Результат интеллектуальной деятельности: Рекомбинантная плазмидная ДНК pQE-30_P36GP12_GP57, обеспечивающая синтез рекомбинантного белка P36GP12 в клетках Escherichia coli, штамм бактерий Escherichia coli - продуцент рекомбинантного белка P36GP12, рекомбинантный белок P36GP12, обладающий способностью связывать липополисахариды Escherichia coli
Вид РИД
Изобретение
Группа изобретений относится к биотехнологии, генной и белковой инженерии и предназначена для получения рекомбинантного белка P36GP12 на основе белка хвостовых нитей Т4-подобного бактериофага, генетической конструкции, содержащей ген, кодирующий рекомбинантный белок P36GP12 и ген, кодирующий белок-шаперон этого белка, и обеспечивающей синтез рекомбинантного белка P36GP12 в клетках Escherichia coli и штамма бактерий Escherichia coli - продуцента рекомбинантного белка P36GP12.
Рекомбинантный белок P36GP12 может быть использован для удаления бактериальных липополисахаридов (ЛПС), в том числе эндотоксинов, при производстве биотехнологических препаратов, а также для селективного удаления бактерий и их фрагментов из жидкостей, предназначенных для инъекций, инфузий и других типов введения в том случае, когда фильтрация не позволяет добиться результата удаления бактерий и их фрагментов.
Частью производства биотехнологических субстанции для инъекционного введения является подготовка специализированных белков (антител, факторов роста, цитокинов, вакцин). В общемировой практике для продукции этих белков используются рекомбинантные штаммы Е. coli в качестве продуцентов, на поверхности которых присутствуют липополисахариды. Липополисахариды являются токсичными и смертельно опасными для организма человека, вызывая при высокой концентрации различные патологии. Поэтому при получении рекомбинантных белков необходимо очищать конечный продукт от эндотоксинов до уровня, ниже требуемого фармакопеей.
Бактериальные эндотоксины или липополисахариды - это обязательный компонент клеточной стенки грамотрицательных микроорганизмов, который содержится в количестве 1 миллион молекул липополисахарида на 1 клетку E. coli [1]. Эндотоксины являются токсичными для организма человека, и поэтому существуют допустимые нормы содержания эндотоксинов в фармацевтических продуктах, которые вводятся в организм человека инъекционно. Например, фармакопея Российской федерации регламентирует максимальный уровень содержания эндотоксинов при применении инъекционных лекарственных средств и биологических продуктов в размере 5 единиц эндотоксина (EU/ЕЭ) на кг массы тела в течение часа при внутривенном введении [2].
Бактериофаги рода Myoviridae, специфичные к бактерии Escherichia coli, в частности фаги рода Т4-подобных, являются одними из самых изученных фагов. Известно, что в геноме таких фагов есть открытые рамки трансляции, которые кодируют белок коротких хвостовых нитей и его белок-шаперон. В нативном вирионе фага белок коротких хвостовых нитей обеспечивает необратимое связывание с бактерией. Связывание возможно из-за сродства белка коротких хвостовых нитей к липополисахаридам бактериальной стенки. Белок-шаперон позволяет гомотримеру белка коротких хвостовых нитей принимать нативную переплетающуюся конформацию, необходимую для осуществления связывания липополисахаридов [3]. Таким образом, короткие хвостовые белки бактериофагов Escherichia coli могут с высоким сродством связываться с липополисахаридами Escherichia coli.
На данный момент существуют разные подходы для очистки от эндотоксинов. При очистке фармацевтических продуктов используют хроматографию, в основном аффинную, а также различные варианты ионообменной хроматографии, гидрофобной хроматографии и гель-фильтрации [1, 4]. Используемые в настоящее время в фармацевтическом производстве протоколы хроматографической очистки являются, как правило, узкоспециализированными, например, в случае использования L-гистидина, в результате чего для каждого белка со своей аминокислотной последовательностью требуется трудоемкая оптимизация протокола очистки [5]. Это же относится и к хроматографическому удалению липополисахаридов [6].
Известны способы очистки белковых препаратов от эндотоксинов с использованием ионообменной хроматографии, аффинных сорбентов с иммобилизованными L-гистидином, поли-L-лизином, поли-(γ-метил-L-глутаматом) или полимиксином В, гель-фильтрации, ультрафильтрации, центрифугирования в градиенте сахарозы и фазовое разделение тритоном Х-114. Эти способы используются в продуктах таких компаний как «ToxinEraser» (полимиксин В), «Pierce High Capacity Resin» (поли-L-лизин) и «EndoBind-R» (S1 пептид). Однако вышеназванные способы во многих случаях не являются универсальными и обладают серьезными недостатками, например, во время очистки сильно уменьшается выход конечного целевого продукта. Применимость этих способов для очистки белковых субстанций от ЛПС сильно зависит от свойств целевого белка, поэтому для каждого конкретного продукта необходимо подбирать наиболее эффективную методику очистки.
Известен рекомбинантный белок, разработанный на основе хвостового белка изолята бактериофага Т4, который связывается с липополисахаридами Escherichia coli и используется в качестве активного вещества в сорбенте. Очистка с помощью данного белка в составе сорбента дает минимальные потери и высокую степень очистки, однако этот сорбент обладает высокой стоимостью и усложненной логистикой (заявка DE 10228133, опубл. 24.06.2002).
Наиболее ближайшими к заявляемой группе изобретений - прототипом, являются рекомбинантная плазмидная ДНК pT4g57g12, кодирующая белок р12 Т4-подобного фага, штамм бактерий Escherichia coli Е. coli BL21 (DE3) - продуцент рекомбинантного белка р12 и рекомбинантный белок р12 Т4-подобного бактериофага, обладающий способностью связывать липополисахариды Escherichia coli [7].
Недостатками прототипа являются дороговизна и недоступность используемого для получения целевого продукта оборудования и реагентов.
Задачей заявляемого изобретения является получение рекомбинантного белка P36GP12, который обладает способностью связывать липополисахариды Escherichia coli.
Техническим результатом является создание плазмиды pQE-30_P36GP12_GP57, рекомбинантного штамма Escherichia coli, несущего в себе данную конструкцию и рекомбинантного белка P36GP12, обладающего способностью связывать липополисахариды Escherichia coli.
Поставленная задача решается путем конструирования рекомбинантной плазмидной ДНК, которая содержит ген, кодирующий белок-шаперон gp57 и ген, кодирующий белок коротких хвостовых нитей gp12 изолята Т4-подобного бактериофага, обладающего высоким сродством к бактериальным липополисахаридам Escherichia coli.
Поставленная задача решается также путем получения рекомбинантного штамма Escherichia coli, несущего в себе плазмиду pQE-30_P36GP12_GP57, который обеспечивает синтез рекомбинантного белка P36GP12, обладающего способностью связывать липополисахариды Escherichia coli.
Поставленная задача решается также путем получения рекомбинантного белка P36GP12, обладающего способностью связывать липополисахариды Escherichia coli.
Сущность группы изобретений заключается в следующем:
Методами генетической инженерии получают плазмидную ДНК pQE-30_P36GP12_GP57, несущую гены, кодирующие рекомбинантный белок P36GP12, содержащий N-концевой олигопептид MRGSHHHHHHGSAN, а также рекомбинантный белок P36GP57, выполняющий роль шаперона для приобретения рекомбинантным белком P36GP12 нативной конформации.
Последовательность ДНК, кодирующую оба белка, получают на основе геномной ДНК изолята Т4-подобного бактериофага.
Исходным генетическим материалом для конструирования целевых плазмид служат следующие генно-инженерные исходные конструкции:
а) Плазмидная ДНК pQE-30 (QIAGEN, Германия);
б) Геномная фаговая ДНК изолята Р36 Т4-подобного бактериофага, содержащая гены, кодирующие рекомбинантный белок P36GP12 и его белок-шаперон P36GP57.
Олигонуклеотидные последовательности для конструирования плазмидной ДНК, содержащей ген, кодирующий рекомбинантный белок P36GP12 (в направлении 5' - 3'):
1. PH36_gp12_dir:
5' АТС ACG GAT CCG САА АСА АТА СТА ТТА АСС ATG ТАА AAG ACG ATG С 3'
2. PH36_gp12_rev:
5' AGT ТАА ТТТ СТС СТС ТТА ТТА GCG САС ССТ ТАТ GAT ATA GTT ТАА TGC 3'
3. PH36_gp57_dir:
5' GTG CGC ТАА ТАА GAG GAG AAA ТТА ACT ATG ТСС AAT CAG CAT GAA С 3'
4. PH36_gp57_rev:
5' СТА ATT AAG CTT ATT AAG CCT CCG TGA TCA GTT СТА CTT CCT CTT С 3'
Далее получают ДНК-фрагмент, кодирующий рекомбинантный белок P36GP12 и его белок-шаперон с нативной геномной ДНК изолята Р36 Т4-подобного бактериофага следующим образом:
1. С использованием геномной ДНК изолята Р36 Т4-подобного бактериофага в качестве матрицы и олигонуклеотидов PH36_gp12_dir и PH36_gp12_rev, соответствующих началу и концу гена короткого хвостового белка, проводят полимеразную цепную реакцию (ПЦР) для амплификации фрагмента, кодирующего белок коротких хвостовых нитей. Продукты ПЦР разделяют электрофоретически, фрагмент геля с нуклеотидными последовательностями рассчитанной длины вырезают, из фрагмента геля элюируют ДНК-фрагмент, кодирующий белок коротких хвостовых нитей, любым доступным образом, например, с использованием набора GeneJET™ Gel Extraction Kit (Fermentas, Литва). Аналогично с помощью олигонуклеотидов PH36_gp57_dir и PH36_gp57_rev получают ДНК-фрагмент, кодирующий белок-шаперон бактериофага Р36.
2. По матрице полученных ДНК-фрагментов проводят объединяющую ПЦР с использованием специфических олигонуклеотидов PH36_gp12_dir и PH36_gp57_rev. Полученный объединенный ДНК-фрагмент GP12_GP57, кодирующий белок коротких хвостовых нитей и его белок-шаперон, очищают электрофорезом в агарозном геле и элюируют из геля. Для подготовки фрагмента к реакции лигирования объединенный ДНК-фрагмент подвергают гидролизу эндонуклеазами рестрикции BamHI и HindIII по сайтам, по которым будет проходить лигирование объединенного фрагмента и плазмиды pQE-30. Продукты гидролиза также подвергаются очистке с помощью электрофоретического разделения в агарозном геле и последующему элюированию ДНК из геля.
Далее осуществляют сборку плазмидной ДНК pQE-30_P36GP12_GP57, содержащей гены белка коротких хвостовых нитей бактериофага Р36 и его белок-шаперон, следующим образом:
1. Плазмидную ДНК pQE-30 гидролизуют эндонуклеазами рестрикции BamHI и HindIII. Для предотвращения самолигирования проводят дефосфорилирование свободных концов гидролизованной плазмидной ДНК с помощью щелочной фосфатазы (Invitrogen, США). Проводят очистку с помощью агарозного геля и последующую элюцию плазмидной ДНК из геля.
2. ДНК-фрагмент GP12_GP57, гидролизованный эндонуклеазами BamHI и HindIII и очищенный электрофорезом в агарозном геле, а также гидролизованный теми же эндонуклеазами фрагмент плазмидной ДНК pQE-30 с дефосфорилированными "липкими" концами подвергают реакции лигирования с использованием фермента ДНК-лигазы бактериофага Т4. В результате получают плазмидную ДНК pQE-30_P36GP12_GP57.
Полученная рекомбинантная плазмида pQE-30_P36GP12_GP57, кодирующая рекомбинантный белок P36GP12 и его белок-шаперон P36GP57, характеризуется следующими признаками:
- имеет молекулярную массу 3,4 МДа и размер 5132 п.о.;
- кодирует рекомбинантные белки P36GP57 - белок-шаперон и P36GP12 - белок коротких хвостовых нитей бактериофага Р36.
- состоит из следующих элементов:
а) промоторная система транскрипции-трансляции бактериофага Т5, синтетический сайт связывания рибосом RBSII, два терминатора транскрипции: t0 терминатор фага лямбда и Т1 терминатор из rrnB оперона бактерии Е. coli и сайт инициации репликации ColE1;
б) генетические маркеры: AMPr - ген β-лактамазы, обеспечивающий устойчивость клеток Escherichia coli к ампициллину;
в) искусственный ген, кодирующий рекомбинантный белок P36GP12, включающий N-концевой олигопептид MRGSHHHHHHGSAN и белок коротких хвостовых нитей gp12 изолята Р36 Т4-подобного бактериофага в одной рамке трансляции, а также белок-шаперон gp57, содержащий свой сайт связывания рибосом.
г) уникальные сайты узнавания эндонуклеазами рестрикции, имеющими следующие координаты: EcoRI (89), BamHI (146), HpaI (1462), Acc65I (1835), KpnI (1839), HindIII (1859), BglI (4261).
Физическая карта плазмиды представлена на фиг. 1
Трансформация культуры плазмидной ДНК и наработка рекомбинантного белка P36GP12
Клетки E. coli DH5αF' трансформируют сконструированной плазмидой pQE-30_P36GP12_GP57 и культивируют в течение ночи. Ночную культуру переносят в свежую среду LB с ампициллином (50 мкг/мл). Синтез белка индуцируют добавлением изопропилтиогалактазида до концентрации 1 мМ, когда культура достигает среднелогарифмической фазы роста (OD600=0,6-0,8). Индуцированные клетки инкубируют 4 часа при 30°С и качании со скоростью 180 об/мин, после чего биомассу отделяют центрифугированием при 6000 g.
Морфологические признаки рекомбинантного штамма Е. coli, трансформированного плазмидной ДНК pQE-30_P36GP12_GP57 и способного продуцировать рекомбинантный белок P36GP12. Клетки мелкие утолщенной палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаризованной среде LB или 2xYT - колонии круглые, гладкие, прижатые, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или LB бульоне) образуют интенсивную ровную муть. Клетки растут при температуре 37°С при оптимуме рН от 6.8 до 7.0.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к ампициллину (100 мкг/мл), обусловленную наличием плазмиды pQE-30_P36GP12_GP57.
Штамм E. coli DH5αF'/pQE-30_P36GP12_GP57 обеспечивает индуцируемый изопропилтиогалактозидом синтез рекомбинантного белка P36GP12 с уровнем экспрессии более 20% суммарного клеточного белка. Уровень экспрессии определяют с помощью денситометрии полиакриламидного геля, окрашенного Кумасси-G250 с использованием программного обеспечения Image Lab Ver. 3.0, поставляемого с прибором GelDocXR+(Bio-Rad).
Полученный штамм депонирован в Коллекции Экстремофильных Микроорганизмов и Типовых Культур ФГБУН Института химической биологии и фундаментальной медицины СО РАН (КЭМТК ИХБФМ СО РАН), под регистрационным номером №974 в Международной федерации коллекций культур (WDCM/WFCC).
Лизат индуцированных клеток Е. coli DH5αF'/pQE-30_P36GP12_GP57 используют для очистки рекомбинантного белка P36GP12 с помощью аффинной хроматографии. В результате получают рекомбинантный белок P36GP12, представляющий собой рекомбинантный хвостовой белок изолята Т4-подобного бактериофага. Данный белок имеет молекулярную массу в нативной тримерной форме около 160 кДа, состоит из N-концевого олигопептида MRGSHHHHHHGSAN и полноразмерного белка gp12 бактериофага Р36, обладающего способностью связывать бактериальные липополисахариды Е. coli. Рекомбинантный белок P36GP12 имеет аминокислотную последовательность SEQ ID NO: 2, представленную на фиг. 3, кодируемую нуклеотидной последовательностью SEQ ID NO: 1, представленной на фиг. 2.
Таким образом, впервые получена плазмидная ДНК pQE-30_P36GP12_GP57 и штамм-продуцент E. coli DH5αF'/pQE-30_P36GP12_GP57, обеспечивающие продукцию в бактериальных клетках Е. coli нативного рекомбинантного белка P36GP12, который представляет собой белок коротких хвостовых нитей P36GP12 изолята Р36 Т4-подобного бактериофага, состоящего из N-концевого олигопептида MRGSHHHHHHGSAN и полноразмерного гена белка коротких хвостовых нитей gp12, который с высоким сродством связывается с эндотоксинами бактериальных клеток Е. coli.
Изобретение иллюстрируется следующими графическими материалами, представленными на фигурах 1-6:
Фиг. 1. Общая схема структурной организации плазмиды pQE-30_P36GP12_GP57 (физическая карта), где: p36gp12 - ген, кодирующий рекомбинантный белок P36GP12, p36gp57 - ген, кодирующий белок-шаперон рекомбинантного белка P36GP12, Т5 - промотор фага Т5, AmpR-ген устойчивости к ампициллину; CmR - ген устойчивости к хлорамфениколу, указаны сайты рестрикции для эндонуклеаз BamHI и HindIII.
Фиг. 2. Нуклеотидная последовательность фрагмента плазмиды pQE-30_P36GP12_GP57, кодирующая рекомбинантный белок P36GP12, состоящий из N-концевого олигопептида MRGSHHHHHHGSAN и полноразмерного белка gp12 и белок-шаперон gp57.
Фиг. 3. Аминокислотная последовательность рекомбинантного белка P36GP12, состоящая из N-концевого олигопептида MRGSHHHHHHGSAN и полноразмерного белка gp12.
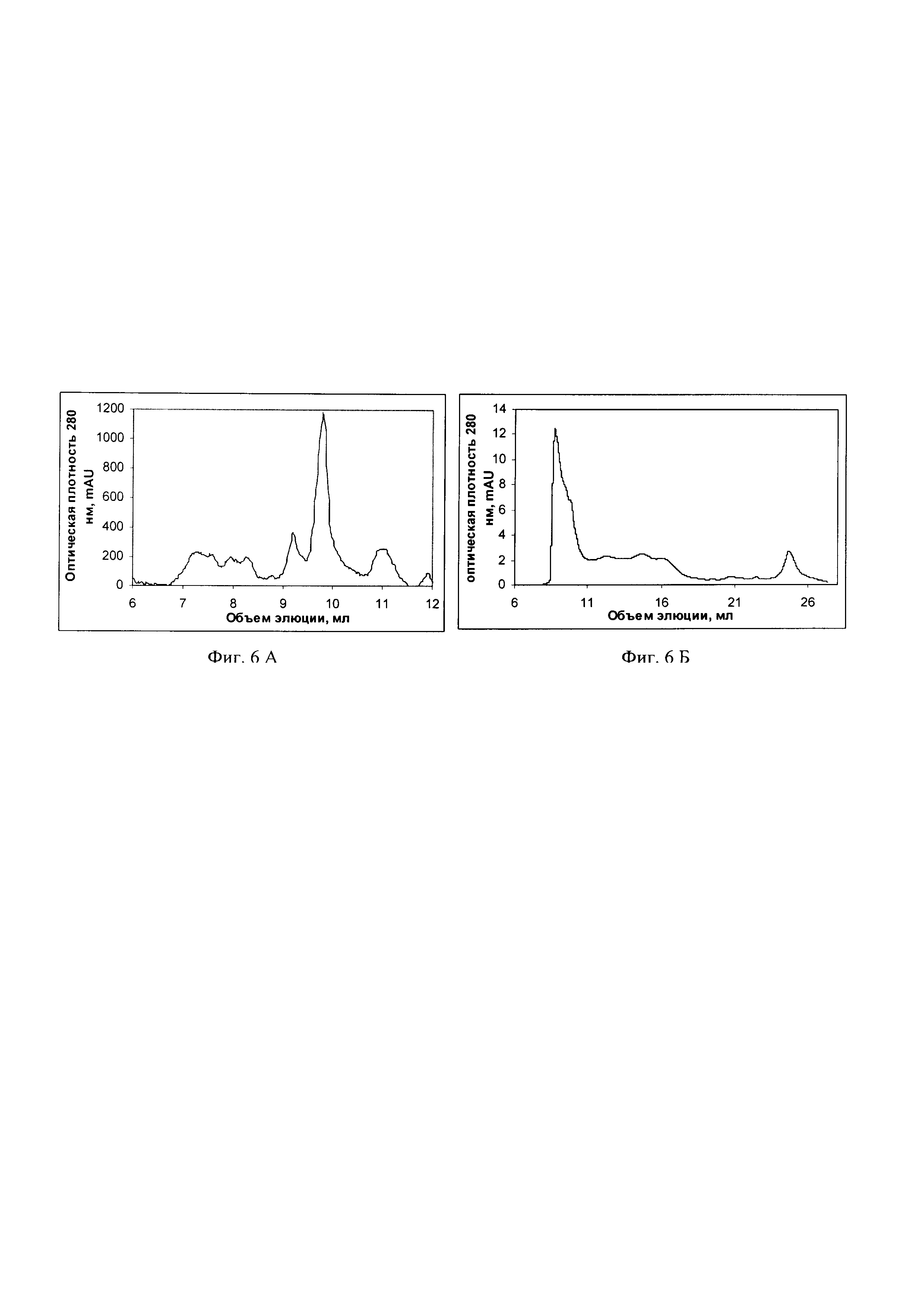
Фиг. 4 Электрофореграмма в 10%-ном полиакриламидном геле (ПААГ) фракций разрушенных клеток DH5αF', несущих плазмиду pQE-30_P36GP12_GP57, в 10%-ном ПААГ, где дорожки: 1 - набор белков с известной молекулярной массой Spectra (Thermo Fisher scientific, США), 2 - растворимая цитоплазматическая фракция без прогрева, 3 - нерастворимая цитоплазматическая фракция без прогрева; 4 - растворимая цитоплазматическая фракция, прогретая при 95°С в лизирующем буфере, содержащем 2% додецилсульфат натрия (ДСН); 5 - нерастворимая цитоплазматическая фракция, прогретая при 95°С в лизирующем буфере, содержащем 2% ДСН. Стрелками указаны тримерная форма (160 кДа) и мономерная форма (55 кДа) хвостового белка P36GP12.
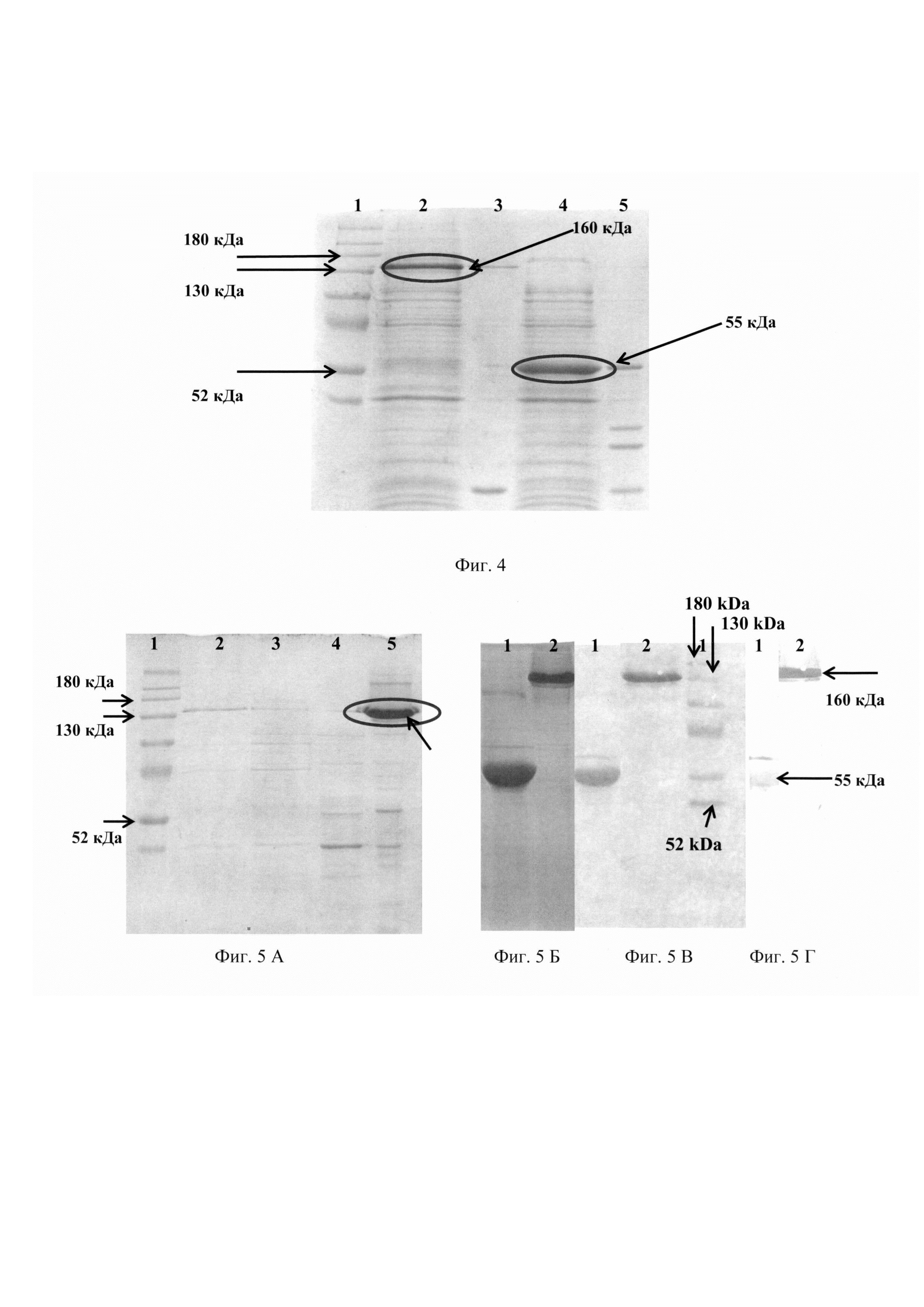
Фиг. 5 А. Электрофоретический анализ в 10%-ном ПААГ фракций, полученых при хроматографической очистке рекомбинантного белка P36GP12: 1 - набор белков с известной молекулярной массой Spectra (Thermo Fisher scientific, США); 2 - растворимая цитоплазматическая фракция до хроматографического разделения и концентрирования; 3 - проскок колонки, содержащей Ni-NTA-сорбент; 4 - фракция белков, элюированная с колонки 25 мМ раствором имидазола; 5 - фракция белков, элюированная с колонки 300 мМ раствором имидазола после концентрирования на фильтре Amicon 30 kDa (Millipore). Стрелкой указан полученный рекомбинантный белок P36GP12 в тримерной форме.
Фиг. 5 Б. Электрофоретический анализ в 10%-ном ПААГ раствора рекомбинантного белка P36GP12, сконцентрированного до концентрации 1-3 мг/мл: 1 - фракция белков, элюированная с колонки 300 мМ раствором имидазола, прогретая при 95°С в лизирующем буфере, содержащем 2% ДСН; 2 - фракция белков, элюированная с колонки 300 мМ раствором имидазола без прогрева.
Фиг. 5 В. Перенос на нитроцеллюлозную мембрану очищенного препарата рекомбинантного белка P36GP12: 1 - фракция, прогретая при 95°С в лизирующем буфере, содержащем 2% ДСН; 2 - фракция без прогрева; 3 - набор белков с известной молекулярной массой Spectra (Thermo Fisher scientific, США). Стрелками указаны молекулярные массы стандарта 52 кДа, 130 кДа и 180 кДа.
Фиг. 5 Г. Вестерн-блот анализ очищенного препарата рекомбинантного белка P36GP12: 1 - фракция, прогретая при 95°С в лизирующем буфере, содержащем 2% ДСН; 2 - фракция без прогрева. Стрелками указаны тримерная форма (160 кДа) и мономерная форма (55 кДа) рекомбинантного белка P36GP12.
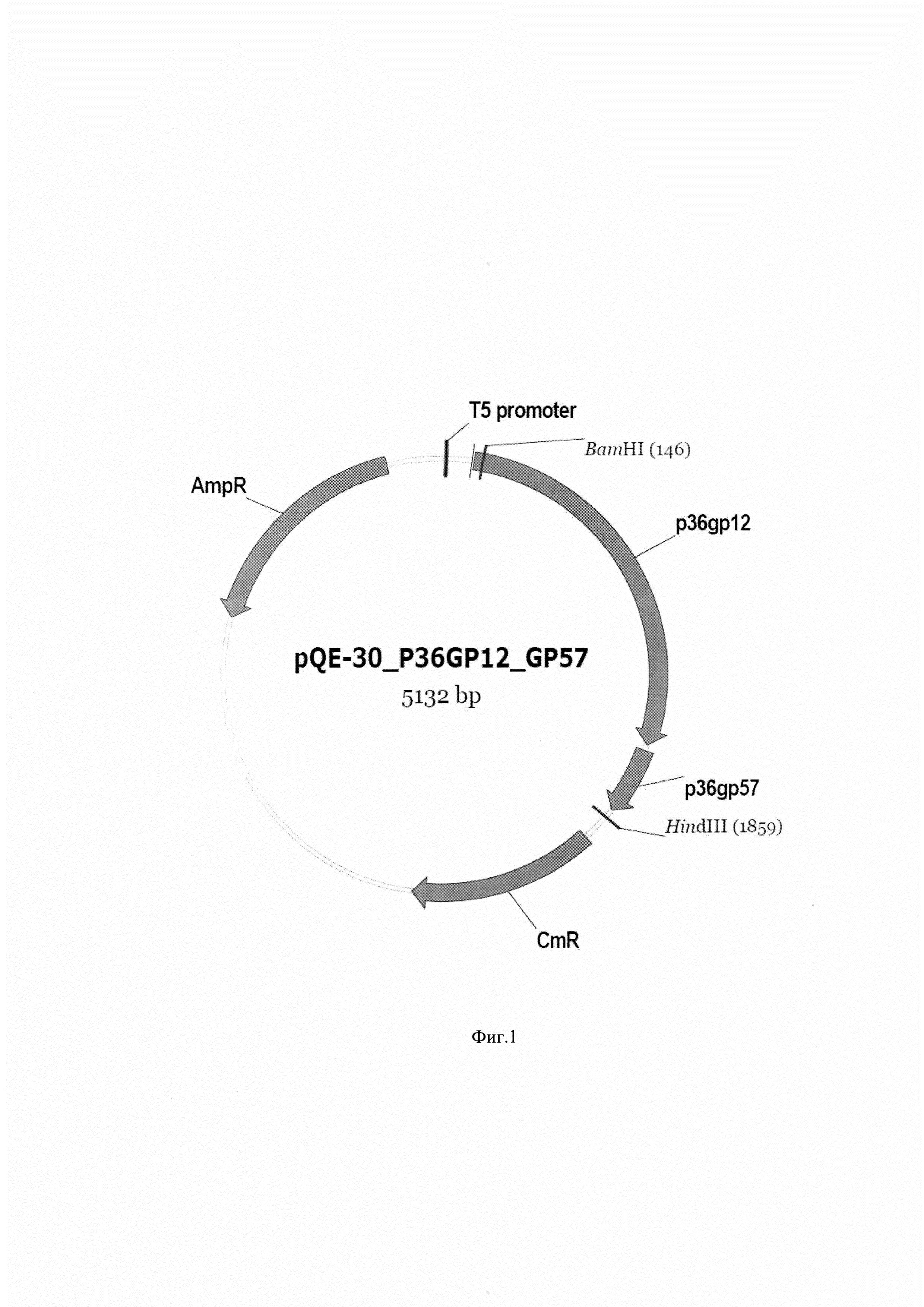
Фиг. 6 А. Хроматограмма элюции рекомбинантного белка P36GP12 с колонки при гель-фильтрации.
Фиг. 6 Б. Хроматограмма элюции рекомбинантного белка P36GP12 с колонки при ионообменной хроматографии.
Для лучшего понимания сущности предлагаемого изобретения изобретение иллюстрируется следующими примерами его осуществления.
Пример 1. Получение фрагментов ДНК, кодирующих белок коротких хвостовых нитей и его белок-шаперон.
В качестве источника генов, кодирующих белок коротких хвостовых нитей и его белок-шаперон, используют геномную ДНК изолята Т4-подобного бактериофага. Амплификацию генов, кодирующих полноразмерные белок коротких хвостовых нитей и его белок-шаперон, проводят с помощью ПЦР, используя в качестве ДНК-зависимой ДНК-полимеразы высокоточную ДНК-полимеразу Phusion (BioLabs, Англия). Используя олигонуклеотидные последовательности, частично комплементарные генам белков коротких хвостовых нитей и его белку-шаперону, PH36_gp12_dir: 5' АТС ACG GAT CCG CAA АСА ATA СТА ТТА ACC ATG ТАА AAG ACG ATG С 3' и PH36_gp12_rev: 5' AGT TAA TTT СТС СТС ТТА TTA GCG CAC CCT TAT GAT ATA GTT TAA TGC 3', PH36_gp57_dir: 5' GTG CGC TAA TAA GAG GAG AAA TTA ACT ATG ТСС AAT CAG CAT GAA С 3' и PH36_gp57_rev: 5' СТА ATT AAG CTT ATT AAG CCT CCG TGA TCA GTT СТА CTT CCT CTT С 3' соответственно, получают фрагменты ДНК, кодирующие белок коротких хвостовых нитей и его белок-шаперон и сайты, необходимые для узнавания эндонуклеазами рестрикции BamHI и HindIII. ПЦР проводят в амплификаторе (Bio Rad, США). Условия проведения реакции: предварительная денатурация - 2 минуты при 94°С; 30 циклов: 30 секунд при 94°С, 30 секунд при 50°С, 50 секунд при 72°С; заключительное инкубирование - 10 минут при 72°С. Продукты реакции анализируют электрофорезом в 1%-ном агарозном геле, вырезают участки геля, содержащие фрагменты ДНК размером около 1431 п.о. и 258 п.о. для гена белка коротких хвостовых нитей и его белка-шаперона, соответственно. Из вырезанных участков геля элюируют ДНК-фрагменты набором GeneJET™ Gel Extraction Kit (Fermentas, Литва) по протоколу производителя. Для подтверждения гомогенности полученные ДНК-фрагменты анализируют электрофорезом в 1%-ном агарозном геле.
Пример 2. Получение объединенного ДНК-фрагмента, кодирующего белок коротких хвостовых нитей и его белок-шаперон.
Отдельные ДНК-фрагменты, кодирующие белок коротких хвостовых нитей и его белок-шаперон, далее используют в объединяющей полимеразной цепной реакции (синтез с помощью удлинения за счет перекрывающихся концов), которая позволяет наработать единый ДНК-фрагмент. Для этого готовят две реакционные смеси. Первая смесь состоит из ДНК-полимеразу Phusion, буфера для проведения реакции, 0,4 мМ каждого дезоксинуклеотидтрифосфата (дНТФ), матрицы в виде 10 нг ДНК каждого ДНК-фрагмента. Вторая смесь содержит ДНК-полимеразу Phusion, буфер для проведения реакции, 0,4 мМ каждого дНТФ, 0,6 мМ специфические олигонуклеотиды. Реакцию проводили по следующему протоколу: предварительно выдерживают пробирки с первой смесью при 94°С в течение 5 минут, далее в течение 10 циклов 30 секунд при 94°С, 2 минуты при 60°С, 3 минуты при 72°С, затем инкубируют при 10°С в течение 10 минут и в это время добавляют равный объем второй смеси, после чего в течение 35 циклов выдерживают при 94°С 30 секунд, при 58°С 30 секунд, при 72°С 2 минуты, затем следует заключительное инкубирование - 10 минут при 72°С. Затем полученные ПЦР-продукты разделяли в течение часа в 1%-ном агарозном геле и вырезают участки, содержащие ДНК-фрагменты размером примерно 1689 п.о. Из вырезанных участков элюируют объединенные ДНК-фрагменты, кодирующие белок коротких хвостовых нитей и его белок-шаперон, и анализируют чистоту ДНК-фрагментов с помощью электрофоретического разделения 1%-ном агарозном геле.
Пример 3. Способ конструирования плазмиды pQE-30_P36GP12_GP57.
Плазмидную ДНК pQE-30 и объединенный ДНК-фрагмент, кодирующий белок коротких хвостовых нитей и его белок-шаперон, подвергают гидролизу, используя эндонуклеазы рестрикции BamHI и HindIII, для образования липких концов. Для этого готовят две реакционных смеси, первая из которых содержит плазмидную ДНК pQE-30, 10 мМ Трис-HCl (рН 8.5), 10 мМ MgCl2, 100 мМ NaCl, 1 мМ дитиотреитол и по 5 ед. активности ферментов BamHI и HindIII, а вторая --объединенный ДНК-фрагмент и те же компоненты буфера и ферменты. Полученные реакционные смеси инкубируют при 37°С в течение часа. Далее продукты реакции гидролиза электрофоретически разделяют в 1%-ном агарозном геле и вырезают участки, приблизительно равные молекулярной массе линеаризованного вектора pQE-30 (3425 п.о.), либо объединенного фрагмента (1707 п.о.). Затем ДНК элюируют из геля с использованием набора GeneJET™ Gel Extraction Kit (Fermentas, Литва) в соответствии с протоколом производителя. Реакцию лигирования проводят с использованием ДНК-лигазы бактериофага Т4 и буфера (Fermentas, Литва), предоставленного производителем, при комнатной температуре в течение 1 часа. Далее лигазной смесью трансформируют компетентные клетки штамма DH5αF' Е. coli. С помощью полимеразной цепной реакции и олигонуклеотидов PH36_gp12_dir и PH36_gp57_rev отбирают клоны, содержащие вставку рассчитанного размера (1707 п.о.). Схема (физическая карта) полученной плазмидной ДНК pQE-30_P36GP12_GP57 представлена на фиг. 1.
Пример 4. Получение штамма-продуцента рекомбинантного белка P36GP12 Т4-подобного бактериофага
Полученной плазмидной ДНК pQE-30_P36GP12_GP57 трансформируют клетки Е. coli штамма DH5αF', и инкубируют при 37°С в течение ночи. Ночную культуру переносят в свежую среду LB в отношении объемов 1:50 и с добавлением ампициллина (50 мкг/мл). Синтез мРНК с плазмидной ДНК индуцируют добавлением изопропилтиогалактазида (ИПТГ) до концентрации 1 мМ, когда культура достигает среднелогарифмической фазы роста (OD600=0,6-0,8). Индуцированную культуру инкубируют при 30°С в течение 6 часов для наработки рекомбинантного белка. Далее клетки осаждают центрифугированием при 6000g в течение 10 минут.
Пример 5. Получение и очистка рекомбинантного белка P36GP12.
Рекомбинантный белок P36GP12 получают из растворимой цитоплазматической фракции индуцированных клеток Е. coli DH5αF'/pQE-30_P36GP12_GP57 в результате аффинной хроматографии на Ni-NTA агарозе (Qiagen, Германия). Индуцированные клетки Е. coli DH5αF'/pQE-30_P36GP12_GP57 осаждают центрифугированием при 6000 g в течение 10 мин. Осадок растворяют в 1/10 объема буфера PBS, содержащего 137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,8 KH2PO4, и разрушают с помощью ультразвукового дезинтегратора в течение 15 мин импульсами мощностью 500 Вт и продолжительностью 20 сек с перерывами 20 сек между импульсами. Полученную суспензию центрифугируют при 16000 g в течение 10 минут, после чего переносят в чистую пробирку супернатант, представляющий собой раствор цитоплазматических белков, а осадок растворяют в PBS. Полученные клеточные фракции анализируют электрофорезом в 10% полиакриламидном геле по Лэммли с предварительным прогреванием проб при 95°С либо без прогревания.
Электрофореграмма фракций разрушенных клеток DH5αF', несущих плазмиду pQE-30_P36GP12_GP57, в 10%-ном ПААГ представлена на фиг. 4. Результаты анализа демонстрируют, что в растворимой цитоплазматической фракции культуры DH5αF'/pQE-30_P36GP12_GP57 содержится белок, по электрофоретической подвижности соответствующий рекомбинантному белку P36GP12 в форме тримера с расчетной молекулярной массой около 160 кДа (дорожка 2). В результате прогревания белок P36GP12 переходит в мономерную форму с молекулярной массой около 55 кДа (дорожка 4).
На хроматографическую колонку, содержащую 4 мл Ni-NTA агарозы (Qiagen, Германия) и уравновешенную буфером А, содержащим 50 мМ Na-фосфатный буфер рН 8.0, 300 мМ NaCl, 5 мМ Трис-HCl, наносят 20 мл раствора, представляющего собой фракцию растворимых цитоплазматических белков индуцированных клеток. Нанесение производят со скоростью потока 1 мл/мин. Колонку промывают 20 мл буфера А, после чего проводят элюцию неспецифически сорбирующихся белков Е. coli 20 мл буфера А, содержащего дополнительно 25 мМ имидазола. Рекомбинантный белок элюируют 10 мл буфера А, содержащего 300 мМ имидазола. Полученные белковые фракции диализуют против 150 мМ NaCl, Трис-HCl рН 7.5 (две смены по 18 ч при 5°С) и анализируют электрофорезом в 10% полиакриламидном геле по Лэммли.
На фиг. 5 А представлен очищенный на хроматографической колонке рекомбинантный белок P36GP12, содержащийся в тримерной форме (дорожка 5). Далее с помощью фильтра Amicon ultra-4 (Millipore) с порогом отсечения 50 кДа, концентрируют раствор белка P36GP12 до концентрации 0,5-3 мг/мл.
Электрофореграмма белка P36GP12 после концентрирования представлена на фиг. 5 Б. Рекомбинантный белок P36GP12 находится в тримерной форме с молекулярной массой 160 кДа (дорожка 2), однако при нагревании при 95°С в присутствии додецилсульфата натрия переходит в мономерную форму (дорожка 1).
Наличие гексагистидиновой последовательности в полученном рекомбинантном белке подтверждают вестерн-блот анализом. Для этого образец, содержащий около 2 мкг рекомбинантного белка P36GP12, разделяют в 10% полиакриламидном геле с предварительным прогреванием при 95°С, либо без прогревания, переносят на нитроцеллюлозную мембрану методом электропереноса при напряжении 80 В в течение 4 часов. Мембрану окрашивают кислым раствором красителя Ponseau S (Понсо С) в течение 15 мин. Результат представлен на фиг. 5 В. Рекомбинантный белок P36GP12 находится в тримерной форме (дорожка 2), либо в мономерной форме (дорожка 1).
Вестерн-блот анализ проводят следующим образом. На первой стадии окрашенную нитроцеллюлозную мембрану, содержащую белок P36GP12 в тримерной и мономерной форме, а также белковый маркер молекулярных масс, блокируют инкубированием при 37°С в течение 1 часа в 4% суспензии нежирного сухого молока, разведенной в PBS с добавлением детергента Твин-20 (PBS-твин). Далее мембрану инкубируют с мышиным моноклональным антителом 6x-His Tag Monoclonal Antibody (Thermo Fisher scientific, США), разведенном в 10000 раз в буфере PBS-твин в течение 1 часа при 37°С на орбитальной шейкерной платформе. После трехкратной серии пятиминутных промывок в буфере PBS-твин мембрану инкубируют в течение 1 часа при 37°С на орбитальной шейкерной платформе в растворе, содержащем в разведении 1/5000 антитела козы против полной молекулы антител мыши Anti-Mouse IgG (whole molecule) antibody produced in goat, конъюгированные с щелочной фосфатазой (Sigma-Aldrich, США). После трехкратной серии пятиминутных промывок в буфере PBS-твин мембрану трижды промывают буфером для щелочной фосфатазы (АР-буфер, состав: 100 мМ Трис рН 9.5; 50 мМ MgCl2, 100 мМ NaCl, 0.1% Твин 20). Образовавшиеся иммунные комплексы выявляют в результате инкубации мембраны в АР-буфере, содержащем красители nitro blue tetrazolium (NBT) и 4-бром-5-хлор-индолилфосфат (BCIP) в конечной концентрации 170 мкг/мл и 330 мкг/мл, соответственно.
Результат вестерн-блот анализа подтверждает, что как тримерная, так и мономерная форма белка содержат гексагистидиновый олигопептид. В совокупности с молекулярной тримерной и мономерной форм белка, оцененной по подвижности в геле, а также с тем обстоятельством, что при нагревании в присутствии додецилсульфата натрия молекулярная масса белка уменьшается приблизительно в 3 раза, можно заключить, что анализируемый белок представляет собой целевой рекомбинантный белок P36GP12.
Пример 6. Проверка способности белка P36GP12 связываться с декстран-содержащими полимерами.
Рекомбинантный белок P36GP12, растворенный до концентрации 1 мг/мл в буфере, содержащем 50 мМ Трис-HCl, 150 мМ NaCl, рН 7,5 при 22°С, используют в качестве тестового образца при проведении гель-фильтрации с использованием колонки "Superdex 200 Increase 10/300 GL" (GE Healthcare) диаметром 10 мм и длиной 300 мм и хроматографа AKTA Explorer 100 (GE Healthcare). В петлю объемом 500 мкл вносят 1 мл исследуемого образца. Хроматографию проводят при 22°С и скорости потока 0,4 мл/мин, используя в качестве системного буфера буфер, содержащий 50 мМ Трис-HCl, 150 мМ NaCl, рН 7,5 при 22°С. Объем элюции после введения пробы из петли на колонку составляет 26,4 мл. Полученную в результате хроматограмму (фиг. 6 А) регистрируют и с помощью программного обеспечения Unicorn 5 интегрируют сигнал, детектируемый при длине волны 280 нм. Полученное значение составляет 40±4 mAU*мл, где mAU - относительная единица оптической плотности, используемая в данном хроматографе.
Далее рекомбинантный белок P36GP12, растворенный до концентрации 1 мг/мл в буфере, содержащем 20 мМ Трис-HCl, рН 9,0 при 22°С, используют в качестве тестового образца при проведении ионообменной хроматографии с использованием колонки "POROS HQ20 4.6/100" (GE Healthcare) диаметром 4.6 мм и длиной 100 мми хроматографа AKTA Explorer 100 (GE Healthcare). В петлю объемом 500 мкл вносят 1 мл исследуемого образца. Хроматографию проводят при 22°С и скорости потока 0.4 мл/мин, используя в качестве буфера А, буфер, содержащий 20 мМ Трис-HCl, рН 9,0 при 22°С, а в качестве буфера Б, буфер, содержащий 20 мМ Трис-HCl, 500 мМ NaCl, рН 9,0 при 22°С. Элюция достигается градиентом от 0 до 100% буфера Б за 10 объемов колонки. Полученную в результате хроматограмму (фиг. 6 Б) регистрируют и с помощью программного обеспечения Unicorn 5 интегрируют сигнал, детектируемый при длине волны 280 нм. Полученное значение составляет 1000±100 mAU*мл, где mAU - относительная единица оптической плотности, используемая в данном хроматографе.
Сопоставляя интегральную оптическую плотность элюата на выходе колонки, полученную в результате гель-фильтрации, с интегральной оптической плотностью элюата на выходе с колонки, полученную в результате ионообменной хроматографии, регистрируют достоверное различие, которое достигает 10-30 раз. Это различие свидетельствует о том, что в случае гель-фильтрации наблюдается аффинное взаимодействие белка P36GP12 с хроматографической матрицей, представляющей собой декстран, сшитый с агарозой, то есть полимер, состоящий из звеньев глюкозы и галактозы. Наличие такого взаимодействия свидетельствует о наличии у белка P36GP12 свойства связываться с липополисахаридами, присущего адгезинам бактериофагов.
Рекомбинантный белок P36GP12 может быть использован для очистки биотехнологических субстанций от бактериальных липополисахаридов.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Petsch D., Anspach F. В. Endotoxin removal from protein solutions // J. Biotechnol. - 2000. - T. 76, №2-3. - C. 97-119.
2. Статья ОФС.1.2.4.0006.15 Государственной фармакопеи Российской Федерации (ГФ РФ) XIV.
3. Anspach F. В. Endotoxin removal by affinity sorbents // J. Biochem Biophys. Methods. - 2001. - T. 49, №1-3. - C. 665-81.
4. 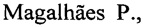 Lopes A., Mazzola P., Rangel-Yagui C, Penna Т., Pessoa A. Methods of endotoxin removal from biological preparations: a review. // J. Pharm Pharm. Sci. - 2007. - T. 10, №3. - C. 388-404.
Lopes A., Mazzola P., Rangel-Yagui C, Penna Т., Pessoa A. Methods of endotoxin removal from biological preparations: a review. // J. Pharm Pharm. Sci. - 2007. - T. 10, №3. - C. 388-404.
5. Matsumae H., Minobe S., Kindan K., Watanabe Т., Sato Т., Tosa T. Specific removal of endotoxin from protein solutions by immobilized histidine // Biotechnol Appl Biochem. - 1990. - T. 12, №2. - C. 129-40.
6. Riede I. Receptor specificity of the short tail fibres (gp12) of T-even type Escherichia coli phages // Mol. Gen Genet. - 1987. - T. 206, №1. - С. 110-5.
7. Burda M. R, Miller S Folding of coliphage T4 short tail fiber in vitro. Analysing the role of a bacteriophage-encoded chaperone // Eur J Biochem. - 1999. - №265. - C. 771-8.




Рекомбинантная плазмидная днк pqe-p35d, обеспечивающая синтез рекомбинантного белка p35d вируса оспы коров, штамм бактерий escherichia coli - продуцент рекомбинантного белка p35d вируса оспы коров и рекомбинантный белок p35d вируса оспы коров, используемый для создания тест-систем и конструирования субъединичных вакцин против ортопоксвирусных инфекций
Рекомбинантная плазмидная днк pg1-rm7, обеспечивающая синтез гибридного белка g1-rm7, и гибридный белок, связывающий фактор некроза опухолей и обладающий биолюминесцентной активностью
Средство для нейтрализации вируса натуральной оспы
Способ получения наноразмерной системы доставки нуклеозидтрифосфатов в клетки млекопитающих
Средство, обладающее антибактериальной активностью
Фосфорсодержащие фторированные производные 1,4-нафтохинона, обладающие цитотоксической активностью по отношению к раковым клеткам человека в культуре
Способ очистки вируса осповакцины или его рекомбинантных вариантов
Рнк-аптамер, обладающий способностью узнавать характерные для рассеянного склероза аутоантитела
Рекомбинантная плазмидная днк pclm4/hygro-14d5, кодирующая полипептид со свойствами легкой цепи химерного антитела против вируса клещевого энцефалита, и рекомбинантная плазмидная днк pchm2-14d5, кодирующая полипептид со свойствами тяжелой цепи химерного антитела против вируса клещевого энцефалита, химерное антитело, обеспечивающее экстренную профилактику клещевого энцефалита у мышей
Способ получения экзосом из крови
Штамм бактериофага staphylococcus aureus sa20, обеспечивающий разрушение биопленок, образуемых бактериями рода staphylococcus
Способ оценки эффективности фаготерапии при лечении инфекционных заболеваний
Рекомбинантная плазмидная днк pyfi-gfp, кодирующая продукцию флюоресцентного белка gfpaav и штамм бактерий escherichia coli jm109-pyfi, продуцирующий флюоресцентный белок gfpaav в присутствии токсических агентов
Рекомбинантная плазмидная днк pсl1, кодирующая полипептид со свойствами легкой цепи антитела человека против вируса эбола, рекомбинантная плазмидная днк рсн1, кодирующая полипептид со свойствами тяжелой цепи указанного антитела, и их применение
Способ определения антагонистической активности штаммов бактерий в отношении pseudomonas aeruginosa

