Результат интеллектуальной деятельности: АНТИТЕЛА К XCR1 ЧЕЛОВЕКА
Вид РИД
Изобретение
Область техники
Настоящее изобретение относится к антителу, которое связывается с XCR1 человека.
Предпосылки изобретения
Хемокины - это собирательное выражение для основных гепарин-связывающих белков, которые влияют на лейкоцитарный хемотаксис и активацию лейкоцитов. На основе сравнения первичных структур различных хемокинов хемокины классифицированы на подсемейства СХС, СС, С и СХ3С в соответствии с положением консервативных цистеиновых остатков. XCL1 (также называемый лимфотактином (Ltn) или лимфотактином-α (Ltn-α)) и XCL2 (также называемый лимфотактином-β (Ltn-β)) являются хемокинами, классифицированными в подсемейство С, описанное выше. XCR1 (также называемый GPR5, SCM-1α или АТАС) является рецептором хемокинов, сопряженным с G-белком, который специфически связывается с XCL1 и XCL2.
Экспрессия XCR1 в различных тканях человека была исследована на уровне мРНК. Как сообщается, уровень экспрессии XCR1 высокий в плаценте, но низкий в селезенке и тимусе (НЕПАТЕНТНАЯ ЛИТЕРАТУРА 1). Кроме того, XCR1 в основном экспрессируется в дендритных клетках. У мышей XCR1 экспрессируется на высоком уровне, особенно в CD8α+ дендритных клетках (НЕПАТЕНТНАЯ ЛИТЕРАТУРА 2 и НЕПАТЕНТНАЯ ЛИТЕРАТУРА 3). CD8α+ дендритные клетки обычно присутствуют во вторичных лимфоидных тканях, напрмер в селезенке и лимфатических узлах и, как известно, осуществляют "кросс-презентацию", которая играет важную роль в реакциях против инфекции и иммунологических ответах на опухолевые клетки. Также известно, что XCR1 экспрессируется на высоком уровне в CD141+ дендритных клетках человека, которые, как полагают, являются гомологами CD8α+ дендритных клеток мыши (НЕПАТЕНТНАЯ ЛИТЕРАТУРА 4).
Антиген, захваченный из внешней среды клетки в антигенпредставляющие клетки, как правило, разрушается до пептидов, представляемых с антигеном главного комплекса гистосовместимости класса II (МНС класса II) и распознаваемых CD4+ Т-клетками. Напротив, есть случай, когда антиген, захваченный из внешней среды клеток, представляется с антигеном главного комплекса гистосовместимости класса I (МНС класса I) через путь, отличающийся от обычного пути, описанного выше. Этот процесс представления антигена называется "кросс-презентацией". В этом процессе антиген, представляемый с МНС класса I, распознается CD8+ Т-клетками, которые затем дифференцируется в цитотоксические Т-клетки (CTL), которые играют роль в филаксии и устранении опухолевых клеток в организме хозяина (НЕПАТЕНТНАЯ ЛИТЕРАТУРА 5).
Миграция различных иммунных клеток происходит во время реакции воспаления. В частности, миграция дендритных клеток к локальному месту воспаления происходит для выполнения фагоцитоза антигенов. Хемокины и рецепторы хемокинов играют важную роль в процессе такой миграции дендритных клеток. После миграции к локальному месту воспаления дендритные клетки представляют антигены Т-клеткам и активизируют Т-клетки. Впоследствии информация передается от Т-клеток многим иммунным клеткам, усиливая иммунный ответ (НЕПАТЕНТНАЯ ЛИТЕРАТУРА 6).
Среди антигенпредставляющих клеток дендритные клетки обладают особенно превосходным антигенпредставляющим свойством и играют очень важную роль в активации Т-клеток. Было выдвинуто предположение, что поскольку Т-клетки участвуют в развитии и усилении различных иммунопатологических заболеваний, включая аутоиммунные заболевания, то управлять дендритными клетками означает управлять активацией Т-клеток, что может привести к улучшению при различных иммунопатологических заболеваниях (непатентная литература 6 и непатентная литература 7).
Кроме того, было показано, что полученные от кролика поликлональные антитела к XCR1 человека имеют эффект ингибирования XCL-индуцируемой миграции нормальных кератиноцитов полости рта и раковых клеток полости рта (непатентная литература 8).
Список цитируемой литературы Непатентная литература
NPL 1: Yoshida Τ, Imai Τ, Kakizaki M, Nishimura M, Takagi S, Yoshie О. "Identification of Single С motif-1/lymphotactin receptor XCR1," J. Biol. Chem. 273: 16551-16554 (1998).
NPL 2: Crozat K, Guiton R, Contreras V, Feuillet V, Dutertre CA, Ventre Ε, Vu Manh TP, Baranek T, Storset AK, Marvel J, Boudinot P, Hosmalin A Schwartz-Cornil I, Dalod M "The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8α+ dendritic cells," J Exp Med, 207:1283-1292 (2010).
NPL 3: Dorner BG, Dorner MB, Zhou X Opitz C, Mora A, Güttler S, Hutloff A, Mages HW, Ranke K, Schaefer M, Jack RS, Henn V, Kroczek RA "Selective expression of the chemokine receptor XCR1 on cross-presenting dendritic cells determines cooperation with CD8+ T-cells," Immunity, 31:823-833 (2009).
NPL 4: Bachem A, Güttler S, Hartung Ε, Ebstein F, Schaefer M, Tannert A, Salama A Movassaghi K, Opitz C, Mages HW, Henn V, Kloerzel PM Gurka S, Kroczek RA "Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells," J Exp Med, 207:1273-1281 (2010).
NPL 5: Kurts C, Robinson BW, Knolle PA "Cross-priming in health and disease," Nat Rev Immunol, 10:403414 (2010).
NPL 6: Kurts C, Robinson BW, Knolle PA "Cross-priming in health and disease," Nat Rev Immunol, 10:403414 (2010).
NPL 7: Waldner H, "The role of innate immune responses in autoimmune disease development," Autoimmun, Rev 8:400404 (2009).
NPL 8: Khurram SA, Whawell SA, Bingle L, Murdoch C, McCabe BM, Farthing PM, "Functional expression of the chemokine receptor XCR1 on oral epithelial cells," J Pathol, 221: 153-63 (2010).
Краткое описание изобретения
Техническая задача
Сведения, что дендритные клетки участвуют в развитии, усилении и подобном, иммунопатологических заболеваний, таким образом, были накоплены в значительной степени при использовании животных моделей заболеваний. Несмотря на это к настоящему времени не были разработаны ни способы эффективного лечения, ни способы предупреждения для многих иммунопатологических заболеваний. Более того, хотя антитело к XCR1 человека, имеющее эффект ингибирования клеточной миграции, известно (Khurram SA, Whawell SA, Bingle L, Murdoch С, McCabe BM, Farthing PM, "Functional expression of the chemokine receptor XCR1 on oral epithelial cells," J Pathol, 221: 153-63 (2010)), но поскольку такое антитело представляет собой полученное от кролика поликлональное антитело, то маловероятно, что такое антитело может иметь прямое клиническое применение в качестве готовой лекарственной формы. Кроме того, вышеуказанный документ не предполагает, что такое антитело ингибирует миграцию дендритных клеток, и невозможно даже предположить, что такое антитело будет эффективным при лечении или предупреждении иммунопатологических заболеваний.
Цель настоящего изобретения состоит в том, чтобы предоставить моноклональное антитело, которое селективно связывается с XCR1 человека; предпочтительно моноклональное антитело, которое селективно связывается с XCR1 человека и ингибирует клеточную миграцию; еще более предпочтительно антитело, которое является эффективным при лечении или предупреждении иммунопатологических заболеваний, в частности иммунопатологических заболеваний кожи, которые основаны на вышеописанном эффекте.
Решение задачи
Авторы настоящего изобретения провели тщательные исследования в попытке решить вышеуказанную задачу. В результате они разработали антитела, которые связываются с XCR1 человека, и обнаружили, что такие антитела имеют эффект ингибирования клеточной миграции, а также значительный эффект при лечении или предупреждении иммунопатологических заболеваний, таких как иммунопатологические заболевания кожи, связанные с миграцией дендритных клеток.
Далее в настоящем документе, в настоящем описании, вышеописанные антитела иногда просто упоминаются как "антитела", "антитела по настоящему изобретению" или "антитела к XCR1 человека".
Преимущественные эффекты изобретения
Антитела по настоящему изобретению связываются с XCR1 человека. Антитела по настоящему изобретению включают антитело, которое ингибирует связывание между XCR1 человека и XCL1 человека. Такое антитело можно использовать в качестве активного ингредиента, который можно добавлять к ингибитору связывания между XCR1 человека и XCL1 человека.
Антитела по настоящему изобретению также включают антитело, которое ингибирует клеточную миграцию, в частности миграцию дендритных клеток. Такое антитело можно использовать в качестве активного ингредиента, который можно добавлять к ингибитору клеточной миграции, в частности к ингибитору миграции дендритных клеток. Кроме того, антитела по настоящему изобретению также включают антитело, которое специфически распознает BDCA3-положительные (также называемые CD141) клетки. Таким образом, фармацевтическую композицию, содержащую антитела по настоящему изобретению, можно использовать в качестве терапевтического средства для лечения иммунопатологических заболеваний, связанных с клеточной миграцией, в частности миграцией дендритных клеток. В частности, фармацевтическую композицию можно использовать в качестве терапевтического средства для лечения иммунопатологических заболеваний кожи, таких как гиперчувствительность замедленного типа, псориаз, парапсориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит, миозит с тельцами включения, аутоиммунное заболевание, проявляющееся появлением волдырей (например, обыкновенная пузырчатка, пемфигоид или приобретенный буллезный эпидермолиз), пустулезные высыпания, системная склеродермия, герпес беременных, линейный IgA-зависимый буллезный дерматоз, очаговая алопеция, вульгарное витилиго, кожные заболевания, связанные с коллагенозом (например, системная красная волчанка, синдром Шегрена или смешанная форма заболевания соединительной ткани), кожные заболевания, связанные с болезнью Аддисона, кожные заболевания, связанные с реакцией "трансплантат против хозяина" (GVHD), экзема и крапивница.
В дополнение к этим иммунопатологическим заболеваниям кожи антитела по настоящему изобретению также можно использовать в качестве терапевтических средств для лечения иммунопатологических заболеваний, таких как сахарный диабет 1 типа гломерулонефрит, аутоиммунный гепатит, рассеянный склероз, анкилозирующий спондилоартрит, тиреоидит, отторжение трансплантата, болезнь Крона, ревматоидный артрит, воспалительное заболевание кишечника, передний увеит, гранулематоз Вегенера или болезнь Бехчета.
Краткое описание графических материалов
[Фиг. 1] На Фиг. 1 показаны результаты FACS-анализа реактивности мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) в отношении клеток В300.19, экспрессирующих XCR1 человека-EGFP.
[Фиг. 2] На Фиг. 2 показаны результаты анализа акцентированного на хемотаксисе исследования нейтрализующего действия мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) на индуцируемую лимфотактином миграцию у человека клеток В300.19, экспрессирующих XCR1 человека-EGFP.
[Фиг. 3] На Фиг. 3 показаны результаты FACS-анализа реактивности гуманизированных мышиных антител к XCR1 человека (HK1L2 и HK5L5) в отношении клеток В300.19, экспрессирующих XCR1 человека-EGFP, в сравнении с реактивностью их исходного мышиного антитела к XCR1 человека (5G7) и их химерного антитела.
[Фиг. 4] На Фиг. 4 показаны результаты FACS-анализа реактивности мышиного антитела к XCR1 человека (5G7) и гуманизированных антител к XCR1 человека (HK1L2 и HK5L5) по отношению к BDCA3+ дендритным клеткам человека.
[Фиг. 5] На Фиг. 5 показаны результаты акцентированного на хемотаксисе исследования нейтрализующего действия гуманизированных антител к XCR1 человека (HK1L2 и HK5L5) на индуцируемую лимфотактином миграцию у человека клеток В300.19, экспрессирующих XCR1 человека-EGFP, в сравнении с нейтрализующим действием их исходного мышиного антитела к XCR1 человека (5G7) и его химерного антитела.
[Фиг. 6] На Фиг. 6 показаны результаты анализа акцентированного на трансэндотелиальной миграции исследования нейтрализующего действия гуманизированных антител к XCR1 человека (HK1L2 и HK5L5) и химерного антитела на индуцируемую лимфотактином миграцию у человека BDCA3+ дендритных клеток человека в сравнении с контрольным изотипическим антителом (IgG2, к человека).
[Фиг. 7] На Фиг. 7 показано сравнение аминокислотных последовательностей CDR1-3 тяжелой цепи и CDR1-3 легкой цепи антител по настоящему изобретению. На фигуре также показаны обобщенные аминокислотные последовательности CDR1-3 тяжелой цепи и CDR1-3 легкой цепи.
[Фиг. 8] На Фиг. 8 показан фармакологический эффект мышиного антитела к XCR1 человека (5G7) по настоящему изобретению на мышиной модели аллергической реакции замедленного типа - контактного дерматита (DTH).
На Фиг. 8A и 8B, соответственно, показаны результаты, полученные при сравнении степени опухания уха (мм) через 24 часа и 48 часов после индукции DNFB для мышиного антитела к XCR1 человека (5G7) по настоящему изобретению и контрольного антитела.
[Фиг. 9] На Фиг. 9 показана специфичность связывания мышиного антитела к XCR1 человека (5G7) по настоящему изобретению с различными рецепторами хемокинов человека. На фигуре на графике по оси абсцисс показана интенсивность флуоресценции фикоэритрина (РЕ).
[Фиг. 10] На Фиг. 10 показаны аминокислотные последовательности XCR1 человека, с которым связываются антитела по настоящему изобретению.
[Фиг. 11] На Фиг. 11 показана цитотоксичность в отношении клеток человека, экспрессирующих XCR1, при применении антител по настоящему изобретению.
[Фиг. 12] На Фиг. 12 показан анализ результата исследования цитотоксичности Т-лимфоцитов, проведенного в отношении мышиного антитела к XCR1 человека (5G7) по настоящему изобретению.
[Фиг. 13] На Фиг. 13 показана реактивность мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) по настоящему изобретению в отношении химерных клеток человека/мыши, экспрессирующих XCR1.
[Фиг. 14] На Фиг. 14 показан анализ результата картирования сайтов связывания мышиных антител к XCR1 человека (2Н6 и 5G7) с внеклеточными доменами XCR1 человека при помощи методики пептидного сканирования ELISA.
[Фиг. 15] На Фиг. 15 показан анализ результата картирования сайтов связывания поликлональных мышиных антител к XCR1 человека с внеклеточными доменами XCR1 человека с применением аланиновых мутантов.
[Фиг. 16] На Фиг. 16 показан анализ результата картирования сайтов связывания мышиного антитела к XCR1 человека (2Н6) с внеклеточными доменами XCR1 человека с применением аланиновых мутантов.
[Фиг. 17] На Фиг. 17 показан анализ результата картирования сайтов связывания мышиного антитела к XCR1 человека (5G7) с внеклеточными доменами XCR1 человека с применением аланиновых мутантов.
[Фиг. 18] На Фиг. 18 показан анализ результата картирования сайтов связывания мышиного антитела к XCR1 человека (11Н2) с внеклеточными доменами XCR1 человека с применением аланиновых мутантов.
[Фиг. 19] На Фиг. 19 показан анализ результата картирования сайтов связывания гуманизированного мышиного антитела к XCR1 человека (HK1L2) с внеклеточными доменами XCR1 человека с применением аланиновых мутантов.
[Фиг. 20] На Фиг. 20 показан анализ результата картирования сайтов связывания гуманизированного мышиного антитела к XCR1 человека (HK5L5) с внеклеточными доменами XCR1 человека с применением аланиновых мутантов.
[Фиг. 21] На Фиг. 21 показан анализ результата конкуренции мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) за связывание с клетками человека, экспрессирующими XCR1.
[Фиг. 22] На Фиг. 22 показана специфичность связывания мышиного моноклонального антитела к XCR1 человека (5G7) и коммерческого поликлонального антитела козы к XCR1 человека с различными рецепторами хемокинов человека. На фигуре на графике по оси абсцисс показана интенсивность флуоресценции фикоэритрина (РЕ).
[Фиг. 23] На Фиг. 23 показана специфичность связывания мышиного антитела к XCR1 человека (5G7) и гуманизированных моноклональных антител к XCR1 человека (HK1L2 и HK5L5) с различными рецепторами хемокинов человека На фигуре на графике по оси абсцисс показана интенсивность флуоресценции фикоэритрина (РЕ).
[Фиг. 24] На Фиг. 24 показан фармакологический эффект мышиного антитела к XCR1 человека (5G7) по настоящему изобретению на мышиной модели реакции замедленного типа - контактного дерматита (DTH), индуцированного Mycobacterium butyricum.
[Фиг. 25] На Фиг. 25 показан фармакологический эффект мышиного антитела к XCR1 человека (5G7) по настоящему изобретению на мышиной модели рассеянного склероза (MS), вызванного экспериментальным аутоиммунным энцефаломиелитом (ЕАЕ).
[Фиг. 26] На Фиг. 26 показан анализ результата исследования конкурентного связывания лиганда, проведенного в отношении мышиных антител к XCR1 человека по настоящему изобретению.
Описание вариантов осуществления
Различные методики, применяемые для выполнения настоящего изобретения, основаны на известных документах и т.п., которые свободно и гарантировано доступны специалисту в данной области техники, за исключением тех методик, источники которых явно указаны в данном документе. Например, в отношении методик генной инженерии и молекулярной биологии может быть представлена ссылка на документы, такие как Sambrook and Russell, "Molecular Cloning: A Laboratory Manual," Cold Spring Harbor Laboratory Press, New York (2001) и Ausubel, FM et al., "Current Protocols in Molecular Biology," John Wiley & Sons, New York, NY.
Кроме того, в отношении методик конструирования антител может быть представлена ссылка на документы, такие как Kabat et al., "Sequences of Proteins of Immunological Interest," U.S. Department of Health and Human Services (1983) и Konterman and Dübel, "Antibody Engineering," Springer.
Разъяснение терминов
Термин "нуклеиновая кислота" охватывает, например, рибонуклеотиды, дезоксирибонуклеотиды и их модифицированные формы. Нуклеиновая кислота может быть либо одно-, либо двухцепочечной и либо полинуклеотидом, либо олигонуклеотидом.
Термин "белок" относится к соединению, в котором две или более аминокислоты связаны пептидными связями.
Термин "моноклональное антитело" относится к антителу, полученному из популяции в значительной мере гомогенных антител. Другими словами, отдельные антитела, входящие в популяцию, идентичны, за исключением встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными и направлены на единственный участок антигена. Кроме того, в отличие от препаратов на основе поликлональных антител, содержащих различные антитела, направленные на различные детерминанты (эпитопы), каждое моноклональное антитело направлено на единственную антигенную детерминанту на антигене. В дополнение к их специфичности, моноклональные антитела также имеют преимущество в том, что их можно синтезировать без примесей других антител. Определение "моноклональный" относится к характеристике антитела, полученного из популяции в значительной мере гомогенных антител, и его не следует интерпретировать как означающее, что антитела должны быть произведены любым конкретным способом.
Например, моноклональное антитело, которое следует использовать в соответствии с настоящим изобретением, можно получить гибридомным способом, впервые описанным в Köhler G and Milstein С, "Continuous cultures of fused cells secreting antibody of predefined specificity," Nature, 256: 495-7 (1975), или способом рекомбинантных ДНК (см. патент США №4816567).
Кроме того, "моноклональные антитела" можно выделять из фаговой библиотеки антител с помощью методики, например, описанной в Clackson Τ, Hoogenboom HR, Griffiths AD, и Winter G, "Making antibody fragments using phage display libraries," Nature, 352: 624-8 (1991) или Marks JD, Hoogenboom HR, и Bonnert TP, McCafferty J, Griffiths AD, Winter G, "By-passing immunization: Human antibodies from V-gene libraries displayed on phage," J Mol Biol, 222: 581-97 (1991).
"Идентичность" аминокислотных последовательностей или нуклеотидных последовательностей относится к степени идентичности аминокислотных последовательностей или нуклеотидных последовательностей между двумя или более сопоставимыми аминокислотными последовательностями или нуклеотидными последовательностями. Таким образом, когда идентичность двух аминокислотных последовательностей или нуклеотидных последовательностей высока, идентичность или сходство этих последовательностей также высока. Уровень идентичности аминокислотных последовательностей или нуклеотидных последовательностей определяют, например, с помощью FASTA, который представляет собой средство анализа последовательностей, с установленными по умолчанию параметрами.
Альтернативно, его можно определить с помощью алгоритма BLAST от Karlin и Altschul (Karlin S, Altschul SF, "Methods for assessing the statistical significance of molecular sequence features by using general scoring schemes," Proc Natl Acad Sci USA 87: 2264-2268 (1990) и Karlin S, Altschul SF, "Applications and statistics for multiple high-scoring segments in molecular sequences," Proc Natl Acad Sci USA, 90: 5873-7 (1993)). Программы, такие как BLASTN и BLASTX, основанные на вышеописанном алгоритме BLAST, разработаны (Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ, “Basic local alignment search tool,” J Mol Biol, 215:403-10 (1990)). Например, BLASTN можно использовать для анализа нуклеотидных последовательностей с установлением в качестве параметров, например, score - 100 и wordlength - 12.
Кроме того, BLASTX можно использовать для анализа аминокислотной последовательности с установлением в качестве параметров, например, score - 50 и wordlength - 3.
При использовании программ BLAST и Gapped BLAST для каждой программы можно использовать параметры по умолчанию. Известны конкретные методики для этих способов анализа. Может быть представлена ссылка на веб-сайт Национальный центр биотехнологической информации (NCBI) (http://www.ncbi.nlm.nih.gov/).
Антитело к XCR1 человека
Антитела по настоящему изобретению являются выделенными антителами.
Антитела по настоящему изобретению связываются с XCR1 человека. Аминокислотная последовательность XCR1 человека представляет собой аминокислотную последовательность, приведенную в NCBI Reference Sequence: NP_001019815.1 или NP_005274.1. В отношении этих аминокислотных последовательностей можно предоставить ссылку на веб-сайты NCBI (соответственно,
http://www.ncbi.nlm.nih.gov/protein/NP_001019815.1 и
http://www.ncbi.nlm.nih.gov/protein/NP_005274.1).
Специфичное антитело по первому варианту осуществления настоящего изобретения представляет собой антитело, содержащее вариабельную область тяжелой цепи, которая содержит
CDR 1 тяжелой цепи, описанный в (А) или (а) ниже,
CDR 2 тяжелой цепи описанный в (В) или (b) ниже, и
CDR 3 тяжелой цепи, описанный в (С) или (с) ниже; и
вариабельную область легкой цепи, которая содержит
CDR 1 легкой цепи, описанный в (D) или (d) ниже,
CDR 2 легкой цепи, описанный в (Е) или (е) ниже, и
CDR 3 легкой цепи, описанный в (F) или (f) ниже.
CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 53,
CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 54,
CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 55;
CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 56,
CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 57, и
CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 58.
CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 41,
CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 42,
CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 43;
CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 44,
CDR 2 легкой цепи, отстоящий из аминокислотной последовательности SEQ ID NO: 45, и
CDR 3 легкой цепи, отстоящий из аминокислотной последовательности SEQ ID NO: 46.
Термин "CDR", определенный по отношению к антителам по настоящему изобретению, представляет собой сокращение от Complementarity Determining Region и также упоминается как гипервариабельный участок. CDR находятся в вариабельной области иммуноглобулина и в значительной степени участвуют в специфическом связывании антитела с антигеном. Кроме того, выражение "CDR легкой цепи" относится к CDR, которые находятся в вариабельной области легких цепей иммуноглобулина, а выражение "CDR тяжелой цепи" относится к CDR, которые находятся в вариабельной области тяжелых цепей иммуноглобулина.
Кроме того, выражение "вариабельная область" относится к области, которая содержит вышеописанные CDR 1-CDR 3 (далее в данном документе называемые просто "CDR 1-3"). Хотя порядок расположения CDR 1-3 конкретно не ограничен, предпочтительно CDR 1, CDR 2 и CDR 3 расположены в таком порядке или в обратном порядке от N-конца до С-конца непрерывным образом или через другие аминокислотные последовательности, которые называются каркасными областями (FR).
Кроме того, "вариабельная область тяжелой цепи" представляет собой область, где расположены вышеописанные CDR 1-3 тяжелой цепи, а "вариабельная область легкой цепи" представляет собой область, где расположены вышеописанные CDR1-3 легкой цепи.
Как описано выше, область, отличная от вышеописанных CDR 1-3 в каждой вариабельной области, называют каркасной областью (FR). В частности, область между N-концом и CDR 1 в каждой вариабельной области определена как FR 1, область между CDR 1 и CDR 2 определена как область FR 2, область между CDR 2 и CDR 3 определена как FR 3, и область между CDR3 и С-концом в каждой вариабельной области определена как FR 4.
FR также выполняют функцию линкерных последовательностей для соединения CDR 1-3, которые особенно важны как антигенраспознающие последовательности. FR представляют собой области, которые вносят вклад в формирование трехмерной структуры всей вариабельной области.
Предпочтительно антитело по первому варианту осуществления согласно настоящему изобретению является антителом, содержащим вариабельную область тяжелой цепи, содержащую
CDR 1 тяжелой цепи из (g) ниже, (m) ниже или (а) выше,
CDR 2 тяжелой цепи из (h) ниже, (n) ниже или (b) выше и
CDR 3 тяжелой цепи из (i) ниже, (о) ниже или (с) выше; и
вариабельную область легкой цепи, содержащую
CDR 1 легкой цепи из (j) ниже, (р) ниже или (d) выше,
CDR 2 легкой цепи из (k) ниже, (q) ниже или (e) выше и
CDR 3 легкой цепи из (l) ниже, (r) ниже или (f) выше.
(g) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 17,
(h) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 18,
(i) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 19;
(j) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 20,
(k) CDR 2 легкой цепи, отстоящий из аминокислотной последовательности SEQ ID NO: 21,
(l) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 22;
(m) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 29,
(n) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 30,
(o) CDR 3 тяжелой цепи, отстоящий из аминокислотной последовательности SEQ ID NO: 31;
(p) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 32,
(q) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 33, и
(r) CDR 3 легкой цепи, отстоящий из аминокислотной последовательности SEQ ID NO: 34.
CDR 3 тяжелой цепи, описанный в (i), и CDR 3 тяжелой цепи, описанный в (о), содержит идентичные аминокислотные последовательности.
Антитело по второму варианту осуществления настоящего изобретения представляет собой антитело, содержащее
вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (А)-(С) выше, или
вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (а)-(с) выше; и
вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (D)-(F) выше, или
вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (d)-(f) выше.
Наиболее предпочтительное антитело по второму варианту осуществления представляет собой антитело, содержащее любое из следующего:
вариабельная область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (g)-(i) выше,
вариабельная область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (m)-(о) выше, и
вариабельная область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (а)-(с) выше; и любое из следующего:
вариабельная область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (j)-(l) выше,
вариабельная область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (p)-(r) выше, и
вариабельная область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (d)-(f) выше.
Антитело по третьему варианту осуществления настоящего изобретения представляет собой антитело, содержащее
вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (А)-(С) выше, и
вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (D)-(F) выше, или антитело, содержащее
вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (а)-(с) выше, и
вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (d)-(f) выше.
Более предпочтительное антитело по третьему варианту осуществления представляет собой антитело, содержащее
вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (g)-(i) выше, и
вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (j)-(l) выше;
антитело, содержащее
вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (m)-(о) выше, и
вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (p)-(r) выше; или
антитело, содержащее
вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (а)-(с) выше, и
вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (d)-(f) выше.
Структуры молекул антител по настоящему изобретению не ограничены таковой иммуноглобулина, поскольку антитела имеют вышеописанные вариабельные области тяжелой и легкой цепи. Примеры конкретных структур охватывают молекулярные структуры F(ab')2, которые не включают Fc-область; Fab, образующегося в результате расщепления папаином молекулы иммуноглобулина и состоящего из СН1- и CL-домена, а так же вариабельной области тяжелой и легкой цепи; Fv, который не включает константную область иммуноглобулина; и scFv, который представляет собой одноцепочечный Fv антитела.
Антитела по настоящему изобретению также могут быть поливалентными антителами, в которых вышеуказанные молекулярные структуры объединены. Такое поливалентное антитело образуют с помощью методики накопительного сбора scFv-конструкций, как в случае scFv-Fc-конструкции, образованной объединением Fc-области и scFv-конструкции, описанной выше, и конструкции, называемой мини-антитело, образованной объединением СН3-домена константной области и scFv-конструкции, описанной выше. Термин "поливалентный" относится к наличию множества антигенсвязывающих участков. Что касается антител по настоящему изобретению, этот термин используется в том же значении и для обозначения наличия множества участков, которые связываются с XCR1 человека.
Антитела по настоящему изобретению также могут иметь константную область человека в дополнение к вышеописанным вариабельным областям тяжелой и легкой цепи.
В иммуноглобулине "константная область" тяжелых цепей содержит домены с названием CH1, СН2 и СН3, а "константная область" легких цепей содержит домен с названием CL.
Как изложено выше, если у антител по настоящему изобретению есть константная область, то предпочтительно, чтобы вариабельная область тяжелой цепи была соединена по меньшей мере с одним из СН1-, СН2- и СН3-доменов, и чтобы вариабельная область легкой цепи была соединена с CL. Кроме того, предпочтительно, чтобы вариабельная область тяжелой цепи была непосредственно соединена с СН1.
Константная область антител по настоящему изобретению является константной областью, полученной из иммуноглобулина человека, предпочтительно константной областью, полученной из иммуноглобулина IgG человека. Изотип иммуноглобулина IgG человека конкретно не ограничен и может быть подходящим образом выбран, например, в зависимости от необходимости наличия ADCC-активности, CDC-активности и другой активности у антител, описанных ниже.
Термин "ADCC-активность" представляет собой сокращение от Antibody-Dependent Cellular Cytotoxicity activity (антителозависимая клеточная цитотоксичность). Она представляет собой активность, при которой клетки, такие как NK-клетки, экспрессирующие рецепторы, специфичные в отношении Fc-области антитела, связываются с антителами и повреждают клетки, находящиеся вблизи антител.
Кроме того, термин "CDC-активность" представляет собой сокращение от Complement-Dependent Cytotoxicity activity (комплементзависимая цитотоксичность). У человека изотипом TgQ, имеющим высокую ADCC- и/или CDC-активность, является IgG1, а изотипом IgG, имеющим низкую ADCC- и/или CDC-активность, является IgG2 или IgG4.
Аминокислотные остатки в Fc-области антител по настоящему изобретению можно мутировать для того, чтобы вызвать изменение ADCC- и/или CDC-активности. Мутации, которые можно вводить, конкретно не ограничены, и можно вводить известные мутации. Например, следующие мутации можно вводить в константную область IgG1 с целью увеличения ADCC-активности: S239D, D32E, S239D/I332E, S239D/I332E7A330L и другие (Lazar GA, Dang W, Karki S, Vafa O, Peng JS, Hyun L, Chan С, Chung HS, Eivazi A, Yoder SC, Vielmetter J, Carmichael DF, Hayes RJ, Dahiyat BL "Engineered antibody Fc variants with enhanced effector function," Proc Natl Acad Sci USA, 103: 4005-10 (2006)) и S298A, K334A, S298A/K334A, S298A/E333A/K334A и т.д. (Shields RL, Namenuk AK, Hong K, Meng YG, Rae J, Briggs J, Xie D, Lai J, Stadlen A, Li B, Fox JA, Presta LG, "High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R," J Biol Chem, 276:6591-604 (2001)).
Примеры мутаций, которые увеличивают CDC-активность, включают S267E, H268F, S324T, S267E/H268F, S267E/S324T, H268F/S324T, S267E/H268F/S324T (Moore GL, Chen H, Karki S, Lazar GA, “Engineered Fc variant antibodies with enhanced ability to recruit complement and mediate effector functions," MAbs, 2:181-9 (2010)).
Кроме того, с целью снижения ADCC-активности можно вводить известные мутации, например V234A/G237A (Cole MS, Anasetti С, Tso JY, “Human IgG2 variants of chimeric anti-CD3 are nonmitogenic to Τ cells," J Immunol, 159:3613-21 (1997)), H268Q/V309L/A330S/P331S (An Z, Forrest G, Moore R, Cukan M, Haytko P, Huang L, Vitelli S, Zhao JZ, Lu Ρ, Hua J, Gibson CR, Harvey BR, Montgomery D, Zaller D, Wang F, Strohl W, "IgG2m4, an engineered antibody isotype with reduced Fc runction," MAbs, 1:572-9 (2009)) и другие.
Нумерация вышеописанных аминокислот, которые можно мутировать, соответствует системе нумерации EU (см. Sequences of proteins of immunological interest, NIH Publication No. 91-3242).
Химерное антитело
Среди антител по настоящему изобретению антитело, у которого вариабельные области тяжелой и легкой цепи содержат аминокислотные последовательности, полученные из видов, не являющихся человеком, а константная область содержит аминокислотные последовательности, полученные из человека, определено как "химерное антитело".
В первом варианте осуществления химерное антитело по настоящему изобретению представляет собой химерное антитело, содержащее тяжелую цепь, состоящую из аминокислотной последовательности SEQ ID NO: 13, и легкую цепь из SEQ ID NO: 14.
Как показано в Таблице 5, аминокислотная последовательность SEQ ID NO: 13 содержит CDR 1-3 тяжелой цепи из SEQ ID NO: 17-19 среди CDR 1-3 тяжелой цепи, приведенные выше в описании вариабельной области тяжелой цепи. Далее, как показано в Таблице 5, аминокислотная последовательность SEQ ID NO: 14 содержит CDR 1-3 легкой цепи из SEQ ID NO: 20-22 из CDR 1-3 легкой цепи, приведенных выше в описании вариабельной области легкой цепи.
Химерное антитело по настоящему изобретению включает варианты, полученные посредством мутаций в тяжелой цепи, состоящей из аминокислотной последовательности SEQ ID NO: 13, и/или легкой цепи, состоящей из аминокислотной последовательности SEQ ID NO: 14, при условии, что такие мутации не лишают химерное антитело способности связываться с XCR1 человека.
Такие варианты тяжелой и легкой цепей предпочтительно получают путем ведения мутаций по меньшей мере в одну из FR 1-FR 4 (далее в настоящем документе называемые просто "FR 1-4") вариабельной области или по меньшей мере в один участок константной области с соответствующими аминокислотным последовательностям SEQ ID NO: 13 и 14.
Определенное число мутаций, вводимых в тяжелую и легкую цепи, конкретно не ограничено. Мутации обычно вводят для получения варианта, характеризующегося 85% или более высокой идентичностью, предпочтительно 90% или более высокой идентичностью, более предпочтительно 95% или более высокой идентичностью и наиболее предпочтительно 99% или более высокой идентичностью аминокислотной последовательности перед мутацией.
Термин "мутация", используемый в данном документе, включает замену, делецие, вставку и т.п. Известный способ, без конкретных ограничений, можно применять в качестве конкретного способа введения мутаций. Например, в случае замены можно использовать консервативную замену. Термин "консервативная замена" относится к замене аминокислотного остатка другим аминокислотным остатком, имеющим сходную боковую цепь.
Например, замена между аминокислотными остатками с основными боковыми цепями, такими как лизин, аргинин и гистидин, соответствует консервативной замене. Кроме того, следующие замены аминокислотных остатков также соответствуют консервативным заменам: замены между аминокислотными остатками с кислотными боковыми цепями, такими как аспарагиновая кислота и глутаминовая кислота; замены между аминокислотными остатками с незаряженными полярными боковыми цепями, такими как глицин, аспарагин, глютамин, серии, треонин, тирозин и цистеин; замены между аминокислотными остатками с неполярными боковыми цепями, такими как аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метеонин и триптофан; замены между аминокислотными остатками с β-разветвленными боковыми цепями, такими как треонин, валин и изолейцин; и замены между аминокислотными остатками с ароматическими боковыми цепями, такими как тирозин, фенилаланин, триптофан и гистидин.
Гуманизированные антитела
Среди антител по настоящему изобретению антитело, включающее вышеописанные CDR 1-3 в вариабельных областях тяжелой и легкой цепи, в которых FR 1-4 содержат аминокислотную последовательность, полученную из человека, или ее вариант, определено как "гуманизированное антитело".
Такие FR, содержащие аминокислотную последовательность, полученную из человека, конкретно не ограничены и могут быть определены, основываясь на известной методике.
Примеры таких FR включают полностью человеческие каркасные области или части областей, при этом предпочтительно, чтобы FR были получены из человеческих последовательностей зародышевого типа. Соответственно, можно предоставить ссылку, например, на веб-сайт NCBI, на котором представлен перечень известных в настоящее время последовательностей FR в качестве примеров полностью человеческих каркасных областей или частей областей.
Неограничивающие примеры последовательностей вариабельных областей тяжелой цепи человека включают VH1-18, VH1-2, VH1-24, VH1-3, VH1-45, VH1-46, VH1-58, VH1-69, VH1-8, VH2-26, VH2-5, VH2-70, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-33, VH3-35, VH3-38, VH3-43, VH348, VH349, VH3-53, VH3-64, VH3-66, VH3-7, VH3-72, VH3-73, VH3-74, VH3-9, VH4-28, VH4-31, VH4-34, VH4-39, VH4-4, VH4-59, VH4-61, VH5-51, VH6-1 и VH7-81.
Неограничивающие примеры последовательностей вариабельных областей легкой цепи человека включают VL1-11, VL1-13, VL1-16, VL1-17, VL1-18, VL1-19, VL1-2, VL1-20, VL1-22, VL1-3, VL-4, VL1-5, VL1-7, VL1-9, VL2-1, VL2-11, VL2-13, VL2-14, VL2-15, VL2-17, VL2-19, VL2-6, VL2-7, VL-8, VL3-2, VL3-3, VL34, VL4-1, VL4-2, VL4-3, VL4-4, VL4-6, VL5-1, VL5-2, VL5-4 и VL5-6.
Полностью человеческие FR выбраны из FR, кодируемых данными функциональными генами зародышевого типа человека. Каждая из этих FR обычно отличается за счет модификации ограниченного количества аминокислот. Эти FR можно использовать в комбинации с CDR, описанными в настоящем описании. Неограничивающие дополнительные примеры FR человека, которые можно использовать в комбинации с вышеописанными CDR, включают KOL, NEWM, REI, EU, TUR, TEL LAY и ТОМ. В отношении примеров таких FR человека можно предоставить ссылку на следующие документы: Kabat, et al., "Sequences of Proteins of Immunological Interest," US Department of Health and Human Services, NIH (1991) USA; Wu TT, Kabat EA "An analysis of the sequences of the variable regions of Bence Jones proteins and myeloma light chains and their implications for antibody complementarity," J Exp Med, 132:211-50 (1970) и другие.
В первом варианте осуществления гуманизированное антитело по настоящему изобретению представляет собой гуманизированное антитело, содержащее вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 60 или SEQ ID NO: 64, и вариабельную область легкой цепи из SEQ ID NO: 68 или SEQ ID NO: 72.
Более предпочтительным вариантом осуществления является гуманизированное антитело, содержащее вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 60, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 68, или гуманизированное антитело, содержащее вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 64, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 72.
Как соответственно показано в Таблицах 11-1 и 12-1, аминокислотные последовательности SEQ ID NO: 60 и SEQ ID NO: 64 содержат CDR 1-3 тяжелой цепи из SEQ ID NO: 17-19 из вышеописанных CDR 1-3 тяжелой цепи в вариабельной области тяжелой цепи. Как соответственно показано в Таблицах 13-1 и 14-1, аминокислотные последовательности SEQ ID NO: 68 и SEQ ID NO: 72 содержат CDR 1-3 легкой цепи из SEQ ID NO: 20-22 из вышеописанных CDR 1-3 легкой цепи в вариабельной области легкой цепи.
Гуманизированное антитело по настоящему изобретению включает варианты, полученные посредством мутаций в вариабельной области тяжелой цепи, содержащей аминокислотные последовательности SEQ ID NO: 60 или 64, и/или в вариабельной области легкой цепи, содержащей аминокислотные последовательности SEQ ID NO: 68 или 72, при условии, что такая мутация не устраняет способность связываться с XCR1 человека. Такие варианты вариабельных областей тяжелой и легкой цепи предпочтительно получают посредством введения мутаций в соответствующие FR1-4.
Определенное число мутаций в вариабельных областях тяжелой и легкой цепи конкретно не ограничено. Мутации обычно вводят для получения варианта, характеризующегося 85% или более высокой идентичностью, предпочтительно 90% или более высокой идентичностью, более предпочтительно 95% или более высокой идентичностью и наиболее предпочтительно 99% или более высокой идентичностью аминокислотной последовательности перед мутацией.
Термин "мутация", используемый в данном документе, включает замену, делецию, вставку и т.п. Как и в случае с химерным антителом, описанным выше, консервативные замены и т.п. можно использовать в качестве конкретного способа для введения мутаций.
Во втором варианте осуществления гуманизированное антитело по настоящему изобретению включает антитело, содержащее константную область человека. Примеры такового включают гуманизированное антитело, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 59 или 63, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 67 или 71.
Более предпочтительным вариантом осуществления является гуманизированное антитело, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 59, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 67, или гуманизированное антитело, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 63, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 71.
Как показано в Таблице 11-1, аминокислотная последовательность SEQ ID NO: 59 содержит аминокислотную последовательность, соответствующую вариабельной области тяжелой цепи из SEQ ID NO: 60, и, таким образом, содержит CDR 1-3 тяжелой цепи из SEQ ID NO: 17-19 из CDR 1-3 тяжелой цепи, описанных выше. Далее, как показано в Таблице 13-1, аминокислотная последовательность SEQ ID NO: 67 содержит аминокислотную последовательность, соответствующую вариабельной области легкой цепи из SEQ ID NO: 68, и, таким образом, содержит CDR 1-3 легкой цепи из SEQ ID NO: 20-22 из CDR1-3 легкой цепи, описанных выше.
Тяжелая и/или легкая цепи, описанные выше, включают варианты, полученные посредством мутаций, при условии, что такие мутации не устраняют способность связываться с XCR1 человека. Такие варианты тяжелых и легких цепей предпочтительно получают путем введения мутаций в FR1-4 или константную область.
Определенное число мутаций, вводимых в тяжелую и легкую цепи, конкретно не ограничено. Мутации обычно вводят для получения варианта, характеризующегося 85% или более высокой идентичностью, предпочтительно 90% или более высокой идентичностью, более предпочтительно 95% или более высокой идентичностью и наиболее предпочтительно 99% или более высокой идентичностью аминокислотной последовательности перед мутацией.
Термин "мутация", используемый в данном документе, включает замену, делецию, вставку и т.п. Как и в случае с химерным антителом, описанным выше, консервативные замены и т.п. можно использовать в качестве конкретного способа для введения мутаций.
Функция антител
Антитела по настоящему изобретению связываются с XCR1 человека. Значение термина "связывается", используемого в данном документе, охватывает по меньшей мере связывание посредством гидрофобных связей и т.п., как можно увидеть в случае взаимодействия между белками. Другими словами, , которые связываются с XCR1 человека по меньшей мере посредством гидрофобного связывания, являются подходящими в качестве антител по настоящему изобретению. Кроме того, комплекс антитела по настоящему изобретению и XCR1 человека может диссоциировать или не диссоциировать после связывания.
Антитела по настоящему изобретению предпочтительно связываются с XCR1 человека. Термин "специфическое связывание", используемый в данном документе, относится к специфическому связыванию с XCR1 человека и означает, что антитела преимущественно связываются с XCR1 человека в том случае, если XCR1 человека присутствует одновременно с другими молекулами, кроме XCR1 человека, в частности с молекулами, имеющими структуру, сходную с таковой XCR1 человека, такими как гомолог XCR1 человека или ортолог XCR1 человека
То, что антитела по настоящему изобретению специфически связываются с XCR1 человека, не означает, что способность связываться с вышеописанным гомологом или ортологом XCR1 человека исключена.
Степень связывания антител по настоящему изобретению с XCR1 человека можно оценить с помощью константы скорости реакции, такой как значение Kd, Koff или Kon. Значение Kd является значением, полученным делением значения Koff на значение Kon.
Константа скорости реакции между антителами по настоящему изобретению и XCR1 человека конкретно не ограничена.
Антитела по настоящему изобретению связываются с внеклеточным доменом XCR1 человека. В частности, антитела связываются с одним или несколькими аминокислотными участками 1-31,90-103,168-190 и 251-267, которые соответствуют области внеклеточного домена с аминокислотной последовательностью (SEQ ID NO: 91, Фиг. 10) из вышеописанной NCBI Reference Sequence: NP_001019815.1 или NP_005274.1.
Более предпочтительно в аминокислотной последовательности SEQ ID NO: 91 антитела связываются по меньшей мере с тремя аминокислотами, выбранными из группы, состоящей из 8-й, 11-й, 12-й, 13-й, 14-й, 16-й, 17-й, 22-й, 23-й, 176-й и 177-й аминокислоты.
Указанные "по меньшей мере три аминокислоты" включают, например, три или более аминокислоты, четыре или более аминокислоты, пять или более аминокислот, шесть или более аминокислот, семь или более аминокислот, восемь или более аминокислот, девять или более аминокислот, десять или более аминокислот или одиннадцать аминокислот.
Нужно отметить, что "эпитоп" в настоящем изобретении также упоминается как "антигенная детерминанта" и включает "линейный эпитоп" и "эпитоп прерывистого типа". "Линейный эпитоп" представляет собой эпитоп, который распознается скорее с помощью первичной структуры аминокислотной последовательности антител, нежели с помощью их конформационной структуры. "Эпитоп прерывистого типа" представляет собой эпитоп, который распознается с помощью конформационной структуры аминокислотной последовательности антител, основанной на структуре более высокого порядка.
Нужно отметить, что специалист в данной области может определить эпитоп антител по настоящему изобретению, соответственно видоизменяя способы, описанные в Примерах настоящего изобретения. Например, эпитоп можно определять посредством синтезирования белка или пептида, состоящего из желаемой аминокислотной последовательности, которая охватывает аминокислотную последовательность внеклеточного домена XCR1 человека, используя известный способ, и подтверждения связывания между полученным белком или пептидом и антителом известным способом. Альтернативно, эпитоп можно определять, получая мутант посредством введения подходящей мутации в желаемые аминокислотные положения аминокислотной последовательности XCR1 человека известным способом и подтверждая, является ли связывание между полученным мутантом и антителом более слабым.
Как описано выше, поскольку антитела по настоящему изобретению связываются с XCR1 человека, антитела по настоящему изобретению также включают антитело, которое ингибирует связывание между XCR1 человека и XCL1 человека. XCL1 человека также упоминается как лимфотактин (Ltn) человека или лимфотактин-α (Ltn-α) человека. Такая ингибиторная активность иногда называется "нейтрализующей активностью", индуцируемой антителами по настоящему изобретению. Поскольку XCR1 человека как рецепторный белок присутствует на поверхности клеток in vivo, ингибирование связывания XCR1 человека и XCL1 с помощью антител по настоящему изобретению преимущественно происходит на поверхности клеток. Не имеет значения, обладают ли антитела по настоящему изобретению ингибирующей активностью в отношении связывания между XCR1 человека и XCL2, если антитела обладают активностью, чтобы по меньшей мере ингибировать связывание между XCR1 человека и XCL1. Таким образом, антитела по настоящему изобретению также включают антитело, которое ингибирует связывание не только между XCR1 человека и XCL1, но также между XCR1 человека и XCL2.
Примеры предпочтительных клеток включают клетки, связанные с иммунной системой, которые активируются связыванием между XCR1 человека и XCL1 человека, при этом дендритные клетки являются особенно предпочтительными. В частности, как показано в нижеприведенных примерах, так как антитела по настоящему изобретению специфически распознают BDCA3+ дендритные клетки, которые представляют собой дендритные клетки, экспрессирующие значительное количество белков XCR1 человека, предпочтительно, чтобы антитела характеризовались эффектом ингибирования связывания между XCR1 человека и XCL1 человека на BDCA3+ дендритных клетках.
Связывание между XCR1 человека и XCL1 человека ингибируется антителами по настоящему изобретению. Неограничивающие примеры форм такого ингибирования включают следующие:
(1) Антитела по настоящему изобретению связываются с XCR1 в участке, с которым первоначально должен связываться XCL1 человека, создавая пространственное препятствие для связывания с XCL1 человека, что приводит к ингибированию связывания между XCR1 человека и XCL1 человека.
(2) Антитела по настоящему изобретению связываются с XCR1 человека, вызывая изменение в трехмерной структуре XCR1 человека, что, как следствие, вызывает изменение в структуре XCR1 человека, с которым должен связываться XCL1 человека, что в результате приводит к ингибированию связывания между XCR1 человека и XCL1 человека.
(3) Антитела по настоящему изобретению связываются с XCR1, вызывая интернализацию рецептора, что приводит к ингибированию связывания между XCR1 человека и XCL1 человека.
Ингибиторную активность антител по настоящему изобретению в отношении связывания XCR1 человека и XCL1 человека оценивают, основываясь на значениях IC50 или IC90. Данные значения можно получить, например, при выполнении эксперимента по конкурентному ингибированию и т.п. для оценки связывания XCL1 человека с XCR1 человека, используя клетки, которые экспрессируют XCR1 человека, в присутствии антител по настоящему изобретению. В качестве конкретного способа для такого эксперимента по конкурентному ингибированию можно использовать известный способ.
Антитела по настоящему изобретению включают антитело, которое характеризуется эффектом ингибирования клеточной миграции. Термин "клеточная миграция" относится к явлению, при котором клетки активно мигрируют в ответ на внешние стимулы, посылаемые клеткам, и индуцируемую стимулом активацию внутриклеточных механизмов передачи сигнала. Эффекты, оказываемые активной клеточной миграцией, варьируют в зависимости от функций клеток. Например, в случае клеточной миграции дендритных клеток такая клеточная миграция представляет собой явление, которое служит одним из механизмов в иммунной системе. В настоящем изобретении ингибиторная активность в отношении клеточной миграции иногда упоминается как "нейтрализующая активность".
Как описано выше, вследствие того, что антитела по настоящему изобретению соответствующим образом ингибируют связывание между XCR1 человека и XCL1 человека в дендритных клетках, в частности в BDCA3+ дендритных клетках, антитела особенно предпочтительно ингибируют миграцию дендритных клеток, в частности BDCA3+ дендритных клеток.
XCR1 человека представляет собой семикомпонентный трансмембранный рецептор, сопряженный с G-белком. Когда XCL1 человека связывается с XCR1 человека, трехмерная структура XCR1 человека изменяется и, как результат, G-белок, сопряженный с внутриклеточным доменом XCR1 человека, высвобождается, и сигнал передается внутрь клеток.
Антитела по настоящему изобретению предупреждают высвобождение G-белков посредством ингибирования связывания между XCR1 человека и XCL1 в соответствии с вышеописанными формами (1), (2) и подобным им. В результате ни один сигнал не передается, что, таким образом, ингибирует явление клеточной миграции.
В качестве альтернативы, явление миграции клеток может быть ингибировано в результате механизма, при котором связывание антител по настоящему изобретению с XCR1 человека усиливает связь между XCR1 человека и G-белком, сопряженным с внутриклеточным доменом XCR1 человека, при этом высвобождение G-белков в результате не происходит, что таким образом ингибирует внутриклеточную передачу сигнала
Ингибиторная активность антител по настоящему изобретению в отношении клеточной миграции клеток человека оценивают на основе значения IC50 или IC90. Определенные значения конкретно не ограничены. Например, значение IC50 обычно составляет приблизительно 0,36 нМ или менее, предпочтительно приблизительно 0,27 нМ или менее и более предпочтительно приблизительно 0,16 нМ или менее. Например, значение IC90 обычно составляет приблизительно 2,38 нМ или менее, предпочтительно приблизительно 1,52 нМ или менее, и более предпочтительно приблизительно 0,86 нМ или менее.
Антитела по настоящему изобретению включают как вариант осуществления антитело, которое характеризуется эффектом снижения активности цитотоксических Т-лимфоцитов (CTL). Механизм снижения активности CTL заключается, например, в том, что антитела по настоящему изобретению ингибируют взаимодействие между XCR1 человека и XCL1 человека в дендритных клетках. Среди дендритных клеток вышеописанные BDCA3+ дендритные клетки являются предпочтительными.
Способ получения антител по настоящему изобретению
Антитела по настоящему изобретению можно получать с помощью способа, включающего следующие три этапа, хотя он не ограничен ими.
(i) Этап 1 введения вектора в хозяина для того, чтобы трансформировать хозяина, при этом вектор содержит нуклеиновую кислоту, включающую нуклеотидную последовательность, кодирующую антитела по настоящему изобретению;
(ii) Этап 2 культивирования трансформированного хозяина, полученных на этапе 1, и сбор фракции, содержащей антитела, которые связываются с XCR1 человека; и
(iii) Этап 3 выделения или очистки вышеуказанных антител из фракции, полученной на этапе 2.
Этап 1
Нуклеиновая кислота, используемая на этапе 1, является нуклеиновой кислотой, которая кодирует антитела по настоящему изобретению. Нуклеотидную последовательность вышеуказанной нуклеиновой кислоты можно определить при помощи методики in silico, основываясь на информации об аминокислотной последовательности антител по настоящему изобретению. В то же время, предпочтительно определить нуклеотидную последовательность в отношении частоты кодонов в хозяине, используемых на этапе 2. Конкретные примеры нуклеотидных последовательностей включают нуклеотидные последовательности SEQ ID NO: 3, 4, 7, 8, 11, 12, 15, 16, 61, 62, 65, 66, 69, 70, 73 или 74 или их вариант.
Вышеуказанный вариант предпочтительно создают посредством введения мутаций (деления, замена, вставка или подобное) в FR или константную область антител.
Определенное число мутаций, вводимых в вариант, конкретно не ограничено. Мутации обычно вводят для получения варианта, характеризующегося 85% или более высокой идентичностью, предпочтительно 90% или более высокой идентичностью, более предпочтительно 95% или более высокой идентичностью и наиболее предпочтительно 99% или более высокой идентичностью аминокислотной последовательности перед мутацией.
Кроме того, вышеуказанная нуклеиновая кислота может включать нуклеотидную последовательность, расположенную у 5'-конца, которая кодирует сигнальный пептид для секреции. Конкретная нуклеотидная последовательность, кодирующая сигнальный пептид для секреции, предпочтительно представляет собой нуклеотидную последовательность, которая эффективно экспрессируется в сигнальный пептид для секреции в клетках-хозяевах, используемых на этапе 2. Термин "сигнальный пептид для секреции" относится к пептиду, содержащему аминокислотную последовательность, которая выполняет роль узнаваемой последовательности для того, чтобы направлять белки или пептиды, продуцируемые в хозяине, в транспортный путь для секреции белков или пептидов во внешнюю среду хозяина.
Примеры нуклеотидных последовательностей, кодирующих сигнальный пептид для секреции, включают:

Вектор, используемый на этапе 1, содержит по меньшей мере одну из вышеуказанных нуклеиновых кислот.
Такой вектор может быть одним из следующих векторов:
(I) вектор, содержащий нуклеиновую кислоту, которая содержит нуклеотидную последовательность, кодирующую по меньшей мере один элемент, выбранный из группы, состоящей из тяжелых цепей, вариабельной области тяжелой цепи и CDR 1-3 тяжелой цепи антител по настоящему изобретению;
(II) вектор, содержащий нуклеиновую кислоту, которая содержит нуклеотидную последовательность, кодирующую по меньшей мере один элемент, выбранный из группы, состоящей из легких цепей, вариабельной области легкой цепи и CDR 1-3 легкой цепи антител по настоящему изобретению; или
(III) вектор, содержащий нуклеиновую кислоту, которая содержит нуклеотидную последовательность, кодирующую по меньшей мере один элемент, выбранный из группы, состоящей из тяжелых цепей, вариабельной области тяжелой цепи и CDR 1-3 тяжелой цепи антител по настоящему изобретению, и нуклеиновую кислоту, которая содержит нуклеотидную последовательность, кодирующую по меньшей мере один элемент, выбранный из группы, состоящей из легких цепей, вариабельной области легкой цепи и CDR 1-3 легкой цепи антитела по настоящему изобретению.
Вышеуказанный вектор может быть вектором экспрессии гена "Вектор экспрессии гена" является вектором, обладающим функцией запуска экспрессии нуклеотидной последовательности вышеуказанной нуклеиновой кислоты. Вектор экспрессии гена может содержать последовательность промотора, последовательность энхансера, последовательность репрессора, последовательность инсулятора и т.п. для того, чтобы осуществлять регуляцию экспрессии нуклеотидной последовательности.
Эти последовательности конкретно не ограничены при условии, что они функционируют в вышеописанном хозяине.
Хозяин, используемый на этапе 1, конкретно не ограничен при условии, что вышеуказанный ген экспрессируется. Примеры хозяев включают клетки насекомых, клетки эукариот и клетки млекопитающих. Из этих клеток клетки HEK, клетки СНО, клетки NS0 или клетки SP2/O, которые являются клетками млекопитающих, особенно предпочтительны с точки зрения более эффективной экспрессии нуклеотидной последовательности, кодирующей антитела.
Методика введения вышеуказанного вектора в хозяина на этапе 1 конкретно не ограничена Можно использовать известную методику. Векторы, приведенные в (I)-(III) выше, можно вводить в хозяина отдельно или в комбинации из двух или более.
Хозяина с вышеуказанным вектором можно получить с помощью такой методики. Вектор может содержаться как в хозяине, так и содержаться в нем таким образом, что нуклеиновая кислота, включающая нуклеотидную последовательность, кодирующую антитела, в векторе внедрена в геном хозяина. Существование полученного хозяина можно поддерживать, используя известную методику, и в случае необходимости можно хранить в течение длительного периода времени.
Этап 2
Этап 2 представляет собой этап культивирования вышеописанного хозяина, полученного на этапе 1, и сбора фракции, содержащей антитела по настоящему изобретению, которые связываются с XCR1 человека. Культивирование хозяина, содержащего вышеописанный вектор, позволяет хозяину экспрессировать нуклеотидную последовательность, кодирующую антитела по настоящему изобретению, на основе нуклеиновой кислоты в векторе, что в результате приводит к продуцированию антител по настоящему изобретению. Продуцируемые антитела накапливаются в хозяине или в среде, используемой для культивирования хозяина.
На этапе 2 можно использовать известный способ в качестве методики для сбора фракции, содержащей антитела по настоящему изобретению. Например, для сбора фракции, содержащей антитела по настоящему изобретению, из хозяев их разрушают физическими или химическими средствами и раствор, полученный в результате разрушения, подвергают обработке для разделения твердой и жидкой фаз, таким образом получая жидкую фракцию. Полученную жидкую фракцию можно использовать как фракцию, содержащую антитела по настоящему изобретению.
Иначе, для сбора фракции, содержащей антитела по настоящему изобретению, из среды, используемой для культивирования хозяина, среду, т.е. культуральный раствор хозяина, полученного на этапе 1, подвергают обработке для разделения твердой и жидкой фаз, таким образом получая жидкую фракцию. Полученную жидкую фракцию можно использовать как фракцию, содержащую антитела по настоящему изобретению.
Для упрощения этапа выделения или очистки в последующем этапе 3 предпочтительно собирать фракцию, содержащую антитела по настоящему изобретению, из культурального раствора хозяина.
Среда, используемая для культивирования на этапе 2, конкретно не ограничена при условии, что среда позволяет хозяину экспрессировать нуклеотидную последовательность, кодирующую антитела по настоящему изобретению, таким образом продуцируя антитела по настоящему изобретению. Однако при сборе фракции, содержащей антитела по настоящему изобретению, из культурального раствора хозяина, как описано выше, предпочтительно использовать бессывороточную среду для максимального упрощения этапа выделения или очистки на последующем этапе 3.
Что касается различных условий, используемых во время культивирования хозяина, таких как контейнер, температура, время, концентрация в среде и культуральные условия, можно применять условия, используемые в известном способе для получения антител.
Этап 3
Этап 3 представляет собой этап выделения или очистки антител по настоящему изобретению, которые связываются с XCR1 человека, из фракции, полученной на этапе 2. Способ выделения и очистки антител по настоящему изобретению конкретно не ограничен. Широкое применение имеет способ, обычно используемый для выделения или очистки белка.
Применение антител по настоящему изобретению в медицине (1) Применение в качестве терапевтических средств для лечения иммунопатологических заболеваний
Как описано выше, антитела по настоящему изобретению характеризуются эффектом ингибирования явления клеточной миграции дендритных клеток, связанных с иммунной системой. Основываясь на этом эффекте, антитела по настоящему изобретению, в частности гуманизированные антитела, можно использовать в качестве активного ингредиента фармацевтической композиции, которая клинически применима для человека.
Заболевания, при которых можно применять антитела по настоящему изобретению, разъяснены ниже.
Подходящие заболевания (иммунопатологические заболевания)
XCR1 человека на высоком уровне экспрессируется в случае человека в CD141+ дендритных клетках и в случае мышей в CD8α+ дендритных клетках. Эти дендритные клетки активируют Т-клетки, используя вышеописанный способ представления антигена, называемый кросс-презентацией (Bachem A, Güttler S, Hartung Ε, Ebstein F, Schaefer M, Tannert A, Salama A, Movassaghi K, Opitz C, Mages HW, Henn V, Kloetzel PM, Gurka S, Kroczek RA, "Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells," J Exp Med, 207: 1273-1281 (2010)).
Кроме того, поскольку источник продуцирования XCL1, который является лигандом для XCR1 человека, включает Т-клетки, в частности CD8+ Т-клетки, система хемокинов, в которую вовлечен комплекс XCL1-XCR1, осуществляет регуляцию индуцируемой дендритными клетками активации CD8+ Т-клеток (Crozat K, Guiton R, Contreras V, Feuillet V, Dutertre CA, Ventre E, Vu Manh TP, Baranek T, Storset AK, Marvel J, Boudinot P, Hosmalin A, Schwartz-Cornil I, Dalod M, "The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8α+ dendritic cells," J Exp Med, 207: 1283-1292 (2010); и Dorner BG, Dorner MB, Zhou X, Opitz C, Mora A, Güttler S, Hutloff A, Mages HW, Ranke K, Schaefer M, Jack RS, Henn V, Kroczek RA, "Selective expression of the chemokine receptor XCR1 on cross-presenting dendritic cells determines cooperation with CD8+ T-cells," Immunity, 31: 823-833 (2009)).
Как описано выше, антитела по настоящему изобретению включают как вариант осуществления антитело, которое проявляет ингибирующий эффект в отношении связывания между XCL1 человека и XCR1 человека в дендритных клетках, в частности BDCA3+ дендритных клетках. Таким образом, антитела по настоящему изобретению можно использовать в качестве терапевтических средств для лечения иммунопатологических заболеваний, в развитии которых участвуют Т-клетки, активируемые миграцией дендритных клеток. В частности, антитела можно использовать в качестве терапевтических средств для лечения заболеваний, связанных с регуляцией активации CD8+ Т-клеток.
Как описано выше, антитела по настоящему изобретению включают как вариант осуществления антитело, которое проявляет ингибирующий эффект в отношении снижения активности CTL. CTL обладают механизмом для активации иммунной системы посредством повреждения клеток или тканей. При различных иммунных заболеваниях, как известно, усиливается активность CTL; таким образом, антитела по настоящему изобретению можно использовать в качестве терапевтического средства для лечения иммунных заболеваний, посредством снижения активности CTL.
Неофаничивающие примеры таких заболеваний включают сахарный диабет 1 типа, псориаз, гломерулонефрит, аутоиммунный гепатит, рассеянный склероз, анкилозирующий спондилоартрит, тиреоидит, отторжение трансплантата, гиперчувствительность замедленного типа, болезнь Крона, дерматомиозит, полимиозит, миозит с тельцами включения, ревматоидный артрит, воспалительное заболевание кишечника, передний увеит, гранулематоз Вегенера, реакцию "трансплантат против хозяина" и болезнь Бехчета (Kurts С, Robinson BW, Knolle PA, "Cross-priming in health and disease," Nat Rev Immunol, 10: 403-114 (2010); Kehren J, Desvignes С, Krasteva M, Ducluzeau MT, Assossou O, Horand F, Hahne M, Kägi D, Kaiserlian D, Nicolas JF. "Cytotoxicity is mandatory for CD8(+) Τ cell-mediated contact hypersensitivity," J Exp. Med. 189:779-786 (1999); Middel P, Thelen P, Blaschke S, Polzien F, Reich K, Blaschke V, Wrede A, Hummel KM, Gunawan B, Radzun HJ, “Expression of the T-cell chemoattractant chemokine lymphotactin in Crohn's disease," Am J Pathol, 159: 1751-1761 (2001); Sugihara T, Sekine C, Nakae T, Kohyama K, Harigai M, Iwakura Y, Matsumoto Y, Miyasaka N, Kohsaka H, "A new murine model to define the critical pathologic and therapeutic mediators of polymyositis," Arthritis Rheum, 56: 1304-1314 (2007); Wang CR, Liu MF, Huang YE, Chen HC, "Up-regulation of XCR1 expression in rheumatoid joints," Rheumatology (Oxford) 43: 569-573 (2004); Muroi E, Ogawa F, Shimizu K, Komura K, Hasegawa M, Fujimoto M, Sato S, "Elevation of serum lymphotactin levels in patients with systemic sclerosis," J Rheumatol, 35: 834-838 (2008); Torrence AE, Brabb T, Viney JL, Bielefeldt-Ohmann H, Treuting P, Seamons A, Drivdahl R, Zeng W, Maggio-Price L, "Serum biomarkers in a mouse model of bacterial-induced inflammatory bowel disease," Inflamm Bowel Dis, 14: 480-190 (2008); Yeh PT, Lin FA, Lin CP, Yang CM, Chen MS, Yang CH, "Expressions of lymphotactin and its receptor, XCR in Lewis rats with experimental autoimmune anterior uveitis," Graefes Arch Clin Exp Ophthalmol, 248:1737-1747 (2010); Blaschke S, Brandt P, Wessels JT, Müller GA, "Expression and function of the C-class chemokine lymphotactin (XCL1) in Wegener's granulomatosis," J Rheumatol, 36:2491-2500 (2009); Asuka H, Okazaki Y, Kawakami Y, Hirakata M, Inoko H, Ikeda Y, Kuwana M, "Autoreactive CD8+ cytotoxic Τ lymphocytes to major histocompatibility complex class I chain-related gene A in patients with Behçet's disease," Arthritis Rheum, 50: 3658-3662 (2004); Serody JS, Burkett SE, Panoskaltsis-Mortari A, Ng-Cashin J, McMahon E, Matsushima GK, Lira SA, Cook DN, Blazar BR, "T-lymphocyte production of macrophage inflammatory protein-1alpha is critical to the recruitment of CD8(+) Τ cells to the liver, lung, and spleen during graft-versus-host disease," Blood, 96:2973-2980 (2000); Sugihara Τ, Sekine С, Nakae T, Kohyama K, Harigai M, Iwakura Y, Matsumoto Y, Miyasaka Ν, Kohsaka H, "A new murine model to define the clinical pathologic and therapeutic mediators of polymyositis," Arthritis & Rheumatism, 56:1304-1314 (2007)); и Dalakas MC, "Review: An update on inflammatory and autoimmune myopathies," Neuropathol Appl Neurobiol, 37:226-242 (2011).
Также было обнаружено, что антитело (мышиное антитело к XCR1 человека (5G7)) по настоящему изобретению значительно ингибирует реакцию DTH в нижеописанном эксперименте, в котором использовалась мышиная модель развития гиперчувствительности замедленного типа (далее в данном документе иногда называемая "DTH"). Как описано выше, гиперчувствительность замедленного типа является заболеванием, которое известно как одно из иммунопатологических заболеваний, в развитии которых участвуют CD8+ Т-клетки, активируемые миграцией дендритных клеток. То обстоятельство, что антитела по настоящему изобретению эффективны при лечении гиперчувствительности замедленного типа, свидетельствует о том, что антитела по настоящему изобретению обладают активностью ингибировать клеточную миграцию, в частности миграцию дендритных клеток, поскольку антитела по настоящему изобретению воздействуют на CD8+ Т-клетки.
Кроме того, в дополнение к гиперчувствительности замедленного типа атопический дерматит и контактный дерматит также известны как иммунопатологические заболевания кожи, в развитии которых участвует реакция DTH (Fabrizi G, Romano A, Vultaggio P, Bellegrandi S, Paganelli R, Venuti A, “Heterogeneity of atopic dermatitis defined by the immune response to inhalant and food allergy," Eur J Dermatol, 9: 380-384 (1999); и Fonacier LS, Dreskin SC, Leung DYM, "Allergic skin diseases," J Allergy Clin Immunol, 125: S138-149 (2010)).
Основываясь на вышесказанном, антитела по настоящему изобретению можно использовать в качестве терапевтических средств для лечения иммунопатологических заболеваний кожи, таких как атопический дерматит или контактный дерматит.
Было указано, что активация CD8+ Т-клеток также может участвовать в реакции DTH (Mody CH, Pain III R, Jackson С, Chen G-H, Toews GB, "CD8 Cells play a critical role in delayed-type hypersensitivity to intact Cryptococcus neoformans," J Immunol, 152: 3970-3979 (1994) и т.д.).
Инвазия CD8+ Т-клеток в эпидерму наблюдается при псориазе, который является аутоиммунным кожным заболеванием, поражающим большое количество пациентов, в частности в форме псориатических очагов. Эти клетки, как полагают, являются главными эффекторными клетками, которые вызывают появление псориатических очагов (Gudjonsson JE, Johnston A, Sigmundsdottir H, Valdimarsson H, "Immunopathogenic mechanisms in psoriasis," Clin Exp Immunol, 135:1-8 (2004)).
Основываясь на вышесказанном, антитела по настоящему изобретению можно использовать в качестве терапевтических средств для лечения иммунопатологических заболеваний кожи, в развитии которых участвует активация CD8+ Т-клеток.
В дополнение к гиперчувствительности замедленного типа, атопическому дерматиту и контактному дерматиту неограничивающие примеры иммунопатологических заболеваний кожи, в развитии которых участвуют CD8+ Т-клетки, также включают дерматомиозит, полимиозит, миозит с тельцами включения, псориаз, парапсориаз, аутоиммунные заболевания, проявляющиеся появлением волдырей (например, обыкновенная пузырчатка, пемфигоид или приобретенный буллезный эпидермолиз), пустулезные высыпания, герпес беременных, линейный IgA-зависимый буллезный дерматоз, очаговую алопецию, вульгарное витилиго, кожные заболевания, связанные с коллагенозом (например, системная красная волчанка, синдром Шегрена или смешанная форма заболевания соединительной ткани), кожные заболевания, связанные с болезнью Аддисона, кожные заболевания, связанные с реакцией "трансплантат против хозяина" (GVHD), экзему и крапивницу.
В этом документе ниже описана взаимосвязь между антителами по настоящему изобретению и различными иммунопатологическими заболеваниями (рассеянный склероз, сахарный диабет 1 типа человека, гломерулонефрит, аутоиммунный гепатит, тиреоидит, реакция "трансплантат против хозяина", дермагомиозит, полимиозит и миозит с тельцами включения). Заболевания, при которых антитела по настоящему изобретению эффективны, не ограничены следующими конкретными заболеваниями.
Рассеянный склероз
Инвазия CD8+ Т-клеток в дополнение к инвазии CD4+ Т-клеток в очаги в центральной нервной системе при рассеянном склерозе у человека была описана некоторое время назад. Кроме того, сообщалось, что в эксперименте с использованием мышей имплантация CD8+ Т-клеток, активизированных антигеном из миелиновой оболочки нервных волокон центральной нервной системы, вызывает развитие экспериментального аутоиммунного энцефаломенингита, который является моделью рассеянного склероза человека. По сравнению с обычной моделью вышеуказанная модель весьма точно имитирует патологию рассеянного склероза человека (повторяющиеся обострения и ремиссии, значительная демиелинизация, инвазия большого количества CD8+ Т-клеток и макрофагов/клеток микроглии в очаги демиелинизации). Как описано выше, было выдвинуто предположение, что CD8+ Т-клетки играют важную роль в развитии рассеянного склероза у человека и в мышиной модели рассеянного склероза (Friese MA, Fugger L, "Autoreactive CD8+ cells in multiple sclerosis: a new target for therapy?" Brain, 128:1747-1763 (2005)).
Таким образом, антитела по настоящему изобретению, которые осуществляют регуляцию активации CD8+ Т-клеток, можно использовать в качестве терапевтических средств для лечения рассеянного склероза.
Сахарный диабет 1 типа человека
На мыши с диабетом без ожирения (NOD), которая представляет модель сахарного диабета 1 типа человека, было показано, что истощение CD8+ Т-клеток приводит к предупреждению возникновения сахарного диабета (Wang В, Gonzales A, Benoist С, Mathis D, "The role CD8+ T-cells in the initiation of insulin-dependent diabetes mellitus," Eur J Immunol, 26: 1762-1769 (1996)). Это означает, что CD8+ Т-клетки также участвуют в развитии патологии сахарного диабета 1 типа.
Таким образом, антитела по настоящему изобретению, которые осуществляют регуляцию активации CD8+Т-клеток, можно использовать в качестве терапевтических средств для лечения сахарного диабета 1 типа у человека.
Гломерулонефрит
На мышиной модели гломерулонефрита было показано, что CD8+ Т-клетки участвуют в процессе формирования почечных повреждений (Heymann F, Meyer-Schwesinger С, Hamilton-Williams ЕЕ, Hammerich L, Panzer W, Kaden S, Quaggin SE, Floege J, Gröne H-J, Kurts C, "Kidney dendritic cell activation is required for progression of renal disease on a mouse model of glomerular injury," J Clin Invest, 119: 1286-1297 (2009)). Инвазия большого количества CD8+ Т-клеток в ткани почки наблюдается у пациентов с тяжелой формой аутоиммунного волчаночного нефрита. Сообщалось о наличии корреляции между количеством CD8+ Т-клеток, и увеличением индекса активности процесса в почках, и уровнем креатинина в сыворотке крови, которые указывают на ухудшение функции почек (Couzi L, Merville Ρ, Deminière С, Moreau J-F, Combe С, Pellegrin J-L, Viallard J-F, Blanco P, "Predominance of CD8+ Τ lymphocytes among periglomerular infiltrating cells and link to the prognosis of class III and class IV lupus nephritis," Arthritis Rheum, 56:2362-2370 (2007)). Как описано выше, CD8+ Т-клетки, как полагают, участвуют в возникновении аутоиммунного гломерулонефрита или прогрессировании этой патологии у человека и в мышиных моделях.
Таким образом, антитела по настоящему изобретению, которые осуществляют регуляцию активации CD8+ Т-клеток, можно использовать в качестве терапевтических средств для лечения гломерулонефрита.
Аутоиммунный гепатит
Было выдвинуто предположение, что инфицирование вирусом гепатита С (HCV) участвует в процессе развития аутоиммунного гепатита. Также предполагалось, что CD8+ CTL, активированные в отношении HCV, участвуют в развитии аутоиммунного гепатита, путем удаления HCV и повреждения инфицированных клеток печени (Kammer AR, van der Burg SH, Grabscheid B, Hunziker IP, Kwappenberg KMC, Reichen J, Melief CJM, Cerny A, "Molecular mimicity of human cytochrome P450 by hepatitis С virus at the level of cytotoxic Τ cell recognition," J Exp Med, 190:169-175 (1999)).
Таким образом, антитела по настоящему изобретению, которые осуществляют регуляцию активации CD8+ Т-клеток, можно использовать в качестве терапевтических средств для лечения аутоиммунного гепатита.
Тиреоидит
CD8+ CTL, как известно, участвуют в развитии экспериментального аутоиммунного тиреоидита (EAT), который является мышиной моделью тиреоидита человека (например, болезнь Хашимото). Также сообщалось, что у мышей в модели обнаруживаются повреждения, подобные тиреоидиту человека (в периферической крови обнаруживаются антитела к тиреоглобулину, и наблюдается инвазия CD8+ Т-клеток и CD4+ Т-клеток в ткани щитовидной железы). Как описано выше, было выдвинуто предположение, что CD8+ Т-клетки участвуют в развитии тиреоидита у человека и в мышиных моделях (Brazillet М-Р, Batteux F, Abehsira-Amar O, Nicoletti F, Charreire J, "Induction of experimental autoimmune thyroiditis by heat-denatured porcine thyroglobulin: a Tc1-mediated disease," Eur J Immunol, 29: 1342-1352(1999)).
Таким образом, антитела по настоящему изобретению, которые осуществляют регуляцию активации CD8+ Т-клеток, можно использовать в качестве терапевтических средств для лечения тиреоидита у человека
Ревматоидный артрит
Как описано ниже в Примерах, 5G7, которое является одним из антител по настоящему изобретению, проявляет значительный эффект при лечении ревматоидного артрита в экспериментальной DTH, индуцированной Mycobacterium butyricum. Таким образом, антитела по настоящему изобретению можно использовать в качестве терапевтических средств для лечения ревматоидного артрита.
Отторжение трансплантата
CD8+ Т-клетки играют важную роль в отторжении трансплантата после трансплантации органа человека. Трансплантат отторгается в организме хозяина с помощью CD8+ Т-клеток, которые распознают МНС класса I, экспрессирующиеся в клетках трансплантата. Кроме того, сообщалось об инвазии большого количества CD8+ Т-клеток в ткани почки у пациентов, которым проводили трансплантацию почки и у которых происходило отторжение трансплантата. Как описано выше, было выдвинуто предположение, что центральную роль в отторжении трансплантата при трансплантации человеческих органов также играют CD8+ Т-клетки (Bueno V, Pestana JOM, "The role of CD8+ T-cells during allograft rejection," Braz J Med Biol Res, 35:1247-1258 (2002)).
Таким образом, антитела по настоящему изобретению, которые осуществляют регуляцию активации CD8+ Т-клеток, можно использовать в качестве терапевтических средств для лечения реакции "трансплантат против хозяина".
Дерматомиозит, полимиозит и миозит с тельцами включения
Когда лимфоциты, которые инвазируют место повреждения у пациентов с дерматомиозитом и полимиозитом, были установлены как клеточные линии, то клеточные линии CD8+ Т-клеток проявляли цитотоксичность в отношении своих собственных культивированных мышечных клеток. Это указывает на то, что поражение мышечных клеток у пациентов с вышеописанным миозитом вызвано CD8+ Т-клетками с антигенспецифической цитотоксичностью (Hohlfeld R, Engel AG, "Coculture with autologous myotubes of cytotoxic Τ cells isolated from muscle in inflammatory myopathies," Ann Neurol, 29: 498-507 (1991)). Кроме того, инвазия CD8+ Т-клеток в место повреждения наблюдалась у пациентов с миозитом с тельцами включения (Dalakas МС, "Review: An update on inflammatory and autoimmune myopathies," Neuropathol Appl Neurobiol, 37: 226-242 (2011)). Таким образом, антитела по настоящему изобретению, которые осуществляют регуляцию активации CD8+ Т-клеток, можно использовать в качестве терапевтических средств для лечения дерматомиозита, полимиозита или миозита с тельцами включения.
Как описано выше, поскольку антитела по настоящему изобретению можно использовать в качестве терапевтических средств для лечения иммунопатологических заболеваний, в частности иммунопатологических заболеваний кожи, настоящее изобретение предоставляет фармацевтическую композицию, содержащую антитела по настоящему изобретению.
Такую фармацевтическую композицию можно использовать в качестве терапевтического средства для лечения иммунопатологического заболевания, средства для целей лечения иммунопатологических заболеваний, в частности иммунопатологических заболеваний кожи.
Термин "лечение", используемый в данном документе, означает достижение желаемых фармакологических и/или физиологических эффектов. Эффекты включают эффект частичного или полного излечения заболевания и/или неблагоприятных эффектов, вызванных заболеванием (проявлений патологии и симптомов). Вышеуказанные эффекты также включают эффект торможения или замедления прогрессирования заболевания и/или неблагоприятных эффектов, вызванных заболеванием (проявлений патологии и симптомов); эффект смягчения проявлений патологии и симптомов (т.е. уменьшение интенсивности заболевания или симптомов или способность вызывать обратное развитие симптомов) и эффект предупреждения рецидивов заболевания. Вышеуказанные эффекты также включают эффект частичного или полного предупреждения возникновения заболевания и/или неблагоприятных эффектов, вызванных заболеванием (проявлений патологии и симптомов) у субъектов, которые имеют предрасположенность к заболеванию и/или неблагоприятным эффектам, вызываемых заболеванием (проявлений патологии и симптомов), но у которых наличие предрасположенности не было диагностировано.
В настоящем изобретении фармацевтическую композицию, содержащую антитела по настоящему изобретению, можно соответствующим образом применять для лечения иммунопатологических заболеваний человека, в частности иммунопатологических заболеваний кожи. Подразумевается, что вышеуказанная фармацевтическая композиция способна обеспечивать, например, эффект частичного или полного торможения различных симптомов иммунопатологических заболеваний (т.е. торможение или замедление прогрессирования); эффект смягчения различных симптомов иммунопатологических заболеваний (т.е. уменьшение интенсивности заболевания или симптомов или способность вызывать обратное развитие симптомов) или эффект предупреждения рецидивов различных симптомов иммунопатологических заболеваний.
Конкретные примеры целевых заболеваний описаны выше, при этом иммунопатологические заболевания кожи являются предпочтительными.
Содержание антител по настоящему изобретению в вышеуказанной фармацевтической композиции конкретно не ограничено при условии, что фармацевтическая композиция содержит эффективное количество антител по настоящему изобретению. Содержание антител можно специальным образом установить, например, таким образом, чтобы антитела по настоящему изобретению содержались в фармацевтической композиции в количестве от 0,001 до 99,99 вес. % относительно 100 вес. % композиции, принимая во внимание тип целевого иммунопатологического заболевания, лекарственной формы, способа введения и т.п.
Термин "эффективное количество", используемый в данном документе, относится к количеству, которое позволяет антителам по настоящему изобретению проявлять эффект ингибирования клеточной миграции дендритных клеток, или количеству, которое позволяет антителам демонстрировать вышеописанные желаемые фармакологические и/или физиологические эффекты (лечебный эффект в отношении иммунопатологических заболеваний).
Фармацевтически приемлемые носители или добавки можно добавлять к фармацевтической композиции в комбинации с антителами по настоящему изобретению. Термин "фармацевтически приемлемые носители или добавки", используемый в данном документе, относится к необязательным носителям, разбавителям, наполнителям, суспендирующим средствам, смазывающим веществам, адъювантам, связующим веществам, системам доставки, эмульгаторам, разрыхлителям, абсорбентам, консервантам, поверхностно-активным веществам, красителям, ароматизирующим веществам или подслащивающим веществам. Можно использовать известные носители или добавки.
Неограничивающие примеры лекарственных форм фармацевтической композиции включают таблетки, сиропы, линименты, инъекционные растворы и инфузионные растворы, при этом инъекционные растворы и инфузионные растворы являются предпочтительными. Такие инъекционные растворы и инфузионные растворы могут быть в водной, неводной или суспензионной форме. Кроме того, у фармацевтической композиции может быть лекарственная форма, которую готовят непосредственно перед введением.
Фармацевтическую композицию по настоящему изобретению, а именно терапевтическое средство для лечения иммунопатологического заболевания, можно использовать в способах лечения иммунопатологических заболеваний, включающих этап введения композиции субъекту, являющемуся человеком, с иммунопатологическим заболеванием, в частности иммунопатологическими заболеванием кожи. Как описано выше, фармацевтическую композицию также можно использовать в способах предупреждения иммунопатологических заболеваний, включающих введение композиции субъекту, являющемуся человеком, у которого не развились проявления патологии или симптомы иммунопатологического заболевания, в частности иммунопатологического заболевания кожи, но который может иметь предрасположенность к иммунопатологическому заболеванию.
Размер дозировки и способ введения фармацевтической композиции (терапевтического средства для лечения иммунопатологического заболевания) можно соответствующим образом установить в пределах диапазона от 0,001 до 100 мг/кг/день, учитывая тип иммунопатологического заболевания, пол субъекта, являющегося человеком, расу, возраст и общее состояние, тяжесть заболевания и т.п.
Антитела по настоящему изобретению можно вводить в вышеописанной дозировке один раз в день или в разделенной дозировке несколько раз в день. Кроме того, интервал введения антител в диапазоне, в котором они имеют лечебный эффект в отношении вышеописанных заболеваний, может быть следующим: один раз в день, один раз в два дня, один раз в неделю, один раз в две недели, один раз в 2-3 недели, один раз в месяц или один раз в 2-3 месяца. Неограничивающие примеры способов введения включают пероральное, внутримышечное, внутривенное, внутриартериальное, интратекальное, внутрикожное, внутрибрюшинное, интраназальное, внутрилегочное, внутриглазное, вагинальное, интрацервикальное, ректальное и подкожное введение.
Применение в качестве антитоксина
Антитела по настоящему изобретению могут быть конъюгированными с цитотоксическими молекулами. Поскольку такие антитела связываются с XCR1 человека, который экспрессируется в значительном количестве в дендритных клетках, которые связаны с иммунной системой, то антитела можно использовать в качестве антитоксинов, направленных против дендритных клеток.
Термин "цитотоксические молекулы", используемый в данном документе, относится к молекулам, которые проявляют эффекты, такие как апоптоз и/или некроз, которые вызывают гибель клеток.
Примеры таких молекул включают сапорин, рицин, эндотоксин Pseudomonas, дифтерийный токсин и химиотерапевтические средства. Связывание между антителом и токсическим веществом может быть осуществлено способом, который используется для получения обычных антитоксинов.
Другие применения антител по настоящему изобретению
Поскольку антитела по настоящему изобретению также включают как вариант осуществления антитело, которое связывается с XCR1, который экспрессируется в значительном количестве в дендритных клетках, то антитела можно использовать в способе обнаружения дендритных клеток. В этом случае для использования предпочтительно метить антитела по настоящему изобретению. Термин "метить", используемый в данном документе, относится к связыванию антител с мечеными молекулами, такими как флуоресцентные молекулы, люминесцентные молекулы, хромогенные молекулы и молекулы радиоизотопа.
Особенности связывания не ограничены при условии, что не происходит диссоциации связи на этапе обнаружения. Известный способ можно использовать в качестве конкретного способа обнаружения. Например, можно использовать методику проточной цитометрии.
Кроме того, антитела по настоящему изобретению могут также быть соответствующим образом применимыми в способах выделения и/или удаления дендритных клеток после обнаружения дендритных клеток. Известные способы можно также использовать для этих способов. Например, известное устройство для сортировки клеток можно соответствующим образом использовать в сочетании с методикой проточной цитометрии.
Настоящее изобретение относится к антителам, указанным выше, и широко охватывает варианты осуществления изобретения, которые описаны ниже.
Пункт 1
Антитело, связывающееся с XCR1 человека, где антитело связывается с линейными эпитопами или эпитопами прерывистого типа, которые содержат по меньшей мере три аминокислоты, выбранные из группы, состоящей из 8-й, 11-й, 12-й, 13-й, 14-й, 16-й, 17-й, 22-й, 23-й, 176-й и 177-й аминокислот в аминокислотной последовательности SEQ ID NO: 91.
Пункт 2
Антитело согласно вышеуказанному пункту 1, где антитело является
антителом, содержащим вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (g)-(i) ниже, и вариабельную область легкой цепи, которая содержит CDR1-3 легкой цепи описанные в (j)-(l) ниже;
антителом, содержащим вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (m)-(о) ниже, и вариабельную область легкой цепи, которая содержит CDR1-3 легкой цепи, описанные в (p)-(r) ниже; или
антителом, содержащим вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (a)-(c) ниже, и вариабельную область легкой цепи, которая содержит CDR1-3 легкой цепи, описанные в (d)-(f) ниже;
(a) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 41,
(b) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 42,
(c) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 43;
(d) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 44,
(e) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 45, и
(f) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 46;
(g) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 17,
(h) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 18,
(i) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 19;
(j) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 20,
(k) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 21,
(l) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 22;
(m) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 29,
(n) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 30,
(о) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 31;
(p) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 32,
(q) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 33, и
(r) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 34.
Пункт 3
Антитело согласно вышеуказанному пункту 1 или 2, где антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 60 или 64, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 68 или 72.
Пункт 4
Антитело согласно любому из вышеуказанных пунктов 1-3, где антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 60, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 68.
Пункт 5
Антитело согласно любому из вышеуказанных пунктов 1-3, где антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 64, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 72.
Пункт 6
Антитело согласно любому из вышеуказанных пунктов 1-5, где антитело содержит константную область человека.
Пункт 7
Антитело согласно любому из вышеуказанных пунктов 1-6, где антитело содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 59, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 67.
Пункт 8
Антитело согласно любому из вышеуказанных пунктов 1-6, где антитело содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 63, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:71.
Пункт 9
Антитело согласно любому из вышеуказанных пунктов 1-8, содержащее Fc-область, где Fc-область мутирована для того, чтобы вызвать изменение ADCC-активности.
Пункт 10
Антитело согласно вышеуказанному пункту 9, где Fc-область мутирована для снижения ADCC-активности.
Пункт 11
Антитело согласно любому из вышеуказанных пунктов 1-10, где антитело конъюгировано с цитотоксической молекулой.
Пункт 12
Антитело согласно любому из вышеуказанных пунктов 1-11, где антитело ингибирует взаимодействие между XCR1 человека и XCL1 человека
Пункт 13
Антитело согласно любому из вышеуказанных пунктов 1-12, где антитело ингибирует клеточную миграцию дендритных клеток.
Пункт 14
Антитело согласно любому из вышеуказанных пунктов 1-13, где антитело подавляет активность цитотоксических Т-лимфоцитов.
Пункт 15
Фармацевтическая композиция, содержащая антитело согласно любому из вышеуказанных пунктов 1-14 и фармацевтически приемлемый носитель или добавку.
Пункт 16
Фармацевтическая композиция согласно вышеуказанному пункту 15, где фармацевтическая композиция является терапевтическим средством для лечения иммунопатологического заболевания.
Пункт 17
Фармацевтическая композиция согласно вышеуказанному пункту 16, где иммунопатологическим заболеванием является иммунопатологическое заболевание кожи.
Пункт 18
Фармацевтическая композиция согласно вышеуказанному пункту 17, где иммунопатологическим заболеванием кожи является псориаз, парапсориаз, атопический дерматит, контактный дерматит, дермагомиозит, полимиозит, миозит с тельцами включения, аутоиммунное заболевание, проявляющееся появлением волдырей (обыкновенная пузырчатка, пемфигоид или приобретенный буллезный эпидермолиз), пустулезное высыпанием, герпес беременных, линейный IgA-зависимый буллезный дерматоз, очаговая алопеция, вульгарное витилиго, кожное заболевание, связанное с коллагенозом (системная красная волчанка, синдром Шегрена или смешанная форма заболевания соединительной ткани), кожное заболевание, связанное с болезнью Аддисона, кожное заболевание, связанное с реакцией "трансплантат против хозяина" (GVHD), экзема или крапивница.
Пункт 19
Фармацевтическая композиция согласно вышеуказанному пункту 17, где иммунопатологическим заболеванием кожи является псориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит или миозит с тельцами включения.
Пункт 20
Фармацевтическая композиция согласно вышеуказанному пункту 17, где иммунопатологическим заболеванием кожи является атопический дерматит или контактный дерматит.
Пункт 21
Фармацевтическая композиция согласно вышеуказанному пункту 16, где иммунопатологическим заболеванием кожи является тиреоидит, ревматоидный артрит, сахарный диабет 1 типа или рассеянный склероз.
Пункт 22
Нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую антитело согласно любому из вышеуказанных пунктов 1-14.
Пункт 23
Способ лечения иммунопатологического заболевания, включающий введение эффективного количества антитела согласно любому из вышеуказанных пунктов 1-14 или фармацевтической композиции согласно вышеуказанному пункту 15 человеку, пораженному иммунопатологическим заболеванием.
Пункт 24
Способ согласно вышеуказанному пункту 23, при котором иммунопатологическим заболеванием является иммунопатологическое заболевание кожи.
Пункт 25
Способ согласно вышеуказанному пункту 24, при котором иммунопатологическим заболеванием кожи является псориаз, парапсориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит, миозит с тельцами включения, аутоиммунное заболевание, проявляющееся появлением волдырей (обыкновенная пузырчатка, пемфигоид или приобретенный буллезный эпидермолиз), пустулезное высыпание, герпес беременных, линейный IgA-зависимый буллезный дерматоз, очаговая алопециея, вульгарное витилиго, кожное заболевание, связанное с коллагенозом (системная красная волчанка, синдром Шегрена или смешанная форма заболевания соединительной ткани), кожное заболевание, связанное с болезнью Аддисона, кожное заболевание, связанное с реакцией "трансплантат против хозяина" (GVHD), экзема или крапивница.
Пункт 26
Способ согласно вышеуказанному пункту 24, где иммунопатологическим заболеванием кожи является псориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит или миозит с тельцами включения.
Пункт 27
Способ согласно вышеуказанному пункту 23, при котором иммунопатологическим заболеванием кожи является тиреоидит, ревматоидный артрит, сахарный диабет 1 типа или рассеянный склероз.
Настоящее изобретение также охватывает варианты осуществления, описанные ниже.
Пункт 1-А
Антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (g)-(i) ниже, и вариабельную область легкой цепи, которая содержит CDR1-3 легкой цепи описанные от (j) до (1) ниже;
антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи описанные в (m)-(o) ниже, и вариабельную область легкой цепи, которая содержит CDR1-3 легкой цепи, описанные в (p)-(r) ниже; или
антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (a)-(c) ниже, и вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (d)-(f) ниже;
(a) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 41,
(b) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 42,
(c) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 43;
(d) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 44,
(e) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 45, и
(f) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 46;
(g) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 17,
(h) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 18,
(i) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 19;
(j) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 20,
(k) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 21,
(l) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 22;
(m) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 29,
(n) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 30,
(o) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 31;
(p) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 32,
(q) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 33, и
(r) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 34.
Пункт 2-А
Антитело согласно вышеуказанному пункту 1-А, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 60 или 64, и вариабельную область легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 68 или 72.
Пункт 3-А
Антитело согласно вышеуказанному пункту 1-А или 2-А, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 60, и вариабельную область легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 68.
Пункт 4-А
Антитело согласно вышеуказанному пункту 1-А или 2-А, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 64, и вариабельную область легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 72.
Пункт 5-А
Антитело согласно любому из вышеуказанных пунктов от 1-А до 4-А, содержащее константную область человека.
Пункт 6-А
Антитело согласно любому из вышеуказанных пунктов от 1-А до 5-А, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 59, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 67.
Пункт 7-А
Антитело согласно любому из вышеуказанных пунктов от 1-А до 5-А, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 63, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 71.
Пункт 8-А
Антитело согласно любому из вышеуказанных пунктов от 1-А до 7-А, содержащее Fc-область, где Fc-область мутирована для того, чтобы вызвать изменение ADCC-активности.
Пункт 9-А
Антитело согласно вышеуказанному пункту 8-А, где Fc-область мутирована для снижения ADCC-активности.
Пункт 10-А
Антитело согласно любому из вышеуказанных пунктов от 1-А до 9-А где антитело ингибирует взаимодействие между XCR1 человека и XCL1 человека.
Пункт 11-А
Антитело согласно любому из вышеуказанных пунктов от 1-А до 10-А, где антитело ингибирует клеточную миграцию дендритных клеток.
Пункт 12-А
Фармацевтическая композиция, содержащая антитело согласно любому из вышеуказанных пунктов от 1-А до 11-А и фармацевтически приемлемый носитель или добавку.
Пункт 13-А
Фармацевтическая композиция согласно вышеуказанному пункту 12-А, где фармацевтическая композиция является терапевтическим средством для лечения иммунопатологических заболеваний.
Пункт 14-А
Фармацевтическая композиция согласно вышеуказанному пункту 13-А, где иммунопатологическим заболеванием является иммунопатологическое заболевание кожи.
Пункт 15-А
Фармацевтическая композиция согласно вышеуказанному пункту 14-А, где иммунопатологическим заболеванием кожи является псориаз, парапсориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит, миозит с тельцами включения, аутоиммунное заболевание, проявляющееся появлением волдырей (обыкновенная пузырчатка, пемфигоид или приобретенный буллезный эпидермолиз), пустулезное высыпание, герпес беременных, линейный IgA-зависимый буллезный дерматоз, очаговая алопеция, вульгарное витилиго, кожное заболевание, связанное с коллагенозом (системная красная волчанка, синдром Шегрена или смешанная форма заболевания соединительной ткани), кожное заболевание, связанное с болезнью Аддисона, кожное заболевание, связанное с реакцией "трансплантат против хозяина" (GVHD), экзема или крапивница
Пункт 16-А
Фармацевтическая композиция согласно вышеуказанному пункту 14-А, где иммунопатологическим заболеванием кожи является псориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит или миозит с тельцами включения.
Пункт 17-А
Фармацевтическая композиция согласно вышеуказанному пункту 14-А, где иммунопатологическим заболеванием кожи является атопический дерматит или контактный дерматит.
Пункт 18-А
Нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую антитело согласно любому из вышеуказанных пунктов от 1-А до 11-А.
Пункт 19-А
Способ лечения иммунопатологического заболевания, включающий этап введения эффективного количества антитела согласно любому из вышеуказанных пунктов от 1-А до 11-А человеку, пораженному иммунопатологическим заболеванием.
Настоящее изобретение, кроме того, охватывает варианты осуществления, описанные ниже.
Пункт 1-В
Антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (g)-(i) ниже, и вариабельную область легкой цепи, которая содержит CDR1-3 легкой цепи, описанные в (j)-(l) ниже;
антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (m)-(о) ниже, и вариабельную область легкой цепи, которая содержит CDR1-3 легкой цепи, описанные в (p)-(r) ниже; или
антитело, содержащее вариабельную область тяжелой цепи, которая содержит CDR 1-3 тяжелой цепи, описанные в (а)-(с) ниже, и вариабельную область легкой цепи, которая содержит CDR 1-3 легкой цепи, описанные в (d)-(f) ниже;
(a) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 41,
(b) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 42,
(c) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 43;
(d) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 44,
(e) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 45, и
(f) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 46;
(g) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 17,
(h) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 18,
(i) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 19;
(j) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 20,
(k) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 21,
(l) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 22;
(m) CDR 1 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 29,
(n) CDR 2 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 30,
(о) CDR 3 тяжелой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 31;
(p) CDR 1 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 32,
(q) CDR 2 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 33, и
(r) CDR 3 легкой цепи, состоящий из аминокислотной последовательности SEQ ID NO: 34.
Пункт 2-В
Антитело согласно вышеуказанному пункту 1-В, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 60 или 64, и вариабельную область легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 68 или 72.
Пункт 3-В
Антитело согласно вышеуказанному пункту 1-В или 2-В, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 60, и вариабельную область легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 68.
Пункт 4-В
Антитело согласно вышеуказанному пункту 1-В или 2-В, содержащее вариабельную область тяжелой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 64, и вариабельную область легкой цепи, которая содержит аминокислотную последовательность SEQ ID NO: 72.
Пункт 5-В
Антитело согласно любому из вышеуказанных пунктов от 1-В до 4-В, содержащее константную область человека.
Пункт 6-В
Антитело согласно любому из вышеуказанных пунктов от 1-В до 5-В, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 59, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 67.
Пункт 7-В
Антитело согласно любому из вышеуказанных пунктов от 1-В до 5-В, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 63, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 71.
Пункт 8-В
Антитело согласно любому из вышеуказанных пунктов от 1-В до 7-В, содержащее Fc-область, где Fc-область мутирована для того, чтобы вызвать изменение ADCC-активности.
Пункт 9-В
Антитело согласно вышеуказанному пункту 8-В, где Fc-область мутирована для снижения ADCC-активности.
Пункт 10-В
Антитело согласно любому из вышеуказанных пунктов от 1-В до 9-В, где антитело конъюгировано с цитотоксической молекулой.
Пункт 11-В
Антитело согласно любому из вышеуказанных пунктов от 1-В до 10-В, где антитело ингибирует взаимодействие между XCR1 человека и XCL1 человека.
Пункт12-В
Антитело согласно любому из вышеуказанных пунктов от 1-В до 11-В, где антитело ингибирует клеточную миграцию дендритных клеток.
Пункт 13-В
Фармацевтическая композиция, содержащая антитело согласно любому из вышеуказанных пунктов от 1-В до 12-В и фармацевтически приемлемый носитель или добавку.
Пункт 14-В
Фармацевтическая композиция согласно вышеуказанному пункту 13-В, где фармацевтическая композиция является терапевтическим средством для лечения иммунопатологического заболевания.
Пункт 15-В
Фармацевтическая композиция согласно вышеуказанному пункту 14-В, где иммунопатологическим заболеванием является иммунопатологическое заболевание кожи.
Пункт 16-В
Фармацевтическая композиция согласно вышеуказанному пункту 15-В, где иммунопатологическим заболеванием кожи является псориаз, парапсориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит, миозит с тельцами включения, аутоиммунное заболевание, проявляющееся появлением волдырей (обыкновенная пузырчатка, пемфигоид или приобретенный буллезный эпидермолиз), пустулезное высыпание, герпес беременных, линейный lgA-зависимый буллезный дерматоз, очаговая алопеция, вульгарное витилиго, кожное заболевание, связанное с коллагенозом (системная красная волчанка, синдром Шегрена или смешанная форма заболевания соединительной ткани), кожное заболевание, связанное с болезнью Аддисона, кожное заболевание, связанное с реакцией "трансплантат против хозяина" (GVHD), экзема или крапивница
Пункт 17-В
Фармацевтическая композиция согласно вышеуказанному пункту 15-В, где иммунопатологическим заболеванием кожи является псориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит или миозит с тельцами включения.
Пункт 18-В
Фармацевтическая композиция согласно вышеуказанному пункту 15-В, где иммунопатологическим заболеванием кожи является атопический дерматит или контактный дерматит.
Пункт 19-В
Нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую антитело согласно любому из вышеуказанных пунктов от 1-В до 12-В.
Пункт 20-В
Способ лечения иммунопатологического заболевания, включающий введение эффективного количества антитела согласно любому из вышеуказанных пунктов от 1-В до 12-В или фармацевтической композиции согласно вышеуказанному пункту 13-В человеку, пораженному иммунопатологическим заболеванием.
Пункт 21-В
Способ согласно вышеуказанному пункту 20-В, при котором иммунопатологическим заболеванием является иммунопатологическое заболевание кожи.
Пункт 22-В
Способ согласно вышеуказанному пункту 21-В, при котором иммунопатологическим заболеванием кожи является псориаз, парапсориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит, миозит с тельцами включения, аутоиммунное заболевание, проявляющееся появлением волдырей (обыкновенная пузырчатка, пемфигоид или приобретенный буллезный эпидермолиз), пустулезное высыпание, герпес беременных, линейный IgA-зависимый буллезный дерматоз, очаговая алопеция, вульгарное витилиго, кожное заболевание, связанное с коллагенозом (системная красная волчанка, синдром Шегрена или смешанная форма заболевания соединительной ткани), кожное заболевание, связанное с болезнью Аддисона, кожное заболевание, связанное с реакцией "трансплантат против хозяина" (GVHD), экзема или крапивница
Пункт 23-В
Способ согласно вышеуказанному пункту 21-В, при котором иммунопатологическим заболеванием кожи является псориаз, атопический дерматит, контактный дерматит, дерматомиозит, полимиозит или миозит с тельцами включения.
Пункт 24-В
Способ согласно вышеуказанному пункту 21-В, при котором иммунопатологическим заболеванием кожи является атопический дерматит или контактный дерматит.
Примеры
Ниже в данном документе настоящее изобретение более подробно описано на основании Примеров. Несомненно, настоящее изобретение не ограничено Примерами.
Пример 1
(1) Получение мышиных моноклональных антител к XCR1 человека
Для получения моноклональных антител к XCR1 человека нокаут по гену XCR1 мышей иммунизировали мембранной фракцией клеток В300.19, которые экспрессируют XCR1 человека. Мембранную фракцию получали согласно следующей процедуре: сначала клетки В300.19, которые экспрессируют XCR1 человека, суспендировали в Но-буфере (0,25 M сахароза, 10 мМ Hepes (pH 7,4), 1 мМ EGTA, 0,5 мМ MgCl2, 1x Complete mini EDTA-free (Roche Applied Science)), разрушали (800 psi, в течение 30 минут на льду) при помощи азот-содержащего газового дезинтегратора для разрушения клеток (Parr Instrument Company), а затем центрифугировали (2000 g, 10 минут). Надосадочную жидкость собирали и повторно центрифугировали (100000 g, 30 минут). Осадок суспендировали в 50 мМ буфере Hepes (pH 7,4) и определяли как мембранную фракцию.
160 мкг или 260 мкг этой мембранной фракции смешивали с равным объемом адъюванта GERBU (GERBU Biotechnik GmbH) и затем инъецировали подкожно в ступни нокаут по гену XCR1 мышей (Deltagen). Пять или шесть дополнительных инъекций проводили с интервалом раз в две недели. Спустя три или четыре дня после заключительной иммунизации мышей умерщвляли и осуществляли слияние клеток периферических лимфатических узлов с миеломными клетками P3U1 в соотношении 2:1 или 5:1 в присутствии GenomeONE-CF (Ishihara Sangyo Kaisha, Ltd.). Слитые клетки культивировали в 96-луночных пластиковых планшетах.
FACS-анализ осуществляли для первичного скрининга. Исходные клетки СНО и клетки СНО, экспрессирующие XCR1 человека-EGFR смешивали в соотношении 1:1 и суспендировали в FACS буфере (PBS- (Sigma), содержащий 1% FBS и 1 мМ EDTA). Клетки инкубировали в течение 20 минут на льду с надосадочной жидкостью культуры от каждой гибридомы. Клетки промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с РЕ-меченым поликлональным антителом к IgG мыши (Jackson, #715-116-151, разбавленными в соотношении 1:100 в FACS буфере). Клетки промывали FACS буфером три раза и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли с помощью анализатора клеток FACSCanto II (BD Bioscience). В результате, надосадочная жидкость, собранная из трех лунок, продемонстрировала высокую реактивность в отношении клеток СНО, экспрессирующих XCR1 человека-EGFP.
Использовали стандартный способ серийных разведений для получения клонов из этих трех положительных лунок (2Н6, 5G7 и 11Н2). Реактивность каждого клона подтверждали FACS-анализом, описанным выше.
Затем проводили анализ хемотаксиса in vitro для того, чтобы оценить нейтрализующую активность этих трех клонов на индуцируемую лимфотактином человека миграцию клеток BaF3 или В300.19, экспрессирующих XCR1 человека. Анализ хемотаксиса выполняли на 24-луночных подложках для культивирования transwell (размер пор 3 мкм, Costar, #3399) или 96-луночных планшетах для культивирования transwell (MultiScreen, размер пор 5 мкм, Millipore, #MAMIC 5S10).
При использовании 24-луночных подложек для культур клетки BaF3, экспрессирующие XCR1 человека (1×106 клеток), суспендировали в смеси 50 мкл буфера для анализа хемотаксиса (среда RPMI 1640 (Invitrogen), содержащая 0,5% BSA, 0,5% FBS и 20 мМ HEPES (pH 7,4)) и 50 мкл надосадочной жидкости каждой из культур и инкубировали при комнатной температуре в течение 30 минут. Затем рекомбинантный лимфотактин человека (Genzyme, #2695), растворенный в буфере для анализа хемотаксиса в концентрации 1 мкг/мл, добавляли в нижние лунки в концентрации 600 мкг/лунка и инкубированные клетки добавляли в верхние лунки. После 4 часов инкубации в инкубаторе с 5% CO2 при 37°C transwell-лунки центрифугировали при 1350 об/мин в течение 5 минут и мигрировавшие клетки собирали в нижние лунки. Собранные клетки фиксировали при помощи параформальдегида (конечная концентрация 1%) и 30 мкл каждого образца наносили на анализатор клеток FACSCanto II, чтобы подсчитать количество клеток.
При использовании 96-луночных планшетов для культур transwell клетки В300.19, экспрессирующие XCR1 человека (2×105 клеток), суспендировали в смеси 25 мкл буфера для анализа хемотаксиса (RPM 1640 (Invitrogen), содержащий 0,5% BSA, 0,5% FBS и 20 мМ HEPES (pH 7,4) и 50 мкМ 2-меркаптозтанола) и 50 мкл надосадочной жидкости каждой из культур и инкубировали при комнатной температуре в течение 30 минут. Затем рекомбинантный лимфотактин человека (Genzyme, #2695), растворенный в буфере для анализа хемотаксиса в концентрации 1 мкг/мл, добавляли в нижние лунки в концентрации 150 мкл/лунка и инкубированные клетки добавляли в верхние лунки. После 4 часов инкубации в инкубаторе с 5% CO2 при 37°C transwell-лунки центрифугировали при 1350 об./мин. в течение 5 минут и мигрировавшие клетки собирали в нижние лунки. 30 мкл каждого образца наносили на анализатор клеток FACSCanto II, чтобы подсчитать количество клеток.
Надосадочная жидкость из клеточных культур, произведенную тремя клонами гибридом (2Н6, 5G7 и 11Н2), продемонстрировала нейтрализующую активность в отношении индуцируемой лимфотактином человека миграции клеток BaF3 и клеток В300.19, экспрессирующих XCR1 человека.
(2) Реактивность мышиных антител к XCR1 человека (2Н6,5G7 и 11Н2) в отношении клеток, экспрессирующих XCR1 человека
Для того, чтобы оценить реактивность и нейтрализующую активность очищенных антител этих трех клонов, антитела очищали из надосадочной жидкости культуры каждого клона при помощи рекомбинантного бежа A (GE Healthcare, #17-5080-01). Изотип каждого клона определяли с помощью набора для изотипирования моноклональных антител (Serotec, #ММТ1). 2Н6 и 5G7 оказались IgG2b, κ и 11Н2 оказался IgG2a, κ.
Реактивность очищенных антител к XCR1 человека оценивали с помощью FACS-анализа Исходные клетки В300.19 и клетки В300.19, экспрессирующие XCR1 человека-EGFP, смешивали в соотношении 1: 1 и суспендировали в FACS буфере (PBS– (Sigma), содержащий 1% FBS). Клетки блокировали в течение 10 минут на льду при помощи FACS буфера, содержащего иммуноглобулина человека в концентрации 100 мкг/мл. После этого клетки инкубировали в течение 20 минут на льду с очищенными антителами (2Н6, 5G7 и 11Н2) при различных концентрациях от 0 до 10 мкг/мл или с изотопическим контролем, мышиным антителом IgG2a (eBioscience, #14-4724-82) или IgG2b (eBioscience, #14-4732-82), при концентрации 10 мкг/мл. Клетки промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с РЕ-меченым поликлональным антителом к IgG мыши (Jackson, #715-116-151, разбавленный при соотношении 1:50 в FACS буфере). Клетки промывали FACS буфером три раза и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли с помощью анализатора клеток FACSCanto II.
Эти три очищенных антитела (2Н6, 5G7 и 11Н2) проявляли реактивность в отношении клеток В300.19, экспрессирующих XCR1 человека-EGFP, но не в отношении исходных клеток В300.19 (Фиг. 1). Напротив, мышиное антитело изотипического контроля не взаимодействовало ни с клетками В300.19, экспрессирующими XCR1 человека-EGFP, ни с исходными клетками (данные не показаны).
(3) Нейтрализующая активность мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) в отношении индуцируемой лимфотактином человека миграции клеток человека, экспрессирующих XCR1 человека
Нейтрализующую активность очищенных антител этих клонов оценивали при анализе хемотаксиса in vitro. Анализ хемотаксиса выполняли с помощью 96-луночных планшетов для культур transwell (MultiScreen, размер пор 5 мкм, Millipore, #MAMIC 5S10). Клетки В300.19, экспрессирующие XCR1 человека (2×105 клеток), суспендировали в 75 мкл буфера для хемотаксиса (среда RPMI1640 (Invitrogen), содержащая 0,5% BSA, 0,5% FBS, 20 мМ HEPES (pH 7,4) и 50 мкМ 2-меркаптоэтанола), содержащего каждые из очищенных антител (2Н6, 5G7 и 11Н2) в различных концентрациях от 0 до 10 мкг/мл, и инкубировали при комнатной температуре в течение 30 минут. Далее рекомбинантный лимфотактин человека (R&D, #695-LT/CF) растворяли в буфере для хемотаксиса в конечной концентрации 1 мкг/мл, в котором были растворены очищенные антитела в различных концентрациях от 0 до 10 мкг/мл. Смесь лимфотактина и очищенных антител добавляли в нижние лунки в концентрации 150 мкл/лунка и инкубировали при комнатной температуре в течение 30 минут.30 минут спустя инкубированные клетки добавляли в верхние лунки и инкубировали в инкубаторе с 5% CO2 при 37°C в течение 4 часов. Затем 30 мкл каждого образца наносили на анализатор клеток FACSCanto II, чтобы подсчитать количество клеток. MAb 2Н6, 5G7 и 11Н2 полностью ингибировали клеточную миграцию при концентрации приблизительно 3 мкг/мл. На Фиг. 2 показан типичный пример зависимого от концентрации ингибирования. Значения IC50 и IC90 вычисляли при использовании трех независимых экспериментов. В Таблице 1 показаны эти значения как среднее значение ± стандартная ошибка.

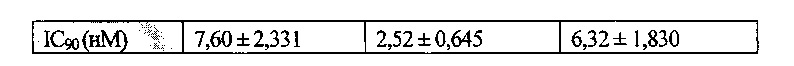
(4) Анализ последовательности мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2)
Полинуклеотиды, содержащие последовательность гена, который кодирует тяжелые и легкие цепи клонов (2Н6, 5G7 и 11Н2), амплифицировали при помощи способа 5'- RACE (5- ПЦР с быстрой амплификацией концов кДНК). Общую РНК получали из гибридомы этих трех клеточных клонов при помощи TRIZOL (Invitrogen) и обрабатывали ДНазой (QIAGEN, RNase tree DNase set). Двухцепочечную кДНК получали из общей РНК с помощью набора для синтеза кДНК (TAKARA). 5'-адаптер, полученный при отжиге ad29S; ACATCACTCCGT (SEQ ID NO: 81) и as29AS;
ACGGAGTGATGTCCGTCGACGTATCTCTGCGTTGATACTTCAGCGTAGCT (SEQ ID NO: 82) добавляли к кДНК. Полученную кДНК амплифицировали с помощью 5'-прямого праймера
(5'-PCR4 праймер, AGCTACGCTGAAGTATCAACGCAGAG: SEQ ID NO: 83) и
3'-обратного праймера
(AGGACAGGGGTTGATTGTTGA: SEQ ID NO: 84 или
CTCAAGTTTTTTGTCCACCGTGGTGC: SEQ ID NO: 85
использовали для амплификации тяжелой цепи IgG2b;
CTCAATTTTCTTGTCCACCTTGGTGC: SEQ ID NO: 86 или
GCCAGTGGATAGACTGATG: SEQ ID NO: 87
использовали для амплификации тяжелой цепи IgG2a и
CTCATTCCTGTTGAAGCTCTTGACAAT: SEQ ID NO: 88,
GATGGATACAGTTGGTGCAGC: SEQ ID NO: 89 или
CAGATCCTCAGCCTCCACTCTGCT: SEQ ID NO: 90
использовали для амплификации легкой цепи Igκ). Амплифицированную кДНК вставляли в вектор pCR2.1 (Invitrogen). Последовательности генов анализировали с помощью ABD130XL. В Таблицах от 2-1 до 4-2 показаны последовательности аминокислотных последовательностей, кодируемых последовательностями генов, которые были идентифицированы с помощью этого анализа




Пример 2
(1) Получение химерного антитела к XCR1 человека и гуманизированных антител к XCR1 человека
Клон 5G7, который продемонстрировал самую высокую нейтрализующую активность среди 2Н6, 5G7 и 11Н2, использовали для получения химерного антитела и гуманизированных антител.
Химерное антитело получали посредством комбинирования при помощи ПЦР с перекрывающимися фрагментами последовательности гена вариабельной области тяжелой цепи 5G7h последовательности гена константной области IgG2 человека, в последовательность, соответствующую тяжелой цепи, которой вводили мутацию V234A/G237A, и последовательности вариабельной области легкой цепи 5G7 и последовательности гена константной области Igκ человека и посредством вставки получившейся последовательности в векторы экспрессии (рЕЕ6.4 или рЕЕ12.4). В Таблице 5 и 6, соответственно, показаны аминокислотные последовательности и нуклеотидные последовательности конкретного химерного антитела.



Антитело гуманизировали посредством пересадки гипервариабельного участка мышиного антитела 5G7 в вариабельную область антитела человека. Гипервариабельный участок определяли согласно системе нумерации по Kabat и способу идентификации гипервариабельного участка (например, Kabat et al., (1991) Sequences of Proteins of Immunological Interest: US Department of Health and Human Services, NIH, USA). Кроме того, гипервариабельные участки 2Н6 и 11Н2 также определяли подобным образом. В Таблицах от 7-1 до 9-2 показаны аминокислотные последовательности и нуклеотидные последовательности гипервариабельных участков этих трех клонов.




Как становится понятно из Таблиц 7-1 и 8-1, идентичность аминокислотных последовательностей CDR между 5G7 и 2Н6 высока; в частности, аминокислотные последовательности CDR 3 тяжелой цепи были абсолютно идентичными. Таким образом, в отношении 5G7 и 2Н6 аминокислотные последовательности можно обобщить, как показано ниже в Таблице 10. Дополнительно, на Фиг. 7 приведено сравнение аминокислотных последовательностей CDR 1-3 этих клонов.
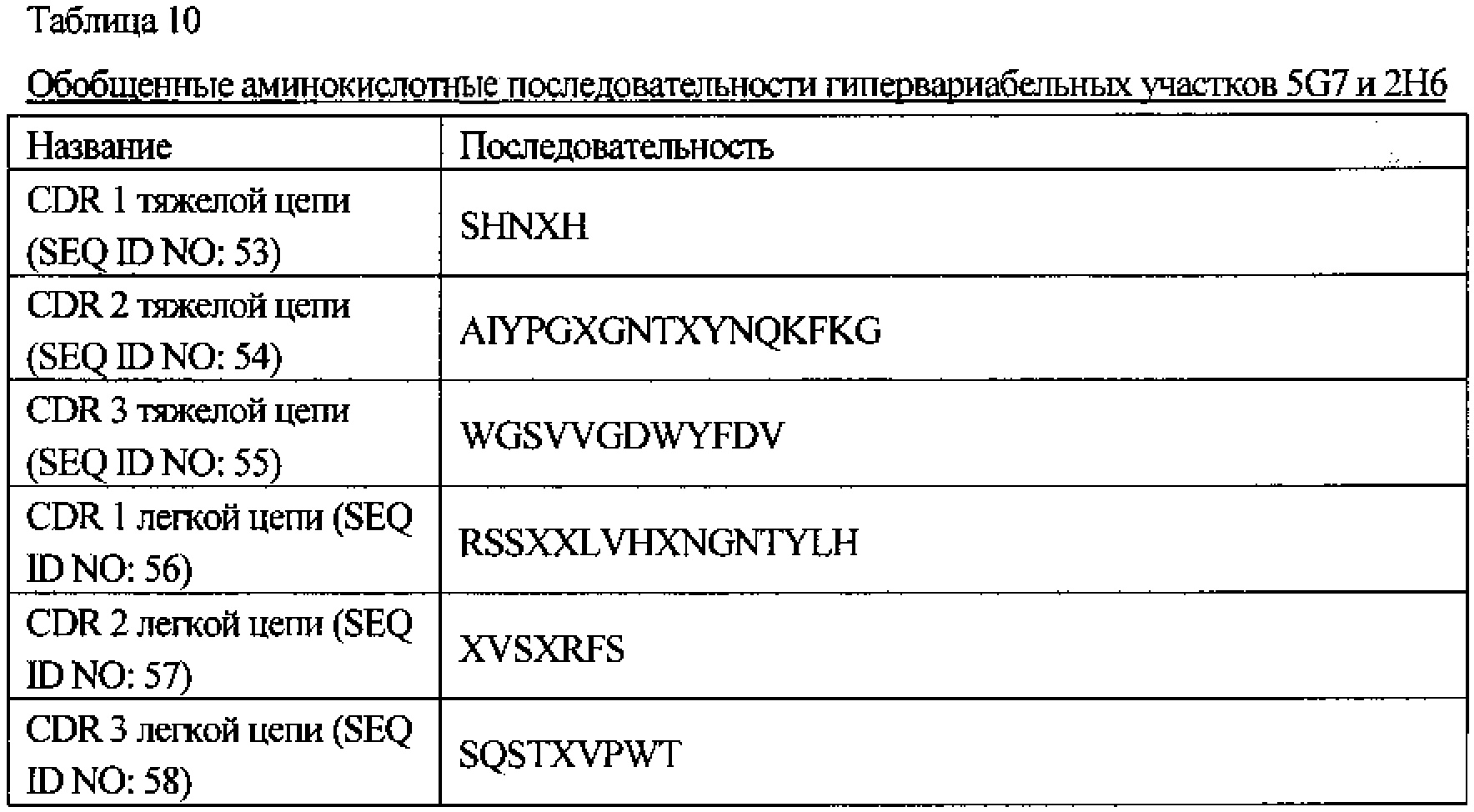
"X" в таблице может представлять собой любое из следующего: аланин (Ala: А), аргинин (Arg: R), аспарагин (Asn: Ν), аспарагиновая кислота (Asp: D), цистеин (Cys: С), глютамин (Gln: Q), глугаминовая кислота (Glu: Ε), глицин (Gly: G), гистидин (His: H), изолейцин (Ile: I), лейцин (Leu: L), лизин (Lys: K), метионин (Met: M), фенилаланин (Phe: F), пролин (Pro: Ρ), серии (Ser: S), треонин (Thr: T), триптофан (Trp: W), тирозин (Tyr: Y) и валин (Val: V).
FR антитела человека с высокой идентичностью по отношению к FR 5G7 выбирали как FR гуманизированных антител. Затем аминокислоты в FR которые взаимодействуют с CDR 5G7, предсказывали при помощи 3D модели полученного в результате антитела и пересаживали с CDR. Константную область IgG2 человека, в которую вводили мутацию V234A/G237A, использовали в качестве константной области. HK1 и HK5 разрабатывали как тяжелые цепи гуманизированного антитела, a L2 и L5 разрабатывали как легкие цепи гуманизированного антитела. В Таблицах от 11-1 до 14-2 показаны аминокислотные последовательности и нуклеотидные последовательности конкретных гуманизированных антител.






Последовательности генов этих гуманизированных антител были полностью синтезированы GenScript USA Inc. и их вставляли в векторы экспрессии (рЕЕ6.4 или рЕЕ12.4, приобретенные у Lonza). Для получения антител векторы экспрессии трансфецировали в клетки HEK293E (Invitrogen) при помощи Lipofectamine 2000 согласно инструкциям для Lipofectamine 2000 (Invitrogen). Надосадочную жидкость собирали и очищали при помощи Белка A (GE Healthcare). Нейтрализующую активность оценивали с помощью этих очищенных гуманизированных антител.
Гуманизированные антитела, обладающие нейтрализующей активностью в отношении индуцируемой лимфотактином человека миграции клеток, экспрессирующих XCR1 человека, идентифицировали путем осуществления анализов хемотаксиса in vitro с помощью клеток В300.19, экспрессирующих XCR1 человека. Анализ хемотаксиса выполняли как описано выше при помощи 96-луночных планшетов для культур transwell (MultiScreen, размер пор 5 мкм, Millipore, #MAMIC 5S10 или Corning #3387 или #3388). Однако в случае использования планшетов для культур Corning transwell количество рекомбинантного лимфотактина человека и очищенных антител, которые добавляют к нижним лункам,составляет 235 мкл на лунку.
Из гуманизированных антител, обладающих нейтрализующей активностью, два типа нижеприведенных антител, HK1L2 и HK5L5, оценивали более детально.
(2) Реактивность гуманизированных антител к XCR1 человека (HK1L2 и HK5L5) в отношении клеток, экспрессирующих XCR1 человека
FACS-анализ выполняли с использованием этих двух гуманизированных антител (HK1L2 и HK5L5), исходного антитела 5G7 и химерного антитела. Исходные клетки В300.19 и клетки В300.19, экспрессирующие XCR1 человека-EGFP, смешивали в соотношении 1:1 и суспендировали в FACS буфере (PBS- (Sigma), содержащий 1% FBS). Клетки инкубировали в течение 20 минут на льду с очищенными антителами при разных концентрациях от 0 до 10 мкг/мл. Клетки промывали три раза FACS буфером и затем инкубировали в течение 20 минут на льду с РЕ-меченым поликлональным антителом к IgG мыши (Jackson, #715-116-151; используемые для клеток, которые окрашивали исходным антителом 5G7, разведенным в соотношении 1:100 в FACS буфере) или с РЕ-меченым поликлональным антителом к IgG человека (Jackson, #709-116-149; используемые для клеток, которые окрашивали химерным антителом или гуманизироваными антителами (HK1L2 и HK5L5), разведенным в соотношении 1:100 в FACS буфере. Клетки промывали три раза FACS буфером и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II (BD Bioscience).
Гуманизированные антитела (HK1L2 и HK5L5) проявляли зависимую от концентрации реактивность в отношении клеток В300.19, экспрессирующих XCR1 человека-EGFP. Исходное антитело 5G7 и химерное антитело продемонстрировали в основном одинаковую реактивность (Фиг. 3).
Реактивность гуманизированных антител (HK1L2 и HK5L5) в отношении XCR1 человека затем исследовали при помощи FACS-анализа с использованием мононуклеарных клеток периферической крови человека. Поскольку ген XCR1 человека, как известно, экспрессируется в BDCA3+ дендритных клетках, которые являются небольшой популяцией мононуклеарных клеток периферической крови человека, то сначала дендритные клетки получали из мононуклеарных клеток периферической крови человека и использовали для FACS-анализа. Мононуклеарные клетки периферической крови человека выделяли из крови здоровых субъектов-людей, с помощью Ficoll-Paque (GE Healthcare, #17-1440-02). CD3-, CD14-, CD19- и CD56-положительные клетки из числа мононуклеарных клеток периферической крови человека метили при помощи микрогранул с антителами к CD3, CD14, CD19 и CD56 (Miltenyi, #130-050-101, #130-050-201, #130-050-301 и #130-050-401) и удаляли при помощи auto-MACS (Miltenyi). Таким образом, собирали дендритные клетки. Собранные дендритные клетки блокировали в течение 10 минут на льду с FACS буфером (PBS- (Sigma), содержащий 1% FBS), включающим 1% сыворотку крови крысы, 1% сыворотку крови мыши, иммуноглобулин человека в концентрации 100 мкг/мл. Затем клетки красили в течение 30 минут на льду отдельно друг от друга при использовании РЕ-меченых 5G7, HK1L2 и HK5L5 и мышиного антитела IgG2b, к изотопического контроля (eBioscience, #14-4732-82) или антитела IgG2, к человека (Sigma, #15404) с FITC-меченого антитела к BDCA3 (Miltenyi, #130-090-513), APC-меченого антитела к CD 123 (Miltenyi, #130-090-901), АРС-Cy7-меченого антитела к HLA-DR (BioLegend, #307617) и А1еха700-меченых антител к CD3, CD14, CD19 и CD56 (BioLegend, #300324, #301822, #302225, и #318316). Клетки промывали три раза FACS буфером и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II.
Как и в случае с исходным антителом 5G7, гуманизированные антитела (HK1L2 и HK5L5) селективно взаимодействовали с BDCA3+ дендритными клетками, экспрессирующими XCR1 человека (Фиг. 4).
(3) Нейтрализующая активность гуманизированных антител к XCR1 человека (HK1L2 и HK5L5) в отношении индуцируемой лимфотактином человека миграции клеток, экспрессирующих XCR1 человека
Нейтрализующую активность этих гуманизированных антител оценивали параллельно с исходным антителом 5G7 и химерным антителом посредством анализа хемотаксиса in vitro, как описано выше.
По сравнению с исходным антителом 5G7 оба гуманизированных антитела, сохраняли нейтрализующую активность. На Фиг. 5 показан типичный пример зависимого от концентоации ингибирования. Значения IC50 и IC90 рассчитывали при проведении трех независимых экспериментов. На Таблице 16 показаны эти значения как среднее значение ± стандартная ошибка.
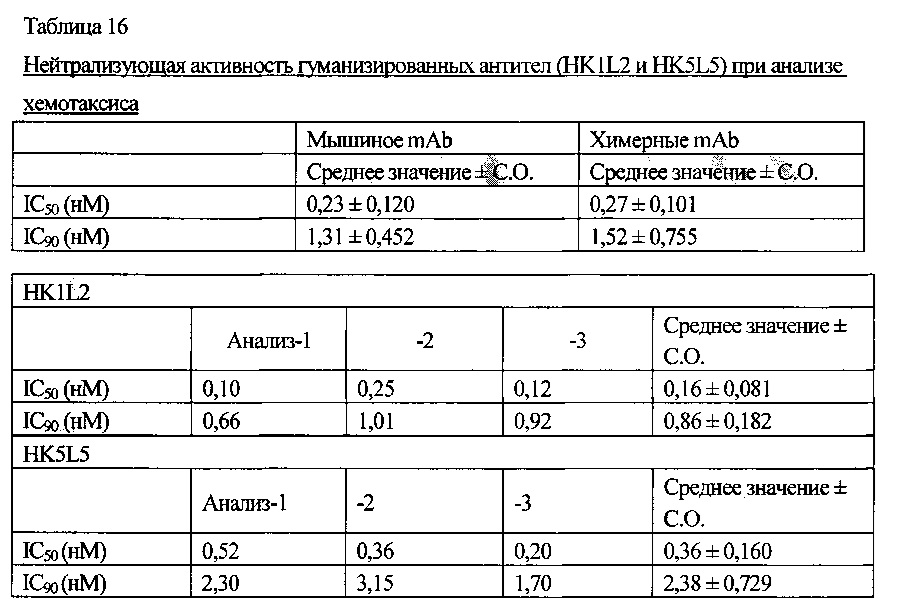
Затем нейтрализующую активность гуманизированных антител (HK1L2 и HK5L5) дополнительно исследовали при помощи анализа трансэндотелиальной миграции, в котором использовали дендритные клетки человека вместо клеток В300.19, экспрессирующих XCR1 человека. Анализ трансэндотелиальной миграции выполняли при использовании 24-луночных подложек для культур transwell (размер пор 5 мкм, Costar, #3421). Сначала клетки ECV304 суспендировали в среде 199 Эрла (Invitrogen), содержащей 10% FBS, и засевали в верхнюю камеру transwell при концентрации 2×105 клеток на лунку с последующей инкубацией в инкубаторе с 5% CO2 при 37°C в течение 3 дней. В день анализа клетки ECV304 промывали буфером для анализа (смесь среды 199 Эрла и среды RPMI 1640 в соотношении 1:1, в которую добавляли 0,5% BSA и 20 мМ HEPES (pH 7,4)). Рекомбинантный лимфотактин человека, растворенный в буфере для анализа при концентрации 1 мкг/мл, к которому добавляли химерное антитело, HK1L2, HK5L5, или антитело изотипического контроля, IgG2, к человека (Sigma), в концентрации 10 мкг/мл, добавляли в нижние лунки в концентрации 600 мкл/лунка. Дендритные клетки человека собирали, как описано выше, суспендировали в буфере для анализа, к которому добавляли химерное антитело, гуманизированные антитела (HK1L2 и HK5L5) и антитело изотипического контроля, IgG2, к человека (Sigma), в концентрации 10 мкг/мл, и добавляли в верхние лунки, содержащие клетки ECV304. После инкубации в течение 4 часов в инкубаторе с 5% CO2 при 37°C клетки в transwell центрифугировали при 1350 об/мин в течение 5 минут и мигрировавшие клетки собирали. Собранные клетки красили в течение 30 минут на льду с помощью маркеров клеточных линий, FITC-меченого антитела к BDCA3 (Miltenyi, #130-090-513), РЕ-меченого антитела к BDCA1 (BioLegend, #331517), АРС-меченого антитела к CD 123 (Miltenyi, #130-090-901) и APC-меченого антитела к HLA-DR (BioLegend, #307617). Затем 170 мкл каждого образца наносили на анализатор клеток FACSCanto II (BD Bioscience) для подсчета количества клеток.
Оба гуманизированных антитела ингибировали миграцию BDCA3+ дендритных клеток так же, как и химерное антитело (Фиг. 6).
Пример 3
Фармакологический эффект мышиного антитела к XCR1 человека
Фармакологический эффект мышиного моноклонального антитела к XCR1 человека (5G7), полученного выше в Примере 2, подтверждали при помощи мышиной модели реакции замедленного типа - контактного дерматита (DTH).
(1) Эффект мышиного антитела к XCR1 человека на опухание уха у мышей, сенсибилизированных DNFB (динитрофторбензолом)
Способ проведения эксперимента
1. Мыши, используемые для применения образца
Для эксперимента использовали происходящих от C57BL/6 нокин по гену XCR1 человека мышей (мышей, у которых ген XCR1 был замещен геном XCR1 человека) в возрасте от 7 до 12 недель.
2. Способ получения DNFB для сенсибилизации и DNFB для индукции
DNFB для сенсибилизации и индукции получали, смешивая DNFB в соотношении 4:1 со смесью ацетона и оливкового масла с получением концентрации 0,5%. Затем смесь ацетона и оливкового масла в соотношении 4:1 использовали в качестве контрольного раствора для индукции.
3. Способ применения DNFB
Шерсть на животе мышей сбривали для того, чтобы обнажить кожу, и наносили на нее 50 мкл 0,5% DNFB для сенсибилизации. На следующий день 50 мкл 0,5% DNFB наносили снова на то же самое место. Спустя 4 дня после нанесения 25 мкл 0,5% DNFB для индукции наносили на наружную сторону правого уха мышей. В то же время в качестве контроля 25 мкл контрольного раствора, полученного при смешивании ацетона и оливкового масла в соотношении 4:1, наносили на наружную сторону левого уха мышей.
4. Способ применения антител
Мышиное моноклональное антитело к XCR1 человека (5G7) и его контрольное антитело, т.е. мышиный IgG (Jackson Laboratory), получали в PBS в конечной концентрации 2 мг/мл. День, когда проводили первую сенсибилизацию, определяли как День 0. Каждое из вышеуказанных антител внутрибрюшинно вводили мышам в количестве 250 мкл/мышь (500 мкг/мышь) в День - 1, День 1 и День 4.
5. Способ измерения опухания уха в сенсибилизированной DNFB мышиной модели
В первый день и на следующий день мышей сенсибилизировали посредством нанесения 50 мкл 0,5% DNFB на обнаженную кожу живота. Спустя 4 дня после сенсибилизации измеряли толщину уха с помощью циркуля. После измерения 25 мкл 0,5% DNFB наносили на наружную сторону правого уха мышей для индукции. Затем в качестве контроля 25 мкл контрольного раствора, полученного путем смешивания ацетона и оливкового масла в соотношении 4:1, наносили на левое ухо мышей. Толщину уха измеряли через 24 часа и через 48 часов после индукции. Опухание уха определяли путем преобразования измеренных значений при помощи следующей формулы.
Формула
Толщина уха, измененная при воздействии DNFB (опухание: мм) = ([А]-[В])-([С]-[D])
[А]: толщина правого уха после индукции (мм)
[B]: толщина правого уха перед индукцией (мм)
[C]: толщина левого уха после нанесения контрольного раствора (мм)
[D]: толщина левого уха перед нанесением контрольного раствора (мм)
Результаты эксперимента и анализ
На Фиг. 8 явно показано значительное подавление опухания уха через 24 часа после индукции DNFB у мышей, которым вводили мышиное моноклональное антитело к XCR1 человека (5G7), по сравнению с мышами, которым вводили контрольное антитело (Фиг. 8А). Эффект также вызвал значительное подавление похожим образом через 48 часов после индукции DNFB (Фиг. 8В).
Хотя антитело регулярно вводили внутрибрюшинно, подавлялось опухание уха, индуцированное DNFB. Поэтому предположили, что антитело транспортировалось из брюшной полости в кровь и вместе с кровотоком достигало места воспаления или лимфатического узла где антитело проявляло эффект подавления опухания уха.
Из этого делали заключение, что антитела по настоящему изобретению обладают особым эффектом в отношении места воспаления локально направленным образом.
Пример 4
Реактивность мышиного моноклонального антитела к XCR1 человека (5G7) по отношению к различным рецепторам хемокинов человека
Реактивность мышиного моноклонального антитела к XCR1 человека (5G7) по отношению к различным рецепторам хемокинов человека оценивали посредством FACS-анализа Исходные клетки В300.19 и клетки В300.19, экспрессирующие рецептор хемокинов-EGFP (XCR1, CXCR1, CXCR3, CXCR4, CXCR5, CXCR6, CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR11 или CX3CR1 и) суспендировали в FACS буфере (PBS- (Sigma), содержащий 1% эмбриональной бычьей сыворотки). Клетки блокировали в течение 20 минут на льду с буфером для блокировки (FACS буфер, содержащий иммуноглобулин человека в концентрации 100 мкг/мл). Затем клетки инкубировали в течение 30 минут на льду с буфером для блокировки, содержащим 5G7 или антитело IgG2b мыши изотипического контроля (eBioscience, #14-4732-82) в концентрации 10 мкг/мл. Клетки промывали три раза FACS буфером и затем инкубировали в течение 20 минут на льду с РЕ-меченым поликлональным антителом к мышиному IgG (Jackson, #715-116-151, разведенным в соотношении 1:50 в буфере для блокировки). Клетки промывали FACS буфером три раза и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли с помощью анализатора клеток FACSCanto II.
Антитело к XCR1 человека 5G7 проявляло высокую реактивность в отношении клеток В300.19, экспрессирующих XCR1 человека-EGFP. Кроме того, антитело к XCR1 человека 5G7 проявляло очень низкую реактивность в отношении клеток В300.19, экспрессирующих CX3CR1-EGFP, и отсутствие реактивности в отношении других клеток, экспрессирующих человеческие рецептор хемокинов-EGFP (Фиг. 9). С другой стороны, мышиное антитело изотипического контроля не проявляло реактивность в отношении каких-либо клеток В300.19.
Пример 5
Цитотоксичность антител к XCR1 человека в отношении клеток, экспрессирующих XCR1 человека, определенная с помощью сапорин-конюгированного Fab вторичного антитела к IgG мыши
Для того чтобы продемонстрировать цитотоксическую активность антител к XCR1 человека в отношении клеток, экспрессирующих XCR1 человека, цитотоксичность мышиных mAb к XCR1 человека в отношении клеток, на которых экзогенным образом экспрессируется XCR1 человека, исследовали с помощью сапорин-конюгированых Fab вторичного антитела к IgG мыши.
2×103 исходных клеток В300.19 или клеток В300.19, экспрессирующих XCR1 человека-EGFP, в 80 мкл RPMH640 (Invitrogen, #11875-093), содержащей 10% эмбриональную бычью сыворотку (Cell Culture Bioscience, #171012), сульфат канамицина в концентрации 100 мкг/мл (Invitrogen, #15160-054) и 50 мкМ 2-меркаптоэтанола (2-МЕ, Invitrogen, #21985-023) добавляли в каждую лунку 96-луночного планшета. Мышиные антитела к XCR1 человека (2Н6, 5G7 или 11Н2), мышиные антитела изотипического контроля, IgG2a, к (eBioscience, #16-4724-85) или IgG2b, к (eBioscience, #164732-85), разводили RPMU640, содержащей 10% эмбриональную бычью сыворотку, сульфатом канамицина в концентрации 100 мкг/мл и 50 мкМ 2-МЕ и 10 мкл разведенных антител добавляли к клеткам в различных концентрациях от 0 до 0,17 мкг/мл. Затем клетки инкубировали в инкубаторе с 5% CO2 при 37°C в течение 20 мин. Далее сапорин-коньюгированные Fab антител к IgG мыши (Advanced Targeting Systems, #IT-48) разводили в концентрации 10 мкг/мл RPMI1640, содержащей 10% эмбриональную бычью сыворотку, сульфатом канамицина в концентрации 100 мкг/мл и 50 мкМ 2-МЕ и 10 мкл разведенных сапорин-конъюгированых Fab антител к IgG мыши добавляли в каждую лунку в конечной концентрации 1 мкг/мл. Затем клетки инкубировали в инкубаторе с 5% CO2 при 37°C в течение 72 часов. Затем количество клеток в каждой лунке измеряли при помощи Cell Count Reagent SF (Nacalai tesque, 07553-15 или -44). В каждую лунку добавляли реактив и клетки инкубировали в инкубаторе с 5% CO2 при 37°C в течение 2 или 3 часов. Затем OD450 измеряли при помощи спектрофотометра для прочтения планшетов (Arvo, PerkinElmer).
Мышиные антитела к XCR1 человека (2Н6, 5G7 или 11Н2) с сапорин-коньюгированым вторичным антителом проявляли подавление роста клеток В300.19, экспрессирующих XCR1 человека-EGFP (Фиг. 11). Значения IC50 для 2Н6, 5G7 и 11Н2, вычисленные при помощи программного обеспечения Graphpad Prism, составляли 0,141 нМ, 0,017 нМ и 0,155 нМ, соответственно. С другой стороны, эти антитела с сапорин-коньюгированым вторичным антителом не проявляли подавление роста исходных клеток В300.19. Контрольные антитела с сапорин-коньюгированым вторичным антителом не подавляли рост ни клеток В300.19, экспрессирующих XCR1 человека-EGFP, ни исходных клеток В300.19 (Фиг. 11). Эти результаты указывают, что мышиные антитела к XCR1 человека 2Н6, 5G7 и 11Н2 интернализировались с сапорин-коньюгированым вторичным антителом и действовали как антитоксин.
Пример 6
Эффекты mAb 5G7 в отношении цитотоксичности Т-лимфоцитов in vivo
Для того, чтобы исследовать ингибиторную активность mAb 5G7 в отношении функции CTL, выполняли анализ CTL.
В день 0 сконструированных нокин по гену hXCR1 мышей, у которых XCR1 человека экспрессируется вместо XCR1 мыши, иммунизировали подкожно овальбумином (200 мкг/мышь), эмульгированным с CFA. MAb 5G7 или контрольный IgG мыши (Jackson Laboratory) инъецировали внутрибрюшино в дозе 500 мкг/мышь в день - 1, день 2 и день 5. Шесть дней спустя спленоциты от интактных мышей C57BL/6 инкубировали в течение 30 мин. при 37°C с пептидом OVA257-264 в концентрации 10 мкг/мл (SIJNFEKL; MBL) или без такового.
Эти инкубированные с пептидом целевые и нецелевые популяции клеток метили при помощи 2,5 и 0,25 мкМ CFSE (Invitrogen Life Technologies), соответственно, затем смешивали в соотношении 1:1 и инъецировали внутривенно иммунизированным мышам.
Спустя один день после инъекции CFSE-меченых спленоцитов целевую цитотоксическую активность оценивали при помощи соотношения CFSE-положительных популяций в селезенке, как изложено ниже.
CFSE-положительные клетки в селезенке у иммунизированных мышей обнаруживали посредством проточной цитометрии и активность CTL у каждой мыши вычисляли по соотношению клеток CFSEhigh и клеток CFSElow следующим образом: активность CTL=(% CFSEhigh/% CFSElow).
Затем относительную активность CTL вычисляли следующим образом: относительная активность CTL = (активность CTL у каждой иммунизированной мыши) / (активность CTL у мыши контроля).
Результаты продемонстрировали, что относительная активность CTL у мышей, обработанных mAb 5G7, проявлялась как более низкая относительная активность CTL по сравнению с таковой у мышей, обработанных контрольными IgG (Фиг. 12).
Эти данные указывали на подавление активности CTL in vivo при обработке антителом к XCR1 и предлагали, что обработка антителами к XCR1 может быть полезной при иммунопатологических заболеваниях, таких как отторжение трансплантата, GVHD и повреждение тканей при аутоиммунных заболеваниях.
Пример 7
Реактивность мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) в отношении клеток, экспрессирующих химерный XCR1 человека/мыши
Чтобы определить эпитопы XCR1 человека, распознаваемые мышиными антителами к XCR1 человека (2Н6, 5G7 и 11Н2), оценивали реактивность этих антител в отношении клеток, экспрессирующих химерный XCR1 человека/мыши.
Поскольку мышиные антитела к XCR1 человека (2Н6, 5G7 и 11Н2) взаимодействовали с XCR1 человека, но не с XCR1 мыши, то подготавливали панель химерных рецепторов XCR1 человека/мыши. В этой панели каждый внеклеточный домен XCR1 человека замещали гомологичной областью XCR1 мыши и наоборот. Векторы экспрессии этой панели создавали с помощью способа полимеразной цепной реакции (ПЦР) с перекрывающимися фрагментами. Каждый химерный рецептор-EGFP экспрессировался в клетках TK-1, и реактивность mAb определяли с помощью FACS-анализа. Исходные клетки TK-1, клетки TK-1, экспрессирующие XCR1 человека-EGFP, XCR1 мыши-EGFP или химерный XCR1-EGFP, суспендировали в FACS буфере (PBS- (Sigma), содержащем 1% эмбриональную бычью сыворотку). Клетки блокировали в течение 10 минут на льду с FACS буфером, содержащим иммуноглобулина человека в концентрации 100 мкг/мл. Затем клетки инкубировали в течение 20 минут на льду с антителами к XCR1 человека (2Н6, 5G7 или 11Н2) при различных концентрациях от 0 до 10 мкг/мл с антителами изотипического контроля, IgG2a или IgG2b мыши (eBioscience, #14-4724-82) или (eBioscience, #144732-82) в концентрации 10 мкг/мл или FACS буфером без антител. Клетки промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с РЕ-мечеными поликлональным антителом к IgG мыши (Jackson, #715-116-151, разведенным в соотношении 1:50 в FACS буфере) или РЕ-меченым поликлональным антителом к XCR1 человека (R&D, #ЕАВ857Р, разведенным в соотношении 2:5 в FACS буфере, используемое для клеток, которые инкубировали с FACS буфером без антител). Клетки промывали FACS буфером три раза и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II.
Антитела мыши к XCR1 человека (2Н6, 5G7 и 11Н2) проявляли реактивность в отношении клеток TK-1, экспрессирующих XCR1 человека-EGFP, но не в отношении исходных клеток TK-1 или клеток TK-1, экспрессирующих XCR1 мыши-EGFP (Фиг. 13; источник четырех внеклеточных доменов был обозначен при помощи четырехбуквенного кода (например, НННН является XCR1 человека дикого типа, в Hmmm имеется N-терминальный внеклеточный домен человека и мышиная первая, вторая и третья внеклеточные петли и т.д.)). Эти три антитела проявляли реактивность в отношении химерных XCR1 с N-концом XCR1 человека, клеток TK-1, экспрессирующих человека-EGFP. В другом эксперименте реактивность в отношении химерного рецептора, mmHm, также исследовали, но реактивности не наблюдали (данные не показаны).
Напротив, мышиные антитела изотипического контроля не проявляли реактивность в отношении любых клеток TK-1 (данные не показаны).
Пример 8
Картирование участков связывания мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) на внеклеточных доменах XCR1 человека с помощью пептидного сканирования ELISA
Чтобы определить аминокислотные остатки антител к XCR1 человека (2Н6, 5G7 и 11Н2), контактирующие с внеклеточными доменами XCR1 человека, проводили анализ в виде пептидного сканирования при помощи наборов 12-мерных пептидов, которые охватывают внеклеточные домены XCR1 человека.
Два набора пептидов, содержащих на N-конце биотин и спейсер GSGS, были синтезированы компанией Sigma. Первый набор из 13 пептидов содержал все возможные 12-меры из N-конечного участка XCR1 человека, каждый с отступом в две аминокислоты. Второй набор из 13 пептидов содержал все возможные 12-меры из внеклеточных петель XCR1 человека, каждый с отступом в три аминокислоты. Пептиды сначала растворяли в 100% диметилсульфоксиде и затем разбавляли 30% раствором диметилсульфоксида с получением конечной концентрации 50 мкг/мл для прямого ELISA.
Покрытые стрептавидином титрационные микропланшеты (Perkin Elmer) покрывали пептидами в концентрации 50 мкг/мл на каждую лунку в объеме 50 мкл и инкубировали при комнатной температуре в течение 1 часа. Раствор пептида удаляли и PBS-, содержащий 4% Block-Ace, добавляли в каждую лунку и инкубировали в течение ночи при 4°С. Каждую лунку промывали три раза промывочным буфером для ELISA (0,02% Tween20 в PES-). Антитела к XCR1 человека (2Н6, 5G7 и 11Н2) добавляли в каждую лунку в концентрации 10 мкг/мл и инкубировали в течение 6 часов при комнатной температуре. Каждую лунку промывали три раза промывочным буфером для ELISA. Конъюгированные с пероксидазой хрена антитела осла к IgG мыши (Jackson, #715-035-150), разбавленные в соотношении 1:5000 в промывочном буфере для ELISA, добавляли в каждую лунку и инкубировали в течение 1 часа при комнатной температуре. Каждую лунку промывали три раза промывочным буфером для ELISA. TMBZ (3,3',5,5' tetramethyl benzidine; Sigma) добавляли в каждую лунку и инкубировали при комнатной температуре. Реакцию останавливали при помощи 2N H2SO4 и А450нм измеряли при помощи спектрофотометра для прочтения планшетов Arvo (PerkinElmer).
Антитела к XCR1 человека 2Н6 и 5G7 проявляли сильное связывание с одним пептидом, содержащим 7PESTTFFYYDLQ18 (SEQ ID NO: 96), и слабое связывание с 11TFFYYDLQSQPC22 (SEQ ID NO: 110) (Фиг. 14). Также 5G7 проявляли слабое связывание с тремя непоследовательными пептидами, содержащими 19SQPCENQAWVFA30 (SEQ ID NO: 101), 172SSGCDYSELTWY183 (SEQ ID NO: 110) и 175CDYSELTWYLTS186 (SEQ ID NO: 111). С другой стороны, 11Н2 не проявляли реактивности в отношении этих пептидов (данные не показаны).
Пример 9
Картирование остатков связывания мышиных антител к XCR1 человека (2Н6, 5G7 и 11Н2) и гуманизированных антител к XCR1 человека (HK1L2 и HK5L51 с внеклеточными доменами XCR1 человека с помощью аланиновых мутантов
Чтобы определить особенно важные остатки XCR1 человека, распознаваемые мышиными антителами к XCR1 человека (2Н6, 5G7 и 11Н2) и гуманизированными антителами к XCR1 человека (HK1L2 и HK5L5), проводили анализ с заменой аланина.
Подготавливали панель аланин-замещенных мутантов XCR1 человека. В этой панели каждую аминокислоту в 7PESTTFFYYDLQSQPCENQAWVFA30 (SEQ ID NO: 118) и 175CDYSELTWYLTS186 (SEQ ID NO: 119) внеклеточных областей XCR1 человека замещали на аланин. Векторы экспрессии для аланин-замещенных мутантов конструировали при помощи сайт-направленного мутагенеза. Каждый мутант экспрессировался на клетках В300.19, и реактивность антитела определяли с помощью FACS-анализа. Исходные клетки В300.19 и клетки В300.19, экспрессирующие XCR1 человека-EGFP или XCR1-EGFP, содержащие каждый из аланин-замещенных мутантов, смешивали в соотношении 1:1 и суспендировали в FACS буфере (PBS- (Sigma), содержащий 1% эмбриональную бычью сыворотку). Клетки блокировали в течение 10 минут на льду с FACS буфером, содержащим 10% сыворотку крови крысы. Затем клетки инкубировали в течение 20 минут на льду с мышиными антителами к XCR1 человека (2Н6, 5G7 и 11Н2), гуманизированными антителами (HK1L2 или HK5L5), мышиными антителами изотипического контроля, IgG2a (eBioscience, #14-4724-82) или IgG2b (eBioscience, #14-4732-82) или антителом человека IgG2 изотипического контроля (Sigma, #15404) в концентрации 10 мкг/мл или инкубировали с FACS буфером без антитела. Клетки промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с РЕ-меченым поликлональным антителом к IgG мыши (Jackson, #715-116-151, разведенным в соотношении 1:50 в FACS буфере, используемом для клеток, которые инкубировали с мышиными антителами), РЕ-меченым поликлональным антителом к IgG человека (Jackson, #709-116-149, разбавленное в соотношении 1:50 в FACS буфере, используемом для клеток, которые инкубировали с гуманизированными антителами или контрольными IgG человека) или РЕ-меченым поликлональным антителом к XCR1 человека (R&D, #FAB857P, разведенным в соотношении 2:5 в FACS буфере, используемом для клеток, которые инкубировали с FACS буфером без антител). Клетки промывали три раза FACS буфером и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II (BD Bioscience).
Каждый аланин-замещенный мутант обнаруживали с помощью РЕ-меченого поликлонального антитела к XCR1 человека, за исключением мутанта С175А (Фиг. 15). Поскольку уровень экспрессии каждого аланинового мутанта на поверхности клеток варьировал среди этих мутантов, как показано на Фиг. 15, то реактивность антител к каждому аланиновому мутанту оценивали по относительному среднему значению РЕ (mAb/pAb), которое вычисляли согласно следующей процедуре. Сначала относительное среднее значение РЕ для каждого антитела вычисляли, устанавливая среднее значение РЕ, которое получали при окраске клеток В300.19 (дикого типа), экспрессирующих XCR1 человека-EGFP, при использовании каждого антитела, как 1,0. Затем относительные средние значения РЕ (mAb/pAb) вычисляли при помощи следующего уравнения: каждое относительное среднее значение РЕ для мышиных антител к XCR1 человека (2Н6, 5G7 или 1H2) или гуманизированных антител (HK1L2 или HK5L5) делили на относительное среднее значение РЕ для РЕ-меченых поликлональных антител к XCR1 человека. Результаты показали, что 2Н6 (Фиг. 16), 5G7 (Фиг. 17), HK1L2 (Фиг. 19) и HK5L5 (Фиг. 20) проявляют более низкую реактивность по отношению ко многим аланиновым мутантам, у которых каждый остаток в N-конечном участке или 2-й петле был замещен аланином. В частности, наблюдалось отсутствие реактивности или слабая реактивность в отношении мутантов Y14A, D16A и L17A. Кроме того, в отношении мутантов Е8А, F13A, С22А и Y177A реактивность была ниже. Взятые вместе, эти результаты означают, что 2Н6, 5G7, HK1L2 и HK5L5 распознают Е8, F13, Y14, D16, L17, С22 и Y177 внеклеточного домена XCR1 человека. 11Н2 (Фиг. 18) проявляло реактивность сходную с реактивностью других mAbs, за исключением F13A и D16A, что указывает на то, что 11Н2 связывается с Е8, Y14, L17, C22hY177.
Пример 10
Конкуренция между мышиными антителами к XCR1 человека (2Н6, 5G7 и 11Н21, распознающими похожие эпитопы, за связывание с клетками, экспрессирующими XCR1 человека.
Чтобы определить, конкурируют ли мышиные антитела к XCR1 человека, распознающие похожие эпитопы, друг с другом за связывание с XCR1 человека, выполняли анализ конкурентного связывания.
Анализ конкурентного связывания проводили согласно следующей процедуре. Исходные клетки В300.19 и клетки В300.19, экспрессирующие XCR1 человека-EGFP, смешивали в соотношении 1:1 и суспендировали в FACS буфере (PBS- (Sigma), содержащий 1% эмбриональную бычью сыворотку). Клетки блокировали в течение 10 минут на льду с FACS буфером, содержащим 10% сыворотку крови крысы. Затем клетки инкубировали с мышиными антителами к XCR1 человека (2Н6, 5G7 или 11Н2), мышиными антителами изотипического контроля, IgG2a (eBioscience, #16-4724-85) или IgG2b (eBioscience, #16-4732-85), при различных концентрациях от 0 до 10 мкг/мл в FACS буфере в течение 20 минут на льду. Затем клетки инкубировали с биотинилированным мышиным антителом к XCR1 человека (5G7) в концентрации 0,3 мкг/мл в FACS буфере в течение 20 минут на льду. Клетки промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с РЕ-меченым стрептавидином (BD Pharmingen, #554061, разведенным FACS буфером с коэффициент разбавления 1:50). Клетки промывали три раза FACS буфером и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II (BD Bioscience).
Само немеченое 5G7, немеченые 2Н6 и 11Н2, распознающие похожие эпитопы на XCR1 человека, конкурировали с биотинилированным мышиным антителом к XCR1 человека (5G7) за связывание с клетками В300.19, экспрессирующими XCR1 человека-EGFP (Фиг. 21). С другой стороны, контрольные антитела не конкурировали с биотинилированным антителом (5G7) за связывание с клетками В300.19, экспрессирующими XCR1 человека-EGFP.
Пример 11
Реактивность мышиного моноклонального антитела к XCR1 человека. 5G7, и гуманизированных моноклональных антител к XCR1 человека, HK1L2 и I1K5L5. по отношению к различным рецепторам хемокинов человека
Реактивность мышиного моноклонального антитела к XCR1 человека, 5G7, и гуманизированных моноклональных антител к XCR1 человека, HK1L2 и HK5L5, по отношению к различным рецепторам хемокинов человека оценивали с помощью FACS-анализа
Исходные клетки В300.19 и клетки В300.19, экспрессирующие рецептор хемокинов человека-EGFP (XCR1, CXCR1, CXCR3, CXCR4, CXCR5, CXCR6, CCR1, CCR2B, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR11 или CX3CR1), суспендировали в FACS буфере (PBS- (Sigma), содержащий 1% эмбриональную бычью сыворотку) в концентрации 1×106 клеток/мл и аликвоты по 100 мкл распределяли в лунки 96-луночного круглодонного планшета. Затем клетки центрифугировали и надосадочную жидкость удаляли. Мышиное mAb к XCR1 человека, 5G7, мышиное антитело изотипического контроля IgG2b (eBioscience, #14-4732-82), гуманизированные моноклональные антитела к XCR1 человека, HK1L2 и HK5L5, и контрольное антитело человека IgG (Mitsubishi, #128-26053-9) разводили FACS буфером в концентрации 5 мкг/мл. РЕ-меченые поликлональные антитела козы к XCR1 человека (R&D, #FAB857P и LifeSpan Bioscience, #LS-C76885) разводили FACS буфером с коэффициентом разбавления 2:5 и 1:5, соответственно. Пятьдесят мкл разбавленных антител добавляли в каждую лунку и клетки инкубировали в течение 20 минут на льду. Затем клетки промывали три раза FACS буфером. РЕ-меченое мышиное поликлональное антитело IgG (Jackson, #715-116-151, разведенное FACS буфером с коэффициентом разбавления 1:50) добавляли к клеткам, которые инкубировали с 5G7 или мышиным антителом изотипического контроля. РЕ-меченое поликлональное антитело к IgG человека (Jackson, #709-116-149, разведенное FACS буфером с коэффициентом разбавления 1:50) добавляли к клеткам, которые инкубировали с HK1L2 и HK5L5 или контрольным антителом IgG человека. FACS буфер добавляли к клеткам, которые инкубировали с поликлональными антителами к XCR1. Затем клетки инкубировали в течение 20 минут на льду. Клетки промывали FACS буфером три раза и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II и выражали как допустимую ошибку среднего значения РЕ. Допустимую ошибку среднего значения РЕ вычисляли, вычитая исходное среднее значение РЕ из каждого среднего значения РЕ, которое получали при окраске каждой клеточной линии с каждым антителом.
Мышиное антитело к XCR1 человека, 5G7, селективно взаимодействовало с клетками В300.19, экспрессирующими XCR1 человека-EGFP, кроме клеток, экспрессирующих CX3CR1 человека-EGFP (Фиг. 22). С другой стороны, поликлональные антитела козы к XCR1 человека взаимодействовали с различными клетками, экспрессирующими рецептор хемокинов человека-EGFP, в дополнение к клеткам, экспрессирующим XCR1 человека-EGFP (Фиг. 22). Гуманизированные антитела к XCR1 человека, HK1L2 и HK5L5, проявляли сниженную реактивность в отношении клеток, экспрессирующих CX3CR1 человека-EGFP, несмотря на их высокую реактивность в отношении клеток, экспрессирующих XCR1 человека-EGFP (Фиг. 23).
Пример 12
Эффект mAb 5G7 на DTH-ответ, индуцированный Mycobacterium butyricum
Известно, что гиперчувствительность замедленного типа (DTH) является одним из главных механизмов, вызывающих аутоиммунные заболевания, такие как тиреоидит, ревматоидный артрит и сахарный диабет 1 типа, если этот ответ направлен против аутоантигенов (Actor, J.K. and Ampel, N.M. (December 2009) Hypersensitivity: Τ Lymphocyte-mediated (Type TV). In: Encyclopedia of Life Sciences (ELS). John Wiley & Sons, Ltd: Chichester). Взаимодействие Т-клеток и дендритных клеток является особенно важным для DTH-ответа. Таким образом, ингибирование взаимодействия Т-клеток и DC, как полагают, может быть полезным при лечении этих заболеваний. Поэтому изучали эффект mAb к XCR1 человека, 5G7, на модели реакции DTH, индуцированной Mycobacterium (M.) butyricum DTH-ответа, у нокин по гену XCR1 мышей (Mihara, M. et al, Immunology Letters 2002, 84: 223-229; Mohan К et al, Eur. J. Immunol. 2005, 35: 1702-1711).
(Способы)
Сконструированных нокин по гену hXCRl мышей, у которых XCR1 человека экспрессируется вместо XCR1 мыши, иммунизировали подкожно убитой нагреванием М. butyricum (100 мкг/особь) с минеральным маслом в День 0. MAb 5G7 или контрольное IgG мыши (Jackson Laboratory) внутрибрюшинно инъецировали в дозе 500 мкг/особь в день 1, день 3, день 7 и день 9. Спустя 10 дней после иммунизации М. butyricum осуществляли контрольное заражение мышей M. butyricum, суспендированным в минеральном масле, в ступню правой лапы (20 мкг/ступня, контрольное заражение M. butyricum) и вводили минеральным маслом само по себе в ступню левой лапы (контрольный стимул). Спустя один день после инъекционного контрольного заражения оценивали DTH-ответ, измеряя толщину каждой лапы. Опухание лапы вычисляли согласно следующей формуле.
Опухание ступни = ([А]-|В])-([С]-[D])
[A] = толщина правой ступни после контрольного заражения M. butyricum
[B] =толщина правой ступни перед контрольным заражением M. butyricum
[C] =толщина левой ступни после контрольного стимула
[D] =толщина левой ступни перед контрольным стимулом.
(Результаты)
Результат продемонстрировал, что DTH-ответ, индуцированный M. butyricum, у мышей, обработанных mAb 5G7, проявлялся как значительно сниженный DTH-ответ по сравнению с мышами, обработанными контрольным IgG (Фиг. 24).
(Вывод)
Результаты продемонстрировали эффективность обработки антителом к XCR1 при DTH-ответе. Предполагается, что применение антител к XCR1 может быть полезным при лечении аутоиммунных заболеваний, связанных с DTH, таких как тиреоидит, ревматоидный артрит и сахарный диабет 1 типа.
Пример 13
Эффект mAb 5G7 на ЕАЕ, опосредованный пептидом MOG 37-50
Рассеянный склероз (MS) является хроническим демиелинизирующим заболеванием центральной нервной системы (CNS) человека, которое может клинически характеризоваться ремиттирующе-рецидивирующим или хронически прогрессирующим течением. Наиболее тщательно изученная животная модель MS, экспериментальный аутоиммунный энцефаломиелит (ЕАЕ), обычно приводит к нарушению моторных функций. Во многих сообщениях указано, что Т-клетки играют важную роль в патогенезе MS и ЕАЕ. Таким образом, осуществляли эксперимент в виде модели ЕАЕ, чтобы исследовать ингибиторную активность mAb 5G7 в патогенезе MS.
(Способ проведения эксперимента)
1. Мыши для применения образца
Для эксперимента использовали нокин по гену XCR1 человека мыши (в возрасте 7-12 недель), у которых XCR1 человека экспрессируется вместо XCR1 мыши, происходящие от C57BIV6.
2. Индукция ЕАЕ
Индукцию ЕАЕ выполняли согласно способу, изложенному в Journal Eur. J. Immunol. 2005, 35: 76-85, в котором указывалась возможная роль CD8+ Т-клеток в развитии ЕАЕ. Кратко, нокин по гену XCR1 человека мышам подкожно инъецировали 200 мкг пептида 37-50 миелина гликопротеина олигодендроцитов (MOG 37-50), эмульгированного в полном адъюванте Фрейнда (CFA), содержащем Mycobacterium tuberculosis H37Ra в концентрации 20 мг/мл. 200 нг токсина коклюша вводили внутривенно в день 0 и 2 после иммунизации.
3. Способ введения антител
Мышиное моноклональное антитело к XCR1 человека (5G7) и его контрольное антитело, т.е. IgG мыши (Jackson Laboratory), подготавливали в PBS в конечной концентрации 2 мг/мл. Каждое из вышеуказанных антител вводили внутривенно мышам в объеме 250 мкл/мышь (500 мкг/мышь) в день 7, день 10, день 14 и день 17.
4. Оценка показателей патологии этой модели
Клинические симптомы ЕАЕ контролировали с первого дня иммунизации и оценивали по шкале от 0 до 5 на основе следующих критериев:
степень 0: отсутствие заболевания, степень 0,5: слабый паралич хвоста, степень 1: паралич хвоста, степень 2: неровная походка, степень 2,5: одна парализованная задняя конечность, степень 3: паралич задних конечностей, степень 4: парализованные передние и задние конечности, и степень 5: агония или смерть.
(Результат эксперимента и вывод)
Полученные клинические показатели у мышей, которым вводили mAb 5G7, были более низкого уровня по сравнению с показателями у мышей, которым вводили контрольный IgG (Фиг. 25). Результаты указывали на то, что обработка антителом к XCR1 вызвала достоверный уровень подавления развития ЕАЕ, также предполагалось, что обработка антителами к XCR1 может быть полезной при лечении MS у человека.
Пример 14
Ингибирование связывания XCL1 человека с клетками, экспрессирующими XCR1 человека, с помощью мышиных антител к XCR1 человека (2Н6,5G7 и 11Н2)
Чтобы определить, ингибируют ли мышиные антитела к XCR1 человека (2Н6,5G7 и 11Н2) связывание XCL1 человека с XCR1 человека, выполняли анализ конкурентного связывания лиганда.
Сначала связывание XCL1 человека-SSS-His(10) с клетками BaF3, экспрессирующими XCR1 человека-EGFP, определяли согласно следующей процедуре. Исходные клетки BaF3 и клетки BaF3, экспрессирующие XCR1 человека-EGFP, смешивали в соотношении 1: 1 и суспендировали в FACS буфере (PBS- (Sigma), содержащий 1% FBS). Клетки инкубировали в течение 30 минут на льду с увеличивающейся концентрацией XCL1 человека-SSS-His(10) в присутствии или в отсутствии 2,5 мкМ растворимого XCL1 (R&D, #695-LT-025/CF) в FACS буфере. Затем клетки промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с антителом к 6xHis-tag (BETHYL, #А 190-1 НА, разведенным в соотношении 1:100 в FACS буфере). Клетки снова промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с РЕ-меченым антителом к IgG кролика (Jackson, #711-166-152, разведенным в соотношении 1:50 в FACS буфере). Затем, клетки еще раз промыли три раза FACS буфером и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II (BD Bioscience). Специфическое связывание определяли, вычитая показатели неспецифического связывания (в присутствие 2,5 мкМ растворимого XCL1) из показателей общего связывания (в отсутствии 2,5 мкМ растворимого XCL1).
Анализ конкурентного связывания лиганда выполняли согласно следующей процедуре. Исходные клетки BaF3 и клетки BaF3, экспрессирующие XCR1 человека-EGFP, смешивали в соотношении 1:1 и суспендировали в FACS буфере (PBS– (Sigma), содержащий 1% FBS). Клетки блокировали в течение 10 минут на льду FACS буфером, авдержащим 10% сыворотку крови крысы. Затем клетки инкубировали в течение 20 минут на льду с мышиными антителами к XCR1 человека (2Н6, 5G7 и 11Н2), мышиными антителами изотипического контроля, IgG2a (eBioscience, #16-4724-85) или IgG2b (eBioscience, #164732-85), при различных концентрациях от 0 до 150 мкг/мл. Затем клетки инкубировали в течение 30 минут на льду с XCL1 человека-SSS-His(10) в насыщающей концентрации 0,12 мкг/мл. Клетки промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с антителом к 6xHis-tag (BETHYL, #А190-114А, разведенным в соотношении 1:100 в FACS буфере). Клетки снова промывали FACS буфером три раза и затем инкубировали в течение 20 минут на льду с РЕ-мечеными антителами к IgG кролика (Jackson, #711-166-152, разбавленными в соотношении 1:50 в FACS буфере). Затем клетки еще раз промывали три раза FACS буфером и затем суспендировали в FACS буфере. Интенсивность флуоресценции измеряли при помощи анализатора клеток FACSCanto II (BD Bioscience).
Происходило ингибирование связывания XCL1 человека с клетками BaF3, экспрессирующими XCR1 человека-EGFP, мышиными антителами к XCR1 человека (2Н6, 5G7 и 11Н2), и IC50 антител составляла 37,0 нМ, 6,9 нМ и 23,8 нМ соответственно. С другой стороны, контрольные антитела не ингибировали связывание XCL1 человека с клетками BaF3, экспрессирующими XCR1 человека-EGFP.


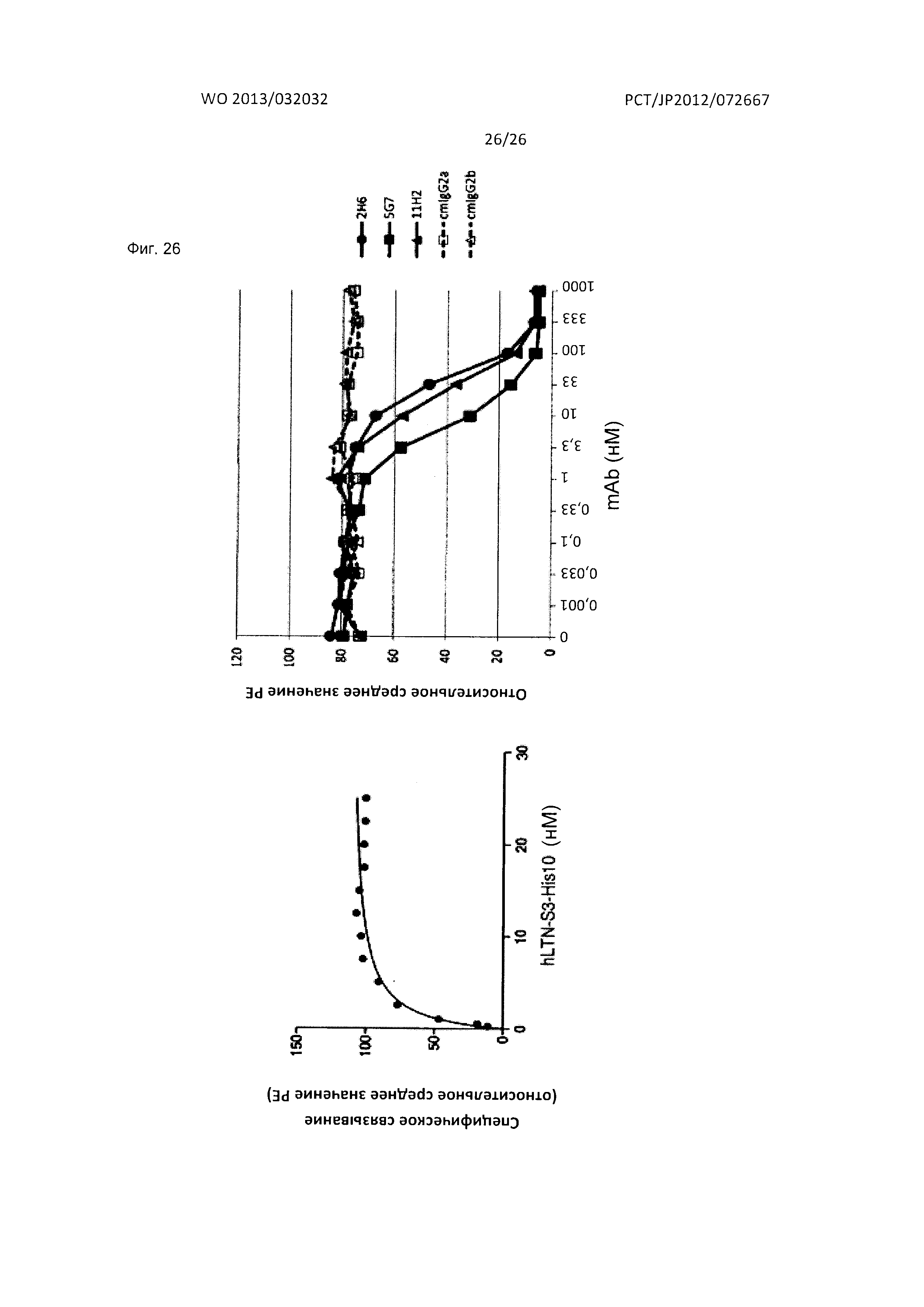

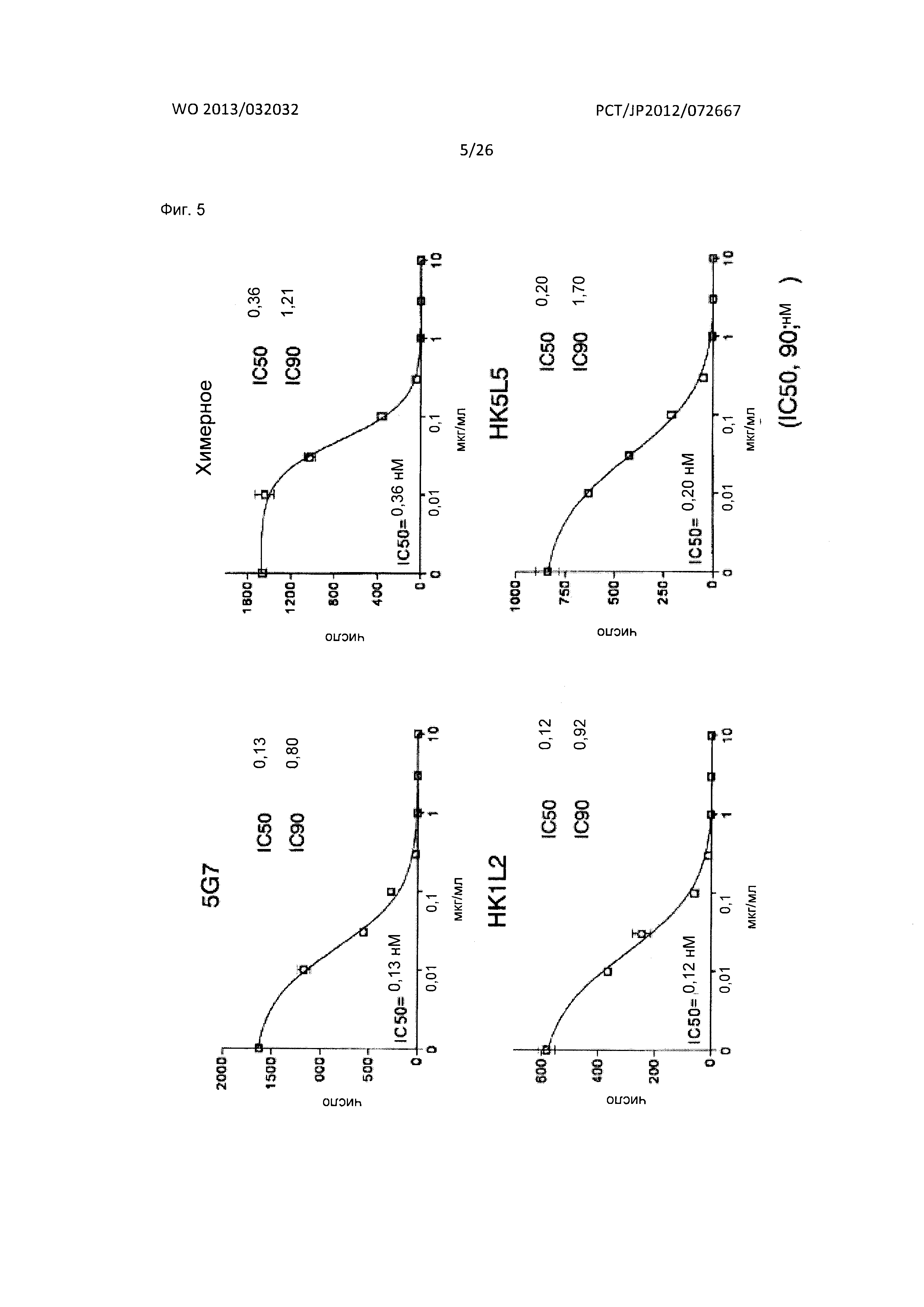






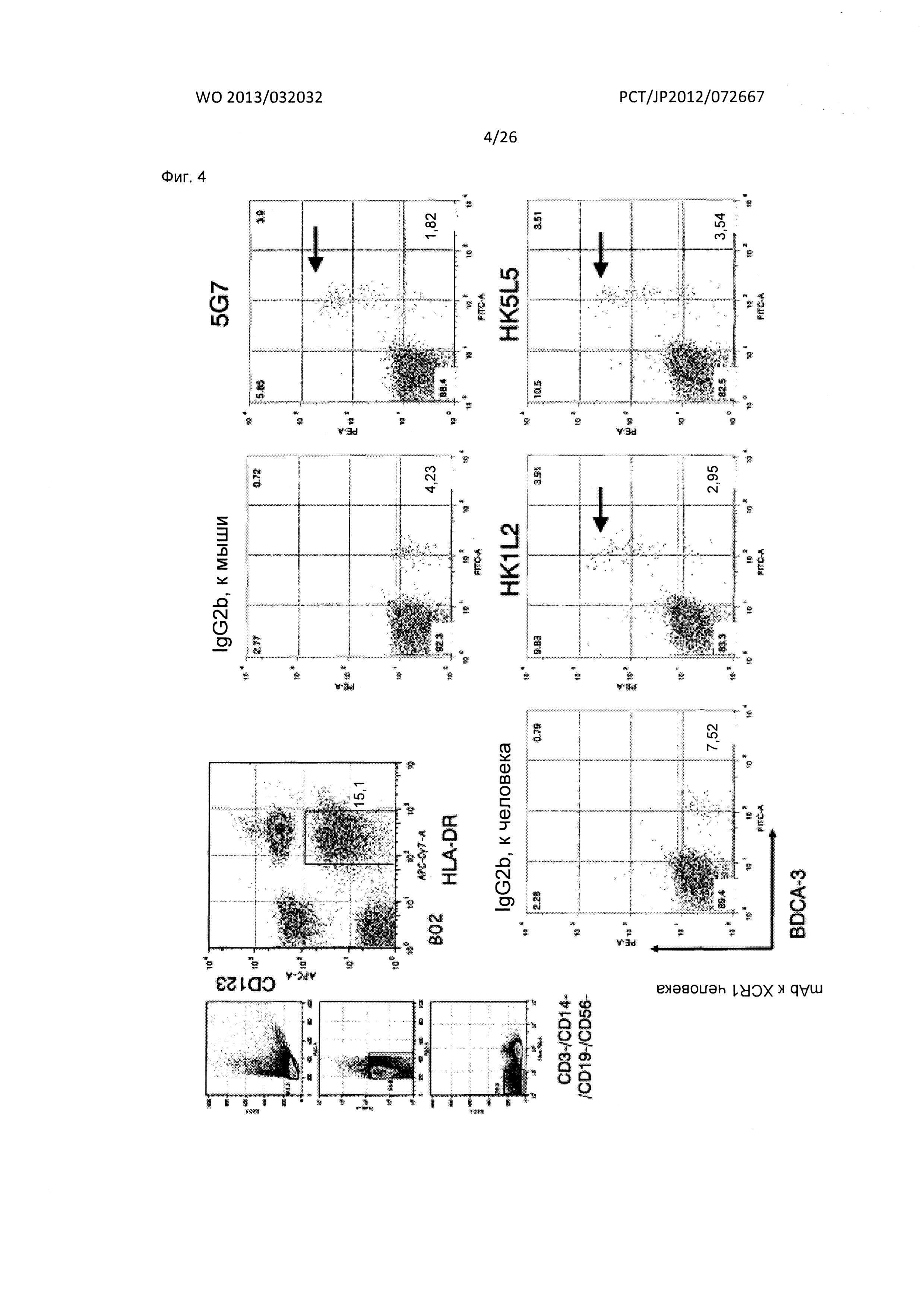



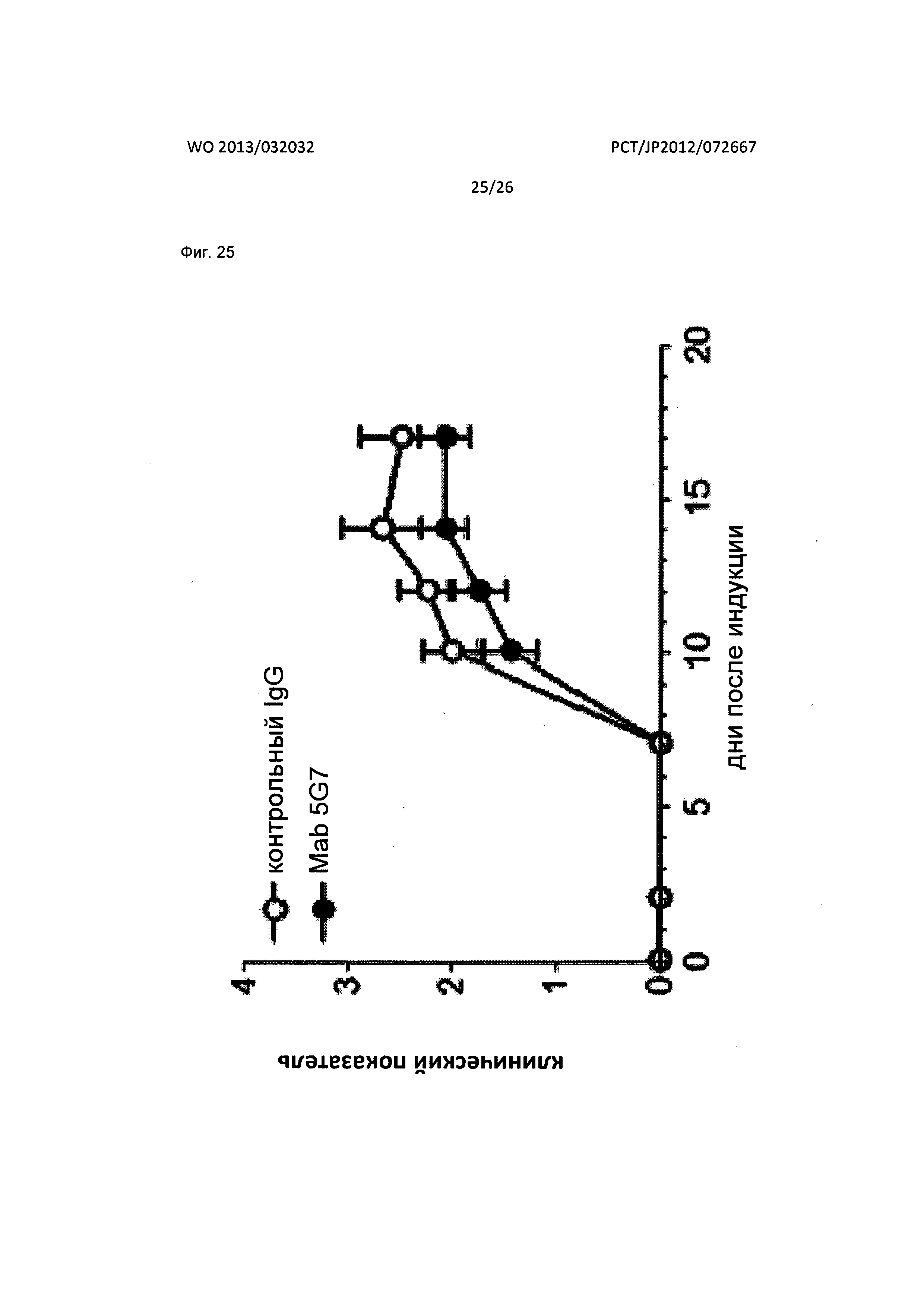
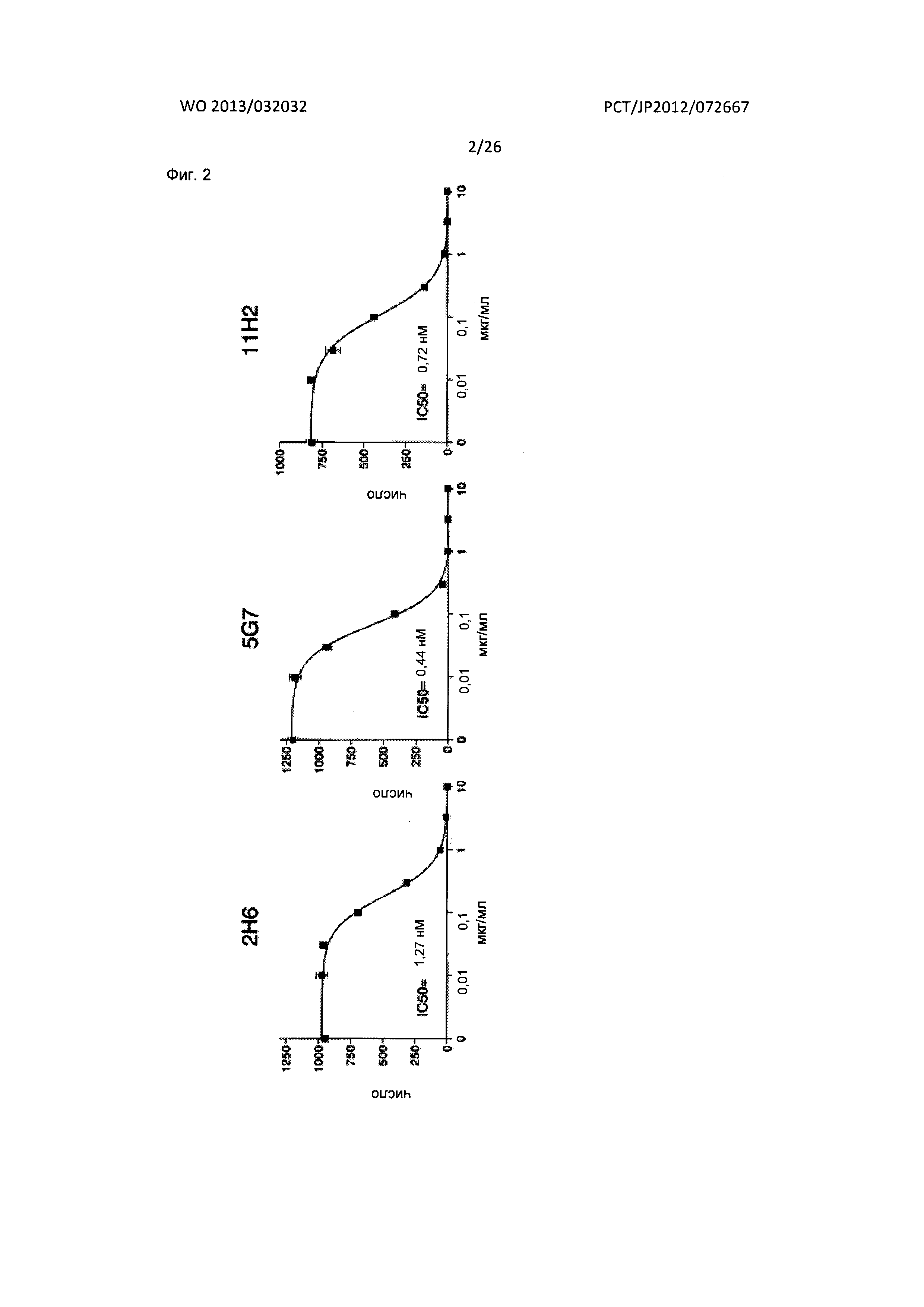
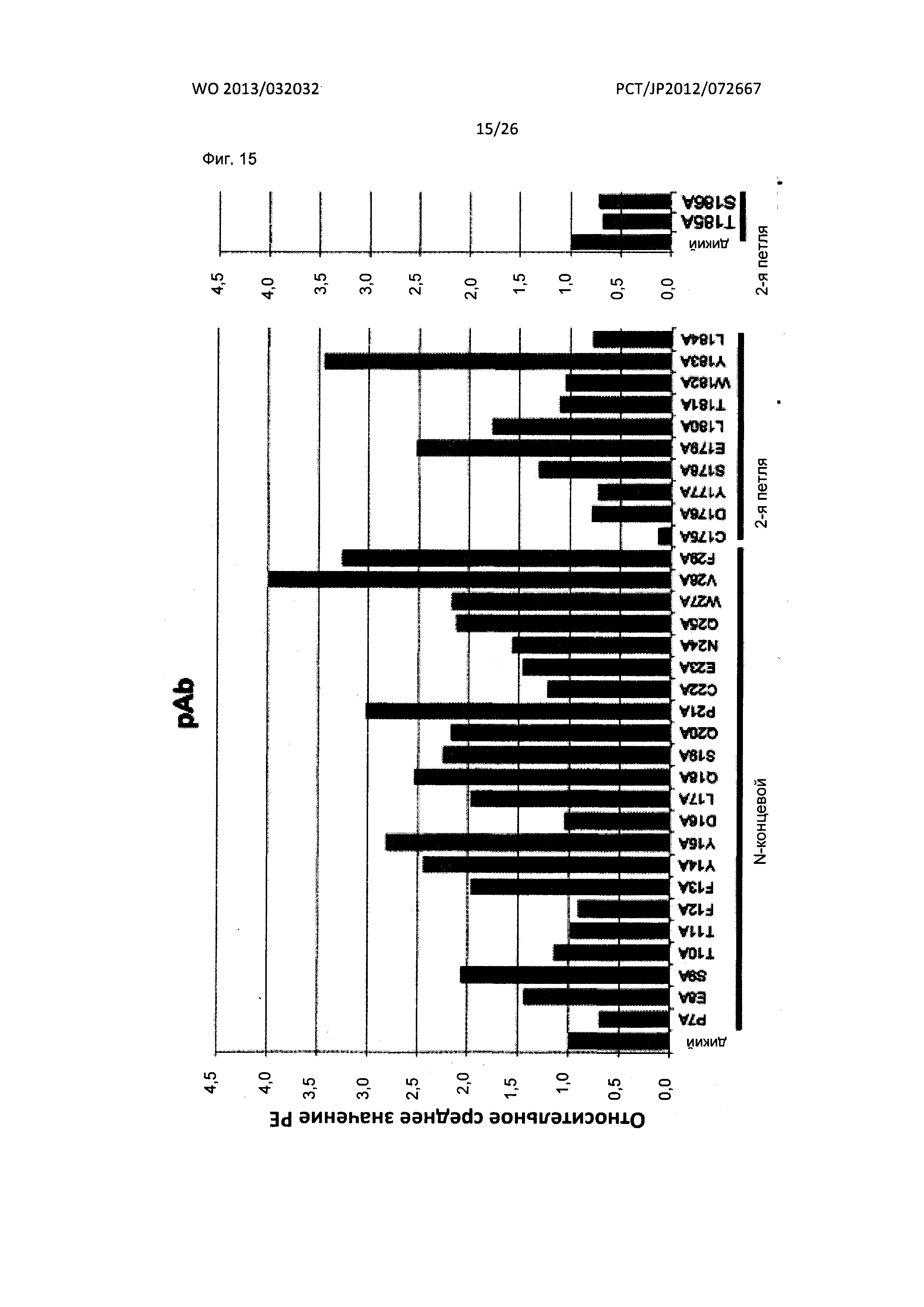
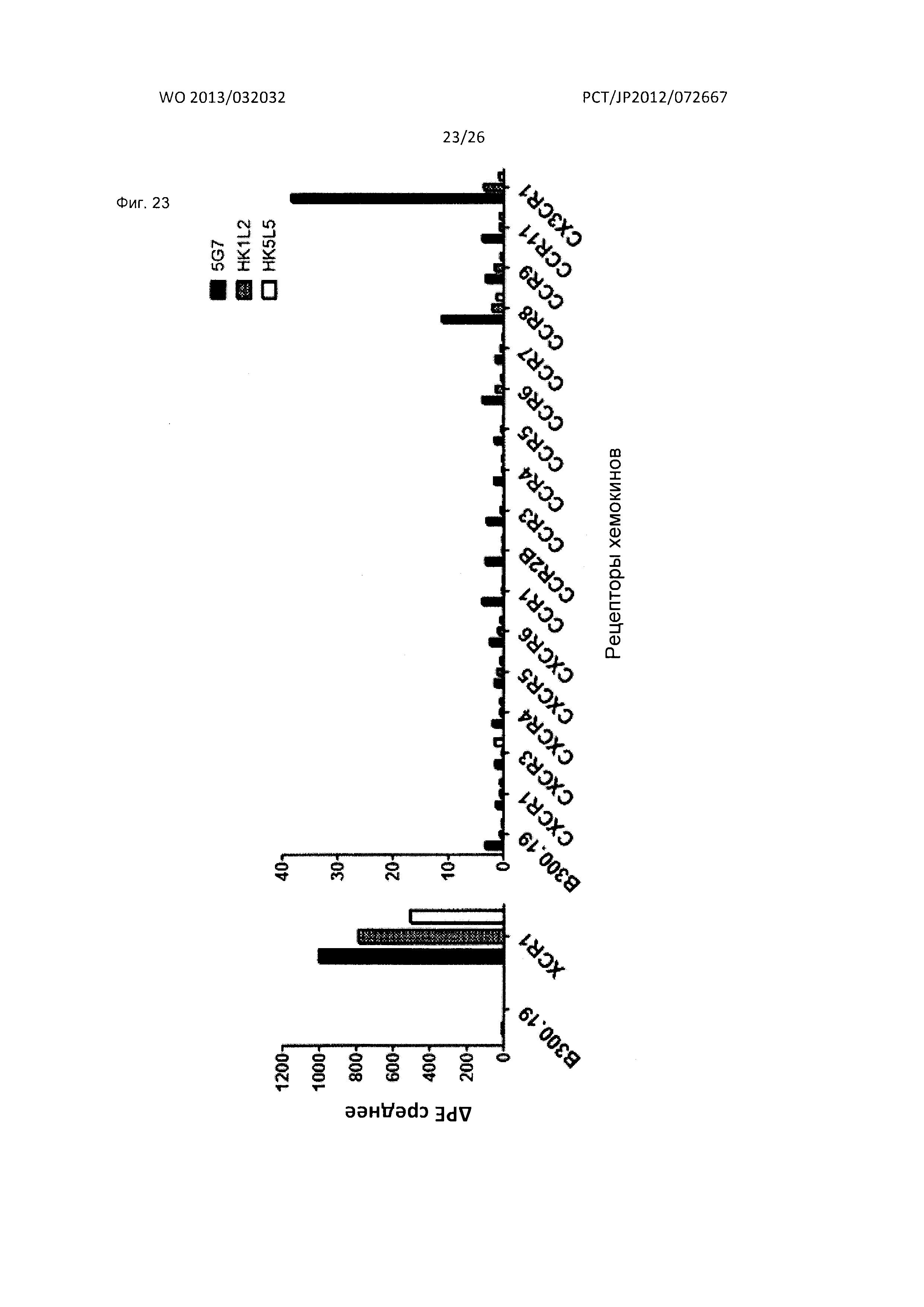

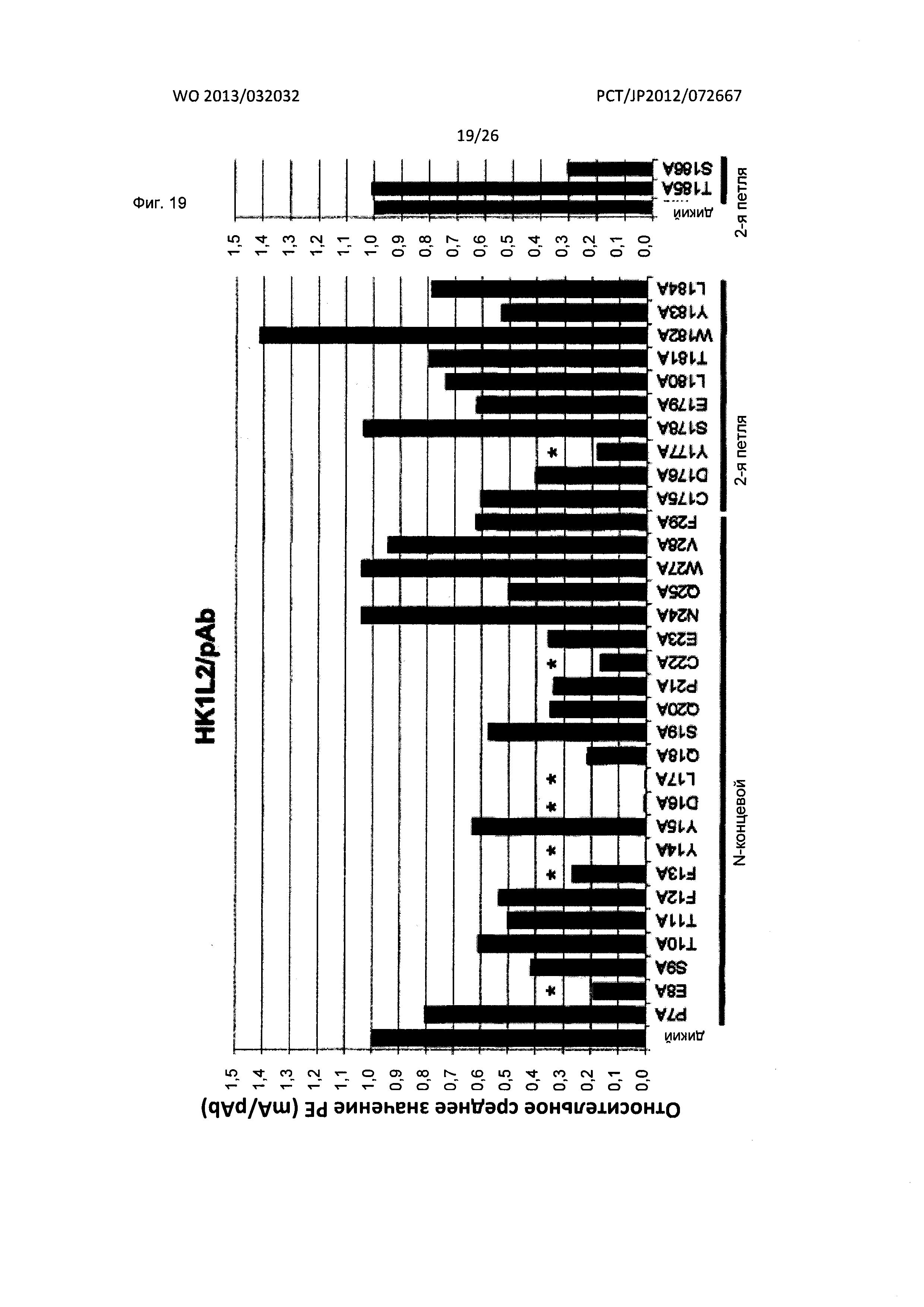



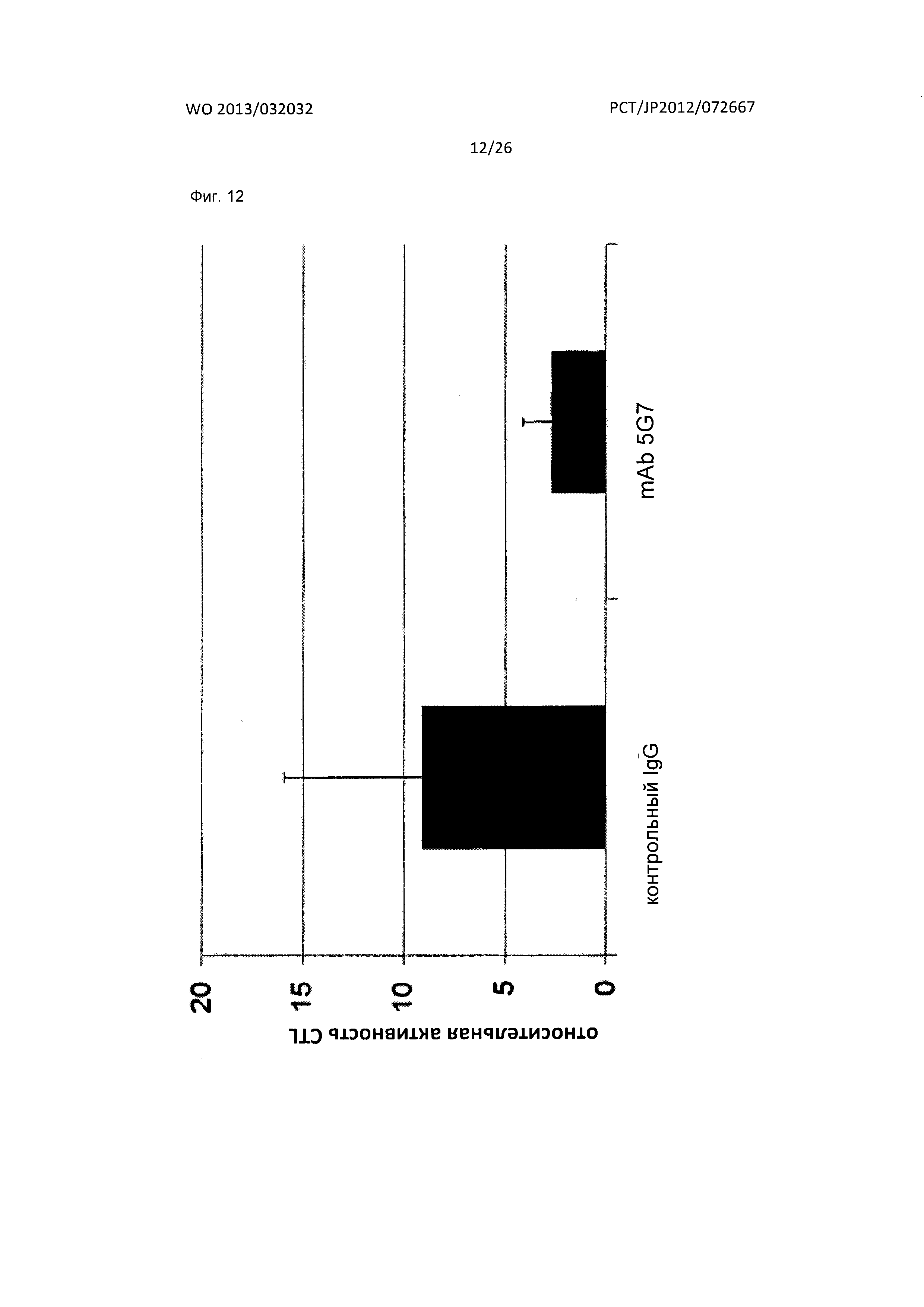
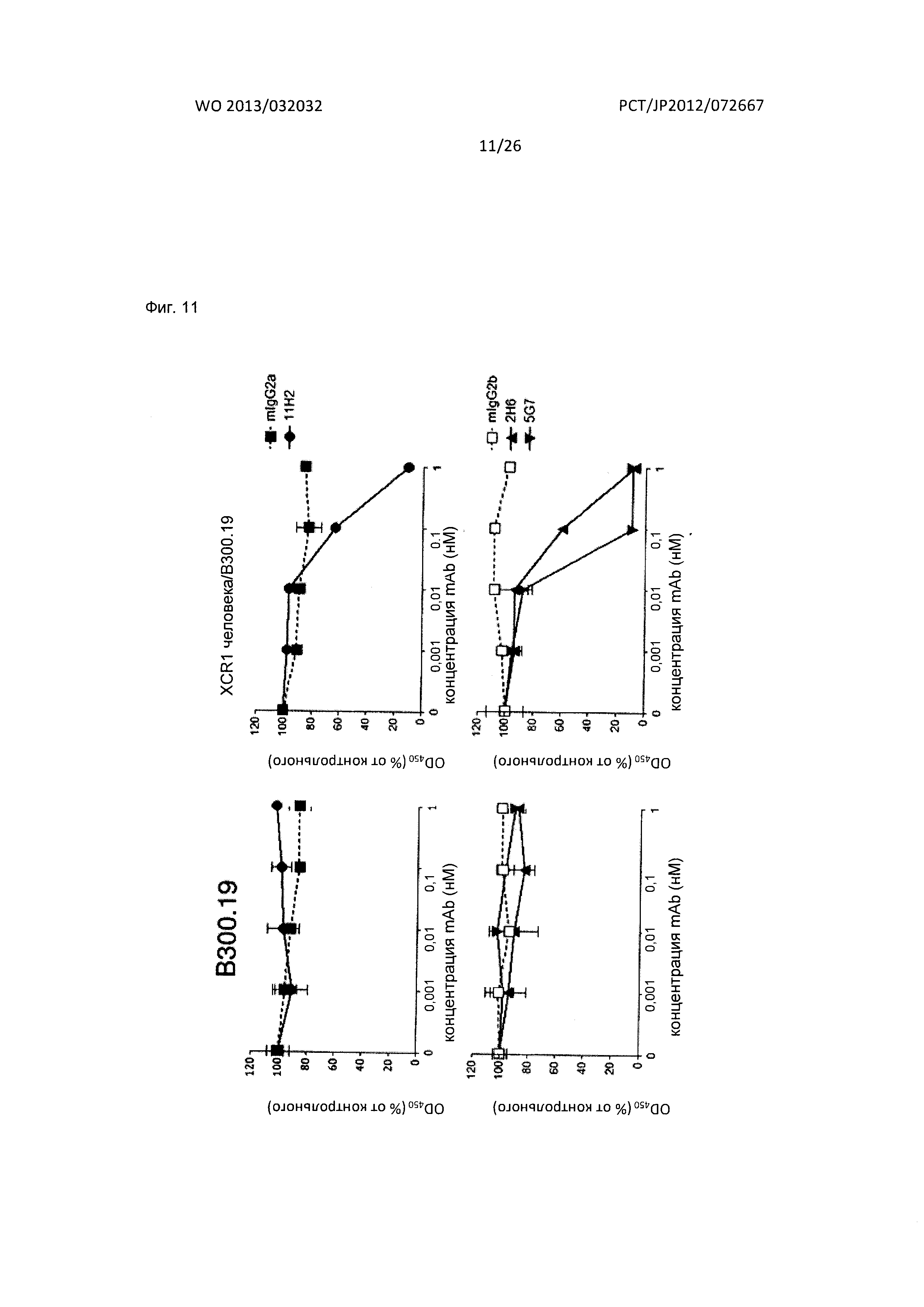



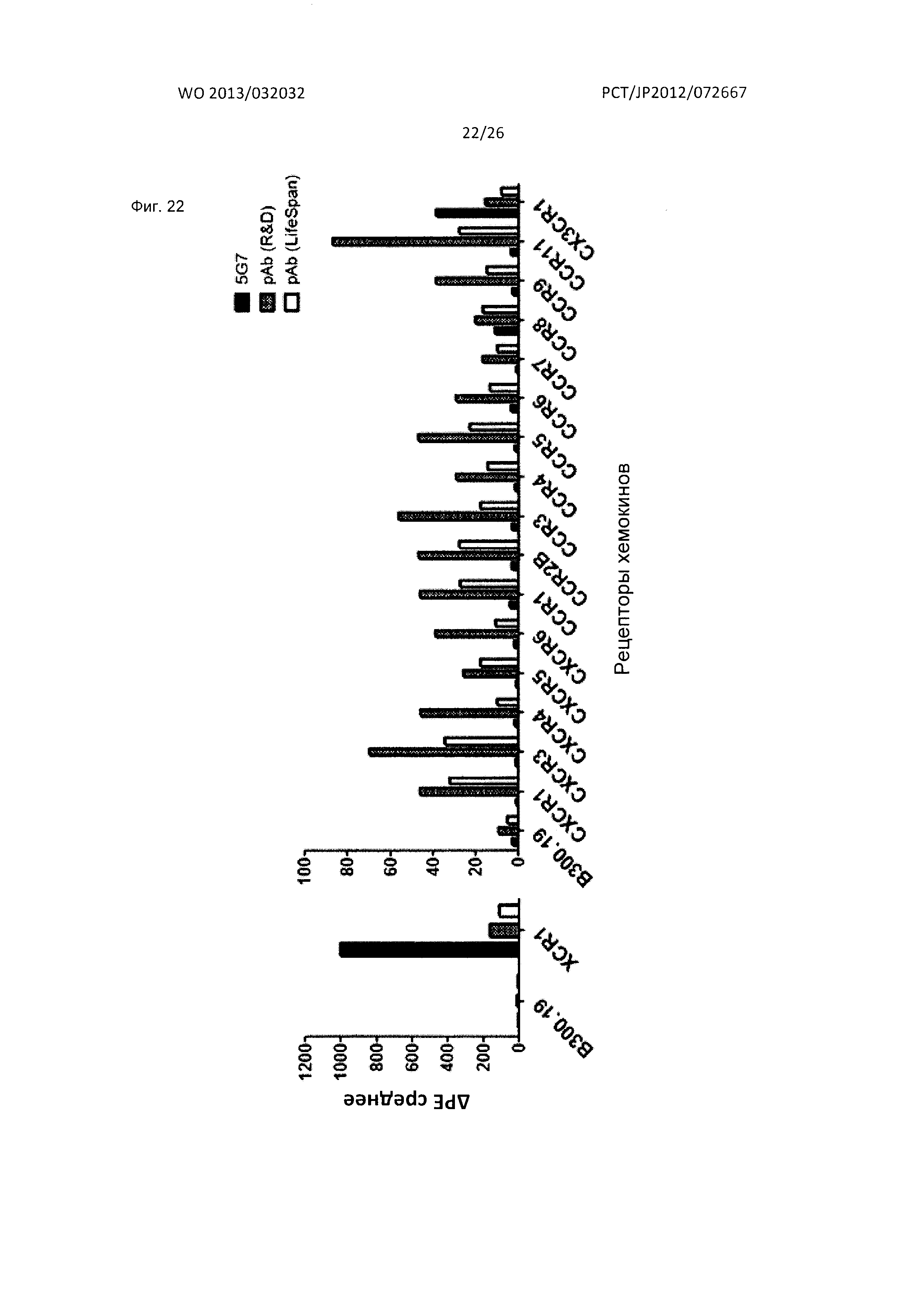




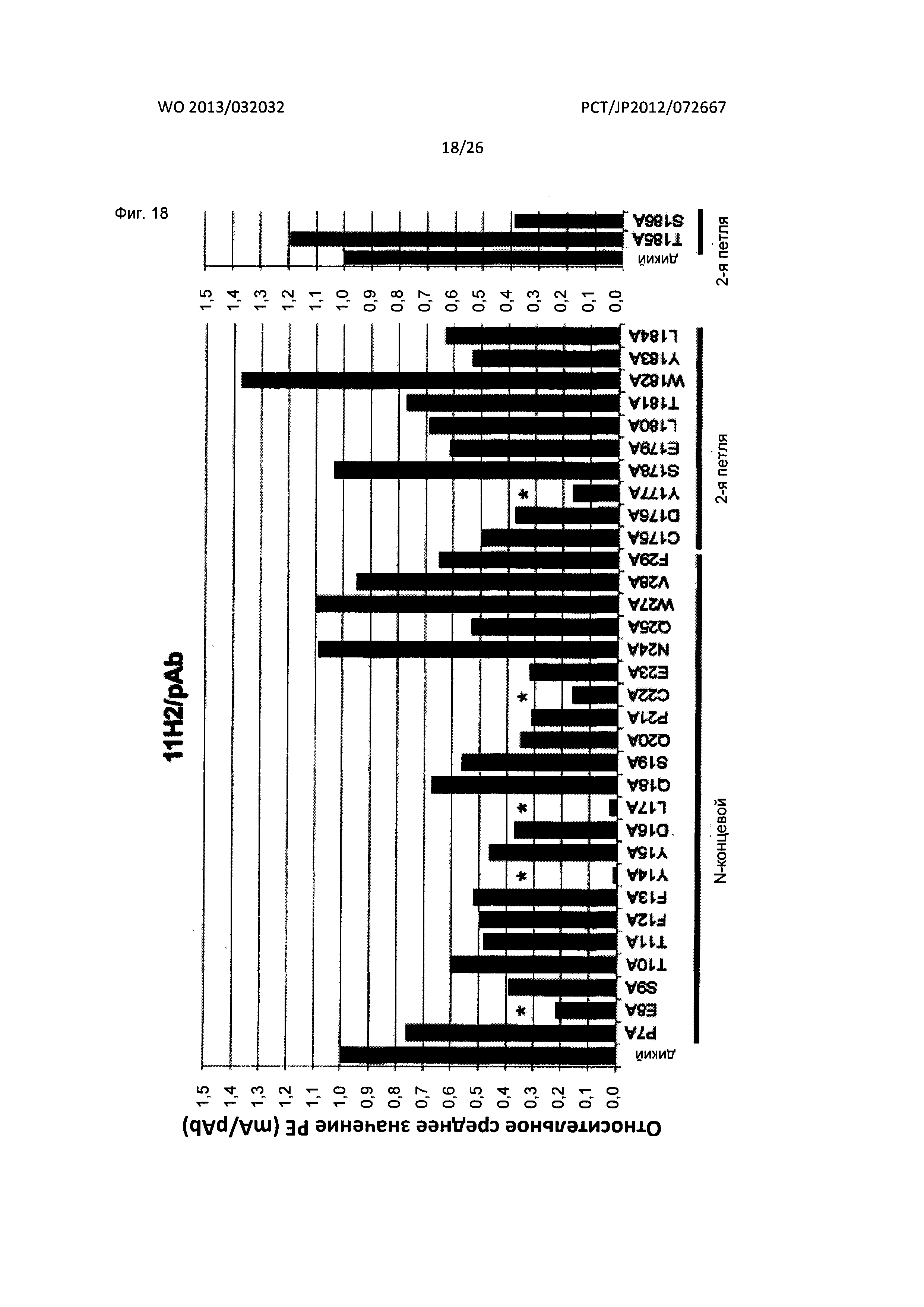







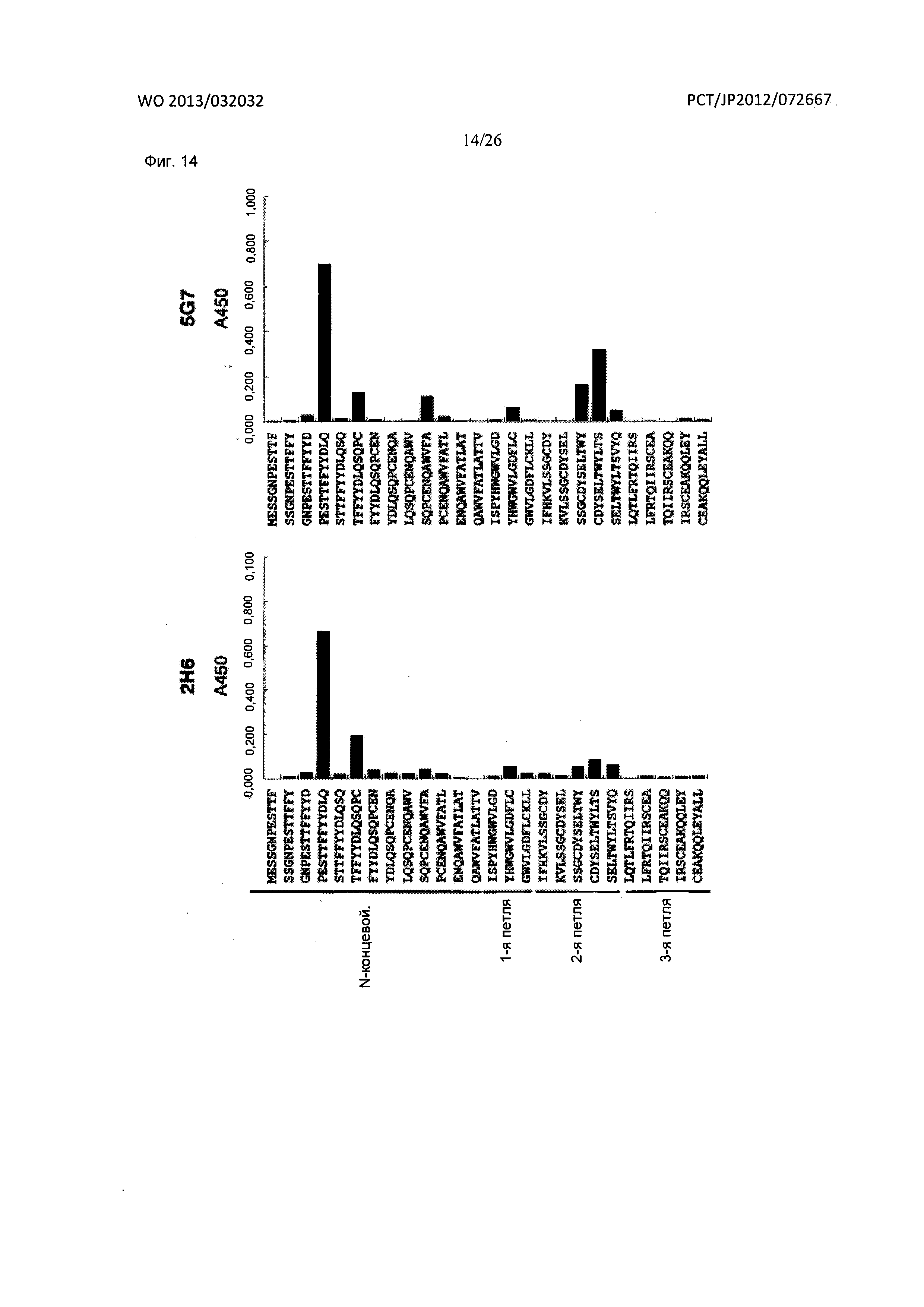
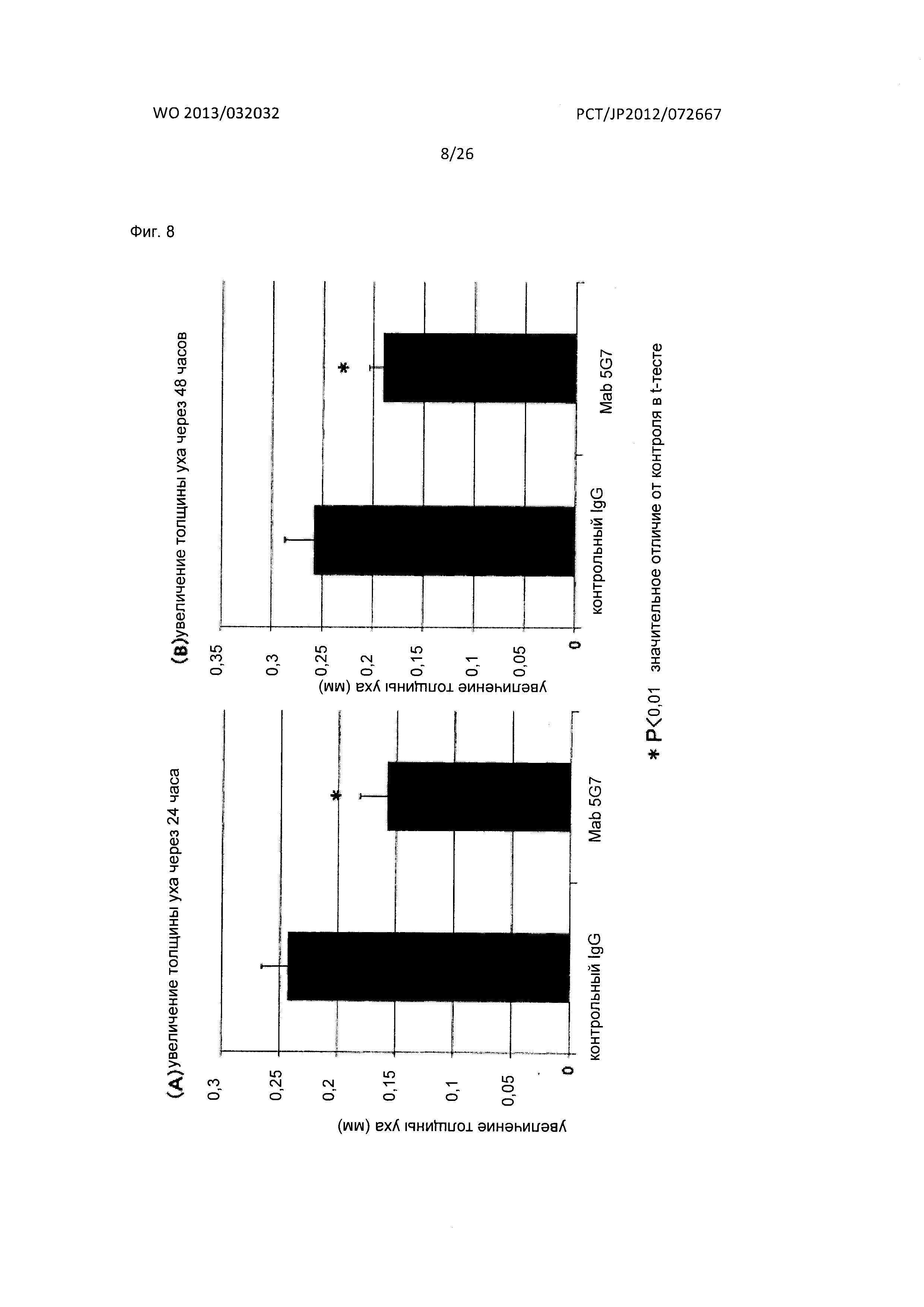

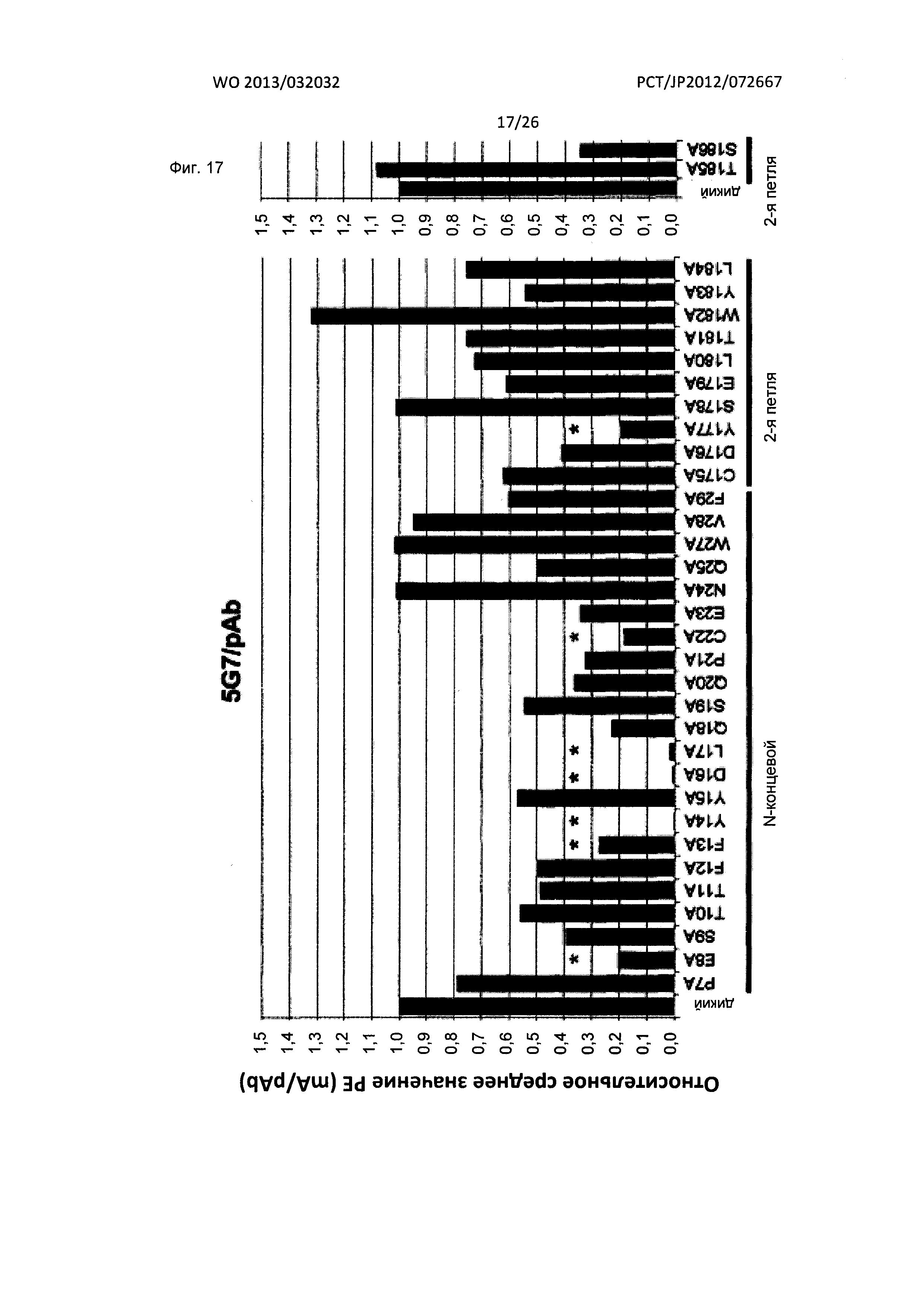
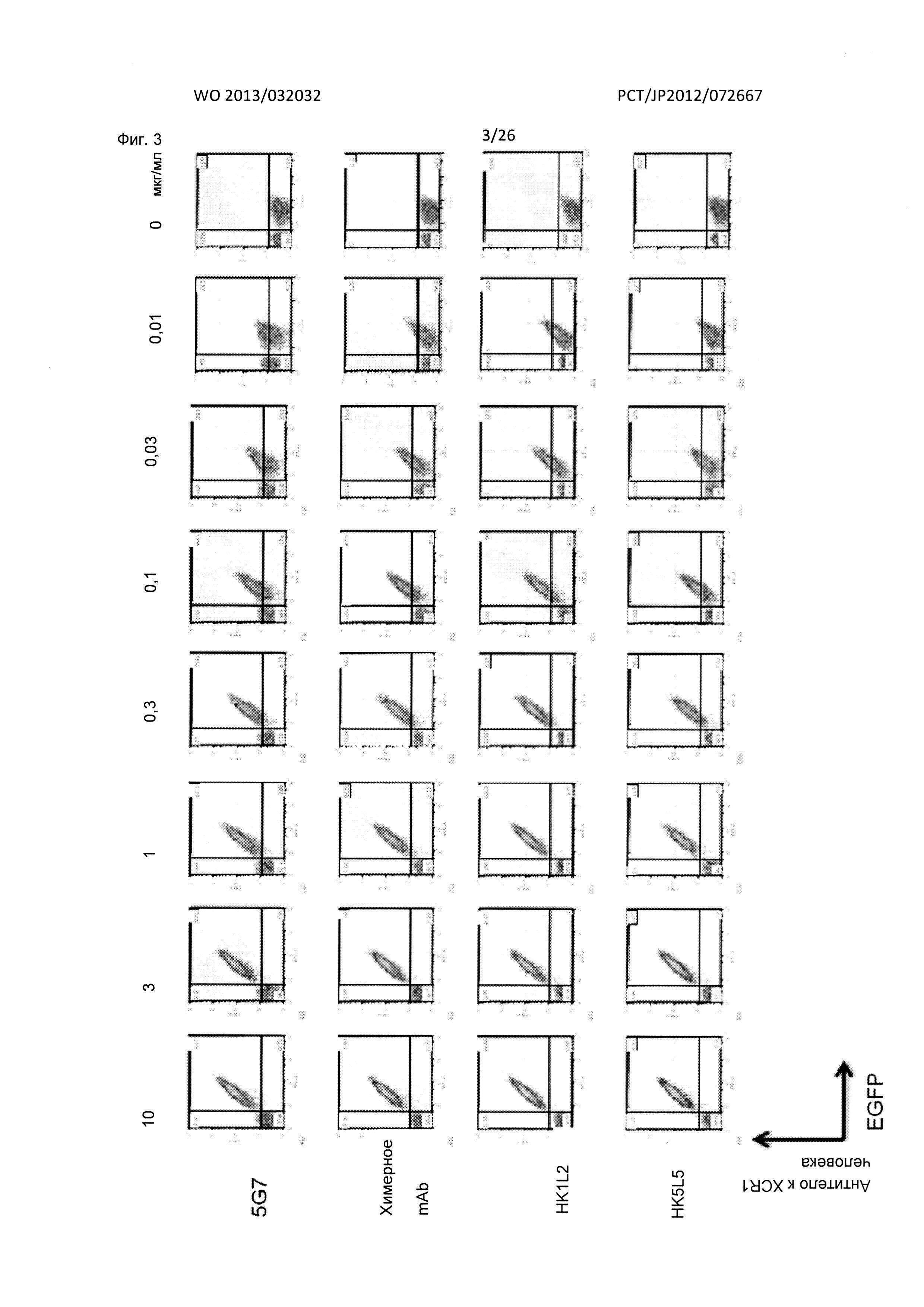




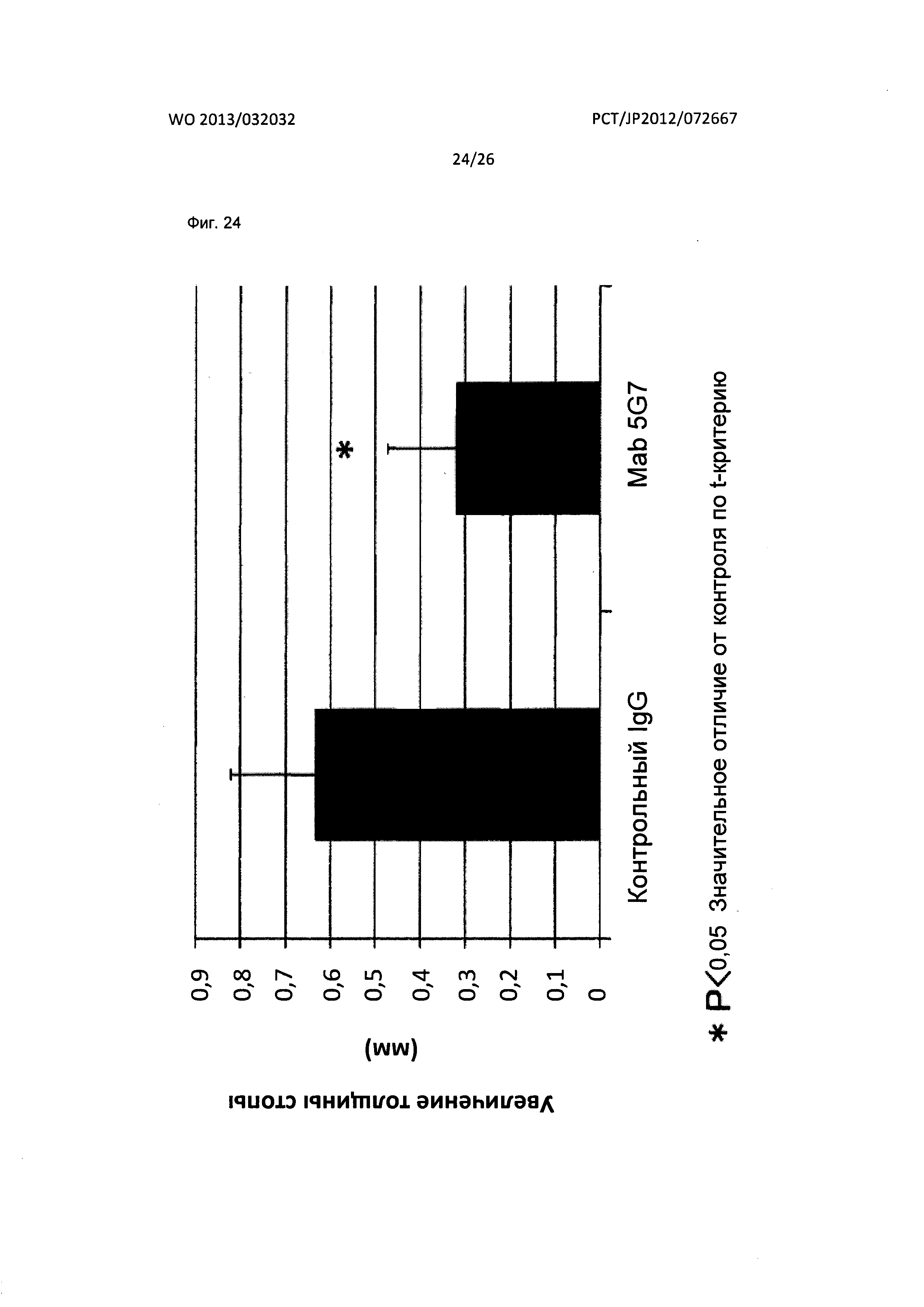

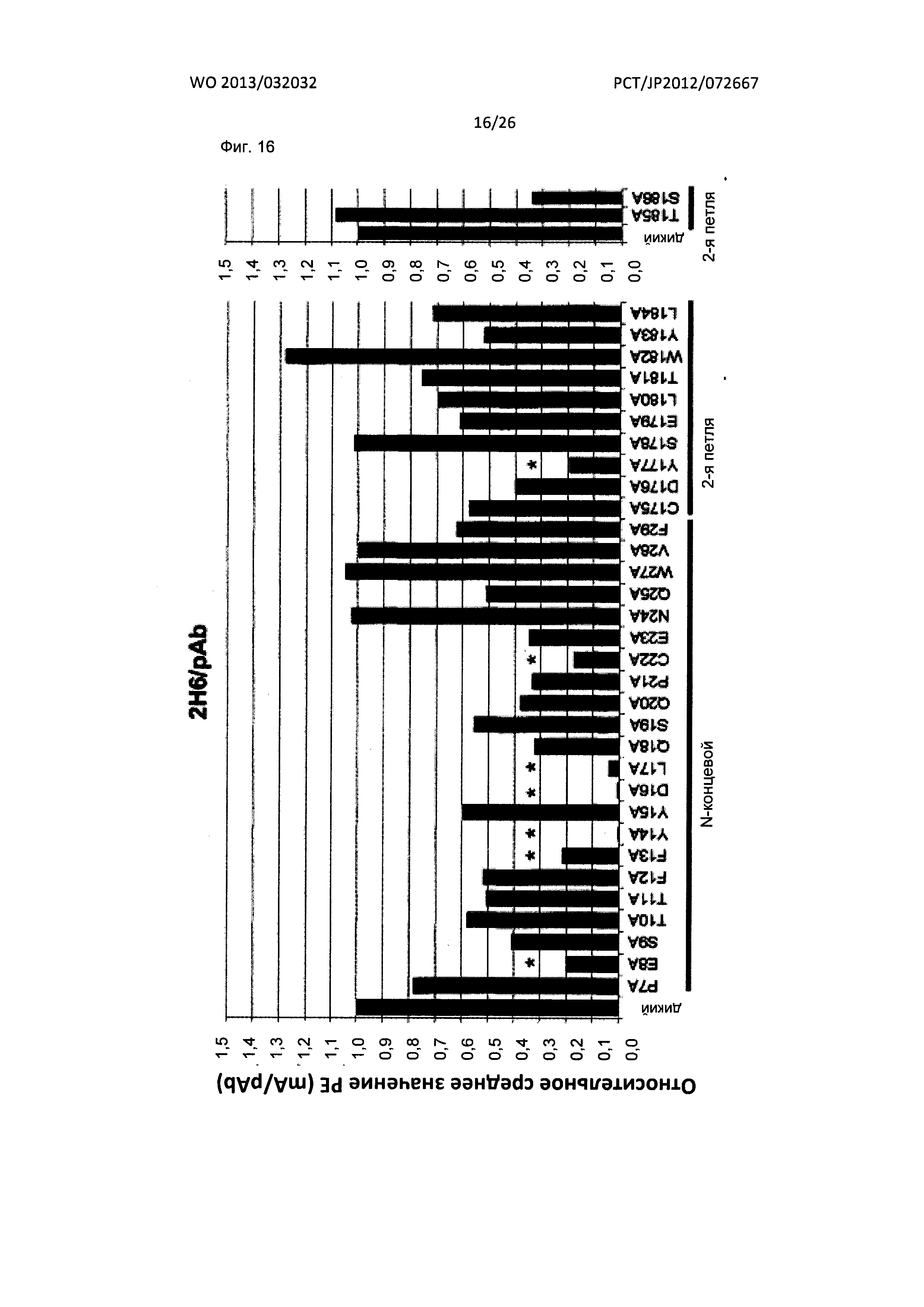

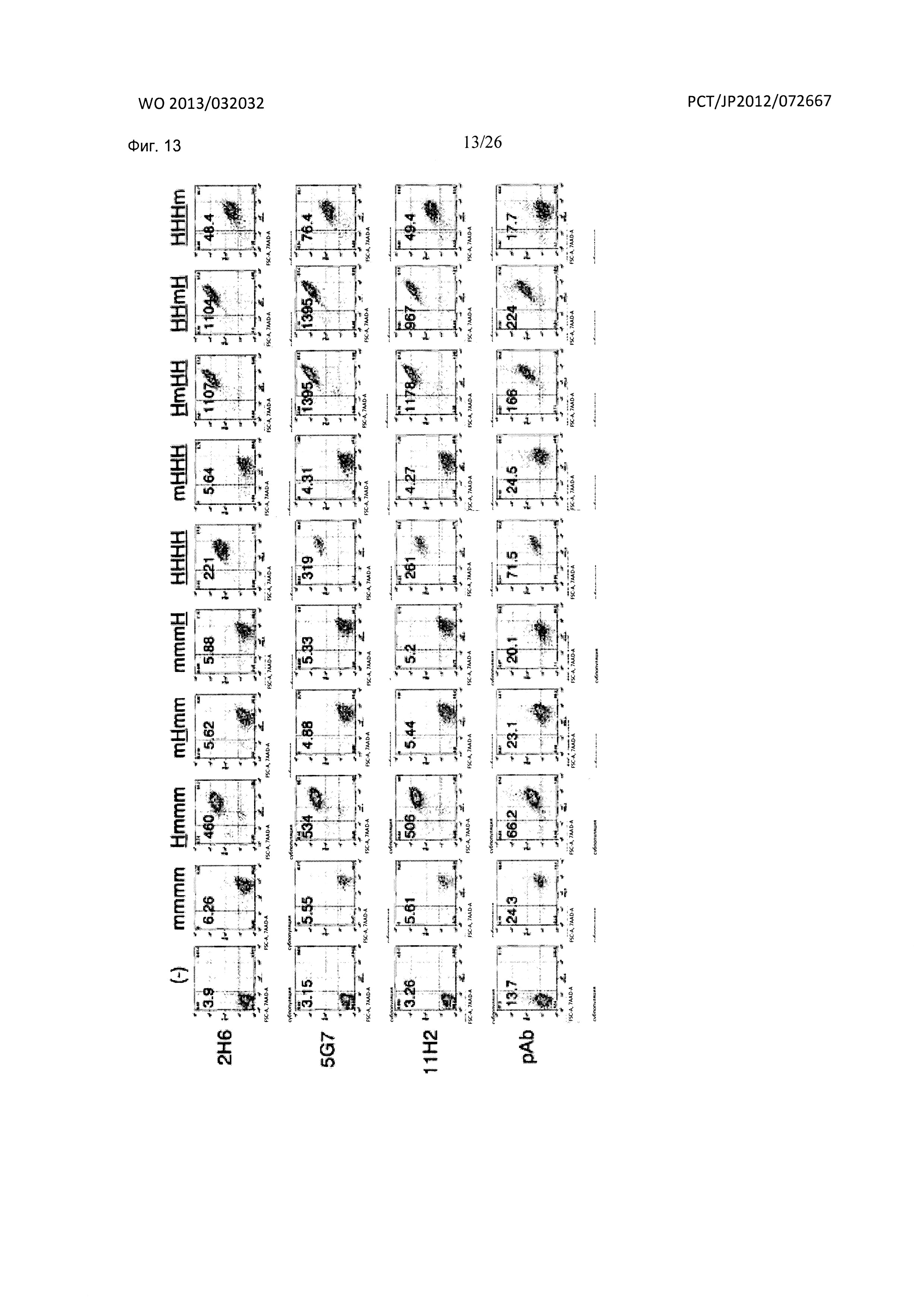

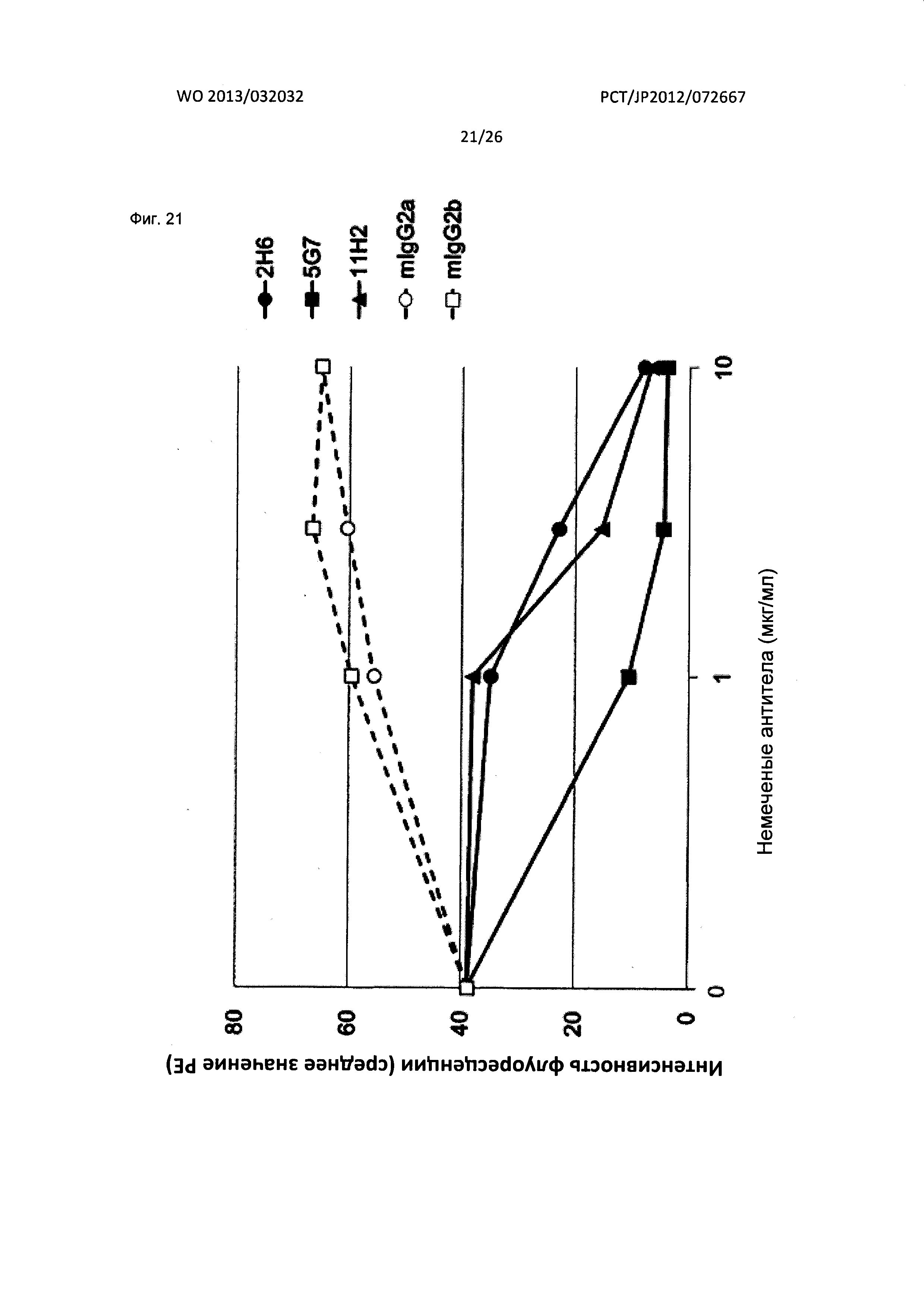








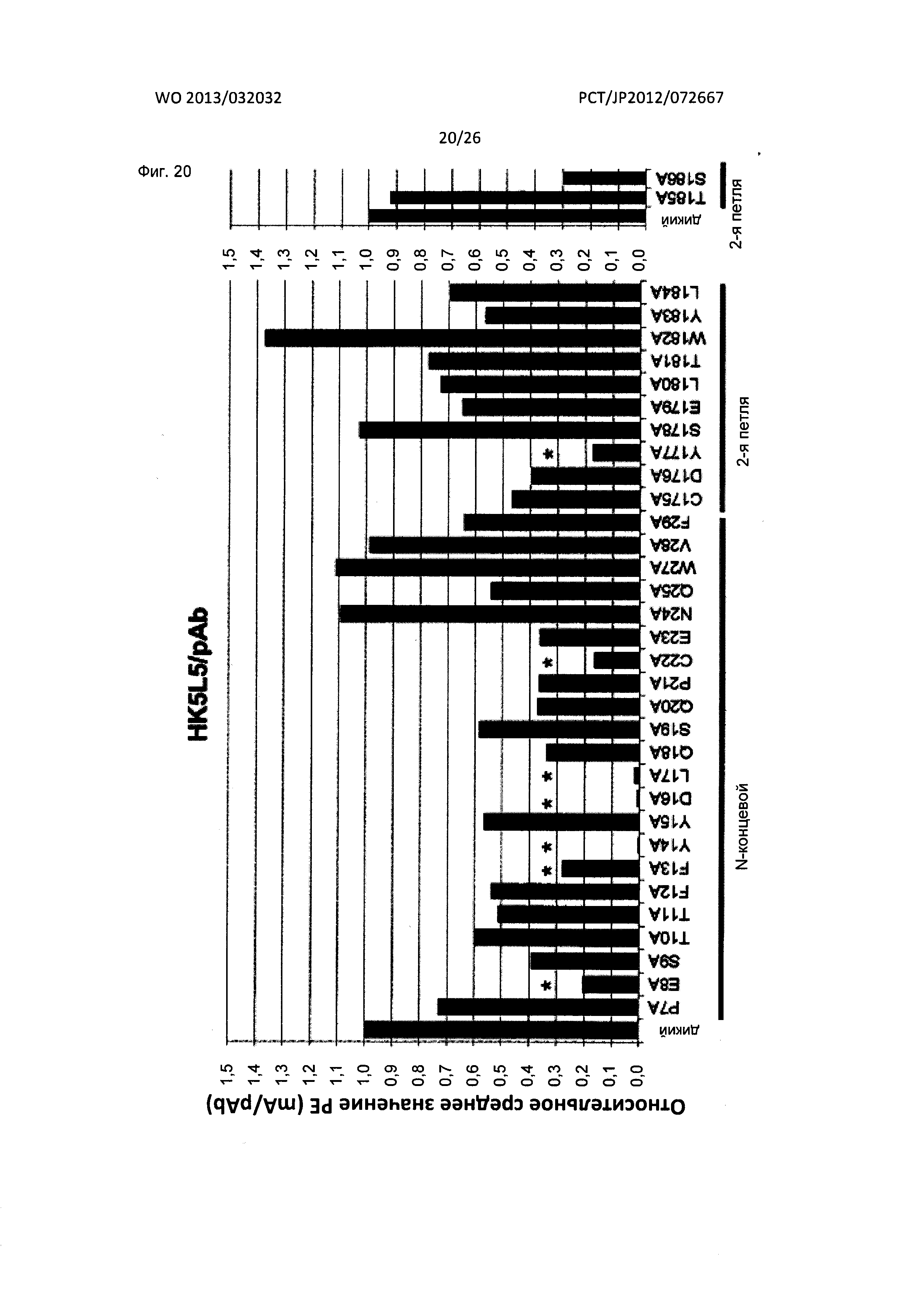
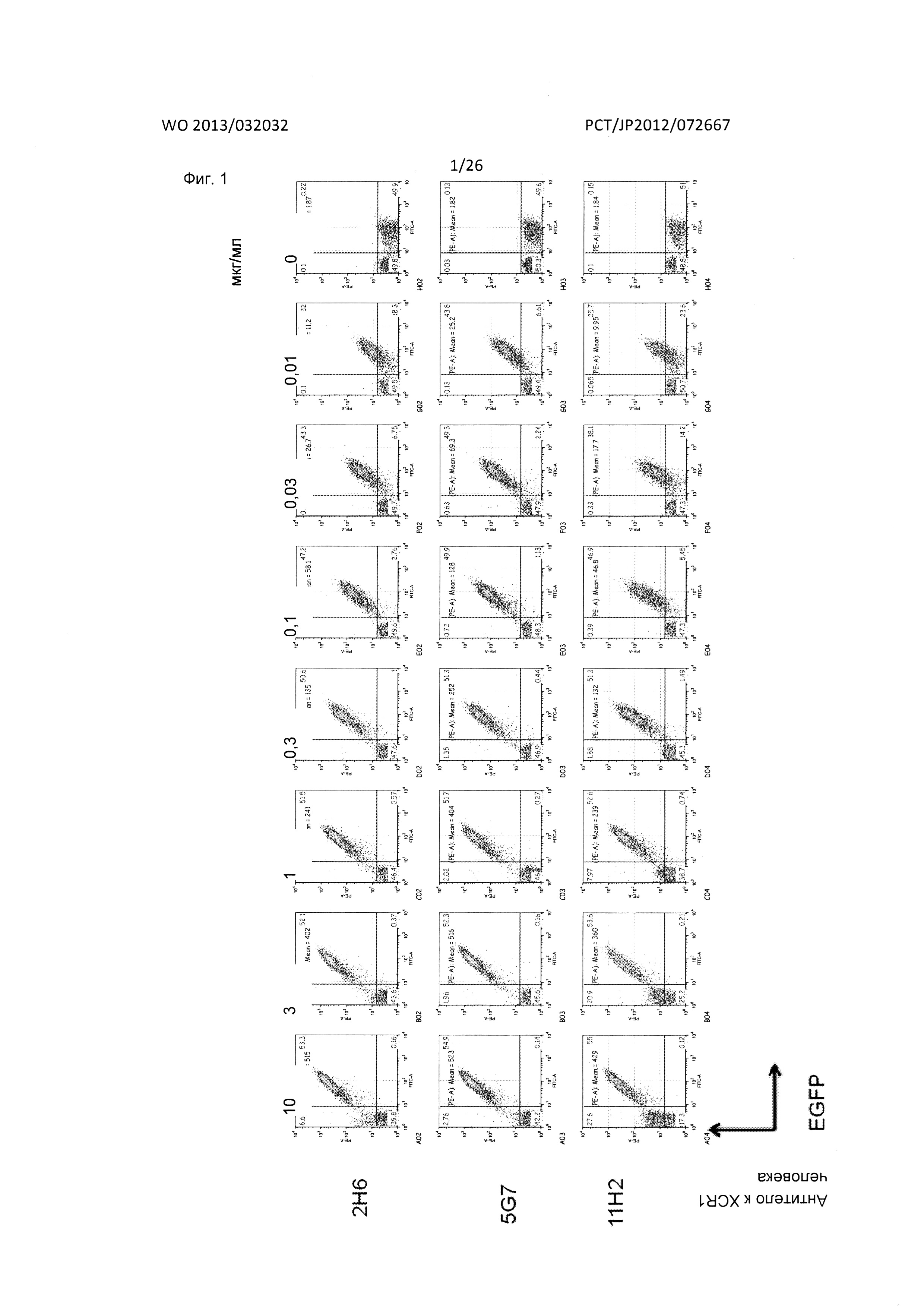
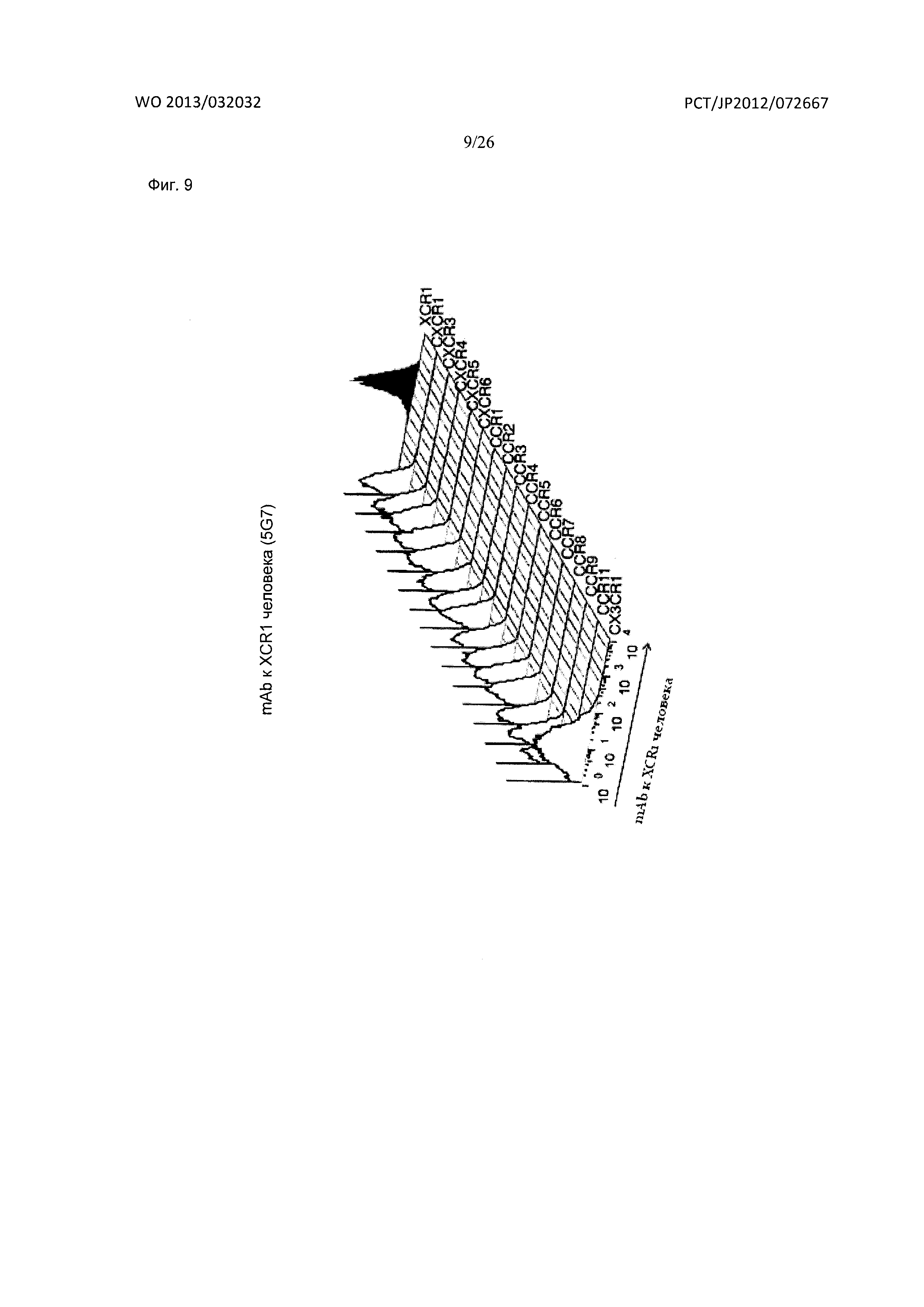
Липосомальная композиция
Конденсированное производное аминодигидротиазина
Производные пиридина, замещенные гетероциклическим кольцом и фосфоноксиметильной группой и содержащие их противогрибковые средства
Промежуточные соединения и способы синтеза аналогов галихондрина в
Реагенты и способы для бета-кетоамидного синтеза синтетического предшественника иммунологического адъюванта е6020
Новое конденсированное производное аминодигидротиазина
Азотсодержащие конденсированные гетероциклические соединения и их применение в качестве ингибиторов продукции бета-амилоида
Аналоги галихондрина в
Противоопухолевое средство, задействующее соединения с ингибирующим эффектом к киназам в комбинации
Биомаркер для болезни альцгеймера или умеренного когнитивного расстройства
Способ получения замещенных гетероциклом производных пиридина
Липосомальная композиция
Конденсированное производное аминодигидротиазина
Производные пиридина, замещенные гетероциклическим кольцом и фосфоноксиметильной группой и содержащие их противогрибковые средства
Промежуточные соединения и способы синтеза аналогов галихондрина в
Реагенты и способы для бета-кетоамидного синтеза синтетического предшественника иммунологического адъюванта е6020
Новое конденсированное производное аминодигидротиазина
Азотсодержащие конденсированные гетероциклические соединения и их применение в качестве ингибиторов продукции бета-амилоида
Аналоги галихондрина в
Противоопухолевое средство, задействующее соединения с ингибирующим эффектом к киназам в комбинации

