Результат интеллектуальной деятельности: КЛЕТКА МИЦЕЛИАЛЬНОГО ГРИБА Penicillium canescens - ПРОДУЦЕНТ КСИЛАНАЗЫ И ЛАККАЗЫ, СПОСОБ ПОЛУЧЕНИЯ КОМБИНИРОВАННОГО ФЕРМЕНТНОГО ПРЕПАРАТА КСИЛАНАЗЫ И ЛАККАЗЫ
Вид РИД
Изобретение
Область техники
Изобретение относится к микробиологической промышленности, генной инженерии и биотехнологии, в частности, к новому штамму Penicillium canescens - продуценту совместного ферментного препарата ксиланазы и лакказы, и к способу получения указанного ферментного препарата с использованием указанного штамма.
Описание предшествующего уровня техники
В настоящее время ферментные препараты в целлюлозно-бумажной промышленности наиболее широко используются при отбелке сульфатной целлюлозы, которая традиционно осуществляется при использовании хлорсодержащих соединений, большинство из которых являются устойчивым токсичными мутагенами. Для этой цели в мировой практике используются коммерческие ферментные препараты ксиланаз, однако постоянно ведутся исследования и других микробных ферментов, перспективных для применения в технологических процессах производства целлюлозы и бумаги. Такие микробные ферменты, как целлюлазы, ксиланазы, пектиназы, липазы, лакказы и их комбинации в совокупности с методами химической и термомеханической обработки могут быть использованы для получения целлюлозы, гемицеллюлозы, лигнина или лигноцеллюлозы из древесной пульпы.
Эндоксиланазы являются ферментами, расщепляющими ксилан - основной компонент гемицеллюлоз. Эндоксиланазы находят промышленное применение в бумажной индустрии в процессах отбеливания, в пищевой и кормовой промышленности и других сферах (Kulkami N., Shendye A., Rao M. FEMS Microbiology Reviews, 1999, v.23, p.411).
Лакказа (ЕС 1.10.3.2 пара-дифенол: кислород оксидредуктаза) является полифенолоксидазой, принадлежащей к семейству голубых оксидаз. Лакказа катализирует окисление редуцирующего субстрата за счет восстановления молекулярного кислорода до воды. Продукты реакции - свободные радикалы редуцирующего субстрата и вода. Обычными субстратами лакказы являются: о-, м-, п-фенолы с различными заместителями, полифенолы, ароматические амины. Недавно были открыты реакции лакказ с участием медиаторов, - низкомолекулярных соединений, повышающих эффективность лакказы в биодеградации лигнина, - устойчивых соединений различной структуры, включая ксенобиотики и вещества, загрязняющие окружающую среду.
В последнее время интенсивно ведутся исследования по разработке стратегии использования ферментных препаратов совместного действия. Так в результате действия ксиланаз лигнин оказывается на поверхности и становится более доступным для воздействия лакказ, в результате чего происходит его деградация, приводя к улучшению качества делигнификации целлюлозно-бумажной пульпы. Однако в этих случаях используют либо смешивание двух отдельных ферментных препаратов лакказ и ксиланаз, либо проводят сокультивирование двух отдельных штаммов продуцентов этих ферментов. Комплексное действие препаратов ксиланаз и лакказ используют также в хлебопечении для повышения реологических свойств теста и качества выпекаемого из него хлеба.
Однако в настоящее время нет сообщений, описывающих получение генно-инженерных конструкций, позволяющих создавать высокоэффективные штаммы-продуценты для ко-экспрессии рекомбинантных ферментов, в частности лакказ и ксиланаз.
Краткое описание изобретения
Целью настоящего изобретения являются создание высокоэффективного штамма Р. canescens - продуцента ксиланазы и лакказы и предоставление способа получения комбинированного ферментного препарата ксиланазы и лакказы.
Поставленная задача была решена путем создания штамма Pemcillium canescens РСА-10-4/I-7/12 niaD-, трансформированного плазмидами pBGlac и pBGxylD, -продуцента ксиланазы и лакказы, а также разработки и оптимизации способа получения комбинированного ферментного препарата ксиланазы и лакказы с использованием указанного штамма.
Целью настоящего изобретения является предоставление клетки мицелиального гриба Penicillium canescens, трансформированная плазмидой, содержащей фрагмент ДНК, кодирующий ксиланазу, и плазмидой, содержащей фрагмент ДНК, кодирующий лакказу, -продуцента ксиланазы и лакказы.
Также целью настоящего изобретения является предоставление клетки, описанной выше, в которой указанными плазмидами являются плазмиды pBGlac и pBGxylD, соответственно.
Также целью настоящего изобретения является предоставление клетки штамма Penicillium canescens РСА-10-4/1-7/12 niaD-, трансформированного плазмидами pBGlac и pBGxylD, - продуцента лакказы С1 лигнинолитического гриба Trametes hirsuta и ксиланазы XylD мицелиального гриба Penicillium canescens.
Также целью настоящего изобретения является предоставление способа получения ферментного препарата ксиланазы и лакказы, включающего стадии выращивания клеток мицелиального гриба Penicillium canescens, описанного выше, в питательной среде и выделения ферментного препарата ксиланазы и лакказы из культуральной жидкости.
Также целью настоящего изобретения является предоставление способа, описанного выше, где в качестве клеток мицелиального гриба используют клетки штамма Penicillium canescens, описанного выше.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором выращивание клеток Penicillium canescens осуществляют в присутствии ингибиторов металлопротеиназ.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором в качестве ингибитора металлопротеиназ используют ЭДТА.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором выращивание клеток Penicillium canescens осуществляют в среде с рН в интервале 5.0-6.5.
Также целью настоящего изобретения является предоставление описанного выше способ, в котором выделение ферментных препаратов ксиланазы и лакказы из культуральной жидкости осуществляют путем концентрирования с использованием ультрафильтрации на мембране с порогом отсечения 5 кДа с последующим вакуум-выпариванием фильтрата культуральной жидкости при температуре 40-70°С.
Настоящее изобретение позволяет реализовать технологию получения отечественных рекомбинантных высокопродуктивных штаммов продуцентов ферментов, в частности ксиланаз и лакказ, предназначенных для использования в крупнотоннажных промышленных процессах, так и технологию получения комбинированного ферментного препарата ксиланазы и лакказы для использования в целлюлозно-бумажной промышленности для отбеливания целлюлозы.
Краткое описание рисунков
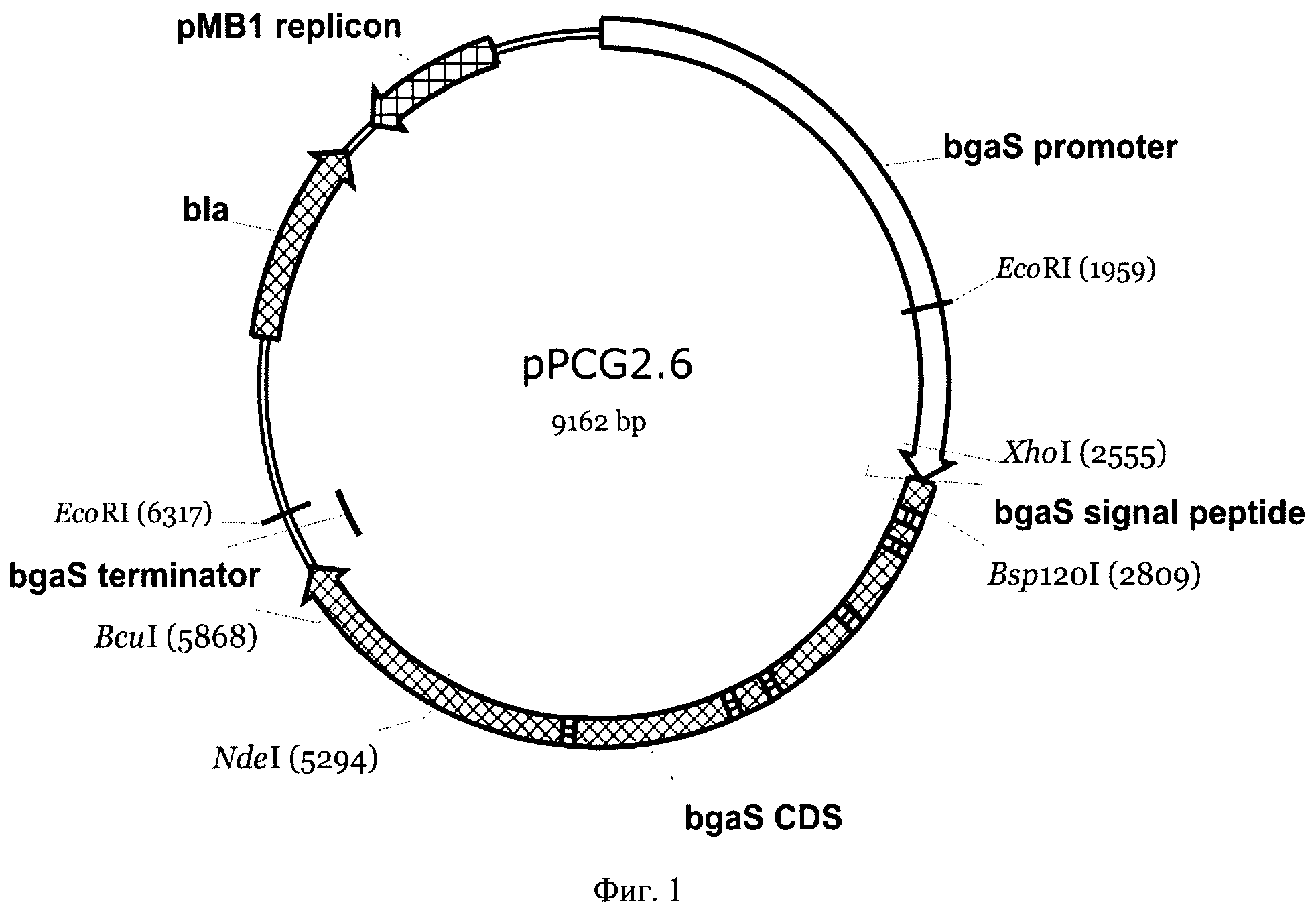
На Фиг.1 приведена карта плазмиды pPCG2.6. Указаны использованные сайты рестрикции, промотор, кодирующая часть, сигнальный пептид и терминатор гена bgaS, ген устойчивости в ампицилину bla и область, обеспечивающая репликацию плазмиды в Е. coli.
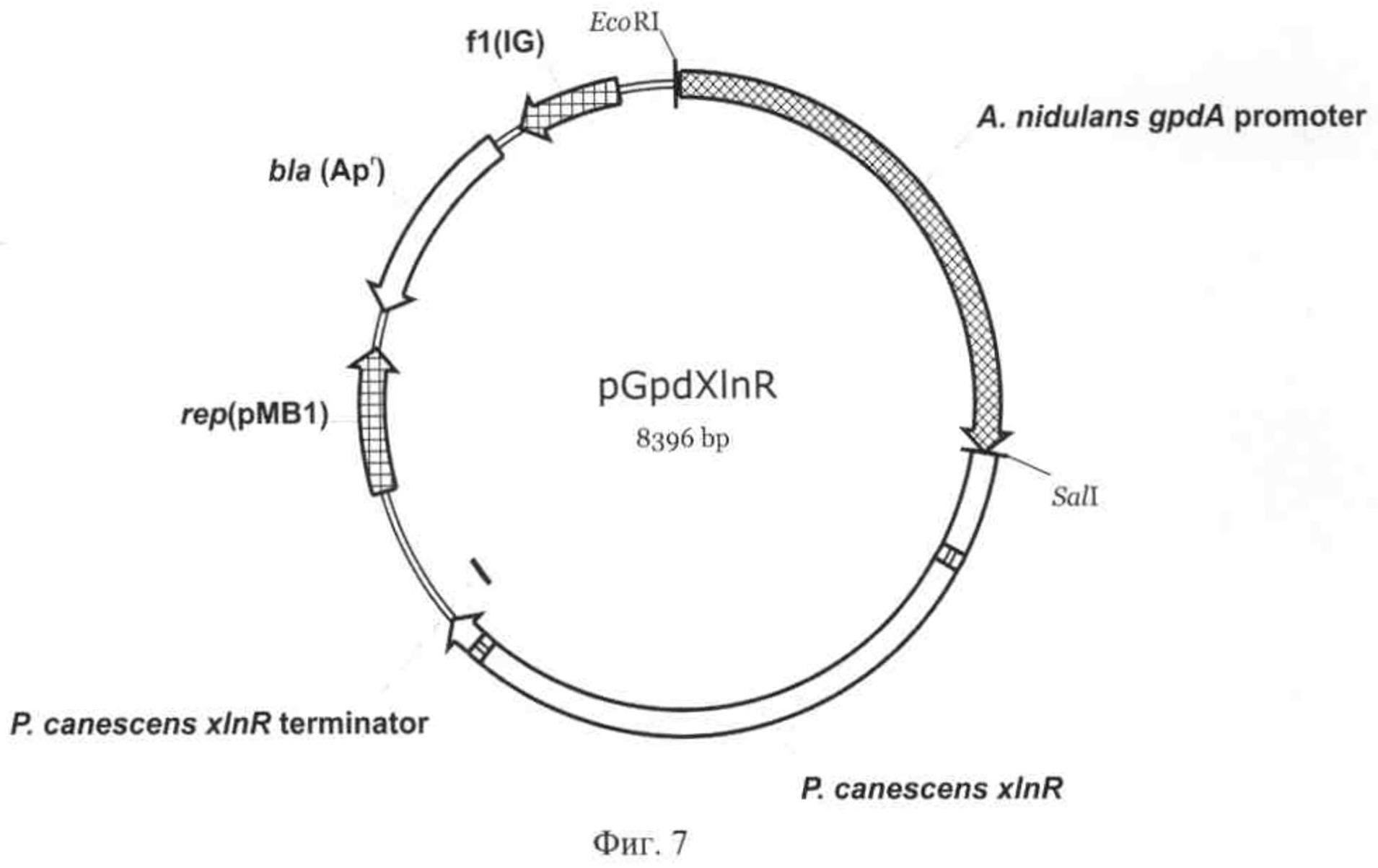
На Фиг.2 приведена карта экспрессионного вектора pPCGMX. Указаны использованные сайты рестрикции, промотор, кодирующая часть, сигнальный пептид и терминатор гена bgaS, ген устойчивости в ампицилину bla и область, обеспечивающая репликацию плазмиды в Е. coli.
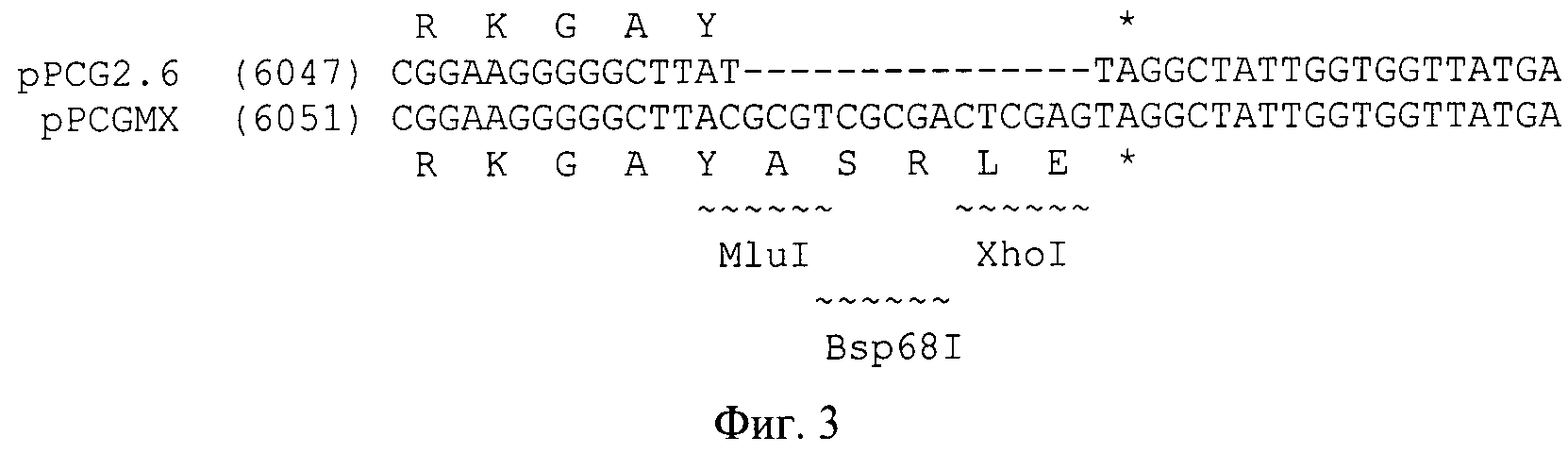
На Фиг.3 изображен поли-линкер перед стоп-кодоном гена bgaS из Р. canescens в экспрессионном векторе pPCGMX.
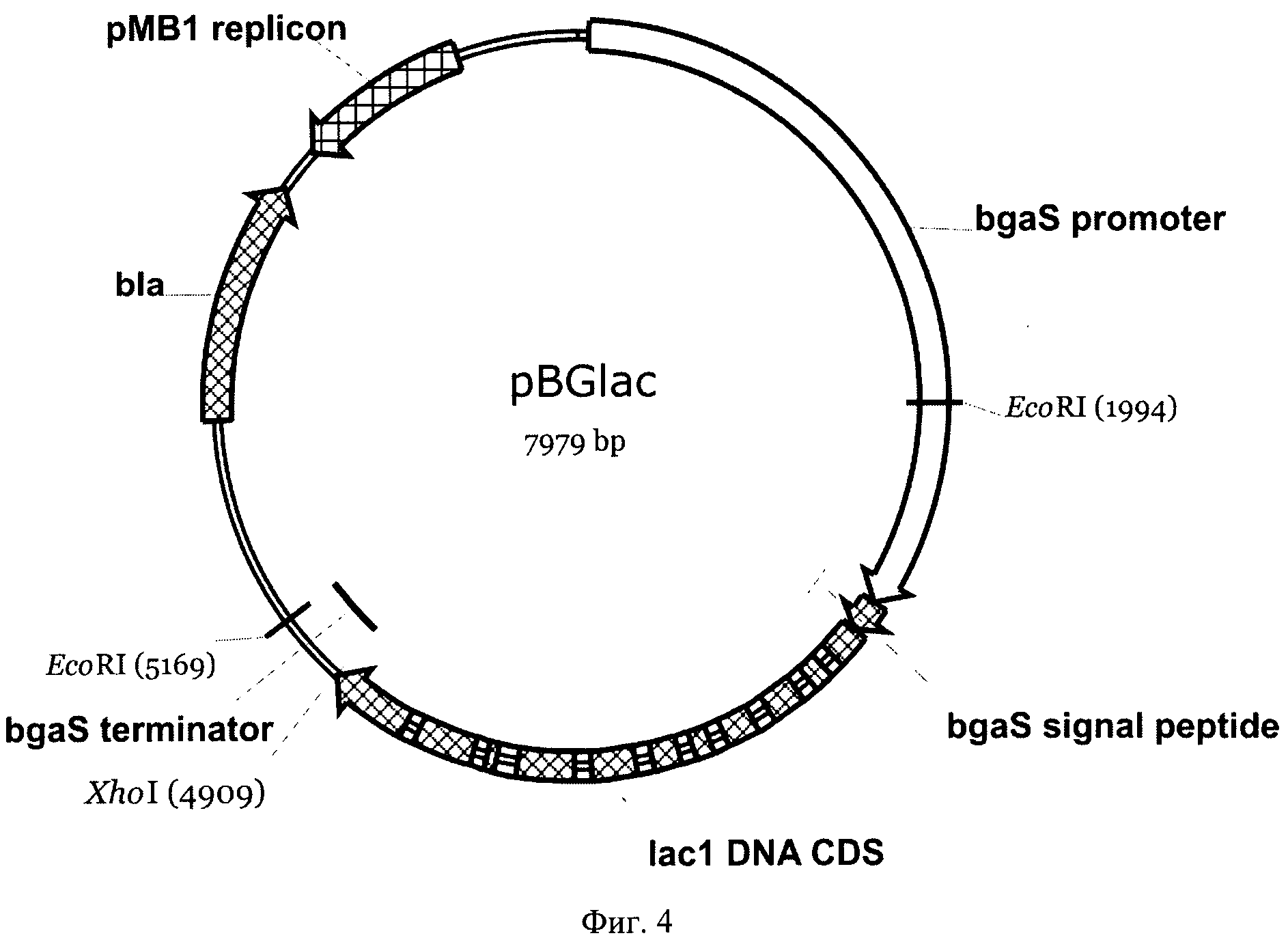
На Фиг.4 приведена карта экспрессионной плазмиды pBGlac. Указаны использованные сайты рестрикции, промотор, сигнальный пептид и терминатор гена bgaS, кодирующая последовательность гена lad из Т. hirsuta без сигнального пептида, ген устойчивости в ампицилину bla и область, обеспечивающая репликацию плазмиды в Е. coli.
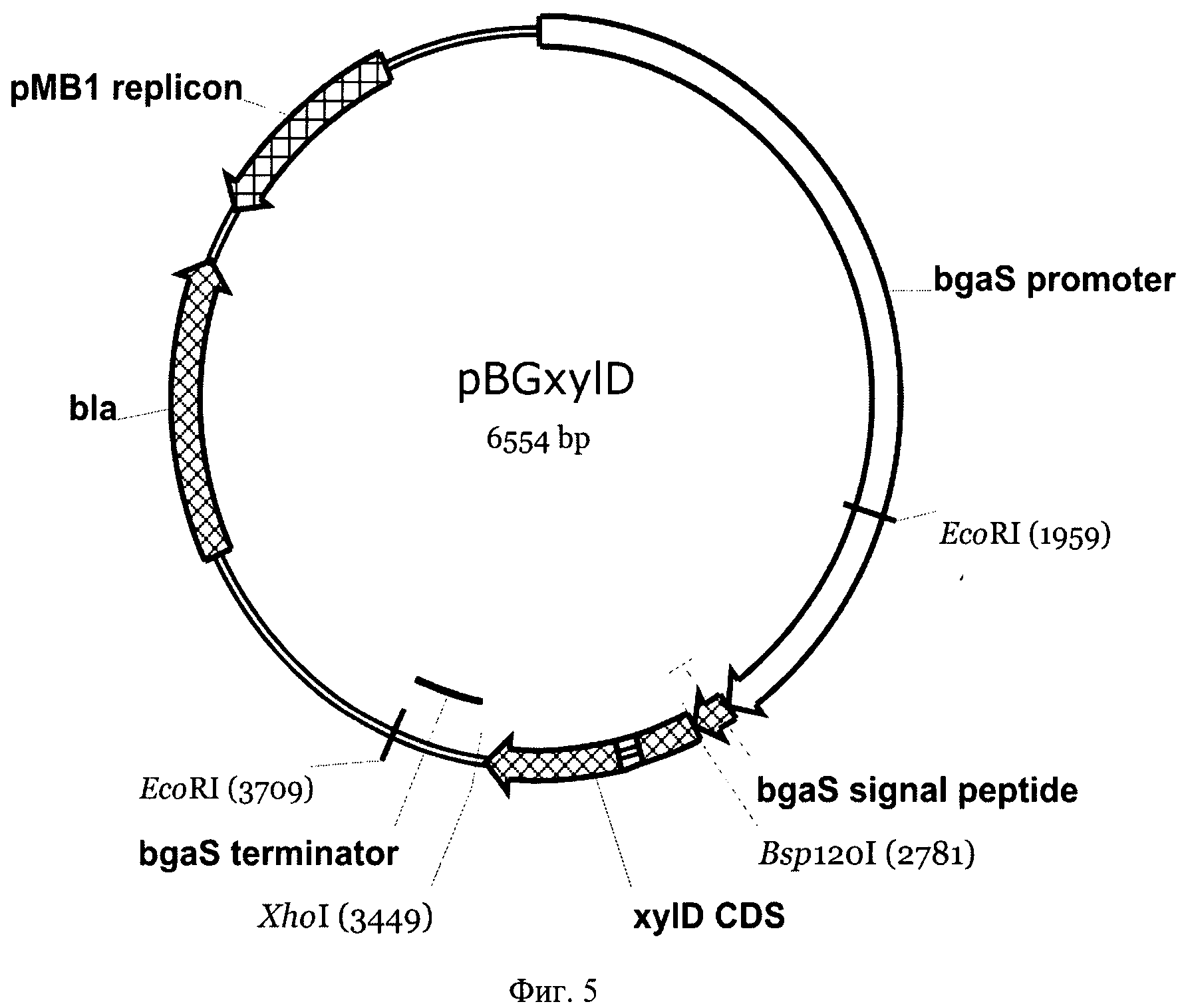
На Фиг.5 приведена карта экспрессионной плазмиды pBGxylD. Указаны использованные сайты рестрикции, промотор, сигнальный пептид и терминатор гена bgaS, кодирующая последовательность гена xylD из Р. canescens без сигнального пептида, ген устойчивости в ампицилину bla и область, обеспечивающая репликацию плазмиды в Е. coli.
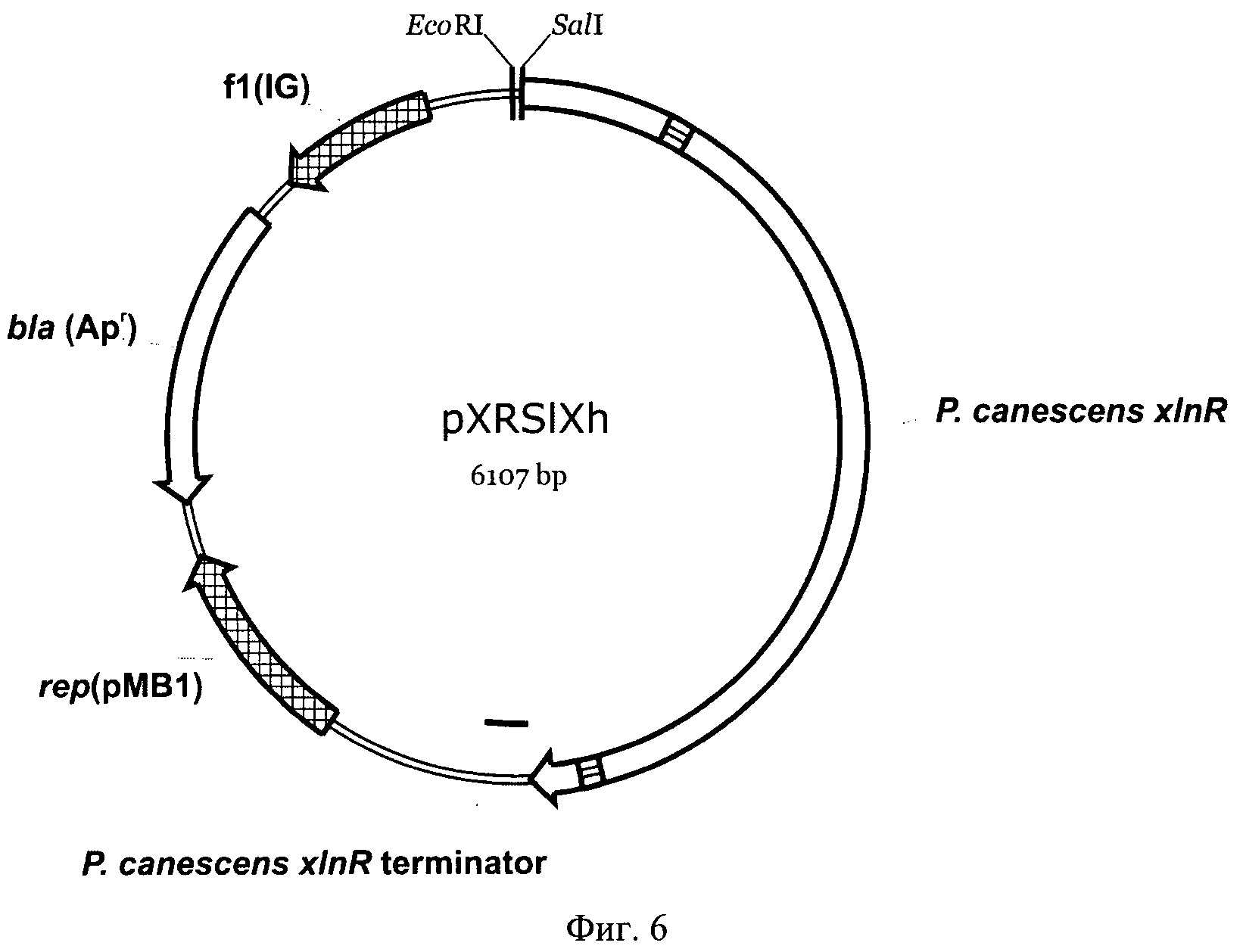
На Фиг.6 приведена карта плазмиды pXRSlXh. Указаны кодирующие области генов xlnR из Р. canescens и bla устойчивости к ампицилину из Е. coli, терминатор гена xlnR, плазмидный и фаговый репликоны rep(pMB1) и f1(IG), соответственно. Также показаны использованные сайты рестрикции.
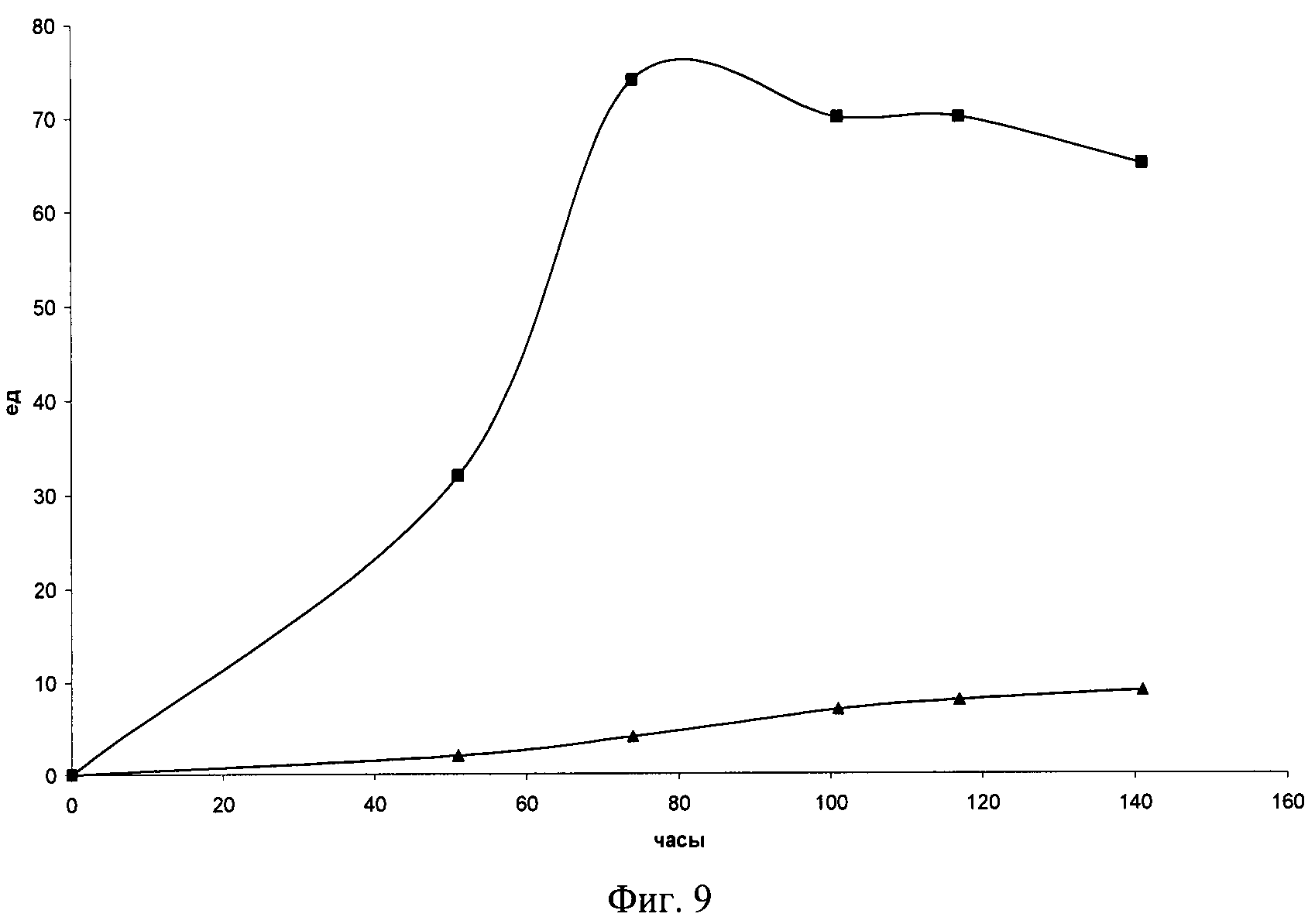
На Фиг.7 приведена карта плазмиды pGpdXlnR. Указаны конститутивный промотор TWugpdA глицеральдегид-3-фосфат-дегидрогеназы из A nidulans, кодирующие области генов xlnR из Р. canescens и bla устойчивости к ампицилину из Е. coli, терминатор генах xlnR, плазмидный и фаговый репликоны rep(pMB1) и f1(IG), соответственно. Показаны использованные сайты рестрикции.
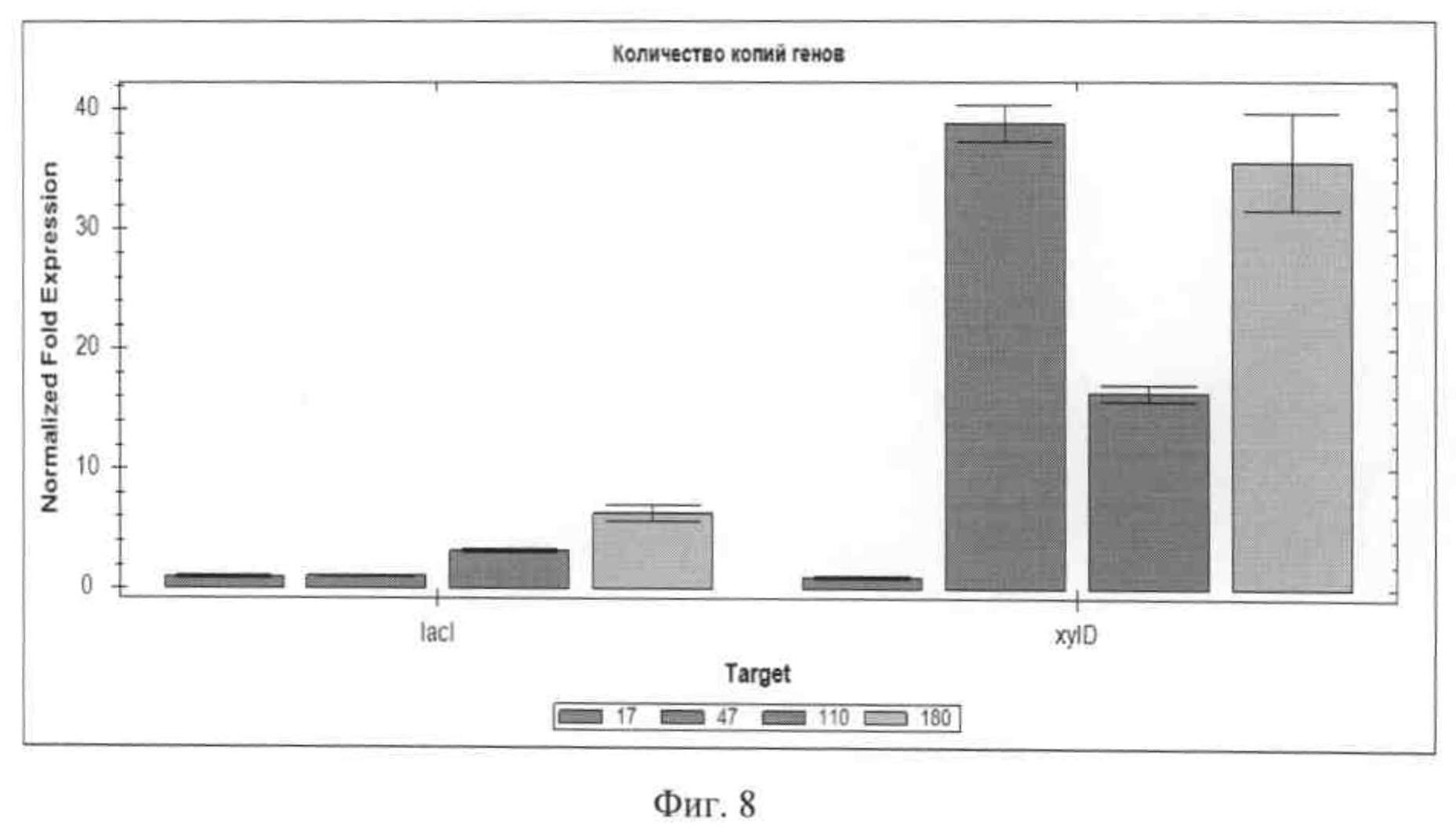
На Фиг.8 приведено графическое представление количества копий генов lac1 из Т. hirsuta и xylD из Р. canescens в трансформантах 17, 47,110,180, определенное методом ПЦР РВ.
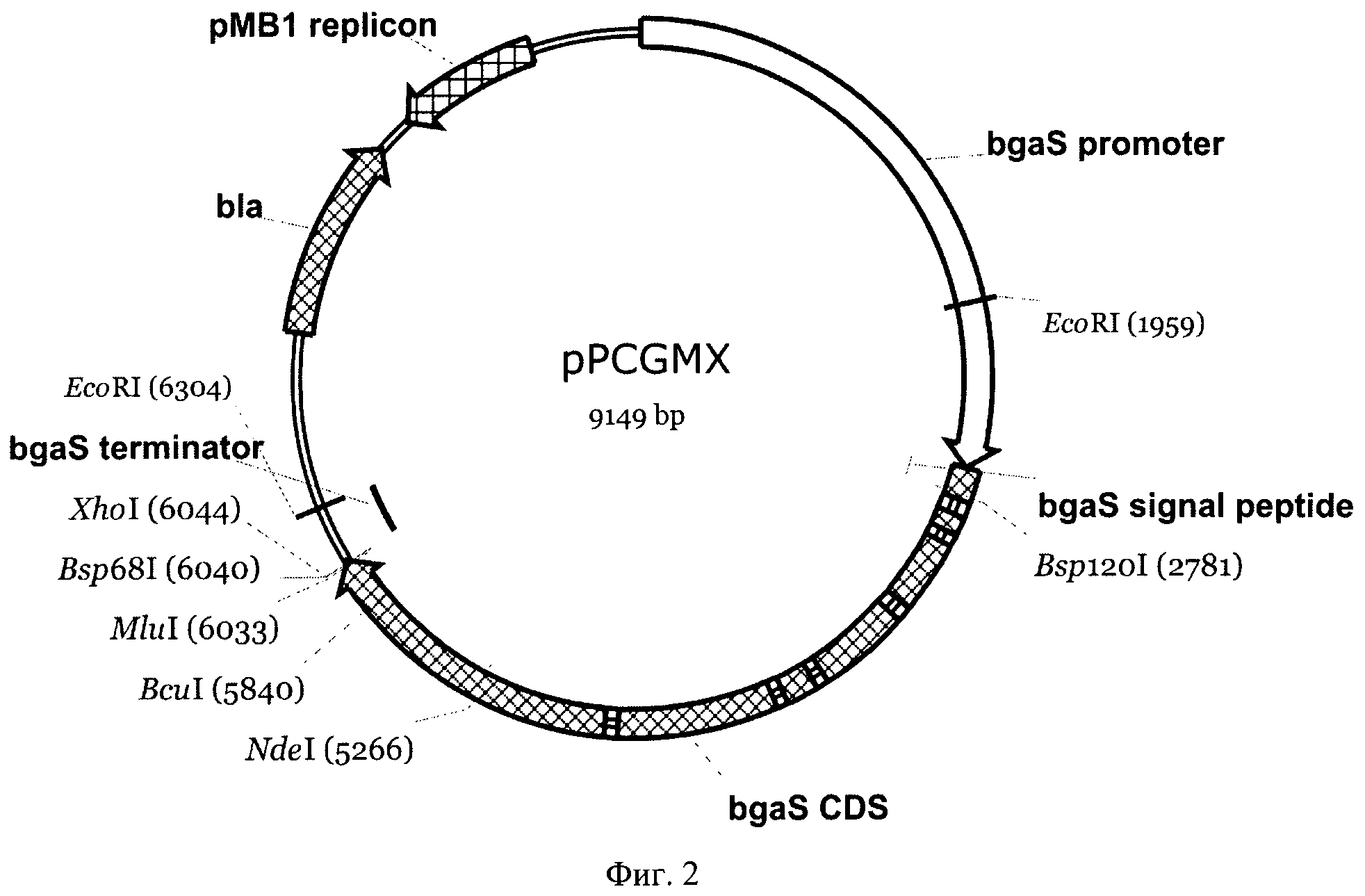
На Фиг.9 показано накопление активностей ксиланазы и лакказы в ходе ферментации.
Подробное описание настоящего изобретения
Для реализации настоящего изобретения главной технической задачей явилось создание штаммов мицелиального гриба Penicillium canescens - продуцентов рекомбинантной лакказы С1 лигнинолитического гриба Trametes hirsuta и ксиланазы XylD мицелиального гриба Penicillium canescens, способных к эффективной продукции и секреции активных целевых белков в культуральную жидкость. Поставленная задача была решена путем конструирования на основе экспрессионного вектора pPCGMX (Пример 1) рекомбинантных плазмид pBGlac и pBGxylD, содержащих гены, кодирующие синтез лакказы С1 лигнинолитического гриба Trametes hirsuta и ксиланазы XylD мицелиального гриба Penicillium canescens, соответственно, а также штамма Penicillium canescens РСА-10-4/1-7/12 niaD-, трансформированного указанными плазмидами pBGlac и pBGxylD и обеспечивающего синтез и продукцию в секретируемой растворимой форме активной лакказы С1 лигнинолитического гриба Trametes hirsuta и активной и ксиланазы XylD мицелиального гриба Penicillium canescens. Поставленная задача также была решена путем разработки и оптимизации способа получения комбинированного ферментного препарата указанных ксиланазы и лакказы с использованием указанного штамма.
Кксиланазы являются ферментами, расщепляющими ксилан - основной компонент гемицеллюлоз. Для мицелиального гриба Penicillium canescens известна эндо-1,4-β-D-ксиланаза (Ху1А), а также описаны штаммы гриба Penicillium canescens ВКПМ F-832 (патент РФ 2197526) и ВКПМ F-912 (патент РФ №2293115) - продуценты секретируемой эндо-(1-4)-β-D-ксиланазы. Недавно авторы настоящего изобретения обнаружили другие эндо-1-4-β-D-ксиланазы, XylB, XylC, XylD и XylE, 10-го и 11-го семейств гликозил-гидролаз мицелиального триба Penicillium canescens (патент РФ №2412246).
Лакказа (ЕС 1.10.3.2 пара-дифенол: кислород оксидредуктаза) является полифенолоксидазой, принадлежащей к семейству голубых оксидаз. Лакказа катализирует окисление редуцирующего субстрата за счет восстановления молекулярного кислорода до воды. Продукты реакции - свободные радикалы редуцирующего субстрата и вода. Обычными субстратами лакказы являются: о-, м-, п-фенолы с различными заместителями, полифенолы, ароматические амины. Основные физико-химические характеристики мажорной изоформы лакказы, секретируемой грибом Trametes hirsuta 072 были недавно описаны, ген этой изоформы был клонирован, и его нуклеотидная последовательность была определена (Ребриков, Д.Н. и др. Прикладная биохимия и микробиология, 42, 6, 645-653 (2006)). Также получение индивидуальной рекомбинантной лакказы С1 лигнинолитического гриба Trametes hirsuta, обладающей высокой каталитической активностью, с использованием штаммов мицелиальных грибов Penicillium canescens недавно описано авторами настоящего изобретения (патент РФ №2394912).
Фрагментом ДНК, кодирующим эндо-1,4-β-D-ксиланазу XylD, является, но не ограничивается им, ген xylD, описанный в патенте РФ №2412246. Структура гена, кодирующего эндо-1,4-β-D-ксиланазу XylD мицелиального гриба Penicillium canescens, его последовательность нуклеотидов и последовательность аминокислот лакказы С1 представлены в патенте РФ №2412246.
Фрагментом ДНК, кодирующим лакказу С1 лигнинолитического гриба Trametes hirsuta, является, но не ограничивается им, ген мажорной лакказы, описанный в статье Ребрикова, Д.Н. и др. (Прикладная биохимия и микробиология, 42, 6, 645-653 (2006)). Структура гена, кодирующего лакказу С1 лигнинолитического гриба Trametes hirsuta, его последовательность нуклеотидов и последовательность аминокислот лакказы С1 представлены в патенте РФ №2394912.
Указанные фрагменты ДНК также могут кодировать варианты эндо-1,4-β-D-ксиланазы XylD мицелиального гриба Penicillium canescens и лакказу С1 лигнинолитического гриба Trametes hirsuta, соответственно.
Термин «вариант» в значении, в котором он используется в настоящем изобретении, означает белок, который имеет изменения в аминокислотной последовательности, а именно делеции, вставки, добавления или замены аминокислот, при условии, что при этом сохраняется необходимый уровень активности белка, например, как минимум 10% от активности целевого фермента. Ряд изменений в варианте белка зависит от положения или от типа аминокислотного остатка в трехмерной структуре белка. Количество изменений может составлять от 1-го до 30-ти, предпочтительно от 1-го до 15-ти и наиболее предпочтительно от 1-го до 5-ти изменений в последовательности соответствующих белков. Эти изменения могут иметь место в областях белка, которые не являются критичными для его функции. Это возможно благодаря тому, что аминокислоты обладают высокой гомологией друг с другом, и поэтому третичная структура или активность белка не нарушаются при таком изменении. Поэтому в качестве варианта белка может выступать белок, который имеет гомологию не менее чем 70%, предпочтительно не менее чем 80%, более предпочтительно, не менее чем 90% и наиболее предпочтительно, не менее чем 95% по отношению к полной аминокислотной последовательности белка при условии, что активность белка сохраняется. Гомология между аминокислотными последовательностями может быть установлена с использованием хорошо известных методов, например с помощью выравнивания последовательностей в компьютерной программе BLAST 2.0, которая вычисляет три параметра: счет, идентичность и сходство.
Замена, делеция, вставка, добавление или замена одного или нескольких аминокислотных остатков будут представлять собой консервативную мутацию или консервативные мутации при условии, что активность белка при этом сохраняется. Примером консервативной мутации(ий) является консервативная замена(ы). Примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Ile на Leu, Met, Val или Phe, замену Leu на Ile, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, Ile или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp и замену Val на Met, Ile или Leu.
Фрагменты ДНК, которые кодируют по существу тот же функциональный белок, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК, кодирующего белок, например, посредством метода сайт-направленного мутагенеза, так, что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации.
Фрагменты ДНК, которые кодируют по существу тот же функциональный белок, могут быть получены путем экспрессии фрагментов ДНК, имеющих мутацию, описанную выше, в соответствующей клетке, и установления активности экспрессируемого продукта. Фрагменты ДНК, которые кодируют по существу тот же функциональный белок, также могут быть получены путем выделения фрагментов ДНК, которые гибридизуются с зондами, имеющими нуклеотидную последовательность, которая содержит, например, нуклеотидную последовательность, кодирующую белок, в жестких условиях, и кодирует белок, обладающие активностью целевого белка. "Жесткие условия", в том значении, которое приписывается этому выражению в рамках настоящего изобретения, означает такие условия, при которых так называемые специфические гибриды образуются, а неспецифические гибриды не образуются. Например, демонстрацией жестких условий могут быть такие условия, при которых фрагменты ДНК, имеющие высокую гомологию, например фрагменты ДНК, имеющие гомологию не менее чем 50%, предпочтительней не менее чем 60%, более предпочтительней не менее чем 70%, еще более предпочтительней не менее, чем 80%, еще более предпочтительней не менее чем 90% и наиболее предпочтительней не менее чем 95% способны гибридизоваться друг с другом, а фрагменты ДНК, имеющие гомологию более низкую, чем описано выше, неспособны гибридизоваться друг с другом. Кроме того, демонстрацией жестких условий могут служить такие условия, при которых фрагменты ДНК гибридизуются при концентрации соли, эквивалентной условиям однократной отмывки при гибридизации по Саузерну, которые составляют 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS, при 60°С. Продолжительность отмывки зависит от типа мембраны, используемой для блотинга и, как правило, рекомендуется производителем набора. Например, рекомендуемая продолжительность отмывки для мембраны HybondTM N+ nylon (Amersham, США), в жестких условиях составляет приблизительно 15 минут. Желательно отмывку повторять 2-3 раза. В качестве зондов может быть также использована неполная нуклеотидная последовательность, кодирующая белок. Зонды могут быть приготовлены с помощью ПЦР с использованием праймеров, синтезированных на основе этой нуклеотидной последовательности и на основе фрагментов ДНК, содержащих эту нуклеотидную последовательность в качестве матрицы. В тех случаях, когда фрагмент ДНК, имеющий длину около 300 п.о., используется в качестве зонда, условия отмывки в процессе гибридизации включают, например, 50°С, 2×SSC и 0.1% SDS.
Замена, делеция, вставка или добавление нуклеотидов, так как они описаны выше, также включает мутации, которые имеют место в природе (мутант или вариант), например, обусловленные видовой изменчивостью.
Показатели функциональной активности, при которой считается, что полученный белок обладает свойствами эндо-1-4-β-D-ксиланазы, определяются по его способности катализировать гидролиз ксилана, расположенного на поверхности целлюлозных волокон. Так, например, активность эндо-1-4-β-D-ксиланазы в культуральной жидкости определяют по количеству восстанавливающих сахаров, выделяемых при гидролизе березового ксилана. Инкубацию субстрата и фермента проводят при 50°С в течение 10 мин. За единицу активности принимают количество фермента, высвобождающее 1 мкмоль/мин восстанавливающих сахаров при указанных выше условиях. Восстанавливающие сахара определяют, например, методом Шомоди-Нельсона (Синицин А.П., Гусаков А.В., Черноглазов В.М. Биоконверсия лигноцеллюлозных материалов. М., Изд-во МГУ, 1995). Считается, что вариант белка обладает свойствами эндо-1-4-β-D-ксиланазы при условии, что активность указанного варианта составляет не ниже 10% активности нативной эндо-1-4-β-D-ксиланазы.
Показатели функциональной активности, при которой считается, что полученный белок обладает свойствами лакказы, определяются по его способности окислять различные субстраты фенольной природы, такие, например, как о-, м-, н-фенолы с различными заместителями, полифенолы, ароматические амины. Также лакказы способны, повышающих эффективность биодеградации лигнина и устойчивых ароматических ксенобиотиков в присутствии медиаторов. Так, например, активность лакказы можно детектировать спектрофотометрически при 410 нм, используя в качестве хромогенного субстрата 10 мМ раствор пирокатехина в 0,1 М Na-ацетатном буфере, рН 4,9, при этом за "условную единицу активности" принимают увеличение оптической плотности в 1 мл реакционной смеси за 1 мин, пересчитанной на 1 мг вносимого белка (Ребриков, Д.Н. и др. Прикладная биохимия и микробиология, 42, 6, 645-653 (2006)).
Активность лакказы также можно определить по измерению начальных скоростей ферментативной реакции с помощью кислородного электрода Кларка в ячейке объемом 0,7 мл при 25°С и постоянном перемешивании. В качестве органического субстрата используют 10 мМ раствор пирокатехина в 0,015 М фосфатном буфере, рН 4,9. Концентрацию кислорода в растворе принимают равной 260 мкМ (согласно значению коэффициента Генри, 773 атм/моль×кг воды). Активность лакказы выражают в мкмолях поглощенного кислорода на 1 мг белка за 1 мин. Считается, что вариант белка обладает свойствами лакказы при условии, что активность указанного варианта составляет не ниже 10% активности нативной лакказы.
Рекомбинантная плазмида согласно настоящему изобретению содержит фрагмент ДНК, кодирующий эндо-1,4-β-В-ксиланазу XylD мицелиального гриба Penicillium canescens или ее вариант, под контролем промотора, функционирующего в эукариотической клетке.
Другая рекомбинантная плазмида согласно настоящему изобретению содержит фрагмент ДНК, кодирующий лакказу С1 лигнинолитического гриба Trametes hirsuta или ее вариант, под контролем промотора, функционирующего в эукариотической клетке.
В качестве рекомбинантных плазмид согласно настоящему изобретению могут быть использованы любые бактериальные плазмиды, такие, например, как pBGxylD и pBGlac, но список плазмид не ограничивается ими.
Рекомбинантная плазмида pBGxylD была сконструирована путем клонирования гена, кодирующего ксиланазу xylD и содержащего кодирующую последовательность без сигнального пептида и стоп-кодона, в плазмиду pPCGMX (см. Пример 1), содержащую полный ген bgaS β-галактозидазы Penicillium canescens, несущий уникальный сайт рестрикции Bspl20I непосредственно после про-области гена bgaS и область линкера перед стоп-кодоном, несущую уникальные сайты MluI, Bsp68I и XhoI.
Аналогичным образом была получена рекомбинантная плазмида pBGlac путем клонирования гена, кодирующего лакказу С1 без сигнального пептида и стоп-кодона, в плазмиду pPCGMX.
При введении данных плазмид в клетку достигается высокий уровень транскрипции генов, кодирующих целевые ферменты, благодаря использованию высокоэффективного промотора гена β-галактозидазы.
В структуру экспрессионных кассет входят нуклеотидная последовательность, содержащая промотор гена β-галактозидазы, последовательность сигнального пептида гена β-галактозидазы Penicillium canescens; последовательность ДНК, кодирующая целевой белок или его вариант; и область терминации транскрипции гена β-галактозидазы Penicillium canescens. При введении данной плазмиды в клетку достигается высокий уровень транскрипции гена, кодирующего целевой белок, благодаря использованию высокоэффективного промотора гена β-галактозидазы.
При помощи созданных плазмид можно трансформировать любую эукариотическую клетку, восприимчивую к подобной трансформации указанными плазмидами. Выбор клетки не является критическим, поскольку методология и приемы трансформации хорошо известны специалисту в данной области техники. И хотя в зависимости от вида клетки и условий культивирования полученного трансформанта уровень экспрессии целевого белка может варьироваться, факт экспрессии целевого белка будет иметь место при условии успешной трансформации клетки-реципиента.
«Трансформация клетки плазмидой» означает введение плазмиды в клетку с помощью методов, хорошо известных специалисту в данной области техники. Трансформация этой плазмидой приводит к усилению экспрессии гена, кодирующего белок согласно настоящему изобретению, и к увеличению активности белка в бактериальной клетке. Методы трансформации включают любые стандартные методы, известные специалисту в данной области техники.
Предпочтительно использование клеток мицелиальных грибов в качестве реципиентов для трансформации рекомбинантной плазмидами, содержащими фрагменты ДНК, кодирующие целевые белки.
Примером штамма-реципиента для получения продуцента рекомбинантной лакказы согласно настоящему изобретению, является, но не ограничивается им, штамм мицелиальных грибов Penicillium canescens PCA-10 (niaD-) (Aleksenko A.Y. et al., Curr. Genet., 28, 474-478 (1995)) или производный штамм Penicillium canescens PCA-10-4/1-7/12 niaD-, трансформированный плазмидами pBGlac и pBGxylD (см. Пример 2).
Способ получения ферментного препарата ксиланазы и лакказы включает стадии выращивания эукариотических клеток, трансформированных описанными выше плазмидами, в питательной среде, подходящей для выращивания указанных эукариотических клеток, и выделения ферментного препарата ксиланазы и лакказы из культуральной жидкости.
Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, различные органические кислоты, а также другие сырьевые материалы, такие как свекловичный жом, отруби и другие подобные материалы. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п. В качестве источника ростовых факторов и аминокислот при культивировании грибных продуцентов также может использоваться кукурузный экстракт.
Авторы настоящего изобретения обнаружили падение активности ксиланазы после 80 ч культивирования вследствие протеолитической деградации ксиланазы секретируемыми протеиназами гриба. Затем авторы настоящего изобретения установили, что мажорная протеолитическая активность в культуральной жидкости связана с присутствием в ней металлопротеиназ, а добавление в ферментационную среду ингибитора металлопротеиназ, ЭДТА, способно предотвращать деградацию ксиланазы и стабилизировать ее активность в ходе ферментации.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как: при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 25 до 35°С. рН среды поддерживают в пределах от 4,0 до 9, предпочтительно от 4,0 до 6,5, наиболее предпочтительно от 4,0 до 5,0. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно, выращивание в течение от 5 до 7 дней приводит к накоплению целевого продукта в культуральной среде.
Очистка целевого продукта может осуществляться любым способом, известным специалисту в данной области техники, таким как, например, фильтрация, мембранная очистка, диализ, осаждение, хроматография с последующим концентрированном и сушкой, например, лиофильной сушкой.
Использование указанного выше способа позволяет проводить синтез и эффективную продукцию в секретируемой растворимой форме комбинированного ферментного препарата ксиланазы и лакказы с высокой активностью.
Примеры
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие примеры, не ограничивающие каким-либо образом рамки настоящего изобретения.
Пример 1. Конструирование плазмид для экспрессии генов ксиланазы и лакказы в Р. canescens.
Для экспрессии гена lac1 из Т. hirsuta (GenBank EU492907) и клонированного ранее авторами настоящего изобретения гена эндо-1-4-бета-ксиланазы xylD из Р. canescens (GenBank FJ 860893, патент РФ №2412246) в мицелиальном грибе Р. canescens использовали сильный промотор гена bgaS β-галактозидазы Р. canescens, ее лидерный пептид и терминатор (GenBank X70408).
На основе плазмиды pPCG2.6 (Вавилова, Е.А. и др. Механизм суперпродукции секретируемых ферментов у мицелиального гриба Penicillium canescens. Прикладная биохимия и микробиология, 2003, т.39, стр.284-292; Фиг.1) был создан экспрессионный вектор pPCGMX (Фиг.2), содержащий полный ген bgaS β-галактозидазы Р. canescens, несущий уникальный сайт рестрикции Bsp120I непосредственно после про-области гена bgaS и область линкера перед стоп-кодоном, несущую уникальные сайты MluI, Bsp68I и XhoI.
На первом этапе в плазмиде pPCG2.6 (Фиг.1) был удален сайт рестрикции XhoI путем рестрикции по этому сайту, достройкой липких концов фрагментом Кленова в присутствии dNTP и последующего лигирования.
На втором этапе был встроен поли-линкер перед стоп-кодоном гена bgaS (Фиг.3). Для этого были получены ПЦР-фрагменты плазмиды pPCG2.6, с использованием следующих пар праймеров N237 (SEQ ID NO:1), N238 (SEQ ID NO:2), N239 (SEQ ID NO:3) и N240 (SEQ ID NO:4).
Полученные фрагменты были элюированы из геля, и подвергнуты второй ПЦР с праймерами N237 (SEQ ID NO:1) и N238 (SEQ ID NO:2). Синтезированный фрагмент был обработан ферментами рестрикции BcuI и EcoRI и клонирован в плазмиде pPCG2.6 с удаленным сайтом XhoI по этим же сайтам вместо интактной последовательности. Так как EcoRI не являлся уникальным сайтом рестрикции в плазмиде, использовали частичное расщепление по этому сайту с последующей обработкой фосфатазой SAP. Таким образом был получен экспрессионный вектор pPCGMX (Фиг.2).
Фрагмент гена lac1, содержащий кодирующую последовательность без сигнального пептида и стоп-кодон был амплифицирован с использованием PfuI полимеразы и пары праймеров с введенными сайтами рестрикции LacEco52 (SEQ ID NO:5) и LacXho2 (SEQ ID NO:6). Геномная ДНК гриба Т. hirsuta была использована в качестве матрицы.
Синтезированный ПЦР фрагмент был обработан рестриктазами Eco52I и XhoI, после чего клонирован в плазмидный вектор pPCGMX, расщепленный по сайтам Bsp120I и XhoI, вместо исходной кодирующей области с сохранением рамки считывания. Таким образом была получена плазмида pBGlac (Фиг.4). Вставленный фрагмент был проверен на отсутствие мутаций секвенированием.
Аналогичным образом была получена плазмида pBGxyID для экспрессии гена xyfD Р. canescens. Фрагмент гена xylD, содержащий кодирующую последовательность без сигнального пептида и стоп-кодон был амплифицирован с использованием PfuI полимеразы и пары праймеров с введенными сайтами рестрикции XlC2ApaI (SEQ ID NO:7) и XlC2XhoI (SEQ ID NO:8). Геномная ДНК гриба Р. canescens была использована в качестве матрицы.
Синтезированный ПЦР фрагмент был обработан рестиктазами Bsp120I и XhoI, после чего клонирован в плазмидный вектор pPCGMX, расщепленный по этим же сайтам, вместо исходной кодирующей области с сохранением рамки считывания. Таким образом была получена плазмида pBGxylD (Фиг.5). Вставленный фрагмент был проверен на отсутствие мутаций секвенированием.
Плазмиды pBGlac и pBGxylD, используемые для трансформации гриба, выделяли с помощью QIAGEN Plasmid Midi Kit.
Пример 2. Получение штамма Р. canescens - продуцента ксиланазы и лакказы.
1. Получение штамма, свободного от углеродной катаболитной репрессии.
В коллекции лаборатории ИНБИ РАН имеется штамм мицелиального гриба Р. canescens F178 (ВКПМ) - природный продуцент (3-галактозидазы и эндо-1,4-β-ксиланазы. Экспрессия генов xylA и bgaS в штамме F178 Р. canescens подвержена репрессии D-глюкозой и индуцируется L-арабинозой. Незначительная индукция экспрессии гена xylA происходит и при росте на D-ксилозе (Николаев И.В., Винецкий Ю.П. Биохимия. 1998, т.63, стр.1523-1528).
Актуальным является получение мутантов, у которых транскрипция генов xylA и bgaS была бы освобождена от углеродной катаболитной репрессии. Для получения таких мутантных штаммов была сконструирована экспрессионная плазмида рХНРН, несущая ген hph гигромицин-Б-фосфотрансферазы Е. coli, которая обеспечивает устойчивость к антибиотику гигромицину Б, под контролем промотора гена xylA из Р. canescens (Чулкин, А. Мутационный анализ углеродной катаболитной репрессии у мицелиального гриба Penicillium canescens. Молекулярная биология, 2011, в печати). Плазмиду вводили в геном штамма Р. canescens РСА-10 (Aleksenko A.Y., et al., Curr. Genet., 1995, 28, 474-478) трансформацией.
После мутагенеза споры выбранного штамма высевали на агаризованную среду, содержащую гигромицин Б в условиях катаболитной репрессии гена xylA. Получена серия мутантных по углеродной катаболитной репрессии штаммов мицелиального гриба Р. canescens. Два мутанта, имеющие наиболее отчетливый фенотип, проанализированы детально. Мутации в исследованных штаммах обусловлены изменениями в гене creA, кодирующим глобальный регулятор углеродной катаболитной репрессии в мицелиальных грибах. Также была показана необходимость присутствия индуктора, арабинозы, для экспрессии генов xylA и bgaS даже в мутантах по углеродной катаболитной репрессии.
Один из этих creA- мутантов (РСА-10/1-7) был выбран как реципиент для дальнейших генетических манипуляций.
2. Изменение уровня экспрессии гена-активатора xlnR в штамме Р. canescens creA-
Ранее был клонирован ген xlnR транскрипционного активатора ксиланолитических генов Р. canescens (Серебряный, В.А. и др., Синтез ксиланолитических ферментов и других карбогидраз в трансформантах гриба Penicillium canescens с измененным числом копий гена транскрипционального активатора xlnR. Прикладная биохимия и микробиология. 2006, т.42, №6, стр.665-673). Показано, что мультипликация этого гена в геноме Р. canescens приводила к значительному увеличению синтеза белка XylA эндоксиланазы Р. canescens, а его делеция к сильному уменьшению.
Как было показано ранее (Mathieu, M. et al., In vivo studies of upstream regulatory cis-acting elements of the alcR gene encoding the transactivator of the ethanol regulon in Aspergillus nidulans, Mol. Microbiol., 2000, v.36, N.1, p.123-131), транскрипция генов-трансактиваторов также может быть подвержена углеродной катаболитной репрессии («механизм двойного замка»). Для снятия углеродной катаболитной репрессии транскрипции гена xlnR была сконструирована плазмидная конструкция, несущая ген xlnR из Р. canescens под контролем конститутивного промотора reHugpdA глицеральдегид-3-фосфат-дегидрогеназы A. nidulans.
Для этого SalI-SalI фрагмент плазмиды рХР53 (Серебряный, В.А. и др. Синтез ксиланолитических ферментов и других карбогидраз в трансформантах гриба Penicillium canescens с измененным числом копий гена транскрипционального активатора xlnR. Прикладная биохимия и микробиология. 2006, т.42, №6, стр. 665-673) длиной 3161 п.н., несущий кодирующую часть гена xlnR без инициирующего ATG кодона и терминатор гена xlnR, был субклонирован в плазмидный вектор pBluescript II KS(+) («Fermentas», Канада), расщепленный эндонуклеазами Sail и XhoI, в ориентации, сохраняющей сайт SalI в кодирующей области xlnR и получена промежуточная плазмида pXRSlXh (Фиг.6).
ПЦР-фрагмент длиной около 2350 п.н., полученный с помощью Pfu ДНК-полимеразы («Fermentas», Канада) с использованием праймеров M13/pUC direct sequencing primer (-46) («Fermentas», Канада) и HPHSL (SEQ ID NO:9), с введенным сайтом рестрикции Sail после инициирующего ATG кодона гена hph из Е. coli, и ДНК плазмиды pAN7-1 (Punt, P.J. et al. Transformation of Aspergillus based on the hygromycin В resistance marker from Escherichia coli. Gene, 1987, v.56, p.117-124) в качестве матрицы, несущий конститутивный промотор гена gpdA из A. nidulans и ATG-кодон, был субклонирован в плазмиду pXRSlXh по сайтам EcoRI и SalI и таким образом была получена плазмида pGpdXlnR (Фиг.7).
Плазмида была введена в геном штамма РСА-10/I-7 (creA-, niaD-) методом ко-трансформации с селективной плазмидой pSTA-10 (Unkles, S.E. et al., The Aspergillus niger niaD gene encoding nitrate reductase: upstream nucleotide and amino acid sequence comparisons. Gene, 1992, v.111, p.149-155), несущей полный ген нитратредуктазы niaD мицелиального гриба Aspergillus niger.
Процедуру трансформации проводили по следующей методике. Штамм Р. canescens PCA-10/I-7 (creA-, niaD-) выращивали 16 часов при 30°С на полной питательной среде: пептон ферментативный - 3 г/л, дрожжевой экстракт - 2 г/л, глюкоза - 10 г/л, NH4Cl - 10 mM, 50× раствор минеральных солей - 20 мл/л. Раствор минеральных солей (50×) имел следующий состав: KCl - 26 г/л, MgSO4×7H2O - 26 г/л, KH2PO4 - 76 г/л, раствор микроэлементов - 50 мл/л. Раствор микроэлементов включал в себя CuSO4×5H2O - 400 мг/л, FeSO4×5H2O - 800 мг/л, MgSO4×2H2O - 800 мг/л, Na2MoO4×2H2O - 800 мг/л, ZnSO4×7H2O - 800 мг/ л, B4Na2O7 - 40 мг/л. Мицелий переносили в раствор 1,2 М MgSO4 и 10 мМ NaH2PO4, pH 5,8 и добавляли лизирующий фермент из Trichoderma harzianum (Sigma, США) (5 мг/мл). Протопластирование проводили в течение 2 часов при 30°С в условиях перемешивания. Суспензию переносили в центрифужную пробирку и наслаивали 1-2 см раствора: 0,6 М сорбитол, 10 мM CaCl, 10 мМ Трис-HCl. После центрифугирования при 3000 об/мин и 4°С в течение 10 мин отбирали интерфазу, содержащую протопласты. Протопласты промывали 2 раза в стабилизирующем растворе SCT (1.2 М сорбитола, 10 мМ Трис, pH 7,5, 10 мМ CaCl2) и ресуспендировали в нем же до концентрации 108 протопластов/мл. Трансформацию проводили следующим образом: к 200 мкл суспензии протопластов добавляли 10 мкг трансформируемой ДНК pGpdXlnR и 1 мкг комплементирующей плазмиды pSTA-10, несущей полный ген niaD нитратредуктазы из A. niger, инкубировали в ледяной бане 20 мин, после чего проводили осмотический шок в течение 5 мин в РСТ (50% PEG 4000, 10 мМ CaCl2, 10 мМ Трис-HCl) и высевали протопласты в верхнем слое на агаризованную селективную минимальную среду, содержащую 1,2 М сорбитола. В качестве селективного маркера использовали источник азота 10 мМ NaNO3, вместо NH4Cl.
Для качественной оценки активности эндоксиланаз полученные трансформанты высевали уколом на агаризованную среду с 5 мМ L-арабинозой и 10 мМ D-глюкозой и выращивали 24 ч при 30°С, после чего заливали верхний слой расплавленной 0,5% агарозой в 0.1М Na-ацетатном буфере рН 5,0, содержащем 1% окрашенный субстрат ксилана Blue Xylan (BX) («BioChemika» 95588) (Chalmers, J.H. et al. Chromogenic and fluorogenic substrates for assaying xylanases of Neurospora. Fungal Genetics Newsletter, 1990, v.37, p.25-31). После инкубации в течение 2-4 ч при 37°С активность эндоксиланаз оценивали по размеру зоны просветления.
Для проверки интеграции плазмиды pGpdXlnR в геном, из 5 штаммов с максимальной зоной просветления была выделена ДНК и использована для ПЦР со специфичными к плазмиде pGpdXlnR праймерами M13/pUC direct sequencing primer (-46) и HPHSL (ЫУЙ ШВ ТЩЖ 9). Результаты ПЦР подтвердили наличие плазмиды в 4 трансформантах. Один из них РСА-10-4/I-7 (pgpdA::xlnR, creA-) был использован в дальнейшей работе.
3. Получение индуцированных мутантов со снятой арабинозной индукцией в штамме Penicillium canescens CreA-.
Как уже упоминалось, экспрессия генов xylA и bgaS в штамме гриба Р. canescens индуцируется L-арабинозой. При этом сама L-арабиноза является дорогостоящим субстратом, использование которого при крупномасштабном производстве нерентабельно. Таким образом, актуальной является задача получения мутантных культур гриба, у которых активационный уровень транскрипции целевых генов достигается в отсутствии L-арабинозы.
Для решения поставленной задачи был использован следующий методический подход. Поскольку штамм РСА-10-4/I-7 (pgpdA::xlnR, creA-) содержит в геноме гибридный ген hph гигромицин-Б-фосфотрансферазы из Е. coli под контролем промотора гена xylA, искомые мутанты могут быть отобраны по фенотипу hphR на среде, не содержащей L-арабинозу.
При получении мутантов была подобрана концентрация гигромицина Б (1000 мкг/мл) на среде ММ с 0,4% глюкозы на которой отсутствовал рост конидий штамма PCA-10-4/I-7 (pgpdA::xlnR, creA-).
Споровая суспензия штамма PCA-10-4/I-7 (титр 3,2×107) приготовили из 14-суточной культуры гриба, выращенной на среде ММ. Суспензию облучали УФ в течение 20 с, достигая эффективности выживания спор 6%. Облучение проводили при постоянном перемешивании. Конидии высевались на агаризованную среду ММ, содержащую глюкозу (0,4%), гигромицин Б (1000 мкг/мл) и X-gal (5-бром-4-хлор-3-индолил-бета-D-галактопиранозид, 100 мг/л) для получения 300 клонов на чашке с учетом эффективности выживания. Чашки инкубировали 7 суток при 30°С.
Получено 3,2×106 клонов с фенотипом hphR. Среди них было отобрано 36 синих клона, продуцирующих β-галактозидазу в данных условиях (фенотип bgs+). Отобранные мутанты фенотипа hphR bgs+ были переколоты на среду для их отбора (см. выше) для повторного тестирования фенотипа. По результатам этого тестирования были выявлены синие клоны с воспроизводимым фенотипом. Мутантый клон PCA-10-4/I-7/12 был использован в дальнейшей работе.
Было подтверждено, что мутация в штамме PCA-10-4/I-7/12 приводит к арабинозо-независимой индукции не только β-галактозидазы и эдоксиланазы, но и ряда других секреторных белков Р. canescens.
4. Получение штамма-реципиента Penicillium canescens со снятой катаболитной репрессией и арабинозной индукцией.
Для последующего использования мутантного штамма PCA-10-4/I-7/12 при создании на его основе штаммов-продуцентов необходимо решить задачу введения селектируемой мутации. В качестве такой мутации была использована мутация по гену нитратредуктазе (niaD-).
Получение мутантов по нитратредуктазе (niaD-) проводили методом индуцированного мутагенеза. Для этого споровую суспензию штамма PCA-10-4/I-7/12 обрабатывали N-нитро-N-нитрозогуанидином (200 мкг/мл) 30 минут при 30°С. Затем отмывали 3 раза в 0,1 М фосфатном буфере, рН 7,0 и высевали на чашки с ММ. Чашки инкубировали 7 суток. Выживаемость спор при такой обработке составила 10%.
Выросшие клоны пересевали на матричные чашки с различными источниками азота (10 мМ NH4Cl, 10 мМ NaNO3, 0,45 М хлорат Na, 5 мМ гипоксантин, 20 мМ NaNO2). Отсутствие роста клонов на среде с 10 мМ NaNO3 и его наличие на других источниках азота говорит о наличие фенотипа niaD-. По результатам этого ростового теста отобрано 18 клонов. Все они были проверены по эффективности секреции β-галактозидазы при выращивании на агаризованной среде ММ, содержащей глюкозу (10 г/л) и X-gal (5-бром-4-хлор-3-индолил-бета-D-галактопиранозид, 100 мг/л).
По результатам проведенного теста отобран мутант с максимальной продукцией β-галактозидазы, обозначенный РСА-10-4/I-7/12 niaD-, который в дальнейшем использовали в качестве реципиентного штамма.
Пример 3. Получение штамма Р. canescens - продуцента ксиланазы и лакказы.
Для получения штамма-продуцента ксиланазы и лакказы провели трансформацию реципиентного штамма гриба РСА-10-4/I-7/12 niaD- плазмидами pBGlac и pBGxylD.
Процедуру трансформации проводили по следующей методике. Штамм Р. canescens РСА-10-4/I-7/12 niaD- выращивали 24 часов при 30°С на полной питательной среде: пептон ферментативный - 3 г/л, дрожжевой экстракт - 2 г/л, глюкоза - 10 г/л, NH4Cl - 10 мМ, 50× раствор минеральных солей - 20 мл/л. Раствор минеральных солей (50×) имел следующий состав: KCl - 26 г/л, MgSO4×7H2O - 26 г/л, KH2PO4 - 76 г/л, раствор микроэлементов - 50 мл/л. Раствор микроэлементов включал в себя CuSO4×5H2O - 400 мг/л, FeSO4×5H2O - 800 мг/л, MgSO4×2H2O - 800 мг/л, Na2MoO4×2Н2О - 800 мг/л, ZnSO4×7H2O - 800 мг/ л, B4Na2O7 - 40 мг/л. Мицелий переносили в раствор 1,2 М MgSO4 и 10 мМ NaH2PO4, pH 5,8 и добавляли лизирующий фермент из Trichoderma harzianum (Sigma, США) (35 мг/мл). Протопластирование проводили в течение 2 часов при 30°С в условиях перемешивания. Суспензию переносили в центрифужную пробирку и наслаивали 1-2 см раствора: 0,6 М сорбитол, 10 мМ Трис-HCl. После центрифугирования при 5000 об/мин и 4°С в течение 10 мин отбирали интерфазу, содержащую протопласты. Протопласты промывали 2 раза в стабилизирующем растворе SCT (1,2 М сорбитола, 10 мМ Трис, pH 7,5, 10 мМ CaCl2) и ресуспендировали в нем же до концентрации 108 протопластов/мл. Трансформацию проводили следующим образом: к 200 мкл суспензии протопластов добавляли 10 мкг трансформируемой ДНК плазмид pBGlac и pBGxylD и 1 мкг комплементирующей плазмиды pSTA-10, несущей полный ген niaD нитратредуктазы A. niger, инкубировали в ледяной бане 20 мин, после чего проводили осмотический шок в течение 5 мин в РСТ (50% PEG 4000, 10 мМ CaCl2, 10 мМ Трис-HCl) и высевали протопласты в верхнем слое на агаризованную селективную минимальную среду, содержащую 1,2 М сорбитола. В качестве селективного маркера использовали источник азота 10 мМ NaNO3, вместо NH4Cl.
Получен 201 клон ко-трансформантов, которые охарактеризованы в ходе последующего 2-этапного скрининга.
Проведение Скрининга №1
Первый этап скрининга проводили по оценке активности продуцируемой лакказы в условиях тестирования на чашках. Для этого споровые культуры грибных клонов выращивали в течение 10 суток на чашках со средой следующего состава:
|
После этого клоны были пересеяны путем перекалывания на чашки с другой средой:
|
После выращивания материала в течение 1 суток при 30°С чашки заливали верхним слоем агара (0,5%), содержащим 1,1 мг/мл АВТС в 0,1 М натрий-ацетатном буфере (рН 5,0) и инкубировали в течение 2 ч при комнатной температуре (openwetware.org/wiki/laccase_protocols).
Клоны, продуцирующие лакказу, образуют в этих условиях зеленый ореол вокруг колоний. По результатам проведенного теста отобрано 44 клона трансформантов.
Проведение Скрининга №2
Целью данного этапа скрининга был отбор клонов, демонстрирующих максимальные уровни продукции ксиланазы и лакказы. Для этого клоны, отобранные на предыдущем этапе, культивировали в плашках с последующим анализом культуральной жидкости.
Культивирование в планшетах проводили после посева уколом при 30°С в течение 5 суток на среде следующего состава:
|
Культуральные жидкости, отобранные после окончания эксперимента были проанализированы методом элекрофореза в ДДС-ПААГ 14%.
Культуральные жидкости, отобранные после культивирования клонов в плашках, были использованы для определения в них активности лакказы. Активность лакказы в культуральной жидкости штаммов детектировали спектрофотометрически (длина волны 450 нм), используя в качестве хромогенного субстрата 10 мМ раствор АБТС в 0,1 М Na-ацетатном буфере рН 5,0. Реакцию проводили в плашках в течение 1 мин. Реакционная смесь состояла из 150 мкл субстрата и 15 мкл культуральной жидкости. Культуральную жидкость штамма Р. canescens PCA-10-4/I-7/12 niaD- использовали в качестве отрицательного контроля. Результаты измерения активности лакказы в культуральных жидкостях клонов-трансформантов приведены в таблице 1.
|
Отбор трансформированных клонов со множественной встройкой экспрессируемых генов ксиланазы и лакказы.
Для определения количества копий генов lad из Т. hirsuta и xylD из Р. canescens в коллекции трансформантов был использован метод ПЦР в реальном времени.
На первом этапе был идентифицирован однокопийный продуцент лакказы. Для этого из трансформантов №№6, 7, 17, 18 и 21, имеющих активность лакказы, была выделена высокомолекулярная геномная ДНК с использованием следующей методики:
1) Споры гриба смывали с чашки, суспендировали в 100 мл минимальной среды (до концентрации 1×106-5×106 спор/мл), и растили 12-24 часа.
2) Мицелий отфильтровывали, промывали буфером и отжимали на фильтровальной бумаге.
Состав буфера:
|
3) 1 г грибного мицелия растирали в ступке, замораживая в жидком азоте, затем переносили в 5 мл буфера и осторожно перемешивали. Добавляли 330 мкл 10% SDS (до 0,5%) и протеиназу K до 0,1 мкг/мл и снова осторожно перемешивали. Инкубировали при 65°С в течение 30 мин.
4) После инкубирования к суспензии мицелия добавляли 1,5 мл 5М ацетата калия pH 5,0, осторожно перемешивали и инкубировали 30-60 мин в ледяной бане.
5) Центрифугировали 10 000 об/мин, 10 мин, 4°С.
6) Надосадочную жидкость отбирали; при необходимости фильтровали через два слоя марли, и добавляли к ней 15 мл этанола. Инкубировали 10-20 мин при -20°С.
7) Центрифугировали 20 мин при 10 000 об/мин и температуре 4°С.
8) Надосадочную жидкость сливали, осадок промывали 70% этанолом, подсушивали 10 мин при 37°С и растворяли в 3 мл деионизованной воды.
9) При необходимости удалить РНК добавляли 300 мкг РНК-азы А и инкубировали 30 мин при 37°С.
10) К раствору добавляли 1 объем (3 мл) изопропанола и аккуратно перемешивали. Выпавшую в осадок ДНК вынимали из раствора, наматывая на наконечник от пипетки, и переносили в 70% этанол, или осаждали центрифугированием 10 мин при 10000 об/мин и промывали 70% этанолом.
11) Осадок подсушивали 10 мин при 37°С и растворяли в 100-500 мкл деионизованной воды.
5 мкг ДНК обрабатывались 10 единицами эндонуклеазы рестрикции SmaI в течение ночи при 30°С, после чего ДНК разделяли в 0,8% агарозном геле, переносили на нейлоновую мембрану и фиксировали ультрафиолетом.
Мембрану гибридизовали с α-32P меченным с помощью DecaLabel™ DNA Labeling Kit (Fermentas, Канада) ПЦР-фрагментом геном lacl, полученным с использованием праймеров LacEco52 (SEQ ID NO:5) и LacXho2 (SEQ ID NO:6).
Для блот-гибридизации ДНК мембраны "HybondTM-N" (Amersham). Гибридизацию проводили при температуре 65°С 18 часов. Концентрации растворов для гибридизации и отмывки фильтров соответствовали рекомендациям производителя мембран:
Раствор для предгибридизации и гибридизации:
5×SSC
5×раствор Денхарда
0,5% ДДС
Денатурированная ДНК из спермы лосося 20 мкг/мл
Отмывка фильтров:
2×SSC и 0,1% ДДС - 10 мин, 65°С
1×SSC и 0,1% ДДС - 15 мин, 65°С
0.1×SSC и 0,1% ДДС - 10 мин, 65°С 2 раза.
Так как сайт рестрикции SmaI уникален в плазмиде pBGlac и находится в середине кодирующей части гена, то однокопийный трансформант 17 показывал на радиоавтографе наличие только двух уникальных полос, в отличие от мультикопийных продуцентов.
Для определения количества копий генов lacl и xylD из нескольких трансформантов с высокими активностями ксиланазы и лакказы выделяли ДНК, разрушая клетки стеклянными шариками размером 0,05 мм на приборе FastPrep®-24 (МР, Великобритания), используя следующую методику:
1) Гриб выращивали на чашке с минимальной средой, до образования спор (5-7 суток).
2) 0,25-1 см2 мицелия вырезали шпателем или скальпелем вместе с агаром и переносили в 2 мл микропробирку с крышкой, содержащую 50-100 мкг стеклянных шариков и 250 мкл буфера следующего состава:
Буфер:
|
3) Пробирку помещали в гомогенизатор FastPrep®-24 и обрабатывали 3 раза по 30 сек при максимальной мощности.
4) Объем раздробленного мицелия измеряли, добавляли 1/20 объема 10% SDS, перемешивали и инкубировали при 65°С в течение 30 мин.
5) После инкубирования к суспензии мицелия добавляли ¼ объема 5М ацетата калия (рН 5,0), перемешивали и инкубировали в ледяной бане 30-60 мин.
6) Суспензию центрифугировали 10 мин при 10000-13000 об/мин, надосадочную жидкость отбирали и добавляли к ней двойной объем этанола, или равный объем изопропанола. Инкубировали 10-20 мин при температуре минус 20°С.
7) Центрифугировали 10 мин при 10000-13000 об/мин, надосадочную жидкость сливали, осадок промывали 70% этанолом, подсушивали 10 мин при 37°С и растворяли в 100 мкл деионизованной воды.
8) При наличии в растворе ДНК нерастворимых фрагментов или взвеси раствор центрифугировали 10 мин при 10000-13000 об/мин, надосадочную жидкость переносили в чистую пробирку.
Полученная ДНК была использована для проведения ПЦР РВ с праймерами на гены lac1 из Т. hirsutus, xylD из Р. canescens и клонированный нами ранее ген gpdA глицеральдегид 3-фосфат дегидрогеназы Р. canescens (GenBank GQ996946): PCRTlacD1 (SEQ ID NO:10), PCRTlacR1 (SEQ ID NO:11), XDRTD (SEQ ID NO:12), XDRTR (SEQ ID NO:13), PCRTGPDD1 (SEQ ID NO:14), PCRTGPDR1 (SEQ ID NO:15).
Для амплификации использовали Комплект реагентов для проведения ПЦР-РВ в присутствии EVA Green («Компания СИНТОЛ», Россия). ПЦР-РВ проводили на приборе Bio-Rad CFX 96 («Bio-Rad Laboratories», США) в трех технических повтора для каждой пробы. Программа ПЦР-РВ: 95°С - 5 мин; (95°С - 15 сек; 60°С - 40 сек) 40 циклов. Анализ производили, используя программное обеспечение Bio-Rad CFX Manager v.1.6.541.1028, («Bio-Rad Laboratories», США). Так как количество копий гена gpdA не изменялось при трансформации, то он был выбран в качестве референсного гена, а изменение копий генов lac1 и xylD измерялось по отношению к однокопийному по гену lac1 штамму. Результаты ПЦР показаны на Фиг.8. Количественные данные приведены в таблице 2.
|
Как видно из полученных данных, количество копий гена lac1 слабо коррелирует с активностью лакказы в культуральной жидкости, что можно объяснить "эффектом положения", когда гены встраиваются в различные участки хроматина. В результате для дальнейшей работы отобраны три клона №47, №110 и №180.
Отбор клона с максимальным уровнем выхода активных белковых продуктов
Для изучения максимальной активности ксиланазы и лакказы отобраны следующие номера клонов-трансформантов 47, 110, 180 и штамм реципиент Р. canescens РСА-10-4/I-7/12 niaD-, которые культивировали в колбах с последующим анализом культуральной жидкости.
Для получения посевного материала штаммы выращивали на агаризованной среде ММ в течение 14 суток. Водной суспензией конидий (105 конидий/мл) инокулировали 100 мл среды, следующего состава:
|
Культивирование проводили в колбах на орбитальной качалке (250 об/мин) при 30°С в течение 6 суток. По завершению выращивания определяли активности эндоксиланазы и лакказы в культуральной жидкости (КЖ). Кроме этого, культуральная жидкость была проанализирована методом электрофореза в ПААГ. Результаты определения активностей ферментов ксиланазы и лакказы в культуральной жидкости представлены в таблице 3.
Активность эндо-1-4-β-D-ксиланазы в культуральной жидкости определяли по ГОСТ Р 53047-2008 с использованием березового ксилана в качестве субстрата. Для чего проводили инкубацию субстрата и фермента при температуре 50°С в течение 10 мин. Проба содержала 0,45 мл 1% раствора ксилана в 0,1 М ацетатном буферном растворе рН 5,0 и 0,05 мл фермента в соответствующем разведении. За единицу активности принимали количество фермента, высвобождающее 1 мкмоль/мин восстанавливающих сахаров при указанных выше условиях. Восстанавливающие сахара определяли методом Шомоди-Нельсона (А.П.Синицин, А.В.Гусаков, В.М.Черноглазов. Биоконверсия лигноцеллюлозных материалов. - М.: Изд-во МГУ, 1995).
Активность лакказы в культуральной жидкости штаммов детектировали спектрофотометрически (длина волны 410 нм), используя в качестве хромогенного субстрата 10 мМ раствор ПКХ в 0,1 М Na-ацетатном буфере рН 4,9. Пирокатехин был предварительно очищен возгонкой в вакууме. Реакционная смесь содержала 2 мл субстрата и 100 мкл культуральной жидкости.
|
В результате клоны №47, №110 и №180 могут считаться эффективными продуцентами ксиланазы и лакказы.
Пример 4. Оптимизация условий культивирования культивирования штамма Р. canescens - продуцента рекомбинантных ксиланазы и лакказы.
В дальнейшей работе был использован штамм гриба Penicillium canscens PCA-10-4/I-7/12 №47.
Для определения оптимума рН среды для культивирования штамм РСА-10-4/I-7/12-47 для получения посевного материала выращивали на агаризованной среде следующего состава:
|
Раствор минеральных солей (50×) имел состав: KCl - 26 г/л, MgSO4×7Н2О - 26 г/л, KH2PO4 - 76 г/л, микроэлементы - 50 мл/л. 50× раствор минеральных солей включал в себя CuSO4×5H2O - 400 мг/л, FeSO4×5H2O - 800 мг/л, MgSO4×2H2O - 800 мг/л, Na2MoO4×2H2O - 800 мг /л, ZnSO4×7H2O - 800 мг/ л, B4Ma2O7 - 40 мг/ л.
Выращивание проводили при 30°С в течение 7-10 суток до стадии конидиального спороношения.
По завершении этой стадии конидии смывали стерильной дистиллированной водой. Водной суспензией конидий инокулировали 100 мл среды (конечный титр засева - 1×105 конидий/мл).
Была использована ферментационная среда следующего состава:
|
В этой среде в качестве источника углерода используется глюкоза, поскольку штамм-продуцент является мутантом creA-. Кроме этого, из состава среды исключен индуктор транскрипции целевых генов - арабиноза, поскольку реципиентный штамм получен как мутант с арабинозо-независимой индукцией. Концентрацию кукурузного экстракта в среде варьировали от 1% до 5%.
Культивирование проводили в ферментационной среде (100 мл) на орбитальной качалке при 240-250 об/мин в качалочных колбах на 750 мл в течение 140 часов. После этого культуральную жидкость отделяли центрифугированием в течение 3 мин при 12000 об/мин. В результате установлено, что уровень активности лакказы не зависит ни от рН среды культивирования, ни от концентрации кукурузного экстракта. В противоположность этому установлено, что продукция ксиланазы xylD сильно зависит от рН среды культивирования, наблюдается в диапазоне рН 4,0-6,0, имея максимум при рН 4,5. По результатам проведенного эксперимента оптимальными условиями признаны: рН среды культивирования 4,5; содержание в среде кукурузного экстракта 3%.
Выбранные условия культивирования был проверены в ходе ферментации в аппарате Biostat Bplus (Sartorius) с рабочим объемом 2,0 л. Для этого штамм Penicillium canscens PCA-10-4/I-7/12 №47 выращивали по следующей схеме:
Посевной материал выращивали на агаризованной среде при 30°С в течение 7-10 суток. Водной суспензией конидий инокулировали 100 мл посевной среды (конечная концентрация - 1×105 конидий/мл) следующего состава:
|
Культивирование проводили в ферментационной среде на орбитальной качалке при 240-250 об/мин в качалочных колбах на 750 мл в течение 24 часов.
Ферментер засевали суточным посевным материалом в количестве 200 мл.
Использовалась ферментационная среда следующего состава:
|
Ферментацию проводили в 1,5 л среды при 30°С. Аэрацию в аппарате осуществляли с помощью двухъярусных грибных импеллеров при 300 об/мин. Подача стерильного воздуха составляла 1 V/V/мин.
Активности лакказы и ксиланазы определяли как описано выше. Результаты представлены на Фиг.9. Как видно из представленных данных, в ходе ферментации происходит постоянное накопление активности лакказы. Активность ксиланазы накапливается после 1 суток культивирования и начинает снижаться после 80 ч культивирования.
Как было далее установлено, одной из причин падения активности ксиланазы являлась протеолитическая деградация ксиланазы секретируемыми протеиназами гриба. При этом мажорная протеолитическая активность в культуральной жидкости связана с присутствием в ней металлопротеиназ.
С учетом полученных по ингибированию данных, приведенных выше, проверена продукция и стабильность ксиланаз при ферментации продуцента в присутствии ингибитора маталлопротеиназ - ЭДТА. Установлено, что максимальная продукция целевых ферментов достигается при добавлении ЭДТА в концентрации 0,2 мМ на 2-е сутки роста.
Пример 5. Подбор методов очистки и концентрирования ферментов
Ксиланаза и лакказа являются экстарцеллюлярными (внеклеточными) ферментами, поэтому в качестве первой стадии очистки и выделения целевых ферментов можно рассматривать стадию отделения биомассы. Наиболее современными методами отделения биомассы являются баромембранные методы, а именно микрофильтрация.
Культуральную жидкость, полученную после культивирования штамма Penicillium canscens PCA-10-4/I-7/12 №47 с активностью ксиланазы 450 Ед/мл и активностью лакказы 1,12 Ед/мл фильтровали на микрофильтрационной мембране 0,45 мкм при давлении 2 Бар (=1,974 Атм) и температуре 20°С. Степень концентрирования составляла K=2. Как показал проведенный опыт, при микрофильтрации культуральной жидкости не происходит разделения биомассы и ферментов. Происходит лишь концентрирование культуральной жидкости. Процесс диафильтрации в данном случае значительно не влияет на вымывание фермента. По-видимому, полученный результат можно объяснить двумя причинами: адсорбцией ферментов на биомассе и образованием высокомолекулярных соединений с фильтрационными характеристиками, не соответствующими ситовой фильтрации. Таким образом, отделение биомассы мембранными методами не эффективно.
Далее проводили изучение эффективности концентрирования фермента на мембранах с молекулярной отсечкой 5 кДа, 10 кДа и 15 кДа (ультрафильтрация).
Осветленный фильтрат культуральной жидкости с рН 5,5 концентрировали на лабораторной ультрафильтрационной установке LabScale производства MILLIPORE (США) в тангенциальном потоке.
Технические характеристики системы:
|
В соответствии с данными электрофореза культуральной жидкости ксиланаза А, ксиланаза Д и лакказа имеют молекулярную массу около 30-и, 25-и 70-и кДа соответственно. В связи с чем для изучения возможности мембранной очистки и концентрирования ферментов в работе использовали кассеты типа Pellicon XL 50 с мембранами, предел отсечения которых составлял 5 кДа, 10 кДа и 30 кДа. Кассеты Pellicon XL 50 подсоединяются к установке LabScale и обладают линейной масштабируемостью, т.е. параметры процесса могут быть пересчитаны для работы на кассетах Pellicon с большей площадью фильтрации.
Технические характеристики кассеты Pellicon XL 50:
|
Процесс ультрафильтрации проводили при давлении Р=3 Бар (=2,961 Атм), с температурой Т=20°С. Степень концентрирования составляла K=5. По достижении степени концентрирования K=3 проводили процесс диафильтрации водой (V=2 л). Затем снова концентрировали до K=5.
В ходе проведенного опыта было показано, что ультрафильтрация культурального фильтрата в испытанных параметрах процесса не оказывает существенного инактивирующего влияния на целевые ферменты. Однако на мембранах предел отсечения которых составлял 10 кДа и 30 кДа наибольшая ксиланазная активность детектировалась в пермеате, в то время как на мембране с пределом отсечения 5 кДа, в пермеате ксиланазная активность детектировалась в минимальных количествах. Таким образом, для дальнейших исследований была выбрана ультрафильтрационная мембрана с пределом отсечения 5 кДа.
Далее изучали возможность получения концентрата целевых ферментов методом вакуум-выпаривания.
В работе также были получены образцы ферментного препарата путем имитации в лабораторных условиях вакуум-выпаривания с использованием ротационного испарителя Hei Vap Advantage HB/G3B производства Heidolph (Германия) в комплекте с вакуумным насосом Rotavac valve control.
Технические характеристики системы:
|
Процесс вакуум-выпаривания осветленного фильтрата проводили при следующих температурах: Т=50, 55 и 60°С. Степень концентрирования составляла K=4 и K=8. В ходе проведенного опыта было показано, что при температурах Т=55 и 60°С происходит значительная (более 30%) инактивация ксиланазы. Проведение процесса вакуум-выпаривания при температуре Т=50°С не оказывает инактивирующего влияния на лакказу, при этом инактивация ксиланазы составляет около 10% при K=4 и около 15% при K=8. Следовательно оптимальной температурой для получения концентрата целевых ферментов является температура процесса вакуум-выпаривания Т=50°С.





Вариабельные домены легкой и тяжелой цепи мышиного моноклонального антитела против фактора некроза опухоли альфа (фно-α) человека (варианты), антигенсвязывающий фрагмент (fab) против фно-α человека, содержащий указанные домены (варианты)
Способ удаления пеногасителя из культуральной жидкости после ферментации
Рекомбинантный фрагмент днк, кодирующий альфа-фетопротеин (афп) человека, содержащий "медленные" кодоны, кодирующие лейцин, экспрессионная плазмида, содержащая указанный фрагмент, клетка saccharomyces cerevisiae, трансформированная указанной плазмидой, и способ получения рекомбинантного афп человека
Участок связывания антигена (fab), в том числе гуманизированный fab, против ботулинического нейротоксина с (варианты), способ получения fab с использованием дрожжей, способ и набор для детекции ботулинического нейротоксина с
Липосома, фармацевтическая композиция и лекарственное средство для лечения местных радиационных поражений кожи, применение липосом и способ для лечения местных радиационных поражений кожи
Вариабельные домены легкой и тяжелой цепи мышиного моноклонального антитела против фактора некроза опухоли альфа (фно-α) человека (варианты), антигенсвязывающий фрагмент (fab) против фно-α человека, содержащий указанные домены (варианты)
Способ удаления пеногасителя из культуральной жидкости после ферментации
Рекомбинантный фрагмент днк, кодирующий альфа-фетопротеин (афп) человека, содержащий "медленные" кодоны, кодирующие лейцин, экспрессионная плазмида, содержащая указанный фрагмент, клетка saccharomyces cerevisiae, трансформированная указанной плазмидой, и способ получения рекомбинантного афп человека
Участок связывания антигена (fab), в том числе гуманизированный fab, против ботулинического нейротоксина с (варианты), способ получения fab с использованием дрожжей, способ и набор для детекции ботулинического нейротоксина с
Липосома, фармацевтическая композиция и лекарственное средство для лечения местных радиационных поражений кожи, применение липосом и способ для лечения местных радиационных поражений кожи
Рекомбинантный штамм мицелиального гриба aspergillus nidulans и его применение для 11α-гидроксилирования прогестерона и для ацетилирования 11α-гидроксипрогестерона
Гуманизированные антигенсвязывающие фрагменты (fab) против вируса бешенства, изолированный фрагмент днк, кодирующий fab против вируса бешенства, клетка дрожжей, трансформированная фрагментом днк, и способ получения fab против вируса бешенства с использованием дрожжей
Гуманизированные антитела и fab, связывающиеся с антигеном f1 из yersinia pestis, и способ их получения с использованием дрожжей
Мышиные моноклональные антитела, связывающиеся с антигеном f1 из yersinia pestis, способ их получения с использованием дрожжей, способ и набор для детекции yersinia pestis
Рекомбинантная лакказа лигнинолитического гриба trametes sp. и способ ее получения

