспрессии генов F3'5'H, полученных из колокольчика, вербены, цинерарии, анютиных глазок, петунии и лобелии, в хризантемах разновидности 94-765, с использованием промотора гена флаванон 3-гидролазы хризантемы и 5'-нетранслируемой области гена алкогольдегидрогеназы табака, гены F3'5'H, полученные из колокольчика, вербены, цинерарии и анютиных глазок, функционируют относительно удовлетворительно, вырабатывая дельфинидин на уровне 25% или больше от общего количества антоцианидинов. В той же публикации утверждается, что среди них, ген F3'5'H, полученный из колокольчика, в хризантеме функционирует наиболее удовлетворительно, вырабатывая 50% и больше дельфинидина.
Mac 1-промотор, который является конститутивным промотором, промотор гена халькон синтазы из львиного зева, который является промотором, специфичным для лепестков, и промотор гена халькон синтазы из петунии, который является промотором, специфичным для лепестков, были использованы для экспрессии гена F3'5'H петунии в гвоздике. В данном случае, два различных промотора, специфичных для лепестков, вырабатывают большие количества дельфинидина, чем промотор Macl (см. PTL 6). Было продемонстрировано, что в лепестках гвоздики промотор гена DFR петунии или промотор гена антоцианидин синтазы являются эффективными для переноса экзогенных генов (см. PTL 7 и 8). В качестве других промоторов, специфичных для лепестков, сообщали о промоторе гена F3'5'H анютиных глазок (см. PTL 9), промоторе гена трансферазы антоцианин 3-ацильных групп (см. PTL 10) и промоторе гена F3'5'H цинерарии (см. PTL 11).
Как упоминалось выше, хотя для выработки дельфинидина ген F3'5'H может быть экспрессирован в различных разновидностях растений, трудно спрогнозировать, какой промотор должен быть использован для экспрессии полученного из растений гена F3'5'H в клетке-мишени для выработки дельфинидина с высокой эффективностью, т.е. с содержанием дельфинидина 80% или больше, предпочтительно 85% или больше, более предпочтительно 90% или больше, еще более предпочтительно 95% или больше, от общего количества антоцианидинов, и необходимо проводить эксперименты методом проб и ошибок или достаточно сложную экспериментальную работу.
Кроме того, для изменения окраски цветов на синий, в общем, необходимо увеличить содержание дельфинидина на 50% или больше, предпочтительно, на 60% или больше, более предпочтительно на 70% или больше, более предпочтительно на 80% или больше, более предпочтительно на 85% или больше, более предпочтительно на 90% или больше, более предпочтительно на 95% или больше, более предпочтительно на 99% или больше, и наиболее предпочтительно на 100%, от общего количества антоцианидинов, а простая сверхэкспрессия гена F3'5'H, как правило, недостаточна для данной цели. В случае гвоздики, например, экспрессия гена F3'5'H и гена DFR петунии в белой гвоздике, которая утратила дигидрофлавонол 4-редуктазу (DFR), дает в результате около 100% дельфинидина, с получением рекомбинантной гвоздики с окраской цветов, измененной на синий (см. PTL 12). Сверхэкспрессия полученного из анютиных глазок гена F3'5'H и полученного из ириса гена DFR в розе, одновременно с подавлением экспрессии гена DFR розы, дает в результате содержание дельфинидина около 100% с получением рекомбинантной розы, с окраской цветов измененной на синюю (см. PTL 3). Повышение содержания дельфинидина также было достигнуто путем сверхэкспрессии полученного из анютиных глазок гена F3'5'H в хризантеме, с одновременным подавлением экспрессии гена F3'H хризантемы (см. PTL 4).
Список процитированной литературы
Документы Патентной Литературы
[PTL 1] Патентная Публикация Японии No. 3403196 (WO 93/18155)
[PTL 2] WO 2004/020637
[PTL 3] WO 2005/017147
[PTL 4] WO 2009/062253
[PTL 5] WO 2010/122849
[PTL 6] WO 94/28140
[PTL 7] WO 96/036716
[PTL 8] WO 2009/062259
[PTL 9] WO 2010/122850
[PTL 10] PCT/JP2010/053886
[PTL 11] PCT/JP2010/070516
[PTL 12] WO 06/36716
[Непатентная литература]
[NPL 1] Biosci. Biotechnol. Biochem., 67(1), 161-165, 2003
[NPL 2] Plant Biotechnology 2006, 23, 5-11
[NPL 3] Plant Cell Physiol. (1996) 37: 49-59
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Проблемы, решаемые изобретением
В свете данных обстоятельств, проблема, решаемая настоящим изобретением, заключается в обеспечении трансформационной плазмиды, содержащей специфическую комбинацию специфического высокоэкспрессирующего промотора, позволяющего осуществить высокую экспрессию F3'5'H, и F3'5'H, имеющего специфическую последовательность, которая позволяет максимизировать содержание дельфинидина в лепестках в жизнеспособной разновидности садовой розы. Средства решения проблем
Авторы провели много исследований и экспериментов, направленных на решение данной проблемы, и в результате реализовали изобретение, обнаружив, что можно получить растение трансформированной розы, запасающей в лепестках дельфинидин на высоком уровне, путем трансформации специфической разновидности растений розы способом с Agrobacterium с использованием плазмиды, содержащей ген F3'5'H колокольчика, имеющего специфическую последовательность, под контролем 35S промотора вируса мозаики цветной капусты.
В частности, настоящее изобретение представляет собой следующее.
[1] Полинуклеотид, выбранный из группы, состоящей из нижеследующих (а)-(d):
(a) полинуклеотид, содержащий нуклеотидную последовательность, указанную как SEQ ID NO: 1;
(b) полинуклеотид, идентичный, по меньшей мере, на 97% по последовательности полинуклеотиду, содержащему нуклеотидную последовательность, указанную как SEQ ID NO: 1, или комплементарной ему нуклеотидной последовательности, кодирующий белок, имеющий активность флавоноид 3',5'-гидроксилазы,
(c) полинуклеотид, кодирующий белок, содержащий аминокислотную последовательность, указанную как SEQ ID NO: 2; и
(d) полинуклеотид, кодирующий белок, имеющий аминокислотную последовательность, идентичную, по меньшей мере, на 97% аминокислотной последовательности, указанной как SEQ ID NO: 2, и имеющий активность флавоноид 3',5'-гидроксилазы.
[2] Плазмида, с которой функционально связан полинуклеотид по [1] выше под контролем 35S промотора вируса мозаики цветной капусты.
[3] Плазмида по [2] выше, с которой дополнительно функционально связан nos-терминатор.
[4] Способ получения трансформированного растения розы или его потомства, имеющего содержание дельфинидина 80% или больше в лепестках, путем трансформации разновидности садовой розы (Rosa hybrida) методом с Agrobacterium с использованием плазмиды по [2] или [3] выше.
[5] Способ по [4] выше, в котором содержание дельфинидина в лепестках составляет 85% или больше.
[6] Способ по [5] выше, в котором содержание дельфинидина в лепестках составляет 90% или больше.
[7] Способ по [6] выше, в котором содержание дельфинидина в лепестках составляет 95% или больше.
[8] Растение трансформированной розы или ее потомство, полученное способом по любому из [4]-[7] выше. Эффект изобретения
Путем трансформации с использованием плазмиды, содержащей ген флавоноид 3',5'-гидроксилазы колокольчика по изобретению под контролем 35S промотора вируса мозаики цветной капусты способом с Agrobacterium, возможно значимо повысить содержание дельфинидина в лепестках разновидностей садовой розы. Лучший вариант осуществления изобретения
Полинуклеотид по изобретению (SEQ ID NO: 1) кодирует полученный из колокольчика F3'5'H, и состоит из 521 остатка. По всему настоящему описанию термин «полинуклеотид» относится к ДНК или РНК. Полинуклеотид изобретения не ограничен полинуклеотидом с нуклеотидной последовательностью, представленной как SEQ ID NO: 1 или кодирующей белок, содержащий соответствующую аминокислотную последовательность (SEQ ID NO: 2), и может быть выбран среди полинуклеотидов, содержащих нуклеотидную последовательность или комплементарную ей последовательность, или полинуклеотиды, кодирующие белки, имеющие гомологию к конкретной последовательности и предпочтительно идентичность последовательности, с аминокислотной последовательностью, и обладающий активностью флавоноид 3',5'-гидролазы. Предпочтительно это полинуклеотид, последовательность которого идентична, по меньшей мере, на 97%.
Полинуклеотид изобретения является молекулой ДНК, которая гибридизуется с нуклеотидной последовательностью, представленной как SEQ ID NO: 1 в жестких условиях и которая кодирует белок с активностью флавоноид 3',5'-гидролазы. В данном документе термин «полинуклеотид, который гибридизуется в жестких условиях» при использовании по всему настоящему описанию означает полинуклеотид, полученный, например, методом гибридизации на колониях, методом гибридизации бляшек или методом гибридизации по Саузерну, с использованием в качестве зонда всего или части полинуклеотида, содержащего нуклеотидную последовательность, представленную как SEQ ID NO: 1 или комплементарную ей нуклеотидную последовательность. Используемый способ гибридизации может быть, например, способом, описанным в «Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor, Laboratory Press 2001» or «Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997".
Термин «жесткие условия», используемый по всему настоящему описанию, может быть слабо-жесткими условиями, умеренно жесткими условиями или сильно жесткими условиями. В данном документе «слабо жесткие условия» являются, например, условиями 5 × SSC, 5 × раствора Денхарта, 0,5% SDS, 50% формамида, 32°С. Также «умеренно жесткие условия» являются, например, условиями 5 × SSC, 5 × раствора Денхарта, 0,5% SDS, 50% формамида, 42°С. «Сильно жесткие условия» являются, например, условиями 5 × SSC, 5 × раствора Денхарта, 0,5% SDS, 50% формамида, 50°С, а особенно 0,1 × SSC, 0,1% SDS, 65°С или 0,1 × SSC, 0,1% SDS, 70°C. В таких условиях можно ожидать, что ДНК, имеющая высокую гомологию, будет получена более эффективно с ростом температуры. Однако есть много потенциальных факторов влияющих на жесткость гибридизации, включая температуру, концентрацию зонда, длину зонда, ионную силу, время и концентрацию соли, и схожие условия жесткости могут быть созданы путем соответствующего выбора этих факторов специалистом в данной области.
Аминокислотная последовательность, кодируемая полинуклеотидом изобретения, может иметь делению, замену и/или вставку одной или нескольких аминокислот, при условии, что активность флавоноид 3',5'-гидроксилазы сохраняется.
По всему настоящему описанию, термин «идентичность» и «гомология» означают для полипептидных последовательностей (или аминокислотных последовательностей) или полшгуклеотидных последовательностей (или нуклеотидных последовательностей) количество (число) аминокислотных остатков или нуклеотидов, составляющих их, которые могут быть определены как идентичные между двумя цепями, с точки зрения взаимного соответствия между ними, что означает степень корреляции последовательности между двумя полипептидными последовательностями или двумя полинуклеотидными последовательностями, а «идентичность» и «гомология» могут быть легко рассчитаны. Известны многочисленные способы измерения гомологии между двумя полинуклеотидными последовательностями или полипептидными последовательностями и термины «идентичность» и «гомология» хорошо известны специалистам (например, см. Lesk, A.M. (Ed.), Computational Molecular Biology, Oxford University Press, New York, (1988); Smith, D.W. (Ed.), Biocomputing: Informatics and Genome Projects, Academic Press, New York, (1993); Grifin, A.M. & Grifin, H.G. (Ed.), Computer Analysis of Sequence Data: Part I, Human Press, New Jersey, (1994); von Heinje, G., Sequence Analysis in Molecular Biology, Academic Press, New York, (1987); Gribskov, M. & Devereux, J. (Ed.), Sequence Analysis Primer, M-Stockton Press, New York, (1991) и другие источники).
Также численные величины для «идентичность» и «гомология», используемые в настоящем описании, если не указано иное, могут быть числовыми значениями, рассчитанными с использованием программы поиска гомологии, известной специалистам в данной области, но предпочтительно они являются числовыми значениями, рассчитанными с использованием программы ClustalW из Mac Vector Application (версия 9.5, Oxford Molecular Ltd., Оксфорд, Англия).
Полинуклеотид (нуклеиновая кислота, ген) изобретения «кодирует» представляющий интерес белок. Здесь «кодирует» означает возможность осуществления экспрессии представляющего интерес белка в состоянии, в котором он проявляется свою активность. Также термин «кодирует» включает как кодирование структурной последовательности (экзона), который является непрерывным участком представляющего интерес белка, так и кодирование через промежуточную последовательность (интрона).
Ген, имеющий естественную нуклеотидную последовательность, может быть получен путем анализа с помощью ДНК-секвенатора, например, как описано в примере ниже. Также ДНК, кодирующие ферменты, обладающие модифицированной аминокислотной последовательностью, могут быть синтезированы с использованием сайт-специфического мутагенеза или ПЦР, на основе ДНК, имеющей естественную нуклеотидную последовательность. Например, модифицируемый фрагмент ДНК может быть получен обработкой рестрикционными ферментами естественной кДНК или геномной ДНК, и использован в качестве матрицы для сайт-специфического мутагенеза или ПЦР с использованием праймеров с искомой мутацией, для получения фрагмента ДНК, обладающего искомой модификацией. Фрагмент ДНК, имеющий мутацию, затем может быть связан с ДНК-фрагментом, кодирующим другую часть целевого фермента.
В ином случае, для получения ДНК, кодирующей фермент, содержащий укороченную аминокислотную последовательность, например, аминокислотная последовательность, которая длиннее, чем целевая аминокислотная последовательность, например, ДНК, кодирующая полноразмерную аминокислотную последовательность, разрезается выбранными рестрикционными ферментами, и если полученный фрагмент ДНК не кодирует целую аминокислотную последовательность, ДНК-фрагмент, содержащий утраченную часть последовательности может быть синтезирован и связан с ним.
Полинуклеотид изобретения вставляется в плазмиду вместе с 35S промотором вируса мозаики цветной капусты. 35S промотор предпочтительно является Е12 35S промотором (выше) обладающим повышенной экспрессией из-за связывания с двумя энхансерными последовательностями в тандеме с 35S промотором, обычно используемым в растительных трансформантах. Полинуклеотид изобретения и 35S промотор связаны функциональным образом, например, с представляющим интерес полинуклеотидом, расположенным за промотором. Плазмида изобретения используется для трансформации разновидности садовой розы.
Для целей изобретения термин «разновидность садовой розы» относится к растениям Rosa из семейства Rosaceae (научное название: Rosa hybridd) в рамках таксономической классификации. Разновидности садовой розы главным образом классифицируются как розы Hybrid Tea, Floribunda и Polyantha на основании их трех форм и размера цветка, при этом основной пигмент (антоцианин) в лепестках во всех линиях принадлежит к двум типам, цианидину и пеларонидину. Изобретение является методом преобразования основного антоцианина в лепестках розы в тип дельфинидина, и такие разновидности и линии пригодны для использования. Изобретение может быть применено к разновидностям цианидинового типа (например, Cool Water, Ocean Song и Fame), или разновидностям пеларгонидинового типа (например, Noblesse, Topless и Peach Avalanche).
Плазмида изобретения может дополнительно содержат непромоторные факторы, например, регуляторные последовательности, такие как терминаторы, последовательности сигналов полиаденилирования и последовательности начала репликации (ori); и селективные маркеры, такие как маркеры лекарственной устойчивости, флуоресцентные белковые маркеры, ферментативные маркеры и маркеры-рецепторы клеточной поверхности. Терминатор предпочтительно является nos-терминатором, расположенным за представляющим интерес полинуклеотидом. По всему настоящему описанию «nos-терминатор» относится к терминаторной последовательности гена нопалин-синтазы из Ti-плазмиды Agrobacterium tumefaciens.
Примеры
Настоящее изобретение далее будет объяснено более подробно с помощью примеров.
Пример 1: выделение кДНК F3'5'H Колокольчика
Два праймера CamF1 (5'-GTGAAGCCACCATGTCTATAG-3', SEQ ID NO: 3) и CamR1 (5'-GCATTTGCCTAGACAGTGTAAG-3', SEQ ID NO: 4) синтезировали на основе транслированной последовательности кДНК F3'5'H колокольчика (Campanula medium), зарегистрированной в базе данных GenBank (Учетный номер D14590). РНК экстрагировали из лепестков в стадии бутонизации коммерчески доступного колокольчика с использованием набора «RNeasy Mini Plant Kit» (Qiagen), а набор для OT-ПЦР использовали для синтеза 1-й цепи ДНК. 1-ю цепь ДНК использовали в качестве матрицы для ПЦР с использованием вышеупомянутых праймеров. Полученный фрагмент ДНК клонировали в pCR-TOPO II. Нуклеотидную последовательность Клона #4 (pSPB2561) анализировали на ДНК-секвенаторе (SEQ ID NO: 1). Хотя F3'5'H, зарегистрированный с учетным номером D14590, состоит из 523 остатков, F3'5'H, кодируемый клоном #4, состоит из 521 остатка, и, таким образом, демонстрирует 96% идентичность последовательности с F3'5'H, зарегистрированной как D14590.
Эталонный Пример 1: Введение pSPB130 в разновидность розы «Cool Water» (Конститутивная экспрессия полученного из анютиных глазок гена F3'5'H и полученного из торении гена 5AT)
Бинарный вектор pSPB130, описанный в PTL 3 (вектор, конститутивно экспрессирующий в растениях ген F3'5'H из анютиных глазок и ген антоцианин 5-ацилтрансферазы из торении) вводили в Agrobacterium (Agrobacterium tumefaciens) Agl0. Трансформированные Agrobacterium вводили в разновидность розы розовато-лилового цвета «Cool Water», способом, описанным в PTL 3, с получением 164 трансформантов. Анализ антоцианидиновых пигментов лепестков подтвердил запасание дельфинидина в 51 из 164 индивидуумов, с максимальным содержанием дельфинидина 76,1% (среднее: 36,6%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 1 ниже.
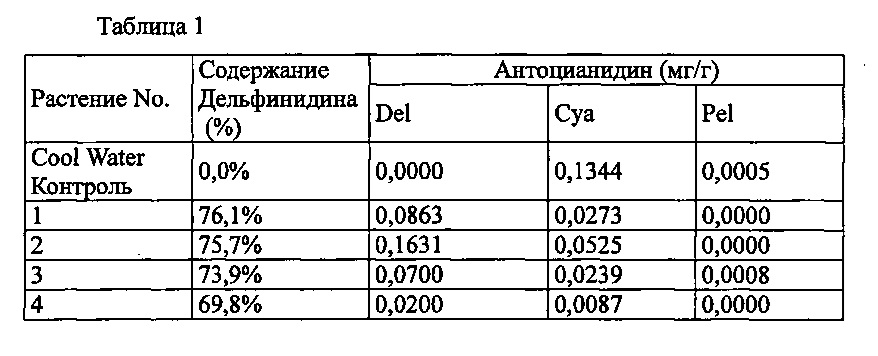

Del: дельфинидин, Cya: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Пример 2: Введение pSPB2564 в разновидность розы «Cool Water» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора Е12 35S)
Экспрессирующую кассету, содержащую кДНК гена колокольчика F3'5'H, полученную в Примере 1, и вставленную между промотором Е12 35S, описанным в PTL 3, и терминатором нопалинсинтазы (nos-терминатором), вставляли в бинарный вектор pBinPLUS (van Engelen et al. Transgenic Research 4, 288-290, 1995), и полученную плазмиду обозначали как pSPB2564. Эту pSPB2564 вставляли в Agrobacterium (Agrobacterium tumefaciens) Agl0. Трансформированные Agrobacterium использовали для трансформации разновидности роз розовато-лилового цвета «Cool Water», способом, описанным в PTL 3, с получением 55 трансформантов. Анализ антопианидиновых пигментов лепестков подтвердил запасание дельфинидина в 48 из 55 индивидуумов, с максимальным содержанием дельфинидина 97,3% (среднее: 61,22%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 2 ниже.
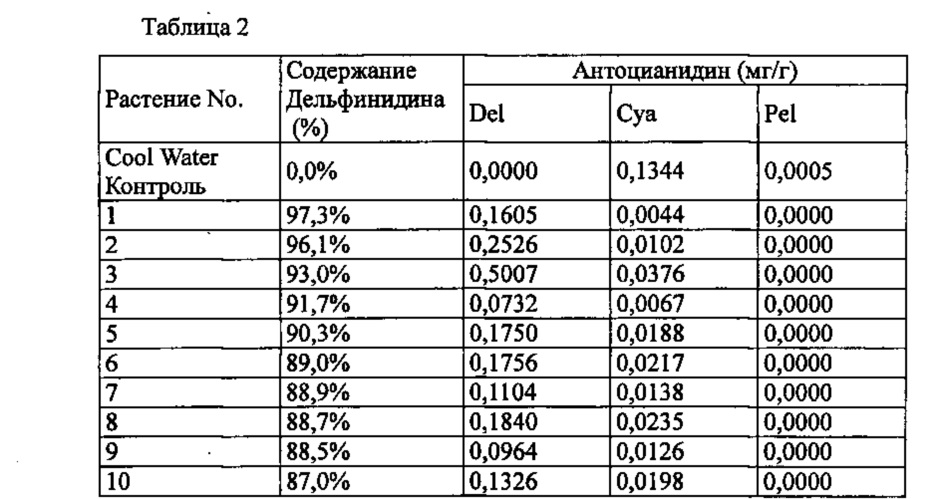
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Пример 3: Введение pSPB2564 в разновидность розы «Fame» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора Е12 35S)
Agrobacterium с введенной pSPB2564 использовали для трансформации розовой разновидности розы «Fame», с получением 114 трансформантов. Запасание дельфинидина было подтверждено в 46 из 107 индивидуумов, подвергнутых анализу пигментов антиоцианидинов в лепестках, с максимальным содержанием дельфинидина 98,9% (среднее: 51,2%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 3 ниже.
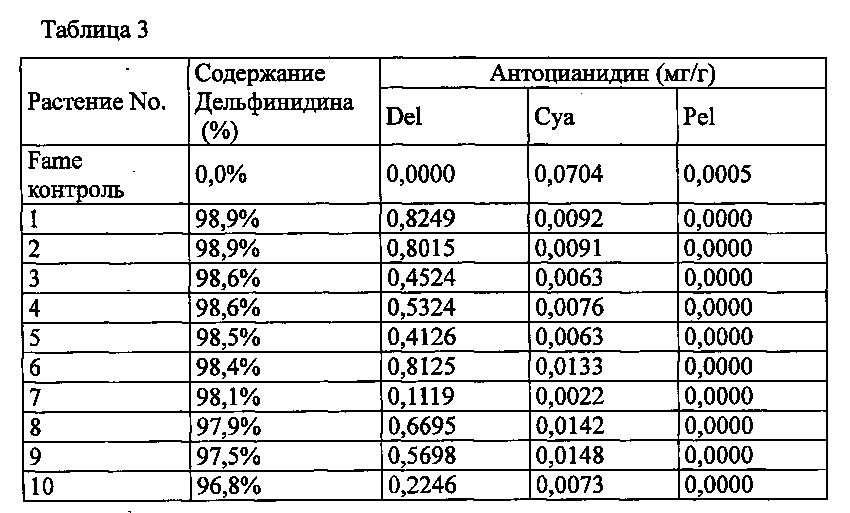
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Пример 4: Введение pSPB2564 в разновидность розы «Fame» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора Е12 35S)
Agrobacterium с введенной pSPB2564 использовали для трансформации розовой разновидности розы «Fame», с получением 55 трансформантов. Анализ антоцианидиновых пигментов лепестков подтвердил запасание дельфинидина в 28 из 55 индивидуумов, с максимальным содержанием дельфинидина 98,9% (среднее: 51,4%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 4 ниже.
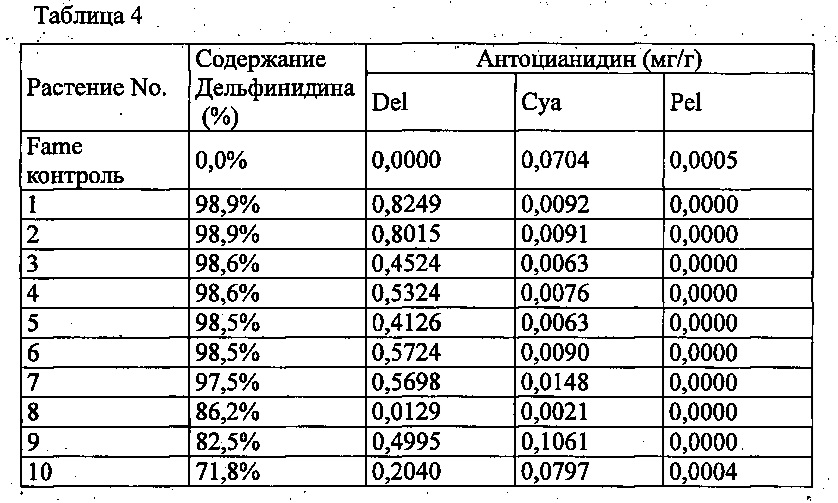
Del: дельфинидин, Cya: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Пример 5: Введение pSPB2564 в разновидность розы «Noblesse» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора Е12 35S)
Agrobacterium с введенной pSPB2564 использовали для трансформации оранжево-розовой разновидности розы «Noblesse», с получением 3 трансформантов. Анализ антоцианидиновых пигментов лепестков подтвердил запасание дельфинидина во всех 3 индивидуумах, с максимальным содержанием дельфинидина 98,9% (среднее: 51,4%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 5 ниже.
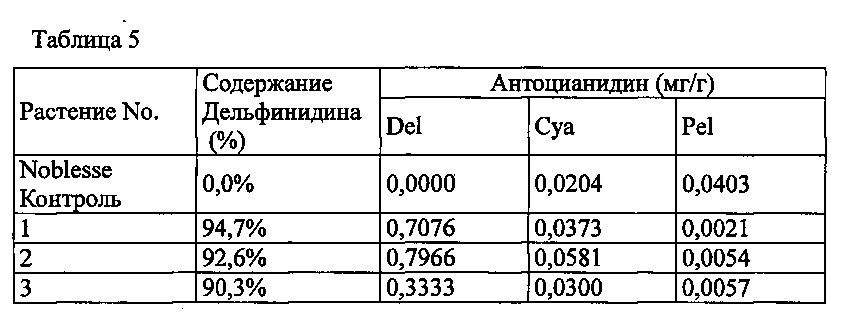
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Пример 6: Введение pSPB2564 в разновидность розы «Ocean Song» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора Е12 35S)
Agrobacterium с введенной pSPB2564 использовали для трансформации разновидности роз розовато-лилового цвета «Ocean Song», с получением 40 трансформантов. Анализ антоцианидиновых пигментов лепестков подтвердил запасание дельфинидина в 33 из 40 индивидуумов, с максимальным содержанием дельфинидина 99,2% (среднее: 39,7%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 6 ниже.
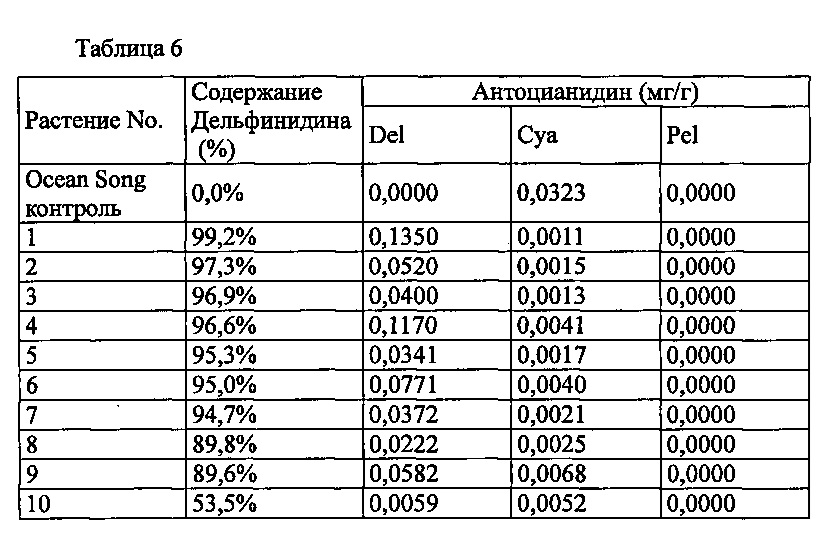
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Пример 7: Введение pSPB2564 в разновидность розы «Peach Avalanche» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора Е12 35S)
Agrobacterium с введенной pSPB2564 использовали для трансформации светло оранжево-розовой разновидности розы «Peach Avalanche», с получением 11 трансформантов. Анализ антоцианидиновых пигментов лепестков подтвердил запасание дельфинидина в 1 из 11 индивидуумов, с максимальным содержанием дельфинидина 99,3%.
Значения анализа для трансформантов, в которых выработка дельфинидина был подтверждена, показаны в Таблице 7 ниже.
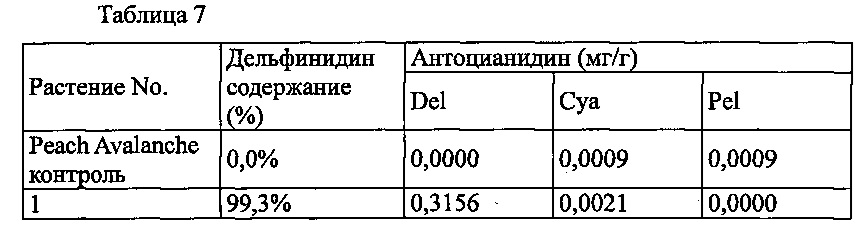
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Пример 8: Введение pSPB2564 в разновидность розы «Topless» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора Е12 35S)
Agrobacterium с введенной pSPB2564 использовали для трансформации оранжево-розовой разновидности розы «Topless», с получением 11 трансформантов. Анализ антоцианидиновых пигментов лепестков подтвердил запасание дельфинидина в 10 из 11 индивидуумов, с максимальным содержанием дельфинидина 99,4% (среднее: 64,4%).
Значения анализа для трансформантов, в которых выработка дельфинидина был подтверждена, показаны в Таблице 8 ниже.
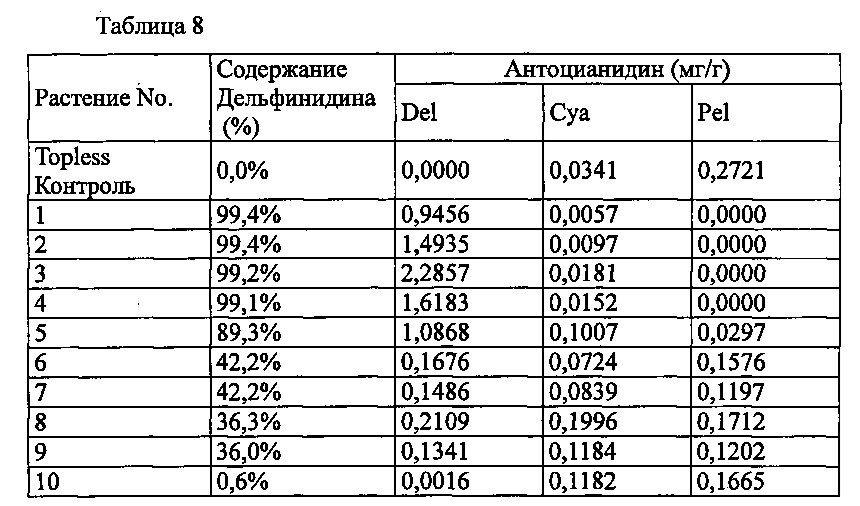
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Эталонный Пример 2: Введение pSPB5202 в разновидность розы «Fame» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора F3'5'H, полученного из анютиных глазок)
Последовательность промотора F3'5'H из анютиных глазок, полученная расщеплением плазмиды pSFL620, описанной в PTL 9, с помощью HindIII и PstI и кДНК последовательность из колокольчика, полученную в Примере 1, объединяли с терминатором белка теплового шока из плазмиды pRI201-AN (Takara Bio, Inc.) и вставляли в бинарный вектор pBinPlus. Полученный бинарный вектор был обозначен как pSPB5202. Этот pSPB5202 вставляли в Agrobacterium (Agrobacterium tumefaciens) Agl0. Трансформированные Agrobacterium использовали для трансформации разновидности розы розовато-лилового цвета «Fame», способом, описанным в PTL 3, с получением 56 трансформантов. Запасание дельфинидина могло быть подтверждено в 4 из 52 индивидуумов, подвергнутых анализу пигмента антиоцианидина в лепестках, с максимальным содержанием дельфинидина 10,8% (среднее: 3,9%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 9 ниже.
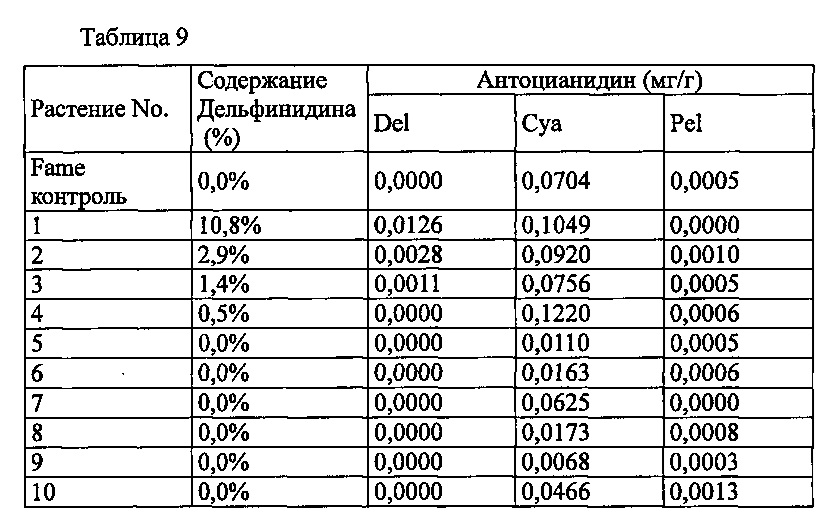
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Несмотря на то, что промотор F3'5'H из анютиных глазок удовлетворительно экспрессирует ген F3'5'H из анютиных глазок в розе (см. PTL 9), а терминатор белка теплового шока очень эффективен для экспрессии инородных белков в растениях (Plant Cell Physiol (2010) 51, 328-332), в трансформированных лепестках розы были обнаружены только очень низкие уровни выработки дельфинидина, что подтверждает, что ген F3'5'H колокольчика не может функционировать достаточным образом. Это указывает на то, что для эффективной выработки дельфинидина в конкретных разновидностях розы необходимо выбрать комбинацию специфического промотора и специфического гена F3'5'H, или комбинацию специфического промотора, специфического гена F3'5'H и специфического терминатора гена.
Эталонный Пример 3: Введение pSPB5204 в разновидность розы «Fame» (Экспрессия полученного из колокольчика гена F3'5'H под контролем промотора F3'5'H, полученного из периллы)
В результате расщепления плазмиды pSFL205, описанной в PTL 10, HindIII и BamHI, был извлечен примерно 1,1 тыс.о. ДНК-фрагмент, содержащий полученный из периллы участок промотора антоцианин 3-ацилтрансферазы, который соединяли с pBinPlus, гидролизованной HindIII и BamHI, с получением pSPB3756. Также с помощью ПНР с использованием плазмиды pSPB3311, описанной в PTL 10, в качестве матрицы и трех различных синтетических нуклеотидных праймеров: Pf3AT-FW-Sal (5'-ATGTCGACTAAATGTATGTAATTAAC-3', SEQ ID NO: 5), Pf3ATt-R1 (5'-ACTCAACACTTTATTAATTG-3', SEQ ID NO: 6) and Pf3ATt-F1 (5'-ATGTAACAATTAATTAAGTG-3', SEQ ID NO: 7), добавляли участок распознавания рестрикционного фермента Sali на 5'-конец полученного из периллы участка терминатора антоцианин ацилтрансферазы, а участок распознавания Paci на 5'-конце в том же участке терминатора разрушали. Фрагмент ДНК, полученный с помощью ПЦР, клонировали в рТОРО. Фрагмент ДНК, полученный расщеплением полученной плазмиды с помощью ΚpnΙ и PacI, связали с фрагментом ДНК, полученным расщеплением pSPB3756 с фрагментом ДНК, полученным расщеплением pSPB3756 с ΚpnΙ и PacI, с получением pSPB5201. Плазмиду pSPB5201, расщепленную SmaI и SpeI, связывали с кДНК колокольчика, полученной в Примере 1, и полученный бинарный вектор обозначили как pSPB5204. Этот pSPB5204 вводили в Agrobacterium (Agrobacterium tumefaciens) Agl0. Трансформированные Agrobacterium использовали для трансформации розовой разновидности розы «Fame» способом, описанным в PTL 3, с получением 63 трансформантов. Запасание дельфинидина могло быть подтверждено только в 8 из 61 индивидуумов, подвергнутого анализу пигмента в лепестках, с максимальным содержанием дельфинидина 2,8% (среднее: 1,3%).
Значения анализа для репрезентативных трансформантов показаны в Таблице 10 ниже.
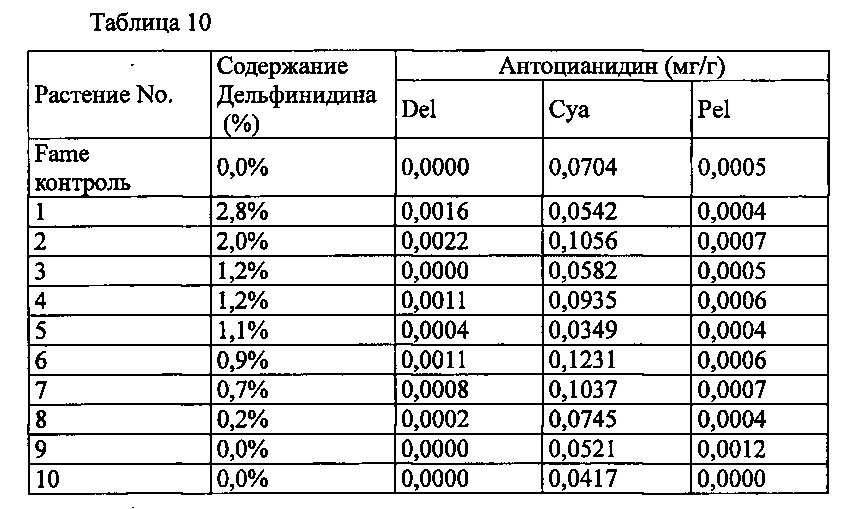
Del: дельфинидин, Суа: цианидин, Pel: пеларгонидин
Содержание дельфинидина: Доля дельфинидина от общего количества антоцианидинов.
Несмотря на то, что промотор антоцианин 3-ацилтрансферазы из периллы удовлетворительно экспрессирует ген антоцианин 3-ацилтрансферазы из периллы в розе, петунии и хризантеме (см. PTL 9), в лепестках таких трансформантов розы вырабатывались только очень низкие уровни дельфинидина, что подтверждает, что ген F3'5'H колокольчика не может функционировать достаточным образом. Это указывает на то, что для эффективной выработки дельфинидина в розе необходимо выбрать комбинацию специфического промотора и специфического гена F3'5'H, или комбинацию специфического промотора, специфического гена F3'5'H и специфического терминатора гена.
Промышленная применимость
Путем трансформации с использованием плазмиды, содержащей ген флавоноид 3',5'-гидроксилазы колокольчика по изобретению под контролем 35S промотора вируса мозаики цветной капусты с помощью способа с Agrobacterium, можно значимо повысить содержание дельфинидина в лепестках разновидностей садовой розы. Таким образом, изобретение может быть соответствующим образом использовано для получения растений розы с измененной окраской цветка.






Содержащая фруктовый сок основа алкогольного напитка и напиток, полученный разведением этой основы
Алкогольный напиток, содержащий фруктовый сок
Гомологи глицерин-3-фосфатацилтрансферазы (гфат) и их использование
Композиции жирных кислот, имеющие новую пропорцию жирных кислот
Глицерол-3-фосфатацилтрансфераза
Новое соединение, содержащееся в голубой розе
Новые гены ацилтрансферазы лизофосфатидной кислоты
Гомологи фосфатазы фосфатидной кислоты и их применение
Гены диацилглицерол-ацилтрансферазы и их использование
Полинуклеотид, кодирующий гомолог ацил-соа-синтетазы, и его применение
Содержащая фруктовый сок основа алкогольного напитка и напиток, полученный разведением этой основы
Алкогольный напиток, содержащий фруктовый сок
Гомологи глицерин-3-фосфатацилтрансферазы (гфат) и их использование
Композиции жирных кислот, имеющие новую пропорцию жирных кислот
Глицерол-3-фосфатацилтрансфераза
Новое соединение, содержащееся в голубой розе
Новые гены ацилтрансферазы лизофосфатидной кислоты
Гомологи фосфатазы фосфатидной кислоты и их применение
Гены диацилглицерол-ацилтрансферазы и их использование
Полинуклеотид, кодирующий гомолог ацил-соа-синтетазы, и его применение

