Результат интеллектуальной деятельности: УСТОЙЧИВЫЙ К НАСЕКОМЫМ-ВРЕДИТЕЛЯМ И ТОЛЕРАНТНЫЙ К ГЕРБИЦИДАМ СЕЛЕКЦИОННЫЙ ГИБРИД ТРАНСФОРМАНТА СОИ pDAB9582.814.19.1 и pDAB4468.04.16.1
Вид РИД
Изобретение
Уровень техники, предшествующий изобретению
[0001] Гены, кодирующие Cry1F и Cry1Ac synpro (Cry1Ac) способны обеспечивать устойчивость трансгенных растений к насекомым-вредителям, например устойчивость к чешуекрылым насекомым, и ген, кодирующий PAT (фосфинотрицинацетилтрансферазу) способен обеспечивать устойчивость трансгенных растений к гербициду фосфинотрицину (глуфосинату). PAT успешно экспрессировали в сое для использования в качестве селектируемуего маркера при получении устойчивых к насекомым-вредителям трансгенных культур и для обеспечения коммерческих уровней толерантности трансгенных культур к гербициду глуфосинату. При экспрессии в трансгенных растениях ген, кодирующий AAD-12 (арилоксиалканоатдиоксигеназу-12), способен обеспечивать коммерческие уровни толерантности к гербицидам на основе феноксиуксусной кислоты, 2,4-D и MCPA и гербицидам на основе пиридилоксиуксусной кислоты, триклопиру и флуроксипиру.
[0002] Известно, что на экспрессию чужеродных генов в растениях влияет их локализация в геноме растения, возможно, вследствие структуры хроматина (например, гетерохроматина) или близости регуляторных элементов транскрипции (например, энхансеров) рядом с участком интеграции (Weising et al., Ann. Rev. Genet., 22:421-477, 1988). В то же время наличие трансгена при различных локализациях в геноме влияет на весь фенотип растения различными способами. По этой причине часто является необходимым проведение скрининга большого числа трансформационных событий для идентификации трансформационного события, характеризующегося оптимальной экспрессией представляющего интерес вводимого гена. Например, наблюдали, что у растений и у других организмов может существовать широкая вариация уровней экспрессии вводимого гена среди трансформационных событий. Также могут существовать различия в пространственных или временных паттернах экспрессии, например, различия относительной экспрессии трансгена в различных тканях растения, которые могут не соответствовать паттернам, прогнозируемым на основании регуляторных элементов транскрипции, содержащихся во вводимой генной конструкции. По этой причине, общепринято получают от сотен до нескольких тысяч различных трансформационных событий и проводят скрининг таких трансформационных событий для поиска одного трансформационного события, которое обладает желаемыми уровнями и паттернами экспрессии трансгена для коммерческих целей. Трансформационное событие, которое обладает желаемыми уровнями или паттернами экспрессии трансгена, является пригодным для интрогрессии трансгена в другие генетические среды, посредством полового ауткроссинга общепринятыми способами скрещивания. Потомство таких скрещиваний сохраняет характеристики экспрессии трансгена исходного трансформационного события. Такую стратегию используют для обеспечения надежной экспрессии гена в ряде сортов, которые хорошо адаптируются к условиям роста в определенной местности.
[0003] Для определения может ли потомство полового скрещивания содержать трансген или группу представляющих интерес трансгенов, желательно иметь возможность детектировать наличие конкретного трансформационного события или многих трансформационных событий. Кроме того, способ детекции конкретного трансформационного события или многих трансформационных событий может оказаться полезным для соблюдения правил, необходимых для предпродажного одобрения и маркировки пищевых продуктов, получаемых, например, из рекомбинантных сельскохозяйственных культур, или для применения в мониторинге окружающей среды, мониторинге следов культур в поле или мониторинге продуктов, получаемых из урожая культуры, а также для применения в обеспечении соблюдения сторон установленных нормативных условий или условий договора.
[0004] Наличие одного или нескольких трансгенных трансформационных событий можно детектировать посредством детекции любой нуклеиновой кислоты известным в данной области способом, включая, но, не ограничиваясь ими, полимеразную цепную реакцию (ПЦР) или гибридизацию ДНК с использованием зондов для нуклеиновых кислот. Такие способы детекции, как правило, направлены на часто используемые генетические элементы, такие как промоторы, терминаторы, маркерные гены и т.д., т.к. для многих ДНК-конструкций кодирующая область является взаимозаменяемой. В результате такие способы могут являться непригодными для идентификации различных трансформационных событий, в частности трансформационных событий, получаемых с использованием аналогичной ДНК-конструкции или очень сходных конструкций, за исключением тех случаев, когда последовательность ДНК, фланкирующая ДНК, смежная со встраиваемой гетерологичной ДНК, является известной. Например, в патентной заявке 2006/0070139 описан специфичный по отношению к трансформационному событию ПЦР анализ для трансформационного события кукурузы DAS-59122-7. Желательно иметь простой и дифференциальный способ идентификации селекционных гибридов трансформантов сои pDAB9582.814.19.1::pDAB4468.04.16.1.
Краткое описание сущности изобретения
[0005] Настоящее изобретение относится к новому устойчивому к насекомым-вредителям и толерантному к гербицидам трансгенному селекционному гибриду трансформанта сои, обозначаемому как трансформант сои pDAB9582.814.19.1::pDAB4468.04.16.1. Этот селекционный гибрид содержит Cry1F, Cry1Ac и pat, как описано в настоящем описании, и aad-12 и pat, как описано в настоящем описании и в международной патентной заявке № WO/2012/075426, введенные в конкретные участки в геном клетки сои. Репрезентативные семена сои депонировали в American Type Culture Collection (ATCC) с № доступа, указанным в абзаце [0021]. ДНК растений сои, содержащих такое трансформационное событие, включают соединительные/фланкирующие последовательности, описываемые в настоящем описании, которые характеризуют локализацию встраиваемой ДНК в геном сои. SEQ ID NO:1, SEQ ID NO:2 и SEQ ID NO:15 являются диагностическими для трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1. Более конкретно, последовательности, окружающие участки соединения при 1400/1401 п.н., 1536/1537 п.н. SEQ ID NO:1, 152/153 п.н. SEQ ID NO:2, 2730/2731 п.н. SEQ ID NO:15 и 9121/9122 п.н. SEQ ID NO:15, являются диагностическими для трансформационного события pDAB9582.814.19.1::pDAB4468.04.16.1. В абзаце [0012] ниже описаны примеры последовательностей, содержащих эти участки соединения, которые представляют собой характерный признак ДНК сои, содержащей трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1.
[0006] В одном из вариантов осуществления изобретение относится к растению сои или его части, которое является устойчивым к Pseudoplusia includens (соевой совке), и геном которой содержит одну или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2, и п.н. 3-303 SEQ ID NO:2, и одну или более последовательностей, выбранных из группы, состоящей из п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15 и их комплементов. В другом варианте осуществления изобретение относится к семенам таких растений.
[0007] В другом варианте осуществления изобретение относится к способу борьбы с насекомыми-вредителями, который включает подвергание воздействию насекомыми-вредителями устойчивых к насекомым-вредителям растений сои, где растения сои обладают геномом, который содержит один или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2 и п.н. 3-303 SEQ ID NO:2, и одну или несколько последовательностей, выбранных из группы, состоящей из п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15, и их комплементы, которые характеризуются содержанием трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1, чтобы таким образом бороться с насекомыми-вредителями. Наличие генов cry1F v3 (cry1F) и cry1Ac synpro (cry1Ac) в трансформационном событии сои pDAB9582.814.19.1::pDAB4468.04.16.1 обеспечивает устойчивость, например, к Pseudoplusia includens (соевой совке), Anticarsia gemmatalis (гусенице бархатных бобов), Epinotia aporema, Omoides indicatus, Rachiplusia nu, Spodoptera frugiperda, Spodoptera cosmoides, Spodoptera eridania, Heliothis virescens, Heliocoverpa zea, Spilosoma virginica и Elasmopalpus lignosellus.
[0008] В одном из вариантов осуществления изобретение относится к растению сои или его части, которое является толерантным к гербицидам на основе феноксиуксусной кислоты, таким как 2,4-D и MCPA. В другом варианте осуществления изобретение относится к растению сои или его части, которое является толерантным к гербицидам на основе пиридилоксиуксусной кислоты, таким как триклопир и флуроксипир. В этих вариантах осуществления растение сои обладает геномом, содержащим одну или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2, п.н. 3-303 SEQ ID NO:2, и одну или более последовательностей, выбранных из группы, состоящей из п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и, п.н. 8921-9321 SEQ ID NO:15, и их комплементов. В другом варианте осуществления изобретение относится к семенам таких растений.
[0009] В другом варианте осуществления изобретение относится к способу борьбы с сорняками соевой культуры, который включает применение гербицидов на основе феноксиуксусной кислоты, таких как 2,4-D и MCPA. В другом варианте осуществления изобретение относится к способу борьбы с сорняками соевой культуры, который включает применение гербицидов на основе пиридилоксиуксусной кислоты, таких как триклопир и флуроксипир, к соевой культуре, где соевая культура содержит растения сои, которые обладают геномом, содержащим одну или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2, п.н. 3-303 SEQ ID NO:2, и одну или более последовательностей, выбранных из группы, состоящей из п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15, и их комплементов. Наличие гена aad-12 в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 обеспечивает толерантность к гербицидам на основе феноксиуксусной кислоты и гербицидам на основе пиридилоксиуксусной кислоты.
[0010] В другом варианте осуществления изобретение относится к способу борьбы с сорняками соевой культуры, который включает применение гербицида глуфосината к соевой культуре, где указанная соевая культура содержит растения сои, которые обладают геномом, содержащим одну или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2, п.н. 3-303 SEQ ID NO:2, и одну или более последовательностей, выбранных из группы, состоящей из п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15, и их комплементов, которые являются диагностическими для содержания трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1. Наличие гена pat v6 (pat) в трансформационном событии сои pDAB9582.814.19.1::pDAB4468.04.16.1 обеспечивает толерантность к гербициду глуфосинату.
[0011] В другом варианте осуществления изобретение относится к способу детекции трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1 в образце, содержащем ДНК сои, где указанный способ включает:
a) контактирование указанного образца с первым праймером длиной по меньшей мере 10 п.н., который селективно связывается с фланкирующей последовательностью в п.н. 1-1400 SEQ ID NO:1 или ее комплементе, и вторым праймером длиной по меньшей мере 10 п.н., который селективно связывается с встраиваемой последовательностью в п.н. 1401-1836 SEQ ID NO:1 или ее комплементом, и анализ ампликона, образуемого между указанными праймерами, или
b) контактирование указанного образца с первым праймером длиной по меньшей мере 10 п.н., который селективно связывается с встраиваемой последовательностью в п.н. 1-152 SEQ ID NO:2 или ее комплементе, и вторым праймером длиной по меньшей мере 10 п.н., который селективно связывается с фланкирующей последовательностью в п.н. 153-1550 SEQ ID NO:2 или ее комплементе, или
c) контактирование указанного образца с первым праймером длиной по меньшей мере 10 п.н., который селективно связывается с встраиваемой последовательностью в п.н. 2731-9121 SEQ ID NO:15 или ее комплементе, и вторым праймером длиной по меньшей мере 10 п.н., который селективно связывается с фланкирующей последовательностью в п.н. 1-2730 SEQ ID NO:15 или ее комплементе, или
d) контактирование указанного образца с первым праймером длиной по меньшей мере 10 п.н., который селективно связывается с встраиваемой последовательностью в п.н. 2731-9121 SEQ ID NO:15 или ее комплементе, и вторым праймером длиной по меньшей мере 10 п.н., который селективно связывается с фланкирующей последовательностью в п.н. 9122-10198 SEQ ID NO:15 или ее комплементе, и
c) анализ ампликона, образуемого между указанными праймерами.
[0012] В другом варианте осуществления изобретение относится к способу детекции трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1, включающему:
a) контактирование указанного образца с первым праймером, который селективно связывается с фланкирующей последовательностью, выбранной из группы, состоящей из п.н. 1-1400 SEQ ID NO:1 и п.н. 153-1550 SEQ ID NO:2 и их комплементом, и вторым праймером, который селективно связывается с SEQ ID NO:3 или ее комплементом, и третьим праймером, который селективно связывается с фланкирующей последовательностью п.н. 1-2730 SEQ ID NO:15 и п.н. 9122-10198 SEQ ID NO:15, и их комплементом; и четвертым праймером, который селективно связывается с встраиваемой последовательностью 2731-9121 SEQ ID NO:15 и ее комплементом.
b) подвергание указанного образца полимеразной цепной реакции и
c) анализ ампликона, образуемого между указанными праймерами.
[0013] В другом варианте осуществления изобретение относится к способу селекции растения сои, включающему: скрещивание первого растения со вторым растением сои с получением третьего растения сои, где указанное первое растение, содержит ДНК, содержащую одну или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2 и п.н. 3-303 SEQ ID NO:2, и одну или более последовательностей, выбранных из группы, состоящей из п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15 и их комплементов, и анализ указанного третьего растения сои на содержание ДНК, содержащей одну или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2, п.н. 3-303 SEQ ID NO:2, п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15 и их комплементов.
[0014] В другом варианте осуществления изобретение относится к выделенной молекуле ДНК, которая является диагностической для трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1. Такие молекулы наряду с SEQ ID NO:1, 2 и 15 включают молекулы длиной по меньшей мере 25 п.н., содержащие п.н. 1400-1401 SEQ ID NO:1 и по меньшей мере 10 п.н. SEQ ID NO:1 в каждом направлении от п.н. 1400/1401 участка соединения; ампликоны длиной по меньшей мере 25 п.н., содержащие 152-153 SEQ ID NO:2 и по меньшей мере 10 п.н. SEQ ID NO:2 в каждом направлении от п.н. 152/153 участка соединения; ампликоны длиной по меньшей мере 25 п.н., содержащие п.н. 2730-2731 SEQ ID NO:15 и по меньшей мере 10 п.н. SEQ ID NO:15 в каждом направлении от п.н. 2730/2731 участка соединения; ампликоны длиной по меньшей мере 25 п.н., содержащие п.н. 9121-9122 SEQ ID NO:15 и по меньшей мере 10 п.н. SEQ ID NO:15 в каждом направлении от п.н. 9121/9122 участка соединения. Примеры представляют собой п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2 и п.н. 3-303 SEQ ID NO:2, п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15, и их комплементы.
[0015] В другом варианте осуществления изобретение относится к способу борьбы с вредителями зерна сои, семян или жмыха, который включает введение трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1 в указанное зерно, семена или жмых, как продемонстрировано указанным зерном, семенами или жмыхом, содержащим ДНК, содержащую одну или более последовательностей, выбранных из группы, состоящей из п.н. 1385-1415 SEQ ID NO:1, п.н. 1350-1450 SEQ ID NO:1, п.н. 1300-1500 SEQ ID NO:1, п.н. 1200-1600 SEQ ID NO:1, п.н. 137-168 SEQ ID NO:2, п.н. 103-203 SEQ ID NO:2 и п.н. 3-303 SEQ ID NO:2, и одну или более последовательностей, выбранных из группы, состоящей из п.н. 2680-2780 SEQ ID NO:15, п.н. 2630-2830 SEQ ID NO:15, п.н. 2530-2930 SEQ ID NO:15, п.н. 9071-9171 SEQ ID NO:15, п.н. 9021-9221 SEQ ID NO:15 и п.н. 8921-9321 SEQ ID NO:15 и их комплементов.
[0016] Изобретение также включает клетки растения и части растения сои, включая, но, не ограничиваясь ими, пыльцу, семязачаток, цветы, побеги, корни и листья, и ядра вегетативных клеток, клеток пыльцы, семян и жмыха и яйцеклетки, которые содержат трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида.
[0017] В некоторых вариантах осуществления трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида можно комбинировать с другими признаками, включая, например, другой ген(ы) толерантности к гербицидам и/или подавляющих рост насекомых белков и регуляторные последовательности транскрипции (т.е. РНК-интерференцию, дцРНК, факторы транскрипции и т.д.). Можно осуществлять стэкинг дополнительных признаков в геноме растения путем селекции растений, повторной трансформации трансгенного растения, содержащего трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида, или добавления новых признаков путем направленной интеграции.
[0018] Другие варианты осуществления включают эксцизию полинуклеотидных последовательностей, которые содержат трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида, включая, например, экспрессионную кассету гена pat. В результате эксцизии полинуклеотидной последовательности модифицированное трансформационное событие можно перенаправлять на конкретный участок хромосомы, где проходит стэкинг дополнительных полинуклеотидных последовательностей совместно с трансформационным событием сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида.
[0019] В одном из вариантов осуществления настоящее изобретение относится к участку-мишени хромосомы сои, локализованному в хромосоме 02 между фланкирующими последовательностями, указанными в SEQ ID NO:1 и 2.
[0020] В одном из вариантов осуществления настоящее изобретение относится к участку-мишени хромосомы сои, локализованному в хромосоме 04 между фланкирующими последовательностями, указанными в SEQ ID NO:15.
[0021] В одном из вариантов осуществления настоящее изобретение относится к способу получения трансгенного растения сои, включающему введение гетерологичной нуклеиновой кислоты в положение в хромосоме 02 между геномными последовательностями, указанными в SEQ ID NO:1 и 2, т.е. между п.н. 1-1400 SEQ ID NO:1 и п.н. 153-1550 SEQ ID NO:2.
[0022] В одном из вариантов осуществления настоящее изобретение относится к способу получения трансгенного растения сои, включающему введение гетерологичной нуклеиновой кислоты в положение в хромосоме 04 между геномными последовательностями, указанными в SEQ ID NO:15, т.е. между п.н. 1-2730 SEQ ID NO:15 и п.н. 9122-10198 SEQ ID NO:15.
[0023] Кроме того настоящее изобретение относится к анализам детекции наличия рассматриваемого трансформационного события в образце (например, сои). Анализы могут быть основаны на последовательности ДНК рекомбинантной конструкции, встраиваемой в геном сои, и на геномных последовательностях, фланкирующих участок вставки. Также предоставлены наборы и условия, пригодные для проведения анализов.
[0024] Настоящее изобретение частично относится к клонированию и анализу последовательностей ДНК граничных областей, получаемых в результате введения Т-ДНК из pDAB9582 и pDAB4468 в трансгенные линии сои. Эти последовательности являются уникальными. На основании встраиваемой и соединительной последовательностей могут существовать специфичные для трансформационного события праймеры и их получали. ПЦР анализ демонстрировал, что эти трансформационные события можно идентифицировать анализом ПЦР-ампликонов, получаемых с использованием таких наборов специфичных для трансформационного события праймеров. Таким образом, эти и другие связанные способы можно использовать для уникальной идентификации линий сои, содержащих трансформационное событие по настоящему изобретению.
Депонирование семян
[0025] В качестве части настоящего изобретения депонировали по меньшей мере 2500 семян линии сои, содержащей трансформационное событие сои pDAB9582.814.19.1, и 2500 семян линии сои, содержащей трансформационное событие сои pDAB4468.04.16.1, и обеспечивали общественный без ограничения (но при условии патентных прав) в Американской коллекции типовых культур (ATCC), 10801 University Boulevard, Manassas, VA, 20110. Депозиты, обозначаемые как № ATCC депозита PTA-10442 (pDAB4468.04.16.1) и № ATCC депозита (pDAB9582.814.19.1), регистрировали от имени Dow AgroSciences LLC 22 октября, 2009 года и соответственно. Эти депозиты вносили, и их будут поддерживать в соответствии и по условиям Будапештского договора в отношении депозитов семян для целей патентной процедуры.
Краткое описание последовательностей
[0026] SEQ ID NO:1 представляет собой 5'-фланкирующую граничную последовательность ДНК для трансформационного события сои pDAB9582.814.19.1. Нуклеотиды 1-1400 представляют собой геномную последовательность. Нуклеотиды 1401-1535 представляют собой реаранжированную последовательность из pDAB9582. Нуклеотиды 1536-1836 представляют собой встраиваемую последовательность.
[0027] SEQ ID NO:2 представляет собой 3'-фланкирующую граничную последовательность ДНК для трансформационного события сои pDAB9582.814.19.1. Нуклеотиды 1-152 представляют собой встраиваемую последовательность. Нуклеотиды 153-1550 представляют собой геномную последовательность.
[0028] SEQ ID NO:3 представляет собой последовательность ДНК pDAB9582, которая аннотирована ниже в таблице 1.
[0029] SEQ ID NO:4 представляет собой олигонуклеотидный праймер 81419_FW3 для подтверждения 5'-граничной геномной ДНК.
[0030] SEQ ID NO:5 представляет собой олигонуклеотидный праймер 81419_RV1 для подтверждения 3'-граничной геномной ДНК.
[0031] SEQ ID NO:6 представляет собой олигонуклеотидный праймер 81419_RV2 для подтверждения 3'-граничной геномной ДНК.
[0032] SEQ ID NO:7 представляет собой олигонуклеотидный праймер 81419_RV3 для подтверждения 3'-граничной геномной ДНК.
[0033] SEQ ID NO:8 представляет собой олигонуклеотидный праймер 5'IREnd-0l для подтверждения 5'-граничной геномной ДНК.
[0034] SEQ ID NO:9 представляет собой олигонуклеотидный праймер 5'IREnd-02 для подтверждения 5'-граничной геномной ДНК.
[0035] SEQ ID NO:10 представляет собой олигонуклеотидный праймер AtUbi10RV1 для подтверждения 5'-граничной геномной ДНК.
[0036] SEQ ID NO:11 представляет собой олигонуклеотидный праймер AtUbi10RV2 для подтверждения 5'-граничной геномной ДНК.
[0037] SEQ ID NO:12 представляет собой олигонуклеотидный праймер 3'PATEnd05 для подтверждения 3'-граничной геномной ДНК.
[0038] SEQ ID NO:13 представляет собой олигонуклеотидный праймер 3'PATEnd06 для подтверждения 3'-граничной геномной ДНК.
[0039] SEQ ID NO:14 представляет собой подтвержденную последовательность трансформационного события сои pDAB9582.814.19.1. Включая 5'-фланкирующую геномную последовательность, pDAB9582T-вставку цепи и 3'-фланкирующую геномную последовательность.
[0040] SEQ ID NO:15 представляет собой подтвержденную последовательность трансформационного события сои pDAB4468.04.16.1. Включая 5'-фланкирующую геномную последовательность, pDAB4468T-вставку цепи и 3'-фланкирующую геномную последовательность.
Краткое описание чертежей
[0041] Фиг.1 представляет собой карту плазмиды pDAB9582, содержащей экспрессионные кассеты cry1F, cry1Ac и pat.
[0042] На фиг.2 проиллюстрированы локализации праймеров для подтверждения 5'- и 3'-граничной последовательности трансформационного события сои pDAB9582.814.19.1.
[0043] На фиг.3 проиллюстрировано расположение геномной последовательности в трансформационном событии сои pDAB9582.814.19.1.
Подробное описание изобретения
[0044] Секвенировали и характеризовали фланкирующие последовательности встраивания трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1. Разрабатывали специфические по отношению к трансформационному событию анализы. Также их картировали в геноме сои (хромосома сои 02 и 04). Трансформационные события можно совместно интрогрессировать в дополнительные элитные линии.
[0045] Как указано выше в разделе уровень техники, предшествующий изобретению, введение и интеграция трансгена в геном растения включает некоторые случайные трансформационные события (таким образом, используют название "трансформационное событие" для данной вставки). Таким образом, многими способами трансформации, такими как опосредованная Agrobacterium трансформация, биолистическая трансформация (т.е. генная пушка) и опосредованная карбидом кремния трансформация (т.е. WHISKERS™), невозможно прогнозировать, где в геноме встроится трансген. Таким образом, определение фланкирующей геномной ДНК растения на обеих сторонах вставки может являться важным для идентификации растения, которое содержит данное трансформационное событие вставки. Например, можно конструировать праймеры для ПЦР, которые образуют ПЦР-ампликон между соединительной областью вставки и генома хозяина. Такой ПЦР-ампликон можно использовать для идентификации уникального или отдельного типа трансформационного события встраивания.
[0046] В настоящем описании предоставлены определения и примеры с целью облегчения описания настоящего изобретения и для руководства для специалистов в данной области для практического осуществления изобретения. Если не указано иное, термины следует понимать в соответствии с их общепринятым использованием специалистами в соответствующей области. Используют такую номенклатуру для оснований ДНК, как указанная в 37 CFR §1.822.
[0047] Как используют в настоящем описании, термин "потомство" означает потомство любой генерации родительского растения, которое содержит трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида.
[0048] Трансгенное "трансформационное событие" получают трансформацией растительных клеток гетерологичной ДНК, т.е. конструкцией нуклеиновой кислоты, которая содержит представляющие интерес трансгены, восстановлением популяции растений, получаемых в результате встраивания трансгена в геном растения, и отбором конкретного растения, характеризующегося конкретной локализацией в геноме. Термин "трансформационное событие" относится к исходному трансформанту и потомство трансформанта, которые содержат гетерологичную ДНК. Термин "трансформационное событие" также относится к потомству, получаемому половым ауткроссингом трансформанта и другого сорта, который содержит геномную/трансгенную ДНК. Даже после повторного обратного скрещивания с рекуррентным родителем, встраиваемая трансгенная ДНК и фланкирующая геномная ДНК (геномная/трансгенная ДНК) от трансформированного родителя содержится у потомства скрещивания в аналогичной локализации в хромосоме. Термин "трансформационное событие" также относится к ДНК от исходного трансформанта и его потомства, содержащего встраиваемую ДНК и фланкирующую геномную последовательность, непосредственно прилегающую к встраиваемой ДНК, для которой предполагают, что она передастся потомству, которое получает встраиваемую ДНК, содержащую представляющий интерес трансген, в результате полового скрещивания одной родительской линии, которая содержит встраиваемую ДНК (например, исходный трансформант и потомство, получаемое в результате самоопыления) и родительской линии, которая не содержит встраиваемую ДНК.
[0049] "Соединительная последовательность" или "граничная последовательность" включает точку, в которой страиваемая в геном ДНК является соединенной с ДНК нативного генома сои, фланкирующей точку встраивания, где идентификация или детекция одной или других соединительных последовательностей в генетическом материале растения является достаточной для диагностики трансформационного события. Также включены последовательности ДНК, которые включают вставки в описываемых в настоящем описании трансформационных событиях сои и аналогичные длины фланкирующей ДНК. В настоящем описании предоставлены конкретные примеры таких диагностических последовательностей, однако, другие последовательности, которые перекрывают участки соединения вставок или соединения вставок и геномной последовательности, также являются диагностическими, и их можно использовать по настоящему изобретению.
[0050] Настоящее изобретение частично относится к идентификации трансформационного события с использованием таких фланкирующих, соединительных и встраиваемых последовательностей. Соответствующие праймеры для ПЦР и ампликоны также входят в изобретение. В соответствие с настоящим изобретением можно использовать способы ПЦР анализа с использованием ампликонов, которые охватывают встраиваемую ДНК и ее границы, для детекции или идентификации находящихся в продаже трансгенных сортов или линий сои, получаемых из трансгенных линий сои объекта собственности.
[0051] Фланкирующие/соединительные последовательности являются диагностическими для трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида. На основании этих последовательностей получали специфичные к трансформационному событию праймеры. ПЦР анализ демонстрировал, что эти линии сои можно идентифицировать в различных генотипах сои посредством анализа ПЦР-ампликонов, получаемых с использованием таких наборов специфичных к трансформационному событию праймеров. Таким образом, эти и другие связанные способы можно использовать для уникальной идентификации этих линии сои. Последовательности, определяемые в настоящем описании, являются уникальными.
[0052] Способы детекции по настоящему изобретению являются особенно пригодными в сочетании с селекцией растений для определения, какие растения потомства содержат данное трансформационное событие после того, как родительское растение, содержащее представляющее интерес трансформационное событие, скрещивают с другой линией растений с целью внесения одного или более дополнительных представляющих интерес признаков в потомство. Такие способы ПЦР анализа являются полезными для программ селекции сои, а также для контроля качества, в частности для находящихся в продаже трансгенных семян сои. В настоящее время также можно получать и использовать наборы для детекции ПЦР для таких трансгенных линий сои. Это также может быть полезным для регистрации продуктов и управления качеством продукта.
[0053] Кроме того, фланкирующие/геномные последовательности сои можно использовать для конкретной идентификации локализации в геноме каждой вставки. Эту информацию можно использовать для получения системы молекулярных маркеров, специфических для каждого трансформационного события. Их можно использовать для способов ускоренной селекции и для установления данных сцепления.
[0054] Кроме того, информацию о фланкирующей последовательности можно использовать для исследования и характеристики способов интеграции трансгена, характеристики участков геномной интеграции, сортировки трансформационных событий, стабильности трансгенов и их фланкирующих последовательностей и экспрессии гена (в частности, связанной с выключением генов, паттернами метилирования трансгенов, эффектов положения и потенциальных связанных с экспрессией элементов, таких как MARS [участки прикрепления к матриксу] и т.п.).
[0055] В свете описания предмета изобретения следует понимать, что настоящее изобретение включает семена, доступные по № ATCC депозита, указанному в абзаце [0021]. Настоящее изобретение также включает толерантное к гербицидам растение сои, выращенное из семян, депонированных с № ATCC депозита, указанным в абзаце [0021]. Настоящее изобретение дополнительно включает части указанного растения, такие как листья, образцы ткани, семена, продуцируемые указанным растением, пыльцу и т.п. (где они содержат cry1F, cry1Ac, pat, и SEQ ID NO:1 и 2 наряду с aad-12, pat и SEQ ID NO:15).
[0056] Кроме того, настоящее изобретение включает растения потомков и/или потомства растений, выращенных из депонированных семян, предпочтительно устойчивого к гербицидам растения сои, где указанное растение, обладает геномом, содержащим детектируемую соединительную последовательность дикого типа, как описано в настоящем описании. Как используют в настоящем описании, термин "соя" означает Glycine max и включает все его сорта, которые можно выводить из растения сои.
[0057] Настоящее изобретение дополнительно включает способы проведения скрещиваний с использованием растения по настоящему изобретению в качестве по меньшей мере одного родителя. Например, настоящее изобретение включает гибридное растение F1, имеющее в качестве одного или обоих родителей любое из растений, иллюстрируемых в настоящем описании, включая, но, не ограничиваясь тем, что оно имеет одного родителя, содержащего трансформационное событие сои pDAB9582.814.19.1, и другого родителя, содержащего pDAB4468.04.16.1. Также настоящее изобретение относится к семенам, продуцируемым такими гибридами F1 по настоящему изобретению. Настоящее изобретение включает способ получения семян гибрида F1 путем скрещивания иллюстративного растения с отличным (например, инбредным родителем) растением и сбора получаемых семян гибрида. Настоящее изобретение включает иллюстративное растение, которое представляет собой материнскую форму или отцовскую форму. Характеристики получаемых растений можно улучшать путем тщательного рассмотрения родительских растений.
[0058] Устойчивое к насекомым-вредителям/толерантное к 2,4-D/ толерантное к глуфосинату растение сои по настоящему изобретению можно размножать сначала половым скрещиванием первого родительского растения сои, состоящего из растения сои, выращенного из семян любой из линий, указанных в настоящем описании, и второго родительского растения сои, таким образом, получая ряд растений первого потомства; затем отбирая растение первого потомства, которое является устойчивым к глуфосинату и/или 2,4-D; проводя самоопыление растения первого потомства, таким образом, получая ряд растений второго потомства, а затем отбирая из растений второго потомства растение, которое является устойчивым к глуфосинату и/или 2,4-D. Эти этапы могут дополнительно включать обратное скрещивание растения первого потомства или растения второго потомства со вторым родительским растением сои или третьим родительским растением сои. Затем можно культивировать культуру сои, содержащую семена сои по настоящему изобретению или их потомство.
[0059] Также следует понимать, что два различных трансгенных растения также можно связывать (mated) с получением потомства, которое содержит два независимо сегрегированных, добавленных экзогенных гена. Самоопылением соответствующего потомства можно получать растение, которое является гомозиготным по обоим добавленным экзогенным генам. Также предусмотрены обратное скрещивание с родительским растением и ауткроссинг с нетрансгенным растением в виде вегетативного размножения. Другие способы селекции, широко используемые для различных признаков и культур, являются известными в данной области. Размножение обратным скрещиванием используют для переноса генов для просто наследуемого, высоконаследуемого признака в желаемый гомозиготный культивар или инбредную линию, которая представляет собой рекуррентного родителя. Источник признака, который необходимо переносить, называют донорным растением. Ожидают, что получаемое растение обладает характеристиками рекуррентного родителя (например, культивара) и желаемым признаком, переносимым от донорного родителя. После первичного скрещивания отбирают индивидуумов, обладающих фенотипом донорного родителя, и повторно скрещивают (проводят обратное скрещивание) с рекуррентным родителем. Ожидают, что получаемый родитель обладает характеристиками рекуррентного родителя (например, культивара) и желаемым признаком, переносимым от донорного родителя.
[0060] Аналогично, устойчивое к насекомым-вредителям/толерантное к 2,4-D/толерантное к глуфосинату растение сои по настоящему изобретению можно трансформировать дополнительными трансгенами способами, известными в данной области. Способы трансформации, такие как опосредованная Agrobacterium трансформация, биолистическая трансформация (т.е. генная пушка) и опосредованная карбидом кремния трансформация (т.е. WHISKERS), можно использовать для введения дополнительного трансгена(ов) в геном селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1. Для идентификации растений, которые содержат стабильный интегрант нового трансгена в дополнение к генам aad-12, cry1F, cry1Ac, pat по настоящему изобретению, можно выполнять отбор и характеристику трансгенного растения, содержащего вновь введенные трансгены.
[0061] Молекулы ДНК по настоящему изобретению можно использовать в качестве молекулярных маркеров в способе скрещивания с использованием маркера (MAB). Молекулы ДНК по настоящему изобретению можно использовать в способах (таких как маркеры AFLP, маркеры RFLP, маркеры RAPD, SNP, и SSR), которыми идентифицируют генетически сцепленные хозяйственно-ценные признаки, как известно в данной области. Признаки устойчивости к насекомым-вредителям и толерантности к гербицидам можно отслеживать у потомства скрещивания с растением сои по настоящему изобретению (или у их потомства и у любого другого культивара или сорта сои) способами MAB. Молекулы ДНК представляют собой маркеры для такого признака, и можно использовать способы MAB, которые хорошо известны в данной области, для отслеживания признака(ов) устойчивости к гербицидам у растения сои, где по меньшей мере одна линия сои по настоящему изобретению или ее потомство, являлась родителем или предком. Способы по настоящему изобретению можно использовать для идентификации любого сорта сои, содержащего рассматриваемое трансформационное событие.
[0062] Способы по настоящему изобретению включают способ получения устойчивого к насекомым-вредителям/толерантного к гербицидам растения сои, где указанный способ включает скрещивание с растением по настоящему изобретению. Более конкретно, указанные способы могут включать скрещивание двух растений по настоящему изобретению или одного растения по настоящему изобретению и любого другого растения. Предпочтительные способы дополнительно включают отбор потомства указанного скрещивания путем анализа указанного потомства в отношении детектируемого трансформационного события по настоящему изобретению и благоприятной сортовой продуктивности (например, урожайности). Например, настоящее изобретение можно использовать для отслеживания рассматриваемого трансформационного события посредством циклов скрещивания с растениями, содержащими другие желаемые признаки, такие как хозяйственно-ценные признаки, толерантность или устойчивость к заболеваниям, толерантность или устойчивость к нематодам и срок созревания. Растения, содержащие рассматриваемое трансформационное событие и желаемый признак, можно детектировать, идентифицировать, отбирать и быстро использовать, например, на последующих этапах размножения. Рассматриваемое трансформационное событие/признак также можно комбинировать с дополнительным признаком(ами) устойчивости к насекомым-вредителям и/или дополнительными признаками толерантности к гербицидам посредством скрещивания и отслеживать согласно настоящему изобретению. Варианты осуществления последнего представляют собой растения, содержащие рассматриваемое трансформационное событие, комбинированное с геном толерантности к глифосату, который обеспечивает толерантность к гербициду глифосату.
[0063] Таким образом, настоящее изобретение можно комбинировать, например, с признаками, кодирующими устойчивость к глифосату (например, устойчивого растения или бактериальных EPSPS, GOX, GAT), устойчивость к глуфосинату (например, pat, bar), устойчивость к гербицидам, ингибирующим ацетолактатсинтазу (ALS) (например, имидазолинонам [таким как имазетапир], сульфонилмочевинам, триазолопиримидинсульфонанилиду, пиримидинилтиобензоатам и другим химическим соединениям [Csr1, SurA и т.д.]), устойчивость к бромоксинилу (например, Bxn), фермент устойчивости к ингибиторам HPPD (4-гидроксифенилпируватдиоксигеназы), устойчивость к ингибиторам фитоендесатуразы (PDS), устойчивость к ингибирующим фотосистему II гербицидам (например, psbA), устойчивость к ингибирующим фотосистему I гербицидам, устойчивость к ингибирующим протопорфириногеноксидазу IX (PPO) гербицидам (например, PPO-1), устойчивость к гербицидам на основе фенилмочевины (например, CYP76B1), разрушающие дикамбу ферменты (см., например, US 20030135879), и другие, для которых можно проводить стэкинг отдельно или во многих комбинациях для обеспечения способности эффективно бороться или предотвращать чередование сорняков и/или устойчивость к любому гербициду указанных выше классов.
[0064] Кроме того, трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1 селекционного гибрида можно комбинировать с одним или более дополнительных агрономических (например, устойчивости к насекомым-вредителям, устойчивости к патогенам или толерантности к стрессовым факторам и т.д.) или потребительских (например, повышенной урожайности, улучшенного масляного профиля, улучшенного качества волокна и т.д.) свойств. Таким образом, настоящее изобретение можно использовать для обеспечения полного агрономического комплекса улучшенного качества культуры наряду с возможностью универсально и экономически эффективно бороться с любым числом сельскохозяйственных вредителей.
[0065] В данной области описаны способы интеграции полинуклеотидной последовательности в конкретный участок хромосомы растительной клетки путем гомологичной рекомбинации. Например, сайт-специфическая интеграция, как описано в публикации патентной заявки США № 2009/0111188 A1, включенной в настоящее описание посредством ссылки, в которой описано использование рекомбиназы или интегразы для опосредования введения донорной полинуклеотидной последовательности в хромосому-мишень. Кроме того, в международной патентной заявке № WO 2008/021207, включенной в настоящее описание посредством ссылки, описана опосредованная цинковым пальцем гомологичная рекомбинация для интеграции одной или более донорных полинуклеотидных последовательностей в конкретные локализации в геноме. Применение рекомбиназ, таких как FLP/FRT, как описано в патенте США № 6720475, включенном в настоящее описание посредством ссылки, или CRE/LOX, как описано в патенте США № 5658772, включенном в настоящее описание посредством ссылки, можно использовать для интеграции полинуклеотидной последовательности в конкретный участок хромосомы. Кроме того, использование мегануклеаз для направленной доставки донорных полинуклеотидов в конкретную локализацию хромосомы описано у Puchta et al., PNAS USA 93 (1996) pp. 5055-5060.
[0066] Другие способы сайт-специфической интеграции в растительные клетки, как правило, известны и применяются (Kumar et al., Trends in Plant Sci., 6(4) (2001) pp. 155-159). Кроме того, системы сайт-специфической рекомбинации, которые идентифицировали у нескольких прокариотических и низших эукариотических организмов, можно применять для использования у растений. Примеры таких систем включают, но не ограничиваются ими, система рекомбиназ R/RS из плазмиды pSR1 дрожжей Zygosaccharomyces rouxii (Araki et al., (1985) J. Mol. Biol., 182: 191-203) и систему Gin/gix мю-фага (Maeser and Kahlmann, (1991) Mol. Gen. Genet., 230: 170-176).
[0067] В некоторых вариантах осуществления настоящего изобретения желательной может являться интеграция или стэкинг нового трансгена(ов) в близости с существующим трансгенным трансформационным событием. Трансгенное трансформационное событие можно рассматривать как предпочтительный геномный локус, который выбирали на основании уникальных характеристик, таких как один участок вставки, обычное менделеевское расщепление и стабильная экспрессия, и превосходная комбинация эффективности, включая толерантность к гербицидам и сельскохозяйственную продуктивность во многих климатических зонах. Вновь интегрированные трансгены должны поддерживать характеристики экспрессии трансгена существующих трансформантов. Кроме того, будет устранена проблема развития анализов детекции и подтверждения вновь интегрированного трансформационного события, т.к. геномные фланкирующие последовательности и локализация в хромосоме вновь интегрированного трансформационного события уже являются идентифицированными. Наконец, интеграция нового трансгена в конкретную локализацию в хромосоме, которая соединена с существующим трансгеном, будет способствовать интрогрессии трансгенов в другие генетические среды посредством полового ауткроссинга общепринятыми способами селекции.
[0068] В некоторых вариантах осуществления настоящего изобретения желательным может являться удаление полинуклеотидных последовательностей из трансгенного трансформационного события. Например, удаление трансгена, как описано в предварительной патентной заявке США № 61/297628, включенной в настоящее описание посредством ссылки, в которой описано использование цинкового пальца нуклеазы для удаления полинуклеотидной последовательности, состоящей из генной экспрессионной кассеты, из хромосомно интегрированного трансгенного трансформационного события. Полинуклеотидная последовательность, которую удаляют, может представлять собой селектируемый маркер. После эксцизии и удаления полинуклеотидной последовательности модифицированное трансгенное трансформационное событие можно перенаправлять посредством вставки полинуклеотидной последовательности. Вставка полинуклеотидной последовательности и последующее перенаправление модифицированного трансгенного трансформационного события обеспечивает преимущества, такие как повторное использование селектируемого маркера или возможность устранения непредусмотренных изменений в транскриптоме растения, которые являются результатом экспрессии конкретных генов.
[0069] В настоящем изобретении в настоящем описании раскрыт конкретный участок в хромосоме 02 в геноме сои, который является превосходным для встраивания гетерологичных нуклеиновых кислот. Таким образом, настоящее изобретение относится к способам введения представляющих интерес гетерологичных нуклеиновых кислот в этот предварительно установленный участок-мишень или вблизи такого участка-мишени. Настоящее изобретение также включает семена сои и/или растение сои, содержащее любую гетерологичную нуклеотидную последовательность, встраиваемую в описываемый участок-мишень или в непосредственной близости от такого участка. Одним из вариантов проведения такой направленной интеграции является удаление и/или замена отличной вставки вместо экспрессирующей кассеты pat, проиллюстрированной в настоящем описании. В этом отношении можно использовать, например и без ограничения, направленную гомологичную рекомбинацию согласно настоящему изобретению.
[0070] Как используют в настоящем описании, ген, трансформационное событие или "стэкинг" признаков представляет собой комбинацию желаемых признаков в одной трансгенной линии. Растениеводы-селекционеры получают стэкинг трансгенных признаков, проводя скрещивание между родителями, каждый из которых обладает желаемым признаком, а затем, идентифицируя потомство, которое обладает этими желаемыми признаками. Другой способ стэкинга генов проводят переносом двух или более генов в ядро клетки растения одновременно во время трансформации. Другой способ стэкинга генов проводят, повторно трансформируя трансгенное растение другим представляющим интерес геном. Например, стэкинг генов можно использовать для комбинации двух или более различных признаков, включая, например, два или более различных признака насекомого, признака(ов) устойчивости к насекомым-вредителям и признака(ов) устойчивости к заболеванию, два или более признака устойчивости к гербицидам и/или признака(ов) устойчивости к насекомым-вредителям и признака(ов) устойчивости к гербицидам. Использование селектируемого маркера в дополнение к представляющему интерес гену также можно рассматривать как стэкинг генов.
[0071] "Гомологичная рекомбинация" относится к взаимодействию между любой парой нуклеотидных последовательностей, содержащих соответствующие участки, содержащие аналогичную нуклеотидную последовательность, в которой могут взаимодействовать (рекомбинировать) две нуклеотидные последовательности с образованием новой рекомбинантной последовательности ДНК. Каждые участки аналогичной нуклеотидной последовательности обозначены в настоящем описании как "гомологичная последовательность". Как правило, частота гомологичной рекомбинации повышается, вследствие того, что длина гомологичной последовательности увеличивается. Таким образом, хотя гомологичная рекомбинация может возникать между двумя нуклеотидными последовательностями, которые являются в меньшей степени идентичными, частота рекомбинации (или эффективность) снижается, т.к. увеличивается дивергенция между двумя последовательностями. Рекомбинацию можно проводить с использованием одной гомологичной последовательности на каждой из донорных молекул и молекул-мишеней, таким образом, получая продукт рекомбинации "единичного кроссинговера". Альтернативно, две гомологичные последовательности могут располагаться на каждой из нуклеотидных последовательностей-мишеней и донорных нуклеотидных последовательностей. Рекомбинация между двумя гомологичными последовательностями в доноре с двумя гомологичными последовательностями в мишени образует продукт рекомбинации "двойного кроссинговера". Если гомологичные последовательности на донорной молекуле фланкируют последовательность, над которой необходимо проводить манипуляцию (например, представляющая интерес последовательность), рекомбинация двойного кроссинговера с молекулой-мишенью приводит к продукту рекомбинации, где представляющая интерес последовательность заменяет последовательностью ДНК, которая первоначально находилась между гомологичными последовательностями в молекуле-мишени. Обмен последовательностями ДНК между мишенью и донором посредством трансформационного события рекомбинации двойного кроссинговера называется "заменой последовательности".
[0072] Предпочтительное растение или семена по настоящему изобретению содержат в своем геноме функционирующие нуклеотидные последовательности aad-12, cry1F v3, cry1Ac synpro и pat v6, как определено в настоящем описании, совместно по меньшей мере с 20-500 или более смежных фланкирующих нуклеотидов на обеих сторонах вставки, как определено в настоящем описании. Если не указано иное, ссылка на фланкирующие последовательности относится к таким, как идентифицированным по отношению к SEQ ID NO:1, 2 и 15. Можно ожидать, что все или часть этих фланкирующих последовательностей передается потомству, которое получает встраиваемую ДНК в результате полового скрещивания родительской линии, которая содержит трансформационное событие.
[0073] Настоящее изобретение включает тканевые культуры регенерирующих клеток растения по настоящему изобретению. Также включены растения, регенерированные из таких тканевых культур, в частности, когда указанное растение способно проявлять все морфологические и физиологические свойства иллюстративного сорта. Предпочтительные растения по настоящему изобретению обладают всеми физиологическими и морфологическими характеристиками растения, выращенного из депонированных семян. Настоящее изобретение дополнительно включает потомство таких семян и семена, обладающие представляющими интерес качественными признаками.
[0074] Как используют в настоящем описании, "линия" представляет собой группу растений, для которых демонстрируют незначительную генетическую вариацию или не демонстрируют ее между индивидуумами по меньшей мере для одного признака. Такие линии можно получать путем нескольких генераций самоопыления и отбора или вегетативного размножения одного родителя способами культивирования тканей или клеток.
[0075] Как используют в настоящем описании, термины "культивар" и "сорт" являются синонимичными и относятся к линии, которую используют для коммерческой продукции.
[0076] "Стабильность" или "стабильный" означает, что в отношении данного компонента, компонент сохраняется от генерации к генерации и предпочтительно по меньшей мере три генерации.
[0077] "Коммерческую пригодность" определяют как наличие хорошей мощности растения и высокой плодовитости, такой что, культуру могут выращивать фермеры с использованием общепринятого сельскохозяйственного оборудования, и из семян можно экстрагировать масло с желаемыми компонентами с использованием общепринятого оборудования для измельчения и экстракции.
[0078] "Агрономически элитный" означает, что линия обладает желаемыми агрономическими характеристиками, такими как урожайность, зрелость, устойчивость к заболеваниям и т.п., в дополнение к устойчивости к инсектицидам и толерантности к гербицидам, вследствие рассматриваемого трансформационного события(й). Любую и все эти агрономические характеристики и измеряемые параметры можно использовать для идентификации таких растений в качестве точки или конца или обоих концов диапазона характеристик, используемых для определения таких растений.
[0079] Как специалисту в данной области будет понятно в свете настоящего описания предпочтительные варианты осуществления наборов для детекции, например, могут содержать зонды и/или праймеры, направленные и/или содержащие "соединительные последовательности" или "переходные последовательности" (где геномная фланкирующая последовательность сои находится рядом с последовательностью вставки). Например, они включают полинуклеотидные зонды, праймеры и/или ампликоны, сконструированные для идентификации одной или обеих соединительных последовательностей (где вставка находится рядом с фланкирующей последовательностью), как указано в таблице выше. Одна из общепринятых конструкций заключается в наличие одного праймера, который гибридизуется во фланкирующей области, и одного праймера, который гибридизуется во вставке. Длина каждого такого праймера часто составляет приблизительно по меньшей мере ~15 остатков. При такой конфигурации праймеры можно использовать для получения/амплификации детектируемого ампликона, который указывает на наличие трансформационного события по настоящему изобретению. Такие праймеры можно использовать для получения ампликона, который охватывает (и включает) соединительную последовательность, как указано выше.
[0080] Праймер(ы), "садящиеся" во фланкирующей последовательности, как правило, не конструируют для гибридизации дальше приблизительно 1200 оснований или дальше области соединения. Таким образом, иллюстративные фланкирующие праймеры конструируют так, чтобы они содержали по меньшей мере 15 остатков цепи в пределах 1200 оснований во фланкирующих последовательностях от начала вставки. Таким образом, праймеры, содержащие последовательность подходящего размера от (или гибридизующиеся с) от 800 до 1400 пар оснований SEQ ID NO:1 и/или от 153 до 753 пар оснований SEQ ID NO:2, и/или от 2130 до 2730 пар оснований SEQ ID NO:15, и/или от 9122 до 9722 пар оснований SEQ ID NO:15 входят в объем настоящего изобретения. Аналогично можно конструировать праймеры для вставки для любого участка вставки SEQ ID NO:3 или любого участка между парами оснований 2731 и 9121 SEQ ID NO:15, и можно использовать, например, неисключительно для конструирования таких праймеров.
[0081] Специалисту в данной области также известно, что праймеры и зонды можно конструировать для гибридизации в диапазоне стандартных условий гибридизации и/или ПЦР, где праймер или зонд не является абсолютно комплементарным иллюстративной последовательности. Таким образом, может быть допустима некоторая степень несоответствия. Для праймера приблизительно 20 нуклеотидов, например, как правило, один или два или т.д. нуклеотидов не должны связываться с противоположной цепью, если неподходящее основание располагается внутри или на концах праймера, который является противоположным по отношению к ампликону. Различные подходящие условия гибридизации приведены ниже. Синтетические аналоги нуклеотидов, такие как инозин, также можно использовать в зондах. Также можно использовать зонды на основе пептидных нуклеиновых кислот (ПНК), а также ДНК- и РНК-зонды. Важным является то, что такие зонды и праймеры являются диагностическими (способны уникально идентифицировать и определять) наличие трансформационного события по настоящему изобретению.
[0082] Следует отметить, что могут возникать ошибки амплификации ПЦР, которые могут приводить, например, к незначительным ошибкам секвенирования. Таким образом, если не указано иное, перечисленные в настоящем описании последовательности определяли, получая длинные ампликоны из геномных ДНК сои, а затем клонируя и секвенируя ампликоны. Обнаружение небольших различий и незначительных несоответствий последовательностей, получаемых и определяемых, таким образом, проводя много этапов амплификации, которая является необходимой для достаточного образования ампликона для секвенирования из геномных ДНК, не является необычным. Специалисту в данной области следует понимать и учитывать, что любые корректировки, необходимые, вследствие этих типов общих ошибок секвенирования или несоответствий, входят в объем настоящего изобретения.
[0083] Также следует отметить, что не является необычным удаление некоторой геномной последовательности, например, когда встраивают последовательность при создании трансформационного события. Таким образом, также могут возникать некоторые отличия между рассматриваемыми фланкирующими последовательностями и геномными последовательностями, перечисленными, например, в GENBANK.
[0084] Компоненты "вставки" последовательности ДНК проиллюстрированы на фигурах и более подробно описаны ниже в примерах. В способах по настоящему изобретению можно использовать полинуклеотидные последовательности ДНК этих компонентов или их фрагменты в качестве ДНК-праймеров или ДНК-зондов.
[0085] Некоторые варианты осуществления изобретения относятся к композициям и способам детекции наличия трансгенного/геномного участка вставки в растениях и семенах и т.п. от растения сои. Предоставлены последовательности ДНК, которые содержат рассматриваемую соединительную последовательность 5'-трансгенного/геномного участка вставки, предоставленную в настоящем описании (между от 800 до 1400 пар оснований SEQ ID NO:1 и SEQ ID NO:3), ее сегменты и комплементы иллюстративных последовательностей, и любые их сегменты. Предоставлены последовательности ДНК, которые содержат рассматриваемую соединительную последовательность 3'-трансгенного/геномного участка вставки, предоставленную в настоящем описании (между от 153 до 753 пар оснований SEQ ID NO:2 и SEQ ID NO:3), ее сегменты и комплементы иллюстративных последовательностей и любые их сегменты. Предоставлены последовательности ДНК, которые содержат рассматриваемую соединительную последовательность 3'-трансгенного/геномного участка вставки, предоставленную в настоящем описании (между от 1 до 2730 пар оснований SEQ ID NO:15 и 2731 до 9121 SEQ ID NO:15), ее сегменты и комплементы иллюстративных последовательностей и любые их сегменты. Предоставлены последовательности ДНК, которые содержат соединительную последовательность, при условии рассматриваемую соединительную последовательность 3'-трансгенного/геномного участка вставки, предоставленную в настоящем описании (между от 9122 до 10198 пар оснований SEQ ID NO:15 и от 2731 до 9121 SEQ ID NO:15), ее сегменты и комплементы иллюстративных последовательностей и любые их сегменты. Соединительная последовательность участка вставки охватывает соединение между гетерологичной ДНК, встраиваемой в геном, и ДНК из клетки сои, фланкирующую участок вставки. Такие последовательности можно диагностировать для данного трансформационного события.
[0086] В зависимости от этих встраиваемых и граничных последовательностей можно получать специфичные по отношению к трансформационному событию праймеры. Анализ ПЦР продемонстрировал, что линии сои по настоящему изобретению можно идентифицировать в различных генотипах сои анализом ампликонов ПЦР, получаемых с использованием наборов таких специфичных по отношению к трансформационному событию праймеров. Эти и другие связанные способы можно использовать для уникальной идентификации таких линий сои. Таким образом, ампликоны ПЦР, получаемые с такими парами праймеров являются уникальными, и их можно использовать для идентификации таких линий сои.
[0087] В некоторых вариантах осуществления последовательности ДНК, которые содержат смежный фрагмент нового трансгенного/геномного участка, представляют собой один из аспектов настоящего изобретения. Включены последовательности ДНК, которые имеют достаточную длину полинуклеотидов трансгенной последовательности вставки и достаточную длину полинуклеотидов геномной последовательности сои от одного или более из трех указанных выше растений сои, и/или последовательности, которые являются пригодными в качестве последовательностей праймера для получения продукта-ампликона, диагностического для одного или более таких растений сои.
[0088] Родственные варианты осуществления относятся к последовательности ДНК, которая содержит по меньшей мере 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более смежных нуклеотидов трансгенного участка последовательности ДНК, определяемой в настоящем описании (такой как SEQ ID NO:1 и ее сегменты) или их комплементов, и имеет аналогичную длину фланкирующей последовательности ДНК сои из таких последовательностей или их комплементов. Такие последовательности являются пригодными в качестве ДНК-праймеров в способах амплификации ДНК. Ампликоны, получаемые с использованием таких праймеров являются диагностическими для любого из трансформационных событий сои, указанных в настоящем описании. Таким образом, изобретение также включает ампликоны, образуемые такими ДНК-праймерами и гомологичными праймерами.
[0089] Это изобретение также включает способы детекции наличия ДНК в образце, которая соответствует трансформационному событию сои, указанному в настоящем описании. Такие способы могут включать: a) контактирование образца, содержащего ДНК, с набором праймеров, который при использовании в реакции амплификации нуклеиновой кислоты с ДНК по меньшей мере от одного из этих трансформационных событий сои, образует ампликон, который является диагностическим для указанного трансформационного события(й); b) проведение реакции амплификации нуклеиновых кислот, таким образом получая ампликон, и c) детекцию ампликона.
[0090] Дополнительные способы детекции по настоящему изобретению включают способ детекции наличия ДНК в образце, соответствующей указанному трансформационному событию, где указанный способ включает: a) контактирование образца, содержащего ДНК, с зондом, который гибридизуется в жестких условиях гибридизации с ДНК по меньшей мере из одного из указанных трансформационных событий сои и который не гибридизуется в жестких условиях гибридизации с контрольным растением сои (не трансформационное событие, представляющей интерес ДНК), b) подвергание образца и зонда жестким условиям гибридизации и c) детекция гибридизации зонда с ДНК.
[0091] В дополнительных вариантах осуществления настоящее изобретение включает способы получения растения сои, содержащего трансформационное событие сои pDAB9582.814.19.1::pDAB4468.04.16.1 по настоящему изобретению, где указанный способ включает этапы: a) полового скрещивания первой родительской линии сои (содержащей экспрессирующую кассету по настоящему изобретению, которая обеспечивает толерантность растений указанной линии к глуфосинату) и второй родительской линии сои (у которой отсутствует такой признак толерантности к гербициду), таким образом, получая ряд растений потомства, и b) отбор растения потомства с использованием молекулярных маркеров. Такие способы могут необязательно включать дополнительный этап обратного скрещивания растения потомства со второй родительской линией сои с получением чистосортного растения сои, которое содержит признак устойчивости к насекомым-вредителям 2,4-D и толерантности к глуфосинату.
[0092] В другом аспекте изобретение относится к способам определения зиготности потомства от скрещивания с указанным трансформационным событием. Указанные способы могут включать контактирование образца, содержащего ДНК сои, с набором праймеров по настоящему изобретению. Указанные праймеры при использовании в реакции амплификации нуклеиновых кислот с геномной ДНК по меньшей мере из одного из указанных трансформационных событий сои образуют первый ампликон, который является диагностическим по меньшей мере для одного из указанных трансформационных событий сои. Такие способы дополнительно включают проведение реакции амплификации нуклеиновых кислот, таким образом, получая первый ампликон; детекцию первого ампликона и контактирование образца, содержащего ДНК сои со вторым набором праймеров (указанный второй набор праймеров при использовании в реакции амплификации нуклеиновых кислот с геномной ДНК из растения сои образует второй ампликон, содержащий нативную геномную ДНК сои, гомологичную геномному участку сои) и проведение реакции амплификации нуклеиновых кислот, таким образом, получая второй ампликон. Способы дополнительно включают детекцию второго ампликона и сравнение первого и второго ампликонов в образце, где наличие обоих ампликонов свидетельствует о том, что образец является гетерозиготным по трансгенной вставке.
[0093] Наборы для детекции ДНК можно разрабатывать с использованием композиций, описываемых в настоящем изобретении, и способы являются известными в области детекции ДНК. Наборы являются пригодными для идентификации ДНК рассматриваемого трансформационного события сои в образце, и их можно применять в способах селекции растения сои, содержащего такую ДНК. Наборы содержат последовательности ДНК, гомологичные или комплементарные ампликонам, например, описываемым в настоящем описании, или последовательностям ДНК, гомологичным или комплементарным ДНК, содержащейся в трансгенных генетических элементах рассматриваемых трансформационных событий. Эти последовательности ДНК можно использовать в реакциях амплификации ДНК или в качестве зондов в способе гибридизации ДНК. Наборы также могут содержать реагенты и вещества, необходимые для проведения способа детекции.
[0094] "Зонд" представляет собой выделенную молекулу нуклеиновой кислоты, к которой присоединяют общепринятую детектируемую метку или репортерную молекулу (такую как радиоактивный изотоп, лиганд, хемилюминесцентное средство или фермент). Такой зонд является комплементарным цепи нуклеиновой кислоты-мишени, в случае по настоящему изобретению цепи геномной ДНК из одного из указанных трансформационных событий сои, неважно из растения сои или из образца, который содержит ДНК из трансформационного события. Зонды по настоящему изобретению включают не только дезоксирибонуклеиновые или рибонуклеиновые кислоты, а также полиамиды и другие вещества для зонда, которые специфически связываются с последовательностью ДНК-мишени, и их можно использовать для детекции наличия последовательности ДНК-мишени.
[0095] "Праймеры" представляют собой выделенные/синтезированные нуклеиновые кислоты, которые отжигаются на комплементарной цепи ДНК-мишени при гибридизации нуклеиновой кислоты с образованием гибрида между праймером и цепью ДНК-мишени, затем удлиняются вдоль цепи ДНК-мишени полимеразой, например, ДНК-полимеразой. Пары праймеров по настоящему изобретению относятся к их применению для амплификации последовательности нуклеиновой кислоты-мишени, например, посредством полимеразной цепной реакции (ПЦР) или другими общепринятыми способами амплификации нуклеиновой кислоты.
[0100] Длина зондов и праймеров, как правило, составляет 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500, или 1000, или 2000, или 5000 полинуклеотидов или более. Такие зонды и праймеры специфически гибридизуются с последовательностью-мишенью в высоко жестких условиях гибридизации. Предпочтительно зонды и праймеры по настоящему изобретению обладают полным сходством последовательности с последовательностью-мишенью, хотя можно конструировать зонды, отличающиеся от последовательности-мишени, и которые сохраняют способность гибридизоваться с последовательностями-мишенями общепринятыми способами.
[0101] Способы получения и применения зондов и праймеров описаны, например, в Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989. Пары праймеров для ПЦР можно получать на основании известной последовательности, например, с использованием компьютерных программ, предназначенных для таких целей.
[0102] Праймеры и зонды на основании фланкирующих последовательностей ДНК и последовательностей ДНК вставки, описываемые в настоящем описании, можно использовать для подтверждения (и при необходимости для коррекции) описываемых последовательностей общепринятыми способами, например, посредством повторного клонирования и секвенирования таких последовательностей.
[0103] Зонды и праймеры на основе нуклеиновых кислот по настоящему изобретению гибридизуются в жестких условиях с последовательностью ДНК-мишени. Любой общепринятый способ гибридизации или амплификации нуклеиновых кислот можно использовать для идентификации наличия ДНК из трансгенного трансформационного события в образце. Молекулы нуклеиновой кислоты или их фрагменты способны специфически гибридизоваться с другими молекулами нуклеиновой кислоты при определенных обстоятельствах. Как используют в настоящем описании, считается, что две молекулы нуклеиновой кислоты способны специфически гибридизоваться друг с другом, если две молекулы способны образовывать антипараллельную двухцепочечную структуру нуклеиновой кислоты. Считается, что молекула нуклеиновой представляет собой "комплемент" другой молекулы нуклеиновой кислоты, если они обладают полной комплементарностью. Как используют в настоящем описании, считается, что молекулы обладают "полной комплементарностью", когда каждый нуклеотид одной из молекул является комплементарным нуклеотиду другой молекулы. Считается, что две молекулы являются "минимально комплементарными", если они могут гибридизоваться друг с другом с достаточной стабильностью, которая позволяет им сохранять способность отжигаться друг с другом по меньшей мере в общепринятых условиях "пониженной жесткости". Аналогично, считается, что молекулы являются "комплементарными", если они могут гибридизоваться друг с другом с достаточной стабильностью, которая позволяет им сохранять способность отжигаться друг с другом в общепринятых условиях "пониженной жесткости". Общепринятые условия жесткости описаны Sambrook et al., 1989. Таким образом, отклонения от полной комплементарности являются допустимыми при условии, что такие отклонения не препятствуют полностью способности молекулы образовывать двухцепочечную структуру. Для того чтобы молекула нуклеиновой кислоты служила в качестве праймера или зонда, ей только необходимо являться достаточно комплементарной по последовательности, чтобы являться способной образовывать стабильную двухцепочечную структуру при конкретных применяемых концентрациях растворителя и солей.
[0104] Как используют в настоящем описании, по существу гомологичная последовательность представляет собой последовательность нуклеиновой кислоты, которая специфически гибридизуется с комплементом последовательности нуклеиновой кислоты, к которой она принадлежит, при сравнении в условиях с высокой жесткостью. Границу "жестких условий" функционально определяют в отношении гибридизации зонда на основе нуклеиновой кислоты с нуклеиновой кислотой-мишенью (т.е. с конкретной представляющей интерес последовательностью нуклеиновой кислоты) способом специфической гибридизации, описанным у Sambrook et al., 1989, в 9.52-9.55. См. также Sambrook et al., 1989 в 9.47-9.52 и 9.56-9.58. Таким образом, нуклеотидные последовательности по изобретению можно использовать на основании их способности избирательно образовывать дуплексные молекулы с комплементарными участками фрагментов ДНК.
[0105] В зависимости от предусматриваемого применения можно использовать различные условия гибридизации для получения различных степеней селективности зонда по отношению к последовательности-мишени. Для применений, где необходима высокая селективность, как правило, применяют относительно жесткие условия для образования гибридов, например, выбирают условия относительно низкого содержания соли и/или высокой температуры, такие как получаемые приблизительно от 0,02M приблизительно до 0,15M NaCl при температурах приблизительно от 50°C приблизительно до 70°C. Жесткие условия, например, могут включать промывание гибридизационного фильтра по меньшей мере два раза буфером для промывания в высоко жестких условиях (0,2X SSC, 0,1% SDS, 65°C). Специалистам в данной области известны подходящие условия жесткости, которые способствуют гибридизации ДНК, например, 6,0X хлорид натрия/цитрат натрия (SSC) приблизительно при 45°C с последующим промыванием 2,0X SSC при 50°C. Например, концентрацию солей на этапе промывания можно выбирать от низкой жесткости приблизительно 2,0X SSC при 50°C до высокой жесткости приблизительно 0,2X SSC при 50°C. Кроме того, температуру на этапе промывания можно повышать от условий низкой жесткости при комнатной температуре приблизительно 22°C до условий с высокой жесткостью приблизительно при 65°C. Температуру и содержание соли можно изменять, или можно поддерживать постоянной температуру или концентрацию солей при изменении другой переменной. Такие избирательные условия мало допустимы, если существует любое несоответствие между пробой и матрицей или цепью-мишенью. Детекция последовательностей ДНК посредством гибридизации хорошо известна специалистам в данной области, и указания в патентах США №№ 4965188 и 5176995 представляют собой иллюстративные способы гибридизационных анализов.
[0106] В особенно предпочтительном варианте осуществления нуклеиновая кислота по настоящему изобретению специфически гибридизуется с одним или более праймеров (или ампликонов, или других последовательностей), иллюстрируемых или рассматриваемых в настоящем описании, включая их комплементы и фрагменты, в условиях с высокой жесткостью. В одном из аспектов настоящего изобретения маркерная молекула нуклеиновой кислоты по настоящему изобретению содержит последовательность нуклеиновой кислоты, как указано в настоящем описании в одной из иллюстративных последовательностей или их комплементов и/или фрагментов.
[0107] В другом аспекте настоящего изобретения маркерная молекула нуклеиновой кислоты по настоящему изобретению обладает от 80% до 100% или 90% и 100% идентичностью последовательности с такими последовательностями нуклеиновых кислот. В дополнительном аспекте настоящего изобретения маркерная молекула нуклеиновой кислоты по настоящему изобретению обладает от 95% до 100% идентичностью последовательности с такой последовательностью. Такие последовательности можно использовать в качестве маркеров в способах селекции растений для идентификации потомства генетических скрещиваний. Гибридизацию пробы с молекулой ДНК-мишенью можно детектировать любым из ряда способов, известных специалистам в данной области, они могут включать, но не ограничиваться ими, флуоресцентные метки, радиоактивные метки, метки на основе антитела и хемилюминесцентные метки.
[0108] В отношении амплификации последовательности нуклеиновой кислоты-мишени (например, ПЦР) с использованием конкретной пары праймеров для амплификации "жесткие условия" представляют собой условия, которые обеспечивают возможность гибридизоваться паре праймеров только с последовательностью нуклеиновой кислоты-мишени, с которой связывается праймер, содержащий соответствующую последовательность дикого типа (или ее комплемент), и предпочтительно с получением уникального продукта амплификации, ампликона.
[0109] Термин "специфичный по отношению к (последовательности-мишени)" означает, что зонд или праймер гибридизуется в наиболее жестких условиях гибридизации только с последовательностью-мишенью в образце, содержащем последовательность-мишень.
[0110] Как используют в настоящем описании, "амплифицированная ДНК" или "ампликон" относится к продукту амплификации последовательности нуклеиновой кислоты-мишени, которая представляет собой часть нуклеиновой кислоты-матрицы. Например, для определения, содержит ли растение сои, получаемое в результате полового скрещивания, геномную ДНК с трансгенным трансформационным событием из растения сои по настоящему изобретению, ДНК, выделяемую из образца ткани растения сои, можно подвергать способу амплификации нуклеиновой кислоты с использованием пары праймеров, которая содержит праймер, получаемый из фланкирующей последовательности в геноме растения, прилежащей к участку вставки встраиваемой гетерологичной ДНК, и второй праймер, получаемый из встраиваемой гетерологичной ДНК, с получением ампликона, который является диагностическим для наличия ДНК с трансформационным событием. Ампликон имеет длину и содержит последовательность, которая также является диагностической для трансформационного события. Длина ампликона может находиться в диапазоне от объединенной длины пары праймеров плюс одна пара оснований нуклеотидов и/или объединенной длины пары праймеров плюс приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, or 500, 750, 1000, 1250, 1500, 1750, 2000 или более пар оснований нуклеотидов (плюс или минус любой из инкрементов, перечисленных выше). Альтернативно, пару праймеров можно получать из фланкирующей последовательности на обеих сторонах встраиваемой ДНК таким образом, чтобы образовывался ампликон, который содержит полную нуклеотидную последовательность вставки. Член пары праймеров, получаемых из геномной последовательности растения, можно располагать на расстоянии от встраиваемой последовательности ДНК. Это расстояние может находиться в диапазоне от одной пары оснований нуклеотидов приблизительно до двадцати тысяч пар оснований нуклеотидов. Использование термина "ампликон" конкретно исключает димеры праймеров, которые могут образовываться в термической реакции амплификации ДНК.
[0111] Амплификацию нуклеиновой кислоты можно проводить любым из ряда известных в данной области способов амплификации нуклеиновой кислоты, включая полимеразную цепную реакцию (ПЦР). Ряд способов амплификации известен в данной области и описан в числе прочего в патенте США № 4683195 и патенте США № 4683202. ПЦР способы амплификации были разработаны для амплификации до 22 т.п.н. геномной ДНК. Эти способы, а также другие известные в данной области способы амплификации ДНК, можно использовать в практическом осуществлении настоящего изобретения. Последовательность вставки гетерологичной трансгенной ДНК или фланкирующую геномную последовательность из рассматриваемого трансформационного события сои можно подтверждать (и корректировать при необходимости) посредством амплификации таких последовательностей из трансформационного события с использованием праймеров, получаемых из последовательностей, предоставленных в настоящем описании, с последующим стандартным секвенированием ДНК ПЦР-ампликона или клонированной ДНК.
[0112] Ампликон, получаемый такими способами, можно детектировать многими способами. Электрофорез в агарозном геле и окрашивание бромистым этидием представляет собой хорошо известный способ детекции ампликонов ДНК. Другой такой способ представляет собой анализ генетических битов (Genetic Bit Analysis), где конструируют олигонуклеотид ДНК, который перекрывает прилежащую фланкирующую последовательность геномной ДНК и последовательность встраиваемой ДНК. Олигонуклеотид иммобилизуют в лунках планшета с микролунками. После ПЦР представляющего интерес участка (с использованием одного праймера во встраиваемой последовательности и одного в прилежащей фланкирующей геномной последовательности) одноцепочечный продукт ПЦР можно подвергать гибридизации с иммобилизованным олигонуклеотидом, и он служит в качестве матрицы для реакции удлинения цепи на одно основание с использованием ДНК-полимеразы и меченых ddNTP, специфичных для удлиняемого следующего основания. Считывание показаний может быть основано на флуоресценции или ELISA. Сигнал означает наличие последовательности вставки/фланкирующей последовательности в результате успешной амплификации, гибридизации и удлинения цепи на одно основание.
[0113] Другой способ представляет собой способ пиросеквенирования, как описано Winge (Innov. Pharma. Tech., 00:18-24, 2000). В этом способе конструируют олигонуклеотид, который перекрывает область соединения прилежащей геномной и встраиваемой ДНК. Олигонуклеотид гибридизуют с одноцепочечным продуктом ПЦР от участка вставки (один праймер во встраиваемой последовательности и один во фланкирующей геномной последовательности) и инкубируют в присутствии ДНК-полимеразы, АТФ, сульфурилазы, люциферазы, апиразы, аденозин-5'-фосфосульфата и люциферина. Отдельно добавляют dNTP, и встраивание приводит к световому сигналу, который измеряют. Световой сигнал свидетельствует о наличие трансгенной вставки/фланкирующей последовательности в результате успешной амплификации, гибридизации и удлинения цепи на одно или много оснований.
[0114] Флуоресцентная поляризация представляет собой другой способ, который можно использовать для детекции ампликона по настоящему изобретению. В соответствии с этим способом конструируют олигонуклеотид, который перекрывает область соединения геномной фланкирующей и встраиваемой ДНК. Олигонуклеотид гибридизуют с одноцепочечным продуктом ПЦР из представляющего интерес участка (один праймер во встраиваемой ДНК и один во фланкирующей геномной последовательности ДНК) и инкубируют в присутствии ДНК-полимеразы и флуоресцентно меченого ddNTP. Удлинение цепи на одно основание приводит к встраиванию ddNTP. Встраивание можно измерять как изменение поляризации с использованием флуорометра. Изменение поляризации свидетельствует о наличие трансгенной вставки/фланкирующей последовательности в результате успешной амплификации, гибридизации и удлинения цепи на одно основание.
[0115] TAQMAN® (PE Applied Biosystems, Foster City, Calif.) представляет собой способ детекции и количественного определения наличия последовательности ДНК. В кратком изложении, конструируют олигонуклеотидный зонд FRET, который перекрывает область соединения геномной фланкирующей ДНК и области вставки. Зонд FRET и праймеры для ПЦР (один праймер в последовательности ДНК ставки и один во фланкирующей геномной последовательности) подвергают циклам гибридизации в присутствии термостабильной полимеразы и dNTP. Во время специфической амплификации ДНК-полимераза Taq отщепляется, и в зонде FRET флуоресцентная группа отделяется от гасящей группы. Флуоресцентный сигнал свидетельствует о наличии фланкирующей/трансгенной последовательности вставки в результате успешной амплификации и гибридизации.
[0116] Описаны молекулярные маяки для применения при детекции последовательности. В кратком изложении, конструируют олигонуклеотидный зонд FRET, который перекрывает область соединения фланкирующей геномной и встраиваемой ДНК. Уникальная структура зонда FRET приводит к тому, что он содержит вторичную структуру, которая удерживает флуоресцентную и гасящую группы в непосредственной близости. Зонд FRET и праймеры для ПЦР (один праймер во встраиваемой последовательности ДНК и один во фланкирующей геномной последовательности) подвергают циклам гибридизации в присутствии термостабильной полимеразы и dNTP. После успешной ПЦР амплификации гибридизация зонда FRET с последовательностью-мишенью приводит к удалению вторичной структуры зонда и к пространственному разделению флуоресцентной и гасящей группы. В результате этого возникает флуоресцентный сигнал. Флуоресцентный сигнал свидетельствует о наличии фланкирующей геномной/трансгенной последовательности вставки в результате успешной амплификации и гибридизации.
[0117] Наряду с описанием локализации в геноме сои, которая является превосходной для встраивания, настоящее изобретение также относится к семенам сои и/или растению сои, содержащему по меньшей мере одну вставку, не являющуюся трансформационным событием сои pDAB9582.814.19.1::pDAB4468.04.16.1, в непосредственной близости к такой локализации в геноме. Один из вариантов заключается в замене отличной вставки вместо вставок из трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1, проиллюстрированного в настоящем описании. В отношении этого согласно настоящему изобретению можно использовать, например, направленную гомологичную рекомбинацию. Этот тип технологии представляет собой объект, например, WO 03/080809 A2 и соответствующей опубликованной заявки США (US 20030232410). Таким образом, настоящее изобретение включает растения и растительные клетки, содержащие гетерологичную вставку (вместо или совместно с многокопийными генами aad-12, cry1F, cry1Ac или pat), фланкированную полными или распознаваемой частью фланкирующих последовательностей, определяемых в настоящем описании (1-1400 п.н. SEQ ID NO:1 и 153-1550 п.н. SEQ ID NO:2 и 1-2730 п.н. SEQ ID NO:15 и 9122-10198 п.н. SEQ ID NO:15). Дополнительная копия (или дополнительные копии) aad-12, Cry1F, cry1Ac или pat также можно направлять для встраивания таким/такими способом(ами).
[0118] Все патенты, патентные заявки, предварительные заявки и публикации, на которые ссылаются, или которые цитируют в настоящем описании, полностью включены посредством ссылки в той мере, в которой они не противоречат явно указанным сведениям настоящего описания.
[0119] Следующие ниже примеры включены для иллюстрации способов практического осуществления изобретения и для демонстрации определенных предпочтительных вариантов осуществления изобретения. Эти примеры не следует интерпретировать как ограничивающие. Специалистам в данной области следует понимать, что способы, описываемые в следующих ниже примерах, представляют собой конкретные подходы, используемые для иллюстрации предпочтительных способов их практического осуществления. Однако в свете настоящего описания специалистам в данной области следует понимать, что в этих конкретных вариантах осуществления можно проводить многие изменения, при этом все еще получая подобные или аналогичные результаты, не выходя за рамки сущности и объема изобретения. Если не указано иное, все проценты являются массовыми, и все пропорции смеси растворителей являются объемными, если не указано иное.
[0120] Используют следующие ниже сокращенные обозначения, если не указано иное.
|
|
[0121] Варианты осуществления настоящего изобретения дополнительно описаны в следующих ниже примерах. Следует понимать, что эти примеры приведены только в качестве иллюстрации. Из указанного выше обсуждения и этих примеров специалист в данной области может установить основные признаки настоящего изобретения и, не выходя за рамки его сущности и объема, может проводить различные изменения и модификации вариантов осуществления изобретения для его адаптации к различному использованию и условиям. Таким образом, различные модификации вариантов осуществления изобретения в дополнение к проиллюстрированным и описываемым в настоящем описании станут понятны специалистам в данной области из указанного выше описания. Подразумевают, что такие модификации также входят в объем прилагаемой формулы изобретения.
[0122] Описание каждой ссылки, указанной в настоящем описании, полностью включено в настоящее описание посредством ссылки.
ПРИМЕРЫ
Пример 1: Трансформация и отбор трансформационного события сои pDAB9582.814.19.1 Cry1F и Cry1Ac
[0123] Трансгенную сою (Glycine max), содержащую трансформационное событие сои pDAB9582.814.19.1 получали посредством опосредованной Agrobacterium трансформации эксплантатов семядольного узла сои. Для инициации трансформации использовали обезвреженный штамм Agrobacterium EHA101 (Hood et al., 1993), несущий бинарный вектор pDAB9582 (фиг.1), содержащий селектируемый маркер, pat v6 и представляющие интерес гены cry1F v3 и cry1Ac synpro, в области T-цепи ДНК. Последовательность ДНК для pDAB9582 приведена в SEQ ID NO:3, которая аннотирована ниже в таблице 1.
[0124]
|
[0125] Опосредованную Agrobacterium трансформацию проводили модифицированным способом Zeng et al. (2004). В кратком изложении, семена сои (cv Maverick) проращивали на базальной среде и выделяли семядольные узлы, и инфицировали Agrobacterium. В среду для закладки побега, роста побега в длину и укоренения добавляли цефотаксим, тиментин и ванкомицин для удаления Agrobacterium. Для ингибирования роста нетрансформированных побегов применяли отбор с использованием глуфосината. Отобранные побеги переносили в среду для укоренения для развития корней, а затем переносили в почвенную смесь для акклиматизации рассады.
[0126] Верхушечные листочки отобранных растений представляли собой лист, с нанесенным глуфосинатом для скрининга предполагаемых трансформантов. Прошедшие скрининг рассады переносили в теплицу, оставляли для акклиматизации, а затем наносили на листья с глуфосинатом для повторного подтверждения толерантности и считали их предполагаемыми трансформантами. У прошедших скрининг растений отбирали образец и проводили молекулярные анализы для подтверждения гена селективного маркера и/или представляющего интерес гена. Растениям обеспечивали возможность самоопыления в теплице для увеличения семян T1.
[0127] Это трансформационное событие, трансформационное событие сои pDAB9582.814.19.1, получали из независимо трансформированного изолята. Растения T1 подвергали обратному скрещиванию и проводили интрогрессию в элитные сорта в течение последующих поколений. Трансформационное событие выбирали на основании его уникальных характеристик, таких как один участок вставки, нормальное менделеевское расщепление, стабильная экспрессия и превосходная комбинация эффективности, включая толерантность к гербициду и агрономическую продуктивность. Следующие ниже примеры содержат данные, которые использовали для характеристики трансформационного события сои pDAB9582.814.19.1.
Пример 2: Характеристика экспрессии белка в трансформанте сои pDAB9582.814.19.1
[0128] Характеризовали биохимические свойства рекомбинантных белков Cry1F, Cry1Ac и PAT, экспрессируемых в трансформанте сои pDAB9582.814.19.1. Количественные твердофазный иммуноферментный анализ (ELISA) представляет собой известный в данной области биохимический анализ, который можно использовать для характеристики биохимических свойств белков и подтверждать экспрессию этих белков в трансформанте сои pDAB9582.814.19.1.
Пример 2.1: Экспрессия белка PAT, Cry1F и Cry1Ac в тканях растений
[0129] Образцы тканей сои выделяли из тестируемых растений и подготавливали для анализа экспрессии. Белок PAT экстрагировали из тканей растения сои раствором фосфатно-солевого буфера, содержащим детергент Tween-20 (PBST), содержащий 0,5% бычьего сывороточного альбумина (BSA). Растительную ткань центрифугировали, собирали водный супернатант, разбавляли подходящим буфером по мере необходимости и анализировали с использованием набора ELISA для определения PAT в формате "сэндвич". Набор использовали согласно протоколу, рекомендуемому производителем (Envirologix, Portland, ME). В этом анализе измеряли экспрессируемый белок PAT.
[0130] Белок Cry1F экстрагировали из тканей растения сои раствором фосфатно-солевого буфера, содержащим детергент Tween-20 (PBST). Растительную ткань центрифугировали, собирали водный супернатант, разбавляли подходящим буфером по мере необходимости и анализировали с использованием набора ELISA для определния Cry1F в формате "сэндвич". Набор использовали согласно протоколу, рекомендуемому производителем (Strategic Diagnostics Inc., Newark, DE). В этом анализе измеряли экспрессируемый белок Cry1F.
[0131] Белок Cry1Ac экстрагировали из тканей растения сои раствором фосфатно-солевого буфера, содержащим детергент Tween-20 (PBST), содержащий 0,5% бычьего сывороточного альбумина (BSA). Растительную ткань центрифугировали, собирали водный супернатант, разбавляли подходящим буфером по мере необходимости и анализировали с использованием набора ELISA для определения Cry1Ac в формате "сэндвич". Набор использовали согласно протоколу, рекомендуемому производителем (Strategic Diagnostics Inc., Newark, DE). В этом анализе измеряли экспрессируемый белок Cry1Ac.
[0132] Анализ детекции проводили для исследования стабильности экспрессии и наследуемости как вертикально (между поколениями), так и горизонтально (между линиями в поколении) в трансформанте сои pDAB9582.814.19.1.
Пример 2.2: Экспрессия белка PAT, Cry1F и Cry1Ac в тканях растений
[0133] Определяли уровни белков Cry1F, Cry1Ac и PAT в трансформанте сои pDAB9582.814.19.1. Растворимые, экстрагируемые белки из ткани листьев сои измеряли способом количественного твердофазного иммуноферментного анализа (ELISA). От T2 до T6 поколений трансформантов сои pDAB9582.814.19.1 экспрессия являлась стабильной (не расщепленной) и однородной для всех линий. В таблице 2 приведен средний уровень экспрессии трансгенных белков в трансформанте сои pDAB9582.814.19.1.
|
Пример 3: Клонирование и характеристика последовательности ДНК во вставке и фланкирующих граничных участках трансформационного события сои pDAB9582.814.19.1
[0134] Для характеристики и описания геномного участка вставки определяли последовательность фланкирующих геномных граничных участков Т-ДНК трансформационного события сои pDAB9582.814.19.1. Подтверждали геномную последовательность трансформационного события сои pDAB9582.814.19.1, содержащую 1400 п.н. 5'-фланкирующей граничной последовательности (SEQ ID NO:1) и 1398 п.н. 3'-фланкирующей граничной последовательности (SEQ ID NO:2). ПЦР амплификация на основе граничных последовательностей трансформационного события сои pDAB9582.814.19.1 подтвердила, что граничные участки являлись соевой природы, и что участки соединения представляют собой уникальные последовательности для трансформационного события сои pDAB9582.814.19.1. Участки соединения можно использовать для специфической в отношении трансформационного события идентификации трансформационного события сои pDAB9582.814.19.1. Кроме того, характеризовали участок вставки T-цепи путем амплификации геномного фрагмента, соответствующего участку идентифицируемых фланкирующих граничных последовательностей из генома нетрансформированной сои. Сравнением трансформационного события сои pDAB9582.814.19.1 с нетрансформированной геномной последовательностью выявляли, что во время интеграции T-цепи происходила делеция приблизительно 57 п.н. из исходного локуса. В целом характеристика вставки и граничной последовательности трансформационного события сои pDAB9582.814.19.1 указывала на то, что в геноме сои содержалась интактная копия T-цепи из pDAB9582.
|
|
|
Пример 3.1: Подтверждение геномных последовательностей сои
[0135] 5'- и 3'-фланкирующие границы выравнивали по последовательности, получаемой при полногеномном секвенировании способом дробовика хромосомы 02 Glycine max, подтверждая, что трансген трансформационного события сои pDAB9582.814.19.1 встраивался в геном хромосомы 02 сои. Для подтверждения участка вставки трансформационного события сои pDAB9582.814.19.1 из генома сои проводили ПЦР с различными парами праймеров (фиг.2, таблица 3, таблица 4 и таблица 5). В качестве матрицы использовали геномную ДНК из трансформационного события сои pDAB9582.814.19.1 и другие трансгенные или нетрансгенные линии сои. Для подтверждения, что 5'-граничные последовательности являются правильными, использовали праймер, сконструированный для связывания с промоторным элементом гена AtUbi10, например, AtUbi10RV1, и праймер, сконструированный для связывания с клонируемой 5'-концом границы хромосомы 02 генома сои, праймер, обозначаемый 81419_FW3, для амплификации сегмента ДНК, который перекрывает промоторный элемент гена AtUbi10 в 5'-конце граничной последовательности. Аналогично, для подтверждения клонированной 3'-граничной последовательности использовали специфический в отношении pat праймер, например, 3'PATEnd05, и три праймера, сконструированных в соответствии с клонированным 3'-концом граничной последовательности, обозначаемых 81419_RV1, 81419_RV2 и 81419_RV3, для амплификации сегментов ДНК, которые перекрывает ген pat до 3'-граничной последовательности. Фрагменты ДНК с ожидаемыми размерами амплифицировали только из геномной ДНК трансформанта сои pDAB9582.814.19.1 с каждой парой праймеров, но не из образцов ДНК от других трансгенных линий сои или нетрансгенного контроля. Результаты свидетельствуют, что клонированные 5'- и 3'-граничные последовательности представляют собой фланкирующие граничные последовательности вставки T-цепи из трансформационного события сои pDAB9582.814.19.1.
[0136] Для дополнительного подтверждения встраивания ДНК в геном сои проводили ПЦР амплификацию, перекрывающую граничные последовательности сои, на геномной ДНК, которая не содержала вставку T-цепи из трансформационного события сои pDAB9582.814.19.1. Праймер 81419_FW3, сконструированный в соответствии с 5'-концом граничной последовательности, и один праймер 81419_RV3, сконструированный для 3'-конца граничной последовательности, использовали для амплификации сегментов ДНК, которые содержали локусы, в которые интегрировали T-цепь pDAB9582. Как и ожидалось, ПЦР амплификацией, проводимой с парой праймеров 81419_FW3 и 81419_RV3, получали фрагмент ДНК приблизительно 1,5 т.п.н. от всех других контрольных линий сои, но не pDAB9582.814.19.1. Выравниванием идентифицированных 5'- и 3'-граничных последовательностей трансформационного события сои pDAB9582.814.19.1 с последовательностью, получаемой при полногеномном секвенировании способом дробовика, из хромосомы 02 Glycine max выявляли делецию приблизительно 57 п.н. в исходном локусе (фигура 3). Эти результаты демонстрировали, что трансген трансформационного события сои pDAB8294 встраивался в участок хромосомы 02 генома сои.
Пример 4: Характеристика посредством саузерн-блоттинга трансформационного событие сои pDAB9582.814.19.1
[0137] Анализ саузерн-блоттинг использовали для установления паттерна интеграции трансформационного события сои pDAB9582.814.19.1. Этими экспериментами получали данные, которые демонстрировали, что интеграция и целостности трансгенов cry1Ac и cry1F в геноме сои. Трансформационное событие сои pDAB9582.814.19.1 характеризовали как первичное простое трансформационное событие интеграции, содержащее одну копию единицы транскрипции (PTU) растения cry1Ac и cry1F из плазмиды pDAB9582.
[0138] Данные саузерн-блоттинга подтверждали, что фрагмент T-цепи встраивался в геном трансформанта сои pDAB9582.814.19.1. Подробный анализ саузерн-блоттинг проводили с использованием зондов, специфичных к гену cry1Ac и cry1F, содержащихся в участке встраивания T-цепи pDAB9582.814.19.1, и иллюстративные ферменты рестрикции, которые содержат участки расщепления, локализованные в плазмиде, и образуют гибридизационные фрагменты, внутренние по отношению к плазмиде или фрагменты, которые перекрывают область соединения плазмиды с геномной ДНК сои (граничные фрагменты). Молекулярные массы, полученные Саузерн-гибридизациией для комбинации рестрикционного фермента и зонда, являлись уникальными для трансформационного события, и устанавливали их идентификационные паттерны. Эти анализы также демонстрировали, что фрагмент плазмиды встраивался в геномную ДНК сои без перестройки PTU cry1Ac и cry1F.
Пример 4.1: Сбор образцов листьев сои и выделение геномной ДНК (гДНК)
[0139] Геномную ДНК экстрагировали из ткани листьев, собранных от индивидуального растения сои, содержащего трансформационное событие сои pDAB9582.814.19.1. Кроме того, гДНК выделяли из общепринятого растения сои Maverick, которое содержит генетическую среду, которая является репрезентативной для конкретной линии, в которой отсутствуют гены cry1Ac и cry1F. Индивидуальную геномную ДНК экстрагировали из лиофилизированной ткани листа стандартным способом CTAB (Sambrook et al. (1989)). После экстракции спектрофотометрически количественно определяли ДНК с использованием реагента PICO GREEN (Invitrogen, Carlsbad, CA). Затем ДНК визуализировали в агарозном геле для подтверждения значений анализа PICO GREEN и для определения качества ДНК.
Пример 4.2: Расщепление и выделение ДНК
[0140] Для молекулярной характеристики саузерн-блоттингом трансформационного события сои pDAB9582.814.19.1 расщепляли десять микрограмм (10 мкг) геномной ДНК. Геномную ДНК от трансформанта сои pDAB9582.814.19.1 и нетрансгенной линии сои Maverick расщепляли, добавляя, приблизительно пять единиц выбранного фермента рестрикции на мкг ДНК и соответствующий реакционный буфер к каждому образцу ДНК. Каждый образец инкубировали приблизительно при 37°C в течение ночи. Ферменты рестрикции AseI, HindIII, NsiI, и NdeI использовали индивидуально для отдельных расщеплений (New England Biolabs, Ipswich, MA). Ферменты рестрикции NotI и ApaLI использовали совместно для двойного расщепления (New England Biolabs, Ipswich, MA). Кроме того, получали контрольные образцы положительной гибридизации, комбинируя плазмиду ДНК, pDAB9582 с геномной ДНК из нетрансгенного сорта сои Maverick. Смесь плазмидной ДНК/геномной ДНК расщепляли аналогичными способами и с использованием аналогичного фермента рестрикции как в тестируемых образцах.
[0141] Затем реакционную смесь расщепления инкубировали в течение ночи, добавляли 25 мкл раствора QUICK-PRECIP PLUS (Edge Biosystems, Gaithersburg, MD) и образцы расщепленной ДНК осаждали изопропанолом. Преципитированный осадок ДНК ресуспендировали в 15 мкл 1X буфера для нанесения (0,01% бромфенолового синего, 10,0 мМ ЭДТА, 10,0% глицерина, 1,0 мМ Tris pH 7,5). Затем образцы ДНК и маркеры молекулярного размера подвергали электрофорезу в 0,85% агарозных гелях с 0,4X буфером TAE (Fisher Scientific, Pittsburgh, PA) при 35 вольт приблизительно в течение 18-22 часов до получения разделения фрагментов. Гели окрашивали бромистым этидием (Invitrogen, Carlsbad, CA) и визуализировали ДНК в ультрафиолетовом (УФ) свете.
Пример 4.3: Перенос по Саузерну и обработка мембраны
[0142] Анализ саузерн-блоттинг проводили по существу, как описано Memelink et al. (1994). В кратком изложении, после электрофоретического разделения и визуализации фрагментов ДНК, гели подвергали депуринации 0,25M HCl приблизительно в течение 20 минут, а затем подвергали действию денатурирующего раствора (0,4M NaOH, 1,5M NaCl) в течение приблизительно 30 минут с последующей обработкой нейтрализующим раствором (1,5M NaCl, 0,5M Tris pH 7,5) в течение по меньшей мере 30 минут. Перенос по Саузерну проводили в течение ночи на нейлоновых мембранах и с использованием капиллярной системы с 10X SSC. После переноса ДНК подвергали связыванию с мембраной путем образования поперечных связей под действием УФ с последующим быстрым промыванием мембраны раствором 2X SSC. Этим способом получали мембраны для саузерн-блоттинга, готовые для гибридизации.
Пример 4.4: Мечение и гибридизация ДНК-зонда
[0143] Фрагменты ДНК, связанные с нейлоновой мембраной, детектировали с использованием меченого зонда (таблица 6). Зонды получали встраиванием на основе ПЦР меченого дигоксигенином (DIG) нуклеотида, [DIG-11]-dUTP, во фрагмент ДНК, амплифицированный из плазмиды pDAB9582 с использованием праймеров, специфичных для элементов генов. Получение ДНК-зондов посредством синтеза ПЦР проводили с использованием набора PCR DIG Probe Synthesis (Roche Diagnostics, Indianapolis, IN) способами, рекомендуемыми производителем.
[0144] Меченые зонды анализировали электрофорезом в агарозном геле для определения их качества и количества. Затем желаемое количество меченого зонда использовали для гибридизации с ДНК-мишенью на нейлоновых мембранах для детекции специфических фрагментов с использованием способов по существу, как описано для раствора DIG EASY HYB (Roche Diagnostics, Indianapolis, IN). В кратком изложении, нейлоновые мембранные блоты, содержащие фиксированную ДНК, быстро промывали 2X SSC и подвергали предварительной гибридизации с 20-25 мл предварительно подогретого раствора DIG EASY HYB в гибридизационных пробирках при приблизительно 45-55°C в течение приблизительно 2 часов в гибридизационной камере. Затем раствор для предварительной гибридизации сливали и заменяли ~15 мл предварительно подогретого раствора DIG EASY HYB, содержащего желаемое количество специфических зондов, денатурированных кипячением на водяной бане в течение приблизительно пяти минут. Затем проводили этап гибридизации при приблизительно 45-55°C в течение ночи в гибридизационной камере.
[0145] В конце гибридизации зонда растворы DIG EASY HYB, содержащие зонды, сливали в чистые пробирки и хранили при приблизительно -20°C. Эти зонды можно повторно использовать до двух раз способом, рекомендуемым производителем. Мембранные блоты быстро ополаскивали и дважды промывали в чистых пластиковых контейнерах буфером для промывания с низкой жесткостью (2X SSC, 0,1% SDS) в течение приблизительно пяти минут при комнатной температуре с последующим двукратным промыванием буфером для промывания с высокой жесткостью (0,1X SSC, 0,1% SDS) в течение 15 минут каждый при приблизительно 65°C. Мембранные блоты быстро промывали 1X буфером на основе малеиновой кислоты из набора DIG WASH AND BLOCK BUFFER (Roche Diagnostics, Indianapolis, IN) в течение приблизительно 5 минут. После этого проводили блокирование в 1X блокирующем буфере в течение 2 часов и инкубацию с конъюгированным с AP (щелочной фосфатазой) антителом против DIG (Roche Diagnostics, Indianapolis, IN) в 1X блокирующем буфере также в течение не менее 30 минут. После 2-3 промываний 1X буфером для промывания специфические ДНК-зонды оставались связанными с мембранными блотами и стандарты меченной DIG ДНК визуализировали с использованием системы CDP-STAR CHEMILUMINESCENT NUCLEIC ACID DETECTION (Roche Diagnostics, Indianapolis, IN) в соответствии с рекомендациями производителя. Блоты подвергали действию хемилюминесцентной пленки в течение одного или более моментов времени для детекции гибридизационных фрагментов и визуализации стандартов молекулярного размера. Пленки проявляли с использованием устройства для проявления пленок ALL-PRO 100 PLUS (Konica Minolta, Osaka, Japan) и анализировали изображения. Для каждого зонда пробы документировали число и размер. Для определения размера гибридизационных фрагментов в саузерн-блоттингах использовали меченый DIG маркер молекулярной массы ДНК II (DIG MWM II) и меченый DIG маркер молекулярной массы ДНК VII (DIG MWM VII), видимые после детекции DIG, как описано.
|
Пример 4.5: Результаты саузерн-блоттинга
[0146] Ожидаемые и наблюдаемые размеры фрагментов с конкретным расщеплением и зондом, основанные на известных участках распознавания рестрикционных ферментов PTU cry1Ac и cry1F, приведены в таблице 7. На основании этих расщеплений и гибридизаций идентифицировали два типа фрагментов: внутренние фрагменты, где известные участки фермента, фланкируют участок зонда и полностью содержаться в области встраивания cry1Ac и cry1F PTU, и граничные фрагменты, где известный участок фермента локализован на одном конце участка зонда, и предполагают, что второй участок находится в геноме сои. Размеры граничных фрагментов изменяются в зависимости от трансформационного события, т.к. в большинстве случаев участки интеграции фрагментов ДНК являются уникальными для каждого трансформационного события. Краевые фрагменты обеспечивают способ локализации участка распознавания рестрикционного фермента относительно интегрируемой ДНК и оценки количества вставок ДНК. Анализом саузерн-блоттинг, проводимым на многих генерациях сои, содержащей трансформационное событие сои pDAB9582.814.19.1, были получены данные, которые подтверждали, что низкокопийная интактная PTU cry1Ac и cry1F из плазмиды pDAB9582 интегрировалась в геном сои трансформанта сои pDAB9582.814.19.1.
[0147]
|
[0148] Ферменты рестрикции AseI и NsiI связываются и расщепляют уникальные участки рестрикции в плазмиде pDAB9582. Впоследствии эти ферменты выбирали для характеристики вставки гена Cry1Ac в трансформационном событии сои pDAB9582.814.19.1. Граничные фрагменты >7286 п.н. или >9479 п.н. теоретически рассчитывали для гибридизации с зондом после расщеплений AseI и NsiI соответственно (таблица 7). Наблюдали отдельные гибридизационные полосы Cry1Ac приблизительно 7400 и >10000 п.н., когда использовали расщепления AseI и NsiI соответственно. Гибридизация зонда с полосами такого размера подтверждает наличие одного участка вставки для гена Cry1Ac в геноме сои трансформанта сои pDAB9582.814.19.1. Ферменты рестрикции NotI и ApaLI выбирали для проведения двойного расщепления и для выделения фрагмента, который содержит единицу транскрипции растения (PTU, промотор/ген/терминатор) Cry1Ac (таблица 7). Теоретически рассчитанные фрагменты 4550 п.н. наблюдали с зондом после двойного расщепления NotI и ApaLI. Результаты, получаемые с ферментативным расщеплением образцов pDAB9582.814.19.1 с последующей гибридизацией с зондом, свидетельствовали о том, что интактная TUP Cry1Ac из плазмиды pDAB9582 встраивалась в геном сои трансформанта сои pDAB9582.814.19.1.
[0149] Ферменты рестрикции NdeI и NsiI связываются и расщепляют участки рестрикции в плазмиде pDAB9582. Впоследствии эти ферменты выбирали для характеристики вставки гена cry1F в трансформационном событии сои pDAB9582.814.19.1. Граничные фрагменты >5569 п.н. и >9479 теоретически рассчитывали для гибридизации с зондом после расщеплений NdeI и NsiI соответственно (таблица 7). Наблюдали отдельные гибридизационные полосы cry1F ~7500 п.н. и >10000 п.н., когда использовали NdeI и NsiI соответственно. Гибридизация зонда с полосами такого размера подтверждала наличие одного участка встраивания гена Cry1F в геном сои трансформанта сои pDAB9582.814.19.1. Фермент рестрикции HindIII выбирали для выделения фрагмента, который содержит единицу транскрипции растения cry1F (PTU, промотор/ген/терминатор) (таблица 7). Теоретически рассчитанный фрагмент 7732 п.н. наблюдали с зондом после расщеплений HindIII. Результаты, получаемые с ферментативным расщеплением образцов pDAB9582.814.19.1 с последующей гибридицией с зондом, свидетельствуют о том, что интактная PTU cry1F из плазмиды pDAB9582 встраивалась в геном сои трансформанта сои pDAB9582.814.19.1.
Пример 4.6: Отсутствие последовательностей основной цепи
[0150] Проводили также анализ саузерн-блоттинг для подтверждения отсутствия гена устойчивости к спектиномицину (specR), элемента Ori Rep и белка начала репликации trfA (элемент trf A) в трансформационном событии сои pDAB9582.814.19.1. Не ожидают специфической гибридизации с геном устойчивости к спектиномицину, элементом Ori Rep или элементом trfA, когда включают соответствующие положительные (с pDAB9582, добавленным в геномную ДНК Maverick) и отрицательные (геномная ДНК Maverick) контроли в анализ по Саузерну. После расщепления NsiI и гибридизации со специфическим зондом specR, наблюдали одну полосу ожидаемого размера 15320 п.н. в образце положительного контроля (с pDAB9582, добавленным в геномную ДНК Maverick). Зонд specR не гибридизовался с образцами отрицательного контроля и трансформационным событием сои pDAB9582.814.19.1. Аналогично, детектировали одну полосу ожидаемого размера 15320 п.н. в образце положительного контроля (pDAB9582 плюс Maverick), но отсутствующую в образцах отрицательного контроля и трансформационного события сои pDAB9582.814.19.1 после расщепления NsiI и гибридизации зондом trfA. Детектировали полосу с другим ожидаемым размером 5329 п.н. в образце положительного контроля (с pDAB9582, добавленным в геномную ДНК Maverick), но отсутствующую в образцах отрицательного контроля и трансформационного события сои pDAB9582.814.19.1 после расщепления NdeI и гибридизации со специфическим зондом OriRep. Эти данные указывают на отсутствие гена устойчивости к спектиномицину, элемента Ori Rep и белка начала репликация trfA в трансформационном событии сои pDAB9582.814.19.1.
Пример 5: Агрономическое и полевое испытание урожайности и толерантности к гербицидам
[0151] Для тестирования агрономических характеристик и эффективности трансформационного события сои pDAB9582.814.19.1 трансформант культивировали в испытании эффективности в Санта Исабель, Пуэрто-Рико в октябре 2010 года и феврале 2011 года. Культивар Maverick, который исходно трансформировали с получением трансформанта pDAB9582.814.19.1, высаживали в каждом рассаднике и включали в качестве контроля в экспериментах. Семена из опытного участка T3 получали из индивидуальных отборов растений на стадии T2 и семена для опытного участка T4 получали из индивидуальных отборов растений на стадии T3. Тестировали каждую генерацию четырех линий трансформанта. Каждую линию высаживали на делянке, которая составляла в ширину 4 грядки и в длину 7,5 футов. Расстояние между грядками составляло 30 дюймов. Делянки выращивали под светом в течение приблизительно 2,5 недель для компенсации короткой длины светового дня в Пуэрто-Рико. Каждый рассадник опрыскивали глуфосинатом со скоростью 411 г экв. кисл./га. Одну делянку контрольных растений Maverick опрыскивали с аналогичной скоростью глуфосинатом, а вторую делянку не опрыскивали и использовали в качестве контрольного сравнения для трансформанта.
[0152] Данные собирали в отношении прорастания, общего внешнего вида, мощности, высоты, полегания и созревания. Толерантность к гербициду оценивали, визуально просматривая хлороз, некроз листьев и гибель растений (таблица 8).
[0153] Для сравнений трансформанта сои pDAB9582.814.19.1 с Maverick использовали данные только из не опрыскиваемого блока Maverick. Для сравнения обработок с опрыскиванием и без него сравнивали данные из блока трансформанта сои pDAB9582.814.19.1, опрыскиваемого данным типов обработки, с данными из блока контрольного неопрыскиваемого Maverick. Трансформант сои pDAB9582.814.19.1 проявлял толерантность к применению гербицида глуфосината. В противоположность этому, ни одно из растений Maverick не являлось толерантным к обработкам гербицидом.
[0154]
|
Пример 6: Характеристика инсектицидной активности трансформанта сои pDAB9582.814.19.1
[0155] Проводили выращивание в поле и теплице для характеристики активности Cry1Ac и Cry1F в трансформанте сои pDAB9582.814.19.1 против выращенных в лаборатории паразитов сои, включая Anticarsia gemmatalis (гусеницу вельветовых бобов), Pseudoplusia includes (соевую совку) и Spodoptera frugiperda (травяную совку). Трансформант сои pDAB9582.814.19.1 сравнивали с нетрансформированным сортом сои Maverick для определения уровня защиты растения, обеспечиваемого белками Cry1F и Cry1Ac.
[0156] Испытания в теплице проводили на растениях приблизительно в возрасте четырех недель. Для оценки трансформанта сои pDAB9582.814.19.1 и контроля Maverick использовали пятнадцать растений. Для каждого тестируемого вида насекомого (Anticarsia gemmatalis, Pseudoplusia includes и Spodoptera frugiperda) получали 3 кусочка листа, получаемых компостированием, от каждого растения всего 45 листовых дисков/растений/видов насекомых. Кусочки листа диаметром 1,4 см (или 1,54 см2) помещали в тестируемую область на вершине 2% водного агара, инфицировали одной новорожденной личинкой и плотно закрывали перфорированной пластиковой крышкой. Через 4 суток после заражения оценивали смертность и потребление листа. Личинку, которая не реагировала на аккуратное прощупывание, считали погибшей. Повреждение листа оценивали посредством визуальной оценки процента съеденного насекомым кусочка листа.
[0157] Полевые оценки проводили, собирая образцы листьев от семян, выращенных в рассадниках в Санта Исабеле (Santa Isabel), Пуэрто-Рико и высылая эти листья в Индианаполис, IN для тестирования. Рассадник для трансформанта сои pDAB9582.814.19.1 высаживали в феврале 2011 года, и он состоял приблизительно из 180 растений, распределенных по четырем грядкам. Каждая грядка составляла 2,3 м в длину и на расстоянии друг от друга 76,2 см; расстояние между индивидуальными растениями составляло 5,1 см на каждой грядке. В марте 2011 года вырезали один полностью распустившийся трехлопастной лист главного стебля, расположенный приблизительно на четыре узла ниже меристемы, у 10 растений с трансформационным событием сои pDAB9582.814.19.1 и у 10 растений "Maverick". Листья помещали в помеченные пластиковые пакеты (один лист на пакет) и запечатывали. Помещенные в мешки листья упаковывали и передавали в лабораторию. В лаборатории один или два диска листа диаметром 3,33 см (1,31 дюйма) получали перфорированием из каждого трехлопастного листа, получая всего 16 листовых дисков. Каждый листовой диск помещали в тестируемую область на вершине 2% агара, инфицировали одной новорожденной личинкой S. frugiperda и плотно закрывали перфорированной пластиковой крышкой. Листовые диски содержали в комнате с контролируемыми условиями в течение 7 суток, во время которых оценивали смертность и потребление листа. Личинку, которая не реагировала на аккуратное прощупывание, считали погибшей. Повреждение листа оценивали визуально посредством оценки процента съеденного насекомым кусочка листа.
[0158] Результаты, получаемые из этих повторяемых экспериментов, указывали на то, что трансформант сои pDAB9582.814.19.1 испытывал значительно меньше повреждений по сравнению с контрольными растениями Maverick от всех тестируемых насекомых. Таким образом, трансформационное событие сои pDAB9582.814.19.1 обладает инсектицидной активностью по отношению к этому широкому кругу хозяев.
Пример 7: Последовательность трансформанта сои pDAB9582.814.19.1
[0159] SEQ ID NO:14 представляет собой последовательность трансформанта сои pDAB9582.814.19.1. Эта последовательность содержит 5'-геномную фланкирующую последовательность, вставку T-цепи pDAB9582 и 3'-геномные фланкирующие последовательности. В отношении SEQ ID NO:14 остатки 1-1400 представляют собой 5'-геномную фланкирующую последовательность, остатки 1401-1536 представляют собой остатки реаранжировки из плазмиды pDAB9582, и 1537-13896 представляют собой остатки вставки T-цепи pDAB9582, и остатки 13897-15294 представляют собой 3'-фланкирующую последовательность. Соединительная последовательность или переход по отношению к 5'-концу вставки, таким образом, возникает при остатках 1400-1401 SEQ ID NO:14. Соединительная последовательность или переход по отношению к 3'-концу вставки, таким образом, возникает при остатках 13896-13897 SEQ ID NO:14.
[0160] Следует отметить, что потомство трансформанта сои pDAB9582.814.19.1 может содержать последовательности, которые немного отличаются от SEQ ID NO:14. Во время интрогрессии и процесса селекции вводимого трансформационного события сои pDAB9582.814.19.1 в геном растительной клетки, нередко возникают делеции или другие изменения вставки. Кроме того, могут возникать ошибки ПЦР амплификации, которые могут приводить к незначительным ошибкам секвенирования. Например, фланкирующие последовательности, перечисленные в настоящем описании, определяли посредством получения ампликонов из геномных ДНК сои, а затем клонировали и секвенировали ампликоны. Часто обнаруживают небольшие отличия и незначительные несоответствия последовательностей, получаемых и определяемых таким образом, обусловленные многими этапами амплификации, которая является необходимой для получения достаточного ампликона из геномных ДНК для секвенирования. Специалисту в данной области следует понимать и учитывать, что любые поправки, необходимые вследствие этого типа общих ошибок секвенирования или несоответствий, входят в объем настоящего изобретения. Таким образом, соответствующий сегмент последовательности плазмиды, предоставленный в настоящем описании, может содержать некоторые незначительные изменения. Таким образом, растение, содержащее полинуклеотид, обладающий некоторым диапазоном идентичности с рассматриваемой последовательностью вставки, входит в объем настоящего изобретения. Идентичность последовательности SEQ ID NO:14 может представлять собой полинуклеотидную последовательность, обладающую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 91%, 98%), 99% или 100% идентичностью последовательности с последовательностью, иллюстрируемой или описываемой в настоящем описании. Таким образом, можно идентифицировать некоторые отличия между SEQ ID NO:14 и растениями потомства трансформанта сои pDAB9582.814.19.1, и они входят в объем настоящего изобретения.
Пример 8: Селекционный гибрид трансформанта сои pDAB4468.04.16.1 и трансформанта сои pDAB9582.814.19.1
Пример 8.1: Половое скрещивание трансформанта сои pDAB4468.04.16.1 и трансформанта сои pDAB9582.814.19.1
[0161] Трансформант сои pDAB4468.04.16.1, как описано в международной патентной заявке № WO/2012/075426, скрещивали половым путем с трансформантом сои pDAB9582.814.19.1. Пыльники трансформанта сои pDAB4468.04.16.1 вручную терли по рыльцу трансформанта сои pDAB9582.814.19.1, таким образом, проводя опыление трансформанта сои pDAB9582.814.19.1. Получаемое потомство F1, которое содержало интеграцию трансформационных событий от трансформанта сои pDAB9582.814.19.1 и трансформанта сои pDAB4468.04.16.1, подвергали скринингу в отношении толерантности к 2,4-D и гербицидам на основе глуфосината для идентификации растений потомства, которые содержали оба трансформационных события интеграции.
Пример 8.2: Характеристика экспрессии белка в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1.
[0162] Проводили характеристику биохимических свойств рекомбинантных белков Cry1F, Cry1Ac, AAD12 и PAT, экспрессируемых в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB8264.44.06.1. Подтверждали, что в целом 51 растение F1 содержит оба трансформационных события сои pDAB9582.814.19.1 и pDAB4468.04.16.1, специфическим в отношении трансформационного события анализом TAQMAN®, анализом на основе такого, как описан в предварительной патентной заявке США № 61/511658 и международной патентной заявке № WO/2011/066360 соответственно. Затем использовали твердофазный иммуноферментный анализ (ELISA) для количественного определения экспрессии белков PAT и AAD12, при этом белки Cry1Ac и Cry1F количественно определяли мультиплексными иммунологическими анализами с использованием технологии электрохемилюминесценции от Meso-Scale Discovery (MSD). В совокупности эти анализы использовали для характеристики биохимических свойств белков и подтверждения экспрессии этих белков в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1.
Экспрессия белка PAT в тканях растений
[0163] В селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 определяли уровни белка PAT. Растворимый экстрагируемый белок PAT измеряли количественным твердофазным иммуноферментным анализом (Envirologix, Portland, ME) для тестируемой ткани листа сои.
[0164] Специфическими в отношении трансформационного события анализами TAQMAN® подтверждали, что в целом 51 растение F1 содержит оба трансформационных события сои pDAB9582.814.19.1 и pDAB4468.04.16.1. У таких выращенных в теплицах тестируемых растений выделяли образцы ткани листа сои на стадии роста V3-V4 и подготавливали для анализа экспрессии генов. Из тканей растения сои экстрагировали белок PAT раствором фосфатно-солевого буфера, содержащего детергент Tween-20 (PBST) и 1% поливинилпирролидон 40 (PVP-40). Затем образцы экстрагировали с использованием GENOGRINDER™ при 1500 об./мин. в течение 5 минут. Экстракт растений центрифугировали, собирали водный супернатант, разбавляли подходящим буфером по мере необходимости и анализировали с использованием набора ELISA для PAT в формате "сэндвич". Набор использовали в соответствии с протоколом, рекомендуемым производителем. Этим анализом измеряли экспрессируемый белок PAT в ткани листьев.
[0165] Для исследования наследования экспрессии в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 проводили анализ детекции. В генерации F1 растения селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 экспрессировали PAT (таблица 9).
|
Экспрессия белков Cry1F и Cry1Ac в тканях растений
[0166] В селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 определяли уровни белков Cry1F и Cry1Ac. Растворимые экстрагируемые белки Cry1F и Cry1Ac измеряли мультиплексным анализом MSD (Meso-Scale Discovery, Gaithersburg, MD) для ткани листа сои.
[0167] Специфическими в отношении трансформационного события анализами TAQMAN® подтверждали, что в целом 51 растение F1 содержит оба трансформационных события сои pDAB9582.814.19.1 и pDAB4468.04.16.1. У таких выращенных в теплицах тестируемых растений выделяли образцы ткани листа сои на стадии роста V3-V4 и подготавливали для анализа экспрессии генов. Белки Cry1F и Cry1Ac экстрагировали из ткани растения сои PBST и 1% PVP40. Затем образцы экстрагировали с использованием GENOGRINDER™ при 1500 об/мин в течение 5 минут. Экстракт растения центрифугировали, собирали водный супернатант, разбавляли подходящим буфером по мере необходимости и анализировали мультиплексным анализом MSD Cry1F/Cry1Ac от Meso-Scale Discovery. В этом анализе измеряли белок Cry1F и Cry1Ac в ткани листа.
[0168] Для исследования наследования экспрессии в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 проводили анализ детекции. В генерации F1 растения селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 экспрессировали Cry1F и Cry1Ac (таблица 10).
|
Экспрессия белков AAD12 в тканях растений
[0169] В селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 определяли уровни белка AAD12. Растворимый экстрагируемый белок AAD12 измеряли количественным твердофазным иммуноферментным анализом (Acadia Bioscience, Portland, ME) для тестируемой ткани листа сои.
[0170] Специфическими в отношении трансформационного события анализами TAQMAN® подтверждали, что в целом 51 растение F1 содержит оба трансформационных события сои pDAB9582.814.19.1 и pDAB4468.04.16.1. У таких выращенных в теплицах тестируемых растений выделяли образцы ткани листа сои на стадии роста V3-V4 и подготавливали для анализа экспрессии генов. Белок AAD12 экстрагировали из тканей растения сои раствором фосфатно-солевого буфера, содержащего детергент Tween-20 (PBST) и 0,5% бычий сывороточный альбумин (BSA). Затем образцы экстрагировали с использованием GENOGRINDER™ при 1500 об/мин в течение 5 минут. Экстракт растения центрифугировали, собирали водный супернатант, разбавляли подходящим буфером по мере необходимости и анализировали с использованием набора ELISA для AAD12 в формате "сэндвич". Набор использовали в соответствии с протоколом производителя (Acadia, Portland, ME). В этом анализе измеряли экспрессируемый белок AAD12 в ткани листа.
[0171] Для исследования наследования экспрессии в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 проводили анализ детекции. В генерации F1 растения селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 экспрессировали AAD12 (таблица 11).
|
Пример 8.3: Толерантность к гербицидам селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1
[0172] Толерантность к гербицидам селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 анализировали в исследовании в теплице. Семена трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 высаживали в горшки и выращивали до зрелости. Пятьдесят три (53) зрелых растения F1 опрыскивали при одном применении гербицида, которое состояло из 840 г экв. кисл./га 2,4-D, при стадии роста V3-V4 (отличающейся тем, что однолопастные и первые три-четыре трехлопастных листа полностью развились). Получаемую толерантность к этим гербицидам измеряли, подсчитывая число выживших растений. Было выявлено, что все пятьдесят три растения являются устойчивыми к применению гербицида. Для сравнения в исследование включали контрольные растения, которые не содержали ген aad-12, и для которых ожидалось, что они являются чувствительными к применению гербицидов на основе 2,4-D.
[0173] В итоге, ген aad-12, который содержится в родительской линии трансформанта сои pDAB4468.04.16.1, обеспечивал толерантность к гербициду 2,4-D. Этот признак передавался и наследовался в сое pDAB9582.814.19.1::pDAB4468.04.16.1, таким образом, обеспечивая толерантность к гербицидам трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1. При сравнении контрольные растения, которые не содержали ген aad-12, являлись чувствительными к применению гербицида 2,4-D.
Пример 8.4: Характеристика инсектицидной активности трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1
[0174] Оценки в теплице проводили для характеристики активности Cry1Ac и Cry1F в селекционном гибриде трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 против выращенных в лаборатории паразитов сои, включая Anticarsia gemmatalis (гусеницу вельветовых бобов) и Pseudoplusia includens (соевую совку). Селекционный гибрид трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 сравнивали с трансформантом сои pDAB9582.814.19.1, трансформантом сои pDAB4468.04.16.1 и нетрансформированным сортом сои Maverick для определения, изменяется ли уровень защиты растения, обеспечиваемый изменениями белков Cry1F и Cry1Ac, при стэкинге. Кроме того, селекционный гибрид трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 и трансформанта сои pDAB4468.04.16.1 опрыскивали одним применением гербицида, содержащим 840 г экв. кисл./га 2,4-D, перед биоанализом для подтверждения отсутствия отрицательного биологического влияния гербицидов на насекомых.
Опыты в теплице проводили на растениях приблизительно в возрасте трех недель. Каждое от пяти до десяти растений использовали для оценки селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1, трансформанта сои pDAB9582.814.19.1 и отрицательных контролей: опрыскиваемых трансформанта сои pDAB4468.04.16.1 и Maverick. Для каждого вида тестируемого насекомого (Anticarsia gemmatalis и Pseudoplusia includes) получали 3 кусочка листа, полученных компостированием, от каждого растения на один вид насекомого. Кусочки листа диаметром 1,4 см (или 1,54 см2) помещали в тестируемую область на вершине 1,5% водного агара, инфицировали одной новорожденной личинкой и плотно закрывали перфорированной пластиковой крышкой. Через 4 суток после заражения оценивали смертность и потребление листа. Личинку, которая не реагировала на аккуратное прощупывание, считали погибшей. Повреждение листа оценивали посредством визуальной оценки процента площади листа, съеденной насекомым.
Результаты, получаемые из таких повторяемых экспериментов (таблица 12) указывают на то, что селекционный гибрид трансформационного события сои pDAB9582.814.19.1::pDAB4468.04.16.1 соответствовал трансформанту сои pDAB9582.814.19.1 и подвергался значительно меньшему повреждению ткани листа (0,6-0,8%) по сравнению с контролями трансформанта pDAB4468.04.16.1 (94-99%) и Maverick (96-97%) со стороны соевой совки и гусеницы вельветовых бобов. Высокую смертность (100%) регистрировали для селекционного гибрида трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 и трансформанта сои pDAB9582.814.19.1, при этом растения pDAB4468.04.16.1 и Maverick приводили к низкой смертности (0%), и все насекомые, помещаемые на эти контрольные растения, выживали. Таким образом, селекционный гибрид трансформанта сои pDAB9582.814.19.1::pDAB4468.04.16.1 обладает сравнимой инсектицидной активностью с трансформантом сои pDAB9582.814.19.1.
|
[0175] Проиллюстрированы и описаны принципы настоящего изобретения, специалистам в данной области следует понимать, что изобретение можно модифицировать в расстановке и деталях, не выходя за рамки таких принципов. Авторы заявляют все модификации, которые входят в сущность и объем прилагаемой формулы изобретения.
[0176] Все публикации и опубликованные патентные документы, цитируемые в этом описании, включены в настоящее описание посредством ссылки в той же степени, как если бы каждую отдельную публикацию или патентную заявку конкретно и индивидуально указывали как включенную посредством ссылки.


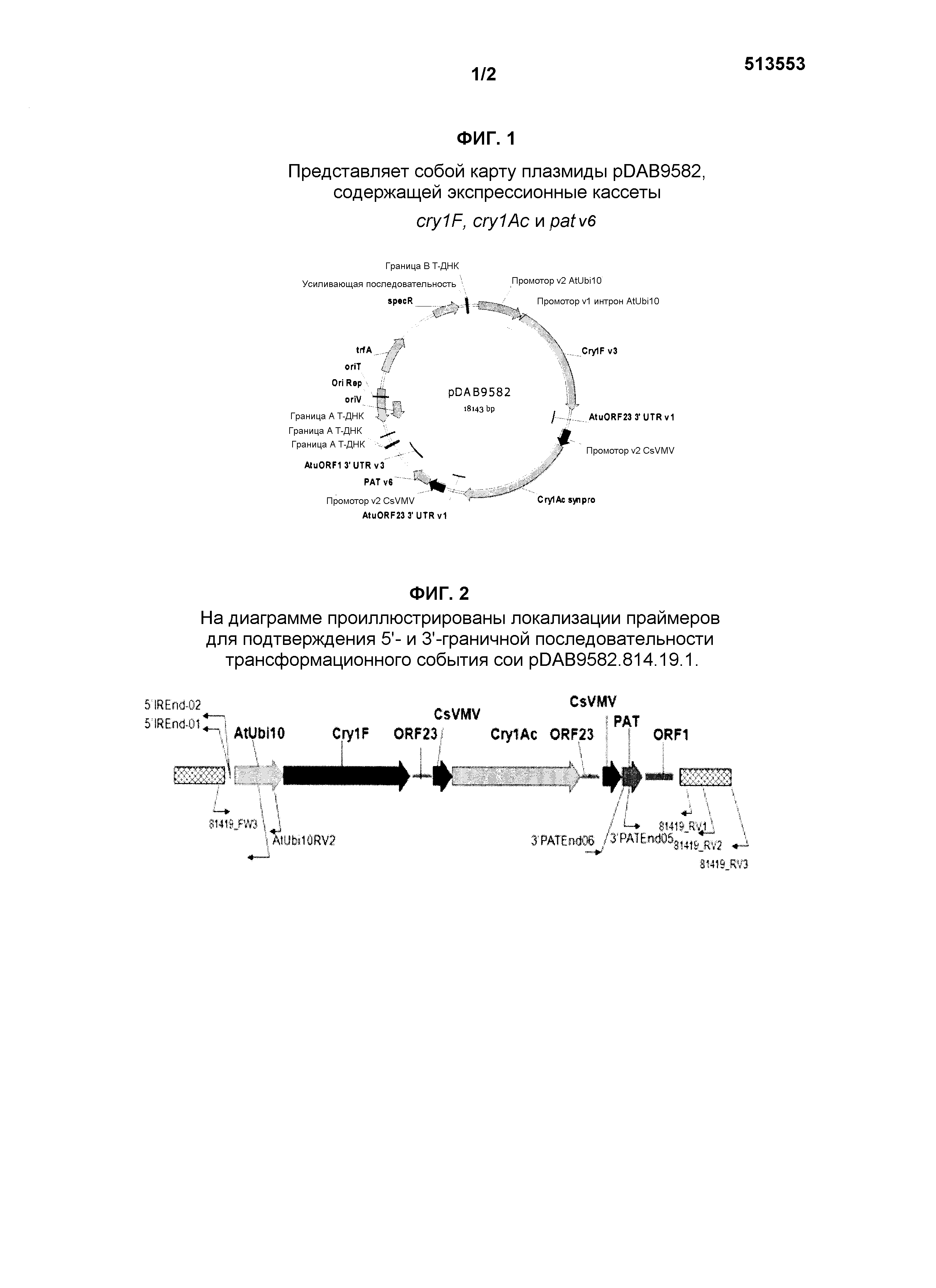
























Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Получение стабилизированной цельнозерновой муки и продуктов из нее
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
5-фторпиримидиновые производные в качестве фунгицидов
Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Получение стабилизированной цельнозерновой муки и продуктов из нее
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
Стабильная пестицидная композиция на основе сульфоксимина и способ борьбы с насекомыми

