Результат интеллектуальной деятельности: НОВЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ ДИМЕРИЗАЦИЮ С-МЕТ, И ИХ ПРИМЕНЕНИЯ
Вид РИД
Изобретение
Область изобретения
Данное изобретение относится к новым антителам, способным специфично связываться с человеческим c-Met-рецептором и/или способным специфично ингибировать тирозинкиназную активность указанного рецептора, особенно к моноклональным антителам мышиного, химерного и гуманизированного происхождения, а также к аминокислотным и нуклеиновокислотным последовательностям, кодирующим эти антитела. В частности, антитела в соответствии с изобретением способны ингибировать димеризацию c-Met. Аналогичным образом изобретение включает в себя применение этих антител в качестве лекарственного средства для профилактики и/или терапевтического лечения раковых заболеваний или любой патологии, связанной со сверхэкспрессией указанного рецептора, а также в способах и наборах для диагностики заболеваний, связанных со сверхэкспрессией c-Met. Наконец, изобретение включает продукты и/или композиции, включающие такие антитела в сочетании с другими антителами и/или химическими соединениями, направленными против других факторов роста, участвующих в опухолевой прогрессии и метастазировании, и/или соединениями и/или противораковыми агентами или агентами, конъюгированными с токсинами, и их применение для профилактики и/или лечения некоторых раковых заболеваний.
Уровень техники
Агенты, мишенями которых является рецепторная тирозинкиназу (receptor tyrosine kinase, RTK), такие как ингибиторы трастуцумаб, цетуксимаб, бевацизумаб, иматиниб и гефитиниб, являются иллюстрацией существующего интереса к выбору этого класса белков в качестве мишеней при лечении отдельных раковых заболеваний.
c-Met является первым членом подсемейства RTK, которое также включает RON и SEA. Семейство c-Met RTK структурно отличается от других семейств RTK и является единственным известным высокоаффинным рецептором для фактора роста гепатоцитов (HGF), называемого также scater-фактором (SF) [D.P. Bottaro et al., Science 1991, 251:802-804; L Naldini et al., Eur. Mol. Biol. Org. J. 1991, 10:2867-2878]. c-Met и HGF широко экспрессируются различными тканями, и их экспрессия, как правило, ограничена клетками эпителиального и мезенхимального происхождения соответственно [M.F. Di Renzo et al., Oncogene 1991, 6:1997-2003; E. Sonnenberg et al., J. Cell. Biol. 1993, 123:223-235]. Оба они необходимы для нормального развития млекопитающих, и, как было показано, особенно важны для клеточной миграции, морфогенной дифференцировки и формирования трехмерных трубчатых структур, а также для роста и ангиогенеза [F. Baldt et al., Nature 1995, 376:768-771; С.Schmidt et al., Nature. 1995:373:699-702; Tsarfaty et al., Science 1994, 263:98-101]. В то время, как было показано, что контролируемое регулирование c-Met и HGF важно для развития млекопитающих, поддержания и репарации тканей [Nagayama Т, Nagayama M, Kohara S, Kamiguchi H, Shibuya M, Katoh Y, Itoh J, Shinohara Y., Brain Res. 2004, 5; 999(2):155-66; Tahara Y, Ido A, Yamamoto S, Miyata Y, Uto H, Hori T, Hayashi K, Tsubouchi H., J Pharmacol Exp Ther. 2003, 307(1):146-51], нарушение их регуляции участвует в прогрессировании раковых заболеваний.
Аберрантная сигнализация, движимая несоответствующей активацией с-Met, является одним из наиболее часто наблюдаемых изменений в раковых опухолях человека и играет важнейшую роль в образовании опухолей и метастазировании [Birchmeier et al., Nat. Rev. Mol. Cell Biol. 2003, 4:915-925; L. Trusolino and Comoglio P. M., Nat Rev. Cancer. 2002, 2(4):289-300].
Несоответствующая активация c-Met может возникнуть путем лиганд-зависимых и лиганд-независимых механизмов, которые включают сверхэкспрессию c-Met и/или паракринную или аутокринную активацию, или путем усиления функции мутации [J.G. Christensen, Burrows J. and Salgia R., Cancer Latters. 2005, 226:1-26]. Однако олигомеризация c-Met-рецептора в присутствии или в отсутствие лиганда необходима для регулирования аффинности связывания и кинетики связывания киназы с АТФ и тирозин-содержащими пептидными субстратами [Hays JL, Watowich SJ, Biochemistry, 2004 Aug 17, 43:10570-8]. Активированный c-Met привлекает сигнальные эффекторы к месту своего скопления, расположенному в цитоплазматическом домене, что приводит к активации ряда ключевых сигнальных путей, в том числе Ras-MAPK, PI3K, Src и Stat3 [Gao CF, Vande Woude GF, Cell Res. 2005, 15(1):49-51; Furge KA, Zhang YW, Vande Woude GF, Oncogene. 2000, 19(49):5582-9]. Эти пути имеют важное значение для пролиферации опухолевых клеток, инвазии и ангиогенеза и для уклонения от апоптоза [Furge KA, Zhang YW, Vande Woude GF, Oncogene, 2000, 19(49):5582-9; Gu H, Neel BG, Trends Cell Biol. 2003 Mar, 13(3):122-30; Fan S, Ma YX, Wang JA, Yuan RQ, Meng Q, Cao Y, Laterra JJ, Goldberg ID, Rosen EM, Oncogene. 2000 Apr 27, 19(18):2212-23]. Кроме того, уникальным аспектом c-Met-сигнализации по сравнению с другими РТК является выявленное взаимодействие его с фокальными адгезионными комплексами и некиназными связывающими партнерами, такими как интегрины α6β4 [Trusolino L, Bertotti A, Comoglio PM, Cell. 2001, 107:643-54], CD44v6 [Van der Voort R, Taher ТЕ, Wielenga VJ, Spaargaren M, Prevo R, Smit L, David G, Hartmann G, Gherardi E, Pals ST, J Biol Chem. 1999, 274(10):6499-506], плексин В1 или семафорины [Giordano S, Corso S, Conrotto P, Artigiani S, Gilestro G, Barberis D, Tamagnone L, Comoglio PM, Nat Cell Biol. 2002, 4(9)720-4; Conrotto P, Valdembri D, Corso S, Serini G, Tamagnone L, Comoglio PM, Bussolino F, Giordano S, Blood. 2005, 105(11):4321-9; Conrotto P, Corso S, Gamberini S, Comoglio PM, Giordano S, Oncogene. 2004, 23:5131-7], которые могут еще больше усложнить регуляцию клеточной функции этим рецептором. Наконец, недавние данные показывают, что c-Met может быть вовлечен в формирование опухолевой резистентности к гефитинибу или эрлотинибу, подтверждая, что комбинация соединения, направленного на EGFR, и c-Met, может представлять значительный интерес [Engelman JA at al., Science, 2007, 316:1039-43].
В последние несколько лет многие различные стратегии были разработаны с целью смягчения c-Met-сигнализации в линиях раковых клеток. Эти стратегии включают i) нейтрализующие антитела против c-Met или HGF/SF [Сао В, Su Y, Oskarsson M, Zhao P, Kort EJ, Fisher RJ, Wang LM, Vande Woude GF, Proc Natl Acad Sci USA. 2001, 98(13):7443-8; Martens T, Schmidt NO, Eckerich C, Fillbrandt R, Merchant M, Schwall R, Westphal M, Lamszus K, Clin Cancer Res. 2006, 12(20):6144-52] или использование HGF/SF-антагониста NK4 для предотвращения связывания лиганда с c-Met [Kuba K, Matsumoto K, Date K, Shimura H, Tanaka M, Nakamura T, Cancer Res., 2000, 60:6737-43], ii) низкомолекулярные ингибиторы сайта связывания АТФ на c-Met для блокирования киназной активности [Christensen JG, Schreck R, Burrows J, Kuruganti P, Chan E, Le P, Chen J, Wang X, Ruslim L, Blake R, Lipson KE, Ramphal J, Do S, Cui JJ, Cherrington JM, Mendel DB, Cancer Res. 2003, 63:7345-55], iii) сконструированный полипептидный домен SH2, который препятствует подходу к месту скопления РНК или рибозима, уменьшая экспрессию рецептора или лиганда. Большинство из этих подходов отображает селективное ингибирование c-Met, приводящее к ингибированию опухоли и показывающее, что c-Met может представлять интерес для терапевтического вмешательства при раке.
Среди молекул, созданных для действия на c-Met, есть некоторые антитела.
Наиболее подробно описанным является антитело анти-c-Met 5D5, созданное Genentech [WO96/38557], которое ведет себя как мощный агонист при добавлении его одного в различных моделях, и как антагонист при использовании его в качестве фрагмента Fab. Моновалентная сконструированная форма этого антитела, описанная как 5D5 с одной ветвью (OA5D5) и полученная как рекомбинантный белок в Е.Coli, также является предметом патентной заявки [WO2006/015371] от Genentech. Однако эта молекула, которая не может рассматриваться в качестве антитела из-за своего особого остова, демонстрирует также мутации, которые могут быть иммуногенными у людей. С точки зрения активности эта негликозилированная молекула лишена эффекторных функций и, наконец, отсутствие четких данных показывает, что OA5D5 ингибирует димеризацию c-Met. Кроме того, при испытании на модели in vivo G55, клеточной линии глиобластомы, которая экспрессирует мРНК и белок c-Met, но не HGF, и которая растет независимо от лиганда, анти-c-Met с одной ветвью не оказывает существенного влияния на рост опухоли G55, подтверждая, что OA5D5 действует главным образом путем блокирования связывания HGF и не способен воздействовать на опухоли, активированные независимо от HGF [Martens Т. et al, Clin. Cancer Res., 2006, 12(20):6144-6152].
Другое антитело, направленное на c-Met, Pfizer описывает как антитело, действующее "по большей части как c-Met-антагонист, а в некоторых случаях как с-Met-агонист" [WO 2005/016382]. Нет данных, свидетельствующих о каком-либо влиянии антител Pfizer'a на димеризацию c-Met, описанную в этой заявке.
Одним из инновационных аспектов данного изобретения является создание мышиных моноклональных антител без внутренней агонистической активности и ингибирующих димеризацию c-Met. Кроме целевых лиганд-зависимых опухолей, этот подход будет также ограничивать лиганд-независимую активацию c-Met благодаря его сверхэкспрессии или мутациям во внутриклеточных доменах, которым по-прежнему необходима олигомеризация для сигнализации. Другим аспектом активности таких антител может быть стерическое препятствие взаимодействию c-Met с его партнерами, что приведет к ослаблению функций с-Met. Эти антитела будут гуманизированными и предпочтительно сконструированными в качестве человеческого lgG1, но не ограничиваясь этим, для получения эффекторных функций, таких как ADCC и CDC, в дополнение к функциям, связанным со специфической блокадой c-Met-рецепторов.
Раскрытие изобретения
Удивительно, но в первый раз изобретателям удалось создать антитело, способное связываться с c-Met, а также способное ингибировать димеризацию с-Met. Если правда, что в предыдущей области иногда полагали, что антитело, способное ингибировать димеризацию c-Met с его партнерами, может быть интересным, то антитело, способное делать это, никогда не было раскрыто или ясно предложено. Кроме того, что касается специфичности антитела, то достижение успеха в получении такого активного антитела не было очевидным для всех.
В первом аспекте предметом данного изобретения является способ получения и отбора антител в соответствии с изобретением.
В частности, изобретение относится к способу отбора анти-с-Met-антитела или одного из его функциональных фрагментов или производных, способного ингибировать как лиганд-зависимую, так и лиганд-независимую активацию c-Met, при этом указанный способ включает следующие этапы:
i) скрининг полученных антител и отбор антител, способных специфично связываться с c-Met;
ii) оценка in vitro отобранных антител этапа i) и отбор антител, способных ингибировать по меньшей мере на 50%, предпочтительно по меньшей мере на 60%, 70% или 80%, опухолевую пролиферацию клеток по меньшей мере одного типа опухоли; а затем
iii) испытание отобранных антител этапа ii) и отбор антител, способных ингибировать димеризацию c-Met.
Как объяснялось ранее, ингибирование димеризации c-Met является главным аспектом изобретения, т.к. такие антитела будут представлять реальный интерес для широкой группы пациентов. Не только лиганд-зависимый активированный c-Met-рак, как это было до данного изобретения, но также и лиганд-независимый активированный c-Met-рак можно лечить с помощью антител, созданных в способе данного изобретения.
Создание антитела может быть реализовано любым способом, известным специалисту в данной области, таким как, например, слияние миеломой клетки с клетками селезенки иммунизированных мышей или других видов, совместимых с отобранными клетками миеломы [Kohler & Milstein, 1975, Nature, 256:495-497]. Иммунизированные животные могут включать трансгенных мышей с человеческими иммуноглобулиновыми локусами, которые затем непосредственно продуцируют человеческие антитела. Другое возможное воплощение может состоять в использовании технологий фагового дисплея для скрининга библиотек.
Этап скрининга i) может быть реализован с помощью любого способа или процесса, известного специалисту в данной области. В качестве не ограничивающих примеров можно упомянуть ИФА, BIAcore, иммуногистохимию, FACS-анализ и функциональную оценку. Предпочтительный способ состоит в скрининге с помощью ИФА по рекомбинантному белку c-Met, а затем в FACS-анализе по меньшей мере опухолевой клеточной линии, чтобы убедиться, что производимые антитела смогут также распознавать нативные рецепторы на опухолевых клетках. Этот способ более точно будет описан в следующих примерах.
Таким же образом этап ii) может быть классически реализован в соответствии с известным способом или процессом, таким как, например, с помощью 3H-тимидина или любого другого ДНК-окрашивающего агента, МТТ-тест, определение АТФ и т.д. Предпочтительной опухолевой клеточной моделью в данном изобретении может быть ВхРС3-модель.
Под ингибированием димеризации c-Met предпочтительно следует понимать гомодимеризацию c-Met.
В предпочтительном воплощении этапа iii) отбора способа изобретения указанный этап iii) состоит в оценке антител на основе BRET-анализа клеток, экспрессирующих c-Met-RLuc/c-Met-YFP, и в отборе антител, способных ингибировать по меньшей мере на 30%, предпочтительно на 35%, 40%, 45%, 50%, 55% и наиболее предпочтительно на 60% BRET-сигнал.
Технология BRET представляет собой технологию, известную в качестве образцовой для димеризации белка [Angers et al., PNAS, 2000, 97:3684-89].
BRET-технология, используемая в этапе iii) способа, хорошо известна специалисту в данной области и будет подробно описана в следующих примерах. В частности, BRET (bioluminescence resonance energy transfer, резонансный перенос энергии биолюминесценции) представляет собой нерадиационный перенос энергии, возникающий между биолюминесцентным донором (люциферазой Renilla (Rluc)) и флуоресцентным акцептором, мутантом GFP (green fluorescent protein, зеленый флуоресцентный белок) или YFP (yellow fluorescent protein, желтый флуоресцентный белок). В данном случае был использован EYFP (enhanced yellow fluorescent protein, усиленный желтый флуоресцентный белок). Эффективность переноса зависит от ориентации и расстояния между донором и акцептором. Затем, передача энергии может происходить только тогда, когда две молекулы находятся в непосредственной близости (1-10 нм). Это свойство используется для проведения анализа белково-белковых взаимодействий. Действительно, в целях изучения взаимодействия между двумя партнерами первый из них генетически сливают с люциферазой Renilla, а второй с желтым мутантом GFP. Гибридные белки, как правило, но не обязательно, экспрессируются клетками млекопитающих. В присутствии своего мембранно-проницаемого субстрата (коэлентеразин) Rluc испускает синий свет. Если мутант GFP находится ближе 10 нм к Rluc, то может возникнуть передача энергии, и будет обнаружен дополнительный желтый сигнал. Сигнал BRET измеряется как соотношение между светом, излучаемым акцептором, и светом, излучаемым донором. Так, BRET-сигнал будет увеличиваться по мере сближения двух гибридных белков, или если конформационное изменение сблизит Rluc и GFP-мутант.
Если BRET-анализ включен в предпочтительное воплощение, то любой способ, известный специалисту в данной области, можно использовать для измерения димеризации c-Met. He ограничивая, можно отметить следующие технологии: FRET (fluorescence resonance energy transfer, резонансный перенос энергии флуоресценции), HTRF (homogenous time resolved fluorescence, гомогенная флуоресценция с временным разрешением), FLIM (fluorescence lifetime imaging microscopy, флуоресцентная микроскопия с изображением по времени жизни флуоресценции) или SW-FCCS (single wavelength fluorescence cross-correlation spectroscopy, кросс-корреляционная спектроскопия флуоресценции одной длины волны).
Также могут быть использованы другие классические технологии, например коиммунопреципитация, Alpha Screen, химическое сшивание, двойной гибрид, аффинная хроматография, ИФА и фар-вестерн-блоттинг.
Во втором аспекте предметом изобретения является изолированное антитело или один из его функциональных фрагментов или производных, полученных в указанном способе. Указанное антитело или один из его указанных фрагментов или производных способно к специфическому связыванию человеческого c-Met, и кроме того, при необходимости предпочтительно способно ингибировать природное присоединение его лиганда HGF и/или способно специфически ингибировать тирозинкиназную активность указанного c-Met; указанное антитело также будет способно ингибировать димеризацию c-Met. В частности, указанные антитела будут способны ингибировать как лиганд-зависимую, так и лиганд-независимую активацию c-Met.
Выражения "функциональные фрагменты и производные" подробно будут определены ниже в данном описании.
Следует понимать здесь, что изобретение не относится к антителам в природной форме, то есть они находятся не в их природной среде, но что они могут быть выделены или получены путем очистки из природных источников, либо получены путем генетической рекомбинации, либо путем химического синтеза, и что они могут содержать неприродные аминокислоты, как будет описано ниже.
В частности, в соответствии с другим аспектом изобретения, заявлено антитело или один из его функциональных фрагментов или производных, охарактеризованное тем, что оно содержит по меньшей мере один участок, определяющий комплементарность (CDR), выбранный среди CDR, содержащих аминокислотную последовательность SEQ ID №№1-17 и 56-61.
Любое антитело или фрагмент или производное, имеющее по меньшей мере один CDR, последовательность которого по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания с последовательностью SEQ ID №№1-17 и 56-61, следует понимать как эквивалент и, следовательно, как часть изобретения.
Термин «CDR-участки», или «CDR», предназначен для обозначения гипервариабельных участков тяжелых и легких цепей иммуноглобулинов, как это определено в IMGT.
Уникальная нумерация IMGT была создана для сравнения вариабельных доменов независимо от антигенного рецептора, типа цепи или вида [Lefranc M.-P., Immunology Today 18, 509 (1997) / Lefranc M.-P., The Immunologist, 7, 132-136 (1999) / Lefranc, M.-P., Pommie, С., Ruiz, M., Giudicelli, V., Foulquier, E., Truong, L., Thouvenin-Contet, V. and Lefranc, Dev. Comp. Immunol., 27, 55-77 (2003)]. В уникальной нумерации IMGT консервативные аминокислоты всегда занимают одну и ту же позицию, например цистеин 23 (1-CYS), триптофан 41 (CONSERVED-TRP), гидрофобная аминокислота 89, цистеин 104 (2-CYS), фенилаланин или триптофан 118 (J-PHE или J-TRP). Уникальная нумерация IMGT предусматривает стандартизированное разграничение каркасных участков (FR1-IMGT: позиции 1-26, FR2-IMGT: 39-55, FR3-IMGT: 66-104 и FR4-IMGT: 118-128) и участков, определяющих комплементарность: CDR1-IMGT: 27-38, CDR2-IMGT: 56-65 и CDR3-IMGT: 105-117. Т.к. разрывы представляют собой незанятые позиции, то длины CDR-IMGT (показаны в скобках и разделенных точками, например [8.8.13]) становятся важной информацией. Уникальная нумерация IMGT используется в 2D-графическом представлении, созданном как IMGT Colliers de Perles [Ruiz, M. and Lefranc, M.-P., Immunogenetics, 53, 857-883 (2002) / Kaas, Q. and Lefranc, M.-P., Current Bioinformatics, 2, 21-30 (2007)], и в 3D-CTpyKrypax в IMGT/3Dstructure-DB [Kaas, Q., Ruiz, M. and Lefranc, M.-P., Т cell receptor and MHC structural data. Nucl. Acids. Res., 32, D208-D210 (2004)].
Существует три тяжелых цепи CDR и 3 легких цепи CDR. Термин «CDR» применяется здесь для обозначения в зависимости от случая одного из этих участков или нескольких, или даже всех этих участков, которые содержат большинство аминокислотных остатков, ответственных за аффинное связывание антитела с антигеном или эпитопом, который он распознает.
Термин "процент идентичности" двух нуклеиновокислотных или аминокислотных последовательностей в контексте данного изобретения применяется для указания процента нуклеотидов или идентичных аминокислотных остатков в двух сравниваемых последовательностях, полученного после лучшего выравнивания (оптимального выравнивания); этот процент не чисто статистический и различия между двумя последовательностями распределяются случайным образом и по всей их длине. Сравнение двух нуклеиновокислотных или аминокислотных последовательностей традиционно проводится путем сравнения этих последовательностей после выравнивания их оптимальным образом; указанное сравнение может осуществляться по сегментам или с помощью "окна сравнения". Оптимальное выравнивание последовательностей для сравнения может быть осуществлено, в дополнение к ручному, с помощью алгоритма локальной гомологии Смита-Уотермана (1981) [Ad. App. Math. 2:482], с помощью алгоритма локальной гомологии Нидлмана-Вунша (1970) [J. Mol. Biol. 48:443], с помощью способа поиска сходства Пирсона и Липмана (1988) [Proc. Natl. Acad. Sci. USA 85:2444), с помощью компьютерного программного обеспечения с использованием этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI, либо с помощью программного обеспечения для сравнения BLAST N или BLAST Р).
Процент идентичности двух нуклеиновокислотных или аминокислотных последовательностей определяют путем сравнения этих двух последовательностей, выравненных оптимальным образом, в котором сравниваемая нуклеиновокислотная или аминокислотная последовательность может включать добавления или делеции по сравнению с исходной последовательностью для оптимального выравнивания этих двух последовательностей. Процент идентичности рассчитывается путем определения числа идентичных позиций, в которых нуклеотид или аминокислотный остаток идентичен для двух последовательностей, деления этого количества идентичных позиций на общее количество позиций в «окне сравнения» и умножения полученного результата на 100, чтобы получить процент идентичности этих двух последовательностей.
Например, программа BLAST "BLAST 2 sequences" (Tatusova et al, "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol, 1999, Lett. 174:247-250), доступная на сайте http://www.ncbi.nlm.nih.gov/gorf/bl2.html, может быть использована с параметрами по умолчанию (в частности с параметрами "штраф за открытие делеции": 5 и "штраф за удлинение делеции": 2; выбранной матрицей будет, например, матрица "BLOSUM 62", предложенная в программе); процентная идентичность двух сравниваемых последовательностей рассчитывается непосредственно в программе.
Среди аминокислотных последовательностей, имеющих по меньшей мере 80%, по меньшей мере 85%, 90%, 95% и 98% идентичности с исходной аминокислотной последовательностью, предпочтительными являются те, которые по сравнению с исходной аминокислотной последовательностью имеют определенные изменения, в частности делецию, добавление или замену по меньшей мере одной аминокислоты, усечения или удлинения. В случае замены одной или более последовательных или непоследовательных аминокислот(ы), предпочтительными являются такие замены, в которых замененные аминокислоты заменены "эквивалентными" аминокислотами. Выражение "эквивалентные аминокислоты" используется здесь для обозначения любой аминокислоты, которая может быть заменена одной аминокислотой из основной структуры, но без значительного изменения биологической активности соответствующих антител, и таких аминокислот, какие будут определены позже, особенно в примерах. Эти эквивалентные аминокислоты могут быть определены либо на основании их структурной гомологии с аминокислотами, которые они заменяют, либо по результатам сравнительных исследований биологической активности различных антител, на которую они способны.
К примеру, упоминается возможность выполнения замены без глубокого изменения биологической активности соответствующего модифицированного антитела.
В качестве неограничивающего примера следующая таблица 1 показывает замены, возможные с сохранением биологической активности модифицированного антитела. Обратные замены, конечно, также возможны в тех же условиях.
|
Следует понимать здесь, что изобретение не относится к антителам в природной форме, то есть они находятся не в их природной среде, но что они могут быть выделены или получены путем очистки из природных источников, либо получены путем генетической рекомбинации, либо путем химического синтеза, и что они могут содержать неприродные аминокислоты, как будет описано ниже.
Согласно первому подходу, антитело будет определяться по последовательности его тяжелой цепи. В частности, антитело изобретения или один из его функциональных фрагментов или производных характеризуется тем, что оно содержит тяжелую цепь по меньшей мере с одним CDR, выбранным из CDR, содержащих аминокислотные последовательности SEQ ID №№1-9 и 56-58.
Упомянутыми последовательностями являются следующие:
|
CDR тяжелой цепи могут быть выбраны случайным образом из предыдущих последовательностей, т.е. SEQ ID №№1-9 и 56-58.
В соответствии с предпочтительным аспектом антитело изобретения или один из его функциональных фрагментов или производных содержит тяжелую цепь по меньшей мере с одним CDR, выбранным среди CDR-H1, CDR-H2 и CDR-H3, где:
- CDR-H1 содержит аминокислотную последовательность SEQ ID №1, 4, 7 или 56,
- CDR-H2 содержит аминокислотную последовательность SEQ ID №2, 5, 8 или 57, и
- CDR-H3 содержит аминокислотную последовательность SEQ ID №3, 6, 9 или 58.
В соответствии с первым воплощением указанного аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, где CDR-H1 содержит аминокислотную последовательность SEQ ID №1, CDR-H2 содержит аминокислотную последовательность SEQ ID №2, а CDR-H3 содержит аминокислотную последовательность SEQ ID №3.
В частности, указанное антитело или один из его функциональных фрагментов или производных в соответствии с этим первым воплощением содержит тяжелую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №18.
SEQ ID №18: EVQLQQSGPELVKPGASVKISCKTSGYIFTAYTMHWVRQSLG ESLDWIGGIKPNNGLANYNQKFKGKATLTVDKSSSTAYMDLRSLTSEDSAVYYCARSEITTEFDYWGQGTALTVSS
В соответствии со вторым воплощением указанного аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, где CDR-H1 содержит аминокислотную последовательность SEQ ID №4, CDR-H2 содержит аминокислотную последовательность SEQ ID №5, а CDR-H3 содержит аминокислотную последовательность SEQ ID №6.
Антитело или один из его функциональных фрагментов или производных в соответствии с указанным вторым воплощением будет предпочтительно содержать тяжелую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №19.
SEQ ID №19: EVQLQQSGPELVKPGASMKISCKASGYSFTDYTLNWVKQSH GKTLEWIGLINPYNGGTTYNQKFKGKATLTVDKSSSTAYMELLSLTSEDSAVYYCAREEITKDFDFWGQGTTLTVSS
В соответствии с третьим воплощением указанного аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, где CDR-H1 содержит аминокислотную последовательность SEQ ID №7, CDR-H2 содержит аминокислотную последовательность SEQ ID №8, а CDR-H3 содержит аминокислотную последовательность SEQ ID №9.
Антитело или один из его функциональных фрагментов или производных в соответствии с указанным третьим воплощением предпочтительно будет содержать тяжелую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №20.
SEQ ID №20: EVLLQQSGPELVKPGASVKIPCKASGYTFTDYNMDWVKQSH GMSLEWIGDINPNNGGTIFNQKFKGKATLTVDKSSSTAYMELRSLTSEDTAVYYCARGRYVGYYYAMDYWGQGTSVTVSS
В соответствии с четвертым воплощением указанного аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, где CDR-H1 содержит аминокислотную последовательность SEQ ID №56, CDR-H2 содержит аминокислотную последовательность SEQ ID №57, а CDR-H3 содержит аминокислотную последовательность SEQ ID №ID №58.
Антитело или один из его функциональных фрагментов или производных в соответствии с указанным четвертым воплощением предпочтительно будет содержать тяжелую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №62.
SEQ ID №62:
QVQLQQSGAELAKPGASVKMSCKASGYTFTSYWMNWVKQRPGQGLEWIGYINPTTGSTDYNQKLKDKATLTADKSSNTAYMQLSSLTSEDSAVYYCAIGGYGSWFAYWGQGTLVTVSA
Во втором подходе антитело теперь будет определяться по последовательности его легкой цепи. В частности, в соответствии со вторым частным аспектом изобретения антитело или один из его функциональных фрагментов или производных характеризуется тем, что содержит легкую цепь по меньшей мере с одним CDR, выбранным из CDR, содержащих аминокислотную последовательность SEQ ID №№10-17 и 59-61.
Упомянутыми последовательностями являются следующие:
SEQ ID №10: ESVDSYANSF
SEQ ID №11: RAS
SEQ ID №12: QQSKEDPLT
SEQ ID №13: ESIDTYGNSF
SEQ ID №14: QQSNEDPFT
SEQ ID №15: ENIYSN
SEQ ID №16: AAT
SEQ ID №17: QHFWGPPYT
SEQ ID №59: SSVSSTY
SEQ ID №60: TTS
SEQ ID №61: HQWSSYPFT
CDR легкой цепи могут быть выбраны случайным образом из предыдущих последовательностей, т.е. SEQ ID №№10-17 и 59-61.
В соответствии с другим предпочтительным аспектом антитело изобретения или один из его функциональных фрагментов или производных содержит легкую цепь по меньшей мере с одним CDR, выбранным среди CDR-L1, CDR-L2 и CDR-L3, где:
- CDR-L1 содержит аминокислотную последовательность SEQ ID №10, 13, 15 или 59;
- CDR-L2 содержит аминокислотную последовательность SEQ ID №11, 16 или 60; а
- CDR-L3 содержит аминокислотную последовательность SEQ ID №12, 14, 17 или 61.
В соответствии с первым воплощением указанного другого аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, где CDR-L1 содержит аминокислотную последовательность SEQ ID №10, CDR-L2 содержит аминокислотную последовательность SEQ ID №11, а CDR-L3 содержит аминокислотную последовательность SEQ ID №12.
В частности, указанное антитело или один из его функциональных фрагментов или производных в соответствии с этим первым воплощением содержит легкую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №21.
SEQ ID №21: DIVLTQSPASLAVSLGQRATISCRASESVDSYANSFMHWYQQ KPGQPPKLLIYRASNLESGIPARFSGSGSRTDFTLTINPVEADDVATYYCQQSKEDPLTFGSGTKLEMK
В соответствии со вторым воплощением указанного другого аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, где CDR-L1 содержит аминокислотную последовательность SEQ ID №13, CDR-L2 содержит аминокислотную последовательность SEQ ID №11, а CDR-L3 содержит аминокислотную последовательность SEQ ID №14.
Антитело или один из его функциональных фрагментов или производных в соответствии с указанным вторым воплощением будет предпочтительно содержать легкую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №22.
SEQ ID №22: GIVLTQSPASLAVSLGQRATISCRVSESIDTYGNSFIHWYQQKPGQPPKLLIYRASNLESGIPARFSGSGSRTDFTLTINPVEADDSATYYCQQSNEDPFTFGSGTKLEMK
В соответствии с третьим воплощением указанного другого аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, где CDR-L1 содержит аминокислотную последовательность SEQ ID №15, CDR-L2 содержит аминокислотную последовательность SEQ ID №16, а CDR-L3 содержит аминокислотную последовательность SEQ ID №17.
Антитело или один из его функциональных фрагментов или производных в соответствии с указанным третьим воплощением будет предпочтительно содержать легкую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №23.
SEQ ID №23: DIQMTQSPASLSVSVGETVTITCRASENIYSNLAWYQQKQGKSPQLLVYAATNLVDGVPSRFSGSGSGTQYSLKINSLQSEDFGSYYCQHFWGPPYTFGGGTKLEIK
В соответствии с четвертым воплощением указанного другого аспекта антитело изобретения или один из его функциональных фрагментов или производных содержит легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, где CDR-L1 содержит аминокислотную последовательность SEQ ID №59, CDR-L2 содержит аминокислотную последовательность SEQ ID №60, а CDR-L3 содержит аминокислотную последовательность SEQ ID №61.
Антитело или один из его функциональных фрагментов или производных в соответствии с указанным четвертым воплощением будет предпочтительно содержать легкую цепь с последовательностью, содержащей аминокислотную последовательность SEQ ID №63.
SEQ ID №63:
QIVLTQSPAIMSASPGEKVTLTCSASSSVSSTYLYWYQQKPGSSPKLWIYTTSILASGVPARFSGSGSGTSYSLTISSMETEDAASYFCHQWSSYPFTFGSGTKLDIK
В соответствии с третьим подходом антитело теперь будет определяться как по последовательности его легкой цепи, так и по последовательности его тяжелой цепи. Антитело изобретения или один из его функциональных фрагментов или производных характеризуется тем, что содержит тяжелую цепь с аминокислотной последовательностью SEQ ID №18, 19, 20 или 62 и легкую цепь с аминокислотной последовательностью SEQ ID №21, 22, 23 или 63.
В частности, предпочтительное антитело, названное 224G11, или один из его функциональных фрагментов или производных в соответствии с изобретением содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, содержащие соответственно аминокислотные последовательности SEQ ID №№1, 2 и 3; а также легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, содержащие соответственно аминокислотные последовательности SEQ ID №10, 11 и 12.
В другом аспекте антитело 224G11 содержит тяжелую цепь с аминокислотной последовательностью SEQ ID №18 и легкую цепь с аминокислотной последовательностью SEQ ID №21.
Другое предпочтительное антитело, названное 227Н1, или один из его функциональных фрагментов или производных в соответствии с изобретением содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, содержащие соответственно аминокислотные последовательности SEQ ID №4, 5 и 6; а также легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, содержащие соответственно аминокислотные последовательности SEQ ID №13, 11 и 14.
В другом аспекте антитело 227Н1 содержит тяжелую цепь с аминокислотной последовательностью SEQ ID №19 и легкую цепь с аминокислотной последовательностью SEQ ID №22.
Еще одно предпочтительное антитело, названное 223С4, или один из его функциональных фрагментов или производных в соответствии с изобретением содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, содержащие соответственно аминокислотные последовательности SEQ ID №7, 8 и 9; а также легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, содержащие соответственно аминокислотные последовательности SEQ ID №15, 16 и 17.
В другом аспекте антитело 223С4 содержит тяжелую цепь с аминокислотной последовательностью SEQ ID №20 и легкую цепь с аминокислотной последовательностью SEQ ID №23.
Еще одно предпочтительное антитело, названное 11Е1, или один из его функциональных фрагментов или производных в соответствии с изобретением содержит тяжелую цепь, содержащую CDR-H1, CDR-H2 и CDR-H3, содержащие соответственно аминокислотные последовательности SEQ ID №56, 57 и 58; а также легкую цепь, содержащую CDR-L1, CDR-L2 и CDR-L3, содержащие соответственно аминокислотные последовательности SEQ ID №59, 60 и 61.
В другом аспекте антитело 11Е1 содержит тяжелую цепь с аминокислотной последовательностью SEQ ID №62 и легкую цепь с аминокислотной последовательностью SEQ ID №63.
В соответствии с другим аспектом изобретение относится к мышиной гибридоме, способной секретировать моноклональные антитела в соответствии с данным изобретением, особенно к гибридоме мышиного происхождения, такой как депонированная в Collection Nationale de Cultures de Microorganismes (CNCM, Национальная коллекция культур микроорганизмов) (Институт Пастера, Париж, Франция).
Моноклональные антитела в соответствии с изобретением или один из их функциональных фрагментов или производных, охарактеризованные как такие указанные антитела, секретируемые гибридомой, хранятся в CNCM с 03/14/2007 под номерами CNCM I-3724 (соответствует 11Е1), I-3731 (соответствует 224G11), I-3732 (соответствует 227Н1) и с 07/06/2007 под номером I-3786 (соответствует 223С4). Эти гибридомы являются мышиными гибридомами, полученными в результате клеточного слияния иммунизированных мышиных спленоцитов с миеломной клеточной линией (Sp20 Ад 14).
В следующей таблице 2 объединены элементы, связанные с предпочтительными антителами.
|
Из таблицы 2 совершенно очевидно, что CDR-L2 из антител 227Н1 и 224G11 аналогичны. Этот пример четко поддерживает пункты данной заявки, охватывающие антитела, содержащие по меньшей мере один CDR, случайно выбранный среди описанных последовательностей CDR.
В соответствии с предпочтительным воплощением изобретение связано к моноклональными антителами.
Термин «моноклональное антитело» используется в соответствии с его обычным значением для обозначения антитела, полученного из популяции существенно однородных антител, т.е. отдельные антитела в составе популяции являются идентичными, за исключением возможных природных мутаций, которые могут присутствовать в небольших количествах. Иными словами, моноклональное антитело представляет собой однородное антитело, полученное в результате пролиферации одного клона клеток (например гибридомных клеток, эукариотических клеток хозяина, трансфицированных ДНК, кодирующей однородное антитело, прокариотических клеток хозяина, трансформированных ДНК, кодирующей однородное антитело и т.д.), которое в целом характеризуется тяжелыми цепями одного класса и подкласса и легкими цепями одного типа. Моноклональные антитела высоко специфичны, будучи направленными против одного антигена. Кроме того, в отличие от получения поликлональных антител, которые обычно включают различные антитела, направленные против различных детерминант, или эпитопов, каждое моноклональное антитело направлено против одной детерминанты на антигене.
В данном описании термины «полипептиды», «полипептидные последовательности», «аминокислотные последовательности», «пептиды» и «белки», применяемые по отношению к соединениям антителам или их последовательностям, являются взаимозаменяемыми.
В соответствии с аналогичным частным аспектом данное изобретение относится к химерному антителу или одному из его функциональных фрагментов в соответствии с изобретением, характеризующемуся тем, что указанное антитело кроме того содержит константные участки легких цепей и тяжелых цепей, полученные из антитела вида, гетерологичного мыши, особенно человека, и в предпочтительном виде характеризующемуся тем, что константные участки легких цепей и тяжелых цепей, полученные из человеческого антитела, являются соответственно участками каппа и гамма-1, гамма-2 или гамма-4.
В данной заявке lgG1 предпочтительно несут эффекторные функции, и наиболее предпочтительно ADCC и CDC.
Специалист поймет, что эффекторные функции включают, например, C1q-связывание; комплемент-зависимую цитотоксичность (complement dependent cytotoxicity, CDC); Fc-рецепторное связывание; антитело-зависимую клеточную цитотоксичность (antibody-dependent cell-mediated cytotoxicity, ADCC); фагоцитоз и отрицательную регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора; BCR).
Антитела в соответствии с данным изобретением предпочтительно представляют собой специфические моноклональные антитела, особенно мышиного, химерного или гуманизированного происхождения, которые могут быть получены в соответствии со стандартным способами, хорошо известными специалистам в данной области.
В общем, для получения моноклональных антител или их функциональных фрагментов или производных, особенно мышиного происхождения, можно сослаться на методики, описанные, в частности, в руководстве «Antibodies» (Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor NY, pp.726, 1988), или на методику получения из гибридом, описанную Kohler и Milstein (Nature, 256:495-497, 1975).
Моноклональные антитела в соответствии с изобретением могут быть получены, например, из животной клетки, иммунизированной против c-Met или одного из его фрагментов, содержащих эпитоп, специфично распознаваемый указанными моноклональными антителами в соответствии с изобретением. Указанный c-Met или один из его указанных фрагментов может в особенности быть получен в соответствии с обычными рабочими способами путем генетической рекомбинации, начиная с нуклеиновокислотной последовательности, содержащейся в последовательности кДНК, кодирующей c-Met, или путем пептидного синтеза, начиная с последовательности аминокислот, содержащейся в пептидной последовательности c-Met.
Моноклональные антитела в соответствии с изобретением могут, например, быть очищены на аффинной колонке, на которой c-Met или один из его фрагментов, содержащий эпитоп, специфично распознается предварительно закрепленными указанными моноклональными антителами в соответствии с изобретением. В частности, указанные моноклональные антитела могут быть очищены путем хроматографии на белке А и/или G, с последующей ионообменной хроматографией для удаления остаточных белковых загрязнителей, а также ДНК и ЛПС, или без нее, с последующей хроматографией на сефарозном геле Sepharose™ для удаления потенциальных агрегатов в связи с наличием димеров или других многомеров или без нее. Еще более предпочтительным образом все эти методики могут быть использованы одновременно или последовательно.
Химерные или гуманизированные антитела тоже включены в антитела в соответствии с данным изобретением.
Термин «химерное антитело» используется для обозначения антитела, которое содержит природный вариабельный участок (легкой цепи и тяжелой цепи), полученный из антитела данного вида, в сочетании с константными участками легкой цепи и тяжелой цепи антитела вида, гетерологичного указанному данному виду (например мыши, лошади, кролика, собаки, коровы, курицы и т.д.).
Антитела или их фрагменты химерного типа в соответствии с изобретением могут быть получены с помощью методик генетической рекомбинации. Например, химерное антитело может быть получено путем клонирования рекомбинантной ДНК, содержащей промотор и последовательность, кодирующую нечеловеческий вариабельный участок, особенно мышиный, моноклональное антитело в соответствии с изобретением и последовательность, кодирующую константный участок человеческого антитела. Химерное антитело изобретения, закодированное таким рекомбинантным геном, будет, например, химерой мыши и человека, специфичность этого антитела будет определяться вариабельным участком, полученным из мышиной ДНК, а его изотип будет определяться константным участком, полученным из человеческой ДНК. Для способов получения химерных антител можно, например, сослаться на документы Verhoeyn et al. (BioEssays, 8:74, 1988), Morrison et al. (Proc. Natl. Acad. Sci. USA 82:6851-6855, 1984) или на патент США 4816567.
Термин «гуманизированное антитело» используется для обозначения антитела, которое содержит CDR-участки, полученные из антитела нечеловеческого происхождения, другие части молекулы антитела получены из одного (или нескольких) человеческих антител. Кроме того, некоторые остатки сегментов скелета (называемые FR, каркасные участки) могут быть изменены в целях сохранения аффинности связывания (Jones et al., Nature, 321:522-525, 1986; Verhoeyen et al., Science, 239:1534-1536, 1988; Riechmann et al., Nature, 332:323-327, 1988).
Гуманизированные антитела в соответствии с изобретением или их фрагменты могут быть получены с помощью методик, известных специалистам в данной области (например таких, которые описаны в документах Singer et al., J. Immun. 150:2844-2857, 1992; Mountain et al., Biotechnol. Genet. Eng. Rev., 10:1-142, 1992; или Bebbington et al., Bio/Technology, 10:169-175, 1992).
Другим способом гуманизации, известным специалистам в данной области, является, например, способ "CDR-прививки", описанный Protein Design Lab (PDL) в патентных заявках ЕР 0451261, ЕР 0682040, ЕР 09127, ЕР 0566647 или US 5530101, US 6180370, US 5585089 и US 5693761. Также можно отметить следующие патентные заявки: US 5639641; US 6054297; US 5886152 и US 5877293.
Термин "функциональный фрагмент" антитела в соответствии с изобретением используется для обозначения, в частности, фрагментов антитела, таких как фрагменты Fv, scFv (sc для одной цепи), Fab, F(ab')2, Fab', scFv-Fc, или димерных антител или любого фрагмента, время полужизни которого было бы увеличено в результате химической модификации, такой как добавление поли(алкилен)гликоля, такого как поли(этилен)гликоль ("ПЭГилирование") (пэгилированные фрагменты называются Fv-ПЭГ, ScFv-ПЭГ, Fab-ПЭГ, F(ab')2-ПЭГ или Fab'-ПЭГ) ("ПЭГ" для поли(этилен)гликоля), либо путем включения в липосому; указанный фрагмент имеет по меньшей мере один характерный CDR из последовательности SEQ ID №№1-17 и 56-61 в соответствии с изобретением, и особенно способен проявлять в общем виде даже частичную активность антитела, из которого он произошел, такую как, в частности, способность распознавать и связывать c-Met, и, при необходимости, ингибировать активность c-Met.
Предпочтительно указанные функциональные фрагменты будут состоять из или включать частичную последовательность тяжелой или легкой вариабельной цепи антитела, из которого они получены; указанная частичная последовательность должна быть достаточной, чтобы сохранить такую же специфичность связывания, как у антитела, из которого она получена, и достаточную аффинность к c-Met, предпочтительно равную по меньшей мере 1/100, в более предпочтительном случае по меньшей мере 1/10 специфичности антитела, из которого она получена. Такой функциональный фрагмент будет содержать как минимум 5 аминокислот, предпочтительно 6, 7, 8, 9, 10, 12, 15, 25, 50 и 100 последовательных аминокислот последовательности антитела, из которого он получен.
Предпочтительно этими функциональными фрагментами будут фрагменты типа Fv, scFv, Fab, F(ab')2, F(ab'), scFv-Fc или димерные антитела, которые в целом имеют такую же специфичность связывания, как и антитела, из которых они получены. В более предпочтительном воплощении изобретения эти фрагменты выбраны среди двухвалентных фрагментов, таких как фрагменты F(ab')2. В соответствии с данным изобретением фрагменты антитела изобретения могут быть получены из антител описанными выше способами, такими как расщепление ферментами, такими как пепсин или папаин, и/или расщепление дисульфидных мостиков путем химического восстановления. Иным образом, фрагменты антитела, входящие в данное изобретение, могут быть получены с помощью методик генетической рекомбинации, известных специалистам в данной области, или также путем пептидного синтеза, например с помощью автоматических пептидных синтезаторов, например поставляемых компанией Applied Biosystems, и т.д.
Под "двухвалентными фрагментами" следует понимать любые фрагменты антител, содержащие две ветви, в частности, F(ab')2-фрагменты.
В частности, изобретение включает антитела или их функциональные фрагменты в соответствии с данным изобретением, особенно химерные или гуманизированные антитела, полученные путем генетической рекомбинации или путем химического синтеза.
Под «производным» антитела в соответствии с изобретением понимается связывающий белок, содержащий белковую матрицу и по меньшей мере один из CDR, выбранных из исходного антитела, для поддержания связывающей способности. Такие соединения хорошо известны специалистам в данной области и будут описаны более подробно в последующем описании.
В частности, антитело или один из его функциональных фрагментов или производных в соответствии с изобретением характеризуется тем, что указанное производное представляет собой связывающий белок, содержащий матрицу, к которой привит по меньшей мере один CDR для сохранения исходных свойств антитела по распознаванию паратопа.
На белковой матрице может присутствовать одна или несколько последовательностей из числа шести CDR-последовательностей, описанных в изобретении. В этом случае белковая матрица воспроизводит белковую основу с соответствующим сворачиванием привитого(ых) CDR, тем самым позволяя ему (или им) сохранять их свойства по распознаванию паратопа антигена.
Специалистам в данной области известно, как выбирать белковую матрицу, на которую можно привить по меньшей мере один CDR, выбранный из исходного антитела. В частности известно, что такая матрица, чтобы быть выбранной, должна обладать следующими несколькими свойствами (Skerra A., J. Mol. Recogn., 13, 2000, 167-187):
- филогенетически хорошей сохранностью,
- надежной архитектурой с известной трехмерной молекулярной организацией (например, с помощью кристаллографии или ЯМР),
- малым размером,
- отсутствием или наличием только в малой степени посттрансляционных изменений,
- легкостью производства, экспрессии и очистки.
Такая белковая матрица может быть структурой, выбранной из группы, включающей фибронектин и предпочтительно десятый домен фибронектина III типа (FNfn10), липокалин, антикалин (Skerra A., J. Biotechnol., 2001, 74(4):257-75), белок Z, полученный из домена В стафилококкового белка А, тиоредоксин А или любой белок с повторяющимся доменом, таким как "анкириновый повтор" ((Kohl et al., PNAS, 2003, vol. 100, No. 4, 1700-1705), "повтор Армадилла", "повтор, богатый лейцином" или "тетратрикопептидный повтор", но не ограничивается ими.
Также можно упомянуть матрицу, полученную из токсинов (таких как, например, токсины скорпионов, насекомых, растений и моллюсков) или белковых ингибиторов нейронной синтазы оксида азота (PIN).
В качестве неограничивающего примера таких гибридных конструкций можно отметить вставление CDR-H1 (тяжелая цепь) анти-CD4-антитела, т.е. антитела 13В8.2, в одну из открытых петель PIN. Связывающие свойства полученного связывающего белка остаются похожими на свойства исходного антитела (Bes et al., BBRC 343, 2006, 334-344). Можно также упомянуть прививку CDR-H3 (тяжелая цепь) анти-лизоцим-антитела VHH в петлю неокарциностатина (Nicaise et al., 2004).
В случае данного изобретения целевой CDR для сохранения может представлять собой CDR-L2, сохраненный в двух определенных антителах изобретения, т.е. 227Н1 и 224G11, но не ограничивается этим.
Как упоминалось выше, такая белковая матрица может содержать от 1 до 6 CDR из исходного антитела. В предпочтительном воплощении, но без каких-либо ограничений, специалист в данной области может выбрать по меньшей мере CDR из тяжелой цепи, о которой известно, что она частично участвует в специфичности антитела. Выбор целевого(ых) CDR известными способами будет очевидным для специалиста в данной области (BES et al., FEBS letters 508, 2001, 67-74).
Для наглядности, эти примеры не являются ограничивающими, и любая другая известная или описанная матрица должна быть включена в данное описание.
В соответствии с новым аспектом данное изобретение относится к выделенной нуклеиновой кислоте, отличающейся тем, что она выбрана из следующих нуклеиновых кислот:
a) нуклеиновая кислота, ДНК или РНК, кодирующая антитело или один из его функциональных фрагментов или производных в соответствии с изобретением;
b) нуклеиновая кислота, содержащая последовательность ДНК, выбранную из группы последовательностей, включающей:
- нуклеиновую последовательность, содержащую последовательности SEQ ID №24, SEQ ID №25, SEQ ID №26 и последовательности SEQ ID №33, SEQ ID №34 и SEQ ID №35;
- нуклеиновую последовательность, содержащую последовательности SEQ ID №27, SEQ ID №28, SEQ ID №29 и последовательности SEQ ID №36, SEQ ID №34 и SEQ ID №37;
- нуклеиновую последовательность, содержащую последовательности SEQ ID №30, SEQ ID №31, SEQ ID №32 и последовательности SEQ ID №38, SEQ ID №39 и SEQ ID №40; и
- нуклеиновую последовательность, содержащую последовательности SEQ ID №64, SEQ ID №65, SEQ ID №66 и последовательности SEQ ID №67, SEQ ID №68 и SEQ ID №69;
c) нуклеиновая кислота, содержащая последовательность ДНК, выбранную из группы последовательностей, включающей:
- нуклеиновую последовательность, содержащую последовательности SEQ ID №41 и SEQ ID №44;
- нуклеиновую последовательность, содержащую последовательности SEQ ID №42 и SEQ ID №45;
- нуклеиновую последовательность, содержащую последовательности SEQ ID №43 и SEQ ID №46; и
- нуклеиновую последовательность, содержащую последовательности SEQ ID №70 и SEQ ID №71;
d) РНК, соответствующая нуклеиновым кислотам, определенным в b) или с);
e) нуклеиновые кислоты, комплементарные нуклеиновым кислотам, определенным в а), b) и с); и
f) нуклеиновая кислота по меньшей мере из 18 нуклеотидов, способная к гибридизации в условиях высокой жесткости по меньшей мере с одним из CDR с последовательностью SEQ ID №№24-40 и 64-69.
Термины «нуклеиновая кислота», «нуклеиновая последовательность» или «нуклеиновокислотная последовательность», «полинуклеотид», «олигонуклеотид», «полинуклеотидная последовательность», «нуклеотидная последовательность», которые будут использоваться равнозначно в данном изобретении, применяются для обозначения четкой связи нуклеотидов, модифицированных или не модифицированных, позволяющей определить фрагмент или участок нуклеиновой кислоты, содержащий или не содержащий неприродные нуклеотиды и способный соответствовать одинаково хорошо двуцепочечной ДНК, одноцепочечной ДНК, как продукты транскрипции указанной ДНК.
Здесь также следует понимать, что данное изобретение не касается нуклеотидных последовательностей в их природной хромосомной среде, т.е. в природном состоянии. Это касается последовательностей, которые были выделены и/или очищены, т.е. были выбраны прямо или косвенно, например путем копирования, их среда была по меньшей мере частично изменена. Таким образом, также здесь следует подразумевать выделенные нуклеиновые кислоты, полученные путем генетической рекомбинации, например с помощью принимающих клеток (клеток-хозяев), или полученные путем химического синтеза.
Гибридизации в условиях высокой жесткости означает, что температурные условия и условия ионной силы выбирают таким образом, чтобы они позволяли сохранять гибридизацию между двумя фрагментами комплементарной ДНК. В качестве примера, условия высокой жесткости этапа гибридизации для определения полинуклеотидных фрагментов, описанные выше, преимущественно являются следующими.
ДНК-ДНК- или ДНК-РНК-гибридизацию осуществляют в два этапа: (1) прегибридизация при 42°С в течение 3 часов в фосфатном буфере (20 мМ, рН 7,5), содержащем 5×SSC (цитратно-солевой буфер) (1×SSC соответствует 0,15 М NaCl+0,015 М раствора цитрата натрия), 50% формамида, 7% додецилсульфата натрия (SDS), 10 × раствор Денхардта, 5% сульфата декстрана и 1% ДНК спермы лосося; (2) собственно гибридизация в течение 20 часов при температуре в зависимости от размера зонда (например: 42°С для зонда размером более 100 нуклеотидов), а затем 2 отмывки в течение 20 минут при температуре 20°С в 2×SSC+2% SDS, 1 отмывка в течение 20 минут при 20°С в 0,1×SSC+0,1% SDS. Последнюю отмывку осуществляют в 0,1х SSC+0,1% SDS в течение 30 минут при температуре 60°С для зонда размером более 100 нуклеотидов. Гибридизационные условия высокой жесткости, описанные выше для полинуклеотида определенного размера, специалист в данной области может адаптировать для олигонуклеотидов большего или меньшего размера в соответствии с руководством Sambrook et al. (1989, Molecular cloning: a laboratory manual. 2nd Ed. Cold Spring Harbor).
Подобным образом изобретение относится к вектору, содержащему нуклеиновую кислоту в соответствии с данным изобретением.
Изобретение особенно нацелено на клонирующие векторы и/или векторы экспрессии, которые содержат нуклеотидную последовательность в соответствии с изобретением.
Векторы в соответствии с изобретением предпочтительно содержат элементы, которые позволяют экспрессировать и/или секретировать нуклеотидные последовательности в определенной принимающей клетке (клетке-хозяине). Поэтому вектор должен содержать промотор, сигналы инициации и терминации трансляции, а также соответствующие участки регуляции транскрипции. Он должен быть способен поддерживать в стабильном состоянии принимающую клетку (клетку-хозяина) и дополнительно может иметь особенные сигналы, которые обуславливают секрецию транслированного белка. Специалисты в данной области выбирают и оптимизируют эти различные элементы в соответствии с функцией используемой принимающей клетки (клетки-хозяина). С этой целью нуклеотидные последовательности в соответствии с изобретением могут быть вставлены в векторах автономной репликации в выбранного хозяина, или могут быть интегративными векторами выбранного хозяина.
Такие векторы изготавливают с помощью способов, использующихся в настоящее время специалистами в данной области, и полученные клоны могут быть введены в соответствующего хозяина с помощью стандартных способов, таких как липофекция, электропорация, тепловой шок или химические способы.
Векторы в соответствии с изобретением представляют собой, например, векторы плазмидного или вирусного происхождения. Они используются для трансформирующихся принимающих клеток (клеток-хозяев), чтобы клонировать или экспрессировать нуклеотидные последовательности в соответствии с изобретением.
Аналогичным образом изобретение включает принимающие клетки (клетки-хозяева), трансформированные с помощью вектора в соответствии с изобретением или содержащие его.
Принимающая клетка (клетка-хозяин) может быть выбрана из прокариотических или эукариотических систем, например бактериальных клеток, а также дрожжевых клеток или клеток животных, в частности клеток млекопитающих. Можно также использовать клетки насекомых или клетки растений.
Аналогичным образом изобретение касается животных, за исключением человека, которые содержат по меньшей мере одну клетку, трансформированную в соответствии с изобретением.
В соответствии с другим аспектом предметом изобретения является способ производства антитела или одного из его функциональных фрагментов в соответствии с изобретением, характеризующийся тем, что он включает в себя следующие этапы:
a) культивирование принимающей клетки (клетки-хозяина) в соответствии с изобретением в среде в соответствующих культуральных условиях; и
b) выделение указанных антител или одного из их функциональных фрагментов, полученных таким образом, из культуральной среды или указанных культивируемых клеток.
Клетки, трансформированные в соответствии с изобретением, могут быть использованы в способах получения рекомбинантных полипептидов в соответствии с изобретением. Способы получения полипептида в соответствии с изобретением в рекомбинантной форме, характеризующиеся использованием вектора и/или клетки, трансформированной вектором в соответствии с изобретением, входят в данное изобретение. Предпочтительно клетку, трансформированную вектором в соответствии с изобретением, культивируют в условиях, которые позволяют экспрессировать указанный полипептид, и указанный рекомбинантный пептид восстанавливают.
Как уже было сказано, принимающая клетка (клетка-хозяин) может быть выбрана из прокариотических или эукариотических систем. В частности, можно определить нуклеотидные последовательности в соответствии с изобретением, содействующие секреции в таких прокариотических или эукариотических системах. Вектор в соответствии с изобретением, несущий такую последовательность, таким образом, может быть выгодно использован для производства рекомбинантных белков, предназначенных для секреции. По сути, очистке этих целевых рекомбинантных белков будет способствовать тот факт, что они находятся в супернатанте клеточной культуре, а не внутри принимающих клеток (клеток-хозяев).
Таким же образом можно получить полипептиды в соответствии с изобретением путем химического синтеза. Такой способ получения также является предметом изобретения. Специалисту в данной области известны способы химического синтеза, например методики с использованием твердых фаз [Steward et al., 1984, Solid phase peptide synthesis, Pierce Chem. Company, Rockford, 111, 2nd ed., (1984)] или методики с использованием частично твердых фаз, путем конденсации фрагментов или путем классического синтеза в растворе. Полипептиды, полученные путем химического синтеза и способные содержать соответствующие неприродные аминокислоты, также входят в изобретение.
Антитела или один из их функциональных фрагментов или производных, которые могут быть получены с помощью способа в соответствии с изобретением, также входят в данное изобретение.
Изобретение также касается антитела изобретения в качестве лекарственного средства.
Аналогичным образом изобретение касается фармацевтической композиции, включающей в качестве активной основы соединение, содержащее антитело или один из его функциональных фрагментов в соответствии с изобретением, предпочтительно в смеси с эксципиентом и/или фармацевтически приемлемым носителем.
Другое дополнительное воплощение изобретения включает композицию, такую как описано выше, в качестве комбинированного продукта для одновременного, раздельного или последовательного применения, содержащую кроме того противоопухолевое антитело.
Наиболее предпочтительно указанное второе противоопухолевое антитело может быть выбрано среди антител анти-IGF-IR, анти-EGFR, анти-HEK2/neu, анти-VEGFR, анти-VEGF и т.д., или любых других противоопухолевых антител, известных специалистам в данной области. Очевидно, что применение в качестве второго антитела функциональных фрагментов или производных вышеупомянутых антител является частью изобретения.
В качестве наиболее предпочтительного антитела выбирают антитела анти-EGFR, такие как, например, антитело С225 (Эрбитукс).
Под "одновременным применением" понимается введение двух соединений композиции в соответствии с изобретением в единой и идентичной фармацевтической форме.
Под "раздельным применением" понимается введение в одно и то же время двух соединений композиции в соответствии с изобретением в различных фармацевтических формах.
Под "последовательным применением" понимается последовательное введение двух соединений композиции в соответствии с изобретением, каждое в отдельной фармацевтической форме.
В общих словах, композиция в соответствии с изобретением значительно повышает эффективность лечения рака. Иными словами, терапевтический эффект антител анти-c-Met в соответствии с изобретением неожиданным образом усиливается при введении цитотоксического агента. Другое важное последующее преимущество, полученное с помощью композиции в соответствии с изобретением, касается возможности использования более низких эффективных доз активной основы, что позволяет избежать или уменьшить риск проявления вторичных эффектов, в частности влияний цитотоксического агента.
Кроме того, эта композиция в соответствии с изобретением позволяет быстрее достигать ожидаемого терапевтического эффекта.
Композиция изобретения также может быть охарактеризована тем, что она в качестве комбинированного продукта для одновременного, раздельного или последовательного применения помимо прочего содержит цитотоксический/цитостатический агент.
Под терминами "противораковые терапевтические агенты" или "цитотоксические/цитостатические агенты" понимаются вещества, которые при введении субъекту лечат или предотвращают развитие рака в теле субъекта. В качестве неограничивающего примера таких агентов можно упомянуть алкилирующие агенты, антиметаболиты, противоопухолевые антибиотики, митотические ингибиторы, ингибиторы функционирования хроматина, антиангиогенные агенты, антиэстрогены, антиандрогены или иммуномодуляторы.
Такими агентами являются, например, агенты, упомянутые в издании Видаль 2001 года на странице, посвященной соединениям, применяемым в онкологии и гематологии, в колонке "Цитотоксические вещества"; эти цитотоксические соединения, цитируемые применительно к данному документу, приводятся здесь в качестве предпочтительных цитотоксических агентов.
В частности, следующие агенты являются предпочтительными в соответствии с изобретением.
"Алкилирующий агент" обозначает любое вещество, которое может связывать или алкилировать любую молекулу, предпочтительно нуклеиновую кислоту (например ДНК) в клетке. Примеры алкилирующих агентов включают азотный аналог горчичного газа, такой как мехлоретамин, хлорамбукол, мелфален, хлоридрат, пипобромен, преднимустин, гидрофосфат натрия или эстрамустин; оксазофорины, такие как циклофосфамид, алтретамин, трофосфамид, сульфофосфамид или ифосфамид; азиридины или иминэтилены, такие как тиотепа, триэтиленамин или алтретамин; нитрозомочевины, такие как кармустин, стрептозоцин, фотемустин или ломустин; алкилсульфонаты, такие как бусульфан, треосульфан или импросульфан; триазены, такие как дакарбазин; или платиновые комплексы, такие как цисплатина, оксалиплатин и карбоплатин.
Термин "антиметаболиты" относится к веществам, которые блокируют клеточный рост и/или метаболизм, вмешиваясь в определенные виды активности, как правило, в синтез ДНК. К примерам антиметаболитов относятся метотрексат, 5-фторурацил, флоксуридин, 5-фтородезоксиуридин, капецитабин, цитарабин, флударабин, цитозинарабинозид, 6-меркаптопурин (6-МП), 6-тиогуанин (6-ТГ), хлородезоксиаденозин, 5-азацитидин, гемцитабин, кладрибин, дезоксикоформицин и пентостатин.
Термин "противоопухолевые антибиотики" относится к соединениям, которые могут предотвращать или замедлять синтез ДНК, РНК и/или белка. Примеры противоопухолевых антибиотиков включают доксорубицин, даунорубицин, идарубицин, валрубицин, митоксантрон, дактиномицин, митрамицин, пликамицин, митомицин С, блеомицин и прокарбазин.
"Митотические ингибиторы" предотвращают нормальное развитие клеточного цикла и митоза. В общем, ингибиторы микротрубочек или таксоиды, такие как паклитаксел и доцетаксел, способны ингибировать митоз. Алкалоид Винка, такой как винбластин, винкристин, виндезин и винорелбин, также способен ингибировать митоз.
К "ингибиторам функционирования хроматина" или "ингибиторам топоизомеразы" относятся вещества, которые ингибируют нормальную функцию белков, моделирующих хроматин, таких как топоизомераза I или топоизомераза II. Примеры ингибиторов функционирования хроматина включают, для топоизомеразы I, камптотецин и его производные, такие как топотекан или иринотекан, и, для топоизомеразы II, этопозид, фосфат этопозида и тенипозид.
Термин "антиангиогенный агент" относится к любому лекарству, соединению, веществу или агенту, который ингибирует рост кровеносных сосудов. Типичные антиангиогенные агенты включают, но отнюдь этим не ограничиваясь, разоксин, маримастат, батимастат, приномастат, таномастат, иломастат, CGS-27023А, галофугинон, COL-3, неовастат, BMS-275291, талидомид, CDC 501, DMXAA, L-651582, скваламин, эндостатин, SU5416, SU6868, интерферон-альфа, EMD121974, интерлейкин-12, IM862, ангиостатин и витаксин.
Термины "антиэстроген" или "антиэстрогенный агент" относятся к любому веществу, которое уменьшает, противодействует или ингибирует действие эстрогена. Примерами антиэстрогенных агентов являются тамоксифен, торемифен, ралоксифен, дролоксифен, иодоксифен, анастрозол, летрозол и экземестан.
Термины "антиандроген" или "антиандрогенный агент" относятся к любому веществу, которое уменьшает, противодействует или ингибирует действие андрогена. Примерами антиандрогенов являются флутамид, нилутамид, бикалютамид, спироналоктон, ацетат ципротерона, финастерид и цимитидин.
"Иммуномодуляторы" представляют собой вещества, которые стимулируют иммунную систему.
Примеры иммуномодуляторов включают интерферон, интерлейкин, такой как алдеслейкин, ОСТ-43, денилейкин дифтитокс и интерлейкин-2, факторы некроза опухоли, такие как тазонермин, или другие иммуномодуляторы, такие как лентинан, сизофиран, рохинимекс, пидотимод, пегадемаза, тимопентин, поли(I:С) (сополимер полиинозиновой и полицитидиловой кислот) или левамизол в сочетании с 5-фторурацилом.
За более подробной информацией специалист в данной области может обратиться к руководству, выпущенному под редакцией "Association Francaise des Enseignants de Chimie Therapeutique" и озаглавленному "traite de chimie therapeutique, vol. 6, Medicaments antitumoraux et perspectives dans le traitement des cancers, edition TEC & DOC, 2003".
В качестве химических агентов или цитотоксических агентов можно также упомянуть все ингибиторы киназ, такие как, например, гефитиниб или эрлотиниб.
В частном предпочтительном воплощении указанная композиция в качестве комбинированного продукта в соответствии с изобретением характеризуется тем, что указанный цитотоксический агент химически соединен с указанным антителом для одновременного использования.
Для облегчения взаимодействия указанного цитотоксического агента и указанного антитела в соответствии с изобретением особенно возможно введение между двумя соединениями, которые должны взаимодействовать, молекул-разделителей, например поли(алкилен)гликолей, таких как полиэтиленгликоль, или аминокислот, или, в другом воплощении, использование активных производных указанных цитотоксических агентов, в которые были бы введены функциональные структуры, способные взаимодействовать с указанным антителом в соответствии с изобретением. Эти методики взаимодействия хорошо известны специалистам в данной области и не будут подробно рассматриваться в данном описании.
В другом аспекте изобретение относится к композиции, которая характеризуется тем, что по меньшей мере одно из указанных антител или один из их функциональных фрагментов или производных конъюгирован с клеточным токсином и/или радиоактивным элементом.
Предпочтительно указанный токсин или указанный радиоактивный элемент способен ингибировать по меньшей мере один вид клеточной активности клеток, экспрессирующих c-Met, в более предпочтительном случае способен предотвращать рост и пролиферацию указанной клетки, особенно полностью инактивируя указанную клетку.
Также предпочтительно указанный токсин представляет собой энтеробактериальный токсин, особенно экзотоксин А от Pseudomonas.
Радиоактивные элементы (или радиоизотопы), предпочтительно конъюгированные с антителами, используемыми для терапии, представляют собой радиоизотопы, которые излучают гамма-лучи, и предпочтительно йод131, иттрий90, золото199, палладий100, медь67, висмут217 и сурьму211. Радиоизотопы, которые излучают бета- и альфа-лучи, также могут быть использованы в терапии.
Под токсином или радиоактивным элементом, конъюгированным по меньшей мере с одним антителом или одним из его функциональных фрагментов в соответствии с изобретением, понимают любое средство, позволяющее указанному токсину или указанному радиоактивному элементу связываться с указанным по меньшей мере одним антителом, особенно путем ковалентного связывания двух соединений, с или без введения связывающей молекулы.
Среди агентов, позволяющих связывать химическим (ковалентным), электростатическим или нековалентным образом все или часть компонентов конъюгата, особенно можно упомянуть те, которые сделаны из бензохинона, карбодиимида и более предпочтительно EDC (1-этил-3-[3-диметил-аминопропил]-карбодиимида гидрохлорида), дималеимида, дитиобис-нитробензойной кислоты (DTNB), N-сукцинимидил-3-ацетилтио-ацетата (SATA), связывающие агенты, имеющие одну или более фенилазидную группу, реагирующую с ультрафиолетом (УФ), и предпочтительно N-[-4-(азидосалициламино)бутил]-3'-(2'-пиридилдитио)-пропионамид (APDP), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), 6-гидразино-никотинамид (HYNIC).
Другая форма связывания, особенно для радиоактивных элементов, может состоять в использовании бифункциональных хелатных ионов.
Среди этих хелатов можно отметить хелаты, полученные из EDTA (этилендиаминтетрауксусной кислоты) или из DTPA (диэтилентриаминпентауксусной кислоты), которые были разработаны для связывания металлов, особенно радиоактивных металлов, и иммуноглобулинов. Таким образом, DTPA и ее производные могут быть заменены различными группами в углеродной цепи в целях повышения стабильности и жесткости комплекса лиганд-металл (Krejcarek et al. (1977); Brechbiel et al. (1991); Gansow (1991); патент США 4831175).
Например, диэтилентриаминпентауксусная кислота (DTPA) и ее производные, которые широко используются в медицине и биологии в течение длительного времени либо в своей свободной форме, либо в виде комплекса с ионом металла, имеет замечательную особенность формировать стабильные хелаты с ионами металла и быть связанной с белками терапевтического или диагностического интереса, такими как антитела, для получения радиоиммуноконъюгатов в терапии рака (Meases et al., (1984); Gansow et al. (1990)).
Также предпочтительно указанное по меньшей мере одно антитело, формирующее указанный конъюгат в соответствии с изобретением, выбирают среди его функциональных фрагментов, особенно фрагментов с удаленным Fc-компонентом, таких как scFv-фрагменты.
Как уже упоминалось, в предпочтительном воплощении изобретения указанный цитотоксический/цитостатический агент или указанный токсин и/или радиоактивный элемент связан химически по меньшей мере с одним из элементов указанной композиции для одновременного применения.
Данное изобретение содержит описанную композицию в качестве лекарственного средства.
Данное изобретение кроме того содержит применение композиции в соответствии с изобретением для получения лекарственного средства.
В другом аспекте изобретение имеет отношение к применению антитела или одного из его функциональных фрагментов или производных и/или композиции, как описано выше, для изготовления лекарственного средства, предназначенного для ингибирования роста и/или пролиферации опухолевых клеток.
Другой аспект изобретения состоит в применении антитела или одного из его функциональных фрагментов или производных и/или композиции, как описано выше, или применении в соответствии с вышеупомянутым, для изготовления лекарственного средства, предназначенного для профилактики или лечения рака.
В данное изобретение также включен способ, который предназначен для замедления роста и/или пролиферации опухолевых клеток у пациента, включающий введение нуждающемуся в этом пациенту антитела или одного из его функциональных фрагментов или производных в соответствии с изобретением, полученного с помощью гибридомы в соответствии с изобретением, или композиции в соответствии с изобретением.
Данное изобретение также содержит способ профилактики или лечения рака у нуждающегося в этом пациента, включающий введение пациенту антитела или одного из его функциональных фрагментов или производных в соответствии с изобретением, полученного с помощью гибридомы в соответствии с изобретением, или композиции в соответствии с изобретением.
В частном предпочтительном аспекте указанный рак представляет собой рак, выбранный среди рака предстательной железы, остеосарком, рака легкого, рака молочной железы, рака эндометрия, глиобластомы или рака толстой кишки.
Как объяснялось выше, преимуществом изобретения является предоставление лечения раковых заболеваний, связанных с HGF-зависимой и HGF-независимой активацией c-Met.
Еще в одном аспекте изобретение включает способ диагностики in vitro заболеваний, индуцированных сверхэкспрессией или недостаточной экспрессией c-Met-рецепторов, начиная с биологического образца, в котором подозревается ненормальное присутствие c-Met-рецептора; указанный способ характеризуется тем, что включает этап, на котором указанный биологический образец контактирует с антителом изобретения, благодаря чему при необходимости указанное антитело можно пометить.
Предпочтительно указанные болезни, связанные с ненормальным присутствием c-Met-рецептора в указанном способе диагностики, представляют собой раковые заболевания.
Указанное антитело или один из его функциональных фрагментов может присутствовать в форме иммуноконъюгата или меченого антитела с тем, чтобы получить выявляемый и/или поддающийся количественной оценке сигнал.
К антителам, меченным в соответствии с изобретением, или их функциональным фрагментам относятся, например, антитела, называемые иммуноконъюгатами, которые могут быть конъюгированы, например, с ферментами, такими как пероксидаза, щелочная фосфатаза, бета-D-галактозидаза, глюкозооксидаза, глюкозоамилаза, карбоангидраза, ацетилхолинэстераза, лизоцим, малатдегидрогеназа или глюкозо-6-фосфат-дегидрогеназа, или с молекулами, такими как биотин, дигоксигенин или 5-бромдезоксиуридин. Флуоресцентные метки можно аналогичным образом конъюгировать с антителами или их функциональными фрагментами в соответствии с изобретением и особенно включать флуоресцеин и его производные, флуорохром, родамин и его производные, GFP («зеленый флуоресцентный белок»), дансил, умбеллиферон и т.д. В таких конъюгатах антитела изобретения или их функциональные фрагменты могут быть изготовлены с помощью способов, известных специалистам в данной области. Они могут быть связаны с ферментами или флуоресцентными метками непосредственно или посредством разделительной группы или связывающей группы, такой как полиальдегид, например глутаральдегид, этилендиаминтетрауксусная кислота (EDTA), диэтиленгликольтриаминпентауксусная кислота (DPTA), либо в присутствии связывающих агентов, таких как упомянутые выше для терапевтических конъюгатов. Конъюгаты, содержащие метку флуоресцеинового типа, могут быть получены путем реакции с изотиоцианатом.
Другие конъюгаты могут также включать хемолюминисцентные метки, такие как люминол и диоксетаны, биолюминесцентные метки, такие как люцифераза и люциферин, либо радиоактивные метки, такие как йод123, йод125, йод126, йод133, бром77, технеций99m, индий111, индий113m, галий67, галий68, рутений95, рутений97, рутений103, рутений105, меркурий107, меркурий203, рений99m, рений101, рений105, скандий47, теллурий121m, теллурий122m, теллурий125m, тулий165, тулий167, тулий168, фтор18, иттрий199, йод131. Способы, известные специалисту в данной области, существующие для связывания терапевтических радиоизотопов с антителами либо непосредственно, либо через хелатирующий агент, такой как упомянутые выше EDTA, DTPA, могут быть использованы для радиоактивных элементов, которые могут быть использованы в диагностике. Также можно упомянуть метку с Na[I125] с помощью хлорамина Т [Hunter W.M. and Greenwood F.C. (1962) Nature 194:495], либо также с технецием99m в методике Crockford et al. (патент США 4424200), либо через DTPA, как описано у Hnatowich (патент США 4479930).
Таким образом, антитела или их функциональные фрагменты в соответствии с изобретением могут быть использованы в способе выявления и/или количественной оценки сверхэкспрессии или недостаточной экспрессии, предпочтительно сверхэкспрессии, c-Met-рецептора в биологическом образце, характеризующемся тем, что он включает следующие этапы:
a) контакт биологического образца с антителом или одним из его функциональных фрагментов в соответствии с изобретением; и
b) выявление возможно сформированного комплекса с-Met/антитело.
В частном воплощении антитела или их функциональные фрагменты в соответствии с изобретением могут быть использованы для выявления и/или количественной оценки c-Met-рецепторов в биологическом образце, для мониторинга эффективности профилактического и/или терапевтического лечения c-Met-зависимого рака.
В более общем плане антитела или их функциональные фрагменты в соответствии с изобретением могут быть выгодно использованы в любой ситуации, когда необходимо следить за экспрессией c-Met-рецептора в качественном и/или количественном виде.
Предпочтительно биологический образец формируют из биологических жидкостей, таких как сыворотка, цельная кровь, клеток, образцов тканей или биоптатов человеческого происхождения.
Для такого выявления и/или определения дозы может быть использована любая процедура или стандартный тест. Указанный тест может быть конкурентным или сэндвич-тестом или любым тестом, известным специалистам в данной области, в зависимости от формирования иммунного комплекса типа антитело-антиген. После применения в соответствии с изобретением антитело или один из его функциональных фрагментов может быть закреплен и помечен. Это закрепление может осуществляться на многочисленных подложках, известных специалистам в данной области. Эти подложки могут в особенности включать стекло, полистирол, полипропилен, полиэтилен, декстран, нейлон, либо природные или модифицированные клетки. Эти подложки могут быть растворимыми или нерастворимыми.
К примеру, предпочтительный способ включает иммуноферментные способы по методике ИФА, путем иммунофлуоресценции, методики радиоиммуноанализа или эквивалентных им.
Таким образом, данное изобретение также включает комплекты или наборы, необходимые для выполнения способов диагностики болезней, индуцированных сверхэкспрессией или недостаточной экспрессией c-Met-рецепторов, или для выполнения способа выявления и/или количественной оценки сверхэкспрессии или недостаточной экспрессии c-Met-рецепторов в биологическом образце, предпочтительно сверхэкспрессии указанного рецептора, характеризующиеся тем, что указанные комплекты или наборы включают следующие элементы:
a) антитело или один из его функциональных фрагментов в соответствии с изобретением;
b) возможно реагенты для формирования среды, благоприятной для иммунологической реакции;
c) возможно реагенты, позволяющие демонстрировать комплексы с-Met/антитело, полученные в иммунологической реакции.
Аналогичным образом предметом изобретения является применение антитела или композиции в соответствии с изобретением для получения лекарственного средства, предназначенного для специфического воздействия биологически активного соединения на клетки, экспрессирующие или сверхэкспрессирующие c-Met-рецептор.
Под биологически активным соединением здесь понимается любое соединение, способное модулировать, особенно ингибировать, активность клеток, в частности их рост, их пролиферацию, транскрипцию или трансляцию гена.
Предметом изобретения является также диагностический реагент in vivo, содержащий антитело в соответствии с изобретением или один из его функциональных фрагментов, предпочтительно меченых, особенно меченных радиоактивной меткой, и его применение в области медицинской визуализации, в частности для выявления рака, связанного с экспрессией или сверхэкспрессией с-Met-рецептора клеткой.
Аналогичным образом изобретение относится к композиции как комбинированному продукту или к конъюгату анти-с-Met/токсин или радиоактивному элементу в соответствии с изобретением в качестве лекарственного средства.
Предпочтительно указанная композиция как комбинированный продукт или указанный конъюгат в соответствии с изобретением будет смешан с эксципиентом и/или фармацевтически приемлемым носителем.
В данном описании под фармацевтически приемлемым носителем понимается соединение или комбинация соединений, входящих в фармацевтическую композицию, не провоцирующих вторичные реакции и позволяющих, например, облегчить введение активного соединения(ий), увеличить время его жизни и/или его эффективность в организме, повысить его растворимость в растворе или улучшить его сохранность. Эти фармацевтически приемлемые носители хорошо известны и будут адаптированы специалистом в данной области в зависимости от природы и способа введения выбранного активного соединениями).
Предпочтительно эти соединения будут введены системным путем, в частности внутривенно, внутримышечно, внутрикожно, внутрибрюшинно или подкожно, или пероральным путем. В более предпочтительном случае композицию, содержащую антитела в соответствии с изобретением, будут вводить несколько раз в последовательном порядке.
Способы введения, дозировки и оптимальные фармацевтические формы можно определить в соответствии с критериями, обычно принимаемыми при установлении лечения, адаптированного для пациентов, такими как, например, возраст или вес тела пациента, тяжесть его/ее общего состояния, толерантность к лечению и отмеченные вторичные эффекты.
Другие характеристики и преимущества изобретения появятся в последующем описании с примерами и фигурами, где:
Фиг.1: Примеры FACS-профилей отобранных анти-с-Met-антител;
Фиг.2А и 2В: Ингибирование in vitro пролиферации ВХРС3 с помощью антител, направленных на c-Met;
Фиг.3: Ингибирование c-Met-димеризации;
Фиг.4: Распознавание белка анти-с-Met-антителами;
Фиг.5А и 5В: "Картирование эпитопов" 11Е1 и 5D5 путем анализа BIAcore;
Фиг.6А и 6В: Влияние моноклональных антител на c-Met-фосфорилирование;
Фиг.7А и 7В: Вытеснение радиомеченого HGF анти-с-Met-антителами;
Фиг.8: Ингибирование инвазии с помощью анти-с-Met-антител [на этой фигуре SVF обозначает эмбриональную телячью сыворотку (FCS)];
Фиг.9: Влияние анти c-Met-антител на заживление ран;
Фиг.10А и 10В: Анализ рассеяния;
Фиг.11: Трехмерный анализ тубулогенеза;
Фиг.12А и 12В: Влияние антител на образование сфероида;
Фиг.13: Активность in vivo моноклональных антител анти-c-Met на модели с ксенотрансплантатом U87MG;
Фиг.14: Экспрессия HGF группой линий опухолевых клеток;
Фиг.15А и 15В: Характеристика клеточной линии NCI-H441; фиг.15А соответствует анализу количественной ОТ-ПЦР, фиг.15В соответствует FACS-анализу;
Фиг.16: Активность in vivo анти-с-Met-антител на модели с ксенотрансплантатом NCI-H441;
Фиг.17А: Выравнивание 224G11 VL с мышиным зародышевым геном IGKV3-5*01;
Фиг.17В: Выравнивание 224G11 VL с мышиным зародышевым геном IGKJ4*01;
Фиг.18А: Выравнивание 224G11 VL с человеческими зародышевыми генами IGKV3-11*01 и IGKV4-1*01;
Фиг.18В: Выравнивание 224G11 VL с человеческим зародышевым геном IGKJ4*02;
Фиг.19А: Основанная на IGKV3-11*01 гуманизированная версия 224G11 VL с вышеупомянутыми мутациями;
Фиг.19В: Основанная на IGKV4-1*01 гуманизированная версия 224G11 VL с вышеупомянутыми мутациями;
Фиг.20А: Выравнивание 224G11 VH с мышиным зародышевым геном IGHV1-18*01;
Фиг.20В: Выравнивание 224G11 VH с мышиным зародышевым геном IGHD2-4*01;
Фиг.20С: Выравнивание 224G11 VH с мышиным зародышевым геном IGHJ2*01;
Фиг.21А: Выравнивание 224G11 VH с человеческим зародышевым геном IGHV1-2*02;
Фиг.21В: Выравнивание 224G11 VH с человеческим зародышевым геном IGHJ4*01;
Фиг.22: Гуманизированный 224G11 VH с вышеупомянутыми мутациями;
Фиг.23А: Выравнивание 227Н1 VL с мышиным зародышевым геном IGKV3-5*01;
Фиг.23В: Выравнивание 227Н1 VL с мышиным зародышевым геном IGKJ4*01;
Фиг.24А: Выравнивание 227Н1 VL с человеческими зародышевыми генами IGKV3-11*01 и IGKV4-1*01;
Фиг.24В: Выравнивание 227Н1 VL с человеческим зародышевым геном IGKJ4*02;
Фиг.25А: Основанная на IGKV3-11*01 гуманизированная версия 227Н1 VL с вышеупомянутыми мутациями;
Фиг.25В: Основанная на IGKV4-1*01 гуманизированная версия 227Н1 VL с вышеупомянутыми мутациями;
Фиг.26А: Выравнивание 227Н1 VH с мышиным зародышевым геном IGHV1-18*01;
Фиг.26В: Выравнивание 227Н1 VH с мышиным зародышевым геном IGHD1-1*02;
Фиг.26C: Выравнивание 227Н1 VH с мышиным зародышевым геном IGHJ2*01;
Фиг.27А: Выравнивание 227Н1 VH с человеческим зародышевым геном IGHV1-2*02;
Фиг.27В: Выравнивание 227Н1 VH с человеческим зародышевым геном IGHJ4*01;
Фиг.28: Гуманизированный 227Н1 VH с вышеупомянутыми мутациями;
Фиг.29А: Выравнивание 22304 VL с мышиным зародышевым геном IGKV12-46*01;
Фиг.29В: Выравнивание 22304 VL с мышиным зародышевым геном IGKJ2*01;
Фиг.30А: Выравнивание 22304 VL с человеческим зародышевым геном IGKV1-NL1*01;
Фиг.30В: Выравнивание 22304 VL с человеческим зародышевым геном IGKJ2*01;
Фиг.31: Гуманизированный 22304 VL с вышеупомянутыми мутациями;
Фиг.32А: Выравнивание 223С4 VH с мышиным зародышевым геном IGHV1-18*01;
Фиг.32В: Выравнивание 223С4 VH с мышиным зародышевым геном IGHD6-3*01;
Фиг.32С: Выравнивание 223С4 VH с мышиным зародышевым геном IGHJ4*01;
Фиг.33А: Выравнивание 223С4 VH с человеческим зародышевым геном IGHV1-2*02;
Фиг.33В: Выравнивание 223С4 VH с человеческим зародышевым геном IGHD1-26*01;
Фиг.33С: Выравнивание 223С4 VH с человеческим зародышевым геном IGHJ6*01; и
Фиг.34: Гуманизированный 223С4 VH с вышеупомянутыми мутациями;
Фиг.35: Противоопухолевая активность мышиного моноклонального антитела 224G11 отдельно или в комбинации с Навельбином® на принятой опухолевой модели с ксенотрансплантатами NCI-H441;
Фиг.36: Оценка моноклональных антител анти-c-Met при пролиферации HUVEC;
Фиг.37: Оценка моноклональных антител анти-c-Met при формировании трубочек HUVEC;
Фиг.38А: Выравнивание 11Е1 VL с мышиным зародышевым геном IGKV4-79*01;
Фиг.38В: Выравнивание 11Е1 VL с мышиным зародышевым геном IGKJ4*01;
Фиг.39А: Выравнивание 11Е1 VL с человеческим зародышевым геном IGKV3D-7*01;
Фиг.39В: Выравнивание 11Е1 VL с человеческим зародышевым геном IGKJ4*02;
Фиг.40: Гуманизированная версия 11Е1 VL с вышеупомянутыми мутациями;
Фиг.41А: Выравнивание 11Е1 VH с мышиным зародышевым геном IGHV1-7*01;
Фиг.41В: Выравнивание 11Е1 VH с мышиным зародышевым геном IGHD4-1*01;
Фиг.41С: Выравнивание 11Е1 VH с мышиным зародышевым геном IGHJ3*01;
Фиг.42А: Выравнивание 11Е1 VH с человеческими зародышевыми генами IGHV1-2*02 и IGHV1-46*01;
Фиг.42В: Выравнивание 11Е1 VH с человеческим зародышевым геном IGHJ4*03;
Фиг.43: Гуманизированный 11Е1 VH с вышеупомянутыми мутациями;
фиг.44А и 44В: Анализ c-Met-фосфорилирования в клетках А549. Оценка очищенных моноклональных антител 11Е1 и 224G11 в отсутствие или в присутствии HGF, либо при 30 мкг/мл (фиг.44А), либо в диапазоне доз от 0,0015 до 30 мкг/мл, для определения значения ЕС50 (фиг.44В);
Фиг.45: Комбинация in vivo моноклонального антитела 224G11 с Навельбином® на модели с ксенотрансплантатом NSCLC NCI-H441;
Фиг.46: Комбинация in vivo моноклонального антитела 224G11 с доксорубицином на модели с ксенотрансплантатом NSCLC NCI-H441;
Фиг.47: Комбинация in vivo моноклонального антитела 224G11 с доцетакселом на модели с ксенотрансплантатом NSCLC NCI-H441;
Фиг.48: Комбинация in vivo моноклонального антитела 224G11 с темозоломидом на модели с ксенотрансплантатом NSCLC NCI-H441;
Фиг.49А, 49В, 49С и 49D: Влияние моноклональных антител анти-c-Met на рост сфероида U87-MG;
Фиг.50А и 50В: Активность in vitro химерных и гуманизированных форм 224G11 в фосфо-с-Met-анализе;
Фиг.51: Параметры анализа Biacore;
Фиг.52: Активность in vivo 224G11 в клетках MDA-MB-231, имплантированных вместе с клетками MRC5 в качестве источника человеческого HGF на атимических голых мышах;
Фиг.53: Основанный на ИФА анализ связывания с Fc-c-Met. Анти-Fc-c-Met-связывающую активность измеряли в анализе, основанном на ИФА, где использовали противомышиные Fc-конъюгаты для обнаружения очищенных мышиных моноклональных антител 11Е1, 224G11 и 227Н1. Дозозависимую связывающую активность на пластике, покрытом рекомбинантным Fc-c-Met, измеряли при 450 нм;
Фиг.54: HGF-c-Met конкурентный анализ. В этом анализе, основанном на ИФА, остаточное связывание рекомбинантного Fc-c-Met с пластиком, покрытым HGF, в присутствии очищенных мышиных моноклональных антител 11Е1, 224G11 и 227Н1 обнаруживали с помощью противомышиных Fc-конъюгатов и измеряли при 450 нм;
Фиг.55: Выравнивание аминокислотных последовательностей рекомбинантных доменов VH, полученных из 227Н1. Аминокислотная последовательность 227Н1 VH выравнена с отобранной принимающей человеческой каркасной последовательностью, с только упомянутыми аминокислотами, которые отличаются от мышиной последовательности 227Н1 VH. Последовательности 227Н1 HZ1, HZ2 и HZ3 VH соответствуют осуществленным гуманизированным версиям мышиного домена VH 227Н1, при этом остающиеся мышиными остатки выделены жирным шрифтом. В HZ3 10 остатков (*) были автоматически заменены на их человеческие аналоги. В HZ2 были изучены семь остатков из третьей группы (3). В HZ1VH девять остатков из второй группы (2) подвергли превращению в их человеческие аналоги, только шесть остатков из первой группы (1) остались мышиными;
Фиг.56: Основанный на ИФА анализ связывания рекомбинантных антител 227Н1 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 227Н1. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антител 227Н1, полученных из гуманизированных VH-доменов, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.57: Основанный на ИФА анализ связывания рекомбинантных антител, полученных из 227Н1, с Fc-c-Met. Анти-Fc-c-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 227Н1. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антитела 227Н1, полученного из гуманизированного HZ4VH, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.58: HGF-c-Met конкурентный анализ мышиных и рекомбинантных антител 227Н1. В этом анализе, основанном на ИФА, остаточное связывание рекомбинантного Fc-c-Met с пластиком, покрытым HGF, в присутствии различных форм антитела 227Н1 обнаруживали с биотинилированным несвязанным анти-с-Met-антителом. Тестировали очищенное мышиное монокпональное антитело 227Н1, химерное антитело и рекомбинантное антитело, полученное из гуманизированного 227Н1, полученного из HZ4VH, и сравнивали их способность конкурировать в связывании с HGF-c-Met, измеренную при 450 нм;
Фиг.59: Последовательность вариабельного домена гуманизированного VH 227Н1-HZ. * соответствует аминокислотам, замененным фактически на их человеческие аналоги; ! соответствует аминокислотам, гуманизированным в ходе осуществления HZ3 в HZ1; § соответствует аминокислотам, гуманизированным в конечной последовательности 227H1-HZ VH;
Фиг.60: Выравнивание аминокислотных последовательностей рекомбинантных VH-доменов, полученных из 11Е1. Аминокислотная последовательность 11Е1 VH выравнена с отобранной принимающей человеческой каркасной последовательностью, с только упомянутыми аминокислотами, которые отличаются от мышиной последовательности 11Е1 VH. Последовательности 11Е1 HZ VH1, VH2 и VH3 соответствуют осуществленным гуманизированным версиям мышиного VH-домена 11Е1, при этом остающиеся мышиными остатки выделены жирным шрифтом. В HZ VH3 7 остатков (*) были автоматически заменены на их человеческие аналоги. В HZ VH2 были изучены семь остатков из третьей группы (3). В HZ VH1 пять остатков из второй группы (2) подвергли превращению в их человеческие аналоги, только пять остатков из первой группы (1) остались мышиными;
Фиг.61: Основанный на ИФА анализ связывания рекомбинантных антител, полученных из 11Е1, с Fc-c-Met. Анти-Fc-c-Met связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 11Е1. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антител 11Е1, полученных из гуманизированных VH-доменов, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.62: Выравнивание аминокислотных последовательностей рекомбинантных VL-доменов, полученных из 11Е1. Аминокислотная последовательность 11Е1 VL выравнена с отобранной принимающей человеческой каркасной последовательностью, с только упомянутыми аминокислотами, которые отличаются от мышиной последовательности 11Е1 VL. Последовательности 11Е1 HZ VL1, VL2 и VL3 соответствуют осуществленным гуманизированным версиям мышиного домена VL 11Е1, при этом остающиеся мышиными остатки выделены жирным шрифтом. В HZ VL3 десять остатков (*) были автоматически заменены на их человеческие аналоги. В HZ VL2 были изучены восемь остатков из третьей группы (3). В HZ VL1 восемь остатков из второй группы (2) подвергли превращению в их человеческие аналоги, только четыре остатка из первой группы (1) остались мышиными;
Фиг.63: Основанный на ИФА анализ связывания рекомбинантных антител 11Е1 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 11Е1. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антител 11Е1, полученных из гуманизированных VL-доменов, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.64: Основанный на ИФА анализ связывания рекомбинантных антител 11Е1 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 11Е1. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антител 11Е1, полученных из однократно или двукратно гуманизированных доменов, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.65: Выравнивание аминокислотной последовательности домена VH 224G11. Аминокислотная последовательность 224G11 VH выравнена с последовательностью 227Н1 VH (подчеркнутые остатки являются негомологичными) и с отобранной принимающей человеческой каркасной последовательностью, с только упомянутыми аминокислотами, которые отличаются от мышиной последовательности 224G11 VH. Последовательность 224G11 HZ VH0 соответствует гуманизированной версии мышиного VH-домена 224G11 «основанный на 227Н1/полностью IMGT». В этой последовательности нет остатков вне-IMGT-CDR, которые остались бы мышиными;
Фиг.66: Основанный на ИФА анализ связывания рекомбинантных антител 224G11 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения рекомбинантных химерных антител и антител, полученных из гуманизированных 224G11, полученных из HZVH0. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антитела 224G11, полученного из гуманизированного VH-домена "полностью IMGT" HZVHO, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.67: HGF-c-Met конкурентный анализ мышиного и рекомбинантного антител 224G11. В этом анализе, основанном на ИФА, остаточное связывание рекомбинантного Fc-c-Met с пластиком, покрытым HGF, в присутствии различных форм антитела 224G11, обнаруживали с помощью биотинилированного несвязанного анти-с-Met-антитела. Очищенное мышиное моноклональное антитело 224G11, рекомбинантные химерное антитело и антитело, полученное из гуманизированного 224G11, полученного из HZVHO, тестировали и сравнивали их способность конкурировать в связывании HGF-c-Met, измеренную при 450 нм;
Фиг.68: Выравнивание аминокислотных последовательностей VL-домена 224G11. Аминокислотная последовательность 224G11 VL выравнена с двумя отобранными принимающими человеческими каркасными последовательностями, с только упомянутыми аминокислотами, которые отличались от мышиной последовательности 224G11 VL. Последовательность 224G11 HZ VL3 соответствует гуманизированной версии мышиного 224G11 VH-домена "короткий CDR1", в то время как HZ VL6 соответствует версии "длинный CDR1", а оставшиеся мышиными остатки выделены жирным шрифтом. Для обеих основных гуманизированных версий остатки, оставшиеся мышиными, упорядочены по способу дальнейшей гуманизации, где * соответствует аминокислотам, гуманизированным в базисных версиях, а 3, 2 и 1 соответствуют группам остатков для разработки осуществленных гуманизированных версий;
Фиг.69: Основанный на ИФА анализ связывания рекомбинантных антител 224G11 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 224G11. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антител 224G11, полученных из гуманизированных доменов VL3 и VL6, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.70: Основанный на ИФА анализ связывания рекомбинантных антител 224G11 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 224G11. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антител 224G11, полученных из гуманизированных VL-доменов, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.71: HGF-c-Met-конкурентный анализ мышиного и рекомбинантного антител 224G11. В этом анализе, основанном на ИФА, остаточное связывание рекомбинантного Fc-c-Met с пластиком, покрытым HGF, в присутствии различных форм антитела 224G11, обнаруживали с помощью биотинилированного несвязанного анти-c-Met антитела. Очищенное мышиное моноклональное антитело 224G11, рекомбинантные химерное и гуманизированное антитела, полученные из 224G11, полученного из HZ VL4, тестировали и сравнивали их способность конкурировать в связывании HGF-c-Met, измеренную при 450 нм;
Фиг.72: Аминокислотная последовательность гуманизированного VL-домена 224G11 VL4. * соответствует аминокислотам, замененным фактически на их человеческие аналоги в базисной версии HZ VL6; ! соответствует аминокислотам, гуманизированным в ходе осуществления HZ VL6 в HZ VL4; § соответствует аминокислотам, которые остаются мышиными в последовательности 224G11-HZ VL4;
Фиг.73: Основанный на ИФА анализ связывания рекомбинантных антител 224G11 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 22G11. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность антител 224G11, полученных из однократно или двукратно гуманизированных доменов, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.74: HGF-c-Met-конкурентный анализ мышиного и рекомбинантного антител 224G11. В этом анализе, основанном на ИФА, остаточное связывание рекомбинантного Fc-c-Met с пластиком, покрытым HGF, в присутствии различных форм антитела 224G11, обнаруживали с помощью биотинилированного несвязанного анти-с-Met-антитела. Очищенное мышиное моноклональное антитело 224G11, химерное и полностью гуманизированное рекомбинантные антитела, полученные из 224G11, тестировали и сравнивали их способность конкурировать в связывании HGF-c-Met, измеренную при 450 нм;
Фиг.75: Основанный на ИФА анализ связывания рекомбинантных антител 224G11 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 22G11. При 450 мм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность одиночных мутантов полностью гуманизированных антител 224G11, полученных из VL4, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.76: Основанный на ИФА анализ связывания рекомбинантных антител 224G11 с Fc-c-Met. Анти-Fc-с-Met-связывающую активность измеряли в основанном на ИФА анализе, где использовали противочеловеческие Fc-конъюгаты для обнаружения химерных и гуманизированных рекомбинантных антител, полученных из 22G11. При 450 нм на пластике, покрытом рекомбинантным Fc-c-Met, измеряли дозозависимую связывающую активность одиночных или множественных мутантов полностью гуманизированных антител 224G11, полученных из VL4, а затем сравнивали с активностью родительского/контрольного химерного антитела;
Фиг.77: HGF-c-Met-конкурентный анализ мышиных и рекомбинантных антител 224G11. В этом анализе, основанном на ИФА, остаточное связывание рекомбинантного Fc-c-Met с пластиком, покрытым HGF, в присутствии различных форм антитела 224G11, обнаруживали с помощью биотинилированного несвязанного анти-с-Met-антитела. Очищенное мышиное моноклональное антитело 224G11, химерные и одиночные или множественные мутанты полностью гуманизированных рекомбинантных антител 224G11, полученных из VL-4, тестировали и сравнивали их способность конкурировать в связывании HGF-c-Met, измеренную при 450 нм.
Пример 1: Создание антител против c-Met
Для создания анти-с-Met-антител 8-недельных BALB/c мышей иммунизировали либо от 3 до 5 раз подкожно трансфицированной клеточной линией СНО (клетки яичников китайских хомячков), которая экспрессирует c-Met на плазматической мембране (20×106 клеток/доза/мышь), либо от 2 до 3 раз гибридным белком внеклеточного домена c-Met (10-15 м кг/доза/мышь) (R&D Systems, каталоговый номер 358МТ) или фрагментами этого рекомбинантного белка, смешанными с полным адъювантом Фрейнда для первой иммунизации и неполным адъювантом Фрейнда для последующих. Также были выполнены смешанные протоколы, в которых мыши получали как CHO-c-Met-клетки, так и рекомбинантные белки. За три дня до слияния клеток мышей бустировали внутрибрюшинно или внутривенно рекомбинантным белком или фрагментами. Затем собирали селезенки мышей, проводили слияние с миеломными клетками SP2/0-Ag14 (АТСС) и подвергали селекции на среде HAT. Проводили четыре слияния. В общем, для получения моноклональных антител или их функциональных фрагментов, особенно мышиного происхождения, можно сослаться на методики, описанные в частности в руководстве "Antibodies" (Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor NY, pp.726, 1988), или на методику получения гибридом, описанную Kohler и Milstein (Nature, 256:495-497, 1975).
Полученные гибридомы вначале проверяли с помощью ИФА на c-Met рекомбинантном белке, а затем с помощью FACS-анализа на А549 NSCLC, панкреатической ВхРС3 и глиобластомной U87-MG линиях клеток (представленные профили представлены в фиг.1), чтобы удостовериться, что полученные антитела также смогут распознавать нативный рецептор на опухолевых клетках. Для этих двух тестов амплифицировали и клонировали положительные контроли, восстанавливали и очищали группу гибридом и проверяли их способность ингибировать in vitro пролиферацию клеток на модели ВхРС3.
Для этой цели 50000 клеток ВхРС3 помещали в 96-луночную плату в среду RPMI, 2 мМ L-глутамина, без SVF (эмбриональной телячьей сыворотки). Через 24 часа после внесения тестируемые антитела добавляли в конечной концентрации в диапазоне от 0,0097 до 40 мкг/мл за 60 минут до добавления 100 нг/мл hHGF. Через 3 дня клетки выдерживали с 0,5 мкКи [3H]-тимидина в течение 16 часов. Количество [3H]-тимидина, включенного в ДНК, нерастворимую в трихлоруксусной кислоте, было подсчитано с помощью жидкого сцинтилляционного счетчика. Результаты представлены в виде исходных данных, чтобы реально оценить внутренний агонистический эффект каждого моноклонального антитела (фиг.2А и 2В).
Затем антитела, ингибирующие по меньшей мере на 50% клеточную пролиферацию, оценивали путем анализа BRET в супернатантах c-Met трансфицированных клеток. Для этой цели создавали стабильные клеточные линии СНО, экспрессирующие c-Met-Rluc или c-Met-Rluc и c-Met-KHOOA-YFP. Клетки распределяли в белых 96-луночных микроплатах в DMEM-F12/FBS 5% культуральной среде за один или два дня до экспериментов BRET. Клетки вначале культивировали при 37°C с 5% СО2, с тем чтобы клетки прикрепились к плате. Затем клетки лишали питания с 200 мкл DMEM на лунку в течение ночи. Непосредственно перед началом эксперимента DMEM удаляли, а клетки быстро промывали PBS. Клетки инкубировали в PBS в присутствии или в отсутствие тестируемых антител или контрольных соединений в течение 10 мин при 37°С перед добавлением коэлентеразина с или без HGF в конечном объеме 50 мкл. После инкубации в течение еще 10 минут при 37°С начинали регистрировать светоизлучение при 485 нм и 530 нм с использованием люминометра Mithras (Berthold) (1 с/длина волны/лунка, повторение 15 раз).
Соотношение BRET было определено ранее [Angers et al., Proc. Natl. Acad. Sci. USA, 2000, 97:3684-3689] как [(излучение при 530 нм) - (излучение при 485 нм) × Cf]/(излучение при 485 нм), где Cf соответствует (излучение при 530 нм)/(излучение при 485 нм) для клеток, экспрессирующих только гибридный белок Rluc в тех же экспериментальных условиях. Упрощение этого уравнения показывает, что соотношение BRET соответствует соотношению 530/485 нм, полученному, когда представлены оба аналога, скорректированному по отношению к 530/485 нм, полученному в тех же экспериментальных условиях, когда в анализе присутствует только аналог, слившийся с люциферазой Р. reniformis. Для удобочитаемости результаты выражены в единицах миллиВРЕТ (mBU); mBU соответствует соотношению BRET, умноженному на 1000.
После этого второго теста in vitro были отобраны 4 антитела i) без внутренней активности в виде целой молекулы в функциональном тесте пролиферации, ii) значительно ингибирующие пролиферацию ВхРС3 (фиг.2А и 2В) и iii) ингибирующие c-Met-димеризацию (фиг.3). Эти 4 антитела IgG1 каппа-изотипа были описаны как 11Е1, 224G11, 223С4 и 227Н1. В экспериментах моноклональное антитело 5D5, полученное Genentech и доступное из АТСС, было добавлено в качестве контроля внутренней агонистической активности.
Фиг.2А и 2В демонстрируют, что 11Е1, 224G11, 223С4 и 227Н1 были без какой-либо агонистической активности в отличие от 5D5, которое индуцировало дозозависимую стимуляцию клеточной пролиферации в отсутствие лиганда. Значительное ингибирование клеточной пролиферации наблюдалось с 4 отобранными антителами. 5D5 не влиял на HGF-индуцированную клеточную пролиферацию в этом тесте.
При оценке блокады димеризации наблюдался значительный эффект, достигающий 32, 55, 69 и 52%-ного ингибирования димеризации для 224G11, 223С4, 11Е1 и 227Н1 соответственно. По сравнению с базальными сигналами в соответствующих экспериментах антитело 5D5 не влияло на эту модель димеризации.
Пример 2: Распознавание белка с помощью анти-c-Met антител
Чтобы охарактеризовать модель распознавания четырех отобранных антител, были поставлены четыре ИФА с рекомбинантным белком c-Met, его мономерным фрагментом (полученным путем расщепления рекомбинантного с-Met-Fc-белка и рекомбинантного домена SEMA).
Результаты, представленные в фиг.4, демонстрируют, что четыре антитела распознают как димерные, так и мономерные белки. Для выполнения этих ИФА человеческий димерный c-Met-белок (R&D Systems, каталоговый номер 358МТ) вносили в платы в концентрации 0,7 мкг/мл в PBS и оставляли в течение ночи при 4°С. После насыщения плат (Costar, номер 3690) 0,5% раствором желатина в течение 2 часов при 37°С, супернатанты гибридом инкубировали в течение 1 часа при 37°С. После промывания в PBS в каждую лунку добавляли противомышиное HRP-антитело (Jackson ImmunoResearch, каталоговый номер 115-035-164) в разведении 1/5000 в буфере для ИФА (0,1% желатин/0,05% Твин 20 в PBS) и инкубировали платы в течение 1 часа при температуре 37°С. После трех промывок в PBS выявляли активность пероксидазы путем добавления 50 мкл ТМБ-субстрата (Uptima). Реакция протекала в течение 5 минут при комнатной температуре. Реакцию останавливали добавлением 50 мкл/лунка 1 М раствора H2SO4 и анализировали на планшетном анализаторе при 450 нм. Такой же протокол выполняли для мономерных доменов c-Met и SEMA, но в этом случае белки вносили в плату в концентрации 5 и 3 мкг/мл соответственно.
Моноклональное антитело 5D5 вводили в качестве положительного контроля, распознающего, как и ожидалось, белок SEMA. 224G11, 227Н1 и 223С4 не связывали домен SEMA. 11Е1 способен связывать SEMA.
Чтобы выявить 11Е1 и 5D5, которые, распознавая домен SEMA, конкурируют за перекрывающиеся эпитопы, проводили анализ BIAcore. Система BIAcore, основанная на феномене резонанса поверхностных плазмонов, поставляла данные путем мониторинга событий связывания в реальном времени. Затем ее использовали для группы антител в так называемых экспериментах "картирования эпитопов". Пара антител, не способных связывать в одно и то же время молекулу антигена, классифицируется в одну и ту же группу (идентичные или соседние сайты связывания). Напротив, когда их соответствующие сайты связывания достаточно далеки, чтобы позволить одновременно связываться обоим антителам, то эти последние классифицируются в две различные группы. В таких экспериментах антиген обычно использовался в качестве лиганда (иммобилизованного на сенсорном чипе), а антитела использовались без какого-либо мечения в качестве аналитов (растворенная фаза).
Все описанные эксперименты были проведены на приборе BIAcore X (GE Healthcare Europe GmbH). Сенсорный чип СМ5 (BIAcore), активируемый мышиным моноклинальным антителом анти-Tag-6His (R&D Systems, см. МАВ050), был изготовлен в соответствии с инструкциями производителя с помощью набора для связывания аминов (BIAcore). Рабочий буфер (HBS-EP) и регенерационный буфер (глицин, HCl) получены от BIAcore. Рекомбинантная растворимая версия человеческого рецептора HGF, полученная как химерная молекула c-Met-Fc-Tag His, была взята от R&D Systems (см. 358-MT-CF). Эксперименты проводились при 25°С при скорости потока 30 мкл/мин. 10 мкг/мл раствор c-Met в рабочем буфере вводили в течение одной минуты в измерительный участок 2 (fc2), обычно 270 RU растворимой формы c-Met было захвачено. Измерительный участок 1 (fc1) использовали в качестве контроля для проверки любого неспецифического связывания антител с матрицей в виде сенсорного чипа.
Проводили последовательные введения тестируемых антител. Антитело вводили в оба измерительных участка в течение двух минут. Затем вводили второе антитело (или то же самое) в тех же условиях. Если не наблюдалось значительное связывание, то проводили третье введение с другим антителом. Сенсорный чип затем восстанавливали с помощью одного введения регенерационного буфера в течение 30 секунд. Либо антитела и c-Met-Fc удаляли на этом этапе.
Анализ результатов:
Способность антитела "А" блокировать связывание антитела "В" рассчитывается как соотношение BIA/C=(R2A/B/R1B)×100: где R2A/B представляет собой ответ, соответствующий связыванию моноклонального антитела "В", когда его вводили после моноклонального антитела "A", a R1B представляет собой ответ, соответствующий связыванию моноклонального антитела "В", когда его вводили в первую очередь. BIA/C ниже 20% означает, что А способен блокировать связывание В, так что А и В имеют соседние сайты связывания.
Картирование эпитопов выполняли с двумя моноклональными антителами 11Е1 и 5D5.
|
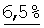
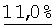
Визуализация связывания примерно 270RU захваченного c-Met-Fc путем последовательных двухминутных введений моноклональных антител 5D5 (первое), 5D5 (второе) и 11Е1 (третье) в концентрации 10 мкг/мл каждый, показали, что 5D5 и 11Е1 четко связывают два удаленных сайта (фиг.5А). Это наблюдение было подтверждено при обратной последовательности антител (фиг.5В).
Таблица 3 обобщает рассчитанные соотношения, полученные для разных последовательностей этих двух антител. Выделение значений черным (более 75%) означает, что моноклональное антитело А не блокирует связывание моноклонального антитела В. Выделение значений жирным шрифтом/курсивом (менее 20%) означает, что сайты связывания обоих антител (А и В) являются идентичными или достаточно близкими, что делает невозможным одновременное связывание.
Пример 3: Влияние моноклональных антител на c-Met фосфорилирование
Для определения влияния анти-с-Met-антител на c-Met фосфорилирование, ставили ИФА с фосфо-c-Met. Кратко говоря, 500000 клеток А549 сеяли в каждую лунку 6-луночной платы в среде F12K+10% FCS. За 16 часов до добавления HGF (100 нг/мл) клетки лишали питания и за 15 минут до стимуляции лигандом каждое анализируемое антитело добавляли до конечной концентрации 30 мкг/мл. Через 15 минут после добавления HGF добавляли холодный лизирующий буфер, соскабливали клетки, собирали клеточный лизат и центрифугировали его при 13000 об/мин в течение 10 мин при 4°С. Супернатанты оценивали количественно с помощью набора ВСА (Pierce) и сохраняли при температуре -20°С. Для ИФА использовали козлиные анти-с-Met-антитела (R&D, см. AF276) в качестве первичных антител (антител захвата) (оставляли в течение ночи при 4°С) и после этапа насыщения (1 ч при комнатной температуре) с TBS-BSA 5% буфером в каждую лунку 96-луночной платы добавляли 25 мкг белка из различных клеточных лизатов. Через 90 минут инкубации при комнатной температуре платы промывали четыре раза и добавляли анти-фосфо-с-Met-антитела (кроличьи анти-pY1230-1234-1235 c-Met). После дополнительной часовой инкубации и четырехкратной промывки добавляли противокроличьи-HRP (Biosource) на 1 час при комнатной температуре, а затем добавляли субстрат Люминол перед оценкой люминесценции с помощью прибора Mithras. Результаты, представленные в фиг.6 В, показали, что 11Е1, 224G11, 223С4 и 227Н1 ингибируют c-Met-фосфорилирование на 68, 54, 80 и 65% соответственно по сравнению с монокпональным антителом 5D5, которое демонстрирует слабое ингибирование c-Met-фосфорилирования (42%). В ходе этого испытания наблюдается слабый базальный эффект (менее 20%) с четырьмя кандидатными антителами (фиг.6А). Как описано в различных примерах, представленных в данном патенте, этот слабый базальный эффект не имеет последствий для активности антител в других испытаниях in vitro и in vivo. 5D5, используемое в качестве контроля, демонстрирует в этом испытании значительный базальный эффект.
Пример 4: Вытеснение радиомеченого HGF анти-с-Met-антителами
Чтобы определить, способны ли анти-с-Met-антитела вытеснять HGF, были поставлены эксперименты по связыванию. Кратко говоря, 96-луночные микроплаты FlashPlate (Perkin Elmer) с белком А насыщали 0,5% желатином в PBS (200 мкл на лунку, 2 ч при комнатной температуре) перед добавлением рекомбинантного c-Met-Fc (R&D Systems) в качестве покрывающего белка. В каждую лунку добавляли 2000 мкл 1 мкг/мл c-Met-Fc раствора в PBS. Затем платы инкубировали в течение ночи при 4°С. Свободные оставшиеся участки с белком А затем насыщали нерелевантным hIgG (0,5 мкг/лунка в PBS) в течение 2 ч при комнатной температуре. Платы промывали PBS после каждого этапа.
В конкурентных анализах связывание [125I]-HGF (специфическая активность ~2000 Ки/ммоль) при 200 пкМ для иммобилизованных c-Met измеряли в присутствии анти-c-Met моноклональных антител 11Е1, 224G11, 223С4, 227Н1 или HGF (R&D Systems) в различных концентрациях в диапазоне от 0,1 пкМ до 1 мкМ в PBS рН 7,4. Платы инкубировали при комнатной температуре в течение 6 ч, затем оценивали на сцинтилляционный счетчике Packard Top Count Microplate Scintillation Counter. Неспецифическое связывание было выявлено в присутствии 1 мкМ HGF. Моноклональное антитело 9G4, которое не направлено на c-Met, но специфично распознает белок E.coli, было использовано в качестве мышиного lgG1 изотипического контроля.
Процент от общего специфического [125I]-HGF-связывания графически представляли как функцию концентрации лиганда на полулогарифмических графиках. Концентрации различных ингибиторов, необходимых для ингибирования связывания радиолиганда на 50% (IC50), определяли графически из полученных сигмоидных конкурентных кривых (фиг.7А и 7В).
Как и ожидалось, немеченый HGF способен полностью вытеснить [125I]-HGF, связанный с иммобилизованным c-Met, в то время как контрольное антитело 9G4 не показывает никакой HGF-блокирующей активности (фиг.7А и 7В).
Моноклональные анти-с-Met-антитела 11Е1, 224G11, 223С4 и 227Н1 способны ингибировать связывание [125I]-HGF с иммобилизованным c-Met со значениями IC50 20 нМ, 3 нМ, 2,7 нМ и 5,8 нМ соответственно. Значения IC50, определенные для антител 224G11, 223С4 и 227Н1, были сопоставимы со значениями IC50, определенными для немеченого HGF, которые включали от 3 до 5 нМ, тогда как антитело 11Е1 демонстрировало более высокое значение IC50.
Пример 5: Ингибирование инвазии анти-с-Met-антителами
Для оценки ингибирующего влияния антител анти-c-Met на процесс инвазии клетки А549 вносили в верхнюю камеру инвазионных камер BD BioCoat™ Matrigel™ (лунки диаметром 6,5 мм с поликарбонатными мембранами для предварительной очистки с диаметром пор 8 мкм). Клетки А459 лишали питания за 24 часа до выполнения анализа инвазии. Затем 500000 клеток А549 помещали в буфере для хемотаксиса (DMEM среда, 0,1% BSA, 12 мМ Hepes) в верхнюю лунку каждой камеры, на покрытие Матригель, с или без тестируемого антитела (конечная концентрация моноклонального антитела 10 мкг/мл). Через 1 час инкубации платы при температуре 37°C с 5% CO2 нижние камеры заполняли либо ростовой средой, содержащей 400 нг/мл rhHGF (рекомбинантного человеческого HGF), либо только ростовой средой. Камеры инкубировали в течение дополнительных 48 часов при температуре 37°C с 5% CO2. В конце времени инкубации клетки, которые остались на верхней поверхности фильтра, аккуратно удаляли с помощью ватного тампона, клетки, которые мигрировали на нижнюю поверхность фильтра, лизировали, метили краской-буфером CyQuant GR (Invitrogen) и подсчитывали с помощью флуоресцентного фотометра Berthold Mithras LB940. Все условия повторяли трижды.
Как и ожидалось, HGF вызывал значительную инвазию опухолевых клеток, сопоставимую с той, которая наблюдается при введении 10% FCS в качестве положительного контроля (фиг.8). Мышиный lgG1 9G4, введенный в качестве изотипического контроля, не оказывал существенного влияния на базальную или HGF-индуцированную инвазию при сравнении с клетками, инкубированными без IgG. Отсутствие агонистического эффекта было отмечено у 11Е1, 224G11, 223С4 и 227Н1, когда их добавляли по одному, а значительное и сопоставимое ингибирование HGF-индуцированной инвазии наблюдалось при использовании трех моноклональных антител.
Пример 6: Ингибирование заживления ран анти-c-Met антителами
HGF стимулирует подвижность. Чтобы определить, способны ли анти-HGF-антитела ингибировать миграцию, NCI-H441-клетки выращивали до высокой плотности и создавали промежуток с помощью наконечника для пипетки Р200. Затем клетки стимулировали к миграции через промежуток с помощью HGF (100 нг/мл) в присутствии или в отсутствие 11Е1. Также оценивали лунки только с 11Е1. Каждое тестируемое условие оценивали шестикратно, и было проведено три независимых эксперимента. После инкубации в течение ночи клетки визуализировали с помощью камеры Axio Vision Camera (объектив х4).
HGF индуцировал значительную миграцию, приводящую к полному закрытию промежутка в течение одной ночи (фиг.9). Нерелевантный lgG1 9G4, представленный в качестве изотипического контроля, не имел никакого влияния на клеточную миграцию. Как и ожидалось, агонистический эффект наблюдался с 5D5 при добавлении его одного, но значительное ингибирование клеточной миграции наблюдалось с этим антителом в присутствии HGF в той части промежутка, который оставался открытым. Фрагмент Fab из 5D5 не имел никакого агонистического эффекта при добавлении его одного. Однако отсутствие активности этого фрагмента наблюдалось в присутствии HGF. Как отмечалось с изотипическим контролем 9G4, моноклональное антитело 11Е1 не имело агонистического эффекта при добавлении его одного, но вело себя как полный антагонист в присутствии HGF.
Пример 7: Анализ рассеяния
Клетки SK-HEP-1 сеяли с низкой плотностью (1×104 клеток/лунка) в 24-луночной плате в DMEM с 10% FCS и выращивали в течение 24 часов до одновременного добавления HGF (100 нг/мл) и тестируемых антител (10 мкг/мл). После 72 часов инкубации колонии фиксировали и окрашивали 0,2% кристаллическим фиолетовым в метаноле и визуально оценивали рассеяние. Каждое тестируемое условие оценивали трехкратно, и было проведено три независимых эксперимента.
Добавление HGF к клеткам SK-HEP-1 вызывало значительное клеточное рассеяние (фиг.10А и 10В). Антитело 9G4, представленное в качестве изотипического контроля, не имело эффекта ни в одиночку, ни в присутствии HGF. Как и ожидалось, антитело 5D5 проявляло значительный агонистический эффект в одиночку, а при добавлении HGF ингибирующий эффект 5D5 не наблюдался (фиг.10А). Агонистический эффект отсутствовал и у 11Е1 (фиг.10А), и у 224G11 (фиг.10В), добавленных в одиночку. Очень значительное ингибирующее действие этих антител было продемонстрировано в присутствии HGF (фиг.10А и 10В).
Пример 8: Трехмерный анализ тубулогенеза
Клетки SK-HEP-1 сеяли с плотностью 1×104 клеток/лунка в 24-луночную плату в DMEM с 10% FCS/Матригель (50/50) и инкубировали в течение 30 мин до одновременного добавления HGF (100 нг/мл) и тестируемых антител (10 мкг/мл). После семи дней инкубации клетки визуально оценивали на формирование трубочек. Каждое тестируемое условие оценивали трехкратно, и было проведено три независимых эксперимента.
Добавление HGF вызвало значительное образование трубочек в SK-HEP-1 (фиг.11). Антитело 9G4, представленное в качестве изотипического контроля, не имело эффекта ни в одиночку, ни в присутствии HGF. Как и ожидалось, антитело 5D5 демонстрировало значительный агонистический эффект в одиночку, а при добавлении HGF ингибирующий эффект 5D5 не наблюдался. Агонистический эффект отсутствовал и у 11Е1, 223С4 и 224G11, добавленных по одному, а полное ингибирующее действие 11Е1 и 223С4 было продемонстрировано в присутствии HGF. Частичное, но значительное ингибирование наблюдалось у моноклонального антитела 224G11.
Пример 9: Образование сфероида
Чтобы оценить способность анти-с-Met-антител ингибировать опухолевый рост in vitro на модели, близкой к ситуации in vivo, был создан сфероид из человеческих клеток глиобластомы U-87MG (АТСС №НТВ-14). Клетки, растущие в виде монослоя, отделяли с помощью трипсин-ЭДТА и ресуспендировали в полной клеточной культуральной среде (DMEM), дополненной 10% FBS. Сфероиды инициировали путем введения 625 клеток в отдельные лунки с круглым дном в 96-луночной плате в DMEM-10% FCS. Для предотвращения адгезии клеток к субстрату платы предварительно покрывали polyHEMA в 95% этаноле и высушивали на воздухе при комнатной температуре. Платы инкубировали в стандартных условиях культивирования клеток при 37°С, 5% CO2 в термостате с увлажнением. Очищенные моноклональные антитела (10 мкг/мл) добавляли через 3 и 7 дней культивирования сфероида. HGF (400 нг/мл) добавляли однократно через 4 дня культивирования. Сфероиды сохраняли в культуре по меньшей мере 10 дней. Затем рост сфероидов контролировали путем измерения площади сфероидов с помощью модуля автоизмерения программного обеспечения Axiovision. Площадь выражали в мкм2. Для каждого условия оценивали 8-16 сфероидов.
Фиг.12А и 12В демонстрируют, что в присутствии 10% FCS не наблюдалось стимуляции, когда HGF добавляли в полную среду. Как и ожидалось, изотипический контроль 9G4 не влиял на рост сфероида. 11Е1 и 223С4 значительно подавляли рост сфероида как в присутствии, так и в отсутствие HGF. Влияние фрагмента Fab 5D5 не наблюдалось.
Пример 10: Активность in vivo моноклональных антител анти-c-Met на модели с ксенотрансплантатом U87-MG
Атимических мышей в возрасте от шести до восьми недель поселяли в стерильных покрытых фильтром клетках, содержали в стерильных условиях и проводили над ними манипуляции в соответствии с французским и европейским руководствами. Для анализа in vivo была отобрана глиобластомная клеточная линия U87-MG, экспрессирующая c-Met и аутокрин для лиганда HGF. Мышам вводили подкожно 5×106 клеток. Затем, через шесть дней после имплантации клеток, опухоли измеряли (около 100 мм3), животных делили на группы по 6 мышей с сопоставимым размером опухоли и воздействовали на них два раза в неделю дозой 1 мг каждого тестируемого антитела. У мышей наблюдали темпы роста ксенотрансплантата и изменения веса тела. Объем опухоли рассчитывали по формуле: π(Pi)/6 × длина × ширина × высота.
Полученные результаты были обобщены в фиг.13 и продемонстрировали, что все протестированные антитела значительно ингибируют in vivo рост клеток U87-MG. Использование нейтрализующего анти-IGF-1R-антитела (lgG1) на панели А показывает, что наблюдаемое ингибирование in vivo специфично связано с модуляцией оси HGF-c-Met.
Пример 11: Активность in vivo моноклональных антител анти-c-Met на модели с ксенотрансплантатом NCI-H441
NCI-H441, полученная из папиллярной аденокарциномы легких, экспрессирует высокие уровни c-Met и демонстрирует постоянное фосфорилирование c-Met RTK.
Чтобы определить, может ли эта линия клеток экспрессировать высокие уровни c-Met и продуцировать HGF, были выполнены и количественные ОТ-ПЦР, и FACS или ИФА (Quantikine HGF, R&D Systems). Для количественных ОТ-ПЦР уровни экспрессии транскриптов общего HGF или c-Met в клеточных линиях оценивали путем количественной ПЦР с использованием стандартной методики TaqMan™. Уровни транскриптов HGF или c-Met нормировали по гену домашнего хозяйства рибосомального белка, большого, РО (RPL0), и результаты выражали в виде нормированных значений экспрессии (способ 2-ddCT).
Набор праймер/зонд для RPLO включал прямой праймер, 5'-gaaactctgcattctcgcttcctg-3' (SEQ ID №47); обратный праймер, 5'-aggactcgtttgtacccgttga-3' (SEQ ID №48); а также зонд, 5'-(FAM)-tgcagattggctacccaactgttgca-(TAMRA)-3' (SEQ ID №49). Набор праймер/зонд для HGF включал прямой праймер, 5'-aacaatgcctctggttcc-3' (SEQ ID №50); обратный праймер, 5'-cttgtagctgcgtcctttac-3' (SEQ ID №51); а также зонд, 5'-(FAM)-ccttcaatagcatgtcaagtggagtga-(TAMRA)-3' (SEQ ID №52). Набор праймер/зонд для с-Met включал прямой праймер, 5'-cattaaaggagacctcaccatagctaat-3' (SEQ ID №53); обратный праймер, 5'-cctgatcgagaaaccacaacct-3' (SEQ ID №54), а также зонд, 5'-(FAM)-catgaagcgaccctctgatgtccca-(TAMRA)-3' (SEQ ID №55). Протокол для термоциклера включал плавление при 50°С в течение 2 минут и 95°С в течение 10 минут, затем 40 циклов при 95°С в течение 15 секунд и 62°С в течение 1 минуты.
Было выявлено отсутствие мРНК HGF в NCI-H441 (фиг.14), и HGF не обнаруживался с помощью ИФА в супернатантах NCI-H441. В этих экспериментах U87-MG, глиобластомная клеточная линия, аутокринно действующая на HGF, была введена в качестве положительного контроля. ОТ-ПЦР-анализ показал значительный уровень мРНК HGF в U87-MG, в супернатанте клеток U87-MG был выявлен уровень 1,9 нг HGF/млн клеток. И количественный ОТ-ПЦР, и FACS-анализ (фиг.15А и 15 В) показали, что, как и ожидалось, клетки NCI-H441 значительно сверхэкспрессируют c-Met, и что эта экспрессия значительно выше, чем наблюдаемая в и87-МС-клетках. В этом эксперименте клеточную линию MCF-7 вводили в качестве отрицательного контроля. Взятые вместе, клетки NCI-H441 проявляли себя в качестве неаутокринной постоянно активированной клеточной линии, способной расти независимо от лиганда HGF, в которой лиганд-независимая димеризация c-Met произошла вследствие сверхэкспрессии рецептора.
Оценка влияния антител анти-c-Met на активность in vivo этой неаутокринной клеточной линии может дать некоторое представление об их способности вызывать c-Met-димеризацию.
Фиг.16 демонстрирует, что 224G11, 11Е1 и 227Н1 значительно ингибировали in vivo рост NCI-H441, подтверждая, что в дополнение к лиганд-зависимому ингибированию эти антитела, способные ингибировать димеризацию, также могут вызывать лиганд-независимое ингибирование c-Met. Как уже упоминалось выше в документе, по этому последнему качеству 224G11, 11Е1 и 227Н1 отличаются от антитела анти-c-Met 5D5 с одной ветвью (OA-5D5).
Пример 12: Способ гуманизации антитела 224G11 путем CDR-прививки
I - Гуманизация вариабельного домена легкой цепи
Сравнение нуклеотидной последовательности 224G11VL с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 224G11 VL сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGKV3-5*01 и IGKJ4*01 с идентичностью последовательности 99,31% для V-области и 94,28% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 224G11VL для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.17А для V-гена и 17В для J-гена.
Сравнение нуклеотидной последовательности 224G11VL с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 224G11VL. С этой целью нуклеотидную последовательность 224G11VL выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT. Для оптимизации отбора проводили выравнивания белковых последовательностей для поиска лучших гомологии.
Эти два взаимодополняющих способа привели к выявлению двух возможных человеческих последовательностей V, получающих мышиные 224G11 VL CDR. Нуклеотидное выравнивание дает человеческий зародышевый ген GKV3-11*01 с идентичностью последовательности 75,99%, тогда как белковое выравнивание дает человеческий зародышевый ген IGKV4-1*01 с идентичностью последовательности 67,30%. Следует отметить, что в обоих случаях два ближайших зародышевых гена и проанализированные последовательности демонстрировали различную аминокислотную длину CDR1 (10 аминокислот в 224G11 VL; 6 аминокислот в IGKV3-11*01; 12 аминокислот в IGKV4-1*01).
Для области J лучший показатель гомологии с человеком был впервые получен с человеческим IGKJ3*01, демонстрирующим идентичность последовательности 80%. Но более высокое количество последовательных идентичных нуклеотидов и лучшая аминокислотная сборка была найдена при выравнивании с человеческим зародышевым геном IGKJ4*02 (идентичность последовательности 77,14%). Таким образом, зародышевый ген IGKJ4*02 был отобран в качестве человеческой J-области, получающей мышиные 11Е1 VL CDR.
Выравнивания представлены в фиг.18А для V-области и 18 Вдля J-области.
Гуманизированная версия 224G11 VL
Учитывая возможность двух человеческих V-областей, получающих мышиные 224G11 VL CDR, будут описаны две гуманизированные версии 224G11 VL-домена. Первая соответствует первоначальному испытанию человеческого каркасного участка с более коротким CDR1 (IGKV3-11*01), вторая с более длинным CDR1 (IGKV4-1*01).
а) Основанная на IGKV3-11*01 гуманизированная версия 224G11 VL
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGKV3-11*01 и IGKJ4*02, a также CDR из мышиного 224G11 VL, с каркасным участком этих последовательностей зародышевых генов.
Как показано в фиг.19А, выделенные жирным шрифтом остатки в последовательности 224G11 VL соответствуют двадцати пяти аминокислотам, которые различаются в домене VL 224G11 и отобранном человеческом каркасном участке (человеческий FR, т.е. IGKV3-11*01 и IGKJ4*02).
Что касается нескольких критериев, таких как их известное участие в VH/VL интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что три из двадцати пяти различных остатков в конечном итоге мутируют. Этими тремя наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного М39 в человеческий L, H40 в А и R84 в G. Эти остатки первого ранга показаны в фиг.19А как выделенные жирным шрифтом остатки в последовательности 224G11 HZ1VL, где они остались мышиными.
Конечно, приведенные выше остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 15 (L/P), 49 (Р/А), 67 (L/R), 68 (E/A), 93 (P/S) и 99 (V/F), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении могут быть пересмотрены все шестнадцать других остатков третьего ранга среди двадцати пяти различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.19А представляет осуществленный основанный на IGKV3-11*01 гуманизированный 224G11 VL с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
б) Основанная на IGKV4-1*01 гуманизированная версия 224G11 VL
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGKV4-1*01 и IGKJ4*02, a также CDR из мышиного 224G11 VL, с каркасными участками этих последовательностей зародышевых генов.
Как показано в фиг.19В, выделенные жирным шрифтом остатки в последовательности 224G11 VL соответствуют двадцати двум аминокислотам, которые различаются в домене 224G11 VL и отобранном человеческом каркасном участке (человеческий FR, т.е. IGKV4-1*01 и IGKJ4*02).
Что касается нескольких критериев, таких как их известное участие в VH/VL интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что четыре из двадцати двух различных остатков в конечном итоге мутируют. Этими четырьмя наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного L4 в человеческий М, М39 в L, H40 в А и R84 в G. Эти остатки первого ранга показаны в фиг.19 В как выделенные жирным шрифтом остатки в последовательности 224G11 HZ2VL, где они остались мышиными.
Конечно, приведенные выше протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 25 (A/S), 66 (N/T), 67 (L/R) и 93 (P/S), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении могут быть пересмотрены все четырнадцать других остатков третьего ранга среди двадцати двух различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.19В представляет осуществленный основанный на IGKV4-1*01 гуманизированный 224G11 VL с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
II - Гуманизация вариабельного домена тяжелой цепи
Сравнение нуклеотидной последовательности 224G11 VH с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 224G11 VH сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGHV1-18*01, IGHD2-4*01 и IGHJ2*01 с идентичностью последовательности 92,70% для V-области, 75,00% для D-области и 89,36% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 224G11 VH для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.20А для V-гена, 20 В для D-гена и 20С для J-гена.
Сравнение нуклеотидной последовательности 224G11 VH с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 224G11 VH. С этой целью нуклеотидную последовательность 224G11 VH выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT. Для оптимизации отбора проводили выравнивания белковых последовательностей для поиска лучших гомологии.
Эти два взаимодополняющих способа привели к выявлению такой же человеческой последовательности V IGHV1-2*02, получающей мышиный 224G11 VH CDR с идентичностью последовательности 75,00% на нуклеотидном уровне и 64,30% на белковом уровне.
Следует отметить, что D-область строго принадлежит CDR3-области в VH-домене. Способ гуманизации основан на подходе «CDR-прививки». Анализ ближайших человеческих D-генов не используется в этой стратегии.
Поиск гомологии для J-областей привел к выявлению человеческого зародышевого гена IGHJ4*04 с идентичностью последовательности 78,72%.
Таким образом, человеческий зародышевый ген V IGHV1-2*02 и человеческий зародышевый ген J IGHJ4*01 были отобраны в качестве человеческих последовательностей, получающих мышиные 224G11 VH CDR.
Выравнивания представлены в фиг.21А для V-области и 21В для J-области.
Гуманизированная версия 224G11 VH
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGHV1-2*02 и IGHJ4*01, a также CDR из мышиного 224G11 VH, с каркасным участком этих последовательностей зародышевых генов.
Как показано в фиг.22, выделенные жирным шрифтом остатки в последовательности 224G11 VH соответствуют тридцати аминокислотам, которые различаются в домене VH 224G11 и отобранном человеческом каркасном участке (человеческий FR, т.е. IGHV1-2*02 и IGHJ4*01).
Что касается нескольких критериев, таких как их известное участие в VH/VH интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что четыре из тридцати различных остатков в конечном итоге мутируют. Этими четырьмя наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного D51 в человеческий Е, G55 в W, V80 в R и K82 в Т. Эти остатки первого ранга показаны в фиг.22 как выделенные жирным шрифтом остатки в последовательности 224G11 HZVH, где они остались мышиными.
Конечно, приведенные выше протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 25 (Т/А), 48 (E/Q), 49 (S/G), 53 (I/M), 76 (A/V), 78 (L/M) и 90 (D/E), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении могут быть пересмотрены все девятнадцать других остатков третьего ранга среди тридцати различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.22 представляет гуманизированный VH 224G11 с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
Пример 13: Способ гуманизации антитела 227Н1 путем CDR-прививки
I - Гуманизация вариабельного домена легкой цепи
Сравнение нуклеотидной последовательности 227Н1 VL с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 227Н1 VL сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGKV3-5*01 и IGKJ4*01 с идентичностью последовательности 96,90% для V-области и 97,29% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 227Н1 VL для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.23А для V-гена и 23В для J-гена.
Сравнение нуклеотидной последовательности 227Н1 VL с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 227Н1 VL. С этой целью нуклеотидную последовательность 227Н1 VL выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT. Для оптимизации отбора проводили выравнивания белковых последовательностей для поиска лучших гомологии.
Эти два взаимодополняющих способа привели к выявлению двух возможных человеческих V-последовательностей, получающих мышиные 227Н1 VL CDR. Нуклеотидное выравнивание дает человеческий зародышевый ген IGKV3-11*01 с идентичностью последовательности 74,91%, тогда как белковое выравнивание дает человеческий зародышевый ген IGKV4-1*01 с идентичностью последовательности 64,00%. Следует отметить, что в обоих случаях два ближайших зародышевых гена и проанализированные последовательности демонстрировали различную аминокислотную длину CDR1 (10 аминокислот в 227Н1 VL; 6 аминокислот в IGKV3-11*01; 12 аминокислот в IGKV4-1*01).
Для J-области лучший показатель гомологии с человеком был впервые получен с человеческим IGKJ3*01, демонстрирующим идентичность последовательности 78,38%. Но более высокое количество последовательных идентичных нуклеотидов и лучшая аминокислотная сборка была найдена при выравнивании с человеческим зародышевым геном IGKJ4*02 (идентичность последовательности 75,68%). Таким образом, зародышевый ген IGKJ4*02 был отобран в качестве человеческой J-области, получающей мышиные 227Н1 VL CDR.
Выравнивания представлены в фиг.24А для V-области и 24В для J-области.
Гуманизированная версия 227Н1 VL
Учитывая возможность двух человеческих V-областей, получающих мышиные 227Н1 VL CDR, будут описаны две гуманизированные версии 227Н1 VL-домена. Первая соответствует первоначальному испытанию человеческого каркасного участка с более коротким CDR1 (IGKV3-11*01), вторая с более длинным CDR1 (IGKV4-1*01).
а) Основанная на IGKV3-11*01 гуманизированная версия 227Н1 VL
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGKV3-11*01 и IGKJ4*02, a также CDR из мышиного 227Н1 VL с каркасным участком этих последовательностей зародышевых генов.
Как показано в фиг.25А, выделенные жирным шрифтом остатки в 227Н1 VL последовательности соответствуют двадцати шести аминокислотам, которые различаются в домене 227Н1 VL и отобранном человеческом каркасном участке (человеческий FR, т.е. IGKV3-11*01 и IGKJ4*02).
Что касается нескольких критериев, таких как их известное участие в VH/VL интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что три из двадцати шести различных остатков в конечном итоге мутируют. Этими тремя наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного 139 в человеческий L, H40 в А и R84 в G. Эти остатки первого ранга показаны в фиг.25А как выделенные жирным шрифтом остатки в последовательности 227Н1 HZ1VL, где они остались мышиными.
Конечно, приведенные выше протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 15 (L/P), 25 (V/A), 49 (P/A), 67 (L/R), 68 (E/A), 93 (P/S) и 99 (S/F), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении могут быть пересмотрены все шестнадцать других остатков третьего ранга среди двадцати пяти различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.25А представляет осуществленный основанный на IGKV3-11*01 гуманизированный 227Н1 VL с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
б) Основанная на IGKV4-1*01 гуманизированная версия 227Н1 VL
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGKV4-1*01 и IGKJ4*02, a также CDR из мышиного 227Н1 VL с каркасными участками этих последовательностей зародышевых генов.
Как показано в фиг.25В, выделенные жирным шрифтом остатки в 227Н1 VL последовательности соответствуют двадцати четырем аминокислотам, которые различаются в домене 227Н1 VL и отобранном человеческом каркасном участке (человеческий FR, т.е. IGKV4-1*01 и IGKJ4*02).
Что касается нескольких критериев, таких как их известное участие в VH/VL интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что четыре из двадцати четырех различных остатков в конечном итоге мутируют. Этими четырьмя наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного L4 в человеческий М, 139 в L, H40 в А и R84 в G. Эти остатки первого ранга показаны в фиг.25В как выделенные жирным шрифтом остатки в последовательности 227Н1 HZ2VL, где они остались мышиными.
Конечно, приведенные выше протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 25 (V/S), 66 (N/T), 67 (L/R) и 93 (P/S), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении могут быть пересмотрены все шестнадцать других остатков третьего ранга среди двадцати двух различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.25В представляет осуществленный основанный на IGKV4-1*01 гуманизированный 227Н1 VL с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
II - Гуманизация вариабельного домена тяжелой цепи
Сравнение нуклеотидной последовательности 227Н1 VH с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 227Н1 VH сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGHV1-18*01, IGHD1-1*02 и IGHJ2*01 с последовательностью, идентичной на 92,70% для V-области, 63,63% для D-области и 91,48% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 227Н1 VH для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.26А для V-гена, 26В для D-гена и 26С для J-гена.
Сравнение нуклеотидной последовательности 227Н1 VH с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 224G11 VH. С этой целью нуклеотидную последовательность 227Н1 VH выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT. Была выявлена человеческая V-последовательность IGHV1-2*02, получающая мышиные 224G11 VH CDR, с идентичностью последовательности 72,92%.
Следует отметить, что D-область строго принадлежит CDR3-области в VH-домене. Способ гуманизации основан на подходе «CDR-прививки». Анализ ближайших человеческих D-генов не используется в этой стратегии.
Поиск гомологии для J-областей привел к выявлению человеческого зародышевого гена IGHJ4*04 с идентичностью последовательности 78,72%.
Таким образом, человеческий зародышевый ген V IGHV1-2*02 и человеческий зародышевый ген J IGHJ4*01 были отобраны в качестве человеческих последовательностей, получающих мышиные 227Н1 VH CDR.
Выравнивания представлены в фиг.27А для V-области и 27В для J-области.
Для оптимизации отбора специалист в данной области также может провести выравнивания белковых последовательностей с целью облегчить себе отбор.
Гуманизированная версия 227Н1 VH
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGHV1-2*02 и IGHJ4*01, a также CDR из мышиного 227Н1 VH, с каркасным участком этих последовательностей зародышевых генов.
Как показано в фиг.28, выделенные жирным шрифтом остатки в 227Н1 VH последовательности соответствуют тридцати двум аминокислотам, которые различаются в домене VH 227Н1 и отобранном человеческом каркасном участке (человеческий FR, т.е. IGHV1-2*02 и IGHJ4*01).
Что касается нескольких критериев, таких как их известное участие в VH/VH интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что шесть из тридцати двух различных остатков в конечном итоге мутируют. Этими шестью наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного L39 в человеческий М, N40 в Н, L55 в W, T66 в N, V80 в R и K82 в Т. Эти остатки первого ранга показаны в фиг.28 как выделенные жирным шрифтом остатки в последовательности 227Н1 HZVH, где они остались мышиными.
Конечно, приведенные выше тестируемые остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 48 (K/Q), 49 (T/G), 53 (I/M), 76 (A/V) и 78 (L/M), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении может быть пересмотрен весь двадцать один другой остаток третьего ранга среди тридцати различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.28 представляет гуманизированный 227Н1 VH с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
Пример 14: Способ гуманизации антитела 223С4 путем CDR-прививки
I - Гуманизация вариабельного домена легкой цепи
Сравнение нуклеотидной последовательности 223С4 VL с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 223С4 VL сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGKV12-46*01 и IGKJ2*01 с идентичностью последовательности 99,64% для V-области и 94,59% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 223С4 VL для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.29А для V-гена и 29В для J-гена.
Сравнение нуклеотидной последовательности 223С4 VL с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 223С4 VL. С этой целью нуклеотидную последовательность 223С4 VL выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT.
Были выявлены человеческие зародышевые гены IGKV1-N1-1*01 и IGKJ2*01 с идентичностью последовательности 78,49% для V-области и 81,08% для J-области соответственно. Таким образом, зародышевые гены IGKV1-NL1*01 для V-области и IGKJ2*01 для J-области были отобраны в качестве человеческих последовательностей, получающих мышиные 223С4 VL CDR.
Выравнивания представлены в фиг.30А для V-области и 30В для J-области.
Для оптимизации отбора специалист в данной области также может провести выравнивания белковых последовательностей с целью облегчить себе отбор.
Гуманизированная версия 223С4 VL
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGKV1-NL1*01 и IGKJ2*01, a также CDR из мышиного 223С4 VL с каркасным участком этих последовательностей зародышевых генов.
На данном этапе способа можно разработать молекулярную модель 223С4 мышиных Fv-доменов и использовать в отборе мышиных остатков, которые будут сохранены из-за их роли в поддержании трехмерной структуры молекулы или в сайте связывания антигена и функционировании. В частности, было выявлено 9 остатков, которые в конечном итоге мутируют.
На первом этапе будут протестированы остатки, участвующие в CDR-заякоривании или структуре. Такими остатками являются остаток 66 (R/N) и остаток 68 (E/V).
На втором этапе проверяют остатки, открытые для растворителя, а также такие, которые могут вызывать иммуногенность. Этими остатками являются 49 (A/S), 51 (K/Q), 69 (S/D), 86 (D/Q) и 92 (S/N).
Затем, на третьем этапе, также могут мутировать остатки, участвующие в структуре/сворачивании вариабельного домена. Этими остатками являются остаток 46 (P/Q) и остаток 96 (P/S).
Конечно, приведенные выше остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 9 (S/A), 13 (A/V), 17 (D/E), 18 (R/T), 54 (L/V), 88 (T/S), 90 (Т/K), 100 (A/G) и 101 (T/S), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.31 представляет гуманизированный 223С4 VL с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
II - Гуманизация вариабельного домена тяжелой цепи
Сравнение нуклеотидной последовательности 223С4 VH с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 223С4 VH сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGHV1-18*01, IGHD6-3*01 и IGHJ4*01 с идентичностью последовательности 98,95% для V-области, 72,72% для D-области и 98,11% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 223С4 VH для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.32А для V-гена, 32В для D-гена и 32С для J-гена.
Сравнение нуклеотидной последовательности 223С4 VH с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 223С4 VH. С этой целью нуклеотидную последовательность 223С4 VH выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT.
Были выявлены зародышевые гены IGHV1-2*02, IGHD1-26*01 и IGHJ6*01 с идентичностью последовательности 76,38% для V-области, 75,00% для D-области и 77,41% для J-области соответственно. Таким образом, зародышевые гены IGHV1-2*02 для V-области и IGHJ6*01 для J-области были отобраны в качестве человеческих последовательностей, получающих мышиные 223С4 VH CDR.
Выравнивания представлены в фиг.33А для V-области, 33В для D-области и 33С для J-области.
Для оптимизации отбора специалист в данной области также может провести выравнивания белковых последовательностей с целью облегчить себе отбор.
Гуманизированная версия 223С4 VH
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGHV1-2*02 и IGHJ6*01, a также CDR из мышиного 223С4 VH, с каркасным участком этих последовательностей зародышевых генов.
На данном этапе способа можно разработать молекулярную модель 223С4 мышиных Fv-доменов и использовать в отборе мышиных остатков, которые будут сохранены из-за их роли в поддержании трехмерной структуры молекулы или в сайте связывании антигена и функции. В частности, было выявлено 14 остатков, которые в конечном итоге мутировали.
На первом этапе будут протестированы остатки, участвующие в CDR-заякоривании или структуре. Такими остатками являются остатки 40 (H/D), 45 (A/S), 55 (W/D), 66 (N/I) и 67 (Y/F).
На втором этапе проверяют остатки, открытые для растворителя, а также такие, которые могут вызывать иммуногенность. Этими остатками являются 1 (Q/E), 3 (Q/L), 5 (V/Q), 48 (Q/M) и 80 (R/V).
Затем, на третьем этапе, также могут мутировать остатки, участвующие в структуре/сворачивании вариабельного домена. Этими остатками являются остатки 9 (А/Р), 13 (K/V), 22 (S/P) и 46 (Р/Н).
Конечно, приведенные выше протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки, т.е. остатки 12 (V/L), 21 (V/I), 43 (R/K), 49 (G/S), 53 (M/I), 68 (A/N), 72 (Q/K), 75 (R/K), 76 (V/A), 78 (M/L), 82 (Т/K), 84 (I/S), 92 (S/R), 93 (R/S), 95 (R/T) и 97 (D/E), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.34 представляет гуманизированный 223С4 VH с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
Пример 15: Противоопухолевая активность мышиного моноклонального антитела 224G11 в одиночку или в сочетании с химиотерапевтическим агентом Навельбин® на принятой опухолевой модели с ксенотрансплантатом NCI-H441
Успешные химиотерапевтические подходы отчасти зависят от клеточного ответа на апоптотические индукторы и от баланса между про- и антиапоптотическими путями в клетке. Был подтверждено защитное влияние активированного c-Met на выживаемость клеток. Оно в основном связано с увеличенной экспрессией антиапоптотических белков Bcl-xl и Bcl-2 вследствие PI3-K-опосредованной сигнализации, что в свою очередь препятствует митохондриально-зависимому апоптозу (каспаза 9). Действительно, можно предположить, что система HGF/c-Met с ее отмеченным регуляторным влиянием на апоптотический процесс также может влиять на чувствительность раковых клеток к химиотерапевтическим препаратам. Эта гипотеза была проверена с Навельбином®, рыночным химиотерапевтическим агентом, используемым для лечения рака легкого (Aapro et al., Crit.Rev.Oncol.Hematol. 2001, 40:251-263; Curran et al., Drugs Aging. 2002, 19:695-697). Модель с ксенотрансплантатом NCI-H441 NSCLC (немелкоклеточный рак легкого) использовали потому что, как было описано ранее, эта клеточная линия чувствительна и к навельбину (Kraus-Berthier et al., Clin. Cancer Res., 2000; 6:297-304), и к терапии, нацеленной на c-Met (Zou Н. Т. et al., Cancer Res. 2007, 67: 4408-4417).
Кратко говоря, клетки NCI-H441 из АТСС культивировали стандартным способом в среде RPMI 1640, 10% FCS и 1% L-глутамин. За два дня до вживления трансплантата клетки разделяли, чтобы они находились в экспоненциальной фазе роста. Десять миллионов клеток NCI-H441 в PBS вживляли домовым голым мышам в возрасте до 7 недель. Через три дня после имплантации измеряли опухоли и делили животных на 4 группы по 6 мышей с сопоставимым размером опухоли. Мышам вводили внутрибрюшинно 224G11 в нагрузочной дозе 2 мг/мышь, а затем дважды в неделю в течение 43 дней 1 мг антитела/мышь. Моноклональное антитело 9G4 использовали в качестве изотипического контроля.
Навельбин® вводили путем внутрибрюшинных инъекций в дозе 8 мг/кг на 5, 12, 19 день после введения клеток. При комбинированной терапии с 224G11 и Навельбином® два соединения вводили отдельно. В этом опыте два соединения использовали в их оптимальной дозе. Объем опухоли измеряли дважды в неделю и рассчитывали по следующей формуле: Р/6 × длина × ширина × высота.
Фиг.35 демонстрирует, что 224G11 так же эффективен, как Навельбин®, когда используется в одиночку при терапии одним агентом. При сочетании обеих терапий наблюдалось значительное преимущество с полной регрессией опухоли, наблюдаемой у 3 из 6 мышей на 63 день.
Пример 16: Ингибиторы c-Met и ангиогенез
В дополнение к своей прямой роли в регуляции целого ряда функций опухолевых клеток, активация c-Met также была вовлечена в опухолевый ангиогенез. Эндотелиальные клетки экспрессируют c-Met, a HGF стимулирует рост, инвазию и подвижность эндотелиальных клеток (Nakamura Y. et al., Biochem. Biophys. Res., Commun. 1995, 215:483-488; Bussolino F. et al., J. Cell Biol. 1992, 119:629-641). Была продемонстрирована согласованная регуляция роста, инвазии и подвижности сосудистых эндотелиальных клеток HGF/c-Met, приведшая к формированию in vitro трехмерных капиллярных эндотелиальных трубочек (Rosen Е.М. et al., Supplementum to Experientia 1991, 59:76-88).
Для определения потенциального вмешательства моноклональных антител анти-c-Met в HGF-индуцированный ангиогенез проводили две группы экспериментов, в том числе i) оценку влияния моноклональных антител на пролиферацию HUVEC (человеческих клеток эндотелия пуповинной вены) и ii) оценку влияния моноклональных антител на формирование трубочек в HUVEC.
Для экспериментов с пролиферацией 7500 HUVEC сеяли в каждую лунку 96-луночной платы, заранее покрытой ламинином. Клетки выращивали в течение 24 часов на среде для анализа ЕМВ-2, дополненной 0,5% FBS и гепарином. Затем тестируемые монокпональные антитела (от 0,15 до 40 мкг/мл) добавляли за 1 час до добавления 20 нг/мл HGF. Через дополнительные 24 часа на клетки воздействовали 0,5 мкКи [3H]-тимидина. Количество включенного [3Н]-тимидина подсчитывали с помощью жидкого сцинтилляционного счета. В этом эксперименте моноклональное антитело 9G4 представляет собой нерелевантное антитело, использованное в качестве lgG1 изотипического контроля.
Результаты, показанные на фиг.36, демонстрируют, что, как и ожидалось, HGF является мощным индуктором клеточного роста HUVEC. Антитела, оцениваемые в отсутствие HGF, не проявляли агонистического пролиферативного влияния на HUVEC ни в одной из испытанных доз. В присутствии HGF резкое дозозависимое ингибирование наблюдалось для обоих моноклональных антител 11Е1 и 224G11.
Для оценки формирования трубочек в HUVEC 25000 клеток, инкубированных в течение 30 минут с тестируемыми антителами, сеяли в 48-луночные платы, покрытые Матригелем. Затем добавляли 50 нг/мл HGF и инкубировали платы при 37°С. Затем собирали среду и добавляли 5 мкМ CMFDA (5-хлорметилфлуоресцеина диацетат) за 15 мин до микроскопического исследования.
Результаты, показанные в фиг.37, демонстрируют, что, как и ожидалось, HGF вызывал значительное образование трубочек. Антитело 9G4, представленное в качестве lgG1 изотипического контроля, не имело какого-либо влияния на HGF-индуцированное формирование трубочек, тогда как 11Е1 и 224G11 резко ингибировали формирование трубочек.
Пример 17: Способ гуманизации антитела 11Е1 путем CDR-прививки
I - Гуманизация вариабельного домена легкой цепи
Сравнение нуклеотидной последовательности 11Е1 VL с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 11Е1 VL сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGKV4-79*01 и IGKJ4*01 с идентичностью последовательности 98,58% для V-области и 97,22% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 11Е1 VL для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.38А для V-гена и 38В для J-гена.
Сравнение нуклеотидной последовательности 11Е1 VL с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 11Е1 VL. С этой целью нуклеотидную последовательность 11Е1 VL выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT.
Были выявлены человеческие IGKV3-7*02 и IGKV3D-7*01 с идентичностью последовательности для обоих зародышевых генов 69,86% для V-области. Человеческий зародышевый ген IGKV3-7*02 известен в базе данных как «ORF», что означает, что эта последовательность была найдена в человеческом геноме, но может представлять некоторые рекомбинантные проблемы, приводящие к нефункциональным природным антителам, полученным из IGKV3-7*02. Таким образом, зародышевый ген IGKV3D-7*02 был отобран в качестве человеческой V-области, получающей мышиные 11Е1 VL CDR.
Для J-области лучший показатель гомологии с человеком был впервые получен с человеческим IGKJ3*01, демонстрирующим идентичность последовательности 78,38%. Но более высокое количество последовательных идентичных нуклеотидов и лучшая аминокислотная сборка была найдена при выравнивании с человеческим зародышевым геном IGKJ4*02 (идентичность последовательности 75,68%). Таким образом, зародышевый ген IGKJ4*02 был отобран в качестве человеческой J-области, получающей мышиные 11Е1 VL CDR.
Выравнивания представлены в фиг.39А для V-области и 39В для J-области.
Для оптимизации отбора специалист в данной области также может провести выравнивания белковых последовательностей с целью облегчить себе отбор.
Гуманизированная версия 11Е1 VL
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGKV3D-7*01 и IGKJ4*02, a также CDR из мышиного 11Е1 VL, с каркасным участком этих последовательностей зародышевых генов.
Как показано в фиг.40, выделенные жирным шрифтом остатки в последовательности 11Е1 VL соответствуют тридцати аминокислотам, которые различаются в домене 11Е1 VL и отобранном человеческом каркасном участке (человеческий FR, т.е. IGKV3D-7*01 и IGKJ4*02).
Что касается нескольких критериев, таких как их известное участие в VH/VL интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что четыре из тридцати различных остатков в конечном итоге мутируют. Этими четырьмя наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного L4 в человеческий М, Y40 в S, Y87 в F и Т96 в Р. Эти остатки первого ранга показаны в фиг.40 как выделенные жирным шрифтом остатки в последовательности 11Е1 HZVL, где они остались мышиными.
Конечно, приведенные выше протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 24 (S/R), 53 (W/L), 66 (I/T), 67 (L/R), 86 (S/D), 95 (Q/E), 99 (A/F) или 121 (E/D), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении могут быть пересмотрены все восемнадцать других остатков третьего ранга среди тридцати различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.40 представляет осуществленный гуманизированный 11Е1 VL с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
II - Гуманизация вариабельного домена тяжелой цепи
Сравнение нуклеотидной последовательности 11Е1 VH с мышиными зародышевыми генами
В качестве предварительного этапа нуклеотидную последовательность 11Е1 VH сравнивали с участком последовательностей мышиных зародышевых генов из базы данных IMGT (http://imgt.cines.fr).
Были определены мышиные зародышевые гены IGHV1-7*01, IGHD4-1*01 и IGHJ3*01 с идентичностью последовательности 94,10% для V-области, 66,67% для D-области и 100% для J-области соответственно. Что касается полученной идентичности, было решено непосредственно использовать последовательности 11Е1 VH для поиска человеческих гомологии.
Эти выравнивания представлены в фиг.41А для V-гена, 41В для D-гена и 41С для J-гена.
Сравнение нуклеотидной последовательности 11Е1 VH с человеческими зародышевыми генами
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с 11Е1 VH. С этой целью нуклеотидную последовательность 11Е1 VH выравнивали с участком последовательностей человеческих зародышевых генов из базы данных IMGT. Для оптимизации отбора проводили выравнивания белковых последовательностей для поиска лучших гомологии.
Эти два взаимодополняющих способа привели к выявлению двух возможных человеческих V-последовательностей, получающих мышиные 11Е1 VH CDR. Нуклеотидное выравнивание дает человеческий зародышевый ген IGKV1-2*02 с идентичностью последовательности 75,69%, тогда как белковое выравнивание дает человеческий зародышевый ген IGHV1-46*01 с идентичностью последовательности 71,10%.
Следует отметить, что D-область строго принадлежит CDR3-области в VH-домене. Способ гуманизации основан на подходе «CDR-прививки». Анализ ближайших человеческих D-генов не используется в этой стратегии.
Поиск гомологии для J-областей привел к выявлению человеческого зародышевого гена IGHJ4*03 с идентичностью последовательности 80,85%.
Таким образом, при поиске всех сходств и выравниваний последовательностей человеческий зародышевый ген V IGHV1-46*01 и человеческий зародышевый ген J IGHJ4*03 были отобраны в качестве человеческих последовательностей, получающих мышиные 11Е1 VH CDR.
Выравнивания представлены в фиг.42А для V-области и 42В для J-области.
Гуманизированная версия 11Е1 VH
Следующие этапы способа гуманизации заключаются в связывании отобранных последовательностей зародышевых генов IGHV1-46*01 и IGHJ4*03, a также CDR из мышиного 11Е1 VH с каркасным участком этих последовательностей зародышевых генов.
Как показано в фиг.43, выделенные жирным шрифтом остатки в 11Е1 VH-последовательности соответствуют двадцати шести аминокислотам, которые различаются в домене VH 11Е1 и отобранном человеческом каркасном участке (человеческий FR, т.е. IGHV1-46*01 и IGHJ4*03).
Что касается нескольких критериев, таких как их известное участие в VH/VH интерфазе, в связывании антигена или в CDR-структуре, изменения классов аминокислот между мышиным и человеческим остатками, локализация остатка в 3D-структуре вариабельного домена, то было выявлено, что пять из двадцати шести различных остатков в конечном итоге мутируют. Этими пятью наиболее важными определенными остатками и мутациями в их человеческих аналогах являются мутация мышиного N40 в человеческий Н, Y55 в I, D66 в S, A80 в R и K82 в Т. Эти остатки первого ранга показаны в фиг.43 как выделенные жирным шрифтом остатки в последовательности 11Е1 HZVH, где они остались мышиными.
Конечно, приведенные выше протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций.
С помощью молекулярной модели могут быть выявлены другие мутации. Можно упомянуть следующие остатки второго ранга, т.е. остатки 53 (I/М), 71 (L/F), 76 (A/V), 78 (L/M) и 87 (A/V), в которых также могут быть предусмотрены мутации в другом предпочтительном воплощении.
Конечно, приведенные выше в итоге протестированные остатки не являются ограничивающими, но должны рассматриваться в качестве предпочтительных мутаций. В другом предпочтительном воплощении могут быть пересмотрены все шестнадцать других остатков третьего ранга среди двадцати шести различных аминокислот.
Все вышеупомянутые мутации будут протестированы в индивидуальном порядке или в соответствии с различными сочетаниями.
Фиг.43 представляет гуманизированный VH 11E1 с вышеупомянутыми мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
Пример 18: Влияние очищенных моноклональных антител на c-Met-фосфорилирование
В примере 3 влияние моноклональных антител анти-c-Met на фосфорилирование оценивали с дозированными супернатантами от каждой гибридомы, которую нужно было оценить. Испытание проводили снова с очищенными моноклональными антителами 11E1 и 224G11, которые оценивали или с конечной концентрацией 30 мкг/мл (200 нМ), или в дозовом диапазоне от 0,0015 до 30 мкг/мл (0,01-200 нМ) с целью определения IC50 каждого антитела. Используемый протокол был таким же, как описанный в примере 3.
Результаты трех независимых экспериментов представлены в фиг.44 и демонстрируют, что один раз очищенные 11E1 и 224G11 не имеют агонистического эффекта при добавлении их к клеткам А549 в одиночку, а в присутствии HGF имеют антагонистический эффект в 87 и 75% соответственно. Как и ожидалось, моноклональное антитело 5D5, введенное в качестве агонистического положительного контроля, имеет значительное (58%) агонистическое влияние при добавлении его в одиночку и лишь умеренное антагонистическое влияние (39%) в присутствии HGF. Что касается расчетов EC50, то и 11E1, и 224G11 имели наномолярные показатели IC50.
Пример 19: Комбинирование in vivo 224G11 и Навельбина® на модели с ксенотрансплантатом NCI-H441
Клетки NCI-H441 из АТСС культивировали стандартным способом в среде RPMI 1640, 10% FCS, 1% L-глутамин. За два дня до вживления трансплантата клетки разделяли, чтобы они находились в экспоненциальной фазе роста. Десять миллионов клеток NCI-H441 в PBS вживляли атимическим голым мышам. Через пять дней после имплантации измеряли опухоли и делили животных на группы по 6 мышей с сопоставимым размером опухоли. Мышам вводили внутрибрюшинно или моноклональное антитело 224G11 в нагрузочной дозе 2 мг/мышь и затем дважды в неделю до 38 дня 1 мг антитела/мышь, или Навельбин® 3 раза (в 5, 12 и 19 день) по 8 мг/кг. Также включили третью группу, в которой проводили комбинированную терапию. Навельбин® вводили путем внутрибрюшинных инъекций. Объем опухоли измеряли дважды в неделю и рассчитывали по следующей формуле: π/6 × длина × ширина × высота, а вес животного контролировали каждый день в течение периода введения Навельбина®. Статистический анализ проводили для каждого периода измерения с использованием либо t-теста, либо теста Манна-Уитни. В этом эксперименте средний объем опухоли в группе, обработанной единственным способом воздействия, уменьшался на 72%, 76% и 99,8% для 224G11, Навельбина® и Навельбин® + 224G11 соответственно на 41 день после первой инъекции. На 41 день комбинированной терапии значительно уменьшился опухолевый рост по сравнению с обработкой одним агентом (р≤0,041 по сравнению с одним Навельбином® и p≤0,002 по сравнению с одним 224G11 на 41 день), 4 из 6 мышей в группе комбинированной терапии не имели опухолей. Результаты представлены в фиг.45.
Эти результаты были подтверждены через 50 дней после окончания лечения (день 88), когда 66% мышей, получавших комбинированное лечение, оставались без опухолей.
Пример 20: Комбинирование in vivo 224G11 и доксорубицина на модели с ксенотрансплантатом NCI-H441
Клетки NCI-H441 из АТСС культивировали стандартным способом в среде RPMI 1640, 10% FCS, 1% L-глутамин. За два дня до вживления трансплантата клетки разделяли, чтобы они находились в экспоненциальной фазе роста. Десять миллионов клеток NCI-H441 в PBS вживляли атимическим голым мышам. Через пять дней после имплантации измеряли опухоли и делили животных на группы по 6 мышей с сопоставимым размером опухоли. Мышам вводили внутрибрюшинно или моноклональное антитело 224G11 в нагрузочной дозе 2 мг/мышь и затем дважды в неделю 1 мг антитела/мышь, или доксорубицин 4 раза (в 5, 12, 19 и 26 день) по 5 мг/кг. Также включили третью группу, в которой проводили комбинированную терапию. Доксорубицин вводили путем внутривенных инъекций. Объем опухоли измеряли дважды в неделю и рассчитывали по следующей формуле: π/6 × длина × ширина × высота, а вес животного контролировали каждый день в течение периода введения доксорубицина. Статистический анализ проводили для каждого периода измерения с использованием либо t-теста, либо теста Манна-Уитни. Обе терапии одним агентом и комбинированное лечение демонстрировали значительную противоопухолевую активность по сравнению с контрольной группой (p≤0,002 с 11 по 39 день). Результаты представлены в фиг.46.
Комбинированное лечение также демонстрировало значительную противоопухолевую активность по сравнению с лечением одним агентом с 11 по 39 день, показывая, что выгодно сочетать доксорубицин с лечением анти-c-Met.
Пример 21: Комбинирование in vivo 224G11 и доцетаксела на модели с ксенотрансплантатом NCI-H441
Клетки NCI-H441 из АТСС культивировали стандартным способом в среде RPMI 1640, 10% FCS, 1% L-глутамин. За два дня до вживления трансплантата клетки разделяли, чтобы они находились в экспоненциальной фазе роста. Девять миллионов клеток NCI-H441 вживляли атимическим голым мышам. Через пять дней после имплантации измеряли опухоли и делили животных на группы по 6 мышей с сопоставимым размером опухоли. Мышам вводили внутрибрюшинно или моноклональное антитело 224G11 в нагрузочной дозе 2 мг/мышь и затем дважды в неделю 1 мг антитела/мышь, или доцетаксел 4 раза (в 5, 12, 19 и 26 день) по 7,5 мг/кг. Также включили третью группу, в которой проводили комбинированное лечение. Доцетаксел вводили путем внутрибрюшинных инъекций. Объем опухоли измеряли дважды в неделю и рассчитывали по следующей формуле; π/6 × длина × ширина × высота, а вес животного контролировали каждый день в течение периода введения доцетаксела. Статистический анализ проводили для каждого периода измерения с использованием либо t-теста, либо теста Манна-Уитни. Обе терапии одним агентом и комбинированное лечение демонстрировали значительную противоопухолевую активность по сравнению с контрольной группой (p≤0,002 с 11 по 35 день). Результаты представлены в фиг.47.
Комбинированное лечение также демонстрировало значительную противоопухолевую активность по сравнению с лечением одним агентом с 18 по 35 день, показывая, что выгодно сочетать доцетаксел с лечением анти-c-Met.
Пример 22: Комбинирование in vivo 224G11 и темозоломида на модели с ксенотрансплантатом U87-MG
Клетки U87-MG из АТСС культивировали стандартным способом в среде RPMI 1640, 10% FCS, 1% L-глутамин. За два дня до вживления трансплантата клетки разделяли, чтобы они находились в экспоненциальной фазе роста. Пять миллионов клеток U87-MG вживляли атимическим голым мышам. Через девятнадцать дней после имплантации измеряли опухоли и делили животных на группы по 6 мышей с сопоставимым размером опухоли. Мышам вводили внутрибрюшинно или моноклональное антитело 224G11 в нагрузочной дозе 2 мг/мышь и затем дважды в неделю 1 мг антитела/мышь, или темозоломид 3 раза (в 19, 26 и 33 день) по 5 мг/кг. Также включили третью группу, в которой проводили комбинированное лечение. Темозоломид вводили путем внутрибрюшинных инъекций. Объем опухоли измеряли дважды в неделю и рассчитывали по следующей формуле: π/6 × длина × ширина × высота, а вес животного контролировали каждый день в течение периода введения темозоломида. Статистический анализ проводили для каждого периода измерения с использованием либо t-теста, либо теста Манна-Уитни. Обе терапии одним агентом и комбинированное лечение демонстрировали значительную противоопухолевую активность по сравнению с контрольной группой (p≤0,002 с 22 по 32 день (где контрольные мыши были умерщвлены по этическим причинам)). Результаты представлены в фиг.48.
Комбинированное лечение также демонстрирует значительную противоопухолевую активность по сравнению с лечением одним агентом темозоломидом (p≤0,002 с 22 по 43 день (где контрольные мыши были умерщвлены по этическим причинам)) для темозоломида и с 29 по 53 день (последний день лечения) для 224G11. Взятые вместе, эти данные показывают, что выгодно сочетать темозоломид с лечением анти-c-Met.
Пример 23: Образование сфероида
Как уже описывалось в примере 9 для других моноклональных антител, мы оценивали способность моноклонального антитела 224G11 ингибировать опухолевый рост in vitro на сфероидной модели U87-MG. С этой целью клетки U87-MG, растущие в виде монослоя, отделяли с помощью трипсин-ЭДТА и ресуспендировали в полной клеточной культуральной среде. Сфероиды инициировали путем введения по 625 клеток в отдельные лунки с круглым дном в 96-луночной плате в DMEM-2,5% PCS. Для предотвращения адгезии клеток к субстрату платы предварительно покрывали polyHEMA в 95% этаноле и высушивали на воздухе при комнатной температуре. Платы инкубировали в стандартных условиях культивирования клеток при 37°С, 5% CO2 в термостате с увлажнением. Очищенные моноклональные антитела (10 мкг/мл) добавляли через 4 и 10 дней культивирования сфероида. Сфероиды сохраняли в культуре в течение 17 дней. Затем рост сфероида контролировали путем измерения площади сфероида с помощью модуля автоизмерения программного обеспечения Axiovision. Площадь выражали в мкм2. Для каждого условия оценивали 8-16 сфероидов. Размер сфероидов оценивали до добавления антител, через 10 дней культивирования и через 17 дней культивирования.
В этих условиях были получены гомогенные сфероиды, и статистической разницы до добавления антител не наблюдалось (фиг.49А).
Как показано в фиг.49B-49D, изотипический контроль 9G4 не влиял на рост сфероидов через 10 или 17 дней культивирования. В то время как добавление 5D5 не имело большого влияния на размер сфероидов, добавление либо 224G11, либо 11Е1 заметно ингибировало рост опухоли.
Пример 24: Активность in vitro химерных и гуманизированных форм 224G11 в фосфо-c-Met анализе
Для того, чтобы сравнить эффективность in vitro мышиной, химерной и гуманизированной форм в функциональном анализе, культуральные супернатанты, полученные из 224G11-гибридомы и HEK293-трансфицированных клеток, дозировали и тестировали, как описано в примере 3. Данные, приведенные в фиг.50, показали ожидаемые результаты для неочищенного мышиного антитела, как уже было описано в фиг.6В. И химерные, и гуманизированные неочищенные антитела демонстрировали сопоставимую активность при добавлении в одиночку (фиг.50А) или при инкубации в присутствии HGF (фиг.50В).
Пример 25: Определение констант аффинности (KD) анти-с-Met-антител с помощью анализа Biacore
Связывающую аффинность очищенных антител 11Е1 и 224G11 исследовали с помощью BIAcore X с использованием рекомбинантного c-Met-внеклеточного домена (ECD), слившегося с человеческим Fc-доменом lgG1 (R&D Systems) в качестве антигена (молекулярный вес = 129 кДа). Так как и гибридные белки c-Met-Fc, и антитела являются двухвалентными, то Fab-фрагменты моноклональных антител 11Е1 и 224G11 (молекулярный вес = 50 кДа) были получены путем расщепления папаином, очищенным и используемым в этом анализе, чтобы избежать влияния параметра авидитета (степени сродства антитела к антигену). Для анализа сенсорный чип СМ5 покрыли антителами захвата анти-Tag-гистидин. Рабочим буфером был HBS-EP, скорость потока составляла 30 мкл/мин, а тест проводили при 25°С. Растворимый антиген c-Met (ECD_M1)2-Fc-(HHHHHH)2 был захвачен сенсорным чипом (около 270 RU), а тестируемые антитела использовали в растворе в качестве анализируемых веществ. Сенсорный чип регенерировали с помощью буфера глицин, HCl рН 1,5, в обоих измерительных участках в течение полминуты.
Фиг.51 иллюстрирует принцип этого анализа. Полученные кинетические параметры приведены в таблице 4. Они указывают, что и 11Е1, и 224G11 анти-с-Met-антитела связывают рекомбинантный гибридный белок c-Met-Fc с сопоставимой аффинностью в размере примерно 40 пкМ.
|
Пример 26: Влияние in vivo 224G11 на клетки MDA-MB-231, имплантированные совместно с MRC5-клетками в качестве источника человеческого HGF, у атимических голых мышей
Клетки MDA-MB-213 и MRC5 из АТСС культивировали в среде DMEM, 10% FCS, 1% L-глутамин. За два дня до вживления трансплантата клетки разделяли, чтобы они находились в экспоненциальной фазе роста. Пять миллионов клеток MDA-MB-213 и пятьсот тысяч клеток MRC5 совместно вводили путем подкожной инъекции атимическим голым мышам. Через двенадцать дней после имплантации измеряли опухоли и делили животных на группы по 6 мышей с сопоставимым размером опухоли. Мышам вводили внутрибрюшинно моноклональное антитело 224G11 в нагрузочной дозе 2 мг/мышь и затем дважды в неделю 1 мг антитела/мышь. Объем опухоли измеряли дважды в неделю и рассчитывали по следующей формуле: π/6 × длина × ширина × высота.
Результаты, описанные в фиг.52, демонстрировали значительное различие медианы роста опухолей у мышей с введением 224G11 по сравнению с медианой у контрольной группы.
Пример 27: Дополнительные элементы, касающиеся гуманизации антител 227Н1, 11Е1 и 224G11
Общая процедура
Гуманизацию анти-с-Met-антител выполняли независимо для каждой цепи и последовательно в отношении анализируемых аминокислот в каждом вариабельном домене. Способ гуманизации оценивали в первом опыте в основанном на ИФА анализе связывания для рекомбинантного Fc-c-Met; связывающую активность гуманизированных антител сравнивали с активностью рекомбинантного химерного антитела. Во втором опыте гуманизированные антитела анти-c-Met оценивали по их способности вытеснять Fc-c-Met, связанный с пластиком, покрытым рекомбинантным HGF; этот конкурентный анализ позволяет непосредственно сравнить мышиные, химерные и гуманизированные версии анти-c-Met-антител.
Фиг.53 и 54 иллюстрируют типичную анти-с-Met-связывающую активность мышиных моноклональных антител 227Н1, 11Е1 и 224G11.
Фиг.53 показывает прямую анти-с-Met-связывающую активность обнаруженных очищенных мышиных антител. В этом анализе мышиные моноклональные анти-с-Met-антитела показывают различную, но все еще дозозависимую анти-с-Met-связывающую активность.
Фиг.54 показывает HGF-c-Met-связывающую конкурентную активность очищенных мышиных антител. Конкурентный анализ показывает достоверные различия между этими анти-c-Met моноклональными антителами с умеренной, неполной, но надежной конкурентой активностью моноклонального антитела 11Е1, в то время как мышиные 224G11 и 227Н1 показывают аналогичный характер конкурентной активности со 100% от максимума вытеснения HGF из места связывания при высокой концентрации антител. Моноклональное антитело 224G11 показывает лучшие значения IC50.
Следует отметить, что прямая связывающая активность мышиных антител не отражает их внутренних HGF-связывающих конкурентных свойств.
Эти два анализа были использованы для характеристики рекомбинантных химерных и гуманизированных версий мышиных анти-с-Met-антител. Для этой цели, коротко говоря, анти-с-Met-вариабельные домены, либо мышиные, либо гуманизированные, клонировали в ряд векторов экспрессии pCONplus от компании LONZA, и рекомбинантные антитела, полученные из lgG1/k, экспрессировались в клетках СНО. Экспрессированные культуральные супернатанты концентрировали и значительно диализировали против PBS, а затем дозировали по концентрации экспрессированных антител и непосредственно использовали для оценки соответствующей анти-с-Met-связывающей активности. И прямой связывающий, и HGF-конкурентный анализы проводили, чтобы лучше охарактеризовать рекомбинантные химерные или гуманизированные версии.
Пример 27-1: Гуманизация вариабельного домена тяжелой цепи 227Н1
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с мышиной последовательностью 227Н1 VH. Таким образом, с помощью базы данных IMGT были отобраны человеческий зародышевый ген V IGHV1-2*02 и человеческий зародышевый ген J IGHJ4*01 в качестве человеческих последовательностей, получающих мышиные 227Н1 VH CDR.
Фиг.55 представляет аминокислотное выравнивание мышиного домена 227Н1 VH с отобранным человеческим каркасным участком. В ряду «человеческий каркасный участок» отображены только те аминокислоты, которые отличаются от 227Н1 мышиного VH-домена. Ряды HZ3VH, HZ2VH и HZ1VH соответствуют осуществленным гуманизированным версиям домена 227Н1 VH с вышеупомянутыми (ряд «изменен в») мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
В первой серии экспериметов мы создали и проанализировали анти-c-Met-связывающую активность трех первых гуманизированных версий VH-домена мышиного 227Н1, экспрессированного в сочетании с химерной легкой цепью 227Н1. Результаты, полученные в прямом анализе анти-с-Met-связывания, показаны в фиг.56. В этом эксперименте не было отмечено никаких различий в связывающей способности тестированных полученных из 227Н1 химерных или частично гуманизированных рекомбинантных антител. На данный момент проанализированы 26 из 32 аминокислот, которые различаются в мышином домене VH 227Н1 и отобранном человеческом каркасном участке, и найдено, что они не имеют отношения к анти-с-Met-связывающей активности гуманизированного VH-домена 227Н1, когда сочетаются с химерной легкой цепью.
В сочетании с анализом сайт-направленного мутагенеза последних шести мышиных остатков в HZ1VH-гуманизированной версии домена VH 227Н1, мы создали оригинальную HZ4VH «полностью IMGT-гуманизированную» версию и испытали ее анти-c-Met связывающие свойства. Результаты представлены в фиг.57 для прямого связывающего анализа и в фиг.58 для HGF-связывающего конкурентного анализа. Следует отметить, что и химерная, и гуманизированная рекомбинантные версии 227Н1 демонстрируют лучшую конкурентную активность по сравнению с родительским мышиным антителом.
Тем не менее, с учетом экспериментальных данных, полученных в отношении анти-с-Met-связывающих свойств «полностью IMGT» гуманизированного 227Н1 VH-домена, отобрали полученную аминокислотную последовательность, изображенную в фиг.59, и затем провели биоинформативный анализ для оценки уровня «человечности» так называемого 227Н1-HZ VH гуманизированного вариабельного домена.
Для этого выполняли простое сравнение каркасных последовательностей человеческой базы данных с использованием IMGT-инструментов. С учетом уровня гуманизации, которого мы достигли в ходе этого способа, из 89 проанализированных аминокислот, соответствующих каркасным остаткам, 89 были признаны достоверно человеческого происхождения. Только остатки из CDR могут быть признаны отличающимися, но если такие существуют, то они отличаются от соответствующих человеческих зародышевых генов, и, очевидно, находятся в гипервариабельных позициях. На основании системы нумерации IMGT и инструментов анализа гомологии, мы сначала полностью гуманизировали вариабельный домен антитела мышиного происхождения.
Пример 27-2: Гуманизация моноклинального антитела 11Е1
I - Гуманизация вариабельного домена тяжелой цепи 11Е1
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с мышиной последовательностью VH 11Е1. Таким образом, с помощью базы данных IMGT были отобраны человеческий зародышевый ген V IGHV1-46*01 и человеческий зародышевый ген J IGHJ4*03 в качестве человеческих последовательностей, получающих мышиные 11Е1 VH CDR.
Фиг.60 представляет аминокислотное выравнивание мышиного 11Е1 VH-домена с отобранным человеческим каркасным участком. В ряду «человеческий каркасный участок» отображены только те аминокислоты, которые отличаются от мышиного 11Е1 VH-домена. Ряды HZ VH3, HZ VH2 и HZ VH1 соответствуют осуществленным гуманизированным версиям 11Е1 VH-домена с вышеупомянутыми (ряд «измененные в») мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
В первой серии экспериметов мы создали и проанализировали анти-c-Met-связывающую активность трех первых гуманизированных версий мышиного 11Е1 VH-домена, экспрессированного в сочетании с химерной легкой цепью 11Е1. Результаты, полученные в анализе прямого c-Met-связывания, показаны в фиг.61. В этом эксперименте была отмечена сходная связывающая способность тестированных полученных из 11Е1 химерных или частично гуманизированных рекомбинантных антител. На данный момент проанализировали 19 из 24 аминокислот, признанных различающимися в мышином 11Е1 VH-домене и отобранном человеческом каркасном участке, и выявили, что они не имеют отношения к анти-с-Ме1-связывающей активности гуманизированного VH-домена 11Е1, когда сочетаются с химерной легкой цепью.
II - Гуманизация вариабельного домена легкой цепи 11Е1
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с мышиной последовательностью 11Е1 VL. Таким образом, с помощью базы данных IMGT были отобраны человеческий зародышевый ген V IGKV3D-7*01 и человеческий зародышевый ген J IGKJ4*01 в качестве человеческих последовательностей, получающих мышиные 11Е1 VL CDR.
Фиг.62 представляет собой аминокислотное выравнивание мышиного 11Е1 VL-домена с отобранным человеческим каркасным участком. В ряду «человеческий каркасный участок» были отображены только те аминокислоты, которые отличаются от мышиного 11Е1 VL-домена. Ряды HZ VL3, HZ VL2 и HZ VL1 соответствуют осуществленным гуманизированмым версиям 11Е1 VL-домена с вышеупомянутыми (ряд «измененные в») мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена.
В первой серии экспериметов мы создали и проанализировали анти-c-Met-связывающую активность трех первых гуманизированных версий мышиного 11Е1 VL-домена, экспрессированного в сочетании с химерной тяжелой цепью 11Е1. Результаты, полученные в анализе прямого c-Met-связывания, показаны в фиг.63. В этом эксперименте была отмечена сходная связывающая способность тестированных полученных из 11Е1 химерных или частично гуманизированных рекомбинантных антител. На данный момент проанализировали 26 из 30 аминокислот, признанных различающимися в мышином 11Е1 VL-домене и отобранном человеческом каркасном участке, и выявили, что они не имеют отношения к анти-с-Met-связывающей активности гуманизированного VL-домена 11Е1, когда сочетаются с химерной тяжелой цепью.
III - Гуманизация антитела 11Е1
На данном этапе гуманизации моноклонального антитела 11Е1, теоретическая полученная последовательность гуманизированного антитела содержит только пять вне-CDR остатков, исходящих из родительского мышиного VH-домена, и четыре вне-CDR остатка, исходящих из родительской мышиной VL-последовательности (см. фиг.60, ряд HZ VH1, и фиг.62, ряд HZ VL1). Затем было решено немедленно охарактеризовать полученную гуманизированную версию антитела 11Е1, комбинирующую тяжелую и легкую цепи. Результаты представлены в фиг.64 для анализа прямого c-Met-связывания.
В этом эксперименте была отмечена похожая связывающая способность тестированных полученных из 11Е1 химерного или гуманизированного рекомбинантных антител. Анализ HGF-связывающих конкурентных свойств и анализ путем сайт-направленного мутагенеза вклада девяти оставшихся мышиных остатков выполнены независимо или в сочетании в этой отобранной VH1/VL1 "предварительно гуманизированной" версией моноклонального антитела 11Е1.
Пример 27-3: Гуманизация моноклонального антитела 224G11
I - Гуманизация вариабельного домена тяжелой цепи 224G11
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с мышиной последовательностью 224G11 VH.
Что касается высокой гомологии последовательностей VH-доменов 224G11 и 227Н1, также подтвержденной использованием инструментов базы данных IMGT, то те же человеческий зародышевый ген V IGHV1-2*02 и человеческий зародышевый ген J IGHJ4*01 таким образом были отобраны в качестве человеческих последовательностей, получающих мышиные 224G11 VH CDR.
Основываясь на этой высокой гомологии, было решено непосредственно перенести информацию о гуманизации, накопленную в ходе гуманизации VH-домена 227Н1 (см. пример 27), и разработать "полностью IMGT" гуманизированную версию, изображенную в фиг.65, которая представляет аминокислотное выравнивание мышиных 227Н1 и 224G11 VH-доменов с отобранным человеческим каркасным участком. В ряду «человеческий каркасный участок» отображены только те аминокислоты, которые были признаны отличными от мышиного 224G11 VH-домена. Ряд HZVHO соответствует «полностью IMGT» гуманизированной версии 224G11 VH-домена, полученной для «полностью IMGT» 227H1-HZVH домена.
Была создана «полностью IMGT» гуманизированная версия мышиного 224G11 VH-домена и проанализирована ее анти-с-Met-связывающая активность, когда она экспрессирована в комбинации с химерной легкой цепью 224G11. Результаты, полученные в анализе прямого c-Met-связывания, представлены в фиг.66, а фиг.67 иллюстрирует HGF-связывающий конкурентный анализ. С учетом экспериментальных данных, полученных в отношении анти-c-Met-связывающих свойств «полностью IMGT» гуманизированного 224G11 VH-домена, отобрали полученную аминокислотную последовательность, изображенную в фиг.65, и затем провели биоинформативный анализ для оценки уровня «человечности» так называемого 224G11-HZ VHO-домена.
С учетом применяемой здесь стратегии гуманизации, на примере 27 необходимо рассмотреть анализ «человечности» последовательности 224G11 HZ VHO. Как описано для гуманизации VH-домена 227Н1, мы подтверждаем достоверность системы нумерации IMGT и инструментов анализа гомологии, а также демонстрируем возможность передачи стратегии гуманизации между антителами в пределах их внутренней гомологии.
II - Гуманизация вариабельного домена легкой цепи 224G11
В целях выявления лучшего человеческого кандидата для CDR-прививки проводили поиск человеческого зародышевого гена, показывающего лучшую идентичность с мышиной последовательностью 224G11 VL. С помощью инструментов аналаза базы данных IMGT были выявлены две возможных человеческих V-области, получающие мышиные 224G11 VL CDR. Таким образом, были запланированы две стратегии гуманизации VL-домена 224G11. Первая соответствует начальному испытанию человеческого каркасного участка с более коротким CDR1 (IGKV3-11*01), второй с более длинным CDR1 (IGKV4-1*01)
Фиг.68 представляет аминокислотное выравнивание мышиного 224G11 VL-домена с двумя отобранными человеческими каркасными участками. В рядах и более короткого («короткий Hu-FR»), и более длинного («длинный Hu-FR») человеческого каркасного участка были отображены только те аминокислоты, которые отличаются от мышиного VL-домена 224G11. Ряды HZ VL3 и HZ VL6 соответствуют основным гуманизированным версиям 224G11 VL-домена с вышеупомянутыми (ряд "ранг") мутациями, четко определенными. Номер под каждой предлагаемой мутацией соответствует рангу, в котором указанная мутация будет осуществлена, когда будет отобран базисный каркасный участок с «коротким» или «длинным» CDR.
В первой серии экспериметов мы создали и проанализировали анти-c-Met-связывающую активность двух базисных гуманизированных версий мышиного VL-домена 224G11, экспрессированного в сочетании с химерной тяжелой цепью 224G11. Результаты, полученные в анализе прямого c-Met-связывания, показаны в фиг.69. В этом эксперименте была отмечена сходная связывающая способность тестированных химерной версии и версии HZ VL6 («длинный» CDR1), в то время как рекомбинантное полученное из 224G11 антитело HZ VL3 («короткий» CDR1) почти не демонстрировало связывания. На данный момент проанализировали 26 из 30 аминокислот, признанных различающимися в мышином 224G11 VL-домене и отобранном человеческом каркасном участке, и выявили, что они не имеют отношения к анти-с-Met-связывающей активности гуманизированного VL-домена 224G11, когда сочетаются с химерной легкой цепью.
Во второй серии экспериментов мы создали и проанализировали анти-с-Met-связывающую активность осуществленных гуманизированных версий VH-домена 224G11, полученного из HZ VL6, когда он экспрессируется в сочетании с химерной тяжелой цепью 224G11. Были проанализированы две дополнительные гуманизированные формы; в версии HZ VL5 гуманизированными являются семь остатков из третьей группы (3 ранг), а в версии HZ VL4 только четыре остатка из первой группы (остатки 1 ранга) остаются мышиными. Результаты, полученные в анализе прямого c-Met-связывания, показаны в фиг.70. В этом эксперименте не было отмечено никаких различий в связывающей способности тестируемых полученных из 224G11 химерных или частично гуманизированных рекомбинантных антител. На данный момент проанализировано 18 из 22 аминокислот, которые были признаны различающимися в мышином 224G11 VL-домене и отобранном человеческом каркасном участке с «длинным CDR1», и обнаружено, что они не имеют отношения к анти-с-Met-связывающей активности гуманизированного VL-домена 224G11, когда сочетаются с химерной тяжелой цепью.
Затем протестировали HZ VL4 гуманизированную версию 224G11 VL-домена в HGF-связывающем конкурентном анализе. Как показано в фиг.71, полученные результаты демонстрируют схожую конкурентную активность мышиных и рекомбинантных химерных и HZ VL4-гуманизированных антител, полученных из 224G11.
На данном этапе гуманизации VL-домена 224G11 полученная последовательность содержит только четыре вне-CDR остатка, исходящих из мышиной родительской последовательности. Как показано в фиг.72, этими четырьмя §-меченными остатками являются L4, М39, Н40 и R84.
На основании системы нумерации IMGT и инструментов анализа гомологии мы показали, что человеческий каркасный участок, демонстрирующий структурные различия в плане длины CDR, по-прежнему может подходит для способа гуманизации. Затем было решено охарактеризовать полученную гуманизированную версию тяжелой и легкой цепи антитела 224G11. Затем с помощью сайт-направленного мутагенеза выполняли анализ вклада сохранившихся четырех мышиных остатков, когда они экспрессированы в сочетании с VHO-гуманизированной версией тяжелой цепи.
III - Гуманизация антитела 224G11
В первой серии экспериметов мы создали и проанализировали анти-c-Met-связывающую активность полностью гуманизированной версии антитела 224G11. Эта рекомбинантная версия охватывает как VHO, так и VL4 гуманизированные VH-и VL-домены соответственно. Результаты, полученные из анализа прямого анти-с-Met-связывания, показаны в фиг.73. В этом эксперименте анти-c-Met связывающая активность полностью человеческого 224G11 была признана схожей с активностью «одноцепочечной» гуманизированной и химерной рекомбинантных версий 224G11.
Затем протестировали полностью гуманизированную версию VL-домена 224G11 в HGF-связывающем конкурентном анализе. Как показано в фиг.74, полученные результаты демонстрируют схожую конкурентную активность родительских мышиных и рекомбинантных химерных и полностью гуманизированных антител, полученных из 224G11.
На данном этапе гуманизации антитела 224G11 полученная последовательность содержит только четыре вне-CDR остатка, исходящих из мышиной родительской последовательности вариабельного домена легкой цепи. Затем с помощью сайт-направленного мутагенеза мы анализировали одиночные варианты VL-4 гуманизированного VL-домена, когда они экспрессированы в сочетании с VH0-гуманизированной версией тяжелой цепи. Как показано в фиг.75 для прямого связывающего анализа, среди четырех протестированных мы выявили потенциальные релевантные остатки, которыми являются М39 и Н40.
Было решено проанализировать множественные мутанты HZ VL4 гуманизированного 224G11 VL-домена, когда он экспрессируется в сочетании с HZ VHO гуманизированным 224G11 VH-доменом. Как показано в фиг.76 для прямого связывающего анализа и в фиг.77 для HGF-связывающего конкурентного анализа, множественные аминокислотные мутанты VL4-домена были проанализированы с целью выявления лучшего гуманизированного сочетания. На основании анализа одиночных мутантов были изучены двойные и тройные мутанты, которые могут демонстрировать лучшую анти-с-Met-активность. Мутант VH0/VL4-2x соответствует HZ VH0 224G11 гуманизированному VH-домену, экспрессированному с HZ VL4 224G11 гуманизированным VL-доменом с двойной мутацией L4M/R84G. Мутант VH0/VL4-3X соответствует HZ VH0 224G11 гуманизированному VH-домену, экспрессированному с HZ VL4 224G11 гуманизированным VL-доменом с тройной L4M/M39L/R84G мутацией.
С учетом экспериментальных данных, полученных в отношении анти-c-Met-связывающих свойств полностью гуманизированного антитела 224G11, выполняли биоинформативный анализ последовательностей вариабельного домена как тяжелой, так и легкой цепей для оценки уровня «человечности» лучших гуманизированных версий VH0/VL4-2x и VH0/VL4-3x. Ранее была продемонстрирована «полностью IMGT» гуманизация VH0 224G11 VH-домена. Что касается уровня «человечности» гуманизированных версий 224G11 VL-домена VL4-2x и -3х, то они содержат только мышиные остатки М39 и/или Н40. Эти два потенциальных ключевых остатка находятся в конце CDR1, М39 является N-концевым CDR-якорем. Учитывая проблемы с длиной CDR, с которыми мы столкнулись во время гуманизации VL-домена 224G11, и учитывая эти позиции как часть определения Кабата VL CDR1, уровень «человечности» полностью гуманизированного антитела 224G11 должен демонстрировать сильно сокращенную иммуногенность из-за минимальных сохранившихся мышиных остатков.
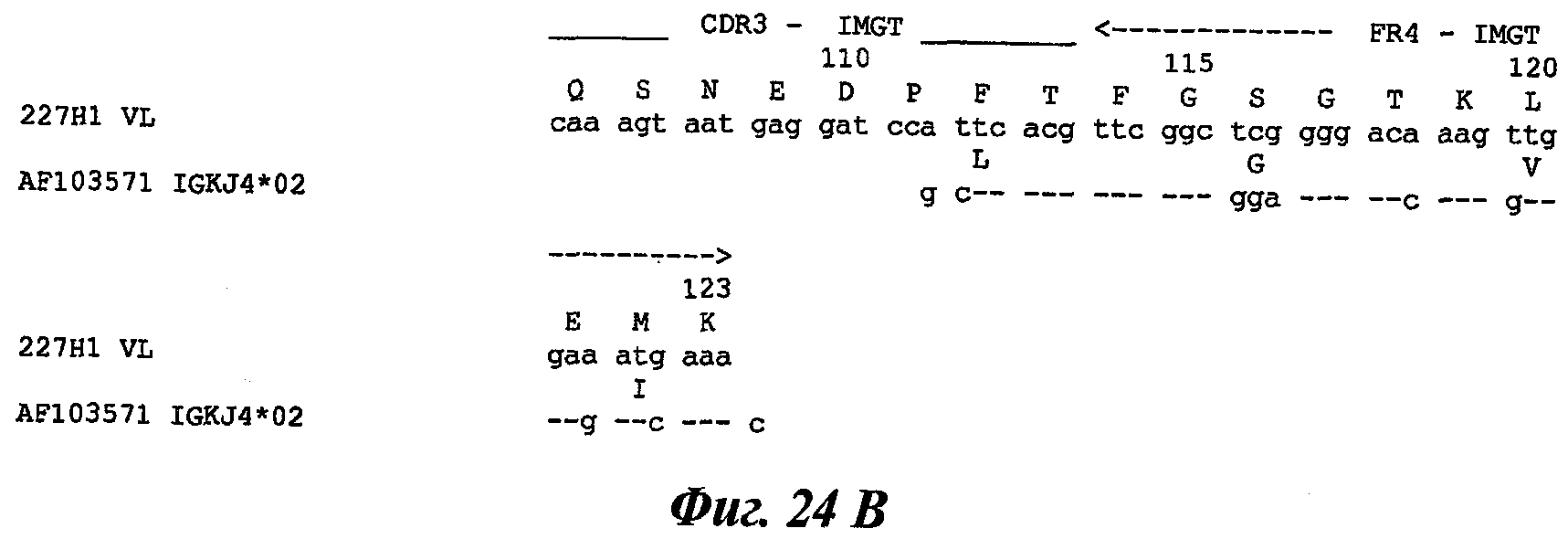
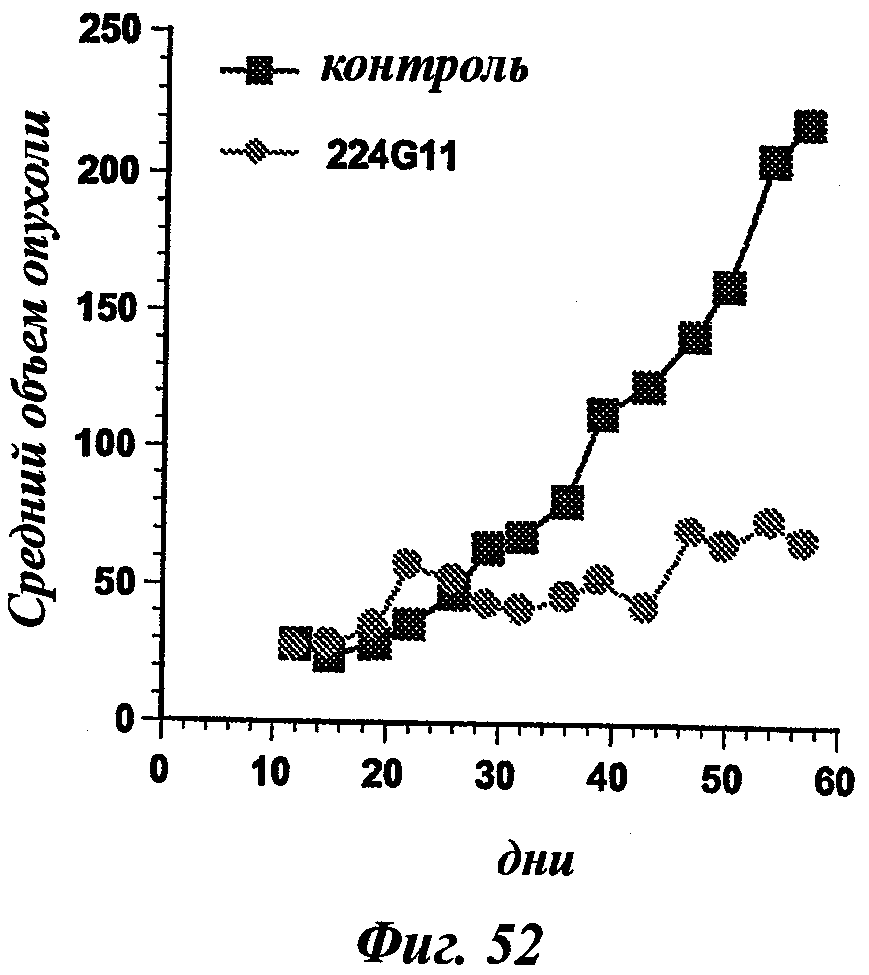
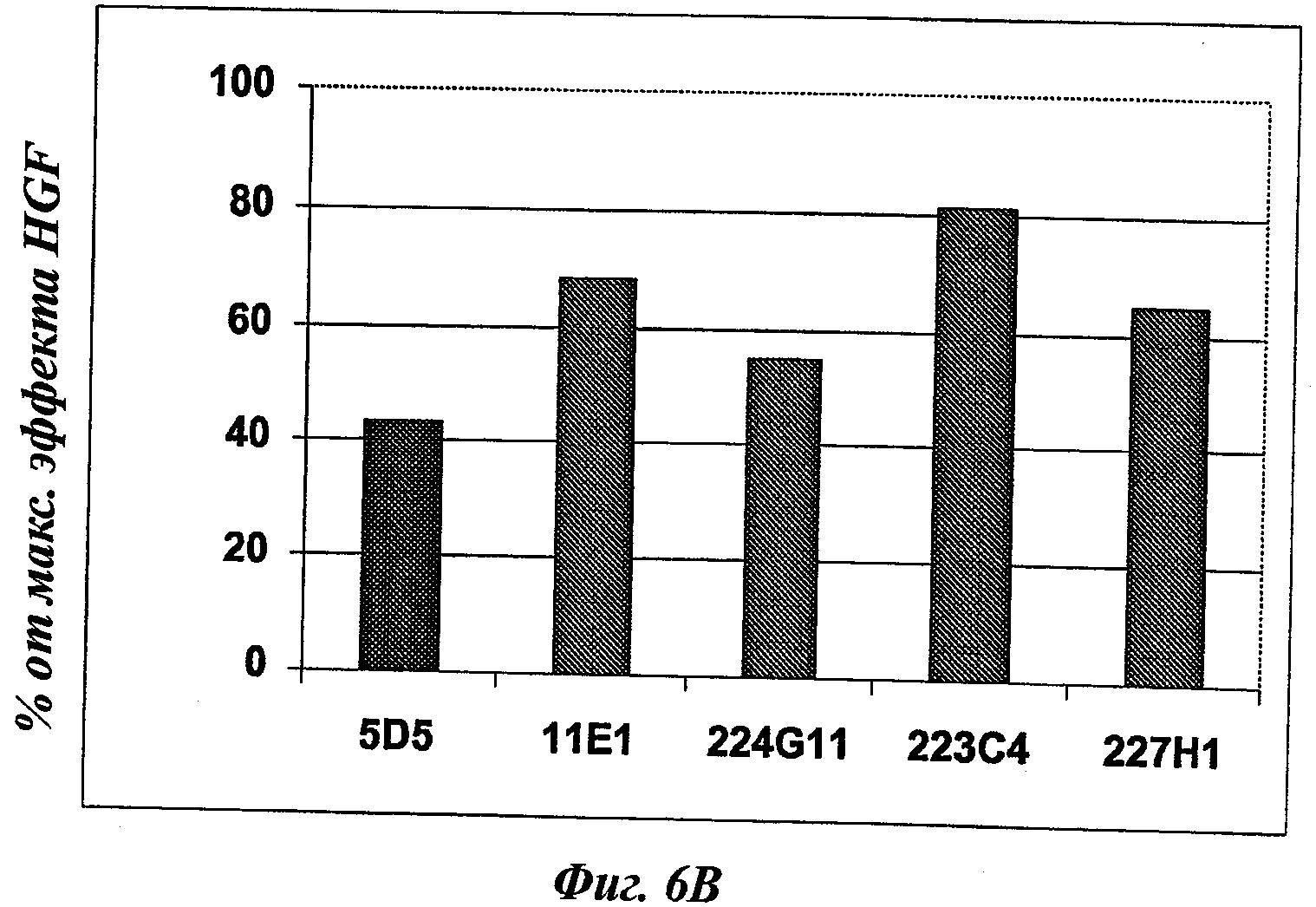
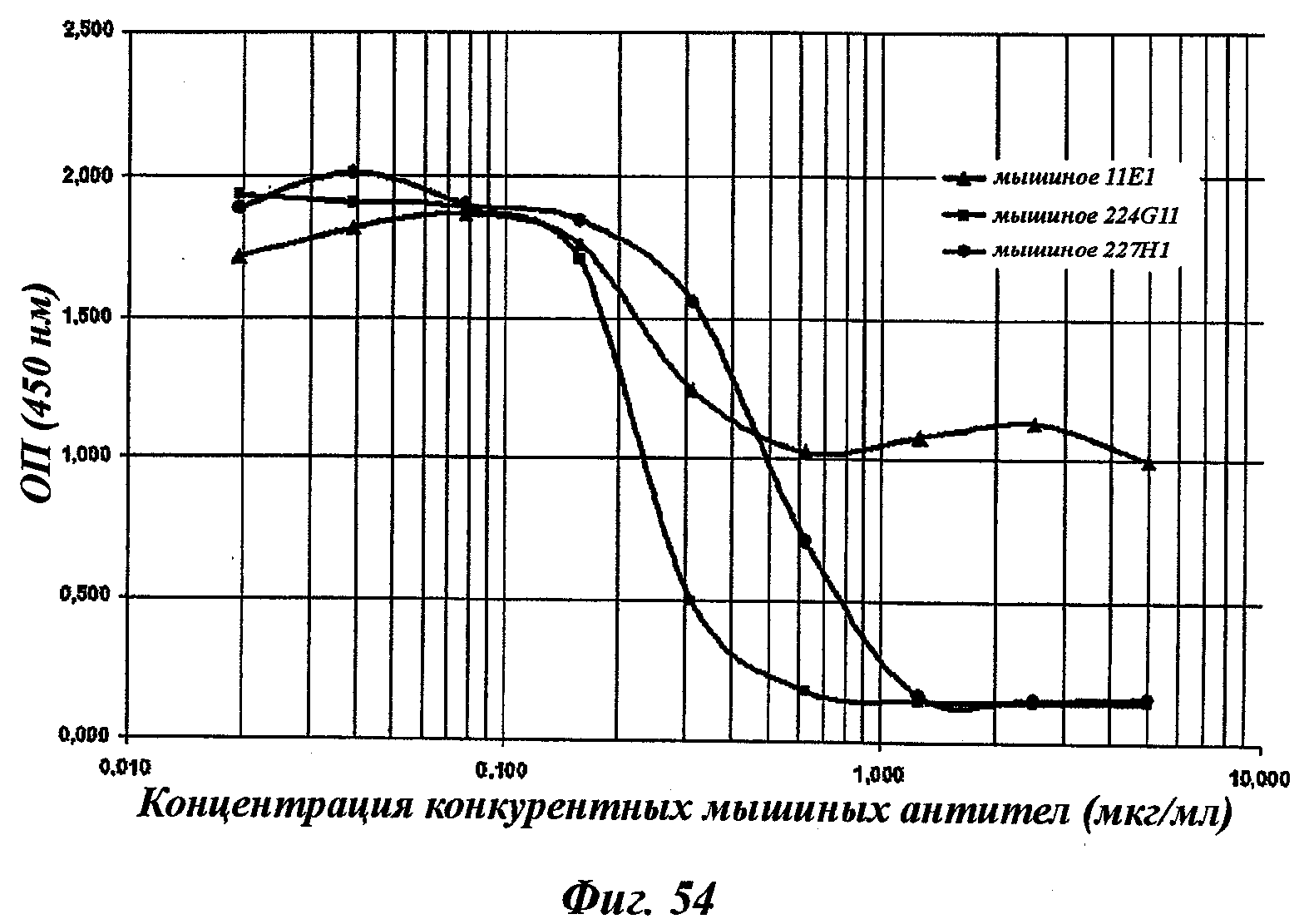
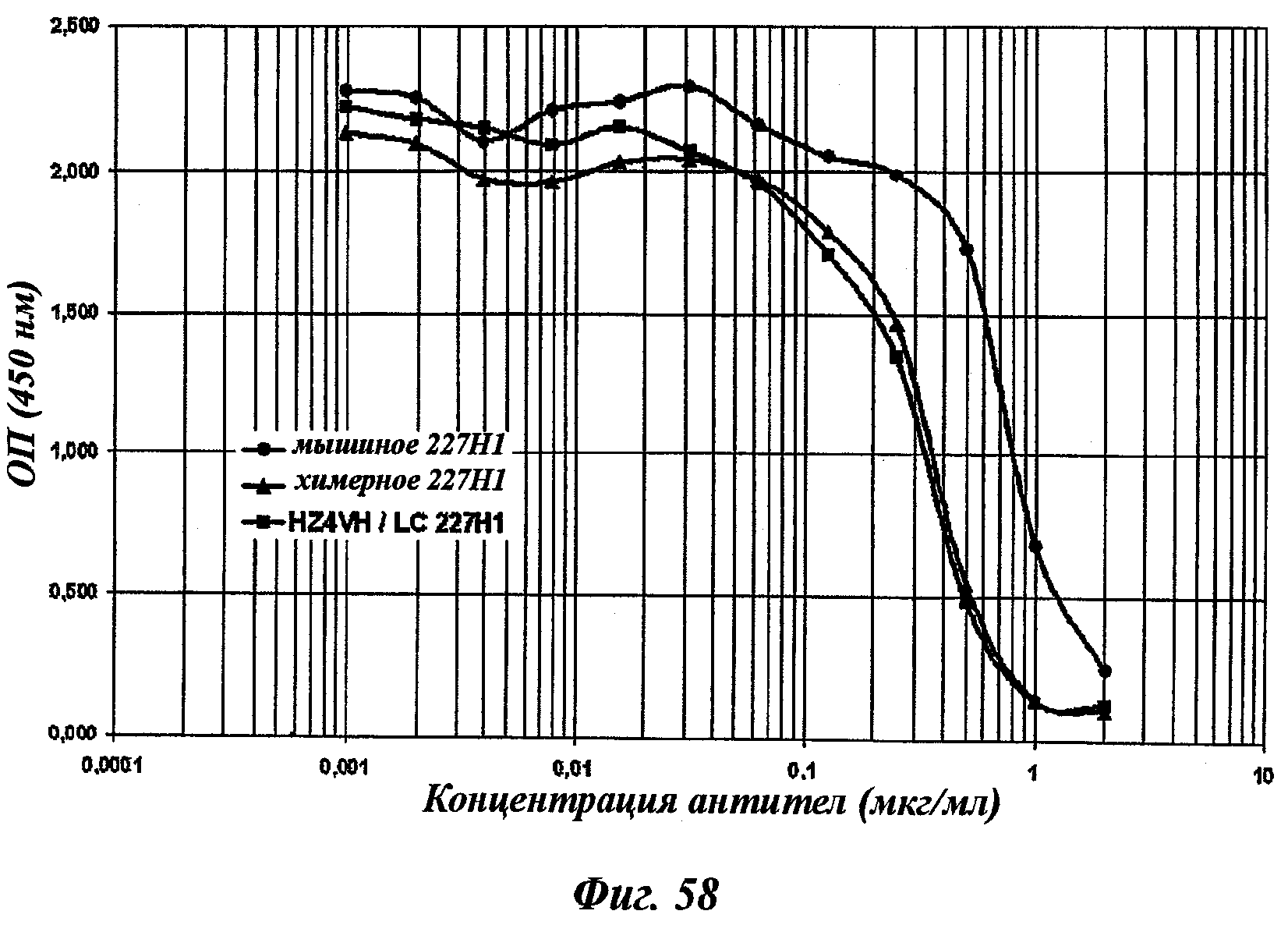
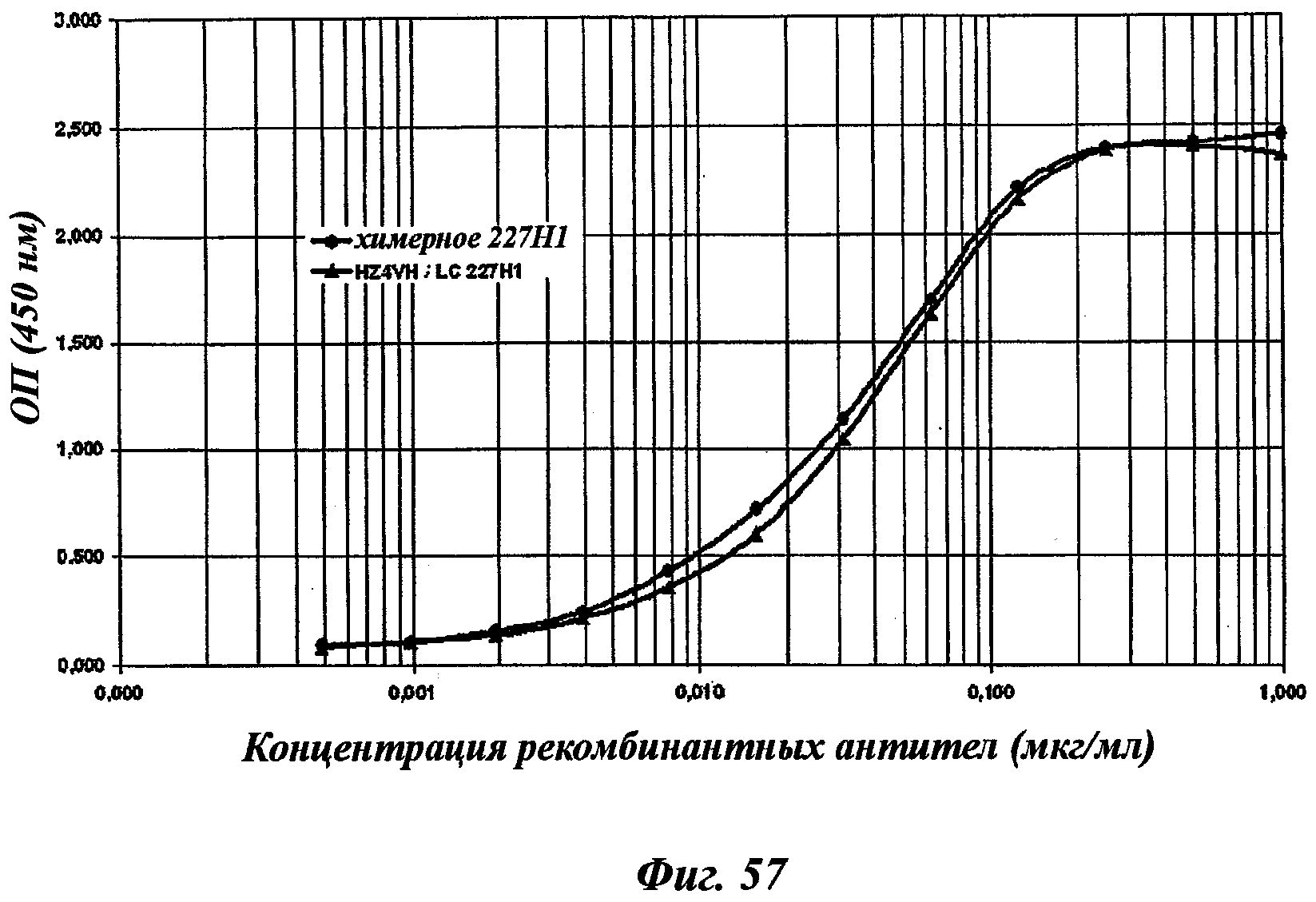

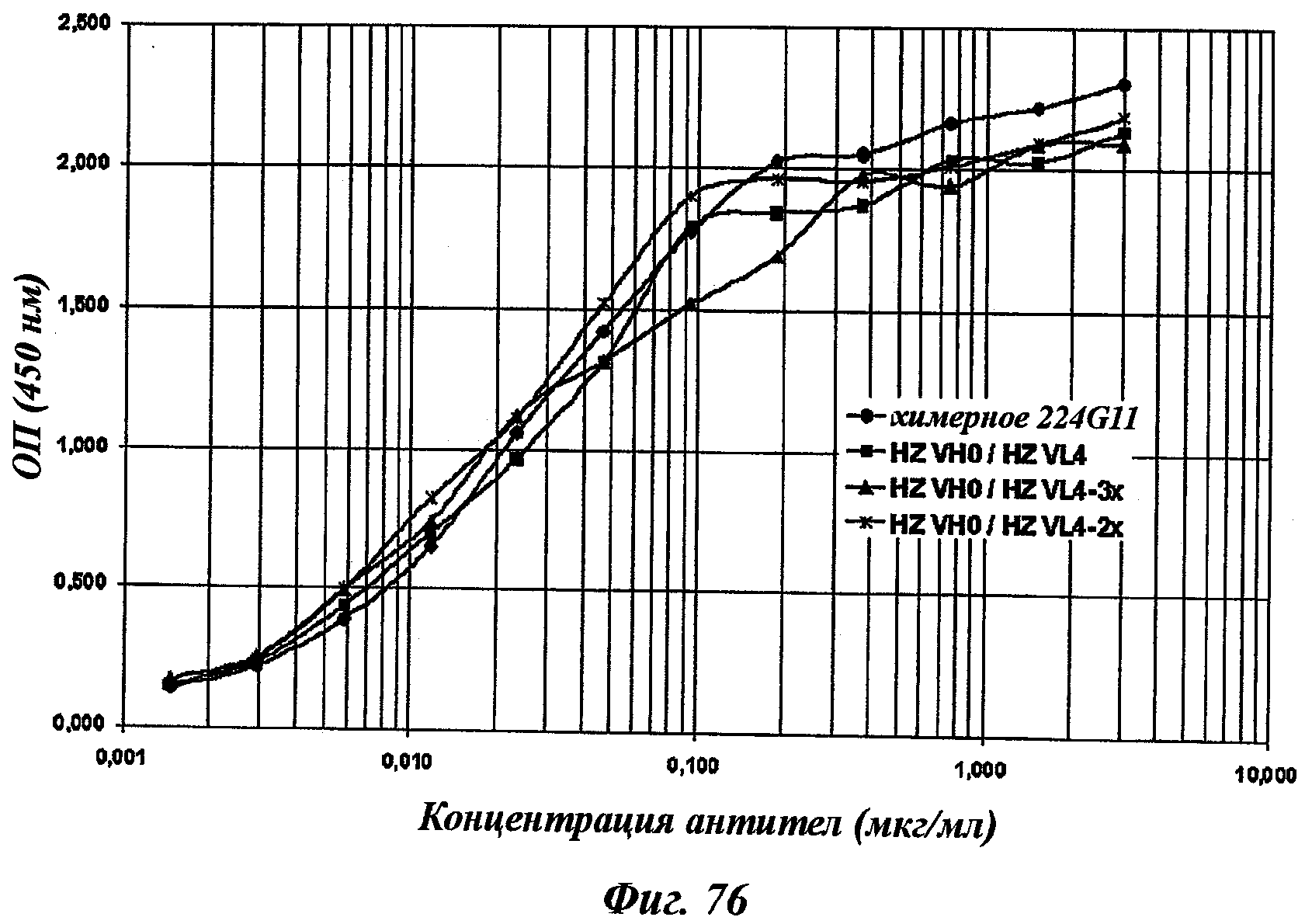
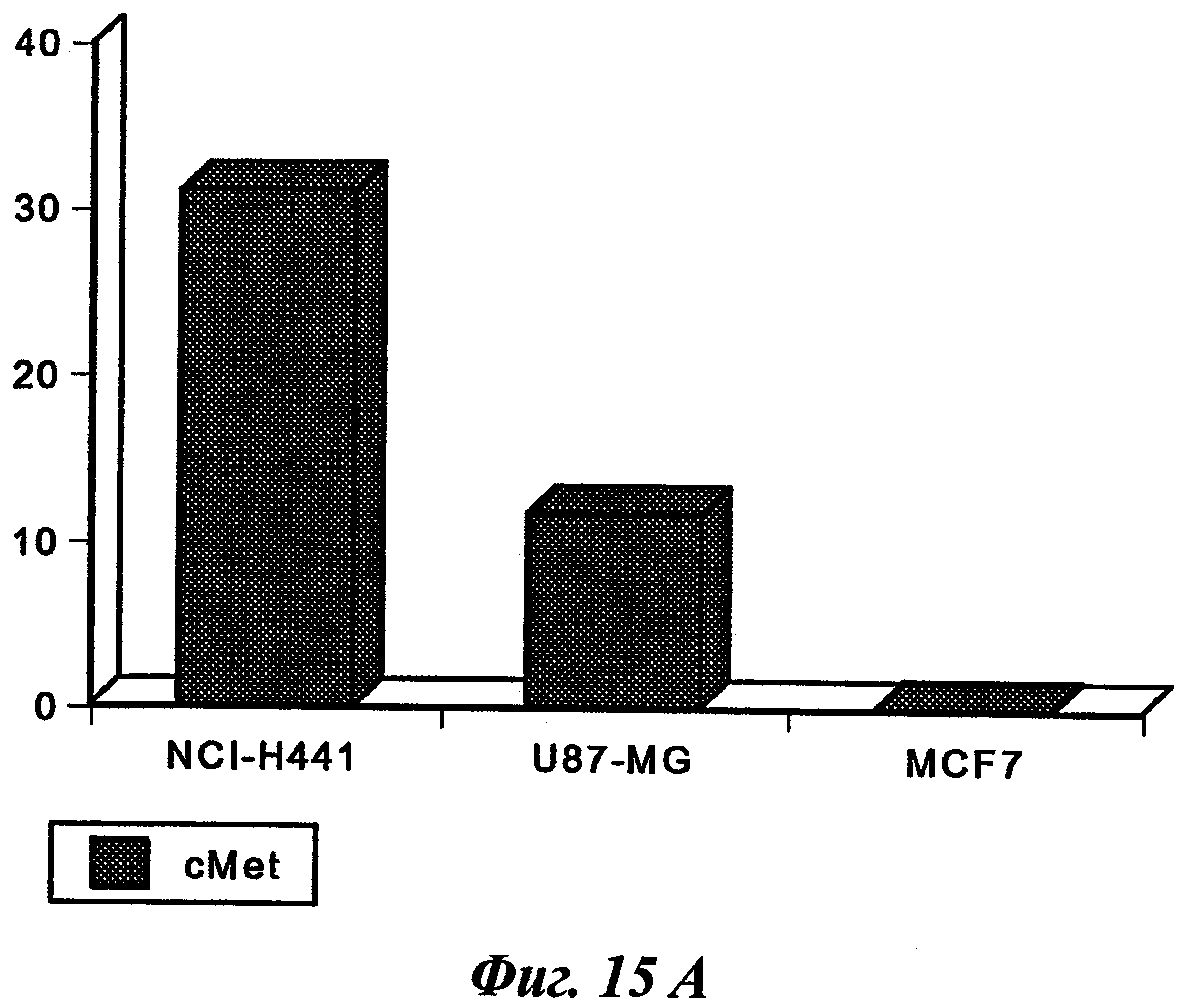
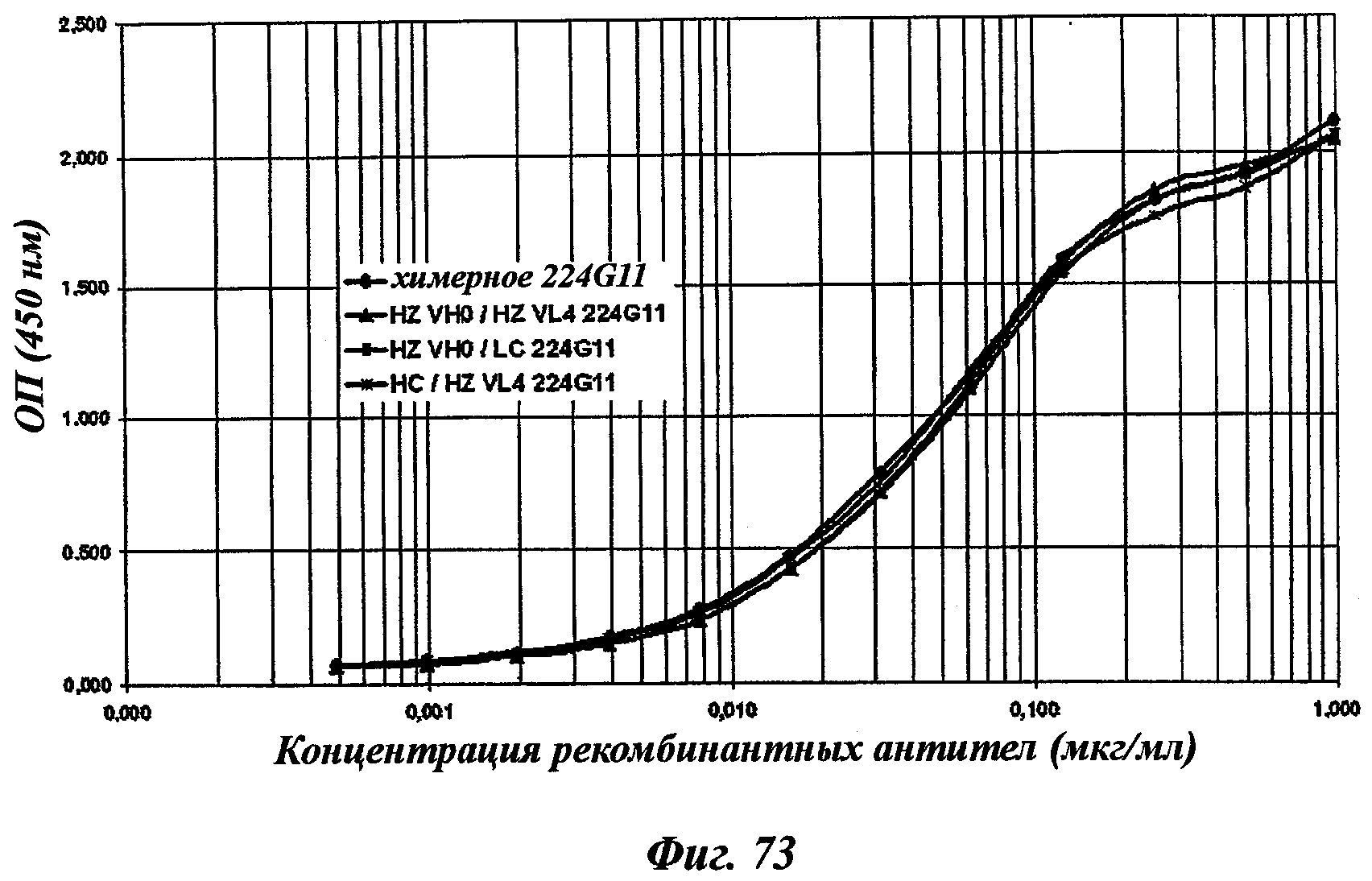
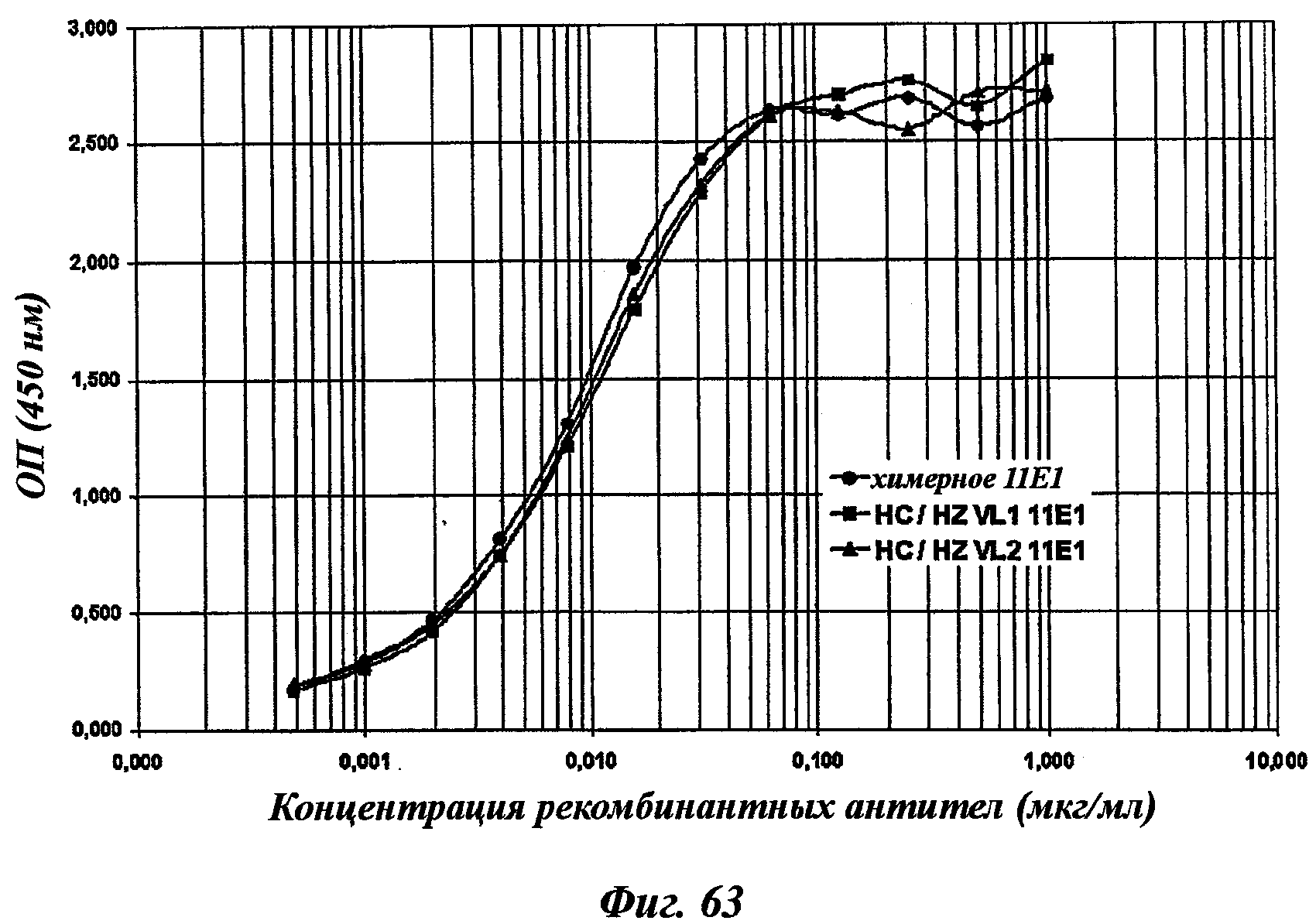
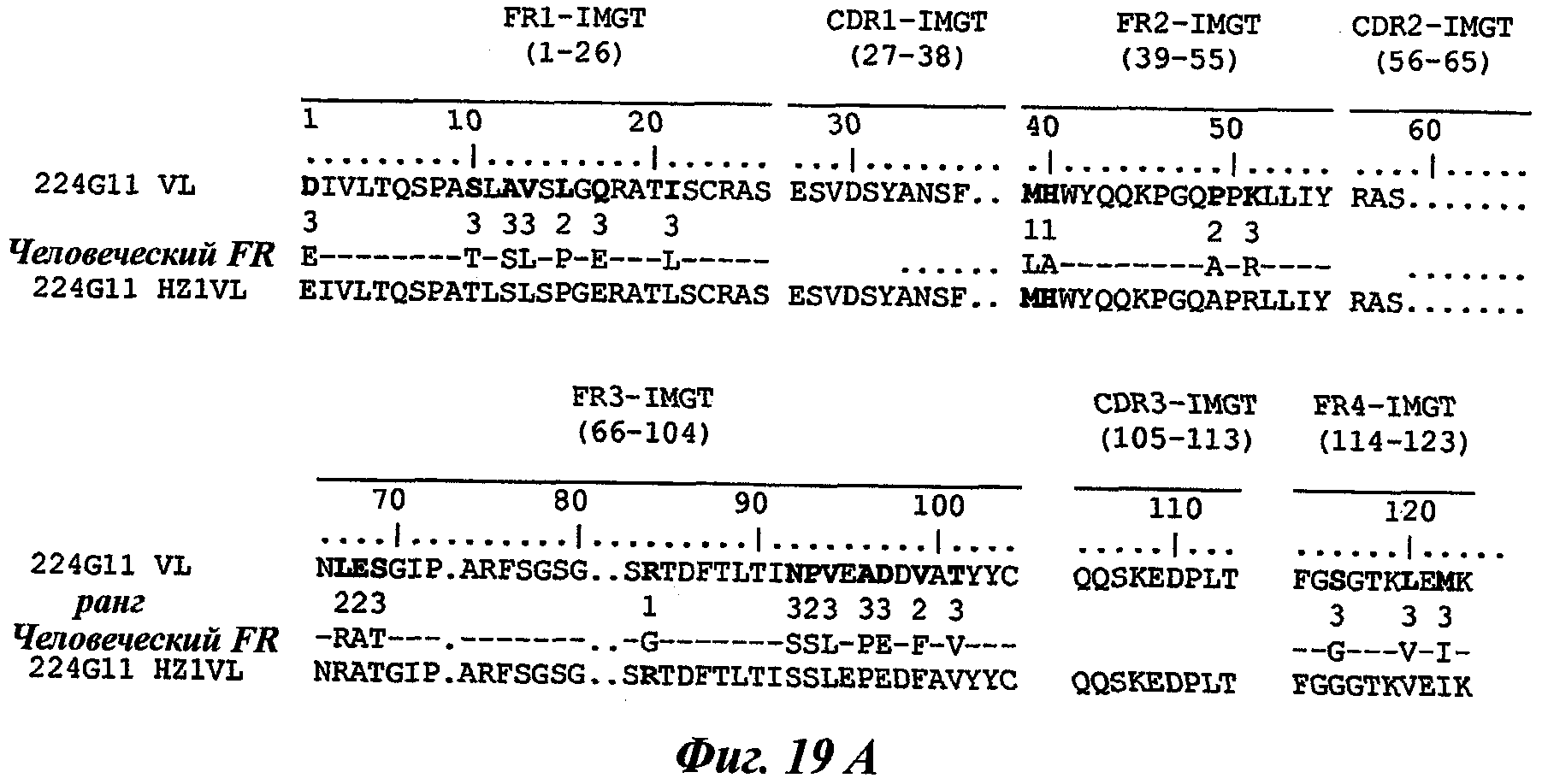
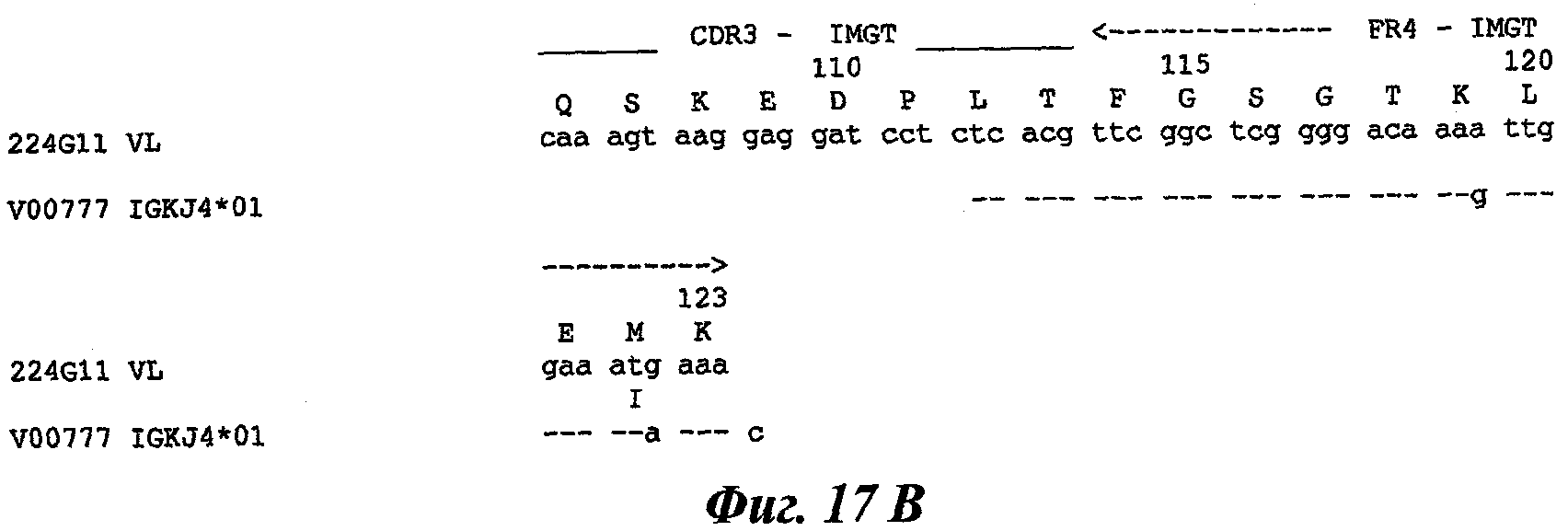
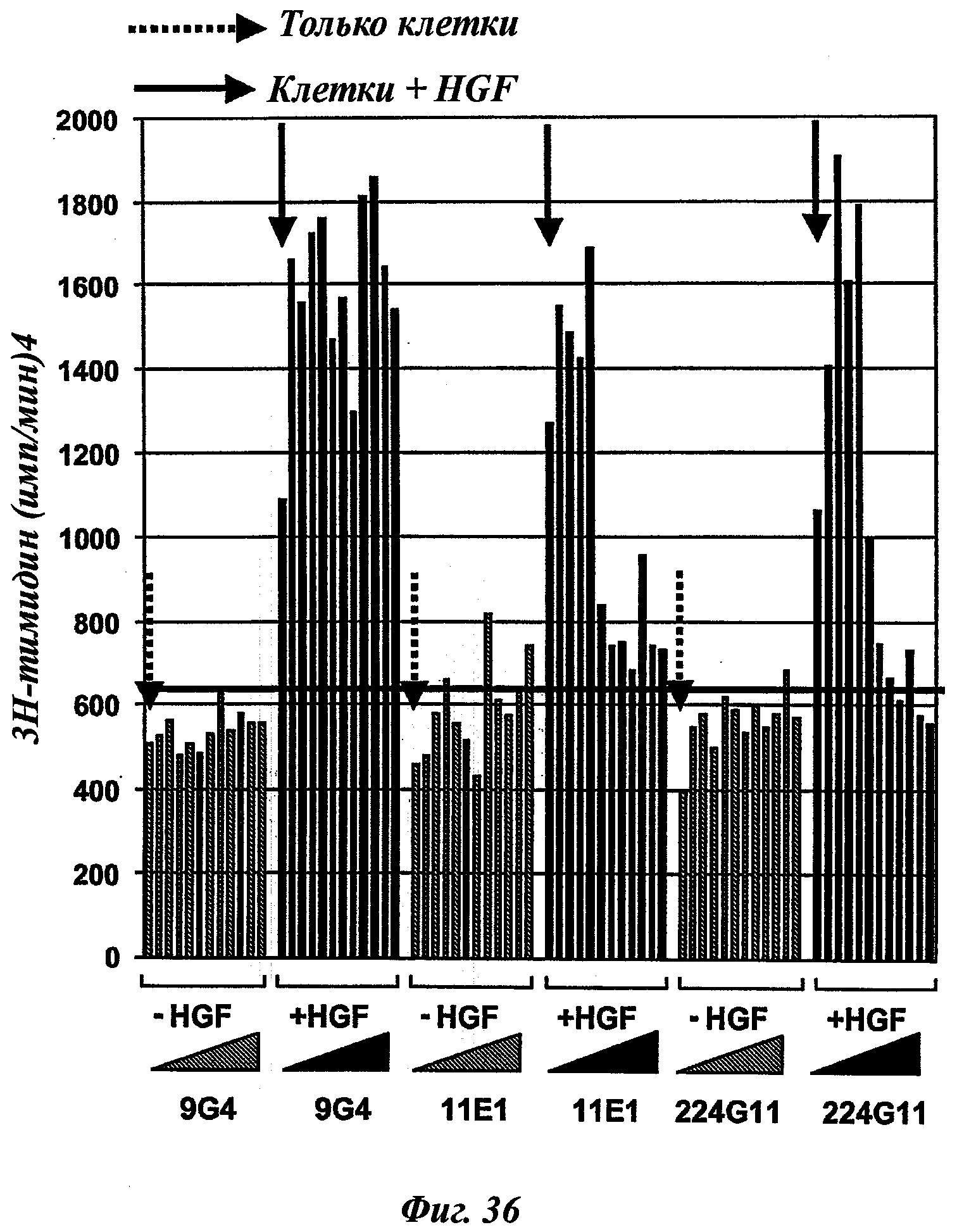
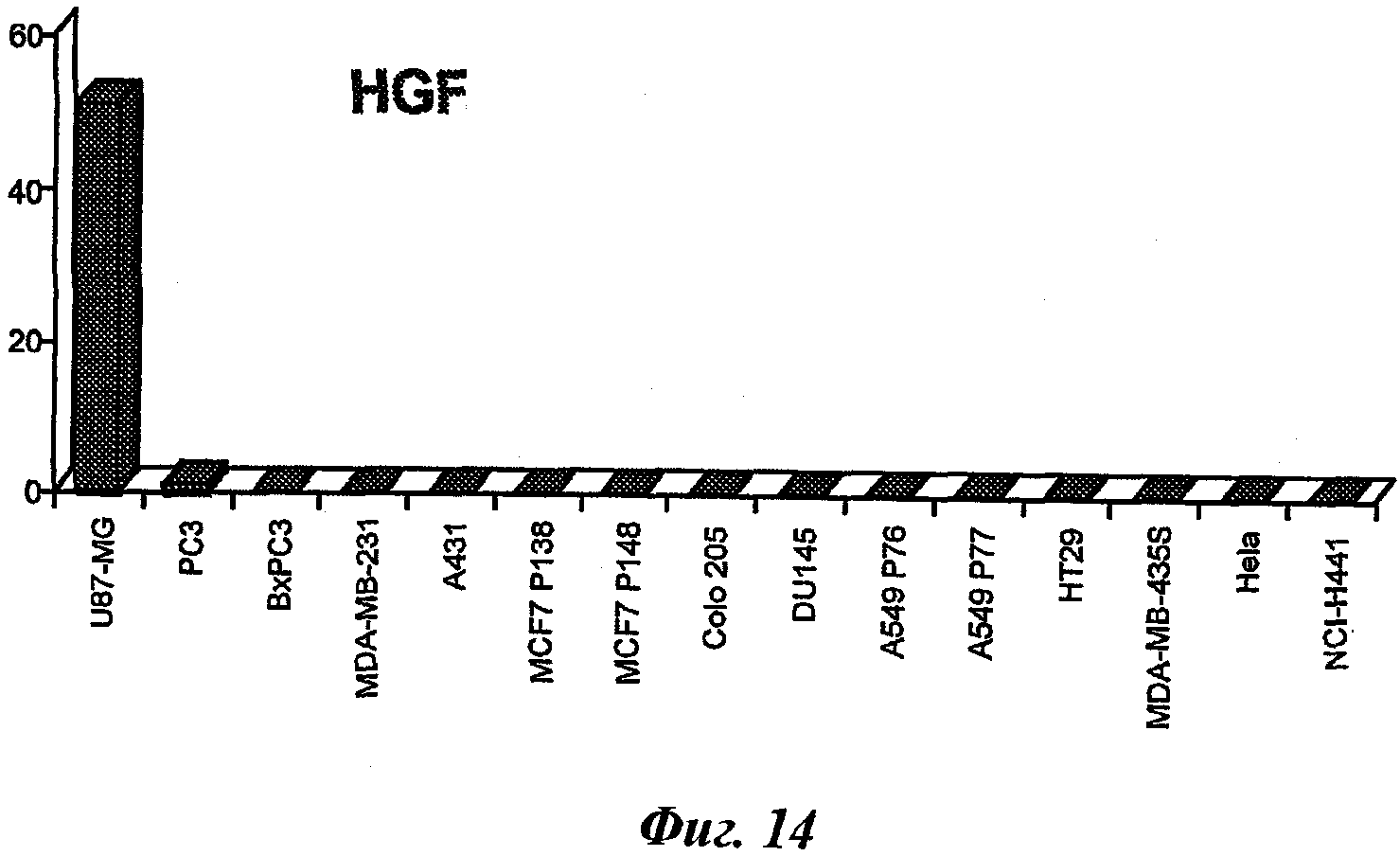
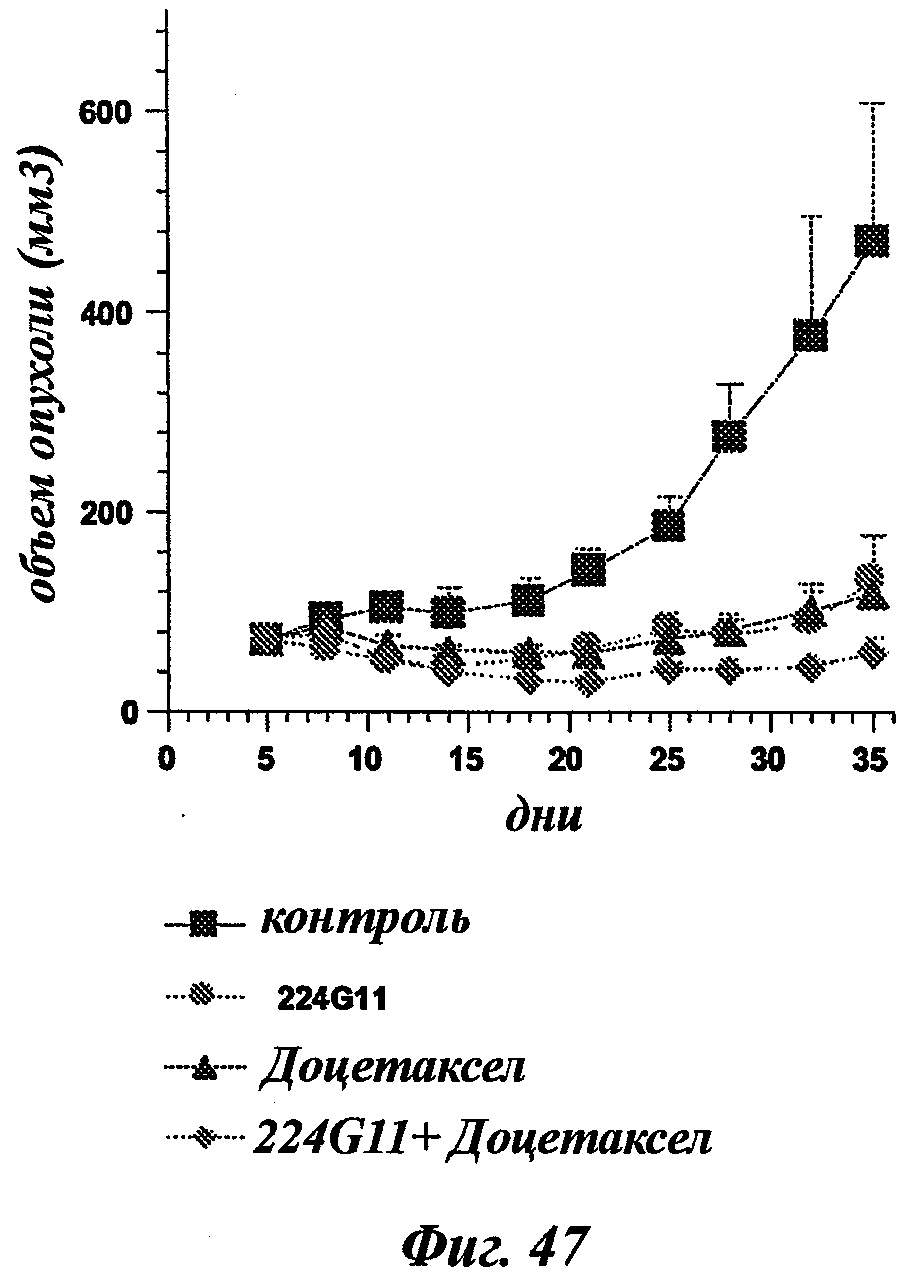

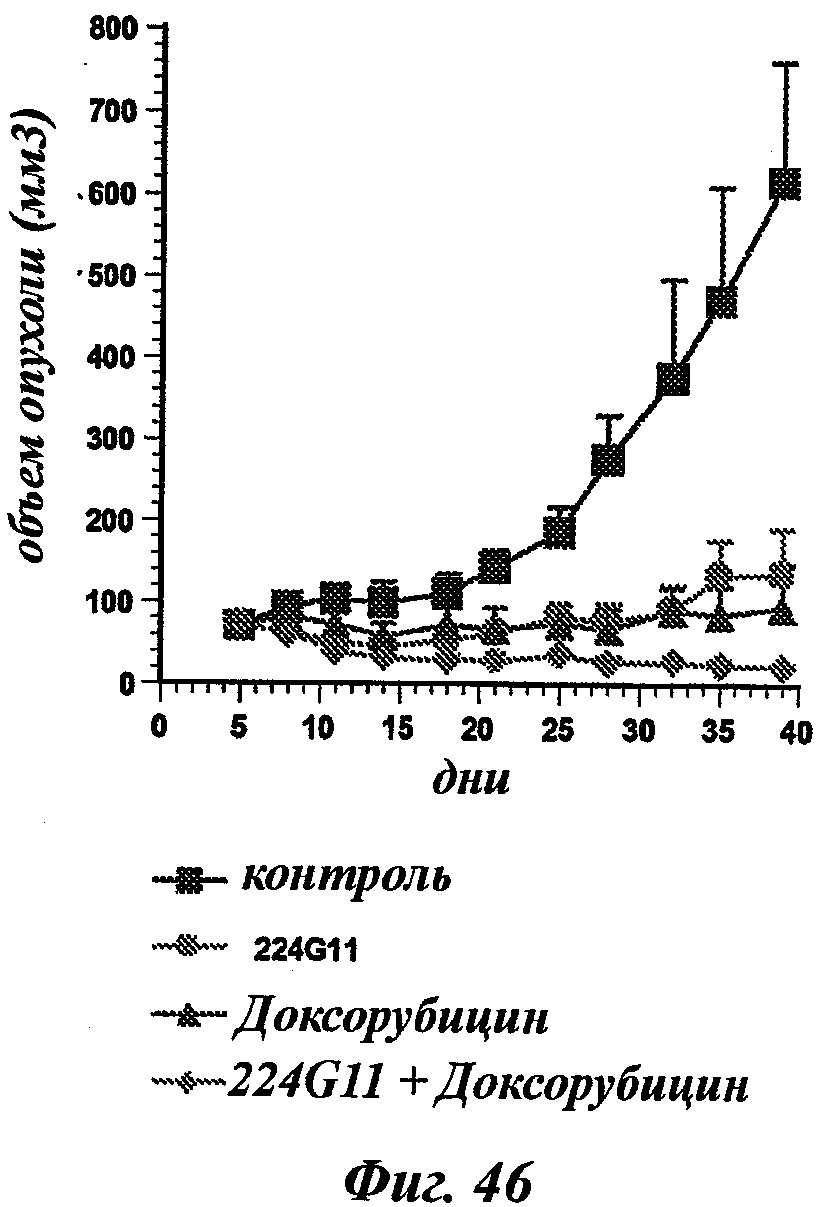
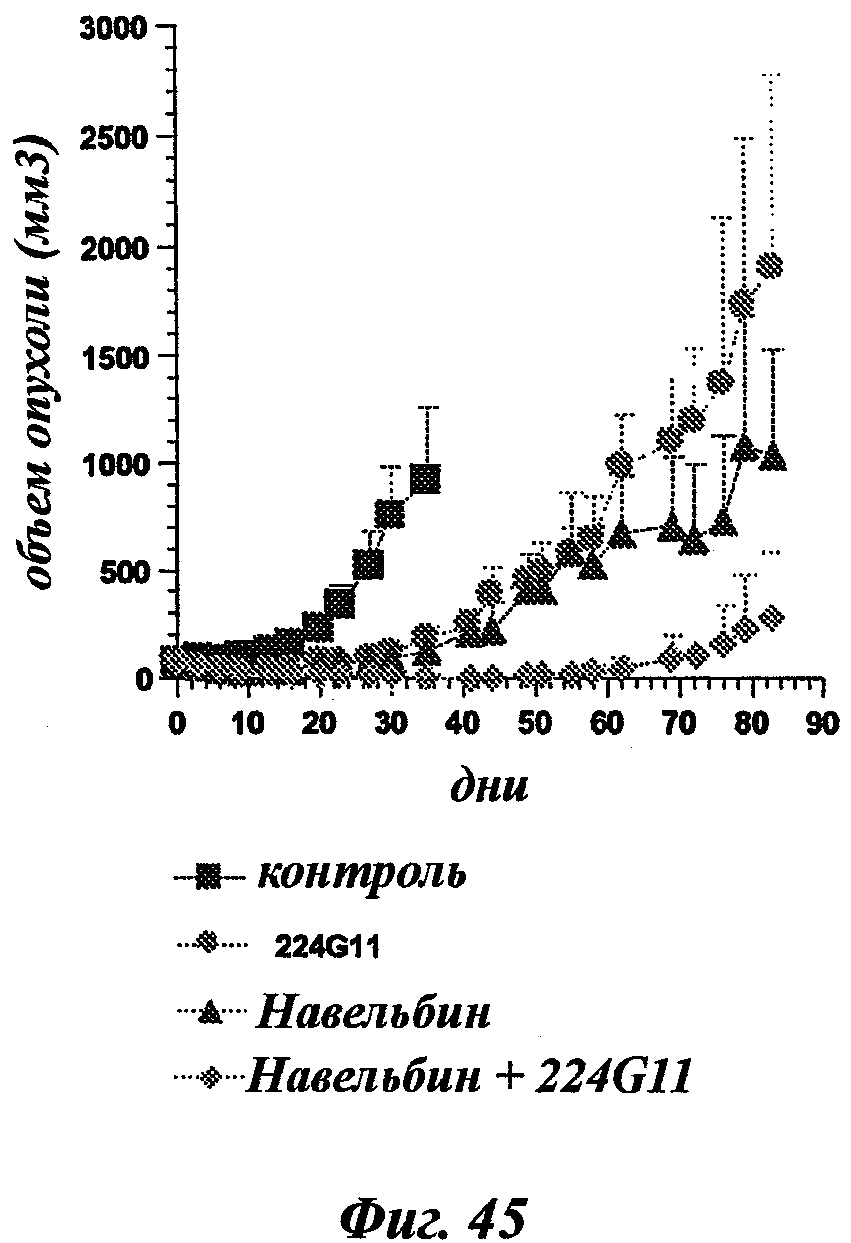
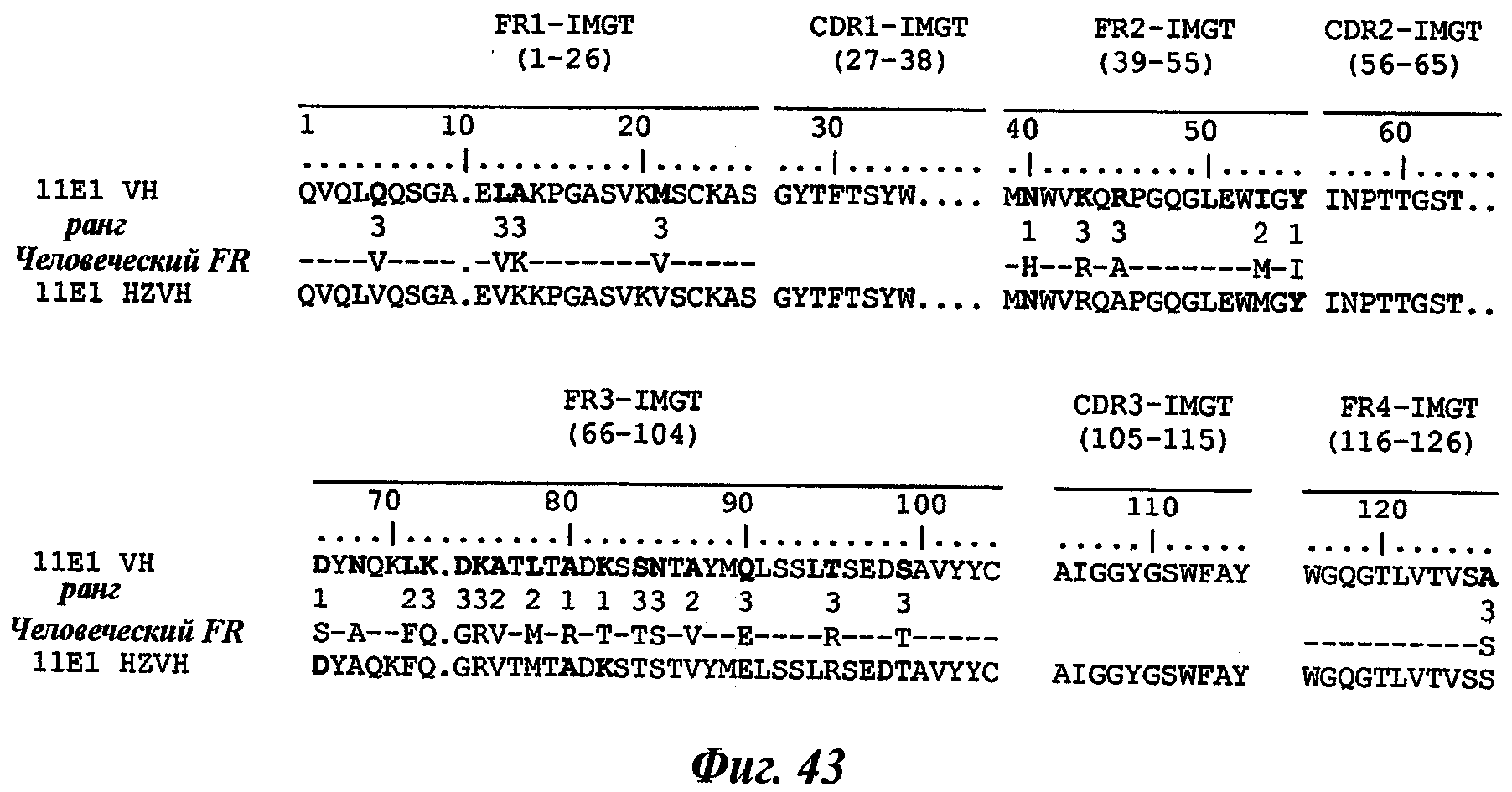
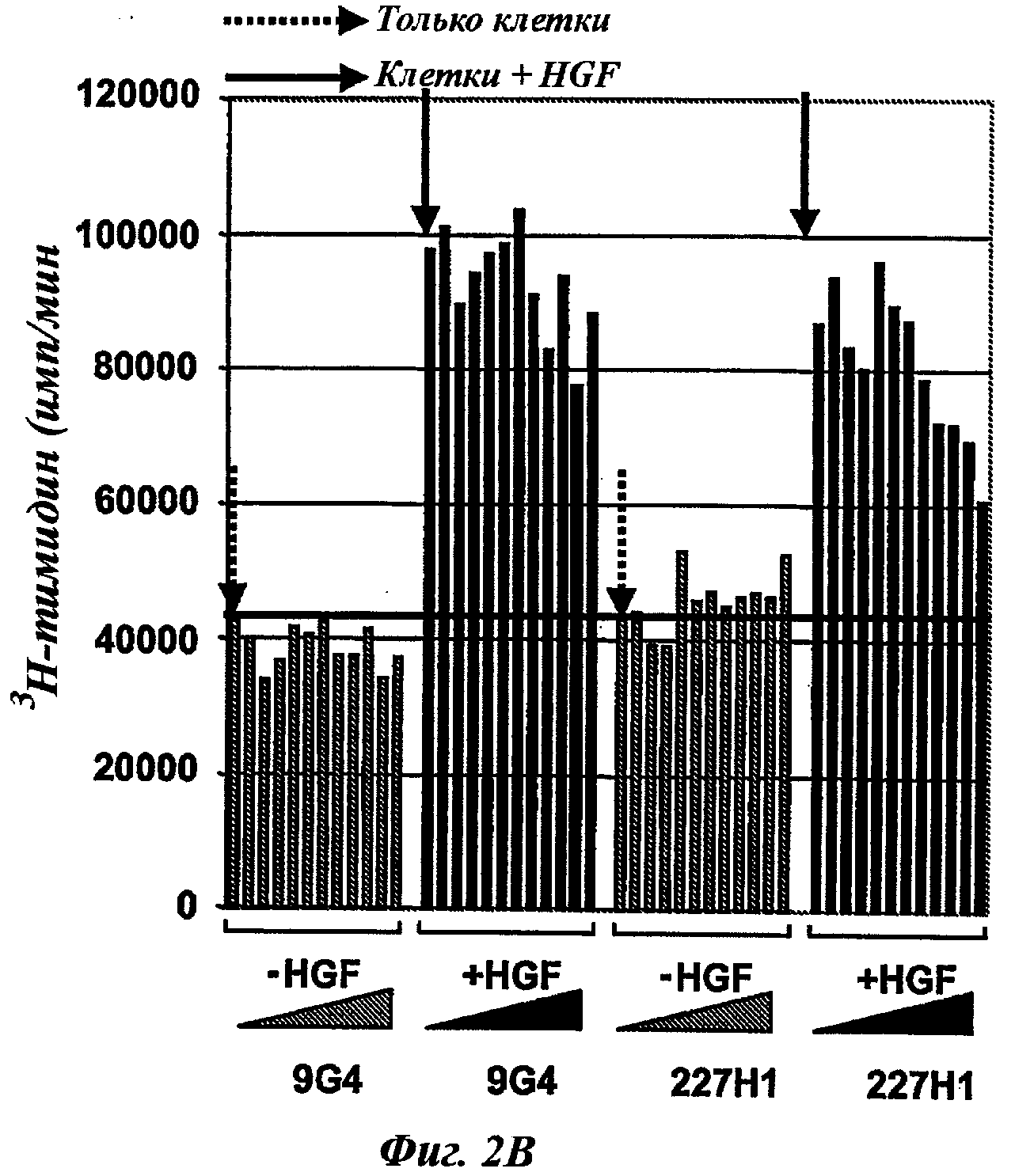
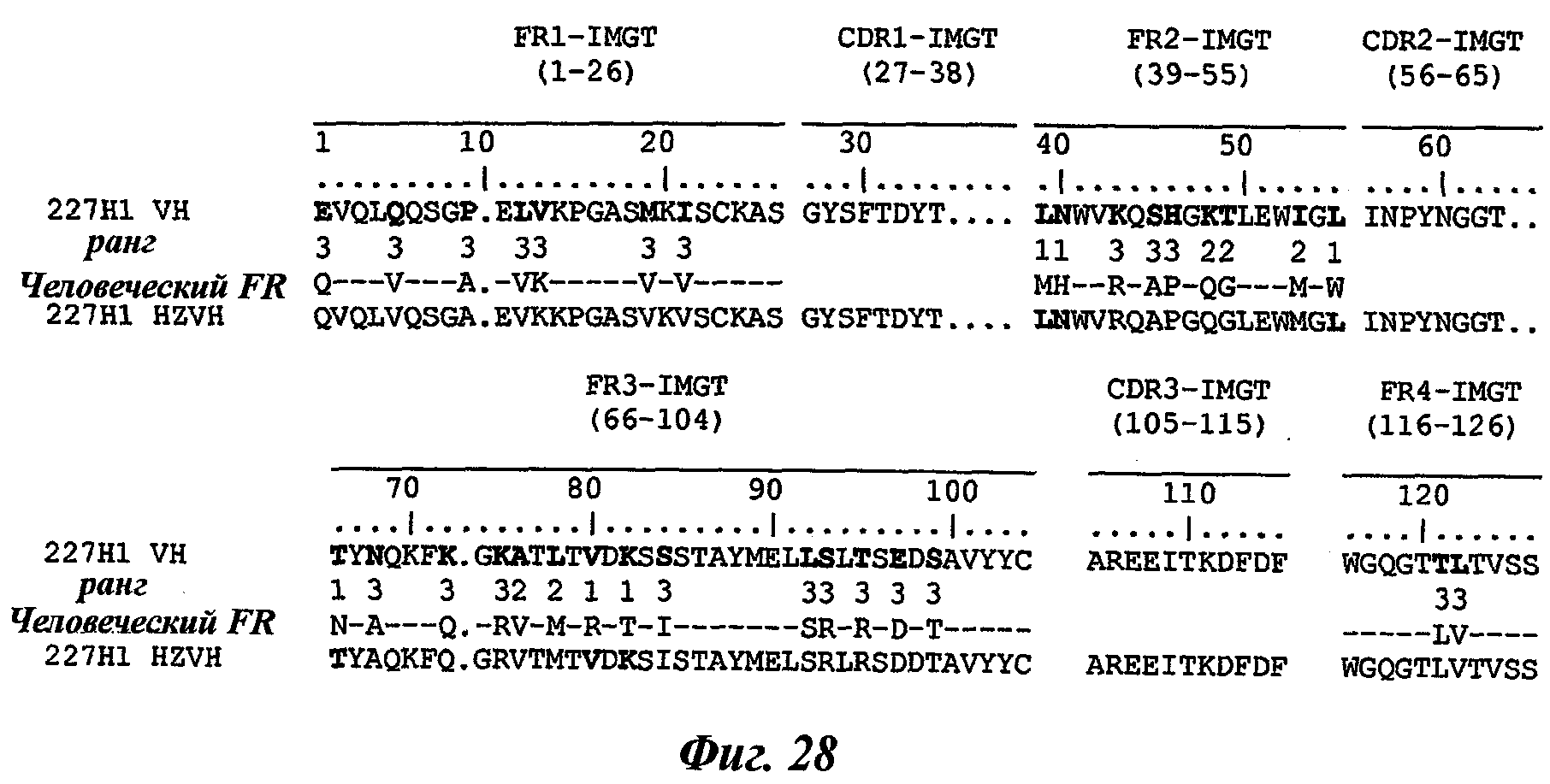

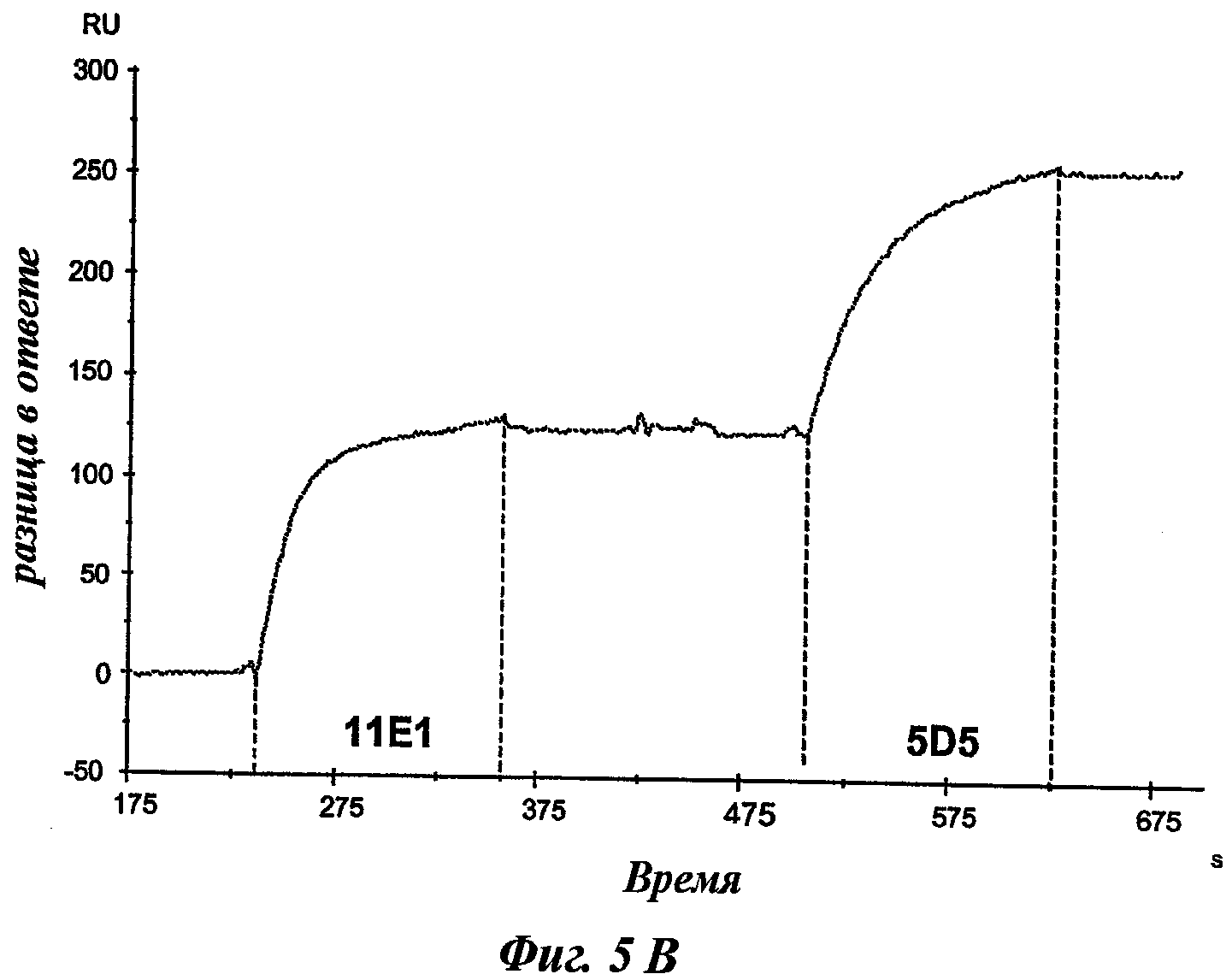
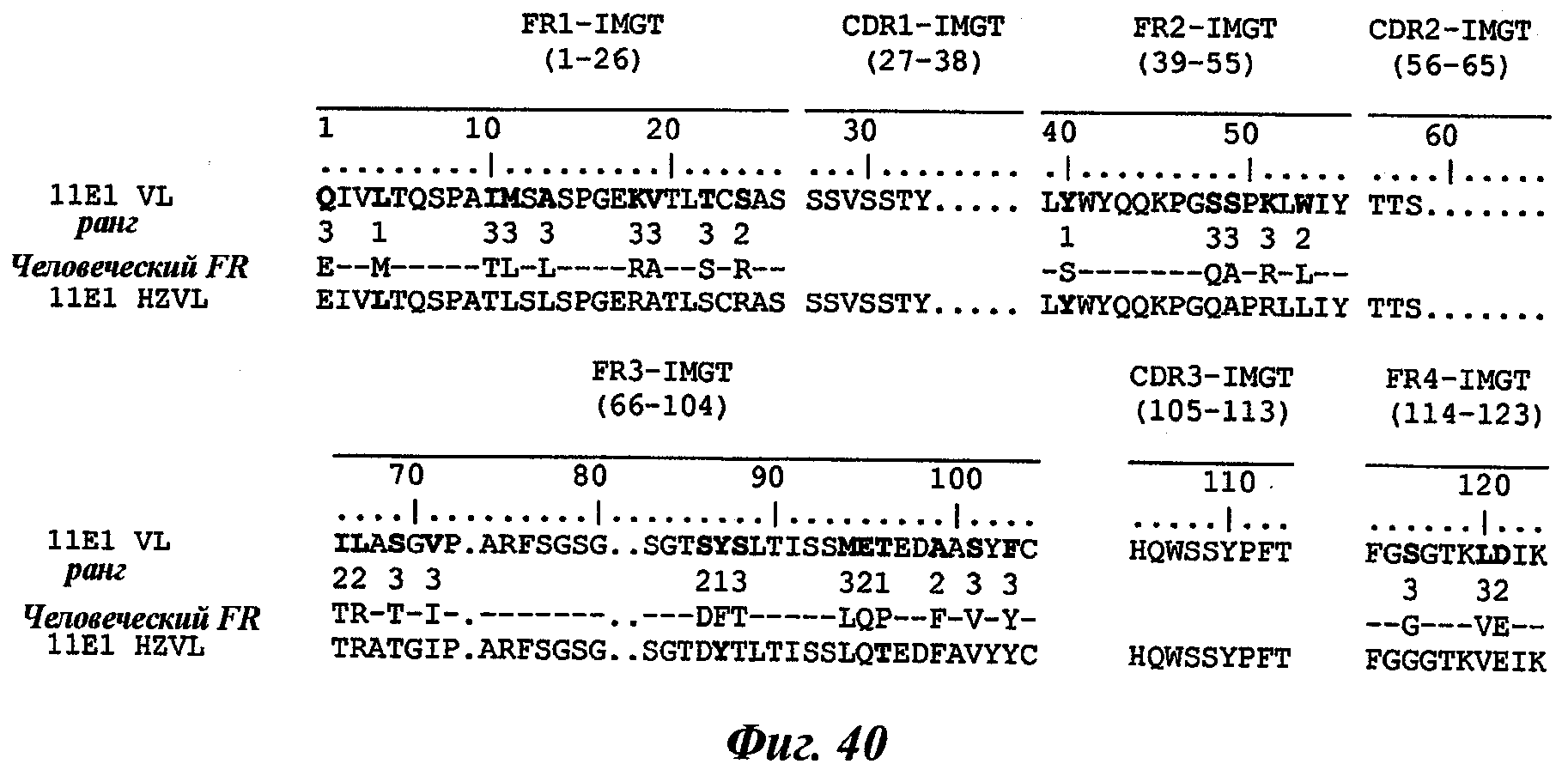



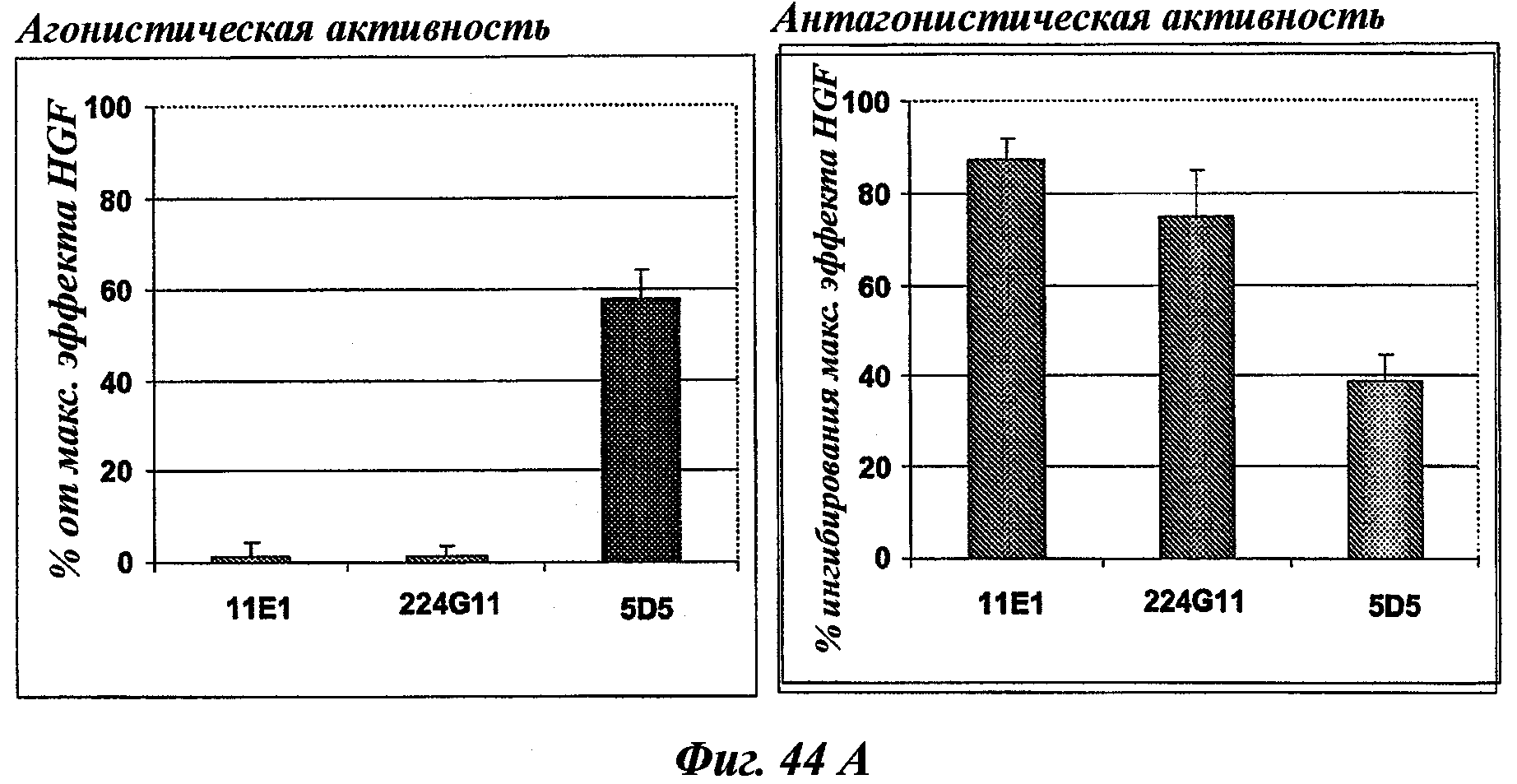
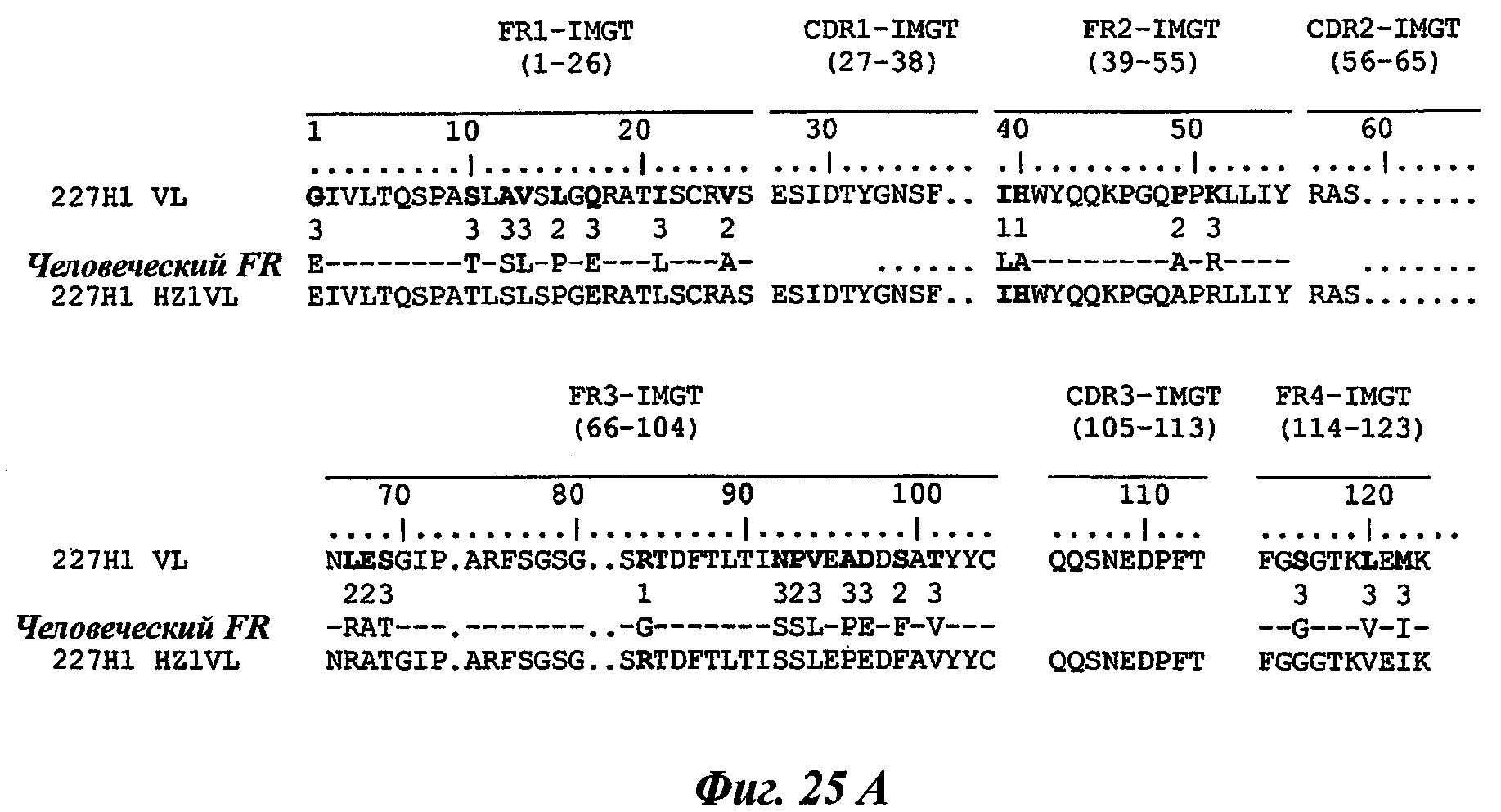


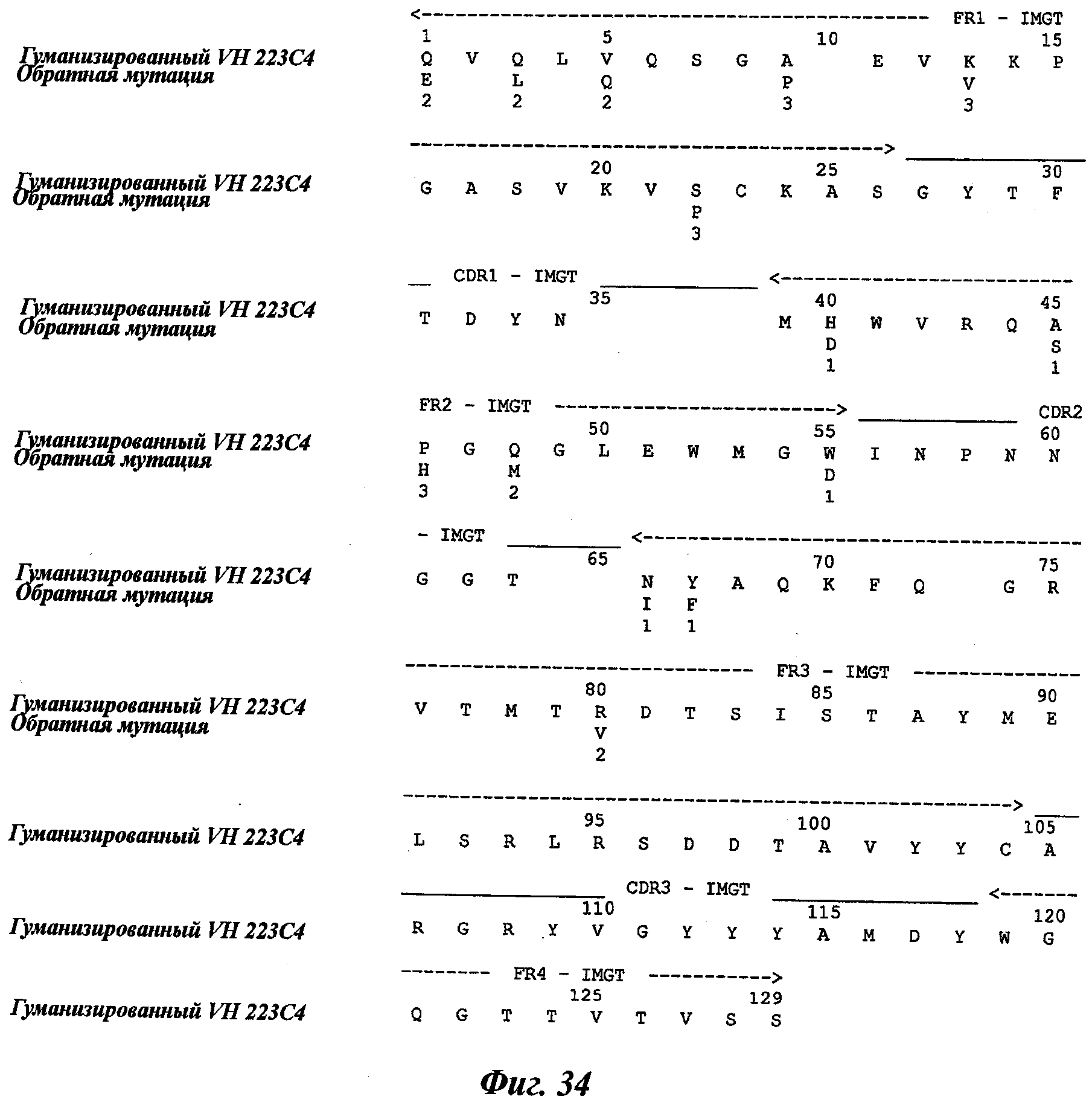
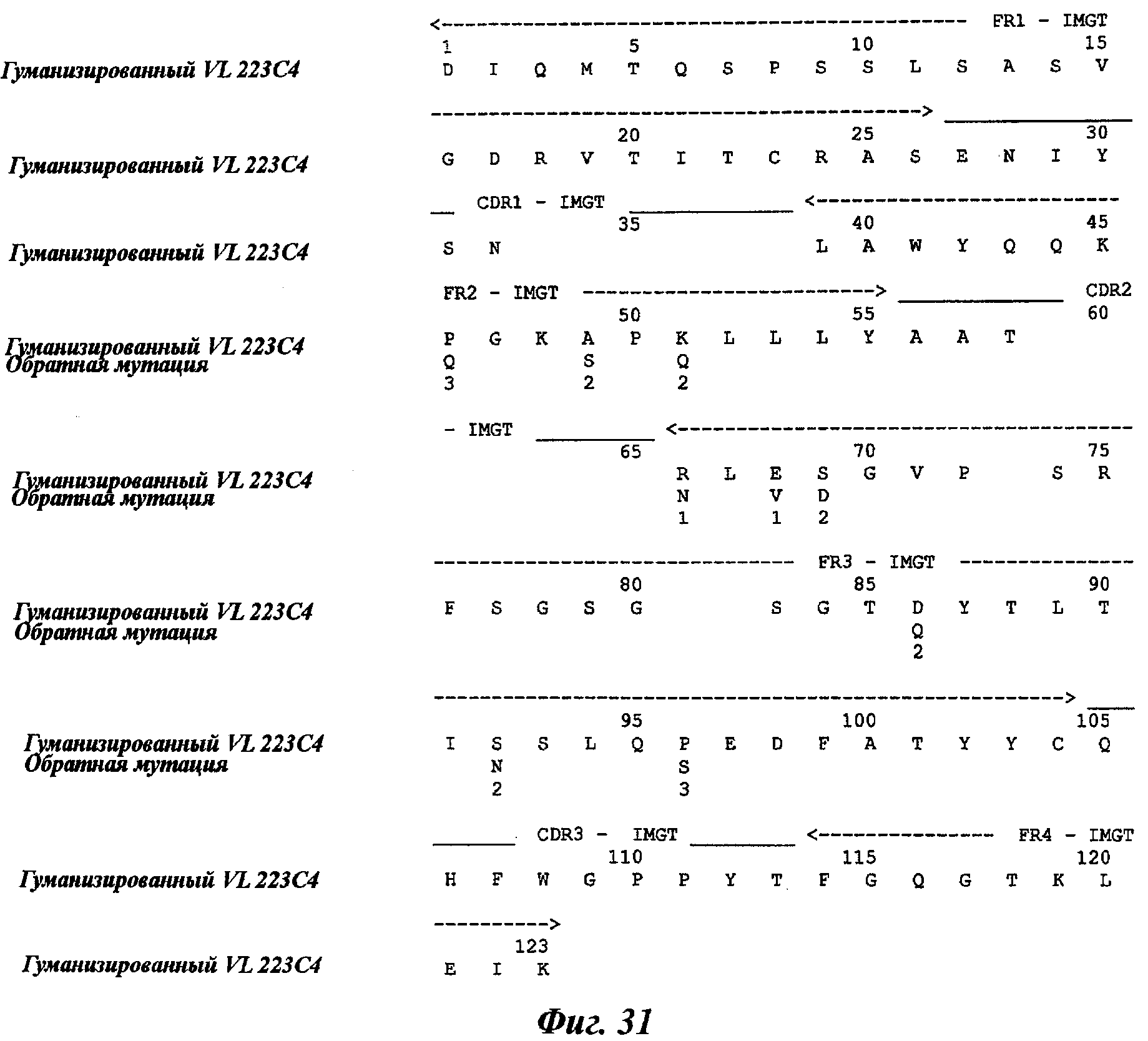
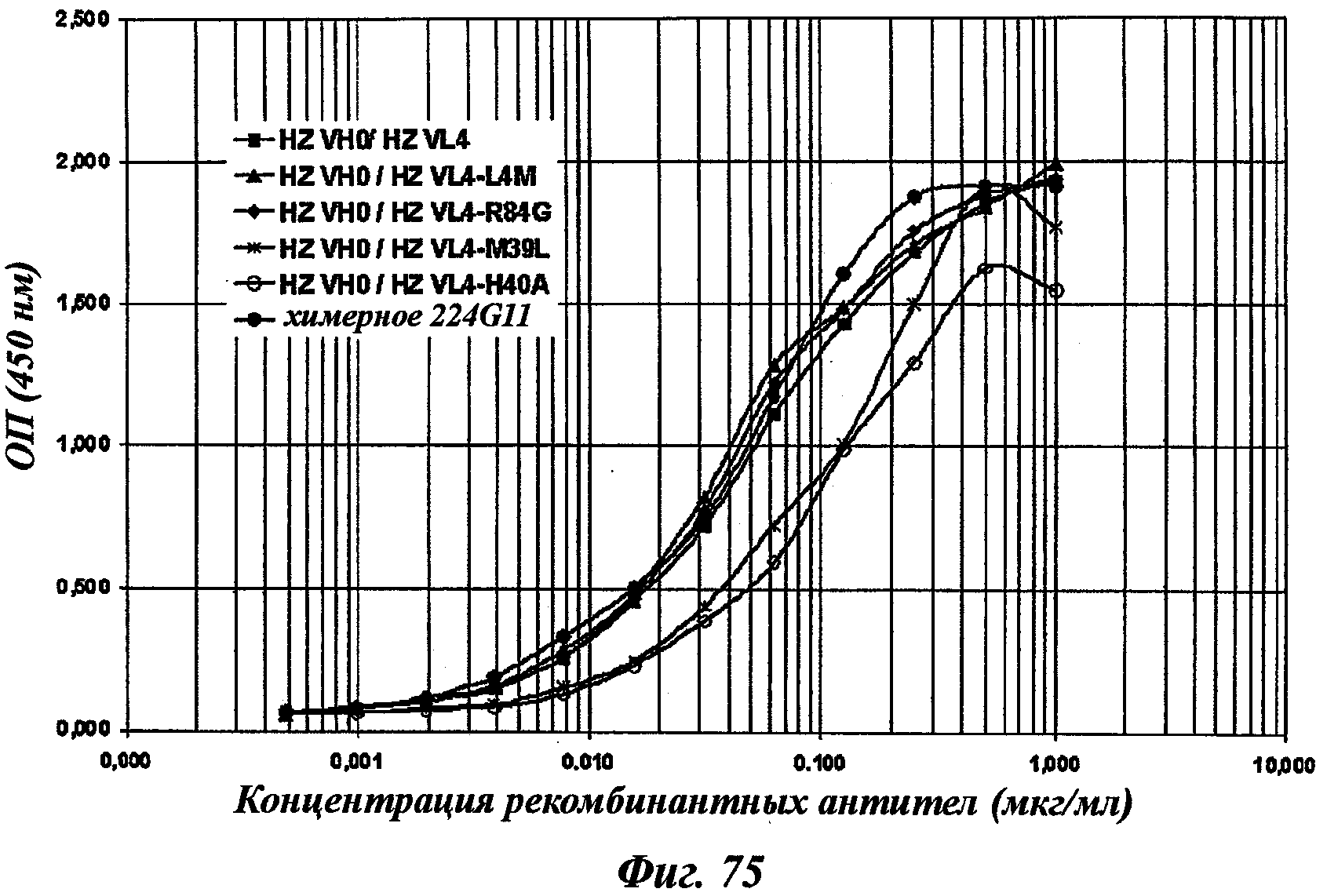



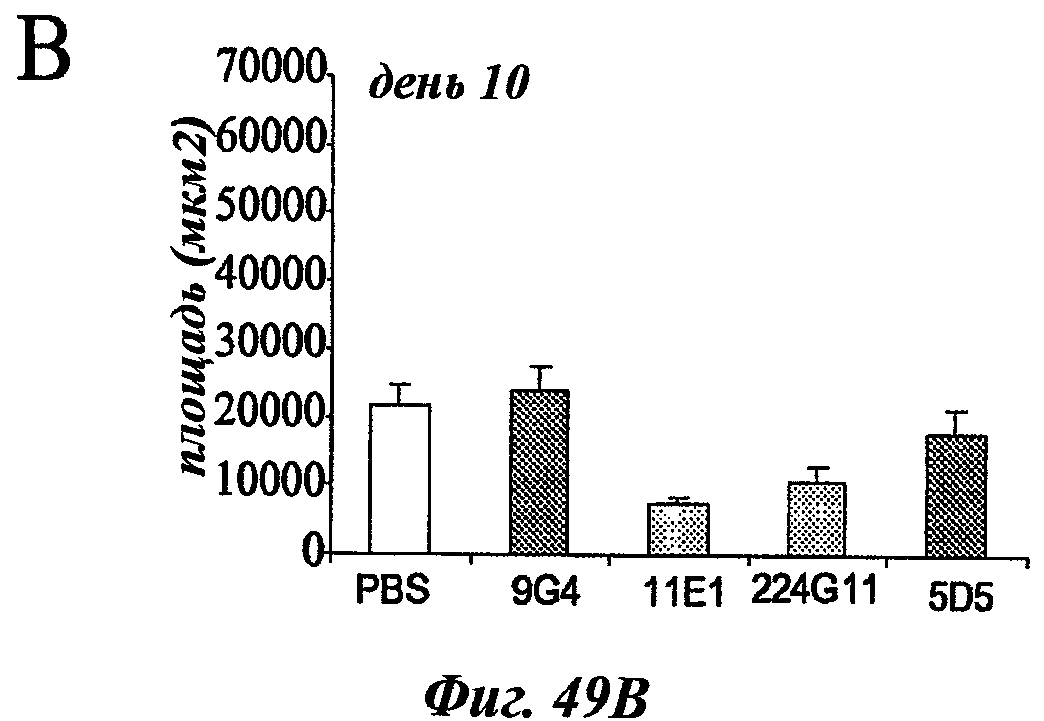
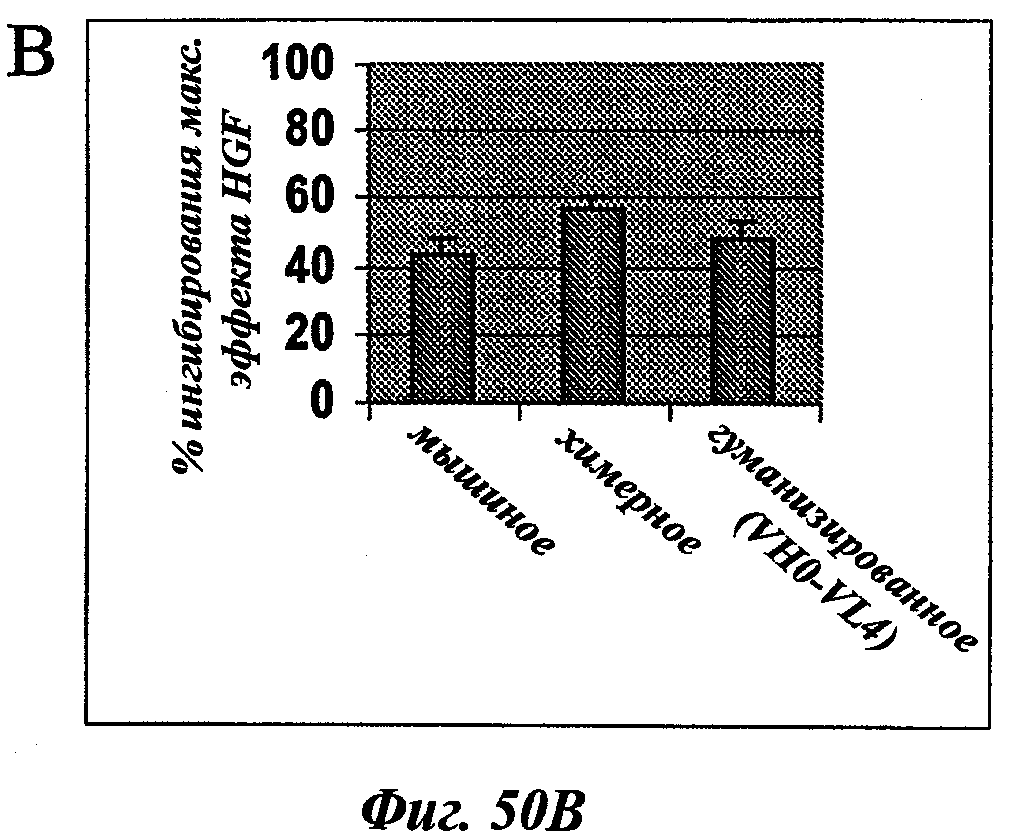
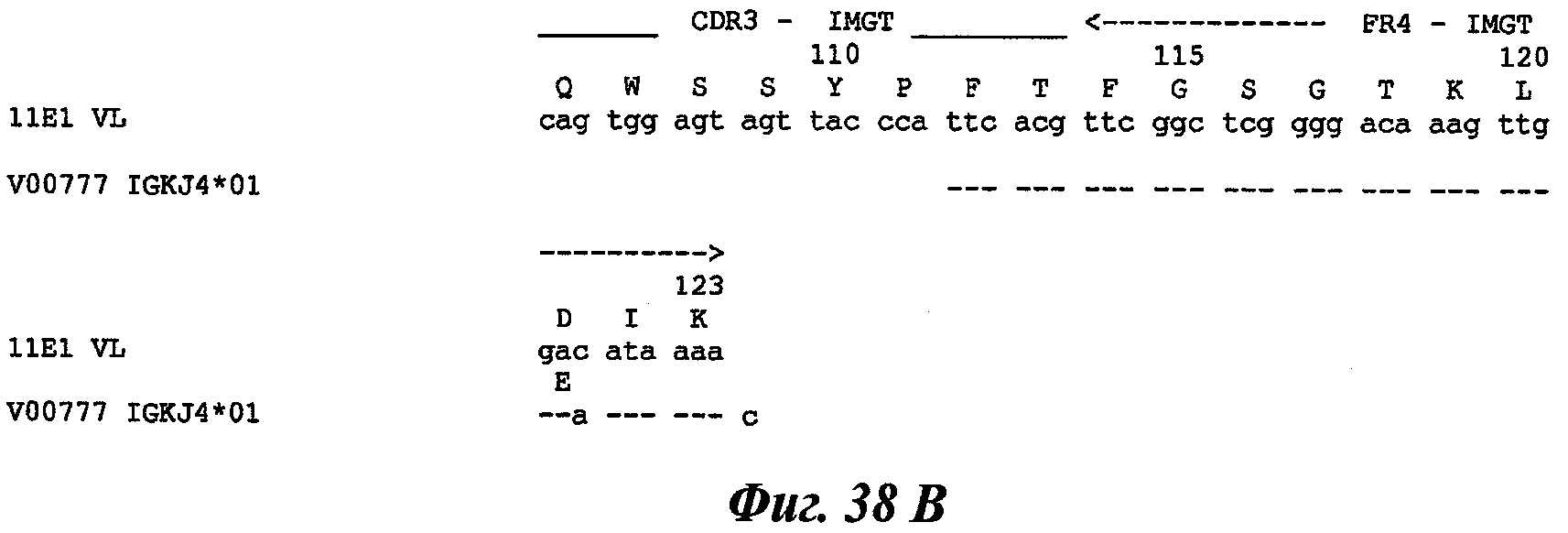
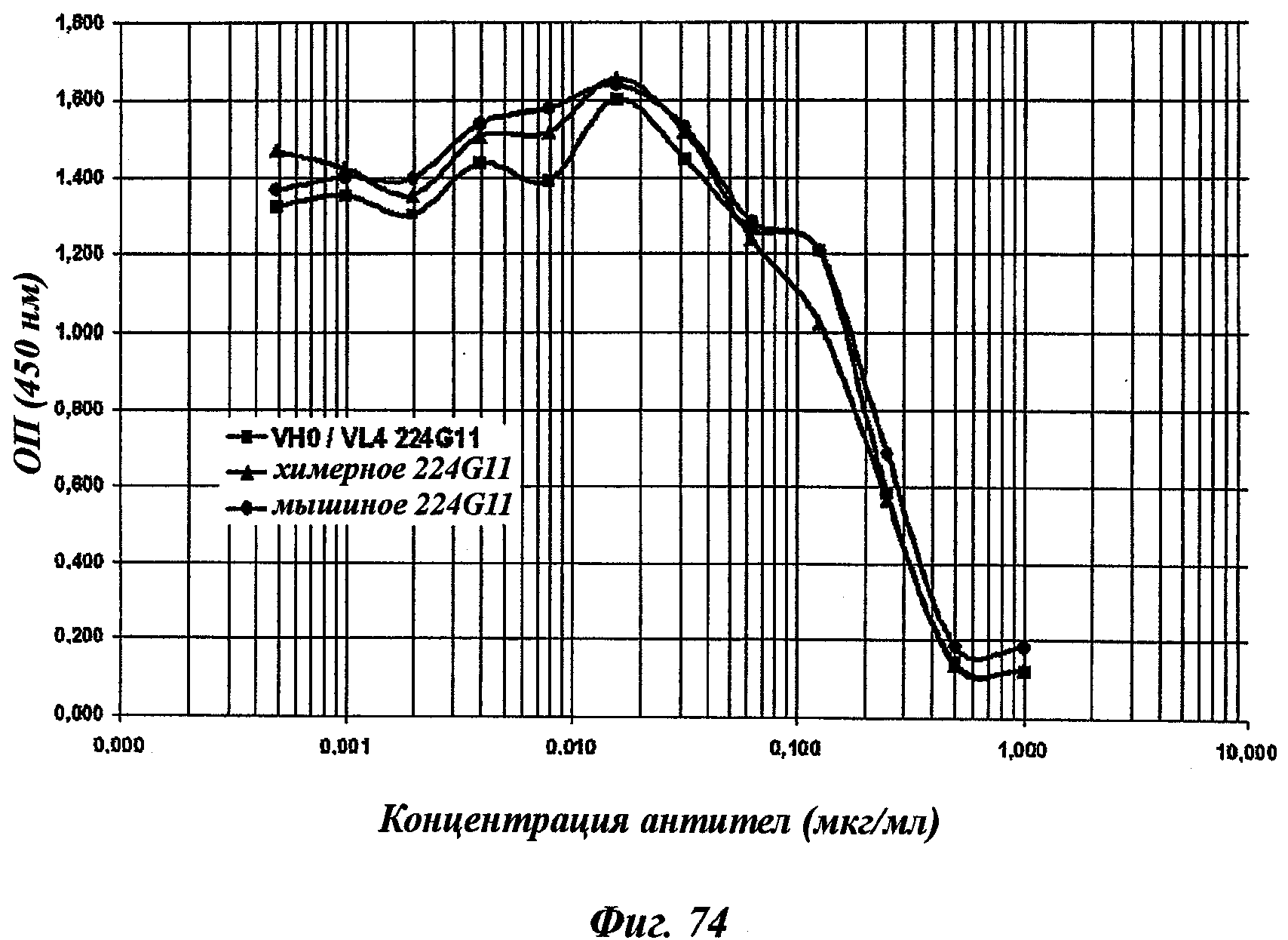
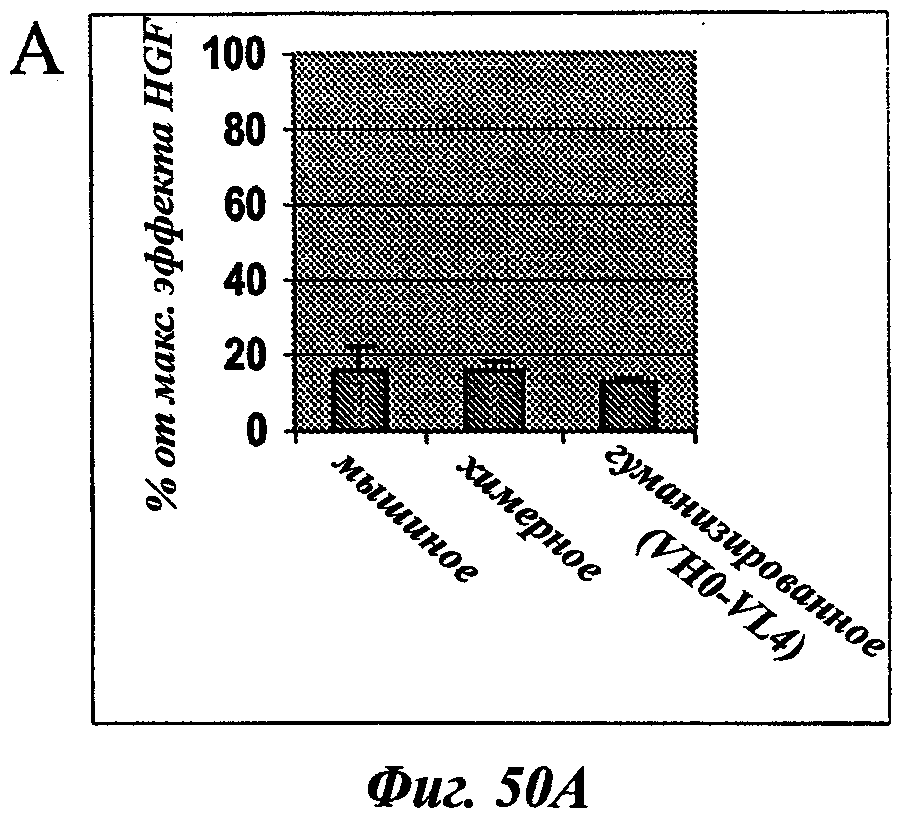

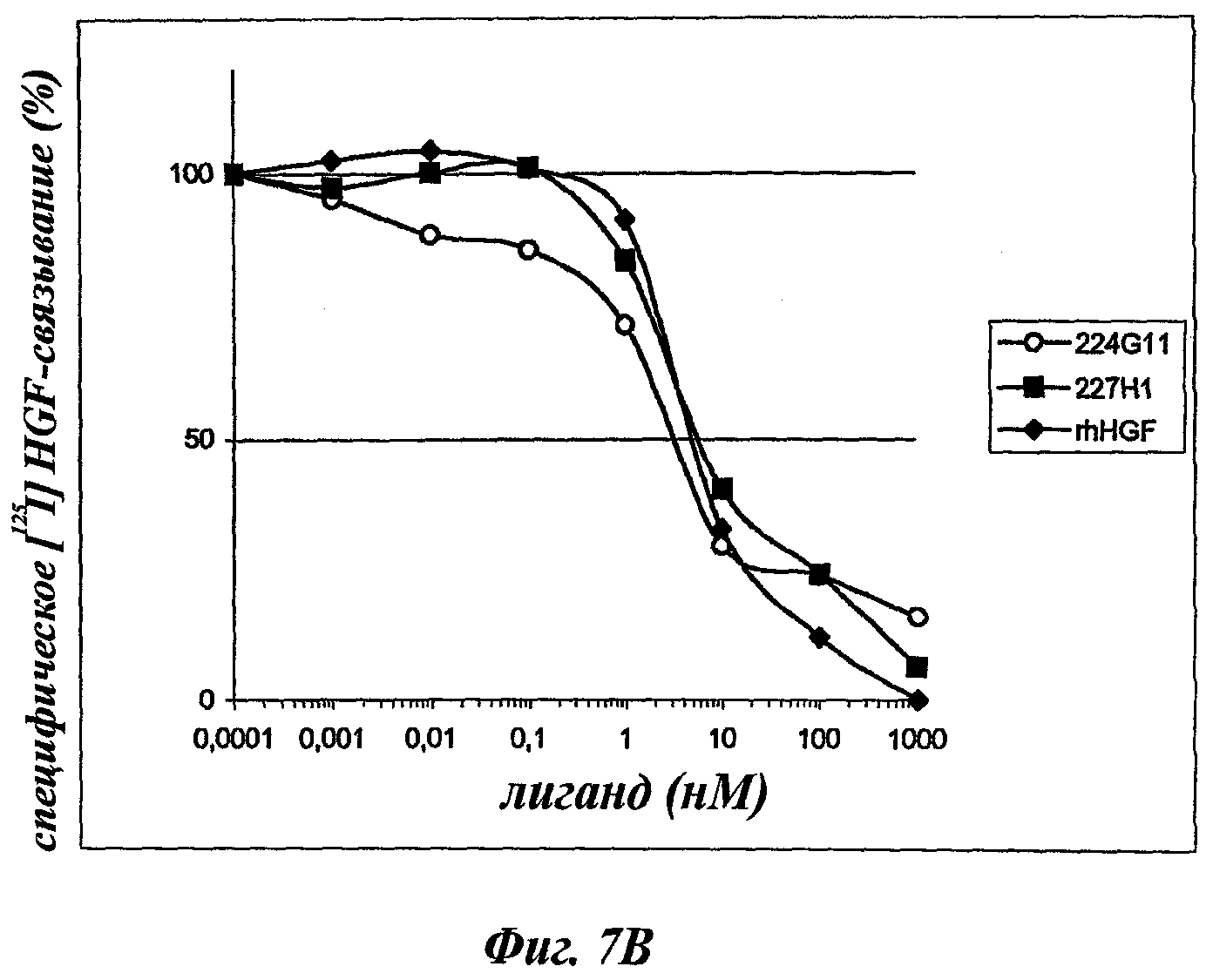

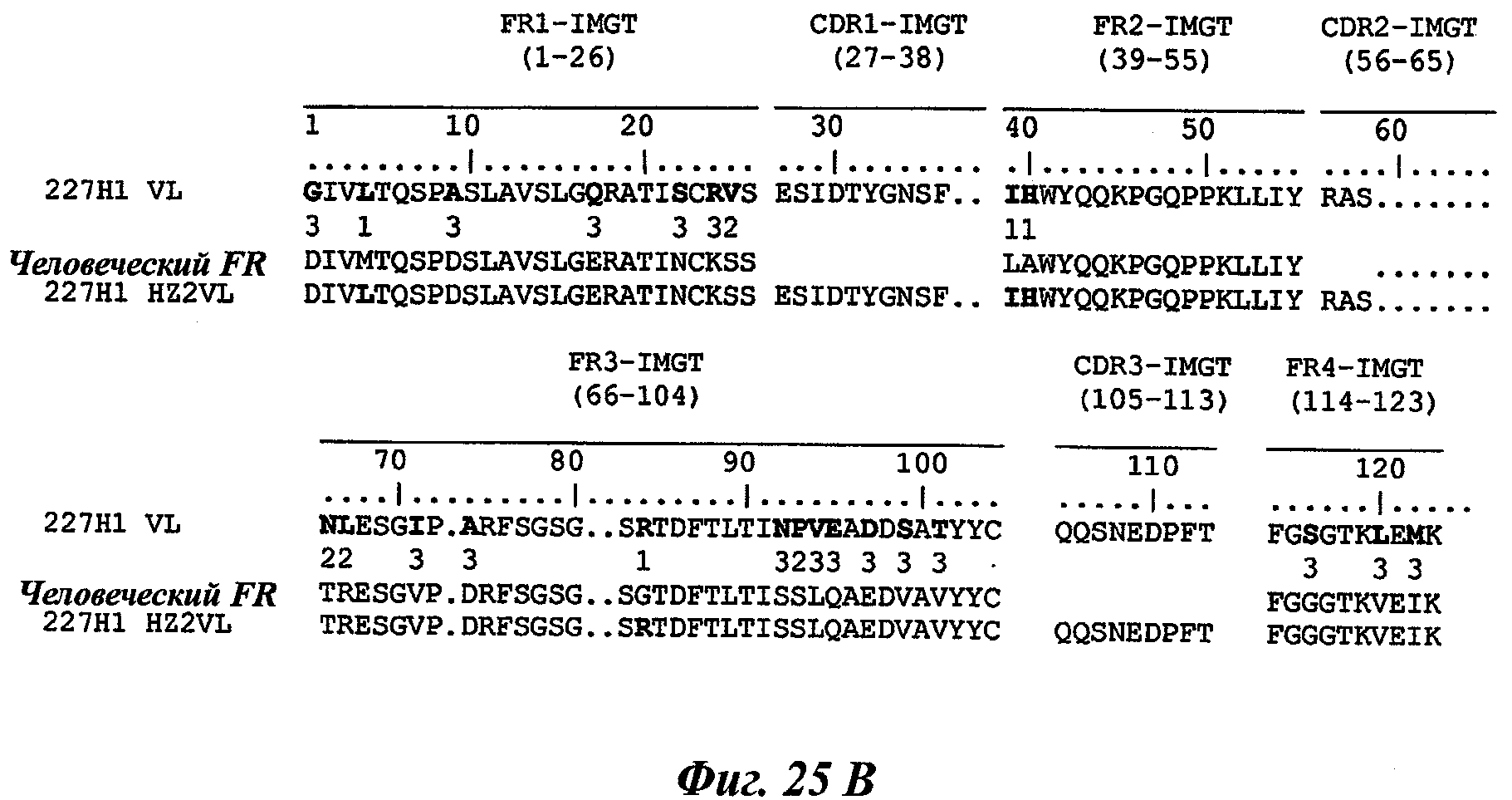
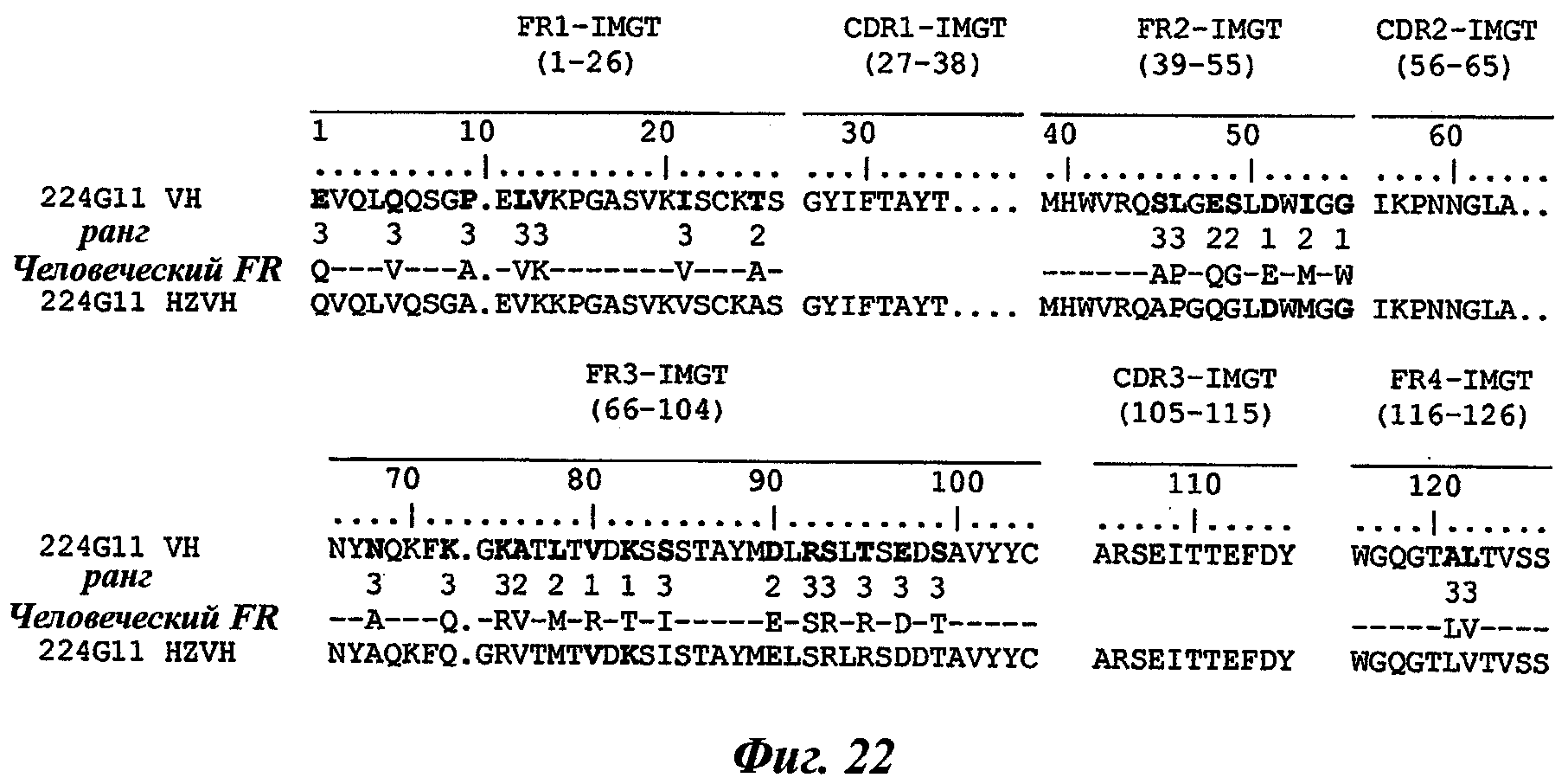

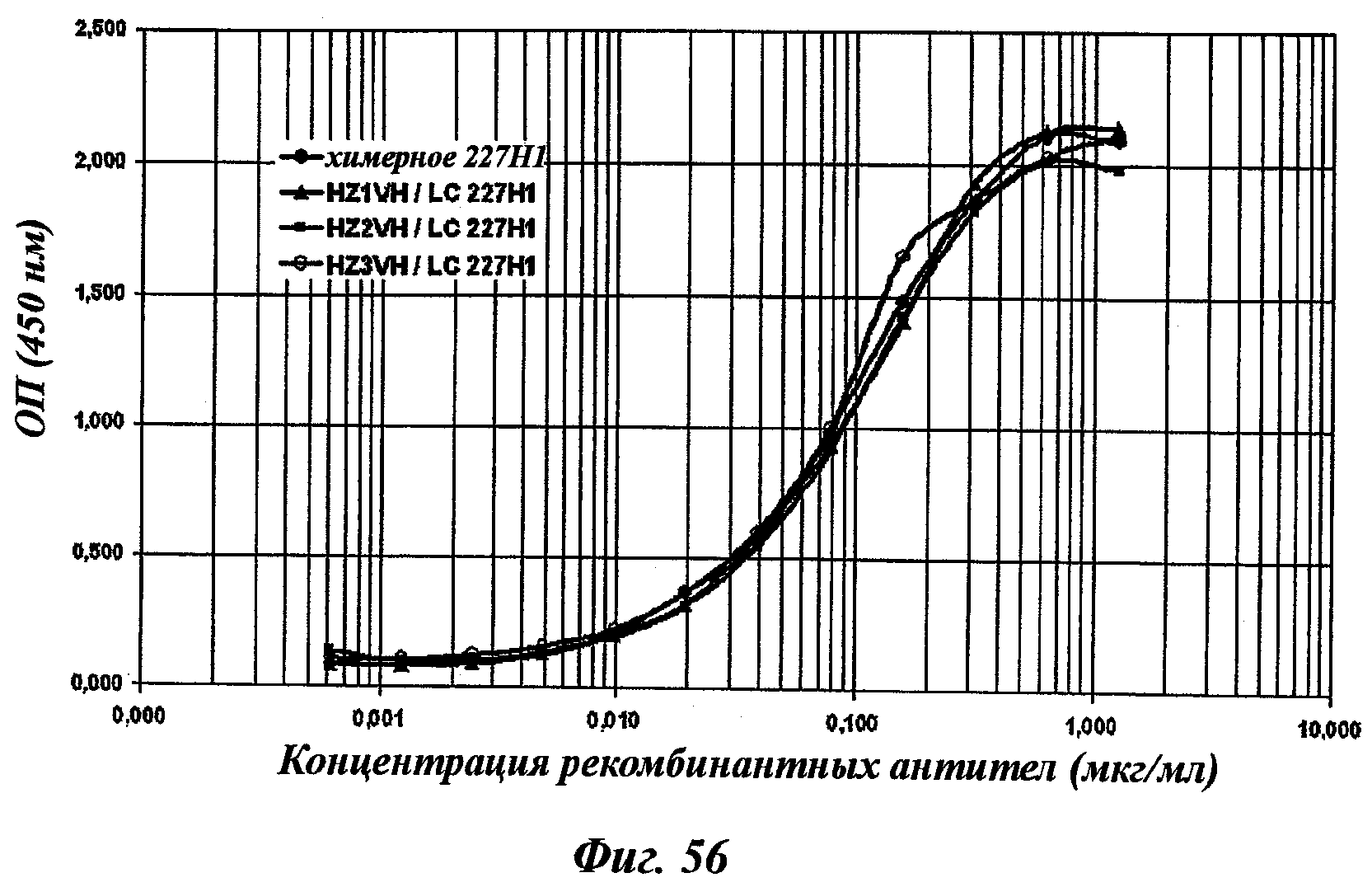
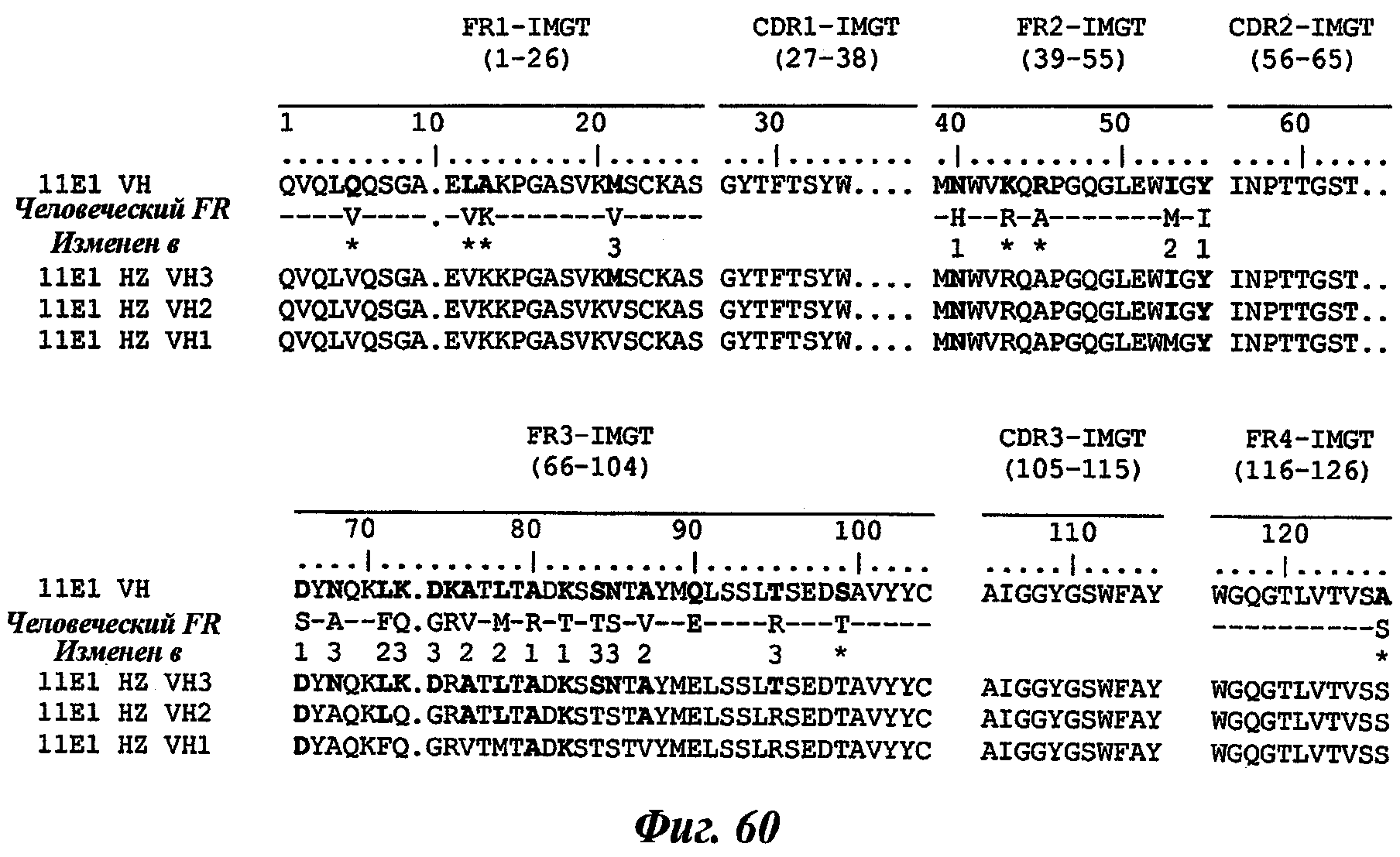
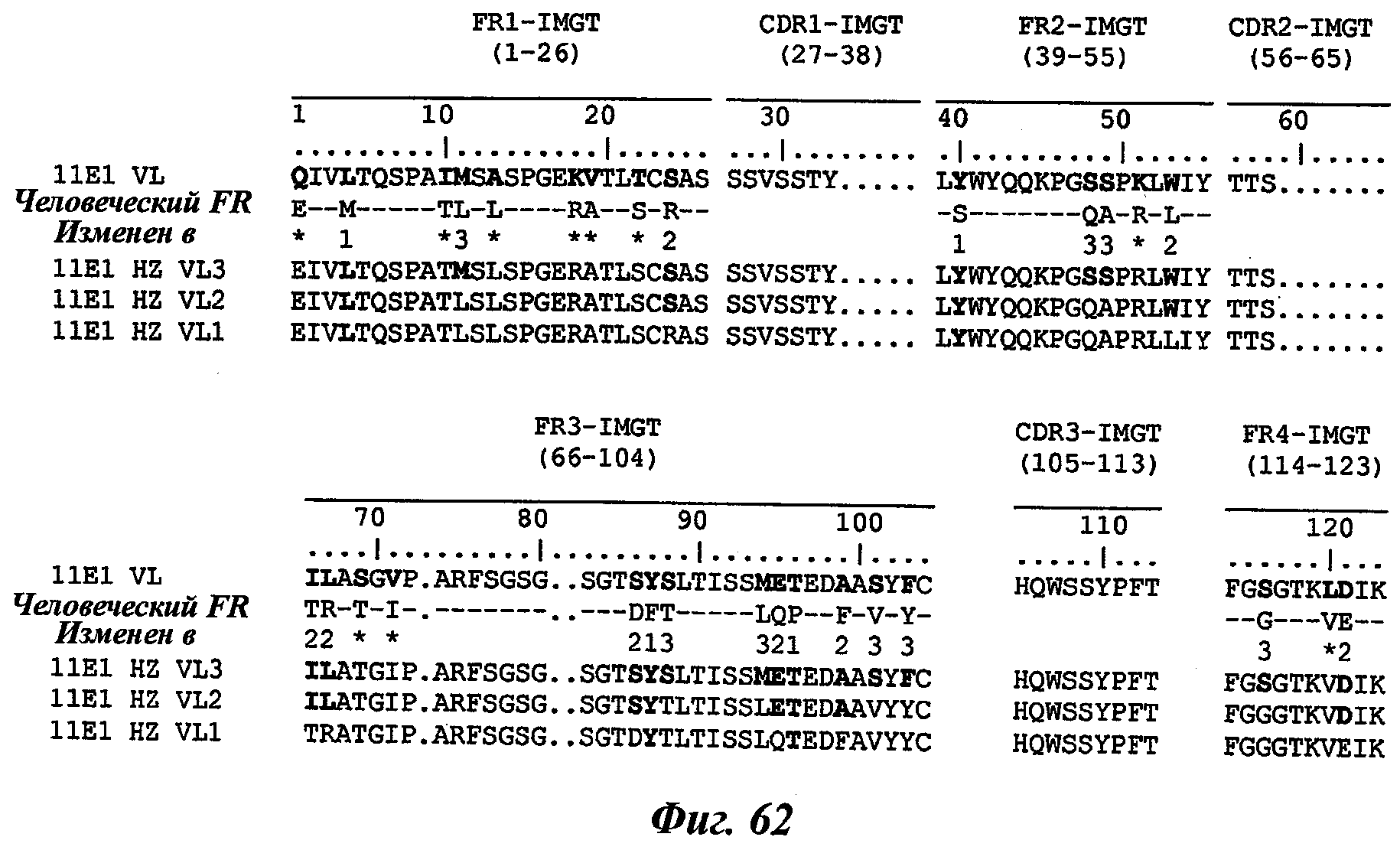
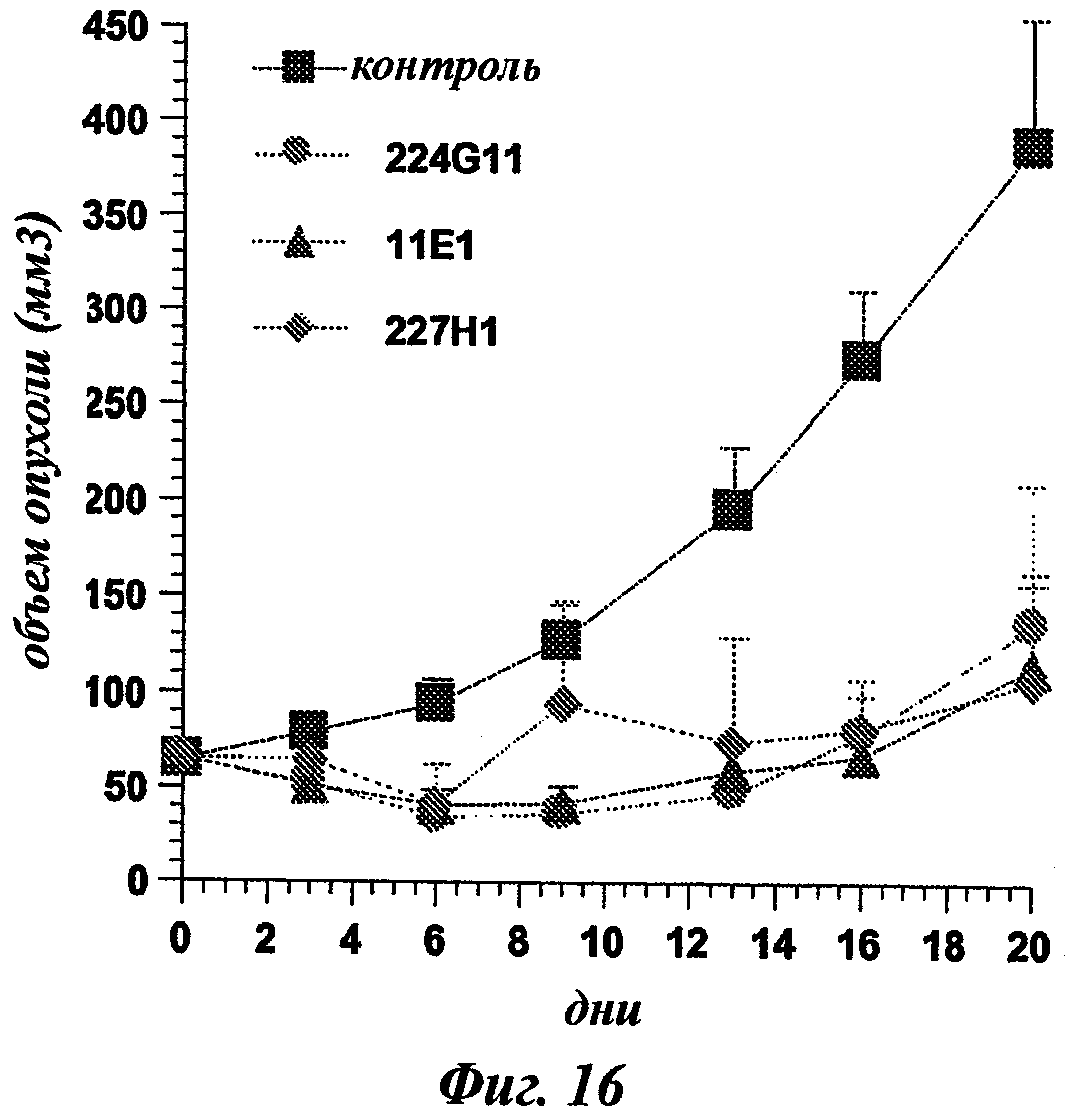
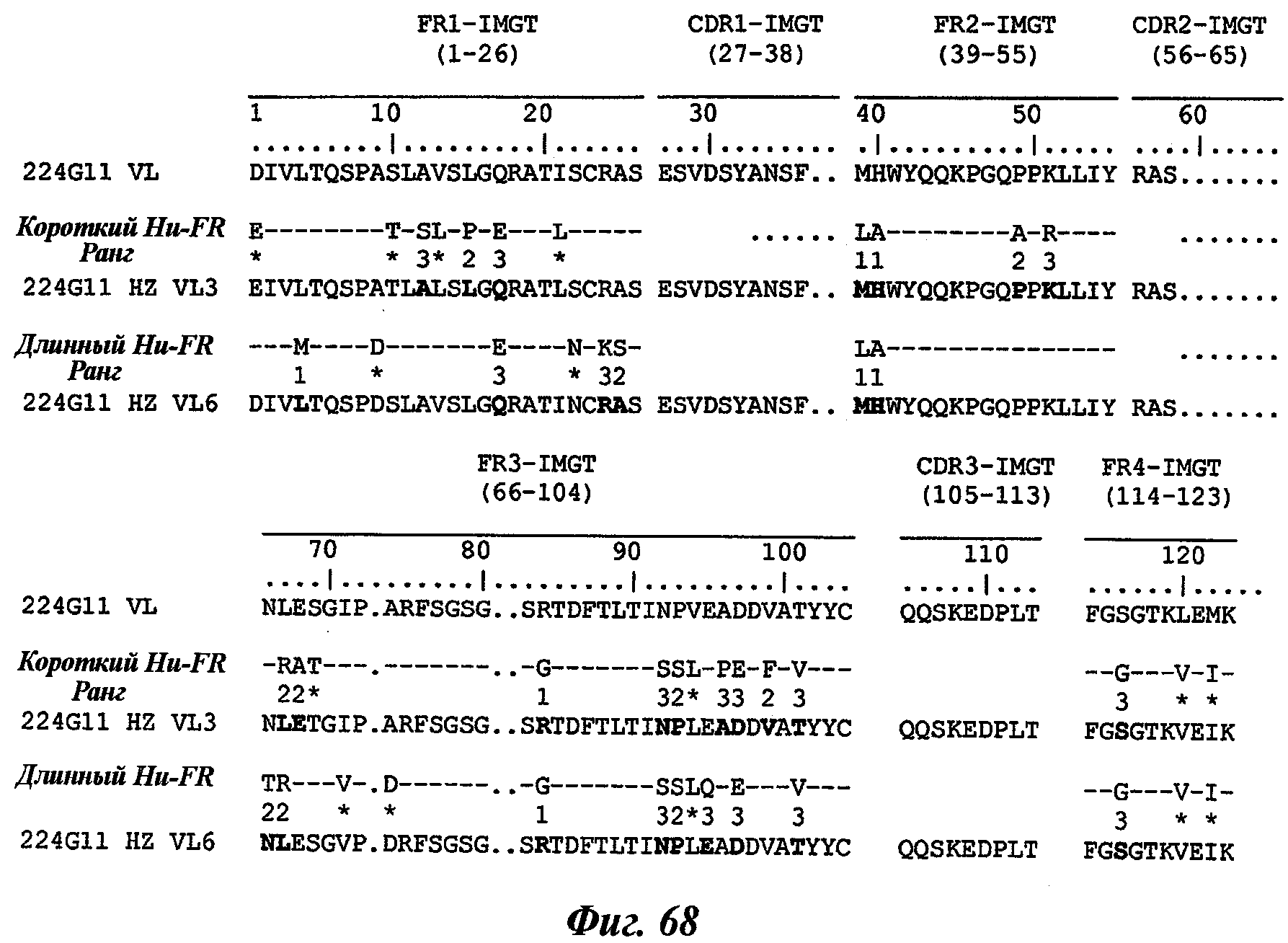
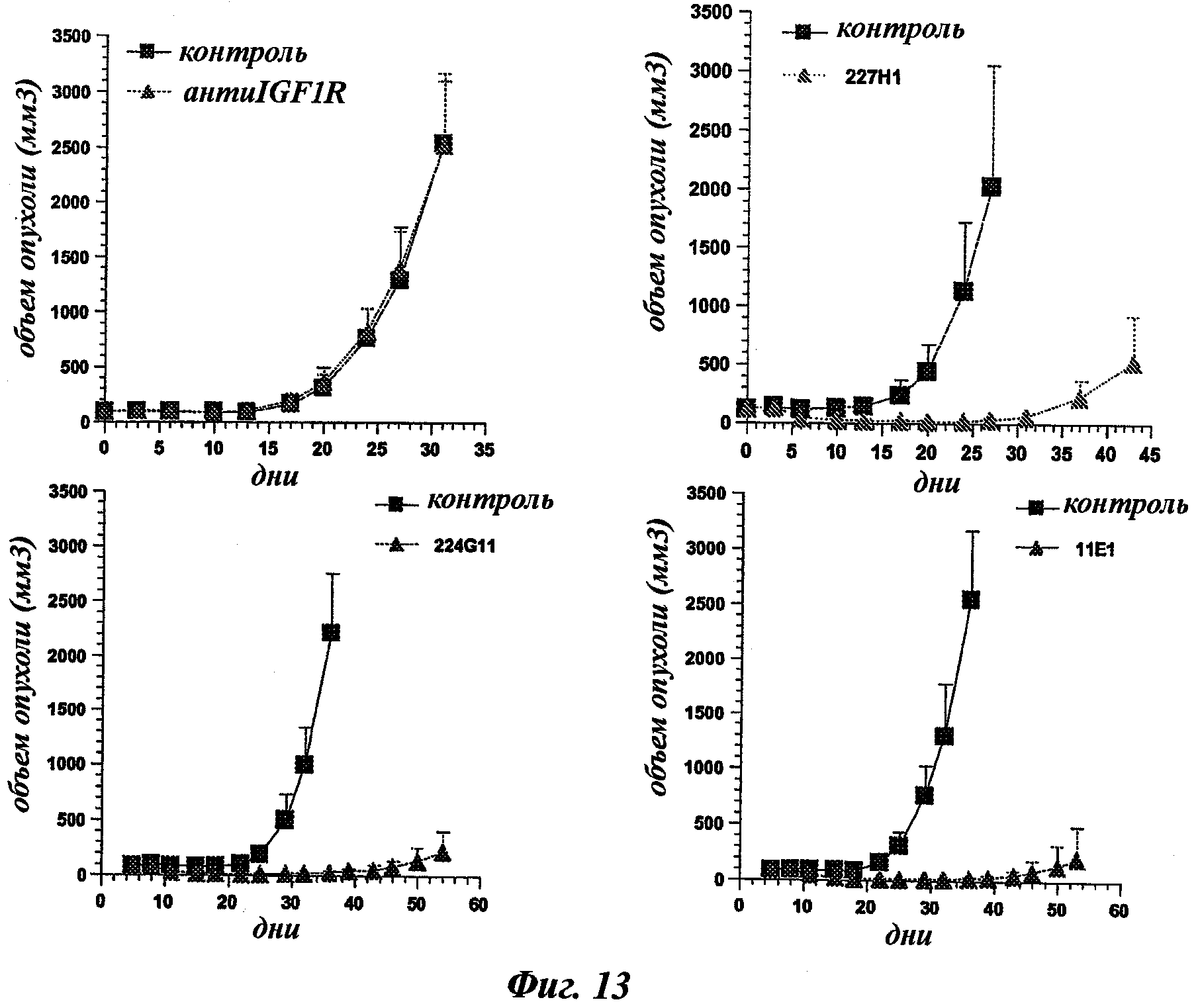

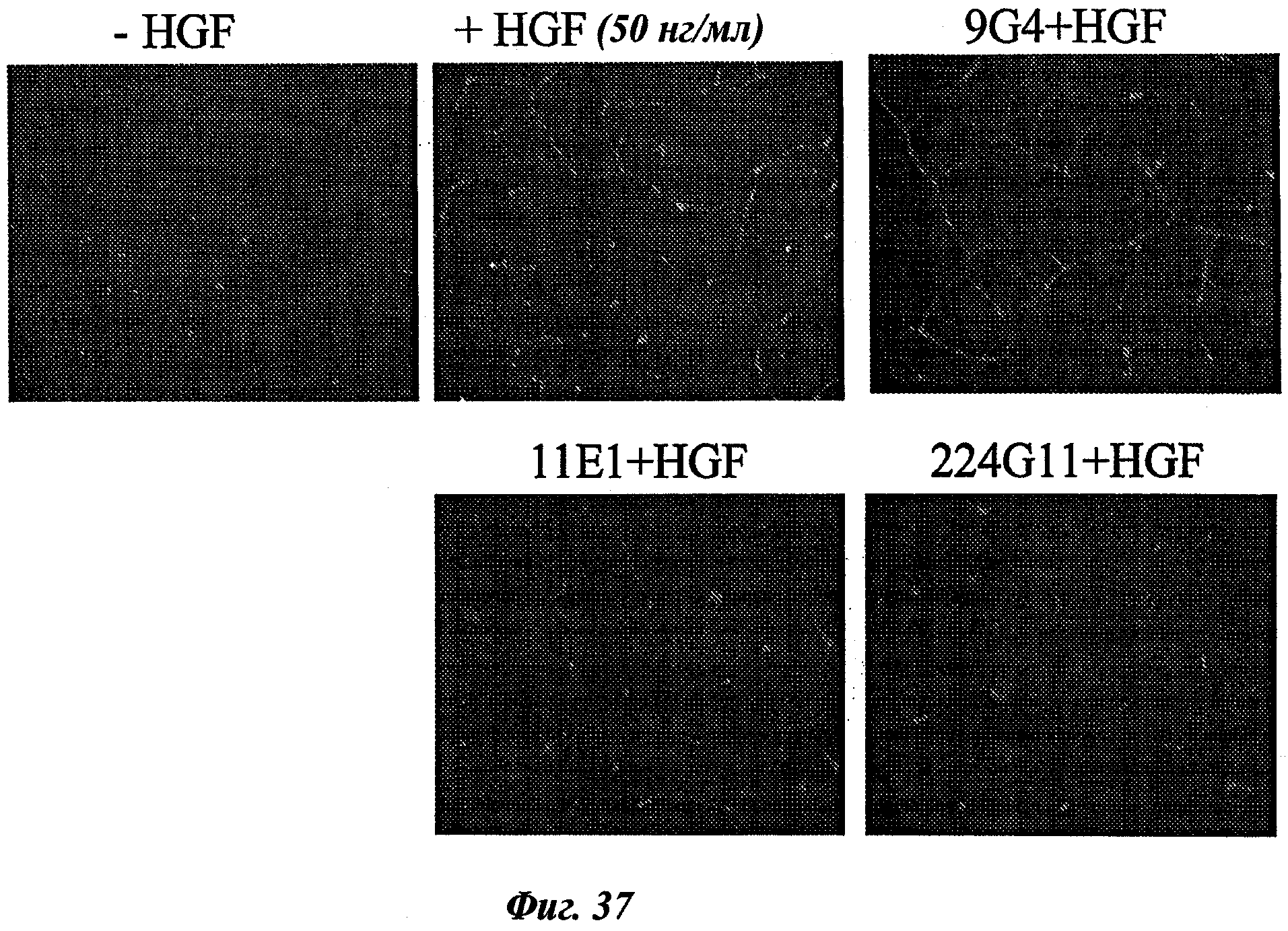


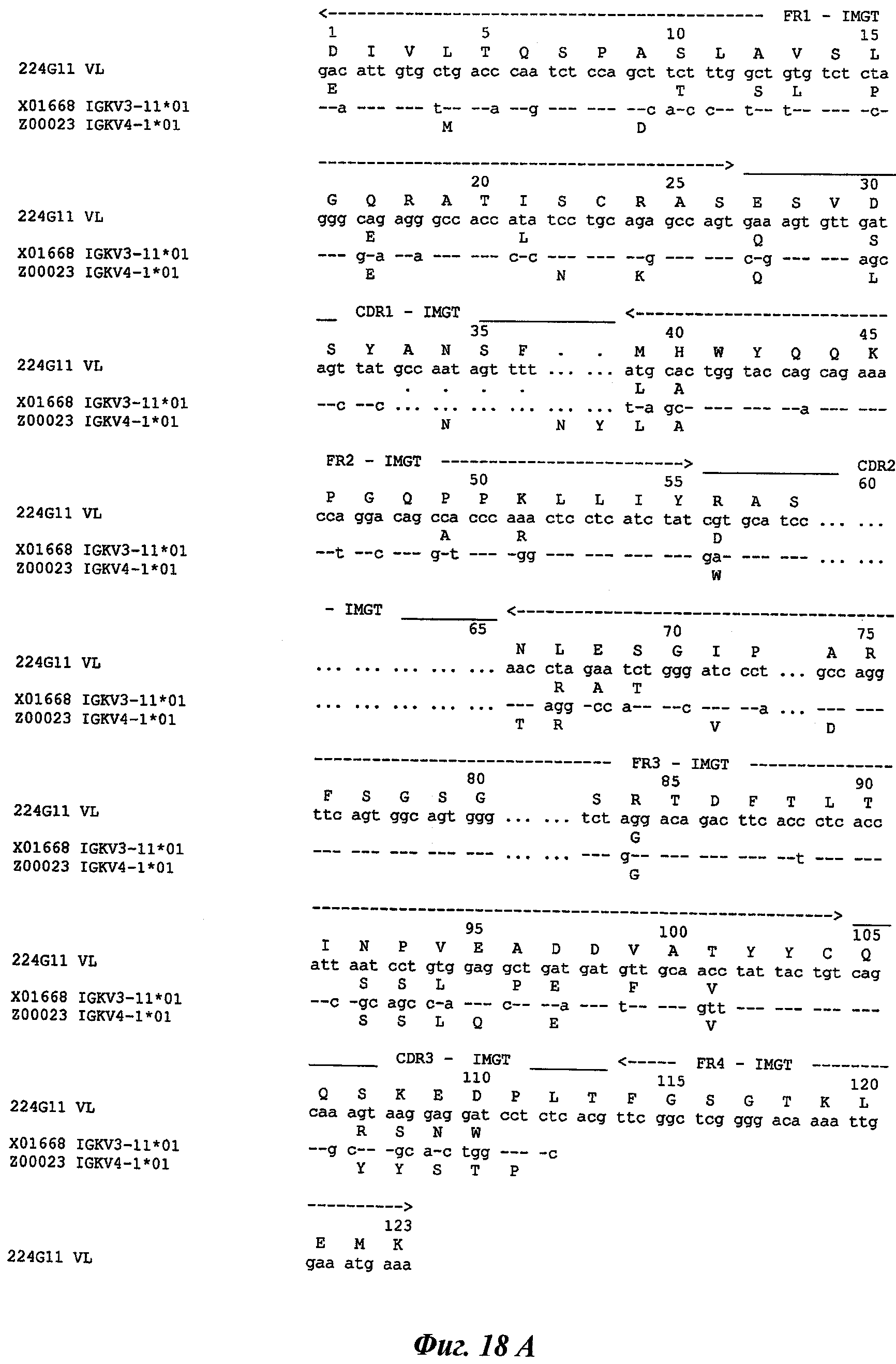


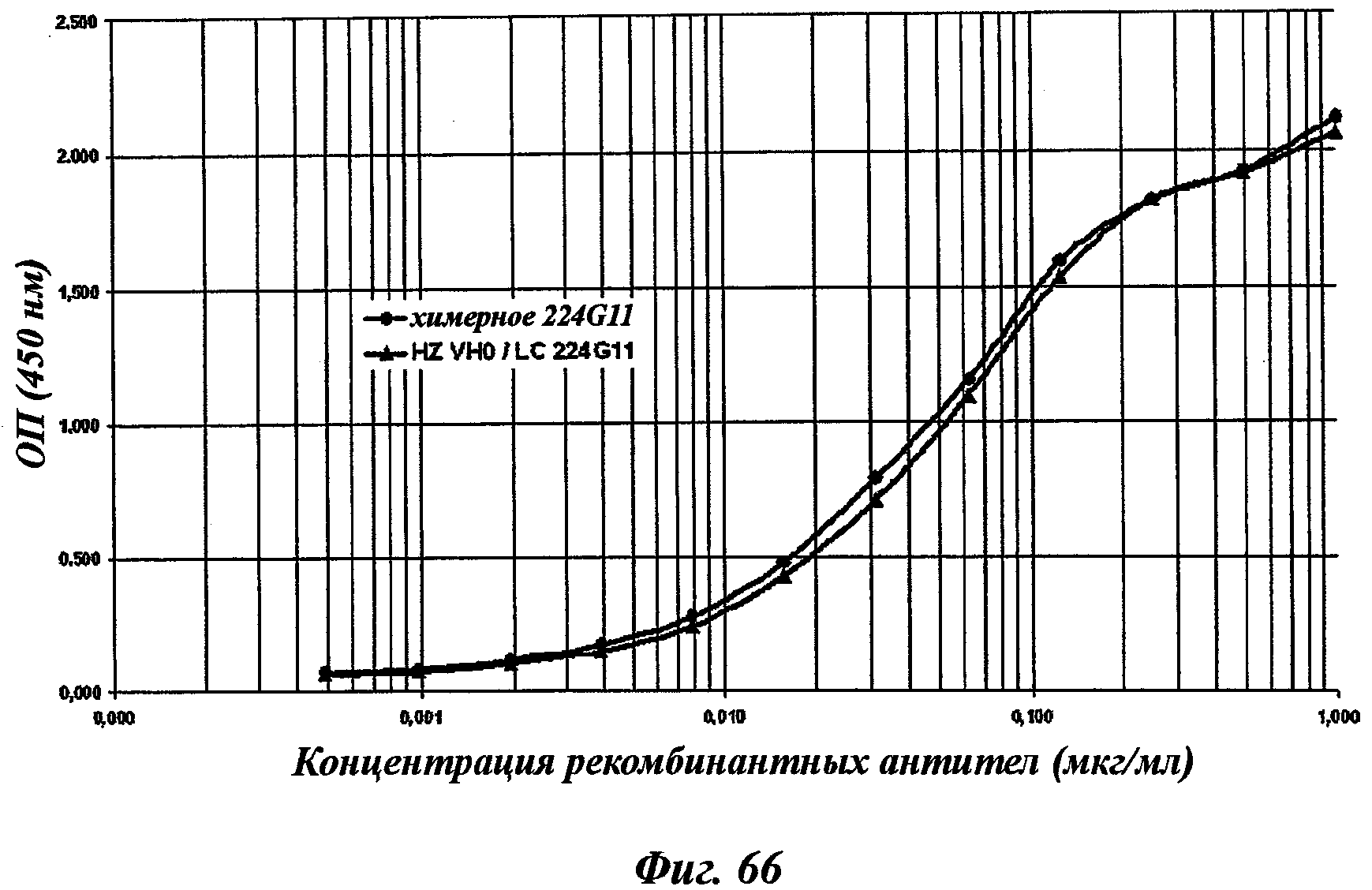
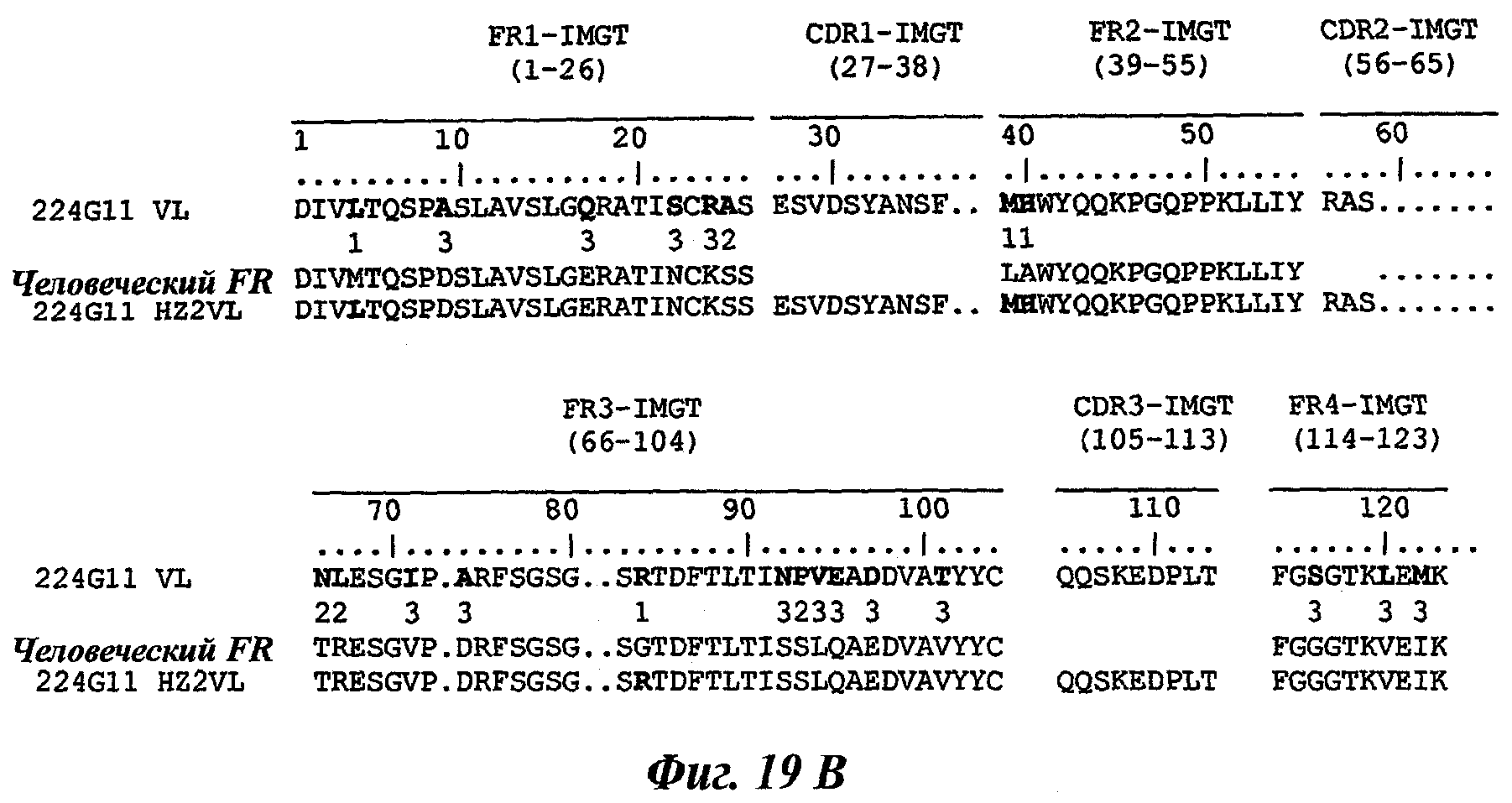
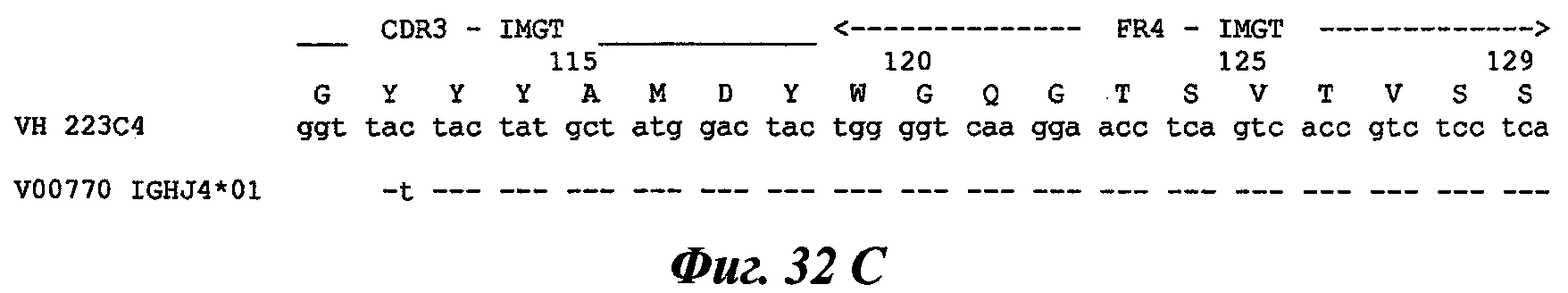
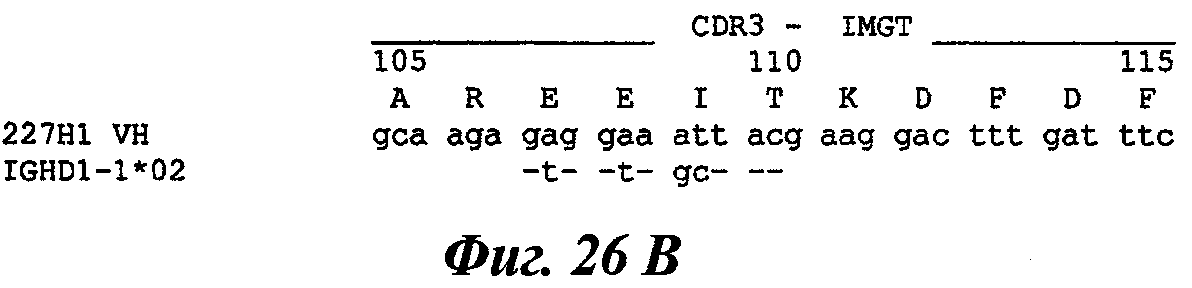
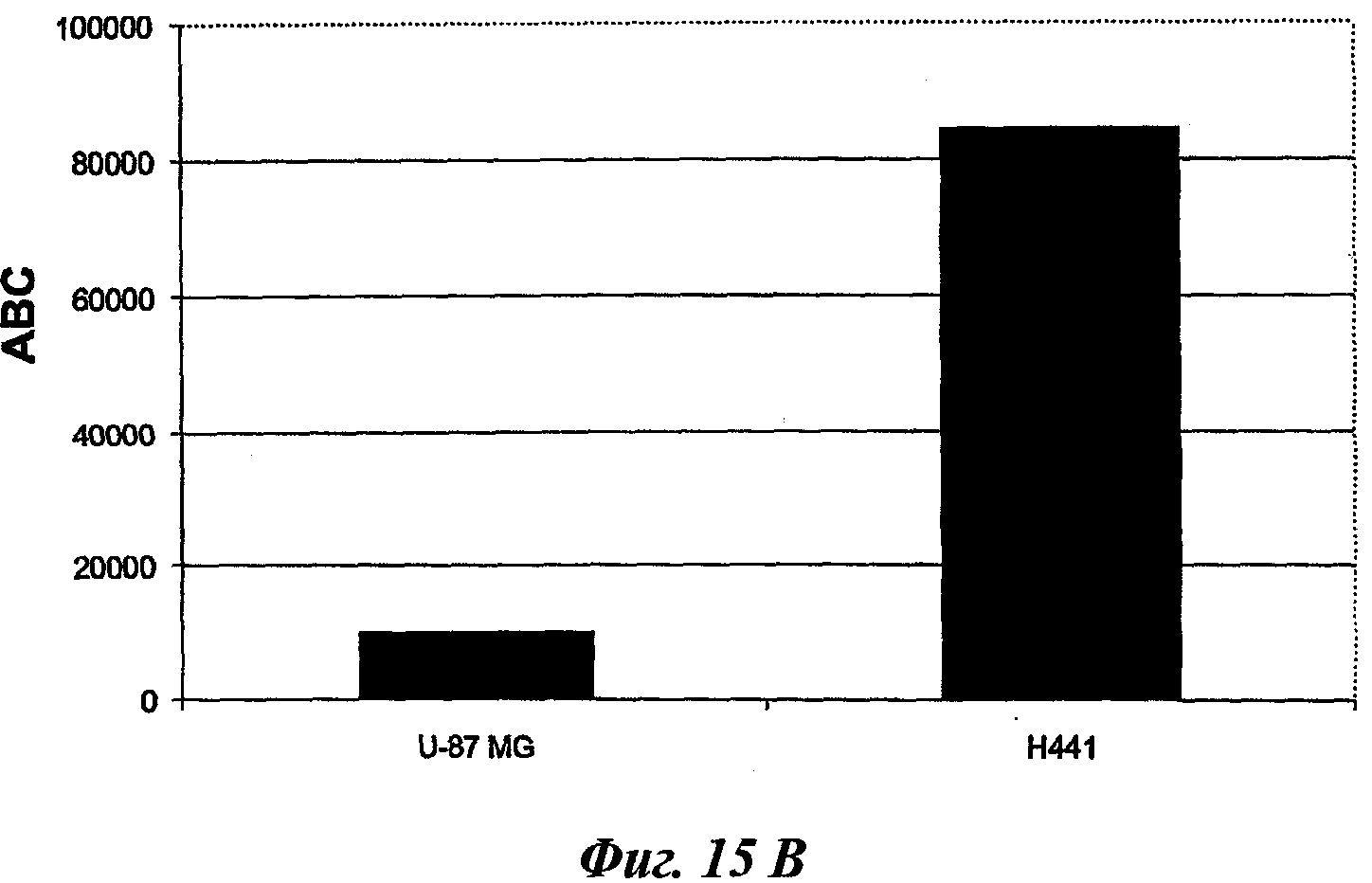
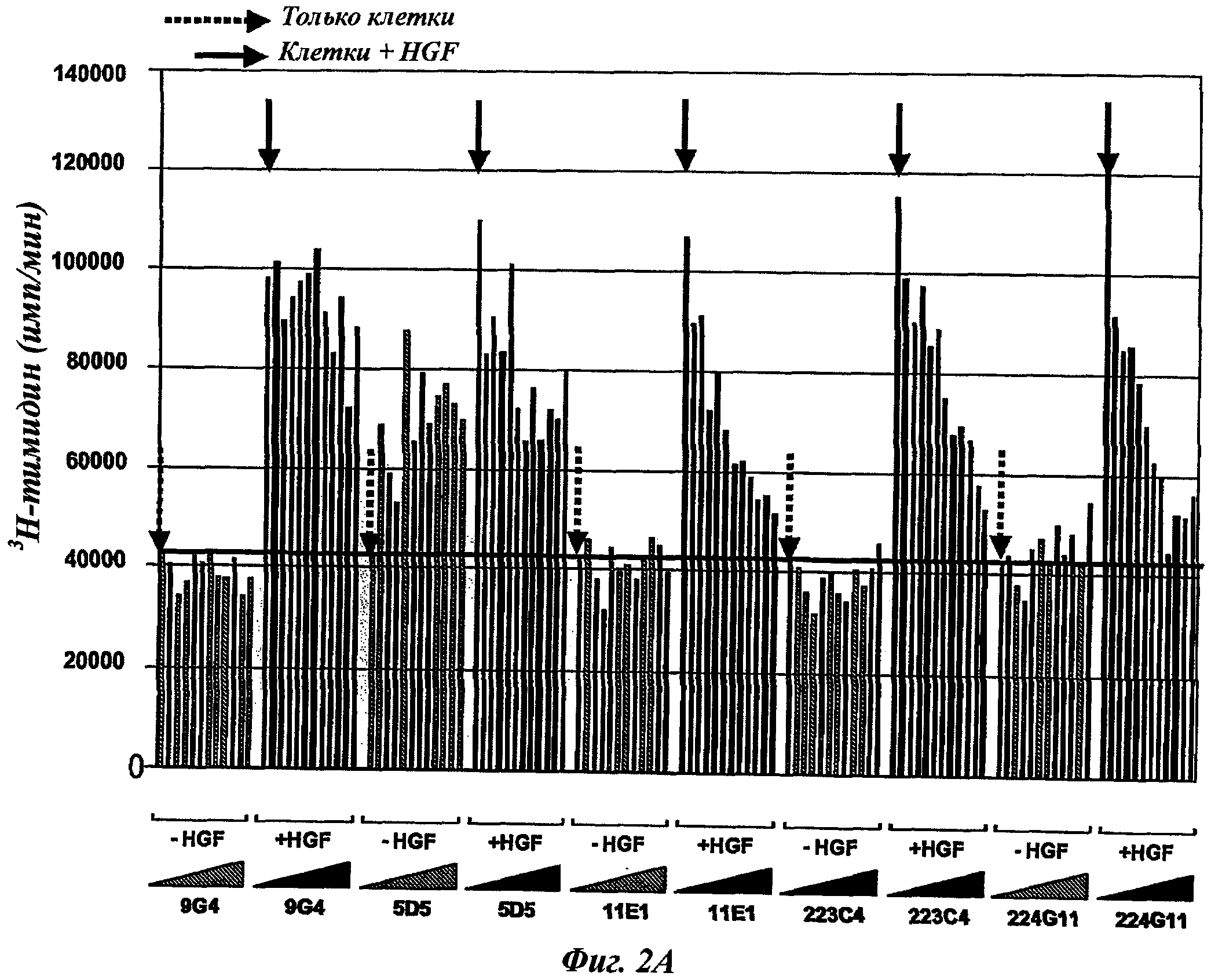
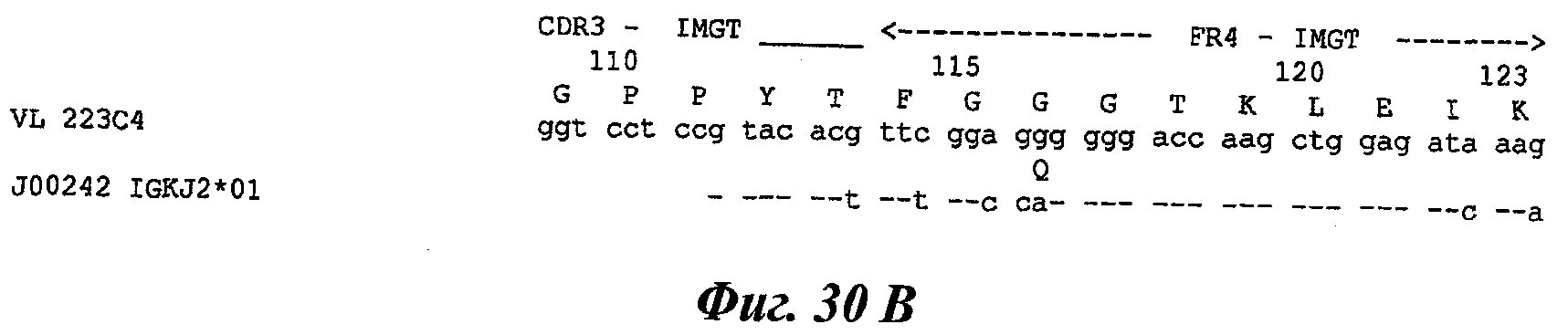
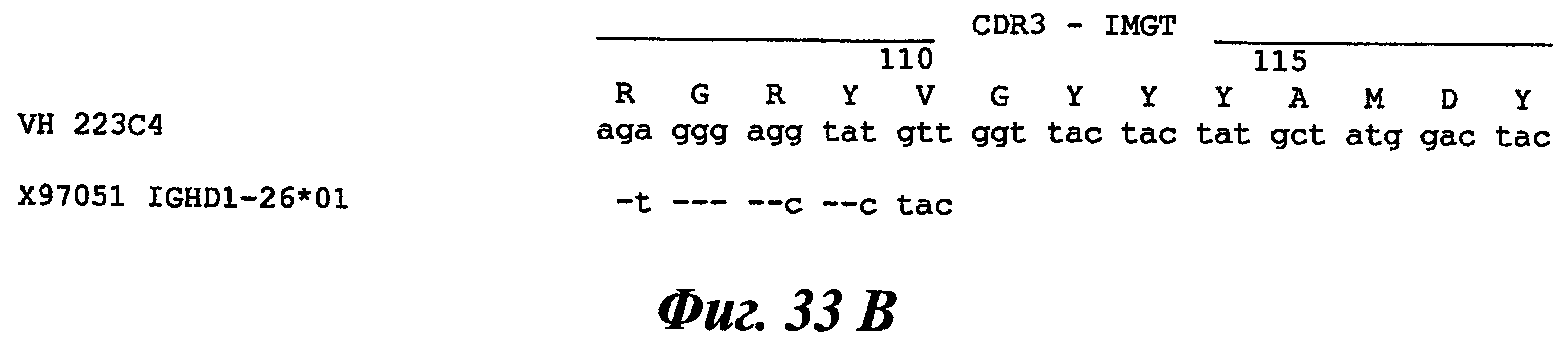
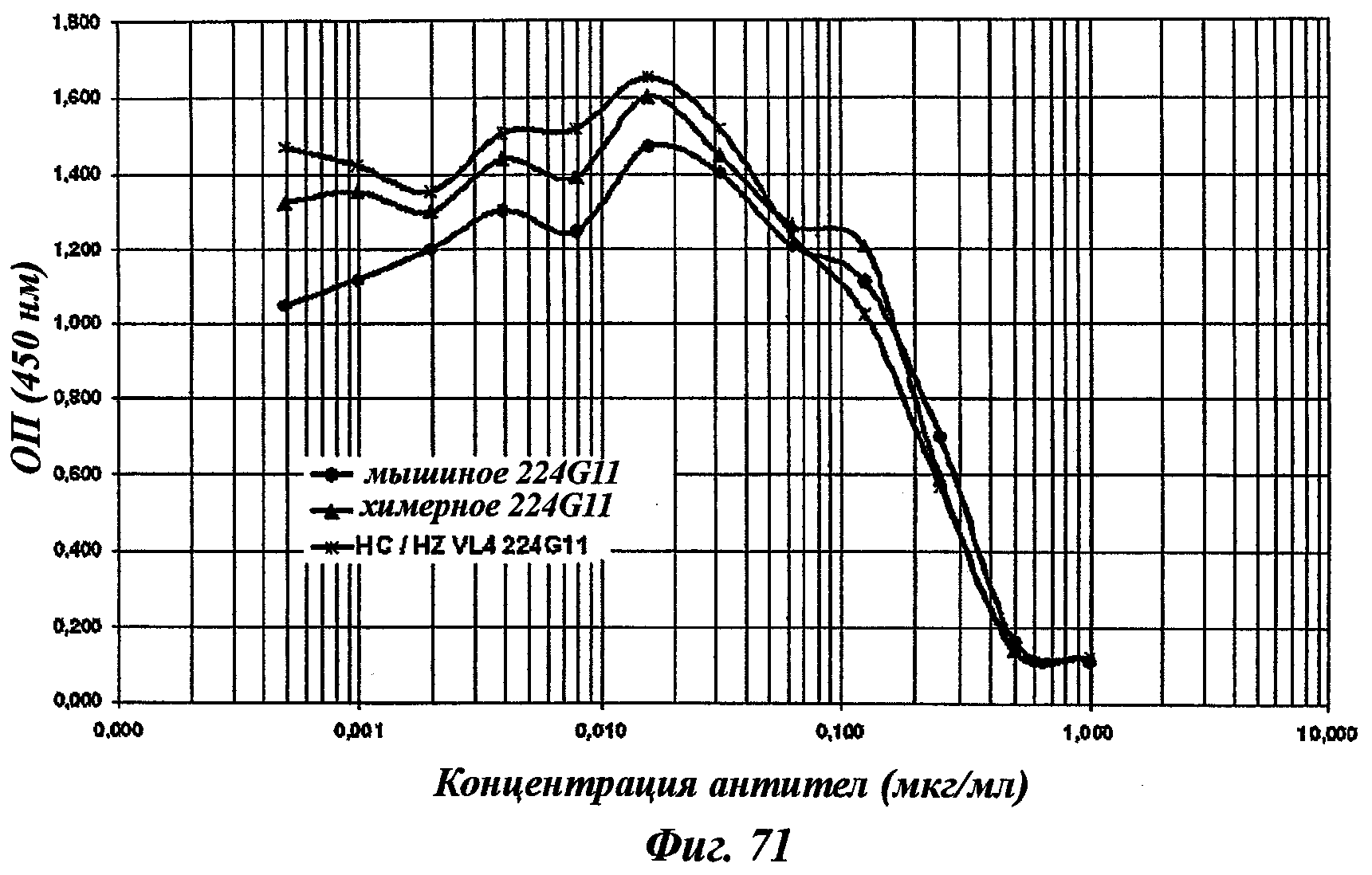
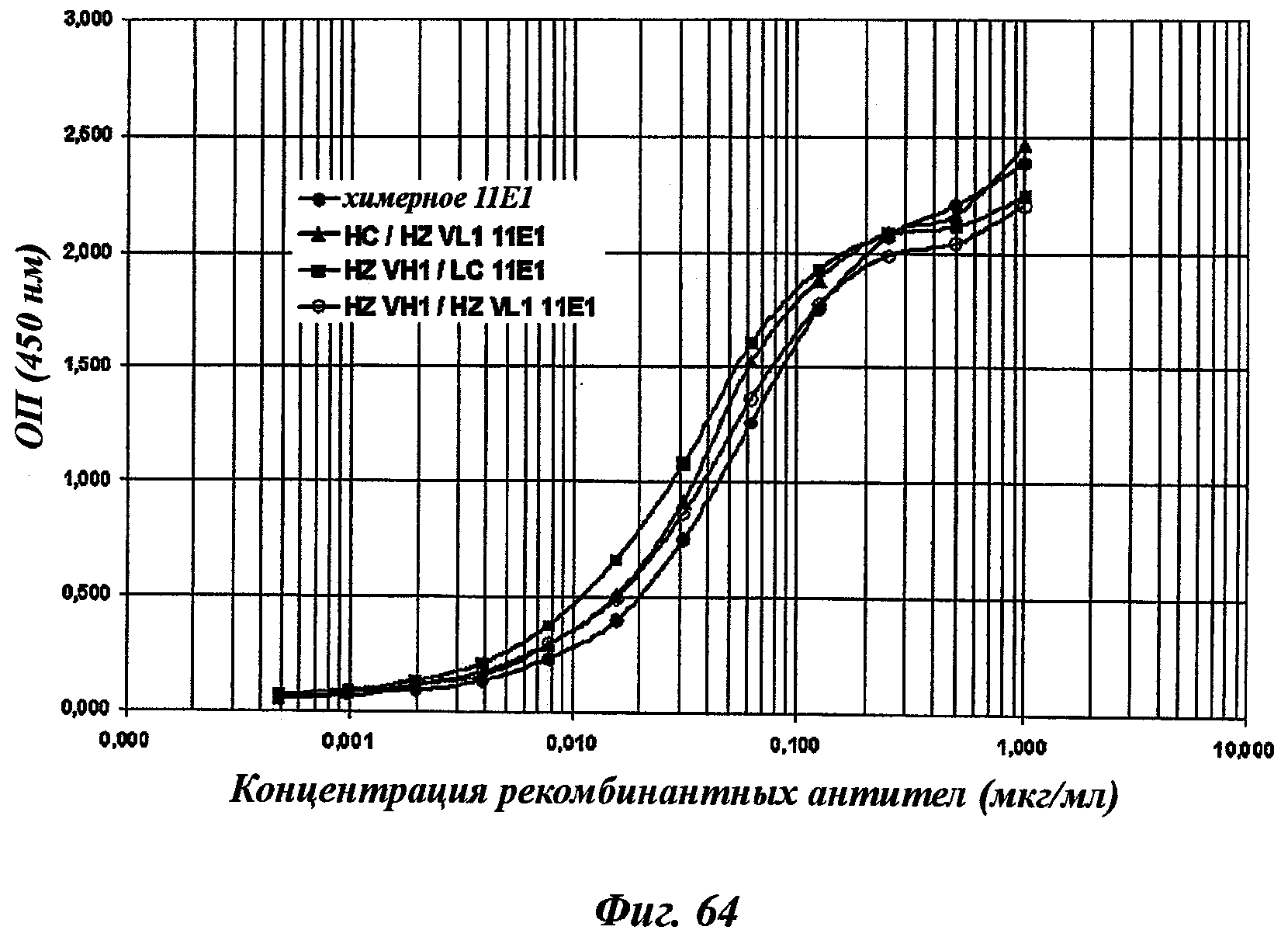
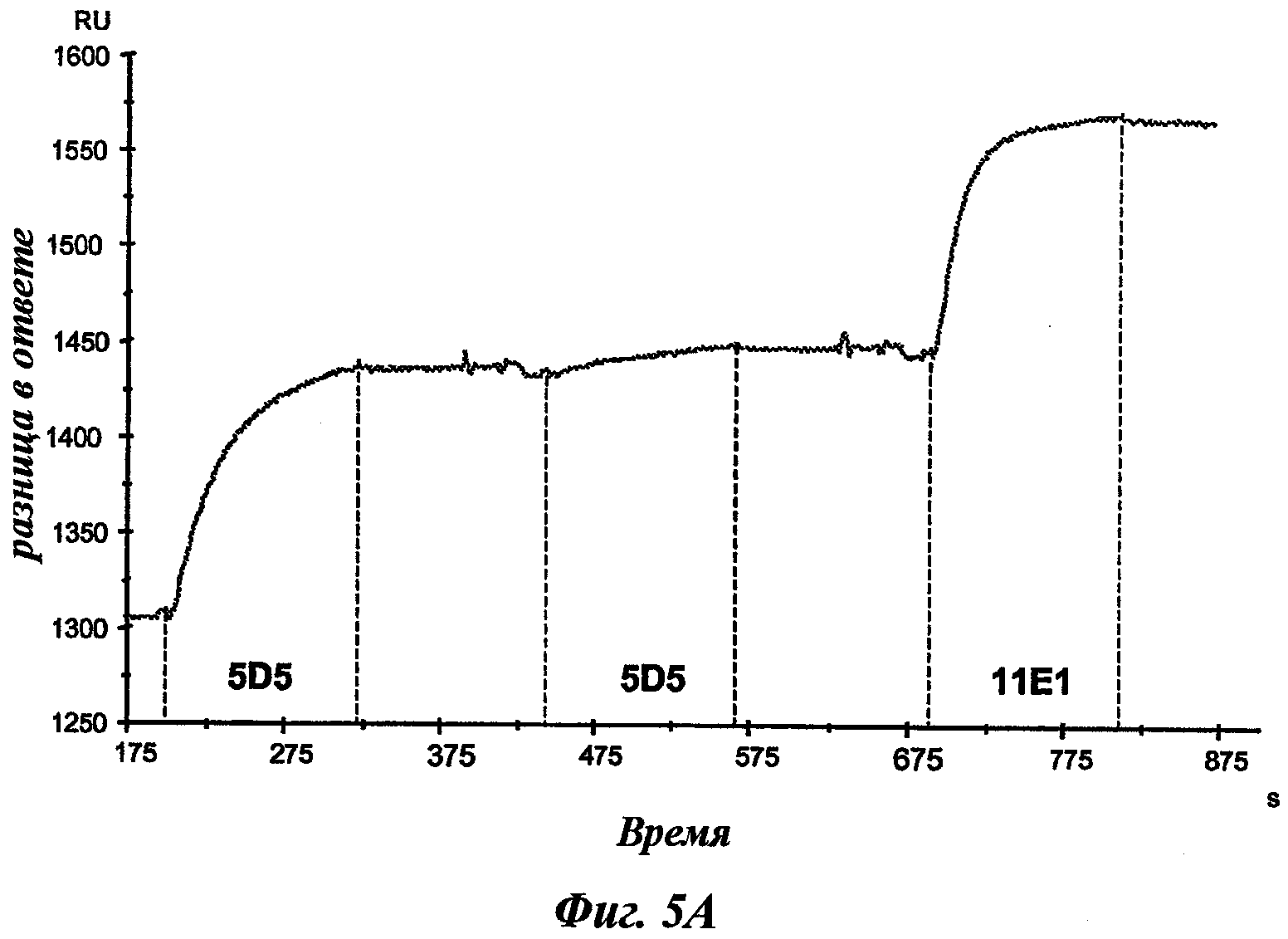
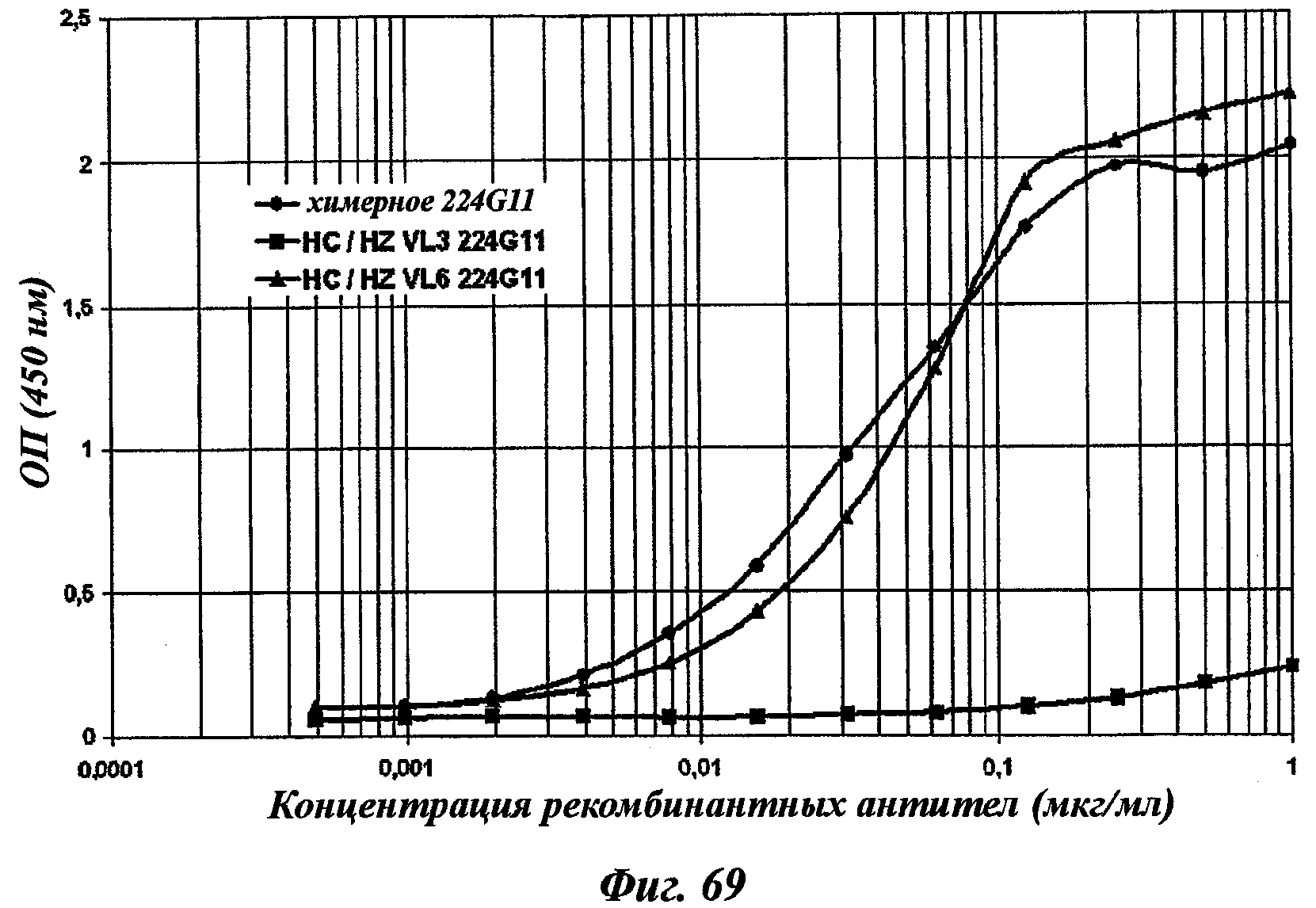
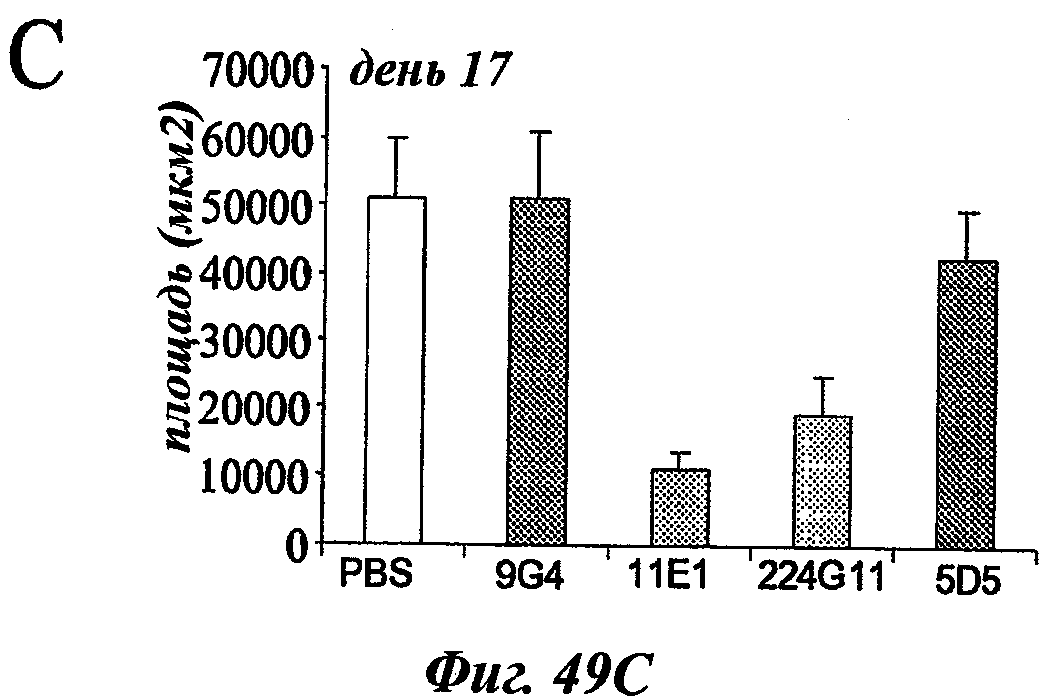
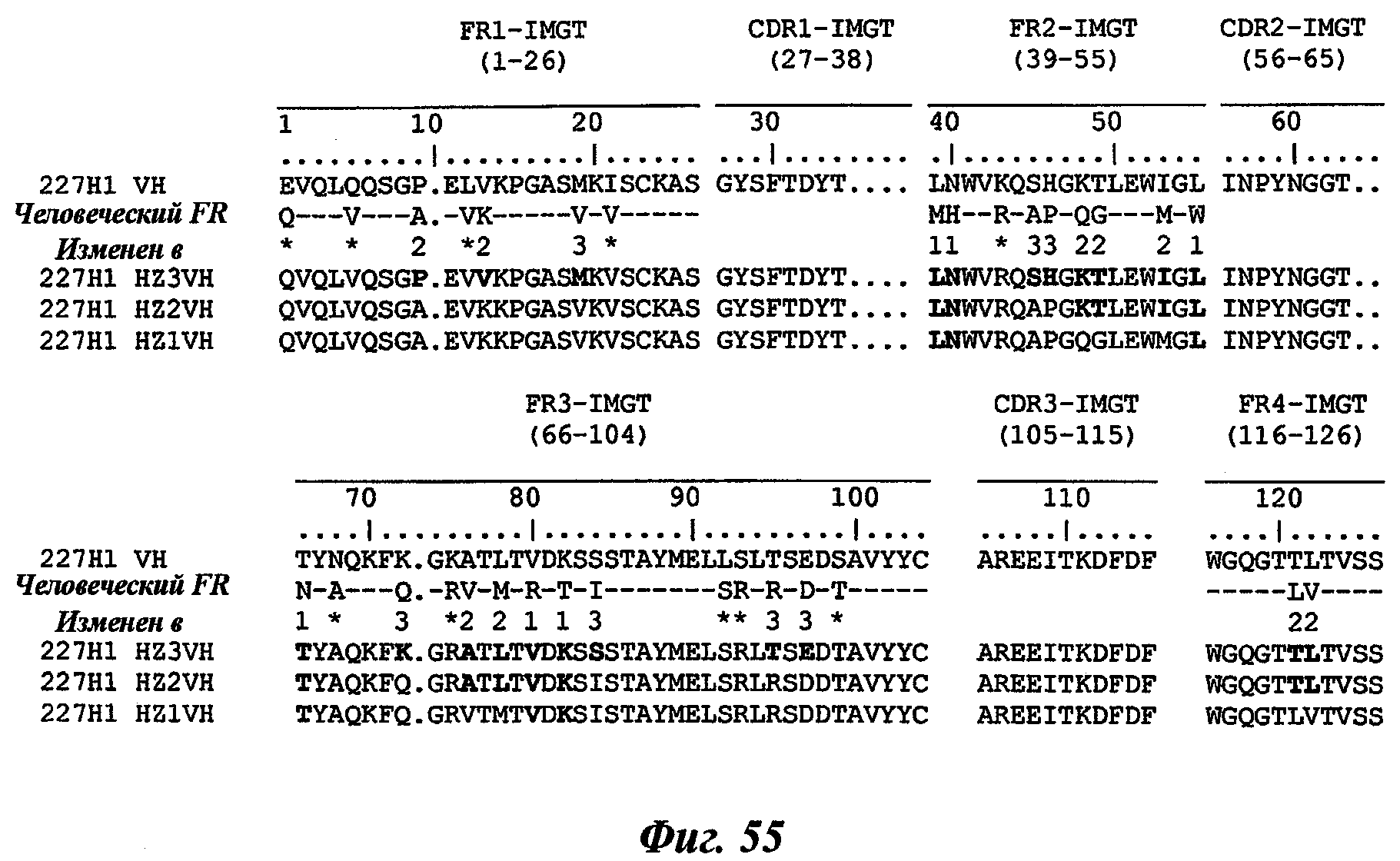

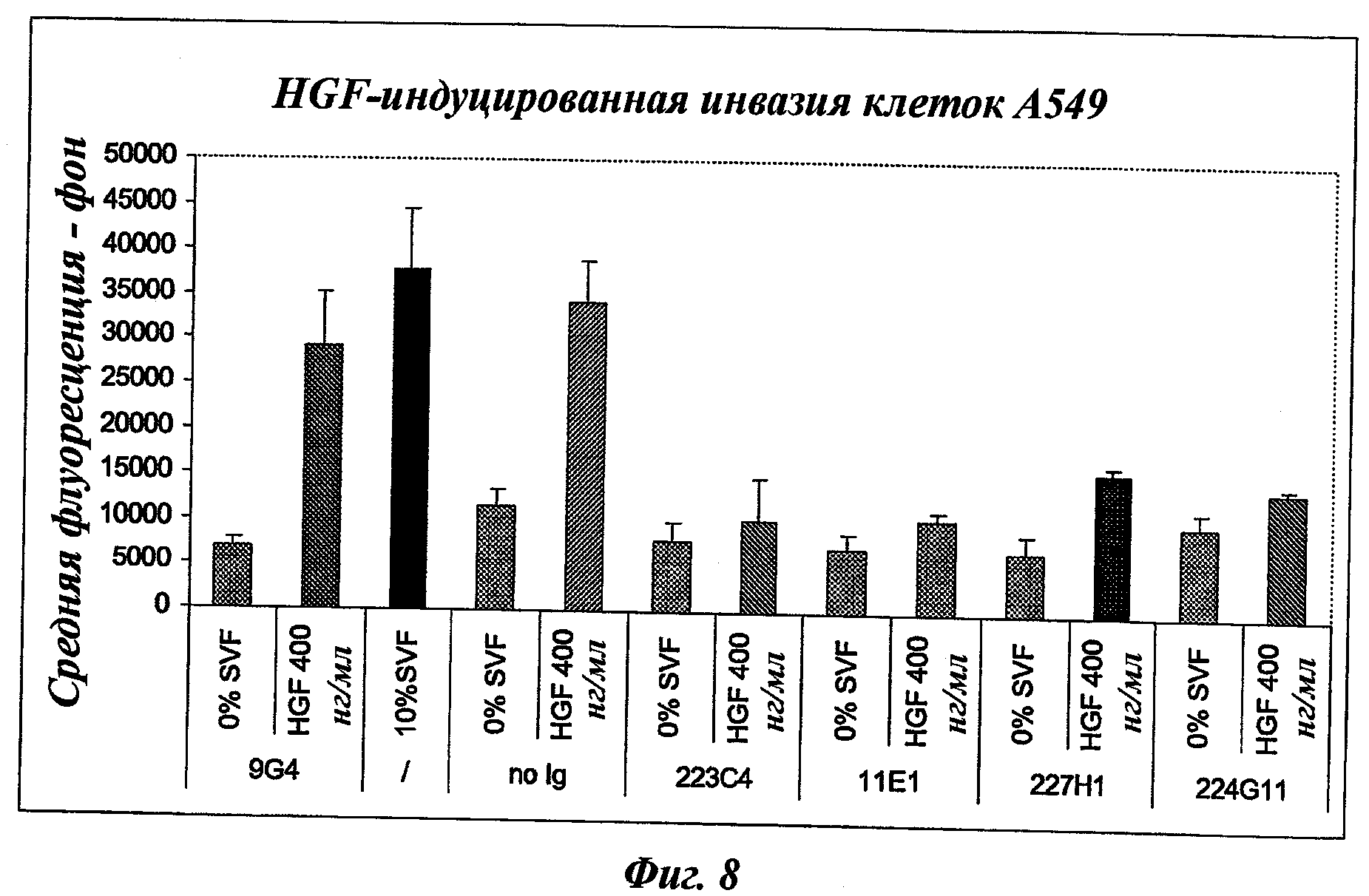
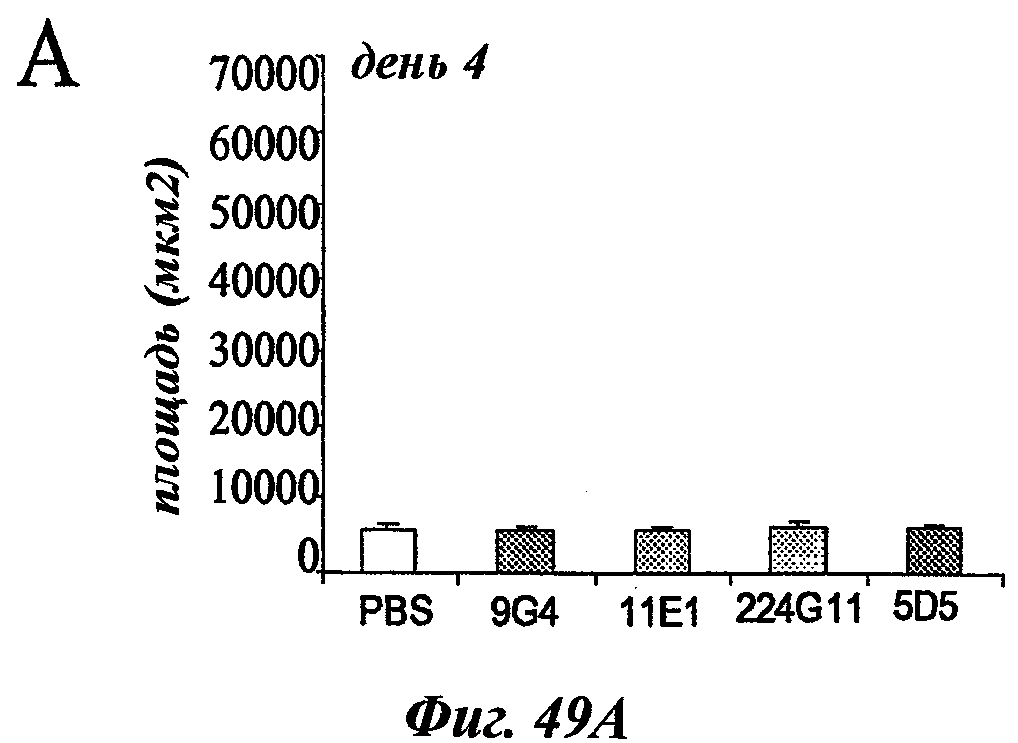
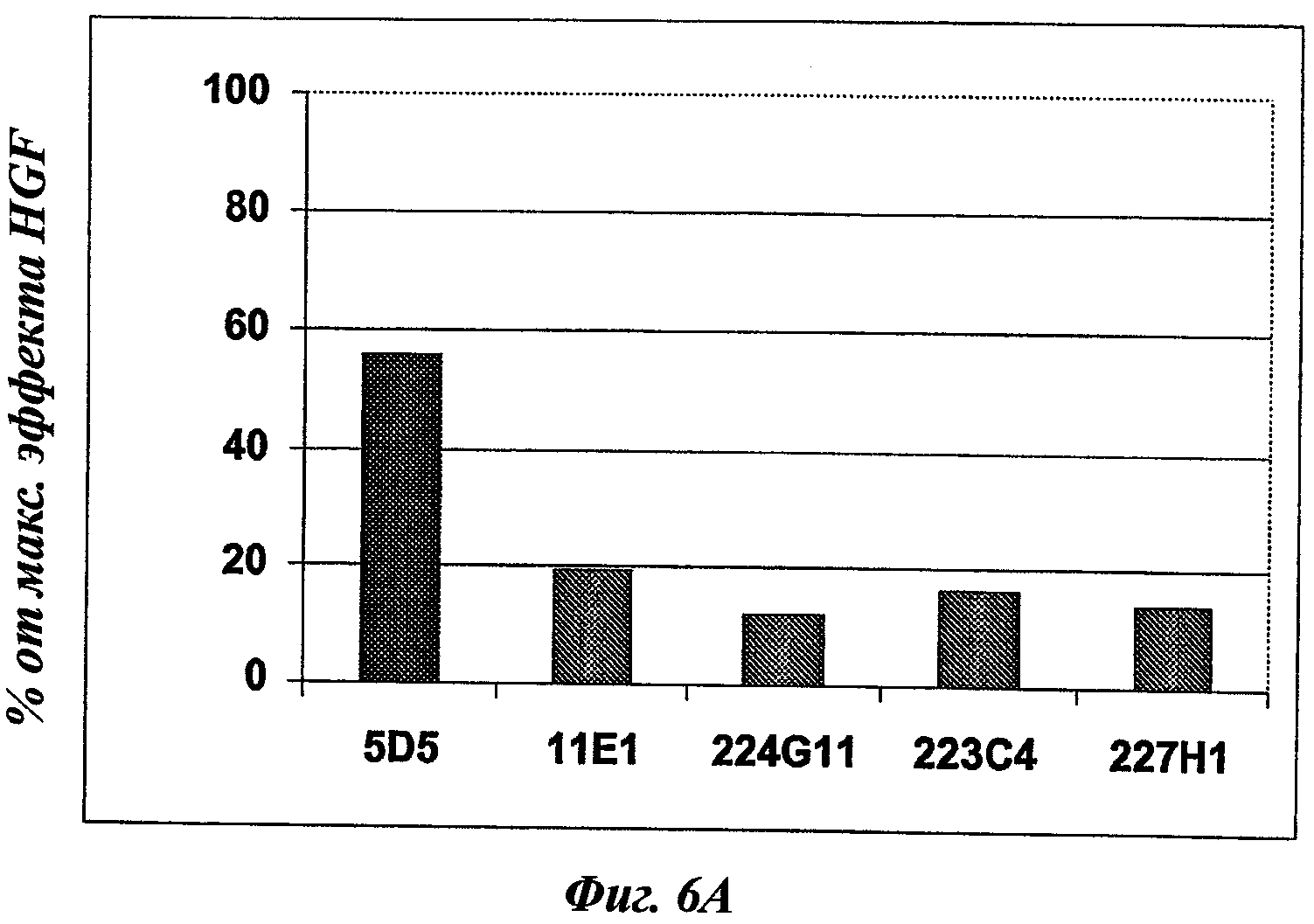
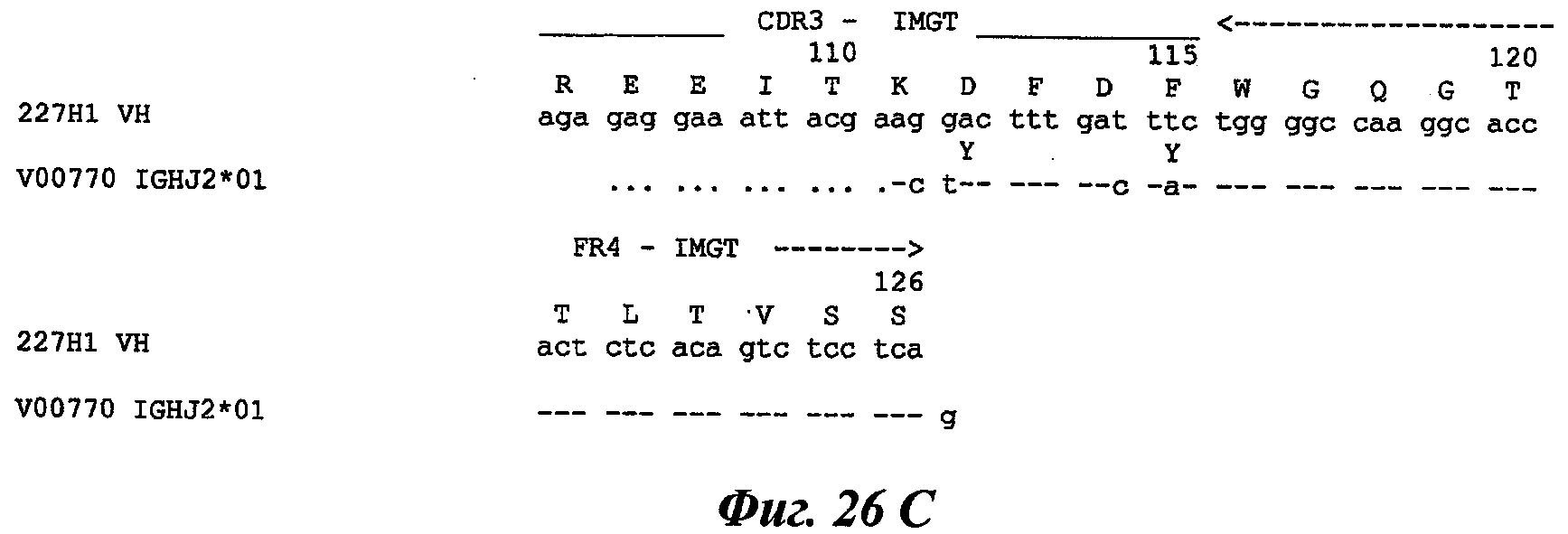


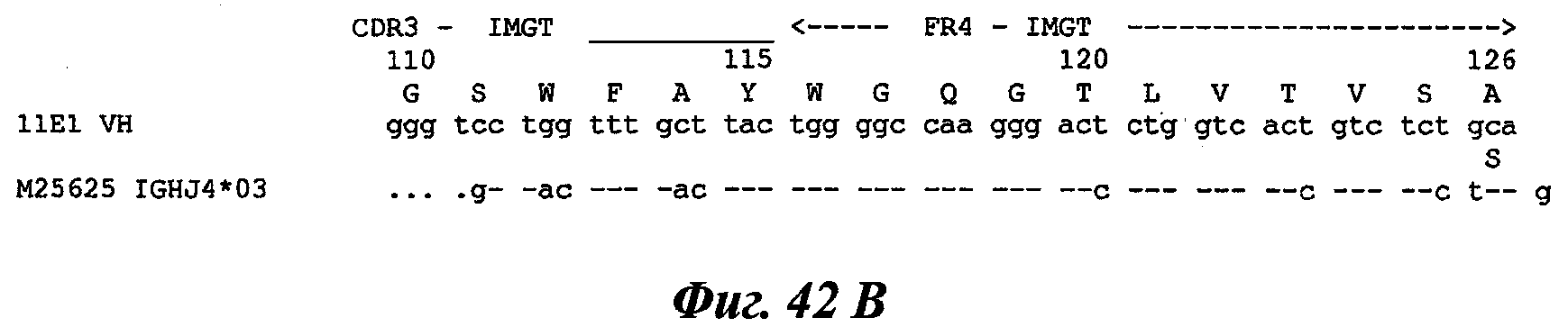
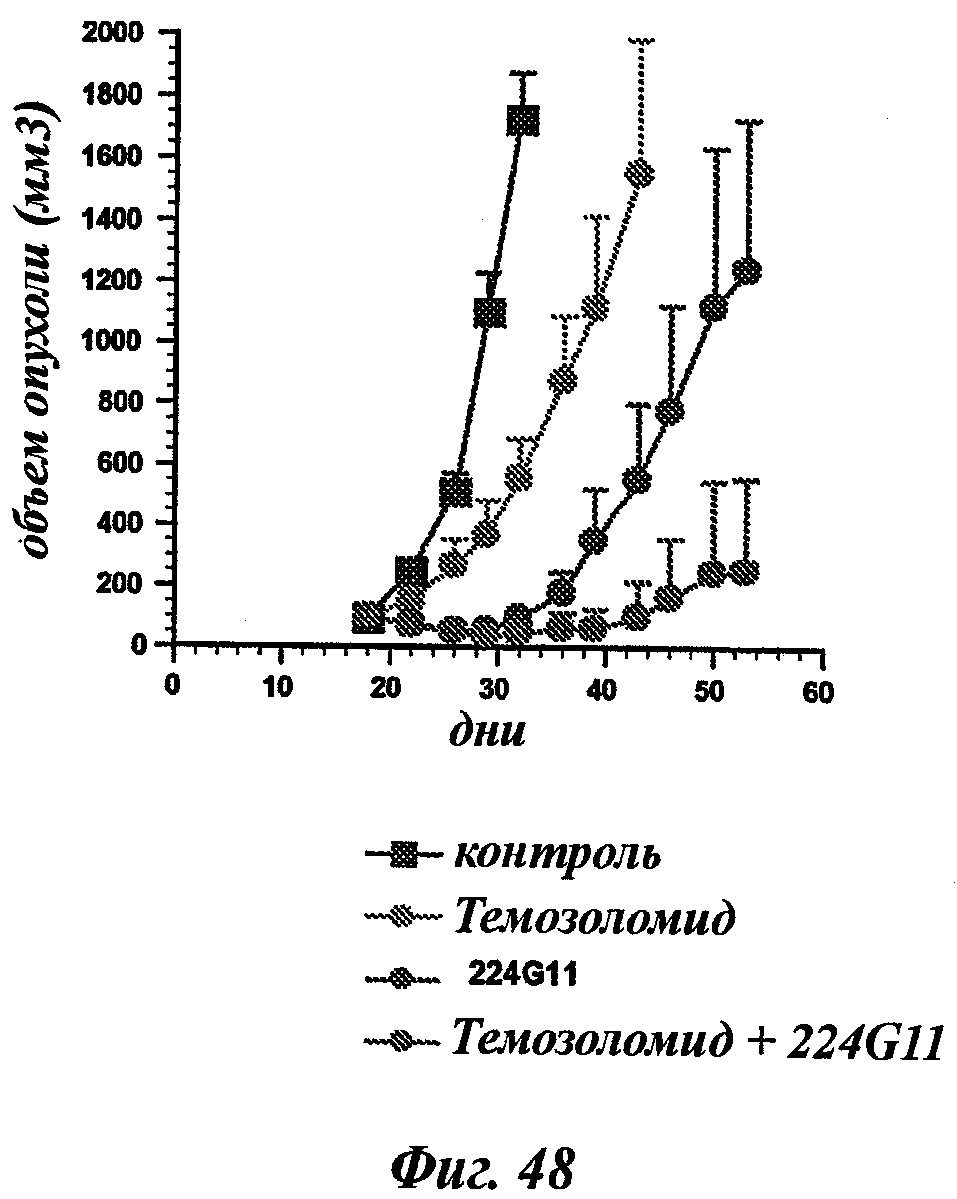
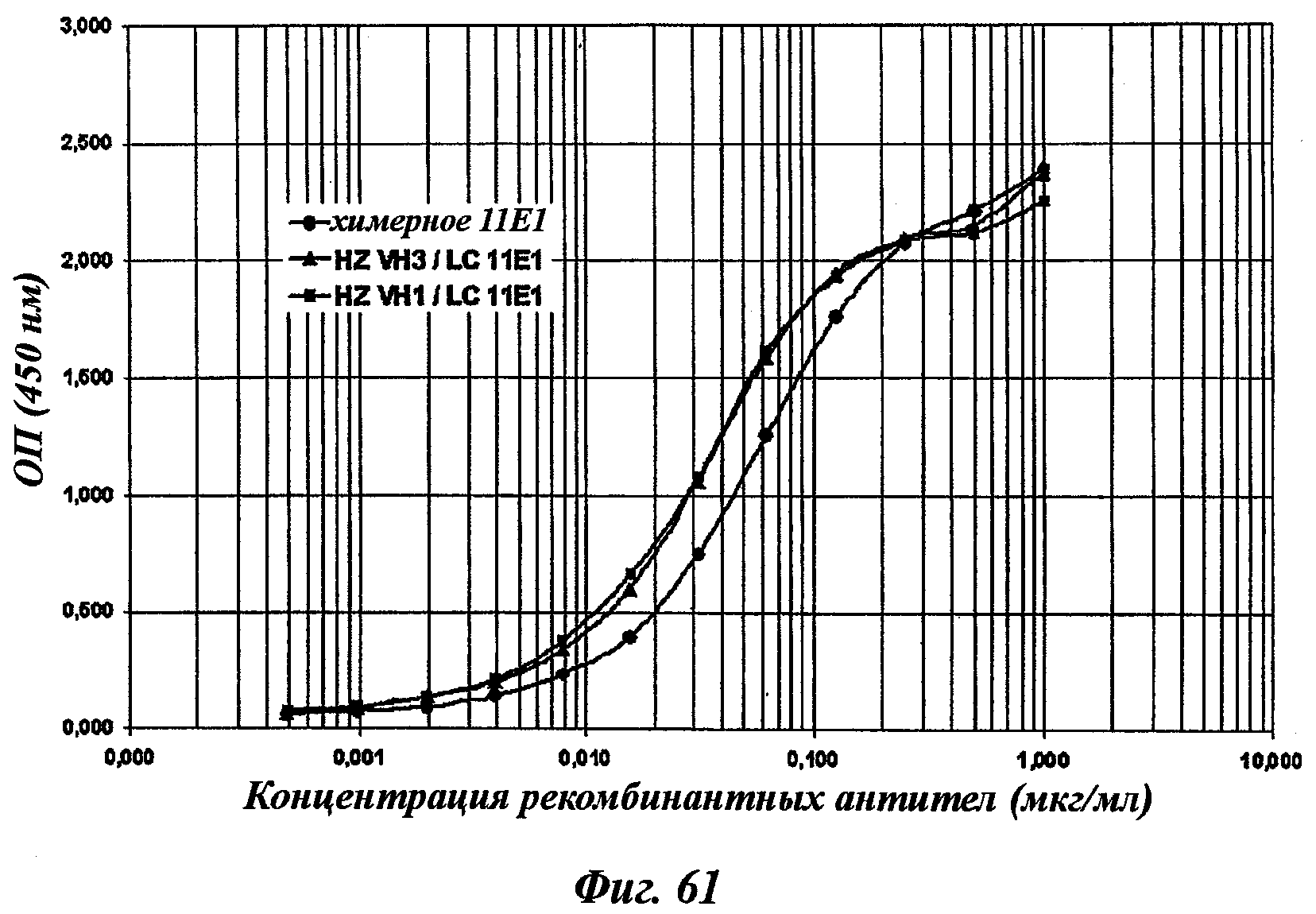

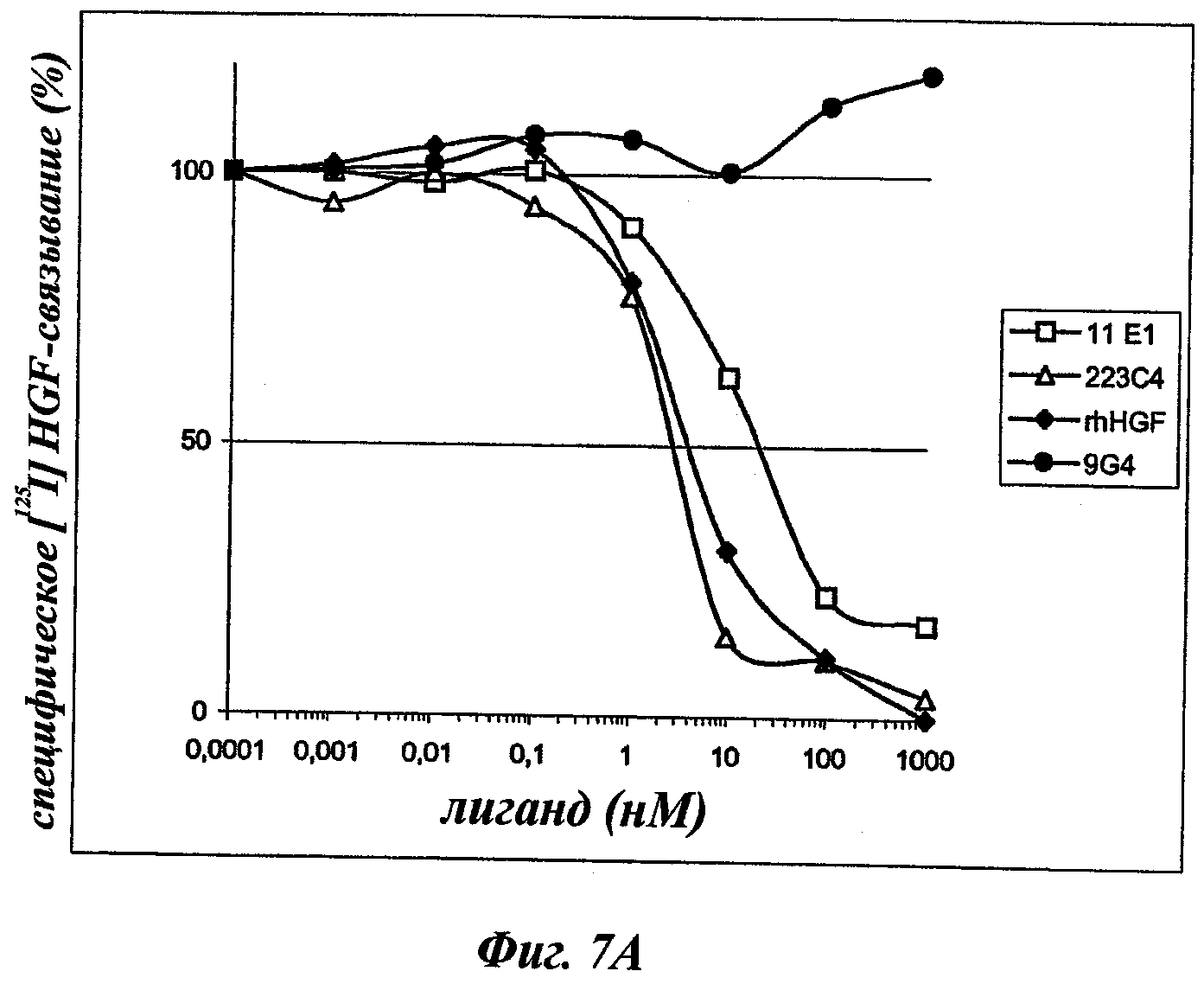




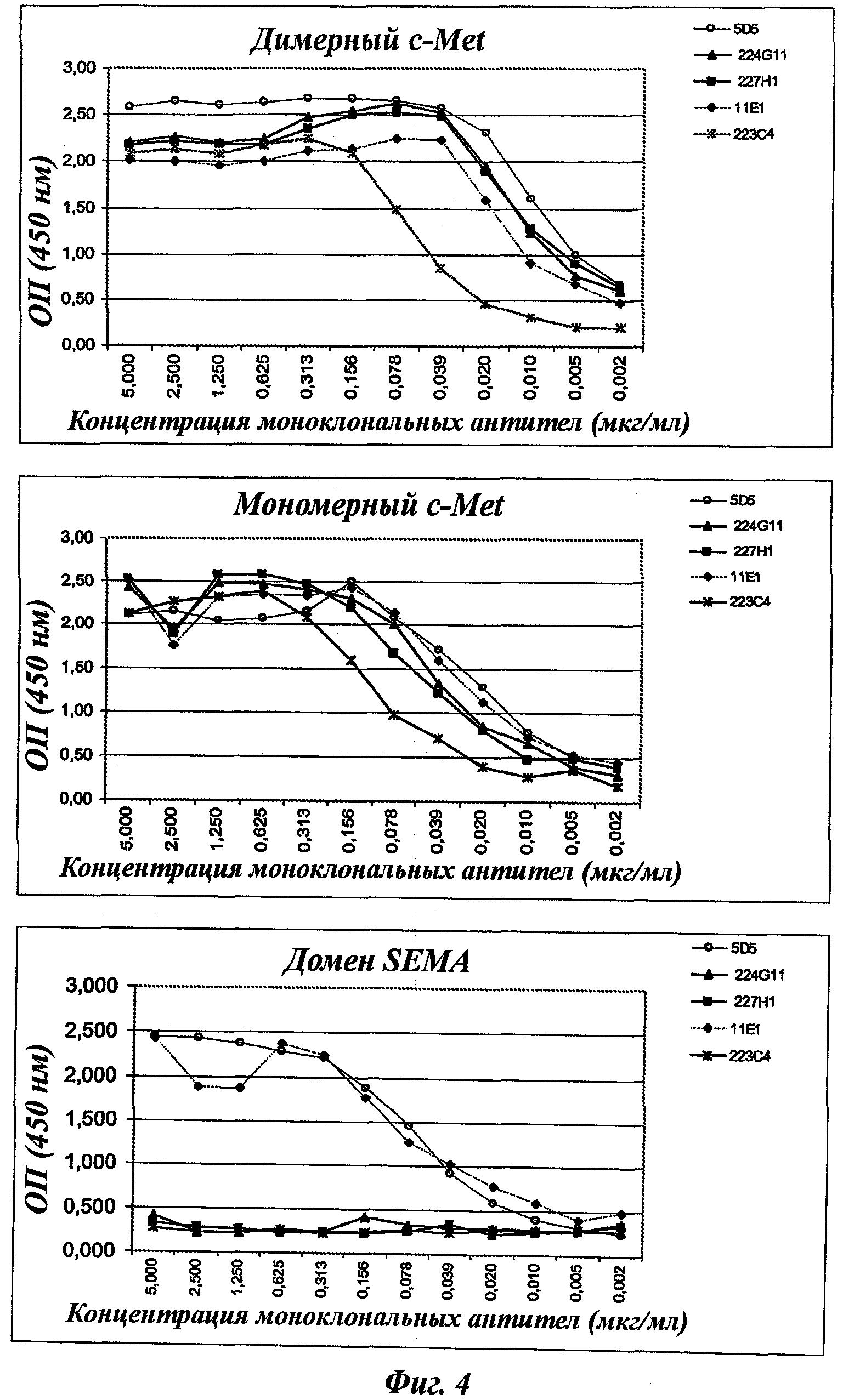

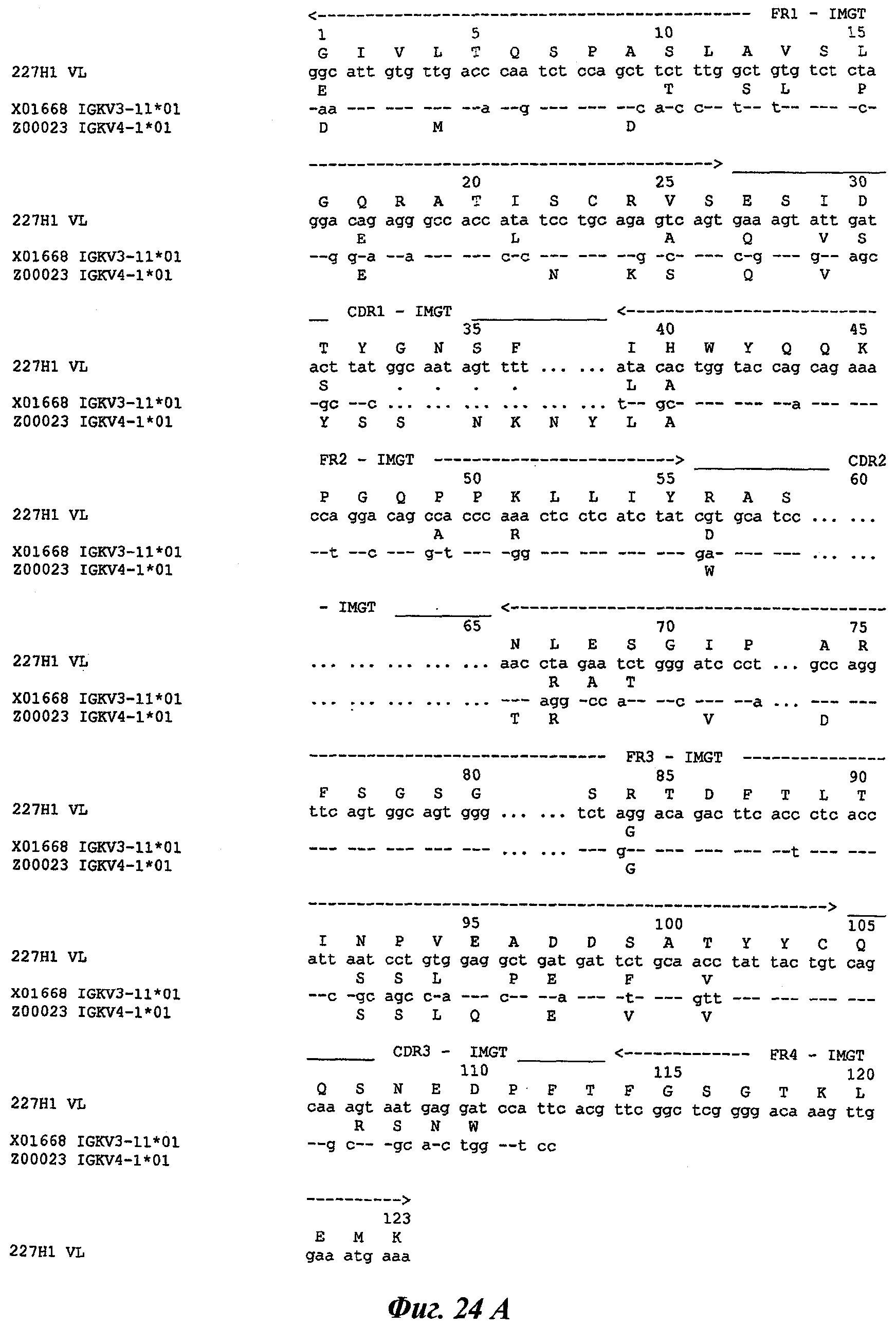
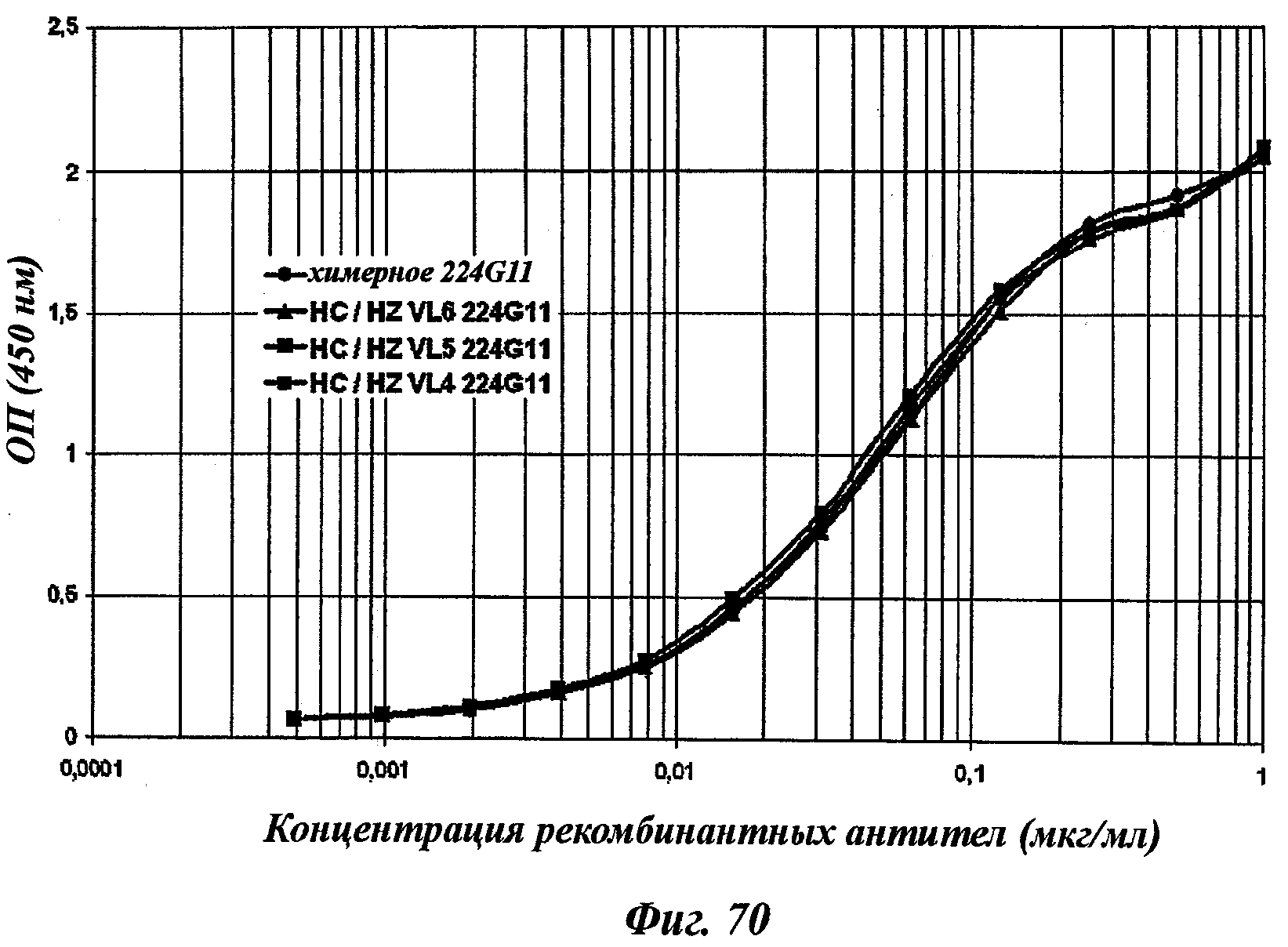
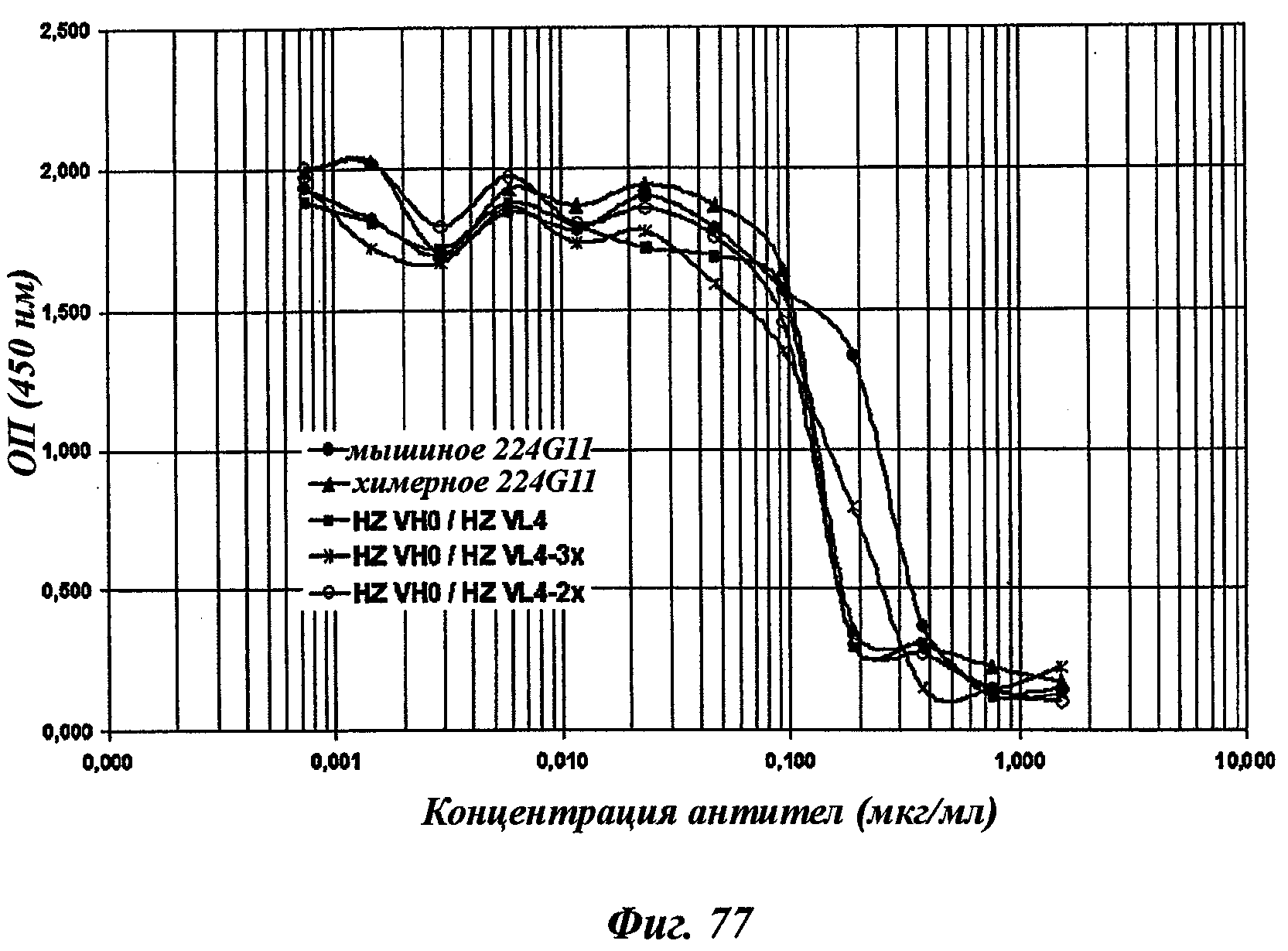
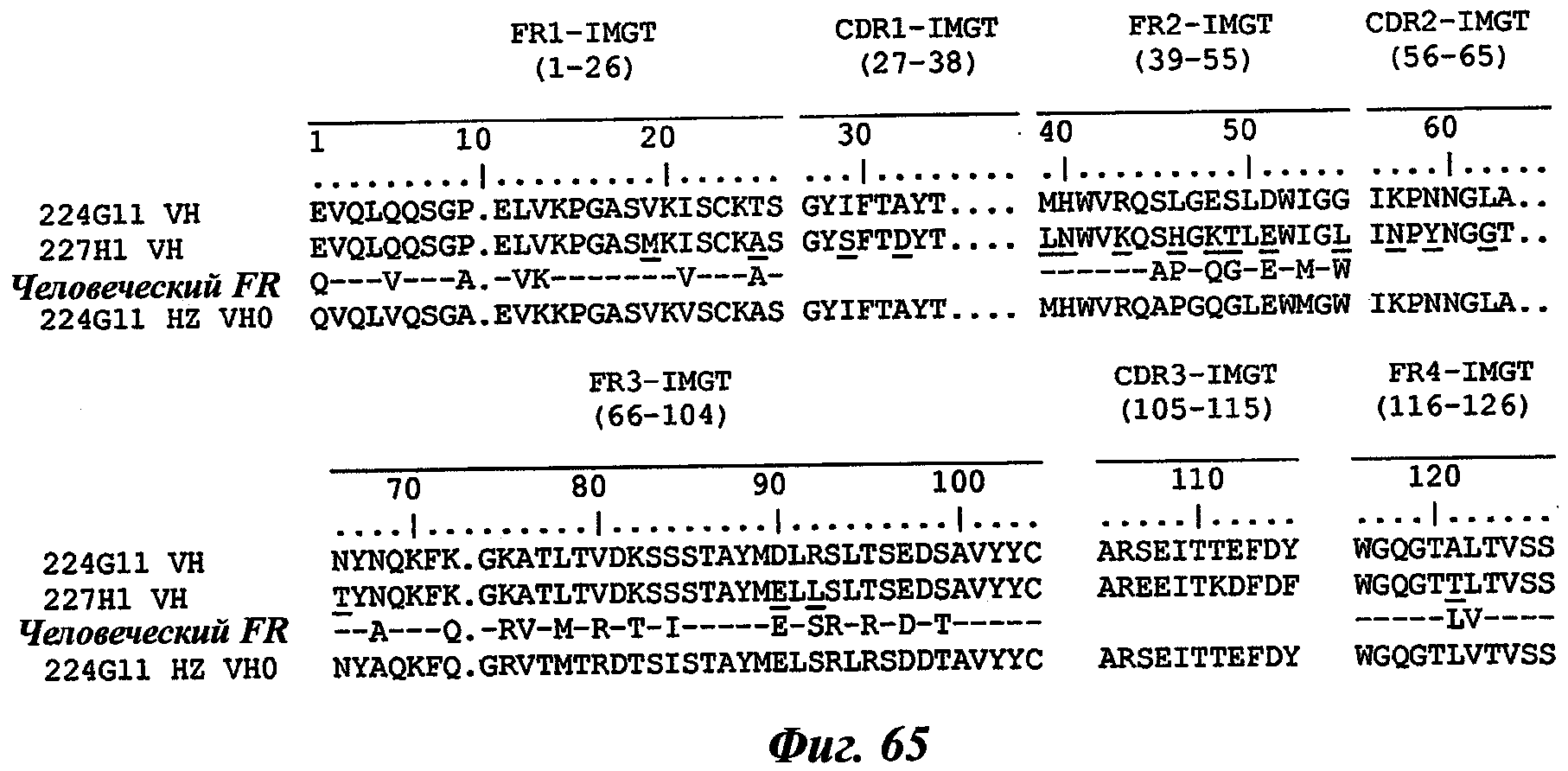
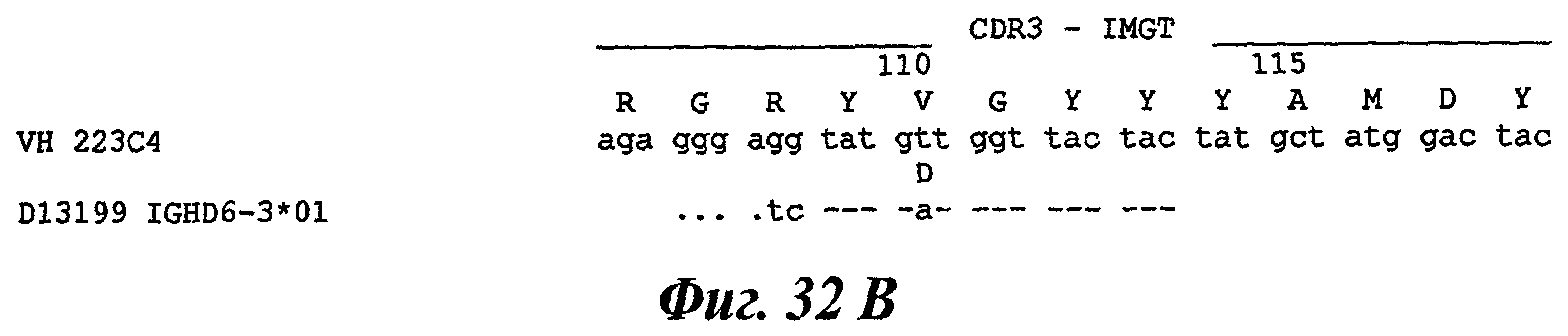
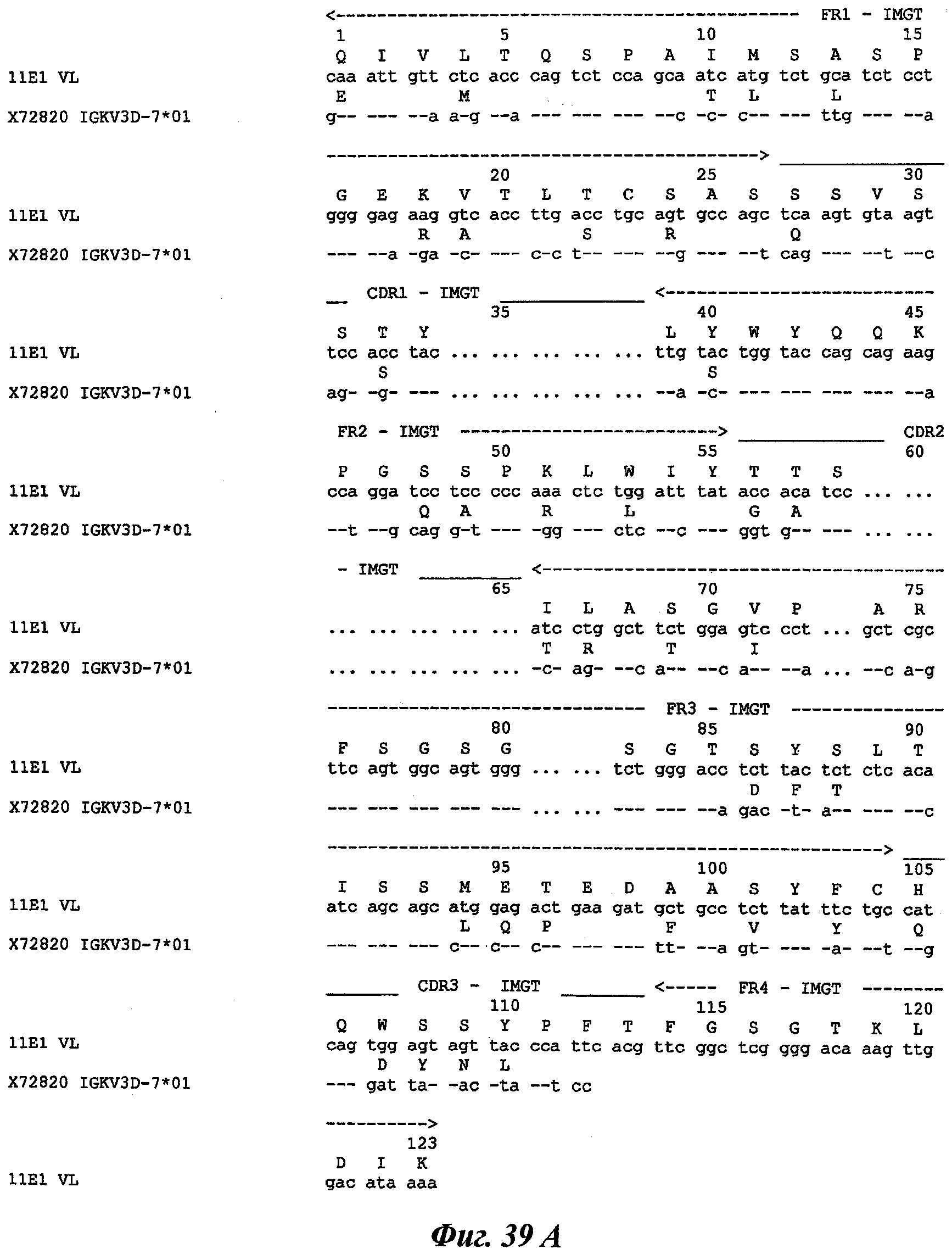
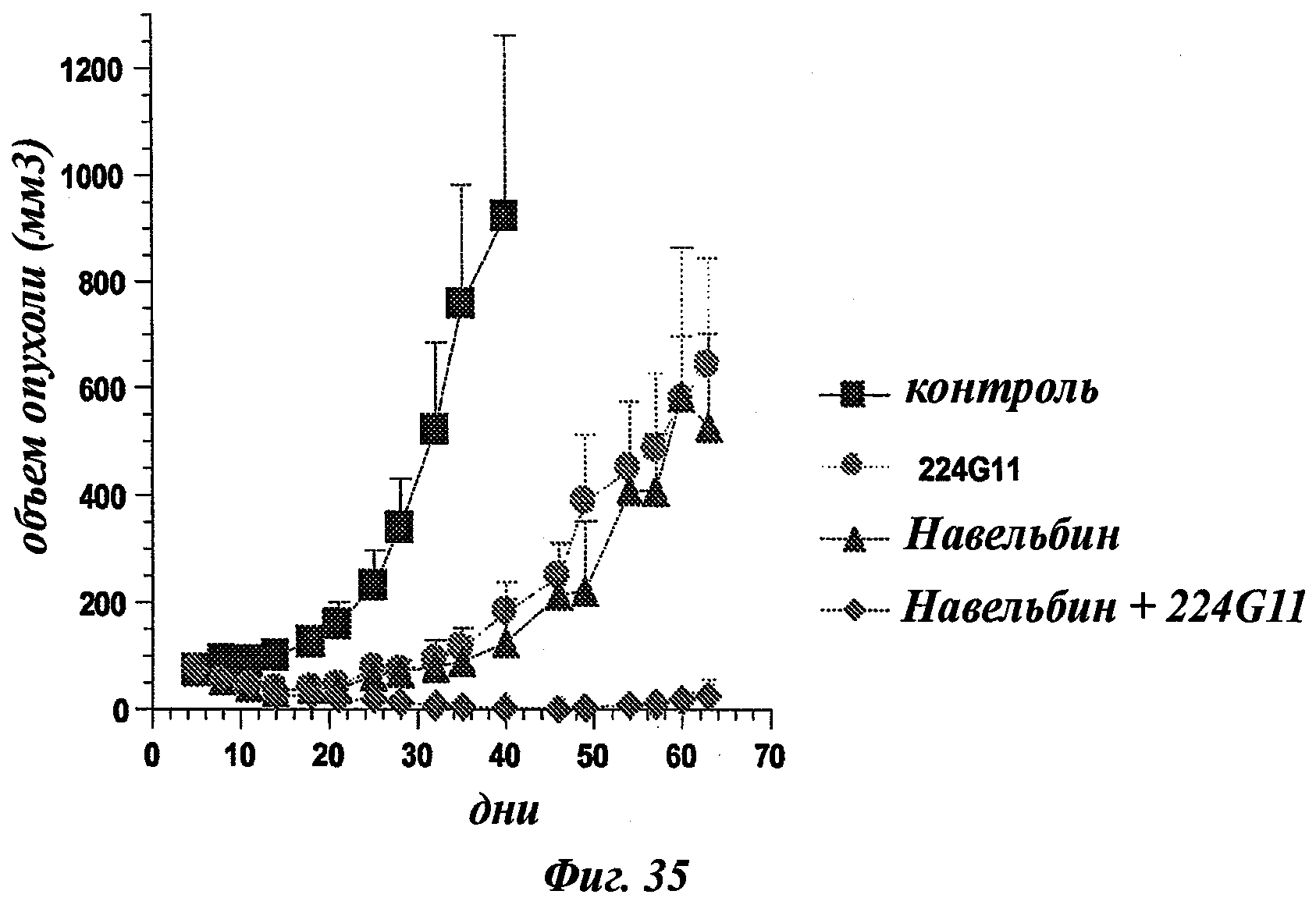
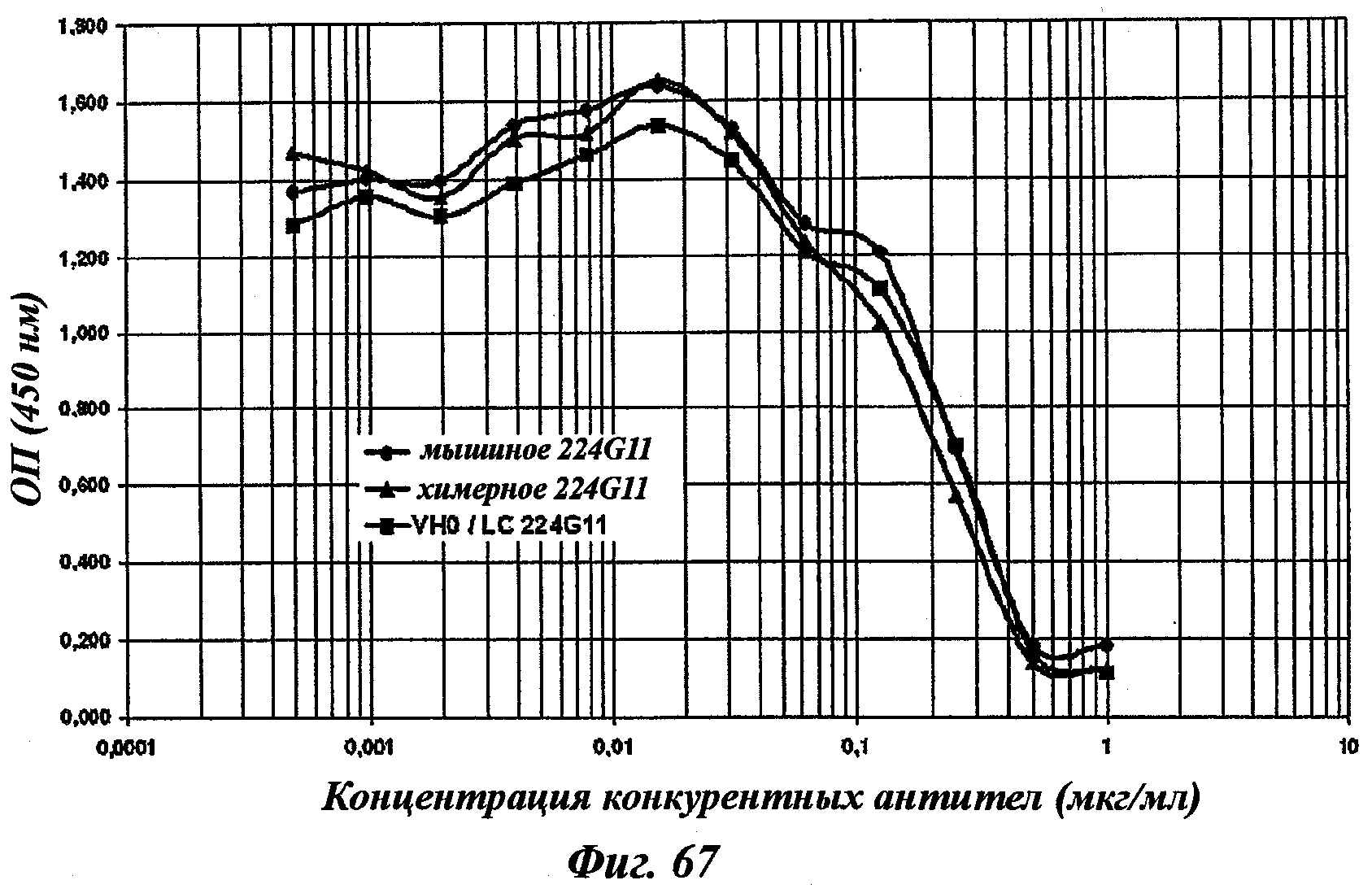

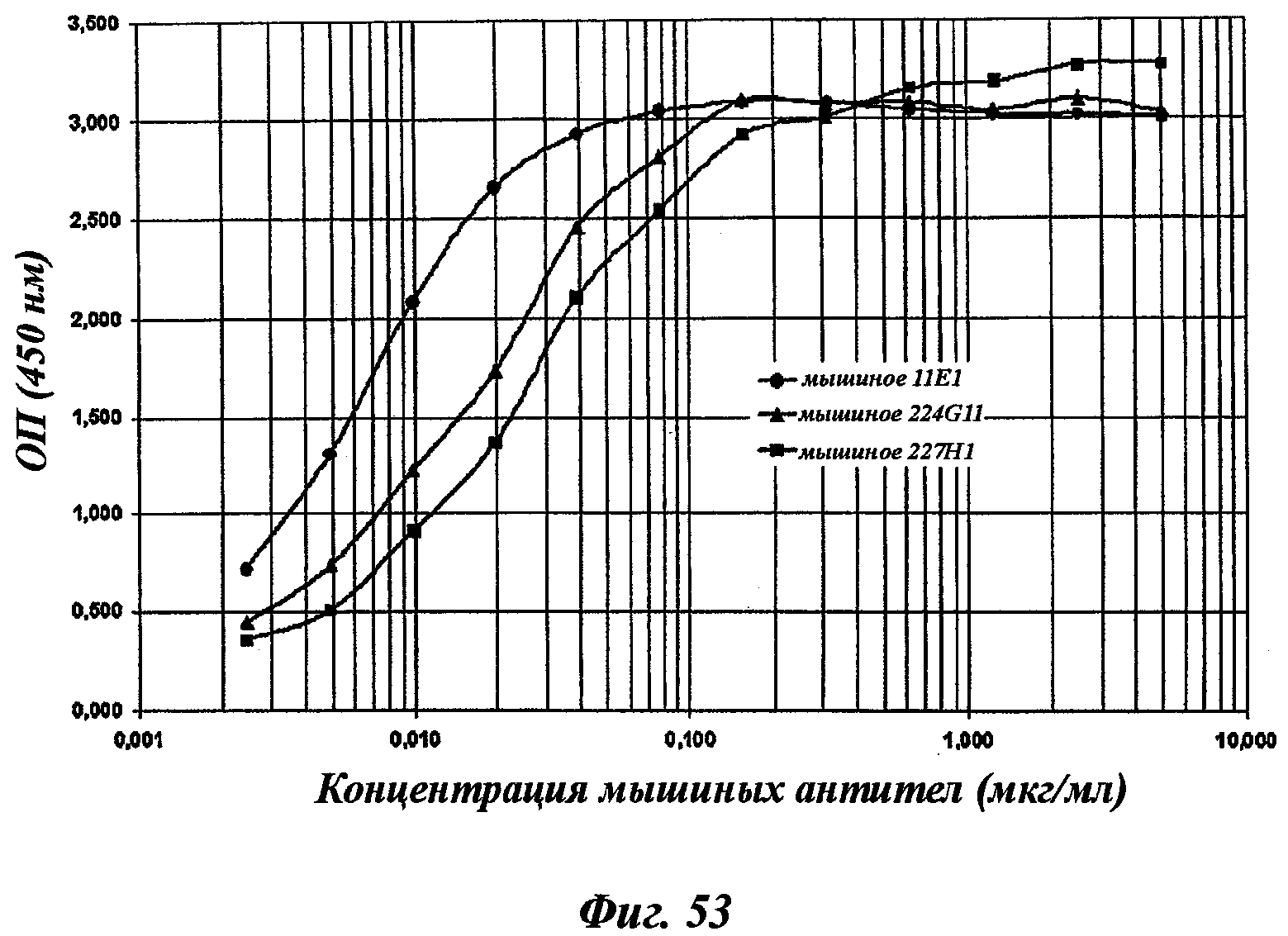
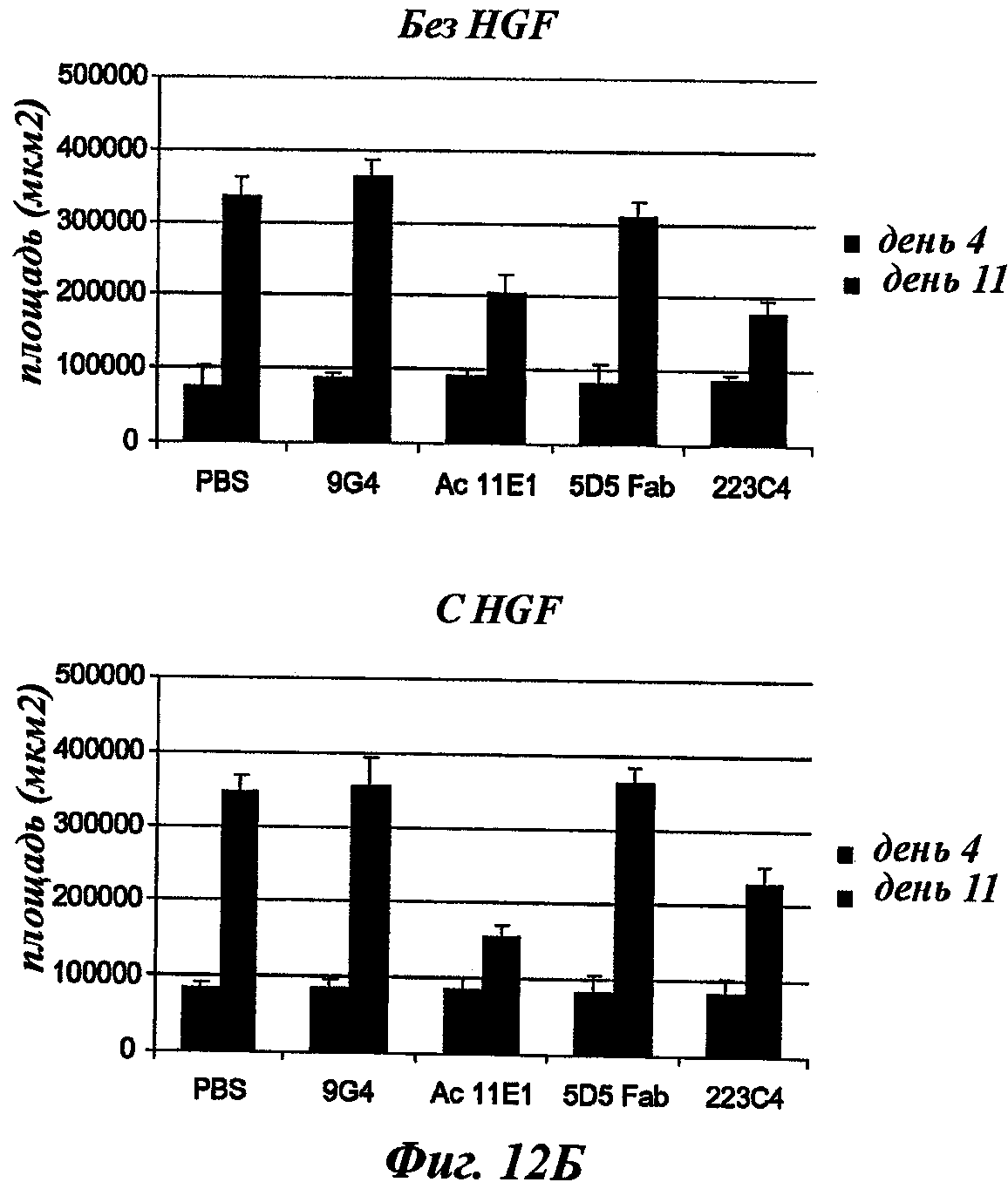
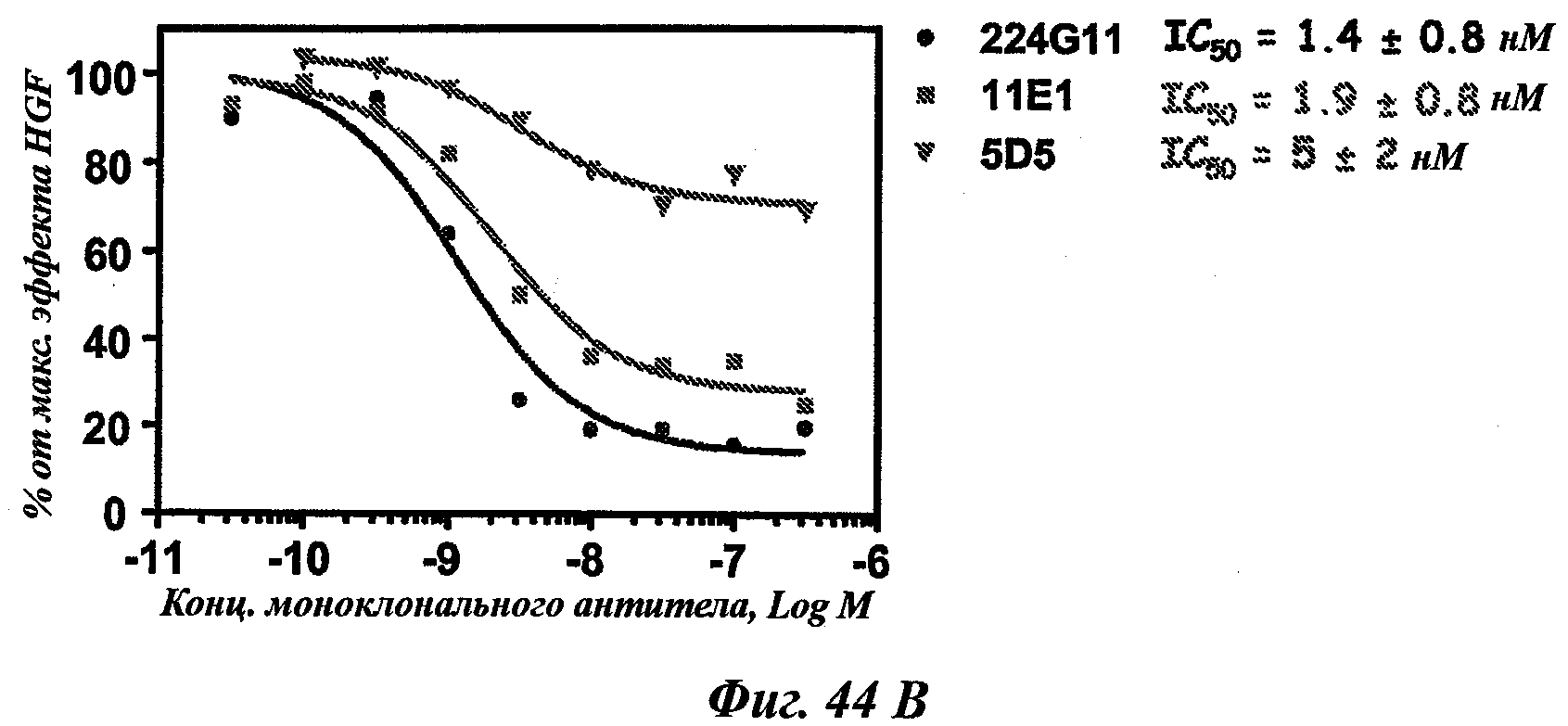


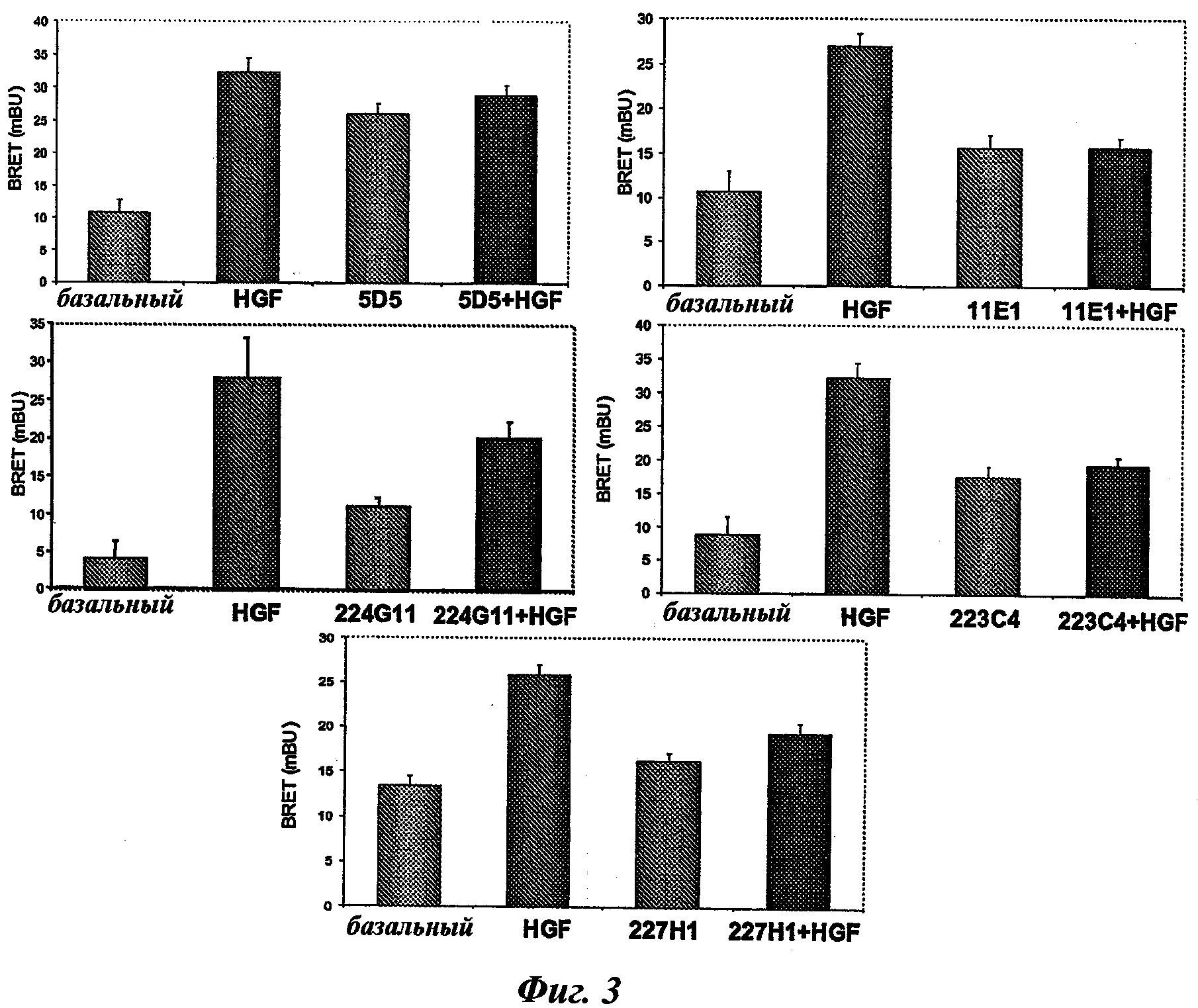
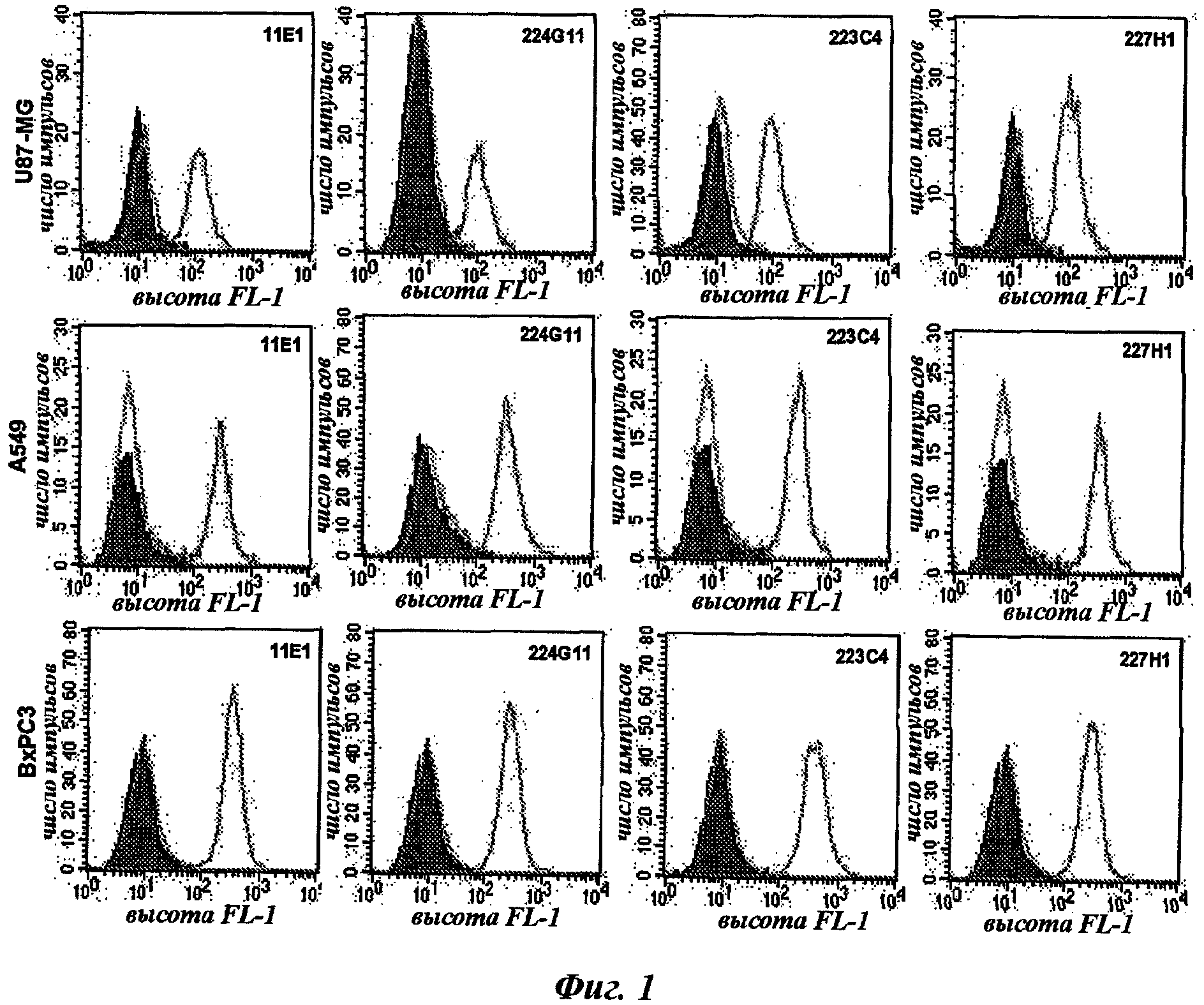
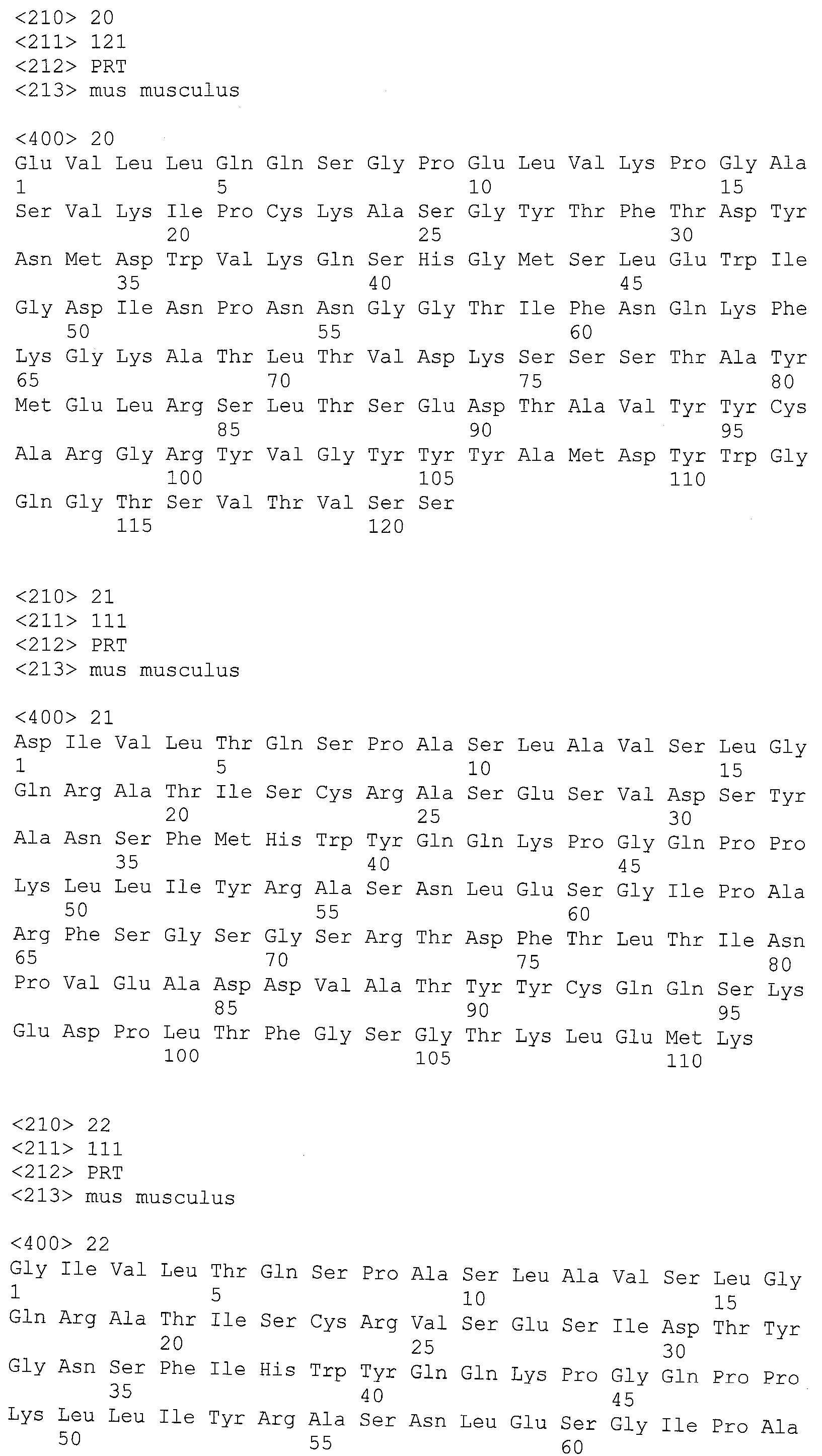


Меквитазин для лечения или предотвращения патологий, в которых задействованы н4 рецепторы гистамина
Способ генерирования активных антител к антигену устойчивости, антитела, полученные этим способом, и их применения
Способ синтеза (1s,2r)-милнаципрана
Производные бензотиазинов, их получение и применение в качестве лекарств
Комбинация антагониста с-мет и аминогетероарила для лечения рака
Новые (поли)аминоалкиламиноалкиламидные, алкил-мочевинные или алкил-сульфонамидные производные эпиподофиллотоксина, способ их получения и их применение в терапии в качестве противораковых средств
Фармацевтическая композиция, содержащая эфир дгк, для парентерального введения
Композиция, содержащая комбинацию экстракта бузины и штамма l. paracasel, l. casei, l. bulgaricus или s. thermophilics
Производные хромонов, способ их получения и их терапевтические применения
Антитело против c-met
Стабильная фармацевтическая композиция водорастворимой соли винорелбина
Меквитазин для лечения или предотвращения патологий, в которых задействованы н4 рецепторы гистамина
Способ генерирования активных антител к антигену устойчивости, антитела, полученные этим способом, и их применения
Способ синтеза (1s,2r)-милнаципрана
Производные бензотиазинов, их получение и применение в качестве лекарств
Комбинация антагониста с-мет и аминогетероарила для лечения рака
Новые (поли)аминоалкиламиноалкиламидные, алкил-мочевинные или алкил-сульфонамидные производные эпиподофиллотоксина, способ их получения и их применение в терапии в качестве противораковых средств
Фармацевтическая композиция, содержащая эфир дгк, для парентерального введения
Композиция, содержащая комбинацию экстракта бузины и штамма l. paracasel, l. casei, l. bulgaricus или s. thermophilics
Производные хромонов, способ их получения и их терапевтические применения

