Результат интеллектуальной деятельности: ПЛАЗМИДА 40NaGal, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ α-N-АЦЕТИЛГАЛАКТОЗАМИНИДАЗЫ α-AlNaGal, ШТАММ E.coli Rosetta(DE3)/40NaGal - ПРОДУЦЕНТ ХИМЕРНОГО БЕЛКА, ВКЛЮЧАЮЩЕГО АМИНОКИСЛОТНУЮ ПОСЛЕДОВАТЕЛЬНОСТЬ РЕКОМБИНАНТНОЙ α-N-АЦЕТИЛГАЛАКТОЗАМИНИДАЗЫ α-AlNaGal, И СПОСОБ ЕЕ ПОЛУЧЕНИЯ
Вид РИД
Изобретение
Изобретение относится к биотехнологии, в частности к генетической инженерии, и касается способа получения рекомбинантного белка α-N-ацетилгалактозаминидазы морской бактерии Arenibacter latericius (α-AlNaGal), а также плазмиды для его получения и рекомбинантного штамма Escherichia coli. Изобретение позволяет производить высокоактивную рекомбинантную α-N-ацетилгалактозаминидазу для использования в трансфузионной и клинической медицине, генной инженерии и молекулярной биологии.
Фермент α-N-ацетилгалактозаминидазу (ЕС 3.2.1.49.) используют в качестве инструмента для исследования структуры сложных гликоконъюгатов и синтеза новых олигосахаридов с N-ацетамидо-2-дезокси-α-D-галактопиранозой (NAcGal), связанных α-O-гликозидной связью, которую трудно синтезировать химическими методами [1]. α-N-ацетилгалактозаминидаза катализирует гидролитическое отщепление N-ацетамидо-2-дезокси-α-D-галактозидных остатков от невосстанавливающих концов различных сложных углеводов и гликоконъюгатов. Его физиологическими субстратами являются гликолипиды, гликопептиды и гликопротеины, содержащие структуры с O-гликозидным кором, олигосахариды и антигены иммунодоминант эритроцитов группы крови A.
Позднее интерес к ферменту возник в связи с возможностью использования его в биотехнологии создания универсальной крови группы O из донорской крови группы A для решения проблем трансфузионной медицины. Известно, что кровь группы А более широко распространена среди доноров, чем группы B и AB, поэтому интерес для медицины представляют ферменты, проявляющие специфичность к антигену группы A. Проблема заключается в том, что кровь группы A состоит из двух основных подгрупп: A1 и A2 [2]. Эритроциты группы A1 содержат на поверхности примерно в 5 раз больше А антигенов, чем эритроциты подгруппы A2. Кроме того, красные клетки подгруппы A1 имеют гликолипиды с иммунными детерминантами, составленными из повторяющейся A структуры.
На сегодняшний день α-N-ацетилгалактозаминидазы были выделены и всесторонне охарактеризованы лишь в некоторых наземных моллюсках, грибах и некоторых патогенных бактериях наземного происхождения Clostridium perfringens, Ruminococcus torques, а также успешно выделены из печени человека, мыши и цыпленка [3-7].
Однако конверсия эритроцитов A в эритроциты группы O была достигнута лишь с помощью α-N-ацетилгалактозаминидаз из печени цыпленка и из патогенных для человека бактерий Clostridium perfringens, причем конверсия возможна только для эритроцитов крови подгруппы A2 при pH 5,5, т.е. в условиях, травмирующих красные клетки [3, 4]. Такая специфичность не является достаточным условием для полной конверсии A-эритроцитов, поэтому эритроциты, обработанные этими ферментами, остаются не пригодными для переливания реципиентам. К тому же ферменты из вышеназванных источников имеют многомерную структуру с молекулярной массой, достигающей 500 кДа, что затрудняет их использование в молекулярной биологии и биотехнологии.
В 2007 году появилась публикация иностранных коллег, в которой сообщалось о ферменте из патогенного штамма бактерии Elisabethkingia meningosepticum, с помощью которого при pH 7,0 успешно конвертировали как эритроциты A2, так и A1 в эритроциты группы О [7]. Фермент оказался NAD+-зависимым, реализующим механизм гидролиза, неродственный механизму действия классических гликозидгидролаз семейств GH 27 и 36. Данный фермент не имел аналогов среди известных α-галактозидаз и α-N-ацетилгалактозаминидаз и открыл новое 109 семейство гликозидгидролаз в классификации, основанной на общих чертах аминокислотных последовательностей.
Нами была обнаружена α-N-ацетилгалактозаминидаза 109 семейства гликозидгидролаз из непатогенной аэробной морской бактерии Arenibacter latericius 426T, ВКМ В-2137Д, способная инактивировать серологическую активность эритроцитов А обеих подгрупп [8]. Фермент имеет нейтральный pH оптимум действия (7,0-7,3), комфортный для жизнеспособности эритроцитов, и проявляет максимальную активность в присутствии 0,4 М NaCl и 1 мМ Mg2+.
Разработка эффективного способа получения универсальных по групповым свойствам эритроцитов для неотложной трансфузиологической помощи пострадавшим продолжает быть весьма актуальной. Клинические испытания конвертированных эритроцитов на добровольцах продемонстрировали, что ферментативное преобразование эритроцитов человека выполнимо, что конвертированные ферментом эритроциты группы О жизнеспособны и могут функционировать так же, как необработанные эритроциты в соответствии с правилами, принятыми в клинической медицине переливания.
Однако необходимость больших количеств фермента в этих исследованиях, даже с современной эффективной рекомбинантной технологией экспрессии, составляет экономическое препятствие для использования их в медицине переливания, так как на одну упаковку (200 мл) эритроцитов требуется 1-2 г фермента. Кроме того, огромное значение имеет pH-оптимум фермента и узкая субстратная специфичность.
Известен способ получения природного белка α-N-ацетилгалактозаминидазы из дикого штамма-продуцента Flavobacterium sp. 426T, ВКМ В-2137Д, с удельной активностью 22 ед./мг белка [9]. Основными недостатками этого способа получения являются высокие технологические затраты при хранении и выращивании дикого штамма и относительно низкая удельная активность.
Недавно была установлена нуклеотидная последовательность зрелого белка α-N-ацетилгалактозаминидазы Arenibacter latericius 426T, ВКМ В-2137Д (код GenBank HQ 108058). Однако способы получения активного рекомбинантного аналога α-N-ацетилгалактозаминидазы Arenibacter latericius 426T (α-AlNaGal) пока еще не известны. Получение растворимой высокоактивной α-AlNaGal в гетерологичных бактериальных системах сопряжено с рядом трудностей, так как функциональный белок является психрофильным и состоит из двух субъединиц.
Задача изобретения - конструирование рекомбинантного штамма Е. coli, плазмиды, кодирующей синтез рекомбинантного белка α-AlNaGal, и разработка способа его получения.
Поставленная задача решена созданием генетической конструкции в виде рекомбинантной плазмиды 40NaGal и штамма Е.coli Rosetta(DE3)/40NaGal, обеспечивающих индуцируемый синтез с высоким и стабильным выходом активной растворимой рекомбинантной α-N-ацетилгалактозаминидазы α-AlNaGal в периплазму клетки кишечной палочки.
Технический результат заявленного изобретения - получение активной рекомбинантной α-N-ацетилгалактозаминидазы α-AlNaGal с высоким выходом и уровнем очистки.
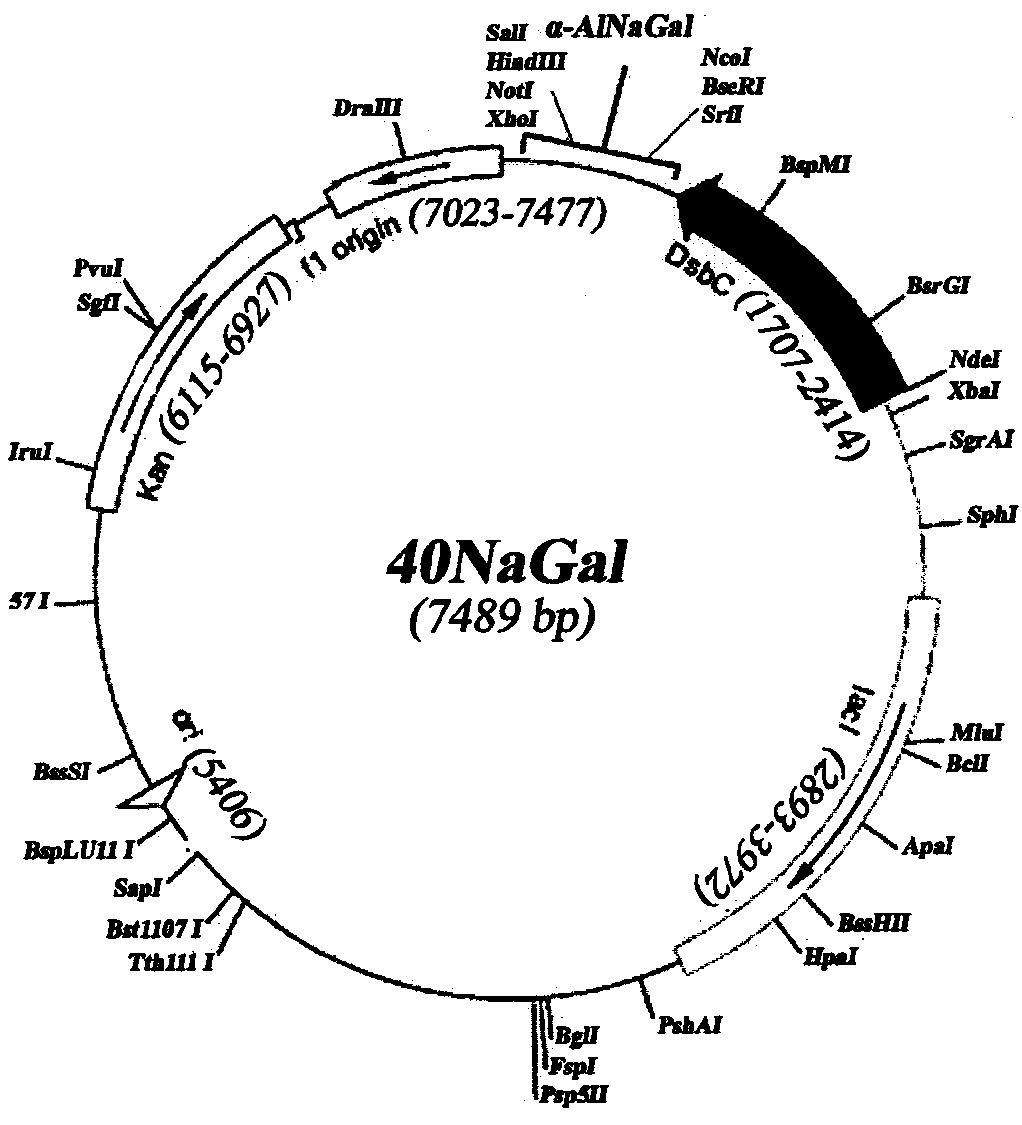
Плазмида 40NaGal имеет 7489 пар оснований (п.о.) и характеризуется наличием NcoI/SalI-фрагмента плазмиды pET-40b(+) (Novagen) и последовательности фрагмента ДНК размером 1299 п.о., адаптированной по N-концу для экспрессии в клетках E.coli, содержащего химерный ген, состоящий из структурной части гена α-AlNaGal и двух специфических последовательностей для протеазы TEV с целью удаления дополнительных аминокислотных остатков с N- и C-концевой части молекулы рекомбинантного белка при необходимости.
На фигуре представлена физическая карта плазмиды 40NaGal и область плазмиды, ответственная за экспрессию рекомбинантного белка α-AlNaGal.
Нуклеотидная последовательность фрагмента плазмиды 40NaGal, фланкированная сайтами NcoI и SaiI, содержит последовательность структурного гена α-AlNaGal с адаптированным N-концом для экпрессии в E.coli, соответствующую открытой рамке считывания для белка α-AlNaGal, и две последовательности с N- и C-конца, специфичные для расщепления протеазой TEV (SEQ ID N 1).
Штамм E.coli Rosetta(DE3)/40NaGal получен трансформацией клеток E.coli Rosetta(DE3) (Novagen) плазмидой 40NaGal с использованием традиционной генно-инженерной технологии [10].
Рекомбинантный штамм E.coli Rosetta(DE3)/40NaGal характеризуется следующими признаками.
Культурально-морфологические признаки.
Клетки штамма образуют крупные круглые с ровными краями выпуклые колонии до 5 мм в диаметре, поверхность колоний гладкая, консистенция слизистая. Пигмент не накапливается. Грамотрицательны, спор не образуют, капсулы не имеют. Колонии хорошо растут на простых питательных средах (LB). При росте в жидких средах образуют интенсивную ровную муть.
Физико-биологические признаки.
Штамм E.coli Rosetta(DE3)/40NaGal видотипичен по своим биохимическим свойствам. Штамм не обладает желатиназной активностью, не ферментирует лизин; расщепляет глюкозу, лактозу, маннит, сахарозу до кислоты и газа. Имеет мутацию в гене lac, обеспечивающую контроль уровня экспрессии, а также трансляцию редких кодонов. Оптимальной для роста является температура 37°C, а для продукции психрофильной α-AlNaGal - 16°C.
Устойчивость к антибиотикам.
Клетки штамма характеризуются устойчивостью к хлорамфениколу (34 мкг/мл) и канамицину (25 мкг/мл).
Патогенность и токсичность.
Рекомбинантный штамм E.coli Rosetta(DE3)/40NaGal не патогенен и не токсичен для теплокровных животных.
Штамм хранится обычным способом в суспензии с глицерином (30%) при -20°C.
Заявляемый способ получения рекомбинантной α-AlNaGal заключается в культивировании клеток штамма E.coli Rosetta(DE3)/40NaGal в питательной среде LB, содержащей канамицин, отделении биомассы от культуральной жидкости, разрушении микробных клеток с последующим выделением целевого продукта водной экстракцией и хроматографической очисткой ферментного препарата. Очистку целевого продукта осуществляют хроматографией на металлоафинной смоле и гель-фильтрацией.
Выход рекомбинантной α-AlNaGal в результате применения описанного способа составляет не менее 10 мг рекомбинантного белка с 1 л культуры с удельной активностью 60 ед./мг по п-нитрофенил-N-ацетил-галактозаминиду, что превышает удельную активность природного аналога из морской бактерии Arenibacter latericius как минимум в 3 раза и рекомбинантного аналога из патогенной бактерии Elizabethkingia meningoseptica - в 5 раз [7].
Рекомбинантный белок α-AlNaGal является гомодимером с молекулярной массой 164 кДа или 96 кДа после удаления аминокислотных довесков протеазой TEV, имеет оптимальную температуру реакции 20-37°C, стабилен до 50°C, оптимум pH 7,0-7,2, и его активность не зависит от присутствия ионов двухвалентных металлов в инкубационной среде. Аминокислотные довески не влияют принципиально на ферментативные свойства белка α-AlNaGal, поэтому стадией их удаления при очистке рекомбинантного белка можно пренебречь.
Рекомбинантная α-AlNaGal может быть успешно использована для удаления групповой специфичности эритроцитов человека группы крови A(II) и получения универсальной донорской крови группы O(I), а также в структурном анализе сложных гликопротеинов и протеогликанов и в синтезе уникальных гликозидов для целей биотехнологии и медицины.
Существенными преимуществами заявляемого способа являются:
- использование штамма-продуцента E.coli Rosetta(DE3)/40NaGal, что позволяет получать при биосинтезе большое количество полноразмерной и высокоактивной рекомбинантной α-AlNaGal;
- использование двухстадийной хроматографической очистки фермента, что позволяет получить чистый рекомбинантный белок за короткое время и с малыми потерями.
Способ получения функционально активного белка на основе использования гена, кодирующего α-N-ацетилгалактозаминидазу морской бактерии α-AlNaGal, иллюстрируется следующими примерами.
Пример 1. Конструирование плазмиды 40NaGal.
Рекомбинантную плазмиду 40NaGal, содержащую структурный ген α-AlNaGal, кодирующий зрелый белок α-N-ацетилгалактозаминидазу A. latericius, и последовательности, специфичные для расщепления протеазы TEV, фланкированные сайтами рестрикции Ncol и Sail, конструируют на основе коммерческой плазмиды рЕТ-40b(+) (Novagen).
Фрагмент ДНК, содержащий полноразмерный ген α-AlNaGal, получают при помощи полимеразной цепной реакции с использованием геномной ДНК штамма морской бактерии Arenibacter latericius 426T, ВКМ В-2137Д, в качестве матрицы и пары праймеров Nac40TEV_NcoFn Nac40TEV_SalIR, где Nac40TEV_NcoF - праймер, специфичный по отношению к N-концевой последовательности α-AlNaGal, включающий последовательность для протеазы TEV, Nac40TEV_SalIR - обратный праймер, специфичный по отношению к C-концевой последовательности α-AlNaGal, включающий последовательность для протеазы TEV:
Nac40TEV_NcoF: 5'-ttaaccatgGAAAATCTTTATTTTCAGGGTGGGGCTAAGTACATGGGCGGTTTTTCTGCT-3'
Nac40TEV_SalIR: 5'-ttaagtcgacACCCTGAAAATAAAGATTTTCGCTTACAATATCTAATGGTGCAGTGGT-3'
Данную реакцию проводят в следующих условиях: 10х Encyclo буфер, 50х смесь полимераз Dream Taq (Ферментас), 50х смесь dNTP (10 mM каждого), смесь праймеров (5 µM каждого), 50 нг ДНК. Процесс амплификации состоит из следующих стадий: прогревание при 95°C - 2 мин, 35 циклов ПЦР (15 сек - 95°C, 1 мин - 72°C) и инкубация 10 мин при 72°C. После амплификации фрагмент ДНК очищают электрофоретически в 1% агарозном геле. Фрагмент (1 мкг) обрабатывают рестриктазами NcoI и SalI в оптимальном буфере (Fermentas) в течение 3 час, затем ферменты удаляют из реакционной среды по стандартной методике фенолом (1:1). В водную фракцию, содержащую фрагмент, добавляют 1/10 объема 0,3 М ацетата натрия, pH 5,2, и 1/2 объема изопропанолового спирта и оставляют при -20°C в течение 30 мин. Затем центрифугируют при 14000 об/мин в течение 20 мин, осадок промывают 75% этанолом и высушивают при комнатной температуре. Осадок растворяют в 20 мкл деионизованной воды.
5 мкг плазмидной ДНК рЕТ-40b(+) обрабатывают рестриктазами NcoI и SalI в соответствии с методикой, описанной выше, и из полученного гидролизата выделяют векторную часть плазмиды в 1% геле легкоплавкой агарозы.
Полученный фрагмент и векторную часть плазмиды pET-40b(+) сшивают при помощи лигазной реакции в 50 мкл буфера для лигирования согласно инструкции (Fermentas). 10 мкл реакционной смеси используют для трансформации компетентных клеток E.coli Rosetta(DE3). Трансформанты высевают на LB-агар, содержащий 25 мкг/мл канамицина. После инкубирования в течение 12 час при 37°C клоны отсевают, выделяют плазмидную ДНК и анализируют на наличие мутаций при помощи автоматического секвенирования. Отбирают ДНК, содержащую необходимую последовательность, представляющую собой плазмиду 40NaGal размером 7489 п.о.
Пример 2. Получение штамма E.coli Rosetta(DE3)/40NaGal, трансформированного плазмидой 40NaGal - продуцента химерного белка, включающего аминокислотную последовательность рекомбинантной α-AlNaGal.
Штамм-продуцент получают путем трансформации клеток штамма E.coli Rosetta(DE3) рекомбинантной плазмидой 40NaGal. Ночную культуру (0,5 мл LB) штамма-продуцента рекомбинантной α-AlNaGal выращивают в литровой колбе в жидкой среде LB, содержащей на литр 10 г бакто-триптона, 5 г бакто-дрожжевого экстракта и 10 г NaCl, 25 мг/мл канамицина, pH 7,7, на шейкере при 200 об/мин при температуре 37°C в течение 2 час до оптической плотности 0,6-0,8 (OD600), затем добавляют индуктор экспрессии IPTG до конечной концентрации 0,2 мМ и инкубируют далее при 16°C в течение 12 час.
Для определения продуктивности штамма клеточные водные экстракты анализируют электрофорезом в 12% полиакриламидном геле с додецилсульфатом натрия. Гель окрашивают Кумасси R-250 по стандартной методике и определяют относительное количество белка в полосе целевого продукта. Содержание рекомбинантного белка в растворимой клеточной фракции составляет не менее 30% от всех белков этой фракции.
Пример 3. Выделение и характеристика рекомбинантной α-AlNaGal.
Штамм-продуцент рекомбинантной α-AlNaGal - E.coli Rosetta(DE3)/40Gal, инкубируют в литровой колбе в жидкой среде LB, содержащей на литр 10 г бакто-триптона, 5 г бакто-дрожжевого экстракта и 10 г NaCl, 0,2 мМ IPTG, 25 мг/мл канамицина, pH 7,7, на шейкере при 250 об/мин в течение 12 час при 16°C. Бактериальные клетки осаждают на проточной центрифуге при 5000 об/мин в течение 10 мин. Суспензию клеток дезинтегрируют в 100 мл буфера А (0,01 М NaH2PO4, pH 7,7, 0.01%) NaN3) в течение 5×30 сек, охлаждая во льду. Затем центрифугируют при 5000 об/мин в течение 10 мин.
Надосадочную жидкость собирают и помещают на колонку с металлоафинной смолой (Qiagen), предварительно уравновешенную буфером А. Элюцию белка проводят буфером А, содержащим 50 мМ ЭДТА. Активные фракции собирают и концентрируют до 3 мл в буфере А на ионообменной смоле ДЕАЕ-целлюлоза (Watman) и инкубируют с протеазой TEV (Invitrogen) при 21°C в течение 12 час. Раствор белка наносят на колонку для гель-фильтрации с Sephacryl S-200 HR (Sigma). Выход рекомбинантного белка составляет около 10 мг с 1 л культуры.
Полученный рекомбинантный полипептид определяют по первым 10 аминокислотам на автоматическом секвенаторе. Проведенное секвенирование препарата рекомбинантной α-AlNaGal, выделенной из клеток штамма Е.coli Rosetta(DE3)/40NaGal, показало, что N-концевая аминокислотная последовательность (GAKYMGGFSAPKLDT) соответствует первым 15 аминокислотам полноразмерного природного белка α-N-ацетилгалактозаминидазы морской бактерии Arenibacter latericius 426T, ВКМ В-2137Д, - аналога α-AlNaGal.
Активность α-N-ацетилгалактозаминидазы определяют по расщеплению п-нитрофенил-альфа-N-ацетилгалактозаминида. Реакционная смесь в объеме 400 мкл содержит 10 мМ NaH2PO4 (pH 7,2), 3 мМ субстрата и фермент. После 20 мин инкубации при 20°C реакцию останавливают добавлением 0,6 мл 1 М Na2CO3. Количество образовавшегося в процессе ферментативной реакции продукта определяют спектрофотометрически при 400 нм. За единицу активности принимают количество фермента, катализирующего освобождение 1 мкМ субстрата в течение 1 мин инкубации. Удельную активность выражают в единицах активности фермента на 1 мг белка. Концентрацию белка в растворе определяют по методу Брэдфорда.
Полученные данные по физико-химическим характеристикам и ферментативной активности продукта экспрессии искусственного химерного гена α-AlNaGal в клетках штамма E.coli Rosetta(DE3)/40NaGal свидетельствуют о соответствии исследуемого полипептида его природному аналогу.
Как следует из приведенных примеров, заявляемая группа изобретений позволяет получать активную рекомбинантную α-N-ацетилгалактозаминидазу α-AlNaGal с высоким выходом при относительно простой и надежной технологии.
Заявленное изобретение позволяет:
- с помощью использования штамма-продуцента E.coli Rosetta(DE3)/40NaGal получать активную рекомбинантную a-N-ацетилгалактозаминидазу α-AlNaGal;
- использование штамма-продуцента E.coli Rosetta(DE3)/40NaGal позволяет получать при биосинтезе большое количество полноразмерной рекомбинантной α-N-ацетилгалактозаминидазы α-AlNaGal;
- использование металлоафинной и гель-фильтрационной хроматографии при очистке фермента из водного экстракта клеток штамма-продуцента позволяет получать фермент с чистотой более 98%.
Литература
1. Naundorf A., Ajisaka K. Purification of α-N-acetyl-galactosaminidase from Aspergillus niger and its use in the synthesis of GalNAc-α-(1→O)-serine // Enz. Microbial. Technol. - 1999. - Vol.25, N 6. - P.483-488.
2. Clausen H., Hakomori S.-i. ABH and related histo-blood group antigens; immunochemical differences in carrier isotypes and their distribution // Vox Sang. - 1989. - Vol.56, N 1. - P.1-20.
3. Hata J., Dhar M., Mitra M., Harmata M., Haibach P., Sun P., Smith D. Purification and characterization of N-acetyl-α-D-galactosaminidase from Gallus domesticus II Biochem. Int. - 1992. - Vol.28, N 1. - P.77-86.
4. Hsieh H.Y., Mitra M., Wells D.C., Smith D. Purification and characterization of α-N-acetylgalactosaminidase from Clostridium perfringens IIIUBMB Life. - 2000. - Vol.50, N 2. - P.91-97.
5. Hoskins L.C., Boulding E.T., Larson G. Purification and characterization of blood group A-degrading isoforms of α-N-acetylgalactosaminidase from Ruminococcus torques strain IX-70 // J. Biol. Chem. - 1997. - Vol.272, N 12. - P.7932-7939.
6. Kadowaki S., Ueda Т., Yamamoto K., Kumagai H., Tochikura T. Isolation and characterization of a blood group A substance-degrading α-N-acetylgalactosaminidase from an Acremonium sp. // Agric. Biol. Chem. - 1989. - Vol.53, N 1. - P.111-120.
7. ChengYu Y.U., Hua X.U., LiSheng W., JianGeng Z., YangPei Z. Human RBCs blood group conversion from A to О using a novel -N-acetylgalactosaminidase of high specific activity // Chinese Science Bulletin. - 2008. - Vol.53, N 13. - P.2008-2016.
8. Бакунина И.Ю., Кульман P.A., Лихошерстов Л.М., Мартынова М.Д., Недашковская О.И., Михайлов В.В., Елякова Л.А. α-N-Ацетилгалактозаминидаза из морской бактерии Arenibacter latericius КММ 426Т, устраняющая групповую специфичность A-эритроцитов // Биохимия. - 2002.- Т.67, N 6. - С.830-837.
9. RU 2141526 С1, 20.11.1999.
10. Sambrook J., Fritsch E.F., Maniatis Т. // Molecular Cloning. A. Laboratory Manual. 2bd ed. Cold Spring Harbor, NY, 1989.


Адъювант
Средство для лечения атопического дерматита
Биологически активный продукт из бурой водоросли, биологически активная добавка к пище, безалкогольный напиток, парфюмерно-косметическое средство
Способ интегральной оценки состояния загрязнения морской и пресной воды
Плазмида 40ph, определяющая синтез щелочной фосфатазы cmap, штамм e.coli rosetta(de3)/40ph - продуцент химерного белка, включающего аминокислотную последовательность рекомбинантной щелочной фосфатазы cmap, и способ ее получения
Способ коррекции эндотелиальной дисфункции

