Результат интеллектуальной деятельности: РЕКОМБИНАНТНЫЙ МИКРООРГАНИЗМ ДЛЯ УЛУЧШЕННОГО ПОЛУЧЕНИЯ ХИМИЧЕСКИХ ПРОДУКТОВ ТОНКОГО ОРГАНИЧЕСКОГО СИНТЕЗА
Вид РИД
Изобретение
В соответствии с настоящей заявкой испрашивается приоритет согласно заявкам с номерами US 61/915517, US 61/915527, US 61/915518, US 61/915534 и ЕР 13197432.1, все из которых включены с помощью ссылки в полном их объеме.
Область изобретения
Настоящее изобретение относится к рекомбинантной молекуле нуклеиновой кислоты, рекомбинантному микроорганизму, к способу получения пирувата, сукцината, аспартата, малата, лактата, валина, лейцина и/или аланина и к применению рекомбинантной молекулы нуклеиновой кислоты или рекомбинантного микроорганизма для ферментативного получения пирувата, сукцината, аспартата, малата, лактата, валина, лейцина и/или аланина.
Сущность изобретения
Аминокислоты представляют собой органические соединения с карбоксильной группой и аминогруппой. Наиболее важными аминокислотами являются альфа-аминокислоты, у которых аминогруппа расположена после карбоксильной группы. В основе белков лежат альфа-аминокислоты.
Аланин привлекает значительный интерес, так как его применяют в качестве добавки к продукту питания, корму и в фармацевтической промышленности. Кроме того, аланин является сырьевым материалом для промышленного получения тринатриевой соли N,N-бис(карбоксиметил)-аланина (MGDA, под торговым названием Trilon М), которая является сильным хелатирующим средством, обладающим отличными функциональными характеристиками при растворении органических и неорганических солевых отложений (WO 94/29421, WO 2012/150155). По данным стандартных тестов OECD различные марки Trilon М легко биоразлагаются. В связи с отличным экологическим и токсикологическим профилем различные марки Trilon М особенно хорошо подходят для применения в продуктах для конечных потребителей, и спрос на такие биоразлагаемые комплексные компоненты постоянно повышается.
Аланин может быть произведен посредством ферментации при помощи дифтериеподобных бактерий (Hermann, 2003: Industrial production of amino acids by Coryneform bacteria, J. of Biotechnol, 104, 155-172.) или E. coli. (WO 2007/120198, WO 2008/119009).
Недавно было описано, что сверхэкспрессия гена ygaW повышает ферментативную производительность микроорганизма по аланину (WO 2012/172822).
Производство аланина у Е. coli является более эффективным, и его широко применяют для промышленного получения аланина в качестве сырьевого материала для химической промышленности. По мере роста спроса на аланин в химической промышленности существует потребность в повышении производительности ферментативного получения аланина.
Одной задачей настоящего изобретения является создание микроорганизмов, которые можно применять для ферментативного получения аланина с высоким выходом и эффективностью.
Подробное описание изобретения
Один вклад в достижение описанной выше задачи обеспечивается рекомбинантным микроорганизмом из семейства Escherichia coli (Е. coli), характеризующимся, по сравнению с соответствующим эталонным микроорганизмом, по меньшей мере одной из i) уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, и/или ii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, и/или iii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, и/или iv) уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, и/или v) измененной активности гена lpxD.
Термин «уменьшенная, подавленная или исключенная экспрессия и/или активность фермента» означает значимо уменьшенную, подавленную или исключенную экспрессию и/или активность, а также охватывает не поддающуюся детекции экспрессию и/или активность соответствующих ферментов.
Термин «более высокий», «увеличение» или «повышенный», например, в отношении экспрессии и/или активности фермента или выхода или производительности, означает значимо более высокую, увеличенную или повышенную экспрессию и/или активность или выход или производительность.
Термин «измененная» экспрессия и/или активность фермента означает экспрессию и/или активность фермента у рекомбинантного микроорганизма, которая значимо отличается от экспрессии и/или активности соответствующего фермента у нерекомбинантного микроорганизма дикого типа.
Неожиданным образом было обнаружено, что микроорганизм, характеризующийся по меньшей мере одной из i) уменьшенной, подавленной или исключенной экспрессии и/или активности белка, кодируемого геном brnQ, и/или ii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, и/или iii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, и/или iv) уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, и/или v) измененной активности гена lpxD, характеризуется более высоким выходом и/или производительностью по аланину при ферментативном производстве по сравнению с таким же микроорганизмом, не характеризующимся уменьшенной, подавленной или исключенной экспрессией и/или активностью соответствующего гена brnQ, и/или внесенной, увеличенной или повышенной активностью и/или экспрессией гегна argP, и/или внесенной, увеличенной или повышенной активностью и/или экспрессией гена gcvA, и/или уменьшенной, подавленной или исключенной активностью и/или экспрессией гена gcvB, и/или измененной активностью гена lpxD.
Соответственно, согласно одному варианту осуществления настоящее изобретение, с одной стороны, относится к рекомбинантному микроорганизму, характеризующемуся по сравнению с соответствующим эталонным микроорганизмом по меньшей мере одной из i) уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или ii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или iii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или iv) уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или v) измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, и характеризующемуся по сравнению с соответствующим эталонным микроорганизмом более высоким выходом и/или производительностью по аланину при ферментативном производстве.
Термин «эталонный микроорганизм» в контексте настоящей заявки означает контрольный микроорганизм, с которым сравнивают рекомбинантный микроорганизм. Такой эталонный микроорганизм имеет практически такой же генотип, как у рекомбинантного микроорганизма, за исключением подлежащих анализу отличий. Предпочтительно эталонный микроорганизм представляет собой штамм, от которого происходит рекомбинантный микроорганизм. Например, если некоторый ген был внесен в микроорганизм дикого типа, создавая таким образом рекомбинантный микроорганизм, то дикий тип будет подходить в качестве эталонного микроорганизма для такого рекомбинантного микроорганизма. Также возможно, что в рекомбинантный микроорганизм А вносят дополнительную мутацию, тем самым создавая рекомбинантный микроорганизм В. Тогда рекомбинантный микроорганизм А может быть подходящим эталонным микроорганизмом для рекомбинантного микроорганизма В. В том случае, если будут сравнивать функциональные характеристики рекомбинантного микроорганизма и соответствующего эталонного микроорганизма, оба микроорганизма выращивают в практически идентичных условиях.
Специалисту в настоящей области очевидно, что микроорганизм, характеризующийся увеличенным выходом и/или производительностью по аланину, также можно применять для получения других метаболитов, которые являются близкородственными для аланина, например метаболитов, которые являются промежуточными продуктами в аланиновом метаболическом пути, у которых есть общие промежуточные продукты с аланиновым путем, или которые являются метаболитами, которые используют аланин в качестве промежуточного продукта в своем метаболическом пути. Микроорганизмы по настоящему изобретению также можно легко адаптировать для того, чтобы они характеризовались увеличенным выходом и/или производительностью по таким родственным метаболитам, посредством повышения или внесения некоторых ферментативных активностей или посредством нокаутирования или снижения некоторых ферментативных активностей.
Такими метаболитами, например, являются пируват, сукцинат, аспартат, малат, лактат, валин и лейцин.
Например, для того, чтобы применять рекомбинантный микроорганизм по настоящему изобретению для получения сукцината, гены ldh, pfl, pta и adhE необходимо нокаутировать, а ген карбоксилазы PEP и/или ген пируваткарбоксилазы необходимо внести в геном микроорганизма по настоящему изобретению. Соответствующий метаболический путь и необходимые мутации описаны, например, в работе Zhang et al. (2009), PNAS (106) pp. 20180-20185.
Соответственно, согласно другому варианту осуществления настоящее изобретение, с одной стороны, относится к рекомбинантному микроорганизму, характеризующемуся по сравнению с соответствующим эталонным микроорганизмом по меньшей мере одной из i) уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или ii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или iii) привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или iv) уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или v) измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, и характеризующемуся по сравнению с соответствующим эталонным микроорганизмом более высоким выходом и/или производительностью по пирувату, сукцинату, аспартату, малату, лактату, валину и/или лейцину при ферментативном производстве.
В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой прокариотическую клетку. Подходящие прокариотические клетки включают грамположительные, грамотрицательные и грамвариабельные бактериальные клетки, предпочтительно являются грамотрицательными.
Таким образом, микроорганизмы, которые можно применять по настоящему изобретению, включают без ограничения Gluconobacter oxydans, Gluconobacter asaii, Achromobacter delmarvae, Achromobacter viscosus, Achromobacter lacticum, Agrobacterium tumefaciens, Agrobacterium radiobacter, Alcaligenes faecalis, Arthrobacter citreus, Arthrobacter tumescens, Arthrobacter paraffineus, Arthrobacter hydrocarboglutamicus, Arthrobacter oxydans, Aureobacterium saperdae, Azotobacter indicus, Brevibacterium ammoniagenes, Brevibacterium divaricatum, Brevibacterium lactofermentum, Brevibacterium flavum, Brevibacterium globosum, Brevibacterium fuscum, Brevibacterium ketoglutamicum, Brevibacterium helcolum, Brevibacterium pusillum, Brevibacterium testaceum, Brevibacterium roseum, Brevibacterium immariophilium, Brevibacterium linens, Brevibacterium protopharmiae, Corynebacterium acetophilum, Corynebacterium glutamicum, Corynebacterium callunae, Corynebacterium acetoacidophilum, Corynebacterium acetoglutamicum, Enterobacter aerogenes, Erwinia amylovora, Erwinia carotovora, Erwinia herbicola, Erwinia chrysanthemi, Flavobacterium peregrinum, Flavobacterium fucatum, Flavobacterium aurantinum, Flavobacterium rhenanum, Flavobacterium sewanense, Flavobacterium breve, Flavobacterium meningosepticum, Micrococcus sp. CCM825, Morganella morganii, Nocardia opaca, Nocardia rugosa, Planococcus eucinatus, Proteus rettgeri, Propionibacterium shermanii, Pseudomonas synxantha, Pseudomonas azotoformans, Pseudomonas jluorescens, Pseudomonas ovalis, Pseudomonas stutzeri, Pseudomonas acidovolans, Pseudomonas mucidolens, Pseudomonas testosteroni, Pseudomonas aeruginosa, Rhodococcus erythropolis, Rhodococcus rhodochrous, Rhodococcus sp. ATCC 15592, Rhodococcus sp. ATCC 19070, Sporosarcina ureae, Staphylococcus aureus, Vibrio metschnikovii, Vibrio tyrogenes, Actinomadura madurae, Actinomyces violaceochromogenes, Kitasatosporia parulosa, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces flavelus, Streptomyces griseolus, Streptomyces lividans, Streptomyces olivaceus, Streptomyces tanashiensis, Streptomyces virginiae, Streptomyces antibioticus, Streptomyces cacaoi, Streptomyces lavendulae, Streptomyces viridochromogenes, Aeromonas salmonicida, Bacillus pumilus, Bacillus circulans, Bacillus thiaminolyticus, Escherichia freundii, Microbacterium ammoniaphilum, Serratia marcescens, Salmonella typhimurium, Salmonella schottmulleri, Xanthomonas citri, Synechocystis sp., Synechococcus elongatus, Thermosynechococcus elongatus, Microcystis aeruginosa, Nostoc sp., N. commune, N. sphaericum, Nostoc punctiforme, Spirulina platensis, Lyngbya majuscula, L. lagerheimii, Phormidium tenue, Anabaena sp., Leptolyngbya sp. и тому подобное.
В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой эукариотическую клетку. Подходящие эукариотические клетки включают клетки дрожжей, как например: виды рода Saccharomyces, такие как Saccharomyces cerevisiae, виды рода Hansenula, такие как Hansenula polymorpha, виды рода Schizosaccharomyces, такие как Schizosaccharomyces pombe, виды рода Kluyveromyces, такие как Kluyveromyces lactis и Kluyveromyces marxianus, виды рода Yarrowia, такие как Yarrowia lipolytica, виды рода Pichia, такие как Pichia methanolica, Pichia stipites и Pichia pastoris, виды рода Zygosaccharomyces, такие как Zygosaccharomyces rouxii и Zygosaccharomyces bailii, виды рода Candida, такие как Candida boidinii, Candida utilis, Candida freyschussii, Candida glabrata и Candida sonorensis, виды рода Schwanniomyces, такие как Schwanniomyces occidentalis, виды рода Arxula, такие как Arxula adeninivorans, виды рода Ogataea, такие как Ogataea minuta, виды рода Klebsiella, такие как Klebsiella pneumonia.
Особо подходят для применения в раскрываемом в настоящей заявке способах многочисленные бактериальные промышленные штаммы. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Corynebacterium, например, С. acetophilum, С. glutamicum, С. callunae, С. acetoacidophilum, С. acetoglutamicum. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Bacillus, например, В. thuringiensis, В. anthracis, В. megaterium, В. subtilis, В. lentils, В. circulans, В. pumilus, В. lautus, В. coagulans, В. brevis, В. firmus, В. alkaophius, В. licheniformis, В. clausii, В. stearothermophilus, В. halodurans, В. subtilis, В. pumilus и В. amyloliquefaciens. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Erwinia, например, Е. uredovora, Е. carotovora, Е. ananas, Е. herbicola, Е. punctata и Е. terreus. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Escherichia, например, Е. coli. В соответствии с другими вариантами осуществления микроорганизм представляет собой вид из рода Pantoea, например, P. citrea или P. agglomerans. В соответствии с еще одними вариантами осуществления микроорганизм представляет собой вид из рода Streptomyces, например, S. ambofaciens, S. achromogenes, S. avermitilis, S. coelicolor, S. aureofaciens, S. aureus, S. fimgicidicus, S. griseus или S. lividans. В соответствии со следующими вариантами осуществления микроорганизм представляет собой вид из рода Zymomonas, например, Z. mobilis или Z. lipolytica. В соответствии со следующими вариантами осуществления микроорганизм представляет собой вид из рода Rhodococcus, например, R. opacus.
Предпочтительно микроорганизм выбран из семейства Enterobacteriaceae, предпочтительно рода Escherichia, например, Escherichia coli (Е. coli), предпочтительно штамм Е. coli W, соответствующий DSMZ 1116, соответствующий АТСС9637.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (а) уменьшенной, подавленной или исключенной активностью и/или экспрессией гена pflB, кодирующего пируват-формиат-лиазу I, причем уменьшенная, подавленная или исключенная активность и/или экспрессия гена pflB определена в сравнении с соответствующим эталонным микроорганизмом.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-О-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (b) уменьшенной, подавленной или исключенной активностью и/или экспрессией гена adhE, кодирующего бифункциональный фермент на основе ацетальдегид-СоА-дегидрогеназы/зависимой от железа алкогольдегидрогеназы/пируват-формиат-лиаза-дезактивазы, причем уменьшенная, подавленная или исключенная активность и/или экспрессия гена adhE определена в сравнении с соответствующим эталонным микроорганизмом.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-О-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (с) уменьшенной, подавленной или исключенной активностью и/или экспрессией гена ldhA, кодирующего зависимую от NAD ферментативную D-тактат дегидрогеназу, причем уменьшенная, подавленная или исключенная активность и/или экспрессия гена ldhA определена в сравнению с соответствующим эталонным микроорганизмом.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (d) уменьшенной, подавленной или исключенной активностью и/или экспрессией гена pta, кодирующего фосфат-ацетилтрансферазу, причем уменьшенная, подавленная или исключенная активность и/или экспрессия гена pta определена в сравнении с соответствующим эталонным микроорганизмом.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-О-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (е) уменьшенной, подавленной или исключенной активностью и/или экспрессией гена frdA, кодирующего фумарат редуктазу, причем уменьшенная, подавленная или исключенная активность и/или экспрессия гена frdA определена в сравнении с соответствующим эталонным микроорганизмом.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-О-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (f) внесенной, увеличенной или повышенной активностью и/или экспрессией гена alaD, кодирующего аланин-дегидрогеназу, причем увеличение или повышение активности и/или экспрессии гена alaD определено в сравнении с соответствующим эталонным микроорганизмом.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-О-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (g) внесенной, увеличенной или повышенной активностью и/или экспрессией гена ygaW, кодирующего аланиновый транспортер, причем увеличение или повышение активности и/или экспрессии гена ygaW определено в сравнении с соответствующим эталонным микроорганизмом, как описано в WO 2012/172822 и РСТ/IB2014/064426, причем последняя включена с помощью ссылки.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (h) внесенной, увеличенной или повышенной активностью и/или экспрессией гена zipA, кодирующего белок клеточного деления, участвующий в сборке Z-кольца, причем увеличение или повышение активности и/или экспрессии гена zipA определено в сравнении с соответствующим эталонным микроорганизмом, как описано в PCT/IB2014/064426, которая включена в настоящее описание при помощи ссылки.
В дополнение к уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, рекомбинантный микроорганизм по настоящему изобретению может дополнительно характеризоваться (j) внесенной, увеличенной или повышенной активностью и/или экспрессией гена lpd, кодирующего липоамид-дегидрогеназу, причем увеличение или повышение активности и/или экспрессии гена lpd определено в сравнении с соответствующим эталонным микроорганизмом, как описано в WO 2012/172822 и РСТ/IB2014/064426.
Предпочтительно рекомбинантный микроорганизм по настоящему изобретению, характеризующийся по меньшей мере одной из уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, дополнительно характеризуется по меньшей мере четырьмя, еще более предпочтительно по меньшей мере пятью, еще более предпочтительно по меньшей мере шестью, еще более предпочтительно по меньшей мере семью, еще более предпочтительно по меньшей мере восемью, наиболее предпочтительно всеми признаками, выбранными из группы, состоящей из
(a) уменьшенной, подавленной или исключенной активности и/или экспрессии гена pflB, кодирующего пируват-формиат-лиазу I, и
(b) уменьшенной, подавленной или исключенной активности и/или экспрессии гена adhE, кодирующего бифункциональный фермент на основе ацетальдегид-СоА-дегидрогеназы/зависимой от железа алкогольдегидрогеназы/пируват-формиат-лиаза-дезактивазы, и
(c) уменьшенной, подавленной или исключенной активности и/или экспрессии гена ldhA, кодирующего зависимую от NAD ферментативную D-лактатдегидрогеназу, и
(d) уменьшенной, подавленной или исключенной активности и/или экспрессии гена pta, кодирующего фосфат-ацетилтрансферазу, и
(e) уменьшенной, подавленной или исключенной активности и/или экспрессии гена frdA, кодирующего фумаратредуктазу, и
(f) внесенной, увеличенной или повышенной активности и/или экспрессии гена alaD, кодирующего аланин-дегидрогеназу,
(g) внесенной, увеличенной или повышенной активности и/или экспрессии гена ygaW, кодирующего аланиновый транспортер, и
(h) внесенной, увеличенной или повышенной активности и/или экспрессии гена zipA, кодирующего белок клеточного деления, участвующий в сборке Z-кольца, и
(j) привнесенной, увеличенной или повышенной активности и/или экспрессии гена lpd, кодирующего липоамид-дегидрогеназу,
причем уменьшение, подавление, исключение, увеличение или повышение активности и/или экспрессии гена определено в сравнении с соответствующим эталонным микроорганизмом.
Ген alaD может быть получен от любого организма или может быть синтетическим геном, который сконструирован человеком, например, имеющим частоту использования кодона, оптимизированную для экспрессии в рекомбинантном микроорганизме по настоящему изобретению, или оптимизированным для ферментативной активности, например, имеющим повышенный Vmax или Km. Предпочтительно ген alaD получен из микроорганизма одного из родов Bacillus, Geobacillus, Paenibacillus, Halobacillus, Brevibacillus. В соответствии с более предпочтительным вариантом осуществления ген alaD получен из микроорганизма рода Geobacillus. В соответствии с наиболее предпочтительным вариантом осуществления ген alaD получен из Geobacillus stearothermophilus.
В соответствии с предпочтительным вариантом осуществления ген alaD был подвергнут оптимизации кодона для экспрессии в рекомбинантном микроорганизме по настоящему изобретению.
Микроорганизм по настоящему изобретению может содержать дополнительные генетические модификации, такие как мутации, нокауты или повышенные или привнесенные ферментативные активности, которые дополнительно повышают выход и/или производительность по аланину, пирувату, сукцинату, аспартату, манату, лактату, валину и/или лейцину, предпочтительно сукцинату или аланину, более предпочтительно аланину.
В соответствии со следующим вариантом осуществления ген brnQ, кодирующий белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, с уменьшенной, подавленной или исключенной активностью и/или экспрессией в рекомбинантном микроорганизме по настоящему изобретению, выбран из группы, состоящей из
(i) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 1, или
(ii) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 1, или
(iii) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 1 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(iv) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 2, или
(v) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 2,
причем полипептид, кодируемый (ii), (iii) или (v), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 2, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
В соответствии с одним примером ген brnQ, кодирующий белок brnQ, обладающий транспортной активностью в отношении аминокислоты с разветвленной нитью, с уменьшенной, подавленной или исключенной активностью и/или экспрессией в рекомбинантном микроорганизме по настоящему изобретению, имеет последовательность SEQ ID NO: 3, кодирующую белок с SEQ ID NO: 4.
В соответствии со следующим вариантом осуществления ген argP, кодирующий белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, с привнесенной, увеличенной или повышенной активностью и/или экспрессией в рекомбинантном микроорганизме по настоящему изобретению, выбран из группы, состоящей из
(i) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 45, или
(ii) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 45, или
(iii) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 45 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(iv) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 46, или
(v) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 46,
причем полипептид, кодируемый (ii), (iii) или (v), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 46, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
В соответствии с одним примером ген argP, кодирующий белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, с привнесенной, увеличенной или повышенной активностью и/или экспрессией в рекомбинантном организме по настоящему изобретению, имеет последовательность SEQ ID NO: 47, кодирующую белок с SEQ ID NO: 48.
В соответствии со следующим вариантом осуществления ген gcvA, кодирующий ДНК-связывающий белок с привнесенной, увеличенной или повышенной активностью и/или экспрессией в рекомбинантном организме по настоящему изобретению, выбран из группы, состоящей из
(i) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 53, или
(ii) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 53, или
(iii) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 53 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(iv) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 54, или
(v) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 54,
причем полипептид, кодируемый (ii), (iii) или (v), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 54, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
В соответствии со следующим вариантом осуществления ген gcvB, кодирующий РНК, которая не кодирует белок, с уменьшенной, подавленной или исключенной активностью и/или экспрессией в рекомбинантном организме по настоящему изобретению, выбран из группы, состоящей из
(i) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 58, или
(ii) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 58, или
(iii) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 58 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
причем или РНК, которая не кодирует белок, кодируемая (ii), (iii) или (v), обладает по меньшей мере 10%, 20%, предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере на 95% такой же активности, что и у РНК, которая не кодирует белок, с SEQ ID NO: 58, и
причем микроорганизм, содержащий мутантный ген, который определен выше, характеризуется повышенным выходом аланина при ферментации.
В соответствии со следующим вариантом осуществления ген lpxD, кодирующий UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок с измененной активностью и/или экспрессией в рекомбинантном организме по настоящему изобретению, выбран из группы, состоящей из
(i) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 49, или
(ii) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 49, или
(iii) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 49 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(iv) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 50, или
(v) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 50, и
причем кодон генов, обозначенных в (i)-(v), соответствующий положению 43-45 в SEQ ID NO: 49, не кодирует аминокислоту аланин и не является стоп-кодоном, или аминокислота белков, кодируемых генами, обозначенными в (i)-(v), соответствующая положению 15 в SEQ ID NO: 50, не является аланином, и
причем белок, кодируемый геном, который определен выше в (1)-(5), характеризуется измененной активностью и/или экспрессией по сравнению с белком с SEQ ID NO: 50, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
В соответствии с одним примером ген lpxD, кодирующий UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок с измененной активностью и/или экспрессией в рекомбинантном организме по настоящему изобретению, имеет последовательность SEQ ID NO: 51, кодирующую белок с SEQ ID NO: 52.
Рекомбинантный микроорганизм по настоящему изобретению, характеризующийся по меньшей мере одной из уменьшенной, подавленной или исключенной активности и/или экспрессии гена brnQ, кодирующего транспортный белок для аминокислоты с разветвленной нитью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена argP, кодирующего белок argP, обладающий связывающей ДНК / активирующей транскрипцию активностью, и/или привнесенной, увеличенной или повышенной активности и/или экспрессии гена gcvA, кодирующего ДНК-связывающий белок, и/или уменьшенной, подавленной или исключенной активности и/или экспрессии гена gcvB, кодирующего РНК, которая не кодирует белок, и/или измененной активности гена lpxD, кодирующего UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацилтрансферазный белок, может дополнительно характеризоваться любой одной, двумя, тремя, четырьмя, пятью или всеми признаками, которые определены выше под (а)-(j),
причем ген pflB выбран из группы, состоящей из
(A) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 5, или
(B) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 5, или
(C) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 5 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(D) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 6, или
(E) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 6,
причем полипептид, кодируемый (В), (С) или (Е), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 6, и
причем ген adhE выбран из группы, состоящей из
(F) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 7, или
(G) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 7, или
(H) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 7 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(I) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 8, или
(J) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 8,
причем полипептид, кодируемый (G), (Н) или (J), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 8, и
причем ген ldhA выбран из группы, состоящей из
(K) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 9, или
(L) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 9, или
(М) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 9 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(N) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 10, или
(О) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 10,
причем полипептид, кодируемый (L), (М) или (О), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 10, и
причем ген pta выбран из группы, состоящей из
(Р) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 11, или
(Q) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 11, или
(R) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 11 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(S) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 12, или
(Т) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 12,
причем полипептид, кодируемый (Q), (R) или (Т), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 12, и
причем ген frdA выбран из группы, состоящей из
(U) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 13, или
(V) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 13, или
(W) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 13 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(X) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 14, или
(Y) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 14,
причем полипептид, кодируемый (V), (W) или (Y), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 14, и
причем ген alaD выбран из группы, состоящей из
(Z) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 15, или
(АА) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 15, или
(ВВ) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 15 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(СС) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 16, или
(DD) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 16,
причем полипептид, кодируемый (АА), (ВВ) или (DD), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 16, и
причем ген ygaW выбран из группы, состоящей из
(FF) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 109, или
(GG) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 109, или
(НН) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 109 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(JJ) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 110, или
(KK) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 110,
причем полипептид, кодируемый (GG), (НН) или (KK), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 110, и
причем ген zipA выбран из группы, состоящей из
(LL) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 111, или
(ММ) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 111, или
(NN) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 111 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(ОО) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 112, или
(РР) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 112,
причем полипептид, кодируемый (MM), (NN) или (РР), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 112, и
причем ген lpd выбран из группы, состоящей из
(QQ) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 113, или
(RR) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 113, или
(SS) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 113 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(ТТ) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 114, или
(UU) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 114,
причем полипептид, кодируемый (RR), (SS) или (UU), обладает по меньшей мере 10%, 20% предпочтительно по меньшей мере 30% или 50%, более предпочтительно по меньшей мере 60% или 70%, еще более предпочтительно по меньшей мере 75%, 80%, 85% или 90%, наиболее предпочтительно по меньшей мере 95% такой же активности, что и у полипептида с SEQ ID NO: 114.
Предпочтительно молекула нуклеиновой кислоты, которая определена в (Z)-(DD), находится под контролем последовательности, функционирующей как промотор в микроорганизме с последовательностью
(1) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 115 или 116, или
(2) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 115 или 116, или
(3) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 115 или 116 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(4) фрагмента по меньшей мере из 10 нуклеотидов, предпочтительно по меньшей мере из 20 нуклеотидов, по меньшей мере из 30 нуклеотидов или по меньшей мере из 40 нуклеотидов, более предпочтительно фрагмента по меньшей мере из 50 нуклеотидов, по меньшей мере из 75 нуклеотидов или по меньшей мере из 100 нуклеотидов, еще более предпочтительно по меньшей мере из 150 или по меньшей мере из 200 нуклеотидов молекулы нуклеиновой кислоты с SEQ ID NO: 115 или 116. Предпочтительно фрагмент SEQ ID NO: 115 или 116 представляет собой фрагмент, содержащий 3' участок SEQ ID NO: 115 или 116, таким образом, фрагмент характеризуется делецией на 5' конце SEQ ID NO: 115 или 116.
В соответствии со следующим вариантом осуществления настоящее изобретение относится к композиции, содержащей один или более описанных выше рекомбинантных микроорганизмов по настоящему изобретению. Композиция может дополнительно содержать среду, которая обеспечивает рост рекомбинантного микроорганизма по настоящему изобретению. Среда может дополнительно содержать источник углерода, такой как гексозы, пентозы или полиолы, например, сахароза, глюкоза, фруктоза, галактоза, манноза, раффиноза, ксилоза, арабиноза, ксилулоза, глицерин, маннит, арабит, ксилит, крахмал, целлюлоза, лигноцеллюлоза или их комбинации. Предпочтительно источник углерода представляет собой глюкозу или сахарозу, более предпочтительно источником углерода является глюкоза.
В соответствии с предпочтительным вариантом осуществления композиция содержит микроорганизм по настоящему изобретению и среду NBS, среду AM1 или среду РРМ01. Более предпочтительно композиция дополнительно содержит источник углерода, предпочтительно сахар. Ингредиенты таких сред известны специалисту в настоящей области техники.
Предпочтительно среда NBS содержит на литр
1-5 г, предпочтительно 3,5 г KH2PO4, и
1-10 г, предпочтительно 5,0 г K2HPO4, и
1-5 г, предпочтительно 3,5 г (NH4)2HPO4, и
0,1-1 г, предпочтительно 0,25 г MgSO4 - 7 Н2О, и
5-25 мг, предпочтительно 15 мг CaCl2 - 2 Н2О, и
0,1-1 мг, предпочтительно 0,5 мг тиамина, и
0,1-5 мл, предпочтительно 1 мл исходного раствора металлических микроэлементов,
причем исходный раствор металлических микроэлементов содержит 0,5-5 г, предпочтительно 1,6 г FeCl3-6 Н2О; 0,05-0,5 г, предпочтительно 0,2 г CoCl2-6 Н2О; 0,01-0,5 г, предпочтительно 0,1 г CuCl2-2 Н2О; 0,1-0,5 г, предпочтительно 0,2 г ZnCl2; 0,05-0,5 г, предпочтительно 0,2 г NaMoO4-2 H2O; 0,001-0,1 г, предпочтительно 0,05 г Н3ВО3 на литр 0,01-1 М, предпочтительно 0,1 М HCl.
Предпочтительным источником углерода в среде NBS является глюкоза или сахароза, предпочтительно 2%-18% глюкоза или 2%-16% сахароза.
Предпочтительно среда AM 1 содержит на литр 0,1-10 мМ, предпочтительно 1 мМ раствор бетаина,
1-10 г, предпочтительно 2,6 г (NH4)2HPO4, и
0,1-5 г, предпочтительно 0,87 г NH4H2PO4, и
0,05-2,5 г, предпочтительно 0,15 г KCl, и
0,05-5 г, предпочтительно 0,37 г MgSO4-7H2O, и
0,1-5 мл, предпочтительно 1 мл исходного раствора металлических микроэлементов,
причем исходный раствор металлических микроэлементов содержит на литр 0,01-1 М, предпочтительно 0,12 М HCl, 1-5 г, предпочтительно 2,4 г FeCl3-6Н2О; 0,1-1 г, предпочтительно 0,3 г CoCl2-6H2O; 0,1-1 г, предпочтительно 0,21 г CuCl2-2H2O; 0,1-1 г, предпочтительно 0,3 г ZnCl2; 0,1-1 г, предпочтительно 0,27 г NaMoO4-2Н2О; 0,01-0,5 г, предпочтительно 0,068 г Н3ВО3 и 0,1-1 г, предпочтительно 0,5 г MnCl2-4H2O,
и необязательно 1-30 г, предпочтительно 15 г (NH4)2SO4.
Предпочтительным источником углерода в среде NBS является глюкоза или сахароза, предпочтительно 2%-18% глюкоза или 2%-16% сахароза.
Предпочтительно среда РРМ01 содержит на литр
0,05-5 г, предпочтительно 0,37 г MgSO4-7H2O, и
0,1-10 г, предпочтительно 1 г (NH4)2SO4, и
0,05-5 г, предпочтительно 0,46 г бетаина, и
0,001-0,5 г, предпочтительно 0,05 г цианокобаламина (В12), и
1-10 г, предпочтительно 3,74 г KH2PO4, и
0,1-5 мл, предпочтительно 1 мл исходного раствора металлических микроэлементов,
причем исходный раствор металлических микроэлементов содержит на литр 10-100 мМ, предпочтительно 60 мМ серной кислоты, 1-10 г, предпочтительно 3,48 г (NH4)2Fe(II)(SO4)2-7H2O; 0,1-1 г, предпочтительно 0,35 г CoSO4-7H2O; 0,1-1 г, предпочтительно 0,31 г CuSO4-5H2O; 0,1-5 г, предпочтительно 0,63 г ZnSO4 - 7Н2О; 0,1-1 г, предпочтительно 0,27 г MnSO4-H2O; 0,01-1 г, предпочтительно 0,07 г NaMoO4-2Н2О и 0,1-5 г, предпочтительно 0,43 г Н3ВО3.
Предпочтительным источником углерода в среде РРМ01 является моногидрат глюкозы, предпочтительно 10-500 г, более предпочтительно 140 г моногидрата глюкозы на литр среды.
В соответствии со следующим вариантом осуществления настоящее изобретение относится к способу получения рекомбинантного микроорганизма с повышенным выходом или производительностью по аланину, пирувату, сукцинату, аспартату, малату, лактату, валину и/или лейцину, предпочтительно сукцинату или аланину, более предпочтительно аланину, который включает следующие стадии:
(I) i) уменьшение, подавление или исключение одной или более активности и/или экспрессии гена brnQ или согласно определению ранее под (i)-(v), и/или ii) внесение, увеличение или повышение одной или более активности и/или экспрессии гена argP, согласно определению ранее под (i)-(v), и/или iii) внесение, увеличение или повышение одной или более активности и/или экспрессии гена gcvA, согласно определению ранее под (i)-(v), и/или iv) уменьшение, подавление или исключение одной или более активности и/или экспрессии гена gcvB, согласно определению под (i)-(v), и/или v) изменение активности гена lpxD, согласно определению ранее под (i)-(v), в микроорганизме; и
(II) получение, выявление и выделение рекомбинантного микроорганизма с повышенным выходом и производительностью по аланину, пирувату, сукцинату, аспартату, малату, лактату, валину и/или лейцину, предпочтительно сукцинату или аланину, более предпочтительно аланину, по сравнению с соответствующим микроорганизмом без модификации, которая определена выше под (I).
В соответствии с предпочтительным вариантом осуществления ген brnQ с уменьшенной, подавленной или исключенной активностью и/или экспрессией имеет последовательность SEQ ID NO: 3 и/или кодирует полипептид с SEQ ID NO: 4.
В соответствии с предпочтительным вариантом осуществления ген argP с привнесенной, увеличенной или повышенной активностью и/или экспрессией имеет последовательность SEQ ID NO: 47 и/или кодирует полипептид с SEQ ID NO: 48.
В соответствии с предпочтительным вариантом осуществления ген gcvA с привнесенной, увеличенной или повышенной активностью и/или экспрессией функционально связан с промотором, имеющим последовательность SEQ ID NO: 56 или 57.
В соответствии с предпочтительным вариантом осуществления ген gcvB с уменьшенной, подавленной или исключенной активностью и/или экспрессией функционально связан с промотором, имеющим последовательностью с SEQ ID NO: 60 или 61.
В соответствии с предпочтительным вариантом осуществления ген lpxD с измененной активностью и/или экспрессией имеет последовательность SEQ ID NO: 51 и/или кодирует полипептид с SEQ ID NO: 52.
В соответствии с предпочтительным вариантом осуществления способа получения рекомбинантного микроорганизма по настоящему изобретению способ дополнительно включает стадию уменьшения, подавления или исключение активности и/или экспрессии по меньшей мере одного, по меньшей мере двух, по меньшей мере трех, по меньшей мере четырех или всех из гена pflB, гена adhE, гена ldhA, гена pta или гена frdA, например, которые определены выше под (А)-(Y), и/или стадию внесения, увеличения или повышения активности и/или экспрессии по меньшей мере одного, по меньшей мере двух, по меньшей мере трех или всех из гена alaD, гена ygaW, гена zipA или гена lpd, например, которые определены выше под (Z)-(UU).
В соответствии с дополнительным предпочтительным вариантом осуществления способа получения рекомбинантного микроорганизма по настоящему изобретению молекула нуклеиновой кислоты, которая определена выше в (Z)-(DD), находится под контролем последовательности, функционирующей как промотор в микроорганизме, имеющем последовательность
(1) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 115 или 116, или
(2) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 115 или 116, или
(3) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 115 или 116 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(4) фрагмента по меньшей мере из 10 нуклеотидов, предпочтительно по меньшей мере из 20 нуклеотидов, по меньшей мере из 30 нуклеотидов или по меньшей мере из 40 нуклеотидов, более предпочтительно фрагмента по меньшей мере из 50 нуклеотидов, по меньшей мере из 75 нуклеотидов или по меньшей мере из 100 нуклеотидов, еще более предпочтительно по меньшей мере из 150 или по меньшей мере из 200 нуклеотидов молекулы нуклеиновой кислоты с SEQ ID NO: 115 или 116. Предпочтительно фрагмент SEQ ID NO: 115 или 116 представляет собой фрагмент, содержащий 3' участок SEQ ID NO: 115 или 116, таким образом, фрагмент характеризуется делецией на 5' конце SEQ ID NO: 115 или 116.
Наиболее предпочтительный способ получения рекомбинантного микроорганизма по настоящему изобретению включает стадию уменьшения, подавления или исключения активности и/или экспрессии всех из гена brnQ, гена gcvB, гена pflB, гена adhE, гена ldhA, гена pta и гена frdA, и стадию внесения, увеличения или повышения активности и/или экспрессии всех из гена alaD, гена ygaW, гена zipA, гена lpd, гена argP и гена gcvA, и стадию изменения активности и/или экспрессии гена lpxD.
В соответствии с одним вариантом осуществления способа получения рекомбинантного микроорганизма по настоящему изобретению микроорганизм выбран из группы, состоящей из прокариотических микроорганизмов, включая Gluconobacter oxydans, Gluconobacter asaii, Achromobacter delmarvae, Achromobacter viscosus, Achromobacter lacticum, Agrobacterium tumefaciens, Agrobacterium radiobacter, Alcaligenes faecalis, Arthrobacter citreus, Arthrobacter tumescens, Arthrobacter paraffineus, Arthrobacter hydrocarboglutamicus, Arthrobacter oxydans, Aureobacterium saperdae, Azotobacter indicus, Brevibacterium ammoniagenes, Brevibacterium divaricatum, Brevibacterium lactofermentum, Brevibacterium flavum, Brevibacterium globosum, Brevibacterium fuscum, Brevibacterium ketoglutamicum, Brevibacterium helcolum, Brevibacterium pusillum, Brevibacterium testaceum, Brevibacterium roseum, Brevibacterium immariophilium, Brevibacterium linens, Brevibacterium protopharmiae, Corynebacterium acetophilum, Corynebacterium glutamicum, Corynebacterium callunae, Corynebacterium acetoacidophilum, Corynebacterium acetoglutamicum, Enterobacter aerogenes, Erwinia amylovora, Erwinia carotovora, Erwinia herbicola, Erwinia chrysanthemi, Flavobacterium peregrinum, Flavobacterium fucatum, Flavobacterium aurantinum, Flavobacterium rhenanum, Flavobacterium sewanense, Flavobacterium breve, Flavobacterium meningosepticum, Micrococcus sp. CCM825, Morganella morganii, Nocardia opaca, Nocardia rugosa, Planococcus eucinatus, Proteus rettgeri, Propionibacterium shermanii, Pseudomonas synxantha, Pseudomonas azotoformans, Pseudomonas jluorescens, Pseudomonas ovalis, Pseudomonas stutzeri, Pseudomonas acidovolans, Pseudomonas mucidolens, Pseudomonas testosteroni, Pseudomonas aeruginosa, Rhodococcus erythropolis, Rhodococcus rhodochrous, Rhodococcus sp. ATCC 15592, Rhodococcus sp. ATCC 19070, Sporosarcina ureae, Staphylococcus aureus, Vibrio metschnikovii, Vibrio tyrogenes, Actinomadura madurae, Actinomyces violaceochromogenes, Kitasatosporia parulosa, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces flavelus, Streptomyces griseolus, Streptomyces lividans, Streptomyces olivaceus, Streptomyces tanashiensis, Streptomyces virginiae, Streptomyces antibioticus, Streptomyces cacaoi, Streptomyces lavendulae, Streptomyces viridochromogenes, Aeromonas salmonicida, Bacillus pumilus, Bacillus circulans, Bacillus thiaminolyticus, Escherichia freundii, Microbacterium ammoniaphilum, Serratia marcescens, Salmonella typhimurium, Salmonella schottmulleri, Xanthomonas citri, Synechocystis sp., Synechococcus elongatus, Thermosynechococcus elongatus, Microcystis aeruginosa, Nostoc sp., N. commune, N. sphaericum, Nostoc punctiforme, Spirulina platensis, Lyngbya majuscula, L. lagerheimii, Phormidium tenue, Anabaena sp., Leptolyngbya sp и тому подобное.
В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой эукариотическую клетку. Подходящие эукариотические клетки включают клетки дрожжей, как, например: виды рода Saccharomyces, такие как Saccharomyces cerevisiae, виды рода Hansenula, такие как Hansenula polymorpha, виды рода Schizosaccharomyces, такие как Schizosaccharomyces pombe, виды рода Kluyveromyces, такие как Kluyveromyces lactis и Kluyveromyces marxianus, виды рода Yarrowia, такие как Yarrowia lipolytica, виды рода Pichia, такие как Pichia methanolica, Pichia stipites и Pichia pastoris, виды рода Zygosaccharomyces, такие как Zygosaccharomyces rouxii и Zygosaccharomyces bailii, виды рода Candida, такие как Candida boidinii, Candida utilis, Candida freyschussii, Candida glabrata и Candida sonorensis, виды рода Schwanniomyces, такие как Schwanniomyces occidentalis, виды рода Arxula, такие как Arxula adeninivorans, виды рода Ogataea, такие как Ogataea minuta, виды рода Klebsiella, такие как Klebsiella pneumonia.
Особо подходят для применения в раскрываемых в настоящей заявке способах многочисленные бактериальные промышленные штаммы. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Corynebacterium, например, С. acetophilum, С. glutamicum, С. callunae, С. acetoacidophilum, С. acetoglutamicum. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Bacillus, например, В. thuringiensis, В. anthracis, В. megaterium, В. subtilis, В. lentils, В. circulans, В. pumilus, В. lautus, В. coagulans, В. brevis, В. firmus, В. alkaophius, В. licheniformis, В. clausii, В. stearothermophilus, В. halodurans, В. subtilis, В. pumilus и В. amyloliquefaciens. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Erwinia, например, Е. uredovora, Е. carotovora, Е. ananas, Е. herbicola, Е. punctata и Е. terreus. В соответствии с некоторыми вариантами осуществления микроорганизм представляет собой вид из рода Escherichia, например, Е. coli. В соответствии с другими вариантами осуществления микроорганизм представляет собой вид из рода Pantoea, например, P. citrea или P. agglomerans. В соответствии с еще одними вариантами осуществления микроорганизм представляет собой вид из рода Streptomyces, например, S. ambofaciens, S. achromogenes, S. avermitilis, S. coelicolor, S. aureofaciens, S. aureus, S. fungicidicus, S. griseus или S. lividans. В соответствии со следующими вариантами осуществления микроорганизм представляет собой вид из рода Zymomonas, например, Z. mobilis или Z. lipolytica. В соответствии со следующими вариантами осуществления микроорганизм представляет собой вид из рода Rhodococcus, например R opacus.
Предпочтительно микроорганизм выбран из семейства Enterobacteriaceae, предпочтительно рода Escherichia, например, Escherichia coli (Е. coli), предпочтительно штамм Е. coli W, соответствующий DSMZ 1116, соответствующий АТСС9637.
Следующий вариант осуществления по настоящему изобретению относится к способу получения аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, наиболее предпочтительно L-аланина, предусматривающему культивирование одного или более рекомбинантных микроорганизмов, которые определены выше, в условиях, которые обеспечивают производство аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, наиболее предпочтительно L-аланина.
В соответствии с некоторыми вариантами осуществления рекомбинантные микроорганизмы, к которым относится настоящее изобретение, выращивают в условиях периодической или непрерывной ферментации. Классическая периодическая ферментация представляет собой закрытую систему, в которой варианты состава среды задают в начале ферментации и не подвергают искусственным изменениям в ходе ферментации. Вариантом периодической системы является периодическая ферментация с добавлением субстрата. В соответствии с таким вариантом субстрат добавляют частями по мере прогрессирования ферментации. Системы с добавлением субстрата пригодны в том случае, когда катаболитная репрессия вероятно ингибирует метаболизм клеток, и там, где желательно наличие ограниченных количеств субстрата в среде. Периодическая ферментация и периодическая ферментация с добавлением субстрата являются распространенными и хорошо известными в настоящей области техники. Непрерывная ферментация, которая также находит применение в настоящем изобретении, представляет собой систему, в которую определенную среду для ферментации непрерывно добавляют в биореактор, а равное количество кондиционированной среды (например, содержащей необходимый конечный продукт) одновременно удаляют для переработки. При непрерывной ферментации культуры, как правило, сохраняются с постоянной высокой плотностью, при которой клетки преимущественно находятся в фазе роста, в которой повышено производство конечных продуктов. Системы непрерывной ферментации направлены на сохранение условий стационарного роста. Способы модулирования питательных веществ и факторов роста для процессов непрерывной ферментации, а также методики максимизации скорости образования продукта хорошо известны в области промышленной микробиологии.
В соответствии с некоторыми вариантами осуществления ферментации осуществляют при температуре в рамках диапазона от приблизительно 10°С до приблизительно 60°С, от приблизительно 15°С до приблизительно 50°С, от приблизительно 20°С до приблизительно 45°С, от приблизительно 25°С до приблизительно 45°С, от приблизительно 30°С до приблизительно 45°С и от приблизительно 25°С до приблизительно 40°С. В соответствии с предпочтительным вариантом осуществления температура составляет приблизительно 34°С, 35°С или 36°С. В соответствии с наиболее предпочтительным вариантом осуществления температура составляет приблизительно 37°С или 38°С.
В соответствии с некоторыми другими вариантами осуществления ферментацию осуществляют в течение периода времени в рамках диапазона от приблизительно 8 часов до 240 часов, от приблизительно 8 часов до приблизительно 168 часов, от приблизительно 10 часов до приблизительно 144 часов, от приблизительно 15 часов до приблизительно 120 часов или от приблизительно 20 часов до приблизительно 72 часов. Предпочтительно ферментацию осуществляют от приблизительно 20 часов до приблизительно 40 часов.
В соответствии с некоторыми другими вариантами осуществления ферментацию осуществляют при рН в диапазоне от приблизительно 4 до приблизительно 9, в диапазоне от приблизительно 4,5 до приблизительно 8,5, в диапазоне от приблизительно 5 до приблизительно 8 или в диапазоне от приблизительно 5,5 до приблизительно 7,5. Предпочтительно ферментация будет осуществляться при рН 7.
В соответствии с одним вариантом осуществления способа получения аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, микроорганизм культивируют в среде, содержащей от 1% до 30% (масса/объем) сахара, от 5% до 25% (масса/объем) сахара, от 10% до 20% (масса/объем) сахара, от 11% до 18% (масса/объем) сахара. Предпочтительно микроорганизм культивируют в среде, содержащей от 12% до 16% (масса/объем) сахара. Более предпочтительно микроорганизм культивируют в среде, содержащей от 13% до 15% (масса/объем) сахара, наиболее предпочтительно микроорганизм культивируют в среде, содержащей 14% (масса/объем) сахара.
В соответствии с другим вариантом осуществления способа получения аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, выход аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина составляет по меньшей мере 80%, например, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84% или по меньшей мере 85%. Предпочтительно выход составляет по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89% или по меньшей мере на 90%. Более предпочтительно выход составляет по меньшей мере 90,5%, по меньшей мере 91%, по меньшей мере 91,5%, по меньшей мере 92%, по меньшей мере 92,5%, по меньшей мере 93%, по меньшей мере 93,5%, по меньшей мере 94% или по меньшей мере 94,5%. В соответствии с еще более предпочтительным вариантом осуществления выход составляет по меньшей мере 95% или по меньшей мере 95,5%. В соответствии с наиболее предпочтительным вариантом осуществления выход составляет по меньшей мере 96%. Процент выхода рассчитывают в виде грамма продукта, произведенного из грамма глюкозы, в среде. Таким образом, если среда содержала 100 г глюкозы, а на выходе ферментации было 98 г аланина, то выход составит 98%.
В соответствии с другим вариантом осуществления способа получения аланина производят предпочтительно L-аланин, причем хиральная чистота L-аланина составляет по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93% или по меньшей мере 94%. В соответствии с предпочтительным вариантом осуществления хиральная чистота L-аланина составляет по меньшей мере 95% или по меньшей мере 95,5%. В соответствии с более предпочтительным вариантом осуществления хиральная чистота L-аланина составляет по меньшей мере 96% или по меньшей мере 96,5%, или по меньшей мере 97%. В соответствии с еще более предпочтительным вариантом осуществления хиральная чистота L-аланина составляет по меньшей мере 97,5% или по меньшей мере 98%, или по меньшей мере 98,5%, или по меньшей мере 99%. Еще более предпочтительно хиральная чистота L-аланина составляет по меньшей мере 99,5% или по меньшей мере 99,6%, например, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9%. В соответствии с наиболее предпочтительным вариантом осуществления вырабатывается хирально чистый L-аланин.
В соответствии с другим вариантом осуществления настоящее изобретение относится к способу культивирования или выращивания любого из генетически модифицированных микроорганизмов, которые определены выше, причем способ включает инокуляцию культуральной среды одним или более генетически модифицированными микроорганизмами и культивирование или выращивание указанного генетически модифицированного микроорганизма в культуральной среде в условиях, которые определены выше.
Еще одним дополнительным вариантом осуществления является применение рекомбинантного микроорганизма, который определен выше, или композиции, которая определена выше, для ферментативного получения аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, наиболее предпочтительно L-аланина.
Рекомбинантный микроорганизм в соответствии с настоящим изобретением характеризуется тем, что, по сравнению с соответствующим эталонным микроорганизмом, например дикого типа, понижена экспрессия и/или активность фермента, который кодируется геном brnQ, и/или РНК, которая кодируется геном gcvB, и/или увеличена экспрессия и/или активность фермента, который кодируется геном argP и/или геном gcvA, и/или изменена активность фермента, кодируемого геном lpxD.
Термин «пониженная экспрессия и/или активность» также охватывает микроорганизм дикого типа, у которого нельзя детектировать экспрессию и/или активность brnQ и/или gcvB.
В соответствии с одним вариантом осуществления понижение экспрессии и/или активности гена достигают посредством дезактивации, мутации или нокаута гена. Это можно осуществлять при помощи делеции части или полного кодирующего участка и/или промотора гена, при помощи мутации гена, такой как вставка или делеция некоторого количества нуклеотидов, например, одного или двух нуклеотидов, что приводит к сдвигу рамки в кодирующем участке гена, введение стоп-кодонов в кодирующий участок, инактивация промотора гена при помощи, например, делеции или мутирования промоторных областей, таких как стороны входа в рибосому, ТАТА-бокс и т.п. Понижение также может быть достигнуто в результате деградации транскрипта гена, например, путем внесения рибозимов, dsRNA, антисмысловой РНК или антисмысловых олигонуклеотидов. Понижение активности гена может быть достигнуто в результате экспрессии в клетке антител или аптамеров, которые специфически связывают целевой фермент. Специалисту в настоящей области известны другие способы понижения экспрессии и/или активности гена.
В соответствии с предпочтительным вариантом осуществления понижение экспрессии и/или активности гена brnQ достигают посредством внесения в ген мутации, предпочтительно делеции. В соответствии с дополнительным предпочтительным вариантом осуществления делецию вносят между положениями 667 и 764 в SEQ ID NO: 1, таким образом исключая 97 нуклеотидов из brnQ. Полученная в результате укороченная нуклеиновая кислота имеет последовательность, которая отображена в SEQ ID NO: 3 и кодирует укороченный белок, который отображен в SEQ ID NO: 4.
В соответствии с предпочтительным вариантом осуществления повышение экспрессии и/или активности гена argP достигают посредством внесения мутации в ген, предпочтительно точечной мутации. Более предпочтительно этого достигают посредством мутации кодона в положениях 286-288 гена argP с SEQ ID NO: 45 или соответствующего кодона функционального гомологичного гена. Еще более предпочтительно кодон подвергают мутации так, чтобы он действительно кодировал такую аминокислоту, как глутаминовая кислота, или другую кислую аминокислоту или их амид, или аминокислоту, аналогичную глутаминовой кислоте, но не аланин. В соответствии с наиболее предпочтительным вариантом осуществления соответствующий кодон подвергают мутации так, чтобы он кодировал такую аминокислоту, как глутаминовая кислота.
Предпочтительно увеличение экспрессии и/или активности гена argP достигают посредством внесения мутации в ген argP, причем мутантный argP имеет последовательность SEQ ID NO: 47, кодирующую белок с SEQ ID NO: 48.
Предпочтительно увеличение экспрессии и/или активности гена gcvA достигают посредством внесения мутации в промотор гена gcvA, причем мутантный промотор предпочтительно имеет последовательность SEQ ID NO: 56 или SEQ ID NO: 57.
В соответствии с предпочтительным вариантом осуществления понижение экспрессии и/или активности гена gcvB достигают посредством внесения мутации в промотор. Например, промотор можно подвергнуть мутации путем внесения делеции по любому одному или более оснований Т в положении 62-68 в SEQ ID NO: 59 или посредством внесения точечной мутации в положении 60 в SEQ ID NO: 59, изменяя А в данном положении на любой из G, С или Т. Предпочтительно мутантный промотор имеет последовательность SEQ ID NO: 60 или 61.
Предпочтительно понижение экспрессии и/или активности гена gcvB достигают посредством внесения мутации в промотор гена gcvB. Предпочтительно промотор дикого типа с SEQ ID NO: 59 подвергают мутации так, чтобы он имел последовательность SEQ ID NO: 60 или 61.
Предпочтительно измененную экспрессию и/или активность гена lpxD достигают посредством внесения мутации в ген lpxD, причем мутантный lpxD имеет последовательность SEQ ID NO: 51, кодирующую белок с SEQ ID NO: 52.
Уменьшенная экспрессия и/или активность РНК или ферментов, соответственно раскрываемых в настоящем документе, в частности, уменьшенная экспрессия и/или уменьшенная активность РНК или фермента, кодируемого посредством лактатдегидрогеназы (ldhA), пируват-формиат-лиазы I (pflB), бифункционального фермента на основе ацетальдегид-СоА-дегидрогеназы/зависимой от железа алкогольдегидрогеназы/пируват-формиат-лиаза-дезактивазы (adhE), фосфат-ацетилтрансферазы (pta), фумаратредуктазы (frdA), gcvB и/или brnQ, может представлять собой уменьшение экспрессии и/или активности по меньшей мере на 50% по сравнению с экспрессией и/или активностью указанных РНК или фермента у соответствующего эталонного микроорганизма, например, дикого типа микроорганизма, или уменьшение экспрессии и/или активности по меньшей мере на 90%, или более предпочтительно уменьшение экспрессии и/или активности по меньшей мере на 95%, или более предпочтительно уменьшение экспрессии и/или активности по меньшей мере на 98%, или еще более предпочтительно уменьшение экспрессии и/или активности по меньшей мере на 99%, или еще более предпочтительно уменьшение экспрессии и/или активности по меньшей мере на 99,9%. В соответствии с наиболее предпочтительным вариантом осуществления экспрессию и/или активность РНК или ферментов нельзя детектировать в микроорганизме по настоящему изобретению.
Потеря экспрессии и/или активности гена gcvB и/или гена brnQ и привнесенная или увеличенная экспрессия и/или активность гена argP и/или гена gvcA, а также измененная активность и/или экспрессия гена lpxD приводит к повышенному выходу и/или производительности по аланину, пирувату, сукцинату, аспартату, малату, лактату, валину и/или лейцину, предпочтительно сукцинату или аланину, более предпочтительно аланину, в рекомбинантном организме по настоящему изобретению по сравнению с соответствующим эталонным микроорганизмом. Таким образом, утрата экспрессии и/или активности гена brnQ и/или гена gcvB и привнесение или увеличение экспрессии и/или активности гена argP и/или гена gvcA, а также изменение активности и/или экспрессии гена lpxD можно определить посредством измерения выхода и/или производительности по аланину, пирувату, сукцинату, аспартату, малату, лактату, валину и/или лейцину, предпочтительно по сукцинату или аланину, более предпочтительно по аланину у рекомбинантного микроорганизма по настоящему изобретению в сравнении с соответствующим эталонным микроорганизмом. Способы ферментативного получения метаболитов, например аланина, известны специалистам в настоящей области, а также описаны в настоящем документе. Повышенный выход, например аланина, при ферментации микроорганизмом по настоящему изобретению по сравнению с выходом аланина при ферментации соответствующим эталонным микроорганизмом является показателем утраты, уменьшения, внесения, или увеличения, или изменения экспрессии и или активности соответствующего гена.
Способы определения экспрессии или активности лактатдегидрогеназы (ldhA), например, раскрыты в работе "The ldhA gene encoding the fermentative lactate de hydrogenase of Escherichia Coli", Microbiology (1997), Vol. 143, pages 187-155; или Bergmeyer, H.U., Bergmeyer J. and Grassl, M. (1983-1986) in "Methods of Enzymatic Analysis", 3rd Edition, Volume III, pages 126-133, Verlag Chemie, Weinheim; или Enzymes in Industry: Production and Applications, Second Edition (2004), Wolfgang Aehle, page 23. Предпочтительным является последний способ.
Способы определения экспрессии или активности пируват-формиат-лиазы I (pflB), например, раскрыты в работе Knappe J, Blaschkowski HP, Grobner Р, Schmitt Т (1974). "Pyruvate formate-lyase of Escherichia coli: the acetyl-enzyme intermediate." Eur J Biochem 1974; 50(1); 253-63. PMID: 4615902; в работе KNAPPE, Joachim, et al. "Pyruvate Formate-Lyase of Escherichia coli: the Acetyl-Enzyme Intermediate." European Journal of Biochemistry 50.1 (1974): 253-263; в работе Wong, Kenny K., et al. "Molecular properties of pyruvate formate-lyase activating enzyme." Biochemistry 32.51 (1993): 14102-14110 и в работе Nnyepi, Mbako R., Yi Peng, and Joan B. Broderick. "Inactivation of E. coli pyruvate formate-lyase: Role of AdhE and small molecules." Archives of biochemistry and biophysics 459.1 (2007): 1-9.
Способы определения экспрессии или активности бифункционального фермента на основе ацетальдегид-СоА-дегидрогеназы/зависимой от железа алкогольдегидрогеназы/пируват-формиат-лиаза-дезактивазы (adhE), например, раскрыты в работе Membrillo-Hernandez, Jorge, et al. "Evolution of the adhE Gene Product of Escherichia coli from a Functional Reductase to a Dehydrogenase GENETIC AND BIOCHEMICAL STUDIES OF THE MUTANT PROTEINS." Journal of Biological Chemistry 275.43 (2000): 33869-33875, и в работе Mbako R. Nnyepi, Yi Peng, Joan B. Broderick, Inactivation of E. coli pyruvate formate-lyase: Role of AdhE and small molecules, Archives of Biochemistry and Biophysics, Volume 459, Issue 1, 1 March 2007, Pages 1-9.
Способы определения экспрессии или активности фосфат-ацетилтрансферазы (pta), например, раскрыты в работе Dittrich, Cheryl R., George N. Bennett, and Ka-Yiu San. "Characterization of the Acetate-Producing Pathways in Escherichia coli." Biotechnology progress 21.4 (2005): 1062-1067, и в работе Brown, Т. D. K., М.С. Jones-Mortimer, and Н.L. Kornberg. "The enzymic interconversion of acetate and acetyl-coenzyme A in Escherichia coli." Journal of general microbiology 102.2 (1977): 327-336.
Способы определения экспрессии или активности фумаратредуктазы (frdA), например, раскрыты в работе Dickie, Peter, and Joel H. Weiner. "Purification and characterization of membrane-bound fumarate reductase from anaerobically grown Escherichia coli." Canadian journal of biochemistry 57.6 (1979): 813-821; in Cecchini, Gary, et al. "Reconstitution of quinone reduction and characterization of Escherichia coli fumarate reductase activity." Journal of Biological Chemistry 261.4 (1986): 1808-1814, или в работе Schroder, I., et al. "Identification of active site residues of Escherichia coli fumarate reductase by site-directed mutagenesis." Journal of Biological Chemistry 266.21 (1991): 13572-13579.
Способ определения экспрессии или активности аланин-дегидрогеназы (alaD), например, раскрыт в работе Sakamoto, Y., Nagata, S., Esaki, N., Tanaka, H., Soda, K. "Gene cloning, purification and characterization of thermostable alanine dehydrogenase of Bacillus stearothermophilus" J Fermen. Bioeng. 69 (1990):154-158.
Термин «уменьшенная экспрессия фермента» включает, например, экспрессию фермента указанным подвергнутым генетическим манипуляциям (например, генетически сконструированным) микроорганизмом на более низком уровне, чем экспрессируемый соответствующим эталонным микроорганизмом, например, диким типом указанного микроорганизма. Генетические манипуляции для уменьшения экспрессии фермента могут включать без ограничения делению гена или его частей, который кодирует фермент, изменение или модификацию регуляторных последовательностей или сайтов, ассоциированных с экспрессией гена, кодирующего фермент (например, посредством удаления сильных промоторов или репрессируемый промоторов), модификацию белков (например, регуляторных белков, супрессоров, энхансеров, транскрипционных активаторов и т.п.), вовлеченных в транскрипцию гена, кодирующего фермент, и/или трансляцию продукта гена, или любые другие традиционные средства снижения экспрессии конкретного гена, которые являются общепринятыми в настоящей области (включая без ограничения применение антисмысловых молекул нуклеиновой кислоты или другие способы нокаута или блокировки экспрессии целевого белка). Кроме того, в мРНК можно внести дестабилизирующие элементы или внести генетические модификации, приводящие к нарушению сайтов связывания с рибосомами (RBS) у РНК. Также можно изменить частоту использования кодона у гена таким образом, чтобы снизить эффективность и скорость трансляции.
Уменьшенную активность фермента также можно получить посредством внесения одной или более вредных мутаций в ген, которые приводят к уменьшенной активности фермента. Кроме того, уменьшение активности фермента также может включать инактивацию (или уменьшенную экспрессию) активирующих ферментов, которые необходимы для активации фермента, активность которого необходимо уменьшить. Согласно последнему подходу фермент, активность которого необходимо уменьшить, предпочтительно поддерживают в инактивированном состоянии.
Вредная мутация в соответствии с настоящей заявкой представляет собой любую мутацию в гене, содержащем промотор и кодирующего участок, которая приводит к пониженной или исключенной белковой активности у белка, кодируемого кодирующим участком гена. Такие вредные мутации включают, например, сдвиги рамки считывания, внесение стоп-кодонов кодирующий участок, мутацию промоторных элементов, таких как ТАТА-бокс, что предотвращает транскрипцию, и т.п.
Микроорганизмы с уменьшенной экспрессией и/или активностью фермента, кодируемого геном brnQ, или РНК, кодируемой геном gcvB, или с повышенной или увеличенной экспрессией и/или активностью белков, кодируемых геном argP или геном gcvA, или с измененной активностью и/или экспрессией белка, кодируемого геном lpxD, могут встречаться в природе, т.е. появляться в результате спонтанных мутаций. Посредством различных методик, таких как химическая обработка или облучение, микроорганизм можно модифицировать так, чтобы у него отсутствовала или была значимо уменьшенная, повышенная или измененная активность фермента или РНК, которая кодируется одним или более указанных генов. Для этого микроорганизмы будут обрабатывать при помощи, например, мутагенного химического средства, рентгеновского или УФ излучения. На следующей стадии будут производить отбор микроорганизмов, которые имеют уменьшенную, повышенную или измененную экспрессию и/или активность фермента или РНК, которые кодируются одним или более из указанных генов. Рекомбинантные микроорганизмы также можно получить посредством методик гомологичной рекомбинации, которые направлены на мутацию, нарушение структуры или исключение одного или более из указанных генов в геноме микроорганизма или на замену одного или более из указанных генов на соответствующий ген, который кодирует фермент или РНК, которые, по сравнению с ферментом или РНК, кодируемым геном дикого типа, имеет уменьшенную, повышенную или измененную экспрессию и/или активность.
В соответствии с одним вариантом осуществления рекомбинантного микроорганизма согласно настоящему изобретению уменьшение экспрессии и/или активности фермента или РНК, кодируемых при помощи гена brnQ или гена gcvB достигают посредством модификации гена brnQ, причем эту/эти модификацию(модификации) гена предпочтительно осуществляют посредством делеции одного или более из указанных генов или по меньшей мере их части, делеции регуляторного элемента одного или более из указанных генов или их частей, таких как промоторная последовательность, или посредством внесения по меньшей мере одной вредной мутации в один или более из указанных генов.
В соответствии с одним вариантом осуществления рекомбинантного микроорганизма согласно настоящему изобретению увеличение экспрессии и/или активности фермента, кодируемого посредством гена argP и/или гена gcvA, может быть достигнуто посредством модификации гена argP и/или гена gcvA, причем эту/эти модификацию(модификации) гена предпочтительно осуществляют посредством умножения числа копий гена argP и/или гена gcvA в геноме микроорганизма, посредством внесения гена на самореплицирующемся векторе экспрессии в микроорганизм, посредством замены промотора гена argP и/или гена gcvA на более сильный промотор или посредством преобразования эндогенного промотора гена в более сильный промотор, например, посредством внесения точечной мутации в промоторную последовательность.
Кроме того, активность гена argP и/или гена gcvA и/или гена lpxD можно повысить или изменить посредством мутации гена для осуществления аминокислотных замен в белке, которые повышают или изменяют активность гена. Такие способы известны специалисту в настоящей области.
Мутация в указанном выше гене может быть внесена, например, посредством сайт-направленного или случайного мутагенеза с последующим внесением модифицированного гена в геном микроорганизма при помощи рекомбинации. Варианты генов можно создать с помощью мутации генетических последовательностей посредством ПЦР. Для осуществления направленного мутагенеза можно применять набор «Quickchange Site-directed Mutagenesis Kit» (Stratagene). Неспецифический мутагенез по всей длине кодирующей последовательности, или же только ее части, можно осуществлять при помощи набора «GeneMorph II Random Mutagenesis Kit» (Stratagene). Частоту возникновения мутаций устанавливают как необходимое количество мутаций на количество используемой матричной ДНК. Множественные мутации получают при помощи целенаправленного сочетания отдельных мутаций или при помощи последовательного осуществления нескольких циклов мутагенеза.
Далее описана подходящая методика для рекомбинации, в частности, для внесения мутации или для исключения последовательностей.
Эту методику также иногда в настоящем документе называют «рекомбинацией Campbell (Leenhouts et al., Appl Env Microbiol. (1989), Vol. 55, pages 394-400). В контексте настоящей заявки «Campbell in» относится к трансформанту исходной клетки-хозяина, у которой вся кольцевая двухцепочечная молекула ДНК (например, плазмида) была интегрирована в хромосому в результате однократного события гомологичной рекомбинации (события «cross in»), и это эффективно приводит в результате к вставке линеаризированного варианта указанной кольцевой молекулы ДНК в первую последовательность ДНК хромосомы, которая гомологична первой последовательности ДНК указанной кольцевой молекулы ДНК. «Введенная Campbell in» относится к линеаризированной последовательности ДНК, которая была интегрирована в хромосому трансформанта «Campbell in». «Campbell in» содержит дупликацию первой гомологичной последовательности ДНК, каждая копия которой включает и окружает копию кроссоверного сайта гомологичной рекомбинации.
В контексте настоящей заявки «Campbell out» относится к клетке, происходящей от трансформанта «Campbell in», у которой второе событие гомологичной рекомбинации (событие «cross out») имело место между второй последовательностью ДНК, которая содержится на линеаризированной вставленной ДНК в «введенной Campbell in» ДНК, и второй последовательностью ДНК хромосомного происхождения, которая гомологична второй последовательности ДНК указанной линеаризированной вставки, причем второе событие рекомбинации приводит в результате к делеции (удалению) части интегрированной последовательности ДНК, но, что также важно, также приводит в результате к тому, что часть (это может быть всего лишь одно основание) интегрированной введенной Campbell in ДНК остается в хромосоме, так чтобы, по сравнению с исходной клеткой-хозяином, клетка «Campbell out» содержала один или более преднамеренных изменений в хромосоме (например, замену одного основания, замены множества оснований, вставку гетерологичного гена или последовательности ДНК, вставку дополнительной копии или копий гомологического гена или модифицированного гомологичного гена или вставку последовательности ДНК, содержащей более одного из таких упомянутых примеров, которые приведены выше). Клетку «Campbell out» предпочтительно получают при помощи контротбора по гену, который содержится в части (части, которую необходимо удалить) «введенной Campbell in» последовательности ДНК, например, гена sacB Bacillus subtilis, который является летальным при экспрессии в клетке, которую выращивают в присутствии приблизительно 5%-10% сахарозы. Либо с контротбором, либо без него, необходимую клетку «Campbell out» можно получить или выявить посредством скрининга на необходимую клетку, при помощи любого подвергаемого скринингу фенотипа, такого как без ограничения структура колонии, цвет колонии, наличие или отсутствие устойчивости к антибиотикам, наличие или отсутствие заданной последовательности ДНК посредством полимеразной цепной реакции, наличие или отсутствие ауксотрофии, наличие или отсутствие фермента, гибридизация нуклеиновых кислот колонии, скрининг антител и т.д. Термины «Campbell in» и «Campbell out» также можно применять в качестве глаголов в различных временах для указания описанного выше способа или процесса.
Понятно, что события гомологичной рекомбинации, которые приводят к «Campbell in» или «Campbell out», могут происходить на протяжении некоторого диапазона оснований ДНК в гомологичной последовательности ДНК, и поскольку гомологичные последовательности будут идентичны друг другу на протяжении по меньшей мере части такого диапазона, обычно невозможно установить точно, где произошло кроссоверное событие. Иными словами, невозможно установить в точности, какая последовательность исходно была от вставленной ДНК, а какая исходно была от хромосомной ДНК. Кроме того, первая гомологичная последовательность ДНК и вторая гомологичная последовательность ДНК обычно разделены участком частичной негомологии, и именно этот участок негомологии остается в хромосоме клетки «Campbell out».
Предпочтительно первая и вторая последовательность ДНК имеет длину по меньшей мере приблизительно 200 пар оснований и может достигать до нескольких тысяч пар оснований. Тем не менее такую процедуру можно адаптировать для работы с более короткими или с более длинными последовательностями. Например, длина первой и второй гомологичных последовательностей может варьировать от приблизительно 500 до 2000 оснований, а получение «Campbell out» из «Campbell in» облегчают путем приведения первой и второй гомологичных последовательностей примерно к одной длине, предпочтительно с отличием на менее 200 пар оснований, и причем наиболее предпочтительно, чтобы более короткая из двух имела по меньшей мере 70% длины от более длинной по парам оснований.
В соответствии с одним вариантом осуществления уменьшение экспрессии и/или активности brnQ и/или гена gcvB достигают при помощи инактивации гена brnQ и/или гена gcvB, который кодирует соответственно транспортер для аминокислот с разветвленной боковой нитью или РНК, которая не кодирует белок.
В соответствии с одним вариантом осуществления инактивацию генов предпочтительно осуществляют при помощи делеции гена или по меньшей мере его частей, при помощи делеции регуляторного элемента гена или по меньшей мере его части или при помощи внесения по меньшей мере одной вредной мутации в ген.
В соответствии с одним вариантом осуществления индукцию экспрессии и/или активности argP осуществляют при помощи активации гена argP, который кодирует белок, обладающий связывающей ДНК / активирующей транскрипцию активностью.
В соответствии с одним вариантом осуществления индукцию экспрессии и/или активности gcvA осуществляют посредством активации гена gcvA, который кодирует ДНК-связывающий белок.
Термины «аланин, пируват, сукцинат, аспартат, малат, лактат, валин и/или лейцин», которые применяют в контексте настоящего изобретения, следует понимать в их наиболее широком смысле, а также они охватывают их соли, как например: соли щелочных металлов, такие как соли Na+ и K+-, или соли щелочноземельных металлов, такие как соли Mg2+ и Са2+, или соли аммония или ангидриды аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина.
Предпочтительно аланин, пируват, сукцинат, аспартат, малат, лактат, валин и/или лейцин, предпочтительно сукцинат или аланин, более предпочтительно аланин получают в микроаэробных условиях. Также можно использовать аэробные или анаэробные условия.
Микроаэробный означает, что концентрация кислорода меньше, чем в воздухе. В соответствии с одним вариантом осуществления микроаэробный означает давление кислорода от 5 до 27 мм рт.ст., предпочтительно от 10 до 20 мм рт.ст. (Megan Falsetta et al. (2011), The composition and metabolic phenotype of Neisseria gonorrhoeae biofilms, Frontiers in Microbiology, Vol 2, page 1 to 11). Предпочтительно микроаэробные условия устанавливаются при 0,1-1 vvm потока воздуха.
Анаэробные условия можно установить посредством традиционных методик, как, например, путем дегазации составных компонентов реакционной среды и поддержания анаэробных условий при помощи внесения двуокиси углерода или азота или их смесей и необязательно водорода при скорости потока, например, 0,1-1 или 0,2-0,5 vvm. Аэробные условия можно установить посредством традиционных методик, как, например, путем внесения воздуха или кислорода при скорости, составляющей, например, 0,1-1 или 0,2-0,5 vvm. При необходимости в таком способе можно прилагать небольшое избыточное давление в 0,1-1,5 бар.
В соответствии с одним вариантом осуществления способа согласно настоящему изобретению усваиваемый источник углерода может представлять собой глюкозу, глицерин, глюкозу, мальтозу, мальтодекстрин, фруктозу, галактозу, маннозу, ксилозу, сахарозу, арабинозу, лактозу, раффинозу и их комбинации.
В соответствии с предпочтительным вариантом осуществления усваиваемый источник углерода представляет собой глюкозу, сахарозу, ксилозу, арабинозу, глицерин или их комбинации. Предпочтительными источниками углерода являются глюкоза, сахароза, глюкоза и сахароза, глюкоза и ксилоза и/или глюкоза, арабиноза и ксилоза. В соответствии с одним вариантом осуществления способа согласно настоящему изобретению усваиваемым источником углерода может быть сахароза, глицерин и/или глюкоза.
Исходную концентрацию усваиваемого источника углерода, предпочтительно исходную концентрацию, предпочтительно, корректируют к значению в диапазоне от 5 до 250 г/л, предпочтительно от 50 до 200 г/л и более предпочтительно от 125 до 150 г/л, наиболее предпочтительно приблизительно 140 г/л, и ее можно поддерживать в указанном диапазоне в ходе культивирования. рН реакционной среды можно контролировать путем добавления подходящих оснований, как например: газообразный аммиак, NH4OH NH4HCO3, (NH4)2CO3, NaOH, Na2CO3, NaHCO3, KOH, K2CO3, KHCO3, Mg(OH)2, MgCO3, Mg(HCO3)2, Ca(OH)2, СаСО3, Са(HCO3)2, CaO, CH6N2O2, C2H7N и/или их смеси.
В соответствии с другим вариантом осуществления настоящего изобретения, оно относится к способу ферментативного получения аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, наиболее предпочтительно L-аланина, предусматривающему стадии
I) выращивания микроорганизма в соответствии с настоящим изобретением, как описано выше, в ферментере;
II) выделения аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, наиболее предпочтительно L-аланина из ферментативного бульона, полученного на I).
Стадию ферментирования I) в соответствии с настоящим изобретением можно, например, осуществлять в ферментерах с мешалкой, барботажных колоннах и петлевых реакторах. Полный обзор возможных типов способа, в том числе типы мешалок и геометрические схемы, можно найти в работе Chmiel: "Bioprozesstechnik: Einfuhrung in die Bioverfahrenstechnik", Volume 1. В способе согласно настоящему изобретению типичными возможными вариантами являются описанные далее варианты, которые известны специалистам в настоящей области техники, или раскрыты, например, в работе Chmiel, Hammes and Bailey: "Biochemical Engineering", такие как периодическая, периодическая с добавлением субстрата или же непрерывная ферментация с рециркуляцией биомассы или без нее. В зависимости от производственного штамма можно производить барботирование воздухом, кислородом, диоксидом углерода, водородом, азотом или соответствующими смесями газов для достижения хорошего выхода (YP/S).
Особенно предпочтительными условиями для получения аланина, пирувата, сукцината, аспартата, малата, лактата, валина и/или лейцина, предпочтительно сукцината или аланина, более предпочтительно аланина, наиболее предпочтительно L-аланина на стадии I) способа являются:
усваиваемый источник углерода: глюкоза;
температура: 30-45°С;
рН: 6,0-7,0;
микроаэробные условия.
На стадии II) способа продукт выделяют из ферментативного бульона, полученного на стадии I) способа.
Обычно способ выделения включает стадию отделения рекомбинантных микроорганизмов от ферментативного бульона, которые также называют «биомассой». Способы удаления биомассы известны специалистам в настоящей области и включают фильтрацию, осаждение, флотацию или их комбинации. Соответственно, биомассу можно удалять, например, при помощи центрифуг, препаративных центрифуг, отстойников, фильтров или во флотационном аппарате. Для максимального выделения имеющего ценность продукта часто рекомендуют промывание биомассы, например, в форме диафильтрации. Выбор способа зависит от содержимого биомассы в ферментативном бульоне и свойств биомассы, а также взаимодействия биомассы с органическим соединением (т.е. имеющим ценность продуктом). В соответствии с одним вариантом осуществления ферментативный бульон можно простерилизовать или пастеризовать. В соответствии со следующим вариантом осуществления ферментативный бульон подвергают концентрированию. В зависимости от необходимости, такое концентрирование можно осуществлять порционно или непрерывно. Диапазон давлений и температур следует выбирать так, чтобы, во-первых, не происходило повреждение продукта, а, во-вторых, было необходимо минимальное применение приборов и энергии. Умелый выбор уровней давления и температуры для многоэтапного выпаривания, в частности, обеспечивает сохранение энергии.
Способ выделения дополнительно может предусматривать дополнительные стадии очистки, на которых дополнительно очищают продукт ферментации. Однако, если продукт ферментации преобразовывают во вторичный органический продукт посредством химических реакций, то дополнительная очистка продукта ферментации, в зависимости от типа реакции и реакционных условий, может не быть обязательно необходимой. Для очистки продукта ферментации, полученного на стадии II) способа, можно применять известные специалистам в настоящей области способы, как например: кристаллизацию, фильтрацию, электродиализ и хроматографию. Полученный в результате раствор можно дополнительно очистить посредством ионнообменной хроматографии для удаления нежелательных остаточных ионов.
В соответствии с одним вариантом осуществления уменьшенную, подавленную или исключенную экспрессию и/или активность гена brnQ достигают посредством внесения делеции в ген дикого типа. Предпочтительно это осуществляют посредством внесения специфической мутации между позициями 667 и 764 гена дикого типа с SEQ ID NO: 1 или его функционального варианта.
С учетом вышесказанного, следующий вариант осуществления настоящего изобретения относится к рекомбинантной молекуле нуклеиновой кислоты, имеющей последовательность, выбранную из группы, состоящей из
(6) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 3, или
(7) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 3, или
(8) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 3 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(9) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 4, или
(10) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 4,
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, но не ген и/или белок дикого типа, характеризуется повышенным выходом аланина в ферментации.
В соответствии с одним вариантом осуществления повышенную или увеличенную экспрессию и/или активность гена argP достигают при помощи внесения мутации в ген дикого типа. Предпочтительно этого достигают посредством внесения специфической мутации в положении 286-288 гена дикого типа с SEQ ID NO: 45 или его функционального варианта.
С учетом вышесказанного следующий вариант осуществления настоящего изобретения относится к рекомбинантной молекуле нуклеиновой кислоты, имеющей последовательность, выбранную из группы, состоящей из
(1) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 45, или
(2) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 45, или
(3) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 45 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(4) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 46, или
(5) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 46, и
причем кодон генов, обозначенных в (1)-(5), соответствующий положению 286-288 в SEQ ID NO: 45, не кодирует аминокислоту аланин и не является стоп-кодоном, или аминокислота белков, кодируемых генами, обозначенными в (1)-(5), соответствующая положению 96 в SEQ ID NO: 46, не является аланином, и
причем белок, кодируемый геном, который определен выше в (1)-(5), имеет повышенную или увеличенную активность по сравнению с белком с SEQ ID NO: 46, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
Предпочтительно рекомбинантная молекула нуклеиновой кислоты имеет последовательность, выбранную из группы, состоящей из
(6) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 47, или
(7) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 47, или
(8) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 47 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(9) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 48, или
(10) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 48,
причем кодон генов, обозначенных в (7)-(10), соответствующий положению 286-288 в SEQ ID NO: 47, кодирует такую аминокислоту, как глутаминовая кислота, или родственную аминокислоту, или аминокислота белков, кодируемых генами, обозначенными в (7)-(10), соответствующая положению 96 в SEQ ID NO: 48, представляет собой глутаминовую кислоту или родственную аминокислоту, и
причем белок, кодируемый геном, который определен выше в (7)-(10), имеет повышенную или увеличенную активность по сравнению с белком с SEQ ID NO: 48, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
В соответствии с одним вариантом осуществления повышенную или увеличенную экспрессию и/или активность гена gcvA достигают при помощи внесения мутации в промотор гена дикого типа. Предпочтительно этого достигают посредством внесения специфической мутации промотора гена дикого типа с SEQ ID NO: 53 или его функционального варианта, причем мутация соответствует мутации, внесенной в SEQ ID NO: 55, приводящей к созданию SEQ ID NO: 56 или SEQ ID NO: 57.
С учетом вышесказанного, следующий вариант осуществления настоящего изобретения относится к рекомбинантной молекуле нуклеиновой кислоты, содержащей последовательность, выбранную из группы, состоящей из
(11) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 53, или
(12) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 53, или
(13) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 53 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(14) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 54, или
(15) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 54,
функционально связанной с мутантным промотором или промотором, активным в микроорганизме, который является гетерологичным для указанной молекулы нуклеиновой кислоты, и
причем микроорганизм, содержащий сверхэкспрессируемый ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
В соответствии с одним вариантом осуществления повышенный или увеличенный выход и/или производительность по аланину или родственным соединениям достигают посредством внесения мутации в ген lpxD дикого типа.
С учетом вышесказанного, один вариант осуществления настоящего изобретения относится к рекомбинантной молекуле нуклеиновой кислоты, имеющей последовательность, выбранную из группы, состоящей из
(16) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 49, или
(17) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 49, или
(18) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 49 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(19) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 50, или
(20) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 50, и
причем кодон генов, обозначенных в (16)-(20), соответствующий положению 43-45 в SEQ ID NO: 49, не кодирует аминокислоту аланин и не является стоп-кодоном, или аминокислота белков, кодируемых генами, обозначенными в (6)-(10), соответствующая положению 15 в SEQ ID NO: 50, не является аланином, и
причем белок, кодируемый геном, который определен выше в (16)-(20), характеризуется измененной активностью и/или экспрессией по сравнению с белком с SEQ ID NO: 50, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
Предпочтительно рекомбинантная молекула нуклеиновой кислоты имеет последовательность, выбранную из группы, состоящей из
(21) молекулы нуклеиновой кислоты, содержащей последовательность SEQ ID NO: 51, или
(22) молекулы нуклеиновой кислоты, которая по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, например, по меньшей мере на 90%, более предпочтительно по меньшей мере на 95%, например, по меньшей мере на 96%, еще более предпочтительно по меньшей мере на 97%, например, по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% идентична молекуле нуклеиновой кислоты с SEQ ID NO: 51, или
(23) молекулы нуклеиновой кислоты, гибридизирующейся с молекулой нуклеиновой кислоты с SEQ ID NO: 51 в условиях средней жесткости, более предпочтительно в условиях высокой жесткости, наиболее предпочтительно в условиях очень высокой жесткости, или
(24) молекулы нуклеиновой кислоты, кодирующей полипептид с SEQ ID NO: 52, или
(25) молекулы нуклеиновой кислоты, кодирующей полипептид, который по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологичен полипептиду с SEQ ID NO: 52,
причем кодон генов, обозначенных в (21)-(25), соответствующий положению 43-45 в SEQ ID NO: 51, кодирует аминокислоту треонин или родственную аминокислоту, или аминокислота белков, кодируемых генами, обозначенными в (21)-(25), соответствующая положению 15 в SEQ ID NO: 52, представляет собой треонин или родственную аминокислоту, и
причем белок, кодируемый геном, который определен выше в (21)-(25), характеризуется измененной активностью и/или экспрессией по сравнению с белком с SEQ ID NO: 50, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
Термин «родственная аминокислота» или «консервативная аминокислотная замена» означает, что аминокислоту заменяют на аминокислоту, имеющую сходную боковую нить. Перечень родственных аминокислот приведен в таблице 2 ниже. Таблицы консервативных замен хорошо известны из уровня техники (см., например, Creighton (1984) Proteins. W.H. Freeman and Company (Eds) и таблицу 2 ниже).
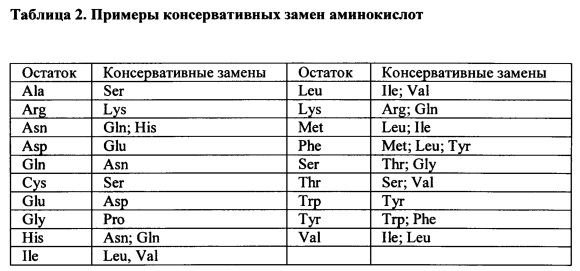
В соответствии с предпочтительным вариантом осуществления рекомбинантная молекула нуклеиновой кислоты имеет SEQ ID NO: 3, 47, 51 и кодирует белок с SEQ ID NO: 4, 48, 52 соответственно.
Следующий вариант осуществления по настоящему изобретению относится к рекомбинантной аминокислотной молекуле, имеющей последовательность, выбранную из группы, состоящей из
(26) молекулы аминокислоты, содержащей последовательность SEQ ID NO: 4, или
(27) молекулы аминокислоты, которая на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологична полипептиду с SEQ ID NO: 4,
причем микроорганизм, содержащий мутантный белок, который определен выше, но не белок дикого типа, характеризуется повышенным выходом аланина в ферментации.
Предпочтительно рекомбинантная молекула аминокислоты по настоящему изобретению имеет SEQ ID NO: 4.
Следующий вариант осуществления по настоящему изобретению относится к рекомбинантной аминокислотной молекуле, имеющей последовательность, выбранную из группы, состоящей из
(28) молекулы аминокислоты, содержащей последовательность SEQ ID NO: 48, или
(29) молекулы аминокислоты, которая на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологична полипептиду с SEQ ID NO: 48,
причем аминокислота белка, обозначенная в (29), соответствующая положению 96 в SEQ ID NO: 48, представляет собой глутаминовую кислоту или родственную аминокислоту, и
причем белок, который определен выше в (29), имеет повышенную или увеличенную активность по сравнению с белком с SEQ ID NO: 46, и
причем микроорганизм, содержащий мутантный белок, который определен выше, но не белок дикого типа, характеризуется повышенным выходом аланина в ферментации.
Предпочтительно рекомбинантная молекула аминокислоты по настоящему изобретению имеет SEQ ID NO: 48.
Следующий вариант осуществления по настоящему изобретению относится к рекомбинантной аминокислотной молекуле, имеющей последовательность, выбранную из группы, состоящей из
(30) молекулы аминокислоты, содержащей последовательность SEQ ID NO: 52, или
(31) молекулы аминокислоты, которая на 60%, предпочтительно по меньшей мере на 70%, например, по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, например, по меньшей мере на 85%, еще более предпочтительно по меньшей мере на 90%, например, по меньшей мере на 95%, наиболее предпочтительно по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологична полипептиду с SEQ ID NO: 52,
причем аминокислота белка, обозначенная в (31), соответствующая положению 15 в SEQ ID NO: 52, представляет собой треонин или родственную аминокислоту, и
причем белок, который определен выше в (31), характеризуется измененной активностью и/или экспрессией по сравнению с белком с SEQ ID NO: 50, и
причем микроорганизм, содержащий мутантный ген и/или белок, которые определены выше, характеризуется повышенным выходом аланина при ферментации.
Предпочтительно рекомбинантная молекула аминокислоты по настоящему изобретению имеет SEQ ID NO: 52.
Определения
Следует понимать, что настоящее изобретение не ограничено конкретной методологией или конкретными протоколами. Также следует понимать, что применяемая в настоящем документе терминология предназначена лишь для описания конкретных вариантов осуществления и не предназначена для ограничения объема настоящего изобретения, которое будет ограничиваться лишь прилагаемой формулой изобретения. Необходимо отметить, что в контексте настоящей заявки и в прилагаемой формуле изобретения формы единственного числа включают и множественное число, если из контекста явно не следует иное. Таким образом, например, отсылка к «вектору» представляет собой отсылку к одному или более векторов и включает их эквиваленты, известные специалистам в настоящей области, и тому подобное. Термин «приблизительно» в настоящем документе применяют для обозначения примерно, приближенно, около или в диапазоне. При использовании термина «приблизительно» в сочетании с числовым диапазоном он модифицирует такой диапазон, расширяя его границы выше и ниже изложенных числовых значений. В целом, термин «приблизительно» в настоящем документе применяют для модификации числового значения выше и нижеуказанного значения на отклонение, составляющее 20 процентов, предпочтительно 10 процентов вверх или вниз (выше или ниже). В контексте настоящей заявки слово «или» означает любой один элемент из конкретного перечня, а также включает любую комбинацию элементов такого перечня. Слова «содержать», «содержащий», «включать», «включающий» и «включает» при использовании в настоящем описании и в приведенной далее формуле изобретения предназначены для описания наличия одного или более указанных признаков, целых чисел, компонентов или стадий, но они не исключают наличие или добавление одного или более других признаков, целых чисел, компонентов, стадий или групп из них. Для наглядности далее приведено определение и применение некоторых терминов, применяемых в настоящем описании.
Кодирующий участок: в контексте настоящей заявки термин «кодирующий участок» при применении в отношении структурного гена относится к нуклеотидным последовательностям, которые кодируют аминокислоты, встречающиеся в образующемся полипептиде в результате трансляции молекулы мРНК. Кодирующий участок связан у эукариот на 5'-стороне нуклеотидным триплетом «ATG», который кодирует инициаторный метионин, прокариоты также используют триплеты «GTG» и «TTG» в качестве старт-кодонов. На 3'-стороне он связан одним из трех триплетов, которые определяют стоп-кодоны (т.е. ТАА, TAG, TGA). Кроме того, ген может включать последовательности, расположенные как на 5'-, так и на 3'-конце последовательностей, которые представлены на РНК-транскрипте. Такие последовательности называются «фланкирующими» последовательностями или участками (такие фланкирующие последовательности расположены 5' или 3' к нетранслируемым последовательностям, присутствующим на мРНК-транскрипте). 5'-фланкирующий участок может содержать регуляторные последовательности, такие как промоторы и энхансеры, которые контролируют транскрипцию гена или оказывают на нее влияние. 3'-фланкирующий участок может содержать последовательности, которые управляют окончанием транскрипции, посттрансляционным расщеплением и полиаденилированием.
Комплементарный: «комплементарный» или «комплементарность» относится к двум нуклеотидным последовательностям, которые представляют собой антипараллельные нуклеотидные последовательности, способные спариваться друг с другом (по правилам спаривания оснований) при образовании водородных связей между остатками комплементарных оснований в антипараллельных нуклеотидных последовательностях. Например, последовательность 5'-AGT-3' является комплементарной последовательности 5-АСТ-3'. Комплементарность может быть «частичной» или «полной». «Частичная» комплементарность имеет место в том случае, когда одно или более оснований нуклеиновой кислоты не соответствуют согласно правилам спаривания оснований. «Полная» или «абсолютная» комплементарность между молекулами нуклеиновых кислот имеет место в том случае, когда все без исключения основания нуклеиновых кислот соответствуют друг другу согласно правилам спаривания оснований. Степень комплементарности между цепями молекулы нуклеиновых кислот оказывает значительное влияние на эффективность и силу гибридизации между цепями молекулы нуклеиновой кислоты. «Комплемент» с последовательностью нуклеиновой кислоты в контексте настоящей заявки относится к нуклеотидной последовательности, молекулы нуклеиновой кислоты с которой характеризуются полной комплементарностью к молекулам нуклеиновой кислоты с указанной последовательностью нуклеиновой кислоты.
Эндогенная: «эндогенная» нуклеотидная последовательность относится к нуклеотидной последовательности, которая присутствует в геноме микроорганизма дикого типа.
Повышенная экспрессия: «повышать» или «увеличивать» экспрессию молекулы нуклеиновой кислоты в микроорганизме в настоящем документе используют эквивалентно, и они означают, что уровень экспрессии молекулы нуклеиновой кислоты в микроорганизме выше по сравнению с эталонным микроорганизмом, например, дикого типа. Термины «повышенная» или «увеличенная» в контексте настоящей заявки означают более высокую, предпочтительно значительно более высокую экспрессию подлежащей экспрессии молекулы нуклеиновой кислоты. В контексте настоящей заявки «повышение» или «увеличение» уровня такого средства, как белок, мРНК или РНК, означает что уровень является увеличенным по сравнению с практически идентичным микроорганизмом, выращенным в практически идентичных условиях. В контексте настоящей заявки «повышение» или «увеличение» уровня такого средства, как например: преРНК, мРНК, рРНК, тРНК, экспрессировавшегося целевым геном, и/или кодируемого им белкового продукта означает, что уровень увеличен на 50% или более, например на 100% или более, предпочтительно на 200% или более, более предпочтительно в 5 раз или более, еще более предпочтительно в 10 раз или более, наиболее предпочтительно в 20 раз или более, например в 50 раз, относительно соответствующего эталонного микроорганизма. Повышение или увеличение можно определить с помощью способов, с которыми знаком специалист в настоящей области. Так, повышение или увеличение количества нуклеиновой кислоты или белка можно определить, например, с помощью иммунологической детекции белка. Кроме того, для измерения конкретного белка или РНК у микроорганизма можно использовать такие методики, как белковый анализ, флуоресценция, нозерн-гибридизация, анализ защиты от нуклеаз, обратная транскрипция (количественная RT-PCR), ELISA (твердофазный иммуноферментный анализ), вестерн-блоттинг, радиоиммунологический анализ (RIA) или другие иммуноанализы и сортировка флуоресцентно-активированных клеток (FACS). В зависимости от типа индуцированного белкового продукта, также можно определить его активность или влияние на фенотип микроорганизма. Специалисту в настоящей области известны способы определения количества белка. Примерами, которые могут быть упомянуты, являются микробиуретовый способ (Goa J (1953) Scand J Clin Lab Invest 5:218-222), способ Фолина-Чокальтеу (Lowry OH et al. (1951) J Biol Chem 193:265-275) или измерение абсорбции CBB G-250 (Bradford MM (1976) Analyt Biochem 72:248-254).
Экспрессия: «экспрессия» относится к биосинтезу продукта гена, предпочтительно к транскрипции и/или трансляции нуклеотидной последовательности, например эндогенного гена или гетерологического гена, в клетке. Например, в случае структурного гена, экспрессия включает транскрипцию структурного гена в мРНК и необязательно последующую трансляцию мРНК в один или более полипептидов. В других случаях экспрессия может относиться только к транскрипции ДНК, несущей информацию о молекуле РНК.
Чужеродная: термин "чужеродная" относится к любой молекуле нуклеиновой кислоты (например, последовательности гена), которая внесена в клетку с помощью манипуляций в ходе эксперимента и может включать последовательности, встречающиеся в такой клетке, при условии, что внесенная последовательность содержит некую модификацию (например, точечную мутацию, наличие селектируемого маркерного гена и т.д.) и поэтому отличается относительно встречающейся в природе последовательности.
Функциональная связь: термин «функциональная связь» или «функционально связанный» является эквивалентным термину «связь с возможностью выполнения своих функций» или «связанный с возможностью выполнения своих функций», и его следует понимать как обозначающий, например, последовательное расположение регуляторного элемента (например, промотора) с подлежащей экспрессии последовательностью нуклеиновой кислоты и, при необходимости, дополнительных регуляторных элементов (таких как, например, терминатор) таким образом, чтобы каждый из указанных регуляторных элементов мог выполнять свою целевую функцию, делая возможной, модифицируя, способствуя или иным образом влияя на экспрессию указанной последовательности нуклеиновой кислоты. Выражение «связь с возможностью выполнения своих функций» или «связанный с возможностью выполнения своих функций» можно использовать в качестве синонима. Экспрессия в зависимости от расположения последовательностей нуклеиновой кислоты может привести к образованию смысловой или антисмысловой РНК. Поэтому непосредственная связь в химическом смысле не является обязательно необходимой. Последовательности генетического контроля, такие как, например, энхансерные последовательности, также могут проявлять свою функцию на целевую последовательность из позиций, которые достаточно удалены, или практически из других молекул ДНК. Предпочтительными расположениями являются такие, в которых подлежащая рекомбинантной экспрессии последовательность нуклеиновой кислоты размещена после последовательности, выступающей в качестве промотора, так чтобы эти две последовательности были ковалентно связаны друг с другом. В соответствии с предпочтительным вариантом осуществления подлежащая транскрипции последовательность нуклеиновой кислоты расположена после промотора таким образом, чтобы точка начала транскрипции была идентичной требуемому началу химерной РНК по настоящему изобретению. Функциональная связь и экспрессионная конструкция могут быть созданы посредством описанных общепринятых рекомбинационных методик и методик клонирования (например, в работе Maniatis Т, Fritsch EF and Sambrook J (1989) Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor (NY); Silhavy et al. (1984) Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor (NY); Ausubel et al. (1987) Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience; Gelvin et al. (Eds) (1990) Plant Molecular Biology Manual; Kluwer Academic Publisher, Dordrecht, The Netherlands). Тем не менее, между двумя последовательностями также могут быть размещены дополнительные последовательности, которые, например, выполняют функцию линкера с сайтами специфического расщепления для рестрикционных ферментов или функцию сигнального пептида. Вставка последовательностей также может привести к экспрессии гибридных белков. Предпочтительно экспрессионная конструкция, состоящая из связи регуляторного участка, например промотора, и подлежащей экспрессии последовательности нуклеиновой кислоты, может существовать в интегрированной в вектор форме или может быть вставлена в геном, например, путем трансформации.
Ген: термин «ген» относится к участку, связанному с возможностью выполнения своих функций с соответствующими регуляторными последовательностями, способными определенным способом регулировать экспрессию продукта гена (например, полипептида или функциональной РНК). Ген включает нетранслируемые регуляторные участки ДНК (например, промоторы, энхансеры, репрессоры и т.д.), предшествующие (находящиеся выше) и следующие после (находящиеся ниже) кодирующего участка (открытой рамки считывания, ORF). Термин «структурный ген» в контексте настоящей заявки предназначен для обозначения последовательности ДНК, которая транскрибируется в мРНК, которая затем транслируется в последовательность аминокислот, характерную для конкретного полипептида.
Геном и геномная ДНК: термины «геном» или «геномная ДНК» относятся к наследственной генетической информации организма-хозяина. Указанная геномная ДНК содержит ДНК нуклеотида, а также ДНК самореплицирующейся плазмиды.
Гетерологичная: термин «гетерологичная» в отношении молекулы нуклеиновой кислоты или ДНК относится к молекуле нуклеиновой кислоты, которая связана с возможностью выполнения своих функций со, или с ней проводят манипуляцию с тем, чтобы сделать ее связанной с возможностью выполнения своих функций со, второй молекулой нуклеиновой кислоты, с которой в природе она не является связанной с возможностью выполнения своих функций или с которой в природе она является связанной с возможностью выполнения своих функций в ином местоположении. Гетерологичная экспрессионная конструкция, содержащая молекулу нуклеиновой кислоты и один или более регуляторных молекул нуклеиновой кислоты (таких как промотор или сигнал окончания транскрипции), связанных с ней, например, представляет собой конструкции, полученные в результате манипуляций в ходе эксперимента, при которых либо а) указанная молекула нуклеиновой кислоты, либо b) указанная регуляторная молекула нуклеиновой кислоты, либо с) обе (т.е. (а) и (b)) не расположены в своем природном (нативном) генетическом окружении или была модифицирована с помощью манипуляций в ходе эксперимента, причем примером модификации является замена, добавление, делеция, инверсия или вставка одного или более нуклеотидных остатков. Природное генетическое окружение относится к природному геномному локусу в организме происхождения или к наличию в геномной библиотеке. В случае геномной библиотеки природное генетическое окружение последовательности молекулы нуклеиновой кислоты предпочтительно сохраняется, по меньшей мере частично. Окружение фланкирует последовательность нуклеиновой кислоты по меньшей мере с одной стороны и имеет последовательность длиной по меньшей мере 50 п.о., предпочтительно по меньшей мере 500 п.о., еще предпочтительнее по меньшей мере 1000 п.о., наиболее предпочтительно по меньшей мере 5000 п.о. Встречающаяся в природе экспрессионная конструкция, например встречающаяся в природе комбинация промотора с соответствующим геном, становится трансгенной экспрессионной конструкцией при модификации ее неприродными, синтетическими "искусственными" способами, такими как, например, мутагенез. Такие способы уже были описаны (US 5565350, WO 00/15815). Например, кодирующую белок молекулу нуклеиновой кислоты, связанную с возможностью выполнения своих функций с промотором, который не является природным промотором для данной молекулы, считают гетерологичной по отношению к промотору. Предпочтительно, гетерологичная ДНК не является эндогенной для клетки, или не ассоциирована с ней в природе, в которую ее вносят, но была получена из другой клетки или была синтезирована. Гетерологичная ДНК также включает последовательность эндогенной ДНК, которая содержит такую же модификацию, не встречающуюся в природе, множество копий последовательности эндогенной ДНК и последовательность ДНК, которая не ассоциирована в природе с другой последовательностью ДНК, которая с ней связана физически. В целом, хотя и не обязательно, гетерологичная ДНК кодирует РНК или белки, которые в норме не вырабатываются клеткой, в которой она экспрессируется.
Гибридизация: термин «гибридизация» в контексте настоящей заявки включает «любой процесс, в результате которого нить молекулы нуклеиновой кислоты соединяется с комплементом путем спаривания оснований». (J. Coombs (1994) Dictionary of Biotechnology, Stockton Press, New York). На гибридизацию и силу гибридизации (т.е. силу ассоциации между молекулами нуклеиновой кислоты) оказывают влияние такие факторы, как степень комплементарности между молекулами нуклеиновой кислоты, жесткость задействованных условий, Tm сформированного гибрида и соотношение G : C в молекулах нуклеиновой кислоты. В контексте настоящей заявки термин «Tm» используют в отношении «температуры плавления». Температура плавления представляет собой температуру, при которой некоторое количество двухцепочечных молекул нуклеиновой кислоты становится наполовину диссоциированными на отдельные цепи. Уравнение для расчета Tm молекул нуклеиновой кислоты известно из уровня техники. Как указано в стандартных литературных источниках, упрощенную оценку значения Tm можно провести путем расчета с помощью уравнения: Tm=81,5+0,41(% G+C), если молекула нуклеиновой кислоты находится в водном растворе с 1 М NaCl [см., например, Anderson and Young, Quantitative Filter Hybridization, in Nucleic Acid Hybridization (1985)]. Другие литературные источники включают более усложненные вычисления, которые для расчета Tm принимают во внимание структурные характеристики, а также характеристики последовательностей. Жесткие условия известны специалистам в настоящей области и могут быть найдены в работе Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6.
Подходящими условиями гибридизации являются, например, гибридизация в условиях, эквивалентных гибридизации в 7% додецисульфата натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с промыванием в 2 X SSC, 0,1% SDS при 50°С (низкая жесткость) с молекулой нуклеиновой кислоты, содержащей по меньшей мере 50, предпочтительно по меньшей мере 100, более предпочтительно по меньшей мере 150, еще более предпочтительно по меньшей мере 200, наиболее предпочтительно по меньшей мере 250 последовательных нуклеотидов комплемента последовательности. Другими подходящими условиями гибридизации являются гибридизация в 7% додецилсульфата натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с промыванием в 1 X SSC, 0,1% SDS при 50°С (средняя жесткость) или 65°С (высокая жесткость) с молекулой нуклеиновой кислоты, содержащей по меньшей мере 50, предпочтительно по меньшей мере 100, более предпочтительно по меньшей мере 150, еще более предпочтительно по меньшей мере 200, наиболее предпочтительно по меньшей мере 250 последовательных нуклеотидов комплемента последовательности. Другими подходящими условиями гибридизации являются гибридизация в 7% додецилсульфата натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с промыванием в 0,1 X SSC, 0,1% SDS при 65°С (очень высокая жесткость) с молекулой нуклеиновой кислоты, содержащей по меньшей мере 50, предпочтительно по меньшей мере 100, более предпочтительно по меньшей мере 150, еще более предпочтительно по меньшей мере 200, наиболее предпочтительно по меньшей мере 250 последовательных нуклеотидов комплемента последовательности.
«Идентичность»: термин «идентичность» при использовании в отношении сравнения двух или более молекул нуклеиновых кислот или аминокислот означает, что последовательности указанных молекул имеют некоторую степень сходства последовательностей, причем последовательности являются отчасти идентичными.
Для определения процента идентичности (термин «гомология» в настоящем документе используют взаимозаменяемо в случае отсылки к последовательностям нуклеиновой кислоты) двух аминокислотных последовательностей или двух молекул нуклеиновой кислоты последовательности записывают одну под другой для проведения оптимального сравнения (например, в последовательность белка или нуклеиновой кислоты могут быть вставлены гэпы для создания оптимального выравнивания с другим белком или другой нуклеиновой кислотой).
Затем проводят сравнение аминокислотных остатков или молекул нуклеиновых кислот по соответствующим позициям аминокислот или позициям нуклеотидов. Если позиция в одной последовательности занята аналогичным аминокислотным остатком или такой же молекулой нуклеиновой кислоты, что и в соответствующем положении в другой последовательности, то молекулы являются идентичными по такому положению. Процент идентичности между двумя последовательностями представляет собой отображение количества идентичных позиций для последовательностей (т.е. % идентичности = количество идентичных позиций/общее количество позиций × 100). Термины «гомология» и «идентичность», таким образом, следует рассматривать как синонимы в отношении последовательностей нуклеиновых кислот. В отношении аминокислотных последовательностей термин «идентичность» относится к идентичным аминокислотам в конкретном положении в последовательности, термин «гомология» относится к гомологичным аминокислотам в конкретном положении в последовательности. Гомологичными аминокислотами являются аминокислоты с одинаковой боковой нитью. В настоящей области техники были описаны семейства аминокислотных остатков с одинаковыми боковыми цепями.
Молекулу нуклеиновой кислоты, кодирующую белок, гомологичный белку по настоящему изобретению, можно создать путем внесения одной или более нуклеотидных замен, добавлений или делеций в нуклеотидную последовательность, так чтобы в кодируемый белок была внесена одна или более аминокислотных замен, добавлений или делеций. Мутации можно вносить в одну из последовательностей по настоящему изобретению с помощью стандартных методик, таких как сайт-направленный мутагенез и PCR-опосредованный мутагенез. Предпочтительно консервативные аминокислотные замены производят по одному или более прогнозируемым, не относящимся к незаменимым аминокислотным остаткам. «Консервативная аминокислотная замена» представляет собой замену, при которой аминокислотный остаток заменяется аминокислотным остатком с подобной боковой нитью. В настоящей области техники были описаны семейства аминокислотных остатков с одинаковыми боковыми цепями. Такие семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Так, прогнозируемый, не относящийся к незаменимому аминокислотный остаток в белке по настоящему изобретению предпочтительно заменяют другим аминокислотным остатком из того же семейства боковых цепей.
В альтернативном варианте, в соответствии с другим вариантом осуществления, мутации могут быть внесены случайным образом по всей или части кодирующей последовательности, к примеру, с помощью насыщающего мутагенеза, а полученные мутанты могут быть подвергнуты скринингу в отношении соответствующей активности, которая описана в настоящем документе, для выявления мутантов, которые сохраняют свою активность. После мутагенеза одной из последовательностей по настоящему изобретению кодируемый белок можно экспрессировать рекомбинантно, а активность такого белка можно определить с помощью, например, описываемых в настоящем документе анализов.
Для определения процента идентичности двух или более аминокислот или двух или более нуклеотидных последовательностей были разработаны несколько компьютерных программных продуктов. Идентичность двух или более последовательностей можно рассчитать с помощью, например, программного обеспечения fasta, которое в настоящее время используется в версии fasta 3 (W.R. Pearson and D.J. Lipman, PNAS 85, 2444(1988); W.R. Pearson, Methods in Enzymology 183, 63 (1990); W.R. Pearson and D.J. Lipman, PNAS 85, 2444 (1988); W.R. Pearson, Enzymology 183, 63 (1990)). Другой используемой программой для расчета значений идентичности различных последовательностей является стандартная программа blast, которая включена в программное обеспечение Biomax pedant (Biomax, Мюнхен, Федеративная Республика Германия). К сожалению, она иногда дает недостаточные результаты, поскольку blast не всегда включает полные рассматриваемые и запрашиваемые последовательности. Тем не менее, поскольку указанная программа очень эффективна, ее можно применять для сравнения с большим количеством последовательностей. Для таких сравнений последовательностей обычно используют приведенные далее установки:
-р Program Name [String]; -d Database [String]; default = nr; -i Query File [File In]; default = stdin; -e Expectation value (E) [Real]; default = 10.0; -m alignment view options: 0 = pairwise; 1 = query-anchored showing identities; 2 = query-anchored no identities; 3 = flat query-anchored, show identities; 4 = flat query-anchored, no identities; 5 = query-anchored no identities and blunt ends; 6 = flat query-anchored, no identities and blunt ends; 7 = XML Blast output; 8 = tabular; 9 tabular with comment lines [Integer]; default = 0; -o BLAST report Output File [File Out] Optional; default = stdout; -F Filter query sequence (DUST with blastn, SEG with others) [String]; default = T; -G Cost to open a gap (zero invokes default behavior) [Integer]; default = 0; -E Cost to extend a gap (zero invokes default behavior) [Integer]; default = 0; -X X dropoff value for gapped alignment (in bits) (zero invokes default behavior); blastn 30, megablast 20, tblastx 0, all others 15 [Integer]; default = 0; -I Show GI's in deflines [T/F]; default = F; -q Penalty for a nucleotide mismatch (blastn only) [Integer]; default = -3; -r Reward for a nucleotide match (blastn only) [Integer]; default = 1; -v Number of database sequences to show one-line descriptions for (V) [Integer]; default = 500; -b Number of database sequence to show alignments for (B) [Integer]; default = 250; -f Threshold for extending hits, default if zero; blastp 11, blastn 0, blastx 12, tblastn 13; tblastx 13, megablast 0 [Integer]; default = 0; -g Perfom gapped alignment (not available with tblastx) [T/F]; default = T; -Q Query Genetic code to use [Integer]; default = 1; -D DB Genetic code (for tblastf[nx] only) [Integer]; default = 1; -a Number of processors to use [Integer]; default = 1; -O SeqAlign file [File Out] Optional; -J Believe the query defline [T/F]; default = F; -M Matrix [String]; default = BLOSUM62; -W Word size, default if zero (blastn 11, megablast 28, all others 3) [Integer]; default = 0; -z Effective length of the database (use zero for the real size) [Real]; default = 0; -K Number of best hits from a region to keep (off by default, if used a value of 100 is recommended) [Integer]; default = 0; -P 0 for multiple hit, 1 for single hit [Integer]; default = 0; -Y Effective length of the search space (use zero for the real size) [Real]; default = 0; -S Query strands to search against database (for blast[nx], and tblastx); 3 is both, 1 is top, 2 is bottom [Integer]; default = 3; -T Produce HTML output [T/F]; default = F; -1 Restrict search of database to list of GI's [String] Optional; -U Use lower case filtering of FASTA sequence [T/F] Optional; default = F; -у X dropoff value for ungapped extensions in bits (0.0 invokes default behavior); blastn 20, megablast 10, all others 7 [Real]; default = 0.0; -Z X dropoff value for final gapped alignment in bits (0.0 invokes default behavior); blastn/megablast 50, tblastx 0, all others 25 [Integer]; default = 0; -R PSI-TBLASTN checkpoint file [File In] Optional; -n MegaBlast search [T/F]; default = F; -L Location on query sequence [String] Optional; -A Multiple Hits window size, default if zero (blastn/megablast 0, all others 40 [Integer]; default = 0; -w Frame shift penalty (OOF algorithm for blastx) [Integer]; default = 0; -t Length of the largest intron allowed in tblastn for linking HSPs (0 disables linking) [Integer]; default = 0.
Результаты с высокой идентичностью получают с помощью алгоритма Needleman и Wunsch или Smith и Waterman. В силу вышесказанного, предпочтительными являются программы, основанные на упомянутых выше алгоритмах. Преимущественно, сравнения последовательностей можно осуществлять с помощью программы PileUp (J. Mol. Evolution., 25, 351 (1987), Higgins et al., CABIOS 5, 151 (1989)) или предпочтительно с помощью программ «Gap» и «Needle», оба из которых основаны на алгоритмах Needleman и Wunsch (J. Mol. Biol. 48; 443 (1970)), и «BestFit», который основан на алгоритме Smith и Waterman (Adv. Appl. Math. 2; 482 (1981)). «Gap» и «BestFit» являются частью пакета прикладных программ GCG (Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711 (1991); Altschul et al., (Nucleic Acids Res. 25, 3389 (1997)), «Needle» является частью пакета The European Molecular Biology Open Software Suite (EMBOSS) (Trends in Genetics 16 (6), 276 (2000)). Таким образом, предпочтительно расчеты для определения процента идентичности последовательностей выполняют с помощью программ «Gap» или «Needle» на протяжении всего диапазона последовательностей. Для «Needle» были использованы следующие стандартные настройки для сравнения последовательностей нуклеиновых кислот: matrix: EDNAFULL, Gap_penalty: 10.0, Extend_penalty: 0.5. Для «Gap» были использованы следующие стандартные настройки для сравнения последовательностей нуклеиновых кислот: gap weight: 50, length weight: 3, average match: 10,000, average mismatch: 0,000.
Например, последовательность, о которой говорят, что она на 80% идентична последовательности SEQ ID NO: 1 на уровне нуклеиновой кислоты, понимают как означающую последовательность, которая, при сравнении с последовательностью, представленной с SEQ ID NO: 1, при помощи упомянутой выше программы «Needle» с указанным выше набором параметров, является на 80% идентичной. Предпочтительно идентичность рассчитывают по полной длине запрашиваемой последовательности, например, SEQ ID NO: 1.
Выделенный: термин «выделенный» в контексте настоящей заявки означает, что материал был удален при помощи человека и существует отдельно от своего исходного природного окружения и, таким образом, не является естественным продуктом. Выделенный материал или молекула (такая как молекула ДНК или фермент) могут существовать в очищенной форме или могут существовать в неприродном окружении, таком как, например, в трансгенной клетке-хозяине. Например, встречающаяся в природе молекула нуклеиновой кислоты или полипептид, присутствующие в живой клетке, не являются выделенными, но та же молекула нуклеиновой кислоты или полипептид, отделенные от некоторых или всех сосуществующих в природной системе материалов, являются выделенными. Такие молекулы нуклеиновых кислот могут быть частью вектора, и/или такие молекулы нуклеиновых кислот или пептиды могут быть частью композиции, и могут быть выделенными в том отношении, что такой вектор или композиция не являются частью исходного окружения. Предпочтительно термин «выделенный» при использовании в отношении молекулы нуклеиновой кислоты, как в случае «выделенная последовательность нуклеиновой кислоты», относится к последовательности нуклеиновой кислоты, которая выявлена и отделена по меньшей мере от одной посторонней молекулы нуклеиновой кислоты, с которой она в норме ассоциирована в ее природном источнике. Выделенная молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, присутствующую в форме или окружающей среде, которая отличается от таковой, которая встречается в природе. И наоборот, невыделенные молекулы нуклеиновых кислот представляют собой такие молекулы нуклеиновых кислот, как ДНК и РНК, которые встречаются в состоянии, в котором они существуют в природе. Например, заданная последовательность ДНК (например, ген) встречается на хромосоме клетки-хозяина вблизи соседских генов; последовательности РНК, такие как последовательность мРНК, кодирующая конкретный белок, встречаются в клетке в виде смеси с множеством других мРНК, которые кодируют множество белков. Тем не менее, выделенная последовательность нуклеиновой кислоты, содержащая, например, SEQ ID NO: 1, включает, к примеру, такие последовательности нуклеиновых кислот в клетках, которые обычно содержат SEQ ID NO: 1, где последовательность нуклеиновой кислоты находится в геномной или плазмидной локализации, отличной от локализации у естественных клеток, или иным образом фланкирована последовательностью нуклеиновой кислоты, отличной от встречающейся в природе. Выделенная последовательность нуклеиновой кислоты может присутствовать в одноцепочечной или двуцепочечной форме. При необходимости использования выделенной последовательности нуклеиновой кислоты для экспрессии белка, последовательность нуклеиновой кислоты будет содержать как минимум по меньшей мере часть смысловой или кодирующей цепи (т.е. последовательность нуклеиновой кислоты может быть одноцепочечной). Альтернативно, она может содержать как смысловую, так и антисмысловую цепи (т.е. последовательность нуклеиновой кислоты может быть двуцепочечной).
Некодирующая: термин «некодирующая» относится к последовательностям молекул нуклеиновых кислот, которые не кодируют часть экспрессируемого белка или весь экспрессируемый белок. Некодирующие последовательности включают без ограничения энхенсеры, промоторные участки, 3' нетранслируемые участки и 5' нетранслируемые участки.
Нуклеиновые кислоты и нуклеотиды: термины «нуклеиновые кислоты» и «нуклеотиды» относятся к встречающимся в природе или синтетическим или искусственным нуклеиновой кислоте или нуклеотидам. термины «нуклеиновые кислоты» и «нуклеотиды» включают дезоксирибонуклеотиды, или рибонуклеотиды, или любой нуклеотидный аналог и их полимеры или гибриды либо в одно-, либо в двухцепочечной, смысловой или антисмысловой форме. Если не указано иное, конкретная последовательность нуклеиновой кислоты также подразумеваемым образом охватывает ее консервативно модифицированные варианты (например, замены в рамках вырожденности кодона) и комплементарные последовательности, а также открытым текстом указанную последовательность. Термин «нуклеиновая кислота» в настоящем документе используют взаимозаменяемо с терминами «ген», «кДНК», «мРНК», «олигонуклеотид» и «молекула нуклеиновой кислоты». Нуклеотидные аналоги включают нуклеотиды, имеющие модификации в химической структуре основания, сахара и/или фосфата, включая без ограничений модификации пиримидина по 5-положениюи, модификации пурина по 8-позиции, модификации в экзоциклических аминах цитозина, замену 5-бромурацила и др.; а также модификации сахаров по 2'-позиции, включая без ограничений рибонуклеотиды с модифицированными сахарами, у которых 2'-ОН заменен группой, выбранной из Н, OR, R, галогена, SH, SR, NH2, NHR, NR2 или CN. Короткие шпилечные РНК (shRNA) также могут содержать неприродные элементы, такие как, например, ионозин и ксантин, неприродные сахара, например, 2'-метоксирибозу, или неприродные фосфодиэфирные мостики, например, метилфосфонаты, фосфортиоаты и пептиды.
Последовательность нуклеиновой кислоты: фраза «последовательность нуклеиновой кислоты» относится к одно- или двухцепочечному полимеру дезоксирибонуклеотидных или рибонуклеотидных оснований, читаемых от 5'- к 3'-концу. Она включает хромосомную ДНК, самореплицирующиеся плазмиды, полимеры ДНК или РНК патогенов и ДНК или РНК, которые выполняют в основном структурную роль. «Последовательность нуклеиновой кислоты» также относится к последовательному перечню аббревиатур, букв, символов или слов, которые символизируют нуклеотиды. В соответствии с одним вариантом осуществления нуклеиновая кислота может представлять собой «зонд», который является относительно короткой нуклеиновой кислотой, обычно длиной менее 100 нуклеотидов. Зачастую зонд для нуклеиновой кислоты составляет в длину от приблизительно 50 нуклеотидов до приблизительно 10 нуклеотидов. «Целевой участок» нуклеиновой кислоты представляет собой часть нуклеиновой кислоты, которая представляет интерес по результатам выявления. «Кодирующий участок» нуклеиновой кислоты представляет собой часть нуклеиновой кислоты, которая транскрибируется и транслируется специфичным для последовательности образом с преобразованием в конкретный полипептид или белок при нахождении под контролем соответствующих регуляторных последовательностей. Считается, что кодирующий участок кодирует такой полипептид или белок.
Олигонуклеотид: термин «олигонуклеотид» относится к олигомеру или полимеру рибонуклеиновой кислоты (РНК) или дезоксирибонуклеиновой кислоты (ДНК) или их миметиков, а также к олигонуклеотидам с не встречающимися в природе частями, которые функционируют сходным образом. Такие модифицированные или замещенные олигонуклеотиды зачастую предпочтительны относительно нативных форм из-за таких необходимых свойств, как, например, повышенное клеточное поглощение, повышенная аффинность к целевой нуклеиновой кислоте и повышенная стабильность в присутствии нуклеаз. Олигонуклеотид предпочтительно включает два или более нуклеомономера, ковалентно связанных с другими при помощи связей (например, фосфодиэфирных) или заменяющих связей.
Выступающая часть: «выступающая часть» представляет собой относительно короткую одноцепочечную нуклеотидную последовательность на 5'- или 3'-гидроксильном конце двухцепочечной олигонуклеотидной молекулы (также называемую «удлиняющим сегментом», «выступающим концом» или «липким концом»).
Полипептид: термины «полипептид», «пептид», «олигопептид», «полипептид», «продукт гена», «продукт экспрессии» и «белок» в настоящем документе используют взаимозаменяемо для отсылки к полимеру или олигомеру последовательных аминокислотных остатков.
Промотор: термины «промотор» или «промоторная последовательность» являются эквивалентами и в контексте настоящей заявки относятся к последовательности ДНК, которая в связанном с возможностью выполнения своих функций состоянии с представляющей интерес нуклеотидной последовательностью может контролировать транскрипцию представляющей интерес нуклеотидной последовательности в РНК. Промотор расположен 5' (т.е. выше) к сайту начала транскрипции представляющей интерес нуклеотидной последовательности, транскрипцию которой в мРНК он контролирует, и представляет собой сайт специфического связывания РНК-полимеразой и другими транскрипционными факторами для инициации транскрипции. Промотор не содержит кодирующих участков или 5' нетранслируемых участков. Промотор может, например, быть гетерологичным или гомологичным для соответствующей клетки. Последовательность молекулы нуклеиновой кислоты является «гетерологичной для» организма или второй последовательности молекулы нуклеиновой кислоты, если она происходит от чужеродного вида, или, если от того же вида, то модифицирована относительно изначальной формы. Например, промотор, связанный с возможностью выполнения своих функций с гетерологичной кодирующей последовательностью, относится к кодирующей последовательности от вида, отличающегося от вида, от которого был получен промотор, или, если от того же вида, то к кодирующей последовательности, которая в норме не ассоциирована с таким промотором (например, генетически сконструированной кодирующей последовательностью или аллелью из отличного экотипа или вариетета). Подходящие промоторы можно получить из генов клеток-хозяев, в которых должна происходить экспрессия, или от патогенов для такого хозяина.
Очищенная: в контексте настоящей заявки термин «очищенная» относится к молекулам, последовательностям либо нуклеиновых кислот, либо аминокислот, которые удалены из их природного окружения, выделены или отделены. «Практически очищенные» молекулы по меньшей мере на 60%, предпочтительно по меньшей мере на 75% и более предпочтительно по меньшей мере на 90% очищены от других компонентов, с которыми они в норме ассоциированы. Очищенная последовательность нуклеиновой кислоты может быть выделенной последовательностью нуклеиновой кислоты.
Рекомбинантная: термин «рекомбинантная» в отношении молекул нуклеиновых кислот относится к молекулам нуклеиновых кислот, получаемым с помощью методик рекомбинантных ДНК. Термин также охватывает молекулы нуклеиновых кислот, которые сами по себе не существуют в природе, но являются модифицированными, измененными, подвергнутыми мутации или другим манипуляциям, осуществляемым человеком. Предпочтительно «рекомбинантная молекула нуклеиновой кислоты» представляет собой не встречающуюся в природе молекулу нуклеиновой кислоты, которая отличается последовательностью от встречающейся в природе молекулы нуклеиновой кислоты по меньшей мере на одну нуклеиновую кислоту. «Рекомбинантная молекула нуклеиновой кислоты» также может охватывать «рекомбинантную конструкцию», которая содержит, предпочтительно связанную с возможностью выполнения своих функций, последовательность молекул нуклеиновых кислот, которые не встречаются в природе в таком порядке. Предпочтительные способы получения указанной рекомбинантной молекулы нуклеиновой кислоты могут охватывать методики клонирования, направленный или ненаправленный мутагенез, методики синтеза или рекомбинационные методики.
Значимое увеличение: увеличение, например, ферментативной активности, экспрессии гена, производительности или выхода для некоторого продукта, которое превышает допустимую погрешность, присущую методике измерения, предпочтительно увеличение на приблизительно 10% или 25% предпочтительно на 50% или 75%, более предпочтительно в 2 или 5 раз или более активности, экспрессии, производительности или выхода контрольного фермента или экспрессии в контрольной клетке, производительности или выхода у контрольной клетки, еще более предпочтительно увеличение приблизительно в 10 раз или более.
Значимое снижение: снижение, например, ферментативной активности, экспрессии гена, производительности или выхода для некоторого продукта, которое превышает допустимую погрешность, присущую методике измерения, предпочтительно снижение по меньшей мере на приблизительно 5% или 10%, предпочтительно по меньшей мере на приблизительно 20% или 25%, более предпочтительно по меньшей мере на приблизительно 50% или 75%, еще более предпочтительно по меньшей мере на приблизительно 80% или 85%, наиболее предпочтительно по меньшей мере на приблизительно 90%, 95%, 97%, 98% или 99%.
Практически комплементарная: в своем наиболее широком смысле термин «практически комплементарная», при использовании в настоящем документе применительно к нуклеотидной последовательности в отношении эталонной или целевой нуклеотидной последовательности, означает нуклеотидную последовательность с процентом идентичности между практически комплементарной нуклеотидной последовательностью и полностью комплементарной последовательностью указанной эталонной или целевой нуклеотидной последовательностью, составляющим по меньшей мере 60%, желательнее по меньшей мере 70%, желательнее по меньшей мере 80% или 85%, предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере 93%, еще более предпочтительно по меньшей мере 95% или 96%, и еще более предпочтительно по меньшей мере 97% или 98%, и еще более предпочтительно по меньшей мере 99% или наиболее предпочтительно 100% (при этом последнее значение в данном контексте эквивалентно термину «идентичная»). Предпочтительно идентичность оценивают на протяжении отрезка, составляющего по меньшей мере 19 нуклеотидов, предпочтительно по меньшей мере 50 нуклеотидов, более предпочтительно по всей длине последовательности нуклеиновой кислоты с указанной эталонной последовательностью (если далее не указано иное). Сравнения последовательностей проводят с помощью анализа GAP с настройками по умолчанию с помощью University of Wisconsin GCG, SEQWEB приложения GAP, в основе которого лежит алгоритм Needleman и Wunsch (Needleman and Wunsch (1970) J Mol. Biol. 48: 443-453; определение которому дано выше). Нуклеотидная последовательность, «практически комплементарная» эталонной нуклеотидной последовательности, гибридизируется с эталонной нуклеотидной последовательностью в условиях низкой жесткости, предпочтительно в условиях средней жесткости, наиболее предпочтительно в условиях высокой жесткости (определение которым дано ранее).
Трансген: термин «трансген» в контексте настоящей заявки относится к любой последовательности нуклеиновой кислоты, которую вносят в геном клетки при помощи манипуляций в ходе эксперимента. Трансген может представлять собой «эндогенную последовательность ДНК» или «гетерологичную последовательность ДНК» (т.е. «чужеродную ДНК»). Термин «эндогенная последовательность ДНК» относится к нуклеотидной последовательности, которая в природе встречается в клетке, в которую ее вносят, при условии, что она не содержит некую модификацию (например, точечную мутацию, наличие селектируемого маркерного гена т.д.) относительно встречающейся в природе последовательности.
Трансгенный: термин трансгенный применительно к организму означает трансформированный, предпочтительно стабильно трансформированный, рекомбинантной молекулой ДНК, которая предпочтительно содержит подходящий промотор, связанный с возможностью осуществления своих функций с представляющей интерес последовательностью ДНК.
Вектор: в контексте настоящей заявки термин «вектор» относится к молекуле нуклеиновой кислоты, способной переносить другую молекулу нуклеиновой кислоты, с которой она связана. Одним типом вектора является интегрируемый в геном вектор или «интегрируемый вектор», который может стать интегрированным в геномную ДНК клетки-хозяина. Другим типом вектора является эписомальный вектор, т.е. плазмида или молекула нуклеиновой кислоты, способная к внехромосомной репликации. Векторы, способные управлять экспрессией генов, с которыми они связаны с возможностью осуществления своих функций, в настоящем документе называют «векторами экспрессии». В настоящем описании «плазмида» и «вектор» используют взаимозаменяемо, если из контекста не очевидно иное.
Дикий тип: термин «дикий тип», «природный» или «природного происхождения» означает применительно к организму, что указанный организм не изменен, не подвергнут мутации или другим манипуляциям, осуществляемым человеком. Что касается полипептидной последовательности или последовательности нуклеиновой кислоты, то такая полипептидная последовательность или последовательность нуклеиновой кислоты является встречающейся в природе или доступна по меньшей мере у одного встречающегося в природе организма, которая не была подвергнута изменениям, мутациям или другим манипуляциям, осуществляемым человеком.
Дикий тип микроорганизма относится к микроорганизму, чей геном находится в состоянии, которое предшествовало внесению генетической модификации некоторого гена. Генетической модификацией может быть, например, делеция гена или его части или точечная мутация или внесение гена.
Термины «производство» или «производительность» известны из уровня техники и включают концентрацию продукта ферментации (например, аланина), образующегося за заданное время и в заданном объеме ферментации (например, кг продукта за час на литр). Термин «эффективность получения» включает время, необходимое для достижения определенного уровня получения (например, как много времени нужно клетке для достижения определенной скорости выхода химического продукта тонкого органического синтеза). Производительность может также означать объемную производительность, которую определяют как количество образовавшегося продукта, деленное на объем реактора и на время.
Термин «выход» или «выход продукт/углерод» известен из уровня техники и включает эффективность преобразования источника углерода в продукт (т.е. химический продукт тонкого органического синтеза). Его обычно записывают как, например, кг продукта на кг источника углерода. При увеличении выхода или получения соединения увеличивается количество получаемых молекул или пригодных получаемых молекул такого соединения в заданном количестве культуры на протяжении заданного количества времени.
Термин «рекомбинантный микроорганизм» включает микроорганизмы, которые были генетически модифицированы так, чтобы у них наблюдали измененный или отличный генотип и/или фенотип (например, когда генетическая модификация затрагивает кодирующие последовательности нуклеиновых кислот микроорганизма) по сравнению с микроорганизмом дикого типа, из которого он был получен. Рекомбинантный микроорганизм содержит по меньшей мере одну рекомбинантную молекулу ДНК.
Термин «рекомбинантная» применительно к ДНК относится к молекулам ДНК, полученным человеком с помощью методик рекомбинантных ДНК. Термин охватывает молекулы ДНК, которые сами по себе не существуют в природе или не существуют в организме, из которого получают ДНК, но являются подвергнутыми модификации, изменению, мутации или другой манипуляции, осуществляемой человеком. Предпочтительно «рекомбинантная молекула ДНК» представляет собой не встречающуюся в природе молекулу нуклеиновой кислоты, которая отличается последовательностью от встречающейся в природе молекулы нуклеиновой кислоты по меньшей мере на одну нуклеиновую кислоту. «Рекомбинантная молекула ДНК» также может охватывать «рекомбинантную конструкцию», которая содержит, предпочтительно связанную с возможностью выполнения своих функций, последовательность молекул нуклеиновых кислот, которые не встречаются в природе в таком порядке. Предпочтительные способы получения указанной рекомбинантной молекулы ДНК могут охватывать методики клонирования, направленный или ненаправленный мутагенез, методики синтеза гена или рекомбинационные методики.
Примером такой рекомбинантной ДНК является плазмида, в которую была вставлена гетерологичная ДНК-последовательность, или ген или промотор, который был подвергнут мутации по сравнению с геном или промотором, из которого была получена рекомбинантная ДНК. Мутацию можно внести посредством методик направленного мутагенеза, известных из уровня техники, или с помощью методик неспецифического мутагенеза, таких как химический мутагенез, мутагенез с помощью УФ-света или рентгеновского излучения, или методик направленной эволюции.
Термин «направленная эволюция» в настоящем документе используют синонимично с термином «метаболическая эволюция», и он включает применение давления отбора, которое способствует росту мутантов с представляющими интерес признаками. В основе давления отбора могут лежать различные условия культивирования, отбор, сопряженный с уровнем АТР и ростом, и отбор, связанный с окислительно-восстановительными процессами. Давление отбора можно оказывать с помощью периодической ферментации с серийным переносом засева или непрерывного культивирования с таким же давлением.
Термин «экспрессия» или «экспрессия гена» означает транскрипцию конкретного гена(генов) или конкретной генетической векторной конструкции. Термин «экспрессия» или «экспрессия гена», в частности, означает транскрипцию гена(генов) или генетической векторной конструкции в мРНК. Процесс включает транскрипцию ДНК и может включать процессинг получаемого в результате РНК-продукта. Термин «экспрессия» или «экспрессия гена» также может включать трансляцию мРНК и вместе с тем синтез кодируемого белка, т.е. экспрессию белка.
Фиг. 1
Проверка клонов после инактивации генов ackA-pta.
А: ПЦР-ампликон, полученный с геномной ДНК Е. coli W ΔackA-pta::FRT с праймерами P395-ackA-pta-check2 и P395-ackA-pta-check5 (338 п.о.). М: маркер размеров ДНК. В: секвенирование ПЦР-ампликона с P395-ackA-pta-check2 и P395-ackA-pta-check5 подтверждало точную модификацию пар оснований по локусу ackA-pta. Нуклеотиды, которые были подтверждены по результатам секвенирования, выделены курсивом. Оставшийся сайт FRT показан зеленым, фланкирующие сайты связывания с праймером показаны красным. Верхним регистром: кодирующая последовательность. Нижним регистром: межгенные участки.
Фиг. 2
Проверка клонов после инактивации гена adhE.
А: ПЦР-ампликон, полученный с геномной ДНК Е. coli W ΔackA-pta::FRT ΔadhE::FRT с праймерами Р395-adhE-check2 и P395-adhE-check5 (569 п.о.). М: маркер размеров ДНК. В: секвенирование ПЦР-ампликона с P395-adhE-check2 и P395-adhE-check5 подтверждало точную модификацию пар оснований по локусу adhE. Нуклеотиды, которые были подтверждены по результатам секвенирования, выделены курсивом. Оставшийся сайт FRT показан зеленым, фланкирующие сайты связывания с праймером показаны красным. Верхним регистром: кодирующая последовательность. Нижним регистром: межгенные участки.
Фиг. 3
Проверка клонов после инактивации генов frdABCD.
А: ПЦР-ампликон, полученный с геномной ДНК Е. coli W ΔackA-pta::FRT ΔadhE::FRT ΔfrdABCD::FRT с праймерами P395-frd-check1 и P395-frd-check4 (797 п.о.). М: маркер размеров ДНК. В: секвенирование ПЦР-ампликона с P395-frd-check1 и P395-frd-check4 подтверждало точную модификацию пар оснований по локусу frd. Нуклеотиды, которые были подтверждены по результатам секвенирования, выделены курсивом. Оставшийся сайт FRT показан зеленым, фланкирующие сайты связывания с праймером показаны красным. Верхним регистром: кодирующая последовательность. Нижним регистром: межгенные участки.
Фиг. 4
Проверка клонов после инактивации гена pflB.
А: ПЦР-ампликон, полученный с геномной ДНК Е. coli W ΔackA-pta::FRT ΔadhE::FRT ΔfrdABCD::FRT ΔpflB::FRT с праймерами P395-pflB-check1 и P395-pflB-check3 (511 п.о.). М: маркер размеров ДНК. В: секвенирование ПЦР-ампликона с P395-pflB-check1 и P395-pflB-check3 подтверждало точную модификацию пар оснований по локусу pflB. Нуклеотиды, которые были подтверждены по результатам секвенирования, выделены курсивом. Оставшийся сайт FRT показан зеленым, фланкирующие сайты связывания с праймером показаны красным. Верхним регистром: кодирующая последовательность. Нижним регистром: межгенные участки.
Фиг. 5
Проверка клонов после встраивания гена alaD-gstear.
А: ПЦР-ампликон, полученный с геномной ДНК Е. coli W ΔackA-pta::FRT ΔadhE::FRT ΔfrdABCD::FRT ΔpflB::FRT ΔldhA::alaD-gstear с праймерами P395-ldhA-check1 и P395-ldhA-check2 (1833 п.о.). M: маркер размеров ДНК. В: секвенирование ПЦР-ампликона с P395-ldhA-check1 и P395-ldhA-check2 подтверждало точную модификацию пар оснований по локусу ldhA и встраивание alaD-gstear. Нуклеотиды, которые были подтверждены по результатам секвенирования, выделены курсивом. ORF alaD-gstear показана голубым, оставшийся сайт FRT показан зеленым, фланкирующие сайты связывания с праймером показаны красным. Верхним регистром: кодирующая последовательность. Нижним регистром: межгенные участки.
Фиг. 6
Метаболическая карта синтеза аланина у микроорганизма по настоящему изобретению.
Красными звездами показаны нокауты ферментативной активности.
Зелеными стрелками показана внесенная ферментативная активность
J7 представляет собой ldh - лактатдегидрогеназу, KO уменьшает выработку лактата.
J6 представляет собой frdABCD - фумаратредуктазу, KO уменьшает выработку сукцината.
J8 представляет собой pfl - пируватформиатлиазу, KO уменьшает выработку ацетата и этанола.
J10 представляет собой ack-pta - фосфотрансацетилазу-ацетаткиназу, KO уменьшает выработку ацетата.
J11 представляет собой adhE - алкогольдегидрогеназу, KО уменьшает выработку этанола.
Фиг. 7
Периодическая ферментация Е. coli QZ20 и Е. coli QZ48 (А96Е в ArgP) в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 8
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ20 и QZ48 (А96Е в ArgP) спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 9
Периодическая ферментация Е. coli QZ20/контрольная плазмида pACYC184 и Е. coli QZ20/pACYC184-argP в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 10
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ20/pACYC184-argP спустя 20 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 11
Периодическая ферментация Е. coli QZ20 и Е. coli QZ58 (SNP в промоторе gcvA/B) в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 12
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ20 и QZ58 (SNP в промоторе gcvA/B) спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 13
Периодическая ферментация Е. coli QZ48 (А96Е в ArgP) и Е. coli QZ66 (А96Е в Arg, SNP в промоторе gcvA/B) в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 14
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ48 (А96Е в ArgP) и Е. coli QZ66 (А96Е в ArgP, SNP в промоторе gcvA/B) спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 15
Относительный анализ транскрипции генов (A) gcvA и (В) gcvB у Е. coli QZ20 и QZ23 через 8 часов, 11 часов и 28 часов в ходе периодической ферментации относительно Е. coli QZ20, через 8 часов. Все полученные в ходе qPCR данные нормализовали по гену rrsA в качестве эталона.
Фиг. 16
Периодическая ферментация Е. coli QZ20/контрольная плазмида pACYC184 и Е. coli QZ20/pACYC184-gcvA и Е. coli QZ20/pACYC184-gcvB в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Образование аланина коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 17
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ20 с контрольными плазмидами pACYC184, pACYC184-gcvA и pACYC184-gcvB спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 18
Периодическая ферментация Е. coli QZ20 и Е. coli QZ71 (нокаут по gcvB) в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 19
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ20 и Е. coli QZ71 (нокаут по gcvB) спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 20
Периодическая ферментация Е. coli QZ20, Е. coli QZ57 (brnQΔ667-764) и Е. coli QZ69 (KO по brnQ) в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 21
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ20, Е. coli QZ57 (brnQΔ667-764) и Е. coli QZ69 (KO по brnQ) спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 22
Периодическая ферментация Е. coli QZ20 и Е. coli QZ56 (LpxD А15Т) в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 23
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ20 и QZ56 (LpxD А15Т) спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Фиг. 24
Периодическая ферментация Е. coli QZ68 (А96Е в ArgP, SNP в промоторе gcvA/B, brnQΔ667-764) и Е. coli QZ70 (А96Е в ArgP, SNP в промоторе gcvA/B, brnQΔ667-764, lpxD А15Т) в 500 мл среды AM 1 с 140 г/л глюкозы. Ферментацию поддерживали при 37°С, 400 об/мин, при рН 6,8 с 5 Н NH4OH, без аэрации. Показано образование аланина, которое коррелировало в зависимости от концентраций аланина в образцах и скорости потребления NH4OH.
Фиг. 25
Волюметрическая производительность по аланину (объемная производительность), определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у Е. coli QZ68 (А96Е в ArgP, SNP в промоторе gcvA/B, brnQΔ667-764) и Е. coli QZ70 (А96Е в ArgP, SNP в промоторе gcvA/B, brnQΔ667-764, lpxD А15Т) спустя 46 часов периодической ферментации в 500 мл среды AM 1 с 140 г/л глюкозы в качестве источника углерода.
Примеры
Химические соединения и общепринятые способы
Если не указано иное, процедуры клонирования, которые осуществляли для реализации настоящего изобретения, в том числе рестрикционное расщепление, электрофорез в агарозном геле, очистка нуклеиновых кислот, лигирование нуклеиновых кислот, трансформацию, отбор и культивирование бактериальных клеток, проводили? как описано (Sambrook et al., 1989). Анализы последовательностей рекомбинантной ДНК проводили с помощью лазерного флуоресцентного секвенатора ДНК (Applied Biosystems, Фостер Сити, Калифорния, США) с применением технологии Сэнгера (Sanger et al., 1977). Если не описано иное, химические соединения и реактивы были получены от Sigma Aldrich (Sigma Aldrich, Сент-Луис, США), от Promega (Мэдисон, Висконсин, США), Duchefa (Харлем, Нидерланды) или Invitrogen (Карлсбад, Калифорния, США). Рестрикционные эндонуклеазы были получены от New England Biolabs (Ипсуич, Массачусетс, США) или Roche Diagnostics GmbH (Пенцберг, Германия). Олигонуклеотиды были синтезированы с помощью Eurofins MWG Operon (Эберсберг, Германия).
Пример 1
Е. coli W (LU17032) конструировали для получения L-аланина путем инактивации ORF ackA-pta, adhE, frdABCD и pflB и замены ORF ldhA на вариант гена alaD с оптимизированным кодоном (alaD-gstear).
ORF ackA-pta, adhE, frdABCD и pflB инактивировали с помощью вставки FRT-фланкированной кассеты с геном устойчивости к канамицину с последующим удалением кассеты с геном устойчивости к канамицину путем FLP-рекомбинации.
Ген ldhA заменяли на alaD-gstear и нижележащую FRT-фланкированную кассету с геном устойчивости к зеоцину, которую в конце удаляли путем FLP-рекомбинации.
Материалы и способы
Бактериальная культура
Е. coli W (LU17032) культивировали в жидкой среде Луриа-Бертани (LB) или на твердой среде Луриа-Бертани. По мере необходимости клоны пересеивали на агаризованную минимальную среду М9, содержавшую 10 мМ сахарозы, для подтверждения идентичности со штаммом W. В соответствующих случаях в жидкую и твердую среды добавляли антибиотики до конечной концентрации 15 мкг/мл (канамицина, хлорамфеникола), 25 мкг/мл (зеоцина) или 3 мкг/мл (тетрациклина).
Red/ET рекомбинация
Red/ET рекомбинацию осуществляли с помощью стандартных протоколов Gene Bridges GmbH (www.genebridges.com). Если кратко, Red/ET-компетентный Е. coli W выращивали в аэробных условиях при 30°С до ОD600нм, составлявшей ~0,3. Экспрессию генов red индуцировали путем добавления 50 мкл 10% (масса/объем) L-арабинозы с последующим увеличением температуры до 37°С. Для неиндуцируемых контрольных культур добавление арабинозы пропускали. Спустя 35 минут инкубирования при 37°С клетки промывали два раза ледяным 10% (объем/объем) глицерином и подвергали электропорации с 500 нг ПЦР-продукта при 1,35 кВ, 10 мкФ, 600 Ом. Затем клетки ресуспендировали в 1 мл ледяной среды LB и выращивали в аэробных условиях при 37°С в течение примерно 1,5 часа. Затем культуры высеивали на агар LB, содержавший 15 мкг/мл канамицина (нокауты) или 25 мкг/мл зеоцина (нокин).
FLP рекомбинация
Фланкирующие сайты FRT делали возможным удаление маркеров устойчивости к антибиотикам путем FLP рекомбинации после модификации хромосомы Е. coli. FLP рекомбинация оставляет один сайт FRT (34 п.о.), а также короткие фланкирующие последовательности (примерно 20 п.о. каждая), которые использовали в качестве сайтов связывания с праймером при амплификации кассет.
Для осуществления FLP рекомбинации плазмиду 708-FLPe (таблица 1), кодирующую FLP рекомбиназу, вносили в Red/ET рекомбинанты путем электропорации. Трансформантов KanR CmR или ZeoR CmR использовали для инокуляции 0,2 мл культур LB, которые инкубировали при 30°С в течение 3 часов. FLP активность затем индуцировали путем сдвига температуры до 37°С с последующей трехчасовой инкубацией при 37°С. Отдельные колонии, полученные из этих культур, впоследствии подвергали скринингу по фенотипу CmS и KanS или ZeoS.
Получение и анализ ДНК
Геномную ДНК (gДНК) Е. coli выделяли из полученных за ночь культур при помощи набора Gentra Puregene Yeast/Bact. Kit В (Qiagen, Хильден, Германия). ПЦР-продукты, несущие нокаут- и нокин-кассеты, амплифицировали с матричных плазмид при помощи высокоточной ДНК-полимеразы PRECISOR (BioCat, Гейдельберг), а аналитические ПЦР-реакции проводили при помощи набора для проведения ПЦР PCR Extender System (5PRIME GmbH, Гамбург, Германия) согласно рекомендациям производителя. ПЦР-ампликоны очищали с помощью набора GeneJET PCR Purification Kit или набора GeneJET Gel Extraction Kit (Fermentas, Санкт-Леон-Рот, Германия), а секвенирование проводили с помощью GATC BioTech (Констанц, Германия) или BioSpring (Франкфурт-на-Майне, Германия).



1.1. Локус ackA-pta - таргетинг ackA-pta
Примерно 500 нг ПЦР-конструкции ΔackA-pta PCR (1737 п.о.) вводили при помощи электропорации в Red/ET-компетентные клетки Е. coli W. Восемь KanR трансформантов анализировали в отношении правильного встраивания кассеты с маркером устойчивости при помощи ПЦР с геном-специфичными праймерами. Три клона подвергали FLP рекомбинации, которую осуществляли, как описано в разделе Материалы и способы (данные не показаны).
Проверка клонов. Инактивацию локуса ackA-pta и удаления кассеты с маркером устойчивости к канамицину подтверждали при помощи ПЦР по оставшемуся следу FRT. Один клон, который давал правильный ПЦР-сигнал, также подтверждали при помощи секвенирования (фиг. 1).
1.2 Локус adhE - таргетинг adhE
Примерно 500 на ПЦР-конструкции ΔadhE (1093 п.о.) вводили при помощи электропорации в Red/ET-компетентные клетки Е. coli W, которые несли модификацию ΔackA-pta::FRT. Восемь KanR трансформантов анализировали в отношении правильного встраивания кассеты с маркером устойчивости при помощи ПЦР с геном-специфичными праймерами. Два клона подвергали FLP рекомбинации, которую осуществляли, как описано в разделе Материалы и способы (данные не показаны).
Проверка клонов. Инактивацию локуса adhE и удаление кассеты с маркером устойчивости к канамицину подтверждали при помощи ПЦР по оставшемуся следу FRT. Один клон, который давал правильный ПЦР-сигнал, также подтверждали при помощи секвенирования (фиг. 2).
1.3 Локус frd - таргетинг frdABCD
Примерно 500 на ПЦР-конструкции ΔfrdABCD (1093 п.о.) вводили при помощи электропорации в Red/ET-компетентные клетки Е. coli W, которые несли модификации ΔackA-pta::FRT и ΔadhE::FRT. Восемь KanR трансформантов анализировали в отношении правильного встраивания кассеты с маркером устойчивости при помощи ПЦР с геном-специфичными праймерами. Один клон подвергали FLP рекомбинации, которую осуществляли как описано в разделе Материалы и способы (данные не показаны).
Проверка клонов. Инактивацию локуса frd и удаления кассеты с маркером устойчивости к канамицину подтверждали при помощи ПЦР по оставшемуся следу FRT. Один клон, который давал правильный ПЦР-сигнал, также подтверждали при помощи секвенирования (фиг. 3).
1.4 Локус pflB - таргетинг pflB
Примерно 500 на ПЦР-конструкции ΔpflB (1093 п.о.) вводили при помощи электропорации в Red/ET-компетентные клетки Е. coli W, которые несли модификации ΔackA-pta::FRT, ΔadhE::FRT и ΔfrdABCD::FRT. Восемь KanR трансформантов анализировали в отношении правильного встраивания кассеты с маркером устойчивости при помощи ПЦР с геном-специфичными праймерами. Четыре клона подвергали FLP рекомбинации, которую осуществляли как описано в разделе Материалы и способы (данные не показаны).
Проверка клонов. Инактивацию локуса pflB и удаления кассеты с маркером устойчивости к канамицину подтверждали при помощи ПЦР по оставшемуся следу FRT. Один клон, который давал правильный ПЦР-сигнал, также подтверждали при помощи секвенирования (фиг. 4).
Локус 1.5 ldhA - нокин alaD-gstear
Примерно 500 на ПЦР-конструкции ΔldhA::alaD-gstear (1783 п.о.) вводили при помощи электропорации в Red/ET-компетентные клетки Е. coli W, которые несли модификации ΔackA-pta::FRT, ΔadhE::FRT, ΔfrdABCD::FRT и ΔpflB::FRT. Четыре ZeoR трансформантов анализировали в отношении правильного встраивания кассеты с маркером устойчивости при помощи ПЦР с геном-специфичными праймерами. Один клон подвергали FLP рекомбинации, которую осуществляли как описано в разделе Материалы и способы (данные не показаны).
Проверка клонов. Встраивание alaD-gstear и удаления кассеты с маркером устойчивости к зеоцину подтверждали при помощи ПЦР по оставшемуся следу FRT. Один клон, который давал правильный ПЦР-сигнал, также подтверждали при помощи секвенирования (фиг. 5).
Пример 2
ВЭЖХ детекция и количественная оценка аланина
Для детекции аланина в среде для выращивания культуры клеток использовали следующий способ ВЭЖХ:
колонка: колонка Aminex НРХ-87С (Bio-Rad), 300×7,8 мм, внутренний диаметр, размер частиц 9 мкм;
подвижная фаза: Са(NO3)2 в количестве 0,1 моль/л - 90%, ацетонитрил - 10%;
скорость потока: 0,6 мл/мин.;
температура колонки: 60°С;
детекция: рефрактометрический детектор.
В соответствии с указанным выше способом основные оцениваемые компоненты в матрице образцов культуры клеток можно полностью отделить от аланина, не препятствуя количественной оценке аланина.
Количество аланина в образце определяли с помощью способа градуировки по внешним стандартам. Образцы стандартов, содержавшие аланин в количестве от 0,5 до 10,0 г/л, вводили инъекцией, а для градуировки использовали площади пиков. Коэффициент линейной регрессии у градуировочной кривой составлял 0,9995.
Образцы вводили инъекцией за один раз в количестве 20 мкл. Площади пиков использовали для расчета присутствовавшего в образце количества с помощью программного обеспечения Waters LC Millenium.
Пример 3
ВЭЖХ детекция и количественная оценка глюкозы, сукцината, лактата, формиата, ацетата и этанола
В способе ВЭЖХ использовали следующее:
колонка: колонка Aminex НРХ-87Н (Bio-Rad), 300×7,8 мм, внутренний диаметр, размер частиц 9 мкм;
подвижная фаза: H2SO4, 4 мМ;
скорость потока: 0,4 мл/мин;
температура колонки: 45°С;
детекция: рефрактометрический детектор.
Количество анализируемых веществ определяли с помощью способа градуировки по внешним стандартам. Образцы стандартов, содержавшие глюкозу в количестве от 0,1 до 38,0 г/л, сукцинат, лактат, формиат, ацетат и этанол в количестве от 0,05 до 10,0 г/л, вводили инъекцией, а для градуировки использовали площади пиков. Коэффициенты линейной регрессии для всех шести градуировочных кривых были точнее 0,999.
Образцы вводили инъекцией за один раз в количестве 20 мкл. Площади пиков использовали для расчета присутствовавшего в образце количества с помощью программного обеспечения Waters LC Millenium.
Пример 4
Метаболическая эволюция линии W Е. coli, полученной в Примере 1, в отношении повышенного выхода аланина
Линию Е. coli, содержавшую все мутации, которые описаны в Примере 1, под названием Е. coli Exl или QZ16, использовали для процедуры метаболической эволюции на предмет повышения выхода аланина у линии Exl Е. coli.
Метаболическую эволюцию осуществляли следующим образом: на первом и втором раунде эволюции непрерывную эволюцию осуществляли в течение соответственно 500 часов и 750 часов в среде NBS с 5% глюкозы.
Среда NBS:
3,5 г KH2PO4;
5,0 г K2HPO4;
3,5 г (NH4)2HPO4;
0,25 г MgSO4-7H2O;
15 мг CaCl2-2H2O;
0,5 мг тиамина;
1 мл исходного раствора металлических микроэлементов.
Исходный раствор металлических микроэлементов готовили в 0,1 М HCl, 1,6 г FeCl3-6H2O; 0,2 г CoCl2-6H2O; 0,1 г CuCl2-2H2O; 0,2 г ZnCl2; 0,2 г NaMoO4-2H2O; 0,05 г Н3ВО3.
Клетки высевали штрихами на слои питательной среды LB и тестировали в отношении выхода аланина. Наилучшая линия Е. coli (Е. coli Ev1 или QZ17) в результате обеспечивала ферментацию со средой NBS, содержавшей 5% глюкозы, за 24 и 48 часов при 37°С с выходом аланина 84%-86% в сравнении с выходом аланина изначальной линии Е. coli Ex1, которая давала 80%-83%.
Е. coli Ev1 использовали для дополнительных стадий эволюции, которые осуществляли в виде периодической эволюции в течение 20 дней. 5% клеток повторно засевали в свежую среду каждые 24 часа, 48 часов, 72 часа и так далее в среду AM1, содержавшую 14% глюкозы, при 37°С.
Среда AM1:
19,92 мм (NH4)2HPO4 = 2,6 г/л молекулярная масса: 132,07;
7,56 мм NH4H2PO4 = 0,87 г/л молекулярная масса: 115;
2,0 мм KCl = 0,15 г/л молекулярная масса: 74,55;
1,5 мм MgSO4-7H2O = 0,37 г/л молекулярная масса: 246,5;
15 г/л сульфата аммония добавляли на последней стадии;
1 мм бетаина;
1 мл исходного раствора металлических микроэлементов.
Для получения 1 л исходного раствора металлических микроэлементов:
исходный раствор металлических микроэлементов готовили в 0,12 М HCl, 2,4 г FeCl3-6H2O; 0,3 г CoCl2-6H2O; 0,21 г CuCl2-2H2O; 0,3 г ZnCl2; 0,27 г NaMoO4-2H2O; 0,068 г H3BO3; 0,5 г MnCl2-4H2O.
В результате такой эволюции была выделена линия Е. coli Ev2, также называемая QZ18. Эту линию тестировали в отношении ферментации, которую проводили в ферментере со средой AM1 с 14% глюкозы. Линия Е. coli Ev2 характеризовалась выходом аланина от 92% до 94% в сравнении с выходом аланина у Е. coli Ev1, который составлял 91%-92% при тех же условиях.
После дополнительных стадий периодической эволюции в течение 300 часов в среде AM1, содержавшей 12% глюкозы, и последующих 10 стадий периодической эволюции в AM1, содержавшей 12% глюкозы, была выделена линия Е. coli Ev3, также называемая QZ20.
Из результатов тестирования на выход аланина видно, что линия Е. coli Ev3 характеризовалась выходом аланина от 94% до 96% в среде AM1, содержавшей 12% глюкозы, в сравнении с выходом аланина у Е. coli Ev2, который составлял 92%-93% при тех же условиях.
Дополнительную последовательную периодическую эволюцию, как описано выше, в течение периода 1000 часов в среде AM1 с 14% глюкозы проводили с Е. coli Ev3, и была выделена линия Е. coli Ev4, также называемая QZ23. Е. coli Ev4 тестировали в сравнении с Е. coli Ev3 в среде AM1 с 14% глюкозы. У линии Ev4 Е. coli наблюдали увеличенную производительность (объемную производительность) по аланину, определяемую как количество образовавшегося продукта, деленное на объем реактора и на время, которая составляла 2,0-2,4 г/(л.ч) по сравнению с 1,0-1,3 г(/л.ч) у линии Е. coli Ev3 спустя 46 ч ферментации.
Пример 5
Определение мутаций у линии Е. coli Ev4 в сравнении с Е. coli Ev3
Секвенировали геном линий Е. coli, Е. coli Ev4 и Е. coli Ev3, а результаты сравнивали с целью определения мутаций, которые приводили к увеличенной производительности по аланину у линии Е. coli Ev4.
Была выявлена мутация в гене brnQ, которая изменяла последовательность гена brnQ с SEQ ID NO: 1, кодирующей белок с SEQ ID NO: 2 у линии Е. coli Ev3, на SEQ ID NO: 3, кодирующую белок с SEQ ID NO: 4 у линии Е. coli Ev4.
Кроме того, была выявлена мутация в гене argP, которая изменяла последовательность гена argP с SEQ ID NO: 45, кодирующей белок с SEQ ID NO: 46 у линии Е. coli Ev3, на SEQ ID NO: 47, кодирующую белок с SEQ ID NO: 48 у линии Е. coli Ev4.
Кроме того, была выявлена мутация промоторе гена gcvA, которая изменяла последовательность промотора гена gcvA с SEQ ID NO: 55 у линии Е. coli Ev3 на SEQ ID NO: 56 у линии Е. coli Ev4. У независимого штамма, у которого также наблюдали повышенный выход аланина, была выявлена другая мутация, изменявшая последовательность промотора гена gcvA с SEQ ID NO: 55 на SEQ ID NO: 57.
Кроме того, была выявлена мутация в промоторе гена gcvB, которая изменяла последовательность промотора гена gcvB с SEQ ID NO: 59 у линии Е. coli Ex1 на SEQ ID NO: 60 у линии Е. coli Ev1. У другого независимого штамма, у которого наблюдали увеличенный выход аланина, была выявлена другая мутация в промоторе гена gcvB, которая изменяла последовательность промотора с SEQ ID NO: 59 на SEQ ID NO: 61.
Кроме того, была выявлена мутация в гене lpxD, которая изменяла последовательность гена lpxD с SEQ ID NO: 49, кодирующей белок с SEQ ID NO: 50 у линии Е. coli Ev3, на SEQ ID NO: 51, кодирующую белок с SEQ ID NO: 52 у линии Е. coli Ev4.
Для определения важности выявленных мутаций для выхода и производительности по аланину, мутации последовательно вносили в штамм Е. Coli, который содержал мутации, описанные в Примере 1, и мутации, описанные в PCT/IB2014/064426, включая мутации в гене ygaW, гене zipA, гене lpd и мутацию в промоторе, который контролирует экспрессию гена alaD, который также описан выше. Эти мутации оценивали в отношении их влияния на производительность по аланину. Уровни экспрессии мутантных генов или генов, находящихся под контролем мутантных промоторных участков, отслеживали при помощи qPCR.
Пример 6
Подтверждение влияния SNP в гене argP (iciA)
ArgP (или iciA) является регулятором транскрипции. Он контролирует гены, вовлеченные в систему транспорта аргинина, и гены, вовлеченные в репликацию ДНК. SNP, приводивший к мутации А96Е в белке ArgP, выявляли у Е. coli QZ23 и оценивали в отношении его влияния на производительность по аланину.
Построение штамма Е. coli QZ48
Кассету argP-cat-sacB с селектируемым маркером устойчивости к хлорамфениколу и обратно-селектируемым маркером sacB (придает чувствительность к сахарозе) амплифицировали с матричного вектора pQZ11 (Genescript) с праймерами argP_1_F и argP_1_R (см. таблицу 1) при помощи высокоточной ДНК-полимеразы Phusion для ПНР с «горячим стартом» (Thermo). ПЦР-продукт представлял собой DpnI (NEB), который расщепляли при 37°С в течение 1 часа для уменьшения фонового сигнала от матрицы плазмиды и экстрагировали в геле, выделяя из 1% агарозного геля при помощи набора для выделения из геля QIAquick Gel Extraction Kit (Qiagen). Кассету с SNP argP (543 п.о.) амплифицировали с геномной ДНК QZ23 с праймерами argP_2_F и argP_2_R (см. таблицу 1) при помощи высокоточной ДНК-полимеразы Phusion для ПЦР с «горячим стартом» (Thermo) и очищали при помощи набора для очистки продуктов ПЦР QIAquick PCR Purification Kit (Qiagen).
Для Red/ET рекомбинации использовали набор Genebridges Red/ET Recombination Kit согласно протоколу производителя. Примерно 200 нг argP-cat-sacB вводили при помощи электропорации в Red/ET-компетентные клетки Е. coli QZ20. Культуры высеивали на агаровые слои питательной среды LB с 10 мкг/мл хлорамфеникола для отбора положительных трансформантов после электропорации. Несколько колоний подвергали скринингу в отношении встраивания маркерной кассеты при помощи ПЦР с геном-специфичными праймерами argP_seq_F и argP_seq_R (см. таблицу 1). Подтвержденный при помощи ПЦР клон использовали для второй Red/ET рекомбинации с кассетой с SNP argP для замены маркерной кассеты cat-sacB. Культуры высеивали на агаровые слои питательной среды LB с 6% сахарозы без NaCl для отбора положительных трансформантов после электропорации. Несколько клонов тестировали с геном-специфичными праймерами argP_seq_F и argP_seq_R (см. таблицу 1) в отношении утраты маркерной кассеты cat-sacB. По меньшей мере один клон, который давал ПЦР-продукт правильного размера, также подтверждали при помощи секвенирования (Genewiz). Перед тем, как штаммы тестировали в биореакторе, термочувствительную рекомбинационную плазмиду pRedET (amp) устраняли из штаммов при 42°С за ночь на слоях среды LB. В штамм Е. coli QZ20 вносили SNP, который приводил к появлению мутации А96Е в ArgP. Полученный штамм был обозначен как QZ48.
Ферментационное испытание Е. coli QZ20 в сравнении с Е. coli QZ48
Штамм QZ48 Е. coli тестировали в отношении его функциональных характеристик в ходе ферментации в лабораторном биореакторе. Клеточный рост и образование аланина отслеживали в сравнении со штаммом QZ20 Е. coli.
Прекультуры выращивали во встряхиваемых колбах со средой LB, 20% объем наполнения, при 37°С и 200 об/мин на протяжении ночи. Ферментацию проводили в 1,5-литровой системе параллельных биореакторов DASGIP (Eppendorf) в 500 мл среды AM 1 (2,6 г/л (NH4)2HPO4, 0,87 г/л NH4H2PO4, 0,15 г/л Kill, 0,37 г/л MgSO4 ⋅ 7 H2O, 15 г/л (NH4)2SO4, 1 мМ бетаина, 1 мл/л исходного раствора металлических микроэлементов). Исходный раствор металлических микроэлементов содержал 1,6 г/л FeCl3 ⋅ 6 H2O; 0,2 г/л CoCl2 ⋅ 6 H2O; 0,1 г/л CuCl2 ⋅ 2 H2O; 0,2 г/л ZnCl2; 0,2 г/л NaMoO4 ⋅ 2 H2O; 0,05 г/л Н3ВО3, 0,1 М HCl. В качестве источника углерода в ферментационной среде использовали 140 г/л глюкозы.
Клетки Е. coli, эквивалентные OD600⋅мл, равной 7, собирали путем центрифугирования и ресуспендировали в 5 мл среды AM 1. # OD600⋅мл = (OD600 неразведенной культуры) × (объем культуры в мл). 5 мл ресуспендированных клеток использовали для засеивания 500 мл ферментационной среды в 1,5-л биореакторе DASGIP. Каждый штамм проходил два цикла при 37°С и скорости мешалки 400 об/мин. 5 Н NH4OH использовали для контроля рН до 6,8 и снабжения культуры аммонием в качестве предшественника аланина в течение всей ферментации. В ходе ферментации барботирование воздухом не производили и не увеличивали давление в сосуде, так что после изначального потребления растворенного в среде кислорода клетками ферментацию проводили в микроаэробных условиях. На протяжении всей ферментации забирали образцы и анализировали с помощью ВЭЖХ в отношении концентраций аланина и глюкозы.
Мутация А96Е в ArgP у QZ48 оказывала сильное влияние на образование аланина (фиг. 7). Волюметрическая производительность (объемная производительность) по аланину, определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у QZ20 спустя 46 часов составляла 1,15±0,06 г/(л.ч). У Е. coli QZ48 наблюдали увеличенную волюметрическую производительность по аланину, составлявшую 1,51±0,01 г/(л.ч) спустя 46 часов (фиг. 8).
Построение плазмиды pACYC184-argP
Для тестирования влияния сверхэкспрессии argP строили плазмиду pACYC184-argP (p15 ori, CmR, ~15 копий на клетку) при помощи коммерческой технологии клонирования InFusion (Clontech, Маунтин-Вью, Калифорния, США). Сначала получали вектор pACYC184 (таблица 1) при помощи NEB (Ипсуич, Массачусетс, США) и линеаризировали при помощи рестрикционных эндонуклеаз HindIII и SaiI, также из NEB. Такое расщепление удаляло большую часть гена устойчивости к тетрациклину. Отдельно амплифицировали ORF argP при помощи ПЦР с ДНК Е. coli W дикого типа с применением полимеразы Phusion (Thermo Scientific, Уолтем, Массачусетс) с праймерами argP-pACYC_F и argP-pACYC_R (таблица 1).
Праймеры содержали дополнительные выступающие части длиной 15 п.о., гомологичные концам линеаризированного вектора для облегчения неразрывного клонирования. Затем проводили реакцию InFusion согласно протоколу производителя как с очищенной основой линеаризированного вектора, так и со вставкой argP. Полученные в результате продукты InFusion затем использовали для трансформации QZ20 путем электропорации и отбора на слоях среды LB с хлорамфениколом. Положительные клоны выявляли с помощью ПЦР, подтверждали путем секвенирования ДНК и использовали в ферментациях для исследований сверхэкспрессии.
Сравнение ферментаций между QZ20/pACYC184 и QZ20/pACYC184-argP
Прекультуры выращивали во встряхиваемых колбах со средой LB, 20% объем наполнения, при 37°С и 200 об/мин на протяжении ночи. Ферментацию проводили в 1,5-литровой системе параллельных биореакторов DASGIP с 14% глюкозы в среде AM 1. Все условия ферментации были такими, как описано выше.
Сверхэкспрессия argP приводила к повышенной скорости образования аланина и более высокому титру аланина спустя 20 часов ферментации (фигура 9). Волюметрическая производительность (объемная производительность) по аланину, определяемая как количество образовавшегося продукта, деленное на объем реактора и на время, у QZ20/pACYC-argP спустя 20 часов составляла 0,69±0,04 г/(л.ч) в сравнении с 0,61±0,02 г/(л.ч) штамма с контрольной плазмидой pACYC184 (фиг. 10).
Пример 7
Подтверждение влияния SNP в промоторном участке gcvA / gcvB
Построение штаммов QZ58 и QZ66
Кассету gcvA-cat-sacB амплифицировали с вектора pQZ11 (Genescript) с праймерами gcvA_1_F/R (таблица 1). Кассету для SNP в gcvA/B (320 п.о.) амплифицировали с геномной ДНК штамма QZ23 с праймерами gcvA_2_F/R (таблица 1). Red/ET проводили, как описано ранее. Клоны тестировали с помощью ПЦР колоний с праймерами для секвенирования gcvA_seq_F/R. SNP в промоторном участке gcvA/B вносили в Е. coli QZ20, а полученный в результате штамм обозначали как QZ58. SNP также вносили в QZ48 (SNP argP), а полученный в результате штамм обозначали как QZ66.
Ферментационное испытание QZ58 и QZ66
Штамм QZ58 (SNP в промоторе gcvA/B) тестировали в отношении его функциональных характеристик в ходе ферментации как описано ранее. Образование аланина отслеживали в сравнении со штаммом QZ20.
SNP в промоторе gcvA/B оказывал значимое влияние на образование аланина, что в результате давало более высокую скорость образования аланина и титр аланина, составлявший примерно 76 г/л аланина в сравнении с примерно 54 г/л, вырабатываемыми QZ20 спустя 46 часов (фиг. 11). Волюметрическая производительность по аланину у QZ58 составляла 1,66±0,02 г/(л.ч) в сравнении с 1,15±0,06 г/(л.ч) у QZ20 спустя 46 часов (фиг. 12).
Также добавляли SNP в промоторе gcvA/B вдобавок к SNP в argP у QZ48, а полученный в результате штамм QZ66 тестировали в ходе ферментации с образованием аланина в сравнении с QZ48. Дополнительная мутация в промоторе gcvA/B в дополнение к мутации argP у QZ66 приводила более высокой скорости образования аланина в сравнении с QZ48 и более высокому выходу аланина спустя 46 часов, составлявшему примерно 76 г/л в сравнении с примерно 70 г/л у QZ48 (фиг. 13). Волюметрическая производительность по аланину у QZ58 составляла 1,65±0,08 г/(л.ч) в сравнении с 1,51±0,01 г/(л.ч) у QZ48 спустя 46 часов (фиг. 14).
RT-qPCR анализ уровней транскрипции gcvA и gcvB
Уровни транскрипции gcvA и gcvB определяли с помощью количественной ПЦР с обратной транскрипцией (RT-qPCR). Для одностадийных реакций (RT)-qPCR с обратной транскрипцией с красителем SYBR Green использовали набор iTaq Universal One-Step Kit от компании Biorad. Из параллельной периодической ферментации Е. coli QZ20 и Е. coli QZ23, которую проводили, как описано ранее, забирали образцы культуры через 8 ч, 11 ч и 28 ч. Образцы немедленно обрабатывали реактивом RNAprotect Bacteria Reagent (Qiagen) для стабилизации РНК. РНК экстрагировали из образцов с помощью набора AurumTotal RNA Mini Kit (Biorad) согласно руководству производителя. Выделенную РНК дополнительно обрабатывали при помощи набора DNA-free DNA Removal Kit (lifetechnologies) для удаления загрязняющей геномной ДНК и уменьшения фонового сигнала в ходе qPCR. РНК количественно оценивали с помощью спектрофотометрического способа при λ=260 нм.
7-стадийную серию 10-кратных разведений 100 нг РНК Е. coli QZ20 тестировали с помощью праймеров для RT-qPCR (таблица 1) gcvA_RT_F/R к гену gcvA, gcvB_RT_F/R к регуляторной РНК gcvB и rrsA_RT_F и rrsA_RT_R, специфичных к гену rrsA, который кодирует рибосомальную 16 S РНК, который служил в качестве стандартного гена в ходе qPCR испытаний. Подходящий линейный динамический диапазон разведений РНК, который давал в результате показатели эффективности усиления сигнала 90%<Е<110% и фактор линейной регрессии R2>0,985, определяли для каждого набора праймеров для RT-qPCR. Тестировали rrsA в отношении его пригодности в качестве гена внутреннего стандарта для нормализации, и было обнаружено, что он стабильно экспрессировался у всех тестируемых образцов (данные не показаны). Реакции RT-qPCR осуществляли с помощью системы CFX96 Touch Real-Time PCR Detection System (Biorad) согласно протоколу производителя. Относительную количественную оценку экспрессии гена рассчитывали с помощью 8 часовой РНК Е. coli QZ20 в качестве внутреннего калибровочного стандарта согласно способу ΔΔCt (Livak and Schmittgen 2001).
Результаты qPCR подтверждали сверхэкспрессию gcvA у QZ23 в сравнении с QZ20 в ходе экспоненциальной фазы спустя 8 часов и 11 часов ферментации. Даунрегуляцию gcvA наблюдали у 28-часовых образцов во время снижения показателей плотности клеток (фиг. 15А). Регуляторный белок gcvB подвергался даунрегуляции у QZ23 в сравнении с QZ20 в ходе экспоненциальной фазы спустя 8 часов и 11 часов ферментации. Общую даунрегуляцию транскрипции gcvB наблюдали у 28-часовых образцов во время снижения показателей плотности клеток (фиг. 15В).
Построение плазмиды pACYC184-gcvA и pACYC184-gcvB
Поскольку SNP в промоторе gcvA/B приводил к сверхэкспрессии gcvA, то было необходимо подтвердить, что действительно имела место сверхэкспрессия gcvA, которая приводила к увеличенной производительности по аланину. Поэтому осуществляли построение плазмиды pACYC184-gcvA с помощью коммерческой технологии клонирования InFusion (Clontech, Маунтин-Вью, Калифорния, США). Сначала получали вектор pACYC184 (таблица 1) при помощи NEB (Ипсуич, Массачусетс, США) и линеаризировали при помощи рестрикционных эндонуклеаз HindIII и SalI, также из NEB. Такое расщепление удаляло большую часть гена устойчивости к тетрациклину. Отдельно амплифицировали ORF gcvA при помощи ПЦР с ДНК Е. coli W дикого типа с применением полимеразы Phusion (Thermo Scientific, Уолтем, Массачусетс) с праймерами gcvA-pACYC_F и gcvA-pACYC_R (таблица 1). Аналогично, для тестирования влияния сверхэкспрессии gcvB осуществляли построение плазмиды pACYC184-gcvB. Единицу транскрипции gcvB подвергали ПЦР-амплификации с праймерами gcvB-pACYC_F и gcvB-pACYC_R (таблица 1).
Праймеры содержали дополнительные выступающие части длиной 15 п.о., гомологичные концам линеаризированного вектора для облегчения неразрывного клонирования. Затем проводили реакцию InFusion согласно протоколу производителя как с очищенной основой линеаризированного вектора, так и со вставкой соответственно gcvA и gcvB. Полученные в результате продукты InFusion затем использовали для трансформации QZ20 путем электропорации и отбора на слоях среды LB с хлорамфениколом. Положительные клоны выявляли с помощью ПЦР, подтверждали путем секвенирования ДНК и использовали в ферментациях для исследований сверхэкспрессии.
Сравнение ферментации между QZ20/pACYC184, QZ20/pACYC184-gcvA и QZ20/pACYC-gcvB
Прекультуры выращивали во встряхиваемых колбах со средой LB, 20% объем наполнения, при 37°С и 200 об/мин на протяжении ночи. Ферментацию проводили в 1,5-литровой системе параллельных биореакторов DASGIP с 14% глюкозы в среде AM 1. Все условия ферментации были такими, как описано выше.
Результаты ферментационного испытания подтверждали, что сверхэкспрессия gcvA с плазмиды pACYC184-gcvA приводила в результате к более высокой скорости образования и титру аланина в сравнении с пустой контрольной плазмидой (фиг. 16). В отличие от этого, сверхэкспрессия малой регуляторной РНК gcvB с плазмиды pACYC184-gcvB приводила к значимому снижению скорости образования и титра аланина. У Е. coli QZ20/pACYC184-gcvA наблюдали волюметрическую производительность по аланину, составлявшую 1,50±0,09 г/(л.ч) в сравнении с 1,18±0,08 г/(л.ч) у QZ20 с контрольной плазмидой. У Е. coli QZ20/pACYC184-gcvB наблюдали уменьшенную волюметрическую производительность по аланину, составлявшую 0,89±0,01 г/(л.ч), в сравнении с контрольной плазмидой (фиг. 17).
Построение QZ71 с нокаутом по gcvB QZ20
Поскольку сверхэкспрессия регуляторной РНК gcvB с плазмиды pACYC184-gcvB приводила к значимому уменьшению производительности по аланину, то нокаутировали gcvB у QZ20 и тестировали в отношении его функциональных характеристик. Кассету gcvB-cat-sacB амплифицировали с вектора pQZ11 (Genescript) с праймерами gcvB_1_F/R (таблица 1). Делеционную кассету для gcvB (400 п.о.) заказывали в виде dsDNA gBlock у IDT (SEQ ID NO: 98). Red/ET проводили, как описано ранее. Клоны тестировали с помощью ПЦР колоний с праймерами для секвенирования gcvB_seq_F/R. Делецию по gcvB вносили в Е. coli QZ20, а полученный в результате штамм обозначали как QZ71.
Ферментационное испытание с QZ71
Штамм QZ71 (нокаут по gcvB) тестировали в отношении его функциональных характеристик в ходе ферментации как описано ранее. Образование аланина отслеживали в сравнении со штаммом QZ20.
Удаление регуляторной РНК gcvB из QZ20 приводило в результате к небольшому увеличению титра аланина в сравнении с QZ20 (фиг 18). Волюметрическая производительность по аланину у QZ71 составляла 1,28±0,05 г/(л.ч) в сравнении с 1,15±0,06 г/(л.ч) у QZ20 спустя 46 часов (фиг. 19).
Пример 8
Подтверждение влияния делеции в гене brnQ (Δ667-764)
BrnQ является предполагаемым 439 АА транспортером для аминокислот с разветвленной боковой нитью, который транспортирует лейцин, валин и изолейцин в клетку в качестве симпортера натрий/аминокислота с разветвленной боковой нитью. У QZ23 была выявлена делеция 97 п.о. (Δ667-764), которая вызывала сдвиг рамки считывания. В то время как первые 222 аминокислоты у 439 АА белка оставались неизмененными, 31 АА изменялись в результате сдвига рамки считывания, а остаточная С-концевая нить была укороченной в связи с образовавшимся стоп-кодоном. Поскольку было сделано предположение, что частичная делеция 97 п.о., обнаруженная в гене brnQ у QZ23, приводила к исчезновению активности BrnQ, проводили тестирование делеции всего гена brnQ (нокаут) в дополнение к частичной делеции brnQ.
Построение штаммов QZ57 и QZ69
Кассету brnQ-cat-sacB амплифицировали с вектора pQZ11 (Genescript) с праймерами brnQ_1_F/R (таблица 1). Кассету для частичной делеции brnQ (462 п.о.) амплифицировали с геномной ДНК штамма QZ23 с праймерами brnQ_2_F/R (таблица 1). Кассету для KO по brnQ (500 п.о.) заказывали в виде dsDNA gBlock у IDT (SEQ ID NO: 117). Red/ET проводили, как описано ранее. Клоны тестировали с помощью ПЦР колоний с праймерами для секвенирования brnQ_seq_F/R. Частичную делецию по brnQ вносили в Е. coli QZ20, а полученный в результате штамм обозначали как QZ57. Полную делецию brnQ вносили в Е. coli QZ20, а полученный в результате штамм обозначали как QZ69.
Ферментационное испытание QZ57 и QZ69
Штаммы QZ57 (brnQ Δ667-764) и QZ69 (brnQ KO) тестировали в отношении их функциональных характеристик в ходе ферментации как описано выше. Образование аланина отслеживали в сравнении со штаммом QZ20.
Делеция 97 п.о. brnQ у QZ57 и полный нокаут brnQ имели сравнимые функциональные характеристики. Как первая, так и второй давали в результате более высокое образование аланина и более высокий титр аланина, чем у QZ20 (фиг. 20). Волюметрическая производительность по аланину у QZ57 составляла 1,30±0,04 г/(л.ч), а производительность у QZ69 составляла 1,28 г/(л.ч) в сравнении с 1,15±0,06 г/(л.ч) у QZ20 спустя 46 часов (фиг. 21).
Пример 9
Подтверждение влияния SNP в гене lpxD
У QZ23 был детектирован SNP в гене lpxD, который приводил к мутации А15Т в кодируемом ферменте. UDP-3-O-(3-гидроксимиристоил)-глюкозамин-N-ацетилтрансфераза, кодируемая LpxD, является важным ферментом, принимающим участие в биосинтезе липида А. Липид А является неотъемлемой частью липополисахарида (LPS) внешней мембраны Е. coli.
Построение штаммов QZ56 и QZ70
Кассету lpxD-cat-sacB амплифицировали с вектора pQZ11 (Genescript) с праймерами lpxD_1C_F/R (таблица 1). Кассету для SNP в lpxD (2588 п.о.) амплифицировали с геномной ДНК штамма QZ23 с праймерами lpxD_fix_F/R (таблица 1). Red/ET проводили, как описано ранее. Клоны тестировали с помощью ПЦР колоний с праймерами для секвенирования lpxD_flank_F/R. SNP в lpxD вносили в Е. coli QZ20, а полученный в результате штамм обозначали как QZ56. SNP в lpxD также вносили в QZ68 (SNP в argP, SNP в промоторе gcvA/B, brnQ Δ667-764), а полученный в результате штамм обозначали как QZ70.
Ферментационное испытание QZ56 и QZ70
Штамм QZ56 (SNP в lpxD) тестировали в отношении его функциональных характеристик в ходе ферментации как описано ранее. Образование аланина отслеживали в сравнении со штаммом QZ20. Мутация А15Т в LpxD у QZ56 давала в результате увеличенный титр аланина в сравнении с QZ20 (фиг. 22). Волюметрическая производительность по аланину у QZ56 составляла 1,34±0,06 г/(л.ч) в сравнении с 1,15±0,06 г/(л.ч) у QZ20 спустя 46 часов (фиг. 23).
SNP в lpxD также вносили в QZ68 (SNP в argP, SNP в промоторе gcvA/B, brnQ Δ667-764), а полученный в результате штамм QZ70 тестировали в ходе ферментации с образованием аланина в сравнении с QZ68. Мутация А15Т в LpxD у QZ48 оказывала сильное влияние на образование аланина (фиг. 7). Скорость образования аланина у QZ68 и QZ70 была сравнимой, однако титр аланина у QZ68 переставал расти на уровне около 75 г/л, в то время как у QZ70 образование аланина продолжалось до тех пор, пока вся глюкоза в среде не была потреблена, а титр аланина, который составлял 102 г/л, достигался через примерно 37 часов (фиг. 24). Волюметрическая производительность по аланину у QZ70 составляла 2,24±0,002 г/(л.ч) в сравнении с 1,64±0,03 г/(л.ч) у QZ68 спустя 46 часов (фиг. 25).

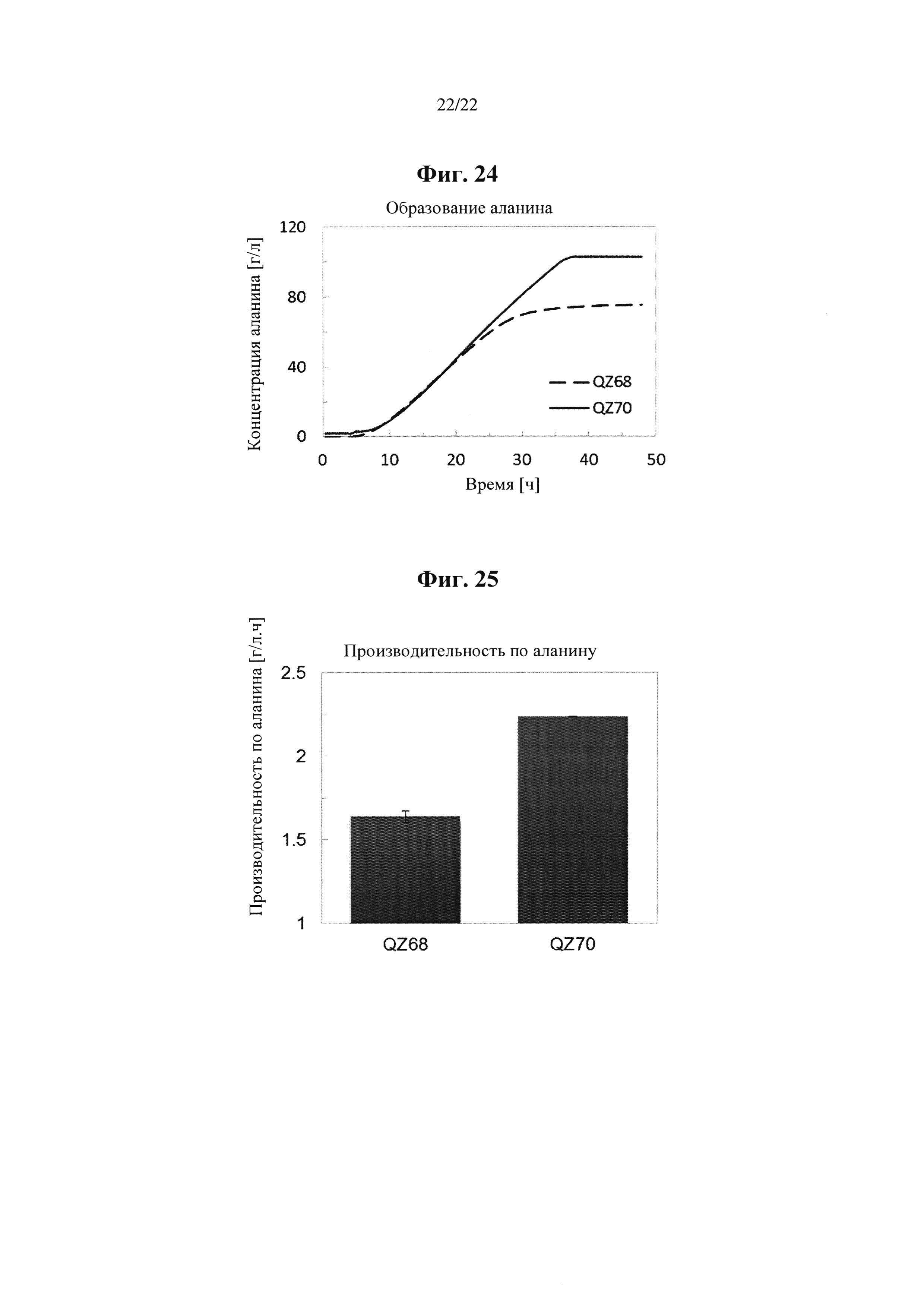

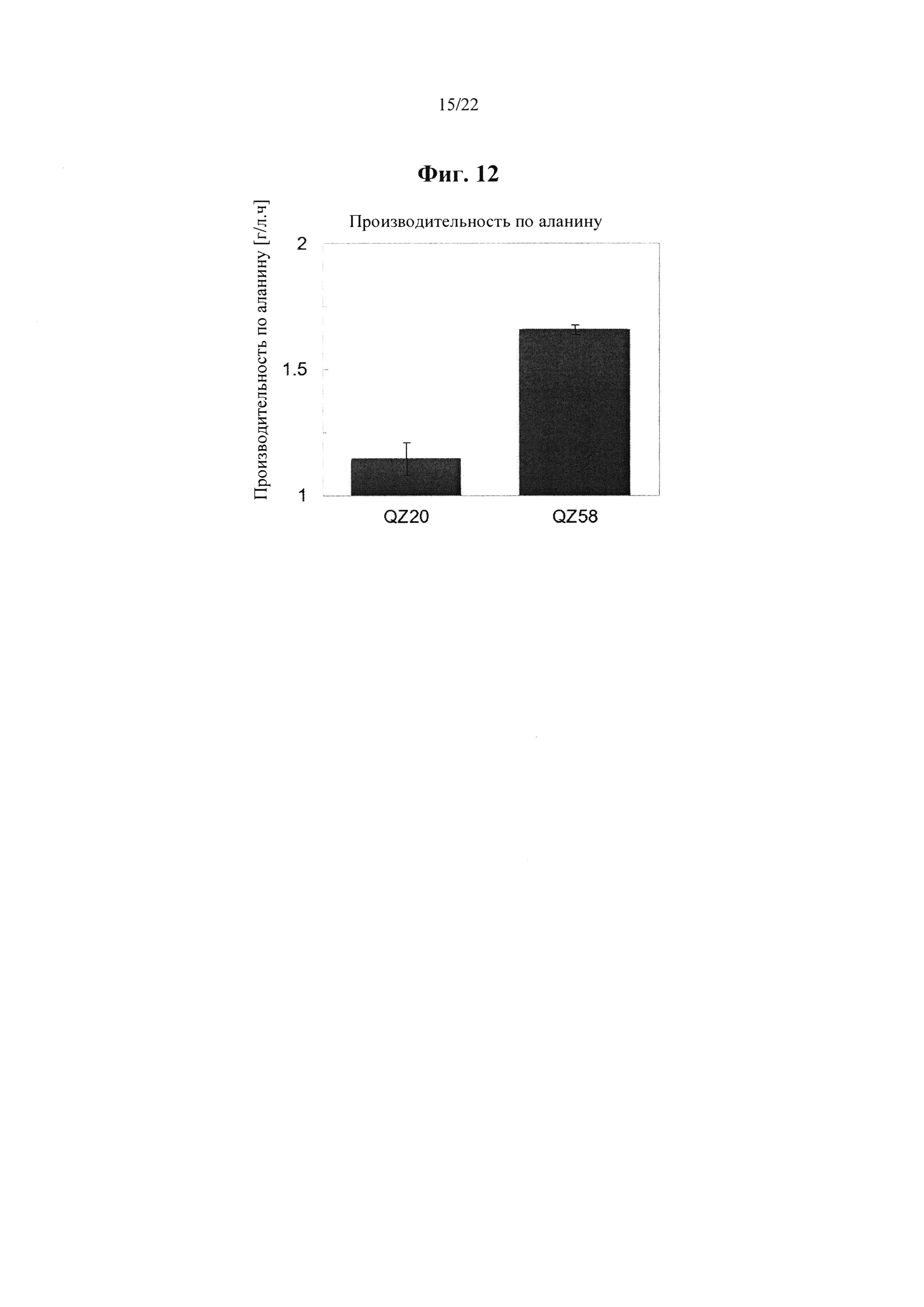
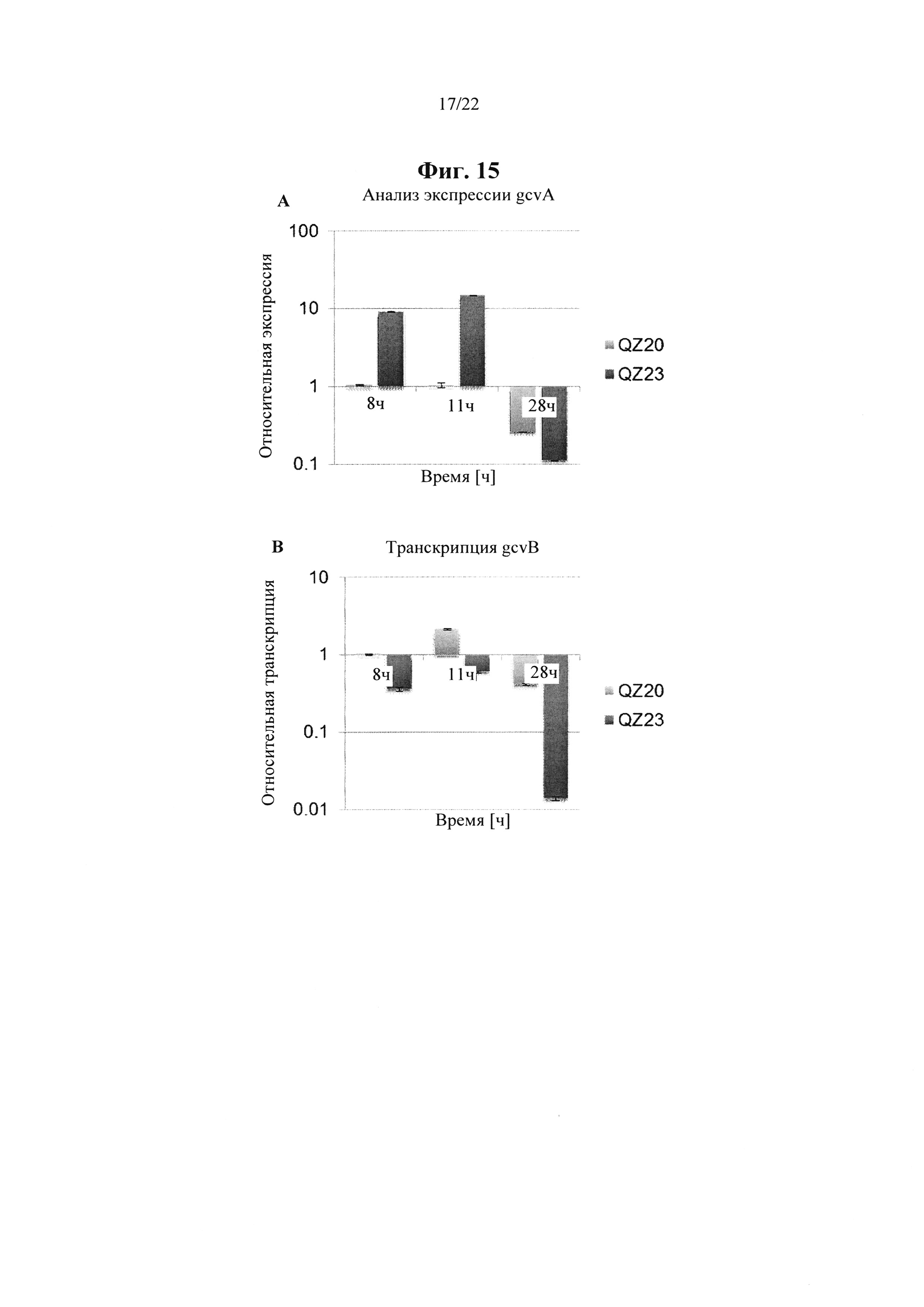


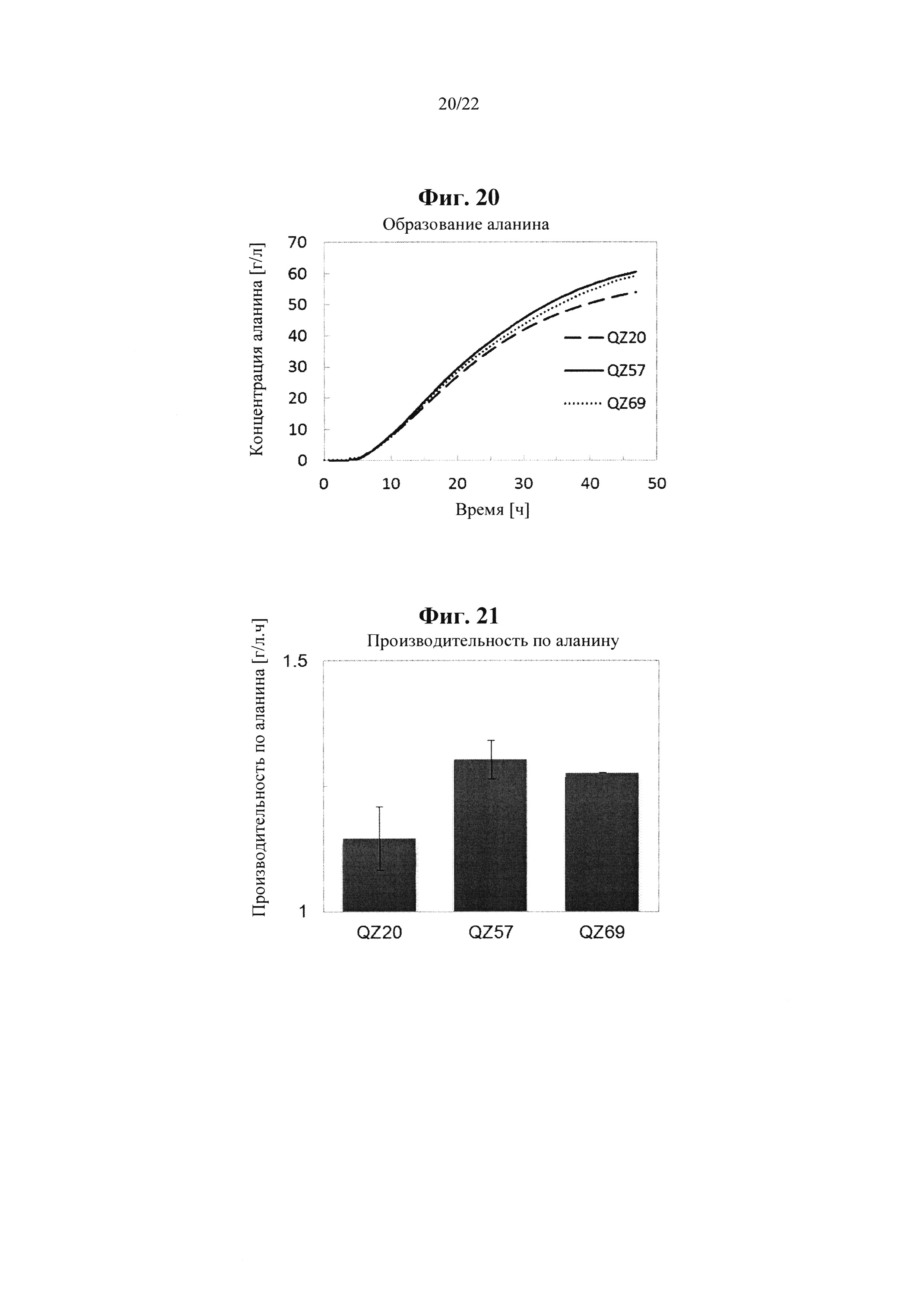


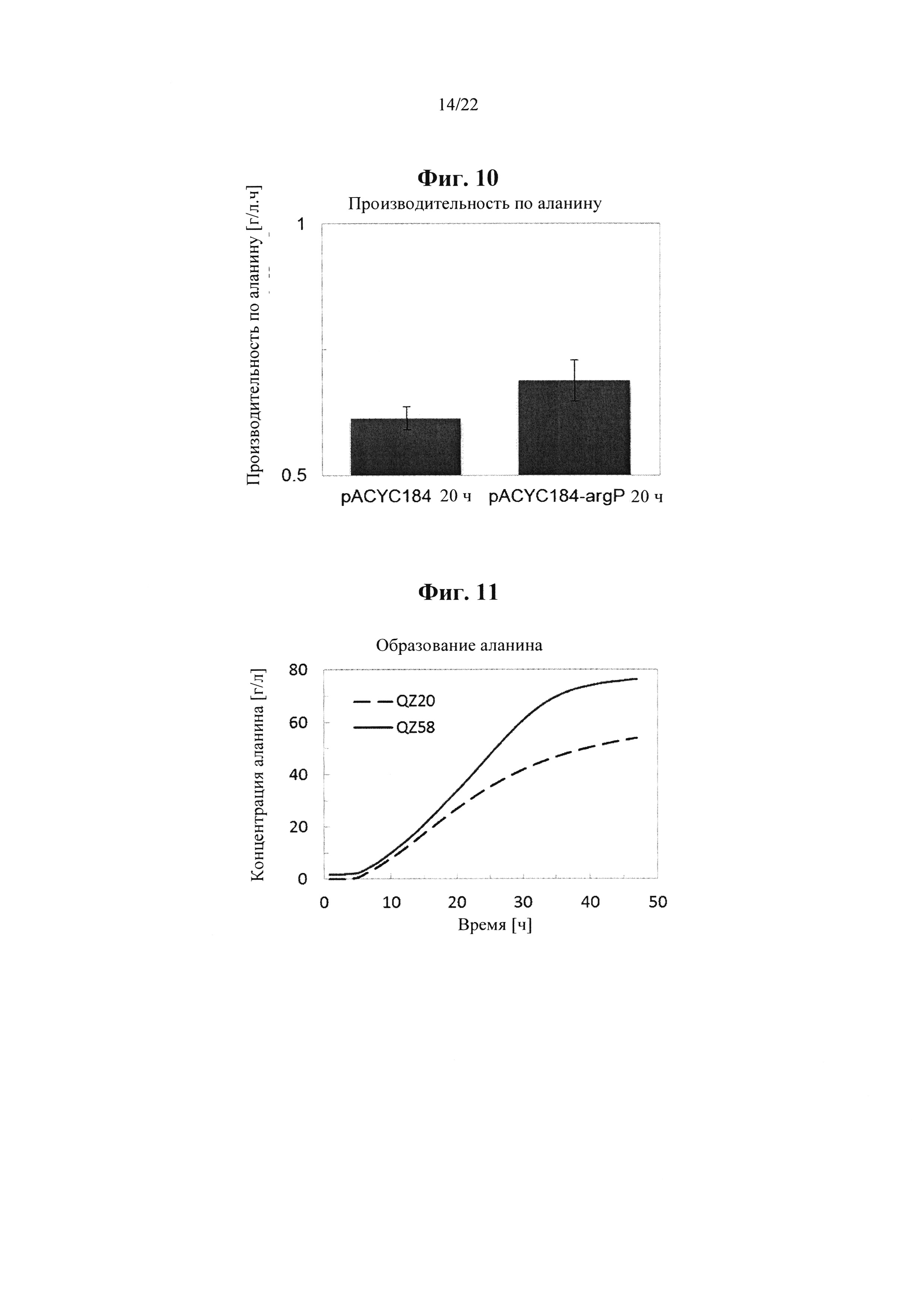











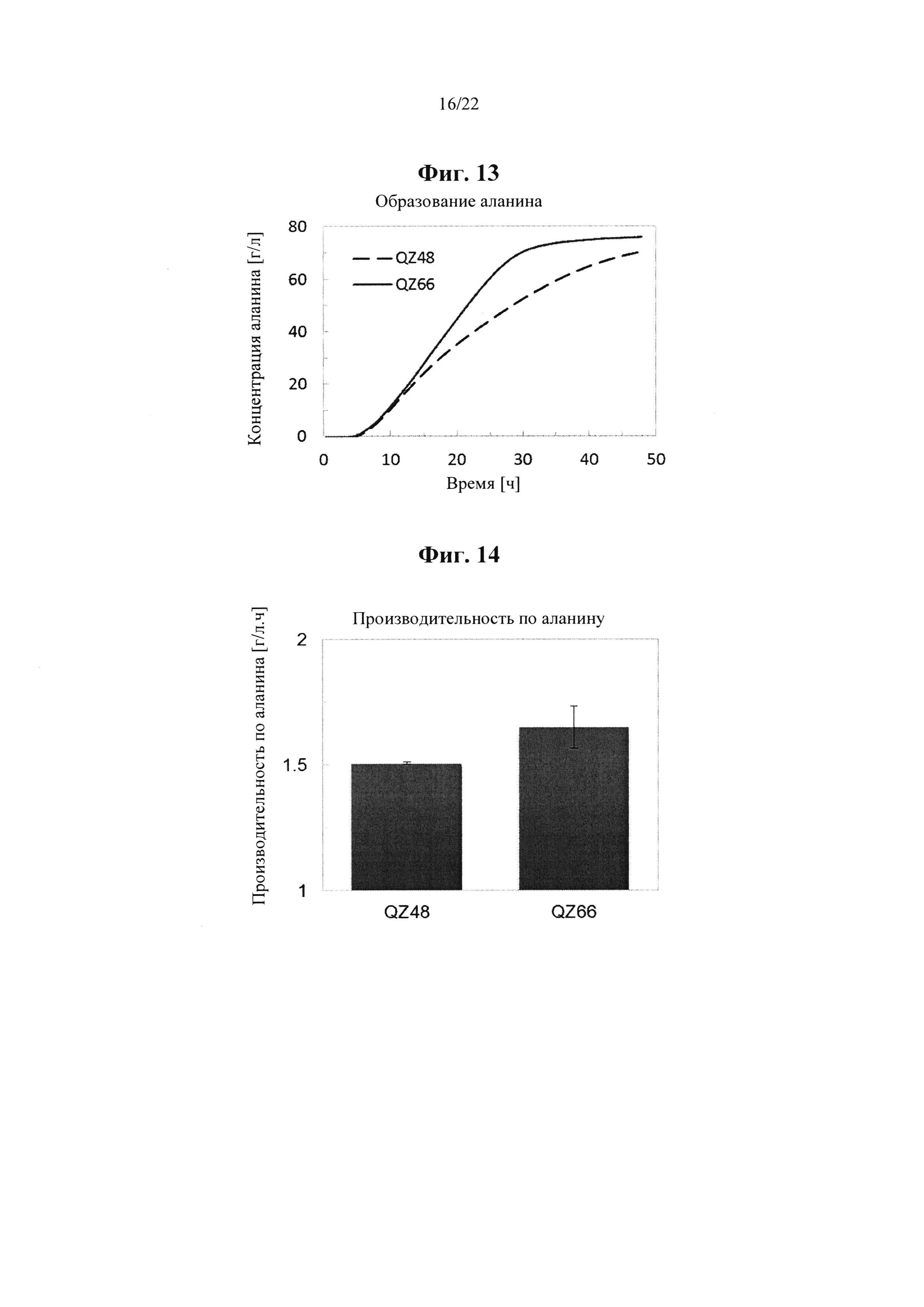










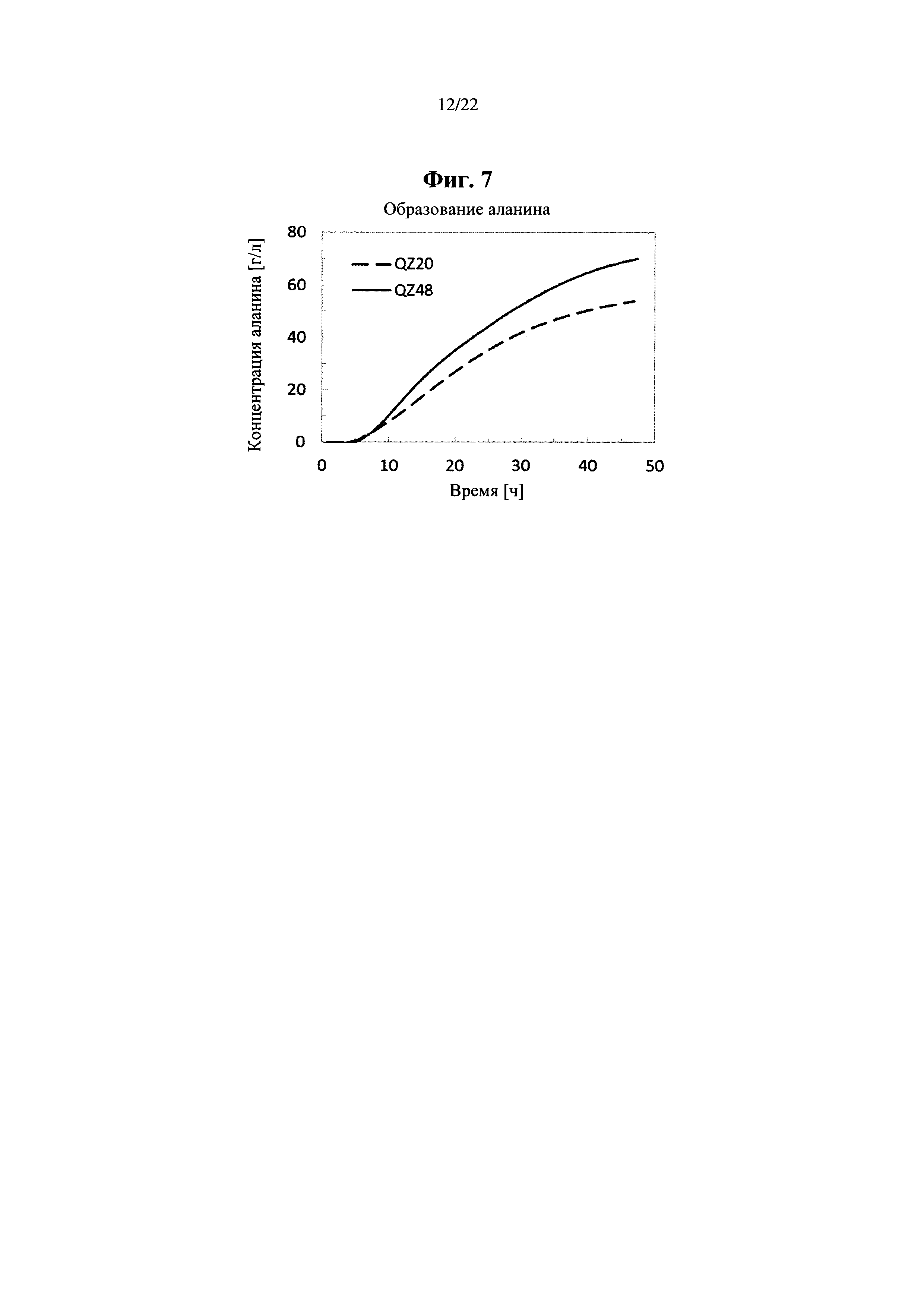
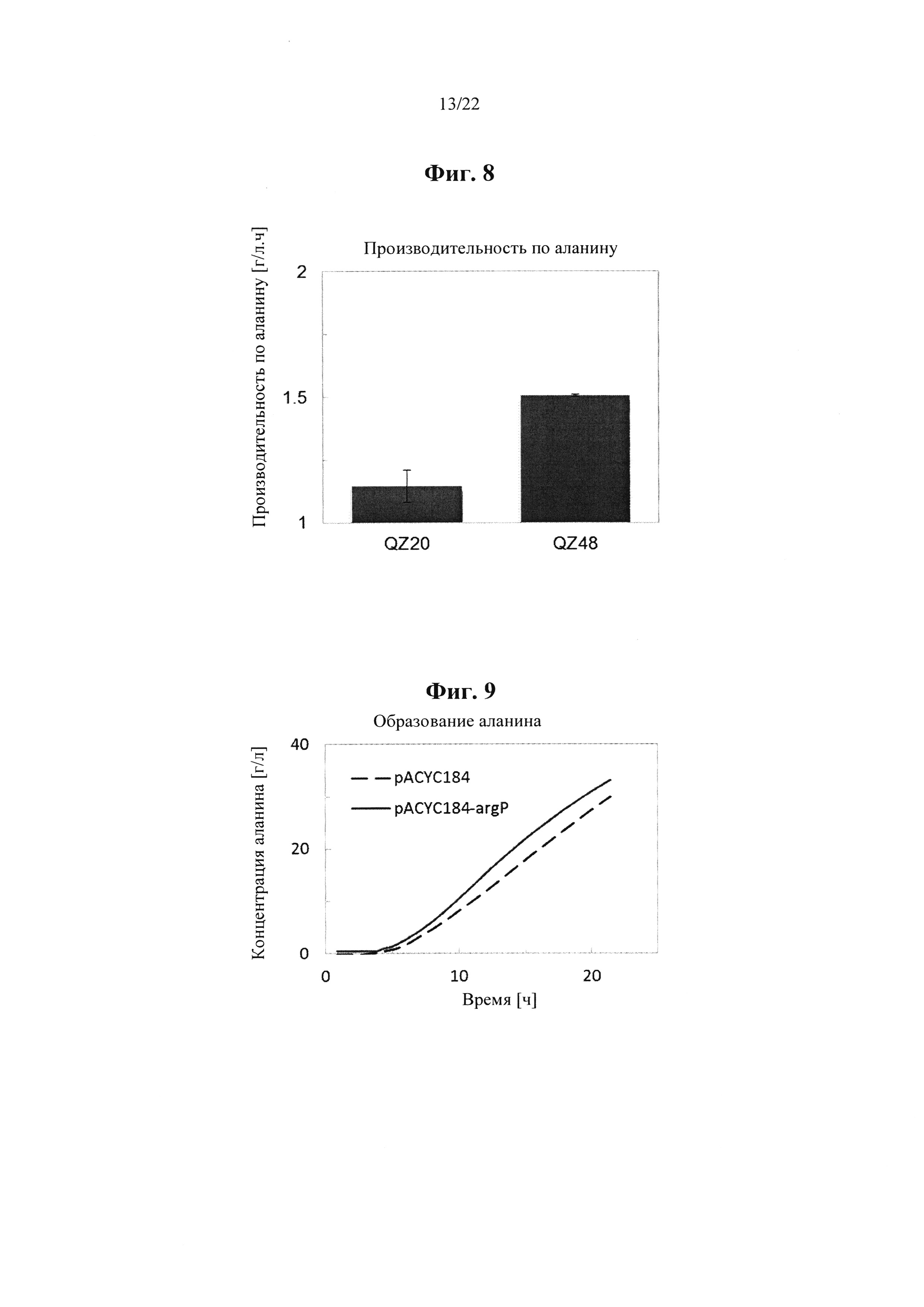
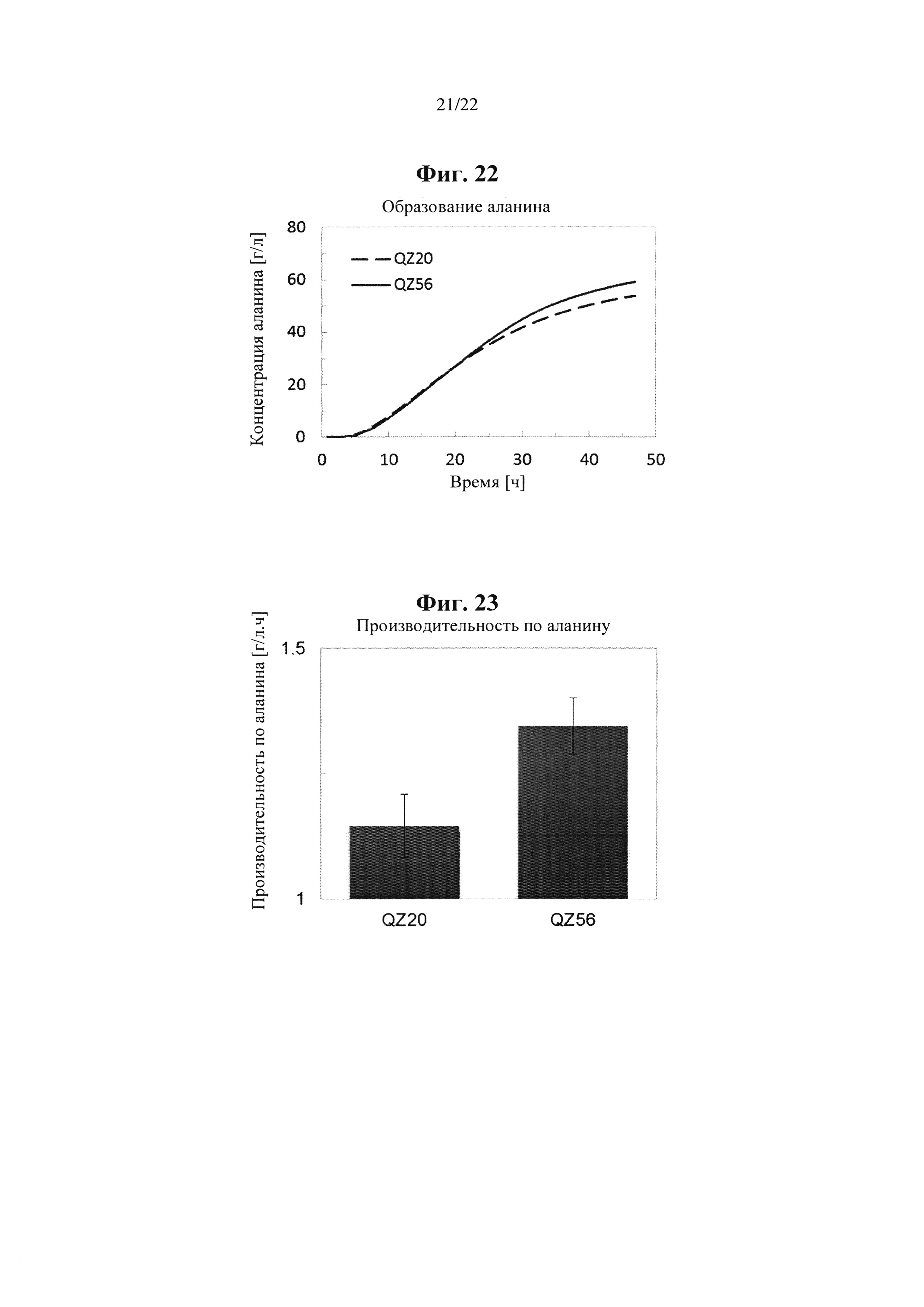

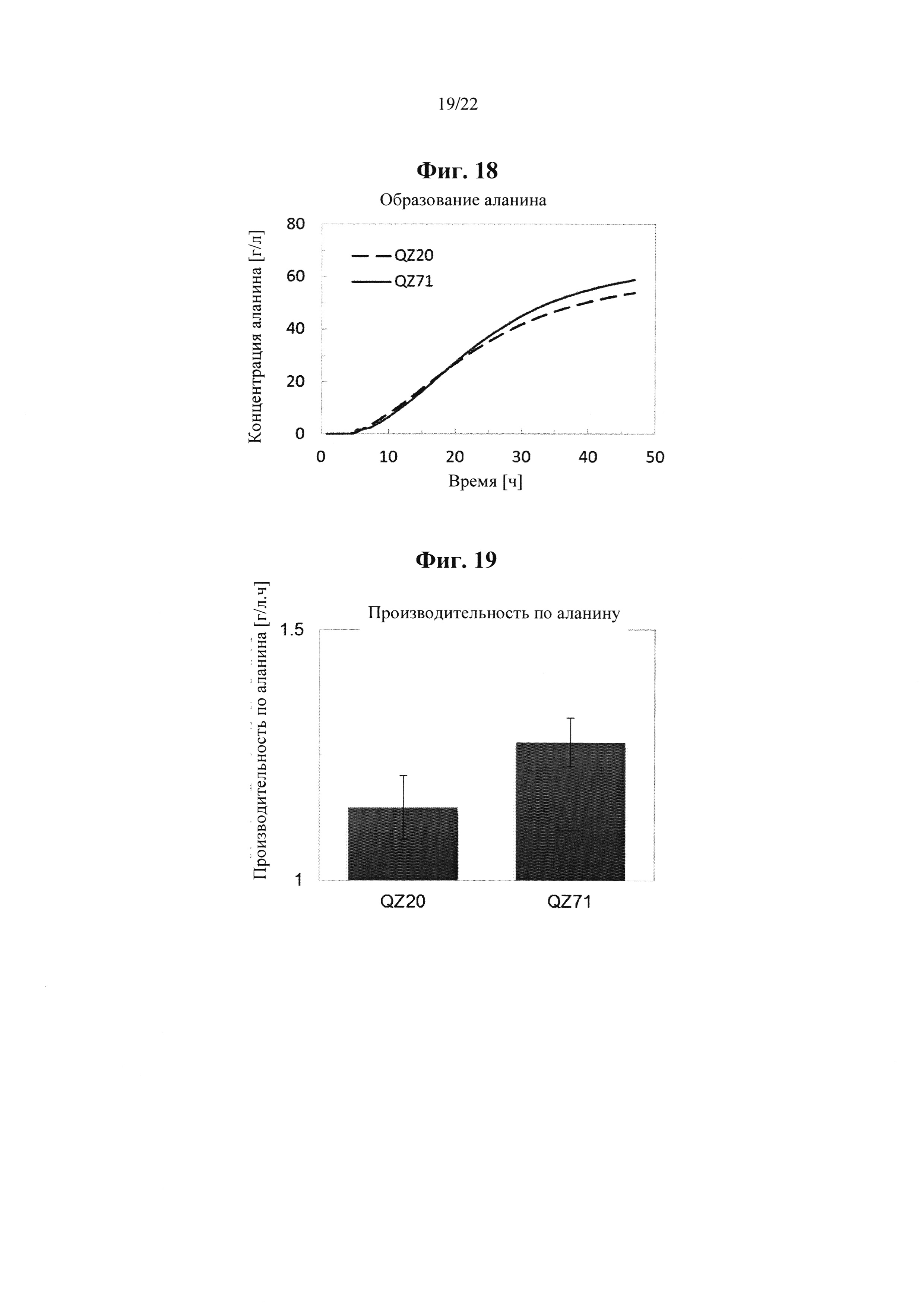






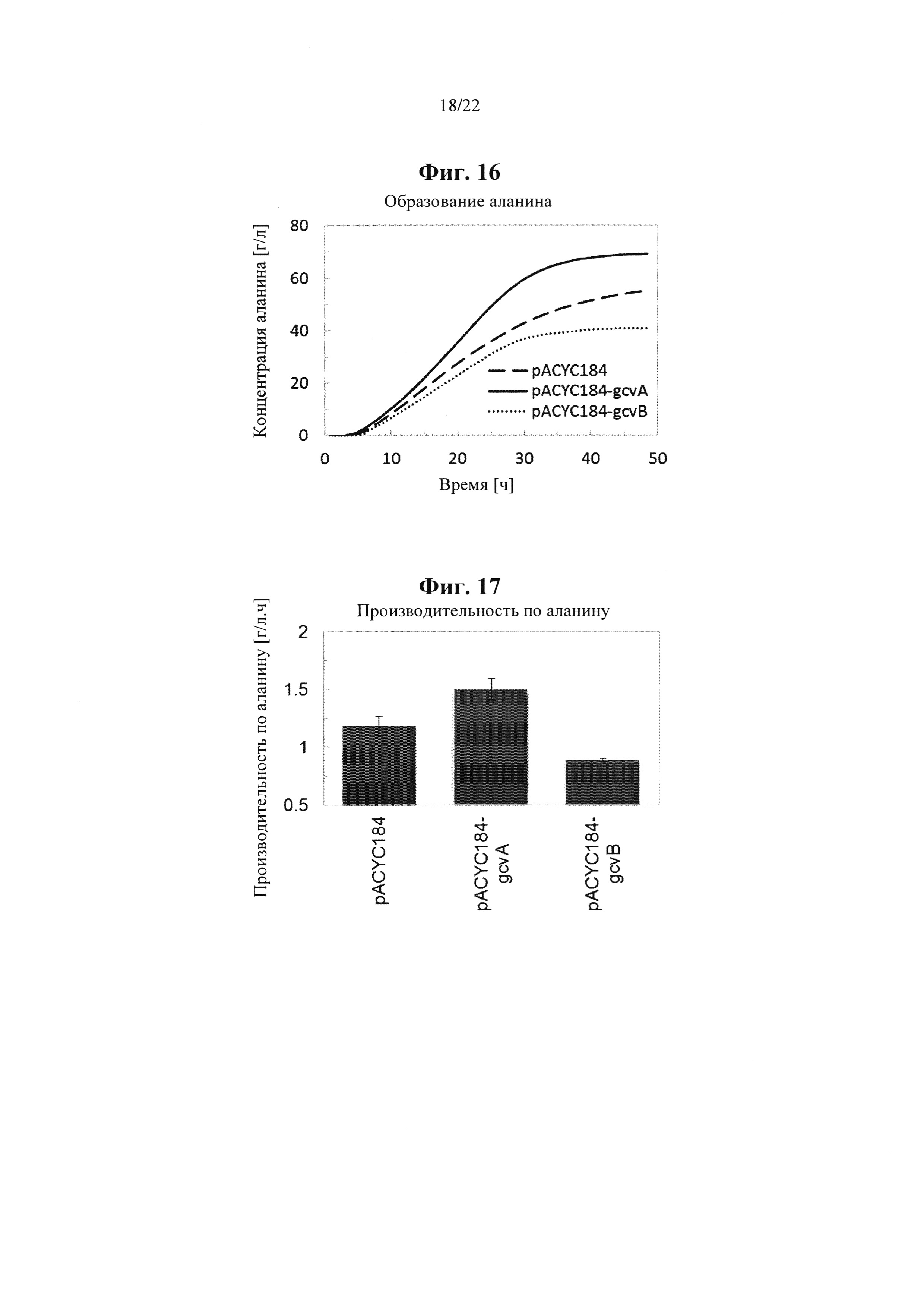
Способ хранения жидкой в условиях хранения мономерной фазы
Способ транспортировки жидкой мономерной фазы, извлеченной из резервуара для хранения, в резервуаре автозаправщика или танкера
Способ получения механически стабильных водопоглощающих полимерных частиц
Способ получения акриловой кислоты
Способ получения триэтилентетрамина (тэта) через этилендиаминдиацетонитрил (эддн)
Способ получения смеси этиленаминов
Средство для нанесения покрытий на вспенивающиеся частицы стирольного полимеризата
Способ для обнаружения и подсчета жизнеспособных микроорганизмов вида legionella pneumophila и набор для его осуществления
Способ получения смесей этиленаминов
Эластичный пеноматериал из частиц на основе смесей полиолефина/полимера стирола
Гены corynebacterium glutamicum, кодирующие белки, участвующие в метаболизме углерода и продуцировании энергии
Гены corynebacterium glutamicum, кодирующие белки метаболических путей
Гены corynebacterium glutamicum, кодирующие белки, участвующие в метаболизме углерода и продуцировании энергии
Рекомбинантный микроорганизм для улучшенного получения химических веществ тонкого органического синтеза
Композиции для улучшения гигиены полости рта

