Результат интеллектуальной деятельности: ВАКЦИНА ПРОТИВ HEV
Вид РИД
Изобретение
Вакцина против HEV
Настоящее изобретение относится к вакцине, содержащей белок открытой рамки считывания 2 (ORF2) вируса гепатита E (HEV) свиней и фармацевтически приемлемый носитель, для применения в защите положительных по MDA поросят против распространения HEV свиней.
На протяжении последнего десятилетия во всем мире наблюдалось увеличение инфекции гепатита E человека. В настоящее время, ежегодно сообщается о приблизительно 20 миллионах случаев инфекции HEV , наблюдалось более трех миллионов случаев острого гепатита и 57000 связанных с HEV случаев смерти.
Вирус гепатита E, этиологический агент гепатита E, принадлежит к роду Hepevirus, единственному члену Hepeviridae. Он представляет собой безоболочечный вирус с диаметром приблизительно 27-34 нм. Геном, положительный смысловой одноцепочечный РНК-геном, имеет длину приблизительно 7,2 т.п.о. Геном содержит три различных открытых рамки считывания (ORF); ORF1, кодирующую неструктурный белок, ORF2, кодирующую полипептид вирусного капсида, и ORF3, кодирующую малый фосфобелок, который связывается с цитоскелетом. Существует только один серотип. Тем не менее, принимая во внимание высокий уровень геномного разнообразия между различными изолятами, проведено подразделение на четыре главных генотипа (генотипы 1-4).
HEV является основной причиной гепатита не-A, не-B у человека. Основные вспышки обнаружены в развивающихся странах, особенно в тропиках и субтропиках. В промышленно развитых странах, инфекции HEV наблюдают в районах, где HEV не является эндемичным, даже в случаях, когда пациенты могут не являться связанными с инфицированными HEV индивидуумами. Инфекцию HEV генотипов 1 и 2 наиболее часто наблюдают у индивидуумов в возрасте между 15 и 35 годами. Инфекцию HEV генотипов 3 и 4 наиболее часто наблюдают у людей с иммунной недостаточностью и иммунодефицитом, и у реципиентов трансплантатов органов. По-видимому, большинство случаев инфекции HEV 3 и 4 у здоровых людей являются субклиническими. (Ling Lu et al., Rev. Med. Virol. 16: 5-36 (2006)).
Вакцины против инфекции HEV человека разрабатывают уже в течение многих лет. В большинстве случаев, такие вакцины основаны на белке ORF 2 HEV или его фрагментах, экспрессированных, например, в бакуловирусных системах экспрессии и бактериальных системах экспрессии. Опубликовано, что такие вакцины являются успешными. (Kamili, S., Virus Research 161: 93-100 (2011), Amini-Bavil-Olyaee, S. et al., Future Virol. 4: 143-154 (2009), Feng-Cai Zhu et al., The Lancet, Published Online August 23, 2010 DOI:10.1016/S0140-6736(10)61030-6.)
В настоящее время считается, что существует зоонозная связь между (по меньшей мере) HEV свиней генотипов 3 и 4 и заболеванием человека. В частности, свиньи являются основным резервуаром HEV генотипов 3 и 4. (Pavio, N. et al., Curr. Opin. In Virology 10: 34-41 (2015), Purcell, R.H. and Emerson, S.U., J. Inf. Dis. 202: 819-821 (2010), Meng, X.J. et al., P.N.A.S. 94: 9860-9865 (1997), WU, J.C. et al., J. Med. Virol. 60: 166-171 (2000), Matsuda, H. et al., J. Inf. Dis. 188, 944 (2003), Matsubayashi, K. et al., Transfusion 48: 1368-1375 (2008), Mishiro, S., Uirusu 54: 243-248 (2004), Dalton, H.R. et al., Lancet Inf. Dis. 8: 698-709 (2008)). Инфекция людей HEV генотипов 3 и 4 является в большинстве случаев вызванной загрязненными пищевыми продуктами, (например, недоваренным или сырым мясом свиней), прямым воздействием свиней и свиных фекалий или загрязнением окружающей среды.
Принимая во внимание эту явную иллюстрацию зоонозного характера HEV, кажется полезным вакцинировать свиней (в дополнение к человеку, или даже вместо человека) против HEV свиней, чтобы исключить HEV свиней в качестве источника инфекции человека. Существует несколько патентных заявок, которые относятся к белку ORF2 вируса гепатита E свиней и к его использованию для детекции вируса и в качестве вакцины. Целью заявки PCT WO 02/089733 и Европейской заявки на патент EP1000168 в основном является вакцинация человека против HEV свиней с использованием полноразмерного вируса HEV свиней или субъединичных вакцин на основе HEV свиней. В EP1000168 предлагается использование HEV свиней в качестве живого ослабленного вируса для вакцинации человека. Вакцинацию свиней в основном предлагают в качестве способов получения антител против HEV свиней для диагностических целей, среди прочего, для проверки того, что органы свиней для использования в ксенотрансплантации являются свободными от HEV. В ней описано, каким образом, с целью получения антител против HEV свиней, свободных от указанного патогена поросят в возрасте 2 недель инфицируют HEV свиней для получения антител против этой инфекции. Такие поросята являются, среди прочего, свободными от происходящих от матери антител против HEV. Однако, в этих патентных заявках не представлены какие-либо примеры вакцинации свиней, не говоря уже о том, что они связаны с возрастом животных, подлежащих вакцинации.
Как правило, может казаться привлекательной вакцинация животных против заболевания так скоро, как только возможно, после их рождения. На практике, однако, и особенно в случае HEV, специалист в данной области не рассматривает это в качестве подходящего способа.
Причиной является следующее: распространенность антител против HEV у свиней является необычайно высокой. Во всем мире, среднюю общую распространенность оценивают как 50% (Vasickova, P, et al., Veterinarni Medicina 52: 365-384 (2007)), но она может быть значительно более высокой во многих странах. Например, в 2013 г. обнаружено, что распространенность антител против HEV у свиней в UK составляет 92,8% (Grierson, S., et al, CDC EID Journal 21 number 8, August 2005). Таким образом, можно заключить, что несомненно, большинство свиноферм являются инфицированными HEV, и что риск инфекции является очень высоким. Вследствие этого, несомненно, большинство свиней испытают инфекцию HEV, и у них образуются антитела против HEV ко времени их первой беременности. Это, в свою очередь, означает, что несомненно, большинство рожденных поросят будут иметь материнские антитела (MDA) против HEV, т.е. большинство рожденных поросят являются положительными по MDA против HEV. Эти материнские антитела защищают поросят в течение первых месяцев жизни. Затем MDA медленно исчезают, таким образом, из-за высокого риска инфекции, в конкретный момент времени поросята являются чувствительными и следовательно, испытают инфекцию HEV, и следовательно, могут начать распространять вирус HEV в окружающую среду до развития своей собственной защиты против HEV, среди прочего, в форме антител против HEV. Следовательно, риск инфекции HEV на свинофермах будет оставаться высоким. В индустрии свиноводства, поросят отлучают от матери между 3 и 4 неделями после рождения. Это означает, что они имеют очень высокие титры MDA в возрасте трех или четырех недель. Затем наблюдают медленное уменьшение титра MDA. В среднем, если отлучение от матери осуществляют через 3 недели после рождения, обычно самое раннее приблизительно через 7-13 недель наблюдают увеличение количества поросят, которые становятся инфицированными HEV, и у которых развивается их собственная иммунная реакция против этой инфекции HEV. Часто проходит между 10 и 25 неделями до того, как количество MDA у поросят уменьшается до уровня, который не предотвращает инфекцию HEV у 90% животных (см. раздел примеры, пример 3 и среди прочего, Andraud, M. et al., PLOS one 9: e105527 (2014)). До этого 10-25-недельного периода MDA защищают определенный процент животных против инфекции, поскольку MDA связываются с вирусом и удаляют его до того, как он сможет инфицировать клетки и запускать иммунную систему.
Следовательно, специалист в данной области может прийти к заключению, что вакцинация стада поросят до 10-25 недель не будет иметь эффекта у высокого процента поросят по той простой причине, что многие поросята в стаде все еще будут иметь достаточно высокие титры MDA, чтобы создавать помехи для иммуногенных компонентов вакцины против HEV до того, как она сможет запустить иммунную систему. Ингибирование вакцинации посредством MDA является общим феноменом, как для вакцин для человека, так и для ветеринарных вакцин (Stefan Niewiesk in Frontiers in Immunology, review article, 16 September 2014; doi:10.3389/fimmu.2014.00446). Таким образом, вакцинацию в этот период нельзя рассматривать как полезную для этих поросят. Однако, с другой стороны, те поросята в стаде, которые уже имеют уменьшенный уровень MDA, являются чувствительными к инфекции HEV и, таким образом, должны быть вакцинированы.
Практически/экономически невозможно ежесуточно определять уровень MDA у каждого индивидуального поросенка и ежесуточно решать, какие индивидуальные поросята должны быть вакцинированы.
Это долгое, но изменчивое время существования MDA против HEV является хорошо известным феноменом, и с точки зрения вакцинации, это является хорошо известной проблемой. Эта проблема существует не только для MDA против HEV, это является проблемой для вакцинации в раннем возрасте в целом. (Andraud, M. (см. выше), Niewiesk, S., Frontiers in Immunology 5, Article 446 (2014), Pastoret, P.P., Journal of Comparative Pathology Volume 137, Supplement 1, Pages S2-S3 (2007)). В Andraud, M. (см. выше) описаны вопрос MDA и эта проблема следующим образом: «Несмотря на то, что вакцинация может преодолевать эту проблему (риск инфекции во время исчезновения MDA), в нескольких исследованиях представлены доказательства антагонистических эффектов между индуцированным вакциной иммунитетом и материнскими антителами посредством ингибирования защиты вакцины, что потенциально может ухудшать динамику инфекции». В Niewiesk, S., (см. выше) даже описано для 11 различных вакцин для человека и 17 различных ветеринарных вакцин, что их ингибируют материнские антитела.
Целью настоящего изобретения является преодоление проблемы присутствия MDA при вакцинации поросят против HEV.
В настоящее время неожиданно обнаружено, что вакцина, содержащая иммуногенно эффективное количество фрагмента белка ORF2 HEV, перекрывающего от самой меньшей области от аминокислоты 125 до 607, до самой большой области от аминокислоты 112 до 660, является способной прорываться через материнский иммунитет и запускать иммунную систему положительных по MDA против HEV поросят, в частности, положительных по MDA против HEV поросят в возрасте до 10 недель.
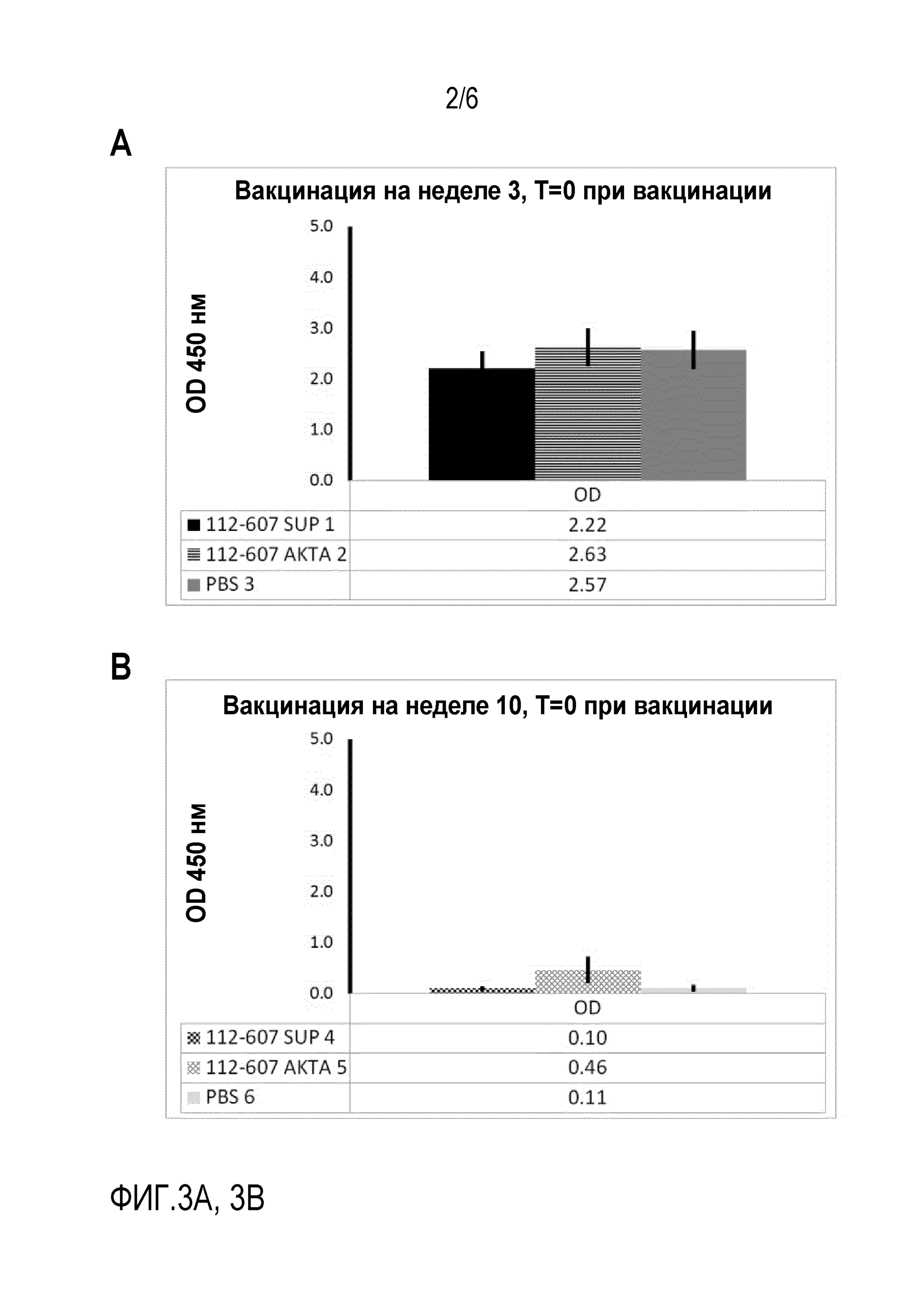
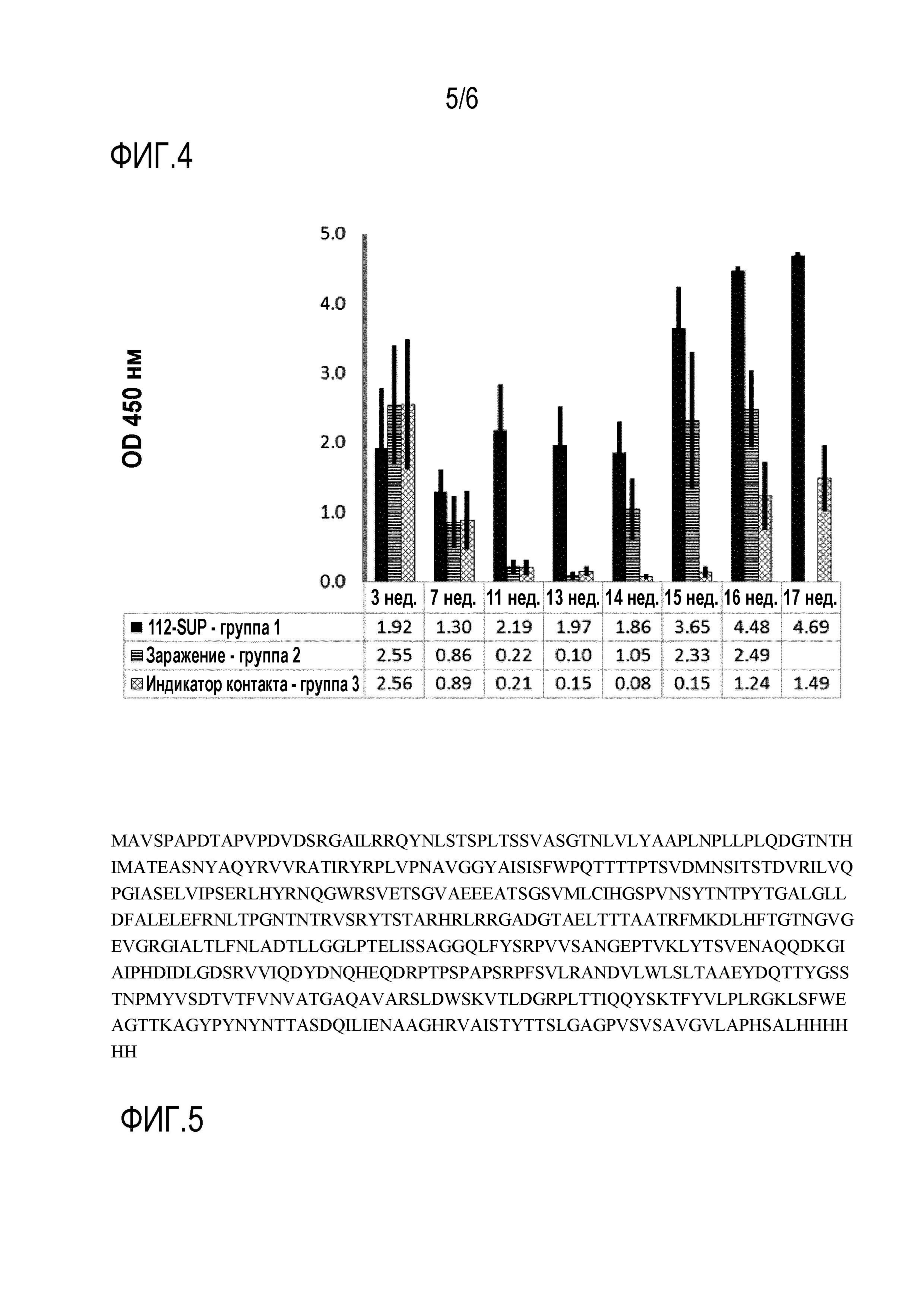
На фигуре 3 проведено сравнение иммунного ответа против HEV у положительных по MDA поросят в возрасте 3 недель (фиг. 3C) и у отрицательных по MDA поросят в возрасте 10 недель (фиг. 3D), измеренное через четыре недели после вакцинации с использованием фрагмента ORF2 HEV по изобретению. Как следует из фигуры, неожиданно, титр антител против HEV, индуцированный у положительных по MDA поросят через четыре недели после вакцинации, является статистически сходным с титром антител против HEV у отрицательных по MDA поросят через четыре недели после вакцинации. Даже через 9 недель после вакцинации, титр антител против HEV у положительных по MDA поросят является только немного более низким, чем титр антител против HEV у отрицательных по MDA поросят (Фигура 3E, 3F).
Белок, кодируемый ORF2 HEV, представляет собой белок капсида HEV 72 кДа. Сравнение между 11 различными последовательностями ORF2 HEV показывает что, несмотря на то, что идентичность последовательности не во всех случаях составляет 100%, длина различных последовательностей ORF2 является практически идентичной во всех случаях. (Meng, X-J. et al., Proc. Natl. Acad. Sci. U S A. 94: 9860-9865 (1997).
ORF2 HEV начинается с нуклеотида (н.) 5147, продолжается 1980 оснований до окончания на н. 7127, и кодирует главный структурный белок из 660 аминокислот (88 кДа), который экспрессируется внутри клетки, так же как на поверхности клетки. Он синтезируется в форме предшественника и подвергается процессингу посредством отщепления сигнальной последовательности до зрелого белка, способного к самоассоциации и гликозилированию. При экспрессии кодирующей последовательности ORF2 в клетках насекомых Sf9 с использованием бакуловирусной системы получают стабильные белковые продукты с расчетной молекулярной массой 72 кДа и 59-62 кДа. Полноразмерные белки из клеток насекомых являются нерастворимыми, в то время как усеченные белки, содержащие аминокислоты между положениями 112-607 и положениями 112-660, собираются в вирусоподобные частицы. Известно, что минимальным требованием для формирования VLP является присутствие аминокислот от положения 125 до 601. (Surjit, M. et al. Journ. Virol, 78: 320-328 (2004), Li, T-C. et al., Journ. Virol. 79, 12999-13006 (2005)). Однако, минимальный нейтрализующий домен ORF2 HEV картирован на аминокислотных остатках 458-607 (Zhou, Y.H. et al., Vaccine 22: 2578-2585 (2004), Ahmad, I. et al., Virus Res. 161: 47-58 (201)). По этой причине, вакцина по изобретению должна содержать по меньшей мере фрагмент белка ORF2 HEV, включающий C-концевые аминокислотные остатки из положений 458-607. В то же самое время, фрагмент белка ORF2 HEV должен содержать N-концевые аминокислотные остатки из положений 125-457, чтобы являться способным формировать VLP.
Таким образом, фрагмент ORF2 HEV в вакцине для использования по изобретению должен содержать по меньшей мере аминокислотные остатки из положений 125-607. Однако, в то же самое время, фрагмент ORF2 HEV не должен содержать дополнительных N-концевых аминокислот из области между 1 и 111, поскольку такие фрагменты белка будут менее растворимыми, особенно при получении в системе экспрессии в клетках насекомых.
Таким образом, предпочтительно, минимальный размер этого фрагмента содержит аминокислотные остатки из положений 112-607, тогда как в то же самое время, максимальный размер этого фрагмента содержит аминокислотные остатки из положений 112-660.
Пример области между аминокислотами 112 и 607 белковой последовательности ORF2 HEV представлен на фигуре 5 (SEQ ID NO: 1).
Термин «иммуногенно эффективное количество», в рамках изобретения, относится к количеству фрагмента белка ORF2 HEV, которое является необходимым для индукции иммунного ответа у поросят до такой степени, что он уменьшает эффекты, вызванные инфекцией HEV дикого типа, по сравнению с эффектами, вызванными инфекцией HEV дикого типа у не иммунизированных поросят.
Следует учитывать, что основной целью вакцинации поросят раннего возраста против HEV является не столько защита поросят от клинических симптомов, сколько уменьшение количества вируса, распространяемого инфицированными поросятами. Это обусловлено тем фактом, что с одной стороны, клинические признаки у инфицированных HEV поросят колеблются от умеренных до отсутствующих, и с другой стороны, конечной целью является защита людей от получения инфекции HEV посредством контакта с фекалиями инфицированных свиней (среди прочих, фермеров) или инфицированным мясом свиней. Таким образом, основной целью вакцинации поросят является уменьшения риска инфекции на свинофермах.
По этой причине специалисту в данной области понятно, что в основном «иммуногенно эффективное количество», в рамках изобретения, относится к количеству фрагмента белка ORF2 HEV, необходимому для индукции иммунного ответа у поросят для защиты против системной инфекции. Эта защита против системной инфекции, в свою очередь, уменьшает количество вируса, распространяемого вакцинированными поросятами после того, как они испытают инфекцию HEV, по сравнению с количеством вируса, распространяемого невакцинированными поросятами после того, как они испытают инфекцию HEV.
Полностью в компетенции специалиста в данной области находится определение того, является ли лечение иммунологически эффективным (т.е., введено ли иммуногенно эффективное количество вакцины), например, посредством проведения экспериментального заражения инфекцией как вакцинированных, так и не вакцинированных животных, и затем определения серологических параметров животных-мишеней, и посредством измерения количества распространяемого вируса, например, посредством отбора ректальных мазков, с последующим сравнением этих обнаружений в двух группах заражения.
Количество введенного фрагмента белка ORF2 зависит от способа введения, присутствия адъюванта, количества вакцинаций и момента введения. Просто в качестве примера; в зависимости от используемого адъюванта, количество около 5 микрограмм белка ORF2 HEV/дозу, введенное в форме однократной внутримышечной вакцинации, является очень хорошо способным индуцировать защитный иммунный ответ у поросят в возрасте трех недель, имеющих высокий титр MDA против HEV.
Предпочтительно, вакцина для использования в соответствии с настоящим изобретением отличается тем, что указанная вакцина содержит по меньшей мере 10 микрограмм/дозу указанного фрагмента белка.
Более предпочтительно, вакцина для использования в соответствии с настоящим изобретением содержит по меньшей мере 20 микрограмм/дозу указанного фрагмента белка.
Предпочтительно, вводят количество приблизительно 15 микрограмм белка ORF2 HEV/дозу, более предпочтительно, даже 20, 30, 40, 50, 60, 70, 80, 90 или даже 100 микрограмм/дозу, в таком порядке предпочтения. Количества более, чем 100 микрограмм/дозу, хотя и являются возможными, как не могут много добавить к уровню защиты, так и не могут являться привлекательными с точки зрения стоимости.
Для целей настоящего изобретения, однократная вакцинация представляет собой вакцинацию, которая имеет место только один раз (в отличие от режима примирующей-бустер вакцинации).
Для целей настоящего изобретения, положительные по MDA поросята представляют собой поросят, имеющих титр антител MDA, имеющий оптическую плотность, равную или превышающую OD(450 нм) 0,25, при измерении в соответствии со способом ELISA, описанным в примере 4.
Таким образом, первый вариант осуществления настоящего изобретения относится к вакцинам, содержащим иммуногенно эффективное количество фрагмента белка открытой рамки считывания 2 вируса гепатита E (ORF2 HEV), перекрывающего от самой меньшей области от аминокислоты 125 до аминокислоты 607, и до самой большой области от аминокислоты 112 до аминокислоты 660, и фармацевтически приемлемый носитель, для использования в защите положительных по MDA поросят против распространения HEV.
Вакцину по изобретению уже можно успешно использовать для поросят в возрасте 2-4 недель. Это обеспечивает преимущества с точки зрения экономики и гуманного обращения с животными, поскольку в этом возрасте поросят часто вакцинируют с использованием вакцины против других заболеваний свиней. Эти вакцины можно, таким образом, вводить в тот же самый момент или около него.
Примеры фармацевтически приемлемых носителей, пригодных для использования в вакцине для применения по изобретению, хорошо известны в данной области и содержат, например, стерильную воду, солевой раствор, водные буферы, такие как PBS и т.п. Кроме того, вакцина для применения по изобретению может содержать другие добавки, такие как адъюванты, стабилизаторы, антиокислители и другие, как описано ниже.
На фигуре 3 показано, что однократная вакцинация с использованием композиции по изобретению уже обеспечивает хорошую защиту против HEV у поросят, особенно против распространения потомства вируса, так что в принципе, бустер-вакцинации не требуется (см. также Пример 7, таблицы A и B ).
Однако, даже более удивительно, в настоящее время обнаружено, что если вакцинированную группу положительных по MDA поросят и вакцинированную группу отрицательных по MDA поросят подвергать второй вакцинации (бустер-вакцинации) через несколько недель после первой вакцинации, титр в обеих группах становится даже выше и достигает насыщения на практически одинаковом титре в обеих группах. Это показывает, что когда поросят как примируют, так и подвергают бустер-инъекции с использованием фрагментов белка ORF2 HEV, как описано по изобретению, они являются даже более защищенными против распространения HEV.
Таким образом, чтобы получить даже более высокий и длительный уровень защиты, можно проводить бустер-введение. В принципе, такая бустер-вакцина может являться наиболее эффективной при введении где-то между 3 и 15 неделями, предпочтительно, между 3 и 10 неделями после первой вакцинации. Более предпочтительно, бустер-введение можно проводить приблизительно через пять недель после первой вакцинации.
Бустер-вакцина можно также содержать количество по меньшей мере 10 микрограмм белка ORF2 HEV/дозу. Предпочтительно, вводят количество приблизительно 15 микрограмм белка ORF2 HEV/дозу, более предпочтительно, даже 20, 30, 40, 50, 60, 70, 80, 90 или даже 100 микрограмм/дозу, в таком порядке предпочтения.
Таким образом, другой вариант осуществления относится к вакцинам, содержащим по меньшей мере 10 микрограмм/дозу фрагмента белка открытой рамки считывания 2 вируса гепатита E (ORF2 HEV), перекрывающего от самой меньшей области от аминокислоты 125 до аминокислоты 607, и до самой большой области от аминокислоты 112 до аминокислоты 660, и фармацевтически приемлемый носитель, для использования в качестве бустер-вакцины для защиты положительных по MDA поросят, вакцинированных с использованием примирующей вакцины против распространения HEV не далее, чем за 15 недель до вакцинации бустер-вакциной.
В более предпочтительной форме этого варианта осуществления, примирующая вакцина представляет собой вакцину, содержащую по меньшей мере 10 микрограмм/дозу фрагмента белка ORF-2 HEV, перекрывающего от самой меньшей области от аминокислоты 125 до аминокислоты 607, и до самой большой области от аминокислоты 112 до аминокислоты 660, и фармацевтически приемлемый носитель.
Фрагменты белка ORF2 HEV, как описано по изобретению, можно легко получать посредством экспрессии фрагмента ДНК, кодирующего эти фрагменты. Подходящими системами экспрессии, хорошо известными в данной области, являются, например, системы экспрессии на основе бактерий, бакуловирусов, дрожжей и эукариотических клеток. Такие системы экспрессии подробно описаны в данной области и являются коммерчески доступными. В Kamili (см. выше) процитированы, например, примеры экспрессии ORF2 HEV в системах экспрессии на основе как бакуловирусов, так и бактерий.
Системы экспрессии на основе бакуловирусов и бактерий могут являться предпочтительными системами с учетом простоты их использования и их относительно большого выхода. Из этих двух, система экспрессии на основе бакуловирусов может являться немного более предпочтительной.
В разделе Примеры представлены примеры экспрессии фрагментов белка ORF2 HEV, перекрывающих от самой меньшей области от аминокислоты 112 до аминокислоты 607, и до самой большой области от аминокислоты 112 до аминокислоты 660, в системе экспрессии на основе бакуловирусов.
Вакцина для использования по изобретению предпочтительно содержит адъювант. Общепринятыми адъювантами, хорошо известными в данной области для использования в вакцинах для свиней, являются, например, неполный адъювант Фрейнда, витамин E, неионные блок-полимеры, мурамил-дипептиды, Quill A(R), минеральное масло например, Bayol(R) или Markol(R), растительное масло и Carbopol(R) (гомополимер) или Diluvac(R) Forte.
Предпочтительно, вакцина для использования по изобретению содержит адъювант вода-в-масле или масло-в-воде. Из них, адъюванты масло-в-воде являются немного более предпочтительными. Масляный компонент в адъювантах вода-в-масле и масло-в-воде может представлять собой минеральное масло. Однако, в настоящее время существует тенденция заменять минеральные масла биоразлагаемыми маслами, например, такими как сквалан, сквален и токоферол.
Таким образом, вакцина для использования по изобретению, более предпочтительно, содержит адъювант масло-в-воде.
Вакцина может также содержать так называемый «носитель». Носитель представляет собой соединение, к которому полипептид проявляет адгезию, без ковалентного связывания с ним. Часто используемые соединения-носители представляют собой, например, гидроксид, фосфат или оксид алюминия, диоксид кремния, каолин и бентонит.
В практике свиноводства, свиней вакцинируют против ряда патогенных вирусов или микроорганизмов. Таким образом, является очень привлекательным, как по практическим, так и по экономическим причинам, комбинировать вакцину по изобретению для свиней, например, с дополнительным иммуногеном из вируса или микроорганизма, патогенного для свиней, или генетическим материалом (РНК/ДНК), кодирующим иммуноген из указанного вируса или микроорганизма.
Таким образом, предпочтительная форма этого варианта осуществления относится к вакцине для использования по изобретению, где эта вакцина содержит по меньшей мере один другой патогенный для свиней микроорганизм или патогенный для свиней вирус, и/или по меньшей мере один другой иммуногенный компонент и/или генетический материал, кодирующий указанный другой иммуногенный компонент, из указанного патогенного для свиней микроорганизма или патогенного для свиней вируса. Иммуноген или иммуногенный компонент представляет собой компонент, индуцирующий иммунный ответ у животного. Он может представлять собой, например, живые ослабленные или убитые цельный вирус, бактерию или паразита, или белок или группу сахара из такого вируса, бактерии или паразита.
Наиболее распространенными патогенными вирусами и микроорганизмами, патогенными для свиней, являются Brachyspira hyodysenteriae, вирус африканской лихорадки свиней, вирус Нипах, цирковирус свиней, вируса свиного гепатита TTV, вирус псевдобешенства, вирус свиного гриппа, свиной парвовируса, вирус свиного репродуктивного и респираторного синдрома (PRRS), вирус свиной эпидемической диареи (PEDV), вирус ящура, вирус трансмиссивного гастроэнтерита, ротавирус, Escherichia coli, Erysipelo rhusiopathiae, Bordetella bronchiseptica, Salmonella cholerasuis, Haemophilus parasuis, Pasteurella multocida, Streptococcus suis, Mycoplasma hyopneumoniae и Actinobacillus pleuropneumoniae.
Таким образом, более предпочтительная форма этого варианта осуществления относится к вакцине для использования по изобретению, где вирус или микроорганизм, патогенный для свиней, выбран из группы, состоящей из Brachyspira hyodysenteriae, вируса африканской лихорадки свиней, вируса Нипах, цирковируса свиней, вируса свиного гепатита TTV, вируса псевдобешенства, вируса свиного гриппа, свиного парвовируса, вируса свиного репродуктивного и респираторного синдрома (PRRS), вируса свиной эпидемической диареи (PEDV), вируса ящура, вируса трансмиссивного гастроэнтерита, ротавируса, Escherichia coli, Erysipelo rhusiopathiae, Bordetella bronchiseptica, Salmonella cholerasuis, Haemophilus parasuis, Pasteurella multocida, Streptococcus suis, Mycoplasma hyopneumoniae и Actinobacillus pleuropneumoniae.
Предпочтительным способом введения является внутримышечный или внутрикожный способ. Для внутрикожного введения, безыгольное устройство, такое как IDAL® (MSD AH), может являться предпочтительным устройством.
Краткое описание чертежей.
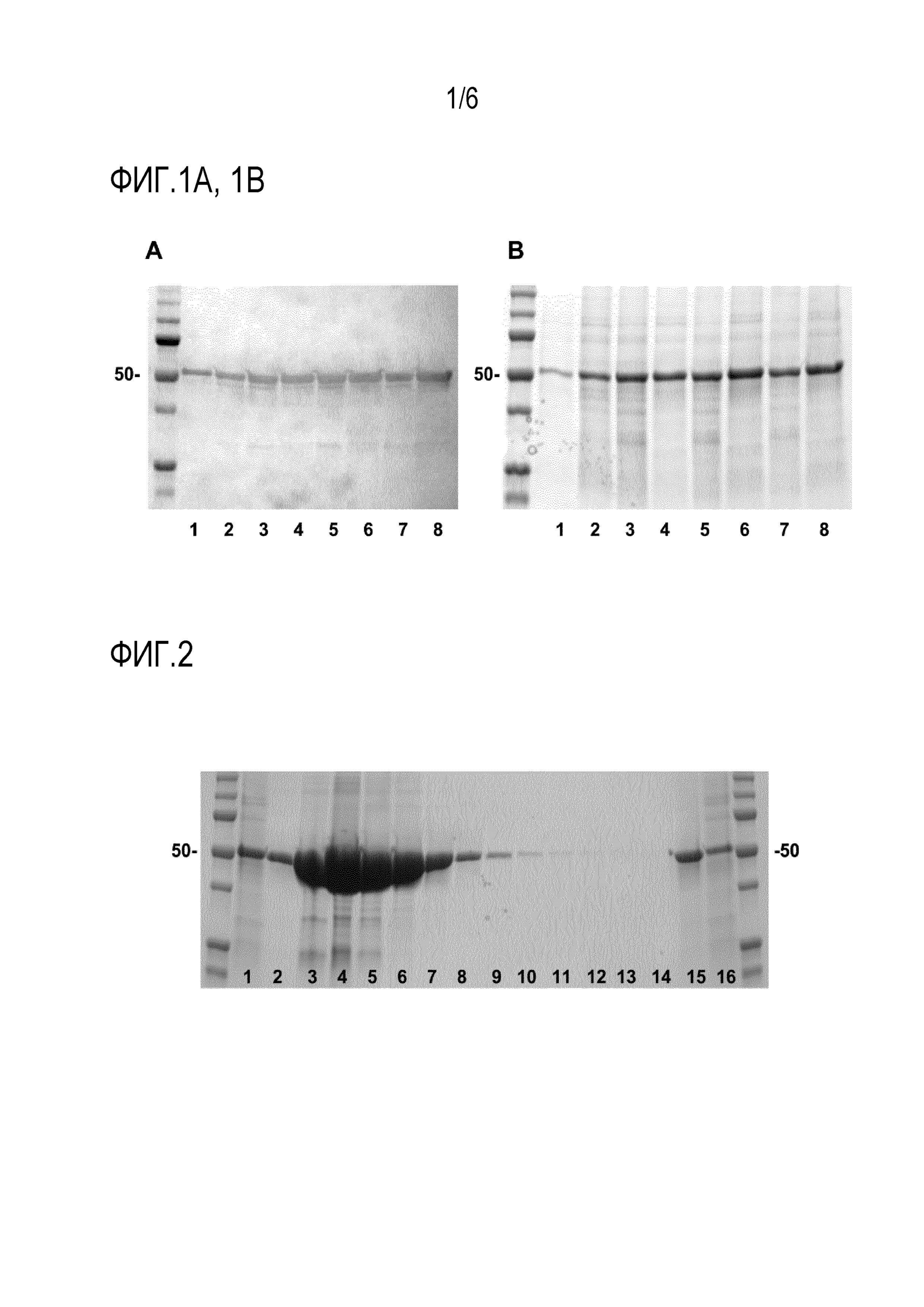
Фигура 1: Экспрессия белка HEV 112-607 в культуральном супернатанте SF9 и в клетках SF9 в различных временных точках сбора после инфекции бакуловирусом:
A) Результаты Вестерн-блоттинга после окрашивания блота с использованием антитела αHIS. После блоттинга геля после SDS-PAGE, мембрану окрашивали в течение 1 часа с использованием αHIS-MCA, разведенного 1:500 в блокирующем буфере. Затем, мембрану промывали буфером для промывки и инкубировали в течение 1 час с вторичным антителом GAM/IgG(H+L)/PO, разведенным 1:1000 в блокирующем буфере. Мембрану снова промывали буфером для промывки и инкубировали с субстратом Vector SG, разведенным в 15 мл 3M ацетата натрия.
B) Окрашивание красителем Instant blue белка в геле после SDS-PAGE. Для окрашивания instant blue, гель после SDS page инкубировали в течение 2 часов в 10 мл красителя для белка InstantBlue.
Маркер молекулярной массы с размером белков в кДа нанесен на левую дорожку. Указан маркер размера 50 кДа. Ожидаемый размер HEV 112-607 составляет 55 кДа.
Нанесение образцов обозначено номерами:
1. сбор на 3 сутки, среда
2. сбор на 3 сутки, клетки
3. сбор на 4 сутки, клетки
4. сбор на 4 сутки, среда
5. сбор на 5 сутки, клетки
6. сбор на 5 сутки, среда
7. сбор на 6 сутки, клетки
8. сбор на 6 сутки, среда
Фигура 2: Окрашивание красителем Instant blue в геле после SDS-PAGE фракций элюата 1-14 HEV 112-607 после очистки с помощью AKTA Avant белка, собранного из культурального супернатанта. Элюат собирали в фракциях по 2 мл. Дорожки 15, 16 соответствуют несвязавшейся фракции и фракции отмывки, соответственно. Для окрашивания instant blue, гель после SDS page инкубировали в течение 2 часов в 10 мл красителя для белка InstantBlue.
Маркер молекулярной массы с размером белков в кДа нанесен на самую левую и самую правую дорожки. Ожидаемый размер HEV 112-607 составляет 55 кДа.
Элюаты 1-7 объединяли после очистки.
Фигура 3: Количество специфических для HEV антител, измеренное в различные временные точки в исследовании вакцинации, как определено в ELISA. Данные представлены с планками погрешностей (стандартная ошибка среднего, SEM)
A: Поросята в возрасте 3 недель в начале эксперимента имели значительные титры материнских антител против HEV
B: Поросята в возрасте 10 недель в начале эксперимента имели уменьшенные титры (материнских) антител против HEV, что согласуется с уменьшенным материнским иммунитетом в этом возрасте.
C: Вакцинация с использованием вакцин 112-607 SUP и 112-607 AKTA приводила к сероконверсии до специфических антител против ORF2 HEV в группах возраста «3 недели» на T=4 недели после вакцинации. Материнские антитела не исчезали полностью.
D: Вакцинация с использованием вакцин 112-607 SUP и 112-607 AKTA приводила к сероконверсии до специфических антител против ORF2 HEV в группах возраста «10 недель» на T=4 недели после вакцинации.
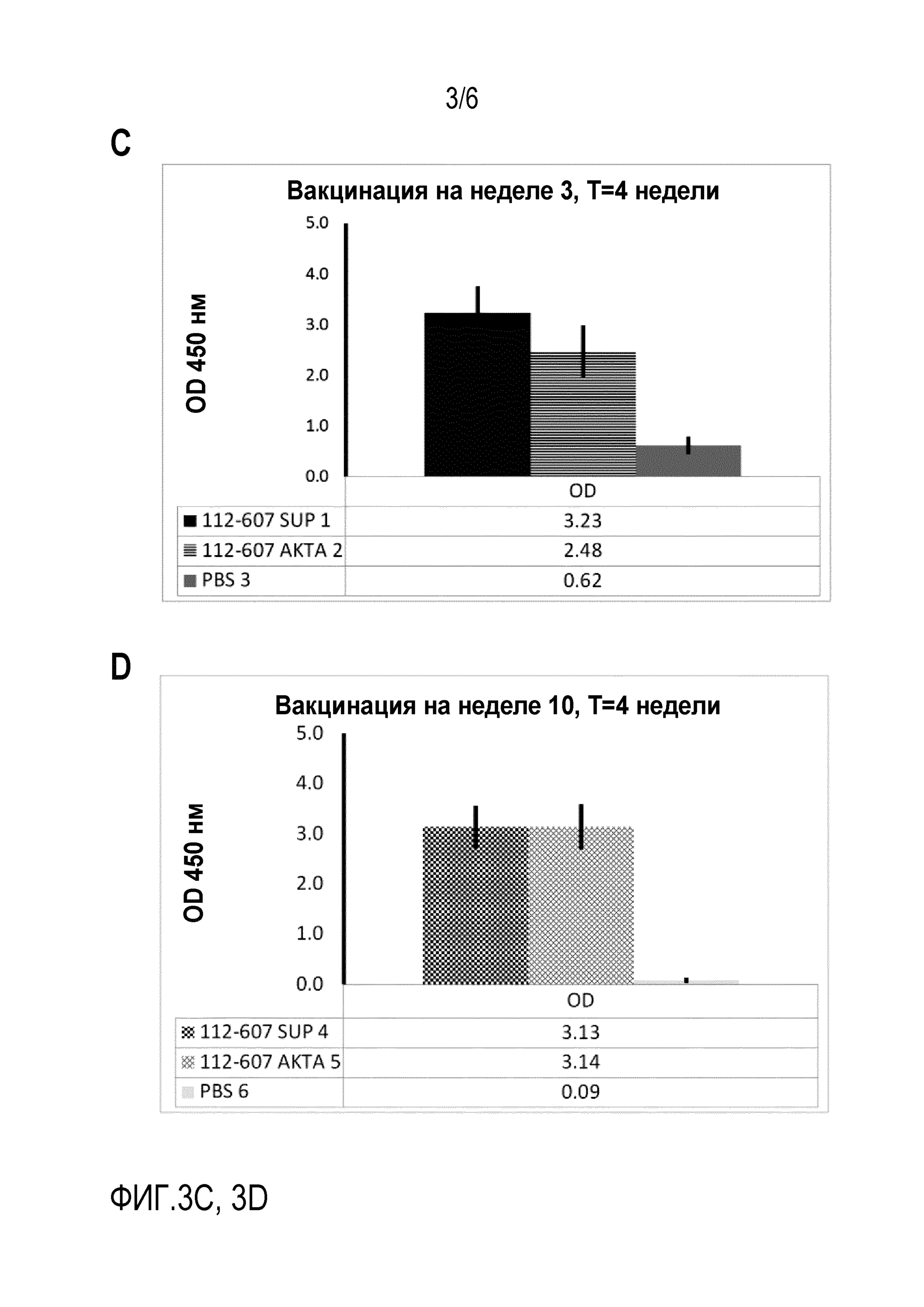
E: Вакцинация с использованием вакцин 112-607 SUP и 112-607 AKTA приводила к сероконверсии до специфических антител против ORF2 HEV в группах возраста «3 недели» на T=9 недель после вакцинации (подгруппы, отмеченные A). Бустер-вакцинация через 5 недель после примирующей вакцинации приводила к увеличению титров антител на T=9 недель после примирующей вакцинации (подгруппы, отмеченные B), по сравнению только с примирующей вакцинацией. Уровень материнских антител упал ниже уровня детекции.
F: Вакцинация с использованием вакцин 112-607 SUP и 112-607 AKTA приводила к сероконверсии до специфических антител против ORF2 HEV в группах возраста «10 недель» на T=9 недель после вакцинации (подгруппы, отмеченные A). Бустер-вакцинация через 5 недель после примирующей вакцинации приводила к увеличению титров антител на T=9 недель после примирующей вакцинации (подгруппы, отмеченные B) по сравнению только с примирующей вакцинацией.
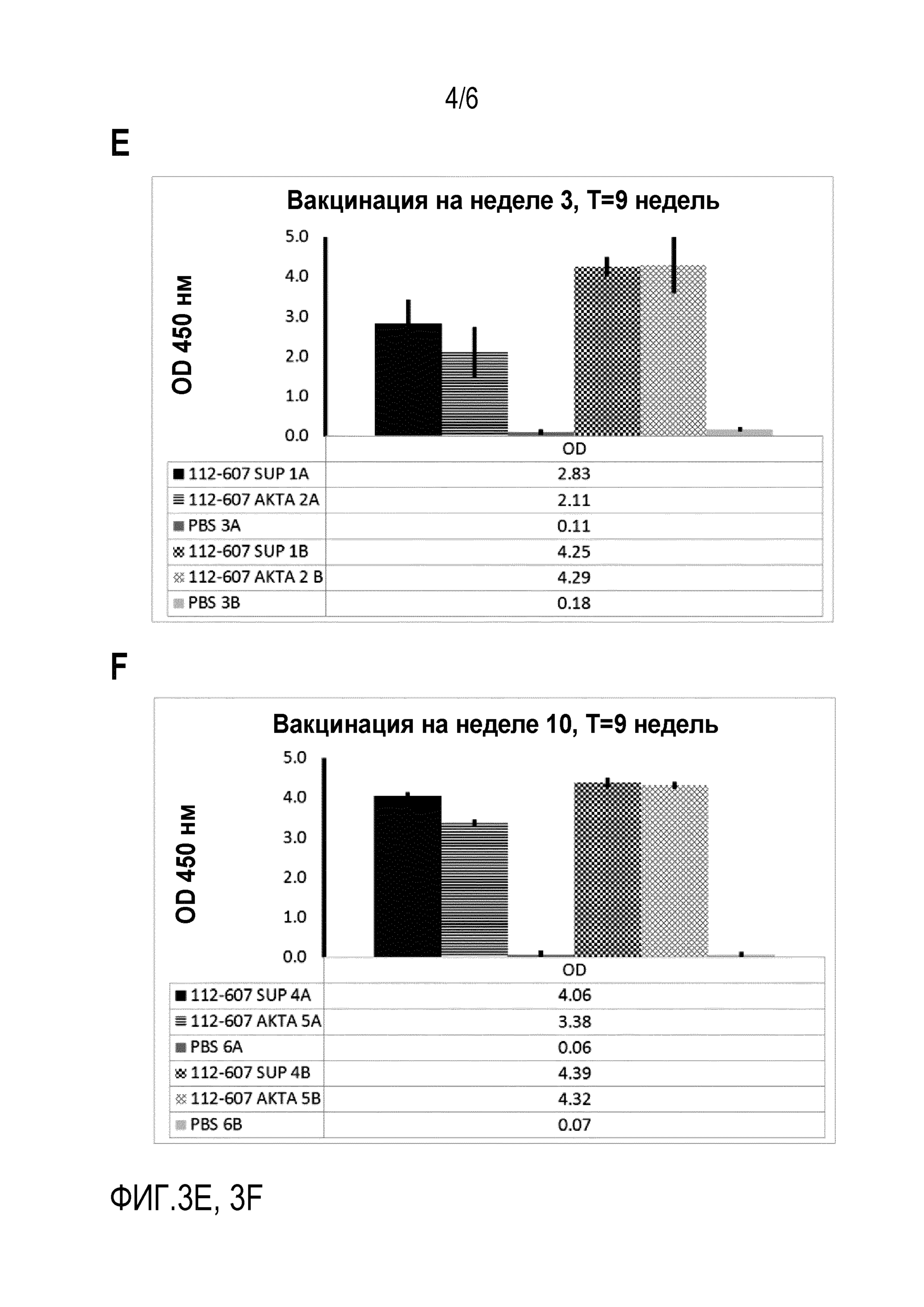
Фигура 4: Количество специфических для HEV антител, измеренное в различные временные точки в эксперименте вакцинации-заражения, как определено в ELISA. Данные представлены с планками погрешностей (стандартная ошибка среднего, SEM)
Фигура 5: На этой фигуре показана аминокислотная последовательность ORF 2 из ак последовательности 112-607 (SEQ ID NO: 1).
Фигура 6: На этой фигуре показана оптимизированная по кодонному составу последовательность ДНК, кодирующая аминокислотную последовательность из фигуры 5 (SEQ ID NO: 2).
Примеры.
Пример 1: Экспрессия: ORF2 HEV в бакуловирусной системе экспрессии BAC to BAC.
Домен белка капсида ORF2 HEV генотипа 3 [вирус гепатита E свиней, на основании GenBank: AFJ06417.1), как изображено на фигуре 5 (SEQ ID NO: 1), экспрессировали в бакуловирусной системе экспрессии BAC to BAC (Invitrogen). Белок сконструирован с C-концевой меткой HIS для очистки.
Экспрессированный фрагмент белка содержит коровый структурный домен (аминокислоты 112-367) и прогнозируемый нейтрализующий эпитоп (аминокислоты 456-607), но лишен связывающего РНК домена ORF2, N-концевых 111 аминокислот ORF2 HEV.
Кодирующую последовательность HIS-меченного белка, как изображено на фигуре 6 (SEQ ID NO: 2), клонировали в плазмиду pFastbac1 (Invitrogen). Кодирующая последовательность сконструирована с фланкирующим 5'-участком рестрикции BamHI, инициирующим кодоном, оптимизированной по кодонному составу нуклеотидной последовательностью для белка, меткой HIS, стоп-кодоном и 3'-участком рестрикции EcoRI. Эта информация послана в Genscript (Piscataway, NJ, USA), где нуклеотидную последовательность синтезировали, расщепили ферментами рестрикции BamHI и EcoRI, и клонировали в плазмиду pFastbac1, которую расщепляли такими же ферментами рестрикции.
Затем белок экспрессировали с использованием бакуловирусной системы экспрессии Bac-to-Bac®, согласно соответствующему руководству пользователя от Invitrogen: Bac-to-Bac® Baculovirus Expression System. An efficient site-specific transposition system to generate baculovirus for high-level expression of recombinant proteins. Invitrogen. Каталожные номера 10359-016, 10360-014, 10584-027, 10712-024. Номер поддокумента 10359, Номер публикации MAN0000414).
Кратко, компетентные клетки E.coli DH10Bac трансформировали с использованием плазмидной ДНК, содержащей нуклеотидную последовательность, как описано выше. Затем вставленную последовательность переносили с помощью транспозона в последовательность бакмиды, присутствующей в клетках DH10Bac. Затем бактерии высевали на чашки с агаром, содержащим антибиотики и IPTG. Отбор проводили посредством бело-синего скрининга, поскольку белые колонии содержат рекомбинантную бакмиду. Из белой колонии получали жидкую культуру. Затем проводили выделение в минимасштабе для выделения бакмидной ДНК из клеток E.coli. Клетки насекомых SF9 трансфицировали (Cellfectin, Invitrogen) этой рекомбинантной бакмидной ДНК, получая в результате формирование рекомбинантных бакуловирусных частиц. Эти частицы собирали и использовали для инфекции свежих монослоев клеток насекомых SF9. Инфекция индуцирует экспрессию рекомбинантного гена и продукцию HIS-меченного белка 112-607 HEV-ORF2.
Для определения временной точки оптимального выхода белка, белок собирали в различные временные точки, через 3, 4, 5 и 6 суток после инфекции. Во всех этих экспериментах инфекции использовали оптимизированную MOI 0,1.
Собирали рекомбинантный белок, экспрессированный в культуральной среде клеток SF9 или в клетках SF9. Клетки освобождали от культурального флакона посредством постукивания, и суспензию клеток центрифугировали при 3000 x g в течение 10 минут. Среду собирали и хранили до анализа при -70°C. Осадки клеток хранили до анализа при -20°C. Сбор как супернатанта, так и клеток анализировали по экспрессии белка. Осажденные клетки ресуспендировали в PBS перед анализом экспрессии белка. Образцы обоих сборов разделяли посредством SDS-PAGE вместе с предварительно окрашенным белковым маркером protein all blue (Biorad) и анализировали посредством окрашивания Instant Blue (Expedeon) и Вестерн-блоттинга (αHIS-MCA).
Для окрашивания посредством instant blue, гель после SDS page инкубировали в течение 2 часов в 10 мл красителя для белка InstantBlue.
Для Вестерн-блоттинга использовали Trans Blot Turbo Transfer Pack (Biorad), согласно соответствующему руководству пользователя. После блоттинга, мембраны блокировали в течение одного часа блокирующим буфером (1% обезжиренное молоко в PBS 0,04 M, полисорбат 20 0,05%). Затем мембрану окрашивали в течение 1 часа с использованием αHIS-MCA (MSD Animal или коммерчески доступный эквивалент), разведенного 1:500 в блокирующем буфере. Затем мембрану промывали буфером для промывки (PBS 0,04 M, полисорбат 20 0,5%) и инкубировали в течение 1 часа с вторичным антителом GAM/IgG(H+L)/PO (Nordic), разведенным 1:1000 в блокирующем буфере. Мембрану снова промывали буфером для промывки и кратко инкубировали с 3 каплями субстрата Vector SG (Вектор Laboratories), разведенными в 15 мл 3M ацетата натрия.
Рекомбинантный белок 112-607 секретировался в культуральную среду в форме растворимого белка, но также присутствовал в осадке клеток (Фигура 1, A: Вестерн-блоттинг, B: окрашивание белка). Наивысшего выхода белка достигали через 5 суток после инфекции в культуральной среде. Культуральную среду, содержащую растворимый белок, использовали для получения вакцины после инактивации бакуловируса посредством гамма-облучения=112-607 SUP (см.: Пример 3, получение вакцины). Из культуральной среды получали также фракцию очищенной субъединицы белка. HIS-меченный белок очищали из культуральной среды с использованием системы очистки белка AKTA Avant=112-607 AKTA (см. Пример 2, способ очистки с использованием AKTA).
Пример 2: Способ очистки с использованием системы очистки белка AKTA Avant
HIS-меченный белок 112-607, присутствующий в культуральной среде, очищали с использованием системы очистки AKTA Avant (GE Healthcare, Germany).
HIS-меченный белок в культуральном супернатанте очищали в системе очистки AKTA Avant (GE Healthcare, Germany) с использованием колонки HIStrap FF (GE Healthcare арт. 17-5255-01 (5 мл). Перед очисткой, фракцию фильтровали на фильтре 0,45 мкм (Millipore).
Использовали следующую программу:
|
Буфер для промывки AKTA, не денатурирующий: 300 мМ KCl, 50 мМ TRIS, 5 мМ имидазол
Буфер для элюции, не денатурирующий: 300 мМ KCl, 50 мМ TRIS, 500 мМ имидазол
Элюат собирали в фракциях по 2 мл, и их разделяли в геле для SDS page с последующим окрашиванием посредством instant blue staining. Гель для SDS page инкубировали в течение 2 часов в 10 мл красителя для белка InstantBlue (Expedeon) (Фигура 2). Элюаты 1-7 were объединяли после очистки.
После очистки проводили диализ очищенного белка. Очищенный белок наносили на диализную мембрану Spectra Por3, MWCO 3,5 кДа (Spectrum Labs), и ее помещали во внешнюю камеру с буфером для диализа (150 мМ KCl+150 мМ TRIS) в объеме, приблизительно в 400 раз превышающем объем очищенного белка. Буфер постоянно перемешивали в течение 2 суток при 4°C, и через 1 сутки буфер для диализа заменяли.
Пример 3: Получение вакцинных составов
Состав X-solve представляет собой эмульсию масло-в-воде на основе минерального масла в комбинации с солюбилизированным ацетатом витамина E. Размер капель эмульсии на основе минерального масла составляет приблизительно 0,5 мкм, и размер капель солюбилизированного ацетата витамина E составляет приблизительно 150 нм. Вязкость конечной вакцины составляет ниже 15 мПа⋅с.
Концентрацию белка антигена определяли следующим образом: Водную фазу, используемую для вакцинного состава, разделяли посредством SDS-PAGE вместе с предварительно окрашенным белковым маркером protein all blue (Biorad) и стандартом BSA с известной концентрацией белка (25-50-100 мкг/мл), и анализировали посредством окрашивания Instant Blue (Expedeon). Для окрашивания посредством instant blue, гель после SDS page инкубировали в течение 2 часов в 10 мл красителя для белка InstantBlue. Плотность сигналов измеряли посредством программного обеспечения GeneTools от Syngene. Концентрацию белка 112-607 AKTA и 112-607 SUP оценивали на основании линейной регрессии по результатам для стандарта BSA.
112-607 AKTA: 40 мкг/мл белка в конечном вакцинном составе
112-607 SUP: 40 мкг/мл белка в конечном вакцинном составе
Пример 4: Определение уровней (материнских) антител в сыворотке
Собирали индивидуальные образцы крови свиней (Vacuolette 8 мл Sep Clot Activator (Greiner-Bio One)). После свертывания при комнатной температуре и в течение ночи при 4°C, из образцов крови получали сыворотку. Это осуществляли посредством центрифугирования пробирок при 2500 x g в течение 25 минут при 4°C. Образцы сыворотки для ELISA инактивировали нагреванием в течение 30 минут при +56°C и наконец, сохраняли при -20°C или ниже до использования. Сыворотку тестировали по присутствию антител против HEV с использованием коммерческого набора для теста HEV ELISA 4.0 от MP Biomedicals, Santa Ana, CA, USA. Это осуществляли согласно соответствующему протоколу для набора HEV ELISA 4.0.
Пороговое значение для ELISA составляет OD (450 нм) 0,2+среднее для отрицательных контрольных образцов (0,05). Образцы с OD ниже 0,250 считали отрицательными по HEV.
Пример 5: Детекция HEV в ректальных мазках, желчи и сыворотке.
Собирали индивидуальные образцы крови свиней (Vacuolette 8 мл Sep Clot Activator (Greiner-Bio One)). После свертывания при комнатной температуре и в течение ночи при 4°C, из образцов крови получали сыворотку. Это осуществляли посредством центрифугирования пробирок при 2500 x g в течение 25 минут при 4°C. Образцы сыворотки для ПЦР не инактивировали нагреванием и сохраняли при -20°C или ниже до анализа.
Желчь отбирали из желчного пузыря при некропсии с использованием шприца и сохраняли при -20°C или ниже до анализа.
Ректальные мазки отбирали в различные временные точки в ходе эксперимента. Тампоны помещали в пробирки, содержащие 1 мл PBS и антибиотики. В лаборатории, содержимое пробирок перемешивали на встряхивателе и переливали в криопробирки. Образцы сохраняли при -20°C или ниже до анализа.
Образцы ректальных мазков, желчи и сыворотки тестировали по присутствию РНК HEV посредством q-RT-ПЦР.
РНК выделяли посредством устройства MagNA Pure 96 (Roche) с использованием внешнего протокола лизиса, как описано производителем. Кратко, 200 мкл неразведенного образца смешивали с 250 мкл буфера для внешнего лизиса (Roche), и нуклеиновые кислоты выделяли в соответствии со связанным протоколом.
Для количественной детекции РНК HEV в этих образцах, использовали набор для RT-ПЦР QuantiTect Probe (Qiagen). Реакционная смесь для qПЦР состояла из 4,5 мкл не содержащей РНКазы воды, 12,5 мкл готовой реакционной смеси для RT-ПЦР QuantiTect Probe RT-PCR mastermix, 400 нМ прямого и обратного праймера, 300 нМ зонда, 0,75 мкл смеси QuantiTect Probe RT-mix (фермента) и 5 мкл РНК.
Использовали следующие праймеры и зонд:
Прямой праймер; GGT GGT TTC TGG GGT GAC (SEQ ID NO: 3)
Обратный праймер; AGG GGT TGG TTG GAT GAA (SEQ ID NO: 4)
Меченный 6FAM TAMRA зонд; TGA TTC TCA GCC CTT CGC (SEQ ID NO: 5)
Обратную транскрипцию проводили при 50°C в течение 30 мин, с последующей денатурацией при 95°C в течение 15 мин. ДНК немедленно амплифицировали с использованием 40 циклов ПЦР при 94°C (15 с), 56°C (30 с) и 76°C (30 с). Реакции проводили в системе с детекцией в реальном времени CFX96 (Biorad), и анализ с использованием программного обеспечения проводили с CFX manager версии 3.1. Данные представлены в копиях/мл образца. Количественная оценка была основана на анализе набора стандартных образцов, содержащих 101-108 копий ORF2 HEV/5мкл в каждой реакционной смеси для q-RT-ПЦР в CFX96.
Пример 6: Эксперимент вакцинации
Всего 30 поросят в возрасте 3 недель, являющихся сероположительными по материнским антителам против HEV, использовали в этом исследовании. Поросят случайным образом выбирали из различных пометов.
Всего 30 свиней в возрасте 10 недель использовали в этом исследовании. Поросят выбирали случайным образом. К возрасту 10 недель, уровни материнских антител против HEV уменьшались по сравнению с поросятами в возрасте 2 недель (Фигура 3A, B).
Поросят в возрасте 3 недель приписывали к 3 группам обработки по 10 поросят каждая (группы 1-2-3). Поросят из группы 1 вакцинировали (IM вакцинация, 1 мл, в шею) в возрасте 3 недель с использованием вакцины ORF2 HEV 112-607 SUP, составленной в X-Solve. Поросят из группы 2 вакцинировали (IM вакцинация, 1 мл, в шею) в возрасте 3 недель с использованием вакцины ORF2 HEV 112-607 AKTA, составленной в X-Solve. Поросят из группы 3 вакцинировали (IM вакцинация, 1 мл, в шею) в возрасте 3 недель с использованием PBS.
Свиней в возрасте 10 недель приписывали к 3 группам обработки по 10 поросят каждая (группы 4-5-6). Поросят из группы 4 вакцинировали (IM вакцинация, 1 мл, в шею) в возрасте 10 недель с использованием вакцины ORF2 HEV 112-607 SUP, составленной в X-Solve. Поросят из группы 5 вакцинировали (IM вакцинация, 1 мл, в шею) в возрасте 10 недель с использованием вакцины ORF2 HEV 112-607 AKTA, составленной в X-Solve. Поросят из группы 6 вакцинировали (IM вакцинация, 1 мл, в шею) в возрасте 3 недель с использованием PBS.
Каждую группу из 10 поросят подразделяли на группы A и B через 5 недель после примирующей вакцинации (возраст 8/15 недель). 5 поросят не подвергали бустер-вакцинации (подгруппа A), в то время как других 5 поросят подвергали бустер-вакцинации, идентичной примирующей вакцинации (подгруппа B).
Сыворотку собирали до вакцинации (возраст 3/10 недель), в возрасте 7/14 недель (4 недели после вакцинации) и в возрасте 12/19 недель (9 недель после вакцинации). Уровни антител против HEV определяли в сыворотке, как описано в примере 4. Отсутствие реплицирующегося HEV подтверждали во всех образцах сыворотки посредством анализа q-RT-ПЦР.
В возрасте 3 недель, поросята имели значительные титры антител против HEV, которые в большой степени исчезали у свиней в возрасте 10 недель (Фигура 3A, B). Вакцинация с использованием вакцины 112-607 SUP (группа 1) и вакцины 112-607 AKTA (группа 2) приводила к сероконверсии до специфических антител против ORF2 HEV в обеих возрастных группах через 4 (Фигура 3C, D) и 9 недель после вакцинации (Фигура 3E, F; подгруппы A). Бустер-вакцинация через 5 недель после примирующей вакцинации приводила к увеличенным титрам антител через 9 недель после примирующей вакцинации. (Фигура 3E, F; подгруппы B)
Пример 7: Эксперимент вакцинации-заражения у поросят,
Всего 12 поросят в возрасте 3 недель, являющихся сероположительными по материнским антителам против HEV, использовали в этом исследовании. Поросят случайным образом выбирали из различных пометов и содержали вместе в одном загоне.
Поросят приписывали к 3 группам обработки по 4 поросят каждая (группы 1-2-3). Поросят из группы 1 вакцинировали (IM вакцинация, 1 мл, в шею) в возрасте 3 недель с использованием вакцины ORF2 HEV 112-607 SUP, составленной в X-Solve. Сыворотку собирали до вакцинации (в возрасте 3 недель), в возрасте 7 недель (4 недели после вакцинации) и до экспериментального заражения (в возрасте 11 недель, 8 недель после вакцинации).
Через 8 недель после вакцинации (в возрасте 11 недель), 4 свиней из группы 2 подвергали экспериментальному заражению с использованием гомогената печени положительного по HEV животного с голландской фермы. Материал для заражения представлял собой 10% гомогенат печени в 50/50 (об./об.) среды Уильяма E/среды Хэма F-12. Гомогенат печени для заражения вводили посредством внутримышечной (IM) инъекции (2×2,5 мл, левая и правая часть шеи), и пероральную дозу 15 мл вводили с использованием шприца. Для перорального введения, гомогенат центрифугировали при 3200 x g в течение 1 часа при 4°C, и супернатант использовали для заражения. Этот материал содержал 2,12×106 копий РНК HEV/мл. Для IM инъекции, этот супернатант центрифугировали снова при 10000 x g в течение 1 часа при 4 °C, и затем фильтровали через фильтр 0,22 мкм (Millipore) и инъецировали. Этот материал содержал 1,22×106 копий РНК HEV/мл. Использованный положительный по HEV материал содержал HEV генотипа 3i, который определили на основании анализа последовательности РНК из нуклеотидной последовательности вируса. Для этого вируса показана наивысшая гомология с номером доступа в Genbank: KC618403.1. Свиньи из группы 3 служили в качестве индикаторов контакта.
Выделение HEV посредством фекалий после экспериментального заражения мониторировали два раза в неделю (возраст 12-17 недель) посредством анализа q-RT-ПЦР ректальных мазков. Присутствие вируса в сыворотке мониторировали еженедельно (возраст 13-17 недель) посредством анализа q-RT-ПЦР. Некропсии проводили в возрасте 17 недель. Желчь и кровь собирали во время некропсии, за исключением животных из группы 2. Присутствие вируса HEV анализировали с использованием q-RT-ПЦР (Пример 5) в ректальных мазках, желчи и сыворотке.
Уровень антител против HEV определяли в сыворотке, как описано в примере 4. На фигуре 4 показаны результаты ELISA. Вакцинация поросят в возрасте 3 недель индуцировала сероконверсию до специфических антител против HEV у вакцинированных животных, которая поддавалась детекции с возраста 7 недель и далее. Для групп 2 и 3 показано уменьшение титров антител из-за исчезновения материнских антител до экспериментального заражения группы 2 в возрасте 11 недель. Заражение животных из группы 2 приводило к индукции возникновения специфических антител против HEV в этой группе в возрасте между 13 и 14 неделями, а также в группе индикаторов 3, которые подвергались сероконверсии вследствие инфекции HEV из-за прямого контакта с экспериментально зараженными свиньями. В группе индикаторов титры антител развивались в возрасте между 15 и 16 неделями, с задержкой приблизительно 2 недели по сравнению с группой экспериментального заражения 2, что соответствует инкубационному периоду вируса. Группа вакцинации 1 проявляла реакцию на воздействие HEV через экспериментально зараженных свиней (группа 2) с увеличением титров специфических антител против HEV, начиная с недель между 14 и 15.
Успешное экспериментальное заражение группы 2 и инфекцию индикаторных свиней в группе 3 подтверждали посредством проведения анализов q-RT-ПЦР желчи и сыворотки экспериментальных животных (Таблица A). У трех из четырех экспериментально зараженных свиней, и у четырех из четырех индикаторных свиней показаны поддающиеся детекции уровни HEV в сыворотке или желчи. Ни одна из вакцинированных свиней не имела поддающихся детекции уровней HEV в сыворотке или желчи. Ректальные мазки также анализировали посредством анализа q-RT-ПЦР, что показано в таблице B.
Настоящий эксперимент разработан для имитации полевых условий, где HEV инфицирует свиней в заключительной стадии откорма. Вакцинация защищала свиней от виремии в крови и желчи (зараженные животные, индикаторные животные), что показывает, что вакцинированные свиньи не имеют системной инфекции HEV. Кроме того, вакцинированные HEV свиньи проявляют реакцию на воздействие вируса с увеличением уровней антител в сыворотке. Выделение HEV в фекалиях уменьшено у вакцинированных свиней по сравнению с индикаторными свиньями в том же загоне.
Более конкретным, как можно видеть в таблице B, является то, что вакцинированные свиньи, по сравнению с индикаторными свиньями, имеют значительно уменьшенное распространение с фекалиями на неделях 13, 14 и 15 (в 10-50 раз более низкое количество материала ДНК HEV присутствует в фекалиях). Около недели 16 распространение кажется стабилизированным на низком уровне, который является по существу одинаковым для вакцинированных животных, так же как для контрольных животных. Поскольку для вакцинированных животных совсем не показано системной инфекции HEV (см. таблицу A), это должен быть вирус, распространяющийся из-за поросят, съедающих инфицированные HEV фекалии. По-видимому, уровень распространения, достигнутый после недели 15, обусловлен поеданием инфицированных фекалий, но не обусловлен фактической инфекцией животного. Кратко, из объединения данных из таблицы A с данными из таблицы B ясно, что с использованием настоящей вакцины, положительного по MDA против HEV поросенка можно защищать от распространения HEV с фекалиями, в частности, от распространения с фекалиями, являющегося результатом собственно инфекции животного.
Таблица A: Анализ q-RT-ПЦР сыворотки и желчи в различных временных точках в исследовании вакцинации-заражения (копий РНК/мл).
|
Таблица B: Анализ q-RT-ПЦР ректальных мазков в различных временных точках в исследовании вакцинации-заражения (копий РНК/мл).
|
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Intervet International B.V.
<120> Вакцина против HEV.
<130> 24334
<160> 5
<170> PatentIn версии 3.5
<210> 1
<211> 503
<212> PRT
<213> Вирус гепатита E
<400> 1
Met Ala Val Ser Pro Ala Pro Asp Thr Ala Pro Val Pro Asp Val Asp
1 5 10 15
Ser Arg Gly Ala Ile Leu Arg Arg Gln Tyr Asn Leu Ser Thr Ser Pro
20 25 30
Leu Thr Ser Ser Val Ala Ser Gly Thr Asn Leu Val Leu Tyr Ala Ala
35 40 45
Pro Leu Asn Pro Leu Leu Pro Leu Gln Asp Gly Thr Asn Thr His Ile
50 55 60
Met Ala Thr Glu Ala Ser Asn Tyr Ala Gln Tyr Arg Val Val Arg Ala
65 70 75 80
Thr Ile Arg Tyr Arg Pro Leu Val Pro Asn Ala Val Gly Gly Tyr Ala
85 90 95
Ile Ser Ile Ser Phe Trp Pro Gln Thr Thr Thr Thr Pro Thr Ser Val
100 105 110
Asp Met Asn Ser Ile Thr Ser Thr Asp Val Arg Ile Leu Val Gln Pro
115 120 125
Gly Ile Ala Ser Glu Leu Val Ile Pro Ser Glu Arg Leu His Tyr Arg
130 135 140
Asn Gln Gly Trp Arg Ser Val Glu Thr Ser Gly Val Ala Glu Glu Glu
145 150 155 160
Ala Thr Ser Gly Ser Val Met Leu Cys Ile His Gly Ser Pro Val Asn
165 170 175
Ser Tyr Thr Asn Thr Pro Tyr Thr Gly Ala Leu Gly Leu Leu Asp Phe
180 185 190
Ala Leu Glu Leu Glu Phe Arg Asn Leu Thr Pro Gly Asn Thr Asn Thr
195 200 205
Arg Val Ser Arg Tyr Thr Ser Thr Ala Arg His Arg Leu Arg Arg Gly
210 215 220
Ala Asp Gly Thr Ala Glu Leu Thr Thr Thr Ala Ala Thr Arg Phe Met
225 230 235 240
Lys Asp Leu His Phe Thr Gly Thr Asn Gly Val Gly Glu Val Gly Arg
245 250 255
Gly Ile Ala Leu Thr Leu Phe Asn Leu Ala Asp Thr Leu Leu Gly Gly
260 265 270
Leu Pro Thr Glu Leu Ile Ser Ser Ala Gly Gly Gln Leu Phe Tyr Ser
275 280 285
Arg Pro Val Val Ser Ala Asn Gly Glu Pro Thr Val Lys Leu Tyr Thr
290 295 300
Ser Val Glu Asn Ala Gln Gln Asp Lys Gly Ile Ala Ile Pro His Asp
305 310 315 320
Ile Asp Leu Gly Asp Ser Arg Val Val Ile Gln Asp Tyr Asp Asn Gln
325 330 335
His Glu Gln Asp Arg Pro Thr Pro Ser Pro Ala Pro Ser Arg Pro Phe
340 345 350
Ser Val Leu Arg Ala Asn Asp Val Leu Trp Leu Ser Leu Thr Ala Ala
355 360 365
Glu Tyr Asp Gln Thr Thr Tyr Gly Ser Ser Thr Asn Pro Met Tyr Val
370 375 380
Ser Asp Thr Val Thr Phe Val Asn Val Ala Thr Gly Ala Gln Ala Val
385 390 395 400
Ala Arg Ser Leu Asp Trp Ser Lys Val Thr Leu Asp Gly Arg Pro Leu
405 410 415
Thr Thr Ile Gln Gln Tyr Ser Lys Thr Phe Tyr Val Leu Pro Leu Arg
420 425 430
Gly Lys Leu Ser Phe Trp Glu Ala Gly Thr Thr Lys Ala Gly Tyr Pro
435 440 445
Tyr Asn Tyr Asn Thr Thr Ala Ser Asp Gln Ile Leu Ile Glu Asn Ala
450 455 460
Ala Gly His Arg Val Ala Ile Ser Thr Tyr Thr Thr Ser Leu Gly Ala
465 470 475 480
Gly Pro Val Ser Val Ser Ala Val Gly Val Leu Ala Pro His Ser Ala
485 490 495
Leu His His His His His His
500
<210> 2
<211> 1512
<212> ДНК
<213> Вирус гепатита E
<400> 2
atggctgttt ctcccgctcc cgacactgct cccgttcccg acgtggactc tcgtggtgcc 60
atcctgcgtc gtcaatacaa cctttctacc tcacccctga ctagctctgt tgctagcgga 120
accaacctgg tgctttacgc tgctcccctg aaccccctgc ttccccttca agacggcact 180
aacacccaca tcatggccac cgaggctagc aactacgccc aataccgtgt tgtgcgtgct 240
accatccgtt accgtcccct ggttcccaac gctgtgggcg gttacgctat cagcatctct 300
ttctggcccc aaaccactac cactcccacc agcgttgaca tgaactctat cacttcaacc 360
gacgttcgta tcctggtgca acccggcatc gctagcgaac tcgttatccc ctctgagcgt 420
ctgcactacc gtaaccaagg ctggcgtagc gttgaaacct ctggtgtggc cgaggaagag 480
gctacttcag gaagcgttat gctgtgcatc catggctcac ccgtgaacag ctacactaac 540
accccctaca ccggagctct gggtctcctg gacttcgctc tcgaactgga gttccgtaac 600
cttacccccg gcaacactaa cacccgtgtt tctcgttaca cttcaaccgc tcgtcaccgt 660
cttcgtcgtg gagccgacgg aaccgctgaa ctcaccacta ccgctgccac tcgtttcatg 720
aaagacctgc atttcactgg caccaacggc gttggcgaag tgggacgtgg catcgccctt 780
actctcttca accttgctga cacccttctc ggaggcctcc ccactgaact gatctcaagc 840
gctggtggac aactcttcta ctcacgtccc gttgtgagcg ctaacggcga acccactgtt 900
aaactgtaca ccagcgtgga gaacgcccaa caagacaagg gtatcgctat cccccacgac 960
atcgaccttg gagactctcg tgttgtgatc caagactacg acaaccaaca tgagcaagac 1020
cgtcccaccc ccagccccgc tccctctcgt cccttctcag ttctgcgtgc taacgacgtg 1080
ctgtggctta gcctcaccgc tgccgaatac gaccaaacta cctacggctc ttcaactaac 1140
cccatgtacg ttagcgacac tgtgaccttc gttaacgtgg ctaccggcgc tcaagccgtt 1200
gctcgtagcc tcgactggtc taaggtgacc cttgacggtc gtcccctcac taccatccaa 1260
caatactcta agaccttcta cgttctgccc cttcgtggta aactgtcatt ctgggaggcc 1320
ggtactacca aggctggata cccctacaac tacaacacta ccgctagcga ccaaatcctt 1380
atcgaaaacg ctgccggcca tcgtgttgct atctctacct acactacctc actcggtgct 1440
ggacccgtga gcgtgtcagc cgttggcgtg cttgctcccc actctgccct ccatcatcat 1500
caccatcatt aa 1512
<210> 3
<211> 18
<212> ДНК
<213> Вирус гепатита E
<400> 3
ggtggtttct ggggtgac 18
<210> 4
<211> 18
<212> ДНК
<213> Вирус гепатита E
<400> 4
aggggttggt tggatgaa 18
<210> 5
<211> 18
<212> ДНК
<213> Вирус гепатита E
<400> 5
tgattctcag cccttcgc 18
<---

Твердые формы макролидов
Способ лиофилизации частиц, имеющих содержащийся в них фармацевтический состав, и фармацевтическая упаковка, содержащая такие частицы
Изоксазолиновые композиции и их применение в качестве противопаразитарных средств
Композиции, включающие антибиотик и кортикостероид
Вакцина для защиты от lawsonia intracellularis, mycoplasma hyopneumoniae, цирковируса свиней
Вакцина для защиты от haemophilus parasuis серотипа 4 у поросят
Рекомбинантный вирус классической чумы свиней (csfv), содержащий модифицированный белок е2, и способы создания указанного рекомбинантного csfv
Способ получения токсинов actinobacillus pleuropneumoniae apxi или apxiii в жидкой культуральной среде, дополненной воздухом, обогащенным углекислым газом
Способ получения токсина actinobacillus pleuropneumoniae apxi, используя культуральную среду, содержащую комплекс кальций-бороглюконат
Вирус гриппа, способный инфицировать собачьих, и его применение

