Результат интеллектуальной деятельности: СПОСОБ И КОНСТРУКТ ДЛЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА SCBV
Вид РИД
Изобретение
По данной заявке испрашивается приоритет по предварительной патентной заявке США, серийный номер 61/582148, поданной 30 декабря 2011. По данной заявке также испрашивается приоритет по предварительной патентной заявке США, серийный номер 61/641956, поданной 3 мая 2012.
Область техники, к которой относится изобретение
Данное изобретение в основном относится к области молекулярной биологии растений, в частности, к области стабильной экспрессии разнообразных генов в трансгенных растениях.
Предпосылки создания изобретения
Многие виды растений обладают способностью трансформироваться с трансгенами из других видов для привнесения желаемых, с агрономической точки зрения, свойств или характеристик, например, улучшения качества пищевой ценности, повышения урожайности, придания устойчивости к сельскохозяйственным вредителям или заболеваниям, повышения устойчивости к засухе и стрессу, улучшения плодовых качеств (таких как пигментация и рост), придания устойчивости к гербицидам, обеспечивая производство полезных в промышленности соединений и/или веществ из растений, и/или обеспечивая производство фармацевтических средств. Введение трансгенов в клетки растений и последующее выявление плодоносящих трансгенных растений, которые содержат стабильно интегрированную копию трансгена, может быть использовано для получения трансгенных растений, обладающих заданными свойствами.
Контроль и регуляция экспрессии гена могут происходить посредством различных механизмов. Инициация транскрипции гена является доминирующим механизмом контроля экспрессии гена. Инициация транскрипции в основном контролируется полинуклеотидными последовательностями, локализованными в 5’-фланкирующей или расположенной выше области транскрибируемого гена. Эти последовательности в совокупности называют промоторами. Промоторы, как правило, содержат сигналы для РНК-полимеразы начать транскрипцию для синтеза матричной РНК (мРНК). На зрелой мРНК происходит трансляция рибосомой с образованием, таким образом, белков. ДНК-связывающие белки взаимодействуют специфически с промоторными ДНК-последовательностями для стимуляции образования транскрипционного комплекса и инициации процесса экспрессии гена. Существует большое разнообразие эукариотических промоторов, выделенных и охарактеризованных из растений, которые являются функциональными для управления экспрессией трансгена в растениях. Были выделены и охарактеризованы промоторы, влияющие на экспрессию гена в ответ на стимулы окружающей среды, усвояемость питательных веществ или неблагоприятные условия, включая тепловой шок, анаэробиоз или наличие тяжелых металлов. Существуют также промоторы, которые контролируют экспрессию генов во время развития или в ткани, или органоспецифическим образом. Кроме того, были выделены и охарактеризованы прокариотические промоторы из бактерий и вирусов, которые являются функционально активными в управлении экспрессией трансгена в растениях.
Конкретный эукариотический промотор состоит из минимального промотора и других цис-элементов. Минимальный промотор является по существу ТАТА-боксобластью, где РНК-полимераза II (polII), ТАТА-связывающий белок (TBP) и ТВР-ассоциированные факторы (TAF) могут связываться для инициации транскрипции. Однако, в большинстве случаев, элементы последовательности, отличные от ТАТА-мотива, необходимы для точной транскрипции. Было обнаружено, что такие элементы последовательности (например, энхансеры) повышают общий уровень экспрессии расположенных поблизости генов часто способом, не зависящим от расположения и/или ориентации. Другие последовательности рядом с сайтом начала транскрипции (например, INR-последовательности) некоторых генов polII могут предоставлять альтернативный сайт связывания для факторов, которые также вносят вклад в активацию транскрипции, альтернативно, даже предоставляя основные сайты связывания промотора для транскрипции в промоторах, у которых отсутствуют функциональные ТАТА-элементы. См., например, Zenzie-Gregory et al. (1992) J. Biol. Chem. 267:2823-30.
Другие регуляторные элементы гена включают последовательности, которые взаимодействуют со специфическими ДНК-связывающими факторами. Эти мотивы последовательностей в некоторых случаях называют цис-элементами, и они, как правило, зависят от расположения и ориентации, хотя они могут быть обнаружены с 5' или 3' стороны кодирующей последовательности гена или в интроне. Такие цис-элементы, с которыми связываются тканеспецифичные или специфичные для развития транскрипционные факторы, индивидуально или в комбинации, могут определять пространственно-временной паттерн экспрессии промотора на транскрипционном уровне. Расположение в определенном порядке идущих против хода транскрипции цис-элементов вслед за минимальным промотором, как правило, задает полярность определенного промотора. Промоторы в растениях, которые были клонированы и широко использованы как для фундаментальных исследований, так и в биотехнологическом масштабе, как правило, являются однонаправленными, управляя только одним геном, который был соединен с их 3'-концом (то есть, по ходу транскрипции). См., например, Xie et al. (2001) Nat. Biotechnol. 19(7):677-9; патент США 6388170.
Множество цис-элементов (или "расположенные против хода транскрипции регуляторные последовательности") было идентифицировано в промоторах растений. Эти цис-элементы широко варьируют по типу контроля, который они осуществляют на функционально связанные гены. Некоторые элементы увеличивают транскрипцию функционально связанных генов в ответ на стимулы окружающей среды (например, температура, влажность и повреждение). Другие цис-элементы могут отвечать на сигналы развития (например, зарождение, созревание семени и цветение) или пространственную информацию (например, тканевую специфичность). См., например, Langridge et al. (1989) Proc. Natl. Acad. Sci. USA 86:3219-23. Тип контроля специфических элементов промотора, как правило, является внутренним качеством промотора; то есть, гетерологичный ген под контролем такого промотора, по-видимому, экспрессируется в соответствии с контролем нативного гена, из которого был выделен элемент промотора. Также эти элементы, как правило, могут быть заменены другими элементами и поддерживать их характерный внутренний контроль над экспрессией гена.
Часто необходимо вводить множество генов в растения для метаболической инженерии и создания характерной многоуровневой структуры, гены которой часто контролируются идентичными или гомологичными промоторами. Однако основанное на гомологии подавление экспрессии гена (HBGS), по-видимому, возникает в том случае, когда множество введенных трансгенов имеют гомологичные промоторы, управляющие ими. См., например, Mol et al. (1989) Plant Mol. Biol. 13:287-94. По литературным данным HBGS активно происходит в трансгенных растениях. См., например, Vaucheret and Fagard (2001) Trends Genet. 17:29-35. Было предложено несколько механизмов, объясняющих феномен HBGS, все из которых затрагивают характерное свойство гомологии последовательностей в промоторе запускать клеточные механизмы распознавания, приводящие к выключению транскрипции повторенных генов. См., например, Matzke and Matzke (1995) Plant Physiol. 107:679-85; Meyer and Saedler (1996) Ann. Rev. Plant Physiol. Plant Mol. Biol. 47:23-48; Fire (1999) Trends Genet. 15:358-63; Hamilton and Baulcombe (1999) Science 286:950-2; and Steimer et al. (2000) Plant Cell 12:1165-78.
Способы предотвращения HBGS в трансгенных растениях часто включают создание синтетических промоторов, которые функционально эквивалентны, но имеют минимальную гомологию последовательности. Когда такие синтетические промоторы используются для экспрессирующихся в сельскохозяйственных культурах трансгенов, они могут помочь предотвратить или снизить уровень HBGS. См., например, Mourrain et al. (2007) Planta 225(2):365-79; Bhullar et al. (2003) Plant Physiol. 132:988-98. Такие промоторы могут быть созданы путем введения известных цис-элементов в новую или синтетическую нить ДНК или, альтернативно, путем "доменной замены", где домены одного промотора заменяют функционально эквивалентными доменами из других гетерологичных промоторов.
Таким образом, существует необходимость в создании конструктов и способов эффективной стабильной экспрессии множества трансгенов с минимальным риском рекомбинации или потери трансгенов вследствие селекции или многократного воспроизведения у трансгенных растений.
Описание изобретения
В настоящем описании описаны определенные синтетические промоторы, включающие минимальный промотор Ubi1. В вариантах осуществления изобретения синтетический промотор, включающий минимальный промотор Ubi1, дополнительно содержит по меньшей мере один элемент последовательности промотора SCBV или его функциональный элемент. В некоторых примерах, такой синтетический промотор ("синтетический промотор SCBV") может представлять собой промотор, обладающий способностью контролировать транскрипцию функционально связанной нуклеотидной последовательности в растительной клетке. В других примерах синтетический промотор SCBV может представлять собой синтетический двунаправленный промотор SCBV, например, нуклеиновую кислоту, содержащую нуклеотидные последовательности элемента минимального промотора Ubi1, ориентированные в противоположном направлении относительно элементов промотора SCBV, который обладает способностью контролировать транскрипцию в растительной клетке двух функционально связанных нуклеотидных последовательностей, фланкирующих промотор. Дополнительные элементы, которые могут быть сконструированы для включения в синтетический двунаправленный промотор SCBV, включают интроны (например, интрон алькогольдегидрогеназы (ADH), экзоны и/или всю или часть расположенной против хода транскрипции области промотора. В определенных примерах синтетический двунаправленный промотор может содержать более чем один из любых вышеперечисленных элементов.
Конкретные варианты осуществления изобретения включают клетки (например, растительные клетки), содержащие синтетический промотор SCBV или его функциональный элемент. Например, конкретные варианты осуществления могут включать клетку, содержащую синтетический двунаправленный промотор SCBV или его функциональный элемент. Растительные клетки, согласно конкретным вариантам осуществления, могут присутствовать в клеточной культуре, ткани, части растения и/или целом растении. Таким образом, растение (например, однодольное или двудольное растение), содержащее клетку, имеющую синтетический промотор SCBV или его функциональный элемент, включено в некоторые варианты осуществления изобретения.
Другие варианты изобретения включают способы инициации транскрипции двух, представляющих интерес функционально связанных нуклеотидных последовательностей. Средства инициации транскрипции двух, представляющих интерес функционально связанных нуклеотидных последовательностей включают синтетический двунаправленный промотор SCBV, имеющий последовательность SEQ ID NO:5.
Также представлены конструкты и способы экспрессии множества генов в растительных клетках и/или растительных тканях. Представленные конструкты включают по меньшей мере один двунаправленный промотор, связанный с экспрессионными кассетами множества генов, где двунаправленный промотор содержит нуклеотидную последовательность функционального промотора из промотора бациллярного вируса сахарного тростника (SCBV). В некоторых вариантах осуществления в представленных конструктах и способах используют двунаправленный промотор, основанный на элементе минимального корового промотора из гена убиквитина-1 Zea mays или его функциональном эквиваленте и элементах нуклеотидной последовательности из промотора бациллярного вируса сахарного тростника. В некоторых вариантах осуществления представленные конструкты и способы обеспечивают экспрессию от трех до двадцати генов.
В одном аспекте предоставлен синтетический полинуклеотид, содержащий элемент минимального корового промотора из гена убиквитина-1 Zea mays или Zea luxurians и нуклеотидную последовательность функционального промотора из промотора бациллярного вируса сахарного тростника. В одном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность, которая по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентична SEQ ID NO:1 или ее комплементарной цепи. В дополнительном или альтернативном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность, выбранную из группы, состоящую из SEQ ID NO:1 и 16-40. В дополнительном варианте осуществления элемент минимального корового промотора содержит SEQ ID NO:1 или ее комплементарную цепь. В дополнительном варианте осуществления элемент минимального корового промотора состоит по существу из SEQ IN NO:1 или ее комплементарной цепи. В другом варианте осуществления представленный синтетический полинуклеотид дополнительно содержит экзон из гена убиквитина-1 и интрон из гена убиквитина-1. В дополнительном варианте осуществления экзон получают из гена убиквитина-1 Zea mays или Zea luxurians. В другом варианте осуществления представленный синтетический полинуклеотид дополнительно содержит интрон из гена алкогольдегидрогеназы. В другом варианте осуществления изобретения представленный синтетический полинуклеотид дополнительно содержит расположенную против хода транскрипции регуляторную последовательность из промотора бациллярного вируса сахарного тростника. В другом варианте осуществления нуклеотидная последовательность функционального промотора из полинуклеотидной последовательности промотора бациллярного вируса сахарного тростника и гена алкогольдегидрогеназы по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентична SEQ ID NO:6 или ее комплементарной цепи. В дополнительном или альтернативном варианте осуществления функциональная нуклеотидная последовательность промотора из промотора бациллярного вируса сахарного тростника и гена алкогольдегидрогеназы содержит SEQ ID NO:6 или ее комлементарную цепь. В дополнительном варианте осуществления функциональная нуклеотидная последовательность промотора из промотора бациллярного вируса сахарного тростника и гена алкогольдегидрогеназы состоит по существу из SEQ ID NO:6 или ее комлементарной цепи.
В одном варианте осуществления представленный синтетический полинуклеотид дополнительно содержит по меньшей мере один элемент, выбранный из списка, включающего расположенную против хода транскрипции регуляторную последовательность (URS), энхансерный элемент, экзон, интрон, сайт старта транскрипции, TATA-бокс, консенсусный элемент теплового шока, нуклеотидную последовательность начала и/или окончания трансляции. В другом варианте осуществления представленный синтетический полинуклеотид дополнительно включает элемент, выбранный из группы, состоящей из расположенной против хода транскрипции регуляторной последовательности (URS), энхансерного элемента, экзона, интрона, сайта начала транскрипции, ТАТА-бокса, консенсусного элемента теплового шока, нуклеотидной последовательности начала и/или окончания трансляции, и их комбинаций. В другом варианте осуществления представленный синтетический полинуклеотид дополнительно содержит представляющую интерес нуклеотидную последовательность, функционально связанную с элементом минимального корового промотора. В другом варианте осуществления элемент минимального корового промотора из гена убиквитина-1кукурузы обыкновенной и нуклеотидная последовательность функционального промотора из промотора бациллярного вируса сахарного тростника расположены в обратной комплементарной ориентации относительно друг друга в полинуклеотиде.
В другом варианте осуществления представленный синтетический полинуклеотид содержит экзон из гена убиквитина-1, интрон из гена убиквитина-1, интрон из гена алкогольдегидрогеназы. В дополнительном или альтернативном варианте осуществления представленный синтетический полинуклеотид содержит вторую представляющую интерес кодирующую нуклеотидную последовательность, функционально связанную с нуклеотидной последовательностью функционального промотора из промотора бациллярного вируса сахарного тростника. В дополнительном варианте осуществления представленный синтетический полинуклеотид включает полинуклеотидную последовательность, которая по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентична SEQ IN NO:5 или ее комплементарной цепи. В дополнительном варианте осуществления представленный синтетический полинуклеотид содержит SEQ ID NO:5 или ее комплементарную цепь. В дополнительном варианте осуществления представленный синтетический полинуклеотид состоит по существу из SEQ ID NO:5 или ее комплементарной цепи. В дополнительном варианте осуществления экзон или интрон получают из гена убиквитина-1 Zea mays или Zea luxurians.
В дополнительном варианте осуществления представленный синтетический полинуклеотид содержит представляющую интерес первую кодирующую нуклеотидную последовательность, функционально связанную с элементом минимального корового промотора из гена убиквитина-1 Zea mays. В другом дополнительном варианте осуществления представленный синтетический полинуклеотид содержит представляющую интерес вторую кодирующую нуклеотидную последовательность, функционально связанную с нуклеотидной последовательностью функционального промотора из промотора бациллярного вируса сахарного тростника.
В другом аспекте предоставлен способ получения трансгенной клетки, включающий трансформацию клетки полинуклеотидом, представленном в настоящем описании. В одном варианте осуществления клетка представляет собой растительную клетку. В другом аспекте предоставлена растительная клетка, содержащая полинуклеотид, представленный в настоящем описании. В другом аспекте предоставлено растение, содержащее растительную клетку, представленную в настоящем описании.
В другом аспекте предоставлен способ экспрессии представляющей интерес нуклеотидной последовательности в растительной клетке, включающий введение в растительную клетку представляющей интерес нуклеотидной последовательности, функционально связанной со структурами для инициации транскрипции двух функционально связанных представляющих интерес нуклеотидных последовательностей. В одном варианте осуществления предоставленный способ включает введение в растительную клетку нуклеиновой кислоты, содержащей (а) представляющую интерес нуклеотидную последовательность, функционально связанную со структурами для инициации транскрипции двух функционально связанных представляющих интерес нуклеотидных последовательностей; и (b) представляющую интерес вторую нуклеотидную последовательность, функционально связанную со структурами для инициации транскрипции двух представляющих интерес функционально связанных нуклеотидных последовательностей.
В одном варианте осуществления средства инициации транскрипции двух представляющих интерес функционально связанных нуклеотидных последовательностей включают SEQ IN NO:5 или ее комплементарную цепь. В другом варианте осуществления средства инициации транскрипции двух представляющих интерес функционально связанных нуклеотидных последовательностей включает SEQ ID NO:5. В другом варианте осуществления средства инициации транскрипции двух представляющих интерес функционально связанных нуклеотидных последовательностей включает обратную комплементарную SEQ ID NO:5 нить. В другом варианте осуществления нуклеиновую кислоту вводят в растительную клетку с тем, чтобы воздействовать на предопределенный сайт в ДНК растительной клетки представляющей интерес нуклеотидной последовательности, функционально связанной со структурами для инициации транскрипции двух представляющих интерес функционально связанных нуклеотидных последовательностей. В дополнительном или альтернативном варианте осуществления представляющую интерес нуклеотидную последовательность, функционально связанную со средствами инициации транскрипции двух представляющих интерес функционально связанных нуклеотидных последовательностей, направляют на предопределенный сайт, используя рекомбинацию, опосредованную нуклеазой "цинковые пальцы".
В некоторых вариантах осуществления экзон получают из гена убиквитина-1 Zea spp. В некоторых вариантах осуществления интрон получают из гена убиквитина-1 Zea spp. В некоторых вариантах осуществления Zea spp. представляет собой Zea mays или Zea luxurians.
В другом аспекте предоставлен конструкт нуклеиновой кислоты для экспрессии множества генов в растительных клетках и/или тканях. Конструкт нуклеиновой кислоты включает (а) двунаправленный промотор, где двунаправленный промотор содержит нуклеотидную последовательность функционального промотора из промотора бациллярного вируса сахарного тростника (SCBV); и (b) две генные экспрессионные кассеты на противоположных концах двунаправленного промотора; где по меньшей мере одна из генных экспрессионных кассет включает два или более генов, соединенных через переключатель трансляции.
В одном варианте осуществления двунаправленный промотор содержит по меньшей мере один энхансер. В другом варианте осуществления двунаправленный промотор не содержит энхансер. В другом варианте осуществления конструкт нуклеиновой кислоты содержит бинарный вектор для опосредованной Agrobacterium трансформации. В одном варианте осуществления двунаправленный промотор содержит элемент, выбранный из группы, состоящей из расположенной против хода транскрипции регуляторной последовательности (URS), энхансерного элемента, экзона, интрона, сайта начала транскрипции, TATA-бокса, консенсусного элемента теплового шока и их комбинаций. В другом варианте осуществления двунаправленный промотор содержит элемент минимального корового промотора из гена убиквитина-1 Zea mays или Zea luxurians. В другом варианте осуществления изобретения элемент минимального корового промотора из гена убиквитина-1 и промоторная нуклеотидная последовательность из промотора бациллярного вируса сахарного тростника (SCBV) расположены в обратной комплементарной ориентации относительно друг друга. В дополнительном или альтернативном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичную SEQ ID NO:1 или ее комплементарной цепи. В дополнительном или альтернативном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность, выбранную из группы, состоящую из SEQ ID NO:1 и 16-40. В дополнительном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность, выбранную из группы, состоящую из SEQ IN NO:1 и 16-35. В дополнительном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность, выбранную из группы, состоящую из SEQ ID NO:1 и 16-30. В дополнительном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность, выбранную из группы, состоящую из SEQ ID NO:1 и 16-25. В дополнительном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность, выбранную из группы, состоящую из SEQ ID NO:1 и 16-20. В дополнительном варианте осуществления элемент минимального корового промотора содержит полинуклеотидную последовательность SEQ ID NO:1.
В дополнительном или альтернативном варианте осуществления двунаправленный промотор содержит экзон из гена убиквитина-1 и/или интрона из гена убиквитина. В другом варианте осуществления двунаправленный промотор содержит интрон из гена алкогольдегидрогеназы. В одном варианте осуществления трансгенные растения стабильно трансформированы конструктом нуклеиновой кислоты. В одном варианте осуществления изобретения растения представляют собой односемядольные растения. В другом варианте осуществления растения являются двудольными растениями. В другом варианте осуществления растения не являются односемядольными растениями. В другом варианте осуществления растения не являются двудольными растениями.
В дополнительном или альтернативном варианте осуществления двунаправленный промотор содержит расположенную против хода транскрипции регуляторную последовательность из гена убиквитина или промотора бациллярного вируса сахарного тростника (SCBV). В дополнительном варианте осуществления двунаправленный промотор содержит расположенную против хода транскрипции регуляторную последовательность из гена убиквитина. В другом варианте осуществления двунаправленный промотор содержит расположенную против хода транскрипции регуляторную последовательность из гена убиквитина или промотора бациллярного вируса сахарного тростника (SCBV).
В дополнительном варианте осуществления двунаправленный промотор содержит полинуклеотид, который по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентичен SEQ ID NO:5 или ее комплементарной цепи. В дополнительном варианте осуществления двунаправленный промотор содержит полинуклеотид SEQ ID NO:5 или ее комплементарную цепь. В дополнительном варианте осуществления двунаправленный промотор содержит полинуклеотид, который по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентичен SEQ IN NO: 6 или ее комплементарной цепи. В дополнительном варианте осуществления двунаправленный промотор содержит полинуклеотид SEQ ID NO:6 или ее комплементарную цепь.
В одном варианте осуществления обе генные экспрессионные кассеты содержат два или более генов, соединенных посредством переключателя трансляции. В дополнительном или альтернативном варианте осуществления переключатель трансляции выбран из группы, состоящей из участка внутренней посадки рибосомы (IRES), альтернативного сайта сплайсинга, сайта расщепления рибозимом, полинуклеотидной последовательности, кодирующей пептид 2А, полинуклеотидной последовательности, кодирующей 2А-подобный пептид, полинуклеотидной последовательности, кодирующей интеин, полинуклеотидной последовательности, кодирующей сайт расщепления протеазой или их комбинаций. В дополнительном или альтернативном варианте осуществления переключатель трансляции содержит цис-действующий гидролазный элемент (CHYSEL). В дополнительном варианте осуществления CHYSEL представляет собой 2А или 2А-подобную пептидную последовательность. В другом варианте осуществления ген, расположенный против хода транскрипции относительно переключателя трансляции, не содержит стоп кодон трансляции. В другом варианте осуществления конструкт нуклеиновой кислоты обеспечивает или дает возможность экспрессии по меньшей мере четырех генов. В дополнительном варианте осуществления все четыре гена являются трансгенами. В другом варианте осуществления конструкт нуклеиновой кислоты обеспечивает экспрессию от трех до двадцати генов. В другом варианте осуществления конструкт нуклеиновой кислоты обеспечивает экспрессию от четырех до восьми генов. В дополнительном или альтернативном варианте осуществления гены являются трансгенами. В другом варианте осуществления изобретения по меньшей мере одна генная экспрессионная кассета содержит полинуклеотидную последовательность, кодирующую химерный белок. В дополнительном варианте осуществления химерный белок содержит от трех до пяти генов.
В некоторых вариантах осуществления экспрессия генов с двунаправленного промотора по меньшей мере в четыре раза выше по сравнению с экспрессией с однонаправленного промотора. В некоторых вариантах осуществления экспрессия генов с двунаправленного промотора в три-десять раз выше по сравнению с экспрессией с однонаправленного промотора. В некоторых вариантах осуществления экспрессия генов с двунаправленного промотора в четыре-восемь раз выше по сравнению с экспрессией с однонаправленного промотора. В некоторых вариантах осуществления маркерный ген селекции расположен на дальнем конце от промотора (то есть, на 3’-конце генной экспрессионной кассеты по ходу транскрипции другого гена).
В другом аспекте предоставлен способ получения трансгенного растения, включающий трансформацию растительной клетки конструктом нуклеиновой кислоты, представленным в настоящем описании. В другом аспекте предоставлена растительная клетка, содержащая конструкт нуклеиновой кислоты, представленный в настоящем описании. В дополнительном или альтернативном варианте осуществления растительная клетка стабильно трансформирована конструктом нуклеиновой кислоты. В другом аспекте представлено трансгенное растение, содержащее конструкт нуклеиновой кислоты, представленный в настоящем описании. В дополнительном или альтернативном варианте осуществления клетки трансгенного растения стабильно трансформированы конструктом нуклеиновой кислоты. В другом аспекте предоставлен способ экспрессии множества генов в растительных клетках и/или тканях, включающий введение в растительные клетки и/или ткани конструкта нуклеиновой кислоты, представленного в настоящем описании. В дополнительном или альтернативном варианте осуществления растительные клетки и/или ткани стабильно трансформированы конструктом нуклеиновой кислоты, представленном в настоящем описании. В другом аспекте предоставлен бинарный вектор для трансформации, опосредованной Agrobacterium. В одном варианте осуществления бинарный вектор содержит конструкт нуклеиновой кислоты, представленный в настоящем описании. В другом варианте осуществления бинарный вектор содержит синтетический полинуклеотид, представленный в настоящем описании. В другом аспекте, предоставлено применение двунаправленного промотора, представленного в настоящем описании, для экспрессии множества трансгенов в растениях.
Краткое описание фигур и последовательностей
На фиг. 1 показан конкретный (не в масштабе) промотор Ubi1 кукурузы (ZmUbi1), который содержит приблизительно 900 п.о. идущего против хода транскрипции элемента, расположенного с 5'-конца сайта инициации транскрипции (TSS). Идущий против хода транскрипции элемент содержит ТАТА-бокс (расположенный приблизительно на расстоянии -30 п.о. относительно TSS) и два перекрывающихся консенсусных элемента теплового шока (расположенных приблизительно на расстоянии -200 п.о. относительно TSS). Этот промотор также содержит приблизительно 1100 п.о., расположенных с 3'-конца участка TSS. 3'-участок содержит прилегающую лидерную последовательность (экзон ZmUbi1) и интрон.
На фиг. 2 показан пример осуществления представленного в настоящем описании синтетического двунаправленного промотора Ubi1, который содержит элемент минимального корового промотора minUbi1P, клонированный в участке, предшествующем промотору ZmUbi1.
На фиг. 3 показано конкретное схематическое изображение экспрессионных кассет генов YFP и GUS, каждая из которых функционально связана с синтетическим двунаправленным промотором Ubi1.
На фиг. 4 показана репрезентативная карта плазмиды pDAB105801.
На фиг. 5 показано схематическое изображение конкретного двунаправленного промотора бациллярного вируса сахарного тростника (SCBV), который содержит элемент минимального корового промотора minUbi1P, клонированный в участке, предшествующем промотору SCBV.
На фиг. 6 показана репрезентативная карта плазмиды pDAB105806.
На фиг. 7 показано конкретное схематическое изображение экспрессионных кассет генов YFP и GUS, каждая из которых функционально связана с синтетическим двунаправленным промотором SCBV.
На фиг. 8 показано конкретное схематическое представление мультигенных конструктов, представленных в настоящем описании. Переключатели трансляции изображены, используя специальный (вертикальный гантелеобразный) символ.
На фиг. 9 показаны репрезентативные карты плазмид pDAB108708 и pDAB101556.
На фиг. 10А показана SEQ ID NO:1, которая содержит участок длиной 215 п.о. элемента минимального корового промотора убиквитина 1 Zea mays (minUbi1P). На фиг.10В показана SEQ ID NO:2, которая содержит обратную комплементарную цепь полинуклеотида, содержащую элемент минимального корового промотора minUbi1P Z. mays (подчеркнут); лидерную последовательность Ubi1 Z. Mays (экзон ZmUbi1; жирный шрифт) и интрон Ubi1 Z. Mays (строчный шрифт).
На фиг. 11 показана SEQ ID NO:3, которая содержит конкретный синтетический двунаправленный промотор Ubi1, в котором обратная цепь первого minUbi1Р и второй minUbi1Р подчеркнуты.
На фиг. 12 показана SEQ ID NO:4, которая содержит конкретную нуклеиновую кислоту, включающую экспрессионные кассеты генов YFP и GUS, управляемые синтетическим двунаправленным промотором Ubi1.
На фиг. 13 показана SEQ ID NO:5, которая содержит конкретный двунаправленный промотор SCBV, содержащий элемент минимального корового промотора minUbi1P, где обратная комплементарная цепь minUbi1P подчеркнута.
На фиг. 14 показана SEQ ID NO:6, которая содержит промотор SCBV, содержащий экзон 6 ADH1 (подчеркнут), интрон 6 (строчный шрифт) и экзон 7 (жирный шрифт).
На фиг. 15 показана SEQ ID NO:7, которая содержит нуклеиновую кислоту, включающую экспрессионные кассеты генов YFP и GUS, управляемые конкретным двунаправленным промотором SCBV.
SEQ ID NO:8 содержит прямой праймер YFP: 5'-GATGCCTCAGTGGGAAAGG-3'. SEQ ID NO:9 содержит обратный праймер YFP: 5'-CCATAGGTGAGAGTGGTGACAA-3'. SEQ ID NO:10 содержит прямой праймер инвертазы: 5'-TGGCGGACGACGACTTGT-3’. SEQ ID NO:11 содержит обратный праймер инвертазы: 5'-AAAGTTTGGAGGCTGCCGT-3'. SEQ ID NO:12 содержит зонд инвертазы: 5'-CGAGCAGACCGCCGTGTACTTCTACC-3'. SEQ ID NO:13 содержит прямой праймер AAD1: 5'-TGTTCGGTTCCCTCTACCAA-3'. SEQ ID NO:14 содержит обратный праймер AAD1: 5'-CAACATCCATCACCTTGACTGA-3'. SEQ ID NO:15 содержит зонд AAD1: 5'-CACAGAACCGTCGCTTCAGCAACA-3' (см. также таблицу 7).
На фиг. 16 показан Вестерн-блот-анализ для стабильной экспрессии YFP, управляемой двунаправленным конструктом промотора SCBV (pDAB108708) в Т0 растениях кукурузы. Репрезентативные растения демонстрировали стабильную экспрессию YFP в листе, управляемую элементом минимального корового промотора Min-Ubi1P. Количество продуцируемого белка указано в частях на миллион (ч./млн.).
На фиг. 17 показан Вестерн-блот-анализ для стабильной экспрессии YFP из контрольного конструкта, содержащего промотор ZmUbi1, который управляет только экспрессией YFP (pDAB101556); кодирующая последовательность GUS не содержится в этом конструкте. Количество продуцируемого белка указано в частях на миллион (ч./млн.).
На фиг. 18 показаны конкретные конструкты для четырехгенных кассетных блоков pDAB105849 (AAD1-2A-YFP плюс Cry34-2A-Cry35) и pDAB105865 (YFP-2A-AAD1 плюс Cry34-2A-Cry35). Затемненные стрелки указывают направление транскрипции с двунаправленного промотора. Ubi1-minP содержит последовательность длиной 200 нуклеотидов, расположенную выше сайта инициации транскрипции промотора кукурузы Ubi1. SCBV-URS содержит расположенную против хода транскрипции регуляторную последовательность промотора SCBV, исключая элемент минимального корового промотора (показано в виде стрелки). Ubi1-Int содержит интрон промотора Ubi1 кукурузы.
На фиг. 19 показаны два дополнительных конкретных конструкта четырехгенных кассетных блоков.
На фиг. 20 показаны репрезентативные карты для плазмид pDAB105818 и pDAB105748.
На фиг. 21А-21Е показаны дополнительные элементы минимального корового промотора (min-Ubi1P или Ubi1-minP), имеющих последовательности SEQ ID NO:16-40.
На фиг. 22 показаны репрезентативные карты для плазмид pDAB105841 и pDAB105847.
На фиг. 23 показаны репрезентативные карты для плазмид pDAB105840 и pDAB105849.
На фиг. 24 показаны репрезентативные карты для плазмид pDAB101917 и pDAB108719.
На фиг. 25 показаны репрезентативные карты для плазмид pDAB105844 и pDAB105848.
На фиг. 26 показаны репрезентативные карты для плазмид pDAB105865 и pDAB108720.
На фиг. 27 показана последовательность нуклеиновой кислоты для генных экспрессионных кассет pDAB108719, где показан каждый ген и элемент.
На фиг. 28 показаны конкретные данные по белковой экспрессии среди различных протестированных конструктов для Cry34 (фиг. 28А), AAD-1 (фиг. 28В) и Cry35 (фиг. 28С).
На фиг. 29 показаны две конкретные последовательности для желтых флуоресцентных белков из Phialidium sp. SL-2003 (PhiYFP, SEQ ID NO:51; и PhiYFPv3, SEQ ID NO:52).
На фиг. 30 показаны конкретные варианты осуществления представленных в настоящем описании синтетического двунаправленного промотора Ubi1 и конструктов, включая pDAB108706 (двунаправленный ZMUbi (-200)), pDAB108707 (двунаправленный ZMUbi (-90)), pDAB108708 (двунаправленный SCBV (-200))и pDAB108709 (двунаправленный SCBV (-90)). pDAB101556 (контроль ZmUbi1-YFP), pDAB108715 (SCBV без минимального промотора) и pDAB108716 (ZmUbi1 без минимального промотора) служили в качестве контрольных конструктов с однонаправленными промоторами.
На фиг. 31А показаны примеры результатов по экспрессии (V6) для семи конструктов, представленных на фиг. 30 для белка YFP (ЖХМС) в нг/см2. На фиг. 31В показаны примеры результатов по относительной экспрессии (V6) для семи конструктов, представленных на фиг. 30 для РНК YFP.
На фиг. 32А показаны примеры результатов по экспрессии (V6) для семи конструктов, представленных на фиг. 30 для белка GUS (ЖХМС) в нг/см2. На фиг. 32В показаны примеры результатов по относительной экспрессии (V6) для семи конструктов, представленных на фиг. 30 для РНК GUS.
На фиг. 33А показаны примеры результатов по экспрессии (V6) для семи конструктов, представленных на фиг. 30 для белка AAD1 (ЖХМС) в нг/см2. На фиг. 33В показаны примеры результатов по относительной экспрессии (V6) для семи конструктов, представленных на фиг. 30 для РНК AAD1.
На фиг. 34А показан статистический анализ результатов по экспрессии (V6) для семи конструктов, представленных на фиг. 30 для белка YFP (ЖХМС) в нг/см2. На фиг. 34В показан статистический анализ результатов по относительной экспрессии (V6) для семи конструктов, представленных на фиг. 30 для РНК YFP. Перечислены значения среднего и статистические результаты.
На фиг. 35А показан статистический анализ результатов по экспрессии (V6) для семи конструктов, представленных на фиг.30 для белка GUS (ЖХМС) в нг/см2. На фиг. 35В показан статистический анализ результатов по относительной экспрессии (V6) для семи конструктов, представленных на фиг. 30 для РНК GUS. Перечислены значения среднего и статистические результаты.
На фиг. 36А показан статистический анализ результатов по экспрессии (V6) для семи конструктов, представленных на фиг. 30 для белка AAD1 (ЖХМС) в нг/см2. На фиг. 36В показан статистический анализ результатов по относительной экспрессии (V6) для семи конструктов, представленных на фиг. 30 для РНК AAD1. Перечислены значения среднего и статистические результаты.
На фиг. 37А, 37В и 37С показаны примеры результатов экспрессии (V10) для семи конструктов, представленных на фиг.30, для белков YFP, AAD1 и GUS (ЖХМС) в нг/см2, соответственно.
На фиг. 38А, 38В и 38С показан статистический анализ результатов экспрессии (V10) для семи конструктов, представленных на фиг. 30, для белков YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Перечислены значения среднего и статистические результаты.
На фиг. 39А, 39В и 39С показаны примеры результатов экспрессии (R3) для семи конструктов, представленных на фиг.30, для белков YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно.
На фиг. 40А, 40В и 40С показан статистический анализ результатов экспрессии (R3) для семи конструктов, представленных на фиг. 30, для белков YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Перечислены значения среднего и статистические результаты.
На фиг. 41 показаны дополнительные мультитрансгенные конструкты, используя промотор Ubi1, включая pDAB108717 и pDAB108718.
На фиг. 42А показаны примеры результатов по относительной экспрессии (V6) РНК Cry34 для шести конструктов pDAB105748 (ZMUbi1-YFP), pDAB105818 (ZMUbi1-Cry34/ZMUbi1-Cry35/ZMUbi1-AAD1), pDAB108717 (YFP/AAD-1-ZMUbi1 двунаправленный-Cry34-Cry35), pDAB108718 (AAD1/YFP-ZMUbi1 двунаправленный-Cry34-Cry35), pDAB108719 (YFP/AAD1-SCBV двунаправленный-Cry34-Cry35) и pDAB108720 (AAD1/YFP-SCBV двунаправленный-Cry34-Cry35). На фиг.42В показаны примеры результатов по относительной экспрессии (V6) белка Cry34 (ЖХМС) для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 43А показаны примеры результатов по относительной экспрессии (V6) РНК AAD1 для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. На фиг.43В показаны примеры результатов по относительной экспрессии (V6) белка AAD1 (ЖХМС) для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 44А показаны примеры результатов по относительной экспрессии (V6) РНК YFP для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. На фиг.44В показаны примеры результатов по относительной экспрессии (V6) белка YFP (ЖХМС) для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 45А показаны примеры результатов по относительной экспрессии (V6) РНК Cry35 для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. На фиг. 45В показаны примеры результатов по относительной экспрессии (V6) белка Cry35 (ELISA) для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 46 показаны примеры результатов по относительной экспрессии (V6) РНК РАТ для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 47А показан статистический анализ результатов по экспрессии (V6) РНК Cry34 для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. На фиг. 47В показан статистический анализ результатов по экспрессии (V6) белка Cry34 для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Перечислены значения среднего и статистические результаты.
На фиг. 48А показан статистический анализ результатов экспрессии (V6) РНК AAD1 для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. На фиг. 48В показан статистический анализ результатов по экспрессии (V6) белка AAD1 для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Перечислены значения среднего и статистические результаты.
На фиг. 49А показан статистический анализ результатов экспрессии (V6) РНК YFP для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. На фиг. 49В показан статистический анализ результатов по экспрессии (V6) белка YFP для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Перечислены значения среднего и статистические результаты.
На фиг. 50А показан статистический анализ результатов экспрессии (V6) РНК Cry35 для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. На фиг. 50В показан статистический анализ результатов по экспрессии (V6) белка Cry35 для тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Перечислены значения среднего и статистические результаты.
На фиг. 51 показан статистический анализ результатов по экспрессии (V6) РНК PAT для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Перечислены значения среднего и статистические результаты.
На фиг. 52А, 52В, 52С и 52D показаны примеры результатов по белковой экспрессии (V10) YFP, AAD1, Cry34 и Cry35, соответственно, для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 53А, 53В, 53С и 53D показан статистический анализ результатов по белковой экспрессии (V10) YFP, AAD1, Cry34 и Cry35, соответственно, для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Перечислены значения среднего и статистические результаты.
На фиг. 54А, 54В, 54С и 54D показаны примеры результатов по белковой экспрессии (R3) YFP, AAD1, Cry34 и Cry35, соответственно, для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 55А, 55В, 55С и 55D показан статистический анализ результатов по белковой экспрессии (R3) YFP, AAD1, Cry34 и Cry35, соответственно, для шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Перечислены значения среднего и статистические результаты.
На фиг. 56 показаны примеры результатов Вестерн-блота по белковой экспрессии Cry34, Cry35 и AAD1 для pDAB108718, pDAB108717, pDAB108719 и pDAB108720.
Способ(ы) осуществления изобретения
Создание трансгенных продуктов становится все более сложным, что требует постепенного увеличения числа вводимых трансгенов в единичный локус. Как правило, для каждого трансгена необходим уникальный промотор для экспрессии, таким образом, для экспрессии различных трансгенов в пределах одной генной кассеты необходимо несколько промоторов. Помимо увеличения размера генной кассеты, это часто приводит к повторному использованию одного и того же промотора для получения сходных уровней экспрессии различных трансгенов, контролирующих один и тот же признак. Известно, что мультигенные конструкты, управляемые одним и тем же промотором, вызывают выключение гена, таким образом, придавая трансгенным продуктам меньшую производительность. Избыток сайтов связывания транскрипционных факторов (TF) вследствие дублирования промотора может вызвать истощение количеств эндогенных ТF, приводя к инактивации транскрипции. Подавление трансгенов, по-видимому, нежелательным образом будет воздействовать на производительность трансгенного растения, выращиваемого для экспрессии трансгенов. Повторяющиеся последовательности внутри трансгена могут привести к внутрилокусной гомологичной рекомбинации генов, приводя к полинуклеотидным перестройкам.
Предоставлены способы и конструкты, комбинирующие систему двунаправленного промотора с бицистронной организацией генов либо на одном, либо на двух концах промотора, например, с использованием последовательности 2А из вируса Thoseaasigna. Белок 2А длиной всего 16-20 аминокислот расщепляет полипротеин на собственном карбоксильном конце. Это свойство "саморасщепления" или "рибосомного скачка" белка 2А или 2А-подобного пептида может быть использовано для процессинга искусственных полипротеинов, продуцируемых в трансгенных растениях. В одном варианте осуществления гены Cry34 и Cry35 сцеплены в одной генной экспрессионной кассете, в то время как гены YFP (или PhiYFP) и AAD1 сцеплены в другой генной экспрессионной кассете (с одной открытой рамкой считывания (ORF) с копией гена белка 2А, расположенного между двумя генами в каждой комбинации). Например, каждая из этих генных экспрессионных кассет (или генных пар) может быть расположена на любом конце двунаправленного промотора для управления 4 трансгенами, используя один промотор. Таким образом, представленные в настоящем описании конструкты и способы пригодны для предотвращения повторного использования одного и того же промотора и значительно уменьшают размер коммерческих конструктов. Кроме того, управление четырех или более генов одним промотором также обеспечивает возможность совместно экспрессировать гены, контролирующие один признак.
Растительные промоторы, используемые для фундаментальных исследований или биотехнологического использования, как правило, однонаправлены, управляя только одним геном, который был сцеплен на его 3'-конце (по ходу транскрипции). Часто необходимо вводить множество генов в растения для метаболической инженерии или создания многоуровневой структуры и, таким образом, необходимо, как правило, множество промоторов в будущих трансгенных культурах для управления экспрессией множества генов. Желательно разработать стратегии, которые могут сохранить число используемых промоторов и дать возможность одновременно регулировать экспрессию для генной пирамиды. В некоторых вариантах осуществления представленные двунаправленные промоторы могут управлять транскрипцией нескольких транскрипционных единиц, включая РНКi, искусственную miРНК или шпилечные последовательности РНК.
Варианты осуществления изобретения в настоящем описании используют процесс, где однонаправленный промотор из гена убиквитина-1 кукурузы (например, ZmUbi1) и промотор SCBV образуют синтетический двунаправленный промотор таким образом, что один промотор может управлять экспрессией двух генов, по одному на каждом конце промотора. Синтетические двунаправленные промоторы могут дать возможность специалистам в данной области техники создавать многоуровневые структуры трансгенов в растительных клетках и растениях, уменьшая повторное использование одного и того же промотора и уменьшая размер трансгенных конструктов. Более того, регуляция экспрессии двух генов одним синтетическим двунаправленным промотором может также обеспечить способность совместной экспрессии двух генов в одних и тех же условиях, которые могут быть полезны, например, когда каждый из двух генов вносит вклад в один признак в клетке-хозяине. Использование двунаправленной функции промоторов в растениях было продемонстрировано в некоторых случаях, включая промоторы CaMV 35 (Barfield and Pua (1991) Plant Cell Rep. 10(6-7):308-14; Xie et al. (2001)), и промотор маннопин-синтазы (mas) (Velten et al. (1984) EMBO J. 3(12):2723-30; Langridge et al. (1989) Proc. Natl. Acad. Sci. USA 86:3219-23).
Инициация транскрипции и управление генной экспрессией в генах растений регулируются разнообразными элементами последовательности ДНК, которые все вместе расположены внутри промотора. Эукариотические промоторы состоят из элемента минимального корового промотора (minP) и дополнительных, расположенных против хода транскрипции регуляторных последовательностей (URS). Элемент минимального корового промотора представляет собой минимальную цепь непрерывной последовательности ДНК, достаточной для управления точной инициации транскрипции. Элементы минимального корового промотора в растениях также содержат канонические области, связанные с инициацией транскрипции, такие как СААТ и ТАТА-боксы. Элемент ТАТА-бокс обычно расположен в области, локализованной против хода транскрипции, на расстоянии приблизительно 20-35 нуклеотидов от сайта инициации транскрипции.
Активация minP зависит от URS, с которой связываются различные белки и в дальнейшем взаимодействуют с комплексом инициации транскрипции. URS содержат последовательности ДНК, которые определяют пространственно-временной паттерн экспрессией промотора, содержащего URS. Полярность промотора часто определяется ориентацией minP, в то время как URS является биполярной (т.е. она функционирует независимо от ее ориентации). Например, синтетический однонаправленный полярный промотор CaMV 35S может быть конвертирован в двунаправленный промотор путем сцепления minP с 5'-концом промотора в противоположной ориентации. См., например, Xie et al. (2001) Nat. Biotechnol. 19(7):677-9.
В конкретных примерах некоторых вариантов осуществления изобретения элемент минимального корового промотора (minUbi1P) модифицированного промотора Ubi1 кукурузы (ZmUbi1), исходно полученного из инбредной линии Z. mays, B73, используется для создания синтетического двунаправленного промотора SCBV, который может функционировать в растениях для обеспечения характерных особенностей контроля экспрессии, которые являются уникальными по отношению к ранее доступным двунаправленным промоторам. Варианты осуществления изобретения включают синтетический двунаправленный промотор SCBV, который дополнительно включает нуклеотидную последовательность, полученную из нативного промотора SCBV. Конкретные варианты осуществления изобретения могут дополнительно включать синтетический двунаправленный промотор SCBV, содержащий интрон (например, интрон ADH) в тесной близости с SCBV и элементами последовательности minUbi1P в синтетическом двунаправленном промоторе SCBV.
Промотор ZmUbi1, исходно происходящий из В73, содержит последовательности, локализованные в геноме кукурузы внутри области протяженностью 899 оснований, расположенной с 5'-стороны сайта начала транскрипции, и дополнительно внутри области протяженностью 1093 оснований, расположенной с 3'-стороны сайта начала транскрипции. Christensen et al. (1992) Plant Mol. Biol. 18(4):675-89 (описание гена B73 ZmUbi1). Модифицированный промотор ZmUbi1, полученный из В73, который используется в некоторых примерах, представляет собой промотор протяженностью приблизительно 2 тысячи оснований, который содержит ТАТА-бокс; два перекрывающихся консенсусных элемента теплового шока; длиной 82 или 83 нуклеотида (в зависимости от исходной цепи) лидерную последовательность, непосредственно примыкающую к сайту начала транскрипции, которая называется в настоящем описании экзоном ZmUbi1; и интрон длиной 1015-1016 нуклеотидов (см. фиг.1 например). Другие варианты промотора убиквитина кукурузы, происходящие из видов Zea и генотипов Zea mays, могут проявлять высокую степень консервативности последовательности вокруг элемента minP, состоящего из элемента ТАТА и расположенных выше против хода транскрипции консенсусных элементов теплового шока. Таким образом, варианты осуществления изобретения проиллюстрированы использованием короткой (~200 нуклеотидов) высоко консервативной области (например, SEQ ID NO:1) промотора ZmUbi1 в качестве элемента минимального корового промотора для создания синтетических двунаправленных промоторов растений.
Приведенные в тексте определенные аббревиатуры перечислены в таблице 1.
|
В данном контексте единственное число включают ссылки на единственное и множественное число, если в контексте ясно и однозначно не указано иное.
Как используется в данном описании, фраза "обратное скрещивание" относится к процессу, в котором селекционер скрещивает гибридное потомство с одним из родителей, например, гибрид F1 первого поколения с одним из родоначальных генотипов гибрида F1.
Как используется в данном описании, фраза "интрон" относится к любой последовательности нуклеиновой кислоты, входящей в состав гена (или представляющей интерес экспрессируемой нуклеотидной последовательности), которая транскрибируется, но не транслируется. Интроны включают нетранслируемую последовательность нуклеиновой кислоты в пределах экспрессируемой последовательности ДНК, а также соответствующую последовательность в молекулах РНК, с них транскрибируемых.
Как используется в данном описании, фраза "изолированный" относится к биологическому компоненту (включая нуклеиновую кислоту или белок), в значительной степени сепарированному, отдельно продуцируемому или очищенному от других биологических компонентов в клетке организма, в которой компонент естественным образом встречается (т.е. других хромосомных и внехромосомных ДНК и РНК, и белках), вызывая химические или функциональные изменения в компоненте (например, нуклеиновая кислота может быть отделена от хромосомы разрушением химических связей, соединяющих нуклеиновую кислоту с оставшейся ДНК в хромосоме). Молекулы нуклеиновых кислот и белки, которые были "изолированы" включают молекулы нуклеиновых кислот и белки, очищенные стандартными методами очистки. Фраза "изолированный" также включает нуклеиновые кислоты и белки, полученные рекомбинантной экспрессией в клетке-хозяине, а также химически синтезированные молекулы нуклеиновых кислот, белков и пептидов.
Как используется в данном описании, фраза "экспрессия гена" относится к процессу, в ходе которого закодированная информация транскрипционной единицы нуклеиновой кислоты (включая, например, геномную ДНК) преобразуется в функциональную, нефункциональную или структурную часть клетки, как правило, включая синтез белка. На экспрессию гена могут влиять внешние стимулы; например, воздействие на клетку, ткань или организм агента, который увеличивает или уменьшает экспрессию гена. Экспрессия гена может также регулироваться на любом участке пути от ДНК к РНК к белку. Регуляция экспрессии гена происходит, например, посредством регуляторов, воздействующих на транскрипцию, трансляцию, транспорт и процессинг РНК, деградацию промежуточных молекул, таких как мРНК, или посредством активации, инактивации, компартментализации или деградации специфических белковых молекул после их синтеза, или путем их комбинации. Экспрессия гена может быть измерена на уровне РНК или уровне белка любым известным в данной области техники способом, включая, но без ограничений, Нозерн-блот, ОТ-ПЦР, Вестерн-блот или способ(ы) оценки активности белка in vitro, in situ или in vivo.
Как используется в данном описании, фраза "основанное на гомологии подавление экспрессии гена" (HBGS) относится к обобщенному термину, который включает как транскрипционное подавление гена, так и посттранскрипционное подавление гена. Подавление локуса-мишени несвязанным подавляющим локусом может быть обусловлено ингибированием транскрипции (транскрипционное подавление гена; TGS) или деградацией мРНК (посттранскрипционное подавление гена; PTGS) в результате образования двухцепочечной РНК (dsРНК), соответствующей промотору или транскрибируемым последовательностям, соответственно. Вовлечение отдельных клеточных компонентов в каждый процесс предполагает, что dsРНК-индуцированное TGS и PTGS, по-видимому, обусловлено расхождением древнего общего механизма. Однако точного сравнения TGS и PTGS было сложно достигнуть, поскольку оно, как правило, опирается на анализ различных молчащих локусов. Отдельно взятый трансгенный локус может быть описан как обладающий способностью инициировать TGS и PTGS в результате образования dsРНК, соответствующей промотору и транскрибируемым последовательностям различных генов-мишеней. См., например, Mourrain et al. (2007) Planta 225:365-79. По-видимому, эти siРНК по существу существующие молекулы, которые инициируют TGS и PTGS на гомологичных последовательностях: siРНК в этой модели будут инициировать подавление и метилирование гомологичных последовательностей в цис- и транс-способах путем распространения метилирования трансгенных последовательностей в эндогенный промотор.
Как используется в данном описании, фраза "молекула нуклеиновой кислоты" (или "нуклеиновая кислота" или "полинуклеотид") относится к полимерной форме нуклеотидов, которые могут включать как смысловые, так и антисмысловые цепи РНК, кДНК, геномной ДНК и синтетических форм и их смешанных полимеров. Нуклеотид может относиться к рибонуклеотиду, дезоксирибонуклеотиду или модифицированной форме любого типа нуклеотида. "Молекула нуклеиновой кислоты" в контексте настоящего описания является синонимом "нуклеиновой кислоте" или "полинуклеотиду". Длина молекулы нуклеиновой кислоты обычно составляет по меньшей мере 10 оснований, если не указано иное. Термин может относиться к молекулам РНК или ДНК промежуточной длины. Термин включает одно- и двухцепочечные формы ДНК. Молекула нуклеиновой кислоты может включать один или оба встречающихся в природе и модифицированных нуклеотида, связанных друг с другом встречающимися в природе и/или не встречающимися в природе нуклеотидными связями.
Молекулы нуклеиновых кислот могут быть модифицированы химически или биохимически, или могут содержать неестественные или являющиеся производными нуклеотидные основания, что будет очевидно специалистам в данной области техники. Такие модификации включают, например, метки, метилирование, замену одного или нескольких встречающихся в природе нуклеотидов аналогом, межнуклеотидные модификации (например, незаряженные соединения: например, метилфосфонаты, фосфотриэфиры, фосфоамидаты, карбаматы и др.; заряженные соединения: например, фосфоротиоаты, фосфородитиоаты и др.; боковые группы: например, пептиды; интеркаляторы: например, акридин, псорален и т.д.; хелаторы; алкилирующие средства; и модифицированные соединения: например, альфа-аномерные нуклеиновые кислоты и т.д.). Термин "молекула нуклеиновой кислоты" также включает любую топологическую конформацию, включая одноцепочечные, двухцепочечные, частично дуплексную, триплексную, шпилечную, кольцевую и запирающие конформации.
Транскрипция происходит в направлении 5'-3' вдоль цепи ДНК. Это означает, что РНК образуется путем последовательного добавления рибонуклеотид-5'-трифосфатов к 3'-концу растущей цепи (с соответствующим высвобождением пирофосфата). Либо в линейной, либо в кольцевой молекуле нуклеиновой кислоты дискретные элементы (например, определенные нуклеотидные последовательности) могут называться "вышерасположенными" относительно дополнительного элемента в случае, если они связаны или могли быть связаны с той же самой нуклеиновой кислотой в 5’-направлении от этого элемента. Аналогичным образом, дискретные элементы могут быть "нижерасположенными" относительно дополнительного элемента в случае, если они связаны или могли быть связаны с той же самой нуклеиновой кислотой в 3’-направлении от этого элемента.
Как используется в данном описании, фраза "положение основания" относится к расположению конкретного основания или нуклеотидного основания в пределах характеризуемой нуклеиновой кислоты. Характеризуемая нуклеиновая кислота может быть определена путем наложения (см. ниже) на референсную нуклеиновую кислоту.
Как используется в данном описании, фраза "гибридизация" относится к процессу, где олигонуклеотиды и их аналоги гибридизуются за счет образования водородных связей, включая Уотсон-Криковское, Хугстеновское или обратное Хугстеновское образование водородных связей, между комплементарными основаниями. Как правило, молекулы нуклеиновых кислот состоят из азотистых оснований, которые являются либо пиримидинами (цитозином (С), урацилом (U) и тимином (Т)), либо пуринами (аденином (А) и гуанином (G)). Эти азотистые основания образуют водородные связи между пиримидином и пурином, и связывание пиримидина с пурином называют "спаривание оснований". В частности, А будет образовывать водородную связь с Т или U, и G будет связываться с С. "Комплементарность" относится к спариванию оснований, происходящему между двумя отдельными последовательностями нуклеиновых кислот или двумя отдельными областями одной и той же последовательности нуклеиновой кислоты.
Как используется в данном описании, фразы "специфически гибридизуемый" и "специфически комплементарный" относится к достаточной степени комплементарности, такой, при которой стабильное и специфическое связывание происходит между олигонуклеотидом и ДНК или РНК-мишенью. Необязательно олигонуклеотид должен быть на 100% комплементарен последовательности-мишени, чтобы быть специфически гибридизуемым. Олигонуклеотид является специфически гибридизуемым, когда связывание олигонуклеотида с молекулой ДНК- или РНК-мишени препятствует нормальной функции ДНК- или РНК-мишени, и существует достаточная степень комплементарности для предотвращения неспецифического связывания олигонуклеотида с последовательностями, не являющимися мишенями, в условиях, где необходимо специфическое связывание, например, в физиологических условиях в случае анализов или систем in vivo. Такое связывание называют специфической гибридизацией.
Условия гибридизации, обусловливающие, в частности, степень точности взаимодействия, варьируют в зависимости от природы выбранного способа гибридизации и состава и длины гибридизующихся последовательностей нуклеиновой кислоты. Как правило, температура гибридизации и ионная сила (особенно концентрация Na+ и/или Mg2+) гибридизационного буфера влияет на точность гибридизации, хотя время промывки также влияет на точность. Вычисления, касающиеся условий гибридизации, необходимых для достижения определенной степени точности, обсуждаются в книге Sambrook et al. (ed.) Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, chs. 9 and 11.
Как используется в данном описании, фраза "жесткие условия" включают условия, при которых гибридизация будет происходить только в том случае, если имеет место менее 50% несоответствий между гибридизующейся молекулой и ДНК-мишенью. "Жесткие условия" включают дополнительные определенные уровни жесткости. Так, например, в данном контексте, условия "умеренной жесткости" являются такими, когда молекулы с более чем 50% несоответствий последовательности не будут гибридизоваться; условия "высокой жесткости" являются такими, когда молекулы с более чем 20% несоответствий последовательности не будут гибридизоваться; и условия "очень высокой жесткости" являются такими, когда молекулы с более чем 10% несоответствий последовательности не будут гибридизоваться.
В конкретных вариантах осуществления жесткие условия могут включать гибридизацию при 65°С с последующими промывками при 65°С с помощью 0,1× SSC/0,1% SDS в течение 40 минут.
Ниже представлены репрезентативные, помимо прочих, условия гибридизации:
Условия очень высокой жесткости: Гибридизация в 5× SSC буфере при 65°С в течение 16 часов; промывка дважды в 2× SSC буфере при комнатной температуре в течение 15 минут каждая; и промывка дважды в 0,5× SSC буфере при 65°С в течение 20 минут каждая.
Условия высокой жесткости: Гибридизация в 5×-6× SSC буфере при 65-70°С в течение 16-20 часов; промывка дважды в 2× SSC буфере при комнатной температуре в течение 5-20 минут каждая; и промывка дважды в 1× SSC буфере при 55-70°С в течение 30 минут каждая.
Условия умеренной жесткости: Гибридизация в 6× SSC буфере в при температуре от комнатных значений до 55°С в течение 16-20 часов; промывка по меньшей мере дважды в 2×-3× SSC буфере при температуре от комнатных значений до 55°С в течение 20-30 минут каждая.
В конкретных вариантах осуществления специфически гибридизуемые молекулы нуклеиновых кислот могут оставаться связанными в условиях очень высокой жесткости взаимодействия. В этих и дополнительных вариантах осуществления специфически гибридизуемые молекулы нуклеиновых кислот могут оставаться связанными в условиях высокой жесткости гибридизации. В этих и дополнительных вариантах осуществления специфически гибридизуемые молекулы нуклеиновых кислот могут оставаться связанными в условиях умеренной жесткости гибридизации.
Как используется в данном описании, фраза "олигонуклеотид" относится к примеру короткой нуклеиновой кислоты. Олигонуклеотиды могут быть образованы путем расщепления более длинных сегментов нуклеиновой кислоты или путем полимеризации индивидуальных нуклеотидных предшественников. Автоматизированные синтезаторы обеспечивают синтез олигонуклеотидов длиной до нескольких сотен пар оснований. Поскольку олигонуклеотиды могут связываться с комплементарной нуклеотидной последовательностью, они могут быть использованы в качестве зондов детекции ДНК или РНК. Олигонуклеотиды, состоящие из ДНК (олигодезоксирибонуклеотиды), могут быть использованы в ПЦР, способа амплификации небольших последовательностей ДНК. В ПЦР олигонуклеотид обычно называют "праймером", который позволяет ДНК-полимеразе удлинять олигонуклеотид и реплицировать комплементарную цепь.
Как используется в данном описании, фраза "идентичность последовательности" или "идентичность" относится к контексту, где две последовательности нуклеиновой кислоты или полипептида могут иметь отношение к остаткам в двух последовательностях, которые являются одинаковыми при наложении в условиях максимального соответствия в рамках установленного окна сравнения.
Как используется в данном описании, фраза "процент идентичности последовательности" относится к значению, определяемому путем сравнения двух оптимальным образом наложенных друг на друга последовательностей (например, последовательностей нуклеиновых кислот и аминокислотных последовательностей) в рамках окна сравнения, где часть последовательности в окне сравнения может включать вставки и делеции (т.е. бреши) при сопоставлении с референсной последовательностью (которая не включает вставки или делеции) для оптимального наложения двух последовательностей. Процент вычисляют путем определения числа положений, при котором имеет место идентичность нуклеотидных или аминокислотных остатков в обеих последовательностях для получения числа совпадающих положений, путем деления числа совпадающих положений на общее число положений в окне сравнения и умножения результата на 100 для получения процента идентичности последовательности.
Способы наложения последовательностей для сравнения хорошо известны в данной области техники. Различные программы и алгоритмы наложения описаны, например, в: Smith and Waterman (1981) Adv. Appl. Math. 2:482; Needleman and Wunsch (1970) J. Mol. Biol. 48:443; Pearson and Lipman (1988) Proc. Natl. Acad. Sci. U.S.A. 85:2444; Higgins and Sharp (1988) Gene 73:237-44; Higgins and Sharp (1989) CABIOS 5:151-3; Corpetetal. (1988) Nucleic Acids Res. 16:10881-90; Huang et al. (1992) Comp. Appl. Biosci. 8:155-65; Pearson et al. (1994) Methods Mol. Biol. 24:307-31; Tatiana et al. (1999) FEMS Microbiol. Lett. 174:247-50. Детальное рассмотрение способов наложения последовательностей и вычисления гомологии можно найти, например, в статье Altschul et al. (1990) J. Mol. Biol. 215:403-10.
Основной локальный инструмент поиска наложений (BLASTTM; Altschul et al. (1990)) Национального Центра Биотехнологической Информации (NCBI) доступен из нескольких источников, включая Национальный Центр Биотехнологической Информации (Bethesda, MD), и в интернете для использования вместе с несколькими программами анализа последовательностей. Описание того, как определить идентичность последовательности, используя эту программу, доступно в интернете в разделе "помощь" для BLAST™. Для сравнения последовательностей нуклеиновых кислот функция "Blast 2 sequences" программы BLAST™ (Blastn) может быть применена, используя параметры, установленные по умолчанию. Последовательности нуклеиновых кислот с гораздо большей степенью схожести с референсной последовательностью покажут увеличение процента идентичности при оценке этим способом.
Как используется в данном описании, фраза "функционально связанный" относится к контексту, где первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты находится в функциональной взаимосвязи со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с кодирующей последовательностью, когда промотор влияет на транскрипцию или экспрессию кодирующей последовательности. Рекомбинантно созданные функционально связанные последовательности нуклеиновых кислот, как правило, непрерывны и, где необходимо объединить две белок-кодирующие области, в одной и той же рамке считывания. Однако элементам необязательно надо быть непрерывными, чтобы быть функционально связанными.
Как используется в данном описании, фраза "промотор" относится к области ДНК, которая, как правило, локализована против хода транскрипции (в направлении 5' области гена), что необходимо для транскрипции. Промоторы могут обеспечить правильную активацию или репрессию гена, который они контролируют. Промотор может содержать специфические последовательности, которые распознаются транскрипционными факторами. Эти факторы могут связываться с ДНК-последовательностями промотора и приводить к рекрутированию РНК-полимеразы, фермента, который синтезирует РНК на кодирующей области гена.
Как используется в данном описании, фраза "трансформирует" или "трансдуцирует" относится к процессу, где вирус или вектор переносит молекулы нуклеиновых кислот в клетку. Клетка "трансформируется" молекулой нуклеиновой кислоты, "трансдуцированной" в клетку, когда молекула нуклеиновой кислоты становится стабильно реплицируемой клеткой, либо путем включения молекулы нуклеиновой кислоты в клеточный геном, либо путем эписомальной репликации. В данном контексте термин "трансформация" охватывает все технологии, с помощью которых молекула нуклеиновой кислоты может быть введена в такую клетку. Примеры включают, но без ограничений: трансфекцию вирусными векторами; трансформацию плазмидными векторами; электропорацию (Fromm et al. (1986) Nature 319:791-3); липофекцию (Felgner et al. (1987) Proc. Natl. Acad. Sci. USA 84:7413-7); микроинъекцию (Mueller et al. (1978) Cell 15:579-85); опосредованный Agrobacterium перенос (Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80:4803-7); прямой захват ДНК; опосредованную микроиглами трансформацию; и бомбардировку микрочастицами (Klein et al. (1987) Nature 327:70).
Как используется в данном описании, фраза "трансген" относится к экзогенной последовательности нуклеиновой кислоты. В одном примере трансген является последовательностью гена (например, гена устойчивости к гербицидам), геном, кодирующим пригодное в промышленности или фармацевтике соединение, или геном, кодирующим необходимое сельскохозяйственное свойство. В еще одном примере трансген является антисмысловой последовательностью нуклеиновой кислоты, где экспрессия антисмысловой последовательности нуклеиновой кислоты ингибирует экспрессию последовательности-мишени нуклеиновой кислоты. Трансген может содержать регуляторные последовательности, функционально связанные с трансгеном (например, промотор). В некоторых вариантах осуществления представляющая интерес последовательность нуклеиновой кислоты является трансгеном. Однако в других вариантах осуществления представляющая интерес последовательность нуклеиновой кислоты является эндогенной последовательностью нуклеиновой кислоты, где желательны дополнительные геномные копии эндогенной последовательности нуклеиновой кислоты, или последовательностью нуклеиновой кислоты, которая находится в антисмысловой ориентации относительно последовательности молекулы-мишени нуклеиновой кислоты в организме-хозяине.
Как используется в данном описании, фраза "вектор" относится к молекуле нуклеиновой кислоты, вводимой в клетку, тем самым образуя трансформированную клетку. Вектор может включать последовательности нуклеиновых кислот, обусловливающие его репликацию в клетке-хозяине, такие как точка начала репликации. Примеры включают, но без ограничений, плазмиду, космиду, бактериофаг или вирус, который доставляет экзогенную ДНК в клетку. Вектор может также содержать один или несколько генов, антисмысловых молекул и/или гены селектируемого маркера и другие генетические элементы, известные в данной области техники. Вектор может трансдуцировать, трансформировать или инфицировать клетку, тем самым заставляя клетку экспрессировать молекулы нуклеиновых кислот и/или белков, кодируемых данным вектором. Вектор может необязательно содержать средства для достижения входа молекулы нуклеиновой кислоты в клетку (например, липосому).
Как используется в данном описании, фраза "растение" включает растения и части растения, включая, но без ограничений, клетки растения и ткани растения, такие как листья, стебли, корнеплоды, цветы, пыльца и семена. Класс растений, который может быть использован в настоящем изобретении, как правило, также широк как класс высших и низших растений, поддающихся мутагенезу, включая покрытосеменные (однодольные и двудольные растения), голосеменные, папоротники и многоклеточные водоросли. Таким образом, "растение" включает двудольные растения и однодольные растения. Примеры двудольных растений включают табак, арабидопсис, сою, томат, папайю, рапс, подсолнечник, хлопок, люцерну, картофель, виноград, голубиный горох, горох, капусту декоративную, нут, сахарную свеклу, рапсу, арбуз, дыню, перец, арахис, тыкву, редис, шпинат, сквош, брокколи, капусту, морковь, цветную капусту, сельдерей, китайскую капусту, огурцы, баклажаны и салат. Примеры однодольных растений включают кукурузу, рис, пшеницу, сахарный тростник, ячмень, рожь, сорго, орхидеи, бамбук, банан, полевой хвощ, лилии, овес, лук, просо, тритикале.
Как используется в данном описании, фраза "растительный материал" относится к листьям, стеблям, корням, цветам или частям цветов, фруктам, пыльце, яйцеклеткам, зиготам, семенам, черенкам, культурам клеток или тканей или любой другой части или продукта растения. В некоторых вариантах осуществления растительный материал включает семядолю и лист.
Как используется в данном описании, фраза "переключение трансляции" относится к механизму в конце гена, обеспечивающему трансляцию близлежащего, расположенного по ходу транскрипции гена. Механизм переключения трансляции может функционировать на уровне нуклеиновой кислоты (например, вирусный или эукариотический участок внутренней посадки рибосомы (IRES), сайт альтернативного сплайсинга или сайт расщепления рибозимом) или на уровне пептида/белка (например, пептид 2А, 2А-подобный пептид, пептид интеин или сайт расщепления протеазой).
Эти механизмы переключения трансляции на уровне нуклеиновой кислоты или на уровне пептида/белка хорошо известны в данной области техники. См., например, Z. Li, H.M. Schumacher, et al. (2010) J. Biotechnol. 145(1): 9-16; Y. Chen, K. Perumal, et al. (2000) Gene Expr. 9(3):133-143; T.D. Dinkova, H. Zepeda, et al. (2005) Plant J. 41(5): 722-731; Y.L. Dorokhov, M.V. Skulachev, et al. (2002) Proc. Natl. Acad. Sci. U. S. A. 99(8): 5301-5306; O. Fernandez-Miragall and C. Hernandez (2011) PLoS One 6(7): e22617; E. Groppelli, G.J. Belsham, et al. (2007) J. Gen. Virol. 88(Pt 5): 1583-1588; S.H. Ha, Y.S. Liang, et al. (2010) Plant Biotechnol J. 8(8): 928-938; A. Karetnikov and K. Lehto (2007) J. Gen. Virol. 88(Pt 1): 286-297; A. Karetnikov and K. Lehto (2008) Virology 371(2): 292-308; M.A. Khan, H. Yumak, et al. (2009) J. Biol. Chem. 284(51): 35461-35470; и D.C. Koh, S.M. Wong, et al. (2003) J. Biol. Chem. 278(23): 20565-20573, содержание которых в полном объеме включено в данное описание посредством ссылок. Конструкты мультигенной экспрессии, содержащие модифицированные интеины, были рассмотрены в патентах США 7026526 и 7741530, а также в патентной заявке США 2008/0115243.
Как используется в данном описании, фраза "селектируемый маркер" или "ген селектируемого маркера" относится к гену, который в некоторых случаях используется в трансформации растений, например, для защиты растительных клеток от селективного агента или обеспечения устойчивости/толерантности к селективному агенту. Только такие клетки или растения, которые получают функциональный селектируемый маркер, способны делиться или расти в условиях, где присутствует селективный агент. Примеры селективных агентов могут включать, например, антибиотики, включая спектиномицин, неомицин, канамицин, паромомицин, гентамицин и гигромицин. Эти селектируемые маркеры включают ген для неомицин-фосфотрансферазы (npt II), который экспрессирует фермент, придающий устойчивость к антибиотику канамицину, и гены для соответствующих антибиотиков неомицина, паромомицина, гентамицина и G418, или ген для гигромицин-фосфотрансферазы (hpt), который экспрессирует фермент, придающий устойчивость к гигромицину. Другие гены селектируемого маркера могут включать гены, кодирующие устойчивость к гербицидам, включая Bar (устойчивость к BASTA® (глюфосинату аммония), или фосфинотрицину (РРТ)), ацетолактат-синтазе (ALS, устойчивость к ингибиторам, таким как сульфонилмочевины (SU), имидазолиноны (IMI), триазолпиримидины (TP), пиримидинилоксибензоаты (POB) и сульфониламинокарбонилтриазолиноны, которые предотвращают первый этап в синтезе аминокислот с разветвленной цепью), глифосат, 2,4-D, и устойчивость или чувствительность к металлам. Фраза "маркер-положительный" относится к растениям, которые были трансформированы для включения гена селектируемого маркера.
Различные селектируемые или детектируемые маркеры могут быть вставлены в выбранный экспрессионный вектор, чтобы обеспечить идентификацию и селекцию трансформированных растений или трансформантов. Имеется в наличии огромное количество способов подтверждения экспрессии маркеров селекции в трансформированных растениях, включая, например, секвенирование ДНК и ПЦР (полимеразная цепная реакция), Саузерн-блоттинг, РНК-блоттинг, иммунологические способы детекции белка, экспрессированного с вектора, например, преципитированного белка, который опосредует устойчивость к фосфинотрицину, или других белков, таких как репортерные гены β-глюкуронидазы (GUS), люциферазы, зеленого флуоресцентного белка (GFP), DsRed, β-галактозидазы, хлорамфеникол-ацетилтрансферазы (САТ), щелочной фосфатазы и т.п. (см. Sambrook et al., Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Press, N.Y., 2001).
Гены селектируемых маркеров используют для селекции трансформированных клеток или тканей. Гены селектируемых маркеров включают гены, кодирующие устойчивость к антибиотикам, такие, которые кодируют неомицинфосфотрансферазу II (NEO) и гигромицинфосфотрансферазу (HPT), а также гены, придающие устойчивость к гербицидным соединениям. Гены устойчивости к гербицидам, как правило, кодируют модифицированный белок-мишень, не чувствительный к гербицидам, или фермент, который расщепляет или обезвреживает гербицид в растении до начала его действия. Например, устойчивость к глифосату была получена путем использования генов, кодирующих мутантные ферменты-мишени, 5-енолпирувилшикимат-3-фосфат-синтазу (EPSPS). Гены и мутанты для EPSPS были рассмотрены в патентах США 4940835, 5188642, 5310667, 5633435, 5633448 и 6566587. Устойчивость к глюфосинату аммония, бромоксинилу и 2,4-дихлорфеноксиацетату (2,4-D) может быть получена путем использования бактериальных генов, кодирующих фосфинотрицин ацетилтрансферазу, нитрилазу или 2,4-дихлорфеноксиацетат монооксигеназу, которые обезвреживают соответствующие гербициды. Ферменты/гены для устойчивости/толерантности к глюфосинату были рассмотрены в патентах США 5273894, 5276268, 5550318 и 5561236. Ферменты/гены для устойчивости к 2,4-D были рассмотрены ранее в патентах США 6100446 и 6153401, а также в патентных заявках США 2009/0093366 и WO 2007/053482. Ферменты/гены для нитрилазы были ранее рассмотрены в патенте США 4810648.
Другие гербициды могут ингибировать точку роста или меристему, включая имидазолинон или сульфонилмочевину, и гены для устойчивости/толерантности синтетазы ацетогидроксикислот (AHAS) и ацетолактатсинтазы (ALS) для этих описанных гербицидов. Гены и мутанты для AHAS и мутантов были рассмотрены в патентах США 4761373, 5304732, 5331107, 5853973 и 5928937. Гены и мутанты для ALS были рассмотрены в патентах США 5013659 и 5141870.
Гены устойчивости к глифосату включают гены мутантной 5-енолпирувилшикимат-3-фосфат-синтазы (EPSP) (путем введения рекомбинантной нуклеиновой кислоты и/или различных форм мутагенеза in vivo нативных генов EPSP), генов aroA и генов глифосатацетилтрансферазы (GAT), соответственно). Гены устойчивости для других фосфоносоединений включают глюфосинат (гены фосфинотрицинацетилтрансферазы (РАТ) из Streptomyces species, включающих Streptomyces hygroscopicus и Streptomyces viridichromogenes), и пиридинокси- или феноксипропионовые кислоты и циклогексоны (кодирующие ингибитор ACC-азы гены). Гены устойчивости/толерантности к гербицидам ацетилкоэнзим А карбоксилазы (ACC-азы) были описаны в патентах США 5162602 и 5498544.
Молекула ДНК, кодирующая мутантный ген aroA, может быть получена под номером доступа АТСС 39256, и нуклеотидная последовательность этого мутантного гена рассмотрена в патенте США 4769061, выданном Comai, европейской патентной заявке № 0333033, поданной Kumada et al., и патенте США 4975374, выданном Goodman et al., раскрывая нуклеотидные последовательности генов глутаминсинтетазы, которые придают устойчивость к гербицидам, таким как L-фосфинотрицин. Нуклеотидная последовательность гена PAT представлена в европейской заявке №0242246, поданной Leemans et al. Также DeGreef et al., Bio/Technology 7:61 (1989) описывают получение трансгенных растений, которые экспрессируют химерные гены bar, кодирующие активность РАТ. Примерами генов, придающих устойчивость к феноксипропионовым кислотам и циклогексонам, включая сетоксидим и галоксифоп, являются гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные Marshall et al., Theor. Appl. Genet. 83:435 (1992). Гены GAT, способные придавать устойчивость к глифосату, описаны в WO 2005012515, выданном Castle et al. Гены, придающие устойчивость к 2,4-D, fop и пиридилоксиауксиновым гербицидам, описаны в WO 2005107437 и в патентной заявке США с серийным номером 11/587893.
Другие гербициды могут ингибировать фотосинтез, включая триазин (гены psbA и ls+) или бензонитрил (ген нитрилазы). Przibila et al., Plant Cell 3:169 (1991), описывают трансформацию плазмидами Chlamydomonas, кодирующими мутантные гены psbA. Нуклеотидные последовательности для генов нитрилазы рассмотрены в патенте США 4810648, выданном Stalker, и молекулы ДНК, содержащие эти гены, доступные под номерами доступа АТСС 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, описаны в работе Hayes et al., Biochem. J. 285:173 (1992).
В контексте настоящего изобретения гены селектируемых маркеров включают, но без ограничений, гены, кодирующие: неомицинфосфотрансферазу II (Fraley et al. (1986) CRC Critical Reviews in Plant Science 4:1-25); цианамид гидратазу (Maier-Greiner et al. (1991) Proc. Natl. Acad. Sci. USA 88:4250-4264); аспартат-киназу; дигидродипиколинат-синтазу (Perl et al. (1993) Bio/Technology 11:715-718); триптофан-декарбоксилазу (Goddijn et al. (1993) Plant Mol. Biol. 22:907-912); дигидродипиколинат-синтазу и десенсибилизированную аспартат-киназу (Perl et al. (1993) Bio/Technology 11:715-718); ген bar (Toki et al. (1992) Plant Physiol. 100:1503-1507; и Meagher et al. (1996), Crop Sci. 36:1367); триптофан-декарбоксилазу (Goddijn et al. (1993) Plant Mol. Biol. 22:907-912); неомицин-фосфотрансферазу (NEO) (Southern et al. (1982) J. Mol. Appl. Gen. 1:327; гигромицин-фосфотрансферазу (HPT или HYG) (Shimizu et al. (1986) Mol. Cell Biol. 6:1074); дигидрофолатредуктазу (DHFR) (Kwok et al. (1986) PNAS USA 4552); фосфинотрицин-ацетилтрансферазу (DeBlock et al. (1987) EMBO J. 6:2513); дегалогеназу 2,2-дихлорпропионовой кислоты (Buchanan-Wollatron et al. (1989) J. Cell. Biochem. 13D:330); синтазу ацетогидроксикислоты (патент США 4761373, выданный Anderson et al.; Haughn et al. (1988) Mol. Gen. Genet. 221:266); 5-енолпирувилшикиматфосфатсинтазу (aroA) (Comai et al. (1985) Nature 317:741); галоарилнитрилазу (опубликованная заявка PCT WO 87/04181, выданная Stalker et al.); ацетил-коэнзим А-карбоксилазу (Parker et al. (1990) Plant Physiol. 92:1220); дигидроптероатсинтазу (sul l; Guerineau et al. (1990) Plant Mol. Biol. 15:127) и полипептид 32 кДа фотосистемы II (psbA) (Hirschberg et al. (1983) Science 222:1346).
Включены также гены, кодирующие устойчивость к: хлорамфениколу (Herrera-Estrella et al. (1983) EMBO J. 2:987-992); метотрексату (Herrera-Estrella et al. (1983) Nature 303:209-213; Meijer et al. (1991) Plant Mol. Biol. 16:807-820); гигромицину (Waldron et al. (1985) Plant Mol. Biol. 5:103-108; Zhijian et al. (1995) Plant Science 108:219-227; и Meijer et al. (1991) Plant Mol. Biol. 16:807-820); стрептомицину (Jones et al. (1987) Mol. Gen. Genet. 210:86-91); спектиномицину (Bretagne-Sagnard et al. (1996) Transgenic Res. 5:131-137); блеомицину (Hille et al. (1986) Plant Mol. Biol. 7:171-176); сульфонамиду (Guerineau et al. (1990) Plant Mol. Biol. 15:127-136); бромоксинилу (Stalker et al. (1988) Science 242:419-423); 2,4-D (Streber et al. (1989) Bio/Technology 7:811-816); глифосату (Shaw et al. (1986) Science 233:478-481); и фосфинотрицину (DeBlock et al. (1987) EMBO J. 6:2513-2518).
Представленный выше список генов селектируемых маркеров и репортеров не ограничен. Любой ген репортера или селектируемого маркера включен в рассмотрение настоящим изобретением. Если необходимо, такие гены могут быть секвенированы способами, известными в данной области техники.
Гены репортера и селектируемого маркера синтезируют для оптимальной экспрессии в растениях. Другими словами, кодирующая последовательность гена модифицируют для повышения экспрессии в растениях. Синтетический маркерный ген создают для экспрессии в растениях на более высоком уровне, обусловливая более высокую эффективность трансформации. Способы синтетической оптимизации генов доступны в данной области техники. Известно, что несколько генов были оптимизированы для повышения экспрессии продукта гена в растениях.
Последовательность маркерного гена может быть оптимизирована для экспрессии в определенном виде растений или, альтернативно, может быть модифицирована для оптимальной экспрессии в семействах растений. Предпочтительные кодоны растений могут быть определены из кодонов высокой встречаемости в белках, экспрессирующихся в наибольшем количестве в определенных представляющих интерес видах растений. См., например, EPA 0359472; EPA 0385962; WO 91/16432; Perlak et al. (1991) Proc. Natl. Acad. Sci. USA 88:3324-3328; and Murray et al. (1989) Nucleic Acids Research 17: 477-498; патент США 5380831 и патент США 5436391. Таким способом нуклеотидные последовательности могут быть оптимизированы для экспрессии в любом растении. Считается, что вся или часть последовательности гена может быть оптимизирована или быть синтетической. Другими словами, полностью оптимизированные или частично оптимизированные последовательности могут быть также использованы.
Гены, которые придают устойчивость к гербициду:
A. Устойчивость/толерантность синтазы ацетогидроксикислот (AHAS) и ацетолактатсинтазы (ALS) к гербицидам имидазолинону или сульфонилмочевине. Гены и мутантны AHAS и мутанты были рассмотрены в патентах США 4761373, 5304732, 5331107, 5853973 и 5928937. Гены и мутанты для ALS были рассмотрены в патентах США 5013659 и 5141870.
В. Гены устойчивости/толерантности ацетилкоэнзима А карбоксилазы (АССазы) к гербицидам циклогександионам и/или арилоксифеноксипропионовой кислоте (включая галоксифоп, диклофоп, феноксипроп, флуазифоп, хизалофоп) были описаны в патентах США 5162602 и 5498544.
С. Гены устойчивости/толерантности к глифосату. Ген 5-енолпирувил-3-фосфошикиматсинтазы (ES3P синтаза) была описана в патенте США 4769601. Гены 5-енолпирувилшикимат-3-фосфатсинтазы (EPSPS) и мутанты были описаны в патентах США 4940835, 5188642, 5310667, 5633435, 5633448 и 6566587.
D. Гены устойчивости/толерантности к глюфосинату (биалофосу, фосфинотрицину (РРТ)). Гены для фосфинотрицинацетилтрансферазы (Pat) были описаны в патентах США 5273894, 5276268 и 5550318; и ген устойчивости к биалафосу (Bar) был описан в патентах США 5561236 и 5646024, 5648477 и 7112665. Ген для глутаминсинтетазы (GS) был описан в патенте США 4975372 и европейской патентной заявке ЕР 0333033 А1.
Е. Гены устойчивости/толерантности гидроксифенилпируватдиоксигеназы (HPPD) к гербицидам изоксазолу, дикетоннитрилам и/или трикетонам, включая сулкотрион и мезотрион, были описаны в патентах США 6268549 и 6069115.
F. Гены устойчивости/толерантности к 2,4-D. Гены 2,4-D-монооксигеназы были описаны в патентах США 6100446 и 6153401. Дополнительные гены устойчивости/толерантности к 2,4-D рассмотрены в US 2009/0093366 и WO 2007/053482.
G. Гены имидазолглицеролфосфатдегидратазы (IGPD) против гербицидов имидазола и/или триазола были описаны в патенте США 5541310. Гены разрушающих дикамба ферментов (оксигеназы, ферредоксина и редуктазы) против гербицида дикамба были рассмотрены в патентах США 7022896 и 7105724.
H. Гены для гербицидов, которые ингибируют фотосинтез, включая триазин (гены psbA и 1s+) или бензонитрил (ген нитрилазы). См., например, Przibila et al., Plant Cell 3:169 (1991), рассматривающих трансформацию плазмидами Chlamydomonas, кодирующими мутантные гены psbA. Нуклеотидные последовательности для генов нитрилазы рассмотрены в патенте США 4810648, и молекулы ДНК, содержащие эти гены, доступны под номерами доступа АТСС 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион S-трансферазу, описаны Hayes et al., Biochem. J. 285:173 (1992).
Если специальным образом не оговорено иное, все технические и научные термины, используемые в настоящем описании, имеют одно и то же значение, как принято понимать специалистами в данной области техники, к которой относится данное описание изобретения. Определения общепринятых терминов в молекулярной биологии можно найти, например, в: Lewin, Genes V, Oxford University Press, 1994 (ISBN 0-19-854287-9); Kendrew et al. (eds.), The Encyclopedia of Molecular Biology, Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и Meyers (ed.), Molecular Biology and Biotechnology: A Comprehensive Desk Reference, VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Данное изобретения предоставляет молекулы нуклеиновых кислот, включающих синтетическую нуклеотидную последовательность, которая может функционировать как двунаправленный промотор. В некоторых вариантах осуществления синтетический двунаправленный промотор может быть функционально связан с одной или двумя представляющими интерес последовательностями. Например, синтетический двунаправленный промотор может быть функционально связан с одной или двумя представляющими интерес нуклеотидными последовательностями (например, двумя генами, по одному на каждом конце промотора) таким образом, чтобы регулировать транскрипцию по меньшей мере одной (например, одной или обеих) из представляющих интерес нуклеотидных последовательностей. Путем вставки URS из промотора SCBV в синтетический двунаправленный промотор может быть достигнута определенная экспрессия или регуляторный паттерн (например, такие, которые демонстрируют гены под контролем промотора SCBV) в отношении представляющей интерес нуклеотидной последовательности, которая функционально связана с синтетическим двунаправленным промотором.
Некоторые варианты осуществления изобретения представлены в настоящем описании в качестве примеров в виде включения элемента минимального корового промотора из однонаправленного промотора гена убиквитина-1 кукурузы (ZmUbi1) в молекулярный контекст, отличающийся от нативного промотора, для создания синтетического двунаправленного промотора. Элемент минимального корового промотора обозначен в настоящем описании как "minUbi1P" и приблизительно имеет длину 200 нуклеотидов. Секвенирование и анализ элементов minUbi1P из множества видов Zea и генотипов Z. mays выявили, что функциональные элементы minUbi1P высоко консервативны, вследствие чего элемент minUbi1P может сохранять свою функцию в качестве инициатора транскрипции, если он обладает, например, по меньшей мере приблизительно 75%; по меньшей мере, приблизительно 80%; по меньшей мере, приблизительно 85%; по меньшей мере, приблизительно 90%; по меньшей мере, приблизительно 91%; по меньшей мере, приблизительно 92%; по меньшей мере, приблизительно 93%; по меньшей мере, приблизительно 94%; по меньшей мере, приблизительно 95%; по меньшей мере, приблизительно 96%; по меньшей мере, приблизительно 97%; по меньшей мере, приблизительно 98%; по меньшей мере, приблизительно 99% и/или по меньшей мере приблизительно 100% идентичностью последовательности элемента minUbi1P в виде SEQ ID NO:1. Характерные свойства элементов minUbi1P, которые могут быть пригодны в некоторых вариантах осуществления могут включать, например, но без ограничений, указанную выше высокую консервативность нуклеотидной последовательности; присутствие по меньшей мере одного ТАТА-бокса; и/или присутствие по меньшей мере одного (например, двух) консенсусного элемента теплового шока. В определенных элементах minUbi1P более чем один консенсусный элемент теплового шока может перекрываться с последовательностью minUbi1P.
В некоторых вариантах осуществления процесс включения элемента minUbi1P в молекулярный контекст, отличный от контекста нативного промотора, для создания синтетического двунаправленного промотора, может заключаться во введении элемента minUbi1P в нуклеиновую кислоту промотора SCBV, изменяя ориентацию элемента minUbi1P относительно оставшейся последовательности промотора SCBV. Таким образом, синтетический двунаправленный промотор SCBV может содержать элемент минимального корового промотора minUbi1P, расположенный с 3'-конца нуклеотидной последовательности промотора SCBV и в обратной ориентации относительно последней таким образом, что он может быть функционально связан с представляющей интерес нуклеотидной последовательностью, расположенной с 3'-конца нуклеотидной последовательности промотора SCBV. Например, элемент minUbi1P может быть вставлен на 3'-конце промотора SCBV в обратной ориентации.
Синтетический двунаправленный промотор SCBV может также содержать один или несколько дополнительных элементов последовательности в дополнение к элементу minUbi1P и элементам нативного промотора SCBV. В некоторых вариантах осуществления синтетический двунаправленный промотор SCBV может содержать промоторную URS; экзон (например, лидерный или сигнальный пептид); интрон; последовательность спейсера и/или одну или несколько их комбинаций. Например, но без ограничений, синтетический двунаправленный промотор SCBV может содержать последовательность URS из промотора SCBV; интрон из гена Ubi1; и их комбинацию.
В некоторых таких примерах, включающих синтетический двунаправленный промотор SCBV, содержащий промоторную URS, URS может быть выбрана для придания определенных регуляторных свойств синтетическому промотору. Известные промоторы широко вариабельны по типу контроля, который они осуществляют в отношении функционально связанных генов (например, ответ на факторы окружающей среды, стимуляторы развития и пространственную информацию), и URS, вставленная в гетерологичный промотор обычно сохраняет тип контроля, свойственный URS в отношении ее нативного промотора и функционально связанного(ых) гена(ов). Langridge et al. (1989), см. выше. Примеры эукариотических промоторов, которые были охарактеризованы и могут содержать URS в составе синтетического двунаправленного промотора Ubi1 в соответствии с некоторыми вариантами осуществления включают, например, но без ограничений: промоторы, как описано в патентах США 6437217 (промотор RS81 кукурузы); 5641876 (промотор актина риса); 6426446 (промотор RS324 кукурузы); 6429362 (промотор PR-1 кукурузы); 6232526 (промотор А3 кукурузы); 6177611 (конститутивные промоторы кукурузы); 6433252 (промотор олеозина L3 кукурузы); 6429357 (промотор актина 2 риса, и интрон актина 2 риса); 5837848 (специфичные для корня промотор); 6294714 (индуцибельные светом промоторы); 6140078 (индуцибельные солью промоторы); 6252138 (патоген-индуцибельные промоторы); 6175060 (индуцибельные дефицитом фосфора промоторы); 6388170 (двунаправленный промоторы); 6635806 (промоторы гамма-коиксина); и патентная заявка США с серийным номером 09/757089 (промотор альдолазы хлоропласта кукурузы).
Дополнительные иллюстративные прокариотические промоторы включают промотор нопалинсинтазы (NOS) (Ebert et al. (1987) Proc. Natl. Acad. Sci. USA 84(16):5745-9); промотор октопинсинтазы (OCS) (который работает в опухоль-индуцирующих плазмидах Agrobacterium tumefaciens); промоторы колимовируса, такие как промотор 19S вируса мозаики цветной капусты (CaMV) (Lawton et al. (1987) Plant Mol. Biol. 9:315-24); промотор 35S CaMV (Odell et al. (1985) Nature 313:810-2; 35S-промотор вируса мозаики норичника (Walker et al. (1987) Proc. Natl. Acad. Sci. USA 84(19):6624-8); промотор синтазы сахарозы (Yang and Russell (1990) Proc. Natl. Acad. Sci. USA 87:4144-8); сложный промотор гена R (Chandler et al. (1989) Plant Cell 1:1175-83); CaMV35S (патенты США 5322938, 5352605, 5359142 и 5530196); FMV35S (патенты США 6051753 и 5378619); промотор PC1SV (патент США 5850019); промотор SCP1 (патент США 6677503); и промоторы AGRtu.nos (код доступа в GenBank V00087; Depicker et al. (1982) J. Mol. Appl. Genet. 1:561-73; Bevan et al. (1983) Nature 304:184-7) и т.п.
В некоторых вариантах осуществления синтетический двунаправленный промотор SCBV может дополнительно содержать экзон. Например, в примерах требуется целенаправленно воздействовать на или осуществлять транспорт полипептида, кодируемого представляющей интерес нуклеотидной последовательностью, функционально связанной с промотором, в определенной внутриклеточной структуре и/или компартменте. В этих и других вариантах осуществления кодирующая последовательность (экзон) может быть вставлена в молекулу нуклеиновой кислоты между оставшейся последовательностью синтетического двунаправленного промотора SCBV и нуклеотидной последовательностью, кодирующей полипептид. Эти элементы могут быть размещены на усмотрение практикующего специалиста в данной области таким образом, чтобы синтетический двунаправленный промотор SCBV запускал экспрессию полипептида (или одной, или обеих последовательностей, кодирующих полипептид, которые функционально связаны с промотором), включая пептид, кодируемый вставленной кодирующей последовательностью, обеспечивая функциональную взаимосвязь с оставшейся частью полипептида. В конкретных примерах может быть вставлен экзон, кодирующий лидерный, транспортный или сигнальный пептид (например, лидерный пептид Ubi1).
Пептиды, которые могут кодироваться экзоном, вставленным в синтетический двунаправленный промотор Ubi1, включают, например, но без ограничений: лидерный пептид убиквитина (например, Ubi1); транспортный пептид хлоропласта (СТР) (например, EPSPS CTP A. Thaliana (Klee et al. (1987) Mol. Gen. Genet. 210:437-42), и EPSPS CTP Petunia hybrid (della-Cioppa et al. (1986) Proc. Natl. Acad. Sci. USA 83:6873-7)), как приведено в качестве примера направленного воздействия на дикамба-монооксигеназу в хлоропластах в международной публикации РСТ № WO 2008/105890.
Интроны могут быть также включены в синтетический двунаправленный промотор SCBV в некоторых вариантах осуществления, например, между оставшейся синтетической последовательностью двунаправленного промотора SCBV и представляющей интерес нуклеотидной последовательностью, которая функционально связана с промотором. В некоторых вариантах осуществления интрон, вставленный в синтетический двунаправленный промотор SCBV, может представлять собой, но без ограничений, 5' UTR, которая функционирует в качестве лидерной последовательности трансляции, которая присутствует в полностью процессированной мРНК, расположенной выше последовательности начала трансляции (такая лидерная последовательность трансляции может влиять на процессинг первичного транскрипта с образованием мРНК, стабильность мРНК и/или эффективность трансляции). Примеры лидерных последовательностей трансляции включают лидерные последовательности белков теплового шока кукурузы и петунии (патент США 5362865), лидерные последовательности белка оболочки вирусов растений, лидерные последовательности рибулозо-1,5-бифосфаткарбоксилазы/оксигеназы растений и другие. См., например, Turner and Foster (1995) Molecular Biotech. 3(3):225-36. Неограниченное число примеров 5’ UTR включает GmHsp (патент США 5659122); PhDnaK (патент США 5362865); AtAnt1; TEV (Carrington and Freed (1990) J. Virol. 64:1590-7); и AGRtunos (идентификационный номер в GenBank V00087; и Bevan et al. (1983) Nature 304:184-7). В конкретных примерах, Ubi1 и/или ADH интрон(ы) могут быть включены в синтетический двунаправленный промотор SCBV.
Дополнительные последовательности, которые могут в некоторых случаях быть включены в синтетический двунаправленный промотор SCBV, включают, например, но без ограничений: 3' нетранслируемые последовательности; 3' участки терминации транскрипции и участки полиаденилирования. Существуют генетические элементы, расположенные ниже представляющей интерес нуклеотидной последовательности (например, представляющая интерес последовательность, которая функционально связана с синтетическим двунаправленным промотором SCBV), и включают полинуклеотиды, которые представляют собой сигнал полиаденилирования и/или другие регуляторные сигналы, способные влиять на транскрипцию, процессинг мРНК или экспрессию гена. Сигнал полиаденилирования может функционировать у растений для присоединения полиаденинов к 3'-концу предшественника мРНК. Последовательность полиаденилирования может быть получена из встречающегося в природе гена, из различных генов растений или из генов Т-ДНК. Неограничивающим примером 3'-области терминации транскрипции является 3'-область нопалин-синтазы (nos 3'; Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80:4803-7). Пример использования различных 3’ нетранслируемых областей представлен в Ingelbrecht et al. (1989), Plant Cell 1:671-80. Неограниченные примеры сигналов полиаденилирования включают сигналы из гена RbcS2 Pisum Sativum (Ps.RbcS2-E9; Coruzzi et al. (1984) EMBO J. 3:1671-9) и AGRtu.nos (идентификационный номер E01312 в GenBank).
В некоторых вариантах осуществления синтетический двунаправленный промотор SCBV содержит одну или более нуклеотидных последовательностей, которые облегчают позиционирование нуклеиновой кислоты, включающей промотор, с определенным локусом в геноме организма-мишени. Например, могут быть включены одна или более последовательностей, которые гомологичны сегментам последовательности геномной ДНК в хозяине (например, редким или уникальным последовательностям геномной ДНК). В некоторых примерах эти гомологичные последовательности могут управлять рекомбинацией и интеграцией нуклеиновой кислоты, состоящей из синтетического двунаправленного промотора SCBV, в сайте гомологичной ДНК в геноме-хозяине. В конкретных примерах синтетический двунаправленный промотор SCBV содержит одну или более нуклеотидных последовательностей, которые облегчают позиционирование нуклеиновой кислоты, содержащей промотор, в редком или уникальном участке в геноме хозяина, используя созданные нуклеазные ферменты, которые распознают последовательность в редких или уникальных участках и облегчают интеграцию в редкий или уникальный участок. Такая направленная система интеграции, использующая эндонуклеазы типа цинковые пальцы, как нуклеазный фермент, описана в патентной заявке США 13/011735.
Нуклеиновые кислоты, содержащие синтетический двунаправленный промотор SCBV, могут быть получены, используя любой способ, известный в данной области техники, включая, например, но без ограничений: RCA; ПЦР-амплификация; ОТ-ПЦР-амплификация; OLA и SNuPE. Эти и другие эквивалентные технологии хорошо известны специалистам в данной области техники и дополнительно описаны в деталях, например, но без ограничений, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory, 2001; и Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, 1998.
Доставка и/или трансформация: Настоящее изобретение также предоставляет способы трансформации клетки молекулой нуклеиновой кислоты, содержащей синтетический двунаправленный промотор SCBV. Любая из большого числа технологий, известных в данной области техники, по введению молекул нуклеиновых кислот в растения может быть использована для трансформации растения молекулой нуклеиновой кислоты, содержащей синтетический двунаправленный промотор SCBV в соответствии с некоторыми вариантами осуществления, например, для введения одного или нескольких синтетических двунаправленных промоторов SCBV в хозяйский геном растения, и/или для дополнительного введения одной или более представляющих интерес молекул нуклеиновой кислоты, функционально связанных с промотором.
Пригодные способы трансформации растений включают любой способ, с помощью которого ДНК может быть введена в клетку, например, но без ограничений: электропорацию (см., например, патент США 5384253); бомбардировку микрочастицами (см., например, патенты США 5015580, 5550318, 5538880, 6160208, 6399861 и 6403865); опосредованную Agrobacterium трансформацию (см., например, патенты США 5635055, 5824877, 5591616; 5981840 и 6384301); и трансформацию протопласта (см., например, патент США 5508184). Путем применения технологий, таких как перечисленные выше, клетки практически любых видов растений могут быть стабильно трансформированы, и эти клетки могут быть преобразованы в трансгенные растения способами, известными специалистам в данной области техники. Например, способы, которые могут быть особенно пригодны в контексте трансформации хлопчатника, описаны в патентах США 5846797, 5159135, 5004863 и 6624344; способы трансформации растений Brassica, в частности, описаны, например, в патенте США 5750871; способы трансформации сои описаны, например, в патенте США 6384301; и способы трансформации кукурузы описаны, например, в патентах США 7060876 и 5591616 и международной публикации РСТ WO 95/06722.
После произведенной доставки экзогенной нуклеиновой кислоты в клетку-реципиента, как правило, идентифицируют трансформированную клетку для дальнейшего культивирования и регенерации растения. Для улучшения возможности идентифицировать трансформантов можно использовать маркер селекции или скрининга с использованием трансформационного вектора для образования трансформанта. В этом случае, потенциально трансформированная клеточная популяция может быть проанализирована, подвергая клетки воздействию селективного агента или агентов, или можно провести скрининг клеток по заданному характерному свойству маркерного гена.
Клетки, которые выживают после воздействия селективного агента, или клетки, которым были выставлены положительные бальные оценки в скрининговом анализе, могут быть культивированы в средах, поддерживающих регенерацию растений. В некоторых вариантах осуществления любые подходящие среды культивирования растительной ткани (например, среды MS и N6) могут быть модифицированы добавлением дополнительных веществ, таких как регуляторы роста. Ткань можно поддерживать в основной среде с регуляторами роста, пока не будет образовано достаточно ткани для начала регенерации растения или после повторенных раундов ручной селекции до тех пор, пока морфология ткани не станет пригодной для регенерации (например, по меньшей мере 2 недели), затем ее переносят в среды, способствующие образованию побегов. Культуры периодически пересаживают, пока не будет образовано достаточное количество побегов. Как только сформируются побеги, их пересаживают в среды, способствующие образованию корневой системы. Как только будет сформирована корневая система, растения могут быть пересажены в грунт для дальнейшего роста и созревания.
Для подтверждения присутствия требуемой молекулы нуклеиновой кислоты, содержащей синтетический двунаправленный промотор SCBV, в регенерирующих растениях могут быть проведены различные анализы. Такие анализы включают, например: молекулярно-биологические анализы, такие как Саузерн- и Нозерн-блоттинг и ПЦР; биохимические анализы, такие как детекция наличия белкового продукта, например, иммунологическими способами (ELISA и/или Вестерн-блот) или оценивая ферментативную функцию; анализы частей растения, таких как анализы листьев и корней; и анализы фенотипа всего регенерированного растения.
Направленные интеграционные события могут быть подвергнуты скринингу, например, с помощью ПЦР-амплификации, используя, например, олигонуклеотидные праймеры, специфичные для представляющих интерес молекул нуклеиновых кислот. В понятие ПЦР-генотипирование входят, но без ограничений, амплификация в ходе полимеразной цепной реакции (ПЦР) геномной ДНК, полученной из выделенной хозяйской каллюсной ткани растения, в которой предполагают наличие представляющей интерес молекулы нуклеиновой кислоты, интегрированной в геном, с последующим стандартным клонированием и секвенированием продуктов ПЦР-амплификации. Способы ПЦР-генотипирования подробно описаны (см., например, Rios et al. (2002), Plant J. 32:243-53) и могут быть применены к геномной ДНК, полученной из любых видов растений или типов ткани, включая клеточные культуры. Комбинации олигонуклеотидных праймеров, которые связываются как с последовательностью-мишенью, так и введенной последовательностью, могут быть использованы последовательно или мультиплексировано в реакциях ПЦР-амплификации. Могут быть синтезированы олигонуклеотидные праймеры, рассчитанные для отжига с сайтом-мишенью, введенными нуклеотидными последовательностями и/или их комбинацией. Таким образом, ПЦР-генотипирование может включать, например, но без ограничений: амплификацию специфических последовательностей в растительном геноме; амплификацию множественных специфических последовательностей в растительном геноме; амплификацию неспецифических последовательностей в растительном геноме и их комбинации. Специалист в данной области техники может разработать дополнительные комбинации праймеров и реакций амплификации для детального исследования генома. Например, набор прямых и обратных олигонуклеотидных праймеров может быть создан для отжига с последовательностью(ями) нуклеиновой(ых) кислот(ы), специфичных для мишени за пределами границ введенной последовательности нуклеиновой кислоты.
Прямой и обратный олигонуклеотидные праймеры могут быть сконструированы для специфического отжига с введенной молекулой нуклеиновой кислоты, например, в последовательности, соответствующей кодирующей области внутри представляющей интерес нуклеотидной последовательности, имеющей ее в своем составе, или другие части молекулы нуклеиновой кислоты. Эти праймеры могут быть использованы в сочетании с праймерами, описанными выше. Олигонуклеотидные праймеры могут быть синтезированы в соответствии с представляющей интерес последовательностью, и коммерчески доступны (например, от компании Integrated DNA Technologies, Inc., Coralville, IA). За амплификацией может следовать клонирование или секвенирование, или прямое секвенирование продуктов амплификации. Специалист в данной области техники сможет предусмотреть альтернативные способы анализа продуктов амплификации, полученных в ходе ПЦР-генотипирования. В одном варианте осуществления изобретения олигонуклеотидные праймеры, специфичные для гена-мишени, применяются в ПЦР-амплификациях.
Некоторые варианты осуществления настоящего изобретения также предоставляют клетки, включающие синтетический двунаправленный промотор SCBV, например, который может присутствовать в конструкте нуклеиновой кислоты. В конкретных примерах, синтетический двунаправленный промотор SCBV в соответствии с некоторыми вариантами осуществления изобретения может быть использован в качестве регуляторной последовательности для регуляции экспрессии трансгенов в клетках растений и растениях. В некоторых таких примерах использование синтетического двунаправленного промотора SCBV, функционально связанного с представляющей интерес нуклеотидной последовательностью (например, трансгеном), может уменьшать число гомологичных промоторов, необходимых для регуляции экспрессии заданного числа представляющих интерес нуклеотидных последовательностей, и/или уменьшать размер конструкта(ов) нуклеиновой кислоты, необходимых для введения заданного числа представляющих интерес нуклеотидных последовательностей. Более того, использование синтетического двунаправленного промотора SCBV может обеспечить совместную экспрессию двух представляющих интерес функционально связанных нуклеотидных последовательностей в одних и тех же условиях (т.е. условиях, при которых промотор SCBV является активным). Такие примеры могут быть особенно результативными, например, когда каждая из двух представляющих интерес оперативно связанных нуклеотидных последовательностей вносит вклад в характерный признак в трансгенном хозяине, содержащем представляющие интерес нуклеотидные последовательности, и совместная экспрессия представляющих интерес нуклеотидных последовательностей предпочтительно оказывает воздействие на характерный признак в трансгенном хозяине.
В некоторых вариантах осуществления трансгенное растение, содержащее один или несколько синтетических двунаправленных промоторов SCBV и/или представляющую(ие) интерес нуклеотидную(ые) последовательность(и) могут обусловливать присвоение одного или более желаемых характерных свойств (например, привнесенных, усиленных или способствующих чему-либо) путем экспрессии представляющей(их) интерес нуклеотидной(ых) последовательности(ей) в растении. Такие характерные свойства могут включать, например, но без ограничений: устойчивость к насекомым, другим сельскохозяйственным вредителям и вызывающих заболевания агентам; толерантность к гербицидам; повышенную стабильность, выход или срок годности; устойчивость к условиям окружающей среды; фармацевтическую продукцию; промышленное производство и улучшение питательных веществ. В некоторых примерах, желательное характерное свойство можно придать с помощью трансформации растения молекулой нуклеиновой кислоты, содержащей синтетический двунаправленный промотор SCBV, функционально связанный с представляющей интерес нуклеотидной последовательностью. В некоторых примерах, желаемое характерное свойство можно придать растению, продуцируемому в виде потомства растения посредством селекции, чье желаемое характерное свойство можно придать одной или более представляющими интерес нуклеотидными последовательностями, функционально связанными с синтетическим двунаправленным промотором SCBV, которое(ые) передается(ются) растению от родительского растения, содержащего представляющую интерес нуклеотидную последовательность, функционально связанную с синтетическим двунаправленным промотором SCBV.
Трансгенное растение в соответствии с некоторыми вариантами осуществления изобретения может быть любым растением, способным к трансформированию молекулой нуклеиновой кислоты по изобретению или давать потомство с растением, трансформированным молекулой нуклеиновой кислоты по изобретению. В соответствии с этим, растение может быть двудольным или однодольным. Неограниченные примеры двудольных растений для использования в некоторых примерах включают: люцерну; бобы; брокколи; капусту; канолу; морковь; цветную капусту; сельдерей; китайскую капусту; хлопчатник; огурец; баклажан; салат; дыню; горох; перец; арахис; картофель; тыкву; редис; редьку; шпинат; сою; сквош; сахарную свеклу; подсолнечник; табак; помидор и арбуз. Неограниченные примеры однодольных растений для использования в некоторых примерах включают: кукурузу; лук; рис; сорго; пшеницу; рожь; просо; сахарный тростник; овес; тритикале; просо и газонную траву.
В некоторых вариантах осуществления трансгенное растение может быть использовано или культивировано любым способом, где желательно наличие синтетического двунаправленного промотора SCBV и/или представляющей интерес функционально связанной нуклеотидной последовательности. Соответственно, такие трансгенные растения могут быть созданы, в том числе обладать одним или более характерными свойствами, путем трансформации молекулами нуклеиновых кислот по изобретению, и могут быть посажены и/или культивированы любым способом, известным специалисту в данной области техники.
Несмотря на то, что изобретение было описано со ссылкой на конкретные способы и варианты осуществления следует принимать во внимание, что могут быть сделаны различные модификации и изменения без отступления от сути изобретения.
Приведенные ниже примеры представлены для иллюстрации определенных конкретных характеристик и/или вариантов осуществления изобретения. Примеры не следует рассматривать как ограничение сути изобретения проиллюстрированными конкретными характеристиками или вариантами осуществления изобретения.
ПРИМЕРЫ
ПРИМЕР 1: Трансформация и экспрессия
Трансформация Agrobacterium tumefaciens: трехкомпонентный штамм DAt13192 Agrobacterium tumefaciens трансформировали бинарным вектором pDAB108706 (предварительная патентная заявка США №61/368965). Бактериальные колонии выделяли, и бинарную плазмидную ДНК экстрагировали и подтверждали рестрикционным анализом.
Трансформация кукурузы: стерилизация початка и выделение зародышей. Для получения незрелых зародышей, растения Zea mays (c.v. B104) выращивали в теплице, используя процесс самоопыления или близкородственного скрещивания для образования початка. Початки собирали приблизительно на 9-12 день после опыления. В день эксперимента поверхность початков стерилизовали погружением в 20% раствор хозяйственного отбеливателя, который содержал 5% гипохлорит натрия, встряхивали в течение 20-30 минут с последующим трехкратным ополаскиванием в стерильной воде. После стерилизации, незрелые зиготные зародыши (1,5-2,2 мм) в асептических условиях вырезали из каждого початка и случайным образом распределяли в микроцентрифужных пробирках, содержащих жидкую среду для заражения (базальную среду LS, 4,43 г/л; раствор витаминов N6 [1000X], 1,00 мл/л; L-пролин, 700,0 мг/л; сахарозу 68,5 г/л; глюкозу 36 г/л; 2,4-D 1,5 мг/л). Для данного набора экспериментов объединенные в пулы зародыши от 2-3 початков использовали для каждой обработки.
Закладка культуры Agrobacterium: глицериновые стоковые образцы штаммов Agrobacterium, содержащих бинарные векторы, описанные выше, высевали штрихом в планшеты с минимальной средой АВ, содержащие соответствующие антибиотики, и выращивали при 20°С в течение 3-4 дней. Одну колонию собирали и высевали штрихом в планшеты YEP, содержащие такие же антибиотики, и инкубировали при 28°С в течение 1-2 дней.
Культура Agrobacterium и совместное культивирование: В день эксперимента колонии Agrobacterium собирали с планшетов YEP, суспендировали в 10 мл среды для заражения в одноразовую пробирку объемом 50 мл и доводили плотность клеток до значения OD600=0,2-0,4 нм, используя спектрофотометр. Культуры Agrobacterium помещали в ротационный шейкер при 100 об/мин, при комнатной температуре, пока выполнялась диссекция зародышей. Незрелые зиготные зародыши размером от 1,5 до 2,2 мм выделяли из стерилизованных зерен кукурузы и помещали в 1 мл среды для заражения и однократно промывали в том же буфере. Суспензию Agrobacterium (2 мл) добавляли в каждую пробирку, и пробирки переворачивали приблизительно 20 раз, затем встряхивали в течение 10-15 минут. Зародыши переносили в среду для совместного культивирования (соли MS 4,33 г/л; L-пролин 700,0 мг/л; мио-инозитол 100 мг/л; ферментативный гидролизат казеина 100 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 3,00 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; ацетосирингон 100 мкМ), ориентированных щитком вверх, и инкубировали в течение 3-4 дней при свете при 25°С.
Транзиторная экспрессия GUS и YFP/PhiYFP: транзиторную экспрессию YFP/PhiYFP и GUS можно наблюдать в трансформированных зародышах и после 3 дней совместного культивирования с Agrobacterium. Зародыши обследовали под стереомикроскопом (Leica Microsystems, Buffalo Grove, IL), используя фильтр YFP и 500 нм источник света. Зародыши, демонстрирующие экспрессию YFP/PhiYFP, отбирали для гистохимического анализа GUS. Раствор для окрашивания GUS готовили, как описано Maniatis et al. (1989), и зародыши инкубировали в 1 мл раствора в течение 24 часов при 37°С. Наблюдения за транзиторной экспрессией GUS в зародышах проводили под микроскопом.
Выбор каллюса и регенерация предполагаемых трансформантов: после периода совместного культивирования зародыши переносили в среду покоя (соли MS 4,33 г/л; L-пролин 700,0 мг/л; мио-инозитол 100 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота)] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 2,30 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; карбенициллин 250,0 мг/л) без селективного агента и инкубировали при свете в течение 7 дней при 28°С. Зародыши переносили в среду Селекция 1 (соли MS 4,33 г/л; L-пролин 700,0 мг/л; мио-инозитол 100 мг/л; MES [(2-(н-морфолино)этансульфоновая кислот), свободная кислота)] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 2,30 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; карбенициллин 250,0 мг/л), содержащую 100 нМ галоксифоп, и инкубировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение 7 дней при 28°С.
Зародыши с пролиферирующими эмбриогенными каллюсами переносили в среду Селекция 2 (соли MS 4,33 г/л; мио-инозитол 100 мг/л; L-пролин 700,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота)] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 2,30 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; карбенициллин 250,0 мг/л), содержащую 500 нМ галоксифоп, и инкубировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение следующих 14 дней при 28°С. Эта стадия селекции позволяет трансгенному каллюсу в дальнейшем пролиферировать и дифференцироваться. Период селекции каллюса длится в течение трех недель. Пролиферируя эмбриогенные каллюсы переносили в среду Регенерация 1 (соли MS 4,33 г/л; мио-инозитол 100,0 мг/л; L-пролин 350,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота)] 250,0 мг/л; ферментативный гидролизат казеина 50,0 мг/л; NAA 0,500 мг/л; ABA 2,50 мг/л; BA 1,00 мг/л; сахароза 45,0 г/л; GelzanTM 2,50 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 1,00 мг/л; карбенициллин 250,0 мг/л), содержащую 500 нМ галоксифоп, и культивировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение 7 дней при 28°С. Эмбриогенные каллюсы с побегоподобными почками переносили в среду Регенерация 2 (соли MS 4,33 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; мио-инозитол 100,0 мг/л; сахароза 60,0 г/л; Gellan Gum G434™ 3,00 г/л; карбенициллин 250,0 мг/л), содержащую 500 нМ галоксифоп. Культуры инкубировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение 7-10 дней при 28°С. Небольшие побеги с первичными почками переносили в среду для удлинения побегов и образования корня (соли MS 4,33 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; мио-инозитол 100,0 мг/л; сахароза 60,0 г/л; Gellan Gum G434™ 3,00 г/л; карбенициллин 250,0 мг/л) в коробках MAGENTA™ (Sigma-Aldrich, St. Louis, MO) и инкубировали в режиме свет/темнота в течение 16/8 часов 7 дней при 28°С. Предполагаемые трансгенные саженцы анализировали на предмет числа копий трансгена и переносили в теплицу.
ПРИМЕР 2: Конструирование синтетического двунаправленного промотора SCBV и вектора pDAB108708.
Иллюстративное схематическое изображение промотора убиквитина-1 кукурузы (Ubi1) показано на фиг. 1. Промотор Ubi1 клонировали из кукурузы. Плазмиду, содержащую промотор, амплифицировали в ходе ПЦР, используя ПЦР-амплификационную систему высокой точности. Область длиной приблизительно 200 нуклеотидов промотора кукурузы Ubi1 идентифицировали как элемент минимального корового промотора Ubi1 Zea mays (minUbi1P) (SEQ ID NO:1). MinUbi1P последовательности SEQ ID NO:1 затем добавляли к полинуклеотиду, содержащему экзон убиквитина-1 Zea mays (экзон ZmUbi1) и интрон убиквитина-1 Zea mays (интрон ZmUbi1), используя способы клонирования, общеизвестные в данной области техники, с получением полинуклеотида с последовательностью SEQ ID NO:2. Образующийся полинуклеотид затем клонировали в обратной ориентации в области, расположенной выше нуклеиновой кислоты, содержащей промотор Ubi1 кукурузы (включая URS Ubi1), с получением синтетического двунаправленного промотора Ubi1 с последовательностью SEQ ID NO:3.
Кодирующие последовательности репортерного гена клонировали в область, расположенную ниже каждого конца синтетического двунаправленного промотора Ubi1. Кодирующую последовательность желтого флуоресцентного белка (YFP) вставляли в область, расположенную ниже полинуклеотидного фрагмента, содержащего minUbi1P, экзон ZmUbi1 и элементы промотора интрона ZmUbi1. Кроме того, расположенную по ходу транскрипции лидерную последовательность, содержащую 3-рамочную концевую полинуклеотидную последовательность и консенсусную полинуклеотидную последовательность кукурузы, добавляли к minUbi1P, ZmUbi1, экзону ZmUbi1 и фрагменту элементов промотора интрона ZmUbi1. Кодирующую последовательность uidA (GUS) также вставляли ниже синтетического двунаправленного промотора Ubi1 в обратной ориентации относительно последовательности YFP с получением нуклеиновой кислоты с последовательностью SEQ ID NO:4. Образующийся полинуклеотид, содержащий синтетический двунаправленный промотор Ubi1, функционально связанный с генами YFP и GUS, клонировали в плазмиду pDAB105801. На фиг. 4 показана ориентация экспрессионной кассеты YFP и GUS относительно синтетического двунаправленного промотора Ubi1 в плазмиде pDAB105801.
Последовательность нативного промотора Ubi1 удаляли из двунаправленного промотора Ubi1 плазмиды pDAB105801 и заменяли на ПЦР-амплифицированный фрагмент, содержащий промотор SCBV и интрон ADH (SEQ ID NO:6). Полученный в итоге иллюстративный синтетический двунаправленный промотор SCBV называемый как SEQ ID NO:5 (также см. фиг. 5). Результатом добавления этого промотора SCBV является завершение конструирования вектора pDAB105806 (фиг. 6). Этот вектор содержал кассеты экспрессии генов YFP и GUS, управляемых двунаправленным промотором SCBV (SEQ ID NO:7; см. также фиг. 7).
Завершение конструирования бинарного вектора, содержащего кассеты экспрессии генов GUS и YFP из плазмиды pDAB105806, осуществляли посредством реакции GATEWAY L-R CLONASE (Invitrogen, Carlsbad, CA). Образующийся вектор, pDAB108708, содержал кассеты экспрессии генов GUS, YFP и AAD-1 внутри области Т-цепи (см. фиг. 9).
ПРИМЕР 3: Экспрессия генов, функционально связанных с синтетическим двунаправленным промотором SCBV
Репрезентативные примеры транзиторной экспрессии YFP и GUS в зародышах Zea mays, трансформированных pDAB108708, могут быть визуализированы. Обе стороны двунаправленного промотора SCBV могут управлять стойкой экспрессией функционально связанных кодирующих последовательностей YFP и GUS. Уровни экспрессии YFP сопоставимы с уровнями экспрессии GUS. Эти наблюдения подтверждают, что оба конца двунаправленного промотора SCBV биологически функциональны. Более того, элемент minUbi1P синтетического двунаправленного промотора SCBV может обеспечивать экспрессию YFP на уровне, подобном уровню при сравнении с каллюсом Zea mays, трансформированным бинарной плазмидой (pDAB101556), содержащей только однонаправленный промотор ZmUbi1, управляющий кодирующей последовательностью YFP. Экспрессия YFP или GUS не детектируется в незрелых зародышах отрицательного контроля, которые не трансформированы бинарным конструктом, и не содержали кодирующих последовательностей YFP или GUS.
ПРИМЕР 4: Стабильная экспрессия генов, функционально связанных с синтетическим двунаправленным промотором SCBV
Можно наблюдать изображения клеток каллюса Zea mays, которые стабильно трансформированы бинарным вектором pDAB108708, содержащим кодирующую последовательность YFP. Эти клетки получали из зародышей Z. mays, которые выращивали на среде Селекция 2. Условия микроскопии и протокол являлись такими, которые использовали для получения изображений зародышей. Двунаправленный промотор SCBV может управлять стойкой экспрессией кодирующих последовательностей YFP. Эти результаты подтверждают, что элемент минимального промотора Min-Ubi1P двунаправленного промотора SCBV способен обеспечивать экспрессию репортерного гена в стабильно трансформированных клетках каллюса Z. Mays. Уровни экспрессии белка YFP подобны, если сравнивать с экспрессией YFP в каллюсе Z. Mays, трансформированным контрольным бинарным вектором, содержащим однонаправленный промотор ZmUbi1, управляющий кодирующей последовательностью YFP (pDAB101556). Экспрессия YFP не детектируется в каллюсе отрицательного контроля, который не был трансформирован бинарным конструктом и не содержал кодирующую последовательность YFP или GUS.
ПРИМЕР 5: Определение числа копий трансгена путем использования TaqManTM ПЦР в реальном времени
Зародыши Zea mays трансформировали бинарным вектором, содержащим двунаправленный промотор SCBV, pDAB108708, и другие растения трансформировали контрольным бинарным вектором, pDAB101556. Наличие трансгенов YFP внутри геномов обоих наборов растений Z. Mays подтверждали посредством анализа гидролиза зонда. Стабильно трансформированные трансгенные саженцы Z. Mays, образующиеся из каллюса, получали и анализировали с целью идентификации трансформантов, содержащих низкое число копий (1-2 копии) полноразмерных вставок Т-цепей из бинарного вектора pDAB108708 и контрольного бинарного вектора pDAB101556. Идентифицированные саженцы перемещали в теплицу и выращивали.
Система Roche Light Cycler480™ использовали для определения числа копий трансгенов для трансформантов, трансформированных бинарным вектором pDAB108708. Способ использует биплексную реакцию TAQMAN®, в которой применяются олигонуклеотиды, специфичные для гена YFP и для эндогенного референсного гена Z. Mays, инвертазы (идентификационный номер в GenBank: U16123.1), в одном анализе. Число копий и зиготность определяли измерением интенсивности YFP-специфичной флуоресценции относительно инвертаза-специфичной флуоресценции, сравнивая с известным числом копий стандартов.
В Z. Mays, трансформированных бинарным вектором pDAB108708, специфичный для гена YFP фрагмент ДНК амплифицировали с одним набором TAQMAN® праймер/зонд, содержащим зонд, меченый флуоресцентным красителем FAM, и инвертазу амплифицировали вторым набором TAQMAN® праймер/зонд, содержащим зонд, меченый флуоресцентным красителем HEX (таблица 2). Реакционную смесь ПЦР готовили, как предложено в таблице 3, и ген-специфичные фрагменты ДНК амплифицировали согласно условиям, указанным в таблице 4. Число копий и зиготность образцов определяли измерением интенсивности флуоресценции, специфичной для репортерного гена, YFP, относительно флуоресценции, специфичной для референсного гена, инвертазы, сравнивая с известным числом копий стандартов.
|
Стандарты создавали путем разбавления вектора, pDAB108708, в геномной ДНК (гДНК) В104 Z. Mays с получением стандартов с известным соотношением pDAB108706:гДНК. Например, готовили образцы, имеющие одну; две или четыре копии ДНК вектора на одну копию гДНК В104 Z. Mays. Одно и два разведения копий pDAB108706, смешанные со стандартной гДНК В104 Z. Mays, оценивали в сопоставлении с контрольным трансформантом Z. Mays, известным как гемизиготный, и контрольным трансформантом Z. Mays, известным как гомозиготный (трансформант 278 Z. Mays; см. международную патентную публикацию РСТ № WO 2011/022469 А2). Биплексный способ TAQMAN®, который использует олигонуклеотиды, специфичные для гена AAD1, и олигонуклеотиды, специфичные для эндогенного референсного гена Z. Mays, инвертазы, выполняли путем амплификации и детекции ген-специфичного фрагмента ДНК для AAD1 c одним набором TAQMAN® праймер/зонд, содержащим зонд, меченый флуоресцентным красителем FAM, и путем амплификации и детекции ген-специфичного фрагмента ДНК для инвертазы вторым набором TAQMAN® праймер/зонд, содержащим зонд, меченый HEX флуоресценцией (таблица 2). Реакционную смесь TAQMAN® AAD1 готовили, как указано в таблице 3, и специфические фрагменты амплифицировали согласно условиям, указанным в таблице 4.
|
Уровень флуоресценции, образуемый для каждой реакции, анализировали с помощью Roche LightCycler 480™ Thermocycler в соответствии с инструкциями фирмы-производителя. Флуоресцентная молекула FAM возбуждалась при оптической плотности 465/510 нм, и флуоресцентная молекула HEX возбуждалась при оптической плотности 533/580 нм. Число копий определяли сопоставлением значений мишень/референс для неизвестных образцов (выходные данные LightCycler 480™) к значениям мишень/референс четырех стандартов известного числа копий (ноль, 1-копийный (геми), 2-копийный (гомо) и 4-копийный).
|
Результаты анализа числа копий трансгена в трансгенных растениях, полученных путем трансформации конструктом, содержащим двунаправленный промотор ZmUbi1 (pDAB108706), и в трангенных растениях, полученных путем трансформации контрольным YFP-конструктом с однонаправленным промотором ZmUbi1 (pDAB101556), представлены в таблице 5. Только растения с 1-2 копиями трансгена yfp перемещали в теплицу для дальнейшего анализа экспрессии.
|
ПРИМЕР 6. Стабильная экспрессия генов, функционально связанных с синтетическим двунаправленным промотором SCBV в целом растении.
Целые растения, которые содержали низкое число копий бинарной плазмиды pDAB108708, и растения, которые содержали низкое число копий контрольной бинарной плазмиды pDAB101566, выращивали в теплице. Эти растения анализировали, используя микроскопию, где можно получать изображения, демонстрирующие экспрессию YFP в растениях Т0 Z. mays, которые стабильно трансформируются конкретным конструктом нуклеиновой кислоты, содержащим экспрессионную кассету YTP, функционально связанного с синтетическим двунаправленным промотором SCBV (pDAB108708). Репрезентативные примеры стабильной экспрессии YFP в листьях и корневой ткани трансгенных Т0 растений кукурузы, полученных из зародышей Z. mays, трансформированных pDAB108708, демонстрировали надлежащую экспрессию YFP. Двунаправленный промотор SCBV может управлять стабильной экспрессией кодирующих последовательностей YFP как в тканях листьев, так и тканях корней. Микроскопический анализ также подтвердил, что элемент минимального промотора Min-UbiP1 в двунаправленном промоторе SCBV может управлять экспрессией YFP на том же уровне при сравнении с растениями Z. Mays, трансформированными контрольной бинарной плазмидой (pDAB101566), которая содержит однонаправленный промотор ZmUbi1, управляющий экспрессией кодирующей последовательности YFP. Контрольные растения демонстрировали стабильную экспрессию YFP в тканях листьев и тканях корней.
ПРИМЕР 7: Вестерн-блот-анализ генов, функционально связанных с синтетическим двунаправленным промотором SCBV
Общий растворимый белок: Получали образец трансформированных растений кукурузы Т0 на стадии развития V6. Все четыре вырубки из самых молодых развернутых листьев помещали в пробирку для взятия образцов (matrix tubes) и располагали в коробке для хранения образцов (matrix box). В качестве отрицательного контроля четырехлистные вырубки двух нетрансформированных растений кукурузы В104 на стадии развития V6 помещали в пробирку для взятия образцов (matrix tubes). Стальной шарик помещали в пробирку с образцами, и затем 400 мкл PBST добавляли в каждую пробирку. Пробирки закрывали крышками, и белок выделяли посредством ударов шарика при 1500 об/мин в течение 5 минут в гомогенизаторе ткани Kleco™. Клеточный дебрис осаждали путем центрифугирования.
5 мкл образца из каждой пробирки разводили до 25 мкл с помощью PBST в 96-луночном планшете для микротитрования. Эти образцы анализировали на предмет общего растворимого белка, используя набор BCA для анализа белка (Thermo Scientific Pierce, Rockford, IL) в соответствии с рекомендациями фирмы. Стандарты бычьего сывороточного альбумина (BSA), предоставляемые в наборе, анализировали дважды, и среднее значение использовали для построения стандартной кривой, которая впоследствии была использована для вычисления общего растворимого белка для каждого образца. Общий растворимый белок для каждого образца затем нормировали к виду мг/мкл.
|
Вестерн-блот-анализ YFP/PhiYFP: В 96-луночном планшете для микротитрования каждые 5 мкл образца экстрагированного белка разводили 5 мкл 2× буфера Лэммли с 2-β-меркаптоэтанолом. Контрольные образцы очищенного YFP/PhiYFP в буфере HEPES (50 мМ HEPES, 200 мМ KCl, 10% глицерин) были приобретены в Axxora (San Diego, CA). Образцы приготавливали в том же планшете путем разведения 1:1 буфером Лэммли для построения стандартной кривой следующих концентраций: 0,5 нг/мкл, 0,25 нг/мкл и 0,125 нг/мкл. Образцы нагревали в термоциклере при 95°C в течение 30 минут и затем охлаждали до 4°С. Затем собирали Bio-Rad Criterion gel™, используя буфер MES/SDS. Образцы нагревали до комнатной температуры, и 10 мкл образца вносили в каждую лунку двух гелей. Кроме того, образцы очищенного YFP/PhiYFP, используемые для стандартной кривой, и маркер размера белка вносили в лунки геля. Электрофорез в гелях проводили при 150 В и 150 мА в течение 90 мин. По окончании фореза гелевую установку открывали и белки переносили на нитроцеллюлозную мембрану, используя iBlot System™ (Invitrogen). Белок переносили из геля на мембрану, запуская ток при 20 В в течение 10 минут. Нитроцеллюлозную мембрану извлекали и помещали в блокирующий буфер StartingBlock T20™ blocking buffer на ночь при 4°С. Блокирующий буфер затем удаляли, и мембрану обрабатывали, используя протокол, представленный в таблице 6.
Связывание антитела оценивали системой детекции хемилюминесценции Amersham ECL™ plus, следуя инструкциям фирмы-производителя. Пленку экспонировали в течение 10 минут и 30 минут. Экспонированную 10 минут пленку использовали для количественной оценки белка, и сверхэкспонированную 30 минут пленка использовали для подтверждения отсутствия белка в В104 и других контрольных образцах. Мембрану приклеивали к задней части экспонированной пленки, и белок количественно измеряли путем анализа плотности пикселей. Плотность пикселей очищенных белковых стандартов сначала использовали для получения стандартной кривой, которую использовали для количественной оценки белка в образцах. Хотя на мембране были выявлены полосы, соответствующие мономеру и димеру PhiYFP даже в очищенном стандарте, для количественной оценки экспрессии белка использовали только мономер PhiYFP. Значения для белка затем нормировали к виду нг/мкл. Отношение нормированного общего растворимого белка (TSP) к PhiYFP вычисляли в единицах нг YFP/мг TSP, или альтернативно, частях на миллион (ч./млн.).
Вестерн-блот-анализ GUS: Экспрессию белка GUS количественно оценивали таким же способом, как и PhiYFP, за исключением следующего: 10 мкл образца экстракта разводили 1:1, используя 2× Лэммли с 2-β-меркаптоэтанолом, денатурировали при 95°С в течение 30 минут, и затем 15 мкл наносили на гель. Обработанные пленкой мембраны (1 минута экспозиции) совмещали с мембраной для анализа плотности пикселей.
Результаты Вестерн-блот-анализа 12 трансгенных растений кукурузы Т0, полученных из зародышей Z. mays, трансформированных бинарным вектором, pDAB108708, представлены на фиг.16. Двунаправленный промотор SCBV демонстрировал стабильную экспрессию кодирующих последовательностей YFP и GUS из ткани листьев. Эти наблюдения подтверждают, что элемент минимального промотора Min-UbiP1, выделенный из промотора убиквитина 1 Zea mays и соединенного с промотором SCBV, может обеспечивать экспрессию YFP на том же уровне в сравнении с каллюсом Z. mays, трансформированного бинарной плазмидой, содержащей однонаправленный промотор ZmUbi1, управляющий кодирующей последовательностью YFP (pDAB101556; см. фиг. 17).
ПРИМЕР 8: Конструкт четырехгенного кассетного блока
Конструкт плазмиды pDAB105806 использовали в качестве исходной плазмиды для получения четырехгенного кассетного блока (AAD1-2A-PhiYFP и Cry34(8V6)-2A-Cry35), управляемого одним двунаправленным промотором SCBV. Репрезентативная карта плазмиды pDAB105806 показана на фиг. 6, которая содержит двунаправленный промотор SCBV.
Фрагмент AAD1-2A-PhiYFP, полученный из плазмиды pDAB105841 (фиг. 22), клонировали по сайтам действия рестриктаз PstI и SacI основного вектора плазмиды pDAB105806, используя способы клонирования, общеизвестные в данной области техники. В результате получали промежуточный вариант плазмиды pDAB105847 (фиг. 22). Рестрицированный NotI/XbaI фрагмент Cry34(8V6)-2A-Cry35, полученный из плазмиды pDAB105840, клонировали между сайтами действия рестриктаз NotI/SpeI в плазмиде pDAB105847 с получением плазмиды pDAB105849 (фиг. 23). Плазмида pDAB105849 содержала генные кассеты Cry34(8V6)-2A-Cry35 и AAD1-2A-PhiYFP на каждой стороне двунаправленного промотора SCBV.
Бинарный вектор, содержащий двунаправленный промотор SCBV, и генные экспрессионные кассеты Cry34(8V6)-2A-Cry35 и AAD1-2A-PhiYFP из плазмиды pDAB105849 преобразовывали посредством реакции GATEWAY L-R CLONASE (Invitrogen, Carlsbad, CA) в представляющую интерес плазмиду pDAB101917 (фиг. 24). Конечный вариант вектора, pDAB108719, содержал Cry34(8V6)-2A-Cry35, AAD1-2A-PhiYFP и PAT генные экспрессионные кассеты в пределах границ Т-ДНК (фиг. 24).
ПРИМЕР 9: Конструкт второго четырехгенного кассетного блока
Фрагмент PhiYFP-2A-AAD1, полученный из плазмиды pDAB105844 (фиг. 25), клонировали по сайтам действия рестриктаз PstI и SacI основного вектора плазмиды pDAB105806, используя способы клонирования, общеизвестные в данной области техники. В результате получали промежуточный вариант плазмиды pDAB105848 (фиг. 25). Рестрицированный NotI/XbaI фрагмент Cry34(8V6)-2A-Cry35, полученный из плазмиды pDAB105840, клонировали между сайтами действия рестриктаз NotI/SpeI в плазмиде pDAB105848 с получением плазмиды pDAB105865 (фиг.26). Плазмида pDAB105865 содержала генные кассеты Cry34(8V6)-2A-Cry35 и PhiYFP-2A-AAD1 на каждой стороне двунаправленного промотора SCBV.
Бинарный вектор, содержащий двунаправленный промотор SCBV, и генные кассеты Cry34(8V6)-2A-Cry35 и PhiYFP-2A-AAD1 из плазмиды pDAB105865 преобразовывали посредством реакции GATEWAY L-R CLONASE (Invitrogen, Carlsbad, CA) в представляющую интерес плазмиду pDAB101917 (фиг. 24). Конечный вариант вектора, pDAB108720, содержал Cry34(8V6)-2A-Cry35, PhiYFP-2A-AAD1 и PAT генные экспрессионные кассеты в пределах границ Т-ДНК (фиг. 26).
ПРИМЕР 10: Трансформация штамма DAt13192 Agrobacterium tumefaciens
Бинарными векторами pDAB108719 и pDAB108720Т трансформировали трехкомпонентный штамм DAt13192 Agrobacterium tumefaciens (см. предварительная патентная заявка США №61/368965). Бактериальные колонии выделяли, и бинарную плазмидную ДНК экстрагировали и подтверждали рестрикционным анализом.
ПРИМЕР 11: Трансформация кукурузы
Стерилизация початка и выделение зародышей: Для получения незрелых зародышей кукурузы, растения Zea mays (c.v. B104) выращивали в теплице и использовали процесс самоопыления или близкородственного скрещивания для образования початка. Початки собирали приблизительно на 9-12 день после опыления. В день эксперимента поверхность початков стерилизовали погружением в 20% раствор хозяйственного отбеливателя, который содержал 5% гипохлорит натрия, и встряхивали в течение 20-30 минут с последующим трехкратным ополаскиванием в стерильной воде. После стерилизации, незрелые зиготные зародыши (1,5-2,2 мм) в асептических условиях вырезали из каждого початка и случайным образом распределяли в микроцентрифужных пробирках, содержащих жидкую среду для заражения (базальную среду LS, 4,43 г/л; раствор витаминов N6 [1000X], 1,00 мл/л; L-пролин, 700,0 мг/л; сахарозу 68,5 г/л; глюкозу 36 г/л; 2,4-D 1,5 мг/л). Для данного набора экспериментов объединенные в пулы зародыши от 2-3 початков использовали для каждой обработки.
Закладка культуры Agrobacterium: глицериновые стоковые образцы штаммов Agrobacterium, содержащих бинарные векторы, описанные выше, высевали штрихом в планшеты с минимальной средой АВ, содержащие соответствующие антибиотики, и выращивали при 20°С в течение 3-4 дней. Одну колонию собирали и высевали штрихом в планшеты YEP, содержащие те же антибиотики, и инкубировали при 28°С в течение 1-2 дней.
Культура Agrobacterium и совместная культивирование: В день эксперимента колонии Agrobacterium собирали с планшеты YEP, суспендировали в 10 мл среды для заражения в одноразовой пробирке объемом 50 мл и доводили плотность клеток до значения OD600=0,2-0,4 нм, используя спектрофотометр. Культуры Agrobacterium помещали в ротационный шейкер при 115 об/мин, при комнатной температуре, пока выполнялась диссекция зародышей. Незрелые зиготные зародыши размером от 1,5 до 2,2 мм выделяли из стерилизованных зерен кукурузы и помещали в 1 мл среды для заражения и однократно промывали в той же среде. Суспензию Agrobacterium (2 мл) добавляли в каждую пробирку, и пробирки переворачивали приблизительно 20 раз, затем встряхивали в течение 10-15 минут. Зародыши переносили в среду для совместного культивирования (соли MS 4,33 г/л; L-пролин 700,0 мг/л; мио-инозитол 100 мг/л; ферментативный гидролизат казеина 100 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 3,00 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; ацетосирингон 100 мкМ), ориентированных щитком вверх, и инкубировали в течение 3-4 дней при свете при 25°С.
Транзиторная экспрессия YFP/PhiYFP: транзиторную экспрессию YFP/PhiYFP можно было наблюдать в трансформированных зародышах после 3 дней совместного культивирования с Agrobacterium. Зародыши обследовали под стереомикроскопом (Leica Microsystems, Buffalo Grove, IL), используя фильтр YFP и источник света 500 нм.
Выбор каллюса и регенерация очищенных трансформантов: после периода совместного культивирования зародыши переносили в среду покоя (соли MS 4,33 г/л; L-пролин 700,0 мг/л; мио-инозитол 100 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота)] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 2,30 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; карбенициллин 250,0 мг/л) без селективного агента и инкубировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение 7 дней при 28°С. Зародыши переносили в среду Селекция 1 (соли MS 4,33 г/л; L-пролин 700,0 мг/л; мио-инозитол 100 мг/л; MES [(2-(н-морфолино)этансульфоновая кислот), свободная кислота)] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 2,30 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; карбенициллин 250,0 мг/л), содержащую 3 мг/л биалафоса, и инкубировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение 7 дней при 28°С.
Зародыши с пролиферирующими эмбриогенными каллюсами переносили в среду Селекция 2 (соли MS 4,33 г/л; мио-инозитол 100 мг/л; L-пролин 700,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота)] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба 3,30 мг/л; сахароза 30,0 г/л; Gelzan™ 2,30 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 15,0 мг/л; карбенициллин 250,0 мг/л), содержащую 5 мг/л биалафоса, и инкубировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение следующих 14 дней при 28°С. Эта стадия селекции позволяет трансгенному каллюсу в дальнейшем пролиферировать и дифференцироваться. Период селекции каллюса может длиться три недели. Пролиферируя, эмбриогенные каллюсы переносили в среду Регенерация 1 (соли MS 4,33 г/л; мио-инозитол 100,0 мг/л; L-пролин 350,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота)] 250,0 мг/л; ферментативный гидролизат казеина 50,0 мг/л; NAA 0,500 мг/л; ABA 2,50 мг/л; BA 1,00 мг/л; сахароза 45,0 г/л; Gelzan™ 2,50 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; AgNO3 1,00 мг/л; карбенициллин 250,0 мг/л), содержащую 3 мг/л биалафоса, и культивировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение 7 дней при 28°С.
Эмбриогенные каллюсы с побегами/почками переносили в среду Регенерация 2 (соли MS 4,33 г/л; модифицированный MS-Vitamin [1000X] 1,00 мл/л; мио-инозитол 100,0 мг/л; сахароза 60,0 г/л; Gellan Gum G434™ 3,00 г/л; карбенициллин 250,0 мг/л), содержащую 3 мг/л биалафоса. Культуры инкубировали в течение 24 часов при свете с интенсивностью 50 мкмоль м-2с-1 в течение 7-10 дней при 28°С. Небольшие побеги с первичными корешками переносили в среду для удлинения побегов и образования корня (соли MS 4,33 г/л; раствор N6 Vitamin Solution [1000X] 1,00 мл/л; мио-инозитол 100,0 мг/л; сахароза 30,0 г/л; агар 5,50 г/л) в емкости phytatray и инкубировали в режиме свет/темнота в течение 16/8 часов при 90 мкмоль м-2с-1 в течение 7 дней при 28°С. Проводили селекцию здоровых предполагаемых трансгенных саженцев, затем инкубировали в режиме свет/темнота в течение 16/8 часов при 200 мкмоль м-2с-1 в течение следующих 2-5 дней при 25°С и анализировали на предмет числа копий трансгена, и переносили в теплицу.
ПРИМЕР 12: Транзиторная экспрессия PhiYFP
Выполняли транзиторную экспрессию PhiYFP из зародышей Zea mays, трансформированных pDAB108719. Двунаправленный промотор SCBV мог экспрессировать PhiYFP с генной экспрессионной кассеты AAD1-2A-PhiYFP, и нетрансформированный зародыш не демонстрировал какой-либо флуоресценции PhiYFP. Подобный уровень экспрессии PhiYFP можно было наблюдать в зародышах Zea mays, трансформированных бинарной плазмидой pDAB105748 (фиг.20), содержащей однонаправленных промотор Ubi1 Zea mays (Zm), управляющий одной кодирующей последовательностью PhiYFP, которая демонстрировала ожидаемый уровень экспрессии YFP/PhiYFP. Транзиторную экспрессию PhiYFP можно было наблюдать в зародышах Zea mays, трансформированных pDAB108720, где двунаправленный промотор Zm Ubi1 мог обеспечивать экспрессию PhiYFP с генной экспрессионной кассеты PhiYFP-2A-AAD1.
ПРИМЕР 13: Экспрессия PhiYFP в стабильно трансформированной кукурузе
Экспрессия PhiYFP в стабильно трансформированном каллюсе Zea mays, управляемая двунаправленным промотором Zm Ubi1: зародыши Zea mays, трансформированные бинарным вектором pDAB108719, содержащим генную экспрессионную кассету AAD1-2A-PhiYFP, демонстрировали хорошую экспрессию PhiYFP. Двунаправленный промотор SCBV мог управлять стабильной экспрессией PhiYFP. Эти результаты подтверждают то, что элемент минимального промотора Min-UbiP1 двунаправленного промотора SCBV, способен обеспечить экспрессию репортерного гена, например, PhiYFP или YFP. Уровни экспрессии белка PhiYFP подобны уровням при сравнении с каллюсом Zea mays, трансформированным контрольным бинарным вектором, который содержал однонаправленный промотор Zm Ubi1, управляющий кодирующей последовательностью PhiYFP (pDAB105748). Экспрессия PhiYFP не детектируется в каллюсе отрицательного контроля, который не был трансформирован бинарным конструктом и не содержал кодирующие последовательности PhiYFP.
Зародыши Zea mays, трансформированные бинарным вектором pDAB108720, который содержал генную экспрессионную кассету PhiYFP-2A-AAD1, демонстрировали хорошую экспрессию PhiYFP. Двунаправленный промотор SCBV мог управлять стабильной экспрессией PhiYFP. Эти результаты подтверждают, что элемент минимального промотора Min-UbiP1 двунаправленного промотора SCBV, способен обеспечить экспрессию репортерного гена, например, PhiYFP или YFP.
ПРИМЕР 14: Определение числа копий трансгена
Определение числа копий трансгена, используя TaqMan™ ПЦР в реальном времени: растения Zea mays трансформировали бинарными векторами, содержащими двунаправленный промотор SCBV, pDAB108719 и pDAB108720, и другие растения трансформировали контрольным бинарным вектором, pDAB105748. Наличие кодирующей последовательности (PhiYFP, AAD1, Cry34, Cry35, Pat) в пределах генома растений Z. mays, трансгенных по pDAB108719 и pDAB108720, подтверждали посредством TaqMan-анализа гидролиза зонда. Растения, трансгенные по контрольному вектору pDAB105748, анализировали на присутствие последовательности PhiYFP. Стабильно трансформированные трансгенные саженцы Z. Mays, образованные из каллюса, получали и анализировали для идентификации трансформантов, содержащих низкое число копий (1-2 копии) полноразмерных вставок Т-цепей из бинарных векторов pDAB108719 и pDAB108720, и контрольного бинарного вектора pDAB105748. Верифицированные саженцы перемещали в теплицу и выращивали.
Систему Roche Light Cycler480™ использовали для определения числа копий трансгенов для трансформантов, трансформированных бинарными векторами pDAB108719 и pDAB108720. Данный способ использует биплексную реакцию TAQMAN®, в которой применяются олигонуклеотиды, специфичные к кодирующей последовательности и эндогенному референсному гену Z. Mays, инвертазе (код доступа в GenBank: U16123.1), в одном анализе. Число копий и зиготность определяли измерением интенсивности специфичной для кодирующей последовательности флуоресценции относительно инвертаза-специфичной флуоресценции, сравнивая с известным числом копий стандартов.
[M! Вставить последовательности праймеров со стр.63 оригинала, таблицы 7, второй столбец. Заменить во втором столбце "ROCHE UPL Probe #125 CTTGGAGC (SEQ ID NO:41) Cat #04693604001 (Roche, Indianapolis, IN) " на "Зонд #125 ROCHE UPL CTTGGAGC (SEQ ID NO:41) каталожный номер 04693604001 (Roche, Indianapolis, IN) "]
|
Для образцов Z. Mays, трансформированных бинарными векторами pDAB108719 и pDAB108720, специфичный для кодирующей последовательности фрагмент ДНК амплифицировали с одним набором TAQMAN® праймер/зонд, содержащим зонд, меченый флуоресцентным красителем FAM, и инвертазу амплифицировали вторым набором TAQMAN® праймер/зонд, содержащим зонд, меченый флуоресцентным красителем HEX (таблица 7). Реакционную смесь ПЦР готовили, как указано в таблице 8, и ген-специфичные фрагменты ДНК амплифицировали согласно условиям, указанными в таблице 9. Число копий и зиготность образцов определяли измерением интенсивности флуоресценции, специфичной для кодирующей последовательности относительно флуоресценции, специфичной для референсного гена, инвертазы, сравнивая с известным числом копий стандартов.
Стандарты создавали путем разбавления вектора (pDAB108719 или pDAB108720) в геномной ДНК (гДНК) В104 Z. Mays с получением стандартов с известным взаимоотношением вектор:гДНК. Например, готовили образцы, имеющие одну, две или четыре копии ДНК вектора на одну копию гДНК В104 Z. Mays. Одно и два разведения копий вектора, смешанные со стандартной гДНК В104 Z. Mays, оценивали в сопоставлении с контрольным трансформантом Z. Mays, известным как гемизиготный, и контрольным трансформантом Z. Mays, известным как гомозиготный (трансформант 278 Z. Mays; см. международную патентную публикацию РСТ № WO 2011/022469 А2). Биплексный способ TAQMAN®, который использует олигонуклеотиды, специфичные для кодирующей последовательности, и олигонуклеотиды, специфичные для эндогенного референсного гена Z. Mays, инвертазы, осуществляли путем амплификации и детекции ген-специфичного фрагмента ДНК для кодирующей последовательности c одним набором TAQMAN® праймер/зонд, содержащим зонд, меченый флуоресцентным красителем FAM, и путем амплификации и детекции ген-специфичного фрагмента ДНК для инвертазы вторым набором TAQMAN® праймер/зонд, содержащим зонд, меченый HEX флуоресценцией. Согласно таблице 7, реакционную смесь TAQMAN® для кодирующей последовательности готовили, как указано в таблице 8, и специфические фрагменты амплифицировали согласно условиям, указанным в таблице 9.
|
Уровень флуоресценции, образуемый для каждой реакции, анализировали с помощью Roche LightCycler 480™ Thermocycler в соответствии с инструкциями фирмы-производителя. Флуоресцентную молекулу FAM возбуждали при оптической плотности 465/510 нм, и флуоресцентную молекулу HEX возбуждали при оптической плотности 533/580 нм. Число копий может быть определено сопоставлением значений мишень/референс для неизвестных образцов (выходные данные LightCycler 480™) к значениям мишень/референс четырех стандартов известного числа копий (ноль, 1-копийный (геми), 2-копийный (гомо) и 4-копийный).
|
Результаты анализа числа копий трансгена в трансгенных растениях, полученных путем трансформации конструктами, содержащим двунаправленный промотор SCBV (pDAB108719 и pDAB108720), и в трангенных растениях, полученных путем трансформации контрольным PhiYFP-конструктом с однонаправленным промотором ZmUbi1 (pDAB105748), сведены в таблицу 10. Только растения с 1-2 копиями всех трансгенов перемещали в теплицу для дальнейшего анализа экспрессии.
|
ПРИМЕР 15. Стабильная экспрессия PhiYFP в растениях Т0 кукурузы
Стабильная экспрессия PhiYFP в растениях Т0 Zea mays, управляемая двунаправленным промотором SCBV: зародыши Zea mays, трансформированные бинарным вектором pDAB108719, содержащим генную экспрессионную кассету AAD1-2A-PhiYFP, были подвергнуты обследованию при наблюдении. Двунаправленный промотор SCBV мог управлять стабильной экспрессией PhiYFP как в тканях побегов, так и корней. Результаты подтверждают, что элемент минимального промотора Min-UbiP1 двунаправленного промотора SCBV способен обеспечивать экспрессию репортерного гена, например, PhiYFP или YFP, который связан в виде бистронных конструкций с aad1 с использованием последовательности 2А. Уровень экспрессии белка PhiYFP подобен уровню в зародышах Z. mays, трансформированных контрольным бинарным вектором, содержащим однонаправленный промотор ZmUbi1, управляющий кодирующей последовательностью PhiYFP (pDAB105748). Экспрессия PhiYFP не детектировалась в растениях отрицательного контроля, которые не были трансформированы бинарным конструктом и не содержали кодирующие последовательности PhiYFP.
Экспрессия PhiYFP в тканях листьев и корней растений Т0 Zea mays, трансгенных по бинарному вектору pDAB108720, который содержал генную экспрессионную кассету PhiYFP-2A-AAD1, была подвергнута обследованию. Двунаправленный промотор SCBV мог управлять стабильной экспрессией PhiYFP. Результаты подтверждают, что элемент минимального промотора Min-UbiP1 двунаправленного промотора SCBV способен обеспечивать экспрессию репортерного гена, например, PhiYFP или YFP, связанного с aad1 последовательностью 2А или 2А-подобной последовательностью.
ПРИМЕР 16: Белковый анализ Cry34, Cry35 и AAD1
Образцы растений отбирали в 1-10 колонок коробок для хранения образцов (matrix box) в конические пробирки на 1,5 мл, в которые добавляли 1 стальной шарик с последующим добавлением PBST+0,5% BSA (0,6 мл). Коробку затем встряхивали для измельчения образцов в приборе Geno Grinder в течение 5 минут при 1500 об/мин, затем центрифугировали при 3700 об/мин в течение 7 минут при 4°С.
Твердофазный иммуноферментный анализ (ELISA) Cry34/35: В отдельном 96-луночном планшете с глубокими лунками образец экстракта разводили 1:200 в PBST+1% блокирующий буфер. Два объема 25 мкл разведенного образца затем переносили в отдельные 96-луночные планшеты, которые были сгруппированы по анти-Cry34 и анти-Cry35 (Meso Scale Discovery). В 11 и 12 колонки каждого планшета добавляли (25 мкл) стандартные концентрации Cry34 и Cry35 в PBST+1% блокирующий буфер. Планшеты затем инкубировали при комнатной температуре в течение часа при встряхивании. Затем планшеты промывали PBST (3×300 мкл). Затем добавляли в каждую лунку 25 мкл раствора конъюгированных с SulfoTAG анти-Cry34 и анти-Cry35 и инкубировали при комнатной температуре в течение часа при встряхивании. Затем планшеты промывали PBST 300 мкл). Затем добавляли 150 мкл буфера для прочтения (3Read Buffer T, Meso Scale Discovery), и считывали сигнал на планшет-ридере SECTOR® 6000. Концентрации белков в образце могут быть вычислены, используя стандартную кривую для соответствующего белка, полученную на том же планшете.
Твердофазный иммуноферментный анализ (ELISA) AAD1: В отдельном 96-луночном планшете с глубокими лунками образец экстракта разводили 1:20 в PBST+0,5% BSA. Два объема 200 мкл разведенного образца затем переносили в отдельные 96-луночные планшеты, которые были покрыты анти-AAD1 (предоставляемые компанией Acadia Bioscience LLC). В 11 и 12 колонки каждого планшета добавляли (200 мкл) стандартные концентрации AAD1 в PBST+0,5% BSA. Затем в каждую лунку добавляли 50 мкл биотинилированных анти-AAD1, и планшеты инкубировали при комнатной температуре в течение часа при встряхивании. Затем планшеты промывали PBST (5×300 мкл). Затем добавляли в каждую лунку 100 мкл раствора конъюгата стрептавидин-щелочная фосфатаза и инкубировали при комнатной температуре в течение 30 минут при встряхивании. Затем планшеты промывали PBST (5×300 мкл). Затем добавляли 100 мкл субстрата (п-нитрофенилфосфат, PNPP) и инкубировали при комнатной температуре в течение 45 минут при встряхивании. Затем считывали сигнал при А405 на планшет-ридере SpectraMax M5 (Molecular Devices). Концентрации белков в образце могут быть вычислены, используя стандартную кривую, полученную на том же планшете.
ПРИМЕР 17: Анализ белков в растениях Т0 кукурузы
В данном примере проводили анализ белков в растениях Т0 кукурузы, регулируемых конструктом двунаправленного промотора SCBV Zea mays (pDAB108719).
|
Репрезентативный анализ ELISA 12 трансгенных растений кукурузы Т0, полученных из зародышей Zea mays, трансформированных pDAB108719, содержащей генную экспрессионную кассету Cry34-2A-Cry35, представлен в таблице 11. Двунаправленный промотор SCBV может управлять стабильной экспрессией кодирующих последовательностей Cry34 и Cry35 в листьях. Эти наблюдения продемонстрировали, что один двунаправленный промотор SCBV в конструкте pDAB108719 может обеспечивать экспрессию множества генов (например, Cry34, Cry35 и AAD1).
Анализ белков растений Т0 кукурузы, регулируемых двунаправленным конструктом промотора убиквитина 1 Zea mays (pDAB108720): Репрезентативный анализ ELISA 9 трансгенных растений кукурузы Т0, полученных из зародышей Zea mays, трансформированных pDAB108720, содержащей генную экспрессионную кассету Cry34-2A-Cry35, представлен в таблице 12. Двунаправленный промотор SCBV может управлять стабильной экспрессией кодирующих последовательностей Cry34 и Cry35 в листьях.
|
ПРИМЕР 18. Формирование трансгенной структуры: Синтетические двунаправленные промоторы (данные Т1)
Экспрессия генов растений Т1, управляемая конструктами двунаправленных промоторов: от десяти до двенадцати однокопийных объектов в одном конструкте отбирали для анализа за исключением того, что контрольный конструкт pDAB108716 имел только один объект. Тестировали пять растений/объектов для стадии V6 и три растения/объекта для стадий V10-12 и/R3. Анализ белков осуществляли с использованием ЖХМС или ELISA.
Конструкты, используемые в данном примере, представлены на фиг. 30. pDAB108708 (двунаправленный SCBV (-200)) и pDAB108709 (двунаправленный SCBV (-90)) представляли собой конструкты с репрезентативным двунаправленным промотором по настоящему изобретению в дополнение к конструктам с двунаправленным промотором Ubi1 кукурузы (pDAB108706 [двунаправленный ZMUbi (-200)) и pDAB108707 [двунаправленный ZMUbi (-90))]; pDAB101556 (контроль ZmUbi1-YFP), pDAB108715 (SCBV без минимального промотора) и pDAB108716 (ZMUbi1 без минимального промотора) служили в качестве контрольных конструктов с однонаправленными промоторами.
Иллюстративные результаты экспрессии (V6) из семи конструктов для белка YFP (ЖХМС) в нг/см2 представлены на фиг. 31А. Иллюстративные результаты по относительной экспрессии (V6) из семи конструктов для РНК YFP представлены на фиг. 31В.
Иллюстративные результаты экспрессии (V6) из семи конструктов для белка GUS (ЖХМС) в нг/см2 представлены на фиг. 32А. Иллюстративные результаты по относительной экспрессии (V6) из семи конструктов для РНК GUS представлены на фиг. 32В.
Иллюстративные результаты экспрессии (V6) из семи конструктов для белка AAD1 (ЖХМС) в нг/см2 представлены на фиг. 33А. Иллюстративные результаты по относительной экспрессии (V6) из семи конструктов для РНК AAD1 представлены на фиг. 33В.
Статистический анализ результатов экспрессии (V6) из семи конструктов для белка YFP (ЖХМС) в нг/см2 представлен на фиг. 34А. Статистический анализ результатов относительной экспрессии (V6) из семи конструктов для РНК YFP представлен на фиг. 34В. Средние значения и статистические результаты перечислены.
Статистический анализ результатов экспрессии (V6) из семи конструктов для белка GUS (ЖХМС) в нг/см2 представлен на фиг. 35А. Статистический анализ результатов относительной экспрессии (V6) из семи конструктов для РНК GUS представлен на фиг. 35В. Средние значения и статистические результаты перечислены.
Статистический анализ результатов экспрессии (V6) из семи конструктов для белка AAD1 (ЖХМС) в нг/см2 представлен на фиг. 36А. Статистический анализ результатов относительной экспрессии (V6) из семи конструктов для РНК AAD1 представлен на фиг. 36В. Средние значения и статистические результаты перечислены.
На фиг. 37А, 37В и 37С показаны иллюстративные результаты экспрессии (V10) из семи конструктов для белков YFP, AAD1 и GUS (ЖХМС) в нг/см2, соответственно.
На фиг. 38А, 38В и 38С показан статистический анализ результатов экспрессии (V10) из семи конструктов для белков YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Средние значения и статистические результаты перечислены.
На фиг. 39А, 39В и 39С показаны иллюстративные результаты экспрессии (R3) из семи конструктов для белков YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно.
На фиг. 40А, 40В и 40С показан статистический анализ результатов экспрессии (R3) из семи конструктов для белков YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Средние значения и статистические результаты перечислены.
Результаты демонстрируют, что оба двунаправленных промотора SCBV по настоящему изобретению и двунаправленные промоторы Ubi1 кукурузы могут управлять стабильной экспрессией GUS и YFP. Экспрессия YFP с двунаправленного промотора Ubi1 кукурузы подобна экспрессии YFP, управляемой однонаправленным Ubi1 кукурузы. Экспрессия YFP с двунаправленного промотора SCBV значительно выше экспрессии YFP, управляемой однонаправленным Ubi1 кукурузы или двунаправленным промотором Ubi1 кукурузы. Однако это отличие становится менее значимым на стадии V10. Результаты также предполагают, что двунаправленная транскрипция не оказывает значимого воздействия на экспрессию GUS (экспрессия GUS в сравнении с конструктами, у которых отсутствует минимальный промотор без экспрессии YFP). Двунаправленные промоторы SCBV также обеспечивают значительно более высокую экспрессию GUS по сравнению с двунаправленными промоторами Ubi1 кукурузы.
ПРИМЕР 19. Комбинирование двунаправленного промотора и бицистронной последовательности 2А для управления четырьмя трансгенами с одного единственного промотора (данные Т1)
Экспрессия генов растений Т1, управляемая конструктами двунаправленных промоторов: от десяти до двенадцати однокопийных объектов в одном конструкте отбирали для анализа, за исключением того, что контрольный конструкт имел четыре или пять объектов в конструкте. Тестировали пять растений/объектов для стадии V6 и три растения/объекта для стадий V10-12 и/R3. Анализ белков осуществляли с использованием ЖХМС или ELISA.
pDAB108719 и pDAB108720 показаны на фиг. 19. pDAB105748 и pDAB105818 показаны на фиг. 20. Дополнительные мультитрансгенные конструкты, использующие промотор Ubi1, включая pDAB108717 и pDAB108718, показаны на фиг. 41.
Иллюстративные результаты относительной экспрессии (V6) РНК Cry34 на шести конструктах pDAB105748 (ZMUbi1-YFP), pDAB105818 (ZMUbi1-Cry34/ZMUbi1-Cry35/ZMUbi1-AAD1), pDAB108717 (YFP/AAD-1-ZMUbi1 двунаправленный-Cry34-Cry35), pDAB108718 (AAD1/YFP-ZMUbi1 двунаправленный-Cry34-Cry35), pDAB108719 (YFP/AAD1-SCBV двунаправленный-Cry34-Cry35) и pDAB108720 (AAD1/YFP-SCBV двунаправленный-Cry34-Cry35) представлены на фиг.42А. Иллюстративные результаты относительной экспрессии (V6) белка Cry34 (ЖХМС) с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлены на фиг. 42В.
Иллюстративные результаты относительной экспрессии (V6) РНК AAD1 на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлены на фиг. 43А. Иллюстративные результаты относительной экспрессии (V6) белка AAD1 (ЖХМС) с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлены на фиг. 43В.
Иллюстративные результаты относительной экспрессии (V6) РНК YFP на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлены на фиг. 44А. Иллюстративные результаты относительной экспрессии (V6) белка YFP (ЖХМС) с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлены на фиг. 44В.
Иллюстративные результаты относительной экспрессии (V6) РНК Cry35 на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлены на фиг. 45А. Иллюстративные результаты относительной экспрессии (V6) белка Cry35 (ELISA) с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлены на фиг. 45В.
На фиг. 46 представлены результаты относительной экспрессии (V6) РНК PAT на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
Статистический анализ результатов экспрессии (V6) РНК Cry34 на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг.47А. Статистический анализ результатов экспрессии (V6) белка Cry34 с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг. 47B. Средние значения и статистические результаты перечислены.
Статистический анализ результатов экспрессии (V6) РНК AAD1 на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг. 48А. Статистический анализ результатов экспрессии (V6) белка AAD1 с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг. 48B. Средние значения и статистические результаты перечислены.
Статистический анализ результатов экспрессии (V6) РНК YFP на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг. 49А. Статистический анализ результатов экспрессии (V6) белка YFP с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг. 49B. Средние значения и статистические результаты перечислены.
Статистический анализ результатов экспрессии (V6) РНК Cry35 на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг. 50А. Статистический анализ результатов экспрессии (V6) белка Cry35 с тех же самых шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720 представлен на фиг. 50B. Средние значения и статистические результаты перечислены.
На фиг. 51 представлен статистический анализ результатов экспрессии (V6) РНК PAT на шести конструктах pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Средние значения и статистические результаты перечислены.
На фиг.52А, 52В, 52С и 52D представлены иллюстративные результаты экспрессии (V10) белков YFP, AAD1, Cry34 и Cry35, соответственно, с шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 53А, 53В, 53С и 53D представлен статистический анализ результатов экспрессии (V10) белков YFP, AAD1, Cry34 и Cry35, соответственно, с шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Средние значения и статистические результаты перечислены.
На фиг. 54А, 54В, 54С и 54D представлены иллюстративные результаты экспрессии (R3) белков YFP, AAD1, Cry34 и Cry35, соответственно, с шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720.
На фиг. 55А, 55В, 55С и 55D представлен статистический анализ результатов экспрессии (R3) белков YFP, AAD1, Cry34 и Cry35, соответственно, с шести конструктов pDAB105748, pDAB105818, pDAB108717, pDAB108718, pDAB108719 и pDAB108720. Средние значения и статистические результаты перечислены.
На фиг. 56 представлены иллюстративные результаты Вестерн-блота по экспрессии белков Cry34, Cry35 и AAD1 с pDAB108718, pDAB108717, pDAB108719 и pDAB108720.
Результаты демонстрируют, что все четыре трансгена в конструктах, управляемых одним промотором, функциональны с выраженными уровнями экспрессии. Три гена (Cry34/Cry35/AAD1) в конструкции с двунаправленным Ubi1 продемонстрировали стабильные уровни экспрессии подобные уровням экспрессии, обеспечиваемым генной конструкцией, управляемой одним Ubi1 (DExT).
Хотя ряд иллюстративных аспектов и вариантов осуществления изобретения были описаны выше, специалистам в данной области техники будут очевидны определенные модификации, перестановки, дополнения и их суб-комбинации. Следовательно, предполагается, что прилагаемая формула изобретения и внесенные последующие пункты формулы изобретения интерпретируются для включения всех таких модификаций, перестановок, дополнений и суб-комбинаций, не отходя от их существа и объема.
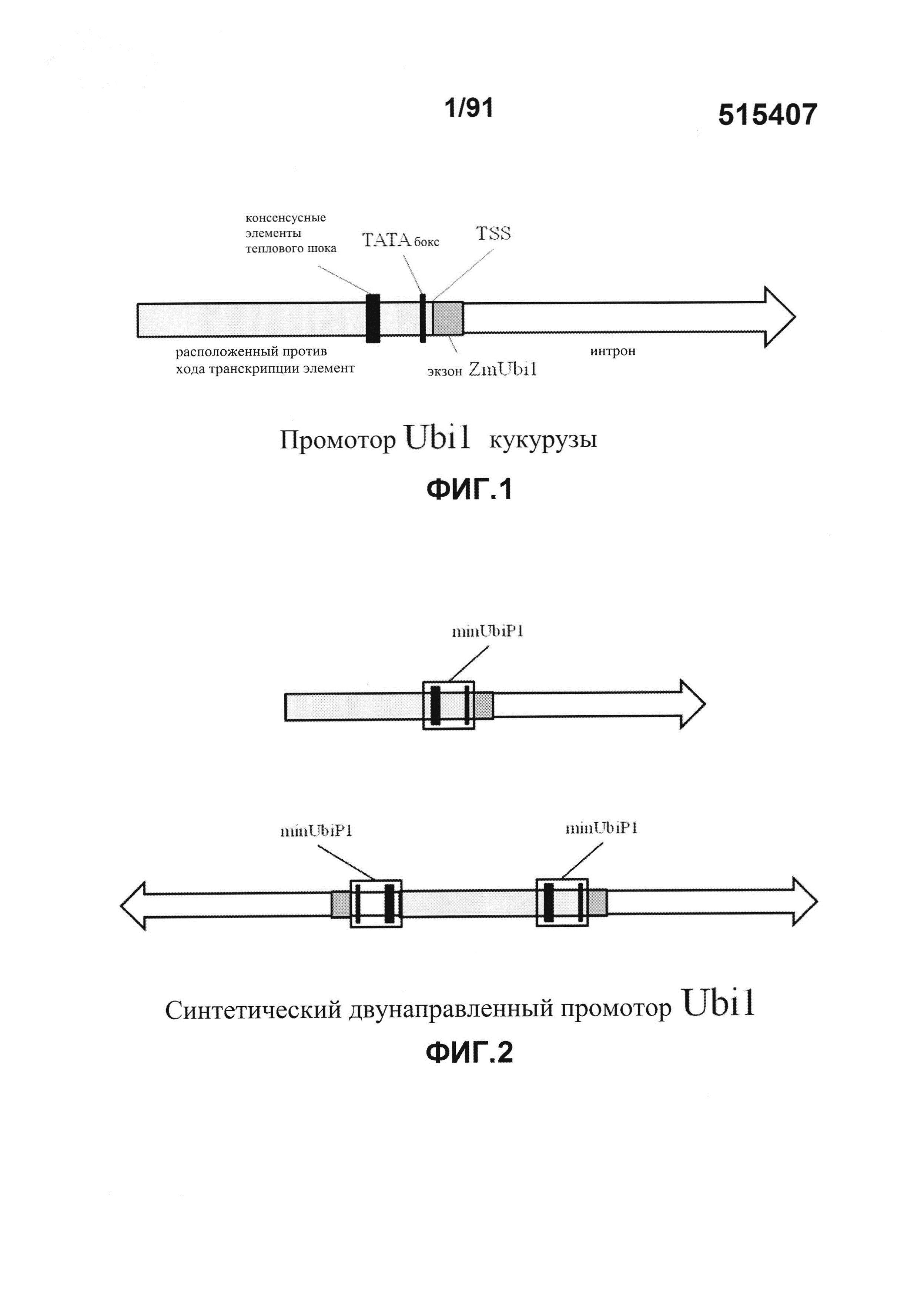
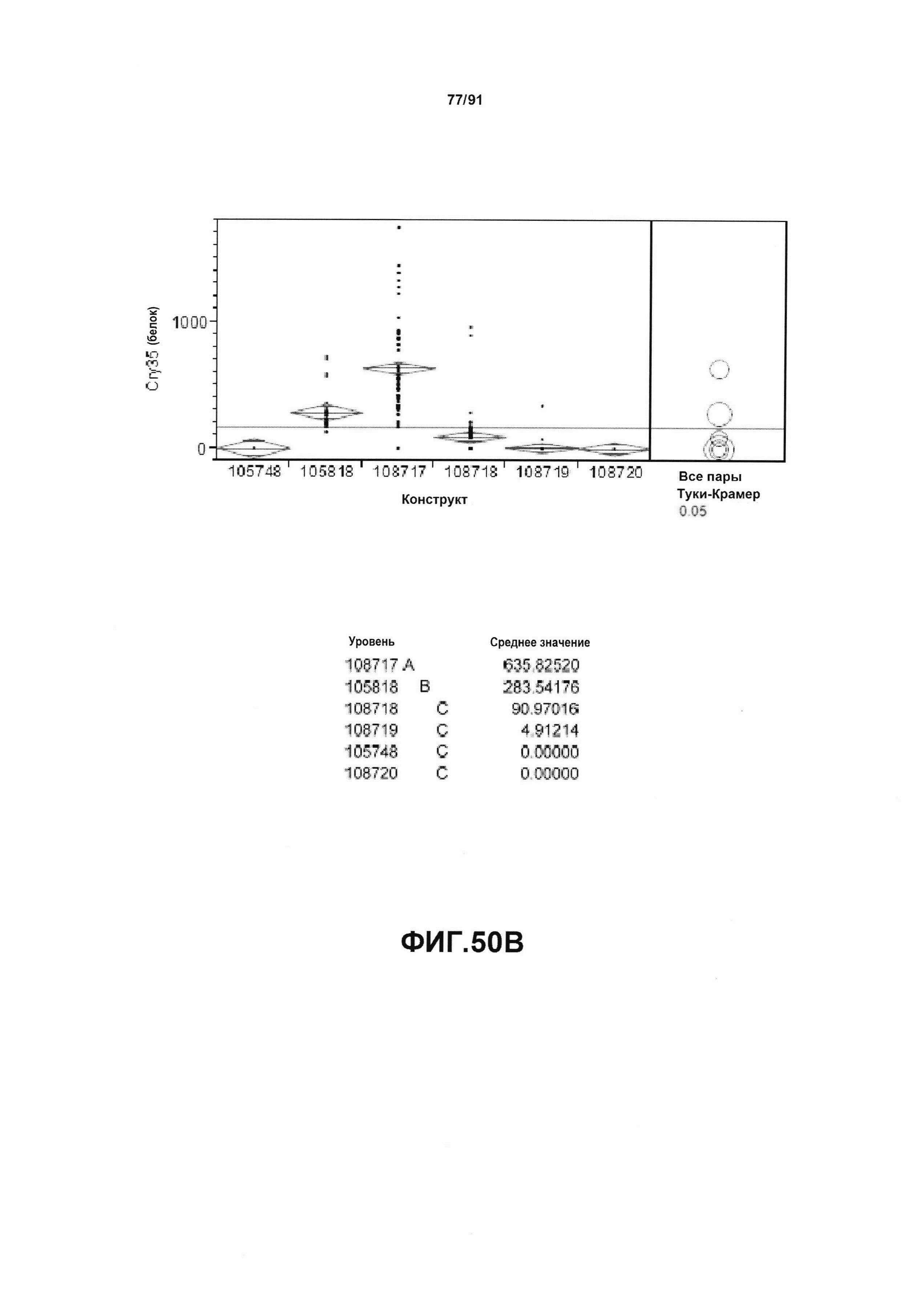

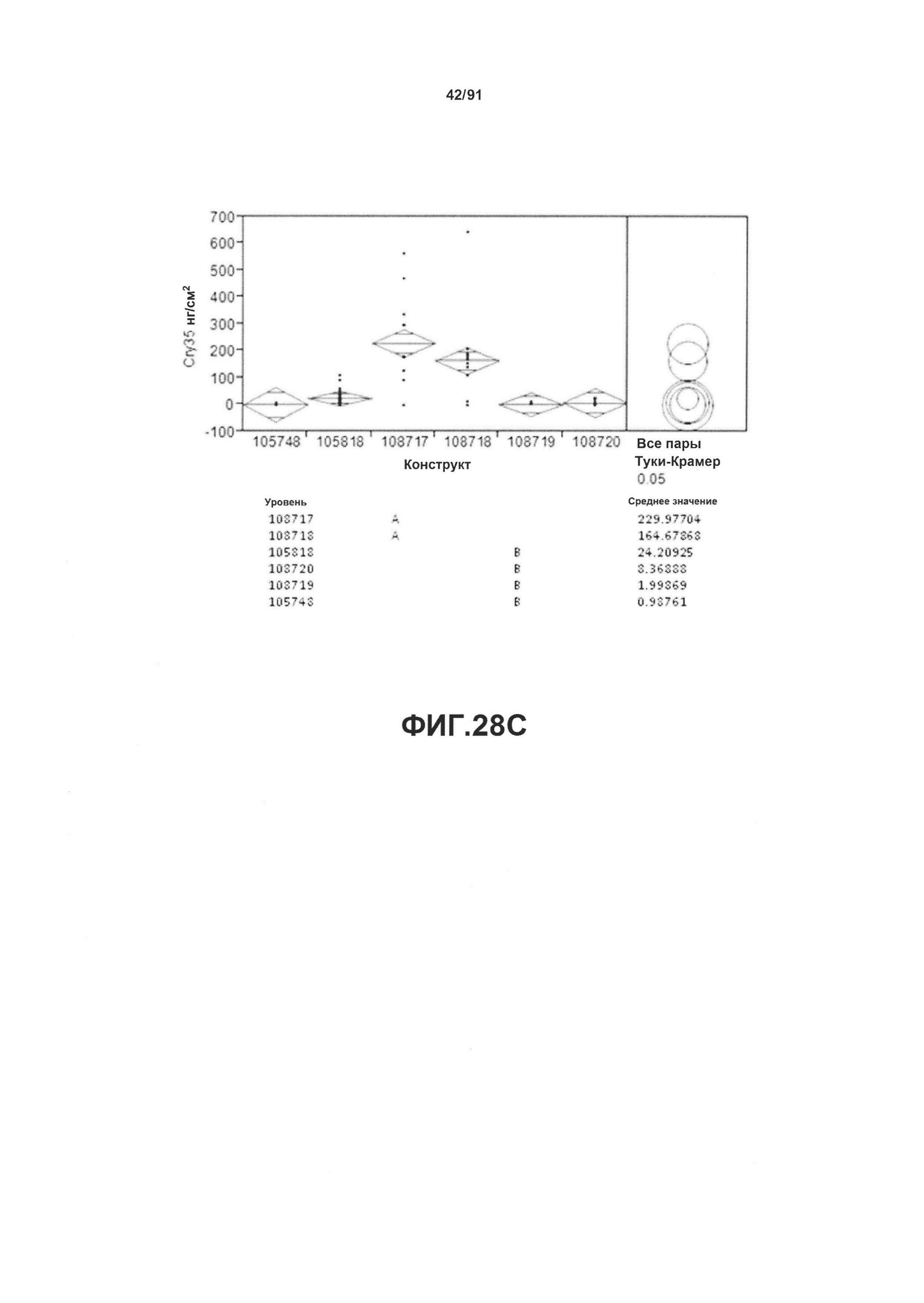





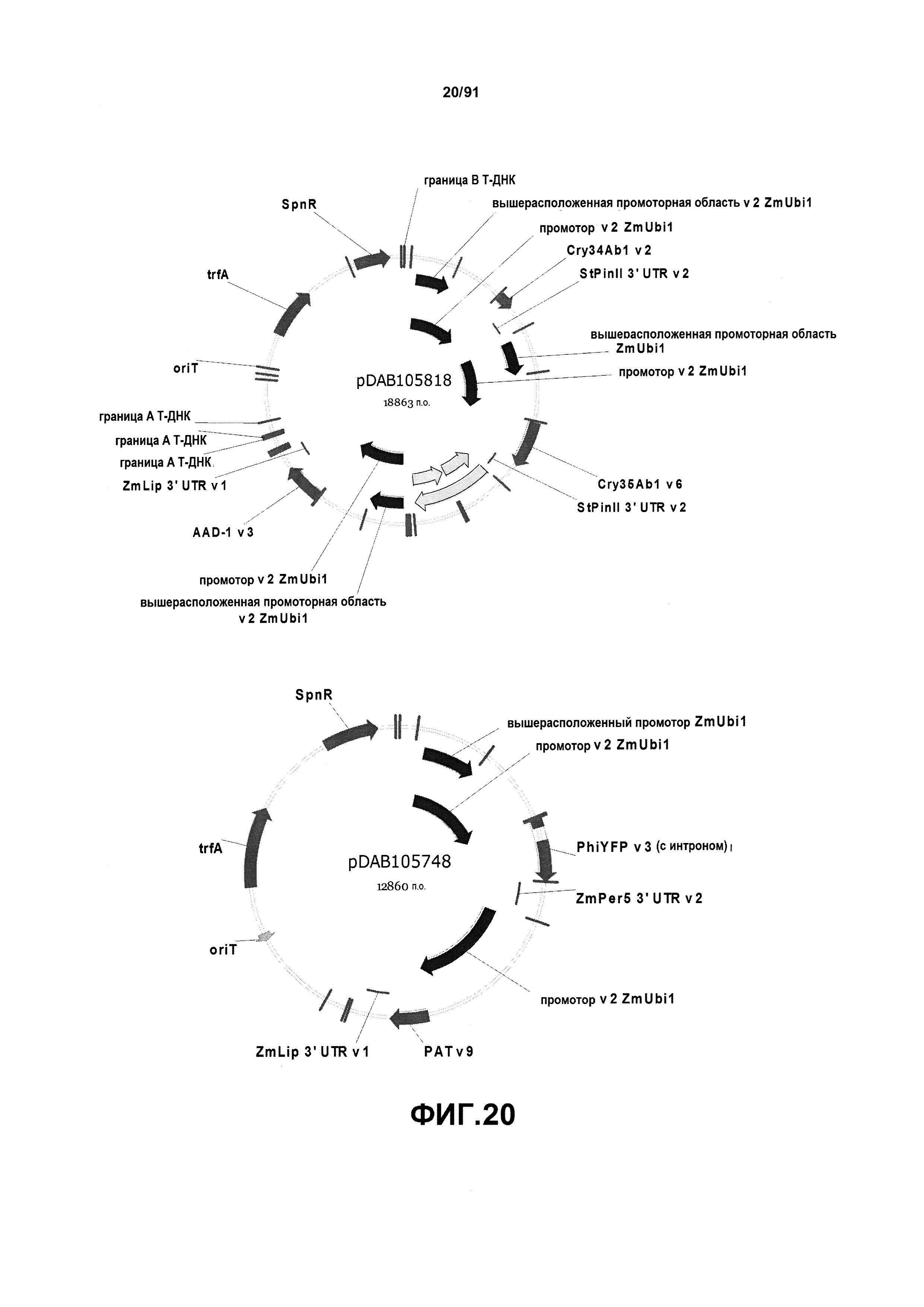
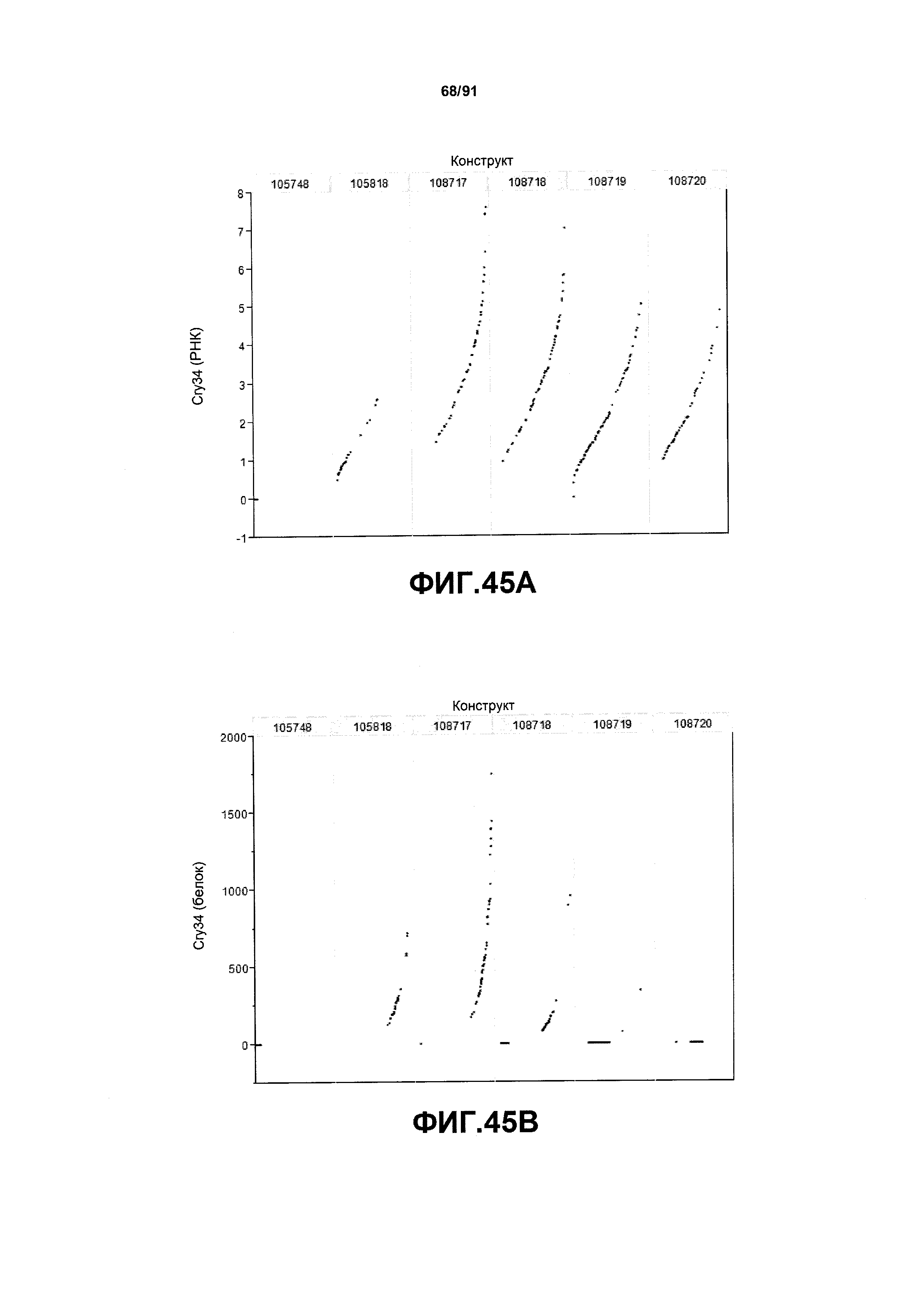


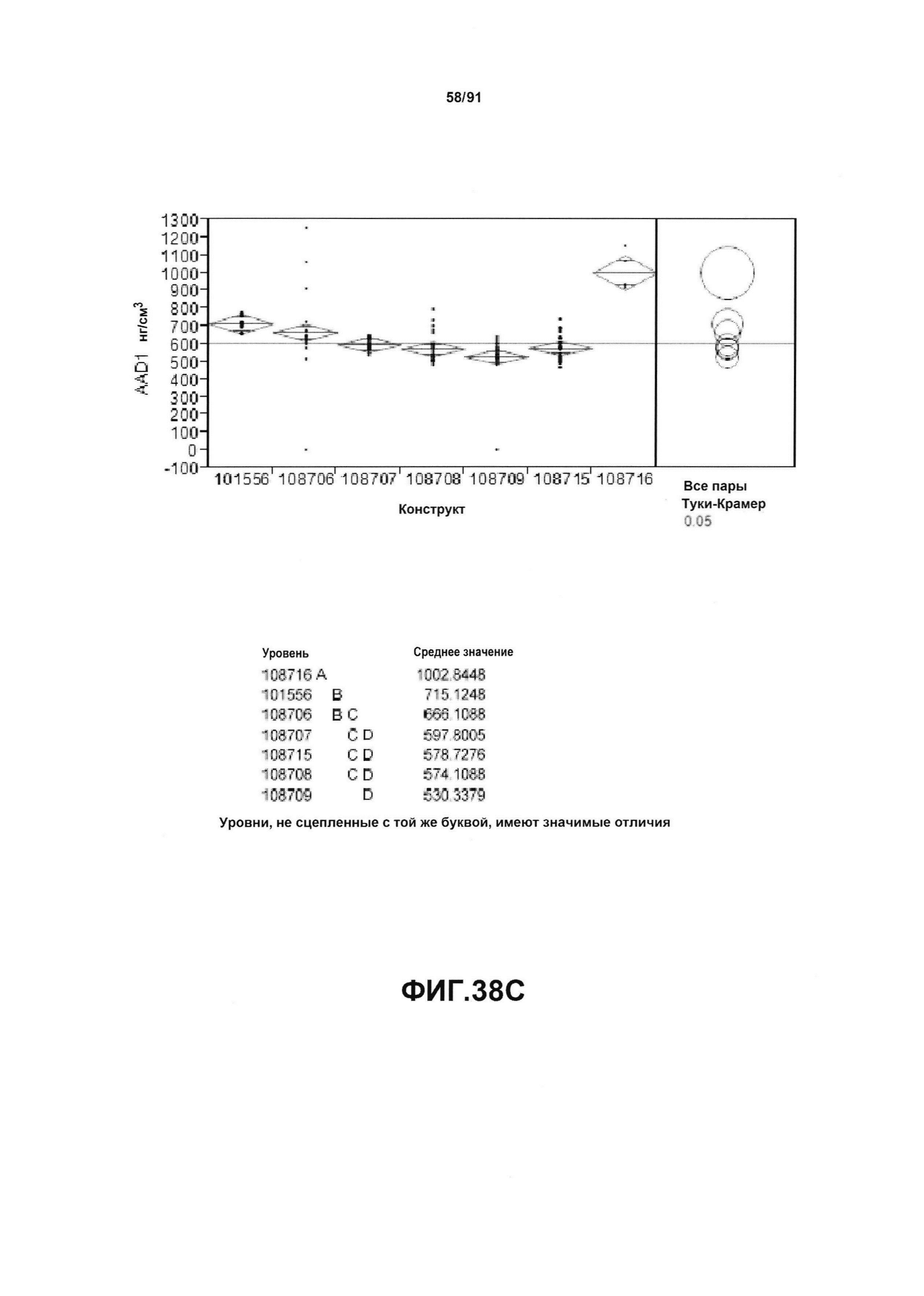


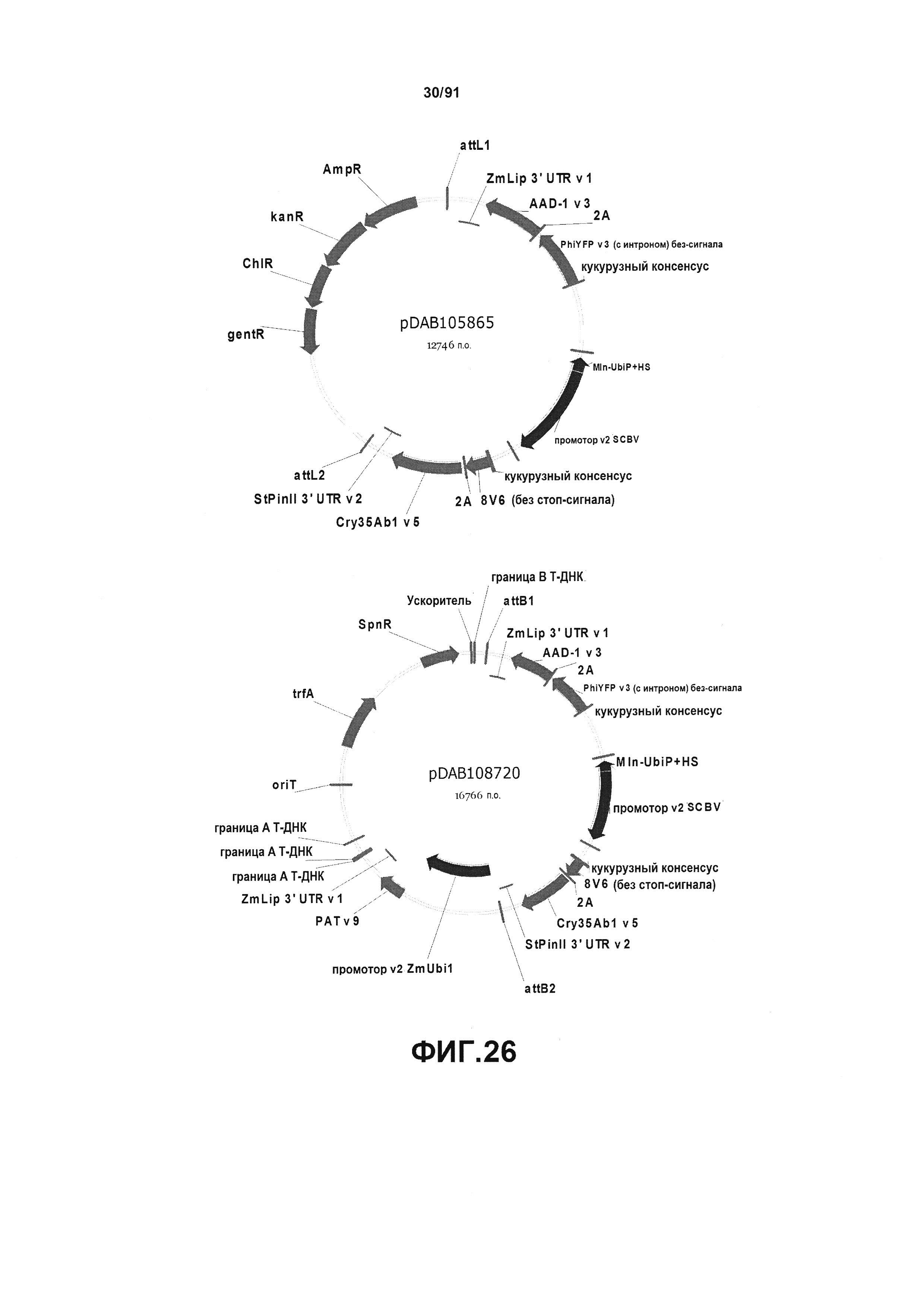


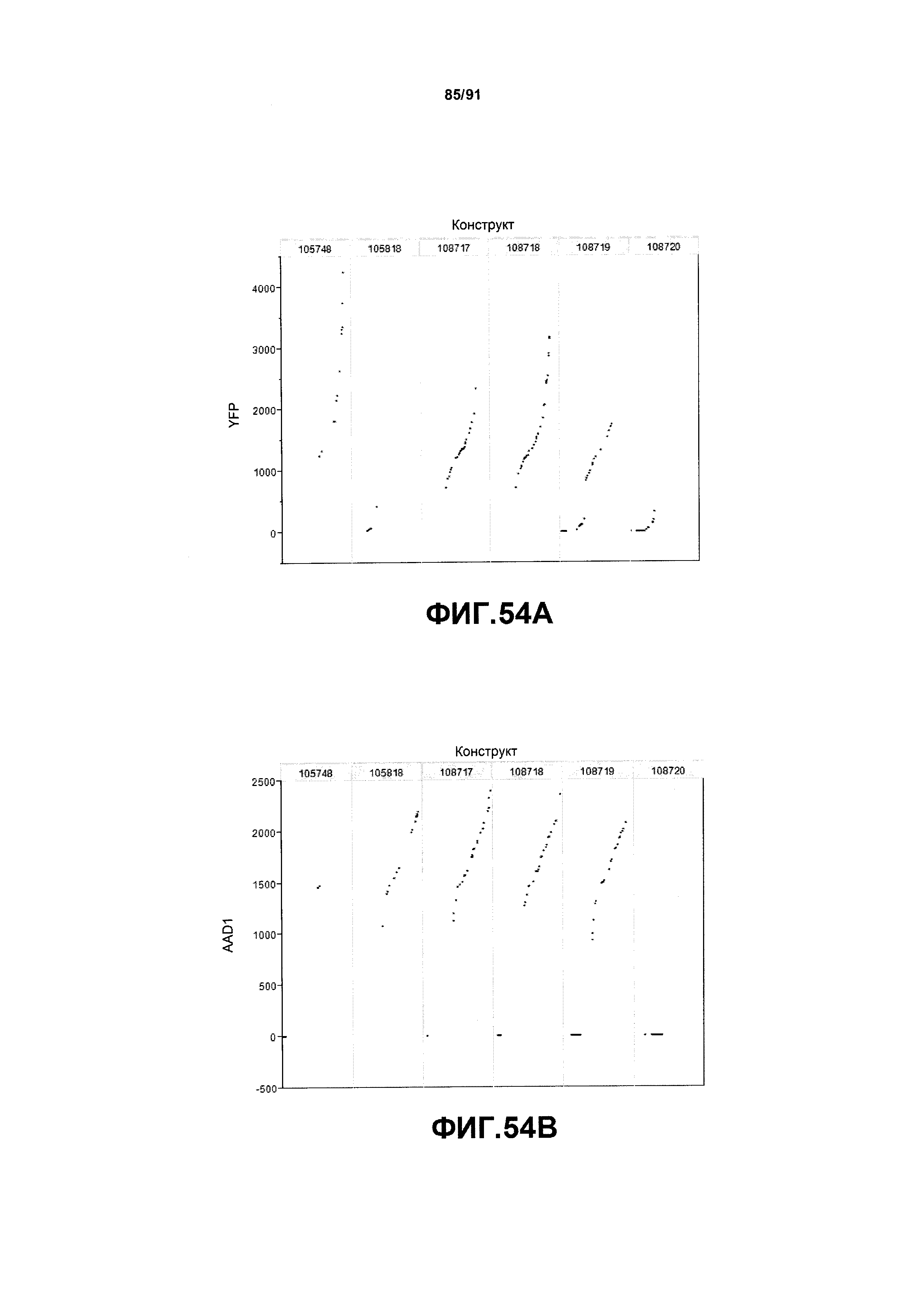


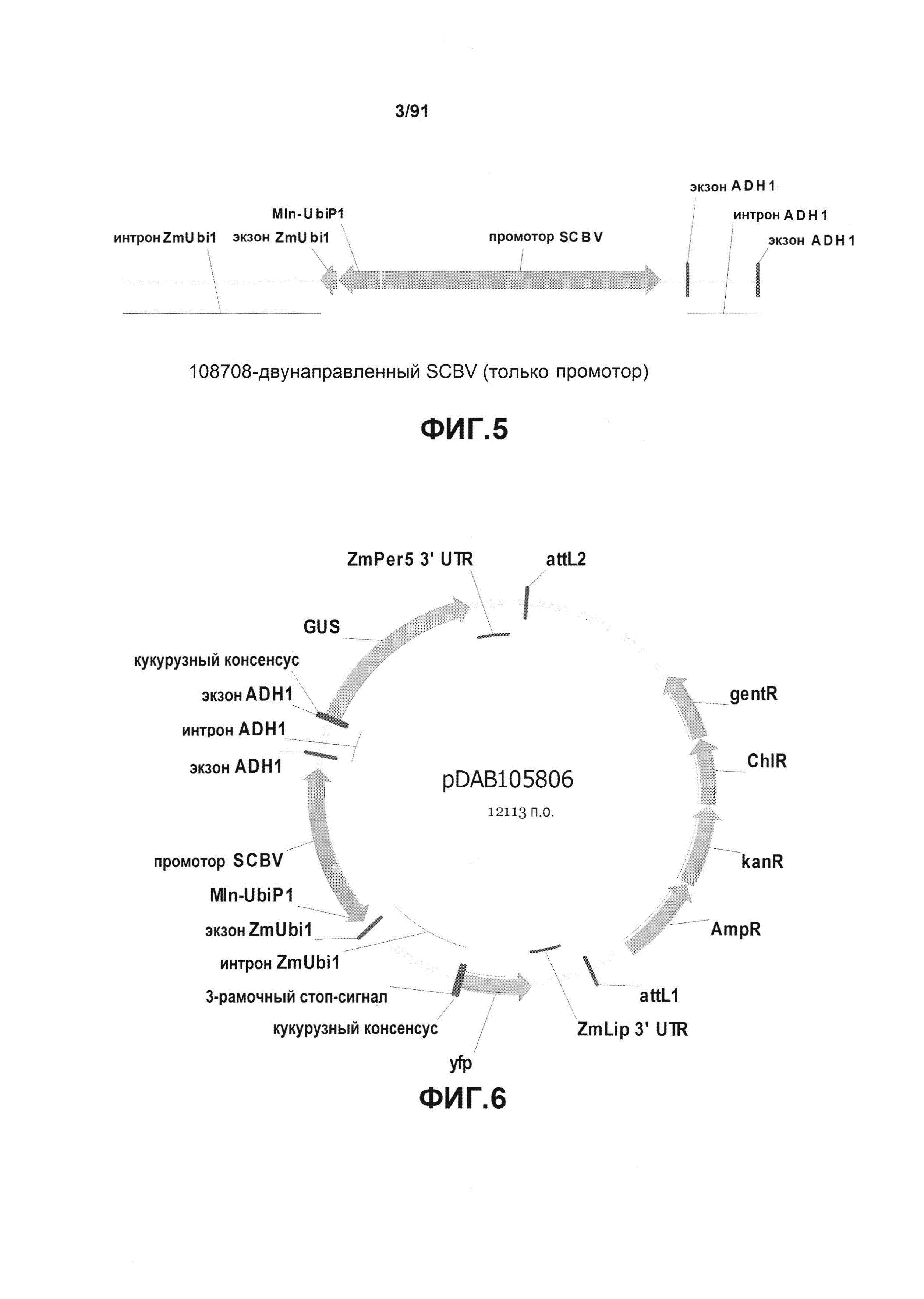
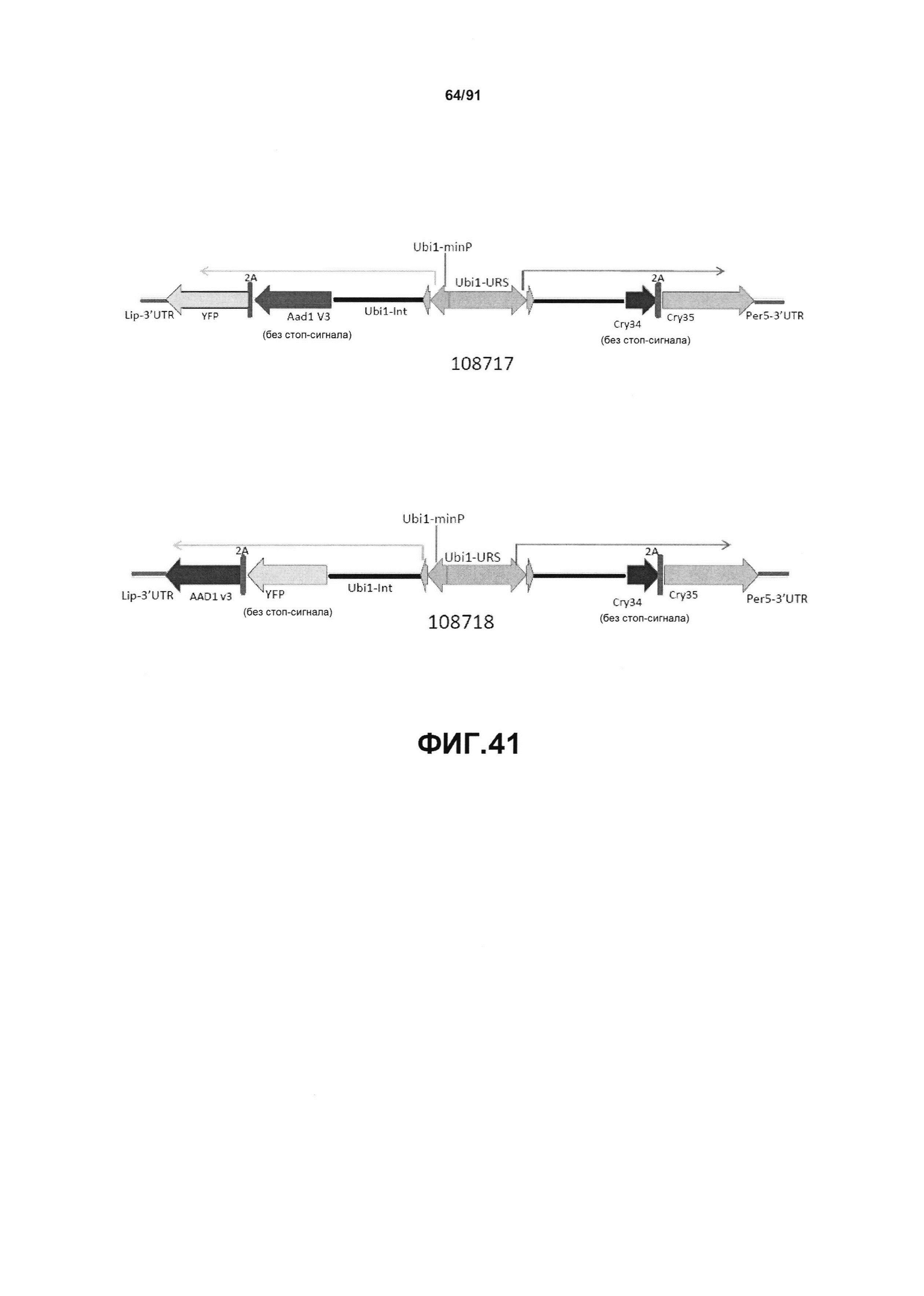
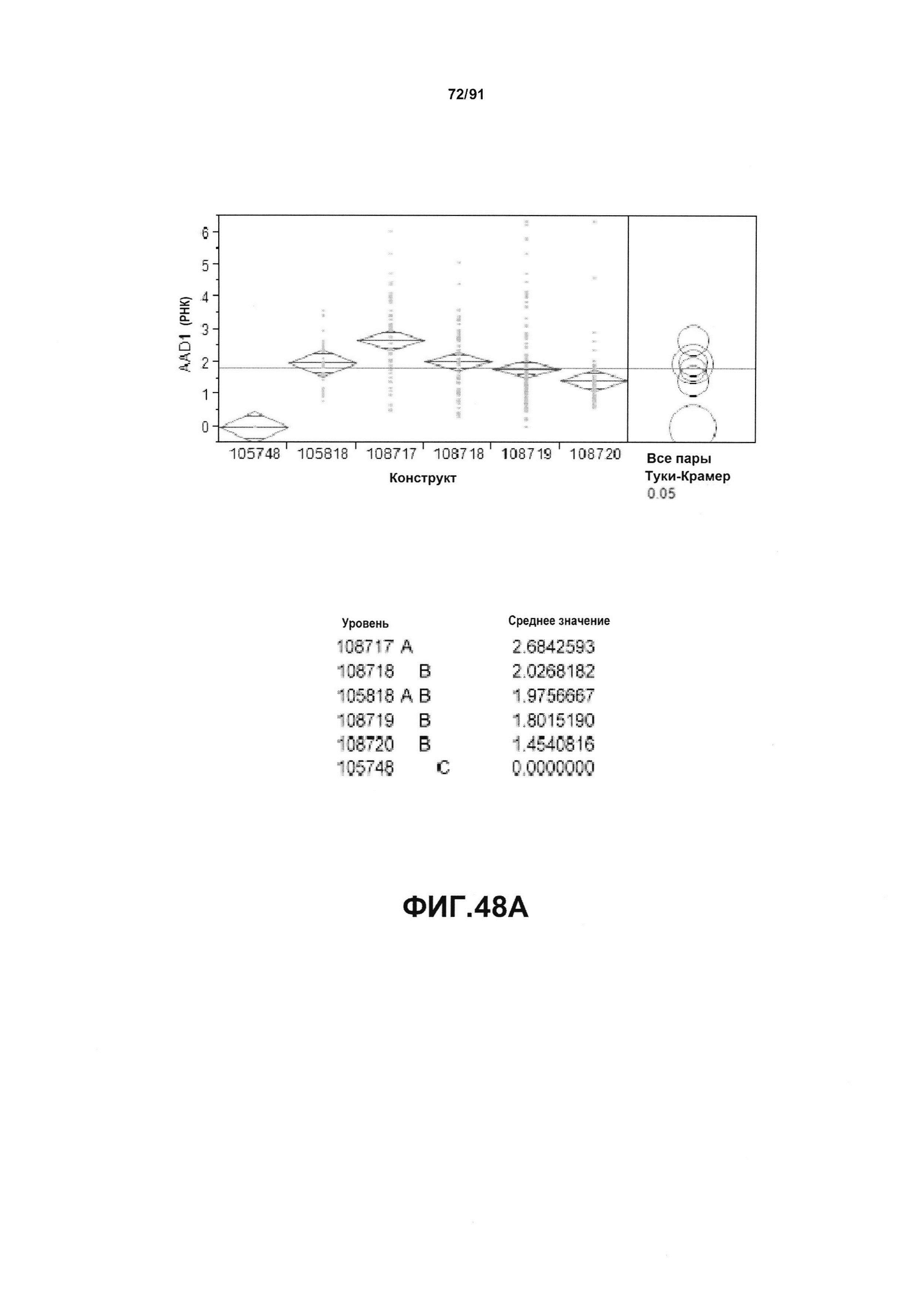




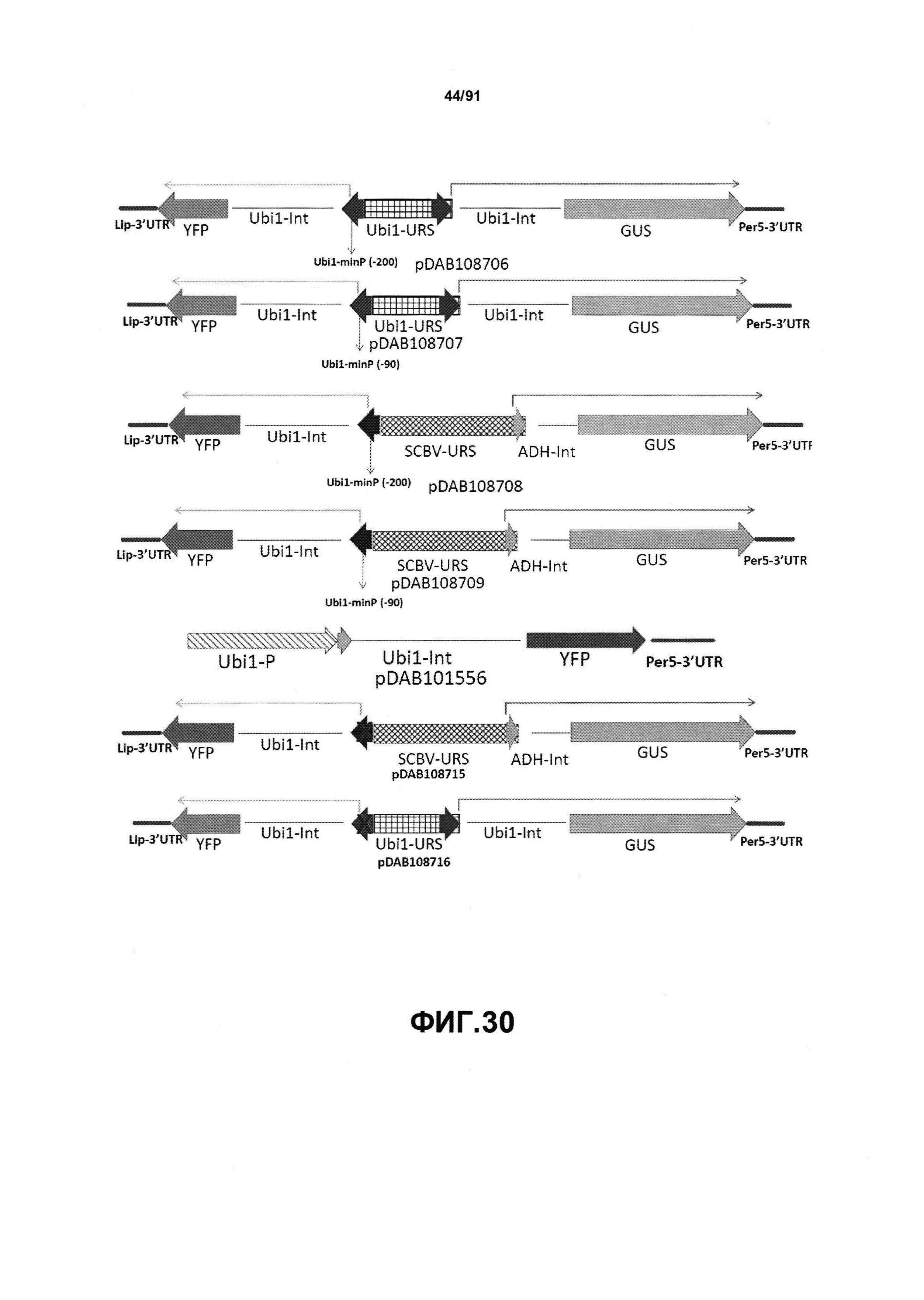
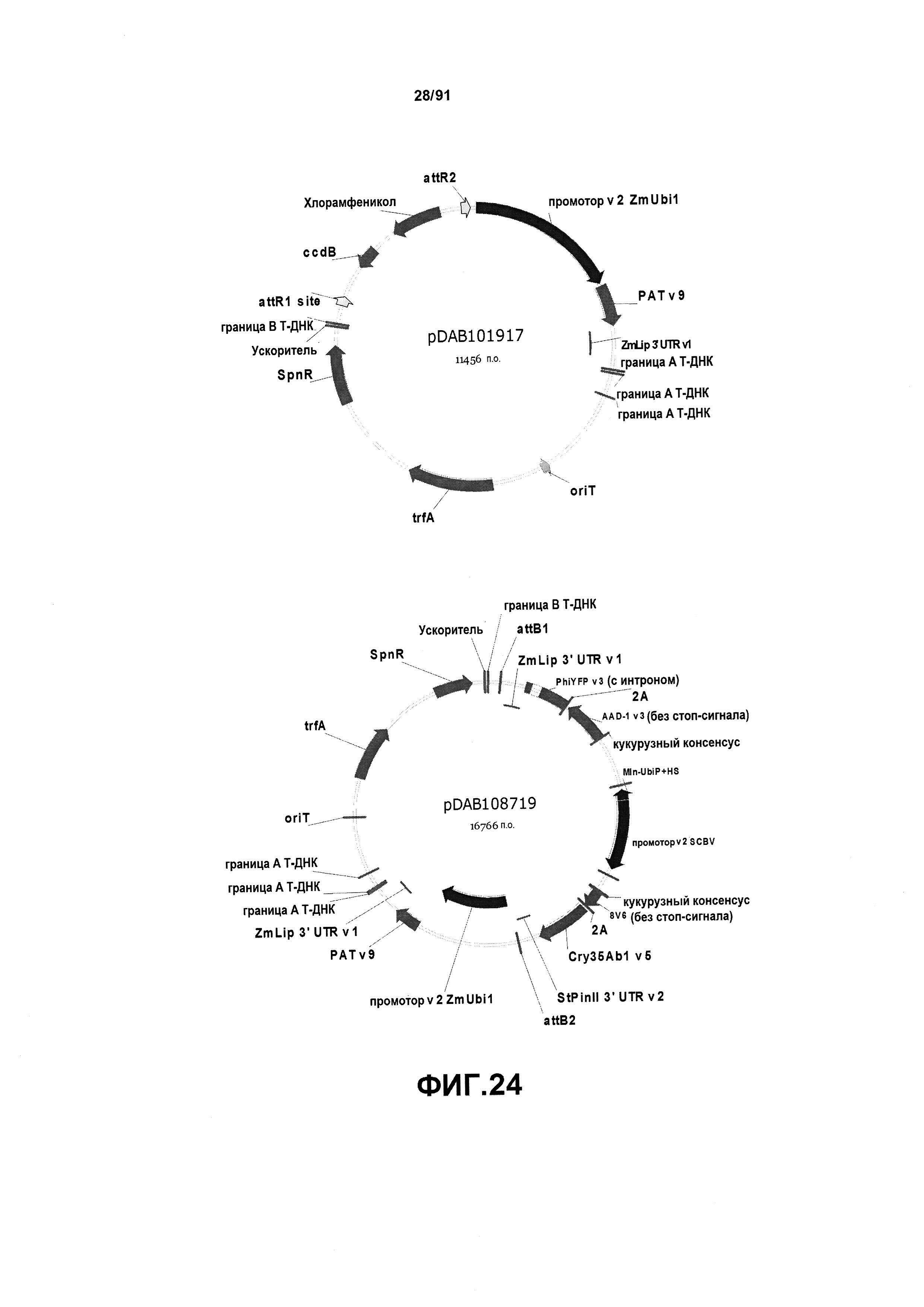
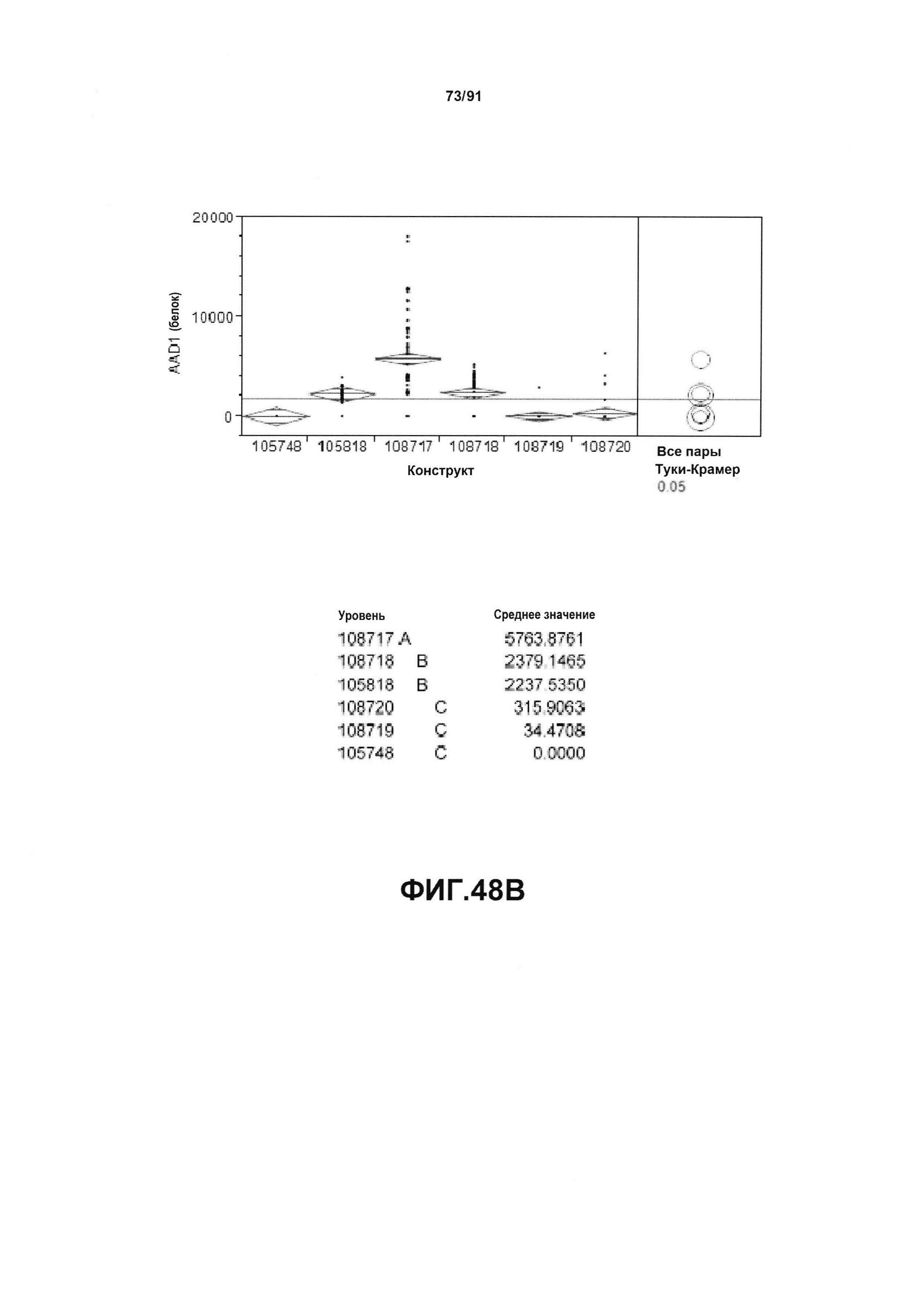
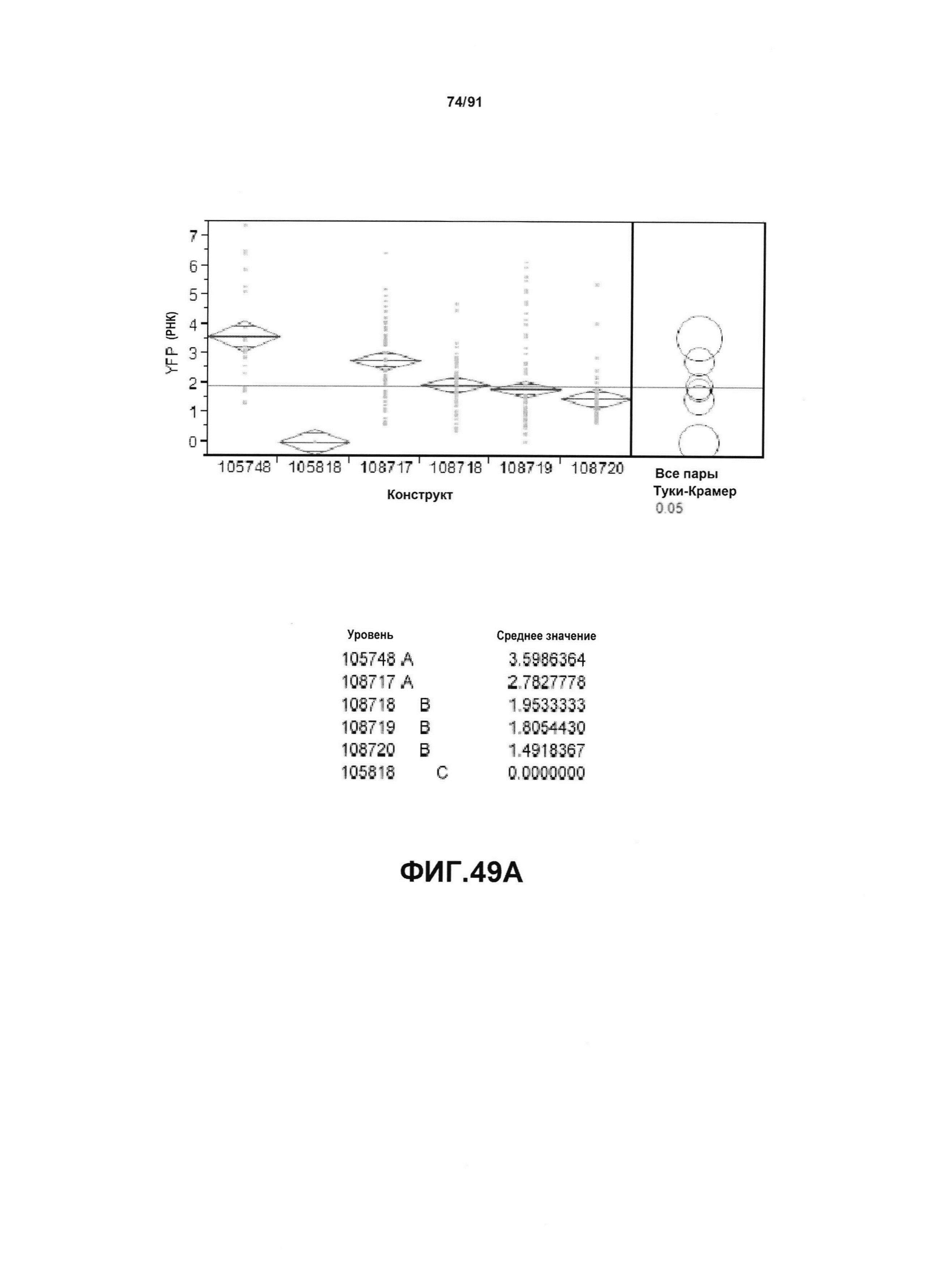



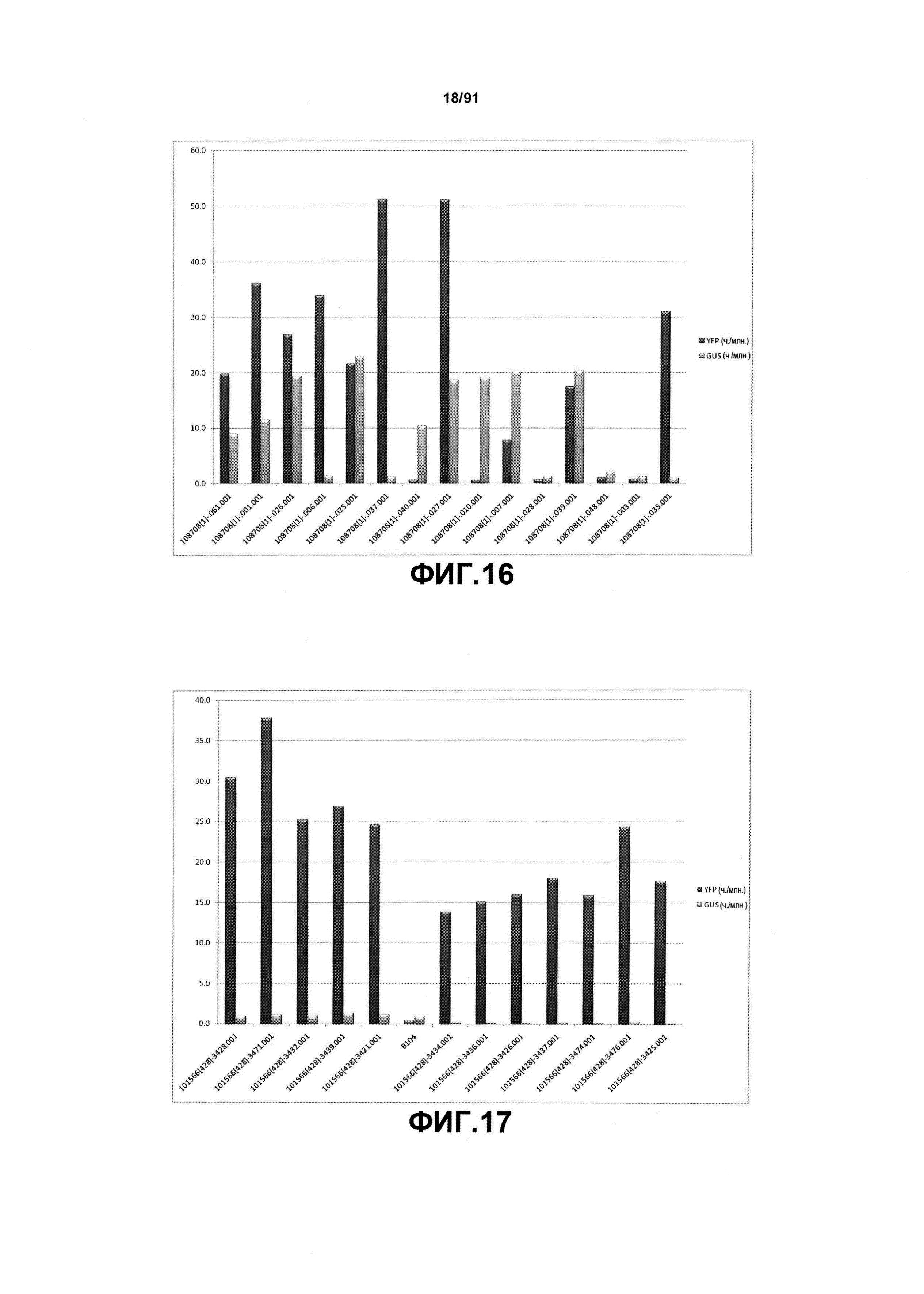






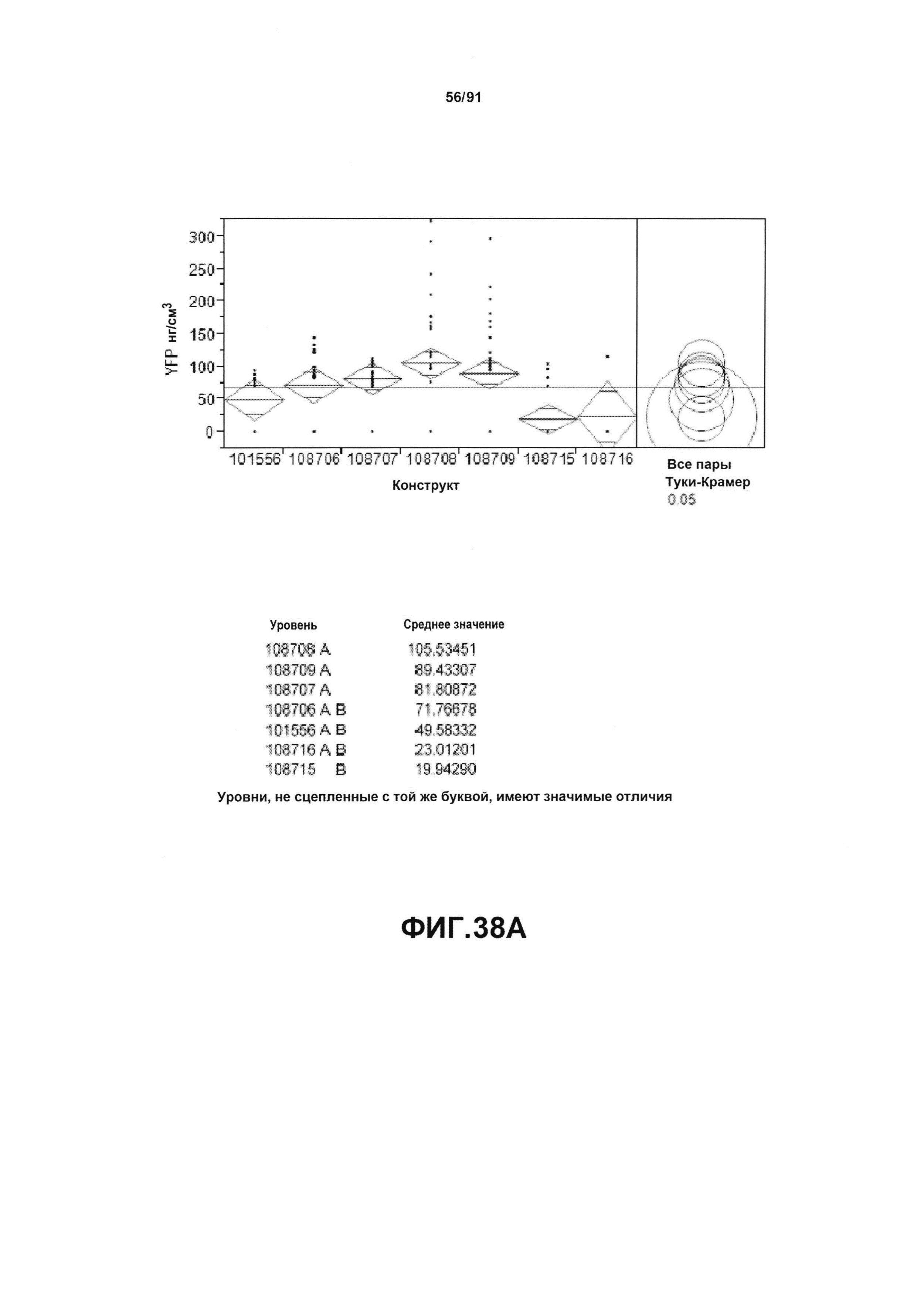



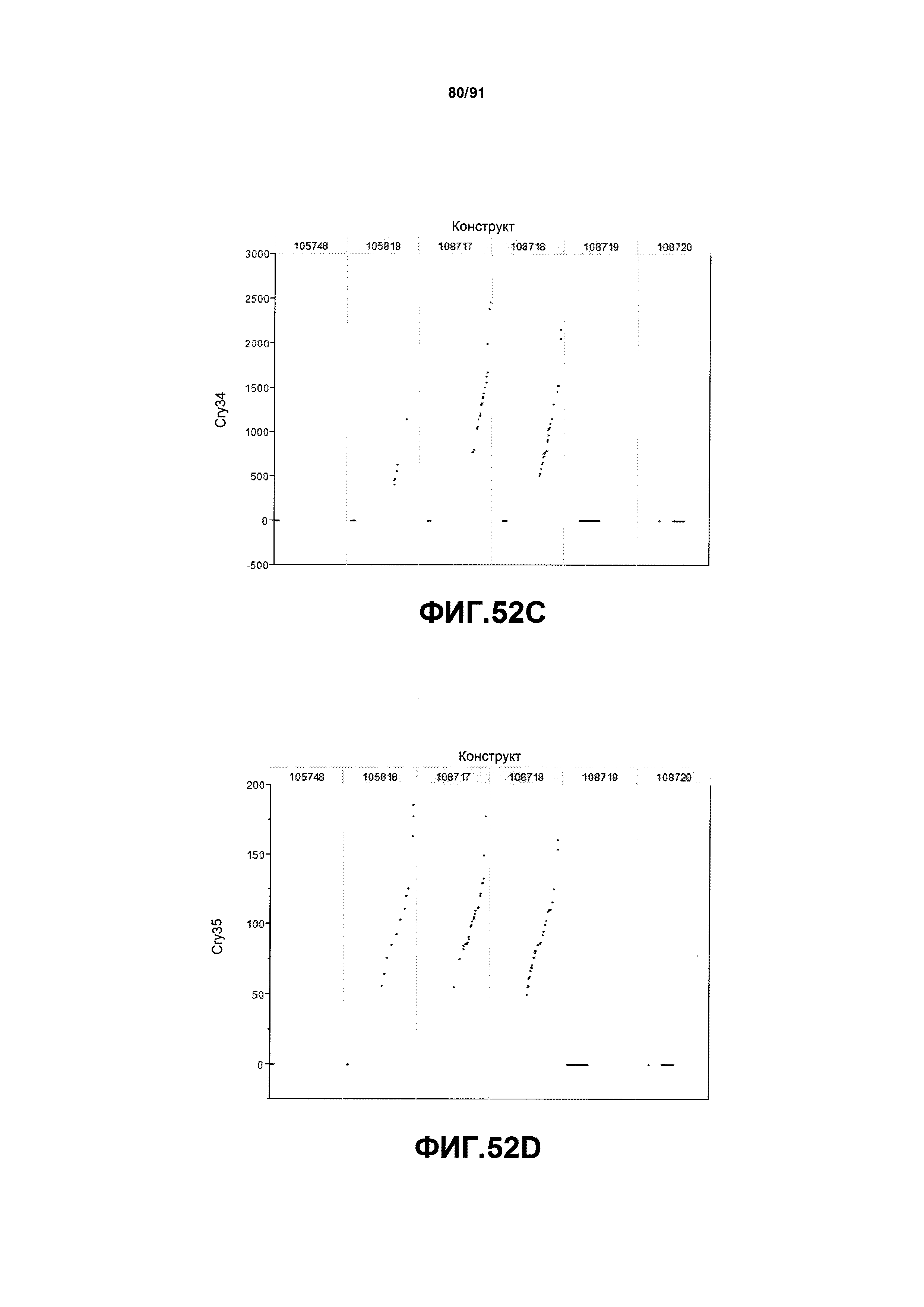

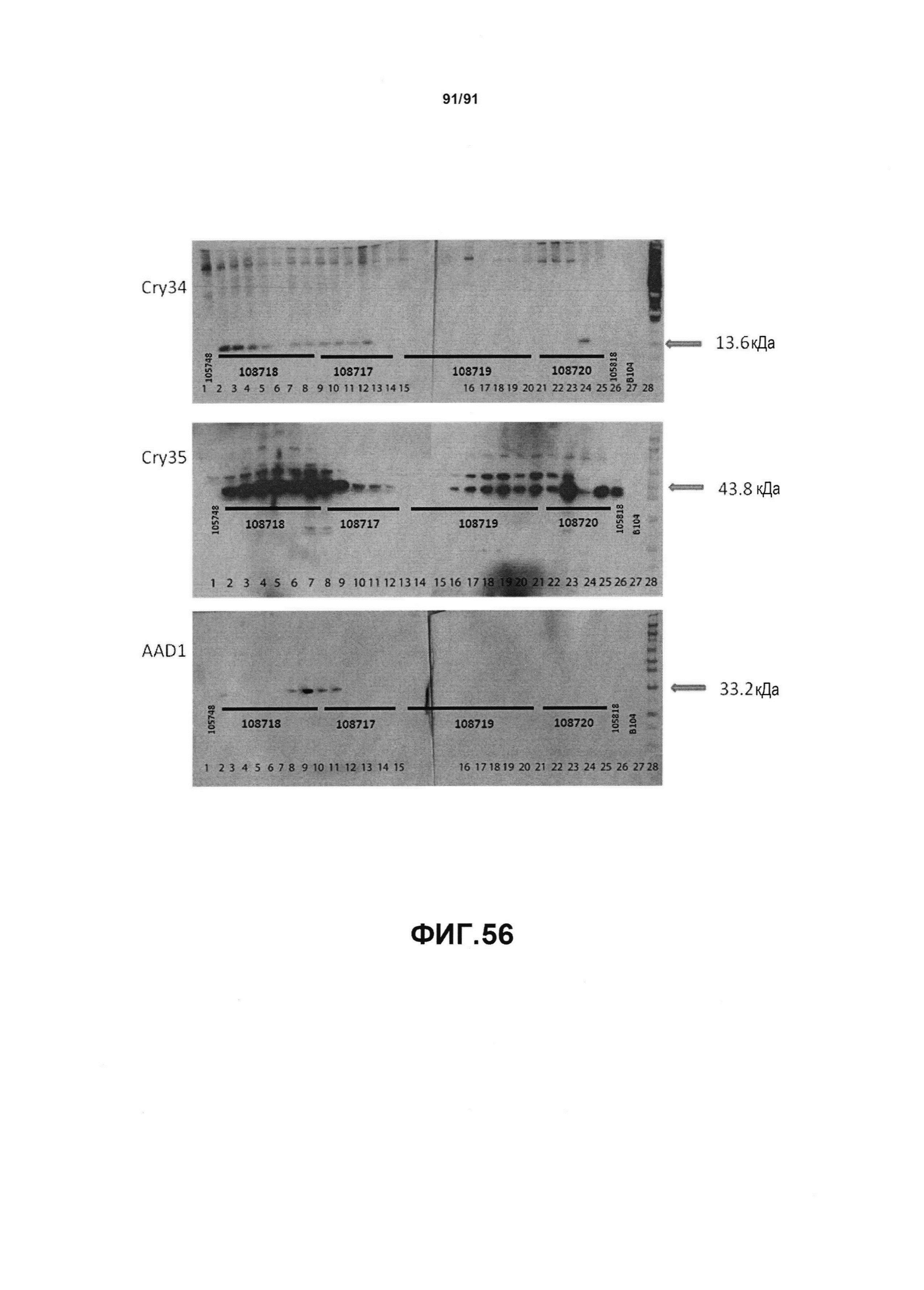
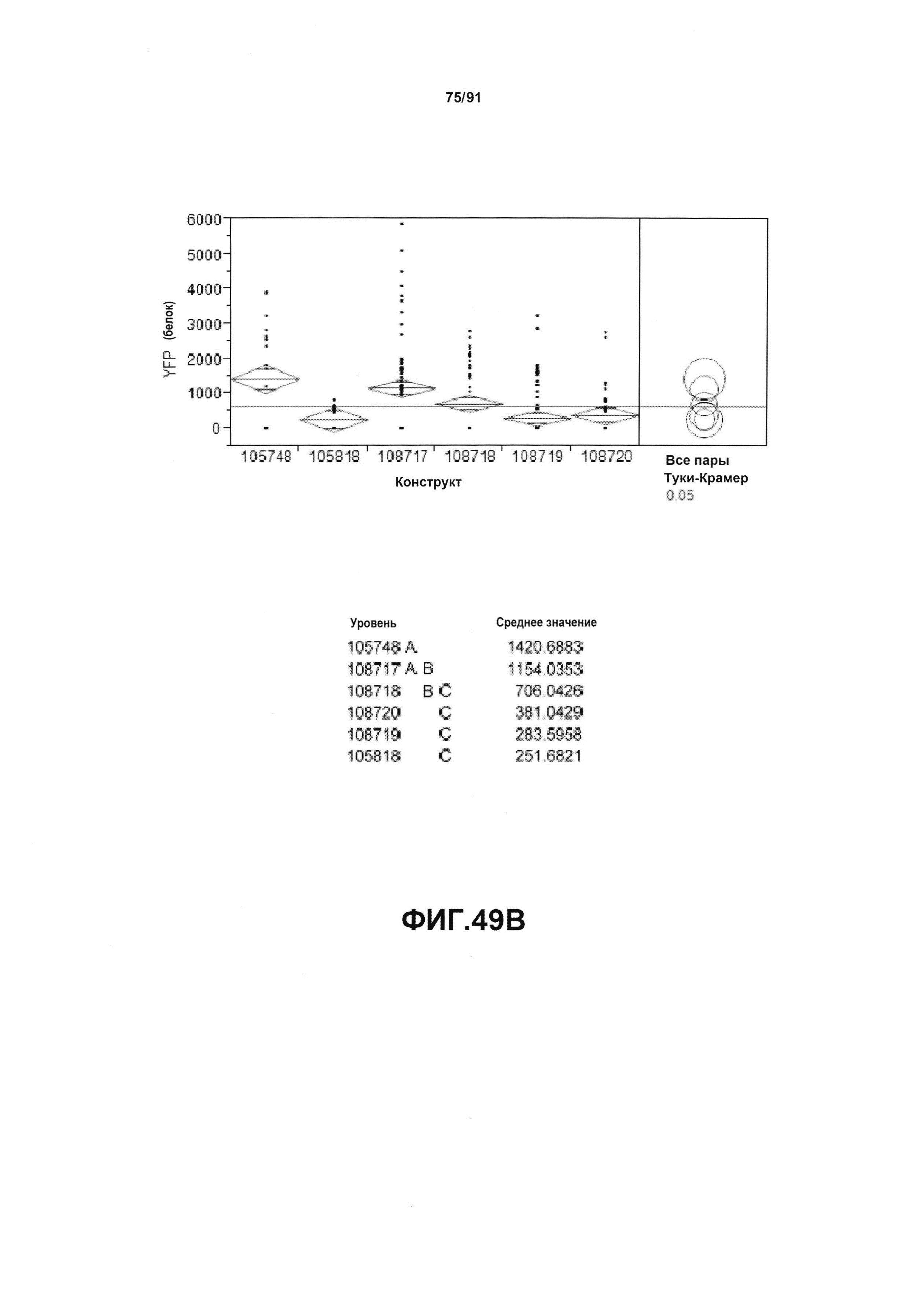
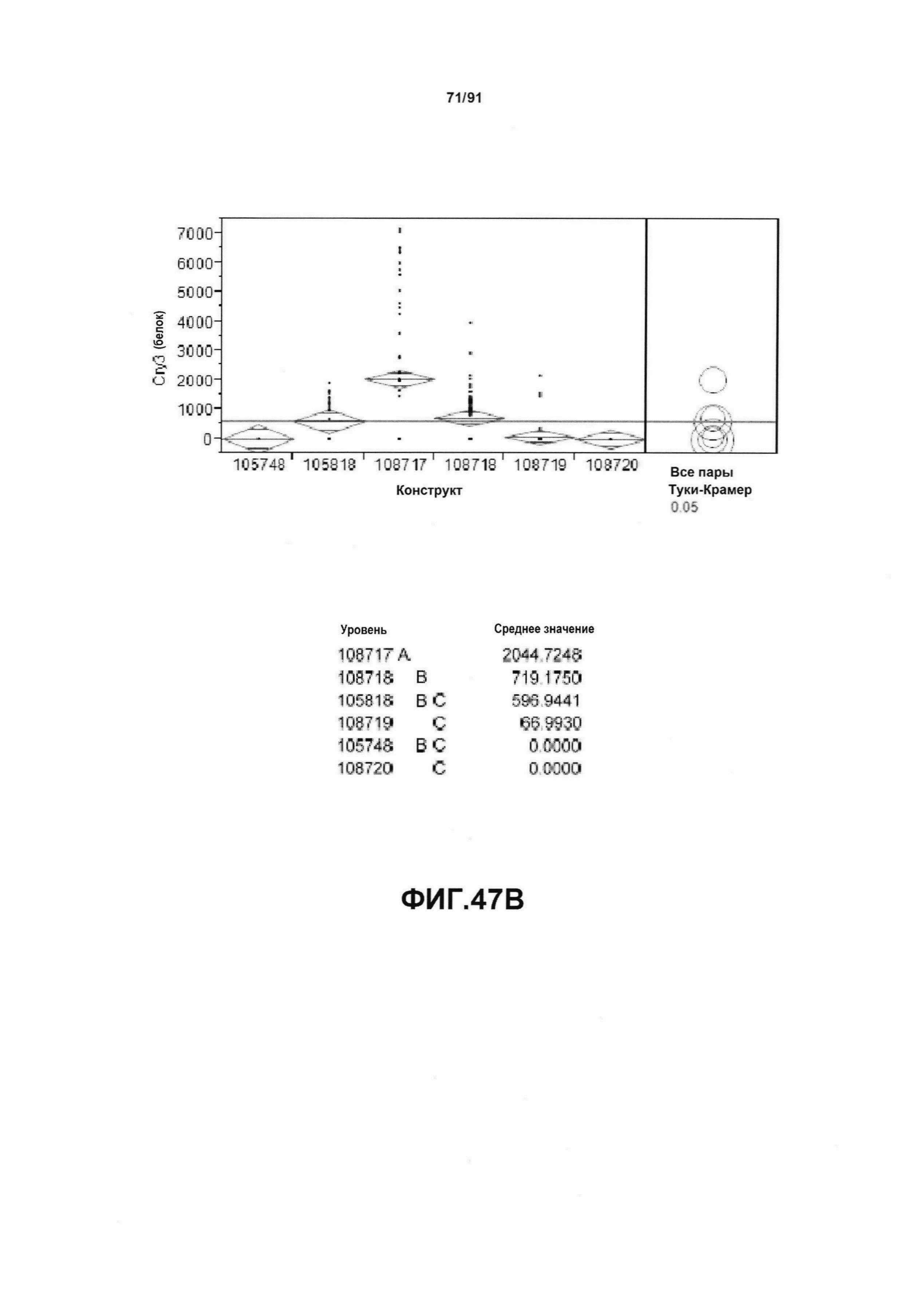
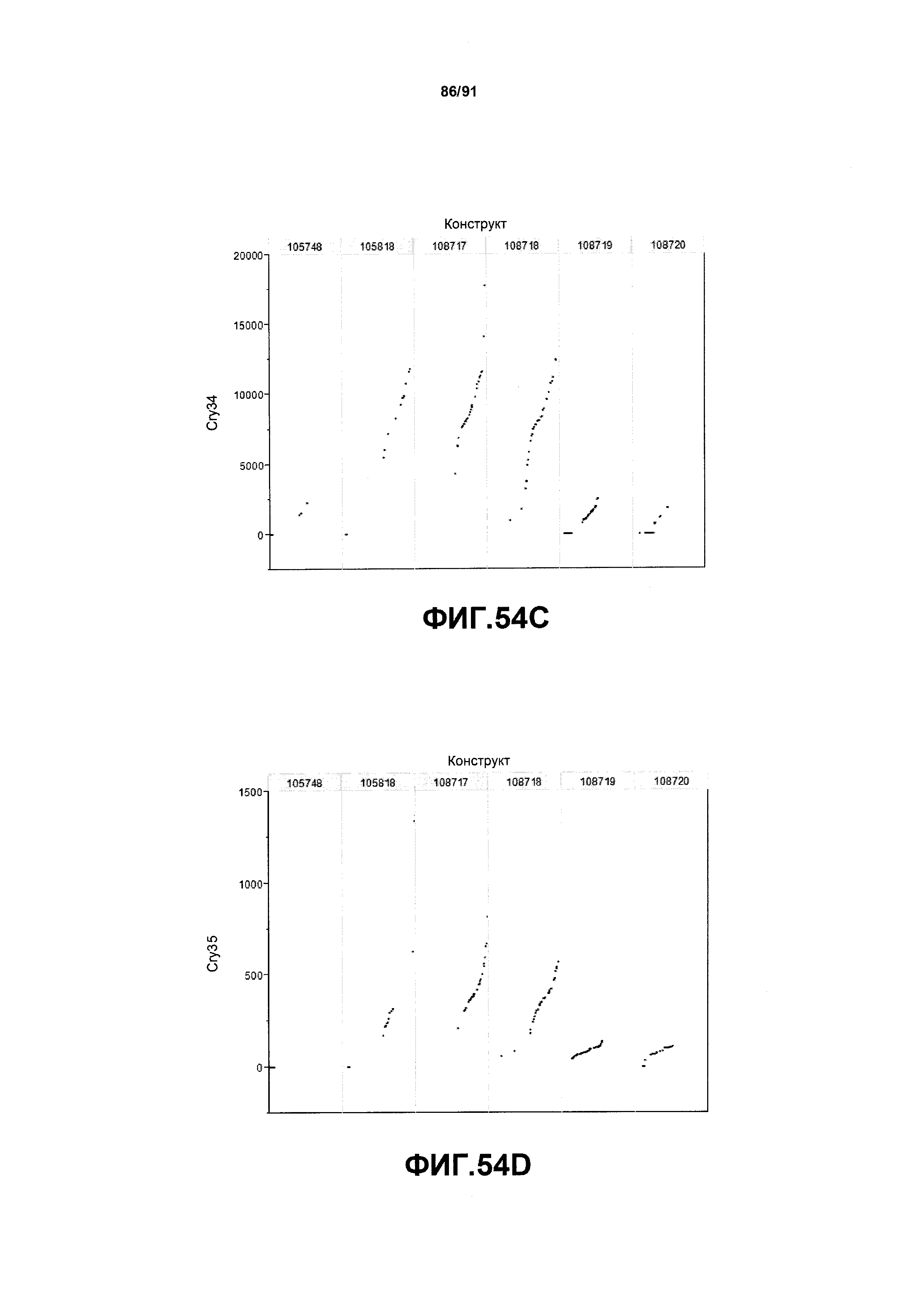

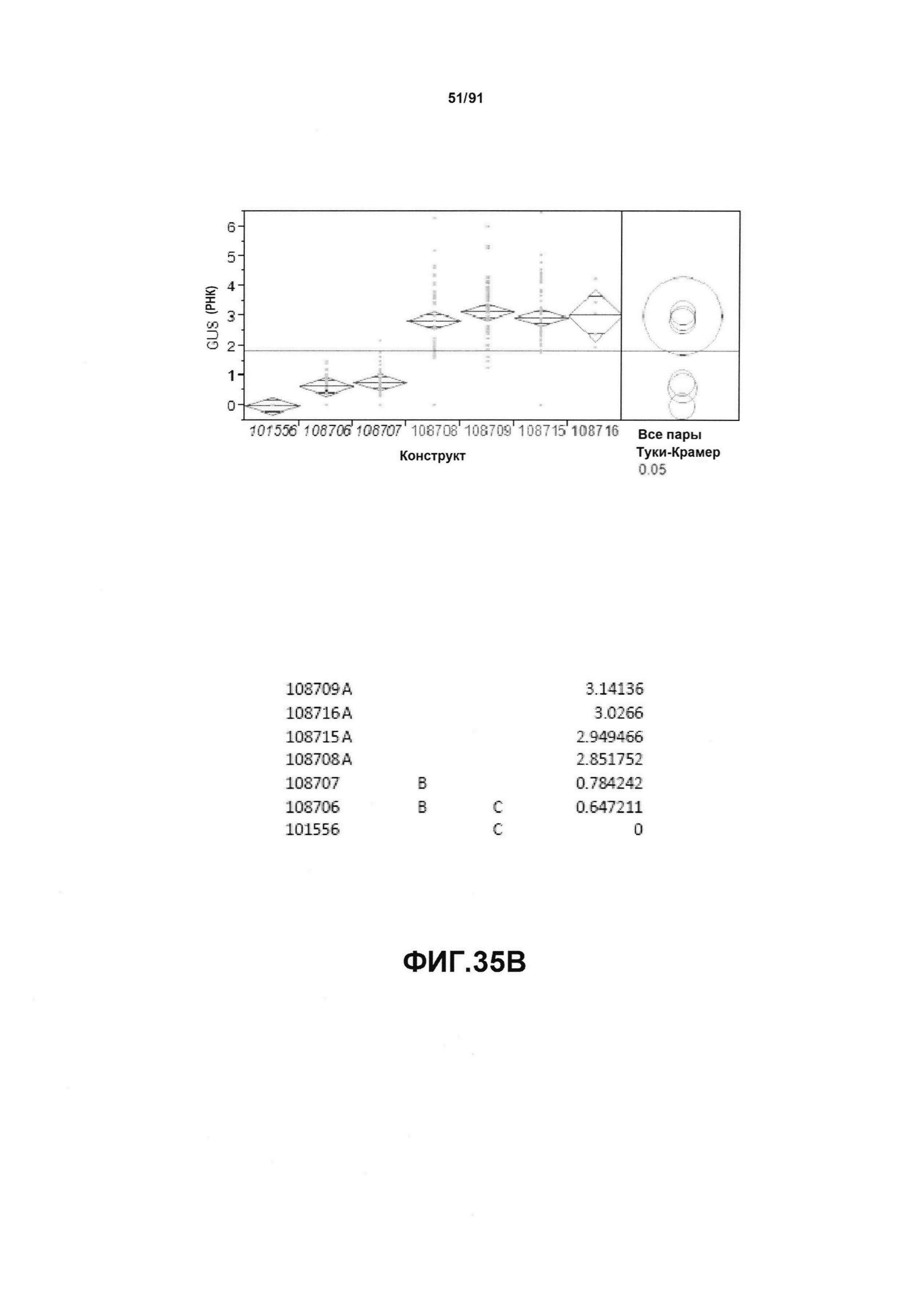

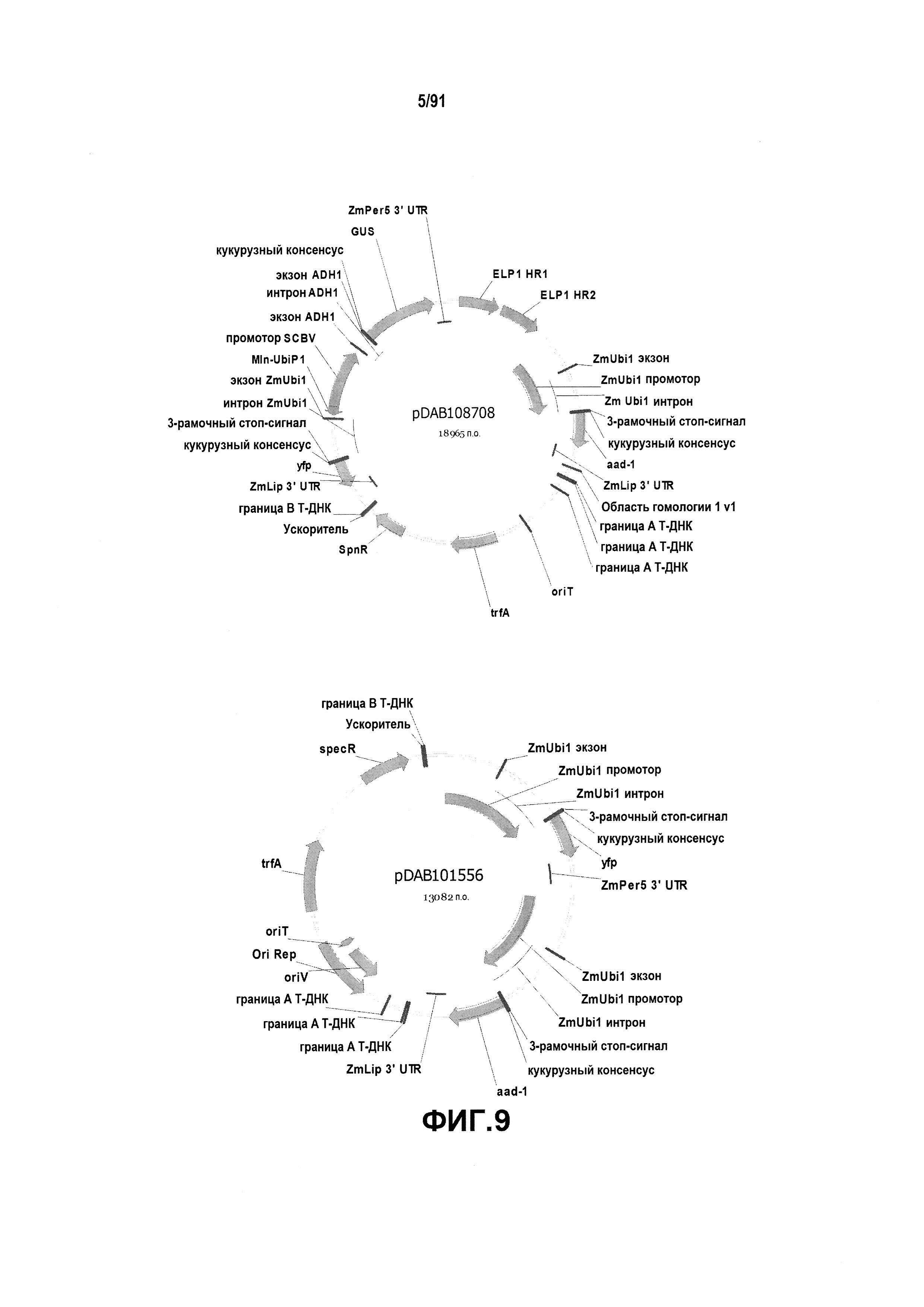
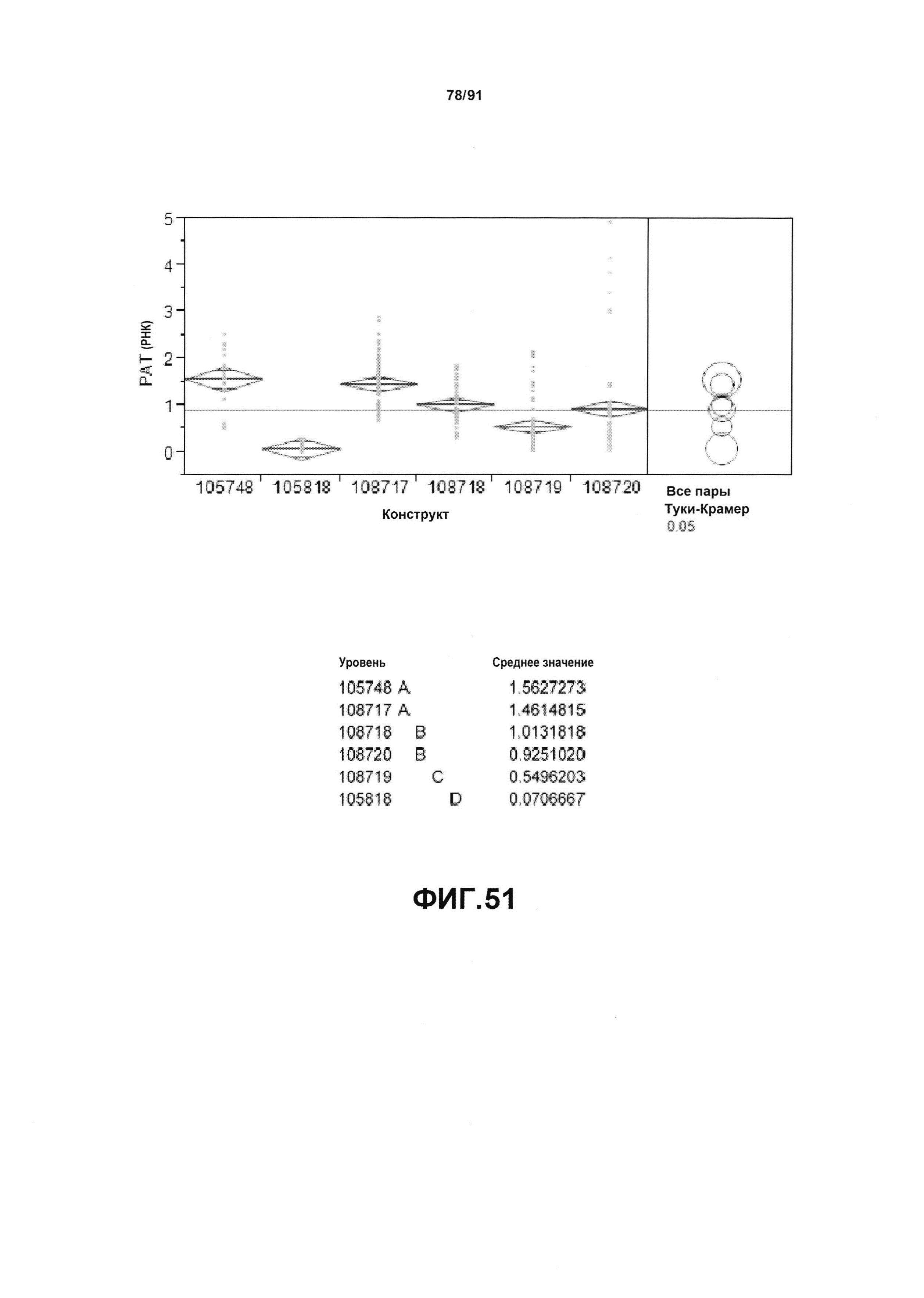
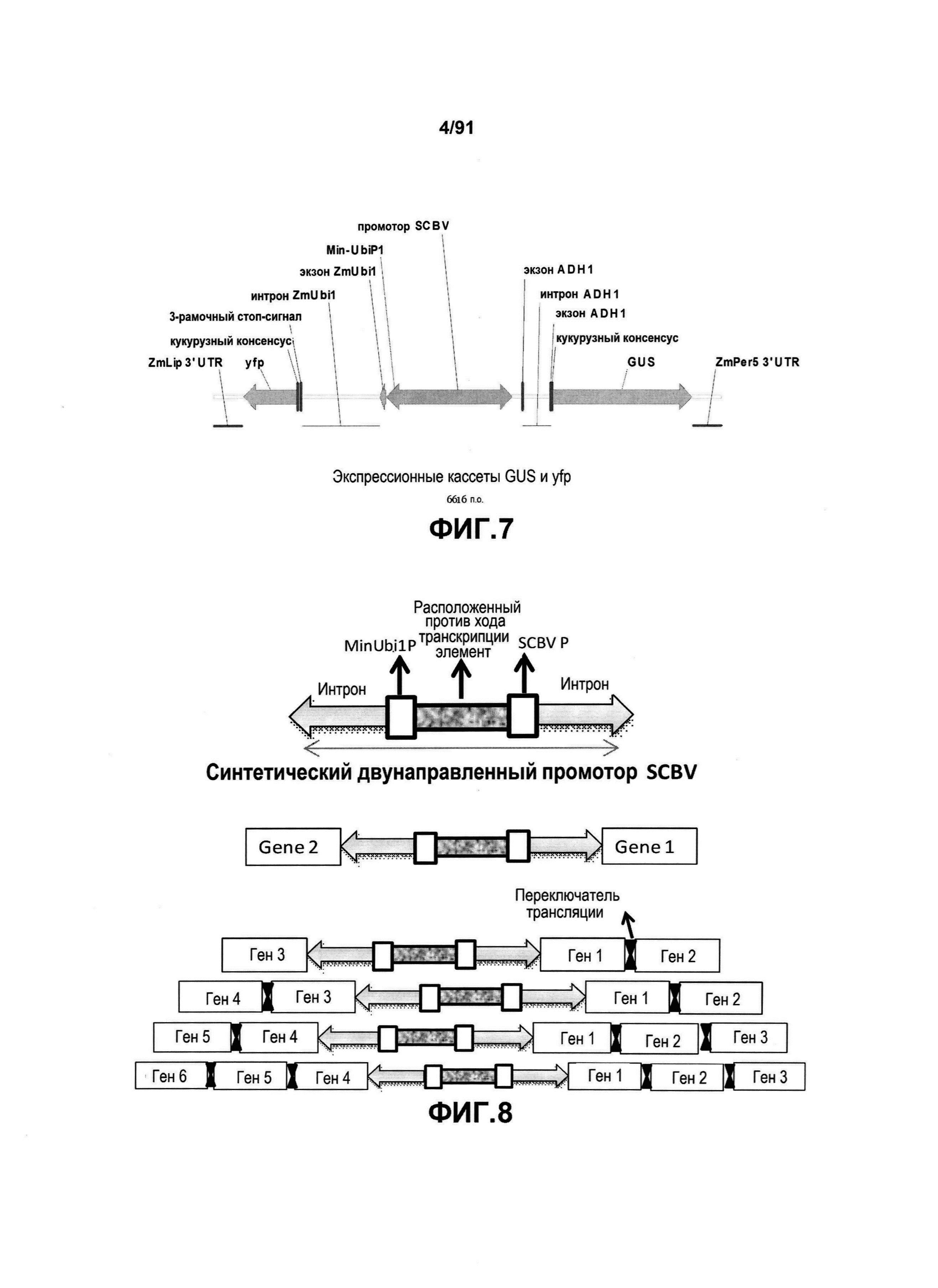

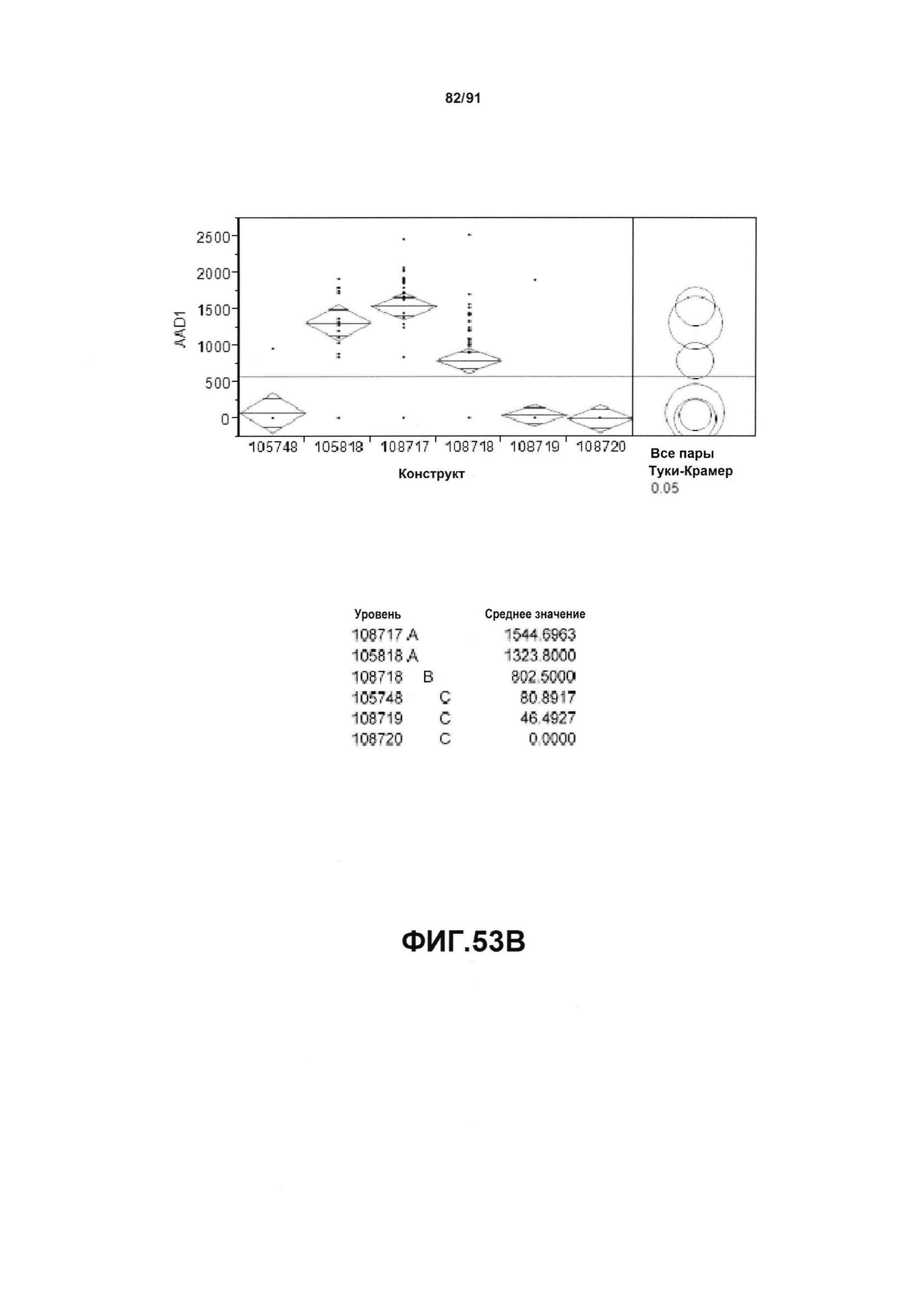


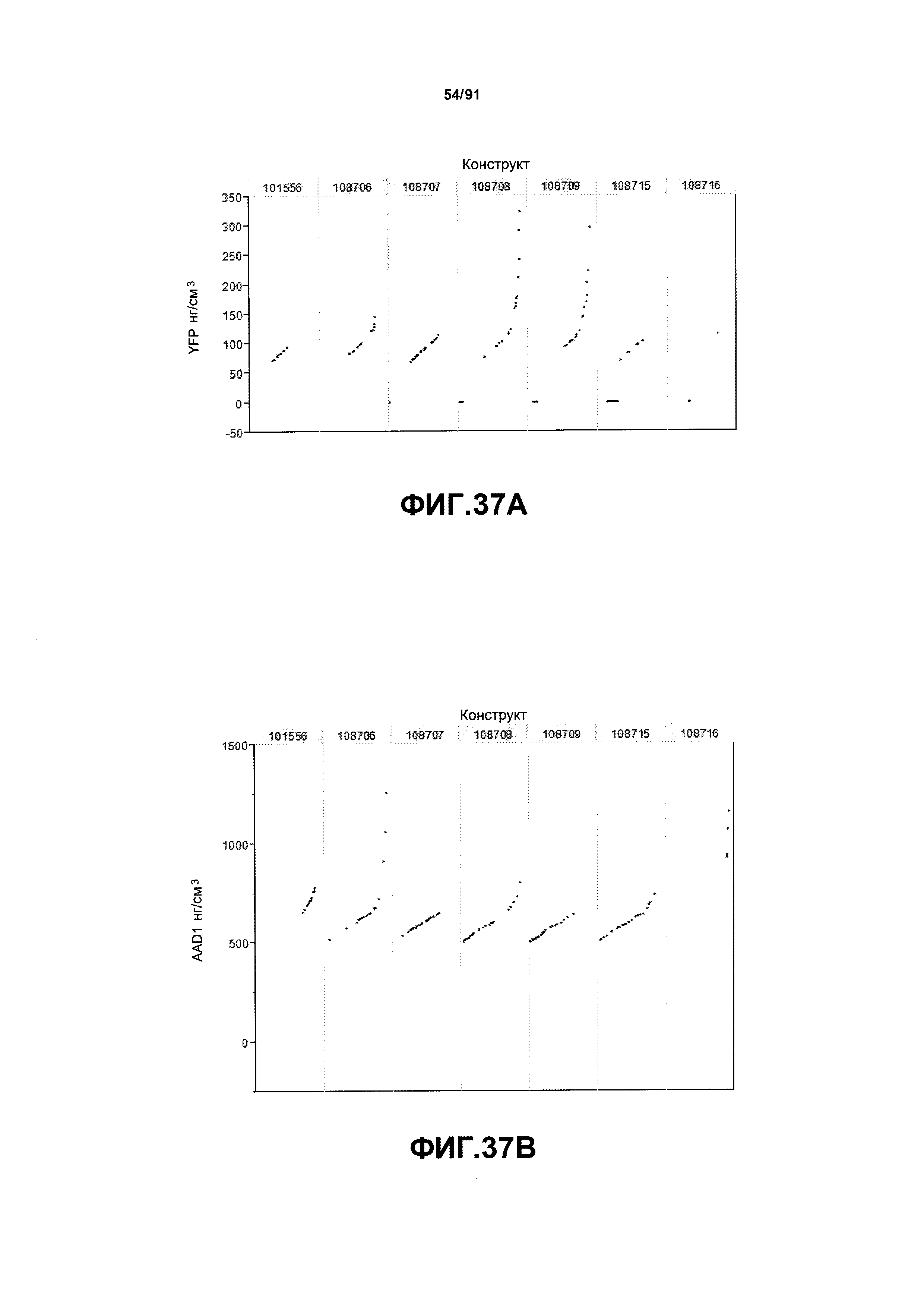
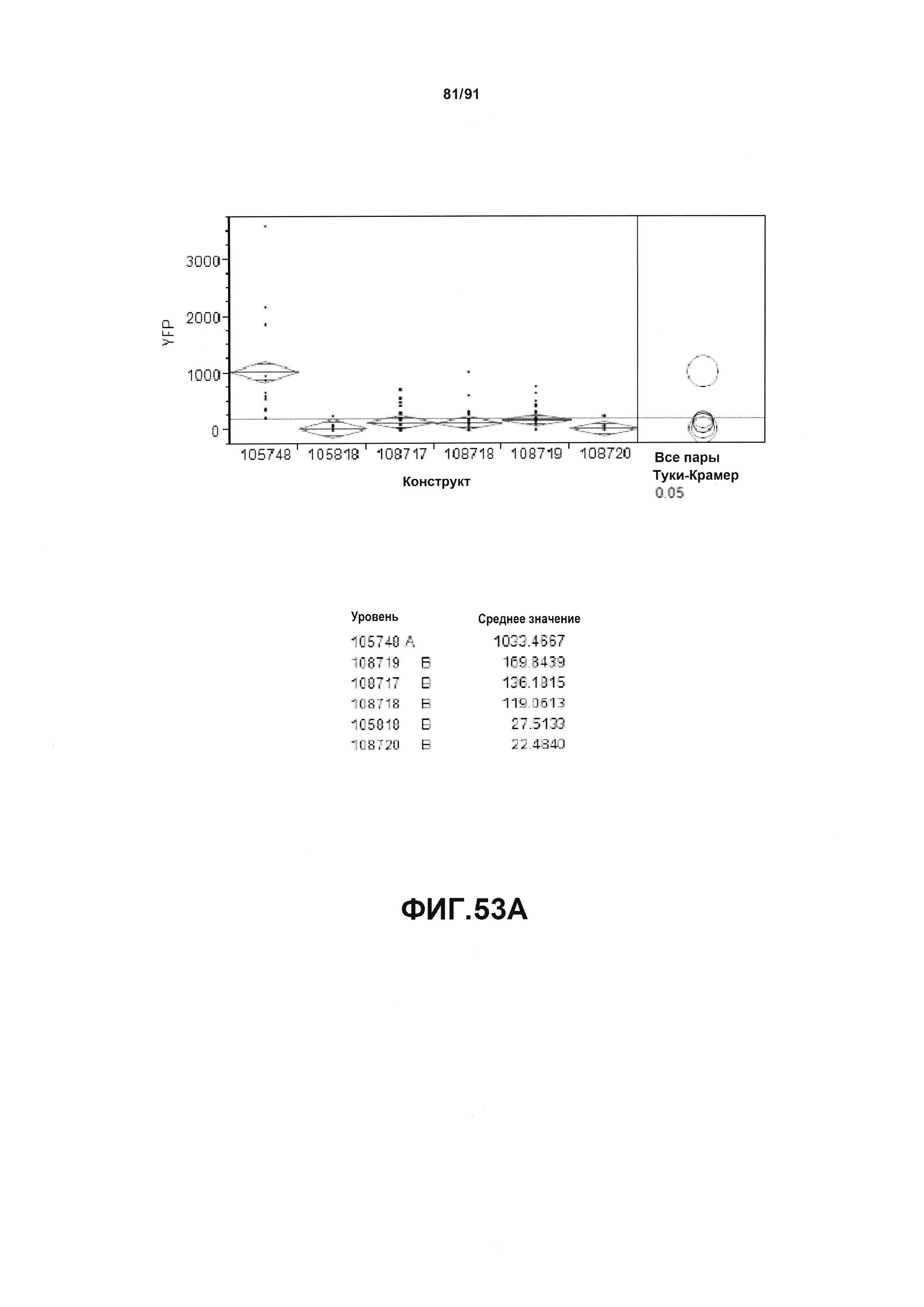
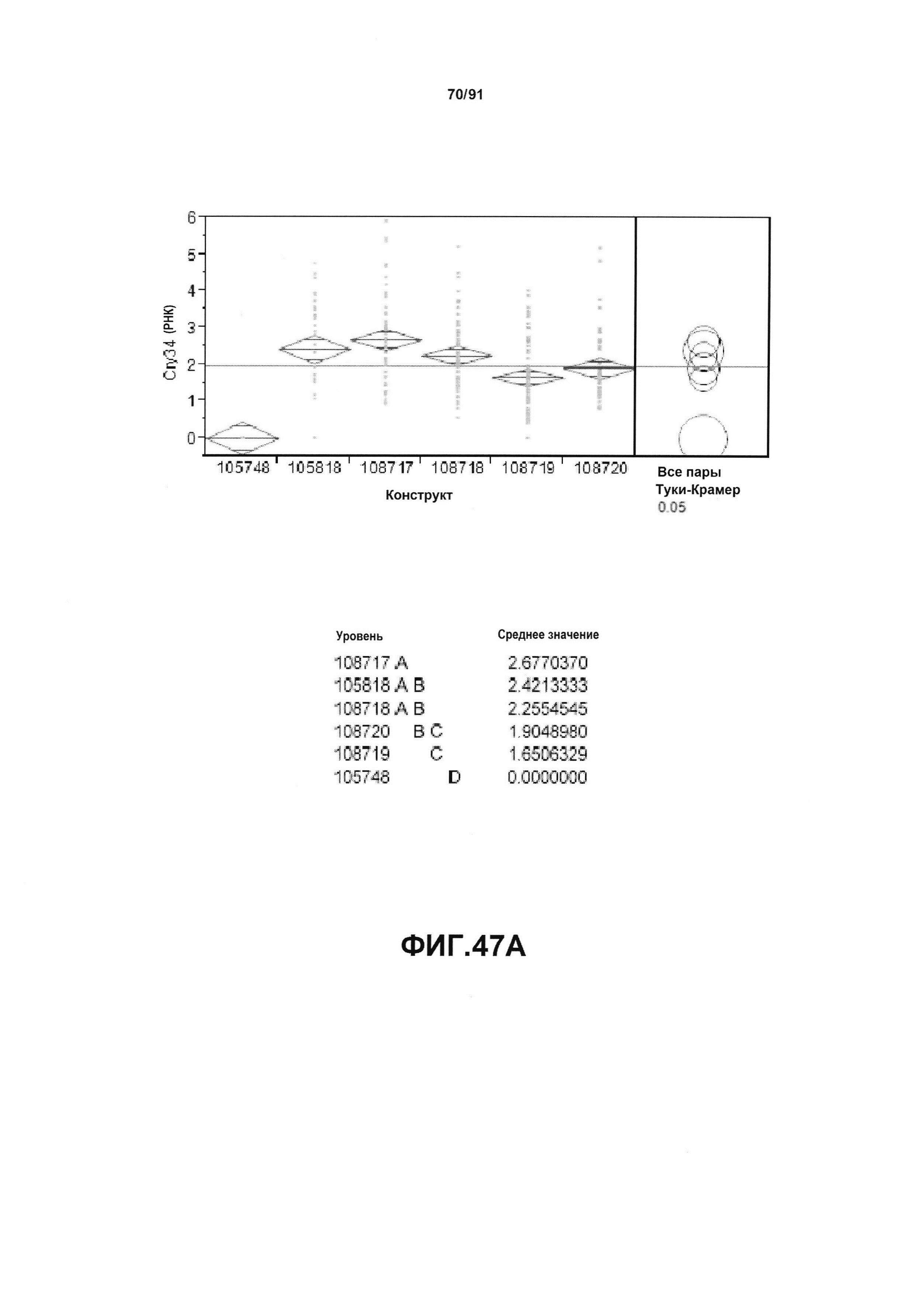

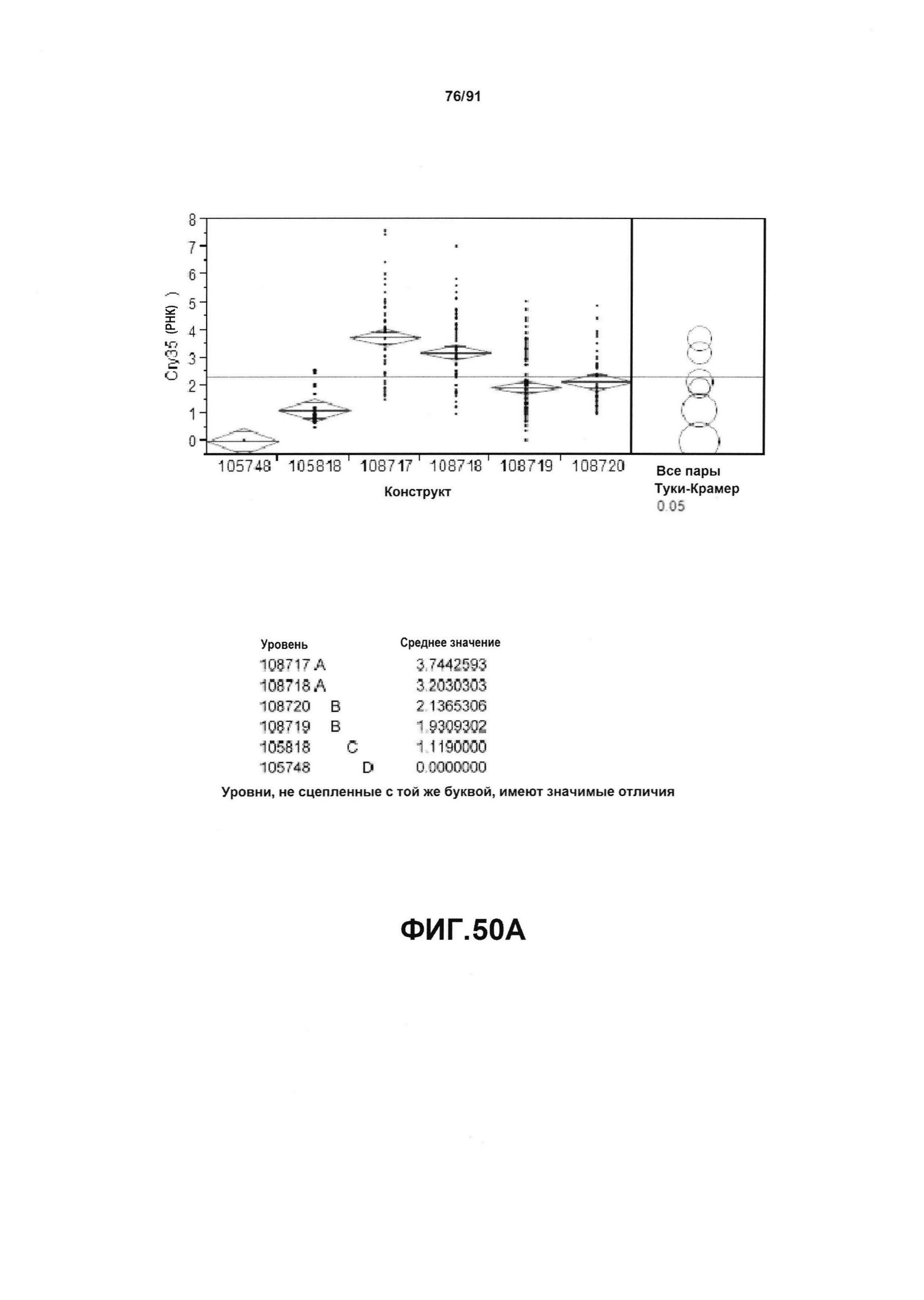
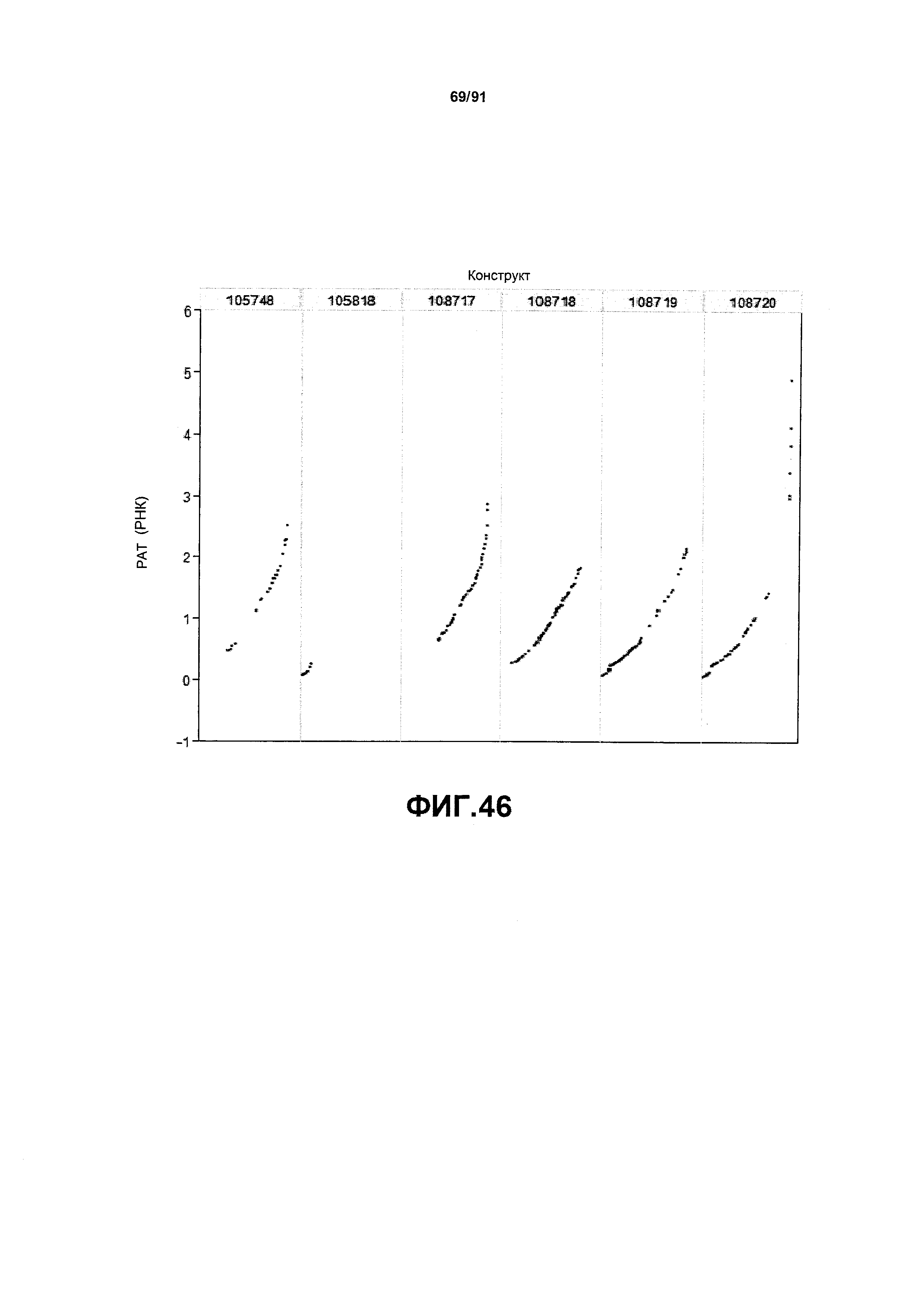
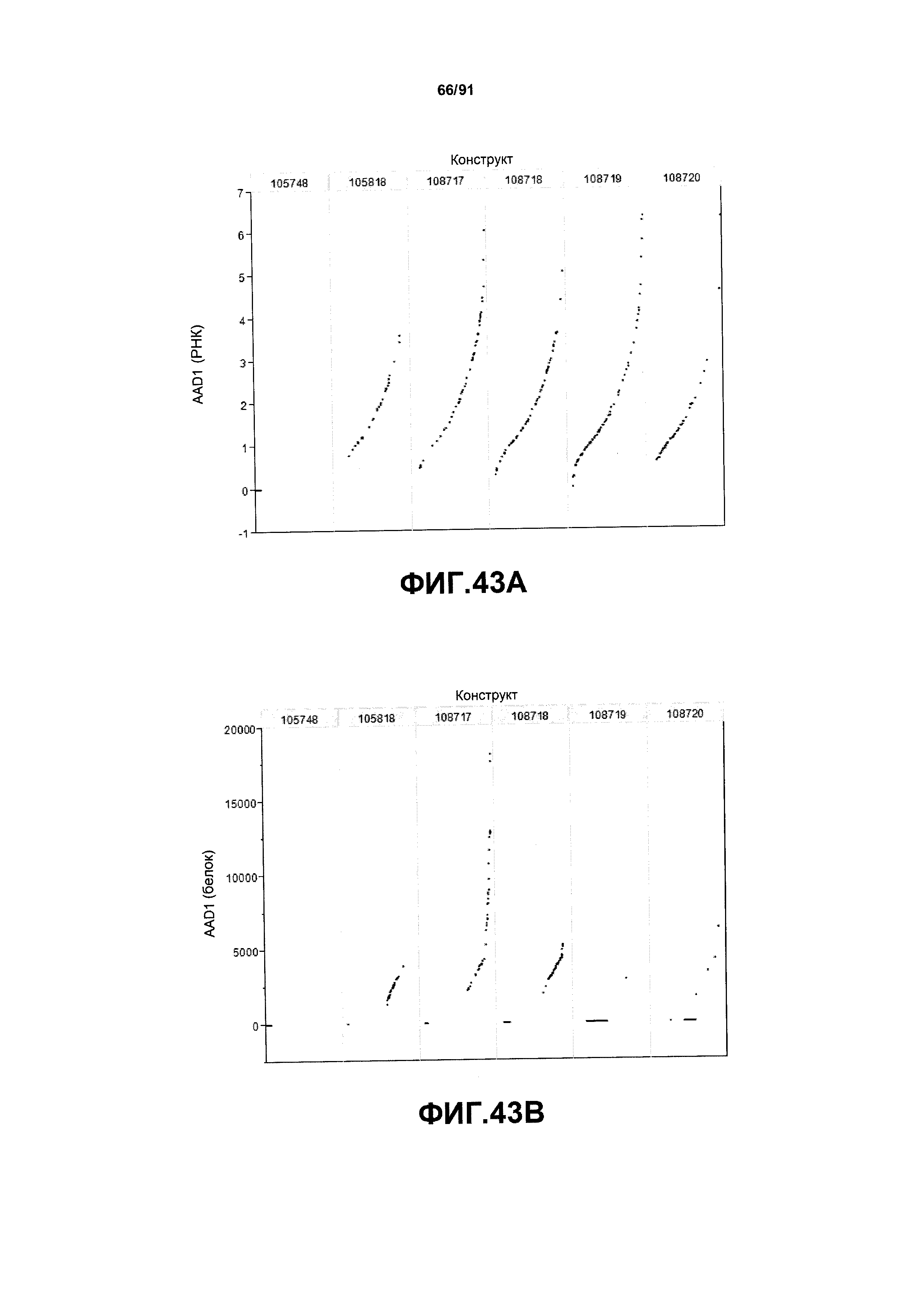
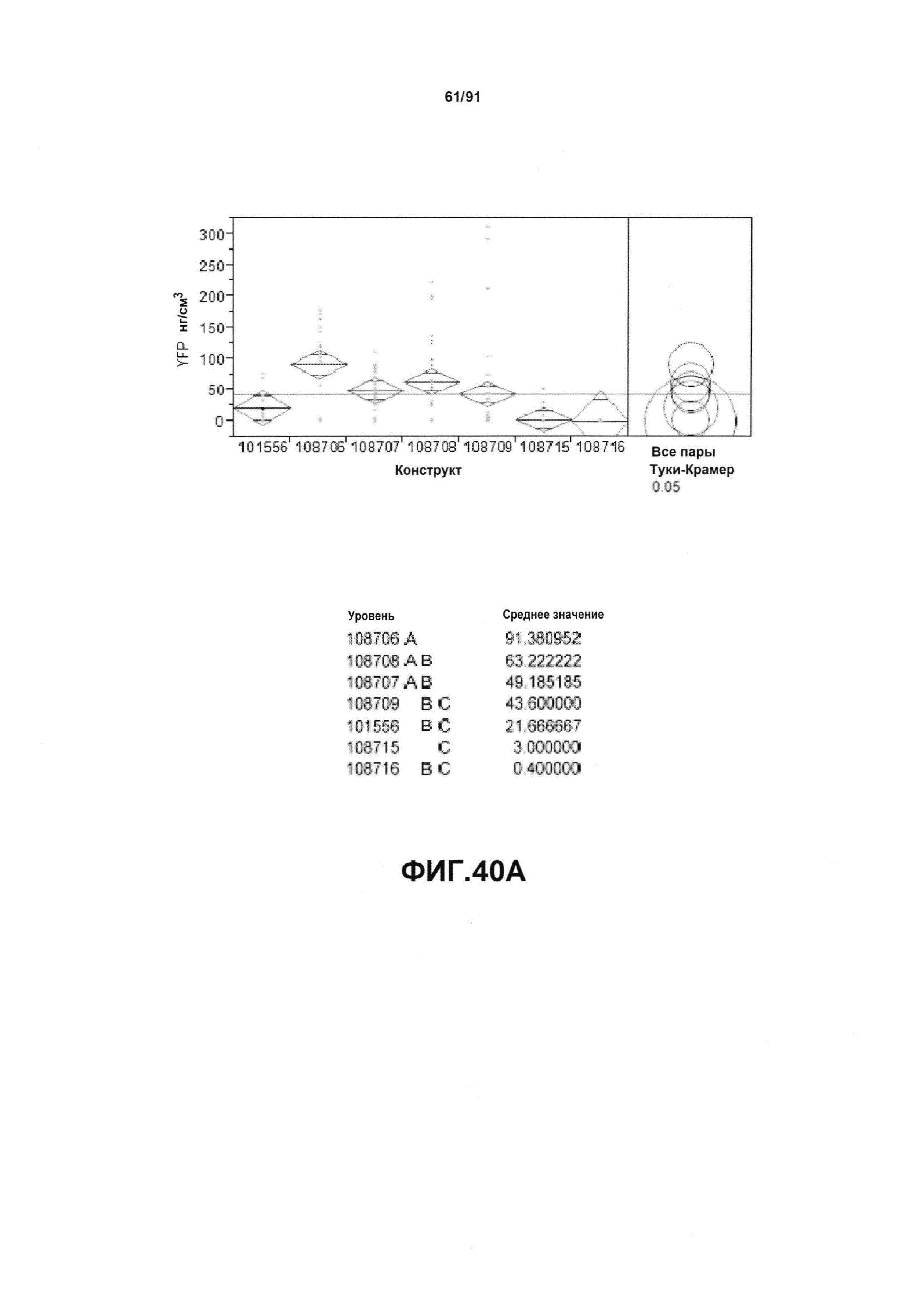


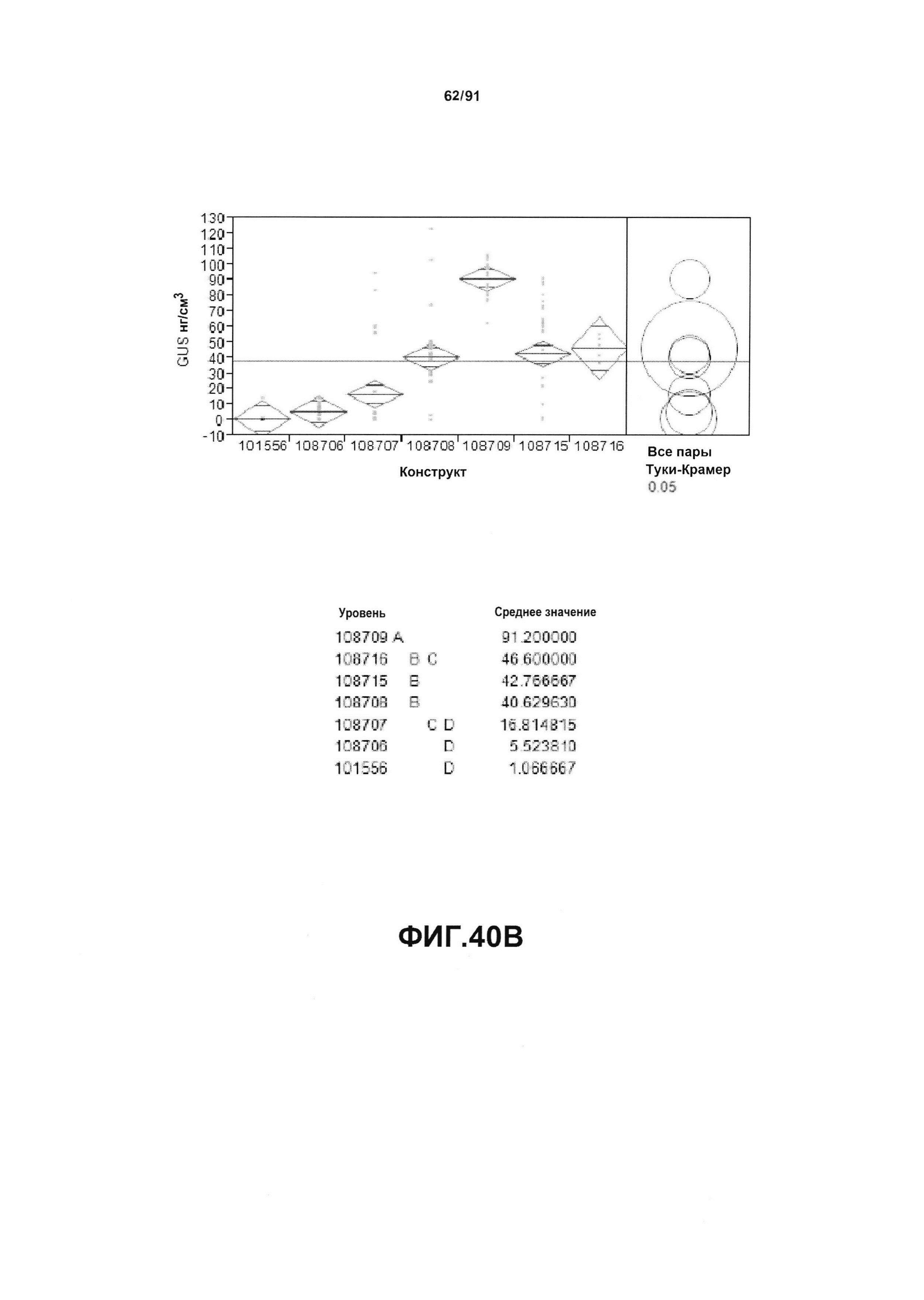



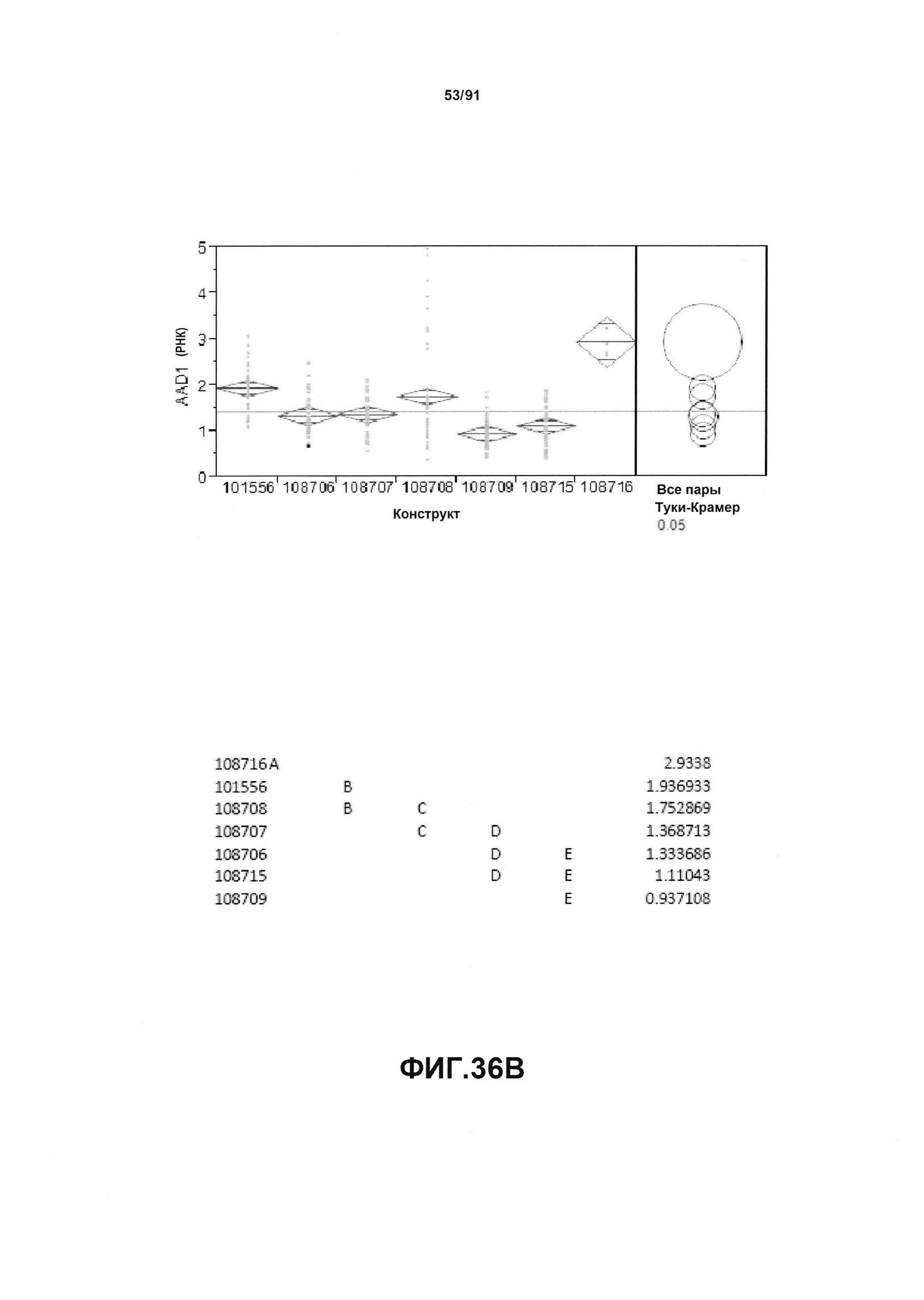
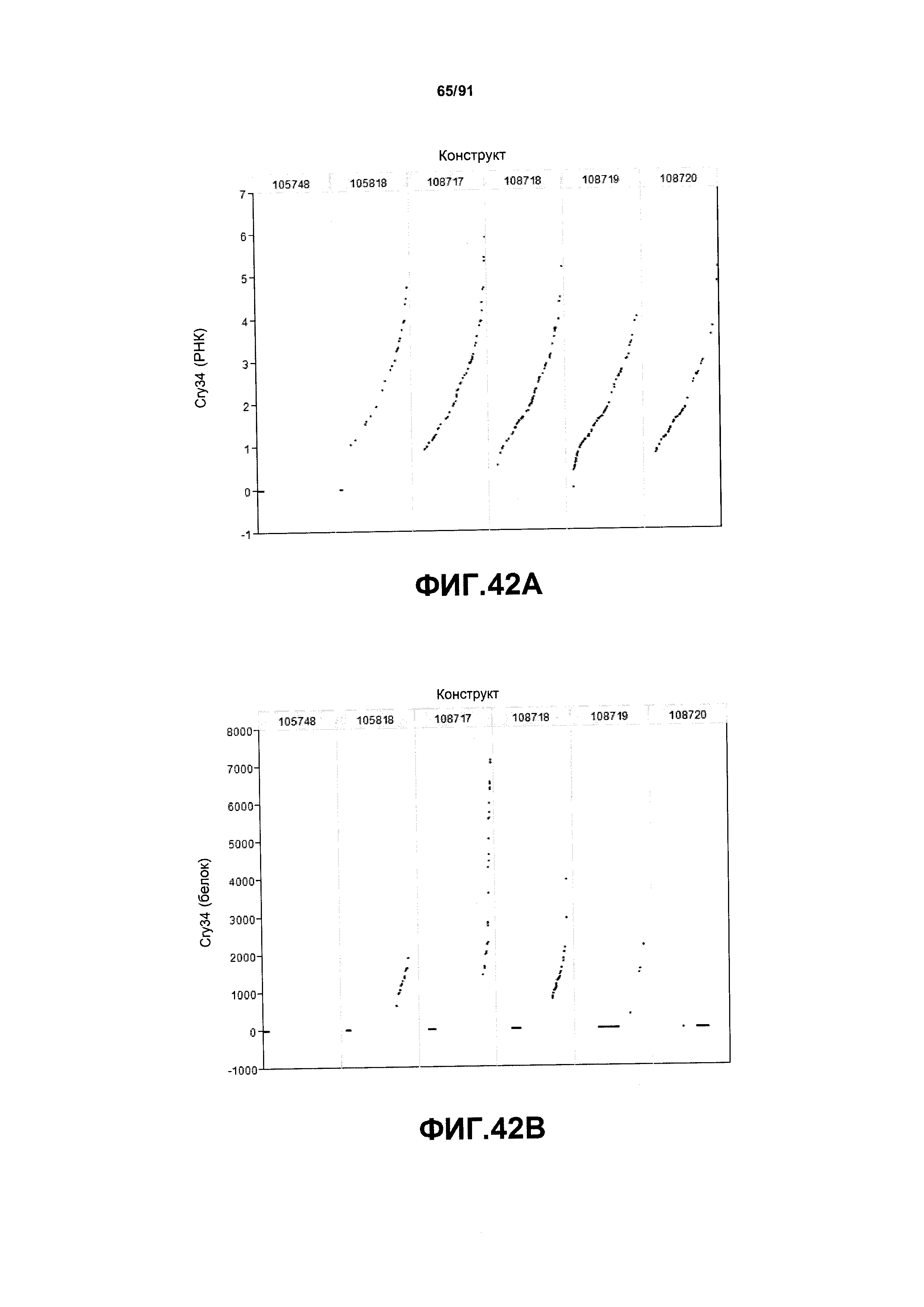
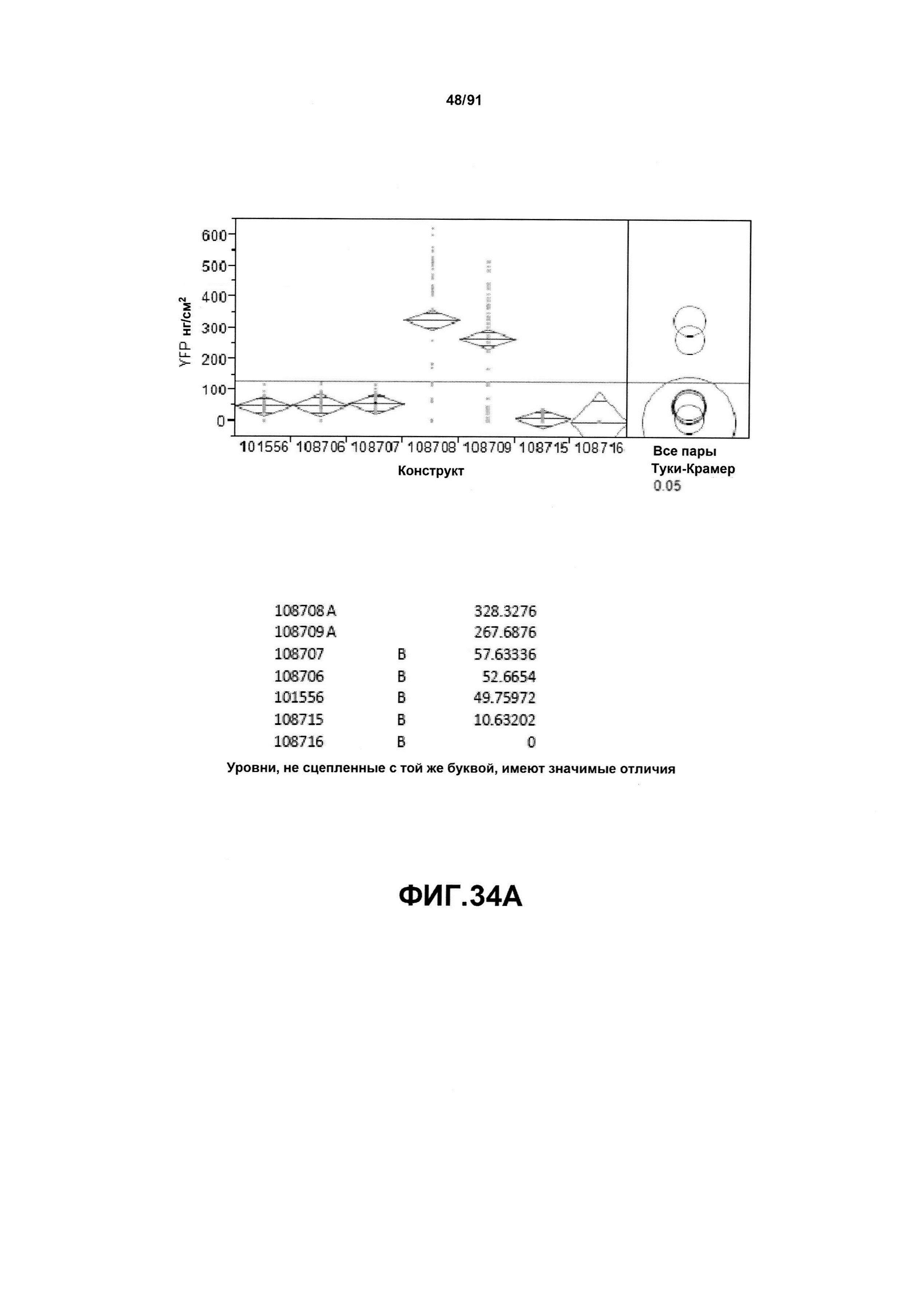
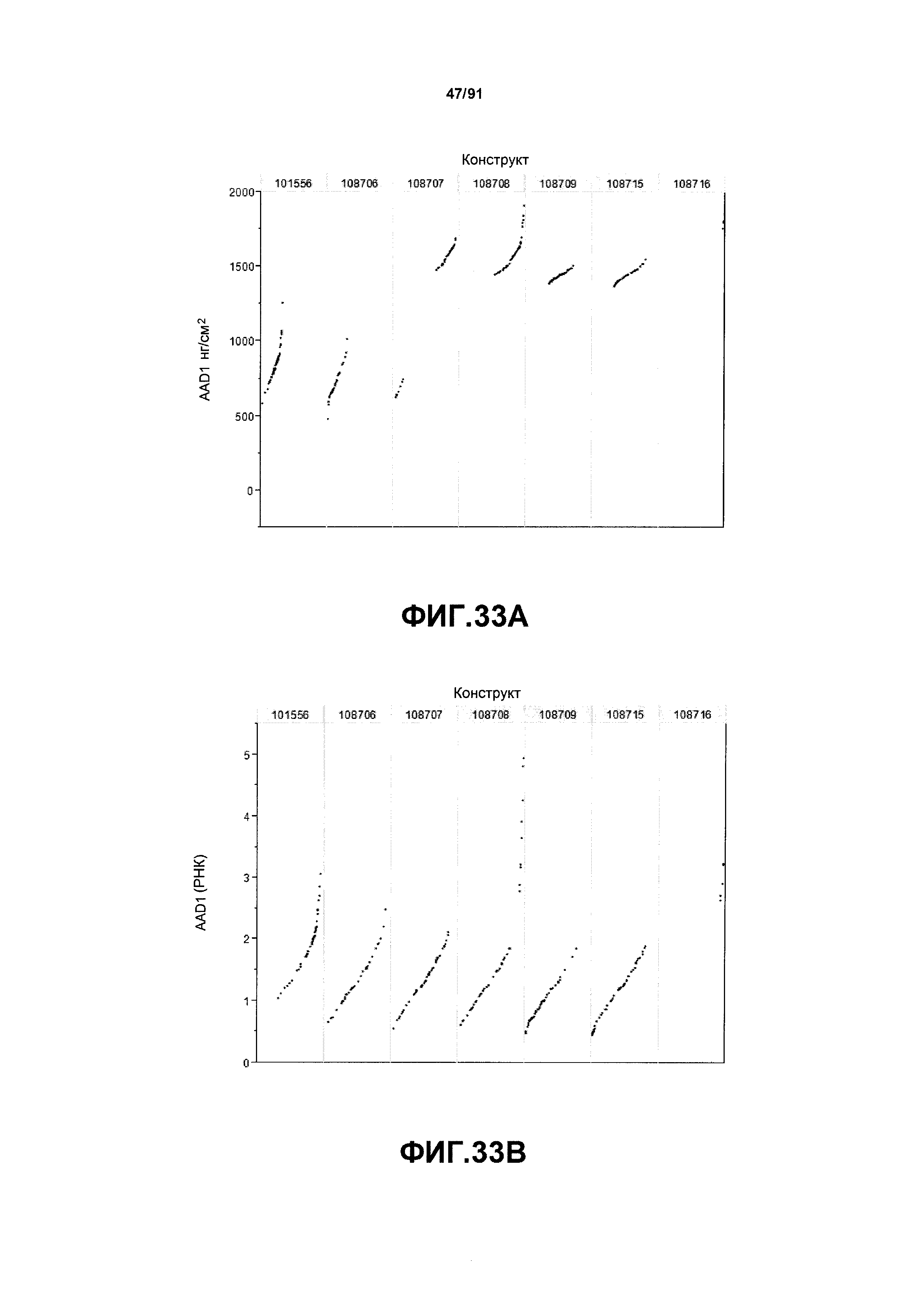

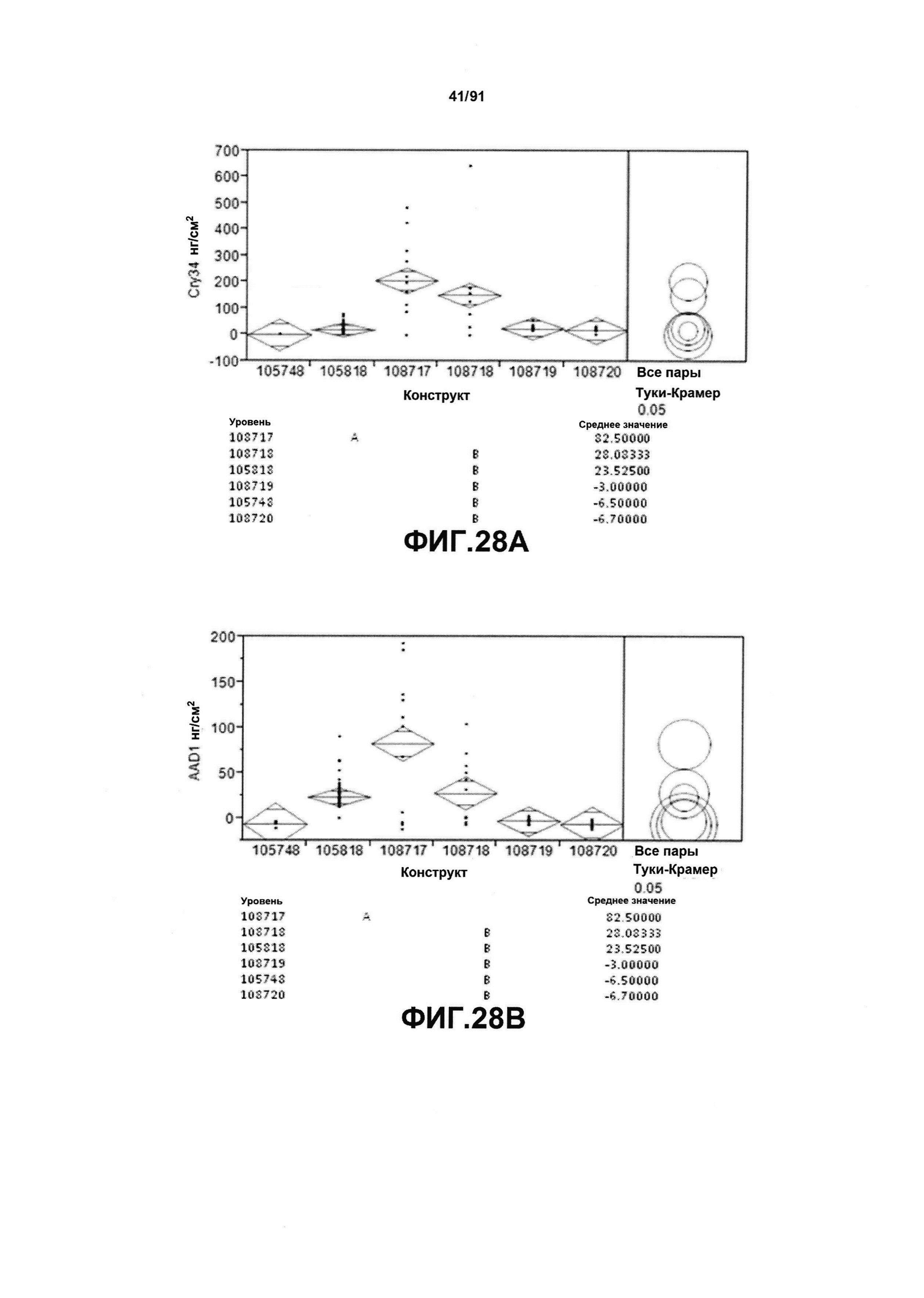
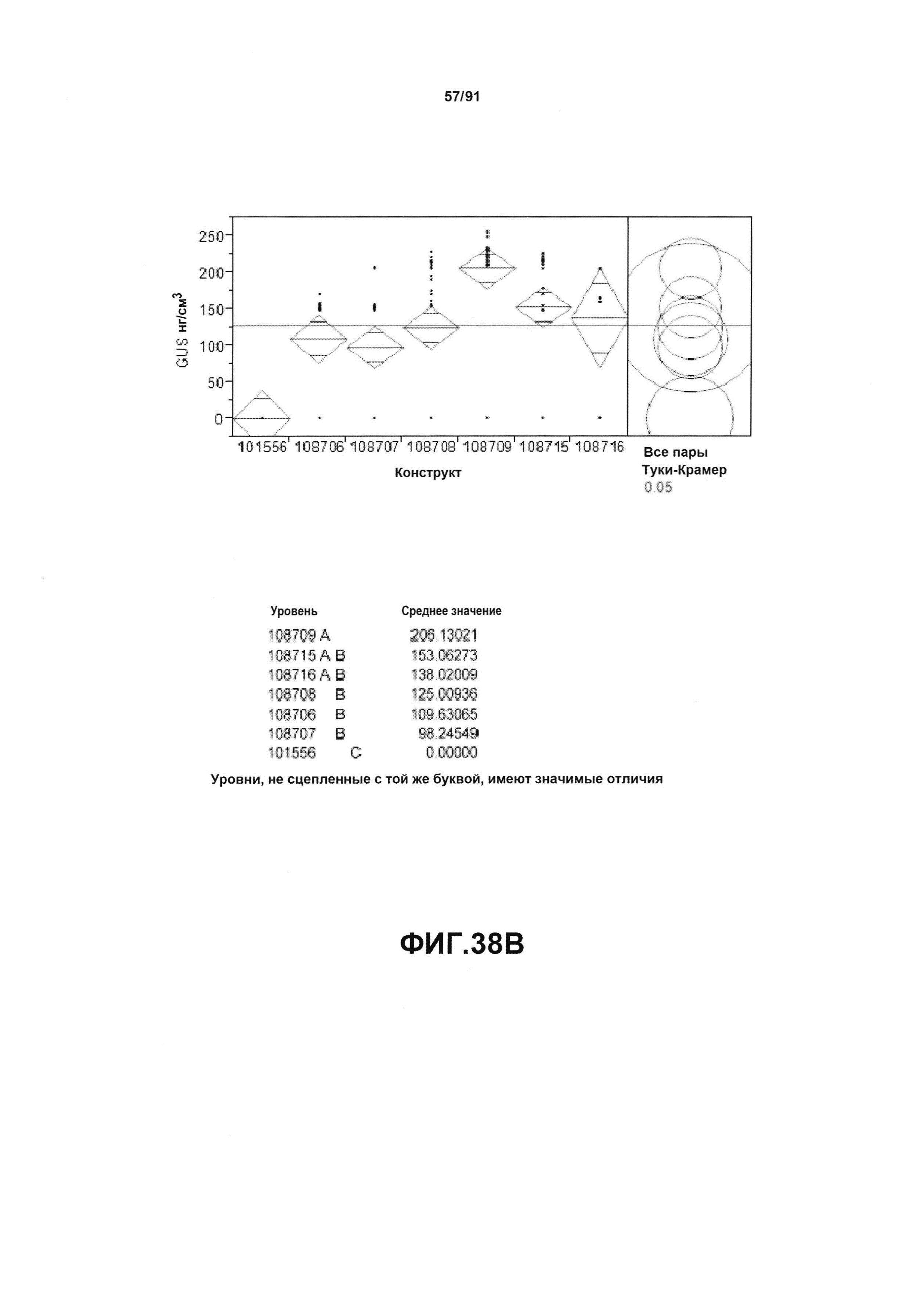

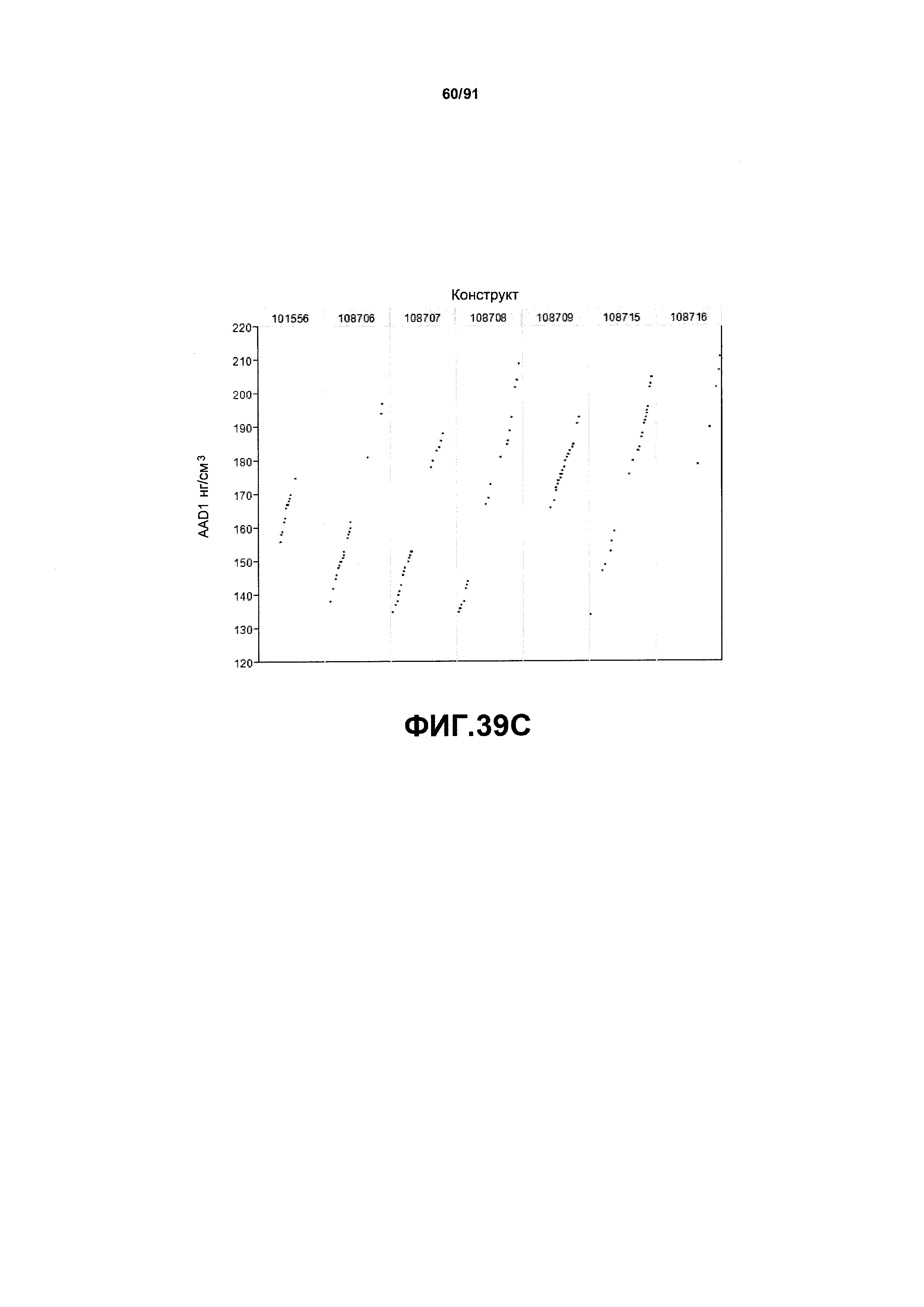
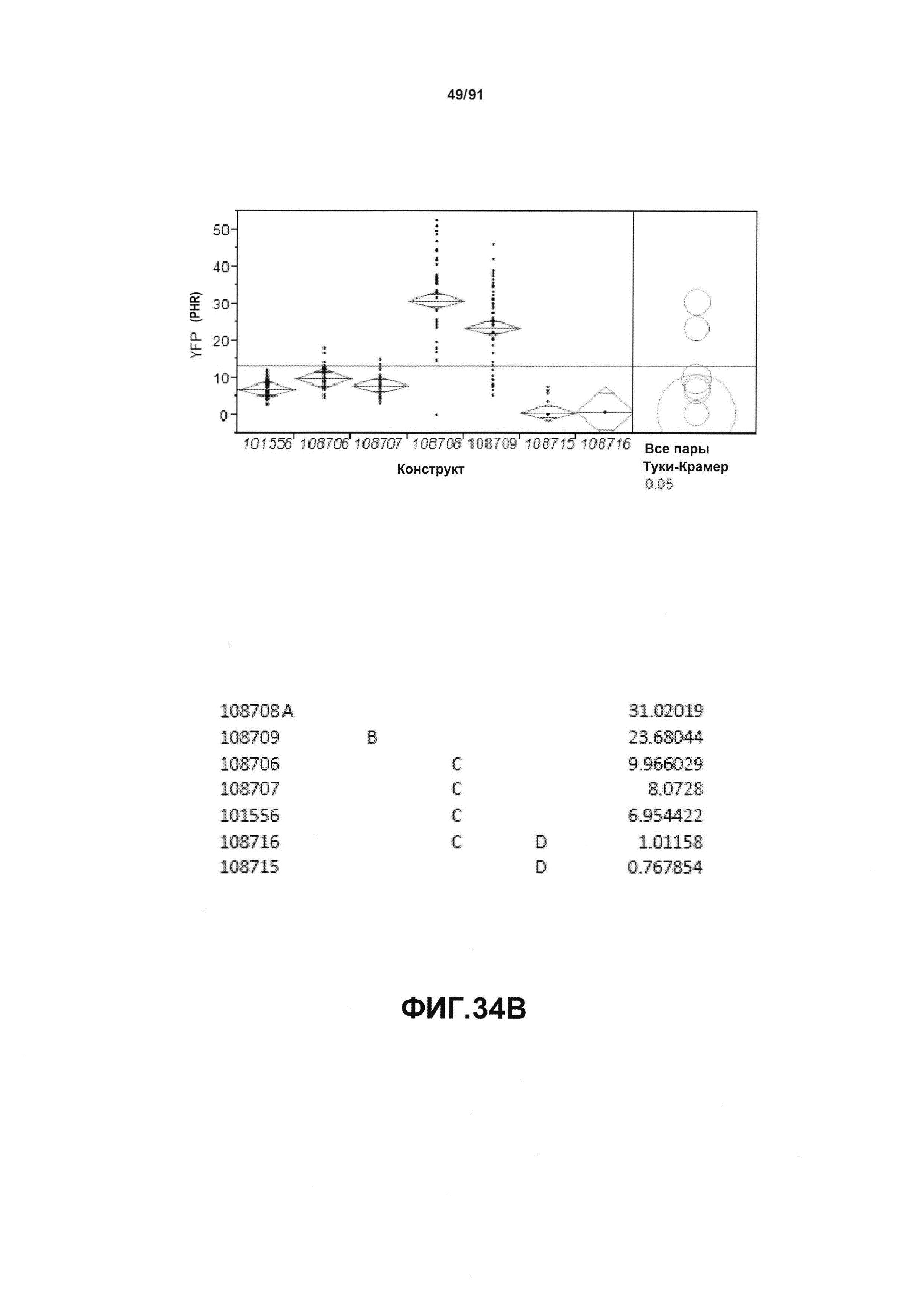

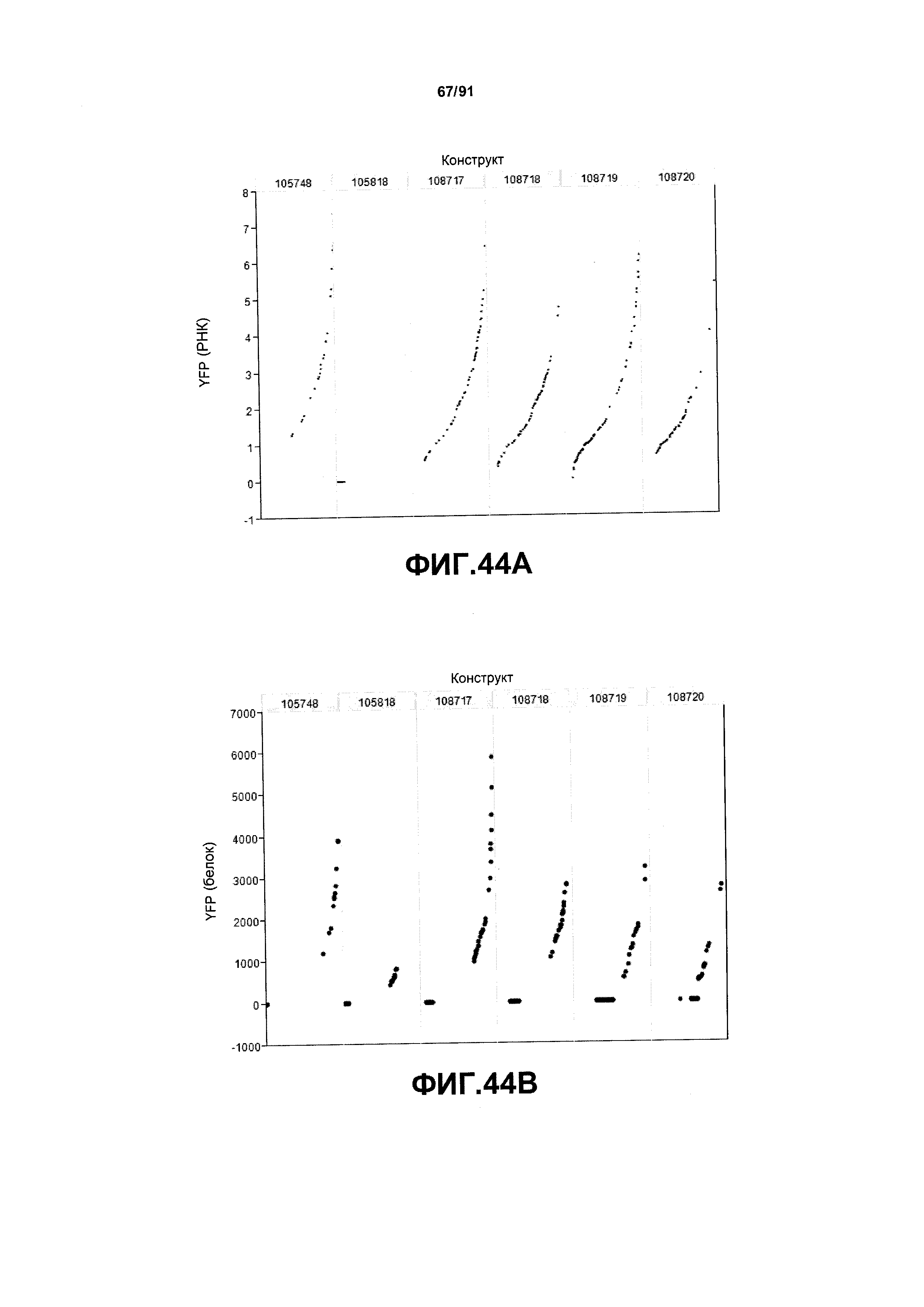



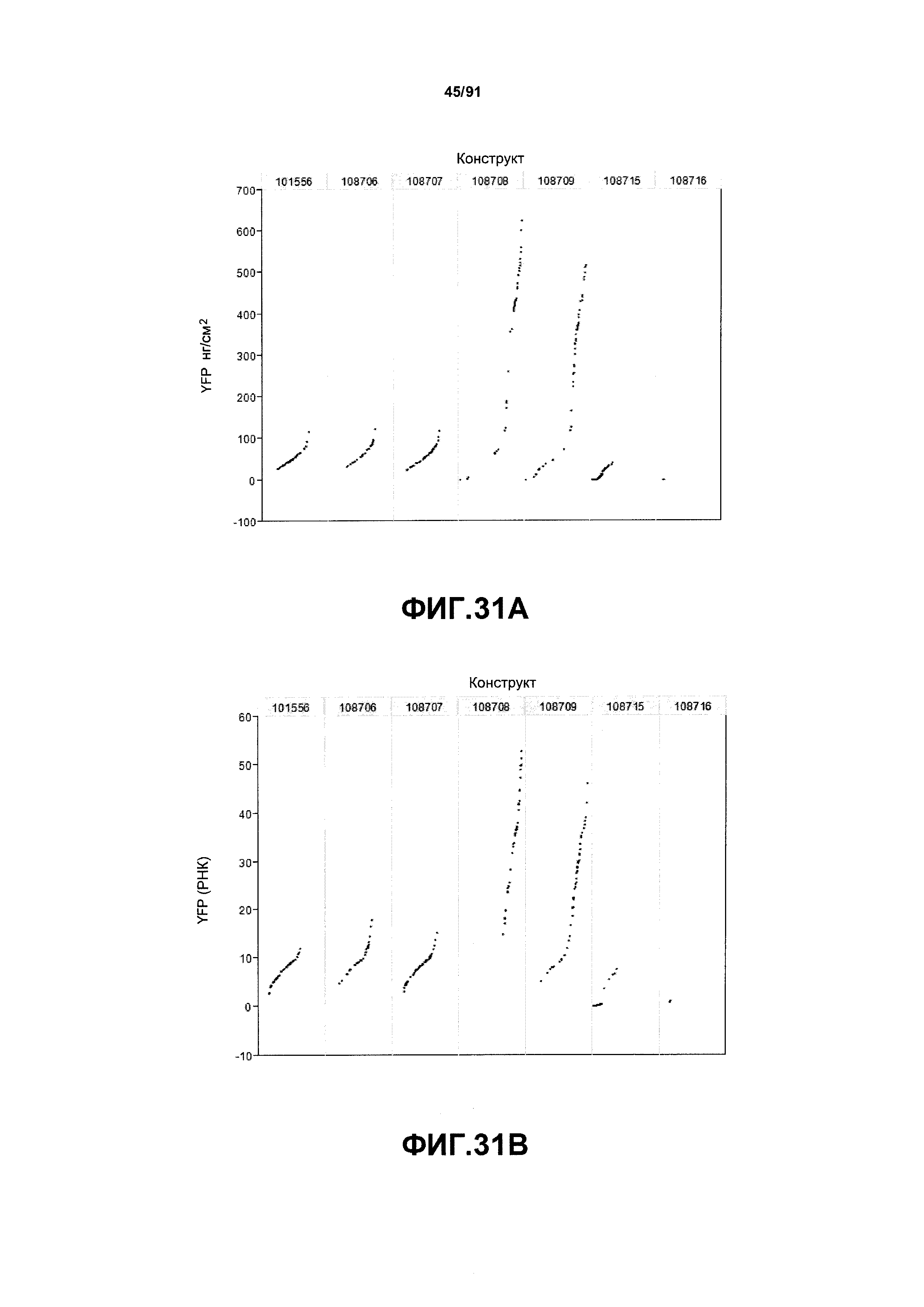
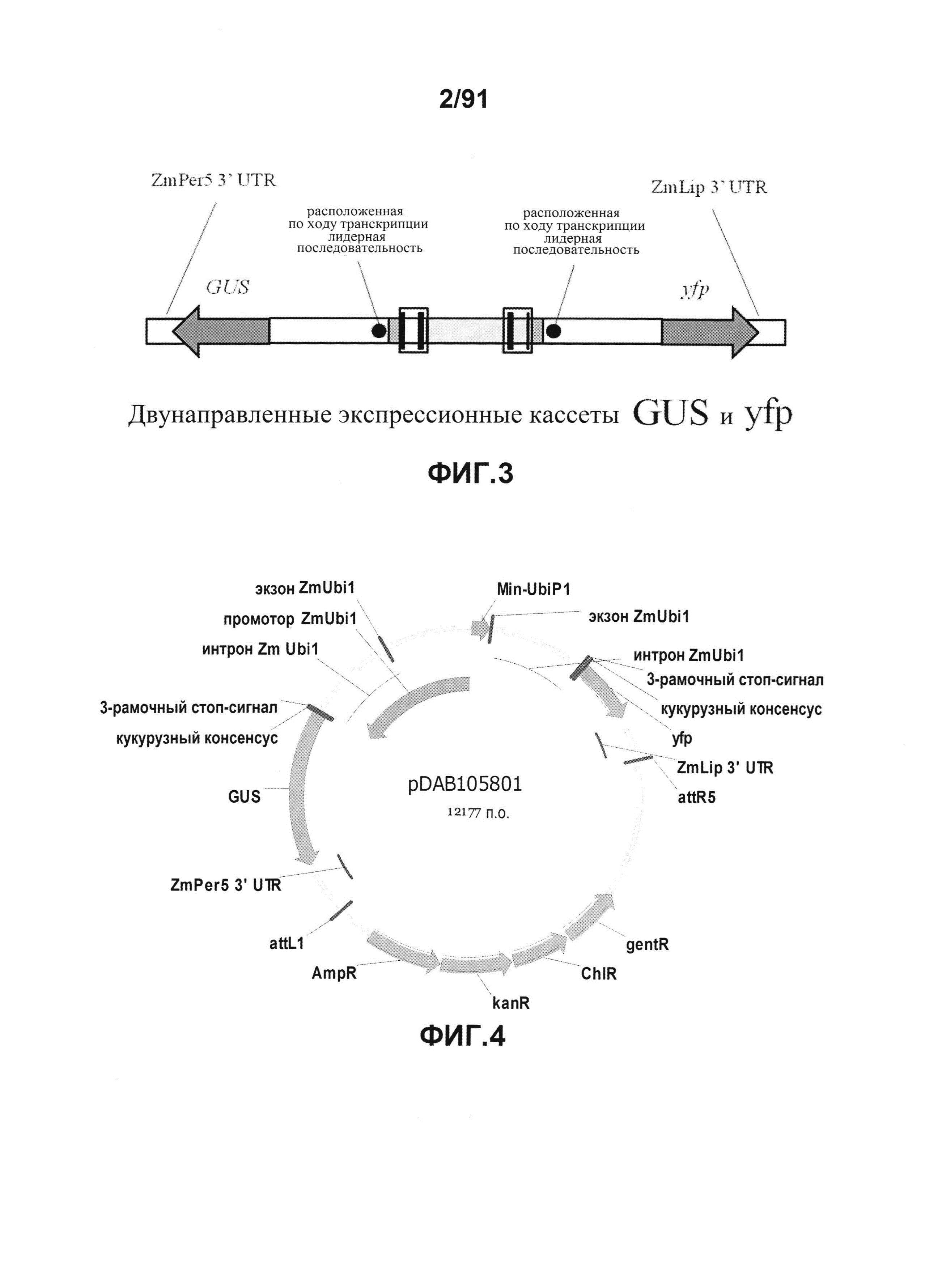
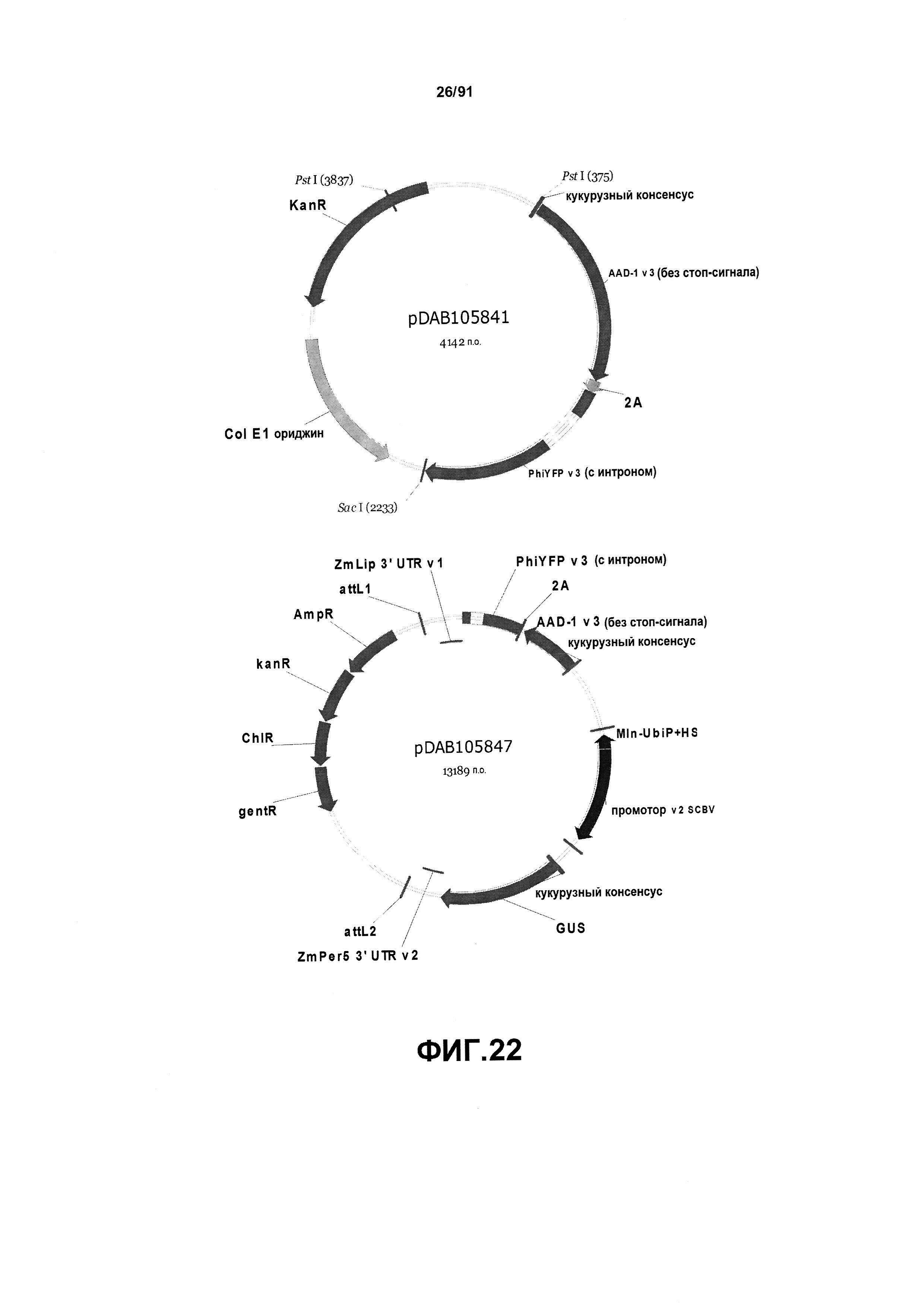
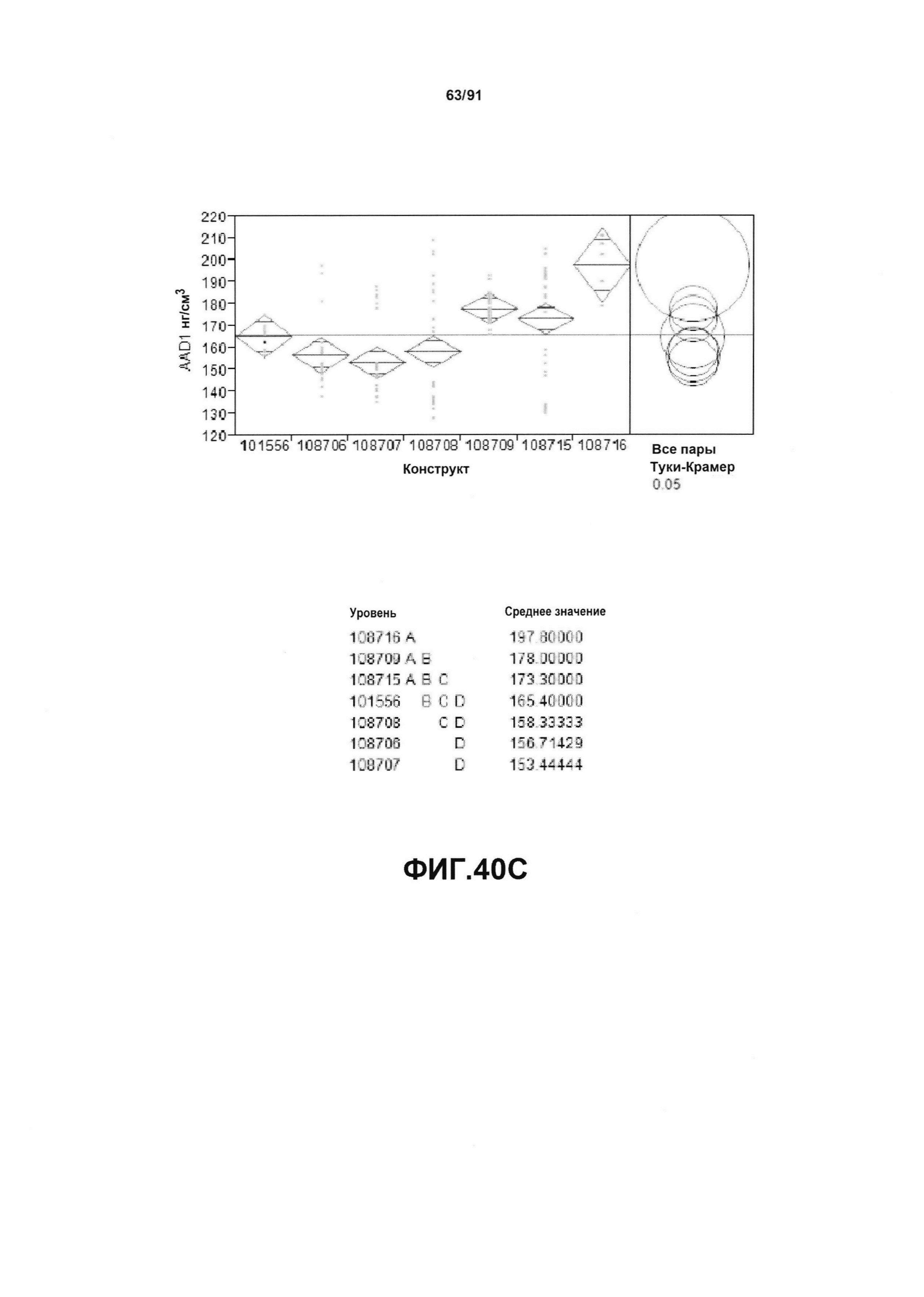

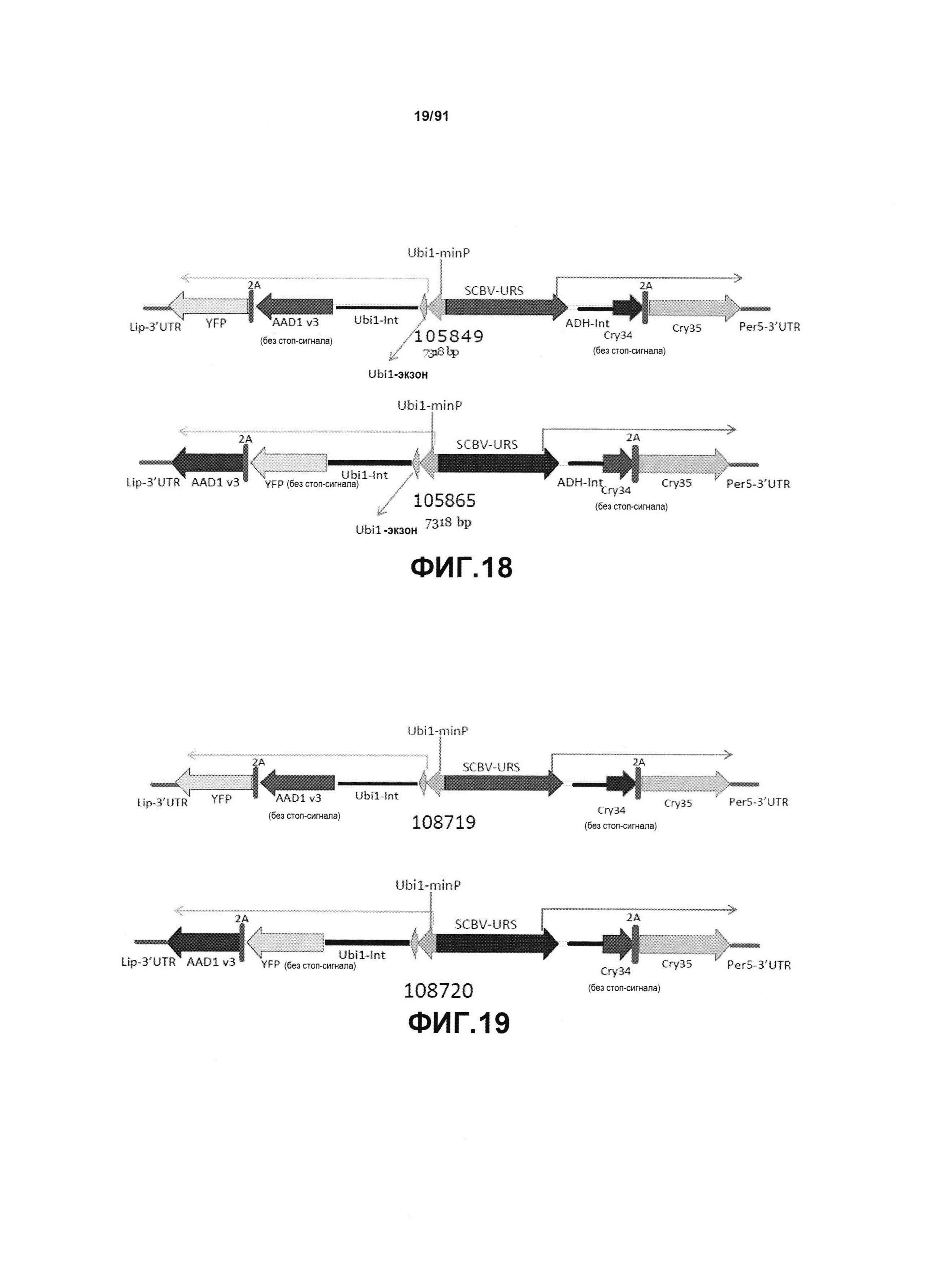

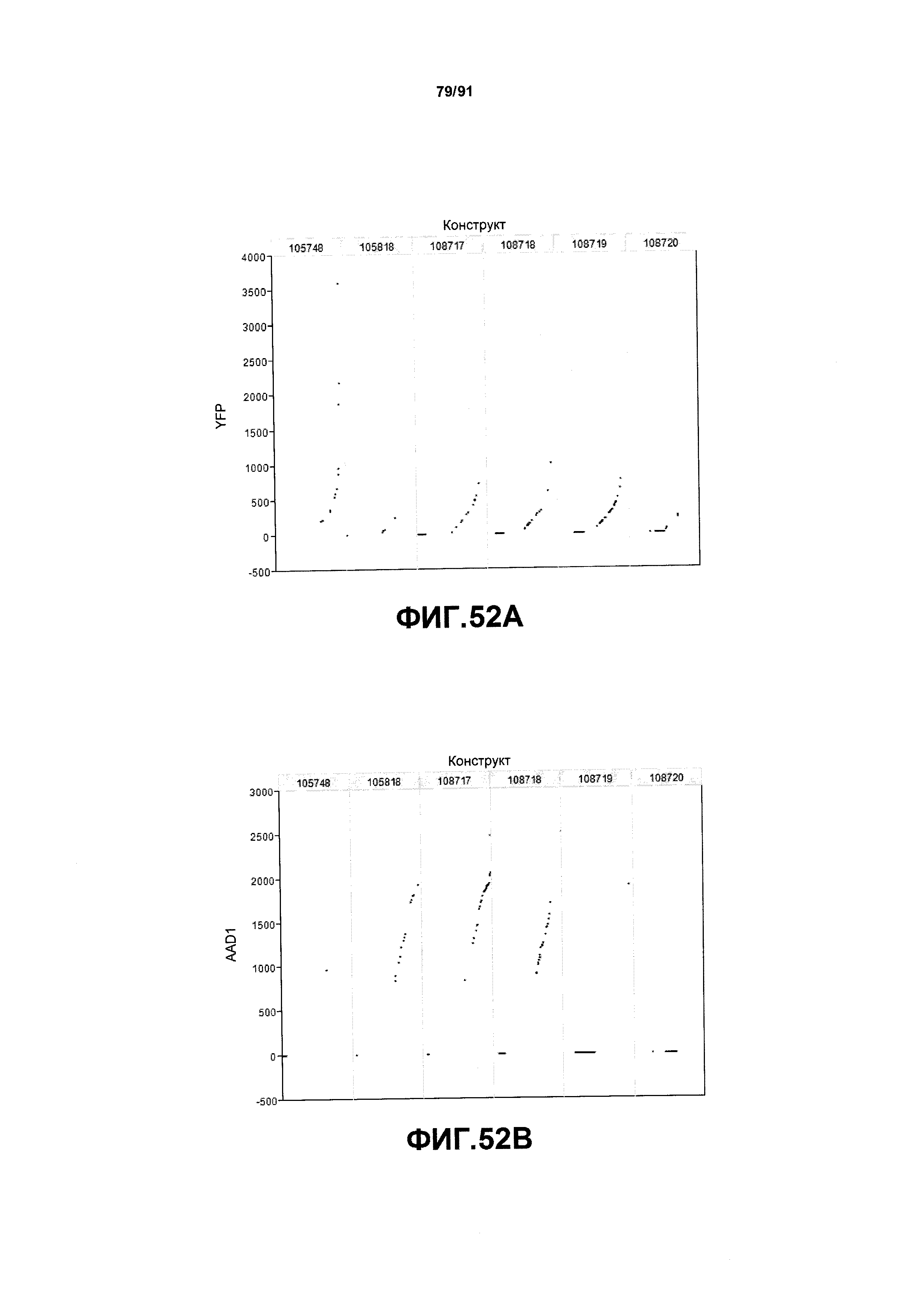






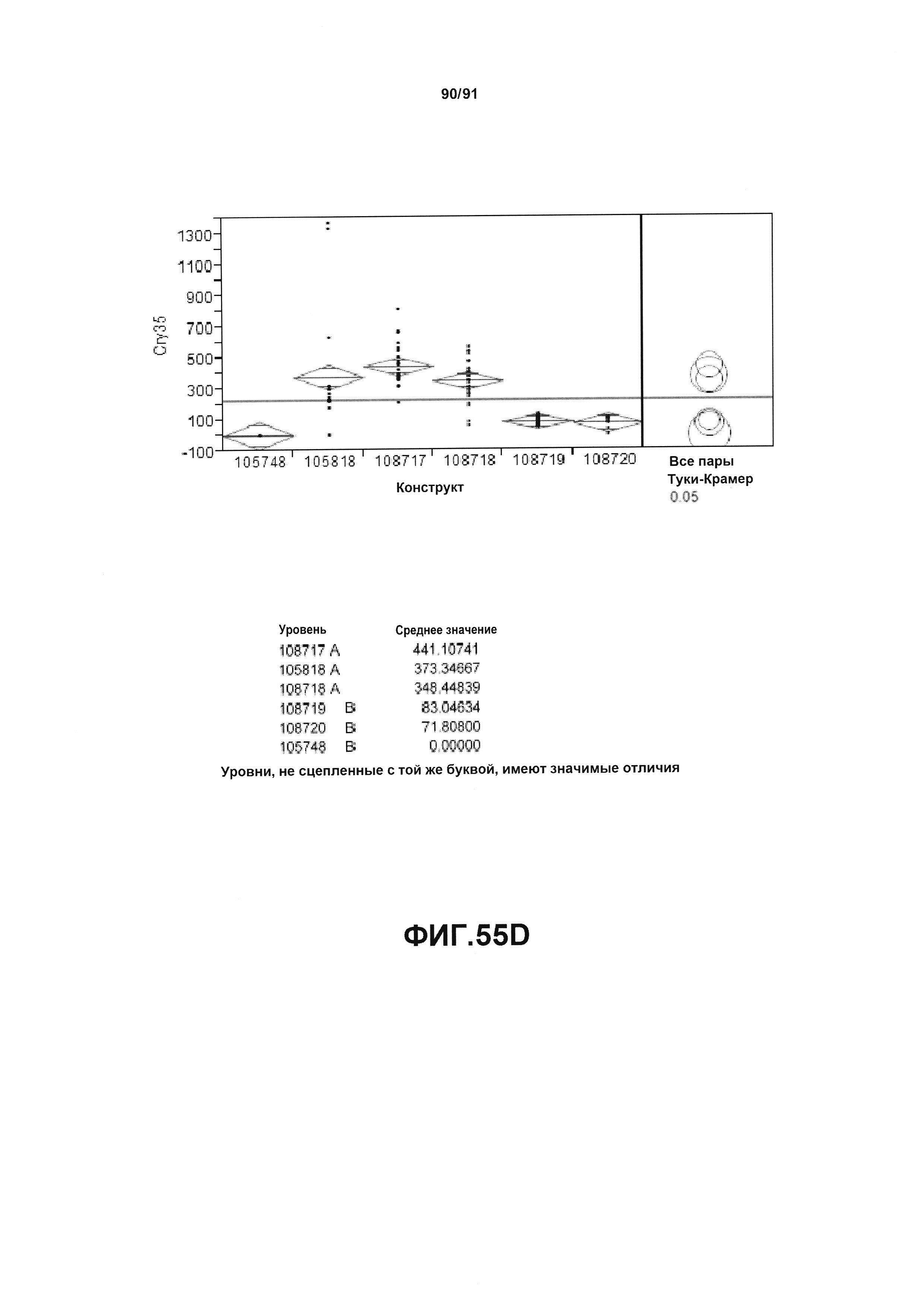



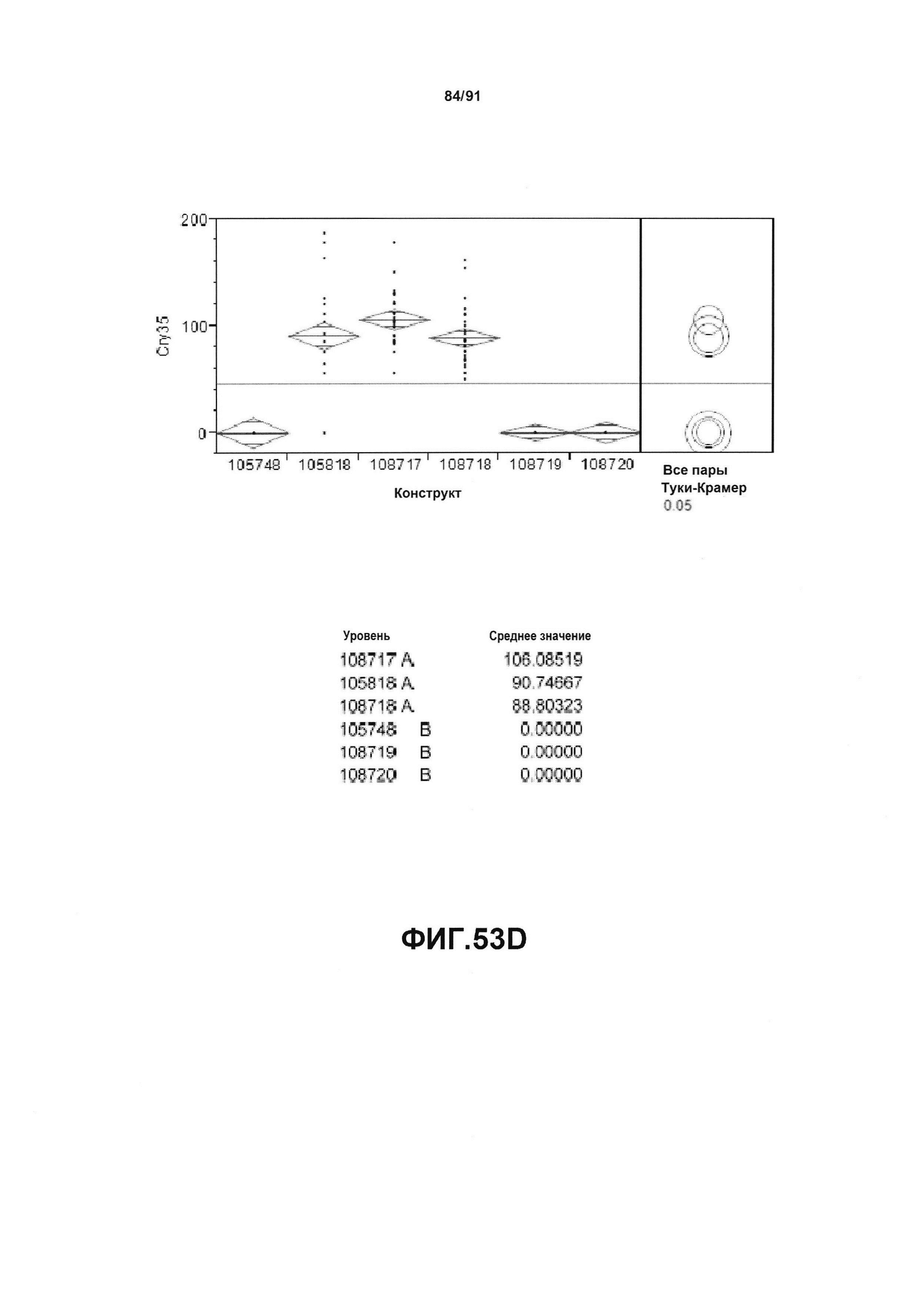
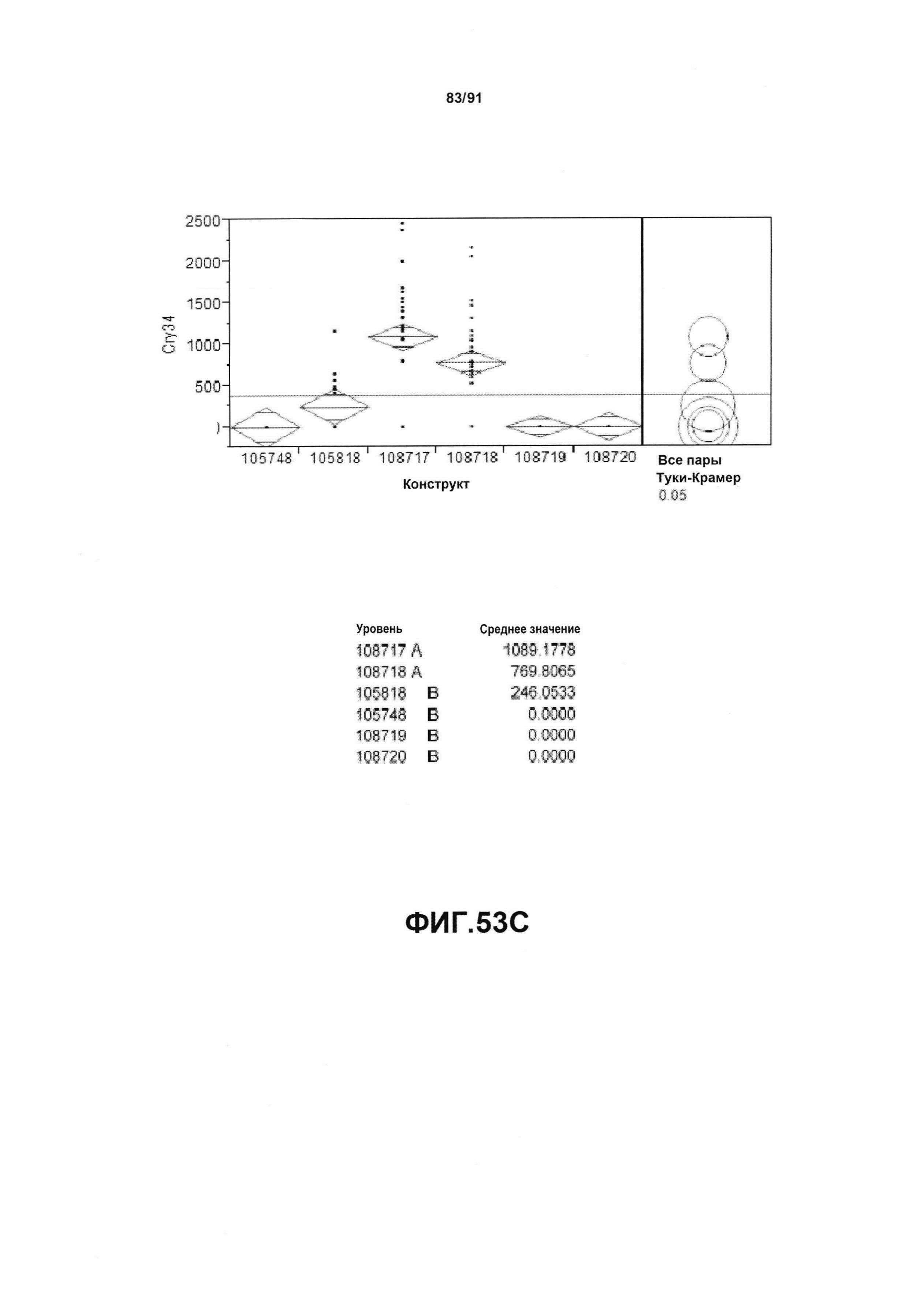


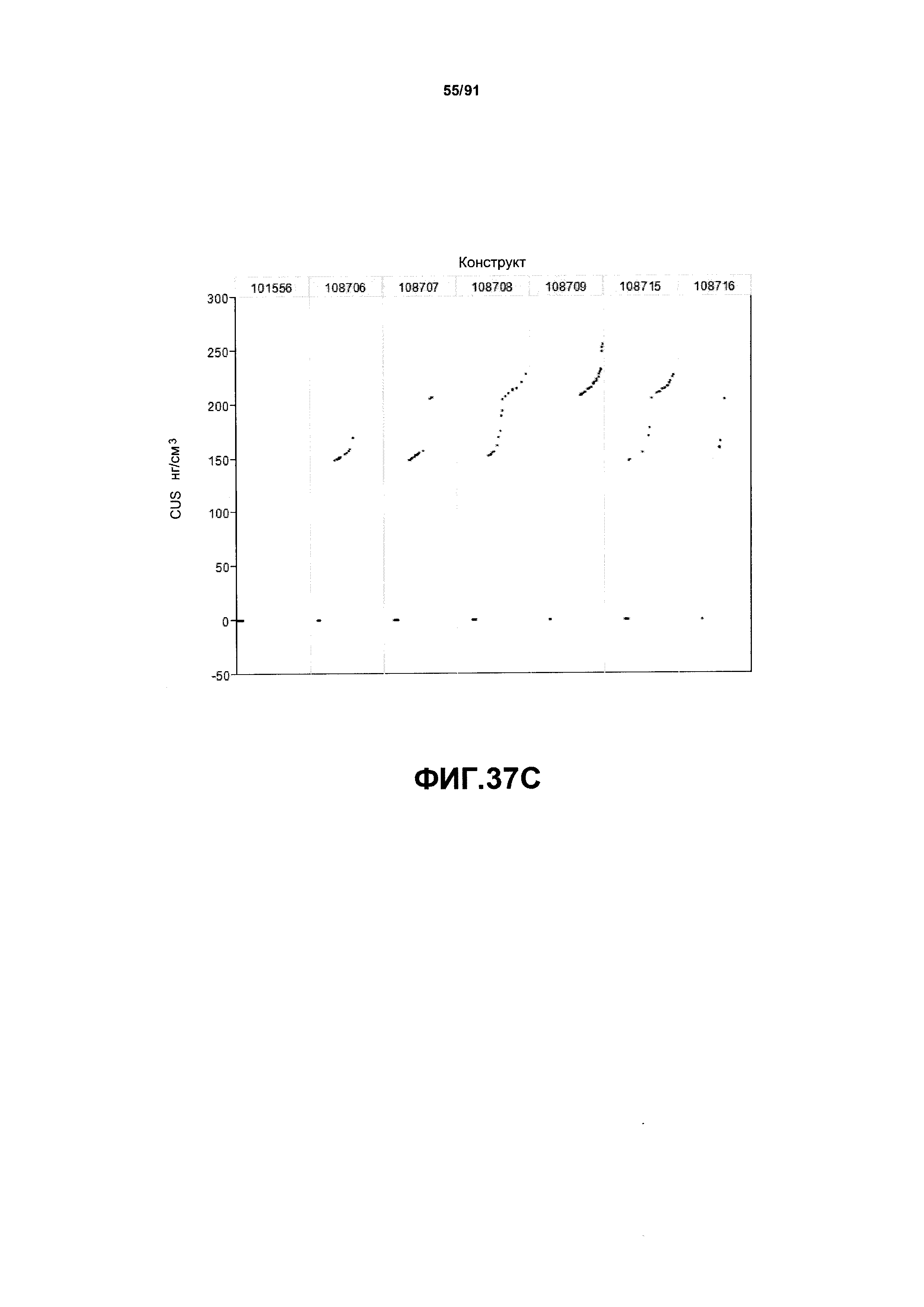
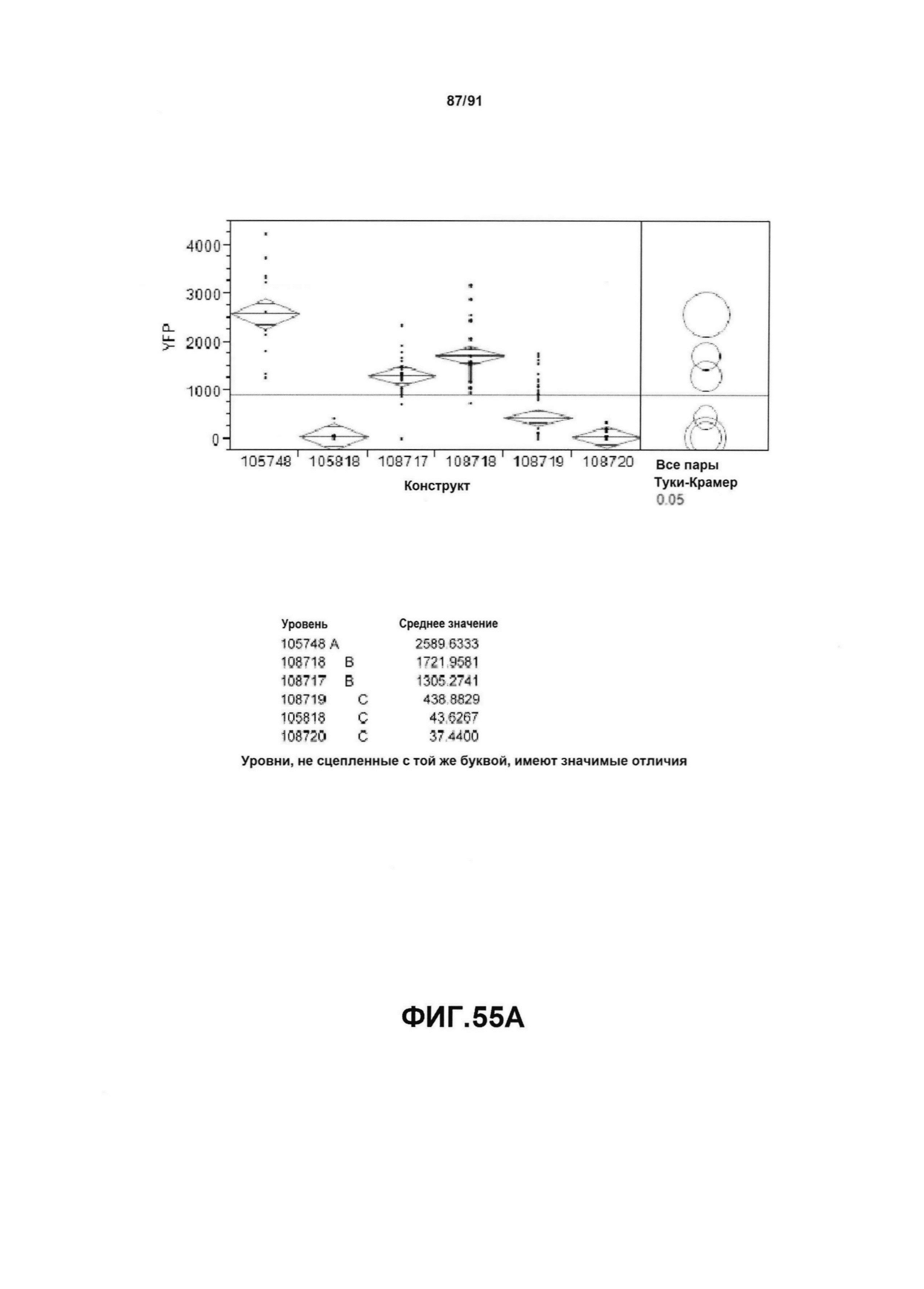
Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
5-фторпиримидиновые производные в качестве фунгицидов
Пестицидные композиции
Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
Стабильная пестицидная композиция на основе сульфоксимина и способ борьбы с насекомыми
5-фторпиримидиновые производные в качестве фунгицидов

