Результат интеллектуальной деятельности: АНТАГОНИСТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА IL-7 И СПОСОБЫ
Вид РИД
Изобретение
Область изобретения
Настоящее изобретение относится к антителам, например к полноразмерным антителам или их антигенсвязывающим участкам, которые антагонизируют активность рецептора интерлейкина-7 (IL-7R), включая его взаимодействие с IL-7. Изобретение также относится к композициям, содержащим антагонист IL-7R, такой как антагонистическое антитело против IL-7R, и способам применения антагониста IL-7R в качестве лекарственного средства. Антагонист IL-7R может быть использован для предупреждения и/или лечения диабета 2 типа, реакции "трансплантат против хозяина" (GVHD) и аутоиммунных расстройств, включая диабет 1 типа, рассеянный склероз, ревматоидный артрит и волчанка.
Предшествующий уровень техники
Комплекс IL-7R представляет собой гетерод и мерный рецептор, состоящий из альфа-цепи IL-7R (IL-7Rα) и общей гамма-цепи (γс) (Mazzucchelli et al., Nat Rev Immunol., 2007, 7:144-54). IL-7R связывается интерлейкином-7 (IL-7), который представляет собой цитокин, необходимый для развития и гомеостатического поддержания Т- и В-лимфоцитов (Fry et al., J Immunol., 2005, 174:6571-6). Связывание IL-7 с IL-7R активирует многочисленные пути, которые регулируют выживание, потребление глюкозы, пролиферацию и дифференцировку лимфоцитов.
IL-7R экспрессируется как на дендритных клетках, так и на моноцитах и, по-видимому, действует во множественных гемопоэтических линиях дифференцировки (Reche PA, et al., J Immunol., 2001, 167:336-43). В дендритных клетках IL-7R играет иммуномодулирующую роль, тогда как лимфоциты нуждаются в IL-7R - сигнальном пути для выживания, пролиферации и дифференцировки. Оба Jak-Stat и PI3K-Akt пути активируются связыванием IL-7 с IL-7R (Jian et al., Cytokine Growth Factor Rev., 2005, 16:513-533). Эти пути вовлекают взаимодействие (crosstalk) путей передачи сигналов, общие домены взаимодействия, петли обратной связи, интегрированную генную регуляцию, мультимеризацию и конкуренцию лигандов. Некоторые мишени IL-7-сигнального пути, включая Вс12 и Pyk2, вносят вклад в клеточное выживание. Другие мишени, такие как PI3 киназа, киназы семейства src (Ick и fyn) и STAT5, вносят вклад в клеточную пролиферацию. Транскрипционный фактор STAT5 вносит вклад в активацию множества различных генов, регулирующих последующие звенья сигнальных каскадов в В- и Т-клетках, и может вносить вклад в VDJ-рекомбинацию посредством изменения структуры хроматина. Сигналы клеточного выживания и клеточной пролиферации, индуцируемые IL-7, объединяются для индукции развития нормальных Т-клеток. Детали сигнальной сети комплекса IL-7 и ее взаимодействия с другими сигнальными каскадами в клетках иммунной системы все еще полностью не выяснены.
Из информации, имеющейся в данной области техники и предшествующей настоящему изобретению, остается неясно, является ли введение антагонистического антитела к IL-7R в кровоток для селективного блокирования IL-7R эффективным для лечения диабета 2 типа, диабета 1 типа, GVHD, волчанки и ревматоидного артрита, и, если это так, то какие свойства антитела против IL-7R необходимы для такой эффективности in vivo.
Краткое изложение сущности изобретения
Предложены антагонистические антитела, которые селективно взаимодействуют и ингибируют функцию IL-7R. Впервые показано, что некоторые антагонистические антитела к IL-7R эффективны in vivo для лечения диабета 1 типа, диабета 2 типа, ревматоидного артрита, GVHD и волчанки.
В некоторых воплощениях предложены антагонистические антитела, которые селективно взаимодействуют и ингибируют функцию IL-7R. В некоторых воплощениях антитело специфически связывается с IL-7R и содержит антигенсвязывающий участок, который конкурирует с моноклональным антителом, выбранным из группы, состоящей из C1GM, С2М3, Р3А9, Р4В3, P2D2, Р2Е11, HAL403a и HAL403b, для связывания с IL-7R. В некоторых воплощениях антитело содержит полипептид, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 42 или SEQ ID NO: 43. В других воплощениях антитело специфически связывается с IL-7R и распознает эпитоп, который перекрывает эпитоп IL-7R, который распознается моноклональным антителом, выбранным из группы, состоящей из C1GM, С2М3, Р3А9, Р4 В3, P2D2, Р2Е11, HAL403a и HAL403b. В некоторых воплощениях антитело связывается с эпитопом, содержащим остатки 182, K84, K100, Т105, и Y192 альфа-цепи рецептора интерлейкина-7 (IL-7Rα). В некоторых воплощениях эпитоп дополнительно содержит один или более дополнительных остатков, выбранных из группы, состоящей из остатков D190, Н191 и K194 человеческого IL-7Rα.
В некоторых воплощениях IL-7R представляет собой человеческий IL-7R.
В некоторых воплощениях антитело специфически связывается с альфа-цепью рецептора интерлейкина-7 (IL-7Rα) и содержит определяющий комплементарность участок один (CDR1) вариабельной области тяжелой цепи (VH), имеющий аминокислотную последовательность X1X2VMH, где X1 представляет собой D или N; Х2 представляет собой S или Y (SEQ ID NO: 50), CDR2 VH, имеющий аминокислотную последовательность X1X2X3X4X5GX6X7TYYADSVKG, где X1 представляет собой L или А; Х2 представляет собой V или I; Х3 представляет собой G или S; Х4 представляет собой W или G; Х5 представляет собой D или S; Х6 представляет собой F, G или S; Х7 представляет собой F, А или S (SEQ ID NO: 51), и CDR3 VH, имеющий аминокислотную последовательность Х1Х2Х3Х4Х5Х6Х7Х8, где X1 представляет собой Q или D; Х2 представляет собой G или I; Х3 представляет собой D или S; Х4 представляет собой Y или G; Х5 представляет собой М, V или G; Х6 представляет собой G или F; Х7 представляет собой N, D или М; Х8 представляет собой N, Y или D (SEQ ID NO: 52), CDR1 вариабельной области легкой цепи (VL), имеющий аминокислотную последовательность TX1SSGX2IX3SSYVQ, где X1 представляет собой R или G; Х2 представляет собой S или R; Х3 представляет собой D или A (SEQ ID NO: 53), CDR2 VL, имеющий аминокислотную последовательность EDX1QRPS, где X1 представляет собой D или N (SEQ ID NO: 54), и CDR3 VL, имеющий аминокислотную последовательность X1X2YX3X4X5X6LX7, где X1 представляет собой Q или М; Х2 представляет собой S или Q; Х3 представляет собой D или А; Х4 представляет собой F или S; Х5 представляет собой Н или S; Х6 представляет собой Н или S; Х7 представляет собой V или W (SEQ ID NO: 55), где указанное антитело блокирует фосфорилирование STAT5 в анализе активации STAT5. В некоторых воплощениях каркасный участок между CDR2 VH и CDR3 VH содержит аминокислотную последовательность аланин-аргинин, где аргинин примыкает к первому аминокислотному остатку CDR3 VH. В некоторых воплощениях каркасный участок между CDR2 VH и CDR3 VH содержит аминокислотную последовательность цистеин-аланин-аргинин, где аргинин примыкает к первому аминокислотному остатку CDR3 VH.
В некоторых воплощениях антитело содержит CDR-контактирующий участок один тяжелой цепи, имеющий аминокислотную последовательность X1X2VMH, где представляет собой D или N; Х2 представляет собой S или Y (SEQ ID NO: 50), CDR-контактирующий участок два тяжелой цепи, имеющий аминокислотную последовательность GWDGFF (SEQ ID NO: 57), и CDR-контактирующий участок три тяжелой цепи, имеющий аминокислотную последовательность ARX1X2X3X4 (SEQ ID NO: 58), CDR-контактирующий участок один легкой цепи, имеющий аминокислотную последовательность SGSIDSSY (SEQ ID NO: 59), CDR-контактирующий участок два легкой цепи, имеющий аминокислотную последовательность EDDQRPSGV (SEQ ID NO: 60), и CDR-контактирующий участок три легкой цепи, имеющий аминокислотную последовательность FHHL (SEQ ID NO: 61), где указанное антитело блокирует фосфорилирование STAT5 в анализе активации STAT5.
В некоторых воплощениях антитело специфически связывается с IL-7Rα и содержит определяющий комплементарность участок один (CDR1) вариабельной области тяжелой цепи (VH), имеющий аминокислотную последовательность DSVMH (SEQ ID NO: 19), GFTFDDS (SEQ ID NO: 46) или GFTFDDSVMH (SEQ ID NO: 47), CDR2 VH, имеющий аминокислотную последовательность LVGWDGFFTYYADSVKG (SEQ ID NO: 23) или GWDGFF (SEQ ID NO: 48), и CDR3 VH, имеющий аминокислотную последовательность QGDYMGNN (SEQ ID NO: 49), или его вариант, имеющий одну или более консервативных аминокислотных замен в CDR1, CDR2 и/или CDR3.
В некоторых воплощениях антитело содержит CDR1 вариабельной области легкой цепи (VL), имеющий аминокислотную последовательность TRSSGSIDSSYVQ (SEQ ID NO: 29), CDR2 VL, имеющий аминокислотную последовательность EDDQRPS (SEQ ID NO: 31), и/или CDR3 VL, имеющий аминокислотную последовательность QSYDFHHLV (SEQ ID NO: 36), или его вариант, имеющий одну или более аминокислотных замен в CDR1, CDR2, и/или CDR3. В некоторых воплощениях антитело дополнительно содержит CDR1 VH, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 19, 46 или 47, CDR2 VH, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 23 или 48, и CDR3 VH, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 49, или его вариант, имеющий одну или более аминокислотных замен в CDR1, CDR2 и/или CDR3.
В некоторых воплощениях антитело специфически связывается с IL-7Rα и содержит определяющий комплементарность участок один (CDR1) вариабельной области тяжелой цепи (VH), имеющий аминокислотную последовательность, представленную в SEQ ID NO: 19, 46 или 47, CDR2 VH, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 23 или 48, и CDR3 VH, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 49, CDR1 вариабельной области легкой цепи (VL), имеющий аминокислотную последовательность, представленную в SEQ ID NO: 29, CDR2 VL, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 31, и CDR3 VL, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 36. В некоторых воплощениях VH-область содержит аминокислотную последовательность EVQLVESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWVSLVGWD GFFTYYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARQGDYMGNNWG QGTLVTVSS (SEQ ID NO: 40), и VL-область содержит аминокислотную последовательность
NFMLTQPHSVSESPGKTVTISCTRSSGSIDSSYVQWYQQRPGSSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDFHHLVFGGGTKLTVL (SEQ ID NO: 41). В некоторых воплощениях антитело содержит легкую цепь, имеющую аминокислотную последовательность
NFMLTQPHSVSESPGKTVTISCTRSSGSIDSSYVQWYQQRPGSSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDFHHLVFGGGTKLTVLQPK AAPSVTLFPPSSEELQANKATLVCLISDFYPGAVWAWKADSSPVKAGVETTTPSKQ SNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 43), и тяжелую цепь, имеющую аминокислотную последовательность EVQLVESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEVWSLVGWD GFFTYYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARQGDYMGNNWG QGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALT SGVHTFPAVLQSSGLYSLSSWTVPSSSLGTQTYICNVNHKPSNTKVDKKVAPELLG GPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEVKFNWYVDGVEVHNAKTKPR EEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQV YTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSF FLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 42), с С-концевым лизином или без С-концевого лизина в SEQ ID NO: 42.
В некоторых воплощениях антитело может представлять собой человеческое антитело, гуманизированное антитело или химерное антитело. В некоторых воплощениях антитело представляет собой моноклональное антитело.
В некоторых воплощениях антитело содержит константную область. В некоторых воплощениях антитело состоит из человеческого IgG1 или IgGΔα подкласса. В некоторых воплощениях антитело содержит гликозилированную константную область. В некоторых воплощениях антитело содержит константную область, имеющую увеличенную аффинность связывания с человеческим Fc-гамма рецептором.
В некоторых воплощениях предложена фармацевтическая композиция, содержащая антитело, которое селективно взаимодействует и ингибирует функцию IL-7R.
В некоторых воплощениях предложена клеточная линия, которая рекомбинантно продуцирует антитело, которое селективно взаимодействует и ингибирует функцию IL-7R.
В некоторых воплощениях предложена нуклеиновая кислота, кодирующая антитело, которое селективно взаимодействует и ингибирует функцию IL-7R.
В некоторых воплощениях предложены способы снижения уровней глюкозы в крови у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагонистического антитела к IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого снижение уровней глюкозы в крови.
В некоторых воплощениях предложены способы повышения толерантности к глюкозе у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагонистического антитела к IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого повышение толерантности к глюкозе.
В некоторых воплощениях предложены способы предупреждения или лечения диабета 1 типа у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагонистического антитела к IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого предупреждение или лечение одного или более симптомов диабета 1 типа.
В некоторых воплощениях предложены способы предупреждения или лечения диабета 2 типа у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагониста IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого предупреждение или лечение одного или более симптомов диабета 2 типа. В некоторых воплощениях антагонист IL-7R представляет собой антагонистическое антитело к IL-7R.
В некоторых воплощениях предложены способы предупреждения или лечения ревматоидного артрита у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагонистического антитела к IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого предупреждение или лечение одного или более симптомов ревматоидного артрита.
В некоторых воплощениях предложены способы предупреждения или лечения реакции "трансплантат против хозяина" (GVHD) у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагонистического антитела к IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого предупреждение или лечение одного или более симптомов GVHD.
В некоторых воплощениях GVHD представляет собой хроническую GVHD или острую GVHD.
В некоторых воплощениях предложены способы предупреждения или лечения волчанки у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагонистического антитела к IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого предупреждение или лечение одного или более симптомов волчанки.
В некоторых воплощениях волчанка представляет собой кожную красную волчанку, системную красную волчанку, медикаментозную красную волчанку или неонатальную волчанку.
В некоторых воплощениях предложены способы предупреждения или лечения рассеянного склероза у индивидуума. В некоторых воплощениях способ включает введение терапевтически эффективного количества антагонистического антитела к IL-7R индивидууму, нуждающемуся в таком лечении, посредством этого предупреждение или лечение одного или более симптомов рассеянного склероза и сокращение и/или истощение "наивных" и/или активированных Т-клеточных популяций у индивидуума. В некоторых воплощениях сокращенные или истощенные Т-клеточные популяции у индивидуума содержат Тн1- и/или Тн17-клетки. В некоторых воплощениях введение антагонистического антитела к IL-7R не приводит к расширению популяции Тн17-клеток у индивидуума.
В некоторых воплощениях антитело можно вводить парентерально. В некоторых воплощениях индивидуум представляет собой человека.
Краткое описание графических материалов
На Фиг.1 показан дозозависимый эффект антагонистических моноклональных антител к IL-7R - P2D2, Р2Е11 и HAL403a - на IL-7-опосредованное фосфорилирование STAT5 в человеческих мононуклеарных клетках периферической крови (PBMCs). На оси x указан процент CD4+ клеток, экспрессирующих фосфо-STAT5 (p-STAT).
На Фиг.2 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на развитие диабета у нетучных диабетических (NOD) мышей.
На Фиг.3 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на (А) уровни глюкозы в крови (мг/дл) и (Б) массу тела (г) у NOD мышей.
На Фиг.4 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на популяции (А) "наивных" CD8+Т - клеток и (Б) CD8+Т-клеток памяти у NOD мышей. На оси x общая популяция CD8+Т - клеток принята за 100%.
На Фиг.5 показан эффект антагонистического моноклонального антитела против к IL-7R - 28G9 - на популяцию "наивных" CD4+Т - клеток у NOD мышей. На оси х общая популяция CD4+Т-клеток принята за 100%.
На Фиг.6 показан эффект антагонистических моноклональных антител к IL-7R - 28 В6 и 28G9 - на клиническую тяжесть ЕАЕ (экспериментальный аутоиммунный энцефаломиелит) у животных. Клиническую тяжесть ЕАЕ оценивали ежесуточно с помощью системы балльной оценки от 0 до 5:0 -нормальный; 1 - вялый хвост; 2 - частичный паралич задних конечностей; 3 - полный паралич задних конечностей; 4 - паралич четырех конечностей; 5 - состояние угасания или смерть.
На Фиг.7 показан дозозависимый эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на клиническую тяжесть ЕАЕ у животных.
На Фиг.8 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на клиническую тяжесть ЕАЕ у животных.
На Фиг.9 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на животных с установленным ЕАЕ.
На Фиг.10 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - в низкой дозе на животных с установленным ЕАЕ.
На Фиг.11 показан эффект антагонистических моноклональных антител к IL-7R - 28G9 и 28В6 - на популяции (A) CD4 Т-клеток и (Б) CD8 Т-клеток из костного мозга (ВМ), селезенки, крови и лимфатических узлов (LNs) у ЕАЕ животных. На оси х общая популяция лимфоцитов принята за 100%.
На Фиг.12А-В показан эффект антагонистического моноклонального антитела к IL-7 - 28G9 - на популяции (А) "наивных" Т-клеток, (Б) Т-клеток памяти и (В) активированных Т-клеток из костного мозга, селезенки и лимфатических узлов у ЕАЕ животных. На оси х популяция CD8+Т - клеток принята за 100%.
На Фиг.13 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на популяции Тек клеток (левый график) и Treg клеток (правый график) из костного мозга, селезенки и лимфатических узлов у ЕАЕ животных. На оси х популяция CD4+Т - клеток принята за 100%. "*" означает Р<0,05 по сравнению с контролем.
На Фиг.14 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на уровни глюкозы в крови (мг/дл) у мышей с ожирением, индуцированным диетой с высоким содержанием жиров (DIO).
На Фиг.15 показан эффект антагонистического моноклонального антитела к IL-7R - 28G9 - на толерантность к глюкозе у мышей с ожирением, индуцированным диетой с высоким содержанием жиров (DIO).
Подробное описание изобретения
В данной заявке раскрыты антитела, которые демонстрируют антагонистическую активность в отношении функции IL-7R, включая его взаимодействие с IL-7. Предложены способы получения антагонистических антител к IL-7R, композиций, содержащих эти антитела, и способы применения этих антител в качестве лекарственного средства. Антагонисты IL-7R, например антагонистические антитела к IL-7R, могут быть использованы для предупреждения и/или лечения диабета 2 типа, GVHD и аутоиммунных расстройств, включая рассеянный склероз (MS), ревматоидный артрит, диабет 1 типа и волчанку.
Общие методы
При практическом осуществлении настоящего изобретения будут использоваться, если не указано иное, обычные методы молекулярной биологии (включая рекомбинантные методы), микробиологии, клеточной биологии, биохимии и иммунологии, которые находятся в пределах компетенции специалистов в данной области техники. Такие методы описаны в полной мере в литературе, например Molecular Cloning: A Laboratory Manual, second edition (Sambrook et al., 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J.Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E.Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I.Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P.Mather and P.E.Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A.Doyle, J.B.Griffiths, and D.G.Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M.Weir and C.C.Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M.Miller and M.P.Calos, eds., 1987); Current Protocols in Molecular Biology (F.M.Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E.Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A.Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D.Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C.Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E.Harlow and D.Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M.Zanetti and J.D.Capra, eds., Harwood Academic Publishers, 1995).
Определения
"Антитело" представляет собой молекулу иммуноглобулина, способную специфически связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и т.д., посредством по меньшей мере одного сайта распознавания антигена, расположенного в вариабельной области молекулы иммуноглобулина. Используемый в данной заявке термин охватывает не только интактные поликлональные или моноклональные антитела, но и их фрагменты (такие как Fab, Fab', F(ab')2 , Fv), одноцепочечные (ScFv) и доменные антитела (включая, например, антитела акул и верблюдовых) и слитые белки, содержащие антитело, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена. Антитело включает антитело любого класса, такое как IgG, IgA или IgM (или его подкласс), и антитело не обязательно должно быть какого-либо конкретного класса. В зависимости от аминокислотной последовательности константной области тяжелых цепей антитела иммуноглобулины можно отнести к различным классам. Имеется пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них также могут быть разделены на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные области тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются альфа, дельта, эпсилон, гамма и мю соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
Как использовано в данной заявке, "моноклональное антитело" относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальных антител, содержащих идентичную популяцию, за исключением возможных природных мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, направленными против одного антигенного сайта. Кроме того, в отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Определение "моноклональное" указывает на характер антитела как полученного по существу из гомогенной популяции антител и не должно толковаться как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела, используемые в соответствии с настоящим изобретением, могут быть получены гибридомным методом, впервые описанным Kohler и Milstein, 1975, Nature 256:495, или могут быть получены методами рекомбинантных ДНК, как описано в патенте США №4816567. Например, моноклональные антитела также могут быть выделены из фаговых библиотек, полученных с использованием методов, описанных в McCafferty et al., 1990, Nature 348:552-554.
Как использовано в данной заявке, "гуманизированное" антитело относится к формам нечеловеческих (например, мышиных) антител, которые представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие последовательности антител), которые содержат минимальную последовательность, происходящую из нечеловеческого иммуноглобулина. Предпочтительно гуманизированные антитела представляют собой человеческие иммуноглобулины (антитело реципиента), в которых остатки из определяющего комплементарность участка (CDR) реципиента заменены остатками из CDR вида, не являющегося человеком (антитело донора), такого как мышь, крыса или кролик, имеющего желаемую специфичность, аффинность и эффективность. В некоторых случаях остатки Fv каркасного участка (FR) человеческого иммуноглобулина заменены нечеловеческими остатками. Кроме того, гуманизированное антитело может содержать остатки, которые не обнаружены ни в антителе реципиента, ни в импортированном CDR или каркасных последовательностях, но включены для дальнейшего совершенствования и оптимизации свойств антитела. В целом, гуманизированное антитело будет содержать по существу все из по меньшей мере одного и обычно двух вариабельных доменов, в которых все или по существу все CDR участки соответствуют таковым в нечеловеческом иммуноглобулине и все или по существу все FR участки представляют собой участки из человеческой консенсусной последовательности иммуноглобулина. Гуманизированное антитело оптимально также содержит по меньшей мере часть константной области или домена (Fc) иммуноглобулина, как правило, человеческого иммуноглобулина. Предпочтительными являются антитела, имеющие Fc-области, модифицированные, как описано в WO 99/58572. Другие формы гуманизированных антител имеют один или более CDRs (CDR L1, CDR L2, CDR L3, CDR Н1, CDR H2 или CDR Н3), которые изменены относительно первоначального антитела, которые также называются как один или более CDRs, "происходящих из" одного или более CDRs из первоначального антитела.
Как использовано в данной заявке, "человеческое антитело" означает антитело, имеющее аминокислотную последовательность, соответствующую аминокислотной последовательности антитела, продуцируемого человеком, и/или которое было получено с использованием любого из методов получения человеческих антител, известных специалистам в данной области или раскрытых в данной заявке. Это определение человеческого антитела включает антитела, содержащие по меньшей мере один полипептид человеческой тяжелой цепи или по меньшей мере один полипептид человеческой легкой цепи. Один такой пример представляет собой антитело, содержащее полипептиды мышиной легкой цепи и человеческой тяжелой цепи. Человеческие антитела можно получать с использованием различных методов, известных в данной области техники. В одном воплощении человеческое антитело селектируют из фаговой библиотеки, где эта фаговая библиотека экспрессирует человеческие антитела (Vaughan et al., 1996, Nature Biotechnology, 14:309-314; Sheets et al., 1998, Proc. Natl. Acad. Sci. (USA) 95:6157-6162; Hoogenboom and Winter, 1991, J. Mol. Biol., 227:381; Marks et al., 1991, J. Mol. Biol., 222:581). Человеческие антитела также могут быть получены посредством иммунизации животных, которым локусы человеческих иммуноглобулинов были трансгенно введены вместо эндогенных локусов, например мышей, у которых эндогенные гены иммуноглобулинов были частично или полностью инактивированы. Такой подход описан в патентах США №№5545807; 5545806; 5569825; 5625126; 5633425; и 5661016. Альтернативно, человеческое антитело может быть получено посредством иммортализации человеческих В-лимфоцитов, которые продуцируют антитело, направленное против целевого антигена (такие В-лимфоциты можно выделить из индивидуума или, возможно, были иммунизированы in vitro). См., например, Cole et al. Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77, 1985; Boerner et al., 1991, J. Immunol., 147 (1):86-95; и патент США №5750373.
Используемый в данной заявке термин "IL-7R" относится к любой форме IL-7R и его вариантам, которые сохраняют по меньшей мере часть активности IL-7R. Если не указано иное, например, посредством конкретной ссылки на человеческий IL-7R, IL-7R включает нативные последовательности IL-7R всех видов млекопитающих, например человека, собак, кошек, лошадей и крупного рогатого скота.
Используемый в данной заявке "антагонист IL-7R" относится к антителу или молекуле, которая способна ингибировать биологическую активность IL-7R и/или нижележащий(е) путь(и), опосредованный(е) IL-7R сигнальным путем, включая связывание с IL-7, фосфорилирование STAT5, Src киназы, PI3 киназу и Рук2 и активацию белка Вс12. Примеры антагонистов IL-7R включают, без ограничения, антагонистические антитела к IL-7R, siRNA (малые интерферирующие РНК) IL-7R, shRNA (короткие РНК, образующие шпильки) IL-7R и антисмысловые олигонуклеотиды IL-7R.
Антагонистические антитела к IL-7R охватывают антитела, которые блокируют, антагонизируют, подавляют или уменьшают (в любой степени, в том числе значительно) биологическую активность IL-7R, включая нижележащие пути, опосредованные IL-7R-сигнальным путем, такие как взаимодействие с IL-7 и/или индукция клеточного ответа на IL-7. Для целей настоящего изобретения будет очевидно, что термин "антагонистическое антитело к IL-7R" (взаимозаменяемо называемое "IL-7R антагонистическим антителом", "антагонистическим антителом против IL-7R" или "анти-IL-7R антагонистическим антителом") охватывает все ранее идентифицированные термины, названия и функциональные состояния и характеристики, в соответствии с которыми IL-7R сам, биологическая активность IL-7R (включая, но не ограничиваясь этим, взаимодействие с IL-7, его способность опосредовать любой аспект фосфорилирования STAT5, активации пути фосфатидилинозитол-3-киназы (PI3K)-Akt, понижающей регуляции p27Kip1, повышающей регуляции Bcl-2, гиперфосфорилирования Rb и повышающей регуляции CXCR4) или последствия биологической активности по существу отменяются, уменьшаются или нейтрализуются в любой значимой степени. В некоторых воплощениях антагонистическое антитело к IL-7R связывает IL-7R и предупреждает взаимодействие с IL-7. Примеры антагонистических антител к IL-7R представлены в данной заявке.
Как использовано в данной заявке, "полный антагонист" представляет собой антагонист, который при эффективной концентрации практически полностью блокирует измеряемый эффект IL-7R. Под "частичным антагонистом" подразумевают антагонист, который способен частично блокировать измеряемый эффект, но который даже при самой высокой концентрации не является полным антагонистом. "Практически полностью" означает по меньшей мере примерно 80%, предпочтительно по меньшей мере примерно 90%, более предпочтительно по меньшей мере примерно 95% и наиболее предпочтительно по меньшей мере примерно 98% измеряемого эффекта блокируется.
Термины "полипептид", "олигопептид", "пептид" и "белок" используются взаимозаменяемо в данной заявке в отношении цепей аминокислот любой длины, предпочтительно относительно коротких (например, 10-100 аминокислот). Цепь может быть линейной или разветвленной, она может содержать модифицированные аминокислоты и/или может прерываться неаминокислотами. Эти термины также охватывают аминокислотную цепь, которая была модифицирована естественно или посредством вмешательства; например, образования дисульфидных связей, гликозилирования, липидизации, ацетилирования, фосфорилирования или любой другой манипуляции или модификации, такой как сопряжение с меченым компонентом. В пределах определения также находятся, например, полипептиды, содержащие один или более аналогов аминокислоты (включая, например, неприродные аминокислоты и т.д.), а также другие модификации, известные в данной области техники. Понятно, что полипептиды могут встречаться в виде одиночных цепей или ассоциированных цепей.
Как известно в данной области техники, "полинуклеотид" или "нуклеиновая кислота", используемые взаимозаменяемо в данной заявке, относятся к цепям нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания и/или их аналоги, или любой субстрат, который может быть включен в цепь с помощью ДНК- или РНК-полимеразы. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. При наличии модификация в нуклеотидной структуре может быть включена до или после сборки цепи. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. Полинуклеотид может быть дополнительно модифицирован после полимеризации, например путем сопряжения с меченым компонентом. Другие типы модификаций включают, например, "кэпы", замену одного или более встречающихся в природе нуклеотидов аналогом, межнуклеотидными модификациями, такими как, например, модификации с незаряженными связями (например, метилфосфонаты, фосфотриэфиры, фосфоамидаты, карбаматы и т.д.) и с заряженными связями (например, фосфоротиоаты, фосфородитиоаты и т.д.), модификации, содержащие боковые группировки, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.д.), модификации с интеркаляторами (например, акридином, псораленом и т.д.), модификации, содержащие хелатирующие агенты (например, металлы, радиоактивные металлы, бор, окислительные металлы и т.д.), модификации, содержащие алкилирующие агенты, модификации с модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и т.д.), а также немодифицированные формы полинуклеотида(ов). Кроме того, любая из гидроксильных групп, обычно присутствующих в сахарах, может быть замещена, например, фосфонатными группами, фосфатными группами, защищена стандартными защитными группами или активирована для получения дополнительных связей с дополнительными нуклеотидами или может быть конъюгирована с твердыми носителями. 5'- и 3'-концевой ОН может быть фосфорилирован или замещен аминами или органическими кэп-группировками, содержащими от 1 до 20 атомов углерода. Другие гидроксилы также могут быть дериватизированы до стандартных защитных групп. Полинуклеотиды также могут содержать аналогичные формы сахаров рибозных или дезоксирибозных сахаров, которые, как правило, известны в данной области техники, включая, например, 2'-O-метил-, 2'-O-аллил, 2'-фтор- или 2'-азидорибозу, карбоциклические аналоги сахара, альфа- или бета-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и аналоги нуклеозидов без азотистых оснований, такие как метилрибозид. Одна или более фосфодиэфирных связей могут быть замещены альтернативными связывающими группами. Эти альтернативные связывающие группы включают, но не ограничиваются этим, воплощения, где фосфат замещен P(О)S ("тиоат"), P(S)S ("дитиоат"), (O)NR2 ("амидат"), P(O)R, P(O)OR', CO или CH2 ("формацеталь"), в котором каждый R или R' независимо представляет собой Н или замещенный или незамещенный алкил (1-20 С), возможно содержащий простую эфирную (-O-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралкил. Не все связи в полинуклеотиде должны быть идентичными. Предшествующее описание относится ко всем полинуклеотидам, упомянутым в данной заявке, включая РНК и ДНК.
"Вариабельная область" антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела, либо одной, либо в комбинации. Как известно в данной области техники, каждая из вариабельных областей тяжелой и легкой цепи состоит из четырех каркасных участков (FR), соединенных тремя определяющими комплементарность участками (CDRs), также известными как гипервариабельные участки. CDRs в каждой цепи находятся вместе в непосредственной близости к FRs и с CDRs из другой цепи, вносят вклад в образование антигенсвязывающего сайта антител. Имеется по меньшей мере два метода определения CDRs: (1) подход, основанный на межвидовой изменчивости последовательностей (т.е. Kabat et al. Sequences of Proteins of Immunological Interest, (5th ed., 1991, National Institutes of Health, Bethesda MD)); и (2) подход, основанный на кристаллографических исследованиях комплексов антиген-антитело (Al-lazikani et al., 1997, J. Molec. Biol. 273:927-948). Как использовано в данной заявке, CDR может относиться к CDRs, определяемым любым из подходов или комбинацией обоих подходов.
Как известно в данной области техники, "константная область" антитела относится к константной области легкой цепи антитела или константной области тяжелой цепи антитела, по отдельности или в комбинации.
Используемое в данной заявке антитело "взаимодействует с" IL-7R, когда равновесная константа диссоциации равна или меньше 20 нм, предпочтительно меньше чем примерно 6 нм, более предпочтительно меньше чем примерно 1 нм, наиболее предпочтительно меньше чем примерно 0,2 нм, как измерено методами, описанными в данной заявке в Примере 2.
Эпитоп, который "предпочтительно связывается" или "специфически связывается" (используются взаимозаменяемо в данной заявке) с антителом или полипептидом, представляет собой термин, хорошо известный в данной области техники, и методы определения такого специфического или предпочтительного связывания также хорошо известны в данной области техники. Считается, что молекула демонстрирует "специфическое связывание" или "предпочтительное связывание", если она взаимодействует или ассоциирует чаще, быстрее, с большей длительностью и/или с большей аффинностью с определенными клетками или веществом по сравнению с альтернативными клетками или веществом. Антитело "специфически связывается" или "предпочтительно связывается" с мишенью, если оно связывается с большей аффинностью, авидностью, более легко и/или с большей длительностью по сравнению с тем, как оно связывается с другими веществами. Например, антитело, которое специфически или предпочтительно связывается с эпитопом IL-7R, представляет собой антитело, которое связывается с этим эпитопом с большей аффинностью, авидностью, более легко и/или с большей длительностью по сравнению с тем, как оно связывается с другими эпитопами IL-7R или эпитопами не-IL-7R. Также понятно при прочтении этого определения, например, что антитело (группировка или эпитоп), которое специфически или предпочтительно связывается с первой мишенью, может специфически или неспецифически или предпочтительно связываться со второй мишенью. Таким образом, "специфическое связывание" или "предпочтительное связывание" не обязательно требует (хотя может включать) исключительное связывание. Как правило, но не обязательно, ссылка на связывание означает предпочтительное связывание.
Как использовано в данной заявке, "по существу чистый" относится к материалу, который является по меньшей мере на 50% чистым (т.е. свободным от загрязнений), более предпочтительно по меньшей мере на 90% чистым, более предпочтительно по меньшей мере на 95% чистым, еще более предпочтительно по меньшей мере на 98% чистым и наиболее предпочтительно по меньшей мере на 99% чистым.
"Клетка-хозяин" включает отдельную клетку или клеточную культуру, которая может быть или была реципиентом для вектора(ов) для включения полинуклеотидных вставок. Клетки-хозяева включают потомство одной клетки-хозяина и данное потомство не обязательно должно быть полностью идентичным (по морфологии или дополнениям геномной ДНК) исходной родительской клетке из-за естественной, случайной или намеренно полученной мутации. Клетка-хозяин включает клетки, трансфицированные in vivo полинуклеотидом(ами) по данному изобретению.
Как известно в данной области техники, термин "Fc-область" используют для определения С-концевой области тяжелой цепи иммуноглобулина. "Fc-область" может иметь нативную последовательность Fc-области или вариант Fc-области. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьироваться, Fc-область тяжелой цепи IgG человека обычно определяют как отрезок от аминокислотного остатка в положении Cys226, или от Pro230, до его карбоксильного конца. Нумерацию остатков в Fc-области соответствует EU индексу по Кабату. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md., 1991. Fc-область иммуноглобулина, как правило, содержит две константные области СН2 и СНЗ.
Как использовано в данной области техники, "Fc-рецептор" и "FcR" описывают рецептор, который связывается с Fc-областью антитела. Предпочтительный FcR имеет нативную последовательность человеческого FcR. Кроме того, предпочтительный FcR представляет собой FcR, который связывается с IgG антителом (гамма-рецептором) и включает рецепторы подклассов FcyRI, FcyRII и FcyRIII, включая аллельные варианты и альтернативно сплайсированные формы этих рецепторов. FcyRII рецепторы включают FcyRIIA ("активирующий рецептор") и FcyRIIB ("ингибирующий рецептор"), которые имеют сходные аминокислотные последовательности, которые различаются в первую очередь их цитоплазматическими доменами. FcRs описаны в Ravetch and Kinet, 1991, Ann. Rev. Immunol., 9:457-92; Capel et al., 1994, Immunomethods, 4:25-34; и de Haas et al., 1995, J. Lab. Clin. Med., 126:330-41. "FcR" также включает неонатальный рецептор FcRn, который отвечает за перенос материнских IgG плоду (Guyer et al., 1976, J. Immunol., 117:587; и Kim et al., 1994, J. Immunol., 24:249).
Термин "конкурировать", используемый в данной заявке в отношении антитела, означает, что первое антитело или его антигенсвязывающий фрагмент связывается с эпитопом способом, достаточно близким к связыванию второго антитела или его антигенсвязывающего фрагмента, так что результат связывания первого антитела с его штатным эпитопом заметно уменьшается в присутствии второго антитела, по сравнению со связыванием первого антитела в отсутствие второго антитела. Может, но не обязательно, иметь место альтернатива, где связывание второго антитела с его эпитопом также заметно уменьшается в присутствии первого антитела. То есть первое антитело может ингибировать связывание второго антитела с его эпитопом без ингибирования вторым антителом связывания первого антитела с его соответствующим эпитопом. Однако если каждое антитело заметно ингибирует связывание другого антитела с его когнатным эпитопом или лигандом с одинаковой, большей или меньшей степенью, то такие антитела называются "перекрестно-конкурирующими" друг с другом за связывание их соответствующего(их) эпитопа(ов). Настоящее изобретение охватывает как конкурирующие, так и перекрестно-конкурирующие антитела. Независимо от механизма, посредством которого происходит такая конкуренция или перекрестная конкуренция (например, стерическое препятствие, конформационное изменение или связывание с общим эпитопом или его частью), специалисту будет понятно на основании информации, представленной в данной заявке, что такие конкурирующие и/или перекрестно-конкурирующие антитела включены и могут быть полезны для способов, раскрытых в данной заявке.
"Функциональная Fc-область" обладает по меньшей мере одной эффекторной функцией Fc-области с нативной последовательностью. Типичные "эффекторные функции" включают связывание C1q; комплемент-зависимую цитотоксичность; связывание Fc-рецептора; антитело-зависимую клеточно-опосредованную цитотоксичность; фагоцитоз; понижающую регуляцию поверхностных рецепторов клетки (например, рецепторов В-клеток) и т.д. Такие эффекторные функции, как правило, нуждаются в Fc-области для объединения со связывающим доменом (например, вариабельным доменом антитела) и могут быть оценены с использованием различных анализов, известных в данной области техники, для определения таких эффекторных функций антител.
"Fc-область с нативной последовательностью" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. "Вариант Fc-области" содержит аминокислотную последовательность, которая отличается от нативной последовательности Fc-области за счет по меньшей мере одной аминокислотной модификации, все еще сохраняет по меньшей мере одну эффекторную функцию нативной последовательности Fc-области. В некоторых воплощениях вариант Fc-области имеет по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-области или с Fc-областью родительского полипептида, например от примерно одного до примерно десяти аминокислотных замен и, предпочтительно от примерно одной до примерно пяти аминокислотных замен в нативной последовательности Fc-области или в Fc-области родительского полипептида. Вариант Fc-области в данной заявке будет предпочтительно иметь по меньшей мере примерно 80%-ную идентичность последовательности с нативной последовательностью Fc-области и/или с Fc-областью родительского полипептида и наиболее предпочтительно по меньшей мере примерно 90%-ную идентичность последовательности с ней, более предпочтительно по меньшей мере примерно 95%, по меньшей мере примерно 96%, по меньшей мере примерно 97%, по меньшей мере примерно 98%, по меньшей мере примерно 99%-ную идентичность последовательности с ней.
Как использовано в данной заявке, "лечение" представляет собой подход для получения полезных или желательных клинических результатов. Для целей настоящего изобретения полезные или желательные клинические результаты включают, но не ограничиваются этим, один или более из следующих: повышение клиренса глюкозы, снижение уровней глюкозы в крови, повышение толерантности к глюкозе, уменьшение частоты случаев высоких уровней глюкозы в крови вследствие диабета 1 или 2 типа, уменьшение частоты случаев или улучшение одного или более симптомов ревматоидного артрита, уменьшение частоты случаев или улучшение одного или более симптомов GVHD, уменьшение частоты случаев или улучшение одного или более симптомов волчанки и уменьшение частоты случаев или улучшение одного или более симптомов рассеянного склероза.
"Уменьшение частоты случаев" означает любое уменьшение тяжести (которое может включать уменьшение необходимости и/или количества (например, воздействия) других лекарственных средств и/или терапий, обычно используемых для этого состояния. Как понятно специалистам в данной области, индивидуумы могут варьироваться в отношении их ответа на лечение и, как таковой, например, "способ уменьшения частоты случаев" отражает введение антагониста IL-7R на основе обоснованного предположения, что такое введение сможет, вероятно, вызвать такое снижение уровня заболеваемости у этого конкретного индивидуума.
"Уменьшение интенсивности" означает ослабление или облегчение одного или более симптомов по сравнению с отсутствием введения антагониста IL-7R. "Уменьшение интенсивности" также включает сокращение или снижение продолжительности симптома.
Как использовано в данной заявке, "эффективная дозировка" или "эффективное количество" лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для воздействия на любой один или более полезных или желательных результатов. Для профилактического применения полезные или желательные результаты включают устранение или снижение риска, уменьшение тяжести или замедление начала заболевания, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические состояния, представленные во время развития заболевания. Для терапевтического применения полезные или желательные результаты включают клинические результаты, такие как уменьшение уровней глюкозы в крови, уменьшение частоты случаев или уменьшение интенсивности одного или более симптомов диабета 1 типа, диабета 2 типа, ревматоидного артрита, GVHD, волчанки или рассеянного склероза, уменьшение дозы других лекарственных средств, необходимых для лечения заболевания, усиление эффекта другого лекарственного средства и/или замедление развития заболевания у пациентов. Эффективную дозировку можно вводить за одно или более введений. Для целей данного изобретения эффективная дозировка лекарственного средства, соединения или фармацевтической композиции представляет собой количество, достаточное для осуществления профилактического или терапевтического лечения, прямо или опосредованно. Как понятно в клиническом контексте, эффективная дозировка лекарственного препарата, соединения или фармацевтической композиции может быть или не быть достигнута в сочетании с другим лекарственным средством, соединением или фармацевтической композиции. Таким образом, "эффективную дозировку" можно рассматривать в контексте введения одного или более терапевтических агентов, и один агент можно рассматривать как подлежащий введению в эффективном количестве, если в сочетании с одним или более другими агентами может быть достигнут или достигается желательный результат.
"Индивидуум" или "субъект" представляет собой млекопитающее, более предпочтительно человека. Млекопитающие также включают, но не ограничиваются этим, сельскохозяйственных животных, животных для спортивных состязаний, домашних животных, приматов, лошадей, собак, кошек, мышей и крыс.
Как использовано в данной заявке, "вектор" означает конструкцию, которая способна доставлять и предпочтительно экспрессировать один или более генов или последовательностей, представляющих интерес, в клетке-хозяине. Примеры векторов включают, но не ограничиваются этим, вирусные векторы, экспрессирующие векторы на основе депротеинизированной ДНК или РНК, плазмидные, космидные или фаговые векторы, ДНК- или РНК-экспрессирующие векторы, ассоциированные с катионными агентами конденсации, ДНК- или РНК-экспрессирующие векторы, инкапсулированные в липосомы, и некоторые эукариотические клетки, такие как клетки-продуценты.
Как использовано в данной заявке, "последовательность, контролирующая экспрессию" означает последовательность нуклеиновой кислоты, которая направляет транскрипцию нуклеиновой кислоты. Последовательность, контролирующая экспрессию, может представлять собой промотор, такой как конститутивный или индуцибельный промотор, или энхансер. Последовательность, контролирующая экспрессию, функциональным образом связана с транскрибируемой последовательностью нуклеиновой кислоты.
Как использовано в данной заявке, "фармацевтически приемлемый носитель" или "фармацевтически приемлемый эксципиент" включает любой материал, который при объединении с активным ингредиентом позволяет ингредиенту сохранять биологическую активность и быть нереактивным в отношении иммунной системы субъекта. Примеры включают, но не ограничиваются этим, любой из стандартных фармацевтических носителей, таких как забуференный фосфатами физиологический раствор, вода, эмульсии, такие как эмульсия масло/вода, и различные типы увлажняющих агентов. Предпочтительными разбавителями для аэрозоля или парентерального введения являются забуференный фосфатами физиологический раствор (PBS) или нормальный (0,9%) физиологический раствор. Композиции, содержащие такие носители, готовят с помощью хорошо известных традиционных методов (см., например, Remington's Pharmaceutical Sciences, 18th edition, A.Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000).
Термин "kon", как использовано в данной заявке, относится к константе скорости ассоциации антитела с антигеном. Конкретно, константы скорости (kon и koff) и равновесные константы диссоциации измеряют с использованием Fab-фрагментов антитела (т.е. одновалентных) и IL-7R.
Термин "koff", как использовано в данной заявке, относится к константе скорости диссоциации антитела из комплекса антитело/антиген.
Термин "KD", как использовано в данной заявке, относится к равновесной константе диссоциации антитело-антигенного взаимодействия.
Ссылка на "примерное" значение или параметр в данной заявке включает (и описывает) воплощения, которые направлены на это значение или параметр как таковые. Например, описание, относящееся к "примерно X", включает описание "X". Числовые диапазоны включают числа, определяющие указанный диапазон.
Следует понимать, что всюду, где воплощения описаны в данной заявке с использованием выражения "содержащий", также предложены иные аналогичные воплощения, описанные терминами "состоящий из" и/или "состоящий по существу из".
Если аспекты или воплощения изобретения описаны в терминах группы Маркуша или других групп альтернатив, то настоящее изобретение охватывает не только всю группу, перечисленную как единое целое, но и каждый член группы отдельно и все возможные подгруппы основной группы, а также основную группу при отсутствии одного или более членов группы. Настоящее изобретение также предусматривает недвусмысленное исключение одного или более любых членов группы в заявленном изобретении.
Если не определено иное, все технические и научные термины, используемые здесь, имеют такое же значение, которое обычно понятно специалисту в данной области техники, к которой относится данное изобретение. В случае конфликта настоящее описание, включая определения, будут контролировать. В данном описании и формуле изобретения слово "содержать", или вариации, такие как "содержит" или "содержащий", следует понимать как предполагающие включение указанного целого или группы целых чисел, но не исключение любого другого целого или группы целых чисел. Если иное не требуется по контексту, термины в единственном числе будут включать множества и термины во множественном числе будут включать форму в единственном числе.
Типичные способы и материалы описаны в данной заявке, хотя способы и материалы, аналогичные или эквивалентные описанным здесь, также могут быть использованы на практике или при тестировании настоящего изобретения. Материалы, способы и примеры являются только иллюстрацией и не предназначены для ограничения.
Способы предупреждения или лечения диабета 2 типа, GVHD и аутоиммунных расстройств
В одном аспекте в изобретении предложен способ лечения или предупреждения диабета 2 типа у индивидуума, включающий введение указанному индивидууму эффективного количества антагониста IL-7R, такого как, например, антагонистическое антитело к IL-7R. В другом аспекте в изобретении предложен способ лечения или предупреждения аутоиммунного заболевания, такого как диабет 1 типа, ревматоидный артрит, волчанка или рассеянный склероз, у индивидуума, включающий введение указанному индивидууму эффективного количества антагониста IL-7R. В другом аспекте в изобретении предложен способ лечения или предупреждения GVHD у индивидуума, включающий введение указанному индивидууму эффективного количества антагониста IL-7R.
В некоторых воплощениях терапевтическое введение антагониста IL-7R преимущественно приводит к снижению уровня глюкозы в крови и повышенной толерантности к глюкозе. В других воплощениях терапевтическое введение антагониста IL-7R преимущественно поддерживает количество глюкозы в крови на желательных уровнях.
В некоторых воплощениях терапевтическое введение антагониста IL-7R преимущественно приводит к пониженной заболеваемости и/или уменьшению интенсивности одного или более симптомов ревматоидного артрита, включая, например, без ограничения, тугоподвижность суставов, опухание суставов, боли в суставах, а также покраснение и нагревание суставов.
В некоторых воплощениях терапевтическое введение антагониста IL-7R преимущественно приводит к снижению заболеваемости и/или уменьшению интенсивности одного или более симптомов волчанки, включая, например, без ограничения, усталость, лихорадку, потерю массы тела, увеличение массы тела, боли в суставах, скованность в суставах, опухание суставов, сыпь на щеках (в виде бабочки), поражения кожи, язвы в полости рта, язвы в носовой полости, выпадение волос, феномен Рейно, одышку, боль в груди, сухость глаз, гематомы, тревогу, депрессию и потерю памяти.
В некоторых воплощениях терапевтическое введение антагониста IL-7R преимущественно приводит к снижению заболеваемости и/или уменьшению интенсивности одного или более симптомов рассеянного склероза, включая, например, без ограничения, паралич конечностей, тремор, трудности при ходьбе, трудности при глотании, слепоту, неясность зрения и мышечную слабость.
В некоторых воплощениях терапевтическое введение антагониста IL-7R преимущественно приводит к снижению заболеваемости и/или уменьшению интенсивности одного или более симптомов GVHD, включая, например, без ограничения, боль в животе, спазмы в животе, лихорадку, желтуху, кожную сыпь, рвоту, потерю массы тела, сухость глаз, сухость во рту, выпадение волос, гепатит, легочные расстройства и расстройства пищеварительного тракта.
В некоторых воплощениях терапевтическое введение антагониста IL-7R преимущественно приводит к снижению заболеваемости и/или уменьшению интенсивности одного или более симптомов острой GVHD, включая, например, без ограничения, пневмонию, воспаление кишечника, диарею, боли в животе, колики в животе, лихорадку, желтуху, тошноту, рвоту, поражение печени, сыпь на коже, повреждение кожи, повреждение слизистой оболочки, отторжение слизистой оболочки, повреждение желудочно-кишечного тракта, потерю массы тела, макулопапулезную сыпь, повышенные уровни билирубина, заболеваемость и смертность.
В некоторых воплощениях терапевтическое введение антагониста IL-7R преимущественно приводит к снижению заболеваемости и/или уменьшению интенсивности одного или более симптомов хронической GVHD, включая, например, без ограничения, сухость глаз, сухость во рту, выпадение волос, гепатит, легочные заболевания, расстройства желудочно-кишечного тракта, кожную сыпь, язву в полости рта, ротовую атрофию, ониходистрофию, синдром сухих глаз, склероз; поражения, напоминающие красный плоский лишай; пойкилодермию, мембраны пищевода, фасциит и облитерирующий бронхиолит и поражение печени, кожи и слизистых оболочек, соединительной ткани, экзокринных желез и/или желудочно-кишечного тракта.
Больного диабетом, нуждающегося в снижении уровней глюкозы в крови, можно лечить с помощью антагониста IL-7R, такого как, например, антагонистическое антитело к IL-7R. Индивидуума, подходящего для терапии антителами, выбирают, используя клинические критерии и прогностические показатели сахарного диабета, хорошо известные в данной области техники. Индивидуум, имеющий риск развития диабета, как оценивают с помощью известных прогностических показателей, таких как семейный анамнез, уровни глюкозы в крови натощак или пониженная толерантность к глюкозе, также нуждается в ведении антагониста IL-7R. Специалист в данной области техники может определить или узнать, как диагностировать индивидуума с диабетом или нарушенным усвоением глюкозы и в зависимости от степени или тяжести заболевания может сделать соответствующее определение, когда вводить антитело, а также выбрать наиболее желательный способ введения.
Индивидуума, страдающего ревматоидным артритом, можно лечить с помощью антагониста IL-7R, такого как, например, антагонистическое антитело к IL-7R. Индивидуума, подходящего для IL-7R антагонистической терапии, выбирают, используя клинические критерии и прогностические показатели ревматоидного артрита, которые хорошо известны в данной области техники. Диагностика и оценка ревматоидного артрита хорошо разработана в данной области техники. Оценку тяжести можно осуществлять на основе мер, известных в данной области техники, таких как шкала тяжести ревматоидного артрита (RASS). Bardwell et al., Rheumatology, 2002, 41:38-45. В некоторых воплощениях уменьшение интенсивности признаков, контролирование, уменьшение частоты случаев или замедление развития или прогрессирования ревматоидного артрита и/или симптомов ревматоидного артрита измеряют с помощью RASS.
Индивидуума, страдающего от волчанки, можно лечить с помощью антагониста IL-7R, такого как, например, антагонистическое антитело к IL-7R. Индивидуума, подходящего для IL-7R антагонистической терапии, выбирают, используя клинические критерии и прогностические показатели волчанки, которые хорошо известны в данной области техники. Специалист в данной области техники может определить или узнать, как диагностировать индивидуума с волчанкой и, в зависимости от степени или тяжести заболевания может соответственно определить, когда вводить антагонист IL-7R, а также выбрать наиболее желательный способ введения.
Индивидуума, страдающего от рассеянного склероза, можно лечить с помощью антагониста IL-7R, такого как, например, антагонистическое антитело к IL-7R. Индивидуума, подходящего для IL-7R антагонистической терапии, выбирают, используя клинические критерии и прогностические показатели рассеянного склероза, которые хорошо известны в данной области техники. Индивидуум, имеющий риск развития рассеянного склероза, как оценивают с помощью известных прогностических показателей, таких как семейный анамнез или история симптомов, также нуждается в ведении антагониста IL-7R. Специалист в данной области техники может определить или узнать, как диагностировать индивидуума с рассеянным склерозом, и в зависимости от степени или тяжести заболевания может соответственно определить, когда вводить антагонист IL-7R, а также может выбрать наиболее желательный способ введения.
Индивидуума, страдающего от GVHD, можно лечить с помощью антагониста IL-7R, такого как, например, антагонистическое антитело к IL-7R. Индивидуума, подходящего для IL-7R антагонистической терапии, выбирают, используя клинические критерии и прогностические показатели GVHD, которые хорошо известны в данной области техники. Диагностика и оценка GVHD хорошо разработана в данной области техники. Тесты для GVHD обычно зависят от симптомов, но могут включать желудочно-кишечную эндоскопию, с биопсией или без нее, тесты на функции печени (уровни AST, ALP и билирубина будут увеличены), биопсию печени, рентген легких и/или биопсию кожи. Признаки, достаточные для установления диагноза хронической GVHD, включают, например, без ограничения, склероз, поражения, подобные красному плоскому лишаю, пойкилодермию, мембраны пищевода, фасцит и облитерирующий бронхиолит (см., например, Leet and Flowers, Hematology, January 2008; 2008:134-141). Острую форму GVHD печени можно измерить с помощью, например, уровня билирубина у пациентов с острой формой. Острая форма GVHD кожи может привести к диффузной макулопапулезной сыпи.
Оценку тяжести GVHD можно осуществить, основываясь на измерениях, известных в данной области техники. В некоторых воплощениях уменьшение интенсивности, контролирование, уменьшение частоты случаев или замедление развития или прогрессирования GVHD и/или симптомов GVHD измеряют с помощью общего балла (overall grade) (кожа-печень-кишечник) для каждого органа, оцениваемого отдельно от низкого значения 1 до высокого значения 4. В некоторых воплощениях уменьшение интенсивности, контролирование, уменьшение частоты случаев или замедление развития или прогрессирования GVHD и/или симптомов GVHD измеряли с помощью мониторинга массы тела.
В отношении всех способов, описанных здесь, ссылка на антагонисты IL-7R также включает композиции, содержащие один или более дополнительных агентов. Эти композиции также могут содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципиенты, включающие буферы, которые хорошо известны в данной области техники. Настоящее изобретение может быть использовано отдельно или в комбинации с другими традиционными способами лечения.
Антагонист IL-7R можно вводить индивидууму любым подходящим путем. Специалисту в данной области будет очевидно, что примеры, описанные в данной заявке, предназначены не для ограничения, но для иллюстрации доступных методов. Соответственно, в некоторых воплощениях антагонист IL-7R вводят индивидууму согласно известным способам, таким как внутривенное введение, например, в виде болюса или посредством непрерывной инфузии в течение определенного периода времени, внутримышечным, внутрибрюшинным, интрацереброспинальным, трансдермальным, подкожным, внутрисуставным, сублингвальным, интрасиновиальным, посредством инсуффляции, интратекальным, пероральным, посредством ингаляции или местным путями. Введение может быть системным, например внутривенное введение, или локализованным. Коммерчески доступные небулайзеры для жидких препаратов, включая струйные небулайзеры и ультразвуковые небулайзеры, полезны для введения. Жидкие препараты можно непосредственно распылять, а лиофилизированный порошок можно распылять после разведения. Альтернативно антагонист IL-7R может быть в форме аэрозоля при использовании фторуглеродной композиции и дозирующего ингалятора или ингалируемого в виде лиофилизированного или измельченного порошка.
В одном воплощении антагонист IL-7R вводят посредством сайт-специфических или направленных локальных методов доставки. Примеры сайт-специфических или направленных локальных методов доставки включают различные имплантируемые депо-источники антагониста IL-7R или катетеры для местной доставки, такие как инфузионные катетеры, постоянные катетеры или игольные катетеры, синтетические трансплантаты, адвентициальные манжеты, шунты и стенты или другие имплантируемые устройства, сайт специфические носители, прямая инъекция или прямая аппликация. См., например, РСТ публикация №WO 00/53211 и патент США №5981568.
Для введения можно использовать различные препараты антагониста IL-7R. В некоторых воплощениях антагонист IL-7R можно вводить без примесей. В некоторых воплощениях антагонист IL-7R и фармацевтически приемлемый эксципиент может быть в различных препаратах. Фармацевтически приемлемые эксципиенты известны в данной области техники и представляют собой относительно инертные вещества, которые облегчают введение фармакологически эффективного вещества. Например, эксципиент может придать форму или консистенцию или действовать в качестве разбавителя. Подходящие эксципиенты включают, но не ограничиваются этим, стабилизирующие агенты, увлажняющие и эмульгирующие агенты, соли для варьирующей осмолярности, инкапсулирующие агенты, буферы и усилители проникновения через кожу. Эксципиенты, а также препараты для парентеральной и непарентеральной доставки лекарственного средства представлены в Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000.
В некоторых воплощениях эти агенты приготовлены для введения посредством инъекции (например, внутрибрюшинно, внутривенно, подкожно, внутримышечно и т.д.). Соответственно, эти агенты можно комбинировать с фармацевтически приемлемыми наполнителями, такими как физиологический раствор, раствор Рингера, раствор декстрозы и тому подобное. Особый режим дозирования, т.е. доза, сроки и повторяемость, будут зависеть от конкретного индивидуума и медицинской истории индивидуума.
Антагонист IL-7R можно вводить с использованием любого подходящего способа, в том числе посредством инъекции (например, внутрибрюшинно, внутривенно, подкожно, внутримышечно и т.д.). Антитела к IL-7R также можно вводить посредством ингаляции, как описано в данной заявке. Как правило, для введения антител к IL-7R начальная возможная дозировка может составлять примерно 2 мг/кг. Для цели настоящего изобретения обычная суточная дозировка может варьироваться в любом диапазоне от примерно 3 мкг/кг до 30 мкг/кг, до 300 мкг/кг до 3 мг/кг, до 30 мг/кг, до 100 мг/кг или более в зависимости от факторов, упомянутых выше. Например, можно использовать дозировку примерно 1 мг/кг, примерно 2,5 мг/кг, примерно 5 мг/кг, примерно 10 мг/кг и примерно 25 мг/кг. Для повторных введений в течение нескольких суток или дольше в зависимости от состояния лечение поддерживают до появления желаемого подавления симптомов или до достижения достаточных терапевтических уровней, например до уменьшения уровней глюкозы в крови. Примерный режим дозирования включает введение начальной дозы примерно 2 мг/кг, последующей еженедельной поддерживающей дозы примерно 1 мг/кг антитела к IL-7R или последующей поддерживающей дозы примерно 1 мг/кг один раз в две недели. Однако могут быть полезны и другие дозовые режимы в зависимости от картины фармакокинетического убывания, которого хочет достичь практикующий врач. Например, в некоторых воплощениях предполагается дозировка от одного до четырех раз в неделю. В других воплощениях предполагается дозировка один раз в месяц или один раз в два месяца или один раз в три месяца. Успешность этой терапии легко контролировать с помощью обычных методов и анализов. Режим дозирования (включая используемый(е) антагонист(ы) IL-7R) может варьироваться с течением времени.
Для цели настоящего изобретения подходящая дозировка антагониста IL-7R будет зависеть от используемого антагониста IL-7R (или его композиций), типа и тяжести симптомов, подлежащих лечению, от того, вводят ли агент для профилактических или терапевтических целей, предшествующей терапии, клинической истории пациента и ответа на агент, скорости клиренса введенного агента у пациента и выбора, сделанного лечащим врачом. Обычно врач будет вводить антагонист IL-7R до тех пор, пока не будет достигаться дозировка, которая дает желаемый результат. Доза и/или частота может варьироваться в процессе лечения. Эмпирические соображения, такие как период полувыведения, обычно вносят вклад в определение дозировки. Например, антитела, которые совместимы с иммунной системой человека, такие как гуманизированные антитела или полностью человеческие антитела, могут быть использованы для увеличения периода полувыведения антитела и для предупреждения атаки на антитело со стороны иммунной системы хозяина. Частоту введения можно определить и скорректировать в процессе терапии и, как правило, но не обязательно, основываясь на лечении и/или подавлении и/или уменьшении интенсивности и/или замедлении симптомов, например высоких уровней глюкозы в крови, боли в суставах и т.д. Альтернативно могут быть целесообразными препараты с замедленным непрерывным высвобождением антагонистических антител к IL-7R. Различные препараты и устройства для достижения замедленного высвобождения известны в данной области техники.
В одном воплощении дозировки для антагониста IL-7R можно определить эмпирически у индивидуумов, которые получали одно или более введений антагониста IL-7R. Индивидуумы получают постепенно возрастающие дозировки антагониста IL-7R. Для оценки эффективности можно отслеживать индикатор заболевания.
Введение антагониста IL-7R согласно способу в настоящем изобретении может быть непрерывным или прерывистым в зависимости, например, от физиологического состояния реципиента независимо от того, является ли введение терапевтическим или профилактическим, и других факторов, известных опытным практикующим врачам. Введение антагониста IL-7R может быть по существу непрерывным в течение заданного периода времени или может быть серией разделенных временными интервалами доз.
В некоторых воплощениях может присутствовать более чем один антагонист IL-7R. Может присутствовать по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять различных или более антагонистов IL-7R. Как правило, эти антагонисты IL-7R могут иметь взаимодополняющие активности, которые не оказывают друг на друга отрицательного влияния. Например, можно использовать один или более следующих антагонистов IL-7R: антагонистическое антитело к IL-7R, антисмысловую молекулу, направленную на IL-7R (включая антисмысловую молекулу, направленную на нуклеиновую кислоту, кодирующую IL-7R), соединение, ингибирующее IL-7R, и структурный аналог IL-7R. Антагонист IL-7R также можно использовать в сочетании с другими агентами, которые служат для увеличения и/или дополнения эффективности агентов.
Терапевтические препараты антагониста IL-7R, используемые в соответствии с настоящим изобретением, готовят для хранения путем смешивания антитела, имеющего желаемую чистоту, с оптимальными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000) в форме лиофилизированных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов при используемых дозировках и концентрациях и могут содержать буферы, такие как фосфатный, цитратный, и другие органические кислоты, соли, такие как хлорид натрия; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид, бензэтония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG).
Липосомы, содержащие антагонист IL-7R, готовят с помощью способов, известных в данной области техники, таких как описаны в Epstein, et al., Proc. Natl. Acad. Sci. USA 82:3688, 1985; Hwang, et al., Proc. Natl Acad. Sci. USA 77:4030, 1980; и патентах США №№4485045 и 4544545. Липосомы с увеличенным временем циркуляции раскрыты в патенте США №5013556. Особенно полезные липосомы могут быть получены с помощью способа обращенно-фазового выпаривания с липидной композицией, содержащей фосфатидилхолин, холестерин и PEG-дериватизированный фосфатидилэтаноламин (PEG-РЕ). Липосомы выдавливают через фильтры с определенным размером пор с получением липосом с желаемым диаметром.
Активные ингредиенты также могут быть заключены в микрокапсулы, полученные, например, методами коацервации или межфазной полимеризации, например гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметакрилат)ные микрокапсулы соответственно в коллоидных системах доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие способы раскрыты в Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000.
Могут быть получены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где эти матрицы находятся в форме профилированных изделий, например пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поливиниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и 7-этил-L-глутамата, неразлагаемый этилен-винилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT ™ (микросферы для инъекций, состоящие из сополимера молочной кислоты-гликолевой кислоты и лейпролидацетата), сахарозы ацетат изобутират и поли-D-(-)-3-оксимасляную кислоту.
Препараты, используемые для введения in vivo, должны быть стерильными. Это легко достигается, например, посредством фильтрации через стерильные фильтрующие мембраны. Композиции терапевтического антагониста IL-7R, как правило, помещают в контейнер, имеющий стерильное входное отверстие, например, контейнер для внутривенного раствора или флакон, имеющий пробку, прокалываемую иглой для подкожных инъекций.
Композиции согласно настоящему изобретению могут находиться в стандартных лекарственных формах, таких как таблетки, пилюли, капсулы, порошки, гранулы, растворы, суспензии или суппозитории для перорального, парентерального или ректального введения или введения посредством ингаляции или инсуффляции.
Для получения твердых композиций, таких как таблетки, основной активный ингредиент смешивают с фармацевтическим носителем, например обычными ингредиентами для таблеток, такими как кукурузный крахмал, лактоза, сахароза, сорбит, тальк, стеариновая кислота, стеарат магния, дикальцийфосфат или камеди, и другие фармацевтические разбавители, например вода, для образования твердой предварительной композиции, содержащей гомогенную смесь соединения по настоящему изобретению или его нетоксичную фармацевтически приемлемую соль. Ссылаясь на эти предварительные композиции как на гомогенные, подразумевают, что активный ингредиент распределен равномерно по всей композиции, так что композицию можно легко разделить на равные эффективные стандартные лекарственные формы, такие как таблетки, пилюли и капсулы. Эту твердую предварительную композицию затем делят на стандартные лекарственные формы вышеописанного типа, содержащие от 0,1 до примерно 500 мг активного ингредиента по настоящему изобретению. Таблетки или пилюли новой композиции могут быть покрыты или иным образом компаундированы для получения лекарственной формы, обеспечивающей преимущество пролонгированного действия. Например, таблетка или пилюля может содержать внутренний дозовый и наружный дозовый компонент, причем последний находится в форме оболочки над первым компонентом. Два компонента могут быть разделены с помощью энтеросолюбильного слоя, который служит для предотвращения распада в желудке и позволяет внутреннему компоненту пройти интактным в двенадцатиперстную кишку или иметь замедленное высвобождение. Для таких энтеросолюбильных слоев или покрытий могут быть использованы различные материалы, такие материалы включают ряд полимерных кислот и смеси полимерных кислот с такими материалами, как шеллак, цетиловый спирт и ацетат целлюлозы.
Подходящие поверхностно-активные вещества включают, в частности, неионные агенты, такие как полиоксиэтиленсорбитаны (например, Tween™ 20, 40, 60, 80 или 85) и другие сорбитаны (например, Span™ 20, 40, 60, 80 или 85). Композиции с поверхностно-активным агентом, как правило, содержат от 0,05 до 5% поверхностно-активного агента и могут содержать от 0,1 до 2,5%. Следует иметь в виду, что могут быть добавлены и другие ингредиенты, например маннит или другие фармацевтически приемлемые наполнители, при необходимости.
Подходящие эмульсии могут быть получены с использованием коммерчески доступных жировых эмульсий, таких как Intralipid™, Liposyn™, Infonutrol™, Lipofundin™ и Lipiphysan™. Активный ингредиент может быть растворен в предварительно смешанной эмульсионной композиции, или, альтернативно, он может быть растворен в масле (например, соевом масле, подсолнечном масле, хлопковом масле, кунжутном масле, кукурузном масле или миндальном масле) и в эмульсии, образующейся при смешивании с фосфолипидом (например, яичными фосфолипидами, соевыми фосфолипидами или соевым лецитином) и водой. Следует иметь в виду, что могут быть добавлены и другие ингредиенты, например глицерин или глюкоза, для регулирования тоничности эмульсии. Подходящие эмульсии, как правило, содержат вплоть до 20% масла, например от 5 до 20%. Жировая эмульсия может содержать жировые капли от 0,1 до 1,0 мкм, особенно от 0,1 до 0,5 мкм, и иметь рН в диапазоне от 5,5 до 8,0.
Эмульсионные композиции могут быть получены путем смешивания антагониста IL-7R с Intralipid™ или его компонентами (соевым маслом, яичными фосфолипидами, глицерином и водой).
Композиции для ингаляции или инсуффляции включают растворы и суспензии в фармацевтически приемлемых, водных или органических растворителях или их смеси и порошки. Жидкие и твердые композиции могут содержать подходящие фармацевтически приемлемые эксципиенты, как указано выше. В некоторых воплощениях композиции вводят пероральным или назальным респираторным путем для локального или системного эффекта. Композиции в предпочтительно стерильных фармацевтически приемлемых растворителях можно распылить с использованием газов. Распыленные растворы можно вдыхать непосредственно из небулайзерного устройства, или небулайзерное устройство может быть присоединено к маске для лица, палатке или машине с перемежающимся положительным давлением. Композиции раствора, суспензии или порошка можно вводить предпочтительно перорально или интраназально из устройств, которые доставляют композиции соответствующим образом.
Антагонисты IL-7R
В способах по изобретению используют антагонист IL-7R, который относится к любой белковой, пептидной или нуклеиновокислотной молекуле, которая блокирует, подавляет или снижает (в том числе значительно снижает) биологическую активность IL-7R, включая нижерасположенные пути, опосредованные передачей сигнала IL-7R, такие как выявление клеточного ответа на IL-7R. Примеры антагонистов IL-7R включают, без ограничения, антагонистические антитела к IL-7R, siRNA IL-7R, shRNA IL-7R и антисмысловые олигонуклеотиды IL-7R.
Антагонист IL-7R должен демонстрировать любую одну или более из следующих характеристик: (а) связываться с IL-7R; (б) блокировать взаимодействие IL-7R с IL-7; (в) блокировать или уменьшать IL-7-опосредованное фосфорилирование STAT5; (г) уменьшать уровни глюкозы в крови in vivo; (д) повышать толерантность к глюкозе in vivo; (е) снижать тяжесть заболевания при экспериментальном аутоиммунном энцефаломиелите (ЕАЕ); (ж) блокировать или уменьшать фосфорилирование PI3K; (з) блокировать или уменьшать фосфорилирование АКТ и (и) блокировать взаимодействие IL-7R с другими, еще не идентифицированными факторами.
В некоторых воплощениях антагонист IL-7R представляет собой антагонистическое антитело к IL-7R. Для целей настоящего изобретения антагонистическое антитело к IL-7R предпочтительно взаимодействует с IL-7Rα, ингибируя сигнальную функцию IL-7R и взаимодействие с IL-7. В некоторых воплощениях антагонистическое антитело к IL-7R специфически распознает IL-7R приматов. В некоторых воплощениях антагонистическое антитело к IL-7R связывается с IL-7R приматов и грызунов.
Антитела, полезные для настоящего изобретения, могут охватывать моноклональные антитела, поликлональные антитела, фрагменты антител (например, Fab, Fab', F(ab')2, Fv, Fc и т.д.), химерные антитела, биспецифические антитела, гетероконъюгатные антитела, одноцепочечные (ScFv), их мутанты, слитые белки, содержащие часть антитела (например, доменное антитело), гуманизированные антитела и любую другую модифицированную конфигурацию иммуноглобулиновой молекулы, которая содержит сайт распознавания антигена необходимой специфичности, включая гликозилированные варианты антител, варианты аминокислотной последовательности антител и ковалентно модифицированные антитела. Антитела могут быть мышиного, крысиного, человеческого или любого другого происхождения (включая химерные или гуманизированные антитела).
В некоторых воплощениях антагонистическое антитело к IL-7R представляет собой моноклональное антитело. Антагонистическое антитело к IL-7R также может быть гуманизированным. В других воплощениях антитело является человеческим.
В некоторых воплощениях антитело содержит модифицированную константную область, такую как, например, без ограничения, константную область, которая имеет повышенный потенциал для индукции иммунного ответа. Например, константная область может быть модифицирована для увеличения аффинности к Fc-гамма-рецептору, такому как, например, FcyRI или FcyRllA.
В некоторых воплощениях антитело содержит модифицированную константную область, такую как константная область, которая является иммунологически инертной, т.е. имеет сниженный потенциал для индукции иммунного ответа. В некоторых воплощениях константная область модифицирована, как описано в Eur. J. Immunol., 1999, 29:2613-2624; РСТ заявке №PCT/GB99/01441; и/или заявке на патент Великобритании №9809951.8. Fc может быть IgG1 человека, IgG2 человека или IgG4 человека. Fc может быть IgG2 человека, содержащего мутацию А330Р331 в S330S331 (IgG2Aa), в которой аминокислотные остатки пронумерованы со ссылкой на последовательность IgG2 дикого типа. Eur. J. Immunol., 1999, 29:2613-2624. В некоторых воплощениях антитело содержит константную область IgG4,, содержащую следующие мутации (Armour et al., 2003, Molecular Immunology 40 585-593): E233F234L235 в P233V234A235 (IgG4Ac), в который нумерация приведена со ссылкой на IgG4 дикого типа. В еще одном воплощении Fc представляет собой человеческий IgG4 E233F234L235 в P233V234A235 с делецией G236 (IgG4Ab). В другом воплощении Fc представляет собой любой человеческий IgG4 Fc (IgG4, IgG4Ab или IgG4Ac), содержащий шарнир-стабилизирующую мутацию S228 в Р228 (Aalberse et al., 2002, Immunology 105, 9-19). В другом воплощении Fc может представлять собой агликозилированный Fc.
В некоторых воплощениях константная область является агликозилированной в результате мутации остатка, присоединяющего олигосахарид (такого как Asn297), и/или фланкирующих остатков, которые являются частью последовательности распознавания гликозилирования в константной области. В некоторых воплощениях константная область является агликозилированной для ферментативного N-связанного гликозилирования. Константная область может быть агликозилированной для ферментативного N-связанного гликозилирования или благодаря экспрессии в клетках-хозяевах, дефектных по гликозилированию.
Аффинность связывания (KD) антагонистического антитела к IL-7R с IL-7R (такого как человеческий IL-7R) может составлять от примерно 0,002 до примерно 200 нМ. В некоторых воплощениях аффинность связывания представляет собой любое значение из примерно 200 нМ, примерно 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ, примерно 60 пМ, примерно 50 пМ, примерно 20 пМ, примерно 15 пМ, примерно 10 пМ, примерно 5 пМ или примерно 2 пМ. В некоторых воплощениях аффинность связывания составляет менее чем любое значение из примерно 250 нМ, примерно 200 нМ, примерно 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ, примерно 50 пМ, примерно 20 пМ, примерно 10 пМ, примерно 5 пМ или примерно 2 пМ.
Один способ определения аффинности связывания антител с IL-7R заключается в измерении аффинности связывания монофункциональных Fab-фрагментов антитела. Для получения монофункциональных Fab-фрагментов, антитело (например, IgG) можно расщепить с помощью папаина или рекомбинантно экспрессировать. Аффинность IL-7R Fab-фрагмента антитела можно определить с помощью поверхностного плазмонного резонанса (Biacore™3000™ surface plasmon resonance (SPR) system, Biacore™, INC, Piscataway NJ), оснащенного предварительно иммобилизованными стрептавидиновыми сенсорными чипами (SA), с использованием HBS-EP подвижного буфера (0,01 М HEPES, рН 7,4, 0,15 NaCI, 3 мМ EDTA, 0,005% об./об. Surfactant Р20). Биотинилированный человеческий IL-7R (или любой другой IL-7R) можно разводить в HBS-EP буфере до концентрации менее 0,5 мкг/мл и вводить через индивидуальные каналы чипов, используя варьирующееся время контакта, для достижения двух диапазонов плотности антигена: либо 50-200 единиц ответа (RU) для детальных кинетических исследований, либо 800-1000 RU для скрининговых анализов. Исследования регенерации показали, что 25 мМ NaOH в 25% об./об. этаноле эффективно удаляет связанный Fab, сохраняя при этом активность IL-7R на чипе в течение более 200 инъекций. Как правило, серийные разведения (охватывающие концентрации 0,1-10х подсчитанного KD) очищенных Fab образцов вводят в течение 1 мин при 100 мкл/мин и допускается время диссоциации вплоть до 2 часов. Концентрации Fab белков определяют с помощью ELISA (иммуноферментный твердофазный анализ) и/или SDS-PAGE электрофореза (электрофорез в полиакриламидном геле с додецилсульфатом натрия) с использованием Fab с известной концентрацией (как определено с помощью аминокислотного анализа) в качестве стандарта. Кинетические скорости ассоциации (kon) и диссоциации (koff) получают одновременно путем подгонки данных на глобальном уровне до 1:1 модели связывания Ленгмюра (Karlsson, R.Roos, Н.Fagerstam, L.Petersson, В. (1994). Methods Enzymology 6. 99-110) с использованием программы В devaluation. Значения равновесных констант диссоциации (KD) рассчитывают как koff/kon - Этот протокол подходит для применения в определении аффинности связывания антитела с любым IL-7R, включая человеческий IL-7R, IL-7R другого млекопитающего (таким как мышиный IL-7R, крысиный IL-7R, IL-7R приматов), а также различные формы IL-7R. Аффинность связывания антитела, как правило, измеряют при 25°С, но также можно было измерять при 37°С.
Антагонистические антитела к IL-7R могут быть получены любым способом, известным в данной области техники, включая способ, предложенный в Примере 1. Для получения гибридомных клеточных линий способ и режим иммунизации животного-хозяина соответствует обычно установленным и традиционным методам стимуляции и продукции антител, как описано далее в данной заявке. Общие методы получения человеческих и мышиных антител известны в данной области техники и/или описаны здесь.
Предполагается, что любое млекопитающее, включая людей, или их антитело-продуцирующие клетки, может служить основой для получения клеточных линий млекопитающих, включая человеческие, гибридомные клеточные линии. Как правило, животное-хозяин подвергают инокуляции внутрибрюшинно, внутримышечно, перорально, подкожно, в подошву и/или внутрикожно количеством иммуногена, в том числе, как описано в данной заявке.
Гибридомы могут быть получены из лимфоцитов и иммортализованных клеток миеломы с использованием общих методов гибридизации соматических клеток согласно Kohler, В. and Milstein, С, 1975, Nature 256:495-497, или как модифицировано в Buck, D. W., et al., In Vitro, 18:377-381, 1982. Для гибридизации могут быть использованы доступные миеломные линии, включая, но не ограничиваясь этим, Х63-Ag8.653 и полученные из Института Солка (Salk Institute, Cell Distribution Center, San Diego, Calif., США). Как правило, методы включают слияние миеломных клеток и лимфоидных клеток с использованием вещества, способствующего слиянию клеток, такого как полиэтиленгликоль, или электрическими средствами, хорошо известными специалистам в данной области. После слияния клетки выделяют из среды для слияния и выращивают в селективной ростовой среде, такой как среда HAT (гипоксантин-аминоптерин-тимидин), для удаления негибридизованных родительских клеток. Любую среду, описанную в данной заявке, дополненную сывороткой или без сыворотки, можно использовать для культивирования гибридом, которые секретируют моноклональные антитела. В качестве другой альтернативы методу слияния клеток можно использовать EBV-иммортализованные В-клетки для продукции IL-7R моноклональных антител субъекта по изобретению. Гибридомы наращивают и субклонируют при желании и супернатанты анализируют на антииммуногенную активность с помощью традиционных процедур иммуноанализа (например, радиоиммуноанализа, иммуноферментного анализа или флуоресцентного иммуноанализа).
Гибридомы, которые могут быть использованы в качестве источника антител, охватывают все производные, клетки-потомки родительских гибридом, которые продуцируют моноклональные антитела, специфичные в отношении IL-7R или его участка.
Гибридомы, которые продуцируют такие антитела, можно выращивать in vitro или in vivo с использованием известных процедур. Моноклональные антитела могут быть выделены из культуральных сред или жидкостей организма с помощью стандартных процедур очистки иммуноглобулинов, таких как осаждение сульфатом аммония, гель-электрофорез, диализ, хроматография и ультрафильтрация, при необходимости. Можно удалить нежелательную активность, если она присутствует, например, путем пропускания препарата через адсорбенты, изготовленные из иммуногена, присоединенного к твердой фазе, и элюирования или отделения желаемых антител от иммуногена. Иммунизация животного-хозяина человеческим IL-7Rα или фрагментом, содержащим целевую аминокислотную последовательность, конъюгированную с белком, который является иммуногенным для видов, подвергнутых иммунизации, таким как, например, гемоцианин лимфы улитки, сывороточный альбумин, бычий тиреоглобулин или соевый ингибитор трипсина, с использованием бифункционального или дериватизирующего агента, например малеимидобензоилсульфосукцинимидного сложного эфира (сопряжение через остатки цистеина), N-гидроксисукцинимида (через остатки лизина), глутаральдегида, янтарного ангидрида, SOCl2 или R1N=C=NR, где R и R1 представляют собой различные алкильные группы, может дать популяцию антител (например, моноклональных антител).
При необходимости антагонистическое антитело к IL-7R (моноклональное или поликлональное), представляющее интерес, можно секвенировать и полинуклеотидную последовательность затем можно клонировать в вектор для экспрессии или размножения. Последовательность, кодирующую интересующее антитело, можно поддерживать в векторе в клетке-хозяине, которую затем можно размножить и заморозить для будущего применения. Продукцию рекомбинантных моноклональных антител в клеточной культуре можно осуществить путем клонирования генов антитела из В-клеток средствами, известными в данной области техники. См., например, Tiller et al., 2008, J. Immunol. Methods 329, 112; патент США №7314622.
В качестве альтернативы полинуклеотидная последовательность может быть использована для генетической манипуляции с целью "гуманизации" антитела или улучшения аффинности или других характеристик антитела. Например, константную область можно сконструировать более похожей на человеческие константные области для того, чтобы избежать иммунного ответа при использовании антитела в клинических исследованиях и в лечении людей. Желательным является генетическое манипулирование последовательностью антитела для получения большей аффинности к IL-7R и большей эффективности в ингибировании IL-7R. Специалисту в данной области будет очевидно, что одно или более полинуклеотидных изменений можно применить к антагонистическому антителу к IL-7R и все же поддерживать его способность связываться с IL-7R.
Существуют четыре основных стадии гуманизации моноклонального антитела. К ним относятся: (1) определение нуклеотидной и предсказанной аминокислотной последовательности вариабельных доменов легкой и тяжелой цепи исходного антитела; (2) конструирование гуманизированного антитела, т.е. определение того, какой каркасный участок антитела использовать в процессе гуманизации; (3) существующие в настоящее время методологии/способы гуманизации; (4) трансфекция и экспрессия гуманизированного антитела. См., например, патенты США №№4816567; 5807715; 5866692; 6331415; 5530101; 5693761; 5693762; 5585089 и 6180370.
Был описан ряд молекул "гуманизированного" антитела, содержащего антигенсвязывающий сайт, происходящий из нечеловеческого иммуноглобулина, в том числе химерные антитела, имеющие V-области грызунов или модифицированные V-области грызунов и связанные с ними CDRs, слитые с человеческими константными областями. См., например, Winter et al. Nature 349:293-299, 1991, Lobuglio et al. Proc. Nat. Acad. Sci. USA 86:4220-4224, 1989, Shaw et al. J Immunol. 138:4534-4538, 1987 и Brown et al. Cancer Res. 47:3577-3583, 1987. Другие ссылки описывают CDRs грызунов, перенесенные на человеческий поддерживающий каркасный участок (FR) перед слиянием с соответствующей константной областью антитела человека. См., например, Riechmann et al. Nature 332:323-327, 1988, Verhoeyen et al. Science 239:1534-1536, 1988 и Jones et al. Nature 321:522-525, 1986. Другая ссылка описывает CDRs грызунов, поддерживаемых рекомбинантно сконструированными каркасными участками грызунов. См., например, Европейскую патентную публикацию №0519596. Эти "гуманизированные" молекулы конструируют для минимизации нежелательного иммунологического ответа в отношении молекул антител грызунов против человека, которые ограничивают продолжительность и эффективность терапевтического применения этих группировок у реципиентов-людей. Например, константную область антитела можно сконструировать таким образом, чтобы она была иммунологически инертной (например, не запускала лизис комплемента). См., например, РСТ публикацию №PCT/GB99/01441; заявку на патент Великобритании №9809951.8. Также могут быть использованы другие способы гуманизации антител, раскрытые в Daugherty et al., Nucl. Acids Res. 19:2471-2476, 1991 и в патенте США №№ 6180377; 6054297; 5997867; 5866692; 6210671; и 6350861; и в РСТ публикации №WO 01/27160.
Очевидно, что хотя вышеописанное обсуждение относится к гуманизированным антителам, обсуждаемые общие принципы применимы к изготавливаемым антителам для применения, например, у собак, кошек, приматов, лошадей и крупного рогатого скота. Кроме того, очевидно, что один или более аспектов гуманизации антитела, описанного в данной заявке, можно комбинировать, например, перенос CDR, мутацию каркасного участка и мутацию CDR.
В еще одной альтернативе полностью человеческие антитела могут быть получены с использованием коммерчески доступных мышей, которые были сконструированы для экспрессии специфических человеческих иммуноглобулиновых белков. Также могут быть использованы трансгенные животные, сконструированные для продукции более желательных антител (например, полностью человеческих антител) или более мощного иммунного ответа для создания гуманизированных или человеческих антител. Примеры такой технологии представляют собой Xenomouse™ от Abgenix, Inc. (Fremont, СА) и HuMAb-Mouse® и ТС Mouse™ от Medarex, Inc. (Princeton, NJ).
В альтернативном варианте антитела можно получать рекомбинантным методом и экспрессировать с использованием любого способа, известного в данной области техники. В другом альтернативном варианте антитела можно получать рекомбинантным методом с помощью технологии фагового дисплея. См., например, патенты США №№5565332; 5580717; 5733743; и 6265150; и Winter et al., Annu. Rev. Immunol. 12:433-455, 1994. Альтернативно технологию фагового дисплея (McCafferty et al., Nature 348:552-553, 1990) можно использовать для получения человеческих антител и фрагментов антител in vitro, из репертуара генов вариабельного домена (V) иммуноглобулина от иммунизированных доноров. Согласно этому методу гены V-доменов антитела клонируют "в рамке" в ген либо мажорного, либо минорного оболочечного белка нитчатого бактериофага, такого как М13 или fd, и экспонируют в виде функциональных фрагментов антитела на поверхности фаговой частицы. Поскольку нитчатая частица содержит одноцепочечную ДНК-копию фагового генома, селекции, основанные на функциональных свойствах антитела, также приводят к селекции гена, кодирующего антитело, демонстрирующее эти свойства. Таким образом, фаг имитирует некоторые из свойств В-клетки. Фаговый дисплей можно осуществлять в различных форматах; для обзора см., например, Johnson, Kevin S. and Chiswell, David J., Current Opinion in Structural Biology 3:564-571, 1993. Для фагового дисплея можно использовать несколько источников V-генных сегментов. Clackson et al., Nature 352:624-628, 1991, выделили разнообразный набор антител против оксазолона из небольшой случайной комбинаторной библиотеки V-генов, происходящих из селезенок иммунизированных мышей. Можно сконструировать репертуар V-генов из неиммунизированных человеческих доноров, и антитела к разнообразному набору антигенов (включая аутоантигены) могут быть выделены по существу в соответствии с методами, описанными Mark et al., J. Mol. Biol. 222:581-597, 1991, или Griffith et al., EMBO J. 12:725-734, 1993. При естественном иммунном ответе гены антитела накапливают мутации с высокой скоростью (соматическое гипермутирование). Некоторые из введенных изменений будут обеспечивать более высокую аффинность, и В-клетки, экспонирующие высокоаффинный поверхностный иммуноглобулин, предпочтительно реплицируются и дифференцируются во время последующей встречи с антигеном. Этот естественный процесс можно имитировать, используя метод, известный как "перетасовка цепей". (Marks et al., Bio/Technol. 10:779-783, 1992). В этом способе аффинность "первичных" человеческих антител, полученных с помощью фагового дисплея, можно улучшить посредством последовательной замены генов V-областей тяжелой и легкой цепи репертуарами встречающихся в природе вариантов (репертуаров) генов V-доменов, полученных от иммунизированных доноров. Этот метод позволяет получать антитела и фрагменты антител с аффинностью в пМ-нМ диапазоне. Стратегия получения очень больших репертуаров фаговых антител (также известных как "мать всех библиотек", от англ. "the mother-of-all libraries") была описана в Waterhouse et al., Nucl. Acids Res. 21:2265-2266, 1993. Перетасовку генов также можно использовать для получения человеческих антител из антител грызунов, где человеческое антитело имеет сходные аффинности и специфичности с исходным антителом грызунов. Согласно этому способу, который также называется "эпитопным импринтингом", ген V-доменов тяжелой или легкой цепи антител грызунов, полученный методом фагового дисплея, заменяют репертуаром генов V-доменов человека, создавая химеры между грызуном и человеком. Селекция антигена приводит к выделению человеческих вариабельных областей, способных восстанавливать функциональный антигенсвязывающий сайт, т.е. эпитоп определяет (фиксирует, от англ. imprints) выбор партнера. При повторении процесса с целью замены оставшегося V-домена грызуна получают человеческое антитело (см. РСТ публикацию №WO 93/06213). В отличие от традиционной гуманизации антител грызунов посредством CDR-переноса (grafting), этот способ обеспечивает полностью человеческие антитела, которые не имеют каркасных или CDR остатков, происходящих из грызунов.
Антитела могут быть получены рекомбинантно сначала путем выделения антител и антитело-продуцирующих клеток из животных-хозяев с получением последовательности гена и использованием этой последовательности гена для экспрессии антитела рекомбинантно в клетках-хозяевах (например, клетках СНО). Другой способ, который может быть использован, заключается в экспрессии последовательности антитела в растениях (например, табаке) или трансгенном молоке. Методы рекомбинантной экспрессии в растениях или молоке были описаны. См., например, Peeters, et al. Vaccine 19:2756, 2001; Lonberg, N. and D.Huszar Int. Rev. Immunol 13:65, 1995; и Pollock, et al., J Immunol Methods 231:147, 1999. Способы получения производных антител, например гуманизированных, одноцепочечных и т.д., известны в данной области техники.
Иммуноанализы и методы проточной цитометрии и сортировки, такие как сортировка клеток с возбуждением флуоресценции (FACS), также могут быть использованы для выделения антител, которые специфичны в отношении IL-7R.
Антитела могут быть связаны со многими различными носителями. Носители могут быть активными и/или инертными. Примеры хорошо известных носителей включают полипропилен, полистирол, полиэтилен, декстран, нейлон, амилазы, стекло, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. Носитель по своей природе может быть либо растворимым, либо нерастворимым для целей изобретения. Специалисты в данной области обычно знают и другие подходящие носители для связывания антител или способны установить их с использованием рутинного экспериментирования. В некоторых воплощениях носитель содержит группировку, которая нацелена на миокард.
ДНК, кодирующую моноклональные антитела, легко выделяют и секвенируют с использованием стандартных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно поместить в экспрессирующие векторы (такие как экспрессирующие векторы, раскрытые в РСТ публикации №WO 87/04462), которые затем трансфицируют в клетки-хозяева, такие как клетки Е. coli, обезьяньи клетки COS, клетки яичника китайского хомячка (СНО) или миеломные клетки, которые в ином случае не продуцируют иммуноглобулиновый белок, для получения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. См., например, РСТ публикацию №WO 87/04462. ДНК также можно модифицировать, например, посредством замещения кодирующей последовательности на человеческие константные области тяжелой и легкой цепей вместо гомологичных мышиных последовательностей, Morrison et al., Proc. Nat. Acad. Sci. 81:6851, 1984, или посредством ковалентного связывания со всей или частью иммуноглобулин-кодирующей последовательности для неиммуноглобулинового полипептида. Таким способом получают "химерные" или "гибридные" антитела, которые имеют специфичность связывания в отношении IL-7R моноклонального антитела в данной заявке.
Антагонистические антитела к IL-7R можно идентифицировать или охарактеризовать с использованием способов, известных в данной области техники, посредством чего определяют и/или измеряют снижение, уменьшение интенсивности или нейтрализацию биологической активности IL-7R. В некоторых воплощениях идентифицируют антагонистическое антитело к IL-7R посредством инкубации кандидатного агента с IL-7R и мониторинга связывания и/или сопутствующего снижения или нейтрализации биологической активности IL-7R. Анализ связывания можно осуществлять с очищенным(и) полипептидом(ами) IL-7R или с клетками, естественно экспрессирующими полипептид(ы) IL-7R или трансфицированными для экспрессии полипептида(ов) IL-7R. В одном воплощении анализ связывания представляет собой конкурентный анализ связывания, где оценивают способность кандидатного антитела конкурировать с известным антагонистом IL-7R за связывание с IL-7R. Анализ можно осуществлять в различных форматах, включая формат ELISA. В других воплощениях антагонистическое антитело к IL-7R идентифицируют посредством инкубации кандидатного агента с IL-7R и мониторинга связывания и сопутствующего ингибирования фосфорилирования STAT5.
После первоначальной идентификации активность кандидатного антагонистического антитела к IL-7R можно дополнительно подтвердить и оценить с помощью биоанализов, известных для проверки целевой биологической активности. Кроме того, биоанализы могут быть непосредственно использованы для скрининга кандидатов. Некоторые из этих способов идентификации и характеристики антагонистических антител к IL-7R подробно описаны в Примерах.
Антагонистические антитела к IL-7R могут быть охарактеризованы с использованием способов, хорошо известных в данной области техники. Например, один способ заключается в идентификации эпитопа, с которым оно связывается, или "картировании эпитопов". Существует много способов, известных в данной области техники, для картирования и определения локализации эпитопов на белках, в том числе определение кристаллической структуры комплекса антитело-антиген, анализы конкуренции, анализы экспрессии фрагментов гена и анализы на основе синтетических пептидов, как описано, например, в Chapter 11 of Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999. В дополнительном примере картирование эпитопов можно использовать для определения последовательности, с которой связывается антагонистическое антитело к IL-7R. Картирование эпитопов является коммерчески доступным из различных источников, например Pepscan Systems (Edelhertweg 15, 8219 РН Lelystad, The Netherlands). Эпитоп может представлять собой линейный эпитоп, т.е. находящийся в одном участке аминокислот, или конформационный эпитоп, образующийся в результате трехмерного взаимодействия аминокислот, которые не обязательно должны содержаться в одном участке. Пептиды различной длины (например, с длиной по меньшей мере 4-6 аминокислот) можно выделять или синтезировать (например, рекомбинантно) и использовать для анализов связывания с антагонистическим антителом к IL-7R. В другом примере эпитоп, с которым связывается антагонистическое антитело к IL-7R, можно определить при систематическом скрининге с использованием перекрывания пептидов, происходящих из последовательности IL-7R, и определения связывания с помощью антагонистического антитела к IL-7R. Согласно анализу экспрессии фрагментов гена, открытая рамки считывания, кодирующая IL-7R, фрагментируется либо случайно, либо с помощью специфических генетических конструкций, и определяется реакционная способность экспрессируемых фрагментов IL-7R с тестируемым антителом. Фрагменты гена, например, можно получить с помощью ПЦР и затем транскрибировать и транслировать in vitro в белок в присутствии радиоактивных аминокислот. Связывание антитела с радиоактивно меченными фрагментами L-7R затем определяют с помощью иммунопреципитации и гель-электрофореза. Некоторые эпитопы также могут быть идентифицированы с использованием больших библиотек случайных пептидных последовательностей, экспонируемых на поверхности фаговых частиц (фаговые библиотеки). Альтернативно, определенную библиотеку перекрывающихся пептидных фрагментов можно проверить на связывание с тестируемым антителом в простых анализах связывания. В дополнительном примере мутагенез антигенсвязывающего домена, эксперименты по обмену доменов и аланин-сканирующий мутагенез могут быть осуществлены для идентификации остатков, требующихся, достаточных и/или необходимых для связывания эпитопа. Например, эксперименты по обмену доменов могут быть осуществлены с использованием мутантного IL-7R, в котором различные фрагменты полипептида IL-7R были заменены (обменены) на последовательности из IL-7R других видов или близкородственных, но антигенно различных белков (таких как другой член семейства пропротеинконвертаз). По оценке связывания антитела с мутантным IL-7R можно определить важность конкретного IL-7R фрагмента для связывания с антителом.
Еще один способ, который может быть использован для характеристики антагонистического антитела к IL-7R, заключается в применении конкурентных анализов с другими антителами, которые, как известно, связываются с тем же антигеном, т.е. различными фрагментами на IL-7R, для определения того, связывается ли антагонистическое антитело к IL-7R с тем же эпитопом, что и другие антитела. Конкурентные анализы хорошо известны специалистам в данной области техники.
Экспрессирующий вектор может быть использован для прямой экспрессии антагонистического антитела к IL-7R. Специалист в данной области знаком с введением экспрессирующих векторов для получения экспрессии экзогенного белка in vivo. См., например, патенты США №№6436908; 6413942; и 6376471. Введение экспрессирующих векторов включает локальное или системное введение, в том числе инъекцию, пероральное введение, генную пушку или введение с помощью катетера и местное введение. В другом воплощении экспрессирующий вектор вводят непосредственно в симпатический ствол или ганглий или в коронарную артерию, предсердие, желудочек сердца или перикард.
Также можно использовать направленную доставку терапевтических композиций, содержащих экспрессирующий вектор или субгеномные полинуклеотиды. Способы рецептор-опосредованной доставки ДНК описаны, например, в Findeis et al., Trends Biotechnol., 1993, 11:202; Chiou et al., Gene Therapeutics: Methods And Applications Of Direct Gene Transfer, J.A.Wolff, ed., 1994; Wu et al., J.Biol. Chem., 1988, 263:621; Wu et al., J.Biol. Chem., 1994, 269:542; Zenke et al., Proc. Natl. Acad. Sci. USA, 1990, 87:3655; Wu et al., J.Biol. Chem., 1991, 266:338. Терапевтические композиции, содержащие полинуклеотид, вводят в диапазоне от примерно 100 нг до примерно 200 мг ДНК для местного введения согласно протоколу генной терапии. Также могут быть использованы диапазоны концентраций от примерно 500 нг до примерно 50 мг, от примерно 1 мкг до примерно 2 мг, от примерно 5 мкг до примерно 500 мкг и от примерно 20 мкг до примерно 100 мкг ДНК во время протокола генной терапии. Терапевтические полинуклеотиды и полипептиды можно доставлять с использованием носителей для генной доставки. Носители для генной доставки могут быть вирусного или невирусного происхождения (см. в основном Jolly, Cancer Gene Therapy, 1994, 1:51; Kimura, Human Gene Therapy, 1994, 5:845; Connelly, Human Gene Therapy, 1995, 1:185; и Kaplitt, Nature Genetics, 1994, 6:148). Экспрессию таких кодирующих последовательностей можно индуцировать с использованием эндогенных или гетерологичных промоторов млекопитающих. Экспрессия кодирующей последовательности может быть либо конститутивной, либо регулируемой.
Векторы на основе вирусов для доставки желаемого полинуклеотида и экспрессии в желаемой клетке хорошо известны в данной области техники. Типичные основанные на вирусах векторы включают, но не ограничиваются этим, рекомбинантные ретровирусы (см., например, РСТ публикацию №№WO 90/07936; WO 94/03622; WO 93/25698; WO 93/25234; WO 93/11230; WO 93/10218; WO 91/02805; патенты США №№5219740 и 4777127; патент Великобритании №2200651; и ЕР патент №0345242), векторы на основе альфавирусов (например, векторы на основе вируса Синдбис, вируса леса Семлики (АТСС VR-67; АТСС VR-1247), вируса Росс-Ривер (АТСС VR-373; АТСС VR-1246) и вируса Венесуэльского конского энцефалита (АТСС VR-923; АТСС VR-1250; АТСС VR 1249; АТСС VR-532)) и аденоассоциированные вирусные векторы (AAV) (см., например, РСТ публикацию №№WO 94/12649, WO 93/03769; WO 93/19191; WO 94/28938; WO 95/11984 и WO 95/00655). Также можно использовать введение ДНК, связанной с убитым аденовирусом, как описано в Curiel, Hum. Gene Ther., 1992, 3:147.
Также могут быть использованы невирусные векторы и способы доставки, включая, но не ограничиваясь этим, поликатионную конденсированную ДНК, связанную или несвязанную только с убитым аденовирусом (см., например, Curiel, Hum. Gene Ther., 1992, 3:147); лиганд-связанную ДНК (см., например, Wu, J.Biol. Chem., 1989, 264:16985); носители для доставки в эукариотические клетки (см., например, патент США №5814482; РСТ публикацию №№WO 95/07994; WO 96/17072; WO 95/30763; и WO 97/42338) и нейтрализацию заряда нуклеиновой кислоты или слияние с клеточными мембранами. Также можно использовать "голую" ДНК. Типичные методы введения "голой" ДНК описаны в РСТ публикации №WO 90/11092 и патенте США №5580859. Липосомы, которые могут действовать в качестве носителей для генной доставки, описаны в патенте США №5422120; РСТ публикациях №№WO 95/13796; WO 94/23697; WO 91/14445; и ЕР 0524968. Дополнительные подходы описаны в Philip, Mol. Cell Biol., 1994, 14:2411 и в Woffendin, Proc. Natl. Acad. Sci., 1994, 91:1581.
В некоторых воплощениях изобретение охватывает композиции, включая фармацевтические композиции, содержащие антитела, описанные в данной заявке или полученные способами и имеющие характеристики, описанные в данной заявке. Используемые в данной заявке композиции содержат одно или более антител, которые антагонизируют взаимодействие IL-7R с IL-7, и/или один или более полинуклеотидов, содержащих последовательности, кодирующие одно или более этих антител. Эти композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципиенты, включающие буферы, которые хорошо известны в данной области техники.
Антагонистические антитела к IL-7R по изобретению характеризуются с помощью любой (одной или более) из следующих характеристик: (а) связывание с IL-7R; (б) блокирование взаимодействия IL-7R с IL-7; (в) блокирование или снижение IL-7-опосредованного фосфорилирования STAT5; (г) снижение уровней глюкозы в крови in vivo; (д) повышение толерантности к глюкозе in vivo; (е) уменьшение тяжести заболевания при ЕАЕ. Предпочтительно антагонистические антитела к IL-7R имеют два или более этих признаков. Более предпочтительно антитела имеют три или более признаков. Более предпочтительно антитела имеют четыре или более признаков. Более предпочтительно антитела имеют пять или более признаков. Наиболее предпочтительно антитела имеют все шесть характеристик.
Соответственно, в изобретении предложено любое из следующего, или композиции (включая фармацевтические композиции), содержащие любое из следующего: (а) антитело, имеющее частичную последовательность легкой цепи:
NFMLTQPHSVSGSPGKTVTISCTRSSGSIDSSYVQWYQQRPGNSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLVTEDEADYYCQSYDSSHLVFGGGTKLTVLC (SEQ ID NO: 1),
NFMLTQPHSVSESPGKTVTISCTGSSGRIASSYVQWYQQRPGSAPTTVIYEDNQRP SGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYASSSLWVFGGGTQLTVLS (SEQ ID NO: 3),
NFMLTQPHSVSGSPGKTVTISCTRSSGSIDSSYVQVVYQQRPGNSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLVTEDEADYYCMQYDSSHLVFGGGTKLTVLC (SEQ ID NO: 5),
NFMLTQPHSVSGSPGKTVTISCTRSSGSIDSSYVQWYQQRPGNSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLVTEDEADYYCQSYDFHHLVFGGGTKLTVLC (SEQ ID NO: 7),
NFMLTQPHSVSGSPGKTVTISCTRSSGSIDSSYVQVVYQQRPGNSPTTVIYEDDQRP
SGVPDRFSGSIDSSSNSASLTISGLVTEDEADYYCQSYDFHHLVFGGGTKLTVLC (SEQ ID NO: 9),
NFMLTQPHSVSGSPGKTVTISCTRSSGSIDSSYVQWYQQRPGNSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLVTEDEADYYCMQYDFHHLVFGGGTKLTVLC (SEQ ID NO: 11),
NFMLTQPHSVSESPGKTVTISCTRSSGSIDSSYVQWYQQRPGSSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCMQYDFHHLVFGGGTKLTVL (SEQ ID NO: 44) или
NFMLTQPHSVSESPGKTVTISCTRSSGSIDSSYVQVVYQQRPGSSPTTVIYEDDQRP SGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDFHHLVFGGGTKLTVL (SEQ ID NO: 41);
и (б) антитело, имеющее частичную последовательность тяжелой цепи:
QVNLRESGGGLVKPGGSLRLSCAASGFTFDDSVMHVWRQAPGKGLEWLSLVGWD GSATYYADSVKGRFTISRDNTKNLLYLQMNSLRAEDTAVYYCARQGDYVFDYWGQ GTLVTVSS (SEQ ID NO: 2),
QVTLKESGGGLVQPGGSLRLSCAASGFTFSNYGMHWVRQAPGKGLEWVSAISGS GGSTYYADSVKGRFTISRDNSKNTVYLQMNSLRDEDTAVYYCARDISGGGMDVWG QGTTVTVSS (SEQ ID NO: 4),
QVNLRESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWLSLVGWD GFFTYYADSVKGRFTISRDNTKNLLYLQMNSLRAEDTAVYYCARQGDYVFNNWGQ GTLVTVSS (SEQ ID NO: 6),
QVNLRESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWLSLVGWD GFFTYYADSVKGRFTISRDNTKNLLYLQMNSLRAEDTAVYYCARQGDYMGDYWGQ GTLVTVSS (SEQ ID NO: 8),
QVNLRESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWLSLVGWD GFFTYYADSVKGRFTISRDNTKNLLYLQMNSLRAEDTAVYYCARQGDYMGNNWGQ GTLVTVSS (SEQ ID NO: 10),
QVNLRESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWLSLVGWD GFFTYYADSVKGRFTISRDNTKNLLYLQMNSLRAEDTAVYYCARQGDYMGNNWGQ GTLVTVSS (SEQ ID NO: 12) или
EVQLVESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWVSLVGWD GFFTYYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARQGDYMGNNWG QGTLVTVSS (SEQ ID NO: 40).
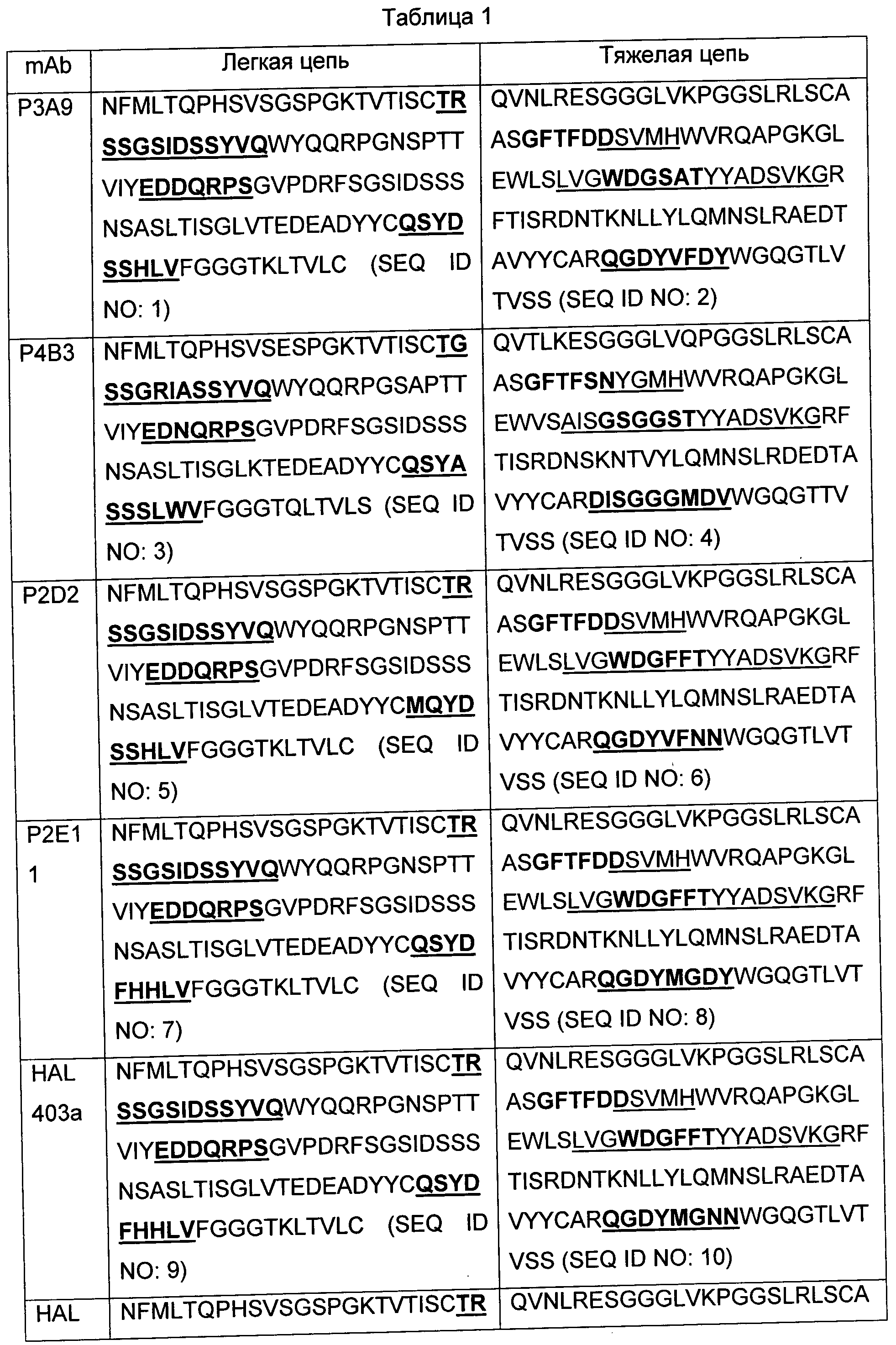

В Таблице 1 подчеркнутые последовательности представляют собой последовательности CDR в соответствии с Kabat, а выделенные жирным шрифтом в соответствии с Chothia.
В изобретении также предложены CDR-участки антител к IL-7R (включая CDRs Chothia, Kabat и CDR-контактирующие области). Определение CDR участков находится в пределах компетентности в данной области техники. Понятно, что в некоторых воплощениях CDRs могут представлять собой комбинацию CDR по Kabat и Chothia (также называемую "комбинированными CDRs" или "расширенными CDRs"). В некоторых воплощениях CDRs представляют собой Kabat CDRs. В других воплощениях CDRs представляют собой CDRs Chothia. Другими словами, в воплощениях с более чем одним CDR CDRs могут быть любыми из CDRs по Kabat, Chothia, их комбинации или комбинаций. В Таблице 2 показаны примеры последовательностей CDR, представленных в данной заявке.
|
CDR-контактирующие области представляют собой области антитела, которые наделяют антитело специфичностью к антигену. В целом, CDR-контактирующие области включают положения остатков в CDRs и зонах Вернье (Vernier), которые ограничены с целью поддержания правильной структуры петли, позволяющей антителу связываться с антигеном. См., например, Makabe et al., 2007, "Thermodynamic Consequences of Mutations in Vernier Zone Residues of a Humanized Anti-human Epidermal Growth Factor Receptor Murine Antibody," Journal of Biological Chemistry, 283:1156-1166. Определение CDR-контактирующих областей известно в данной области техники. В некоторых воплощениях антагонистическое антитело к IL-7R содержит одну или более CDR-контактирующих областей, содержащих аминокислотную последовательность, выбранную из группы, состоящей из FTFDDSVM (SEQ ID NO: 56), GWDGFF (SEQ ID NO: 57), ARX1X2X3X4, где X1, X2, X3 и X4 может быть любой аминокислотой (SEQ ID NO: 58), SGSIDSSY (SEQ ID NO: 59), EDDQRPSGV (SEQ ID NO: 60) и FHHL (SEQ ID NO: 61).
Аффинность связывания (KD) антагонистического антитела к IL-7R с IL-7R может составлять от примерно 0,002 до примерно 200 нМ. В некоторых воплощениях аффинность связывания является любой из примерно 200 нМ, 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ, примерно 60 пМ, примерно 50 пМ, примерно 20 пМ, примерно 15 пМ, примерно 10 пМ, примерно 5 пМ или примерно 2 пМ. В некоторых воплощениях аффинность связывания менее чем любая из примерно 250 нМ, примерно 200 нМ, примерно 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ или примерно 50 пМ.
В изобретении также предложены способы получения любого из этих антител. Антитела по данному изобретению могут быть получены с помощью процедур, известных в данной области техники. Полипептиды могут быть получены с помощью протеолитической или другой деградации антител, рекомбинантными методами (т.е. единичные или слитые полипептиды), как описано выше, или посредством химического синтеза. Полипептиды антител, особенно более короткие полипептиды вплоть до примерно 50 аминокислот, обычно получают посредством химического синтеза. Способы химического синтеза известны в данной области техники и коммерчески доступны. Например, антитело может быть получено с помощью автоматизированного полипептидного синтезатора, использующего твердофазный метод. См. также патенты США №№5807715; 4816567; и 6331415.
В другом альтернативном варианте антитела могут быть получены рекомбинантно с использованием процедур, хорошо известных в данной области техники. В одном воплощении полинуклеотид содержит последовательность, кодирующую вариабельные области тяжелой цепи и/или легкой цепи антитела Р3А9, Р4В3, P2D2, Р2Е11, HAL403a, HAL403b, C1GM или С2М3. Последовательность, кодирующую интересующее антитело, можно поддерживать в векторе в клетке-хозяине, которую можно нарастить и заморозить для будущего применения. Векторы (включая экспрессирующие векторы) и клетки-хозяева дополнительно описаны в данной заявке.
Изобретение также охватывает scFv антител по данному изобретению. Одноцепочечные фрагменты вариабельной области получают путем связывания вариабельных областей легкой и/или тяжелой цепи с использованием короткого связывающего пептида (Bird et al., 1988, Science 242:423-426). Примером связывающего пептида является (GGGGS)3 (SEQ ID NO: 13), мостики которого составляют приблизительно 3,5 нм между карбоксильным концом одной вариабельной области и аминоконцом другой вариабельной области. Линкеры из других последовательностей были сконструированы и использованы (Bird et al., 1988, выше). Линкеры должны быть короткими, гибкими полипептидами и предпочтительно содержать менее чем примерно 20 аминокислотных остатков. Линкеры, в свою очередь, могут быть модифицированы для дополнительных функций, таких как присоединение лекарственных средств или присоединение к твердым подложкам.
Одноцепочечные варианты могут быть получены либо рекомбинантно, либо синтетически. Для синтетического получения scFv может быть использован автоматизированный синтезатор. Для рекомбинантного получения scFv подходящую плазмиду, содержащую полинуклеотид, который кодирует scFv, можно вводить в подходящую клетку-хозяина, либо эукариотическую, такую как клетки дрожжей, растений, насекомых или млекопитающих, либо прокариотическую, такую как Е. coli. Полинуклеотиды, кодирующие интересующий scFv, могут быть получены рутинными манипуляциями, такими как лигирование полинуклеотидов. Полученные scFv можно выделить с использованием стандартных методов очистки белка, известных в данной области техники.
Также включены другие формы одноцепочечных антител, такие как диатела. Диатела являются двухвалентными биспецифическими антителами, в которых вариабельные домены тяжелой цепи (VH) и вариабельные домены легкой цепи (VL) экспрессируются на одной полипептидной цепи, но с использованием линкера, который является слишком коротким для того, чтобы обеспечить спаривание между двумя доменами на одной и той же цепи, посредством этого заставляя домены спариваться с комплементарными доменами другой цепи и создавая два антигенсвязывающих сайта (см., например, Holliger, P., et al., 1993, Ргос.Natl. Acad Sci. USA 90:6444-6448; Poljak, R. J., et al., 1994, Structure 2:1121-1123).
Например, биспецифические антитела, моноклональные антитела, которые обладают специфичностями связывания по меньшей мере с двумя различными антигенами, могут быть получены с использованием антител, раскрытых в данной заявке. Способы получения биспецифических антител известны в данной области техники (см., например, Suresh et al., 1986, Methods in Enzymology 121:210). По традиции рекомбинантное получение биспецифических антител основано на коэкспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулина с двумя тяжелыми цепями, имеющими различную специфичность (Millstein and Cuello, 1983, Nature 305, 537-539).
Согласно одному подходу для получения биспецифических антител вариабельные домены антитела с желаемыми специфичностями связывания (сайты объединения антитело-антиген) подвергают слиянию с последовательностями константной области иммуноглобулина. Слияние осуществляют предпочтительно с константной областью тяжелой цепи иммуноглобулина, содержащей по меньшей мере часть шарнира, СН2- и СН3-области. Предпочтительно иметь первую константную область тяжелой цепи (СН1), содержащую сайт, необходимый для связывания легкой цепи, присутствующий по меньшей мере в одном из слияний. ДНК, кодирующую слияния тяжелой цепи иммуноглобулина и, при необходимости, легкой цепи иммуноглобулина, вставляют в отдельные экспрессирующие векторы и котрансфицируют в подходящий организм-хозяин. Это обеспечивает большую гибкость в корректировке взаимных соотношений трех полипептидных фрагментов в воплощениях, когда неравные соотношения трех полипептидных цепей, используемых в конструкции, обеспечивают оптимальный выход. Однако можно было вставлять кодирующие последовательности для двух или всех трех полипептидных цепей в одном экспрессирующем векторе, когда экспрессия по меньшей мере двух полипептидных цепей в равном соотношении приводит к высоким выходам или когда соотношения не имеют особого значения.
В одном подходе биспецифические антитела состоят из тяжелой цепи гибридного иммуноглобулина с первой связывающей специфичностью в одном плече и пары тяжелая цепь-легкая цепь гибридного иммуноглобулина (обеспечивающей вторую связывающую специфичность) в другом плече. Эта асимметричная структура с легкой цепью иммуноглобулина только в одной половине биспецифической молекулы способствует отделению желаемого биспецифического соединения от нежелательных комбинаций иммуноглобулиновых цепей. Этот подход описан в РСТ публикации №WO 94/04690.
Гетероконъюгатные антитела, содержащие два ковалентно связанных антитела, также находятся в рамках изобретения. Такие антитела были использованы для нацеливания клеток иммунной системы на нежелательные клетки (патент США №4676980) и для лечения ВИЧ-инфекции (РСТ публикации №№WO 91/00360 и WO 92/200373; ЕР 03089). Гетероконъюгатные антитела могут быть получены с использованием любого удобного метода перекрестного сшивания. Подходящие агенты для перекрестного сшивания и методы хорошо известны в данной области техники и описаны в патенте США №4676980.
Химерные или гибридные антитела также могут быть получены in vitro с использованием известных методов синтетической белковой химии, включая методы, вовлекающие перекрестно-сшивающие агенты. Например, иммунотоксины могут быть сконструированы с использованием реакции дисульфидного обмена или образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат.
Гуманизированные антитела могут быть получены с использованием любых методов, известных в данной области техники. Например, четыре основных стадии могут быть использованы для гуманизации моноклонального антитела. Они представляют собой: (1) определение нуклеотидной и предсказанной аминокислотной последовательности исходных вариабельных доменов легкой и тяжелой цепи антитела, (2) конструирование гуманизированного антитела, т.е. принятие решения, какой каркасный участок антитела использовать в процессе гуманизации, (3) существующие в настоящее время методологии/методы гуманизации и (4) трансфекцию и экспрессию гуманизированного антитела. См., например, патенты США №№4816567; 5807715; 5866692; 6331415; 5530101; 5693761; 5693762; 5585089; и 6180370.
В рекомбинантных гуманизированных антителах Fcγ-участок может быть модифицирован для того, чтобы избежать взаимодействия с Fcγ-рецептором и комплементом и иммунными системами. Методы получения таких антител описаны в WO 99/58572. Например, константную область можно конструировать для большего сходства с человеческими константными областями для того, чтобы избежать иммунного ответа, если антитело используют в клинических испытаниях и лечении людей. См., например, патенты США №№5997867 и 5866692.
Изобретение охватывает модификации в антителах и полипептидах вариантов по изобретению, представленных в Таблице 1, включая функционально эквивалентные антитела, которые существенно не влияют на их свойства, и варианты, которые имеют повышенную или пониженную активность и/или аффинность. Например, аминокислотная последовательность может быть мутирована для получения антитела с желательной аффинностью связывания с IL-7R. Модификация полипептидов представляет собой обычную практику в данной области и не обязательно должна быть подробно описана здесь. Примеры модифицированных полипептидов включают полипептиды с консервативными заменами аминокислотных остатков, одной или более делециями или добавлениями аминокислот, которые не приводят к существенному вредному изменению функциональной активности или которые приводят к созреванию (увеличению) аффинности полипептида к его лиганду, или применение химических аналогов.
Инсерции аминокислотной последовательности включают амино- и/или карбокси-концевые слияния, варьирующиеся по длине от одного остатка до полипептидов, содержащих сто или более остатков, а также инсерции внутри последовательности из одного или множества аминокислотных остатков. Примеры концевых инсерции включают антитело с N-концевым метионильным остатком или антитело, слитое с эпитопной меткой. Другие инсерционные варианты молекулы антитела включают слияние с N- или С-концом антитела фермента или полипептида, который увеличивает время полужизни антитела в кровотоке.
Варианты замен имеют по меньшей мере один удаленный аминокислотный остаток в молекуле антитела и другой остаток, вставленный на его место. Сайты, представляющие наибольший интерес в отношении заместительного мутагенеза, включают гипервариабельные участки, но рассматриваются также изменения в FR. Консервативные замены показаны в Таблице 3 под заголовком "консервативные замены". Если такие замены приводят к изменению биологической активности, то более существенные изменения, названные "типичными заменами" в Таблице 3 или дополнительно описанные ниже со ссылкой на классы аминокислот, могут быть введены и продукты подвергнуты скринингу.
|
Существенные модификации в биологических свойствах антитела осуществляются путем выбора замен, которые значительно отличаются по своему эффекту на поддержание: (а) структуры полипептидного остова в области замены, например, в виде складчатой или спиральной конформации, (б) заряда или гидрофобности молекулы в целевом сайте или (в) большей части боковой цепи. Встречающиеся в природе остатки делятся на группы на основании общих свойств боковой цепи:
(1) Неполярные: Норлейцин, Met, Ala, Val, Leu, lie;
(2) Полярные незаряженные: Cys, Ser, Thr, Asn, Gin;
(3) Кислотные (отрицательно заряженные): Asp, Glu;
(4) Щелочные (положительно заряженные): Lys, Arg;
(5) Остатки, которые влияют на ориентацию цепи: Gly, Pro; и
(6) Ароматические: Trp, Tyr, Phe, His.
Неконсервативные замены получают посредством замены члена одного из этих классов на другой класс.
Любой цистеиновый остаток, не вовлеченный в поддержание правильной конформации антитела, также может быть замещен, как правило, серином для улучшения окислительной стабильности молекулы и предотвращения аберрантного сшивания. Напротив, цистеиновая(ые) связь(и) могут быть добавлены к антителу для улучшения его стабильности, особенно там, где антитело представляет собой фрагмент антитела, такой как Fv-фрагмент.
Аминокислотные модификации могут варьироваться от изменения или модификации одной или более аминокислот до полной реконструкции области, такой как вариабельная область. Изменения в вариабельной области могут изменить аффинность связывания и/или специфичность. В некоторых воплощениях не более чем 1-5 консервативных аминокислотных замен делают в пределах CDR-домена. В других воплощениях не более чем 1-3 консервативных аминокислотных замен делают в пределах CDR-домена. В еще одном воплощении CDR-домен представляет собой CDR НЗ и/или CDR L3.
Модификации также включают гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, гликозилирование различными сахарами, ацетилирование и фосфорилирование. Антитела гликозилированы в консервативных положениях в их константных областях (Jefferis and Lund, 1997, Chem. Immunol. 65:111-128; Wright and Morrison, 1997, TibTECH 15:26-32). Олигосахаридные боковые цепи иммуноглобулинов влияют на функцию белка (Boyd et al., 1996, Mol.Immunol. 32:1311-1318; Wittwe and Howard, 1990, Biochem. 29:4175-4180) и внутримолекулярное взаимодействие между частями гликопротеина, которые могут влиять на конформацию и представленную трехмерную поверхность гликопротеина (Jefferis and Lund, выше; Wyss and Wagner, 1996, Current Opin. Biotech. 7:409-416). Олигосахариды также могут служить для нацеливания данного гликопротеина на определенные молекулы на основании специфических структур распознавания. Также сообщалось, что гликозилирование антител влияет на антитело-зависимую клеточную цитотоксичность (ADCC). В частности сообщалось, что клетки СНО с тетрациклин-регулируемой экспрессией (3(1,4)-N-ацетилглюкозаминтрансферазы III (GnTIII), гликозилтрансфераза-катализируемым образованием разделенного GlcNAc, улучшало ADCC активность (Umana et al., 1999, Mature Biotech. 17:176-180).
Гликозилирование антител является, как правило, либо N-связанным, либо О-связанным. N-связанный относится к присоединению углеводной группировки к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин, аспарагин-Х-треонин и аспарагин-Х-цистеин, где X представляет собой любую аминокислоту за исключением пролина, представляют собой последовательности распознавания для ферментного присоединения углеводной группировки к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. О-связанное гликозилирование относится к присоединению одного из сахаров N-ацетилгалактозамина, галактозы или ксилозы к гидроксиаминокислоте, чаще всего к серину или треонину, хотя 5-гидроксипролин или 5-гидроксилизин также могут быть использованы.
Добавление сайтов гликозилирования к антителу обычно осуществляется путем изменения аминокислотной последовательности, такой, которая содержит один или более вышеописанных трипептидных последовательностей (для N-связанных сайтов гликозилирования). Изменение также может быть сделано путем добавления или замены одного или более остатков серина или треонина в последовательности исходного антитела (для О-связанных сайтов гликозилирования).
Картина гликозилирования антител также может быть изменена без изменения основной нуклеотидной последовательности. Гликозилирование в значительной степени зависит от клетки-хозяина, используемой для экспрессии антитела. Поскольку тип клетки, используемой для экспрессии рекомбинантных гликопротеинов, например антител как потенциальных терапевтических агентов, редко представляет собой нативную клетку, можно ожидать вариаций в картине гликозилирования антител (см., например Hse et al., 1997, J.Biol. Chem. 272:9062-9070).
В дополнение к выбору клеток-хозяев факторы, влияющие на гликозилирование во время рекомбинантного получения антител, включают режим роста, состав сред, плотность культуры, оксигенацию, рН, схемы очистки и тому подобное. Были предложены различные способы изменения картины гликозилирования, достигаемой в конкретном организме-хозяине, включая введение или сверхэкспрессию (патенты США №№5047335; 5510261 и 5278299). Гликозилирование или определенные типы гликозилирования можно ферментативно удалить из гликопротеина, например, с использованием эндогликозидазы Н (Endo Н), N-гликозидазы F, эндогликозидазы F1, эндогликозидазы F2, эндогликозидазы F3. Кроме того, рекомбинантная клетка-хозяин может быть генетически сконструирована таким образом, чтобы быть дефектной при процессинге определенных типов полисахаридов. Эти и сходные методы хорошо известны в данной области техники.
Другие способы модификации включают использование методов связывания, известных в данной области техники, включая, но не ограничиваясь этим, ферментативные способы, окислительное замещение и хелатирование. Модификации могут быть использованы, например, для присоединения меток для иммуноанализа. Модифицированные полипептиды получают с использованием хорошо известных в уровне техники методов и можно оценить с помощью стандартных анализов, известных в данной области техники, некоторые из которых описаны ниже и в Примерах.
В некоторых воплощениях изобретения антитело содержит модифицированную константную область, такую как константная область, которая имеет повышенную аффинность к человеческому Fc-гамма-рецептору, является иммунологически инертным или частично инертным, например не индуцирует комплемент-опосредованный лизис, не стимулирует антитело-зависимую клеточную цитотоксичность (ADCC) или не активирует микроглию; или имеет уменьшенную активность (по сравнению с немодифицированным антителом) в любом одном или более из следующего: запуск комплемент-опосредованного лизиса, стимуляция антитело-зависимой клеточной цитотоксичности (ADCC) или активация микроглии. Различные модификации константной области могут быть использованы для достижения оптимального уровня и/или комбинации эффекторных функций. См., например, Morgan et al., Immunology 86:319-324, 1995; Lund et al., J.Immunology 157:4963-9 157:4963-4969, 1996; Idusogie et al., J.Immunology 164:4178-4184, 2000; Tao et al., J.Immunology 143: 2595-2601, 1989; и Jefferis et al., Immunological Reviews 163:59-76, 1998. В некоторых воплощениях константная область модифицирована, как описано в Eur. J. Immunol., 1999, 29:2613-2624; РСТ заявка №PCT/GB99/01441; и/или заявка на патент Великобритании №9809951.8. В других воплощениях антитело содержит человеческую константную область тяжелой цепи IgG2, содержащую следующие мутации: А330Р331 в S330S331 (нумерация аминокислот относительно последовательности IgG2 дикого типа). Eur. J. Immunol., 1999, 29:2613-2624. В еще одном воплощении константная область является агликозилированной для N-связанного гликозилирования. В некоторых воплощениях константная область является агликозилированной для N-связанного гликозилирования путем мутирования гликозилированного аминокислотного остатка или фланкирующих остатков, которые являются частью последовательности распознавания N-гликозилирования в константной области. Например, сайт N-гликозилирования N297 может быть мутирован в А, Q, К или Н. См., Tao et al., J. Immunology 143: 2595-2601, 1989; и Jefferis et al., Immunological Reviews 163:59-76, 1998. В некоторых воплощениях константная область является агликозилированной для N-связанного гликозилирования. Константная область может быть агликозилирована ферментативно для N-связанного гликозилирования (такого как удаление углевода ферментом ПНГазой (PNGase)), или посредством экспрессии в дефектной по гликозилированию клетке-хозяине.
Другие модификации антитела включают антитела, которые были модифицированы, как описано в РСТ публикации №WO 99/58572. Эти антитела содержат в дополнение к связывающему домену, нацеленному на молекулу-мишень, эффекторный домен, имеющий аминокислотную последовательность, по существу гомологичную всей или части константной области тяжелой цепи иммуноглобулина человека. Эти антитела способны связываться с молекулой-мишенью без запуска значительного комплемент-зависимого лизиса или клеточно-опосредованного разрушения мишени. В некоторых воплощениях эффекторный домен способен специфически связываться с FcRn и/или FcyRllb. Они, как правило, основаны на химерных доменах, происходящих из двух или более СН2-доменов тяжелой цепи иммуноглобулина человека. Антитела, модифицированные таким образом, являются особенно подходящими для применения в постоянной терапии антителами для того, чтобы избежать воспалительных и других неблагоприятных реакций на традиционную терапию антителами.
Изобретение включает аффинно зрелые воплощения. Например, аффинно зрелые антитела могут быть получены с помощью процедур, известных в данной области техники (Marks et al., 1992, Bio/Technology, 10:779-783; Barbas et al., 1994, Proc Nat. Acad. Sci, USA 91:3809-3813; Schieretal., 1995, Gene, 169:147-155; Yelton et al., 1995, J.Immunol., 155:1994-2004; Jackson et al., 1995, J.Immunol., 154(7):3310-9; Hawkins et al., 1992, J.Mol.Biol., 226:889-896; и РСТ публикация №WO2004/058184).
Следующие способы могут быть использованы для регулирования аффинности антитела и для характеристики CDR. Один способ характеристики CDR антитела и/или изменения (такого как улучшение) аффинности связывания полипептида, такого как антитело, называется "библиотека-сканирующим мутагенезом". Как правило, библиотека-сканирующий мутагенез работает следующим образом. Одно или более аминокислотных положений в CDR заменяют двумя или более (такими как 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) аминокислотами с использованием принятых в данной области методов. Это создает небольшие библиотеки клонов (в некоторых воплощениях одну для каждого анализируемого аминокислотного положения), каждая с множеством из двух или более членов (если две или более аминокислот замещены в каждом положении). Как правило, библиотека также включает клон, содержащий нативную (незамещенную) аминокислоту. Небольшое число клонов, например примерно 20-80 клонов (в зависимости от сложности библиотеки), из каждой библиотеки подвергают скринингу в отношении аффинности связывания с целевым полипептидом (или другой связывающей мишенью) и идентифицируют кандидаты с увеличенным, таким же, уменьшенным связыванием или без связывания. Методы определения аффинности связывания хорошо известны в данной области техники. Аффинность связывания можно определить с помощью анализа поверхностного плазмонного резонанса с использованием Biacore™, который выявляет различия в аффинности связывания примерно 2-кратные или больше. Biacore™ является особенно полезным, когда исходное антитело уже связывается с относительно высокой аффинностью, например KD составляет примерно 10 нМ или ниже. Скрининг с использованием поверхностного плазмонного резонанса Biacore™ описан в Примерах в данной заявке.
Аффинность связывания можно определить с использованием Kinexa Biocensor, анализов сцинтилляционной близости, ELISA, иммуноанализа ORIGEN (IGEN), гашения флуоресценции, переноса флуоресценции и/или дрожжевого дисплея. Аффинность связывания также можно подвергнуть скринингу с использованием подходящего биоанализа.
В некоторых воплощениях каждое аминокислотное положение в CDR заменено (в некоторых воплощениях поочередно) всеми 20 природными аминокислотами с использованием принятых в данной области методов мутагенеза (некоторые из которых описаны в данной заявке). Это создает небольшие библиотеки клонов (в некоторых воплощениях один для каждого анализируемого аминокислотного положения), каждую с множеством из 20 членов (если все 20 аминокислот замещены в каждом положении).
В некоторых воплощениях библиотека, подлежащая скринингу, содержит замены в двух или более положениях, которые могут быть в том же CDR или в двух или более CDRs. Таким образом, библиотека может содержать замены в двух или более положениях в одном CDR. Библиотека может содержать замену в двух или более положениях в двух или более CDRs. Библиотека может содержать замену в 3, 4, 5 или более положениях, где указанные положения обнаружены в двух, трех, четырех, пяти или шести CDRs. Замену можно получить с использованием кодонов, характеризующихся низкой избыточностью. См., например, Таблицу 2 из Balint et al., 1993, Gene 137(1):109-18.
CDR может представлять собой CDRH3 и/или CDRL3. CDR может представлять собой один или более из CDRL1, CDRL2, CDRL3, CDRH1, CDRH2 и/или CDRH3. CDR может представлять собой CDR по Kabat, CDR по Chothia или расширенный CDR.
Кандидаты с улучшенным связыванием можно секвенировать, посредством этого идентифицируя мутант с заменой в CDR, который приводит к улучшенной аффинности (также названный "улучшенной" заменой). Кандидаты, которые связываются, также можно секвенировать, посредством этого идентифицируя замену в CDR, которая сохраняют связывание.
Может быть проведено несколько раундов скрининга. Например, кандидаты (каждый из которых содержит аминокислотную замену в одном или более положениях одного или более CDR) с улучшенным связыванием также полезны для конструирования второй библиотеки, содержащей по меньшей мере исходную и замещенную аминокислоту в каждом улучшенном положении в CDR (т.е. аминокислотное положение в CDR, в котором мутант с заменой показал улучшенное связывание). Получение, скрининг или селекция этой библиотеки обсуждается ниже.
Библиотека-сканирующий мутагенез также обеспечивает способы характеристики CDR, поскольку частота клонов с улучшенным связыванием, таким же связыванием, уменьшенным связыванием или без связывания также обеспечивает информацию, касающуюся важности каждого аминокислотного положения для стабильности антитело-антигенного комплекса. Например, если положение в CDR сохраняет связывание при замене на все 20 аминокислот, то такое положение идентифицируют как положение, которое, по-видимому, не требуется для связывания с антигеном. Напротив, если положение в CDR сохраняет связывание лишь в небольшом проценте замен, то такое положение идентифицируют как положение, которое является важным для функции CDR. Таким образом, методы библиотека-сканирующего мутагенеза дают информацию о положениях в CDRs, которые могут быть заменены множеством различных аминокислот (включая все 20 аминокислот), и положениях в CDRs, которые нельзя заменять или которые могут быть заменены только на некоторые аминокислоты.
Кандидаты с улучшенной аффинностью можно комбинировать во второй библиотеке, которая включает улучшенную аминокислоту, исходную аминокислоту в этом положении и может также включать дополнительные замены в этом положении в зависимости от желательной сложности библиотеки или позволяют использовать желаемый метод скрининга или селекции. В дополнение к этому при желании соседнее аминокислотное положение можно рандомизировать для по меньшей мере двух или более аминокислот. Рандомизация соседних аминокислот может позволить дополнительную конформационную гибкость в мутантном CDR, которая, в свою очередь, может позволить или облегчить введение большого количества улучшающих мутаций. Библиотека также может содержать замену в положениях, которые не показывают улучшенной аффинности в первом раунде скрининга.
Вторую библиотеку подвергают скринингу или селекции в отношении членов библиотеки с улучшенной и/или измененной аффинностью связывания с использованием любого метода, известного в данной области техники, включая скрининг с использованием анализа поверхностного плазмонного резонанса Biacore™ и селекции с использованием любого метода, известного в данной области техники для селекции, включая фаговый дисплей, дрожжевой дисплей и рибосомный дисплей.
Изобретение также охватывает слитые белки, содержащие один или более фрагментов или участков антител по данному изобретению. В одном воплощении предложен слитый полипептид, который содержит по меньшей мере 10 смежных аминокислот вариабельной области легкой цепи, представленной в SEQ ID NO: 1, 3, 5, 7, 9,11, 41 или 44, и/или по меньшей мере 10 аминокислот вариабельной области тяжелой цепи, представленной в SEQ ID NO: 2, 4, 6, 8, 10, 12 или 40. В других воплощениях предложен слитый полипептид, который содержит по меньшей мере примерно 10, по меньшей мере примерно 15, по меньшей мере примерно 20, по меньшей мере примерно 25 или по меньшей мере примерно 30 смежных аминокислот вариабельной области легкой цепи и/или по меньшей мере примерно 10, по меньшей мере примерно 15, по меньшей мере примерно 20, по меньшей мере примерно 25 или по меньшей мере примерно 30 смежных аминокислот вариабельной области тяжелой цепи. В другом воплощении слитый полипептид содержит вариабельную область легкой цепи и/или вариабельную область тяжелой цепи, как показано в любой из пар последовательностей, выбранных из SEQ ID NO: 1 и 2, 3 и 4, 5 и 6, 7 и 8, 9 и 10, 11 и 12, 41 и 40, и 44 и 40. В другом воплощении слитый полипептид содержит один или более CDR(s). В еще одном воплощении слитый полипептид содержит CDR Н3 (CDR3 VH) и/или CDR L3 (CDR3 VL). Для цели данного изобретения слитый белок содержит одно или более антител и другую аминокислотную последовательность, к которой он не присоединен в нативной молекуле, например гетерологичную последовательность или гомологичную последовательность из другой области. Типичные гетерологичные последовательности включают, но не ограничиваются этим, "метку", такую как FLAG-метка или 6His-метка. Метки хорошо известны в данной области техники.
Слитый полипептид можно создать с помощью методов, известных в данной области техники, например синтетически или рекомбинантно. Как правило, слитые белки по данному изобретению делают посредством получения экспрессии полинуклеотида, кодирующего их с использованием рекомбинантных методов, описанных в данной заявке, хотя они также могут быть получены другими способами, известными в данной области техники, включая, например, химический синтез.
В данном изобретении также предложены композиции, содержащие антитела, конъюгированные (например, связанные) с агентом, который облегчает связывание с твердой подложкой (таким как биотин или авидин). Для простоты обычно будут ссылаться на антитела, подразумевая, что эти способы применимы к любому из воплощений со связыванием IL-7R и/или антагонистических воплощений, описанных в данной заявке. Конъюгирование обычно относится к связыванию этих компонентов, как описано в данной заявке. Связывание (которое обычно представляет собой фиксирование этих компонентов в непосредственной ассоциации по меньшей мере для введения) может быть достигнуто различными способами. Например, прямая реакция между агентом и антителом возможна, когда каждый из них обладает заместителем, способным реагировать с другим заместителем. Например, нуклеофильная группа, такая как амино- или сульфгидрильная группа, может реагировать с карбонил-содержащей группой, такой как ангидрид или галогенангидрид, или с алкильной группой, содержащей хорошую уходящую группу (например, галогенид) на другой группе.
Антитело или полипептид по данному изобретению может быть связано с метящим агентом, таким как флуоресцентная молекула, радиоактивная молекула или любые другие метки, известные в данной области техники. В данной области техники известны метки, которые обычно обеспечивают (прямо или опосредованно) сигнал.
В изобретении также предложены композиции (включая фармацевтические композиции) и наборы, содержащие, как ясно из описания изобретения, любое или все антитела и/или полипептиды, описанные в данной заявке.
В изобретении также предложены выделенные полинуклеотиды, кодирующие антитела по изобретению, и векторы и клетки-хозяева, содержащие данный полинуклеотид.
Соответственно, в изобретении предложены полинуклеотиды (или композиции, в том числе фармацевтические композиции), содержащие полинуклеотиды, кодирующие любое из следующего: антитела C1GM, С2М3, Р3А9, Р4В3, P2D2, Р2Е11, HAL403a и HAL403b, или любой их фрагмент или участок, обладающий способностью антагонизировать IL-7R.
В другом аспекте в изобретении предложены полинуклеотиды, кодирующие любое из антител (включая фрагменты антитела) и полипептиды, описанные в данной заявке, такие как антитела и полипептиды, имеющие нарушенную эффекторную функцию. Полинуклеотиды можно получать и экспрессировать с помощью процедур, известных в данной области техники.
В другом аспекте в изобретении предложены композиции (такие как фармацевтические композиции), содержащие любой из полинуклеотидов по изобретению. В некоторых воплощениях композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий антитело, как описано в данной заявке. В другом воплощении композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий любое из антител, описанных в данной заявке. В еще одном воплощении композиция содержит один или оба полинуклеотида, представленных в SEQ ID NO: 38 и SEQ ID NO: 39 ниже:
Вариабельная область тяжелой цепи C1GM
GAGGTCCAGTTAGTGGAGTCTGGGGGAGGCCTGGTCAAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGATTCTGTCATGCACTGGGTCCGTCAAGCTCCGGGGMGGGTCTGGAGTGGGTTTCTCTTGTTGGTTGGGATGGTTTTTTTACATACTATGCAGACTCAGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCGAAGAACTCTCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCGAGACAAGGGGATTACATGGGGAACAACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA (SEQ ID NO: 38)
Вариабельная область легкой цепи C2GM
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAATCTCCGGGAAAGACGGTGACCATCTCCTGCACCCGCAGCAGTGGCAGCATTGACAGTTCCTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGCTCCCCCACCACTGTGATCTATGAGGATGACCAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAAACTGAGGACGAGGCTGACTACTACTGTCAGTCTTATGATTTTCATCATCTGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA (SEQ ID NO: 39).
В еще одном воплощении композиция содержит один или оба полинуклеотида, представленных в SEQ ID NO: 14 и SEQ ID NO: 15 ниже:
Вариабельная область тяжелой цепи HAL403a
CAGGTCAACTTAAGGGAGTCTGGGGGAGGCCTGGTCAAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGATTCTGTCATGCACTGGGTCCGTCAAGCTCCGGGGAAGGGTCTGGAGTGGCTCTCTCTTGTTGGTTGGGATGGTTTTTTTACATACTATGCAGACTCAGTGAAGGGCCGATTCACCATCTCCAGAGACAACACCAAGAACTTACTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCGAGACAAGGGGATTACATGGGGAACAAC TGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA (SEQ ID NO: 14)
Вариабельная область легкой цепи HAL403a
AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGGGTCTCCGGGAAAGACGGTGACCATCTCCTGCACCCGCAGCAGTGGCAGCATTGACAGTTCCTATGTGCAGTGGTACCAGCAGCGCCCGGGCAATTCCCCCACCACTGTGATCTATGAGGATGACCAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGGTGACTGAGGACGAGGCTGACTACTACTGTCAGTCTTATGATTTTCATCATCTGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTATGT (SEQ ID NO: 15).
Экспрессирующие векторы и введение полинуклеотидных композиций дополнительно описаны в данной заявке.
В другом аспекте в изобретении предложен способ получения любого из полинуклеотидов, описанных в данной заявке.
Настоящее изобретение также охватывает полинуклеотиды, комплементарные любым таким последовательностям. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут представлять собой молекулы ДНК (геномные, кДНК или синтетические) или РНК. Молекулы РНК включают молекулы гяРНК (гетерогенной ядерной РНК), которые содержат интроны и соответствуют молекуле ДНК в соотношении один к одному (in a one-to-one manner), и молекулы мРНК, которые не содержат интронов. Дополнительные кодирующие или некодирующие последовательности могут присутствовать, но не обязательно, в пределах полинуклеотида по настоящему изобретению, и полинуклеотид может быть связан, но не обязательно, с другими молекулами и/или подложками.
Полинуклеотиды могут содержать нативную последовательность (т.е. эндогенную последовательность, которая кодирует антитело или его участок) или может содержать вариант такой последовательности. Полинуклеотидные варианты содержат одну или более замен, добавлений, делеций и/или вставок, так что иммунореактивность кодируемого полипептида не уменьшается по сравнению с нативной иммунореактивной молекулой. Эффект на иммунореактивность кодируемого полипептида можно оценивать обычно, как описано в данной заявке. Варианты предпочтительно демонстрируют по меньшей мере примерно 70%-ную идентичность, более предпочтительно по меньшей мере примерно 80%-ную идентичность, еще более предпочтительно по меньшей мере примерно 90%-ную идентичность и наиболее предпочтительно по меньшей мере примерно 95%-ную идентичность с полинуклеотидной последовательностью, которая кодирует нативное антитело или его участок.
Две полинуклеотидные или полипептидные последовательности называют "идентичными", если последовательность нуклеотидов или аминокислот в двух последовательностях является одинаковой при выравнивании для максимального соответствия, как описано ниже. Сравнение между двумя последовательностями, как правило, осуществляют путем сравнения последовательностей в окне сравнения для идентификации и сравнения локальных участков сходства последовательностей. "Окно сравнения", как использовано в данной заявке, относится к сегменту по меньшей мере из примерно 20 смежных положений, обычно от 30 до примерно 75 или от 40 до примерно 50, в которых последовательность можно сравнивать с эталонной последовательностью из такого же числа смежных положений после оптимального выравнивания двух последовательностей.
Оптимальное выравнивание последовательностей для сравнения можно осуществлять с использованием программы Megalign в пакете программ по биоинформатике Lasergene (DNASTAR, Inc., Madison, WI), с использованием параметров по умолчанию. Эта программа включает несколько схем выравнивания, описанных в следующих ссылках: Dayhoff, М.О., 1978, A model of evolutionary change in proteins - Matrices for detecting distant relationships. В Dayhoff, M.O. (ed.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Washington DC Vol.5, Suppl. 3, pp.345-358; Hein J., 1990, Unified Approach to Alignment and Phylogenes pp.626-645 Methods in Enzymology vol. 183, Academic Press, Inc., San Diego, CA; Higgins, D.G. and Sharp, P.M., 1989, CABIOS 5:151-153; Myers, E.W. and Muller W., 1988, CABIOS 4:11-17; Robinson, E.D., 1971, Comb. Theor. 11:105; Santou, N., Nes, M., 1987, Mol. Biol. Evol. 4:406-425; Sneath, P.H.A. and Sokal, R.R., 1973, Numerical Taxonomy the Principles and Practice of Numerical Taxonomy, Freeman Press, San Francisco, CA; Wilbur, W.J. and Lipman, D.J., 1983, Proc. Natl. Acad. Sci. USA 80:726-730.
Предпочтительно "процент идентичности последовательностей" определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения по меньшей мере из 20 положений, где участок полинуклеотидной или полипептидной последовательности в окне сравнения может содержать добавления или делеции (т.е. пробелы) 20 процентов или меньше, обычно от 5 до 15 процентов, или от 10 до 12 процентов по сравнению с эталонными последовательностями (которые не содержат добавлений или делеции) для оптимального выравнивания этих двух последовательностей. Процент рассчитывают путем определения числа положений, в которых идентичные основания нуклеиновой кислоты или остатки аминокислот в обеих последовательностях дают число совпадающих положений, деля число совпадающих положений на общее число положений в эталонной последовательности (т.е. размер окна) и умножая результаты на 100 для получения процента идентичности последовательностей.
Варианты могут также или, альтернативно, являются по существу гомологичными нативному гену или его участку или комплементу. Такие полинуклеотидные варианты способны гибридизоваться в умеренно жестких условиях с природной последовательностью ДНК, кодирующей нативное антитело (или комплементарной последовательностью).
Подходящие "умеренно жесткие условия" включают предварительную промывку в растворе 5×SSC, 0,5% SDS, 1,0 мМ EDTA (рН 8,0); гибридизацию при 50°С-65°С, 5×SSC в течение ночи; с последующей двухкратной промывкой при 65°С в течение 20 минут каждым из 2×, 0,5× и 0,2×SSC, содержащим 0,1% SDS.
Используемые в данной заявке "жесткие условия" или "условия высокой жесткости" представляют собой условия, при которых: (1) используют низкую ионную силу и высокую температуру для промывки, например, 0,015 М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50°С; (2) используют в процессе гибридизации денатурирующий агент, такой как формамид, например, 50%-ный (об./об.) формамид с 0,1% бычьего сывороточного альбумина/0,1% Ficoll/0,1% поливинилпирролидона/50 мМ натрий-фосфатным буфером при рН 6,5 с 750 мМ хлорида натрия, 75 мМ цитрата натрия при 42°С; или (3) используют 50%-ный формамид, 5×SSC (0,75 М NaCl, 0,075 М цитрата натрия), 50 мМ фосфата натрия (рН 6,8), 0,1% пирофосфата натрия, 5× раствор Денхардта, обработанную ультразвуком ДНК спермы лосося (50 мкг/мл), 0,1% SDS и 10% декстрансульфата при 42°С с промывками при 42°С в 0,2×SSC (хлорид натрия/цитрат натрия) и 50% формамида при 55°С, с последующей промывкой в жестких условиях в растворе, состоящем из 0,1×SSC, содержащего EDTA, при 55°С. Специалист в данной области знает, как подобрать температуру, ионную силу и т.д. в соответствии с необходимостью соответствовать факторам, таким как длина пробы и тому подобное.
Специалисту в данной области понятно, что в результате вырожденности генетического кода существует множество нуклеотидных последовательностей, которые кодируют полипептид, как описано в данной заявке. Некоторые из этих полинуклеотидов имеют минимальную гомологию с нуклеотидной последовательностью нативного гена. Тем не менее, полинуклеотиды, которые варьируются из-за различий в использовании кодонов, специально рассмотрены в настоящем изобретении. Кроме того, аллели генов, содержащих полинуклеотидные последовательности, представленные здесь, находятся в рамках настоящего изобретения. Аллели представляют собой эндогенные гены, которые изменены в результате одной или более мутаций, таких как делеции, добавления и/или замены нуклеотидов. Полученная мРНК и белок может, но не обязательно, иметь измененную структуру или функцию. Аллели можно идентифицировать с использованием стандартных методов (таких как гибридизация, амплификация и/или сравнение последовательностей из базы данных).
Полинуклеотиды по данному изобретению могут быть получены с использованием химического синтеза, рекомбинантных методов или ПЦР. Методы химического синтеза пол и нуклеотидов хорошо известны в данной области техники и не обязательно должны подробно описываться здесь. Специалист в данной области может использовать последовательности, предложенные здесь, и коммерческий синтезатор ДНК для получения желаемой ДНК-последовательности.
Для получения полинуклеотидов с использованием рекомбинантных методов полинуклеотид, содержащий желаемую последовательность, можно вставлять в подходящий вектор, и вектор, в свою очередь, можно вводить в подходящую клетку-хозяина для репликации и амплификации, как дополнительно обсуждается здесь. Полинуклеотиды можно встраивать в клетки-хозяева любыми способами, известными в данной области техники. Клетки подвергают трансформации путем введения экзогенного полинуклеотида посредством прямого поглощения, эндоцитоза, трансфекции, F-спаривания или электропорации. После введения экзогенный полинуклеотид может поддерживаться в пределах клетки в виде неинтегрированного вектора (например, плазмиды) или интегрироваться в геном клетки-хозяина. Амплифицированный таким образом полинуклеотид можно выделить из клетки-хозяина способами, хорошо известными в данной области техники. См., например, Sambrook et al., 1989.
Альтернативно ПЦР позволяет воспроизводить последовательности ДНК. ПЦР-технология хорошо известна в данной области техники и описана в патентах США №№4683195, 4800159, 4754065 и 4683202, а также в PCR: The Polymerase Chain Reaction, Mullis et al. eds., Birkauswer Press, Boston, 1994.
РНК может быть получена с использованием выделенной ДНК в соответствующем векторе и вставки его в подходящую клетку-хозяина. Когда клетка реплицируется и ДНК транскрибируется в РНК, РНК может быть затем выделена с использованием способов, хорошо известных специалистам в данной области, как изложено в Sambrook et al., 1989, выше, например.
Подходящие клонирующие векторы могут быть сконструированы согласно стандартным методам или могут быть выбраны из большого числа клонирующих векторов, доступных в данной области. В то время как выбранный клонирующий вектор может варьироваться в соответствии с используемой клеткой-хозяином, полезные клонирующие векторы обычно будут способны самореплицироваться, могут обладать одной мишенью для конкретной рестриктазы и/или могут нести гены для маркера, который может быть использован в выбранных клонах, содержащих указанный вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например pUC18, pUC19, Bluescript (например, pBS SK+) и его производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговые ДНК и шаттл-векторы, такие как pSA3 и рАТ28. Эти и многие другие клонирующие векторы доступны от коммерческих поставщиков, таких как BioRad, Strategene и Invitrogen.
Экспрессирующие векторы обычно представляют собой реплицируемые полинуклеотидные конструкции, которые содержат полинуклеотид согласно изобретению. Подразумевается, что экспрессирующий вектор должен реплицироваться в клетке-хозяине либо в виде эписом, либо в виде интегральной части хромосомной ДНК. Подходящие экспрессирующие векторы включают, но не ограничиваются этим, плазмиды, вирусные векторы, включая аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды и экспрессирующий(е) вектор(ы), раскрытые в РСТ публикации №WO 87/04462. Векторные компоненты обычно могут включать, но не ограничиваются этим, одно или более из следующего: сигнальную последовательность; точку инициации репликации; один или более маркерных генов; подходящие транскрипционные контролирующие элементы (такие как промоторы, энхансеры и терминатор). Для экспрессии (т.е. трансляции) также обычно требуются один или более трансляционных контролирующих элементов, таких как сайты связывания рибосом, сайты инициации трансляции и стоп-кодоны.
Векторы, содержащие полинуклеотиды, представляющие интерес, можно вводить в клетку-хозяина любым из множества подходящих способов, включая электропорацию, трансфекцию с применением хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию; и инфекцию (например, когда вектор представляет собой инфекционный агент, такой как вирус осповакцины). Выбор вводимых векторов или полинуклеотидов часто зависят от признаков клетки-хозяина.
В изобретении также предложены клетки-хозяева, содержащие любые из полинуклеотидов, описанных в данной заявке. Любые клетки-хозяева, способные сверхэкспрессировать гетерологичные ДНК, могут быть использованы для выделения генов, кодирующих антитело, полипептид или белок, представляющие интерес. Неограничивающие примеры клеток-хозяев млекопитающих включают, но не ограничиваются этим, клетки COS, HeLa и СНО. См. также РСТ публикацию №WO 87/04462. Подходящие хозяйские клетки не млекопитающих включают прокариотов (таких как Е. coli или В. subtillis) и дрожжи (такие как S. cerevisae, S. pombe; или К. lactis). Предпочтительно клетки-хозяева экспрессируют кДНК на уровне примерно в 5 раз выше, более предпочтительно в 10 раз выше, еще более предпочтительно в 20 раз выше по сравнению с соответствующим эндогенным антителом или белком, представляющим интерес, если они присутствуют, в клетках-хозяевах. Скрининг клеток-хозяев в отношении специфического связывания с IL-7R или доменом IL-7R осуществляют с помощью иммуноанализа или FACS. Можно идентифицировать клетку, сверхэкспрессирующую антитело или белок, представляющий интерес.
Композиции
Композиции, используемые в способах по изобретению, содержат эффективное количество антагонистического антитела к IL-7R, полипептида, происходящего из антагонистического антитела к IL-7R, или других антагонистов IL-7R, описанных в данной заявке. Примеры таких композиций, а также ее приготовления в виде препаратов также описаны в предыдущем разделе и ниже. В некоторых воплощениях композиция содержит одно или более антагонистических антител к IL-7R. В других воплощениях антагонистическое антитело к IL-7R распознает человеческий IL-7Rα. В других воплощениях антагонистическое антитело к IL-7R представляет собой человеческое антитело. В других воплощениях антагонистическое антитело к IL-7R представляет собой гуманизированное антитело. В некоторых воплощениях антагонистическое антитело к IL-7R содержит константную область, которая способна запускать желательный иммунный ответ, такой как антитело-опосредованный лизис или ADCC. В других воплощениях антагонистическое антитело к IL-7R содержит константную область, которая не запускает ненужный или нежелательный иммунный ответ, такой как антитело-опосредованный лизис или ADCC. В других воплощениях антагонистическое антитело к IL-7R содержит один или более CDR(s) антитела (таких как один, два, три, четыре, пять или в некоторых воплощениях все шесть CDRs).
Понятно, что композиции могут содержать более чем одно антагонистическое антитело к IL-7R (например, смесь антагонистических антител к IL-7R, которые распознают различные эпитопы IL-7R). Другие типичные композиции содержат более чем одно антагонистическое антитело к IL-7R, которое распознает один и тот же эпитоп(ы), или различные виды антагонистических антител к IL-7R, которые связываются с различными эпитопами IL-7R.
Композиция, используемая в настоящем изобретении, может дополнительно содержать фармацевтически приемлемые носители, эксципиенты или стабилизаторы (Remington: The Science and practice of Pharmacy 20th Ed., 2000, Lippincott Williams and Wilkins, Ed. К. E. Hoover) в форме лиофилизированных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в применяемых дозировках и концентрациях и могут содержать буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид, бензэтония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин, резорцин, циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстраны; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG). Фармацевтически приемлемые эксципиенты дополнительно описаны в данной заявке.
Антагонистическое антитело к IL-7R и его композиции также могут быть использованы в сочетании с другими агентами, которые служат для улучшения и/или дополнения эффективности агентов.
Г. Наборы
В изобретении также предложены наборы для применения в экспресс-методах. Наборы по изобретению включают один или более контейнеров, содержащих антагонист IL-7R (таких как, например, человеческое антитело), описанный в данной заявке, и инструкции по применению согласно любому из способов по изобретению, описанных в данной заявке. Как правило, эти инструкции содержат описание введения антагониста IL-7R для выше описанного терапевтического лечения.
В некоторых воплощениях антагонист IL-7R представляет собой антагонистическое антитело к IL-7R. В некоторых воплощениях антитело представляет собой человеческое антитело. В некоторых воплощениях антитело представляет собой гуманизированное антитело. В некоторых воплощениях антитело представляет собой моноклональное антитело. Инструкции, относящиеся к применению антагонистического антитела к IL-7R, как правило, включают информацию о дозировке, схеме введения и пути введения для предполагаемого лечения. Контейнеры могут представлять собой однократные дозы, нефасованные упаковками (например, мультидозовые упаковками) или субдозы. Инструкции, поставляемые в наборах по изобретению, обычно представляют собой инструкции на этикетке или внутреннем вкладыше (например, листе бумаги, входящем в набор), но машиночитаемые инструкции (например, инструкции, выполненные на магнитных или оптических дисках) также приемлемы.
Наборы по данному изобретению находятся в подходящей упаковке. Подходящие упаковки включают, но не ограничиваются этим, флаконы, бутылки, банки, гибкие упаковки (например, запечатанный майлар или пластиковые пакеты) и тому подобное. Рассматриваются также упаковки для применения в комбинации с конкретным устройством, таким как ингалятор, устройство для назального введения (например, атомайзер) или устройство для инфузии, такое как мини-насос. Набор может иметь стерильное входное отверстие (например, контейнер может представлять собой мешок для внутривенного раствора или флакон, имеющий пробку, прокалываемую иглой для подкожных инъекций). Контейнер также может иметь стерильное входное отверстие (например, контейнер может быть мешком для внутривенного раствора или флаконом, имеющим пробку, прокалываемую иглой для подкожных инъекций). По меньшей мере один активный агент в композиции представляет собой антагонистическое антитело к IL-7R. Контейнер может дополнительно содержать второй фармацевтически активный агент.
Наборы возможно могут обеспечивать дополнительные компоненты, такие как буферы и интерпретирующая информация. Как правило, набор содержит контейнер и этикетку или листок-вкладыш(и) на контейнере или прикрепленный(е) к контейнеру.
Мутации и модификации
Для экспрессии антител к IL-7R по настоящему изобретению ДНК-фрагменты, кодирующие VH- и VL-области, сначала могут быть получены с использованием любого из способов, описанных выше. Различные модификации, например мутации, делеции и/или добавления, также можно вводить в ДНК-последовательности с использованием стандартных методов, известных специалистам в данной области. Например, мутагенез можно осуществлять с использованием стандартных методов, таких как ПЦР-опосредованный мутагенез, при котором мутировавшие нуклеотиды встраиваются в ПЦР-праймеры таким образом, что ПЦР-продукт содержит желательные мутации, или сайт-направленный мутагенез.
Один тип замены, например который может быть осуществлен с целью замены одного или более цистеинов в антителе, который может быть химически реактивным, на другой остаток, такой как, без ограничения, аланин или серин. Например, может быть осуществлена замена неканонического цистеина. Замену можно осуществить в CDR или каркасном участке вариабельного домена или в константной области антитела. В некоторых воплощениях цистеин является каноническим.
Антитела могут быть также модифицированы, например, в вариабельных доменах тяжелой и/или легкой цепи, например, для изменения связывающего свойства антитела. Например, может быть осуществлена мутация в одном или более CDR участках для увеличения или уменьшения KD антитела к IL-7R, для увеличения или уменьшения koff или для изменения специфичности связывания антитела. Методы сайт-направленного мутагенеза хорошо известны в данной области техники. См., например, Sambrook et al. и Ausubel et al., выше.
Модификация или мутация также можно осуществить в каркасном участке или константной области для увеличения периода полувыведения антитела к IL-7R. См., например, РСТ публикацию №WO 00/09560. Мутацию в каркасном участке или константной области также можно осуществить для изменения иммуногенности антитела, для обеспечения сайта для ковалентного или нековалентного связывания с другой молекулой или для изменения таких свойств, как фиксация комплемента, связывание FcR и антитело-зависимая клеточно-опосредованная цитотоксичность. Согласно изобретению единичное антитело может иметь мутации в любом одном или более CDRs или каркасных участках вариабельного домена или в константной области.
В процессе, известном как "germlining" (возвращение к зародышевому генотипу), некоторые аминокислоты в VH- и VL-последовательностях могут быть мутированы для соответствия аминокислотам, обнаруженным в природе в VH- и VL-последовательностях зародышевой линии. В частности, аминокислотные последовательности каркасных участков в VH- и VL-последовательностях могут быть мутированы для того, чтобы соответствовать последовательностям зародышевой линии для уменьшения риска иммуногенности при введении антитела. ДНК-последовательности зародышевой линии для человеческих VH- и VL-генов известны в данной области техники (см., например, the "Vbase" human germline sequence database; см. также Kabat, E.A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, публикация NIH №91-3242; Tomlinson et al., 1992, J.Mol.Biol. 227:776-798; и Cox et al., 1994, Eur. J.Immunol. 24:827-836.
Другой тип аминокислотной замены, который может быть осуществлен, заключается в удалении потенциальных протеолитических сайтов в антителе. Такие сайты могут встречаться в CDR, или каркасном участке вариабельного домена, или в константной области антитела. Замена остатков цистеина и удаление протеолитических сайтов может уменьшить риск гетерогенности в антительном продукте и, таким образом, увеличить гомогенность. Другой тип аминокислотной замены заключается в элиминации аспарагин-глициновых пар, которые образуют потенциальные сайты дезамидирования путем изменения одного или обоих остатков. В другом примере С-концевой лизин тяжелой цепи антитела к IL-7R по изобретению может быть расщеплен. В различных воплощениях изобретения тяжелые и легкие цепи антител к IL-7R, возможно, могут включать сигнальную последовательность.
Как только фрагменты ДНК, кодирующие VH- и VL-сегменты по настоящему изобретению, получены, этими фрагментами ДНК можно дополнительно манипулировать с помощью стандартных методов рекомбинантных ДНК, например, для превращения генов вариабельной области в гены полноразмерных цепей антитела, гены Fab-фрагментов или в ген scFv. При этих манипуляциях VL- или VH-кодирующий ДНК-фрагмент функциональным образом связывают с другим ДНК-фрагментом, кодирующим другой белок, такой как константная область антитела или гибкий линкер. Термин "функциональным образом связанный", используемый в данном контексте, предназначен для обозначения того, что два ДНК-фрагмента соединены так, что аминокислотные последовательности, кодируемые двумя ДНК-фрагментами, остаются "в рамке".
Выделенную ДНК, кодирующую VH-область, можно превратить в полноразмерный ген тяжелой цепи, функциональным образом связывая VH-кодирующую ДНК с другой молекулой ДНК, кодирующей константные области тяжелой цепи (СН1, СН2 и СН3). Последовательности генов константной области тяжелой цепи человека известны в данной области техники (см., например, Kabat, Е. A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, публикация NIH №91-3242), и ДНК-фрагменты, охватывающие эти области, могут быть получены с помощью стандартной ПЦР-амплификации. Константная область тяжелой цепи может представлять собой константную область IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно представляет собой константную область IgG1 или IgG2. Последовательность константной области IgG может представлять собой любую из различных аллелей или аллотипов, которые, как известно, встречаются среди различных индивидуумов, таких как Gm(1), Gm(2), Gm(3) и Gm(17). Такие аллотипы представляют природные аминокислотные замены в константных областях IgG1. Для гена Fab-фрагмента тяжелой цепи VH-кодирующая ДНК может быть функциональным образом связана с другой молекулой ДНК, кодирующей только константную область СН1 тяжелой цепи. Константная область СН1 тяжелой цепи может быть получена из любого гена тяжелой цепи.
Выделенную ДНК, кодирующую VL-область, можно превратить в полноразмерный ген легкой цепи (а также ген Fab легкой цепи) посредством функционального связывания VL-кодирующей ДНК с другой молекулой ДНК, кодирующей константную область CL легкой цепи. Последовательности генов константной области легкой цепи человека известны в данной области техники (см., например, Kabat, Е.A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, публикация NIH №91-3242), и ДНК-фрагменты, охватывающие эти области, могут быть получены с помощью стандартной ПЦР-амплификации. Константная область легкой цепи может представлять собой константную область каппа или лямбда. Константная область каппа может представлять собой любую из различных аллелей, которые, как известно, встречаются среди различных индивидуумов, таких как lnv(1), lnv(2) и lnv(3). Константная область лямбда может быть получена из любого из трех генов лямбда.
Для создания гена scFv VH- и VL-кодирующие ДНК-фрагменты функциональным образом связывают с другим фрагментом, кодирующим гибкий линкер, например кодирующим аминокислотную последовательность (Gly4-Ser)3, (SEQ ID NO: 16), так что VH- и VL-последовательности могут экспрессироваться в виде непрерывного одноцепочечного белка с VL- и VH-областями, соединенными гибким линкером (см., например, Bird et al., 1988, Science 242:423-426; Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., 1990, Nature 348:552-554). Одноцепочечное антитело может быть моновалентным, если используют только одну VH и VL, двухвалентным, если используют две VH и VL, или поливалентным, если используют более чем две VH и VL. Могут быть получены биспецифические или поливалентные антитела, которые специфически связываются с IL-7R и с другой молекулой.
В другом воплощении можно получить слитое антитело или иммуноадгезин, которые содержат все или участок антитела к IL-7R по изобретению, связанное с другим полипептидом. В другом воплощении только вариабельные домены антитела к IL-7R соединены с полипептидом. В другом воплощении VH-домен антитела к IL-7R соединен с первым полипептидом, в то время как VL-домен антитела к IL-7R соединен со вторым полипептидом, который ассоциирован с первым полипептидом, так что VH- и VL-домены могут взаимодействовать друг с другом, образуя антигенсвязывающий сайт. В другом предпочтительном воплощении VH-домен отделен от VL-домена линкером, так что VH- и VL-домены могут взаимодействовать друг с другом. VH-линкер-VL антитело затем соединяют с полипептидом, представляющим интерес. Кроме того, можно создать слитые антитела, в которых два (или более) одноцепочечных антитела соединены друг с другом. Это полезно, если хотят создать двухвалентное или поливалентное антитело на одной полипептидной цепи или если хотят создать биспецифические антитело.
В других воплощениях другие модифицированные антитела могут быть получены с использованием антитела к IL-7R, кодируемого молекулами нуклеиновой кислотой. Например, "Каппа-антитела" (Kappa bodies) (III et al., 1997, Protein Eng. 10:949-57), "Мини-антитела" (Minibodies) (Martin et al., 1994, EMBO J. 13:5303-9), "Диатела" (Diabodies) (Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444-6448), или "Janusins" (Traunecker et al., 1991, EMBO J. 10:3655-3659 и Traunecker et al., 1992, Int. J. Cancer (Suppl.) 7:51-52) могут быть получены с использованием стандартных способов молекулярной биологии в соответствии с описанием.
Биспецифические антитела или антигенсвязывающие фрагменты могут быть получены различными способами, включающими слияние гибридом или соединение Fab'-фрагментов. См., например, Songsivilai & Lachmann, 1990, Clin. Exp.Immunol. 79:315-321, Kostelny et al., 1992, J. Immunol. 148:1547-1553. Кроме того, биспецифические антитела могут быть образованы в виде "диател" или "Janusins". В некоторых воплощениях биспецифическое антитело связывается с двумя различными эпитопами IL-7R. В некоторых воплощениях модифицированные антитела, описанные выше, получают с использованием одного или более вариабельных доменов или CDR участков из человеческого антитела к IL-7R, предложенного в данной заявке.
Получение антиген-специфических антител
Моноклональные антитела, полученные против рекомбинантной мышиной химеры IL-7Rα/CD127/Fc (R&D Systems, кат.№747-MR), и человеческие антитела, полученные посредством биопэннинга (сортировки) библиотеки человеческих "наивных" антител с рекомбинантным IL-7Rα, оценивают по их способности связываться с мышиным и человеческим IL-7R. Антитела подвергают дополнительному скринингу в отношении их способности блокировать IL-7-опосредованное фосфорилирование STAT5 в человеческих мононуклеарных клетках периферической крови (PBMCs) и/или PBMCs обезьяны. Этот способ получения антител давал антагонистические антитела, которые демонстрируют блокирование IL-7-опосредованного фосфорилирования STAT5, как показано в Примере 1.
Репрезентативные материалы настоящего изобретения были депонированы в Американской коллекции типовых культур (АТСС) 9 февраля 2011 г. Вектор C1GM-VH, имеющий № доступа в АТСС РТА-11679, представляет собой полинуклеотид, кодирующий вариабельную область тяжелой цепи C1GM, и вектор C1GM-VL, имеющий № доступа в АТСС РТА-11678, представляет собой полинуклеотид, кодирующий вариабельную область легкой цепи C1GM. Депозиты были сделаны в соответствии с положениями Будапештского договора о международном признании депонирования микроорганизмов для целей процедуры патентования и Инструкциями, принятыми на его основании (Будапештский договор). Это обеспечивает поддержание жизнеспособной культуры депозита в течение 30 лет с даты депонирования. Депозит будет доступен в АТСС по условиям Будапештского договора и в соответствии с соглашением между Pfizer, Inc. и АТСС, который обеспечивает постоянный и неограниченный доступ к потомству культуры депозита для общественности после выдачи соответствующего патента США или обнародовании любой патентной заявки США или иностранной патентной заявки, в зависимости от того, что будет доступным первым, и обеспечивает доступность потомства для лица, определенного комиссаром по патентам и товарным знакам (руководителем Патентного ведомства США) как уполномоченного согласно ст.122, главы 35 Свода законов США (35 U.S.С.Section 122) и правилам комиссара, относящимся к этому (включая ст.1.14, главы 37 Свода законов США (37 C.F.R. Section 1.14) с конкретной ссылкой на 886 OG 638).
Патентовладелец настоящей заявки согласился, что в случае гибели, потери или уничтожения культуры из депонируемого материала при культивировании в подходящих условиях материал будет незамедлительно заменен по уведомлению на такой же другой. Доступность депонируемого материала не следует истолковывать как лицензию на практическое применение изобретения в нарушение прав, предоставляемых ведомством любого государства в соответствии с его патентным законодательством.
Следующие примеры предлагаются только для иллюстрации и в любом случае не предназначены для ограничения объема настоящего изобретения. Действительно, различные модификации изобретения в дополнение к показанным и описанным в данной заявке будут очевидны специалистам в данной области техники из вышеизложенного описания и подпадают под объем прилагаемой формулы изобретения.
Примеры
Пример 1: Получение и скрининг антагонистических антител к IL-7R
Этот пример иллюстрирует получение и скрининг антагонистических антител к IL-7R.
Основные процедуры иммунизации животных для получения моноклональных антител:
2-месячных крыс-самок Sprague Dawley иммунизировали 50 мкг рекомбинантной мышиной химеры IL-7Rα/CD127/Fc, которая включает мышиный IL-7Rα (Glu21-Asp239), сигнальный пептид hCD33 (Met1-Ala16) и человеческий IgG (Pro100-Lys330) (R&D Systems, кат.№747-MR). Антиген для иммунизации готовили путем смешивания 50 мкг антигена в 100 мкл PBS с 100 мкл адъювантной системы Sigma (кат.№S6322). Смесь антигенов встряхивали и вводили в подушечки стоп и брюшину на 0, 2, 5 и 7-е сутки. На 9-е сутки 50 мкг антигена без адъюванта инъецировали внутривенно в общем объеме 150 мкл в физиологическом растворе. На 13-е сутки клетки селезенки готовили в виде одноклеточной суспензии и подвергали слиянию с клетками мышиной миеломы P3×63Ag8.653 согласно стандартному протоколу слияния с использованием 40% PEG 1500 (Boeringer Mannheim Biochemicals #783641). Слитые клетки ресуспендировали в среде, содержащей 18% FBS, 2 мМ L-глутамина, пен/стреп, гипоксантин, аминоптерин и тимидин (HAT) (Sigma Н0262) и 10% добавки для слияния и клонирования гибридомы (HFCS) (кат.№11 363 735 001, Sigma), затем высевали в 54 96-луночных планшета при 200 мкл/лунка. На 7-е сутки после слияния 150 мкл среды отсасывали из каждой лунки и в лунки вносили 200 мкл свежей среды. На 11-13-е сутки супернатант из каждой лунки тестировали в отношении антитела к IL-7R и человеческого Fc с использованием ELISA (описано ниже).
ELISA скрининг антител:
Супернатантные среды от растущих гибридомных клонов подвергали скринингу отдельно на их способность связываться с рекомбинантным мышиным (rm) IL-7R. Анализы проводили с 96-луночными планшетами, покрытыми в течение ночи 50 мкл 1 мкг/мл раствора антигена. 55 покрытых планшетов промывали 4 раза PBS с 0,05% Tween и затем добавляли 50 мкл PBS с 0,5% БСА в каждую лунку. 5 мкл из каждой лунки планшетов с гибридомой добавляли в планшеты для анализа и планшеты инкубировали при комнатной температуре в течение 2 часов для связывания. Излишки реагентов вымывали из лунок между каждой стадией с помощью PBS, содержащего 0,05% Tween-20. 50 мкл Fc-специфического козьего анти-мышиного F(ab')2, конъюгированного с пероксидазой хрена (HRP) (Jackson #115-036-008), добавляли для связывания с мышиными антителами, связанными с антигеном. Для определения добавляли 50 мкл ABTS, 2,2'-азино-бис(3-этилбензотиазолин-6-сульфокислоты) диаммониевой соли, в качестве субстрата. Планшеты считывали через 30 мин при 405 нм с использованием прибора Molecular Devices THERMOmax™. Гибридомные клоны, которые секретировали антитела, способные связываться с мышиным IL-7R, выделяли для дальнейшего анализа. Эти положительные гибридомные супернатанты затем собирали из планшетов с гибридомой и тестировали в ELISA-анализах против человеческого Fc, козьего антикрысиного IgM и рекомбинантного человеческого (rh) IL-7R. Очищенные антитела затем готовили для антител, которые связывались с rm IL-7R, и антител, которые связывались обоими с rm IL-7R, и с rh IL-7R.
Основные процедуры получения полностью человеческих моноклональных антител с использованием фагового дисплея:
Человеческие антитела против человеческого IL-7Rot выделяли из фаг-дисплейной библиотеки человеческих "наивных" scFv антител (Glanville G. et al., 2009, Proc Natl Acad Sci USA, 106(48):20216-20221) с помощью серии из четырех раундов био-пэннинга против человеческого IL-7Rα (R&D Systems®). Для каждого раунда пэннинга 1 мл IL-7Rα (10 мкг/мл в PBS) наносили на иммунопробирку при 4°С в течение ночи. 11-7Ра-покрытую иммунопробирку промывали три раза PBST. 1013 фага (1 мл) добавляли в иммунопробирку и инкубировали при комнатной температуре в течение 1 часа для связывания. После связывания иммунопробирки промывали восемь раз PBST. Связанный фаг элюировали и использовали для заражения свежей культуры клеток TG1. После четвертого раунда пэннинга положительно связавшиеся клетки подвергали скринингу против как человеческого IL-7Rα, так и мышиного IL-7R с помощью ELISA. Антитела, связывающиеся как с человеческим, так и с мышиным IL-7R, дополнительно исследовали в отношении их аффинностей и блокирующей функции и антитела выбирали по созреванию аффинности.
Функциональный анализ in vitro:
Гибридомные клоны, секретирующие антитела, связывающие человеческий или мышиный IL-7R, размножали и собирали супернатанты. Общие IgGs очищали из приблизительно 10 мл супернатанта, используя шарики с протеином А, диализовали в буфере PBS и конечный объем уменьшали до получения растворов с 0,7-1 мг/мл антител. Очищенные антитела затем использовали для тестирования их способности блокировать IL-7-опосредованное фосфорилирование STAT5 в человеческих PBMCs. Для получения РВМС все клетки крови собирали с помощью градиента Ficoll. Клетки поддерживали при 37°С в 5% CO2 в конических пробирках (для предупреждения прикрепления моноцитов/макрофагов) в течение 1-2 ч перед стимуляцией IL-2.
Для функционального скрининга человеческие PBMCs предварительно инкубировали в течение 5 минут с тестируемыми антителами (10 мкг/мл) перед добавлением IL-7. Нереактивное изотипически сходное антитело использовали в качестве отрицательного контроля (изотипический контроль). Клетки стимулировали человеческим IL-7 (0,1 нг/мл, R&D Systems®) в течение 15 минут. Для остановки стимуляции IL-7 добавляли формальдегид непосредственно в культуральную среду до конечной концентрации 1,6%. Клетки фиксировали в течение 15 мин при комнатной температуре. Затем добавляли метанол непосредственно до конечной концентрации 80% и образцы хранили при 4°С в течение периода времени от 30 минут до 1 часа перед иммуноокрашиванием. Клетки окрашивали анти-фосфо-STATS (p-STAT) антителами (BD Pharmingen, Y694 клон 47) и анти-CO4 антителами (BD Pharmingen, RPA-T4). При использовании проточной цитометрии (LSRII, BD™ Biosciences) CD4+ выделенные клетки анализировали на окрашивание р-STAT5. Изотипический контроль устанавливали как 100% p-STAT.
На Фиг.1 показан эффект полностью человеческих моноклональных антител P2D2 и Р2Е11, антагонистических в отношении IL-7R, а также HAL403a на IL-7-опосредованное фосфорилирование STAT5 в человеческих PBMCs. Мышиное моноклональное антитело против человеческого IL-7R 13A2F4 использовали в качестве положительного контроля, и нереактивное изотипически сходное антитело использовали в качестве отрицательного контроля (изотипический контроль). Человеческие PBMCs предварительно инкубировали в течение 5 минут с каждым из тестируемых антител или 13A2F4 в следующих концентрациях: 0,001, 0,01, 0,1, 1 и 10 мкг/мл. Антитело для изотипического контроля использовали при самой высокой концентрации 10 мкг/мл. Клетки стимулировали человеческим IL-7 (0,1 нг/мл) в течение 15 минут, затем фиксировали и подвергали иммуноокрашиванию, как описано выше.
Как измерено по окрашиванию p-STAT5, человеческие антитела P2D2, Р2Е11, HAL403a C1GM, C1GM-2 и С2М3 блокируют дозозависимым образом опосредованный человеческим IL-7 сигнальный путь (Фиг.1 и данные не показаны). Изотипический контроль устанавливали как 100%-ное окрашивание P-STAT5. При 10 мкг/мл антитело HAL403a блокировало фосфорилирование STAT5 очень эффективно (Фиг.1). C1GM, C1GM-2 и С2М3 блокировали фосфорилирование STAT5 сравнимо с HAL403a (данные не показаны).
Аминокислотная последовательность тяжелой цепи антагонистического антитела к IL-7R C1GM (SEQ ID NO: 42) показана ниже.
EVQLVESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWVSLVGWDGFFTYYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARQGDYMGNNWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSWTVPSSSLGTQTYICNVNHKPSNTKVDKKVAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 42)
Аминокислотная последовательность легкой цепи антагонистического антитела к IL-7R C1GM (SEQ ID NO: 43) показана ниже.
NFMLTQPHSVSESPGKTVTISCTRSSGSIDSSYVQWYQQRPGSSPTTVIYEDDQRPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDFHHLVFGGGTKLTVLQPKMPSWLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 43)
Аминокислотная последовательность тяжелой цепи антагонистического антитела к IL-7R C1GM-2 (SEQ ID NO: 45) показана ниже.
EVQLVESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWVSLVGWDGFFTYYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARQGDYMGNNWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSWTVPSSNFGTQTYTCNVDHKPSNTKVDKTVAPPVAGPSVFLFPPKPKDTLMISRTPEWCVWDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTFRWSVLTWHQDWLNGKEYKCKVSNKGLPSSIEKTISKTKGQPREPQNATLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 45)
Аминокислотная последовательность легкой цепи антагонистического антитела к IL-7R C1GM-2 (SEQ ID NO: 43) показана ниже.
NFMLTQPHSVSESPGKTVTISCTRSSGSIDSSYVQvVYQQRPGSSPTTVIYEDDQRPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDFHHLVFGGGTKLTVLQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS (SEQ ID NO: 43)
Аминокислотная последовательность тяжелой цепи антагонистического антитела к IL-7R HAL403a (SEQ ID NO: 17) показана ниже.
QVNLRESGGGLVKPGGSLRLSCAASGFTFDDSVMHWVRQAPGKGLEWLSLVGWDGFFTYYADSVKGRFTISRDNTKNLLYLQMNSLRAEDTAVYYCARQGDYMGNNWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSWTVPSSNFGTQTYTCNVDHKPSNTKVDKTVAPPVAGPSVFLFPPKPKDTLMISRTPEVTCWVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTFRWSVLTWHQDWLNGKEYKCKVSNKGLPSSIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 17)
Аминокислотная последовательность легкой цепи антагонистического антитела к IL-7R HAL403a (SEQ ID NO: 18) показана ниже.
NFMLTQPHSVSGSPGKTVTISCTRSSGSIDSSYVQWYQQRPGNSPTTVIYEDDQRPSGVPDRFSGSIDSSSNSASLTISGLVTEDEADYYCQSYDFHHLVFGGGTKLTVLTVAAPSVFIFPPSDEQLKSGTASWCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ IDNO: 18)
Пример 2: Определение аффинности связывания антитела
Этот пример иллюстрирует определение аффинности связывания антитела для антагонистических антител к IL-7R.
Аффинности антагонистических антител против IL-7R к человеческому IL-7R измеряют на биосенсоре на основе поверхностного плазмонного резонанса Biacore™ 2000 или 3000, оснащенном сенсорным чипом СМ5 для исследований (Biacore™ АВ, Uppsala, Sweden - теперь GE Healthcare). Козьи поликлональные F(ab')2 фрагменты против человека (Fc-специфические) присоединяют по аминам при насыщающих уровнях на всех четырех проточных ячейках с использованием стандартной N-гидроксисукцинимид/этилдиметиламинопропил-карбодиимидной химии (NHS/EDC) в подвижном буфере HBS-P (от Biacore™). Буфер меняют на HBS-P, содержащий 1 мг/мл БСА. Человеческий антиген IL-7R-hFc (R&D systems, Minneapolis, США) разбавляют до примерно 30 мкг/мл и захватывают в течение 3 мин при 5 мкл/мин с получением уровней примерно 500-1000 RU на проточную ячейку, оставляя один пустой контроль, который служит в качестве эталонного канала. Fab, hIgG1 или hIgG2AA форматы антител инъецировали в двух повторностях в виде 5-членных 3-кратных серий, начиная с 2 мкМ и 5-членных 4-кратных серий, начиная с 0,4 мкМ в течение 3 мин при 20-50 мкл/мин. Диссоциацию контролировали в течение 5 мин. Чип для захвата регенерировали после последней инъекции каждого титрования с использованием двух 30-секундных выбросов 75 мМ фосфорной кислотой или 10 мМ Глицин-HCl, рН 1,7. Буферные циклы обеспечивали контроли с двойным стандартом для результатов связывания, которые затем подгоняли в целом к простой обратимой модели связывания с использованием программного обеспечения Biaevaluation v.4.1 для выведения кинетических констант скорости ассоциации и диссоциации, соответственно ka и kd. Аффинность выводили из соотношения (KD=kd/ka). Результаты в Таблице 4 показывают, что эти антитела имеют пикомолярную или наномолярную аффинность к человеческому IL-7R.
|
Пример 3: Антагонистические антитела к IL-7R уменьшают заболеваемость у нетучных диабетических (NOD) животных в мышиной модели диабета 1 типа
Этот пример иллюстрирует эффект антагонистических антител к IL-7R в мышиной модели диабета 1 типа.
Для изучения эффекта in vivo антагонистических антител к IL-7R на диабетогенный процесс крысиное антимышиное антагонистическое антитело к IL-7R, 28G9 (Rinat) тестировали на NOD мышах. NOD мыши демонстрируют склонность к спонтанному развитию аутоиммунного инсулиннезависимого сахарного диабета (IDDM, диабет 1 типа) (Kikutani et al., 1992, Adv. Immunol. 51: 285-322). 28G9 блокирует IL-7 - опосредованное фосфорилирование STAT5 в мышиных спленоцитах и перекрестно конкурирует с антагонистическими антителами к IL-7R человека IL-7R-C1GM, С2М3, HAL403a, HAL403b, Р3А9, Р4В3, P2D2 и Р2Е11 в Biacore™.
6-8 недельным мышам-самкам NOD (The Jackson Laboratory) инъецировали внутрибрюшинно (в/б) еженедельно начиная с 9-недельного возраста (t=0) либо 3, либо 10 мг 28G9 на кг массы тела. PBS или нереактивное сходное по изотипу крысиное моноклональное антитело (изотип) использовали в качестве отрицательных контролей. Сходное по изотипу антитело вводили в концентрации 10 мг/кг массы тела. Мышей подвергали мониторингу два раза в неделю в отношении массы тела и глюкозы в крови. Диабет считали установленным, когда уровень глюкозы в крови был на уровне или выше положительных показаний, т.е. более 250 мг/дл для двух последовательных мониторингов. Начало диабета регистрировали от первого из последовательных измерений.
Как показано на Фиг.2, ни у одной из мышей, обработанных 28G9 в концентрации 10 мг/кг, не развивался диабет даже в возрасте 18 недель. Напротив, у 75-80% PBS- и изотип-обработанных мышей развивался диабет (Фиг.2). Хотя не все мыши, обработанные 28G9 в концентрации 3 мг/кг, не имели диабета в конце исследования, наблюдали значительно уменьшенную заболеваемость диабетом по сравнению с PBS и изотипическим контролями, демонстрируя, что ингибиторный эффект 28G9 на развитие диабета был дозозависимым (Фиг.2). Обработка 28G9 в концентрации 10 мг/кг значительно уменьшала уровень глюкозы в крови по сравнению с изотипическим или PBS контролями (Фиг.3А). Развитие мышей при обработке антагонистическим антителом к IL-7R контролировали путем определения массы тела и смертности. Как показано на Фиг.3В, многократное дозирование 3 или 10 мг/кг 28G9 не оказывало значительного эффекта на рост мышей и смертность не наблюдалась при 10 мг/кг. Таким образом, антагонистические антитела к IL-7R уменьшают уровни глюкозы в крови и ингибируют развитие диабета у NOD животных. Эти результаты демонстрируют, что антагонистические антитела к IL-7R эффективны в предупреждении и замедлении развития диабета 1 типа.
Для исследования эффекта антагонистических антител к IL-7R на регуляцию периферических Т-клеток, CD4+и CD8+Т-клетки подвергали иммуноокрашиванию в отношении маркеров активации CD44 и CD62L и анализировали с помощью проточной цитометрии. CD4+и CD8+Т-клетки выделяли из периферической крови PBS-обработанных, 28С9-обработанных или изотип-обработанных мышей. По сравнению с изотипическим контролем процент "наивных" CD8+Т-клеток (B220-CD8+CD44loCD62Lhi) у мышей, обработанных 28G9 в концентрации 10 мг/кг, был значительно ниже, и процент CD8+Т-клеток памяти (B220-CD8+CD44hiCD62Lhi) был значительно выше (Фиг.4А и 4Б). Напротив, "наивные" CD4+Т-клетки (B220-CD4+CD44loCD62Lhi) не были значительно истощены у мышей, обработанных антагонистическим антителом к IL-7R, по сравнению с изотипическим контролем (Фиг.5). Эти результаты показывают, что антагонистические антитела к IL-7R снижают уровни глюкозы в крови посредством истощения CD8+Т-клеток.
Пример 4: Антагонистические антитела к IL-7R замедляют начало аутоиммунного заболевания
Этот пример иллюстрирует эффект антагонистических антител к IL-7R на мышиную модель рассеянного склероза - экспериментальный аутоиммунный энцефаломиелит (ЕАЕ).
Анализ активации STAT5 использовали для идентификации антагонистических антител к IL-7R. Селезенки от мышей В6 или BALB/c гомогенизировали в PBS и лизировали в лизирующем буфере АСК (Invitrogen) в течение 2 мин и затем фильтровали через сетку с размером пор 100 мкм, осаждали и ресуспендировали в концентрации 5×106 клеток/мл при комнатной температуре в нагретую до 37°С RPM11640, содержащий 10% FBS, пенициллин (100 Ед./мл), стрептомицин (100 мкг/мл) и L - глутамин. Клетки поддерживали при 37°С в 5% CO2 в конических пробирках (для предупреждения прикрепления моноцитов/макрофагов) в течение 1-2 ч перед стимулированием. Клетки предварительно инкубировали с тестируемым антителом в течение 5 минут перед стимуляцией с помощью IL-7. Клетки обрабатывали в течение 15 мин мышиным IL-7 (mlL-7, 0,1 нг/мл). Формальдегид добавляли непосредственно в культуральную среду до конечной концентрации 1,6% и клетки фиксировали в течение 15 мин при комнатной температуре. Метанол затем непосредственно добавляли до конечной концентрации 80% и образцы хранили при 4°С в течение от 30 мин до 1 ч перед иммуноокрашиванием. Следующие антитела использовали для иммуноокрашивания: CD11b-FITC (М1/70), В220-Су5.5РегСР, TCRβ-FITC, p-STAT5 (Y694, клон 47) - Alexa 647 (BD™ Pharmingen). TCRβ+ и CD11b+ клетки окрашивали в отношении фосфо-STAT5. IL-7-стимулированное фосфорилирование STAT5 наблюдали посредством выделения с помощью TCRβ при проточной цитометрии. Идентифицировали ряд антител, которые блокировали фосфорилирование STAT5, включая моноклональные антитела 28В6 и 28G9.
Активный ЕАЕ индуцировали у 6-8-недельных самок мышей линии В6 посредством подкожной иммунизации 100 мкг пептида MOG35-55 (MEVGWYRSPFSRWHLYRNGK (SEQ ID NO: 15)), эмульгированного в CFA, содержащем 1 мг/мл убитых нагреванием Mycobacterium tuberculosis H37RA (Difco), в 0-е сутки (см. Steinman and Zamvil, 2006). Дополнительно мыши получали 400 нг коклюшного токсина (Calbiochem) в/в в 0,1 мл PBS на 0 и 2-е сутки. Начиная с 7-х суток после MOG иммунизации животных обрабатывали два раза в неделю антагонистическим антителом к IL-7R28B6 (10 мг/кг), антагонистическим антителом к IL-7R 28G9 (10 мг/кг) или нереактивным, сходным по изотипу антителом (10 мг/кг). По сравнению с изотипическим контролем обработка либо 28G9, либо 28 В6 значительно уменьшала ЕАЕ активность уже на 15-е сутки после иммунизации (Фиг.6). Этот результат показывает, что антагонистические антитела к IL-7R эффективны для замедления развития ЕАЕ.
Для проверки того, является ли эффект антагонистических антител к IL-7R дозозависимым MOG-иммунизированных ЕАЕ животных обрабатывали либо 1, либо 3 мг/кг 28G9 на 7-е и 14-е сутки после иммунизации. Нереактивное, изотипически сходное антитело (1 мг/кг) использовали в качестве отрицательного контроля. По сравнению с изотипическим контролем обработка 28G9 при обоих уровнях дозировки уменьшала тяжесть ЕАЕ на пике заболевания (Фиг.7). Этот ингибирующий эффект антагонистического антитела к IL-7R длился в течение примерно недели. Этот результат продемонстрировал, что антагонистические антитела к IL-7R обеспечивали защиту как при 1, так и при 3 мг/кг. В отдельном исследовании MOG-иммунизированных ЕАЕ животных обрабатывали еженедельно 1, 3 или 10 мг/кг 28G9, начиная с 7-х суток. Мыши, обработанные 28G9 при 1 мг/кг, продемонстрировали значительную эффективность, без смертности (Фиг.8). Эти результаты показывают, что обработка антагонистическим антителом к IL-7R в концентрации 1 мг/кг является эффективной для замедления развития ЕАЕ и хорошо переносится.
Для исследования эффективности антагонистического антитела к IL-7R при установленном заболевании, MOG-иммунизированных ЕАЕ животных обрабатывали дважды в неделю 28G9 при 10 мг/кг, начиная с 14-х суток после иммунизации. Нереактивное, изотипически сходное антитело (10 мг/кг) использовали в качестве отрицательного контроля. По сравнению с контролем, обработка антагонистическим антителом к IL-7R значительно снижала тяжесть ЕАЕ (Фиг.9). Наблюдали отсутствие смертности при обработке антагонистическим антителом к IL-7R. Этот результат демонстрирует, что антагонистические антитела к IL-7R являются эффективными для снижения интенсивности установленного, продолжающегося ЕАЕ.
Для дополнительного определения, могут ли антагонистические антитела к IL-7R уменьшать ЕАЕ при позднем вмешательстве при низкой дозе, MOG-иммунизированных ЕАЕ животных обрабатывали один раз в неделю 28G9 при 3 мг/кг, начиная с 14-х суток после иммунизации. Нереактивное, изотипически сходное антитело (3 мг/кг) использовали в качестве отрицательного контроля. По сравнению с контролем наблюдали значительное уменьшение тяжести заболевания при обработке антагонистическим антителом к IL-7R (Фиг.10). Этот результат демонстрирует, что обработка антагонистическим антителом к IL-7R является эффективной для уменьшения активности заболевания даже при позднем вмешательстве и более низкой дозе.
Пример 5: Иммунологические изменения после терапии антагонистическим антителом к IL-7R при аутоиммунном заболевании
Этот пример иллюстрирует иммунологические изменения у ЕАЕ мышей после обработки антагонистическим антителом к IL-7R.
Для понимания механизмов, с помощью которых антагонистическое антитело к IL-7R действует для уменьшения интенсивности ЕАЕ в мышиной модели, популяции лимфоцитов из обработанных и контрольных мышей анализировали с помощью иммуноокрашивания и проточной цитометрии. Для иммунологических исследований в этом примере MOG-иммунизированных ЕАЕ животных обрабатывали еженедельно антагонистическим антителом к IL-7R 28G9 (10 мг/кг), 28В6 (10 мг/кг) или носителем (нереактивным, сходным по изотипу антителом, 10 мг/кг). В отдельных исследованиях группу MOG-иммунизированных ЕАЕ животных обрабатывали еженедельно 28В6 (10 мг/кг). Животных умерщвляли на 21-е сутки после иммунизации и собирали центральные лимфоидные органы. Лимфоциты получали из органов и окрашивали, как описано ниже. Иммуноокрашенные лимфоциты анализировали с помощью проточной цитометрии.
Т-клеточные популяции в ВМ, селезенке, крови и тимусе от ЕАЕ животных, обработанных антагонистическими антителами к IL-7R, значительно уменьшались по сравнению с контролями-носителями. Как показано на Фиг.11, обе популяции CD4 Т-клеток (Фиг.11А) и CD8 Т-клеток (Фиг.11Б) из ВМ, селезенки, крови и лимфатических узлов значительно уменьшались у ЕАЕ животных, обработанных антагонистическими антителами к IL-7R. Это согласуется с ролью IL-7R в развитии как CD4, так и CD8 Т-клеток. Однако популяции В-клеток не были значительно уменьшены во всех периферических лимфоидных органах. Этот результат отличается от данных, полученных на IL-7R нокаутных мышах, у которых не имеют ни Т-, ни В-клеток.
Поскольку IL-7Р-сигнальный путь является критичным для выживания "наивных" Т-клеток и пролиферации Т-клеток памяти, эффект антагонистических антител к IL-7R в регуляции периферических Т-клеток анализировали с помощью иммуноокрашивания с использования маркеров активации CD44 и CD62L. CD44loCD62Lhi представляют собой "наивные" Т-клетки, CD44hiCD62L1o представляют собой активированные Т-клетки, и CD44hiCD62Lhi представляют собой Т-клетки памяти. По сравнению с животными, обработанными носителем (нереактивным, сходным по изотипу антителом), мыши, обработанные антагонистическим антителом к IL-7R, имели значительно истощенные популяции "наивных" Т-клеток и активированных Т-клеток (Фиг.12А и 12В). Однако популяции Т-клеток памяти не были значительно истощены (Фиг.12Б). Это селективное истощение популяции "наивных" и активированных Т-клеток может обеспечить преимущество в том, что истощение "наивных" Т-клеток может блокировать активацию образующихся аутоАg-специфических Т-клеток, в свою очередь предупреждая ЕАЕ. Т-клетки памяти не истощаются, и, таким образом, противовоспалительный иммунитет сохраняется.
Снижения NK-клеток у ЕАЕ животных, обработанных антагонистическим антителом к IL-7R, не наблюдалось. Незначительное увеличение процента NK-клеток наблюдалось предположительно благодаря уменьшению процента CD4 и CD8 Т-клеток. Эти данные согласуются с наблюдением, что IL-7/IL-7R сигнальный путь регулирует развитие Т-клеток, но не NK-клеток.
Для определения эффекта обработки антагонистическим антителом к IL-7R на популяцию Treg клеток у ЕАЕ животных лимфоциты окрашивали в отношении Foxp3 для идентификации Treg клеток и тетрамера MOG-MHC класса II (I-Ab) для определения MOG-специфических Т-клеток. Популяция MOG-специфических CD4+Treg клеток, обнаруженная в лимфатических узлах от 2869-обработанных ЕАЕ животных, сходна с таковой у контрольных животных (обработанных нереактивным, сходным по изотипу антителом) (Фиг.13, левый график). Однако увеличение популяции Treg клеток наблюдалось при обработке 28G9 (Фиг.13, правый график). Эти результаты демонстрируют, что обработка ЕАЕ животных антагонистическим антителом к IL-7R приводит к увеличению популяции Treg клеток. Предпочтительно обработка антагонистическим антителом к IL-7R не приводит к развитию воспалительного заболевания, побочного эффекта, наблюдаемого при терапии IL-2Rα антителом.
Получение лимфоцитов и иммунофлуоресцентное окрашивание
Для приведенных выше исследований одноклеточные лейкоцитарные суспензии из селезенок, периферических лимфатических узлов (парных подмышечных, бронхиальных и паховых), тимуса и костного мозга (ВМ) билатеральных бедренных костей получали посредством осторожного иссечения. Мононуклеарные клетки из центральной нервной системы (ЦНС) выделяли после перфузии сердца PBS. Кратко, CNS ткани переваривали коллагеназой D (2,5 мг/мл; Roche Diagnostics) и ДНКазой I (1 мг/мл; Roche Diagnostics) при 37°С в течение 45 минут. Мононуклеарные клетки выделяли посредством пропускания ткани через 70-мкм клеточные фильтры (BD Biosciences) с последующим центрифугированием в градиенте Percoll (70%/37%). Лимфоциты собирали с 37%/70% поверхности раздела и промывали. Следующие антитела использовали для иммуноокрашивания: FITC-, РЕ- или РЕ-Су5-конъюгированный CD3 (17А2), CD4 (Н129.19), CD8 (53-6.7), CD62L (MEL14), CD44 (IM7), В220 (H1.2F3), IgM (II/41), DX5 (CD49b) (все от BD Biosciences). Для внутриклеточного окрашивания цитокинов лимфоциты стимулировали in vitro форбол-12-миристат-13-ацетатом (20 нг/мл; Sigma-Aldrich) и иономицином (1 мкг/мл; Sigma-Aldrich) в присутствии GolgiStop™ (монензин) (5 мкг/мл) в течение 5 часов перед окрашиванием. MOG38-49/IAb тетрамер и контрольный тетрамер (CLIP/lAb) были сконструированы и поставлялись из NIH Tetramer Core Facility. Фоновое окрашивание оценивали с использованием нереактивных, сходных по изотипу контрольных mAbs. Для 2- или 3-цветного иммунофлуоресцентного анализа одноклеточные суспензии (106 клеток) окрашивали при 4°С с использованием заданных оптимальных концентраций mAb в течение 20 минут. Для окрашивания тетрамера лимфоциты окрашивали в течение 3 часов при 37°С.
Пример 6: Антагонистические антитела к IL-7R улучшают толерантность к глюкозе у животных с индуцированным диетой ожирением (DIO)
Этот пример иллюстрирует эффект антагонистических антител к IL-7R в мышиной модели диабета 2 типа.
Для исследования эффекта in vivo антагонистических антител к IL-7R на предварительно установленное жировое воспаление у DIO мышей, мышей-самцов линии C57BL/6 (The Jackson Laboratory) кормили кормом с высоким содержанием жиров (HFD, D12492, 60 ккал% жира, Research Diets) начиная с шестинедельного возраста. После десяти недель диеты с высоким содержанием жиров 16-недельных мышей с ожирением случайным образом распределяли в группы для в/б введения антагонистического антитела к IL-7R 28G9 (10 мг/кг), PBS или нереактивного, изотипически сходного контрольного антитела (10 мг/кг). Через четверо суток после обработки антителом мышей подвергали тестированию на толерантность к глюкозе (в/б 1 г/кг, после 16 часов голодания) для оценки толерантности к глюкозе. В Таблице 5 показаны средняя масса тела и уровни глюкозы для каждой из обработанных групп (PBS-, изотип- или 2809-обработанные мыши). Животные в каждой группе имели сходную массу тела.
|
Результаты теста на толерантность к глюкозе показаны на Фиг.14. Нарушение толерантности к глюкозе, индуцированное диетой с высоким содержанием жиров, облегчали путем обработки антагонистическим антителом к IL-7R. В тесте на толерантность к глюкозе DIO мыши, обработанные 28G9, имели значительно более низкие уровни глюкозы в крови по сравнению с мышами, обработанными изотипом или PBS (Фиг.14). Этот результат демонстрирует, что антагонистические антитела к IL-7R эффективны в животной модели диабета 2 типа.
Пример 7: Антагонистические антитела к IL-7R уменьшают тяжесть заболевания в мышиной модели ревматоидного артрита
Этот пример показывает эффект антагонистических антител к IL-7R в мышиной модели ревматоидного артрита (RA).
Коллаген-индуцированный артрит (CIA) представляет собой широко используемую животную модель, имеющую ряд патологических и гистологических сходных свойств с RA. Для исследования эффекта in vivo антагонистических антител к IL-7R на CIA, 6-8-недельных самцов мышей B10.RIII (линия #000457, The Jackson Laboratory) иммунизировали 150 мкг коллагена II типа (Elastin Products), эмульгированного в полном адъюванте Фрейнда, содержащем 4 мг/мл убитых нагреванием Mycobacterium tuberculosis H37RA (Difco), на 0 и 15-е сутки. Мышам инъецировали в/б 1, 3 или 10 мг/кг антагонистического антитела к IL-7R 28G9 или нереактивного, изотипически сходного контрольного антитела в сутки -1, сутки 1, сутки 8, сутки 15 и сутки 22.
Клинические признаки CIA оценивали ежесуточно с использованием системы балльной оценки от 0 до 5: 0 - норма; 1 - пораженные суставы задних или передних лап или минимальная диффузная эритема и припухлость; 2 -пораженные суставы задних или передних лап или умеренная диффузная эритема и припухлость; 3 - пораженные суставы задних или передних лап или умеренная диффузная эритема и припухлость; 4 - заметная диффузная эритема и припухлость; 5 - диффузная эритема и сильная припухлость всех лап, неспособность согнуть пальцы. Обработка 28G9 при 3 мг/кг значительно уменьшала тяжесть CIA у CII-иммунизированных мышей по сравнению с изотипическим контролем (Фиг.15). Обработка 28G9 при 1 мг/кг не показала значительного уменьшения. Этот результат демонстрирует, что антагонистические антитела к IL-7R эффективны в замедлении развития заболевания в животной модели ревматоидного артрита.
Пример 8: Антагонистические антитела к IL-7R уменьшают тяжесть заболевания в мышиной модели установленного ЕАЕ
Этот пример иллюстрирует эффективность антагонистических антител к IL-7R в мышиной модели установленного ЕАЕ.
ЕАЕ индуцируют у мышей SJL/J посредством иммунизации с помощью 200 мкг PLP(p139-151), растворенного в полном адъюванте Фрейнда, содержащем 4 мг/мл убитых нагреванием Mycobacterium tuberculosis H37Ra (Difco Laboratories). Мышей обследовали ежесуточно путем измерения массы тела и в отношении клинических признаков ЕАЕ и оценивали следующим образом: 0 - отсутствие паралича; 1 - потеря тонуса хвоста; 2 - слабость задних конечностей; 3 - паралич задних конечностей; 4 - паралич задних и передних конечностей; 5 - агония или смерть.
Мышей, имеющих клинические показатели ЕАЕ 2-3, обрабатывали 28G9 (10 мг/кг, в/б), SB/14 (10 мг/кг, в/б) или контрольным IgG (10 мг/кг, в/б) один раз в неделю в течение 2 недель (на 0, 7 и 14-е сутки). 28G9 представляет собой крысиное IgG1 антитело, и SB/14 (BD Biosciences) представляет собой крысиное IgG2a антитело. Клинические показатели оценивали ежесуточно вплоть до 61-х суток.
К 7-м суткам мыши, обработанные 28G9, имели клинические показатели ниже 2 (N=7). Мыши, обработанные 28G9, сохраняли клинические показатели примерно 2 до конца исследования (61-е сутки). По сравнению с ними контрольные IgG-обработанные животные имели клинические показатели между 3 и 4 на протяжении всего исследования. Никакого снижения тяжести заболевания не наблюдалось при обработке SB/14 по сравнению с контролем.
Значительное уменьшение тяжести заболевания наблюдали у животных, обработанных антагонистическим антителом к IL-7R 28G9. Эти результаты демонстрируют, что антагонистические антитела к IL-7R эффективны в уменьшении тяжести заболевания при установленном аутоиммунном заболевании.
Пример 9: Антагонистические антитела к IL-7R уменьшают уровни глюкозы в крови у животных с недавним диабетом
Этот пример иллюстрирует эффективность антагонистических антител к IL-7R в реверсии недавнего диабета в мышиной модели диабета 1 типа.
Панель антагонистических антител к IL-7R тестировали в мышиной модели диабета 1 типа. 28G9 представляет собой крысиное моноклональное антитело IgG1, 28G9-mIgG2a представляет собой антитело, имеющее вариабельные области 28G9 с мышиной константной областью IgG2a, и agly-28G9 представляет собой агликозилированное антитело, имеющее вариабельные области 28G9 с мышиным IgG2a N297A. Для конструирования и экспрессии 28G9-mIgG2a ген VH и Vk крысиного моноклонального антитела 28G9 амплифицировали с помощью ПЦР, клонировали в pARC мышиный IgG2a и pARC мышиный каппа экспрессирующие векторы млекопитающих и котрансфицировали в клетки 293F с помощью Lipofectamin™ (Invitrogen™). Через 5 суток после трансфекции культуральные среды собирали и 28G9 мышиный IgG2a очищали с использованием смолы Mabselect™ (GE). Для конструирования и экспрессии agly-28G9 VH крысиного 28G9 клонировали в сконструированный pARC мышиный IgG2a вектор, в котором Asn-297 из СН2 домена заменяли на Ala (pARC мышиный IgG2a-N297A). Агликозилированное m28G9 (agly-28G9) получали посредством котрансфекции клеток 293F pARC мышиным IgG2a-N297A и pARC-28G9 мышиным каппа вектором.
Мышей NOD со спонтанным недавним началом диабета (т.е. две последовательные концентрации глюкозы в крови были выше 250 мг/дл) обрабатывали в/б еженедельно 28G9-mIgG2a (10 мг/кг, в/б), 28G9 (10 мг/кг, в/б), agly-28G9 (10 мг/кг, в/б) или контрольным IgG (10 мг/кг, в/б). Уровни глюкозы в крови оценивали ежесуточно в течение 140 суток после начала заболевания. У мышей, обработанных 28G9-mIgG2a, наблюдалась 100%-ная ремиссия. У мышей NOD, обработанных 28G9-mIgG2a, уровни глюкозы в крови поддерживали ниже 250 мг/дл при еженедельных инъекциях 28G9-mIgG2a. 28G9 также показало некоторую эффективность в уменьшении уровней глюкозы в крови по сравнению с контрольным IgG. Мыши, обработанные Agly-28G9 и контрольным IgG, имели уровни глюкозы в крови более чем 250 мг/дл на протяжении всего исследования. Эти результаты демонстрируют, что антагонистическое антитело к IL-7R 28G9-mIgG2a является высокоэффективным в уменьшении уровней глюкозы в крови у мышей с установленным диабетом 1 типа.
В отдельном исследовании мышей NOD со спонтанным недавним началом диабета обрабатывали еженедельно начиная с начала заболевания 28G9-mIgG2a (10 мг/кг в/б) для числа доз, указанных в Таблице 6. Уровни глюкозы в крови определяли ежесуточно.
|
Результаты, показанные в Таблице 6, показывают, что две дозы антагонистического антитела к IL-7R были достаточными для поддержания уровней глюкозы в крови ниже 250 мг/дл вплоть до 89-х суток. Три дозы антагонистического антитела к IL-7R были достаточными для поддержания уровней глюкозы в крови ниже 250 мг/дл в течение по меньшей мере 117 суток. В этом исследовании уровни глюкозы в крови, поддерживаемые ниже 250 мг/дл, наблюдались вплоть до пяти месяцев после обработки антагонистическим антителом к IL-7R.
Результаты исследований, описанных выше, показывают, что антагонистическое антитело к IL-7R 28G9-mIgG2a было очень эффективным в снижении уровней глюкозы в крови у животных с недавним началом диабета. Более того, мышей обрабатывали только двумя или тремя дозами антагонистических антител к IL-7R, поддерживая уровни глюкозы в крови ниже 250 мг/дл в течение нескольких месяцев после введения антитела.
Пример 10: Антагонистические антитела к IL-7R уменьшают тяжесть заболевания в мышиной модели реакции "трансплантат против хозяина" (GVHD)
Этот пример иллюстрирует эффект антагонистических антител к IL-7R в мышиных моделях острой и хронической реакции "трансплантат против хозяина" (GVHD).
Острая GVHD
Для мышиной модели острой GVHD, 10×106 человеческих РВМС (свежевыделенных) инъецировали необлученным мышам NOD.SCID IL2Rγ-/-(The Jackson Laboratory, 8-12-недельным). Через 14 суток после инъекции мышей обрабатывали 10 мг/кг антагонистического полностью человеческого IgG1 антитела к IL-7R HAL403b (n=10) или изотипического контроля (n=10) один раз в неделю. Клинические признаки GVHD и массу тела оценивали два раза в неделю. На сороковые сутки после обработки 100% животных, обработанных антагонистическим антителом к IL-7R, оставались живыми, в отличие от лишь 50% выживших животных, обработанных изотипическим контролем. Этот результат показывает, что антагонистические антитела к IL-7R эффективны в уменьшении уровня смертности в животной модели острой GVHD.
Хроническая GVHD
Для мышиной модели хронической GVHD человеческие клетки пуповинной крови, содержащие небольшой (1-5%) процент CD3+Т-клеток, трансплантировали новорожденным облученным мышам NOD.SCID IL2Rγ-/-. Кратко, человеческую CD34+ пуповинную кровь (AHCells, LLC, Emeryville, СА) истощали в отношении CD3+Т-клеток, используя шарики для селекции CD3 человека (Miltenyi Biotec GmBH, Германия, кат. #130-050-101) Для трансплантации от примерно 300000 до 400000 CD34+ клеток, содержащих примерно 1-5% CD3+Т-клеток (в объеме 50 мкл), вводили в полость сердца новорожденной и облученной мыши NOD.SCID IL2Rγ-/- (The Jackson Laboratory). cGVHD развивалась через 16-20 недель после трансплантации.
Начиная с 24-недельного возраста мышам с cGVHD вводили 10 мг/кг антагонистического, полностью человеческого IgG1 антитела к IL-7R HAL403b (n=4) или PBS (n=4) один раз в неделю вплоть до умерщвления.
Мышей умерщвляли в возрасте примерно 28-32 недель, после примерно 4-8 недель обработки антагонистическим антителом к IL-7R или PBS. Мыши, обработанные антагонистическим, полностью человеческим IgG1 антителом к IL-7R, имели значительно меньше потери шерсти, чем мыши, получавшие PBS. Гистологический анализ показал, что почки PBS-обработанных мышей имели более серьезные поражения, чем почки мышей, обработанных антагонистическим антителом к IL-7R. Например, почки контрольных (PBS-обработанных) мышей имели заметное утолщение капиллярных петель с увеличением количества эозинофильного материала. Напротив, почки мышей, обработанных антагонистическим антителом к IL-7R, имели слегка утолщенные капиллярные петли с увеличенным количеством эозинофильного материала. Кроме того, почки мышей, обработанных антагонистическим антителом к IL-7R, имели менее расширенные канальцы по сравнению с почками мышей, обработанных изотипическим контролем, который показал много расширенных канальцев. Гистология легких обнаружила существенное сокращение бронхиальной лимфоидной ткани (BALT) в легких мышей, обработанных антагонистическим антителом к IL-7R, по сравнению с легкими контрольных мышей, у которых присутствовало некоторое количество BALT. Тяжелую атрофию лимфоидной ткани наблюдали в селезенке мышей, обработанных антагонистическим антителом к IL-7R, по сравнению с изменением от легкого до умеренного в селезенке мышей, обработанных PBS.
Эти результаты показывают, что антагонистические антитела к IL-7R эффективны в уменьшении тяжести заболевания в животной модели хронической GVHD.
Пример 11: Антагонистические антитела к IL-7R уменьшают тяжесть заболевания в мышиной модели волчанки
Этот пример иллюстрирует эффект антагонистических антител к IL-7R в мышиной модели волчанки.
Для мышиной модели волчанки использовали мышей MRL/MpJ-Faslpr/J (The Jackson Laboratory). Обычно называемые Ipr мутантами, эти мыши являются гомозиготными по спонтанной мутации лимфопролиферации (Faslpr) и демонстрируют системный аутоиммунитет, массивную лимфаденопатию, связанную с пролиферацией аномальных Т-клеток, артрит и иммунный комплексный гломерулонефроз. Таким образом, мышей MRL/MpJ-Faslpr/J можно использовать в качестве модели системной красной волчанки.
Начиная с момента начала заболевания мыши получали в/б еженедельно 1, 3 или 10 мг/кг антагонистического антитела к IL-7R 28G9-mIgG2a (см. Пример 10), 1 мг/кг антагонистического антитела к IL-7R agly-28G9, изотипический контроль IgG (отрицательный контроль) или циклофосфамид (положительный контроль). Для каждой группы обработки использовали десять мышей (n=10). Тяжесть заболевания контролировали путем измерения уровней протеинурии, уровней активности и оценки рефлекса выпрямления. При оценке рефлекса выпрямления, мышей, которые не могли восстановиться в течение 30 секунд, умерщвляли. Уровень выживаемости представлен в Таблице 7 ниже.
|
Как показано в Таблице 7, мыши, обработанные 1 мг/кг agly-28G9, 3 мг/кг 28G9-mIgG2a или 10 мг/кг 28G9-mIgG2a, имели повышенный уровень выживаемости по сравнению с мышами, обработанными изотипическим контрольным IgG. Эти результаты показывают, что антагонистические антитела к IL-7R эффективны в уменьшении тяжести заболевания в животной модели волчанки.
Пример 12: Эпитопное картирование/связывание антагонистических антител к IL-7R
Этот пример иллюстрирует опосредованный структурой мутагенез для картирования антителосвязывающих эпитопов.
На основе кристаллической структуры комплекса IL-7/IL-7Rα и вероятного участия некоторых остатков в связывании IL-7 (McElroy et al., 2009, Structure, 17(1):54-65), двадцать три мутанта поверхностных остатков IL-7Rα (N29D, V58S, E59R, R66N, K77S, L80S, I82S, K84S, K100S, Т105А, N131S, Q136K, K138S, Y139F, K141S, М144А, D190S, H191N, Y192A, Y192F, K194S, K194A и F196S) были выбраны для мутации для картирования антителосвязывающих эпитопов. Нумерация мутантов (т.е. N29D, V58S, E59R и т.д.) соответствует соглашению о пост-процессированном белке, где первые 20 аминокислот не учитываются.
Панель IL-7Rα одиночных точечных мутантов (his-меченных) готовили следующим образом. Двадцать три одиночных точечных мутанта IL-7Rα, описанных выше, получали из ранее описанной конструкции ДНК дикого типа (McElroy et al., 2009, выше) с использованием стандартных методов ДНК. Мутантные белки были экспрессированы с использованием временной трансфекции в клетках HEK293T и секретировались в клеточные среды. Мутантные белки очищали посредством Ni2+ колоночной хроматографии. Концентрации белка измеряли спектрофотометрически (NanoDrop™).
Анализ взаимодействия IL-7Rα осуществляли при 25°С с использованием биосенсора ProteOn™ XPR36 на основе поверхностного плазмонного резонанса, оснащенного сенсорным чипом GLM (Bio-Rad, Hercules, СА, США). Подвижный буфер HBST (10 мМ Hepes рН 7,4, 150 мМ NaCl, 0,05% об./об. Tween-20) использовали в течение процесса. Полноразмерные антитела к IL-7R (HAL403a или HAL403b) связывали по аминным группам на отдельных "вертикальных" каналах чипа посредством стандартной ЕОС/сульфо-NHS-опосредованной химии до уровней примерно 2000-5000 RU. Панель мутантов IL-7Rα (включая IL-7Rα дикого типа) подвергали скринингу в "горизонтальном" направлении при 100 нМ с использованием фаз ассоциации и диссоциации по 3 и 10 минут соответственно при 30 мкл/мин. Поверхности регенерировали с помощью элюирующего буфера 2/1 об./об. Pierce immunopure (рН 2,8)/4 М NaCl. Большинство инъекций дублировали для подтверждения воспроизводимости анализа.
В Таблице 8 суммирован эффект единичных точечных мутаций у мутантов IL-7Rα на связывание антитела по сравнению с IL-7Rα дикого типа.
|
Мутанты IL-7Rα, демонстрирующие ослабленное связывание антитела по сравнению с IL-7Rα дикого типа, идентифицировали как имеющие точечную мутацию в остатке, вовлеченном в связывание mAb. Связывающие остатки IL-7Rα с антителом НА1403а в порядке убывания эффектов мутации идентифицировали следующим образом: I82 (значительный эффект на связывание), K84 (средний эффект), K100 (средний эффект), Т105 (средний эффект), Y192 (средний эффект), D190 (незначительный эффект), Н191 (незначительный эффект) и K194 (незначительный эффект). Связывающие остатки IL-7Rα с антителом HAL403b в порядке убывания эффектов мутации идентифицировали следующим образом: I82 (значительный эффект на связывание), K84 (средний эффект), K100 (средний эффект), Т105 (средний эффект), Y192 (средний эффект), D190 (незначительный эффект), Н191 (незначительный эффект) и K194 (незначительный эффект).
Хотя раскрытые идеи были описаны со ссылкой на различные применения, способы, наборы и композиции, следует понимать, что различные изменения и модификации могут быть сделаны без отклонения от идей, изложенных в данной заявке, и заявленного изобретения ниже. Представленные выше примеры предложены для лучшей иллюстрации раскрытых идей и не предназначены для ограничения объема идей, представленных здесь. Хотя настоящие идеи были описаны в терминах этих примерных воплощений, специалистам будет понятно, что многочисленные вариации и модификации этих примерных воплощений возможны без чрезмерного экспериментирования. Все такие вариации и модификации находятся в пределах рассматриваемых идей.
Все ссылки, процитированные в данной заявке, включая патенты, патентные заявки, статьи, учебники и тому подобное, включены посредством ссылки во всей своей полноте. В случае если один или более из включенных литературных источников и сходных материалов отличаются или противоречат данной заявке, включая, но не ограничиваясь этим, определенные термины, использование терминов, описанные методы и тому подобное, эта заявка имеет преимущественную силу.
Приведенное выше описание и примеры выделяют некоторые конкретные воплощения изобретения и описывают лучший вариант, рассматриваемый изобретателями. Следует отметить, однако, что независимо от того, насколько подробно вышеизложенное описано в тексте, изобретение можно осуществить разными способами и изобретение следует рассматривать в соответствии с прилагаемой формулой изобретения и любыми ее эквивалентами.
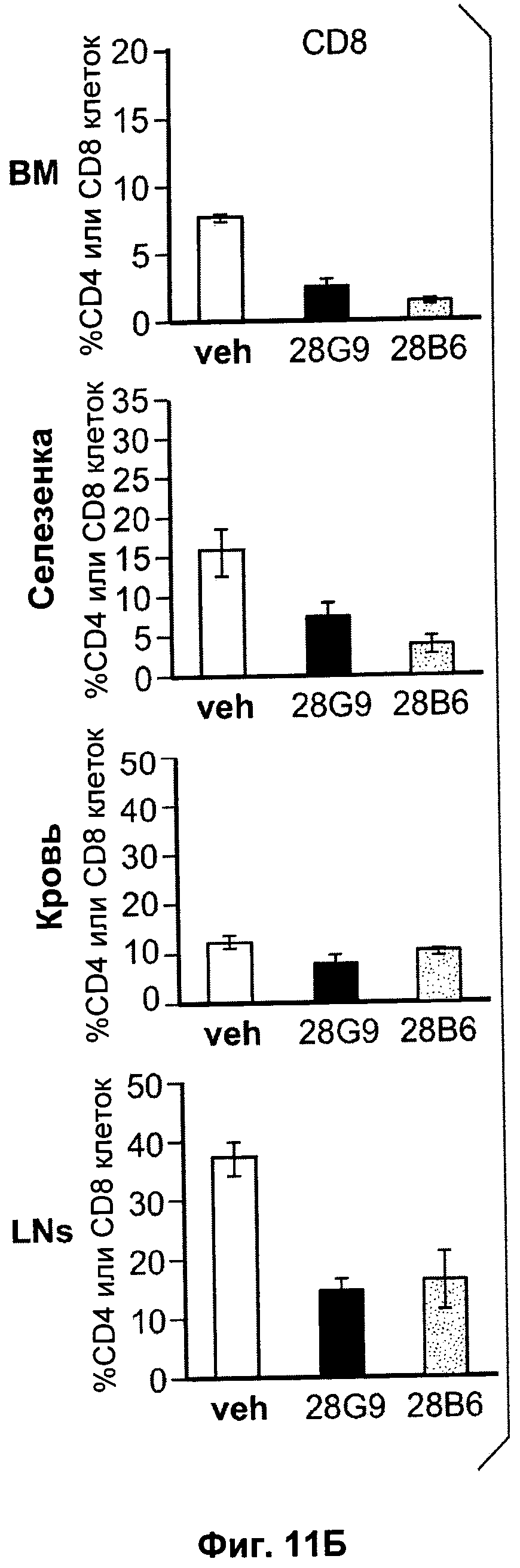
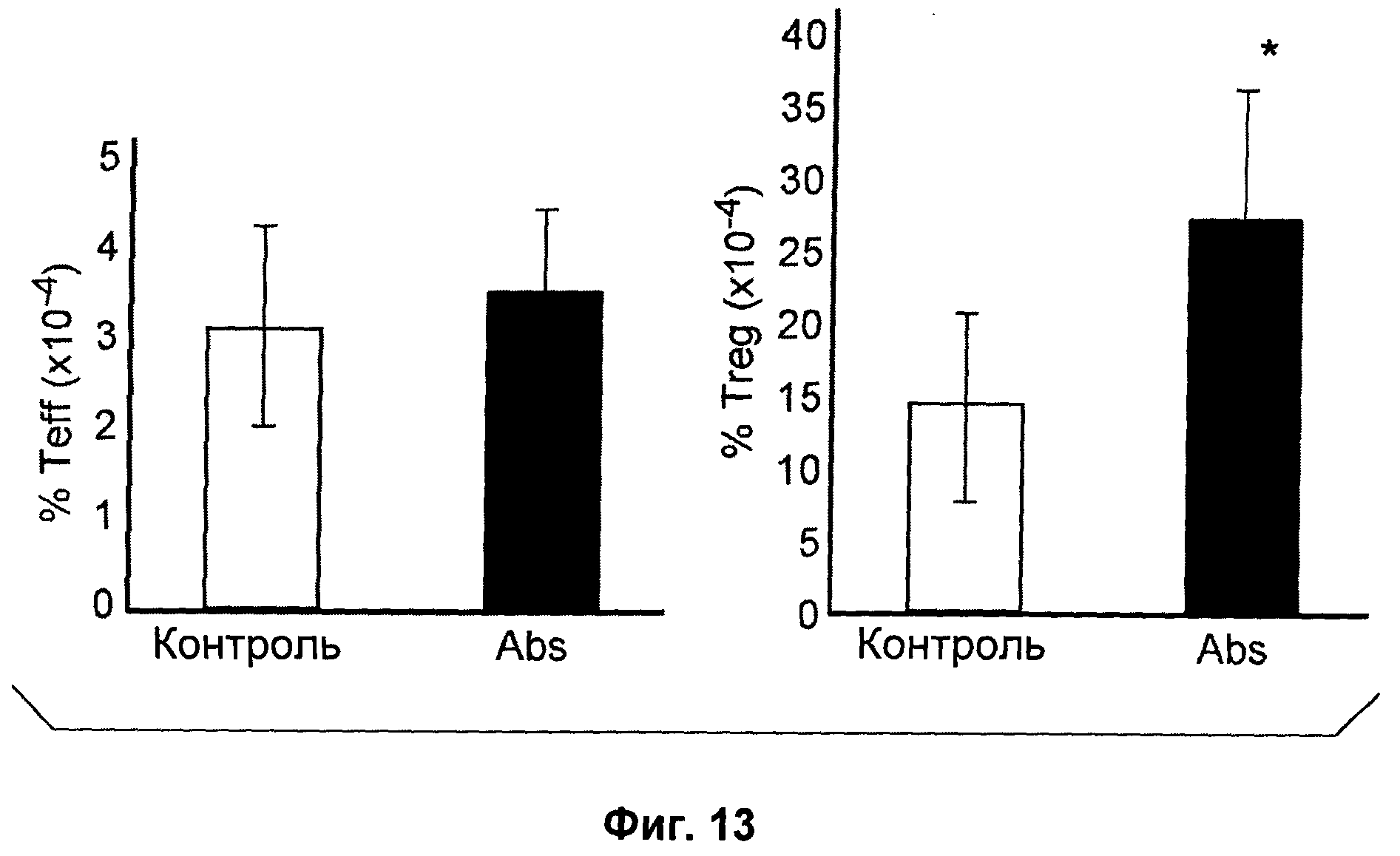
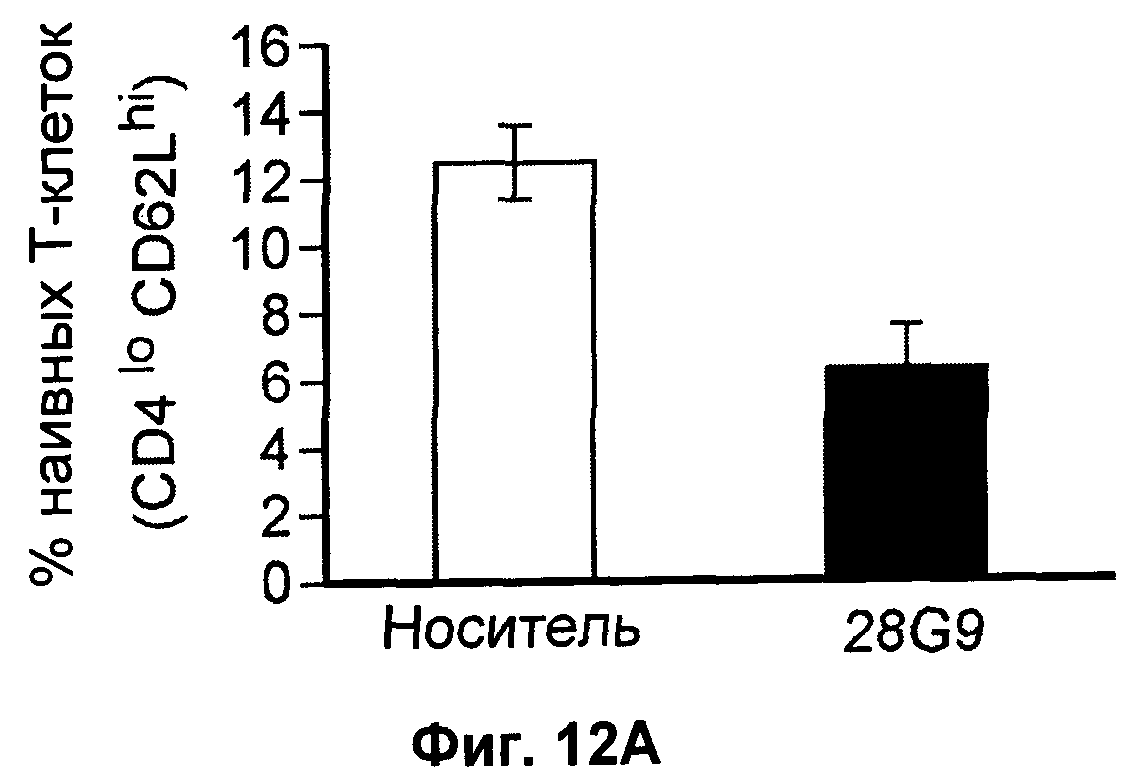
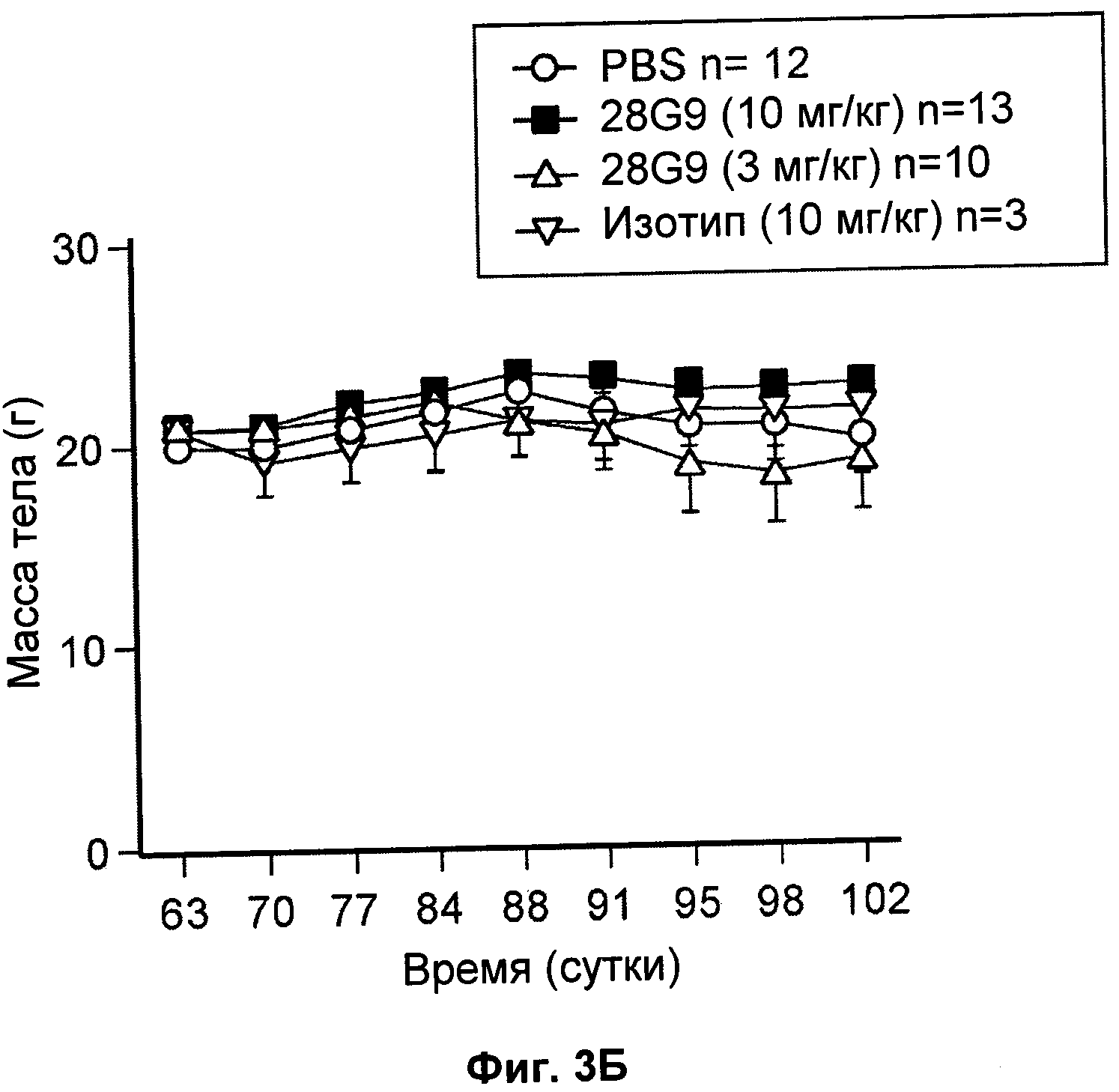
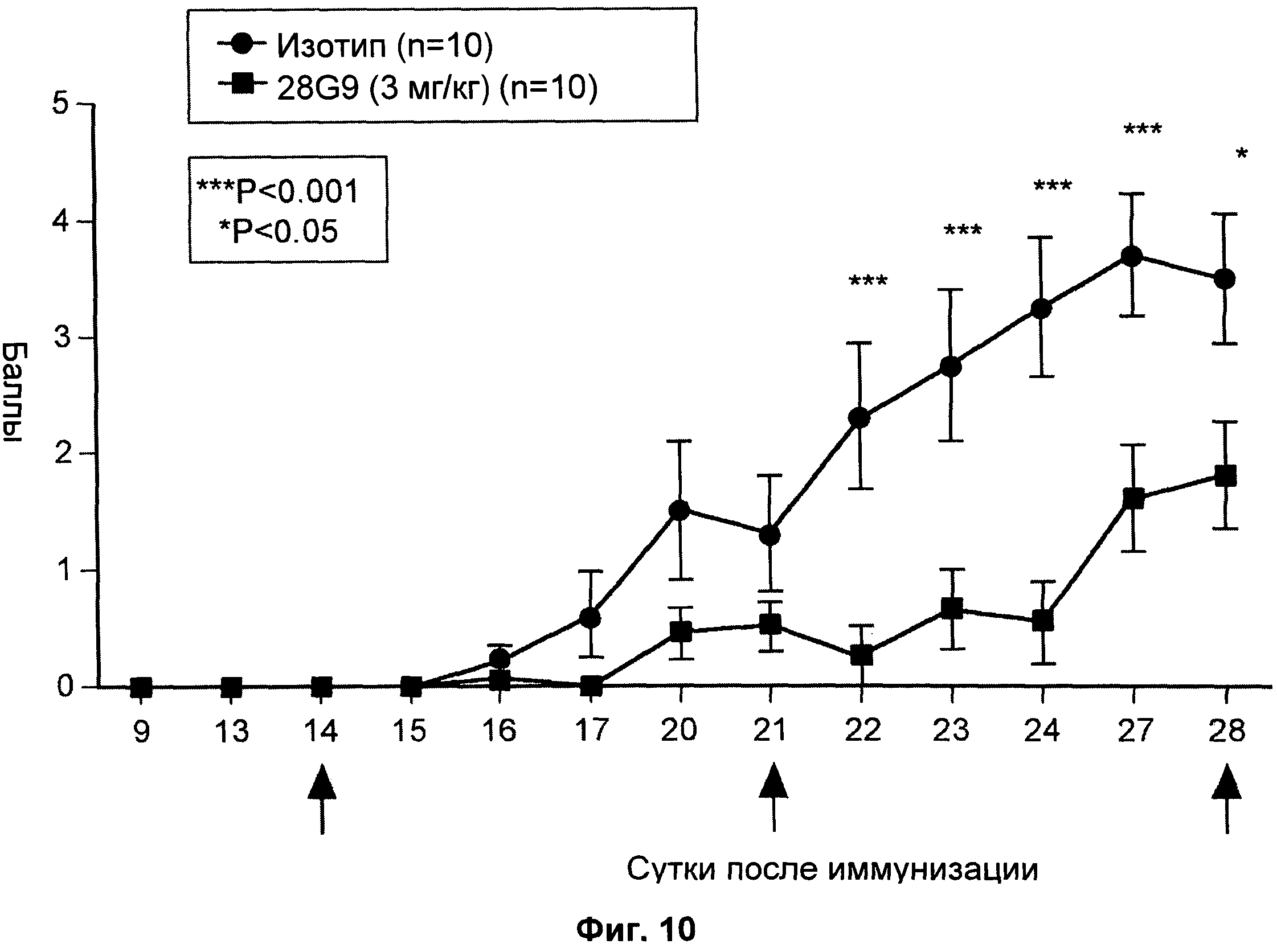
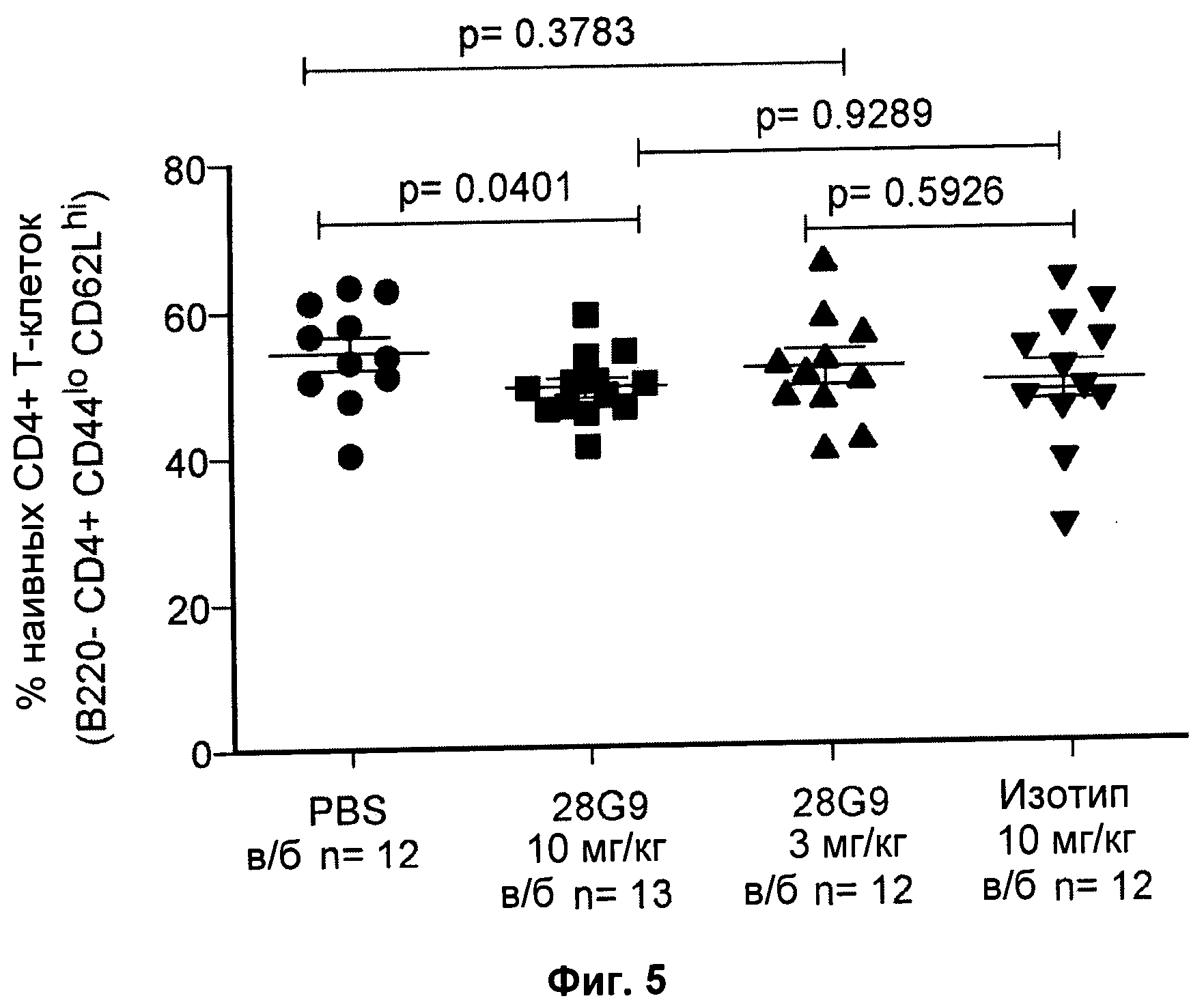
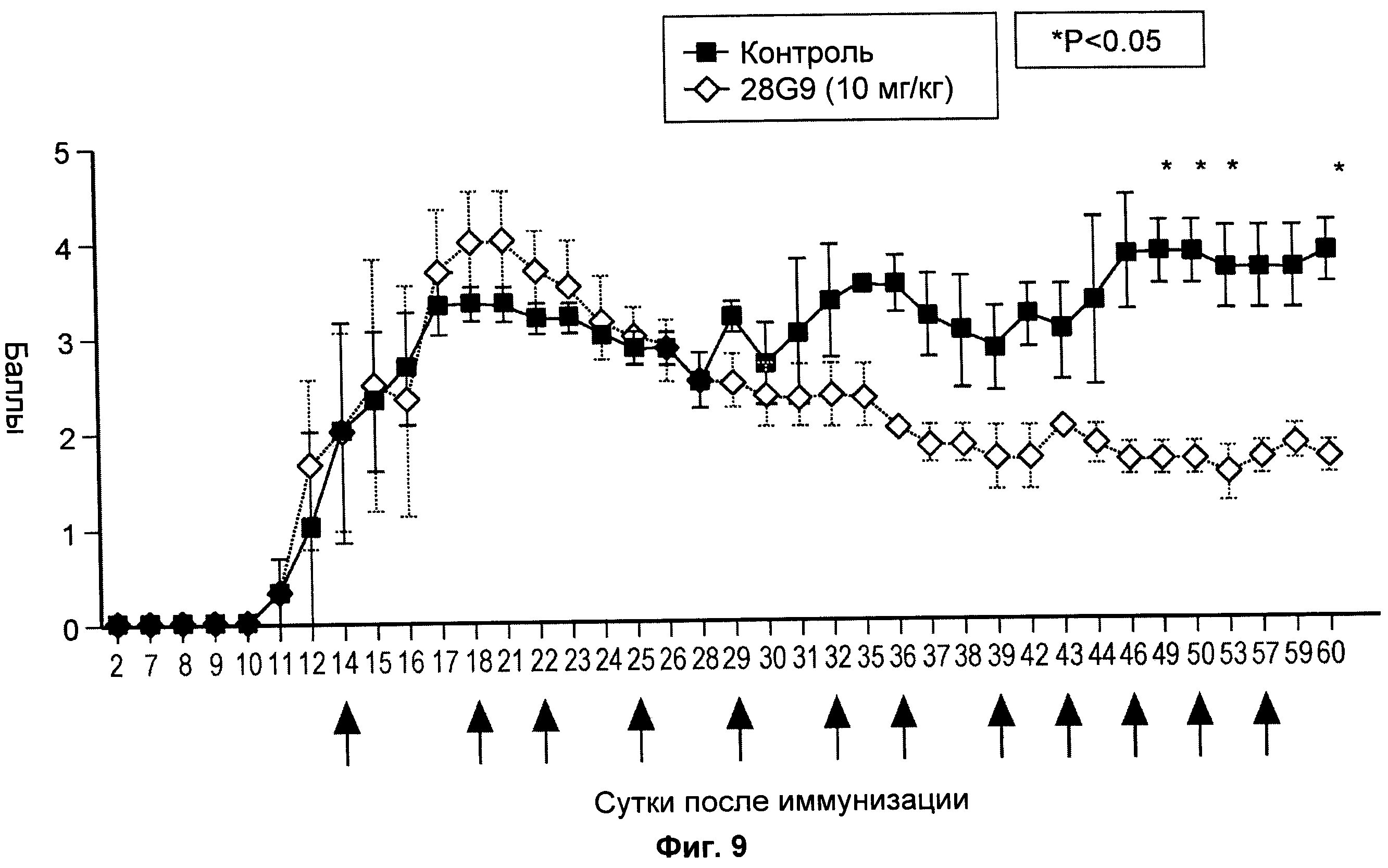
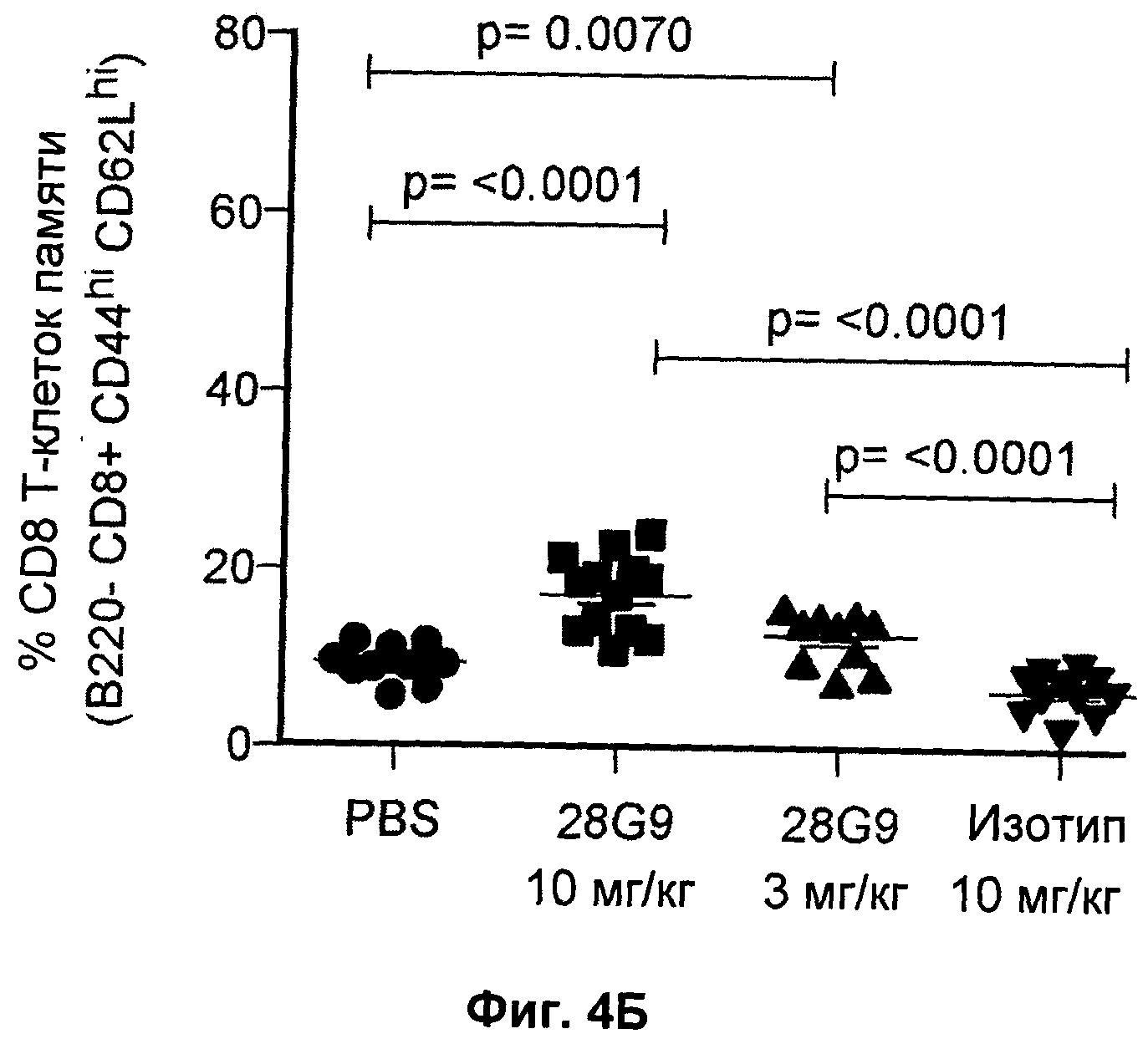
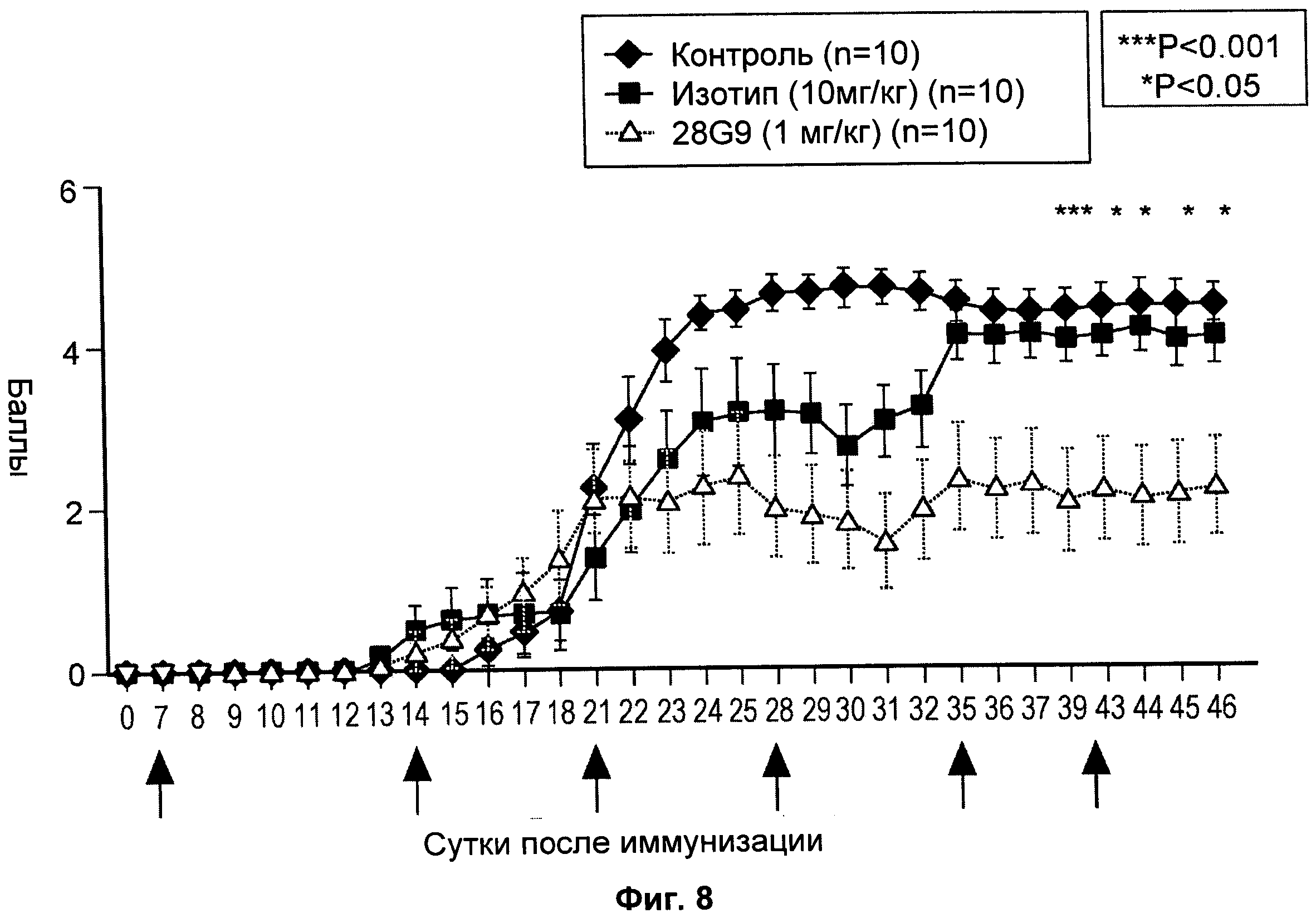
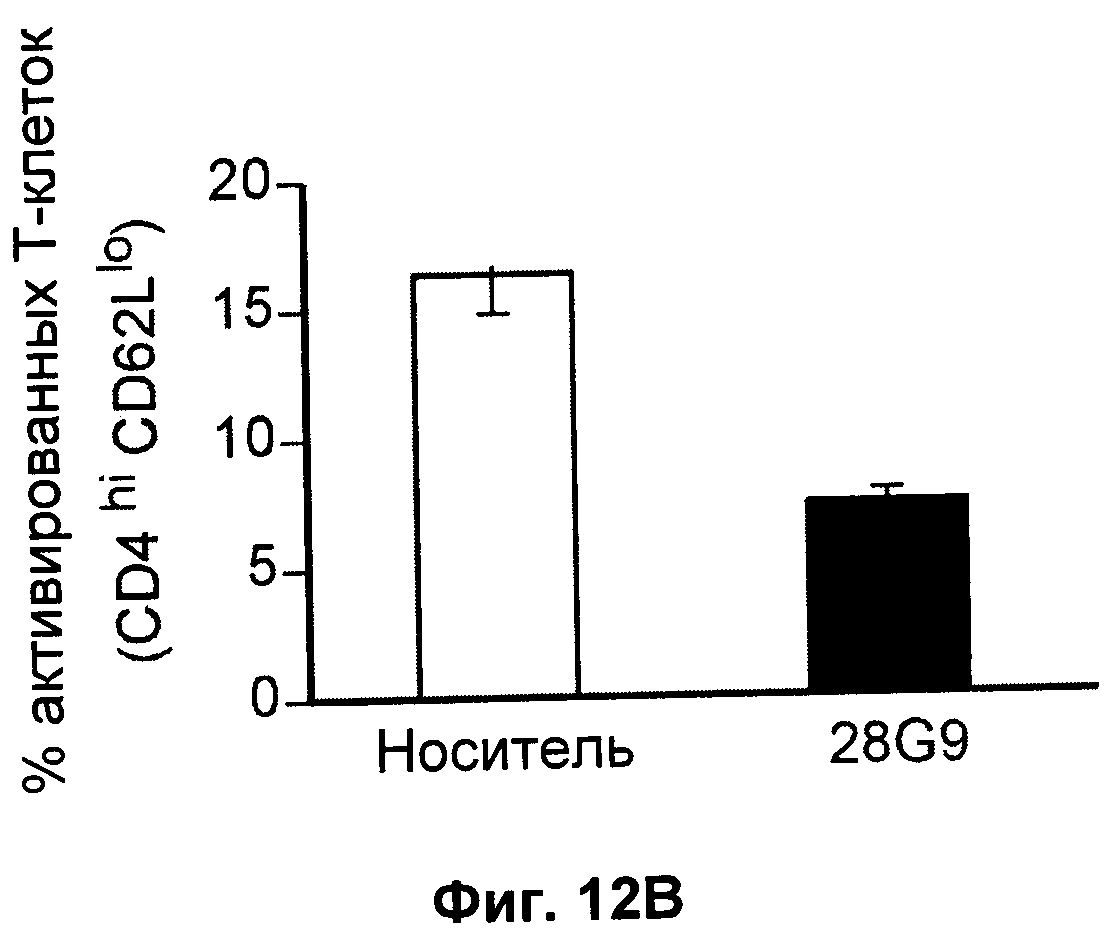
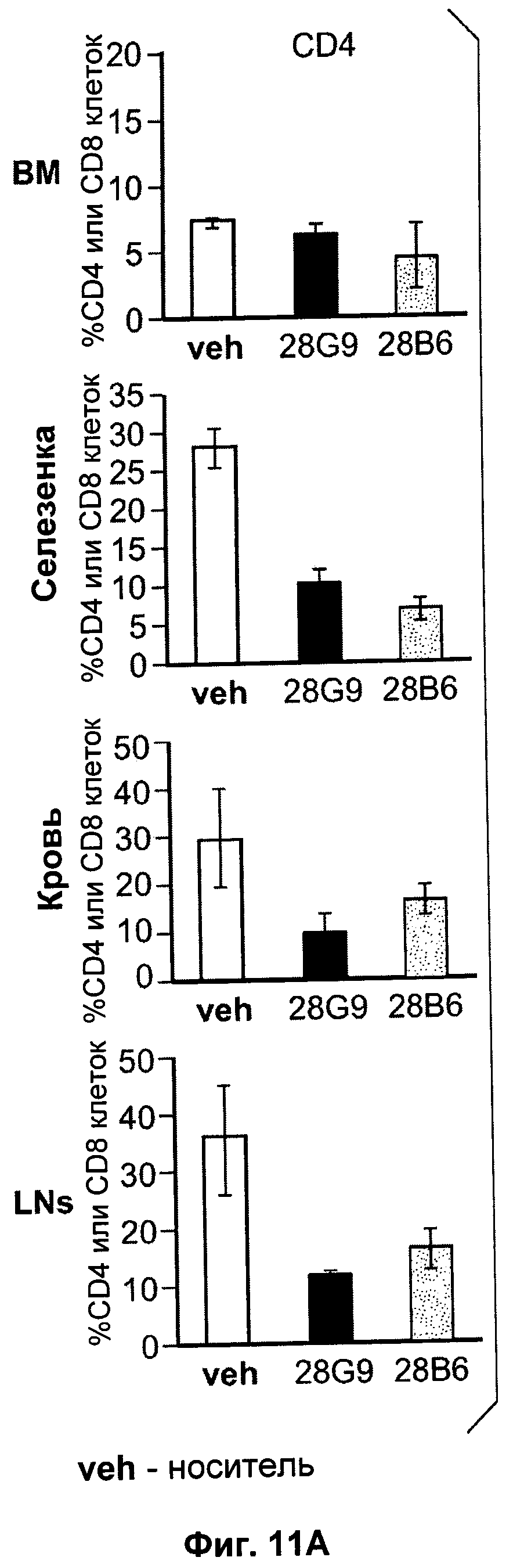
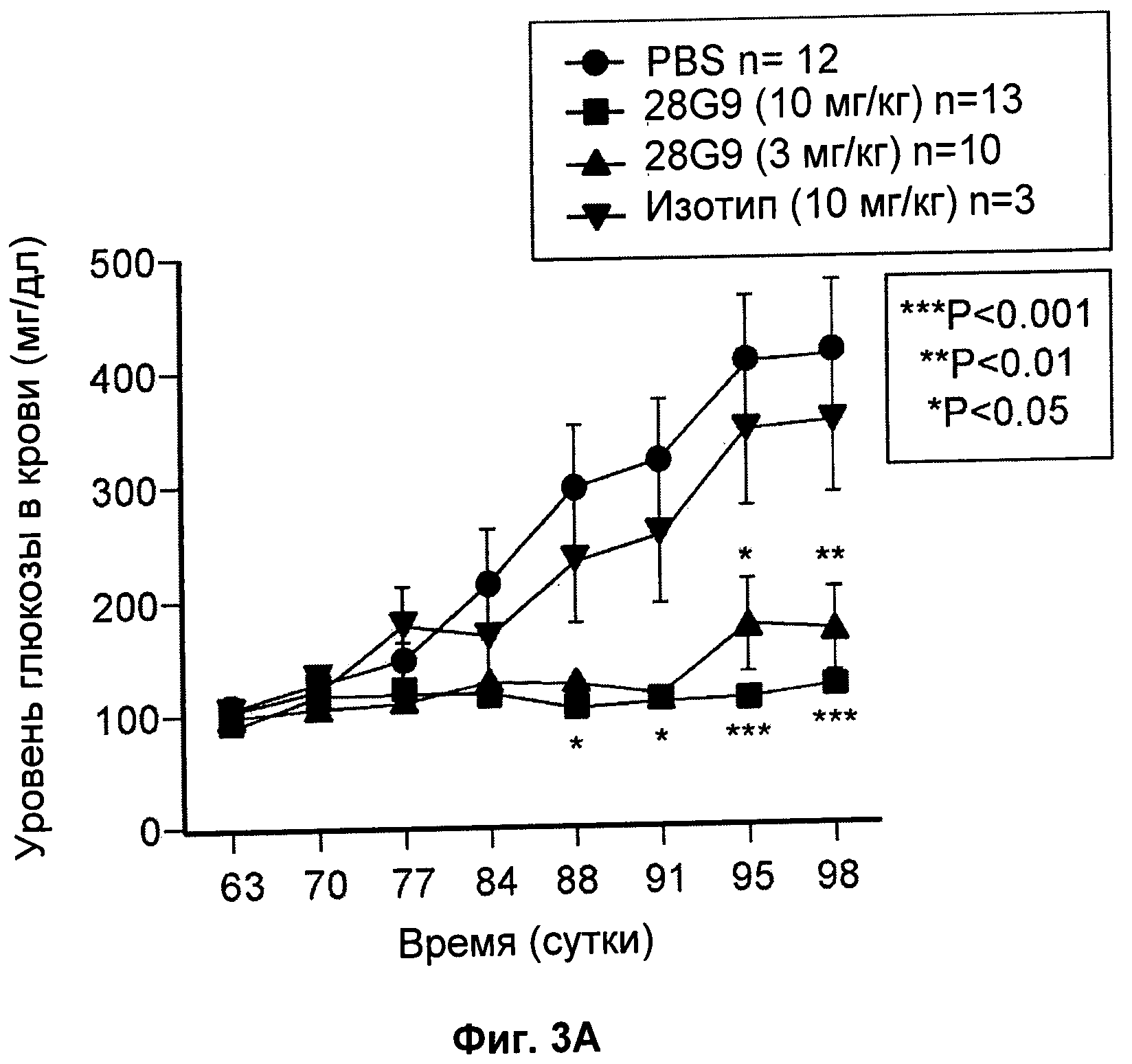
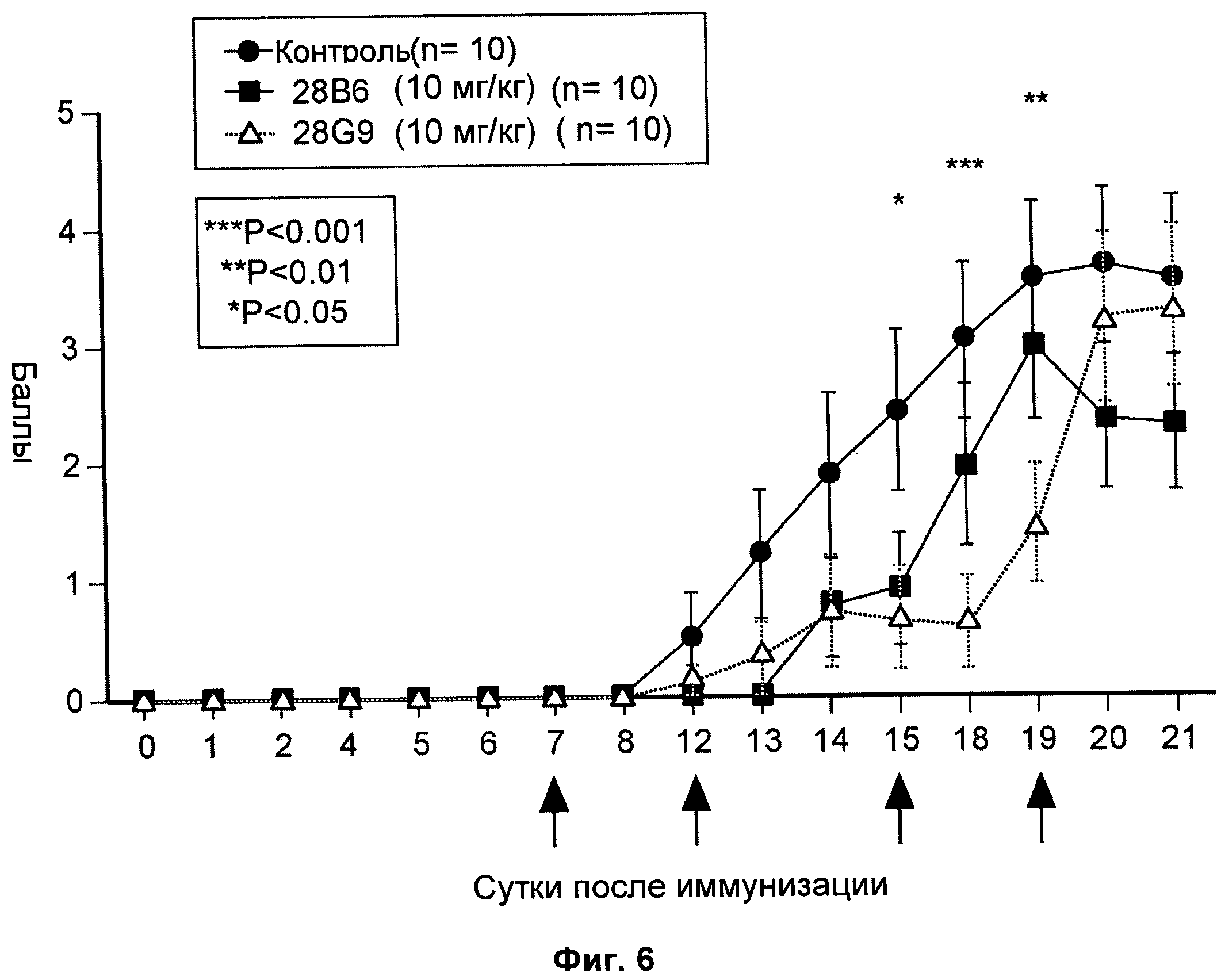
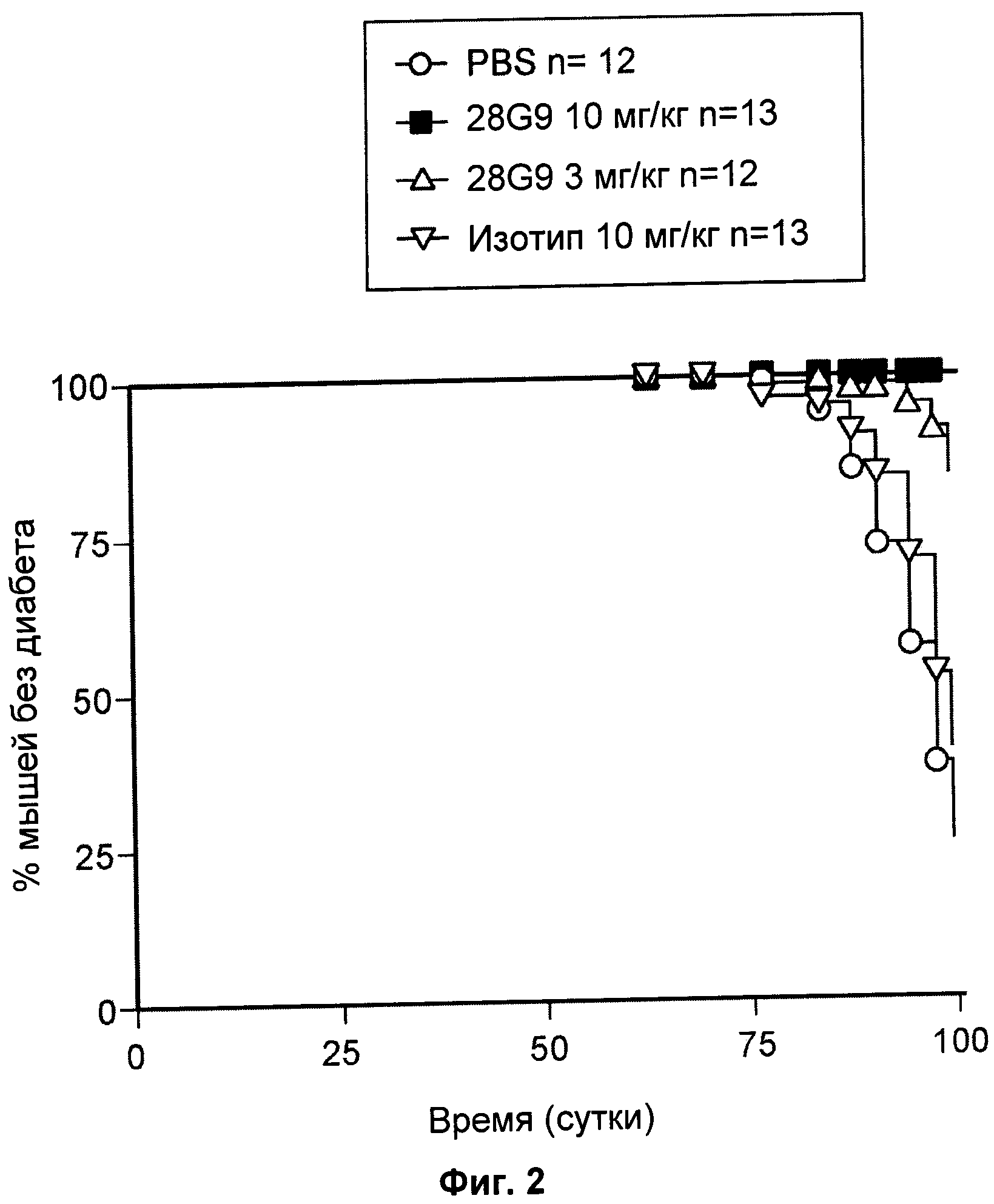
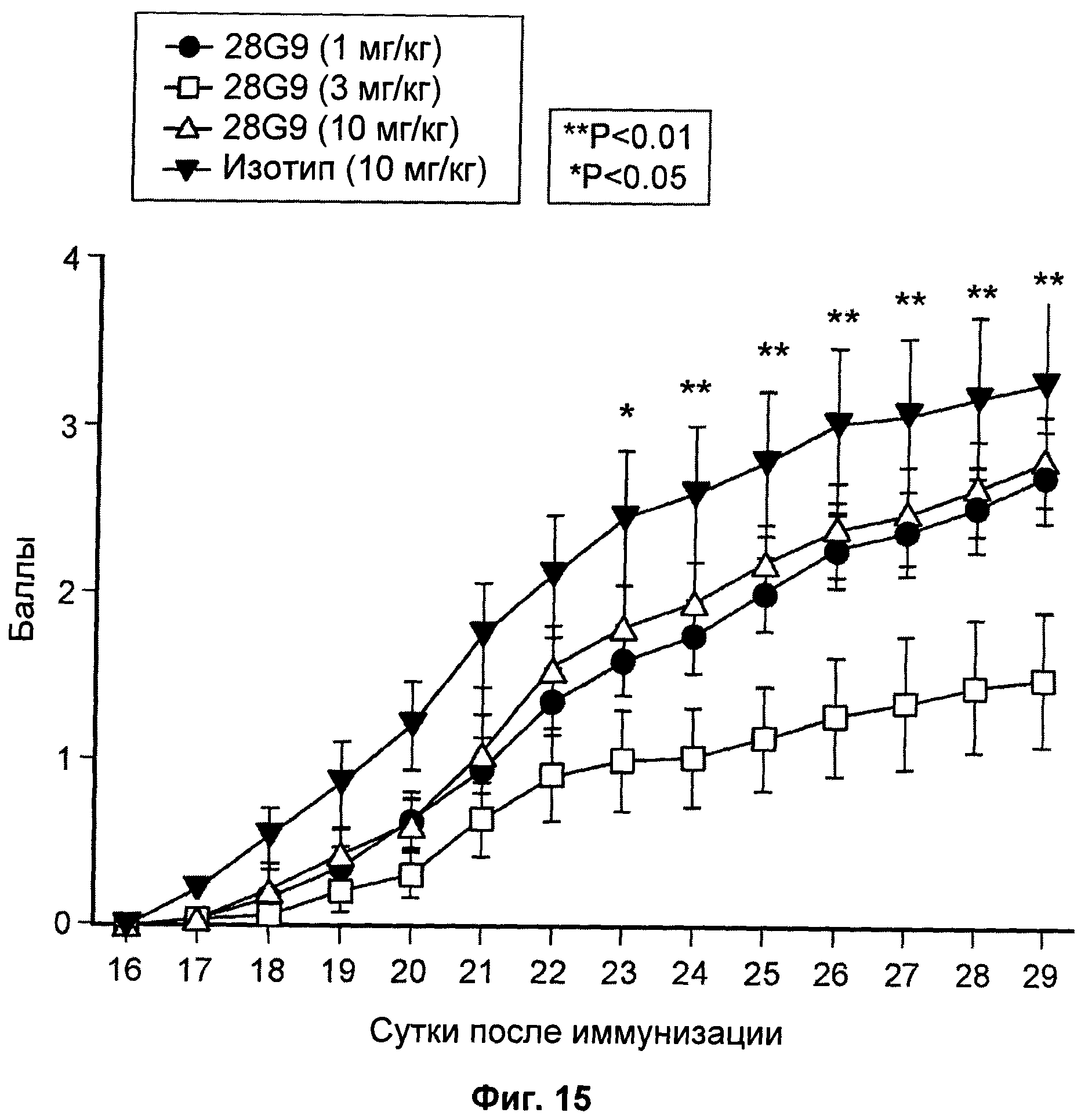
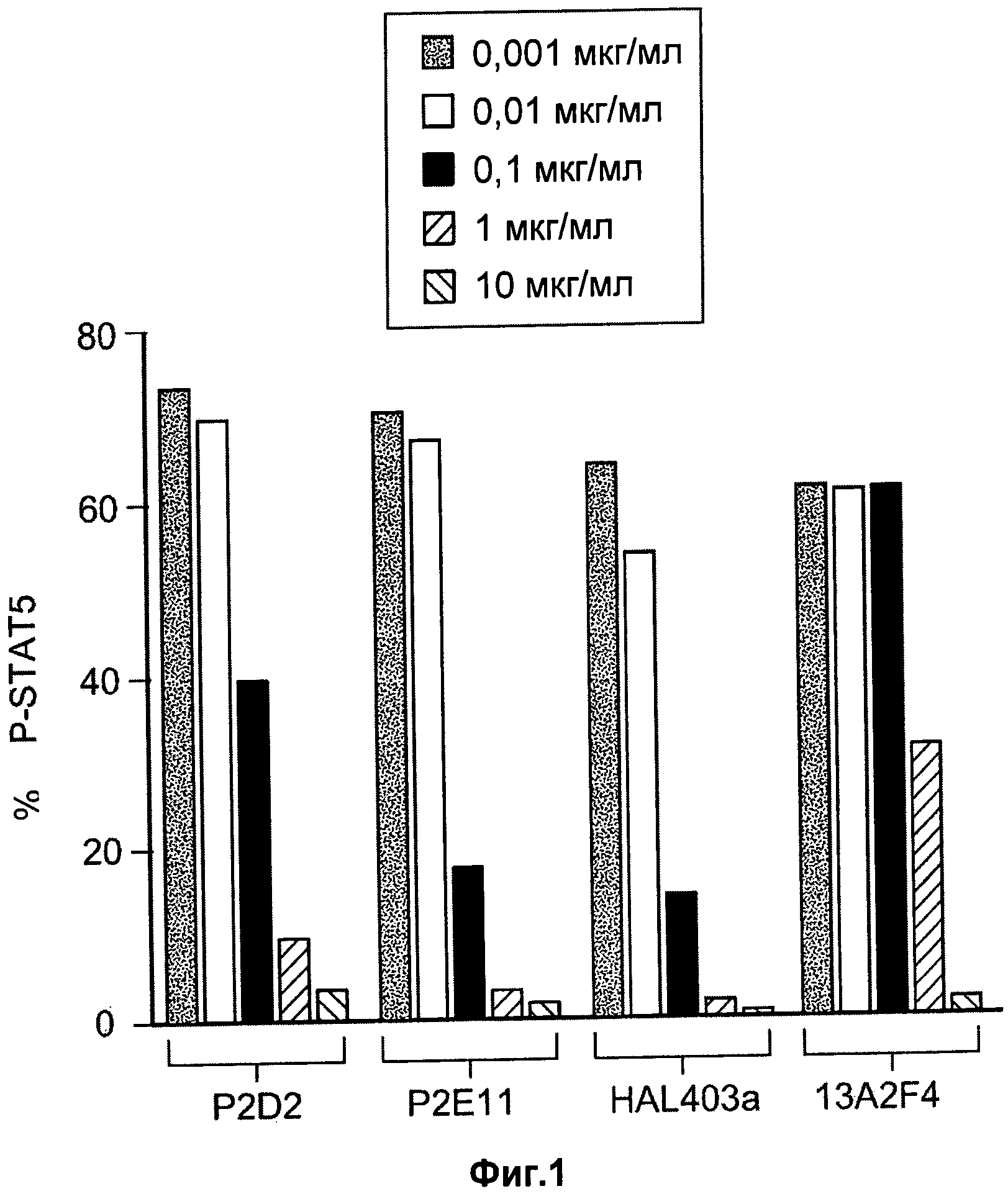
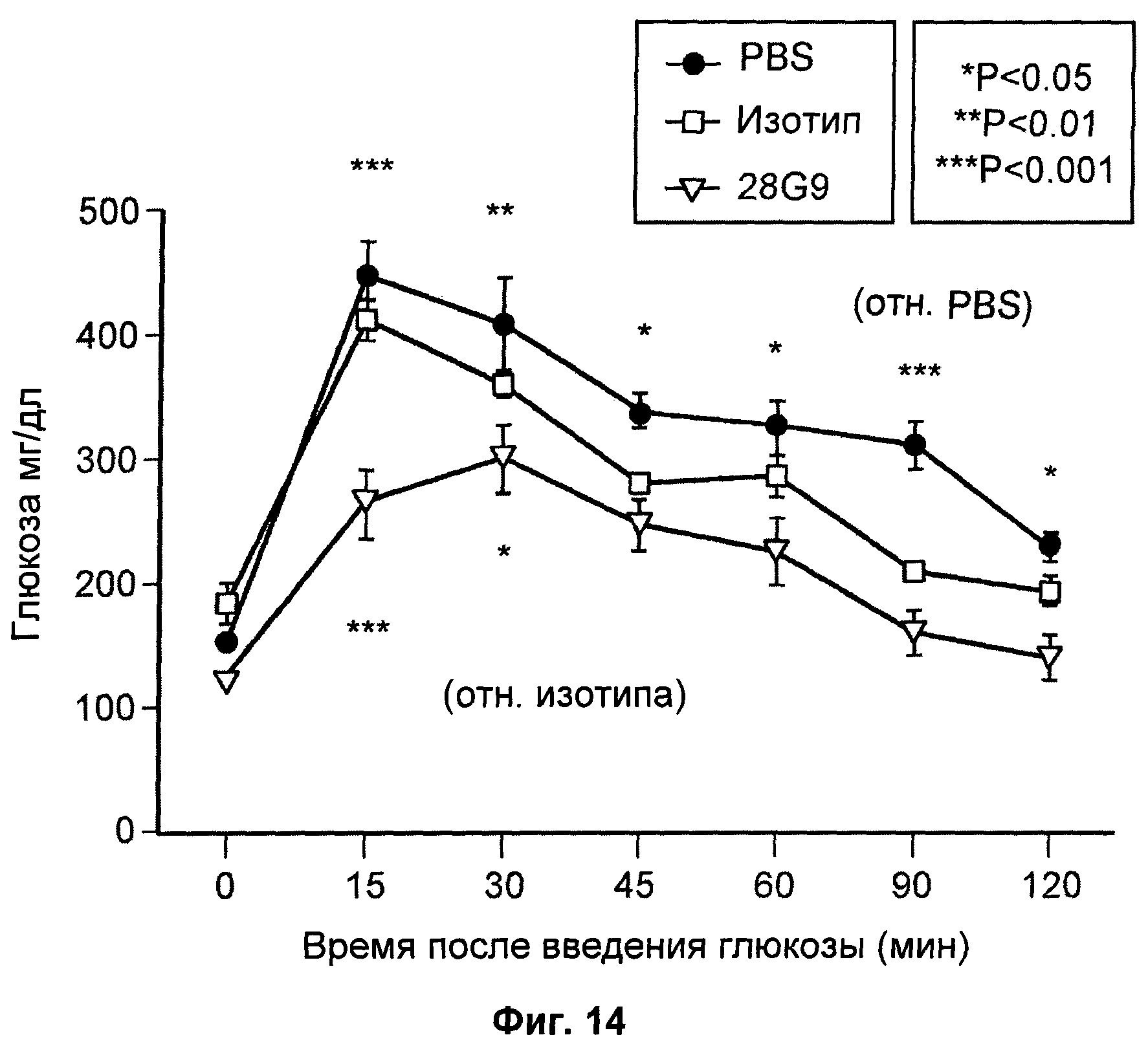
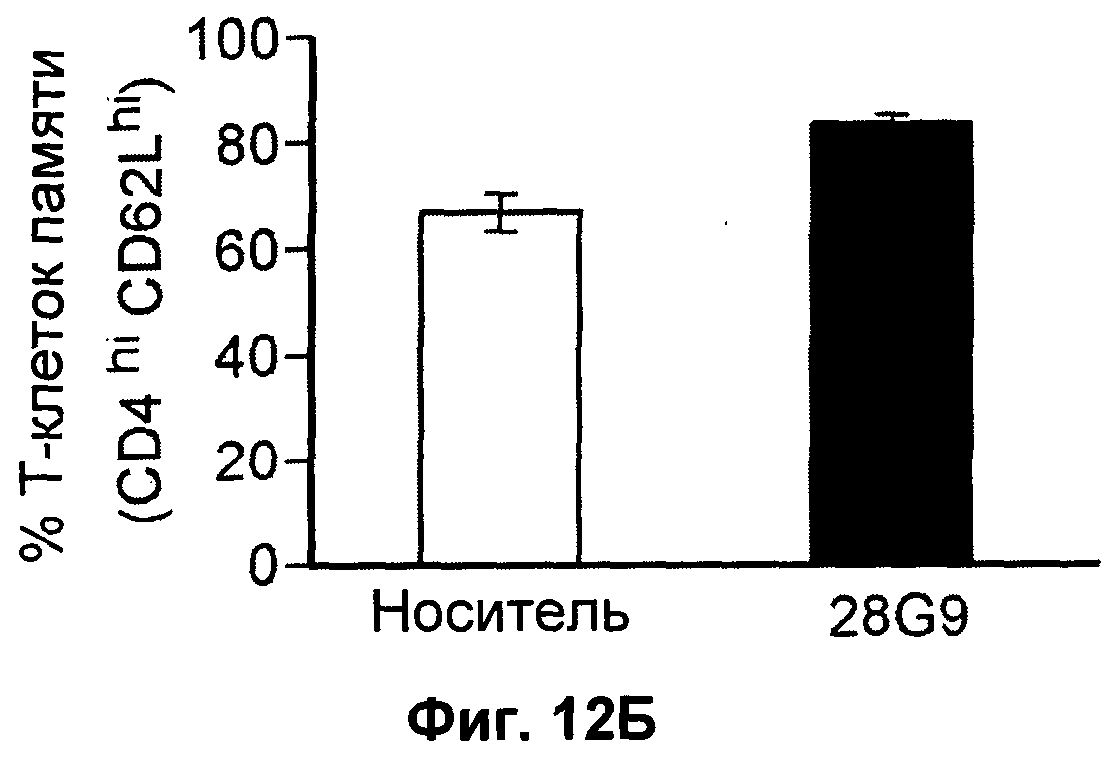
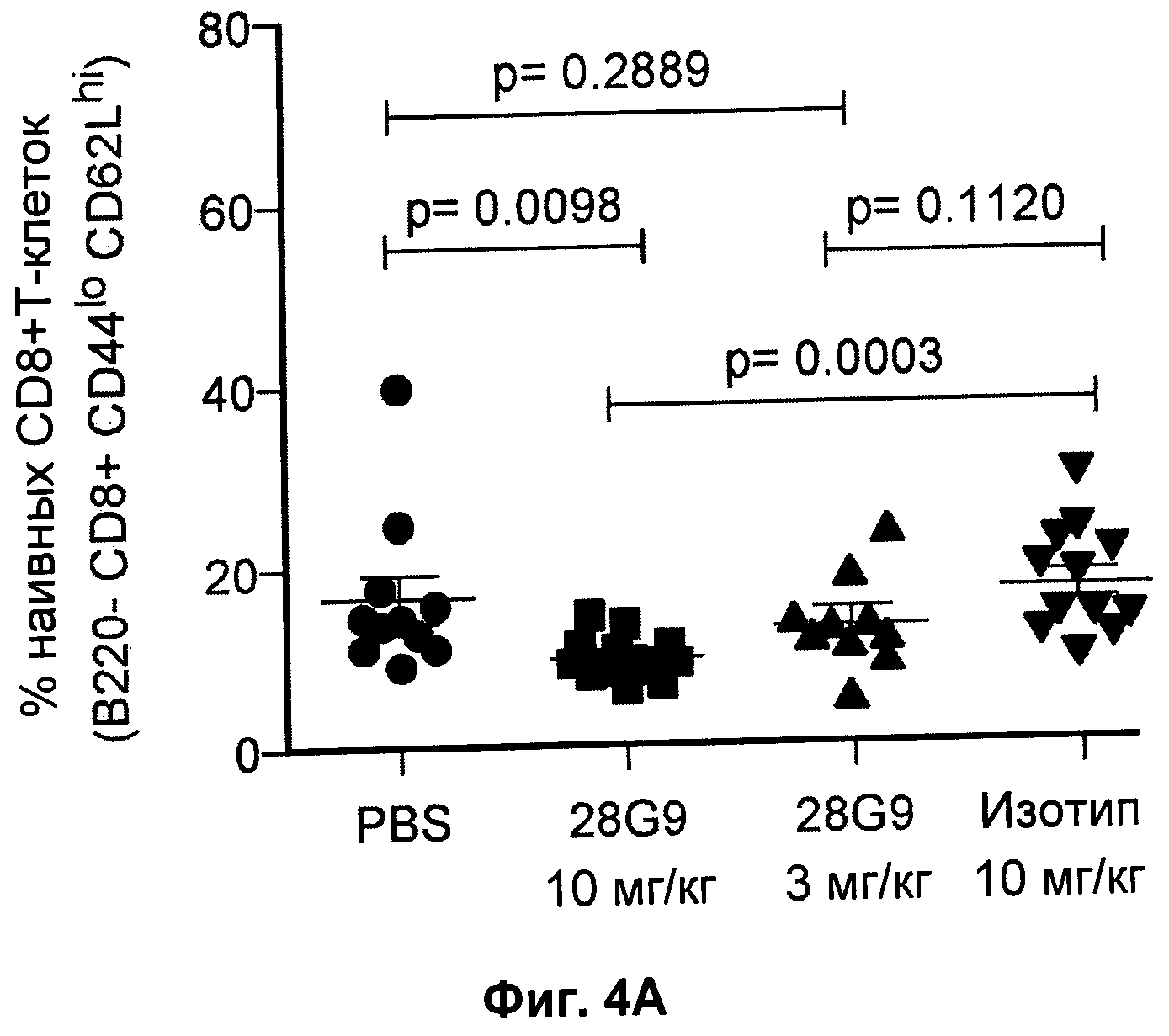
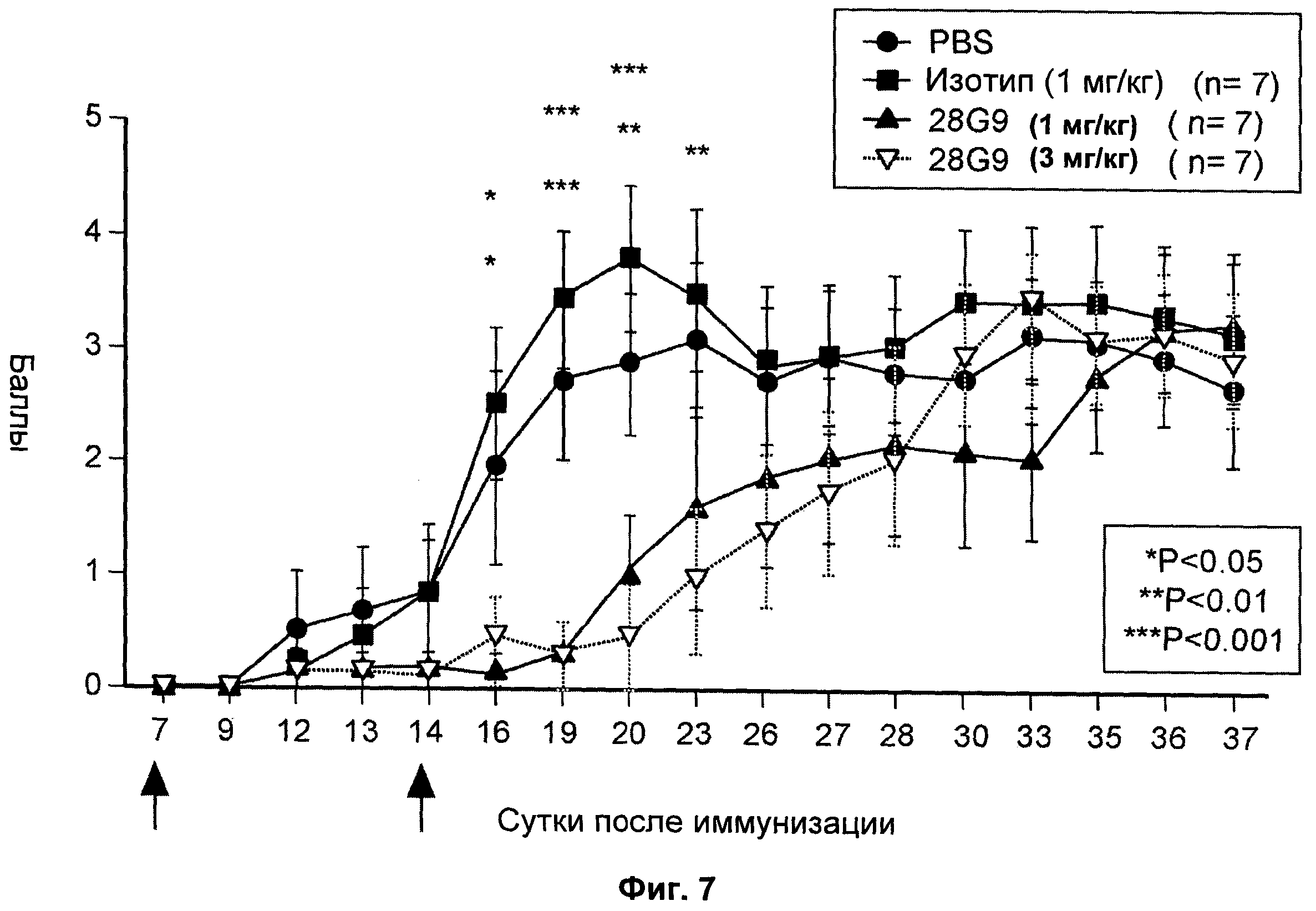
Антагонистические антитела против рецептора il-7 и способы
Конъюгаты "антитело - лекарственное средство" с высокой лекарственной нагрузкой
Модулирующие стабильность линкеры для использования с конъюгатами антитело-лекарственное средство
Способы лечения боли, вызванной остеоартритом, путем введения антагониста фактора роста нервов и содержащих его композиций
Антагонистические антитела против рецептора il-7 и способы
Комбинированные способы лечения антагонистами pd-l1

