Результат интеллектуальной деятельности: КЛЕТКИ В КАЧЕСТВЕ МОДЕЛИ ДЛЯ ИДЕНТИФИКАЦИИ ВОЗМОЖНЫХ МОДУЛЯТОРОВ ВКУСОВЫХ ОЩУЩЕНИЙ
Вид РИД
Изобретение
Каждая ссылка, приведенная в данном описании, включена сюда во всей своей полноте.
Эта заявка включает посредством ссылки текстовый файл размером 13,7 кбайт, созданный 5 мая 2014 года и названный «056943.01231sequencelisting.txt», который представляет собой список последовательностей для этой заявки.
ОБЛАСТЬ ТЕХНИКИ
Это описание относится в целом к линиям клеток и тестам, которые могут использоваться для идентификации модуляторов рецепторов сладкого вкуса.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
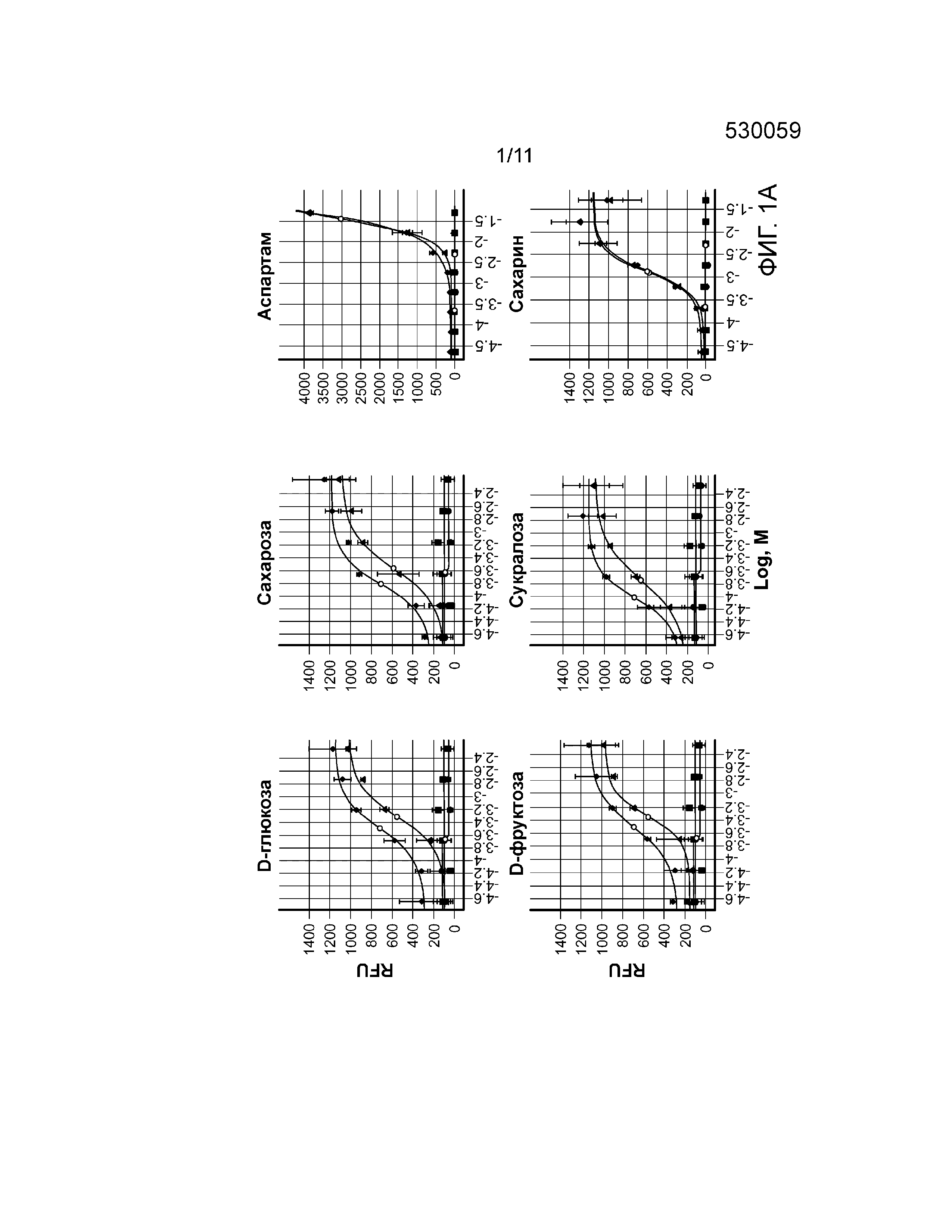
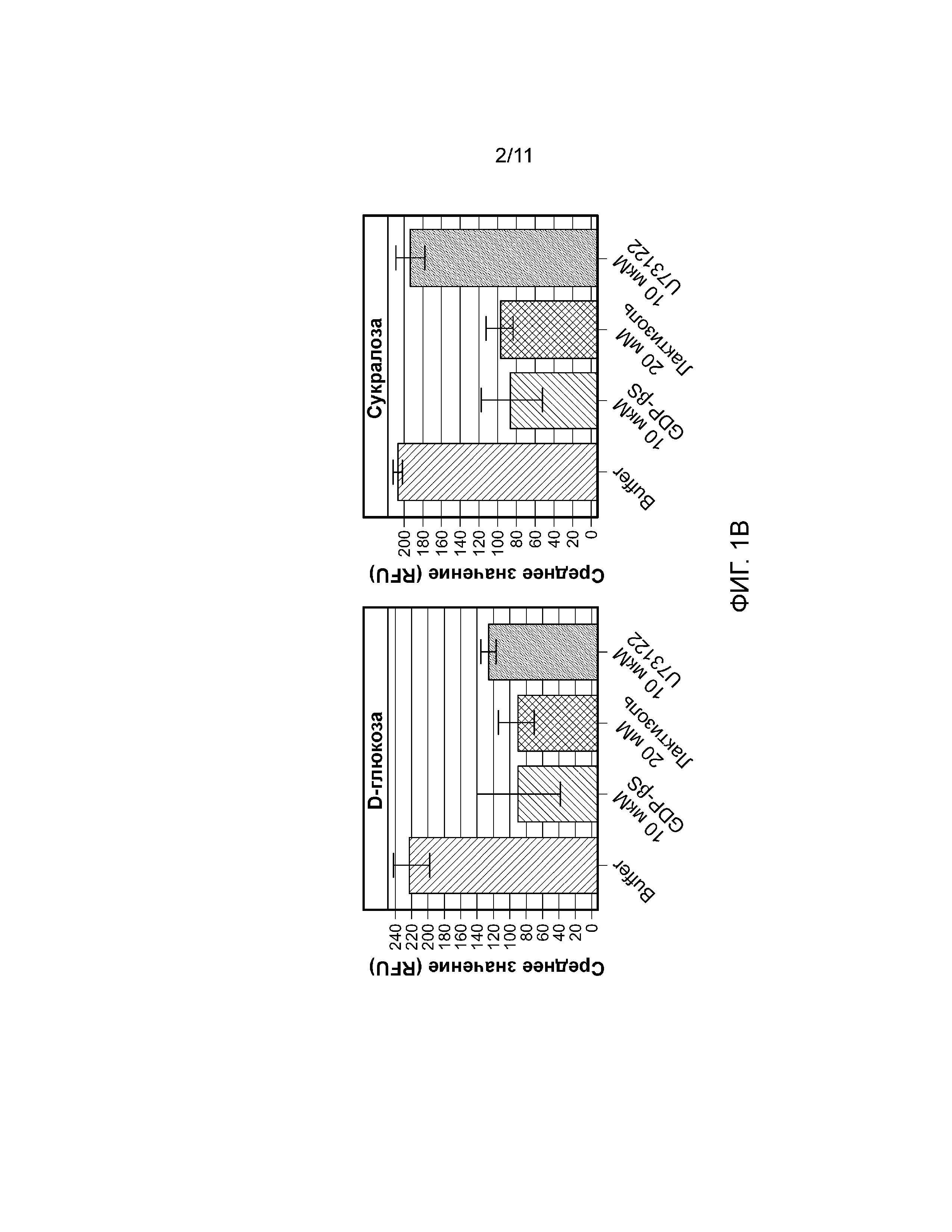
Фиг. 1A-B. Са(2+) ответ на соединения со сладким вкусом. Фиг. 1А, кривая зависимости от дозы высвобождения Са(2+) в JUMP-IN™ T-REX™ U2-OS с (ромб) и без (треугольник) активации T1R3/T1R2/Gα15 (n=3 эксперимента. Фиг. 1В, Эффекты лактизоля и U73122 на Ca(2+) ответ в клетках JUMP-IN™ T-REX™ U2-OS (среднее значение ±SD (среднеквадратическое отклонение).
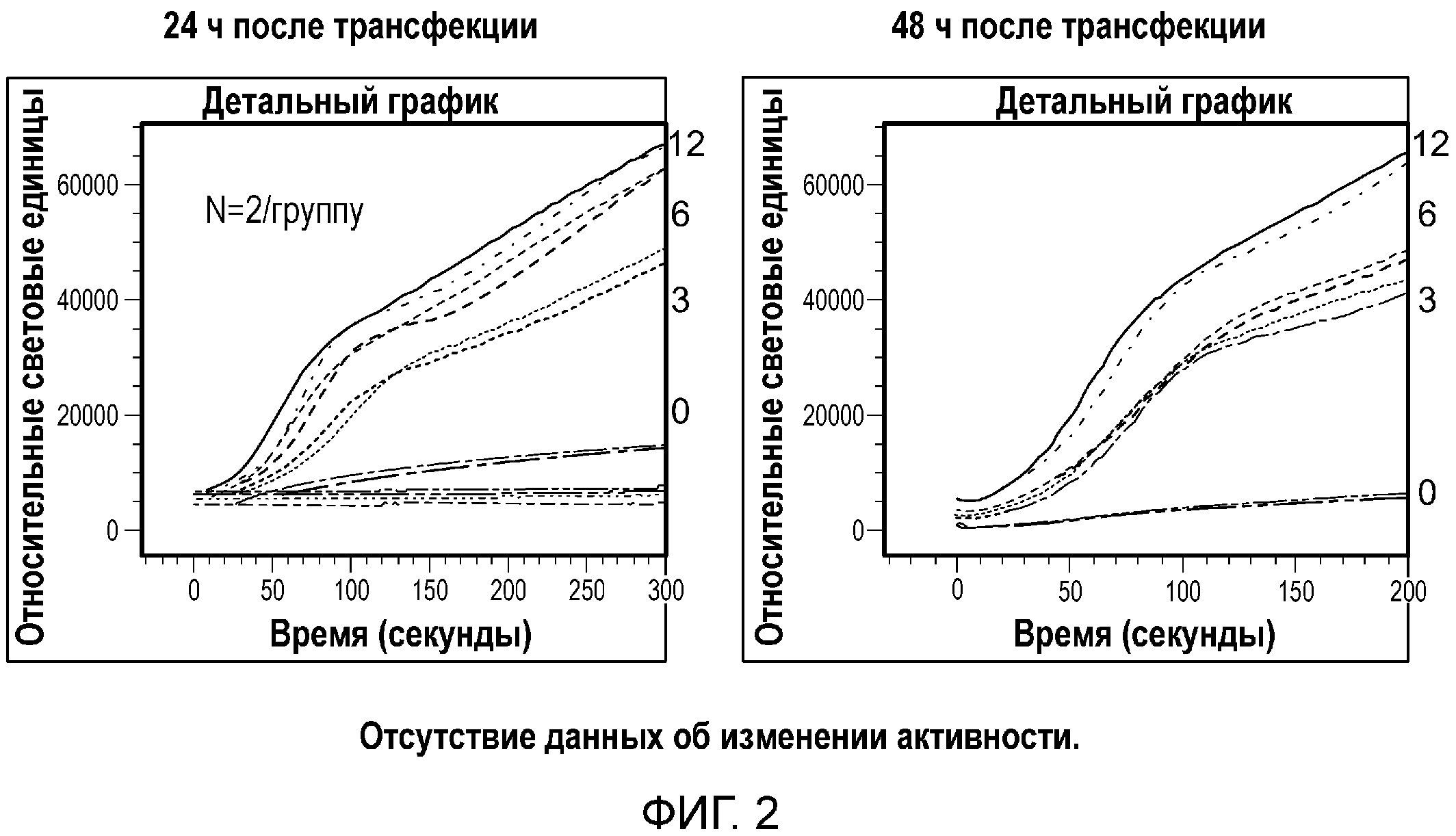
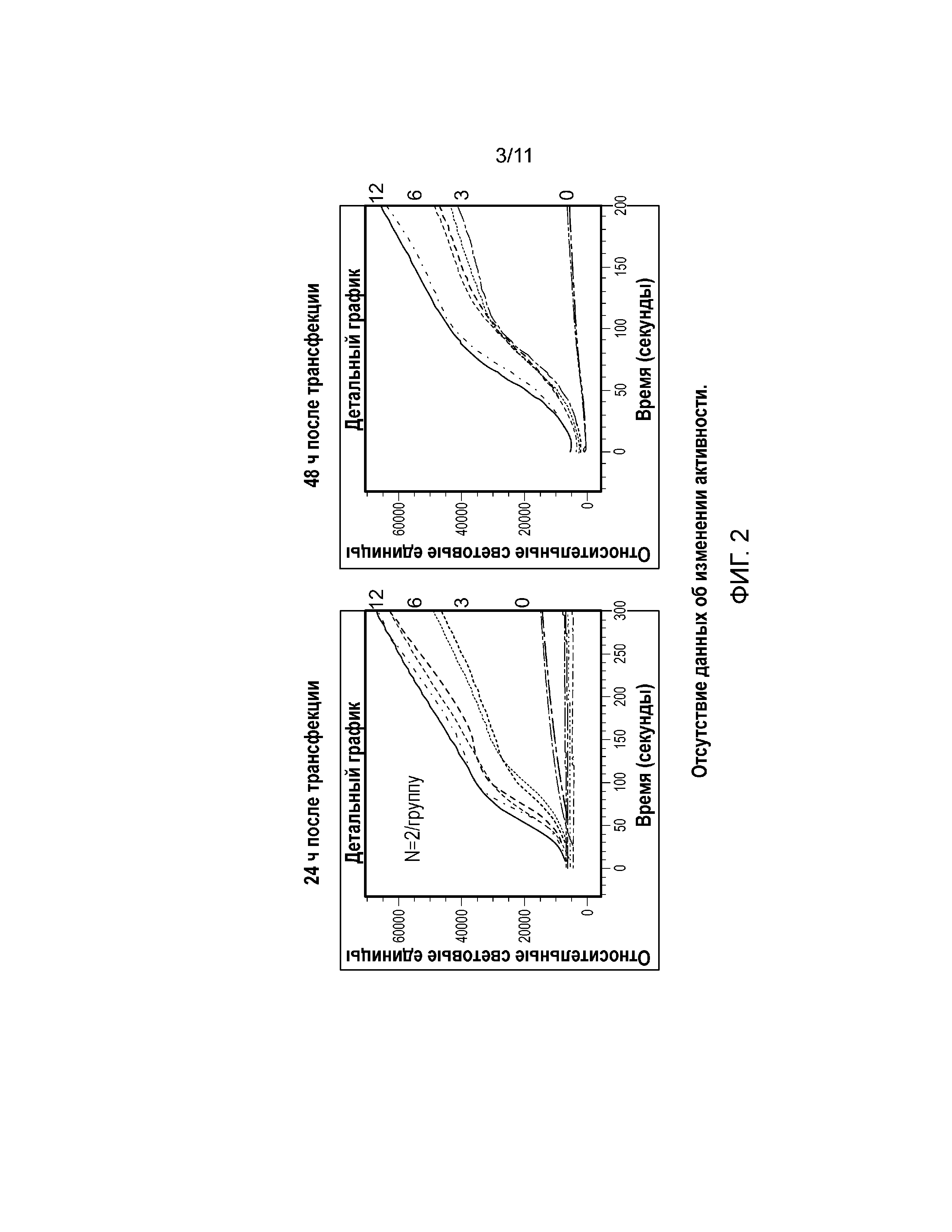
Фиг. 2. Графики, демонстрирующие эффект белка Gα15 на ответ клеток U2-OS на 100 мМ сукралозу. Клетки U2-OS транзиторно трансфецировали с использованием 3, 6 и 12 мкл/40000 клеток BacMam, кодирующего G-белок, гастдуцин (Gα15), в течение 24 ч (слева) или 48 часов (справа).
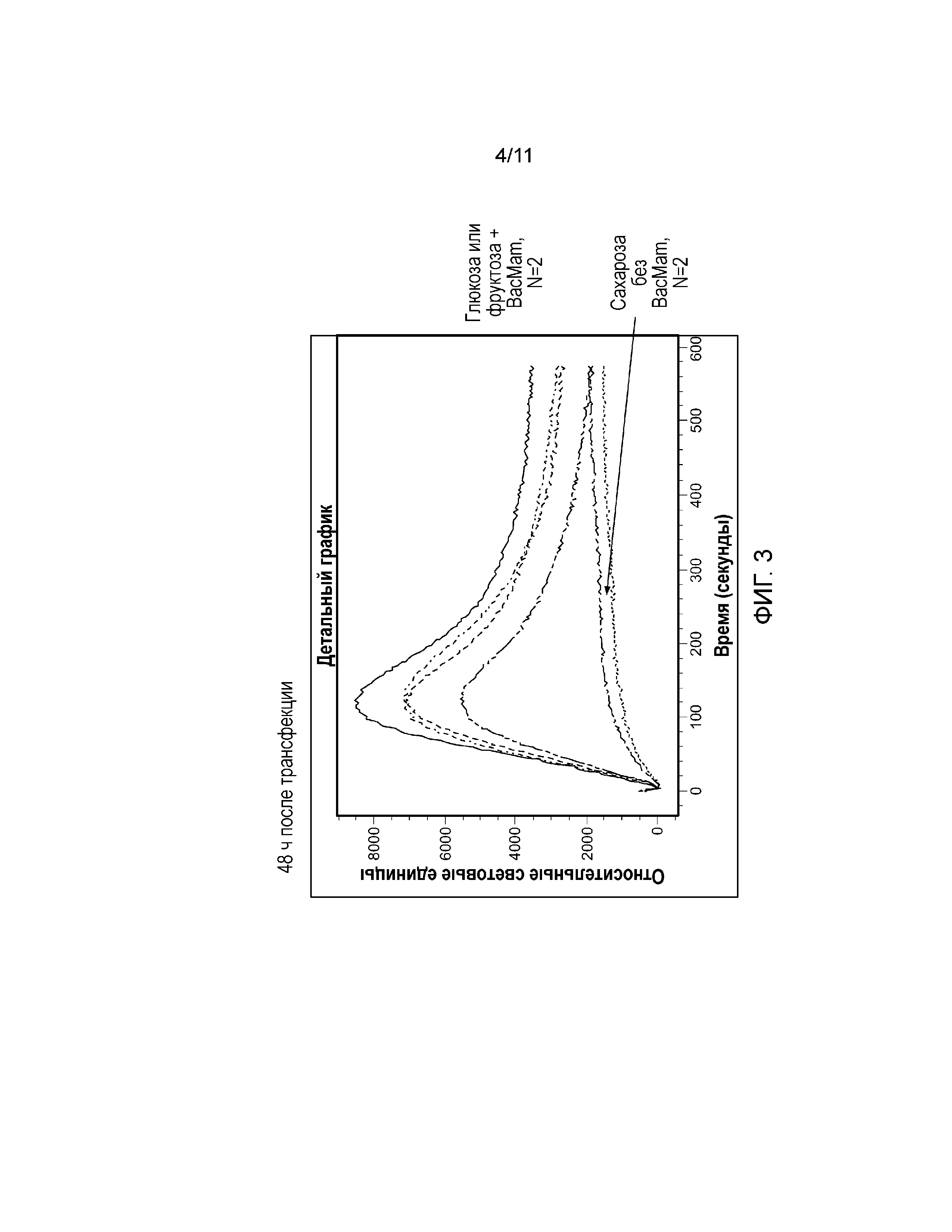
Фиг. 3. График, демонстрирующий эффект природных сахаров (глюкозы, фруктозы и сахарозы) в концентрации 100 мМ на клетки U2-OS, трансфецированные с использованием 0 или 6 мкл/40000 клеток BacMam, кодирующего Gα15.
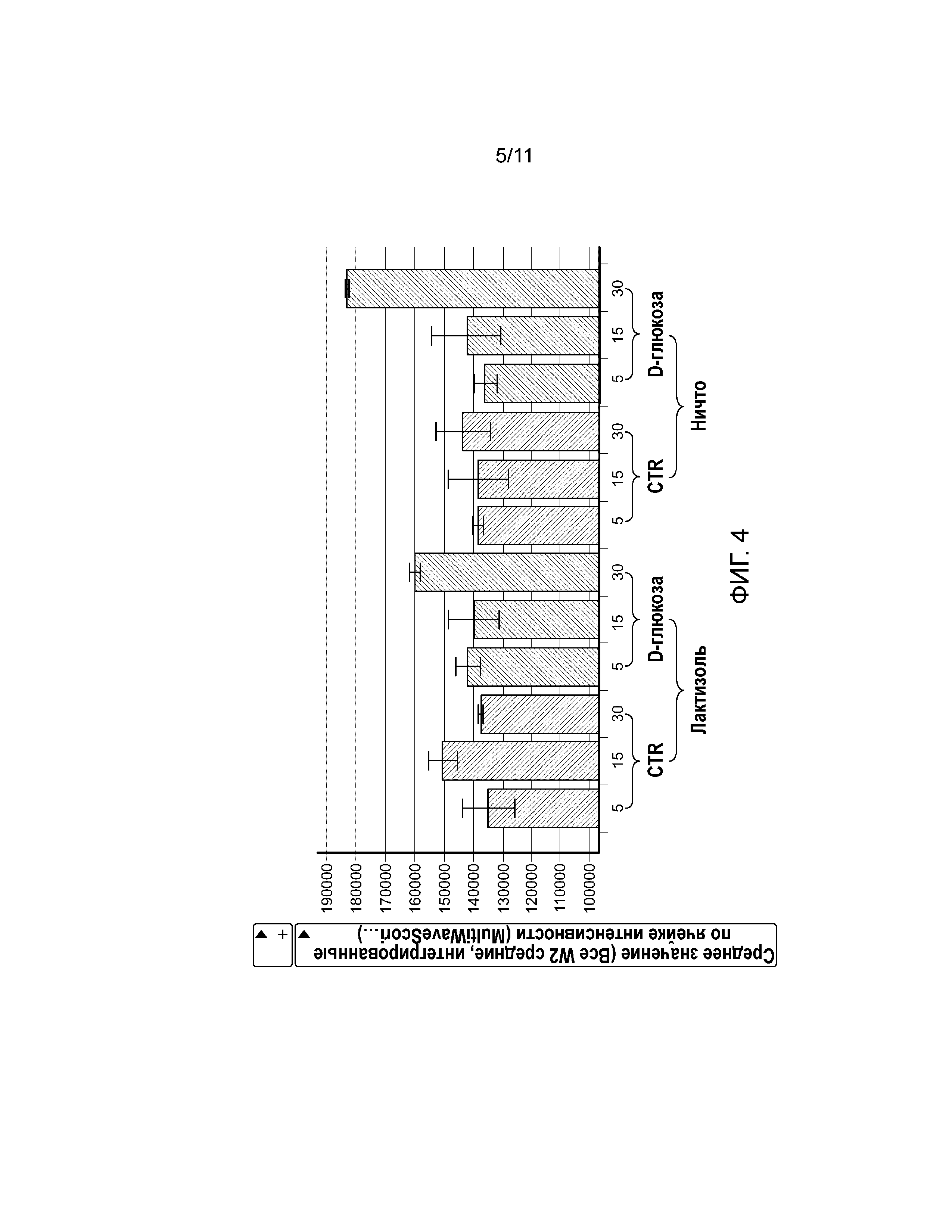
Фиг. 4. График, показывающий, что рецептор сладкого вкуса активирует pERK1/2 в клетках U2-OS при обработке глюкозой в концентрации 100 мМ. Обработка клеток U2-OS лактизолем (20 мМ, левые столбики) блокировала D-глюкозо-опосредованную активацию pERK1/2.
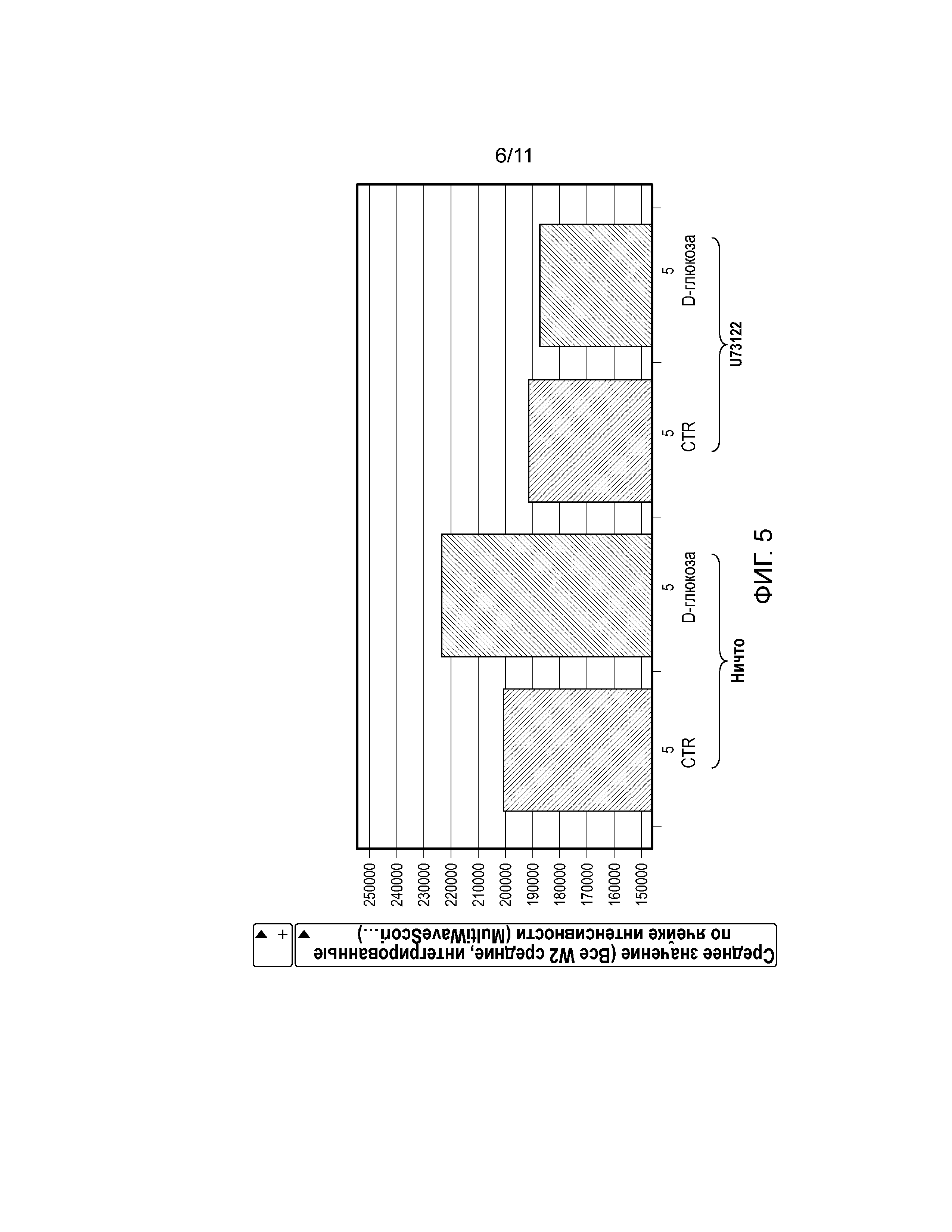
Фиг. 5. График, показывающий, что трансфекция клеток U2-OS с использованием Gα15 (6 мкл/40000 клеток) значительно увеличивает активацию pERK1/2, которая была заблокирована ингибитором PLCβ2 U73122 (10 мкМ).
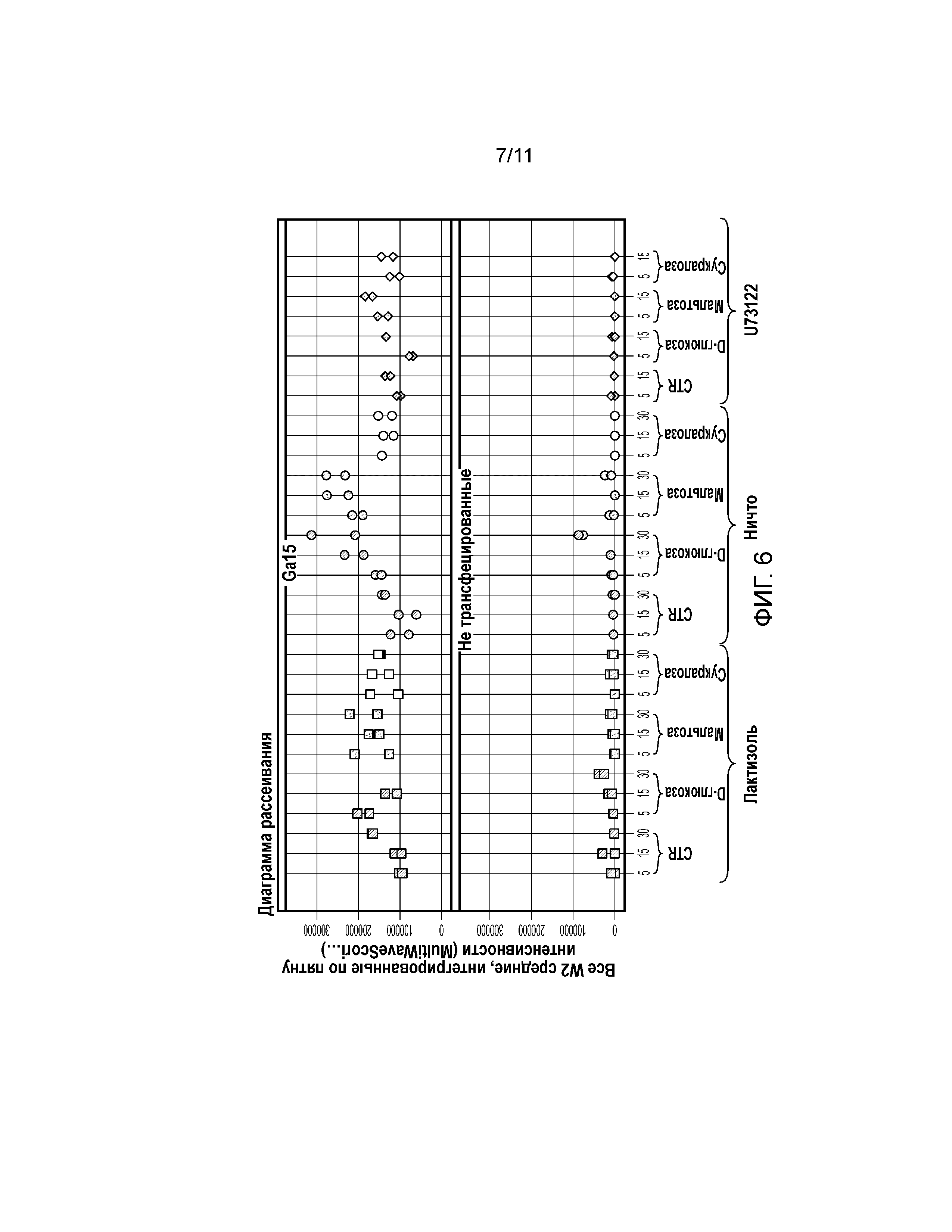
Фиг. 6. График, демонстрирующий активацию pERK1/2 при обработке D-глюкозой и мальтозой в клетках U2-OS, трансфецированных Gα15 (6 мкл/40000 клеток).
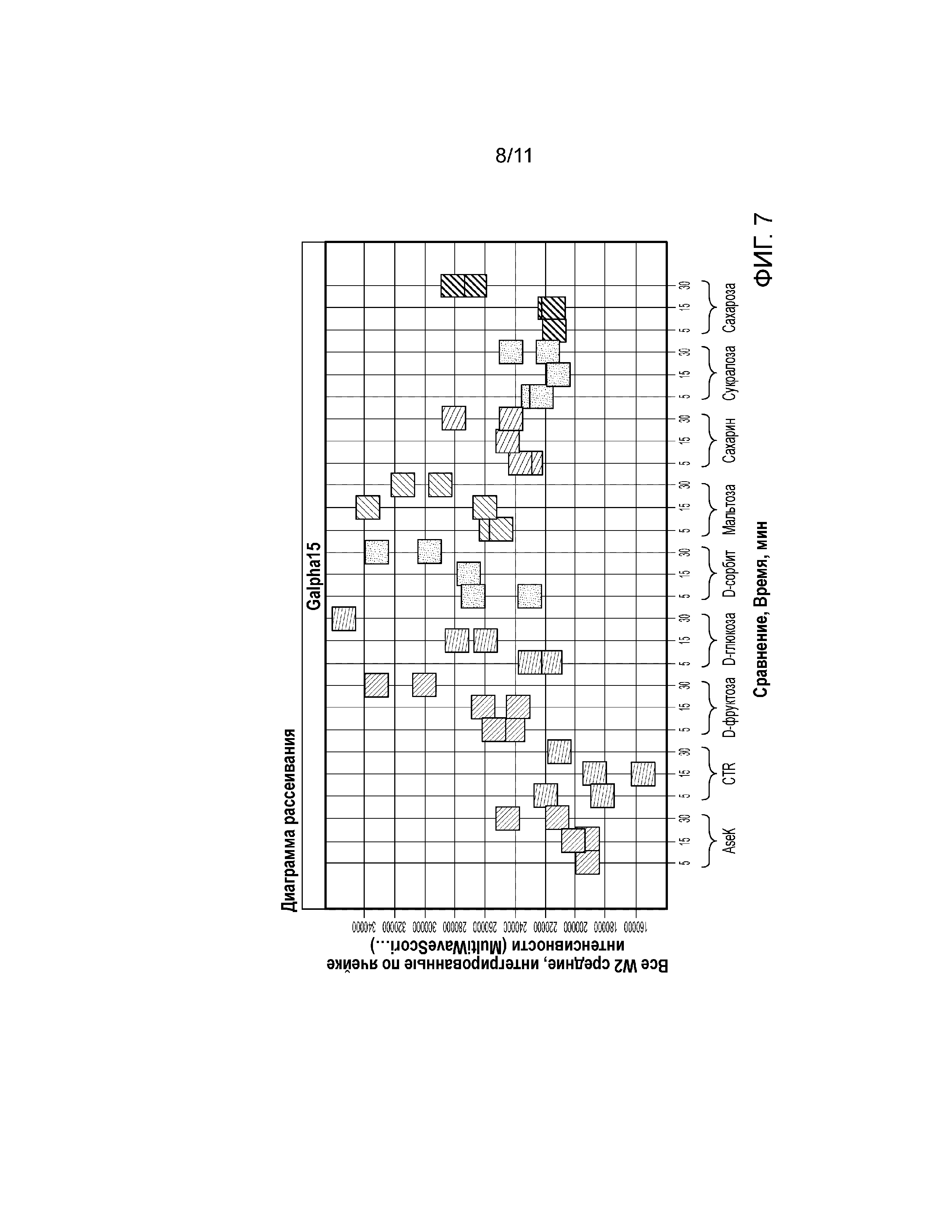
Фиг. 7. График, демонстрирующий активацию pERK1/2 при обработке различными сахарами в клетках U2-OS, трансфецированных Gα15 (6 мкл/40000 клеток).
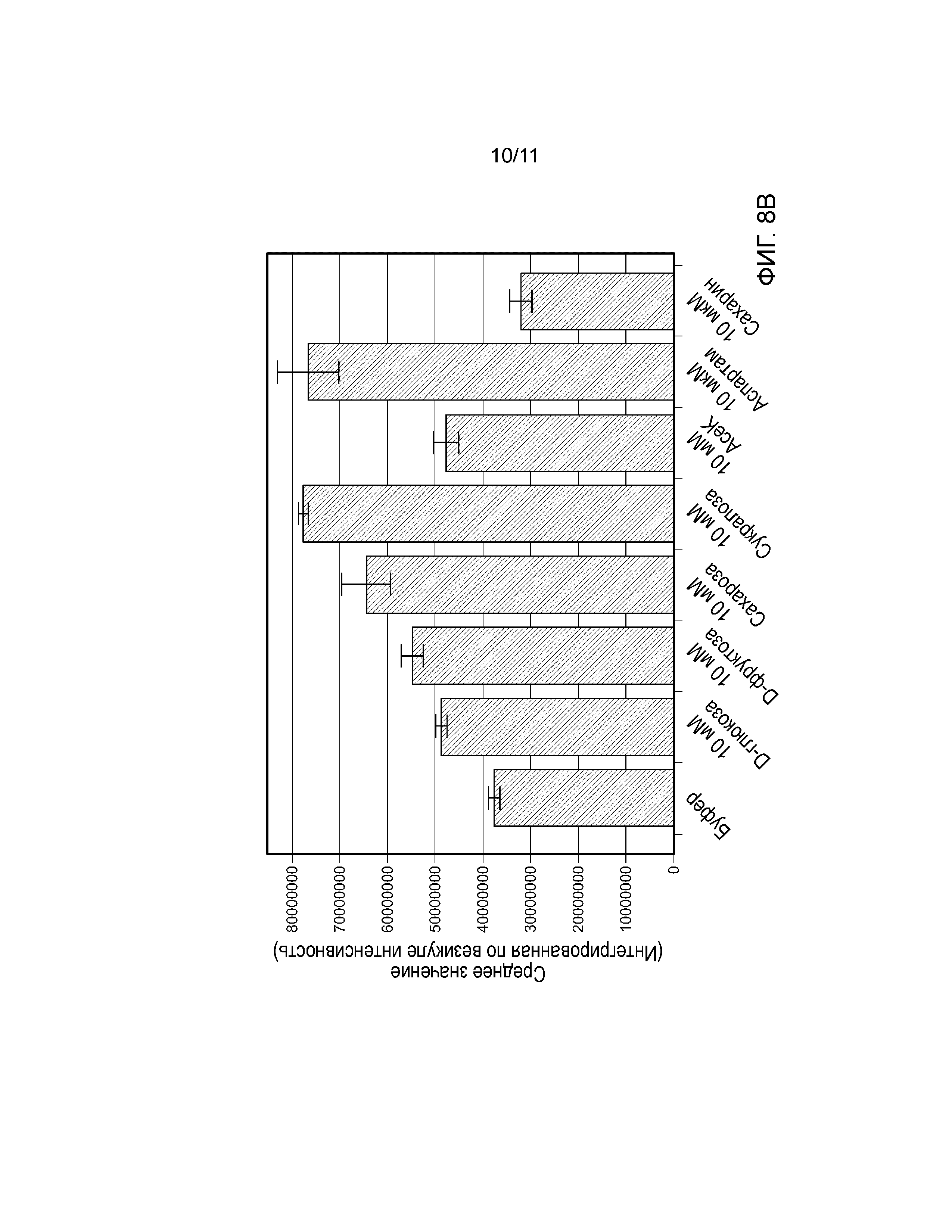
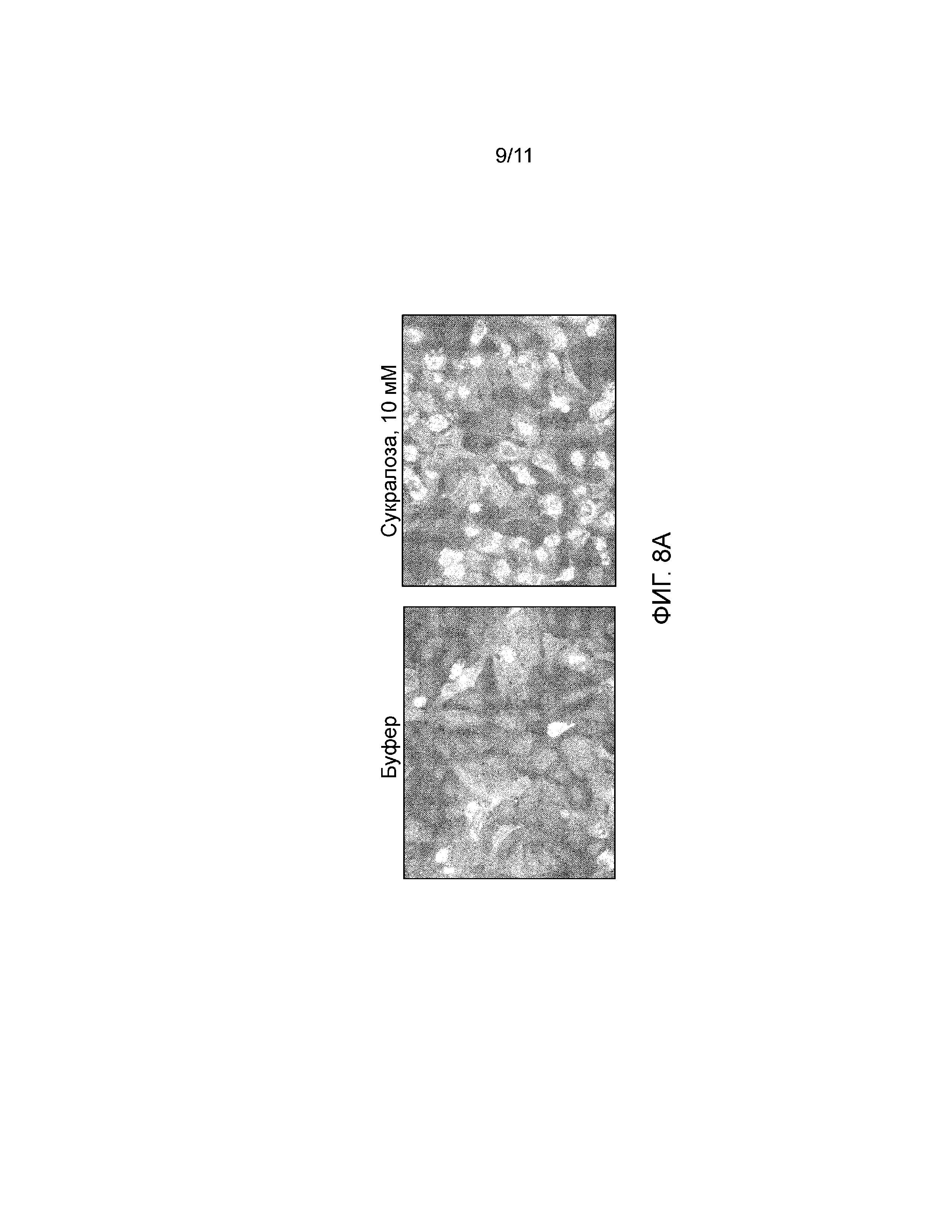
Фиг. 8A-B. Тест Transfluor. Фиг. 8A, Образование зеленых флуоресцентных везикул в клетках Transfluor U2-OS, трансфецированных BacMam, кодирующим Gα15. Ядра подвергают контрастному синему окрашиванию с использованием Hoechst 33342. Фиг. 8B, Количественный анализ интернализации T1R2/T1R3 в модели Transfluor (средние значения ±SD).
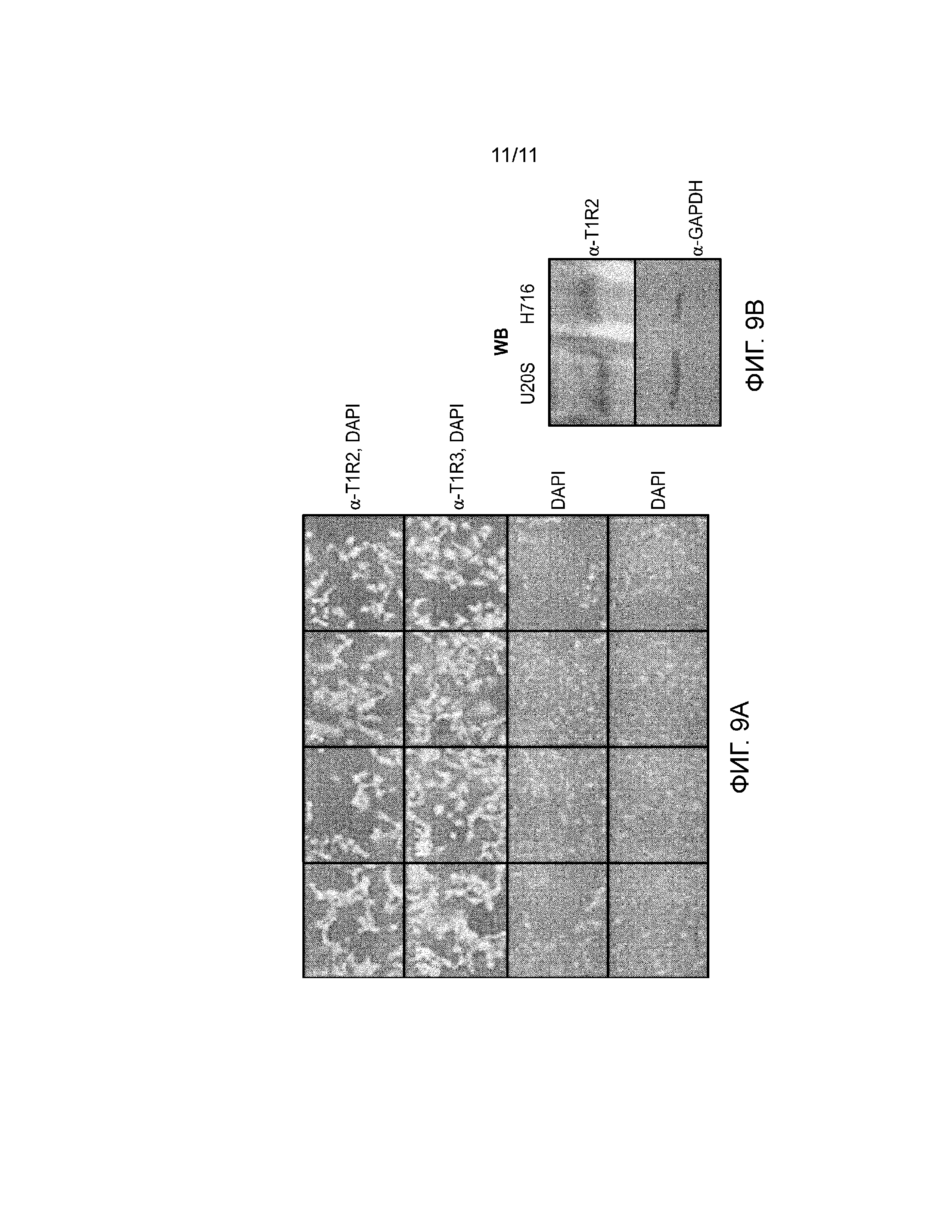
Фиг. 9А-В. Клетки U2-OS экспрессируют эндогенный рецептор сладкого вкуса T1R2/T1R3. Фиг. 9А, клетки U2-OS фиксировали и обрабатывали с целью непрямой иммунофлуоресценции антителами против рецептора сладкого вкуса T1R2 и T1R3. Все изображения были получены с использованием 20-х объектива, используя автоматизированный микроскоп ImageXpress Micro. Окрашивание с использованием Hoechst 33342 синего псевдоцвета, а специфическое окрашивание антителами зеленого псевдоцвета. Фиг. 9В. Вестерн-блоттинг T1R2 в клетках U2-OS и NCI-H716. Клетки лизировали в буфере RIPA (50 мМ Tris-HCl, рН 7,5, 150 мМ NaCl, 1% NP-40, 5 мМ EDTA, 0,1% SDS, и 10 мкг/мл ингибиторов протеиназы). После циклов замораживания-оттаивания буфер для загрузки добавляли в конечной концентрации 2х, образцы нагревали при 95°С в течение 10 минут и центрифугировали в микроцентрифуге в течение 10 мин на самой высокой скорости при комнатной температуре. Супернатант наносили на гель для анализа методом иммуноблоттинга. Первые антитела для T1R2 и GAPDH были от ThermoFisher.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Коэкспрессия T1R2 и T1R3 приводит к вкусовому рецептору, который отвечает на стимулы ощущений сладкого вкуса, в том числе встречающиеся в природе и искусственные подсластители. Лиганды со сладким вкусом связываются с рецептором T1R2/T1R3 и активируют преобразование сигнала в пути с участием G-белка, который включает интернализацию рецептора и внутриклеточную мобилизацию кальция, а также индукцию последующих мишеней, такую как фосфорилирование ERK1/2 (Jang et al., "Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1," Proc Natl Acad Sci U S A. 2007 Sep 18; 104(38): 15069-74).
Любая клетка, которая включает или может быть сконструирована так, что она включает функциональный рецептор сладкого вкуса, может использоваться в описанном анализе. В некоторых вариантах осуществления клетки экспрессируют один или более рецепторов сладкого вкуса. В некоторых вариантах осуществления клетки генетически модифицированы для экспрессии или сверхэкспрессии рецепторов сладкого вкуса. Например, клетка может быть генетически модифицирована в результате введения экзогенной нуклеиновой кислоты, кодирующей T1R2 и T1R3, или двух отдельных экзогенных нуклеиновых кислот, кодирующих T1R2 и T1R3, соответственно. Аминокислотные последовательности этих белков известны; например, № доступа в GenBank: NP_689418.2 (T1R2) и № доступа в GenBank: NP_689414.1 (T1R3).
Можно сконструировать клетки, которые экспрессируют T2R14, T1R2/T1R3, α-гастдуцин и/или GLUT4, используя хорошо известные в данной области техники методы. GLUT4 описан, в частности, в патенте США с № 7799538 и в приведенных там ссылках. См. патент США с № 8338115 и приведенные там ссылки и Adler et al., Cell, 100, 693-702, 2000 ради описаний α-гастдуцина.
В некоторых вариантах осуществления клетки генетически модифицированы для сверхэкспрессии α-гастдуцина (например, Gα16, Gα15, Gα16gust25, Gα15gust25, Gα16gust44, Gα15gust44, Gα15-i/3-5), что увеличивает чувствительность анализа. SEQ ID NO: 1 представляет собой аминокислотную последовательность Gα15. Нуклеотидные последовательности, которые могут использоваться для экспрессии Gα16 (SEQ ID NO: 2), Gα16gust25 (SEQ ID NO: 3), Gα15 (SEQ ID NO: 4), Gα15gust25 (SEQ ID NO: 5), Gα16gust44 (SEQ ID NO: 6), Gα15gust44 (SEQ ID NO: 7) и Gα15-i/3-5 (SEQ ID NO: 8), приведены в списке последовательностей; однако, любые нуклеотидные последовательности, которые кодируют эти α-гастдуцины, могут использоваться для их экспрессии.
В некоторых вариантах осуществления клетки U2-OS используют, как описано ниже. Клетки U2-OS человека (АТСС каталожный # НТВ-96) экспрессируют функциональный рецептор сладкого вкуса (фиг. 9А-B). Поэтому клетки U2-OS могут использоваться для выявления клеточных ответов на потенциальные вещества, стимулирующие вкусовые ощущения, посредством контактирования клеток U2-OS с тестируемым соединением и измерения активности рецептора сладкого вкуса. Можно использовать множество функциональных анализов, в том числе определение изменений внутриклеточной концентрации кальция (например, анализ мобилизации Са2+ на основе спектрофотометра для чтения планшетов для визуализации флуоресценции, FLIPR), активации pERK1/2 (например, визуализацию высокого процентного содержания, HCI) и интернализации рецепторов (например, Transfluor). Изменение активности рецептора сладкого вкуса благодаря подсластителю в присутствии тестируемого соединения указывает на модуляцию рецептора сладкого вкуса с помощью тестируемого соединения, таким образом идентифицируя модулятор ощущения сладкого вкуса (например, молекулу, которая сама по себе обеспечивает сладкий вкус, молекулу, которая блокирует некоторое количество или весь горький вкус, молекулу, которая усиливает сладкий вкус другой молекулы). Увеличение активности рецептора сладкого вкуса указывает на то, что тестируемое соединение представляет собой модулятор ощущения сладкого вкуса. Модуляторы ощущения сладкого вкуса могут быть включены в различные предметы потребления, в том числе пищевые продукты, напитки и лекарственные препараты.
Другие клетки, которые могут использоваться в описанном анализе, включают, но без ограничения, 1A2, ARH-77, RWPE-1, WI-38, EJM, NCI-H1155, L-1236, NCI-H526, JM1, SHP-77, SNU-878, NCI-H2196, C3A, CA46, SNU-466, KS-1, SNU-738, MOLP-2, HDLM-2, Pfeiffer, HCC-15, клетки Alexander, L-540, KMS-12-BM, JK-1, NCI-H1092, SW 1990, NCI-H1184, SU-DHL-1, Hep 3B2.1-7, P3HR-1, NCI-H2029, SU-DHL-5, SNU-1, MOLP-8, SUP-M2, MONO-MAC-1, SNU-1040, KYM-1, HEC-59, HCC1569, OCI-LY3, Hs 819.T, DU4475, CI-1, S-117, OVCAR-8, SNU-626, HL-60, SUIT-2, T3M-4, RKO, MOR/CPR, DK-MG, GA-10, OCUM-1, HCT-15, HT, MONO-MAC-6, G-402, Toledo, COV362, SU-DHL-8, Daoy, NCI-H1435, LS513, Hs 839.T, Hs 172.T, BT-483, KMS-21BM, AGS, NCI-H2172, LC-l/sq-SF, SNU-201, NUGC-4, SK-HEP-1, SUP-B15, SNU-5, HT-1197, SUP-T1, AMO-1, KU812, AN3 CA, AML-193, VMRC-RCW, HLE, HuH28, Hs 751.T, NCI-H2110, MEG-01, MV-4-11, Hep G2, KYSE-30, KALS-1, BICR 6, RMUG-S, JHH-6, Ki-JK, IST- MES1, HCC-95, HPB-ALL, HSC-3, 697, LOU-NH91, KARPAS-299, GI-1, COLO 792, SK-N-FI, D341 Med, HGC-27, SR-786, COLO-818, MHH-CALL-2, SF126, NCI-H322, A-253, NCI-H1623, MCF7, HCC-44, FU97, OCI-LY-19, Hs 766T, NCI-H522, RL, HCC1428, RPMI 6666, U-937, NCI-H460, SW 1088, NCI-H1792, NCI- H1693, UACC-257, JHUEM-2, HuT 78, UACC-893, NCI-H929, A-704, OV56, LN-229, OE19, SK-MEL-24, RD-ES, NCI-H211, KCI-MOH1, NCI-H1963, Hs 706.T, ChaGo-K-1, EPLC-272H, OPM-2, KHM-1B, A549, HuG1-N, NCI-H508, MHH-CALL-3, SNU-1076, A3/KAW, MEL-HO, TO 175.T, Caki-1, Hs 936.T, SK-LU-1, WM-983B, K-562, EFE-184, SNU-520, NCI-H2291, HCC-1195, ABC-1, KE-39, NH-6, HCC2218, CMK, RS4;11, KYSE-450, OV7, KYSE-510, SK-UT-1, SNU-C1, OE33, P12-ICHIKAWA, DLD-1, COV434, HuNSl, SNU-899, SW480, COLO-678, LU99, KOPN-8, NCI-H2227, SW1463, Hs 675.T, JHH-4, NCI-H1703, HEC-1-A, BDCM, MIA PaCa-2, PC-3, TE-15, PK-45H, MKN-45, HCC-366, CAL-29, HEC-50B, CPC-N, KMRC-20, SW1116, EOL-1, COLO 205, EHEB, YD-38, MC116, SK-N-BE(2), BV-173, NCI-H2347, LU65, RT4, U-87 MG, LK-2, KP-N-YN, HEC-251, NCI-H1651, GP2d, RERF-LC-MS, NB-4, NCI-H2286, SNU-61, T-47D, huH-1, KYSE-180, ST486, SW 1353, M-07e, KASUMI-1, YH-13, NCI-H28, GAMG, JeKo-1, GOS-3, SNU-324, PA-TU-8902, MFE-280, SNU-245, NALM-1, RERF-LC-Sq1, BICR 22, ZR-75-1, COR-L23, SW579, COR-L88, KM12, Hs 611.T, OUMS-23, RERF-LC-Adl, NCI-H1385, SK-LMS-1, COLO-320, BL-70, GRANTA-519, MCAS, Pane 08.13, AM-38, KMS-11, SIG-M5, SNU-407, JHOS-2, OVCAR-4, Set-2, OV-90, MeWo, HEL, HT-29, MDA-MB-231, TOV-21G, NCI-H1355, KMS-27, NALM-6, KMS-26, Caov-4, KASUMI-2, UACC-62, U266B1, Hs 695T, HT55, BICR 31, TCC-PAN2, KMS-20, Hs 578T, RI-1, Hs 606.T, NCI-H1341, THP-1, BCP-1, Hs 737.T, SW1417, MOLT-4, Raji, ESS-1, MEL-JUSO, SH-10-TC, Hs 683, ME-1, EB2, PLC/PRF/5, NCI-H1339, A4/Fuk, SEM, HEC-265, IST-MES2, KE-97, NCI-H1437, COLO-704, NCI-H1915, TE-5, NCI-H2023, NCI-H82, T1-73, SNU-840, HuT 102, NCI-H1944, KYSE-520, Kasumi-6, 1321N1, Hs 742.T, IM95, PL45, CL-40, WM1799, KMM-1, SNU-449, JHUEM-1, KARPAS-620, Loucy, SNU-1079, Daudi, HCC-56, HSC-2, COR-L47, PA-TU-8988S, OAW28, COR-L311, L-363, Malme-3M, NOMO-1, Hs 870.T, SU-DHL-10, Hs 229.T, NCI-H810, KYSE-410, RPMI-8402, SNU-175, EBC-1, RVH-421, K029AX, PA-TU-8988T, LXF-289, OVSAHO, CAL-12T, Hs 940.T, MM1-S, SUP-HD1, LNCaP клон FGC, HSC-4, NU-DHL-1, NCI-H2228, BEN, CAL-78, Sq-1, NCI-H1793, SNU-C2A, MDA-MB-134-VI, COV318, KE-37, TYK-nu, MOTN-1, T98G, SW837, EB1, Becker, PE/CA-PJ34 (клон C12), Hs 616.T, NCI-H446, WM-88, CHP-126, Calu-1, SNU-283, NCI-HI 573, SW 1271, SNU-16, JHOS-4, ACHN, Calu-3, KMRC-1, SW 1783, TE-11, TE-9, HuH-6, P31/FUJ, HT-1376, NCI-H520, 786-O, KNS-60, Caki-2, OVK18, PL-21, NCI-H2452, JURL-MK1, TEN, JHH-7, MDA-MB-157, Calu-6, RKN, NUGC-2, ONS-76, J82, OUMS-27, SNU-1196, Hs 739.T, RPMI-7951, NCI-H854, JHH-5, JVM-2, Hey-A8, 5637, KYSE-140, Capan-2, KYSE-150, HEC-1-B, BICR 16, HEL 92.1.7, MHH-NB-11, SNU-387, SK-OV-3, SK-MEL-28, IGROV1, ML-1, HLF-a, CHL-1, YKGl, A-204, OCI-M1, 8505C, JVM-3, NCI-H647, DB, COLO-800, PK-59, FaDu, HLF, OVMANA, EFO-27, PF-382, NCI-H747, LS123, SU-DHL-6, SJRH30, PANC-1, NCI-H2342, KM-H2, DND-41, HH, HuCCT1, F-36P, DMS 454, Hs 274.T, AU565, NCI-H1666, EN, RH-41, NCI-H1373, NCI-H838, SK-MEL-30, MOLM-6, DEL, NCI-H226, NCI-H1648, NCI-H661, 143B, Mino, C32, KMS-34, NCI-H1694, SK-ES-1, UACC-812, GDM-1, NCI-H23, Panc 02.03, CCF-STTG1, LOX IMVI, SJSA-1, MDST8, PK-1, NCI-H716, SU-DHL-4, MPP 89, MJ, COLO 829, PE/CA-PJ15, HD-MY-Z, BxPC-3, WM-793, COLO 668, T84, JHOM-1, PEER, LS411N, GMS-10, KMBC-2, RMG-I, KELLY, SNU-761, NALM-19, HEC-151, G-361, OVTOKO, A-498, SW 900, LCLC-103H, FTC-133, QGP-1, Reh, CMK-11-5, NU- DUL-1, BT-20, Hs 600.T, Hs 604.T, KATO III, SNU-410, NCI-H2126, SK-MEL-5, MDA-MB-468, AsPC-1, HUP-T3, KP-N-SI9s, L-428, SNU-1105, HUP-T4, 769-P, LMSU, NCI-H1869, NCO2, MOLM-16, CAL 27, HCC70, NCI-H1930, COV644, Hs 863.T, HCC-2279, D283 Med, Hs 944.T, HCC1599, MDA-MB-415, HCC2157, NCI-H1618, SNU-308, HCC1954, DMS 153, HPAF-II, T24, CJM, VM-CUB1, UM-UC-3, LAMA-84, NCI-H1734, JHH-2, VMRC-RCZ, MFE-319, MDA-MB-453, SNU-503, TOV-112D, B-CPAP, GSU, HCC-78, NCI-H2171, CAMA-1, HEC-108, HCC4006, CAL-85-1, NCI-H2122, COLO-699, NCI-H196, LUDLU-1, SW 780, RPMI 8226, LP-1, PC-14, HuTu 80, T.T, SW948, 22Rvl, HARA, NCI-H596, IPC-298, SCaBER, NCI-H1838, NB-1, Hs 934.T, Hs 895.T, DMS 114, KYSE-70, KP-3, KP4, DAN-G, NCI-H2009, OC 316, SCC-25, U-138 MG, RCC10RGB, MFE-296, NCI-H1755, RERF-LC-KJ, 8305C, WSU-DLCL2, ES-2, MSTO-211H, SCC-15, ZR-75-30, PSN1, SNU-423, NCI-H2106, TE-1, UT-7, KMS-28BM, NCI-H2081, SK-MM-2, COLO 741, OC 314, HCC1395, MOLT-13, LN-18, Pane 10.05, PE/CA-PJ41 (клон D2), Hs 746T, CW-2, SKM-1, NUGC-3, TE-10, NCI-H358, NCI-H69, BFTC-909, HOS, BICR 18, NCI-H1395, OVKATE, Hs 698.T, EFM-19, COLO-783, MHH-CALL-4, ACC-MESO-1, NCI-H1436, KP-N-RT-BM-1, SK-MEL-31, NCI-H1105, CAL-51, YD-15, NCI-H2085, NCI-H2444, HCC1187, Hs 939.T, CAL-120, SCC-9, TUHR14TKB, KMRC-2, KG-l-C, ECC10, CGTH-W-1, NCI-H841, C2BBel, SUP-T11, RCH-ACV, CADO-ES1, JURKAT, 647-V, SK-MEL-2, MDA-MB-175-VII, MKN74, SNU-C4, LCLC-97TM1, SCC-4, BHY, IGR-37, KYO-1, Hs 281.T, TT, TUHR4TKB, HT-1080, NCI-H660, TE 441.T, LS1034, KNS-42, Pane 04.03, HCC1419, AZ-521, SNG-M, NCI-N87, G-292, клон A141B1, KPL-1, MDA-MB-361, CL-14, NCI-H2170, HuH-7, RD, NCI-H2066, IGR-1, TE-14, VCaP, BL-41, SNU-620, SK-MES-1, MEC-2, NCI-H1299, IGR-39, RT112/84, SF-295, DV-90, A2780, BICR 56, NCI-H510, NCI-H2141, YD-8, NCI-H2405, TF-1, MEC-1, CCK-81, NCI-H1048, Hs 822.T, NCI-H2052, KO52, CAL-54, Hs 840.T, SW620, SK-CO-1, BT-474, CL-11, KNS-62, NCI-H1650, G-401, MOLT-16, SNU-398, COLO-680N, EM-2, Hs 294T, CAL-62, KMRC-3, A101D, KG-1, BT-549, HT115, A-375, SW-1710, WM-115, KLE, JHUEM-3, MKN7, CHP-212, HCC202, BC-3C, NCI-H1568, KMS-18, PE/CA-PJ49, COLO-849, SIMA, OCI-AML3, GSS, EC-GI-10, EFO-21, RCM-1, DMS 273, KU-19-19, RERF-GC-1B, SH-4, SK-MEL-3, RERF-LC-Ad2, M059K, JHOM-2B, MDA PCa 2b, Hs 852.T, RL95-2, Panc 03.27, SNU-216, Pane 02.13, CFPAC-1, SK-N-SH, OCI-AML2, LoVo, SBC-5, NCI-H1876, NCI-H441, SK-N-AS, COR-L24, HCC38, NCI-H1781, DOHH-2, NCI-H1563, U-251 MG, HPAC, JIMT-1, U-2 OS, A-673, TC-71, NCI-H650, NIH:OVCAR-3, CAS-1, JL-1, SK-MEL-1, MDA-MB-435S, Ishikawa (Heraklio) 02 ER-, TE 617.T, SU.86.86, RERF-LC-AI, TT2609-C02, LS 180, YAPC, HDQ-P1, KNS-81, FU-OV-1, KP-2, DMS 53, SNU-1272, Detroit 562, 42-MG-BA, L3.3, COLO-679, NCI-H2087, NCI- H2030, GCT, NCI-H889, Caov-3, MDA-MB-436, NCI-H524, MKN1, KCL-22, Capan-1, CML-T1, H4, NCI-H727, Hs 343.T, MHH-ES-1, NMC-G1, HCC-1171, REC-1, Hs 618.T, A172, YD-10B, SW48, MUTZ-5, TE-6, JHH-1, HCT 116, TE-4, IA-LM, MG-63, NCI-H1975, TALL-1, HCC1806, HMCB, SCLC-21H, HCC1500, CL-34, Panc 05.04, SW403, TM-31, HCC1937, JMSU-1, DMS 79, SNB-19, NCI-H1836, Li-7, HCC827, 639-V, MOLM-13, SK-BR-3, IMR-32, TUHR10TKB, OAW42, SK-N-MC, TGBC11TKB, NCI-H1581, EFM-192A, YMB-1, HCC2935, ECC12, HCC-33, DU 145, NCI-H146, SNU-1214, SNU-1077, 23132/87, HT-144, SNU-182, Hs 888.T, SNU-475, GCIY, Hs 729, JHOC-5, SW 1573, HEC-6, OCI-AML5, Hs 688(A).T, Hs 821.T, PCM6, RT-112, SK-N-DZ, SNU-478, SNU-119, HCC1143, NCI-H209, 8-MG-BA, COR-L105, COR-L95, SNU-46, COV504, CAL-148, SNU-C5, DBTRG-05MG, BHT-101, WM-266-4, BFTC-905, KYSE-270, TE-8, SNU-213 и SH-SY5Y.
Тестируемые соединения
Тестируемые соединения могут быть встречающимися в природе или синтетически полученными. Белки, полипептиды, пептиды, полисахариды, небольшие молекулы и неорганические соли являются примерами тестируемых соединений, которые могут быть подвергнуты скринингу с использованием описанных здесь методов.
Любая молекула нуклеиновой кислоты, которая кодирует эти белки, может быть введена в клетку. Нуклеиновая кислота может быть введена в клетку стабильно или транзиторно. Экзогенная нуклеиновая кислота может находиться в конструкции или векторе, который включает промотор, который функционально связан с кодирующей частью нуклеиновой кислоты.
Клетка(и) может выращиваться на соответствующем субстрате, таком как многолуночный планшет, чашка для культивирования ткани, колба и т.д.; см. примеры 1 и 2, ниже.
Методы скрининга
В некоторых вариантах осуществления способы идентификации модулятора ощущения сладкого вкуса включают контактирование клетки с тестируемым соединением и измерение активности рецептора сладкого вкуса. Изменение активности рецептора сладкого вкуса с помощью тестируемого соединения указывает на модуляцию рецептора сладкого вкуса с помощью тестируемого соединения, таким образом идентифицируя тестируемое соединение в качестве модулятора ощущения сладкого вкуса.
Активность рецептора сладкого вкуса можно определить количественно или качественно любым способом, который является общепринятым в данной области, включая анализы на активность связанных с G-белком рецепторов, изменения уровня вторичного мессенджера в клетке, образование инозитолтрифосфата (IP3) через гидролиз с использованием фосфолипазы С фосфатидилинозитола, изменения цитоплазматических уровней ионов кальция, привлечение β-аррестина и т.п. Примеры таких анализов приведены в конкретных примерах ниже.
Активность связывания также может использоваться для измерения активности вкусового рецептора, например, с помощью анализа конкурентного связывания или поверхностного плазмонного резонанса. Также может быть измерена интернализация рецептора и/или десенсибилизация рецептора, как известно в данной области техники. Зависимая от рецептора транскрипция гена может быть также измерена для оценки активности вкусового рецептора. Уровень транскрипции может быть измерен с помощью любого способа, известного квалифицированным в данной области техники специалистам. Например, экспрессия с мРНК представляющего интерес белка может быть выявлена, используя методы ПЦР, микрочип или Нозерн-блоттинг. Количество полипептида, продуцированного с мРНК, может быть определено с помощью методов, которые являются общепринятыми в данной области для количественного анализа белков в клетке, например, Вестерн-блоттинга, ELISA, ELISPOT, иммунопреципитации, иммунофлуоресценции (например, FACS), иммуногистохимии, иммуноцитохимии и т.д., а также любого другого метода, который известен в настоящее время или разработан позже для количественного анализа белка в клетке или белка, продуцированного клеткой.
Физические изменения клетки также могут быть определены, например, посредством оценки с помощью микроскопа размера, формы, плотности или любого другого физического изменения, опосредованного активацией вкусового рецептора. Проточная цитометрия также может использоваться для оценки физических изменений и/или определения присутствия или отсутствия клеточных маркеров.
В некоторых вариантах осуществления активность рецептора сладкого вкуса измеряют посредством определения уровня внутриклеточного вторичного мессенджера в клетке (например, цАМФ, цАМФ, цГМФ, NO, CO, H2S цГМФ, DAG, IP3). В некоторых вариантах осуществления активность рецептора сладкого вкуса измеряют посредством определения уровня внутриклеточного кальция. В некоторых вариантах осуществления активностью рецептора сладкого вкуса является активность связывания.
В любом из этих вариантов осуществления клетка может быть модифицирована для сверхэкспрессии рецептора сладкого вкуса и/или Gα15, как описано ниже.
Квалифицированным в данной области техники специалистам будет понятно, что существуют многочисленные вариации и перестановки вышеописанных вариантов осуществления, которые подпадают под объем прилагаемой формулы изобретения.
ПРИМЕР 1
Создание линий клеток JUMP-IN™ T-REX™ U2-OS и таргетируемых линий клеток JUMP-IN™ U2-OS
Создание линий клеток JUMP-IN™ T-REX™ U2-OS. Клетки JUMP-IN™ U2-OS трансфецировали вектором pLenti TR puro, используя реагент LIPOFECTAMINE™ LTX (Life Technologies # 15338-100). Трансфецированные клетки отбирали с использованием 1 мкг пуромицина (Life Technologies # А113803), и не трансфецированные родительские клетки использовали в качестве отрицательного контроля при отборе с использованием антибиотика.
Отобранные клоны были размножены и трансфецированы вектором pJTI™-R4 EXP CMV TO GFP pA (Life Technologies). Затем клетки проверяли на тетрациклиновую (Tet) репрессию и Tet-индуцибельность с использованием или без 1 мкг доксициклина, и уровни экспрессии GFP количественно определяли с использованием микропланшет-ридера Safire II.
Восемьдесят четыре клона выжили после сортинга с использованием FACS. Каждый клон высевали в трех повторах в 96-луночные планшеты. Одну лунку использовали с целью сохранения, а две другие лунки использовали для транзиторной трансфекции экспрессионным вектором pJTI™ R4 EXP CMV-TO pA (Life Technologies), кодирующим зеленый флуоресцентный белок (GFP), в течение 24 часов. Затем клетки обрабатывали или не обрабатывали 1 мкг доксициклина в течение 24 часов с последующим количественным анализом GFP. Коэффициент ответа рассчитывали как (показание в лунке с доксициклиом)/(показание в лунке без доксициклина).
Двенадцать клонов JUMP-IN™ T-REX™ U2-OS с коэффициентом ответа свыше 2 были в дальнейшем размножены. Пять из этих клонов были котрансфецированы с использованием конструкции для интегразы R4 и вектора pJTI™-R4 EXP CMV-TO GFP (при условии индуцируемой экспрессии GFP). Клетки, котрансфецированные конструкцией CMV-GFP (при условии конститутивной экспрессии GFP) и конструкцией для интегразы R4, использовали в качестве положительного контроля для трансфекции и отбора с использованием антибиотика. После отбора с использованием 5 мкг/мл бластицидина в течение 28 дней, клетки, которые были ретаргетированы с использованием вектора pJTI™-R4 EXP CMV-TO GFP, были индуцированы с использованием 1 мкг/мл доксициклина в течение 20 часов, а затем проанализированы с помощью FACS.
Создание таргетируемых линий клеток JUMP-IN™ U2-OS. Таргетируемые линии клеток JUMP-IN™ U2-OS были созданы, используя набор JUMP-IN™ TI™ Platform (Life Technologies). Вектор PJI™ PhiC31 Int котрансфецировали в клетки-хозяева U2-OS, используя таргетирующий вектор pJTI™/Blasticidin, который содержит ATT-сайты, которые комплементарны псевдо-ATT-сайтам в геноме млекопитающего-хозяина. Гомологичной рекомбинации между АТТ-сайтами вектора R4 и генома хозяина содействует интеграза phiC31. Таргетирующий вектор PJI™ содержит ген устойчивости к гигромицину, который делает возможным отбор клеток, содержащих стабильные геномные интеграции. Эти клетки затем клонировали при помощи проточной цитометрии и исследовали для определения количества сайтов R4, присутствующих в геноме клетки-хозяина, с помощью анализа с использованием блоттинга по Саузерну.
Клоны с валидизированными одиночными сайтами интеграции R4 были валидизированы для ретаргетирования путем трансфекции pJTI™-R4 EXP CMV-TO-EmGFP-pA и кодирующими интегразу векторами JTI™ R4, с последующим отбором с использованием антибиотиков в течение 4 недель. Клоны высевали при 60-80% конфлюентности в 6-луночную чашку в среду для роста без антибиотиков (McCoy’s 5A плюс 10% подвергнутой диализу фетальной телячьей сыворотки, HEPES, несущественные аминокислоты и пируват натрия) и трансфецировали с использованием соотношения 1:1 pJTI™-R4 EXP CMV-TO-EmGFP-pA и кодирующих интегразу векторов JTI™ R4 (2,5 мкг тотальной ДНК) с использованием LIPOFECTAMINE™ LTX (6,25 мкл) и реагента PLUS™ (2,5 мкл). Положительный контроль CMV-GFP вводили в клетки параллельно. После инкубации в течение ночи клетки отбирали в течение 28 дней с использованием 5 мкг/мл бластицидина в среде для роста (McCoy’s 5A плюс 10% подвергнутой диализу FBS, HEPES, несущественные аминокислоты, пируват натрия и пенициллин/стрептомицин). Через 28 дней отбора с использованием антибиотика отобранные пулы были проанализированы с помощью FACS.
Пять клонов были выбраны для проверки эффективности ретаргетирования и возможности индукции. Все клоны были успешно ретаргетированы. Два клона с наибольшей эффективностью ретаргетирования были, кроме того, проверены на возможность индукции с помощью доксициклина. Один клон продемонстрировал ~88% GFP+ клеток в GFP-ретаргетированных контрольных клетках и ответил очень хорошо на индукцию с использованием доксициклина, при этом >99% GFP-позитивной популяции было в индуцированной культуре, и лишь ~4% GFP-позитивных клеток было в неиндуцированной культуре. Другой клон также хорошо ответил на индукцию с использованием доксициклина, при этом >99% GFP-позитивной популяции было в индуцированной культуре, и лишь ~8% GFP-позитивных клеток было в неиндуцированной культуре; при этом 64% клеток были GFP-позитивными в GFP-ретаргетированных контрольных клетках.
Среда для культивирования клеток. В таблице ниже перечислены компоненты среды, используемой для культивирования клеток Jump-IN™ T-REX™ U2-OS (Life Technologies):http://patentimages.storage.googleapis.com/WO2014183044A1/imgf000012_0001.png
|
ПРИМЕР 2
Методы
Клеточная культура и обработка соединениями. Клетки U2-OS выращивали в среде McCoy с добавлением 10% фетальной телячьей сыворотки. Клетки высевали при плотности 5000 клеток/лунку в 384-луночные планшеты с PDL-покрытием.
Трансфекция BacMam. Клетки U2-OS транзиторно трансфецировали с использованием 0,5-12 мкл BacMam, кодирующего G-белок, гастдуцин (Gα15, Gα16gust44), на 40000 клеток в течение 24 часов или 48 часов. Трансфецированные клетки высевали при плотности 5000 клеток/лунку в 384-луночные планшеты с PDL-покрытием.
Функциональная экспрессия. Клетки JUMP-IN™ T-REX™ U2-OS трансфецировали pJTI™R4EXP-CMV-TO-T1R3/T1R2/Gα15-pA и кодирующими интегразу векторами JTI™ R4 (Life Technology). Доксициклин-индуцированную экспрессию T1R3/T1R2/Gα15 выявляли с помощью ОТ-ПЦР.
Анализ pERK1/2. Для исследований pERK1/2 среду для роста заменяли средой без сыворотки на 2 ч. Клетки U2-OS обрабатывали 100 мМ соединений со сладким вкусом с или без U73122 (10 мкМ) и с лактизолем или без него (20 мМ). Контрольные группы клеток также получали DMSO (0,1%) в среде. Зафиксированные клетки окрашивали кроличьими антителами против фосфо-р44/р42 МАРК (pERK1/2) от Cell Signaling Technologies, а затем вторыми антителами, конъюгированными с Alexa 488 (Life Technology), и Hoechst 33342. Изображения были получены на ImageXpressMicro (MDC) и проанализированы с использованием MWCS модуля.
Анализ кальция. Следовые количества кальция в Fluo-4АМ-нагруженных клетках регистрировании в течение продолжительного периода в FLIPRTetra (MDC) с λвозбуждения=470-495 нм и λэмиссии=515-575 нм. Максимальное увеличение флуоресценции по сравнению с исходным уровнем было определено и представлено в виде относительных единиц флуоресценции (RFU).
Модель Transfluor. Клетки Transfluor U2-OS, стабильно экспрессирующие β-аррестин-GFP, были транзиторно трансфецированы вкусовым G-белком, гастдуцином (Gα15 или Gα16gust44) (Life Technologies), или были транзиторно трансфецированы рецептором сладкого вкуса T1R2/T1R3 и вкусовым G-белком. Обработка клеток соединениями со сладким вкусом вызывала образование флуоресцентных везикул. Интернализацию рецепторов сладкого вкуса количественно анализировали с помощью Transfluor Application Module в программном обеспечении MetaXpress (MDC).
ПРИМЕР 3
Фосфорилирование ERK1/2
Фосфорилирование ERK1/2 является обычным последующим событием после активации GPCR с помощью пути PLCβ2-IP3. Среду для роста заменяли забуференным фосфатом солевым раствором Дульбекко (DPBS) без сыворотки на 2 ч. Клетки U2-OS обрабатывали 100 мМ D-глюкозы с добавлением или без 20 мМ лактизоля в течение 5, 15 и 30 минут, а затем фиксировали и окрашивали антителом против ERK1/2 и Hoechst 33342 до визуализации. Экспрессию pERK1/2 количественно анализировали, используя алгоритм для анализа Multiwaves Cell Scoring Molecular Devices. Для каждой точки измерения, среднее значение для средней, интегрированной по ячейке интенсивности и среднеквадратическое отклонение рассчитывают, исходя из точек измерений в четырех повторах. D-глюкоза индуцировала активацию pERK1/2 зависимым от времени образом в родительских клетках U2-OS (фиг. 4, слева) и в клетках U2-OS, сверхэкспрессирующих Gα15 (фиг. 5, слева). Важно отметить, что активность pERK1/2 блокировал антагонист рецептора сладкого вкуса, лактизоль (фиг. 4, справа), или ингибитор PLCβ2 U73122 (фиг. 5, справа).
ПРИМЕР 4
Сверхэкспрессия Gα15
Сверхэкспрессия Gα15 в клетке U2-OS значительно увеличивает передачу сигналов в ответ на модулятор ощущения сладкого вкуса. Сукралоза индуцировала Са(2+) ответ в линии клеток U2-OS, трансфецированной Gα15, в зависимости от концентрации (фиг. 2). Авторы настоящего изобретения наблюдали наибольший Ca(2+) ответ в клетках U2-OS, трансфецированных с использованием 12 мкл/40000 клеток (фиг. 2). Трансфекция клеток U2-OS с использованием 6 мкл/40000 клеток значительно увеличила Ca(2+) ответ после обработки природными сахарами, такими как глюкоза, фруктоза и сахароза (фиг. 3). Кроме того, сверхэкспрессия Gα15 увеличивала активность pERK1/2 при обработке глюкозой или мальтозой в клетках U2-OS (фиг. 6). Важно отметить, что активность pERK1/2 блокировал ингибитор PLCβ2 U73122 (фиг. 7).
ПРИМЕР 5
С(+2) ответ
Авторы настоящего изобретения обнаружили, что как моносахариды, так и дисахариды индуцировали зависимый от концентрации Са(2+) ответ в линии клеток JUMP-IN™ T-REX™ U2-OS с доксициклин-индуцируемой экспрессией рецептора сладкого вкуса T1R2/T1R3/Gα15 (фиг. 1А). T1R2/T1R3-опосредованное высвобождение Ca(2+) ингибировал лактизоль (фиг. 1B). Кроме того, U73122 отменял высвобождение Ca(2+) при обработке D-глюкозой и D-фруктозой и не оказывал никакого влияния на Ca(2+) ответ после стимуляции с использованием сахарозы и сукралозы (фиг. 1B).
В совокупности эти данные показывают взаимосвязь между молекулярной структуры сахаров и рецептором сладкого вкуса T1R2/T1R3.
ПРИМЕР 6
Искусственные подсластители
Некоторые поразительные наблюдения были сделаны в случае искусственных подсластителей, Асе-К, аспартама и сахарина. Асе-К и аспартам не индуцировали T1R2/T1R3-опосредованный Са(2+) ответ в клетках JUMP-IN™ T-REX™ U2-OS (фиг. 1А). T1R2/T1R3-независимое высвобождение Са(2+) имело место при обработке сахарином (фиг. 1А), указывая на то, что мишенью искусственных подсластителей Асе-К, аспартама и сахарина является общий рецептор, который должен еще быть идентифицирован.
ПРИМЕР 7
Эффекты соединений со сладким вкусом на привлечение β-аррестина к рецептору сладкого вкуса.
Несколько линий доказательств говорят о том, что лиганды GPCR селективно активируют путь передачи сигналов с участием β-аррестина. Для оценки эффектов соединений со сладким вкусом на привлечение β-аррестина к рецептору сладкого вкуса, авторы настоящего изобретения использовали модель Transfluor (MDC), которая является очень чувствительной к перераспределению β-аррестина. Обработка клеток соединениями со сладким вкусом вызывала образование флуоресцентных везикул, что указывает на интернализацию комплексов T1R2/T1R3-β-аррестин в эндосомы (фиг. 8А). Интернализацию рецепторов сладкого вкуса количественно анализировали с помощью Transfluor Application Module в программном обеспечении MetaXpress (MDC). D-глюкоза, D-фруктоза и Асе-К немного увеличивали интернализацию рецептора сладкого вкуса, тогда как сахароза, сукралоза и аспартам оказывали сильный эффект на интернализацию T1R2/T1R3 (фиг. 8B). В противоположность этому, даже минимальная интернализация не происходила при обработке сахарином (фиг. 8B).
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Pepsico
<120> Клетки в качестве модели для идентификации возможных
модуляторов вкусовых ощущений
<130> 056943.012321
<150> US 61/821,943
<151> 2013-05-10
<160> 8
<170> FastSEQ for Windows Version 4.0
<210> 1
<211> 372
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Ala Arg Ser Leu Thr Trp Gly Cys Cys Pro Trp Cys Leu Thr Glu
1 5 10 15
Glu Glu Lys Thr Ala Ala Arg Ile Asp Gln Glu Ile Asn Arg Ile Leu
20 25 30
Leu Glu Gln Lys Lys Gln Glu Arg Glu Glu Leu Lys Leu Leu Leu Leu
35 40 45
Gly Pro Gly Glu Ser Gly Lys Ser Thr Phe Ile Lys Gln Met Arg Ile
50 55 60
Ile His Gly Val Gly Tyr Ser Glu Glu Asp Arg Arg Ala Phe Arg Leu
65 70 75 80
Leu Ile Tyr Gln Asn Ile Phe Val Ser Met Gln Ala Met Ile Asp Ala
85 90 95
Met Asp Arg Leu Gln Ile Pro Phe Ser Arg Pro Asp Ser Lys Gln His
100 105 110
Ala Ser Leu Val Met Thr Gln Asp Pro Tyr Lys Val Ser Thr Phe Glu
115 120 125
Lys Pro Tyr Ala Val Ala Met Gln Tyr Leu Trp Arg Asp Ala Gly Ile
130 135 140
Arg Ala Cys Tyr Glu Arg Arg Arg Glu Phe His Leu Leu Asp Ser Ala
145 150 155 160
Val Tyr Tyr Leu Ser His Leu Glu Arg Ile Ser Glu Asp Ser Tyr Ile
165 170 175
Pro Thr Ala Gln Asp Val Leu Arg Ser Arg Met Pro Thr Thr Gly Ile
180 185 190
Asn Glu Tyr Cys Phe Ser Val Lys Lys Thr Lys Leu Arg Ile Val Asp
195 200 205
Val Gly Gly Gln Arg Ser Glu Arg Arg Lys Trp Ile His Cys Phe Glu
210 215 220
Asn Val Ile Ala Leu Ile Tyr Leu Ala Ser Leu Ser Glu Tyr Asp Gln
225 230 235 240
Cys Leu Glu Glu Asn Asp Gln Glu Asn Arg Met Glu Glu Ser Leu Ala
245 250 255
Leu Phe Ser Thr Ile Leu Glu Leu Pro Trp Phe Lys Ser Thr Ser Val
260 265 270
Ile Leu Phe Leu Asn Lys Thr Asp Ile Leu Glu Asp Lys Ile His Thr
275 280 285
Ser His Leu Ala Thr Tyr Phe Pro Ser Phe Gln Gly Pro Arg Arg Asp
290 295 300
Ala Glu Ala Ala Lys Ser Phe Ile Leu Asp Met Tyr Ala Arg Val Tyr
305 310 315 320
Ala Ser Cys Ala Glu Pro Gln Asp Gly Gly Arg Lys Gly Ser Arg Ala
325 330 335
Arg Arg Phe Phe Ala His Phe Thr Cys Ala Thr Asp Thr Gln Ser Val
340 345 350
Arg Ser Val Phe Lys Asp Val Arg Asp Ser Val Leu Ala Arg Tyr Leu
355 360 365
Asp Glu Ile Asn
370
<210> 2
<211> 1177
<212> ДНК
<213> Homo sapiens
<400> 2
ggatccaggc cggccggtac cgccgccacc atggctagat cccttacttg gcgctgttgt 60
ccttggtgcc tcacggaaga tgaaaaggca gcggctagag tcgatcagga aatcaaccgg 120
attctcttgg agcagaagaa gcaggataga ggagaactca aactgcttct cctcgggcca 180
ggagagagcg gaaaatccac gtttatcaaa cagatgagaa tcattcatgg ggcagggtac 240
agcgaagaag agcgcaaagg gtttcggccc ctcgtctatc agaacatctt tgtctcgatg 300
agggccatga tcgaagcaat ggagcgcctc caaattcctt tctcaaggcc ggagtcgaag 360
caccacgcct cgttggtgat gagccaggac ccgtacaagg tgacaacgtt cgagaaaaga 420
tacgcggcag cgatgcagtg gctgtggcgc gatgcgggta tccgggcctg ttacgagagg 480
aggcgagaat ttcacctctt ggactcggca gtatactatc tgtcccatct tgaacggatc 540
acagaggagg gatacgtgcc tactgctcaa gacgtactgc ggagccgaat gccgacgact 600
gggattaacg agtattgctt ctccgtgcaa aagacaaacc ttaggatcgt agacgtgggc 660
ggacaaaagt cagagcggaa gaagtggatt cactgctttg agaatgtcat tgcgctgatc 720
taccttgcat cgcttagcga gtacgatcag tgtctcgagg agaacaatca ggaaaacagg 780
atgaaagagt ccttggccct gttcgggacc attctggaat tgccctggtt caaatcgaca 840
tcagtgattt tgtttctgaa taagaccgac atcttggaag agaaaatccc cacatcgcat 900
ctcgcgacct atttcccatc atttcagggt cccaaacagg acgcggaagc ggccaagcga 960
ttcatcttgg atatgtacac gcggatgtat accggctgcg tcgacggacc ggagggttcg 1020
aagaaaggtg cgcgatcaag gagactcttc tcgcactata cgtgcgccac tgatacgcag 1080
aatatcagaa aggtattcaa ggacgtccgc gattcggtgc ttgcgcgcta tttggacgaa 1140
atcaatctcc tttaggcggc cgcatattca tggatcc 1177
<210> 3
<211> 1163
<212> ДНК
<213> Homo sapiens
<400> 3
ggatccaggc cggccggtac cgccgccacc atggctagat cccttacttg gcgctgttgt 60
ccttggtgcc tcacggaaga tgaaaaggca gcggctagag tcgatcagga aatcaaccgg 120
attctcttgg agcagaagaa gcaggataga ggagaactca aactgcttct cctcgggcca 180
ggagagagcg gaaaatccac gtttatcaaa cagatgagaa tcattcatgg ggcagggtac 240
agcgaagaag agcgcaaagg gtttcggccc ctcgtctatc agaacatctt tgtctcgatg 300
agggccatga tcgaagcaat ggagcgcctc caaattcctt tctcaaggcc ggagtcgaag 360
caccacgcct cgttggtgat gagccaggac ccgtacaagg tgacaacgtt cgagaaaaga 420
tacgcggcag cgatgcagtg gctgtggcgc gatgcgggta tccgggcctg ttacgagagg 480
aggcgagaat ttcacctctt ggactcggca gtatactatc tgtcccatct tgaacggatc 540
acagaggagg gatacgtgcc tactgctcaa gacgtactgc ggagccgaat gccgacgact 600
gggattaacg agtattgctt ctccgtgcaa aagacaaacc ttaggatcgt agacgtgggc 660
ggacaaaagt cagagcggaa gaagtggatt cactgctttg agaatgtcat tgcgctgatc 720
taccttgcat cgcttagcga gtacgatcag tgtctcgagg agaacaatca ggaaaacagg 780
atgaaagagt ccttggccct gttcgggacc attctggaat tgccctggtt caaatcgaca 840
tcagtgattt tgtttctgaa taagaccgac atcttggaag agaaaatccc cacatcgcat 900
ctcgcgacct atttcccatc atttcagggt cccaaacagg acgcggaagc ggccaagcga 960
ttcatcttgg atatgtacac gcggatgtat accggctgcg tcgacggacc ggagggttcg 1020
aagaaaggtg cgcgatcaag gagactcttc tcgcactata cgtgcgccac tgatacgcag 1080
aatgtcaaat tcgtgtttga cgccgtaacc gatatcatca tcaaggagaa tctcaaggac 1140
tgcggcctgt tctaggcggc cgc 1163
<210> 4
<211> 1177
<212> ДНК
<213> Homo sapiens
<400> 4
ggatccaggc cggccggtac cgccgccacc atggcgcgct cccttacgtg gggctgctgt 60
ccctggtgcc ttactgagga ggaaaagact gcagcgcgca tcgaccagga gatcaaccgc 120
atcttgttgg aacagaagaa gcaggaacga gaagagttga agctcctcct gcttgggcct 180
ggtgaatcgg gaaagtcaac attcatcaag cagatgcgga tcatccacgg ggtgggatat 240
agcgaggagg accgacgggc gtttcgattg cttatctacc agaatatctt tgtgtcgatg 300
caggccatga ttgacgccat ggatcgcttg caaattccat tttcacgacc ggacagcaaa 360
cagcacgcgt cccttgtaat gacgcaagac ccgtacaaag tctcgacatt tgagaaaccc 420
tatgccgtgg cgatgcaata cctttggaga gatgcaggga ttagggcgtg ttatgaacga 480
cggagagagt ttcatctcct cgactccgca gtctactatc tgagccatct ggagcggatc 540
agcgaggata gctacattcc taccgctcaa gacgtgctga gatcaaggat gccgacgact 600
ggcattaacg aatactgctt ctcggtcaag aaaaccaagc tccggatcgt cgatgtcgga 660
gggcagagat cagagaggag aaagtggatt cactgtttcg agaacgtaat cgctttgatc 720
tacttggcga gcctttcgga atacgaccag tgtctggaag agaatgacca ggaaaaccgg 780
atggaggagt cgcttgcact tttctccaca attctggaac tcccctggtt caaatcaacg 840
tccgtgattc tcttcttgaa taagacagac attctcgagg ataagatcca cacgtcccac 900
ctggcgacct actttccatc gtttcagggt ccgaggcgcg acgctgaggc agcgaaatcg 960
ttcatcctcg atatgtatgc cagggtgtat gcctcgtgcg ccgaacccca agatggaggt 1020
aggaaaggat caagagcgag gcgcttcttt gcgcatttca catgcgctac ggatacccag 1080
tcagtgcgat ccgtattcaa agatgtacgc gattcggtcc tcgcccggta tcttgacgag 1140
atcaatctgt tgtaagcggc cgcatattca tggatcc 1177
<210> 5
<211> 1163
<212> ДНК
<213> Homo sapiens
<400> 5
ggatccaggc cggccggtac cgccgccacc atggcgcgct cccttacgtg gggctgctgt 60
ccctggtgcc ttactgagga ggaaaagact gcagcgcgca tcgaccagga gatcaaccgc 120
atcttgttgg aacagaagaa gcaggaacga gaagagttga agctcctcct gcttgggcct 180
ggtgaatcgg gaaagtcaac attcatcaag cagatgcgga tcatccacgg ggtgggatat 240
agcgaggagg accgacgggc gtttcgattg cttatctacc agaatatctt tgtgtcgatg 300
caggccatga ttgacgccat ggatcgcttg caaattccat tttcacgacc ggacagcaaa 360
cagcacgcgt cccttgtaat gacgcaagac ccgtacaaag tctcgacatt tgagaaaccc 420
tatgccgtgg cgatgcaata cctttggaga gatgcaggga ttagggcgtg ttatgaacga 480
cggagagagt ttcatctcct cgactccgca gtctactatc tgagccatct ggagcggatc 540
agcgaggata gctacattcc taccgctcaa gacgtgctga gatcaaggat gccgacgact 600
ggcattaacg aatactgctt ctcggtcaag aaaaccaagc tccggatcgt cgatgtcgga 660
gggcagagat cagagaggag aaagtggatt cactgtttcg agaacgtaat cgctttgatc 720
tacttggcga gcctttcgga atacgaccag tgtctggaag agaatgacca ggaaaaccgg 780
atggaggagt cgcttgcact tttctccaca attctggaac tcccctggtt caaatcaacg 840
tccgtgattc tcttcttgaa taagacagac attctcgagg ataagatcca cacgtcccac 900
ctggcgacct actttccatc gtttcagggt ccgaggcgcg acgctgaggc agcgaaatcg 960
ttcatcctcg atatgtatgc cagggtgtat gcctcgtgcg ccgaacccca agatggaggt 1020
aggaaaggat caagagcgag gcgcttcttt gcgcatttca catgcgctac ggatacccag 1080
aatgtcaaat tcgtgtttga cgccgtaacc gatatcatca tcaaggagaa tctcaaggac 1140
tgcggcctgt tctaggcggc cgc 1163
<210> 6
<211> 1163
<212> ДНК
<213> Homo sapiens
<400> 6
ggatccaggc cggccggtac cgccgccacc atggctagat cccttacttg gcgctgttgt 60
ccttggtgcc tcacggaaga tgaaaaggca gcggctagag tcgatcagga aatcaaccgg 120
attctcttgg agcagaagaa gcaggataga ggagaactca aactgcttct cctcgggcca 180
ggagagagcg gaaaatccac gtttatcaaa cagatgagaa tcattcatgg ggcagggtac 240
agcgaagaag agcgcaaagg gtttcggccc ctcgtctatc agaacatctt tgtctcgatg 300
agggccatga tcgaagcaat ggagcgcctc caaattcctt tctcaaggcc ggagtcgaag 360
caccacgcct cgttggtgat gagccaggac ccgtacaagg tgacaacgtt cgagaaaaga 420
tacgcggcag cgatgcagtg gctgtggcgc gatgcgggta tccgggcctg ttacgagagg 480
aggcgagaat ttcacctctt ggactcggca gtatactatc tgtcccatct tgaacggatc 540
acagaggagg gatacgtgcc tactgctcaa gacgtactgc ggagccgaat gccgacgact 600
gggattaacg agtattgctt ctccgtgcaa aagacaaacc ttaggatcgt agacgtgggc 660
ggacaaaagt cagagcggaa gaagtggatt cactgctttg agaatgtcat tgcgctgatc 720
taccttgcat cgcttagcga gtacgatcag tgtctcgagg agaacaatca ggaaaacagg 780
atgaaagagt ccttggccct gttcgggacc attctggaat tgccctggtt caaatcgaca 840
tcagtgattt tgtttctgaa taagaccgac atcttggaag agaaaatccc cacatcgcat 900
ctcgcgacct atttcccatc atttcagggt cccaaacagg acgcggaagc ggccaagcga 960
ttcatcttgg atatgtacac gcggatgtat accggctgcg tcgacggacc ggagggttcg 1020
aacctcaaga aagaagataa ggaaatctac agccatatga cttgtgcgac cgacacgcag 1080
aatgtgaagt ttgtatttga cgcggtaact gacatcatca ttaaggaaaa ccttaaagat 1140
tgtggtcttt tctgagcggc cgc 1163
<210> 7
<211> 1163
<212> ДНК
<213> Homo sapiens
<400> 7
ggatccaggc cggccggtac cgccgccacc atggcgcgct cccttacgtg gggctgctgt 60
ccctggtgcc ttactgagga ggaaaagact gcagcgcgca tcgaccagga gatcaaccgc 120
atcttgttgg aacagaagaa gcaggaacga gaagagttga agctcctcct gcttgggcct 180
ggtgaatcgg gaaagtcaac attcatcaag cagatgcgga tcatccacgg ggtgggatat 240
agcgaggagg accgacgggc gtttcgattg cttatctacc agaatatctt tgtgtcgatg 300
caggccatga ttgacgccat ggatcgcttg caaattccat tttcacgacc ggacagcaaa 360
cagcacgcgt cccttgtaat gacgcaagac ccgtacaaag tctcgacatt tgagaaaccc 420
tatgccgtgg cgatgcaata cctttggaga gatgcaggga ttagggcgtg ttatgaacga 480
cggagagagt ttcatctcct cgactccgca gtctactatc tgagccatct ggagcggatc 540
agcgaggata gctacattcc taccgctcaa gacgtgctga gatcaaggat gccgacgact 600
ggcattaacg aatactgctt ctcggtcaag aaaaccaagc tccggatcgt cgatgtcgga 660
gggcagagat cagagaggag aaagtggatt cactgtttcg agaacgtaat cgctttgatc 720
tacttggcga gcctttcgga atacgaccag tgtctggaag agaatgacca ggaaaaccgg 780
atggaggagt cgcttgcact tttctccaca attctggaac tcccctggtt caaatcaacg 840
tccgtgattc tcttcttgaa taagacagac attctcgagg ataagatcca cacgtcccac 900
ctggcgacct actttccatc gtttcagggt ccgaggcgcg acgctgaggc agcgaaatcg 960
ttcatcctcg atatgtatgc cagggtgtat gcctcgtgcg ccgaacccca agatggaggt 1020
aacctcaaga aagaagataa ggaaatctac agccatatga cttgtgcgac cgacacgcag 1080
aatgtgaagt ttgtatttga cgcggtaact gacatcatca ttaaggaaaa ccttaaagat 1140
tgtggtcttt tctgagcggc cgc 1163
<210> 8
<211> 1163
<212> ДНК
<213> Homo sapiens
<400> 8
ggatccaggc cggccggtac cgccgccacc atggcgcgct cccttacgtg gggctgctgt 60
ccctggtgcc ttactgagga ggaaaagact gcagcgcgca tcgaccagga gatcaaccgc 120
atcttgttgg aacagaagaa gcaggaacga gaagagttga agctcctcct gcttgggcct 180
ggtgaatcgg gaaagtcaac attcatcaag cagatgcgga tcatccacgg ggtgggatat 240
agcgaggagg accgacgggc gtttcgattg cttatctacc agaatatctt tgtgtcgatg 300
caggccatga ttgacgccat ggatcgcttg caaattccat tttcacgacc ggacagcaaa 360
cagcacgcgt cccttgtaat gacgcaagac ccgtacaaag tctcgacatt tgagaaaccc 420
tatgccgtgg cgatgcaata cctttggaga gatgcaggga ttagggcgtg ttatgaacga 480
cggagagagt ttcatctcct cgactccgca gtctactatc tgagccatct ggagcggatc 540
agcgaggata gctacattcc taccgctcaa gacgtgctga gatcaaggat gccgacgact 600
ggcattaacg aatactgctt ctcggtcaag aaaaccaagc tccggatcgt cgatgtcgga 660
gggcagagat cagagaggag aaagtggatt cactgtttcg agaacgtaat cgctttgatc 720
tacttggcga gcctttcgga atacgaccag tgtctggaag agaatgacca ggaaaaccgg 780
atggaggagt cgcttgcact tttctccaca attctggaac tcccctggtt caaatcaacg 840
tccgtgattc tcttcttgaa taagacagac attctcgagg ataagatcca cacgtcccac 900
ctggcgacct actttccatc gtttcagggt ccgaggcgcg acgctgaggc agcgaaatcg 960
ttcatcctcg atatgtatgc cagggtgtat gcctcgtgcg ccgaacccca agatggaggt 1020
aggaaaggat caagagcgag gcgcttcttt gcgcatttca catgcgctac ggatacccag 1080
tcagtgcgat ccgtattcaa agatgtacgc gattcggtcc tcgcccggta tcttgacgag 1140
tgcgggctgt attaagcggc cgc 1163
Композиция напитка и способ уменьшения деградации монатина
Напиток с консервирующей системой, содержащей пимарицин-циклодекстриновый комплекс
Композиции на основе геля и способы их получения
Устройство для выдачи напитков
Устройство дозирования напитков с обратной связью в виде аудио-и видеоинформации
Видеодисплей для витрин с продуктами
Устройство для выдачи напитков
Газированный желеобразный напиток с включениями
Ненасыщенные кислоты для защиты от обесцвечивания красителей, полученных из натуральных источников, используемых в напитках
Насосная система для упаковок типа мешок-в-коробке
Композиция напитка и способ уменьшения деградации монатина
Напиток с консервирующей системой, содержащей пимарицин-циклодекстриновый комплекс
Композиции на основе геля и способы их получения
Устройство для выдачи напитков
Устройство дозирования напитков с обратной связью в виде аудио-и видеоинформации
Видеодисплей для витрин с продуктами
Устройство для выдачи напитков
Газированный желеобразный напиток с включениями
Ненасыщенные кислоты для защиты от обесцвечивания красителей, полученных из натуральных источников, используемых в напитках
Насосная система для упаковок типа мешок-в-коробке

