Результат интеллектуальной деятельности: ШТАММЫ AGROBACTERIUM, МОДИФИЦИРОВАННЫЕ ДЛЯ УВЕЛИЧЕНИЯ ЧАСТОТЫ ТРАНСФОРМАЦИИ РАСТЕНИЙ
Вид РИД
Изобретение
По настоящей заявке испрашивается приоритет предварительной заявки США № 61/368965, поданной 29 июля 2010 г. Раскрытие предшествующей заявки рассматривается как часть раскрытия данной заявки.
Известный уровень техники
Трансформация растений обычно охватывает методы, требуемые и используемые для введения в растительные клетки способного экспрессироваться в растениях чужеродного гена, так что могут быть получены способные к оплодотворению потомки растений, которые стабильно поддерживают и экспрессируют чужеродный ген. Были трансформированы многочисленные представители, классифицированные как однодольные и двудольные растения. Трансгенные сельскохозяйственные культуры, а также фрукты и овощи представляют коммерческий интерес. Такие культуры включают, но не ограничиваются этим, маис, рис, сою, канолу, подсолнечник, люцерну, сорго, пшеницу, хлопок, арахис, томаты, картофель и тому подобное. Несмотря на развитие систем трансформации растений для введения в растительные клетки способного экспрессироваться в растениях чужеродного гена, желательны дополнительные усовершенствования, которые позволят повысить эффективность трансформации и обеспечат существенные преимущества для преодоления рабочих недостатков при трансформации растений чужеродными генами.
Известно несколько способов введения чужеродного генетического материала в растительные клетки и получения растений, которые стабильно поддерживают и экспрессируют введенный ген. Такие способы включают доставку генетического материала, нанесенного на микрочастицы, с ускорением прямо в клетки (например, в патенте США № 4945050 и патенте США № 5141131). Другой способ трансформации включает технологию с применением карбида кремния или технологию WHISKERS™. См., например, патент США № 5302523 и патент США № 5464765. Метод электропорации также использовался для трансформации растений. См., например, патент WO 87/06614, патент США № 5472869, патент США № 5384253, патенты WO 92/09696 и патент WO 93/21335. Кроме того, может быть использовано слияние протопластов растений с липосомами, содержащими ДНК, предназначенную для доставки, прямое введение ДНК, а также другие возможные способы.
После интеграции ДНК в растительный геном, она обычно становится относительно стабильной на протяжении последующих поколений. Трансформированные клетки растут внутри растений обычным образом. Они могут образовывать половые клетки и передавать характерную(-ые) особенность(-и) трансформации потомкам растений. Такие растения могут быть выращены нормальным образом и могут скрещиваться с растениями, которые имеют те же самые трансформированные наследуемые факторы или другие наследуемые факторы. Полученные в результате гибридные индивидуумы имеют соответствующие фенотипические свойства, например, способность контролировать питание насекомых-вредителей растений.
Для введения ДНК в растительные клетки-хозяева может быть также использован ряд альтернативных методов. Эти методы включают, но не ограничиваются этим, трансформацию с помощью Т-ДНК, доставляемой Agrobacterium tumefaciens или Agrobacterium rhizogenes в качестве агентов трансформации. Растения могут быть трансформированы с использованием метода с применением Agrobacterium, как описано, например, в патенте США № 5177010, патенте США № 5104310, европейской патентной заявке № 0131624 B1, европейской патентной заявке № 120516, европейской патентной заявке № 159418 B1, европейской патентной заявке № 176112, патенте США № 5149645, патенте США № 5469976, патенте США № 5464763, патенте США № 4940838, патенте США № 4693976, европейской патентной заявке № 116718, европейской патентной заявке № 290799, европейской патентной заявке № 320500, европейской патентной заявке № 604662, европейской патентной заявке № 627752, европейской патентной заявке № 0267159, европейской патентной заявке № 0292435, патенте США № 5231019, патенте США № 5463174, патенте США № 4762785, патенте США № 5004863 и патенте США № 5159135. Использование содержащих Т-ДНК векторов для трансформации растительных клеток интенсивно исследовалось и адекватно описано в европейской патентной заявке 120516; An et al., (1985, EMBO J. 4:277-284), Fraley et al., (1986, Crit. Rev. Plant Sci. 4:1-46) и Lee and Gelvin (2008, Plant Physiol. 146: 325-332), и хорошо обосновано в данной области техники.
Известна биология переноса Т-ДНК из Agrobacterium в растительные клетки. См., например, Gelvin (2003) Microbiol. Molec. Biol. Rev. 67:16-37; и Gelvin (2009) Plant Physiol. 150:1665-1676. Как минимум, по меньшей мере, правый пограничный повтор Т-ДНК, но часто как правый пограничный повтор, так и левый пограничный повтор плазмиды Ti или Ri должны быть соединены в виде фланкирующей области генов, которые желательно ввести в растительную клетку. Левый и правый пограничные повторы Т-ДНК являются ключевыми цис-действующими последовательностями, требуемыми для переноса Т-ДНК. В полном геноме Agrobacterium закодированы различные транс-действующие компоненты. Главными среди них являются белки, кодируемые генами vir, которые обычно выявляются в виде серий оперонов в плазмидах Ti или Ri. Различные плазмиды Ti и Ri в некоторой степени отличаются по набору генов vir, например, virF присутствует не всегда. Белки, кодируемые генами vir, выполняют много различных функций, включая, в виде упоминания некоторых характеристик, узнавание и сигнализацию при взаимодействии растительная клетка/бактерии, индукцию транскрипции гена vir, образование секреторного канала типа IV, узнавание пограничных повторов Т-ДНК, образование T-цепей, перенос T-цепей в растительную клетку, импорт T-цепей в ядро растительной клетки и интеграцию T-цепей в хромосоме клеточного ядра растения. См., например, Tzfira and Citovsky (2006) Curr. Opin. Biotechnol. 17:147-154.
Если для трансформации используют штаммы Agrobacterium, ДНК, предназначенную для введения в растительную клетку, можно клонировать в специальных плазмидах, например, либо в промежуточном (челночном) векторе, либо в бинарном векторе. Промежуточные векторы не способны к независимой репликации в клетках Agrobacterium, но манипуляции с ними и их репликацию можно проводить в распространенных штаммах Escherichia coli для молекулярного клонирования. Такие промежуточные векторы включают последовательности, обычно ограниченные рамками областей пограничных правых и левых повторов Т-ДНК, они могут включать маркерный ген селекции для отбора трансформированных растительных клеток, клонирующий линкер, клонирующий полилинкер или другую последовательность, которая может функционировать в качестве сайта для введения генов, предназначенных для трансформации растительной клетки. Клонирование и управление генами, которые желательно перенести в растения, таким образом, может быть легко осуществлено с помощью стандартных методов в E. coli с использованием в качестве клонирующего вектора челночного вектора. После окончания манипуляций челночный вектор может быть впоследствии введен в штаммы Agrobacterium, предназначенные для трансформации растений, для последующей работы. Промежуточный челночный вектор может быть перенесен в Agrobacterium с помощью вспомогательной плазмиды (путем конъюгации бактерий), путем электропорации, с помощью опосредованной химически прямой трансформации ДНК или с помощью других известных методов. Челночные векторы могут быть интегрированы в плазмиду Ti или Ri или в их производные с помощью гомологичной рекомбинации, благодаря последовательностям, которые гомологичны у плазмиды Ti или Ri или их производных и у промежуточной плазмиды. Это событие гомологичной рекомбинации (т.е. интеграции плазмиды) тем самым является средством стабильного поддержания измененного челночного вектора в Agrobacterium с точкой начала репликации и другими функциями поддержания плазмиды, обеспечиваемыми частью плазмиды Ti или Ri в коинтегративной плазмиде. Плазмида Ti или Ri также включает области vir, включающие гены vir, необходимые для переноса Т-ДНК. Плазмида, несущая область vir, обычно представляет собой мутировавшую Ti или Ri плазмиду (вспомогательную плазмиду), из которой удалена область Т-ДНК, включая правые и левые пограничные повторы Т-ДНК. Такие плазмиды, происходящие от pTi, имеющие функционирующие гены vir и лишенные всего или по существу всего из T-области и сопровождающих элементов, при описании в настоящем документе обозначаются как вспомогательные плазмиды.
Супербинарная система является специализированным примером челночного вектора/системы гомологичной рекомбинации (рассмотрено в обзорах Komari et al., (2006) в: Methods in Molecular Biology (K. Wang, ed.) No.343: Agrobacterium Protocols (2nd Edition, Vol. 1) HUMANA PRESS Inc., Totowa, NJ, pp.15-41; и Komori et al., (2007) Plant Physiol. 145:1155-1160). Штамм хозяина Agrobacterium tumefaciens, применяемый с супербинарной системой, представляет собой LBA4404(pSB1). Штамм LBA4404(pSB1) несет две независимо реплицирующиеся плазмиды pAL4404 и pSB1. pAL4404 представляет собой вспомогательную плазмиду, происходящую от Ti плазмиды, содержащую интактный набор генов vir (от Ti плазмиды pTiACH5), но которая не имеет области Т-ДНК (и, следовательно, не имеет последовательностей левых и правых пограничных повторов). Плазмида pSB1 снабжает дополнительным частичным набором генов vir, происходящих из pTiBo542; этот частичный набор генов vir включает оперон virB и оперон virC, а также гены virG и virD1. Одним примером челночного вектора, используемого в супербинарной системе, является pSB11, который содержит клонирующий полилинкер, служащий в качестве сайта введения генов, предназначенных для трансформации растительной клетки, к которому примыкают области правых и левых пограничных повторов Т-ДНК. Челночный вектор pSB11 не способен к независимой репликации в Agrobacterium, но стабильно поддерживается в виде коинтегративной плазмиды при интеграции в pSB1 с помощью гомологичной рекомбинации между общими последовательностями, присутствующими в pSB1 и pSB11. Таким образом, полностью модифицированная область Т-ДНК, введенная в LBA4404(pSB1) в модифицированном векторе pSB11, продуктивно действует в нем и переносится в растительные клетки с помощью белков Vir, происходящих от двух различных источников плазмид Ti Agrobacterium (pTiACH5 и pTiBo542). Супербинарная система, как доказано, является особенно пригодной для трансформации видов однодольных растений. См. Hiei et al., (1994) Plant J. (6:271-282 и Ishida et al., (1996) Nat. Biotechnol. 14:745-750.
Кроме генов vir, которые несут плазмиды Ti Agrobacterium, другие, происходящие из хромосом гены, контролирующие вирулентность (обозначаемые как гены chv), известны как контролирующие определенные аспекты взаимодействий клеток Agrobacterium и растительных клеток и таким образом влияющие на суммарную частоту трансформации растений (Pan et al., (1995) Molec. Microbiol. 17:259-269). Некоторые из происходящих из хромосом генов, необходимых для вирулентности и присоединения, сгруппированы вместе в хромосомном локусе на отрезке в 29 тысяч оснований (Matthysse et al., (2000) Biochim. Biophys. Acta 1490:208-212).
Независимо от конкретной применяемой плазмидной системы, клетки Agrobacterium, трансформированные таким образом, используются для трансформации растительных клеток. Растительные экспланты (например, части листа, сегменты стебля, корней, а также протопласты или культивируемые в суспензии клетки) могут быть успешно прокультивированы с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в растительную клетку. Затем может быть осуществлена регенерация целых растений из инфицированного растительного материала с последующим помещением в подходящие условия роста и культуральную среду, которая может содержать антибиотики или гербициды для селекции трансформированных растительных клеток. Полученные таким образом растения можно затем тестировать на присутствие вставленной ДНК.
Эти методы введения чужеродного генетического материала в растения могут быть использованы для введения благоприятных признаков в растения. Например, миллиарды долларов расходуются каждый год для контролирования насекомых-вредителей и еще миллиарды теряются из-за причиненного ими ущерба. Синтетические инсектициды на основе органической химии являются главными средствами, используемыми для контролирования насекомых-вредителей, но в некоторых областях важную роль играют биологические инсектициды, такие как инсектицидные белки, происходящие от Bacillus thuringiensis (Bt). Способность создавать устойчивые к насекомым растения путем введения генов инсектицидных белков Bt играет революционную роль в современном сельском хозяйстве и повышает ценность инсектицидных белков и их генов.
Для создания устойчивых к насекомым трансгенных растений использовано несколько белков Bt, которые успешно разработаны и во многих случаях зарегистрированы и внедрены в промышленность. Они включают Cry1Ab, Cry1Ca, Cry1Fa и Cry3Bb в кукурузе, Cry1Ac и Cry2Ab в хлопке и Cry3A в картофеле.
Коммерческие продукты, экспрессирующие белки Bt, экспрессируют один белок за исключением случаев, когда желателен объединенный спектр из 2 белков (например, объединение Cry1Ab и Cry3Bb в кукурузе для придания устойчивости к чешуекрылым бабочкам вредителям и длинноусым блошкам, соответственно) или когда независимое действие белков делает их полезными в качестве средства для задержки развития устойчивости у восприимчивых популяций насекомых (например, объединение Cry1Ac и Cry2Ab в хлопке для обеспечения управления устойчивостью к гусенице листовертки-почкоеда табака).
Это значит, что некоторые из качеств трансгенных растений, устойчивых к насекомым, которые привели к быстрому и широкому использованию этой технологии, в результате также приводят к тому, что популяции вредителей будут вырабатывать устойчивость к инсектицидным белкам, продуцируемым этими растениями. Предложено несколько стратегий для сохранения пригодности признаков устойчивости к насекомым, созданных на основе Bt, которые включают развертывание белков при высокой дозе в сочетании с укрытием и чередование с различными токсинами или их совместное развертывание (McGaughey et al. 1998, Nature Biotechnol. 16:144-146).
Если проводят селекцию белков Bt для использования в сочетании, необходимо, чтобы они проявляли свой инсектицидный эффект независимо так, чтобы устойчивость, развившаяся к одному белку, не придавала устойчивости ко второму белку (т.е. чтобы не наблюдалось перекрестной устойчивости к белкам). Оценка надежности перекрестной устойчивости обычно осуществляется с использованием популяций видов вредителей, обычно чувствительных к инсектицидному белку, которые были отобраны по устойчивости к инсектицидным белкам. Если, например, популяция вредителей, отобранная по устойчивости к "белку A", чувствительна к "белку B", то можно заключить, что не существует перекрестной устойчивости, и что сочетание белка A и белка B должно быть эффективным в отношении сдерживания устойчивости к белку A в отдельности.
При отсутствии популяций устойчивых насекомых оценки могут быть сделаны на основе других характеристик, предположительно относящихся к механизму действия и потенциалу перекрестной устойчивости. Предположено использование связывания, опосредованного рецепторами, для идентификации инсектицидных белков, вероятно, не проявляющих перекрестной устойчивости (патент США № 6855873). Ключевым моментом предсказания отсутствия суммарной перекрестной устойчивости в этом подходе является то, что инсектицидные белки не конкурируют за рецепторы у чувствительных видов насекомых.
В том случае, когда два токсина Cry из Bt конкурируют за один и тот же рецептор, то затем, если этот рецептор мутирует у этого насекомого так, что один из токсинов не будет больше связываться с этим рецептором и таким образом не будет больше инсектицидным в отношении этого насекомого, это также должно быть аргументом в пользу того, что насекомое будет также устойчивым ко второму токсину (который конкурентно связывается с тем же рецептором). Однако если два токсина связываются с двумя различными рецепторами, это может служить указанием на то, что насекомое не должно быть устойчиво одновременно к этим двум токсинам.
Cry1Fa является пригодным для контроля многих видов чешуекрылых вредителей, включая европейского кукурузного мотылька (ECB; Ostrinia nubilalis (Hubner)) и травяную совку (FAW; Spodoptera frugiperda), и активен против огневки сахарного тростника (SCB; Diatraea saccharalis).
Белок Cry1Fa при продукции в зерновых растениях с индуцированным событием TC1507 является ответственным за лидирующий в отрасли признак устойчивости для контроля FAW. Cry1Fa дополнительно используют в продуктах HERCULEX®, SMARTSTAX™ и WIDESTRIKE™.
Возможность проведения исследований (конкурентного или гомологичного) связывания с рецептором с использованием белка Cry1Fa ограничена, так как наиболее общепринятый метод, доступный для мечения белков в методах определения связывания с рецептором, ведет к инактивации инсектицидной активности белка Cry1Fa.
Cry1Ab и Cry1Fa представляют собой инсектицидные белки, в настоящее время используемые (по отдельности) в трансгенной кукурузе для защиты растений от различных насекомых-вредителей. Главным вредителем кукурузы, против которого эти белки предоставляют защиту, является европейский кукурузный мотылек (ECB). Патентная заявка США № 2008/0311096 частично относится к использованию Cry1Ab для контролирования устойчивой к Cry1F популяции ECB.
В данной заявке описываются штаммы Agrobacterium tumefaciens, которые модифицированы для повышения частоты трансформации растений. При использовании этих штаммов предоставляются новые системы трансформации растений для введения в растительные клетки чужеродных генов, способных экспрессироваться в растениях. Кроме того, эти штаммы обеспечивают дополнительными усовершенствованиями, которые позволяют повысить эффективность трансформации и предоставляют существенные преимущества в преодолении рабочих недостатков при трансформации растений чужеродными генами.
Краткое изложение сущности изобретения
В настоящем документе описываются штаммы Agrobacterium, которые несут гены, усиливающие трансформацию, в плазмиде, способной реплицироваться независимо от хромосомы Agrobacterium, плазмиде Ti, и бинарные векторы трансформации растений, а также способы их применения. Штаммы Agrobacterium являются дефектными в отношении функций рекомбинации ДНК; что ведет к нестабильности или реорганизации бинарных векторов трансформации растений, и они несут гены, усиливающие трансформацию, в плазмиде, способной реплицироваться независимо от хромосомы Agrobacterium, плазмиде Ti, и бинарные векторы трансформации растений. Описываются также дополнительные штаммы Agrobacterium, которые несут гены, усиливающие трансформацию, интегрированные в хромосому Agrobacterium, в локусе, который не препятствует или каким-либо другим образом не нарушает нормальный рост и способность клеток Agrobacterium к трансформации растений, и их применение.
В одном варианте осуществления описанных в настоящем документе способов растение трансформируется в результате контактирования клетки растения со штаммом Agrobacterium, имеющим, по меньшей мере, одну вспомогательную плазмиду pTi, включающую 14,8 KpnI фрагмент pSB1, и плазмиду pTi, имеющую, по меньшей мере, одну обезоруженную область Т-ДНК, причем область Т-ДНК включает, по меньшей мере, правую границу Т-ДНК и экзогенную ДНК, прилегающую к границе, где плазмиды имеют различные точки начала репликации относительно друг друга.
В дополнительном варианте осуществления описанных в настоящем документе способов растение трансформируется в результате контактирования клетки растения с бактерией рода Agrobacterium, имеющей 14,8 KpnI фрагмент VirBCDG и плазмиду pTi, имеющую, по меньшей мере, одну обезоруженную область Т-ДНК, где 14,8 KpnI фрагмент VirBCDG интегрирован в нейтральный сайт интеграции хромосомы бактерии.
В дополнительном варианте осуществления описанных в настоящем документе способов штамм Agrobacterium включает, по меньшей мере, одну вспомогательную плазмиду pTi, включающую 14,8 KpnI фрагмент pSB1, и плазмиду pTi, имеющую, по меньшей мере, одну обезоруженную область Т-ДНК, где плазмиды имеют различные точки начала репликации относительно друг друга.
В дополнительном варианте осуществления штамм Agrobacterium с усиливающими трансформацию свойствами включает 14,8 KpnI фрагмент VirBCDG, выделенный из pSB1, и плазмиду pTi, имеющую, по меньшей мере, одну обезоруженную область Т-ДНК.
В другом варианте осуществления геномный локус nilA Agrobacterium tumefaciens включает полинуклеотидную последовательность, которая интегрирована в геномный локус nilA.
В дополнительном варианте осуществления штамм LB4404 Agrobacterium включает 14,8 KpnI фрагмент VirBCDG pSB1 во вспомогательной плазмиде pTi и плазмиду pTi, имеющую, по меньшей мере, одну обезоруженную область Т-ДНК и имеющую экзогенную ДНК, прилегающую, по меньшей мере, к одной границе Т-ДНК Agrobacterium, где плазмиды имеют различные точки начала репликации относительно друг друга.
В дополнительном варианте осуществления штамм LB4404 Agrobacterium включает, по меньшей мере, один ген vir из 14,8 KpnI фрагмента VirBCDG, выделенного из pSB1, интегрированный в нейтральный сайт интеграции хромосомы Agrobacterium.
В дополнительных вариантах осуществления предлагаются растения, которые созданы в соответствии с описанными в настоящем документе способами трансформации.
В еще одном варианте осуществления фертильное трансгенное зерновое растение или его потомок экспрессирует инсектицидные количества белка Cry1Ca, инсектицидного белка Cry1F, инсектицидного белка Cry1Ab1 и толерантные к гербицидам количества белка AAD-1, где белки Cry1Ca, Cry1F, Cry1Ab1 и AAD1 совместно экспрессируются с одного локуса рекомбинантной ДНК, стабильно включенной в геном растения.
Описание фигур
Фиг.1. Показана схема клонирования для конструкции плазмиды pDAB9292.
Фиг.2. Показана карта плазмиды pDOW3719.
Фиг.3. Показана схема клонирования для конструкции плазмиды pDAB9698.
Фиг.4. Показаны карты бинарных векторных плазмид pDAB101513 и pDAB101514.
Фиг.5. Показана карта бинарной векторной плазмиды pDAB101556.
Подробное описание изобретения
Штаммы Agrobacterium отличаются от других по своей способности трансформировать растительные клетки. Онкогенные штаммы Agrobacterium дикого типа известны своей способностью индуцировать галлы (опухолевый избыточный рост) у многих растений-хозяев, особенно у видов двудольных. Эта трансформация нормально растущих растительных клеток в опухолевые клетки с отсутствием их саморегуляции возникает как результат переноса специальных последовательностей ДНК (Т-ДНК), которые кодируют гены, экспрессируемые в растениях, которые кодируют растительные гормоны, из индуцирующей опухоль (Ti) плазмиды в растительные клетки, где они стабильно интегрируются в хромосомы растения. Плазмида Ti из штамма Bo542 (т.е. pTiBo542) примечательна тем, что при размещении в хромосомных средах Agrobacterium она стимулирует индукцию особенно больших интенсивно растущих опухолей у некоторых растений (Hood et al., (1986) J. Bacteriol. 168:1291-1301). Гены, ответственные за этот фенотип "супервирулентности", находятся в pTiBo542 вне областей Т-ДНК. При дальнейшем исследовании было обнаружено, что плазмида, содержащая "15,8" тысяч пар оснований (т.п.о.) фрагмента KpnI, происходящего из pTiBo542, которая содержала полные опероны virG, virB и virC, стимулировала усиленное образование опухоли штаммом A281 по сравнению со штаммами, не имеющими плазмиду (Jin et al., (1987) J. Bacteriol. 169:4417-4425). Ген virG плазмиды pTiBo542, как считается, является ответственным за супервирулентный фенотип штамма Agrobacterium A281. virG из pTiBo542 вызывает повышение экспрессии virB в 1,7 раз по сравнению с virG из pTiA6 из-за различий между двумя генами в промоторных областях, кодирующих последовательностях и 3'-нетранслируемых областях (Chen et al., (1991) Molec. Gen. Genet. 230:302-309). Таким образом, ген virG из pTiBo542 может быть выгодно использован для стимуляции более высокой эффективности переноса Т-ДНК, и в результате этого, более высокой частоты трансформации растений, особенно, когда он присутствует в большом фрагменте KpnI плазмиды pTiBo542, которая несет также опероны virB и virC плазмиды pTiBo542.
Полная, аннотированная последовательность pTiBo542 была послана в GENBANK с номером поступления DQ058764 12 мая 2005 г. Анализ карты рестрикции фрагмента KpnI и аннотации генов показывают, что полный оперон virB (который включает гены virB1, virB2, virB3, virB4, virB5, virB6, virB7, virB8, virB9, virB10 и virB11), ген virG, оперон virC (который включает гены virC1 и virC2) и часть оперона virD, включающую ген virD1, возможно выделить из фрагмента KpnI, включающего 14815 пар оснований (п.о.). Предполагается, что размер "15,8 т.п.о." фрагмента KpnI, на который ссылаются в статье Jin et al. (выше), был определен по подвижности фрагмента в агарозном геле, и что реальный размер указанного фрагмента составляет в действительности 14,8 т.п.о. Специалист в области молекулярной биологии должен понимать, что определение размера таких больших фрагментов ДНК с помощью определения подвижности при электрофорезе в агарозном геле может отличаться от реального размера фрагмента, определенного анализом последовательности ДНК на 1 т.п.о. или более. Для облегчения описания этот фрагмент, происходящий из pTiBo542, в настоящем документе будет обозначаться как фрагмент 14,8 KpnI VirBCDG.
Вариант осуществления способов, описанных в настоящем документе, включает использование для трансформации растения усиливающих трансформацию свойств, кодируемых во фрагменте 14,8 KpnI VirBCDG, выделенном из pSB1, в штаммах Agrobacterium, несущих, по меньшей мере, одну обезоруженную вспомогательную плазмиду Ti, где фрагмент 14,8 KpnI VirBCDG несется плазмидой, имеющей точку начала репликации группы несовместимости, отличной от IncP. Дополнительный вариант осуществления включает штамм Agrobacterium, как описано, для использования в этом способе. Область Т-ДНК, предназначенную для введения в растение с использованием штамма Agrobacterium, может переносить плазмида, имеющая область Т-ДНК, примыкающую, по меньшей мере, к одной границе Т-ДНК Agrobacterium, причем плазмида имеет точку начала репликации группы несовместимости IncP или группы несовместимости, которая совместима с группой несовместимости фрагмента 14,8 KpnI VirBCDG, который переносится плазмидой, имеющей точку начала репликации группы несовместимости, отличной от IncP. Область Т-ДНК этой плазмиды может прилегать к правой и левой границам Т-ДНК Agrobacterium.
Плазмиды приписывают к группам несовместимости (генотипическое обозначение: inc; обозначение группы Inc) на основе последовательностей, содержащихся в плазмиде. Детерминанта inc обычно служит в качестве препятствия для сосуществования у одного и того же хозяина других плазмид той же или родственной группы несовместимости и помогает поддерживать определенное количество копий плазмиды в клетке. См., например, Fernandez-Lopez, et al., (2006) FEMS Microbiol. Rev. 30:942-66; и Adamczyk and Jagura-Burdzy (2003) Acta Biochim. Pol. 50:425-53. Две плазмиды несовместимы, если одна из них в присутствии другой менее стабильна, чем сама по себе. Когда две плазмиды одной и той же группы несовместимости находятся в одной и той же клетке, может происходить конкуренция за клеточные ресурсы. Та плазмида, которая способна реплицироваться быстрее или предоставляет какое-либо другое преимущество, будет представлена среди копий в непропорциональной степени, что допускается системой несовместимости. Неожиданно оказалось, что плазмиды могут быть также несовместимыми, когда обе они обладают одними и теми же функциями своего распределения в дочерних клетках.
Плазмиды обычно попадают только в одну из многих существующих групп несовместимости. Насчитывается более 30 известных групп несовместимости. Плазмиды, принадлежащие к группе несовместимости IncP, всесторонне исследованы, и сконструировано большое количество плазмид, которые происходят из этой группы IncP (Schmidhauser et al., (1988) Biotechnology 10:287-332). Примеры плазмид, содержащих группу несовместимости IncP, включают: pMP90RK, pRK2013, pRK290, pRK404 и pRK415. Эти плазмиды могут поддерживаться в многочисленных бактериальных штаммах, включая E. coli и Agrobacterium tumefaciens. Примеры других групп несовместимости включают, но не ограничиваются этим: IncN, IncW, IncL/M, IncT, IncU, IncW, IncY, IncB/O, IncFII, IncII, IncK, IncCom9, IncFI, IncFII, IncFIII, IncHIl, IncHI2, IncX, IncA/C, IncD, IncFIV, IncFV/FO, IncFVI, IncHl 3, IncHII, Incl2, IncI, IncJ, IncV, IncQ и тому подобное, включая их варианты, например, имеющие существенное сходство последовательности или функциональное отношение. В таблице 1 перечислено несколько общеизвестных групп несовместимости и представлены примеры плазмид, которые представляют эти группы несовместимости (это перечисление групп несовместимости и плазмид предлагается только в качестве примера и не предназначено для ограничения групп несовместимости и плазмид, используемых в штаммах Agrobacterium и описанных в настоящем документе способах).
Другой вариант осуществления описанных в настоящем документе способов включает использование усиливающих трансформацию свойств, кодируемых фрагментом 14,8 KpnI VirBCDG, выделенным из pSB1, в штаммах Agrobacterium, имеющих недостаточность функции RecA и несущих, по меньшей мере, одну обезоруженную вспомогательную плазмиду pTi, где фрагмент 14,8 KpnI VirBCDG несется плазмидой, имеющей точку начала репликации группы несовместимости, отличной от IncP. Дополнительный вариант осуществления включает штамм Agrobacterium, как описано в настоящем документе, для использования в этом способе. Область Т-ДНК, предназначенная для введения в растение с использованием этого штамма Agrobacterium, может переноситься плазмидой, имеющей область Т-ДНК, примыкающую, по меньшей мере, к одной границе Т-ДНК Agrobacterium, причем плазмида имеет точку начала репликации группы несовместимости IncP или группы несовместимости, которая совместима с группой несовместимости фрагмента 14,8 KpnI VirBCDG, переносимого плазмидой, имеющей точку начала репликации группы несовместимости, отличной от IncP.
Еще один вариант осуществления описанных в настоящем документе способов включает использование усиливающих трансформацию свойств, кодируемых фрагментом 14,8 KpnI VirBCDG, выделенным из pSB1, и вставленным, по меньшей мере, в одну обезоруженную вспомогательную плазмиду pTi, где фрагмент 14,8 KpnI VirBCDG интегрирован в нейтральный сайт интеграции, локализованный на хромосоме штамма Agrobacterium, отличного от штамма C58. Дополнительный вариант осуществления включает штамм Agrobacterium, как описано, для использования в этом методе. Область Т-ДНК, предназначенная для введения в растение с использованием этого штамма Agrobacterium, дополнительно включает плазмиду, имеющую область Т-ДНК, примыкающую, по меньшей мере, к одной границе Т-ДНК Agrobacterium.
Хотя известны супербинарные системы, например, см. патенты WO 94/00977 A1, WO 95/06722 A1 и WO 95/16031 Al, и они дополнительно описаны Komari et al., (выше) и Komori et al., (выше), эти системы обладают рядом недостатков. Рабочий недостаток супербинарной системы, который преодолевается штаммами Agrobacterium и описанными в настоящем документе способами, заключается в необходимости создания коинтегративной плазмиды из pSB1 и pSB11 (и их производными), в качестве средства, с помощью которого измененная Т-ДНК, переносимая на производных pSB11, стабильно поддерживается в Agrobacterium. Это событие коинтеграции создает пару больших (приблизительно 2,3 т.п.о.) прямо повторяющихся последовательностей из-за рекомбинации между гомологичными областями pSB1 и pSB11. Как хорошо известно специалистам в области молекулярной биологии, большие повторяющиеся последовательности, такие как эти, являются предпочтительными мишенями для внутримолекулярной рекомбинации, что в конечном итоге приводит к делециям ДНК и другим перестройкам, особенно, когда повторы являются частью структуры плазмиды. В супербинарной системе Agrobacterium такие перестройки могут приводить к частичной перестройке или полной потере области Т-ДНК, введенной производными pSB11, приводя в конечном итоге к незначительному переносу или к отсутствию переноса желаемых чужеродных генов в клетки растения-хозяина.
Дополнительный недостаток описанной выше супербинарной системы, который также преодолевается штаммами Agrobacterium и описанными в настоящем документе способами, заключается в том, что создание коинтегративной плазмиды из производных pSB1 и pSB11 генерирует большую плазмиду (минимум более 43 т.п.о.), имеющую две различных точки начала репликации (ori) ColE1-типа (группа несовместимости pMB1/ColE1), а также третий ori, происходящий от плазмиды RK2 (группа несовместимости IncP). Хотя в нормальных условиях ColE1 ori является нефункционирующим в Agrobacterium, известны геномные мутации, которые позволяют стабильно поддерживать плазмиды, имеющие ColE1 ori, в Agrobacterium (Ruslyakova et al., (1999) Russian J. Genet. 35:327-331). В клетках, имеющих такие мутации, плазмида, такая как коинтегративная плазмида производных pSB1::pSB1l, имеющая 3 функционирующих точки начала репликации, как можно ожидать, будет крайне нестабильной. Таким образом, супербинарная система имеет дефекты, на которые выгодно направлены элементы штаммов Agrobacterium и способы трансформации растений, описанные в настоящем документе.
Структура ДНК чужеродного гена или генов, предназначенных для введения и экспрессии в трансгенных растительных клетках с помощью трансформации, опосредованной Agrobacterium, может иметь сильное влияние на стабильность бинарной векторной плазмиды или челночной векторной плазмиды, несущих эти гены в клетках Escherichia coli и Agrobacterium. Нестабильность частично проявляется, когда чужеродные гены включают компоненты генов, которые применяются множество раз в генных конструктах. Например, не является необычным, что конкретный промотор, способный к экспрессии в растениях, может быть использован для стимуляции экспрессии кодирующих областей различных белков в трансгенном растении. Другие компоненты гена, такие как 3'-нетранслируемые области (3'UTR) (т.е. последовательности терминации транскрипции и последовательности, определяющие добавление полиаденилирования) и даже очень сходные кодирующие области белков могут дуплицироваться или присутствовать во множественных копиях в пределах одной области Т-ДНК. Как указывалось выше, эти элементы повторяющихся последовательностей, которые могут существовать либо в инвертированной, либо в прямой ориентации, являются мишенями для внутримолекулярных рекомбинаций, что может приводить к делециям ДНК и другим перестройкам, особенно когда повторы являются частью структуры плазмиды.
Разработано множество специализированных штаммов E. coli, служащих в качестве хозяев для молекулярного клонирования, что помогает преодолеть такие трудности как нестабильность (например, штаммы STBL2™, STBL3™ и STBL4™, предлагаемые INVITROGEN; Carlsbad, CA). Общей характеристикой всех таких клонирующих штаммов E. coli является присутствие геномной мутации в гене recA. Белок RecA представляет собой многофункциональный фермент, который играет роль в гомологичной рекомбинации, репарации ДНК и индукции бактериального ответа SOS. В процессе гомологичной рекомбинации белок функционирует в качестве ДНК-зависимой АТФазы, стимулируя конъюгацию хромосом, образование гетеродуплекса и обмен цепями между гомологичными ДНК. Таким образом, клетка с недостаточностью функции RecA более предрасположена к сохранению устойчивости гомологичных последовательностей ДНК без перестройки или делеции.
Штаммы Agrobacterium с недостаточностью RecA разработаны с целью содействия в решении проблем нестабильности, наблюдаемых при клонировании больших фрагментов ДНК, содержащих повторяющиеся последовательности (Klapwicj et al., (1979) Molec. Gen. Genet. 173:171-175; Farrand et al., (1989) J. Bacteriol. 171:5314-5321; Lazo et al., (1991) Bio/Technology 9:963-967). Эти штаммы, как доказано, пригодны для содействия стабилизации трансформирующих конструктов высокой молекулярной массы в некоторых случаях (Frary and Hamilton, (2001) Transgenic Res. 10:121-132), но не во всех случаях (Song et al., (2003) Theor. Appl. Genet.107:958-964). Таким образом, могут быть выгодно использованы хромосомные среды Agrobacterium, являющиеся дефектными в отношении recA в разрабатываемых штаммах, которые высокоэффективны в поддержании плазмиды и способности трансформировать растение. В дополнение к использованию хромосомных сред Agrobacterium с недостаточностью recA в разрабатываемых штаммах для применения в способах, описанных в настоящем документе, функционирование recA может быть отключено у существующего или полученного штамма с целью сделать штамм, пригодным для способов, описанных в настоящем документе. См., например, Farrand et al. (выше). Например, может быть создан штамм с функционирующим RecA и любыми желаемыми хромосомными добавками, например, добавлением генов vir, затем функция RecA может быть блокирована.
Могут быть использованы векторы BIBAC, сконструированные для возможности эффективной трансформации большими фрагментами ДНК растительных и нерастительных клеток-хозяев. См., например, патент США № 5733744, патент США № 5977439 и патентную заявку США № 2002/0123100 A1. Одним штаммом Agrobacterium, который может быть использован с векторами BIBAC, является штамм UIA143 с недостаточностью RecA, разработанный Farrand et al., (выше). При усовершенствованиях системы BIBAC для повышения способности созданных штаммов Agrobacterium к трансформации растений использованы подгруппы генов, расположенных во фрагменте 14,8 KpnI VirBCDG, в сочетании с другими генами vir. В частности, был использован ген virG из фрагмента 14,8 KpnI VirBCDG, один или в сочетании с генами virE1 и virE2 из pTiA6 в штамме UIA143 с недостаточностью RecA. См., например, Hamilton et al., (1996) Proc. Natl. Acad. Sci. 93:9975-9979; Hamilton, (1997) Gene 200:107-116; Frary and Hamilton (выше).
Кроме того, подходящий вектор, используемый для трансформации растительной клетки с применением способов, описанных в настоящем документе, может содержать ген маркера селекции, кодирующий белок, который придает трансформированным растительным клеткам устойчивость к антибиотику или гербициду. Индивидуально применяемый ген маркера селекции может позволить соответственно отобрать трансформированные клетки, в то время как рост клеток, которые не содержат вставленной ДНК, может быть подавлен с помощью выбранного соединения. Выбор конкретного(-ых) применяемого(-ых) гена(-ов) маркеров селекции может зависеть от дизайна эксперимента или предпочтения, но может быть использован любой из следующих маркеров селекции, также как любой другой ген, не перечисленный в настоящем описании, который может функционировать в качестве маркера селекции. Примеры маркеров селекции включают, но не ограничиваются этим, гены, которые обеспечивают устойчивость или толерантность к антибиотикам, таким как канамицин, G418, гигромицин, блеомицин и метотрексат, или к гербицидам, таким как фосфинотрицин (биалафос), глифосат, имидазолиноны, производные сульфонилмочевины, триазолопиримидины, хлоросульфурон, бромоксинил и DALAPON.
В дополнение к маркеру селекции может быть также использован репортерный ген. В некоторых случаях репортерный ген может быть использован без маркера селекции. Репортерные гены представляют собой гены, которые обычно не предоставляют преимуществ в росте организма или ткани реципиента. Репортерные гены обычно кодируют белок, который придает фенотипическое изменение или ферментативное свойство. Подходящие репортерные гены включают, но не ограничиваются этим, гены, которые кодируют глюкуронидазу (GUS), люциферазу светлячка или флуоресцентные белки, такие как зеленый флуоресцентный белок и желтый флуоресцентный белок.
В дополнение к многочисленным способам трансформации растений, тип ткани, которая вводится в контакт с чужеродными генами, также может варьироваться. Такая ткань может включать, но не ограничиваться этим, эмбриональную ткань, ткань каллуса типов I и II, гипокотиль и меристему. Почти все растительные ткани могут быть трансформированы в процессе дедифференцировки с использованием подходящих методов в пределах данного уровня техники. Специалист в области трансформации растений должен понимать, что для получения трансформированных растений доступны многочисленные методы и что они могут быть модифицированы и специализированы для приспособления к биологическим различиям между различными видами растений-хозяев.
Независимо от конкретного применяемого метода трансформации, чужеродный ген может быть включен в вектор для переноса генов, адаптированный для экспрессии чужеродного гена в растительной клетке в результате включения в вектор растительного промотора. В дополнение к растительным промоторам для экспрессии чужеродных генов в растительных клетках могут быть эффективно использованы промоторы из разнообразных источников. Например, могут быть использованы промоторы бактериального происхождения, такие как промотор октопинсинтазы, промотор нопалинсинтазы, промотор маннопинсинтазы; промоторы вирусного происхождения, такие как промоторы 35S и 19S вируса мозаики цветной капусты (CaMV), промотор из палочковидного вируса сахарного тростника и тому подобное. Промоторы, происходящие от растений, включают, но не ограничиваются этим, промотор малой субъединицы (ssu) рибулозо-1,6-бифосфат (RUBP) карбоксилазы, промотор бета-конглицинина, промотор фазеолина, промотор ADH (алкогольдегидрогеназы), промоторы белков теплового шока, промотор ADF (фактора деполимеризации актина) и тканеспецифические промоторы. Промоторы могут также содержать определенные элементы энхансерных последовательностей, которые могут улучшать эффективность транскрипции. Типичные энхансеры включают, но не ограничиваются этим, интрон 1 алкогольдегидрогеназы 1 (ADH1) и интрон 6 ADH1. Могут быть использованы конститутивные промоторы. Конститутивные промоторы управляют постоянной экспрессией генов почти во всех типах клеток и почти во все периоды времени (например, промотор актина, промотор убиквитина, промотор CaMV 35S). Тканеспецифические промоторы отвечают за экспрессию генов в конкретных типах клеток или тканей, таких как листья или семена. Примеры других промоторов, которые могут быть использованы, включают промоторы, которые активны в течение определенной стадии развития растения, а также активные в конкретных тканях и органах растений. Примеры таких промоторов включают, но не ограничиваются этим, промоторы, которые специфичны для корней, промоторы, специфичные для пыльцы, специфичные для эмбриональной стадии, специфичные для кукурузных рыльцев, специфичные для хлопкового волокна, специфичные для эндосперма семян и специфичные для флоэмы.
При определенных условиях может быть желательно использование индуцибельного промотора. Индуцибельный промотор отвечает за экспрессию генов в ответ на специфический сигнал, такой как физический стимул (например, промоторы генов белков теплового шока); свет (например, промотор рибулоза-бис-фосфат-1,5-карбоксилазы): гормон (например, глюкокортикоид); антибиотик (например, тетрациклин); метаболиты и стресс (например, засуху). Могут быть также использованы другие желательные элементы транскрипции и трансляции, которые функционируют в растениях, такие как, например, 5'-нетранслируемые лидирующие последовательности, последовательности терминации транскрипции РНК и последовательности добавления сигнала полиаденилирования. Может быть использован любой подходящий, специфичный для растений трансферный вектор для гена, известный в данной области техники.
Трансгенные зерновые культуры, содержащие признаки устойчивости к насекомым (IR), преобладают в зерновых и хлопковых растениях по всей северной Америке, и использование этих признаков расширяется по всему миру. Коммерческие трансгенные зерновые культуры, объединяющие признаки IR и устойчивости к гербицидам (HT), созданы многими посевными компаниями. Они включают сочетания признаков IR, придаваемых инсектицидными белками Bt (Bacillus thuringiensis), и признаков HT, таких как толерантность к ингибиторам ацетолактатсинтазы (ALS), таким как производные сульфонилмочевины, имидазолиноны, триазолопиримидин, сульфонанилиды и тому подобное, ингибиторам глутаминсинтетазы (GS), таким как биалафос, глуфосинат и тому подобное, ингибиторам 4-гидроксифенилпируватдиоксигеназы (HPPD), таким как мезотрион, изоксафлутол и тому подобное, ингибиторам 5-енолпирувилшикимат-3-фосфатсинтазы (EPSPS), таким как глифосат и тому подобное, и ингибиторам ацетил-коэнзим А-карбоксилазы (ACCазы), таким как галоксифоп, хизалофоп, диклофоп и тому подобное. Известны другие примеры, в которых предлагаемые в виде трансгенных белки обеспечивают толерантность растения к химическим классам гербицидов, таким как гербициды на основе феноксикислот и гербициды на основе пиридилоксиацетатов (см. патент WO 2007/053482 A2) или гербициды на основе феноксикислот и гербициды на основе арилоксифеноксипропионатов (см. патент WO 2005107437 A2, A3). Способность контролировать множественные проблемы в отношении вредителей с помощью признаков IR является ценной концепцией коммерческого продукта, и преимущество этой концепции продукта увеличивается, если признаки контроля над насекомыми и признаки контроля над сорняками сочетаются в одном и том же растении. Более того, повышенная ценность может быть достигнута путем сочетаний в одном растении признаков IR, придаваемых инсектицидным белком Bt, с одним или более дополнительными признаками HT, такими как указанные выше признаки, плюс один или более дополнительных вводимых признаков (например, другая устойчивость к насекомым, придаваемая происходящими от Bt или другими инсектицидными белками, устойчивость к насекомым, придаваемая такими механизмами как RNAi и тому подобное, устойчивость к заболеваниям, толерантность к стрессу, улучшенное использование азота и тому подобное), или признаков выхода (например, высокое содержание масел, состав полезных для здоровья масел, улучшение питательных свойств и тому подобное). Такие сочетания могут быть получены либо путем обычной селекции (например, с помощью селекционного набора), либо совместно с новым событием трансформации, включающим одновременную индукцию множественных генов (например, с помощью молекулярного набора). Преимущества включают способность управлять насекомыми-вредителями и улучшенный контроль над сорняками у зерновых культур, который предоставляет вторичные преимущества производителю и/или потребителю. Таким образом, штаммы Agrobacterium и способы, описанные в настоящем документе, могут быть использованы для предоставления трансформированных растений с сочетаниями признаков, которые включают полный агрономический пакет, сочетающий улучшенное качество зерновых культур со способностью гибкого и эффективного по цене контроля над количеством выхода агрономической продукции.
Гены virG различных плазмид pTi были исследованы для понимания их способности повышать частоту трансформации растений. Liu et al., (1992, Plant Molec. Biol. 20:1071-1087) обнаружили, что дополнительные копии генов virG из множественных источников (т.е. из различных плазмид pTi, но включая pTiBo542) повышали транзиторную трансформацию некоторых растений, и величина эффекта зависела от особенности вспомогательной плазмиды pTi, с которой был соединен конкретный ген virG. Мутант гена virG (предположительно из pTiA6), названный virGN54D (мутация с заменой аминокислоты Asn54 на Asp), конститутивно экспрессировался в Agrobacterium (индукция генов virG дикого типа требует кислого pH, высокой концентрации моносахаридов и присутствия фенольных индукторов, таких как ацетосирингон). См. Pazour et al., (1992) J. Bacteriol. 174:4169-4174. VirGN54D плазмиды pTiA6 был эффективен в отношении трансформации маиса, в то время как множественные копии родительского virG дикого типа были неэффективны. См. Hansen et al., (1994) J. Bacteriol. 174:4169-4174. Описана "тройная" система (т.е. система из трех плазмид), в которой копия конститутивного мутанта гена virGN54D из pTi15955 одновременно находилась в плазмиде, происходящей от pBBR1, в штамме Agrobacterium tumefaciens, который содержал обезоруженную вспомогательную pTi плазмиду pAL4404, и бинарный вектор, несущий гены для трансформации растений. См. van der Fits et al., (2000) Plant Molec. Biol. 43:495-502. Конститутивно экспрессирующийся ген virGN54D, как обнаружено, значительно повышал эффективность как транзиторной, так и стабильной трансформации у некоторых видов растений. Плазмиды, содержащие область контроля репликации pBRR1, не могут быть классифицированы как принадлежащие к любой известной группе несовместимости и, следовательно, могут сосуществовать с широким спектром других плазмид у одного хозяина. Более того, у табака, хлопка и риса была специально протестирована способность оказывать влияние на эффективность трансформации растений у различных сочетаний генов vir: мутантного гена virGN54D, происходящего из pTiA6, гена virG из pTiBo542, генов VirE1/E2 из pTiA6, и сочетания последних двух наборов генов. См. Park et al., (2000) Theor. Appl. Genet. 101:1015-1020. Повышение эффективности трансформации наблюдали у некоторых видов растений и при дополнительных копиях генов vir.
В европейской патентной заявке № 2042602 A1 и патентной заявке США № 2010/0132068 A1 описываются космидные бинарные векторы и "бустерные" плазмиды, которые в случае присутствия в клетке Agrobacterium, несущей вспомогательную плазмиду pTi, составляют дополнительные примеры тройных плазмидных систем. Бустерные плазмиды, как раскрывается в этих документах, обладают точкой начала репликации группы несовместимости IncW и включают плазмиду pVGW, имеющую ген virGN54D, и плазмиду pVGW2, которая является производной от pVGW, имеющей модификации для облегчения клонирования и селекции.
Функции, кодируемые генами хромосом в Agrobacterium, классически определяются с помощью двух генетических подходов. Первый, или прямой, генетический метод включает получение молекулярного клона гена, подвергаемого исследованию, с последующим помещением клонированного гена в генетическую среду, в которой может быть оценен фенотип "усиления функции". Второй, или "метод обратной генетики", требует нарушения структуры гена путем вставки или делеции последовательностей в гене или вокруг него в хромосоме с последующим определением того, какие белки или фенотипы удалены в результате потери функции гена. Этот подход использовали для конструирования ранее описанного мутанта штамма C58 с недостаточностью RecA. См. Farrand et al., выше. Специалисты в области генетического конструирования в клетках Agrobacterium должны понимать, что для создания возможности осуществления таких экспериментов по нарушению структуры генов описаны разнообразные векторы и многочисленные методы. Как доказано, метод является особенно пригодным при использовании для идентификации генов, которые не вовлечены в жизнеспособность, рост и способность к трансформации мутантного штамма. Один такой генетический локус в штамме C58 Agrobacterium представляет собой локус pgl/picA. См. Lee et al., (2001) Plant Microbe Interact. 14:577-579; и Lee (2006) In: Methods in Molecular Biology (K. Wang, ed.) № 343: Agrobacterium Protocols (2nd Edition, Vol. 1) HUMANA PRESS Inc., Totowa, NJ. pp.55-66. Клетки, в которых ген virD2 интегрирован в этот хромосомный локус с помощью гомологичной рекомбинации, как обнаружено, имеют фенотип трансформированного растения, идентичный фенотипу, возникающему у штаммов A. tumefaciens, несущих ген virD2, локализованный на реплицирующейся плазмиде. См. Lee et al., выше. Кроме того, область Т-ДНК, интегрированная в локус pgl/picA штамма C58, может быть функционально доставлена в растительную клетку (Oltmanns et al., 2010. Plant Physiol. 152: 1 158-1166). Таким образом, в штамме C58 локус pgl/picA может служить в качестве "нейтрального сайта интеграции" для введения генов в хромосому C58. При применении в настоящем описании "нейтральный сайт интеграции" обозначает ген или хромосомный локус, в природе присутствующий в хромосоме клетки Agrobacterium, чья нормальная функция не требуется для роста клетки или для способности клетки осуществлять все ее функции, необходимые для трансформации растения. При нарушении структуры путем интеграции последовательности ДНК, в норме не присутствующей в гене, клетка, несущая ген с нарушенным нейтральным сайтом интеграции, может продуктивно осуществлять трансформацию растения. В качестве примера Hoekema et al. (1984, EMBO J. 3:2485-2490) продемонстрировали, что функциональная T-область, интегрированная в неохарактеризованный локус в хромосоме C58 посредством транспозиции Tn3, продуктивно переносилась в растительные клетки.
Штаммы Agrobacterium, обсуждаемые в настоящем описании, могут быть выгодно использованы для введения одного или более генов в растение, например, для придания растению индивидуальных или множественных инсектицидных или гербицидных свойств. Например, штаммы Agrobacterium могут быть использованы для введения одного или более, двух или более, трех или более, четырех или более, пяти или более, шести или более генов в растение. Путем использования штаммов Agrobacterium, описанных в настоящем документе, полинуклеотид, содержащий последовательности выбранного гена, вставляют в одно местоположение в растительную клетку при трансформации растительной клетки. В отношении размера областей Т-ДНК, используемых для вставки генов, области Т-ДНК могут равняться или составлять более 15000 пар нуклеотидных оснований или равняться или составлять более 20000 пар нуклеотидных оснований, равняться или составлять более 25000 пар нуклеотидных оснований, равняться или составлять более 26000 пар нуклеотидных оснований, равняться или составлять более 27000 пар нуклеотидных оснований, равняться или составлять более 28000 пар нуклеотидных оснований, равняться или составлять более 29000 пар нуклеотидных оснований или равняться или составлять более 30000 пар нуклеотидных оснований. При использовании штаммов Agrobacterium, описанных в настоящем документе, выбранные последовательности генов могут иметь гомологию последовательностей, равную или составляющую более 60%, равную или составляющую более 65%, равную или составляющую более 67%, равную или составляющую более 69,5%, равную или составляющую более 70%, равную или составляющую более 75%, равную или составляющую более 80%, и сохранять идентичность их транскрибируемых последовательностей. Типы генов, которые могут быть введены, могут кодировать инсектицидные белки, гербицидные белки или смесь инсектицидных белков и гербицидных белков. Конкретные примеры генов, которые могут быть введены, включают гены, кодирующие инсектицидный белок Cry1Ca, инсектицидный белок Cry1F, инсектицидный белок Cry1Ab1, и гербицидный белок AAD1, которые могут быть введены в различных сочетаниях или в виде набора, включающего все четыре гена. С использованием этих штаммов Agrobacterium могут быть трансформированы виды однодольных (с одной семядолей) и двудольных (с двумя семядолями) растений.
В настоящем описании раскрывается также геномный локус nilA Agrobacterium tumefaciens, в который может быть интегрирована полинуклеотидная последовательность. Такая интегрированная полинуклеотидная последовательность может включать любой ген vir или оперон vir или другие пригодные гены. В примерах 17-20 представлена идентификация, характеристика и использование геномного локуса nilA Agrobacterium tumefaciens, а также продукция штаммов Agrobacterium tumefaciens с множеством генов vir, локализованных в хромосоме. Геномный локус nilA или любой локус, который характеризуется 85-100% идентичностью последовательности, может быть идентифицирован в других штаммах Agrobacterium, используемых в описанных в настоящем документе методах идентификации и характеристики, и любые такие идентифицированные локусы nilA могут быть использованы сходным с описанным в настоящем документе образом для интеграции генов vir или других подходящих генов, которые могут, например, повышать эффективность трансформации растений. Методы идентификации и характеристики таких геномных локусов, описанных в настоящем документе, могут также использоваться для идентификации других нейтральных сайтов интеграции в хромосоме Agrobacterium, в которую могут быть интегрированы полинуклеотидные последовательности, содержащие гены vir или другие гены так, чтобы штамм Agrobacterium оставался способным к трансформации растений. Уже известны некоторые хромосомные сайты, которые могут быть использованы в качестве нейтральных сайтов интеграции, например, сайт RecA в штамме с недостаточностью RecA, и локус pgl/picA в штамме C58 Agrobacterium tumefaciens. Однако существует необходимость в идентификации новых нейтральных сайтов в штаммах Agrobacterium tumefaciens в дополнение к C58, так как локус pgl/picA не определяется в некоторых других штаммах, например, в штамме LBA4404 (Oltmanns et al., выше). Дополнительные хромосомные сайты, которые могут быть использованы в качестве нейтральных сайтов интеграции, описаны в патенте США № 6323396. Таким образом, раскрыт также штамм Agrobacterium с геном vir, интегрированным в нейтральный сайт интеграции в хромосоме Agrobacterium. Такой штамм Agrobacterium может использовать геномный локус nilA или другой нейтральный сайт интеграции для интеграции генов vir.
Таким способом могут быть добавлены в хромосому множественные типы пригодных генов, обеспечивая отсутствие необходимости в использовании T-вспомогательных плазмид. Например, могут быть использованы дополнительные гены vir и множественные копии пригодных генов vir из различных штаммов.
В настоящем документе раскрыт также штамм Agrobacterium, содержащий гены vir на вспомогательной плазмиде, имеющей точку начала репликации группы несовместимости, отличной от IncP, и плазмиду, имеющую область Т-ДНК, прилегающую, по меньшей мере, к одной границе Т-ДНК Agrobacterium, причем плазмида имеет точку начала репликации группы несовместимости IncP.
Дополнительно раскрыты растения, созданные с помощью описанных в настоящем документе способов, с использованием штаммов Agrobacterium, описанных в настоящем документе. Такие растения стабильно интегрируют любые области Т-ДНК, введенные с использованием раскрытых в настоящем описании способов. Более того, такие растения экспрессируют любые гены и проявляют любые генетические признаки, придаваемые этими областями Т-ДНК. Кроме того, любой потомок растений, созданных с помощью описанных в настоящем документе способов, с использованием штаммов Agrobacterium, описанных в настоящем документе, стабильно экспрессирует любые гены и проявляет любые генетические признаки, придаваемые теми областями Т-ДНК, которые найдены у родительского растения.
В конкретном варианте осуществления описывается растение, которое стабильно экспрессирует инсектицидные белки Cry1Ca, инсектицидные белки Cry1F, инсектицидные белки Cry1Ab1 и гербицидные белки AAD1. Это растение, например, может представлять собой маис.
Наряду с тем, что в настоящем документе описываются определенные примеры штаммов Agrobacterium, обсуждаемая функциональность может быть перемещена в другие штаммы Agrobacterium с теми же самыми критериями, например, в другие штаммы, которые имеют недостаточность RecA или могут быть созданы с недостаточностью RecA. Примеры других штаммов, которые могут быть использованы с описанными в настоящем документе штаммами и способами, включают, но не ограничиваются этим, штамм C58 Agrobacterium tumefaciens, штамм Chry5 Agrobacterium tumefaciens, штаммы Agrobacterium rhizogenes, штамм EHA101 Agrobacterium tumefaciens, штамм EHA105 Agrobacterium tumefaciens, штамм MOG101 Agrobacterium tumefaciens и штамм T37 Agrobacterium tumefaciens.
Все патенты, патентные заявки, предварительные заявки и публикации, на которые ссылаются в настоящем описании или которые цитируются в нем, включены в качестве ссылки в полном объеме в той степени, в какой они не являются несовместимыми с точными указаниями этого описания.
Далее следуют примеры, которые иллюстрируют методы использования штаммов Agrobacterium и использование описанных в настоящем документе методов на практике. Эти примеры не должны рассматриваться как лимитирующие. Все проценты рассчитаны по массе и все пропорции смесей растворителей - по объему, если не указано иначе. Все температуры даны в градусах Цельсия.
Если специально не указано или не обозначено иначе, термины "a", "an" и "the" при применении в настоящем описании обозначают "по меньшей мере, один".
Пример 1: Конструирование делеционного варианта плазмиды pUCD2
Конструкция плазмиды pUCD2 была описана Close et al., (1984, Plasmid 12:111-118), и полная последовательность 13239 п.о. ДНК раскрыта впервые в настоящем описании в виде DNA SEQ ID NO:1. pUCD2 несет четыре гена, придающие бактериальную устойчивость к антибиотикам: конкретно устойчивость к спектиномицину, канамицину, тетрациклину и ампициллину (фиг.1). Стандартные методы молекулярной биологии, как указывается, например, в Sambrook et al., (1989, Molecular Cloning: A Laboratory Manual (2nd Edition., COLD SPRING HARBOR LABORATORY PRESS, Plainview, N.Y.) и Ausubel et al., (1995, Current Protocols in Molecular Biology, (GREENE PUBLISHING AND WILEY-INTERSCIENCE, New York) и их обновлениях, применяли на этой и других стадиях, описанных в этом примере и в других примерах данного раскрытия. Первая модификация в отношении pUCD2 была сделана с помощью расщепления ДНК pUCD2 ферментами рестрикции Sac I и Sac II и лигирования в основном двухцепочечного фрагмента, имеющего подходящие свешивающиеся "липкие концы", совместимые с созданными под действием Sac I- или Sac II свешиваниями. Этот двухцепочечный олигонуклеотид (фиг.1) был создан с помощью отжига двух комплементарных олигонуклеотидных последовательностей, раскрытых как SEQ ID NO:2 и SEQ ID NO:3. Последовательности SEQ ID NO:2 и SEQ ID NO:3 конструируются для восстановления функционирующего гена устойчивости к канамицину при лигировании ДНК pUCD2, расщепленной Sac I и Sac II. С помощью этой манипуляции создана плазмида pDAB9290 (фиг.1), которая отличается от pUCD2 делецией кодирующей области устойчивости к спектиномицину, удалением сайта узнавания ферментом рестрикции Kpn I из кодирующей области устойчивости к канамицину, и созданием нового сайта Kpn I ниже кодирующей области устойчивости к канамицину.
Для придания отсутствия функциональности генам, кодирующим устойчивость к тетрациклину и устойчивость к ампициллину, проводили дальнейшие манипуляции с ДНК плазмиды pDAB9290 с помощью первоначального расщепления ферментами рестрикции Pst I и Sal I, обработки свешивающихся слева концов этими ферментами с помощью набора QUICK BLUNTING™ (NEW ENGLAND BIOLABS; Ipswich, MA) для создания тупых концов и самолигирования для замыкания в круг образованных таким образом фрагментов. Полученная плазмида (pDAB9291) сохраняет только ген устойчивости к бактериальному антибиотику канамицину и имеет уникальный сайт для расщепления с помощью Kpn I ниже гена устойчивости к канамицину. Последовательность pDAB9291 раскрыта как SEQ ID NO:4. Плазмида pDAB9291 имеет две точки начала репликации, одну (группа несовместимости colE1), происходящую от плазмиды pBR322, и вторую, происходящую от плазмиды pSa (группа несовместимости W). Таким образом, плазмида pDAB9291 способна поддерживать среднее количество копий в E. coli и Agrobacterium.
Пример 2: Клонирование фрагмента 14,8 Kpn I virBCDG в pDAB9291
Фрагмент 14,8 т.п.о. Kpn I, содержащий опероны virG, virB и virC и ген virDl из "супервирулентной" pTiBo542 (фиг.1), был выделен из плазмиды pSB1 (Komari et al., выше; и Komori et al., выше) и клонирован в уникальный сайт Kpn I pDAB9291. Были получены плазмиды, содержащие каждую из двух возможных ориентаций вставленного фрагмента, и названы pDAB9292 и pDAB9293. Одна плазмида pDAB9292 (фиг.1) была выбрана для дальнейшей работы. Последовательность ДНК pDAB9292 раскрыта как SEQ ID NO:5.
Пример 3: Конструирование штамма Agrobacterium с недостаточностью RecA, несущего вспомогательную плазмиду pTiEHA105
Штамм UIA143 Agrobacterium представляет собой штамм с недостаточностью RecA, имеющий генетический фон C58, и он был сконструирован и описан Farrand et al., (выше). Хромосомный ген recA был удален и заменен кассетой генов, придающих устойчивость к эритромицину при 150 мкг/мл. Штамм UIA143 не содержит плазмиды Ti или производного плазмиды Ti.
Штамм EHA105 Agrobacterium сконструирован и описан Hood et al., (1993, Transgenic Research 2:208-221), несет вспомогательную плазмиду (называемую в настоящем описании pTiEHA105), происходящую от "супервирулентной" плазмиды pTiBo542. ДНК плазмиды pTiEHA105 получали из штамма EHA105 и вводили с помощью электропорации в клетки штамма UIA143, приготовленные как электрокомпетентные с помощью стандартных методов (Weigel and Glazebrook, (2002) Arabidopsis: A Laboratory Manual. COLD SPRING HARBOR PRESS, Cold Spring Harbor, NY, 354 pages; Mersereau et al., (1990) Gene 90:149-151; Mattanovich, et al., (1989) Nucl. Acids Res. 17:6747)). Клетки штамма UIA143, трансформированные плазмидой pTiEHA105, отбирали по их способности расти в минимальной среде AB (Watson, et al., (1975) J. Bacteriol. 123:255-264) с использованием очищенного агара и маннопина (2 мг/мл) в качестве единственного источника углерода и азота для роста (Guyon et al., (1980) Proc. Natl. Acad. Sci. 77:2693-2697; Dessaux et al., (1987) Molec. Gen. Genet. 208:301-308).
Присутствие pTiEHA105 подтверждали с помощью полимеразной цепной реакции (ПЦР) с использованием праймеров, созданных для амплификации фрагментов генов virD2 и virG плазмиды pTiBo542, и дополнительно характеризовали, анализируя с помощью блоттинга по Саузерну суммарной ДНК, полученной из кандидатных колоний, зондируя 32P-меченной ДНК плазмиды pTiEHA101, очищенной центрифугированием в градиенте хлорида цезия. Этот штамм Agrobacterium (т.е. UIA143, содержащий pTiEHA105) назван DA2552.
Пример 4: Конструирование штамма Agrobacterium с недостаточностью RecA, несущего вспомогательную плазмиду pTiC58∆
Штамм Z707 произошел путем замены полной области Т-ДНК плазмиды pTiC58 штамма C58 Agrobacterium tumefaciens геном npt I Tn903, который придает устойчивость к канамицину. Полная область vir полученной в результате плазмиды, называемой в настоящем описании pTiC58A, была интактной слева (Hepburn et al., (1985) J. Gen. Microbiol. 131:2961-2969). Вспомогательную плазмиду pTiC58∆ из штамма Z707 очищали центрифугированием в градиенте хлорида цезия и вводили электропорацией в электрокомпетентные клетки UIA143. Трансформант отбирали на основе происходящего из плазмиды pTiC58∆ гена устойчивости к канамицину и происходящего из хромосомы гена устойчивости к эритромицину, и штамм был назван DA2569. Присутствие pTiC58∆ в DA2569 подтверждали ПЦР-амплификацией, с использованием праймеров для определения отобранных областей генов vir и анализируя с помощью блоттинга по Саузерну суммарной ДНК, полученной из кандидатных колоний DA2569, зондируя 32P-меченной ДНК плазмиды pTiC58∆, очищенной центрифугированием в градиенте хлорида цезия из клеток штамма Z707.
Пример 5: Конструирование штамма Agrobacterium с недостаточностью RecA, несущего вспомогательную плазмиду pMP90
Штамм GV3101(pMP90) Agrobacterium tumefaciens несет делеционный вариант pTiC58, называемый pMP90, из которого удалена полная область Т-ДНК и заменена геном, придающим устойчивость к гентамицину (Koncz and Schell, (1986) Mol. Gen. Genet. 204:383-396). ДНК плазмиды pMP90 получают такими методами как центрифугирование в градиенте хлорида цезия или MACHEREY-NAGEL NUCLEOBOND XTRA MAXI KIT "LOW COPY" (MACHEREY-NAGEL Inc.; Bethelem, PA) и вводят электропорацией в клетки UIA143. Трансформант отбирают на основе происходящего из плазмиды pMP90 гена устойчивости к гентамицину (100 мкг/мл) и штамм обозначают DAt20538. Присутствие pMP90 в DAt20538 подтверждают ПЦР-амплификацией, с использованием праймеров для определения отобранных областей генов vir и анализируя с помощью блоттинга по Саузерну суммарной ДНК, полученной из DAt20538.
Пример 6: Конструирование штамма Agrobacterium с недостаточностью RecA, несущего вспомогательную плазмиду pMP90RK
Вспомогательную плазмиду pMP90, описанную в пример 5, дополнительно модифицировали с помощью введения (путем двойного кроссинговера с гомологичной рекомбинацией) 42 т.п.о. фрагмента EcoR I, происходящего из плазмиды pRK2013 (Figurski and Helinski, (1979) Proc. Natl. Acad. Sci. USA 79:1648-1652). 42 т.п.о. фрагмент содержит гены, происходящие из плазмиды RK2, для репликации и мобилизации плазмиды (например, trfA, tra1, tra2, tra3 и oriT), и ген, придающий устойчивость к канамицину. С помощью этой манипуляции заменяли ген устойчивости к гентамицину плазмиды pMP90, и полученную в результате плазмиду обозначили как pMP90RK (Koncz and Schell, выше). ДНК плазмиды pMP90RK получают такими методами как центрифугирование в градиенте хлорида цезия или MACHEREY-NAGEL NUCLEOBOND XTRA MAXI KIT "LOW COPY" и вводят электропорацией в электрокомпетентные клетки UIA143. Трансформант отбирают на основе происходящего из плазмиды pMP90RK гена устойчивости к канамицину, и штамм обозначают как DAt20539. Присутствие pMP90RK в DAt20539 подтверждают ПЦР-амплификацией с использованием праймеров для определения отобранных областей генов vir и анализируя с помощью блоттинга по Саузерну суммарной ДНК, полученной из DAt20539.
Пример 7: Электропорация ДНК pDAB9292 в штамм DA2552 Agrobacterium
Электрокомпетентные клетки DA2552 получали, используя стандартный протокол (см. пример 3). 50 мкл компетентных клеток DA2552 оттаивали на льду и трансформировали с использованием от 300 до 400 нг ДНК плазмиды pDAB9292. Смесь ДНК и клеток подвергали электропорации с использованием предварительно охлажденных кювет для электропорации (0,2 см) и электропоратора BIO-RAD GENE PULSER (BIO-RAD Inc.; Hercules, CA.) при следующих условиях: напряжение: 2,5 кВ, длительность импульса: 5 мсек, выходная емкость: 25 мкФ, сопротивление: 200 Ом. После электропорации к кювете добавляли 1 мл бульона YEP (г/л: экстракт дрожжей 10, пептон 10, NaCl 5), и суспензию клеток в YEP переносили в 15 мл пробирку для культивирования. Клетки инкубировали при 28° при осторожном перемешивании в течение 4 часов, после этого культуру высевали на чашки с YEP + агаром, содержащим канамицин 50 мкг/мл и эритромицин 150 мкг/мл. Чашки инкубировали в течение от 2 до 4 дней при 28°, и колонии собирали и высевали штрихом на свежие чашки YEP + агар с антибиотиками, как указано выше, и инкубировали при 28° в течение от 1 до 3 дней. Принадлежность этих колоний к Agrobacterium подтверждали при использовании теста на кетолактозу (Bouzar et al., (1995) In: Methods in Molecular Biology (K. Gartland and M. Davey, eds.) Agrobacterium Protocols. (Vol. 44) HUMANA PRESS, Totowa, NJ. pp. 9-13. Несколько позитивных в отношении кетолактозы колоний отбирали для исходных 3 мл YEP (с антибиотиками) высеваемых культур, которые росли в течение ночи при 28° при перемешивании. 300 мкл каждой высеянной культуры использовали для инокуляции 200 мл YEP (с антибиотиками) с ростом культуры в течение ночи при 28° при перемешивании при 200 об/мин. Плазмидную ДНК получали из 165 мл каждых 200 мл культуры, выраставшей в течение ночи, с использованием набора MACHEREY-NAGEL NUCLEOBOND® XTRA MAXI PLASMID DNA PURIFICATION. Следовали протоколу производителя за исключением использования 30 мл каждого из буферов RES, LYS и NEU. Элюированную ДНК хранили при 4°.
Гидролиз ферментом рестрикции BamH I плазмидной ДНК использовали для подтверждения присутствия pDAB9292 в этих изолятах, и колонии, имеющие корректные паттерны, затем дополнительно очищали, используя два пассажа изолята одиночной колонии. Плазмидную ДНК получали из культур, инкубируемых в течение ночи, как описано выше, и анализ с помощью гидролиза ферментами рестрикции использовали для подтверждения присутствия интактной pDAB9292. В качестве гидролизованного стандарта включали плазмидную ДНК вектора pDAB9292, исходно использованного для трансформации DA2552. Четыре отдельных реакции гидролиза (Pst I, BamH I, Mfe I и Hind III) проходили с использованием от 750 нг до 1 мкг ДНК. Реакции давали проходить в течение от 1 до 2 часов, и результат анализировали с помощью электрофореза в агарозном геле (0,8% масс./об.), и фрагменты ДНК визуализировали окрашиванием бромидом этидия. Этот штамм Agrobacterium (т.е. DA2552, несущий pDAB9292) назван DAt13192. Этот штамм создает основу для рекомбинантно-дефицитной "тройной" системы трансформации растений.
Пример 8: Электропорация ДНК pDAB9292 в штамм GV3101(pMP90) Agrobacterium
Клетки штамма GV3101(pMP90) Agrobacterium tumefaciens (Koncz and Schell, выше) делали электрокомпетентными с помощью стандартного протокола (см. пример 3). 50 мкл компетентных клеток GV3101(pMP90) оттаивали на льду и трансформировали с использованием от 300 до 400 нг ДНК плазмиды pDAB9292. Смесь ДНК и клеток подвергали электропорации с использованием предварительно охлажденных кювет для электропорации (0,2 см) и электропоратора BIO-RAD GENE PULSER при следующих условиях: напряжение: 2,5 кВ, длительность импульса: 5 мсек, выходная емкость: 25 мкФ, сопротивление: 200 Ом. После электропорации к кювете добавляли 1 мл бульона YEP и суспензию клеток в YEP переносили в 15 мл пробирку для культивирования. Клетки инкубировали при 28° при осторожном перемешивании в течение 4 часов, после этого культуру высевали на чашки с YEP + агаром, содержащим канамицин 50 мкг/мл и гентамицин 100 мкг/мл. Чашки инкубировали в течение от 2 до 4 дней при 28°, и колонии собирали и высевали штрихом на свежие чашки YEP + агар с антибиотиками, как указано выше, и инкубировали при 28° в течение от 1 до 3 дней. Принадлежность этих колоний к Agrobacterium подтверждали при использовании теста на кетолактозу. Несколько позитивных в отношении кетолактозы колоний отбирали для исходных 3 мл YEP (с антибиотиками) высеваемых культур, которые росли в течение ночи при 28° при перемешивании. 300 мкл каждой высеянной культуры использовали для инокуляции 200 мл YEP (с антибиотиками) с ростом культуры в течение ночи при 28° при перемешивании при 200 об/мин. Плазмидную ДНК получали из 165 мл каждых 200 мл культуры, выраставшей в течение ночи, с использованием набора MACHEREY-NAGEL NUCLEOBOND® XTRA MAXI PLASMID DNA PURIFICATION. Следовали протоколу производителя за исключением использования 30 мл каждого из буферов RES, LYS и NEU. Элюированную ДНК хранили при 4°.
Гидролиз ферментом рестрикции BamH I плазмидной ДНК использовали для подтверждения присутствия pDAB9292 в этих изолятах, и колонии, имеющие корректные паттерны, затем дополнительно очищали, используя два пассажа изолята одиночной колонии. Плазмидную ДНК получали из культур, инкубируемых в течение ночи, как описано выше, и анализ с помощью гидролиза ферментами рестрикции использовали для подтверждения присутствия интактной pDAB9292. В качестве гидролизованного стандарта включали плазмидную ДНК вектора pDAB9292, исходно использованного для трансформации GV3101(pMP90). Четыре отдельных реакции гидролиза (Pst I, BamH I, Mfe I и Hind III) проходили с использованием от 750 нг до 1 мкг ДНК. Реакции давали проходить в течение от 1 до 2 часов, и результат анализировали с помощью электрофореза в агарозном геле (0,8% масс./об.), и фрагменты ДНК визуализировали окрашиванием бромидом этидия. Изолят GV3101 A. tumefaciens, несущий вспомогательную плазмиду pMP90 Ti и pDAB9292, назван DAt20712.
Пример 9: Электропорация ДНК pDAB9292 в штамм LBA4404 Agrobacterium
Клетки штамма LBA4404 Agrobacterium tumefaciens (Ooms et al., (1982) Plasmid 7:15-29) делали электрокомпетентными с помощью стандартного протокола (см. пример 3). 50 мкл компетентных клеток LBA4404 оттаивали на льду и трансформировали с использованием от 300 до 400 нг ДНК плазмиды pDAB9292. Смесь ДНК и клеток подвергали электропорации с использованием предварительно охлажденных кювет для электропорации (0,2 см) и электропоратора BIO-RAD GENE PULSER при следующих условиях: напряжение: 2,5 кВ, длительность импульса: 5 мсек, выходная емкость: 25 мкФ, сопротивление: 200 Ом. После электропорации к кювете добавляли 1 мл бульона YEP, и суспензию клеток в YEP переносили в 15 мл пробирку для культивирования. Клетки инкубировали при 28° при осторожном перемешивании в течение 4 часов, после этого культуру высевали на чашки с YEP + агаром, содержащим канамицин 50 мкг/мл и стрептомицин 250 мкг/мл. Чашки инкубировали в течение от 2 до 4 дней при 28°, и колонии собирали и высевали штрихом на свежие чашки YEP + агар с антибиотиками, как указано выше, и инкубировали при 28° в течение от 1 до 3 дней. Принадлежность этих колоний к Agrobacterium подтверждали при использовании теста на кетолактозу и их дополнительно очищали, используя два пассажа изолята одиночной колонии.
Несколько позитивных в отношении кетолактозы колоний отбирали для исходных 3 мл YEP (с антибиотиками) высеваемых культур, которые росли в течение ночи при 28° при перемешивании. 300 мкл каждой высеянной культуры использовали для инокуляции 200 мл YEP (с антибиотиками) с ростом культуры в течение ночи при 28° при перемешивании при 200 об/мин. Плазмидную ДНК получали из 165 мл каждых 200 мл культуры, выраставшей в течение ночи, с использованием набора MACHEREY-NAGEL NUCLEOBOND® XTRA MAXI PLASMID DNA PURIFICATION. Следовали протоколу производителя за исключением использования 30 мл каждого из буферов RES, LYS и NEU. Элюированную ДНК хранили при 4°.
Присутствие интактной плазмиды pDAB9292 подтверждали с помощью гидролиза ферментами рестрикции. В качестве гидролизованного стандарта включали плазмидную ДНК вектора pDAB9292, исходно используемого для трансформации LBA4404. Три отдельных реакции гидролиза (Pst I, BamH I и Hind III) проходили с использованием от 750 нг до 1 мкг ДНК. Реакции давали проходить в течение от 1 до 2 часов, и результат анализировали с помощью электрофореза в агарозном геле (0,8% масс./об.), и фрагменты ДНК визуализировали окрашиванием бромидом этидия. Изолят LBA4404 A. tumefaciens, несущий pDAB9292, назван DAt20711. Этот штамм создает основу для рекомбинантно-профицитной "тройной" системы.
Пример 10: Электропорация ДНК pDAB9292 в штамм DAt20538 Agrobacterium
Электрокомпетентные клетки DAt20538 получают с использованием стандартного протокола (см. пример 3). 50 мкл компетентных клеток DAt20538 размораживают на льду и трансформируют с помощью от 300 до 400 нг ДНК плазмиды pDAB9292. Смесь ДНК и клеток электропорируют, используя предварительно охлажденные кюветы для электропорации (0,2 см) и электропоратор BIO-RAD GENE PULSER, при следующих условиях: напряжение: 2,5 кВ, длительность импульса: 5 мсек, выходная емкость: 25 мкФ, сопротивление: 200 Ом. После электропорации в кювету добавляют 1 мл бульона YEP, и суспензию клеток в YEP переносят в 15 мл пробирку для культивирования. Клетки инкубируют при 28°С при легком перемешивании в течение 4 часов, после чего культуру высевают на YEP + агар, содержащий канамицин в концентрации 50 мкг/мл и гентамицин в концентрации 100 мкг/мл. Чашки инкубируют на протяжении от 2 до 4 дней при 28°С, отбирают колонии и наносят их штрихом на свежие чашки с YEP + агар с антибиотиками, как указано выше, и инкубируют при 28°С на протяжении от 1 до 3 дней. С помощью кетолактозного теста подтверждают, что эти колонии представляют собой Agrobacterium, и позитивные в отношении кетолактозы колонии затем выделяют с помощью двух пассажей изолята одиночной колонии.
Отбирают колонии для инициации культур в 3 мл YEP (с антибиотиками) для инокуляции, которые выращивают в течение ночи при 28°С при встряхивании. 300 мкл каждой из культур для инокуляции используют для инокуляции 200 мл YEP (с антибиотиками) ночной культуры, выращиваемой при 28°С со встряхиванием при 200 об/мин. Плазмидную ДНК получают из 165 мл из каждых 200 мл ночной культуры с помощью набора MACHEREY-NAGEL NUCLEOBOND® XTRA MAXI PLASMID DNA PURIFICATION. Следуют протоколу производителя, за исключением того, что используют 30 мл каждого из буферов RES, LYS и NEU. Элюированную ДНК хранят при 4°С.
Для подтверждения наличия интактной плазмиды pDAB9292 используют рестрикционный анализ. В качестве гидролизованного стандарта включают плазмидную ДНК вектора pDAB9292, исходно использованного для трансформации DAt20538. Проводят четыре отдельные реакции гидролиза, такие как Pst I, BamH I, Mfe I и Hind III, с использованием от 750 нг до 1 мкг ДНК. Реакции дают развиться в течение от 1 до 2 час и анализируют с помощью электрофореза в агарозном геле (0,8% масс./об.), и фрагменты ДНК визуализируют окрашиванием этидия бромидом. Изолят A. tumefaciens DAt20538, несущий pDAB9292, назван DAt20538(pDAB9292).
Пример 11: Конструирование векторов для трансформации растений, содержащих многократно повторяющиеся элементы последовательности, и введение в штаммы Agrobacterium
В настоящем описании иллюстрируется пригодность сконструированного штамма Agrobacterium tumefaciens с недостаточностью функции RecA в сочетании со вспомогательными генами, обеспечиваемыми фрагментом 14,8 KpnI VirBCDG. Бинарный вектор для трансформации растений, pDAB101513 (фиг.4A), был сконструирован в штамме E. coli для клонирования STBL2TM с помощью сочетания стандартных методов клонирования (как описано, например, в Sambrook et al., (1989, выше) и Ausubel et al., (1995, выше)) и технологии GATEWAYTM (INVITROGEN). Бинарный вектор pDAB101513 базируется на точке начала репликации типа IncP плазмиды RK2, и основа вектора содержит бактериальный ген, придающий устойчивость у спектиномицину (SpcR на фигуре 4) в концентрации 100 мкг/мл. Граничные повторы Т-ДНК берут начало от области TL pTi15955. Внутри правого граничного участка (Т-ДНК граничного участка B на фиг.4) и тройного левого граничного участка (Т-ДНК граничного участка A на фиг.4) области Т-ДНК плазмиды pDAB101513 расположены 4 экспрессируемые в растениях, содержащие оптимизированные для растений кодоны, кодирующие белки последовательности (CDS), транскрипция каждой из которых управляется промотором убиквитина 1 маиса из 1991 п.н. с ассоциированным интроном 1 (патент США № 5510474). Три кодирующие области кодируют отдельные белки Bt Cry1 (Cry1Ca, SEQ ID NO:7; Cry1Fa, SEQ ID NO:9; и Cry1Ab, SEQ ID NO: 11), причем каждая из них содержит приблизительно 3500 п.н. Эти кодирующие области были оптимизированы для экспрессии в растениях маиса с использованием таблицы предпочтительных для маиса (Zea mays) кодонов, рассчитанной на основе анализа 706 областей, кодирующих белки маиса, которые были получены из депозитов GENBANK. Дополнительные указания, касающиеся конструирования и получения синтетических генов, можно найти, например, в WO 97/13402 A1, патенте США № 6166302 и патенте США № 5380831. Три области, кодирующие белки B.t, соотносятся друг с другом следующим образом: кодирующая область cry1Ca (SEQ ID NO: 6) и кодирующая область cry1Fa (SEQ ID NO:8) имеют 67% гомологию последовательности; кодирующие области cry1Ca (SEQ ID NO: 6) и cry1Ab (SEQ ID NO: 10) имеют 69,5% гомологию последовательности, а кодирующие области cry1Fa (SEQ ID NO:8) и cry1Ab (SEQ ID NO: 10) имеют 67% гомологию последовательности. Кроме того, C-концевые 1600 п.н. CDS cry1Ca, cry1Fa и cry1Ab имеют 73% гомологию последовательности. Каждая из этих трех кодирующих областей заканчивается Per5 3’-нетранслируемой областью маиса (3’UTR) (патент США № 6384207). Четвертый ген включает кодирующую область aad1 с оптимизированными для растений кодонами (SEQ ID NO: 12), которая кодирует маркерный белок селекции AAD1 (SEQ ID NO: 13) (патент США № 7838733). Кодирующая область aad1 не является родственной CDS cry1Ca, cry1Fa или cry1Ab. Кодирующая область aad1 была сконструирована с использованием таблицы предпочтительных для растений кодонов. Таблица предпочтительных для маиса кодонов была рассчитана на основе анализа 706 последовательностей, кодирующих белки маиса, которые были получены из последовательностей, депонированных в GENBANK. Таблицы используемых кодонов для табака (Nicotiana tabacum, 1268 CDS), канолы (Brassica napus, 530 CDS), хлопка (Gossypium hirsutum, 197 CDS) и сои (Glycine max; ca. 1000 CDS) были загружены с сайта http://www.kazusa.or.jp/codon/. Предпочтительный набор кодонов, который включает часто используемые кодоны, общие для наборов данных для маиса и двудольных, при соответствующем изменении масштаба средних относительных величин рассчитывали после удаления любого дополнительного кодона, используемого реже, чем приблизительно 10% от суммарного использования кодонов для данной аминокислоты в одном из типов растений. Ген aad1 заканчивается 3’UTR липазы маиса (патент США № 7179902). Таким образом, внутри области Т-ДНК pDAB101513, содержащей 22729 п.н., четыре копии промотора ubi1 маиса включают в общей сложности 7964 оснований, организованных в виде четырех прямых повторов из почти 2 т.п.н. (тысяч пар нуклеотидов), причем каждый повтор на 100% сходен с другими. Три копии Per5 3’UTR включают в общей сложности 1095 оснований, организованных в виде трех прямых повторяющихся единиц, причем каждая из них на 100% сходна с другими, а три кодирующие области cry1Ca, cry1Fa и cry1Ab организованы в виде прямых повторов, имеющих гомологию друг с другом от 67% до 73%. В общей сложности T-область pDAB101513 включает приблизительно 86% последовательностей с высокой повторяемостью и для удобства может быть изображена следующим образом:
RB> промотор Ubi1:cry1Ca CDS:Per5 3’UTR>промотор Ubi1: cry1Fa CDS:Per5 3’UTR>промотор Ubi1 cry1Ab CDS:Per5 3’UTR>промотор Ubi1: aad1 CDS:Lip 3’UTR>LB
Природа высокой повторяемости этого конструкта необходима для того, чтобы завершить стадии клонирования в штамме E. coli для клонирования STBL2TM, который был специально сконструирован для поддержания целостности клонов, содержащих такие последовательности ДНК с высокой повторяемостью.
Плазмиду pDAB101513 вводили путем электропорации в электрокомпетентные клетки A. tumefaciens штамма EHA105 (приобретшего устойчивость к стрептомицину в результате спонтанной хромосомной мутации), и устойчивые к спектиномицину/стрептомицину изоляты проверяли с помощью рестрикционного анализа на предмет наличия интактной плазмиды pDAB101513 перед получением замороженных в глицерине партий и их хранением при -80°С. Этот штамм назван EHA105(pDAB101513). Было обнаружено, что многочисленные индивидуальные культуры из клеток, полученных из замороженных в глицерине партий EHA105(pDAB101513), содержат перестроенные версии или версии с делециями плазмиды pDAB101513. Для трансформаций маиса собирали все клетки штамма EHA105(pDAB101513) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их точно, как описано в примере 13.
Плазмида pDAB101513 была успешно введена путем электропорации в электрокомпетентные клетки A. tumefaciens штамма DA2552 (по существу являющегося версией штамма EHA105 с недостаточностью RecA) с получением штамма DA2552(pDAB101513). Трансформанты, отобранные на основе устойчивости к эритромицину и спектиномицину, проверяли с помощью гидролиза плазмидной ДНК ферментами рестрикции перед получением замороженных в глицерине партий и их хранением при -80°С. Было обнаружено, что многочисленные индивидуальные культуры из клеток, полученных из замороженных в глицерине партий, содержат интактную плазмиду pDAB101513. Собирали все клетки штамма DA2552(pDAB101513) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их для трансформаций маиса (пример 13).
Плазмида pDAB101513 была успешно введена путем электропорации в электрокомпетентные клетки A. tumefaciens штамма DAt13192 (штамма DA2552, несущего плазмиду pDAB9292) с получением штамма DAt13192(pDAB101513). Трансформанты, отобранные на основе устойчивости к эритромицину, канамицину и спектиномицину, проверяли с помощью гидролиза плазмидной ДНК ферментами рестрикции перед получением замороженных в глицерине партий и их хранением при -80°С. Было обнаружено, что многочисленные индивидуальные культуры из клеток, полученных из замороженных в глицерине партий, содержат интактную плазмиду pDAB101513. Собирали все клетки штамма DAt13192(pDAB101513) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их для трансформаций маиса (см. пример 13).
Сходным образом было сконструировано производное pSB11 (челночного вектора супербинарной системы), содержащее область Т-ДНК, аналогичную области Т-ДНК pDAB101513. Неоднократные попытки сконструировать супербинарную плазмиду с помощью стандартных методов в LBA4404(pSB1) были неудачными. Все попытки приводили к выделению коинтегративных плазмид на основе pSB1 с большими перестройками и делециями.
Пример 12: Конструирование трансформирующего растения вектора pDAB101514, содержащего множественные повторяющиеся элементы последовательности, и введение в штаммы Agrobacterium
В настоящем описании дополнительно иллюстрируется пригодность сконструированного штамма A. tumefaciens с недостаточностью функции RecA в сочетании со вспомогательными генами vir, обеспечиваемыми фрагментом 14,8 KpnI VirBCDG. Бинарный трансформирующий растения вектор pDAB101514 (фиг.4B) был сконструирован в клонирующем штамме STBL2TM E. coli. Структура бинарного вектора pDAB101514 является почти такой же, что и структура pDAB101513 (см. предшествующий пример), за исключением экспрессионных элементов, используемых для управления экспрессией гена cry1Ca. Транскрипция cry1Ca CDS в pDAB101514 управляется промотором бациллоподобного вируса сахарного тростника из 1429 п.н. (SCBV; Tzafrir et al., (1998) Plant Molec. Biol. 38:347-356). 5’UTR состоит по существу из интрона 6 гена алкогольдегидрогеназы маиса (GENBANK номер регистрации X04049), с прилегающими 20 основаниями экзона 6 и 11 основаниями экзона 7. Транскрипция этого гена терминируется pinII 3’UTR картофеля (An et al., (1989) Plant Cell. 1:115-122). Экспрессионные элементы, используемые для контроля экспрессии генов cry1Fa, cry1Ab и aad1, являются теми же, что и использованные в pDAB101513. Таким образом, в пределах области Т-ДНК pDAB101514, состоящей из 22586 п.н., три копии промотора ubi1 маиса включают в общей сложности 5973 оснований, организованных в виде 3 прямых повторов, каждый из которых состоит из почти 2 т.п.н., причем каждый повтор на 100% сходен с другими. Две копии Per5 3’UTR включают в общей сложности 730 оснований, организованных в виде двух прямых повторяющихся единиц, причем каждая из них на 100% сходна с другой, а три кодирующие области cry1Ca, cry1Fa и cry1Ab организованы в виде прямых повторов, имеющих гомологию последовательности ДНК друг с другом от 67% до 73%. В общей сложности T-область pDAB101514 включает приблизительно 76% последовательностей с высокой повторяемостью, и физическая организация для удобства может быть изображена следующим образом:
RB>промотор SCBV: cry1Ca CDS:pinII 3’UTR>промотор Ubi1: cry1Fa CDS:Per5 3’UTR>промотор Ubi1 cry1Ab CDS:Per5 3’UTR>промотор Ubil:aad1 CDS:Lip 3’UTR>LB
Природа высокой повторяемости этого конструкта необходима для того, чтобы завершить стадии клонирования в штамме E. coli для клонирования STBL2TM, который специально сконструирован для поддержания целостности клонов, содержащих такие последовательности ДНК с высокой повторяемостью.
Плазмиду pDAB101514 вводили путем электропорации в электрокомпетентные клетки A. tumefaciens штамма EHA105 (приобретшего устойчивость к стрептомицину в результате спонтанной хромосомной мутации), и устойчивые к спектиномицину/стрептомицину изоляты проверяли с помощью рестрикционного анализа на предмет наличия интактной плазмиды pDAB101514 перед получением замороженных в глицерине партий и их хранением при -80°С. Этот штамм был назван EHA105(pDAB101514). Было обнаружено, что многочисленные индивидуальные культуры из клеток EHA105(pDAB101514), полученных из замороженных в глицерине партий, содержат перестроенные версии или версии с делециями плазмиды pDAB101514. Для трансформаций маиса собирали все клетки штамма EHA105(pDAB101514) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их с помощью методов, раскрытых в примере 13.
Плазмида pDAB101514 была успешно введена путем электропорации в электрокомпетентные клетки A. tumefaciens штамма DA2552 (по существу являющегося версией штамма EHA105 с недостаточностью RecA) с получением штамма DA2552(pDAB101514). Трансформанты, отобранные на основе устойчивости к эритромицину и спектиномицину, проверяли с помощью гидролиза плазмидной ДНК ферментами рестрикции перед получением замороженных в глицерине партий и их хранением при -80°С. Было обнаружено, что многочисленные индивидуальные культуры из клеток, полученных из замороженных в глицерине партий, содержат интактную плазмиду pDAB101514. Собирали все клетки штамма DA2552(pDAB101514) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их для трансформаций маиса с помощью методов, раскрытых в примере 13.
Плазмида pDAB101514 была успешно введена путем электропорации в клетки A. tumefaciens штамма DAt13192 (штамма DA2552, несущего плазмиду pDAB9292) с получением штамма DAt13192(pDAB101514). Трансформанты, отобранные на основе устойчивости к эритромицину, канамицину и спектиномицину, проверяли с помощью гидролиза плазмидной ДНК ферментами рестрикции перед получением замороженных в глицерине партий и их хранением при -80°С. Было обнаружено, что многочисленные индивидуальные культуры из клеток DAt13192(pDAB101514), полученных из замороженных в глицерине партий, содержат интактную плазмиду pDAB101514. Собирали все клетки штамма DAt13192(pDAB101514) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их для трансформаций маиса с помощью методов, раскрытых в примере 13.
Сходным образом было сконструировано производное pSB11 (челночного вектора супербинарной системы), содержащее область Т-ДНК, аналогичную области Т-ДНК pDAB101514. Неоднократные попытки сконструировать супербинарную плазмиду с помощью стандартных методов в LBA4404(pSB1) были неудачными. Все попытки приводили к выделению коинтегративных плазмид на основе pSB1 с большими перестройками и делециями.
Пример 13: Трансформация маиса штаммами Agrobacterium, несущими бинарные векторы pDAB101513 и pDAB101514
Опосредуемая Agrobacterium трансформация маиса. Семена гибрида Hi-II F1 (Armstrong et al., (1991) Maize Genet. Coop. Newslett. 65:92-93) высаживали в 5-галлоновые (18,9 л) горшки, содержащие смесь из 95% среды METRO-MIX 360 для выращивания без почвы (SUN GRO HORTICULTURE; Bellevue, WA) и 5% суглинистой почвы. Растения выращивали в оранжерее с использованием сочетания натриевых и металл-галоидных ламп высокого давления с режимом освещения свет 16 час:темнота 8 час. Производили контролируемое переопыление сестринских растений с получением незрелых зародышей F2 для трансформации. Колосья маиса собирали через приблизительно 8-10 дней после опыления, когда незрелые зародыши имели размер между 1,0 мм и 2,0 мм.
Инфицирование и совместное культивирование. Колосья маиса очищали, и поверхность стерилизовали с помощью щетки и жидкого мыла, погружения в 20% имеющийся в продаже отбеливатель (содержащий 5% гипохлорит натрия) в течение приблизительно 20 минут и последующей трехкратной промывки стерильной водой. Суспензию клеток A. tumefaciens, несущих pDABl01513 или pDAB101514, бинарные векторы, содержащие три гена, кодирующие белки Bt Cry1Ca, Cry1Fa и Cry1Ab и содержащие маркерный ген селекции растений aad-1, получали перенесением 1 или 2 петель бактерий (выращенных в течение 2-3 дней при 28°С в среде агара с YEP, содержащей соответствующие антибиотики) в 5 мл жидкой среды для инфицирования (LS Basal Medium (Linsmaier and Skoog, (1965) Physiologia Plantarum 18: 100-127) с витаминами N6 (Chu et al., (1975) Scientia Sinica 18:659-668), 1,5 мг/л 2,4-дихлорфеноксиуксусной кислотой (2,4-D), 68,5 г/л сахарозой, 36,0 г/л глюкозой, 6 мМ L-пролином, pH 5,2], содержащей 200 мкМ ацетосирингона. Раствор перемешивали до получения однородной суспензии, и концентрацию доводили до конечной оптической плотности приблизительно 0,4 при 550 нм.
Незрелые зародыши выделяли прямо в микроцентрифужную пробирку, содержащую 2 мл среды для инфицирования. Среду удаляли и заменяли 1 мл раствора Agrobacterium, и раствор Agrobacterium/зародыш инкубировали в течение от 5 до 10 минут при комнатной температуре. После этого зародыши переносили в среду для совместного культивирования (LS Basal Medium, витамины N6, 1,5 мг/л 2,4-D, 30,0 г/л сахарозы, 6 мМ L-пролин, 0,85 мг/л AgNO3, 2,8 г/л GELLAN GUMTM (PHYTOTECHNOLOGY LABORATORIES; Lenexa, KS), pH 5,8), содержащую 200 мкМ ацетосирингона, и культивировали в течение 3-4 дней при 20°С в темноте.
После совместного культивирования зародыши переносили в успокаивающую среду, содержащую соли MS и витамины (Frame et al., 2011 , Genetic Transformation Using Maize Immature Zygotic Embryos. В: Plant Embryo Culture Methods and Protocols: Methods in Molecular Biology. T. A. Thorpe and E. C. Yeung, (Eds), SPRINGER SCIENCE AND BUSINESS MEDIA, LLC. pp 327-341), 6 мМ L-пролина, 100 мг/л мио-инозита, 500 мг/л MES (2-(N-морфолино)этансульфокислоты моногидрата; PHYTOTECHNOLOGIES LABR.), 30 г/л сахарозы, 1,5 мг/л 2,4-D, 0,85 AgNO3, 250 мг/л цефотаксима, 2,8 г/л GELLAN GUMTM, pH 5,8. Через приблизительно 7 дней зародыши переносили в ту же среду с добавкой 100 нМ галоксифопа. Трансформированные изоляты были идентифицированы через приблизительно 8 недель и были объединены путем переноса в свежую селективную среду с 2-недельными интервалами для регенерации и анализа.
Регенерация и получение семян. Для регенерации культуры переносили в среду для индукции "28" (соли MS и витамины, 30 г/л сахарозы, 5 мг/л бензиламинпурина, 0,25 мг/л 2,4-D, 250 мг/л цефотаксима, GELLAN GUMTM, pH 5,7) с добавкой 100 нМ галоксифопа. Инкубация происходила в течение 1 недели в условиях слабого освещения (14 мкэйнштейн·м-2·сек-1) и затем в течение 1 недели в условиях сильного освещения (приблизительно 89 мкэйнштейн·м-2·сек-1). Ткани затем переносили в среду для регенерации "36" (такую же, как среда для индукции, исключая отсутствие регуляторов роста растений). Когда проростки достигали длины от 3 см до 5 см, их переносили в стеклянные пробирки для культивирования, содержащие среду SHGA [(соли и витамины Schenk and Hildebrandt, PHYTOTECHNOLOGIES LABR.), 1,0 г/л мио-инозита, 10 г/л сахарозы и 2,0 г/л GELLAN GUMTM, pH 5,8], для обеспечения дальнейшего роста и развития побегов и корней. Растения переносили на ту же почвенную смесь, что и описанная ранее, и выращивали до цветения в оранжерее. Собирали образцы тканей растения и использовали в биотестах с насекомыми с помощью методов, раскрытых в примере 14, и для молекулярных и биохимических анализов.
Специалисты в области техники трансформации маиса должны понимать, что для трансформации маиса и отбора трансформированных растений пригодны другие методы при использовании других экспрессируемых маркерных генов селекции растений (например, генов устойчивости к гербицидам).
Пример 14: In vitro биотесты образцов листьев против насекомых-вредителей маиса
Анализированными видами чешуекрылых были кукурузная уховертка (CEW; Helicoverpa zea (Boddie)), кукурузный мотылек (ECB; Ostrinia nubilalis (Hubner)) и травяная совка (FAW; Spodoptera frugiperda (J.E. Smith)). Яйца этих насекомых были получены от BENZON RESEARCH (Carlisle, PA).
Биотест первого уровня: Высокопроизводительный 96-луночный биотест. 96-луночные лотки (TPP-US; St. Louis, MO) частично заполняли 2% раствором агара (SIGMA-ALDRICH), и давали агару застыть. С помощью стандартного ручного пробойника для бумаг готовили по три образца дисков из листьев с диаметром 1/8 дюйма (3,2 мм) для каждого из двух видов насекомых (CEW и FAW), тестируемых в этом формате. Один диск листа помещали в одну лунку 96-луночного планшета; использовали три планшета для каждого тестируемого насекомого (один для каждой реплики/диск листа). Для введения яиц насекомого в каждую лунку 96-луночного планшета использовали устройство для высева яиц. Планшеты затем закрывали перфорированными клейкими крышками, а также накрывали пластиковой крышкой, прилагаемой к планшету. Планшеты выдерживали при 30°С, 40% относительной влажности (RH) при режиме освещения 16 часов свет:8 часов темнота в течение трех дней. Классификацию проводили с использованием шкалы 0-1-2, в которой 0 обозначает повреждение <25% диска листа, 1 обозначает повреждение 25-50% диска листа, а 2 обозначает повреждение >50% диска листа в каждой лунке. Показатели повреждения для каждого теста усредняли и использовали в паре с анализом экспрессии белка для проведения корреляционного анализа. Активными против тестируемого вредителя считали растения с показателем повреждения насекомым 0,67 или менее.
Биотест второго уровня: 32-луночный биотест. 32-луночные лотки (C-D INTERNATIONAL; Pitman, NJ) частично заполняли 2% раствором агара, и давали агару застыть. Из каждого растения брали фрагменты листьев размером приблизительно 1 квадратный дюйм (6,45 см2) и помещали по одному в лунки 32-луночных лотков. В каждую лунку помещали один кусочек листа, и тестировали по два кусочка листа на одно растение и на одно насекомое. Насекомые (CEW и FAW) в массовом порядке переносили с помощью кисти, помещая в каждую лунку по 10-20 новорожденных личинок. Лотки закрывали перфорированными клейкими крышками, которые обеспечивали вентиляцию во время тестирования. Лотки выдерживали при 28°С, 40% RH при режиме освещения 16 часов свет:8 часов темнота в течение трех дней. После окончания тестирования для каждого кусочка листа получали показатель простого процента повреждения. Показатели повреждения для каждого теста усредняли и использовали в паре с анализом экспрессии белка для проведения корреляционного анализа. Активными против тестируемого вредителя считали растения со средними оценками повреждения насекомым 25% или менее.
Статистический анализ. Все анализы проводили в JMP 8.0.2 (SAS INSTITUTE Inc., Cary, North Carolina). Для выявления значимых различий между воздействиями и растениями негативного контроля для данных с повреждением насекомым использовали односторонний анализ ANOVA. Для дальнейшей оценки значимых различий между воздействиями также использовали метод Таки-Крамера HSD сравнения средних значений. Кроме того, для анализа корреляции между количеством экспрессированного белка и измерениями активности насекомых использовали линейный регрессионный (метод наименьших квадратов) анализ.
Результаты биотестов суммированы в таблице 2.
Пример 15: Биохимическая и молекулярная характеристика тканей маиса, трансформированных pDAB101513
Были проведены многочисленные эксперименты по трансформации сконструированными штаммами A. tumefaciens EHA105(pDAB101513), DA2552(pDAB101513) и DAt13192(pDAB101513). Количество копий четырех трансгенов в трансгенных растениях T0 было установлено с помощью тестов гидролиза зондов (Bubner and Baldwin, (2004) Plant Cell Rep. 23:263-271) с использованием специфичных для генов олигонуклеотидов. Белковые экстракты из растений, содержащих от 1 до 3 копий ("низкая копийность") генов, дополнительно проверяли на предмет продукции белков Bt Cry1Ca, Cry1Fa и Cry1Ab и белка AAD1 с помощью методов ELISA с использованием имеющихся в продаже наборов антител (ENVIROLOGIXTM, Portland, MA). Были обнаружены растения, которые продуцируют все четыре белка (таблица 3). Кроме того, кусочки листьев растений подвергали биотестированию на предмет активности против трех насекомых-вредителей маиса: кукурузной уховертки (CEW, Helicoverpa zea), травяной совки (FAW, Spodoptera frugiperda) и европейского кукурузного мотылька (ECB, Ostrinia nubilalis) в тестах поедания (пример 14). Были обнаружены растения, которые содержат все четыре трансгена в малом количестве копий, продуцируют все четыре белка и обладают активностью против всех трех насекомых-вредителей (таблица 4). В экспериментах со штаммами EHA105(pDAB101513) или DA2552(pDAB101513) не было получено трансформированных растений, которые не соответствовали этим критериям (таблица 4). Таким образом, особенностью штамма DAt13192, имеющего делецию хромосомного гена recA, дополнительно включающего полный набор генов vir из pTiBo542, содержащийся на pTiEHA105, и также дополнительно включающего частичный набор генов vir pTiBo542, содержащийся во фрагменте 14,8 KpnI VirBCDG вектора pDAB9292, служит то, что он способен эффективно продуцировать трансформированные растения маиса с большими областями Т-ДНК, состоящими из элементов последовательности с высокой повторяемостью.
Пример 16: Биохимическая и молекулярная характеристика тканей маиса, трансформированных pDAB101514
Были проведены многочисленные эксперименты по трансформации сконструированными штаммами A. tumefaciens EHA105(pDAB101514), DA2552(pDAB101514) и DAt13192(pDAB101514). Количество копий четырех трансгенов в трансгенных растениях T0 было установлено с помощью тестов гидролиза зондов (Bubner and Baldwin, выше) с использованием специфичных для генов олигонуклеотидов. Белковые экстракты из растений, содержащих от 1 до 3 копий ("низкая копийность") генов дополнительно проверяли на предмет продукции белков Bt Cry1Ca, Cry1Fa и Cry1Ab и белка AAD1 с помощью методов ELISA с использованием имеющихся в продаже наборов антител (ENVIROLOGIXTM, Portland, MA). Кроме того, кусочки листьев растений подвергали биотестированию на предмет активности против трех насекомых-вредителей маиса в тестах поедания (пример 14). Были обнаружены растения, которые содержат все четыре трансгена в малом количестве копий, продуцируют все четыре белка и обладают активностью против всех трех насекомых-вредителей (таблица 5). В экспериментах со штаммами EHA105(pDAB101514) или DA2552(pDAB101514) не было получено трансформированных растений, которые не соответствовали этим критериям. Таким образом, особенностью штамма DAt13192, имеющего делецию хромосомного гена recA, дополнительно включающего полный набор генов vir из pTiBo542, содержащийся на pTiEHA105, и также дополнительно включающего частичный набор генов vir pTiBo542, содержащийся во фрагменте 14,8 KpnI VirBCDG вектора pDAB9292, служит то, что он способен эффективно продуцировать трансформированные растения маиса с большими областями Т-ДНК, состоящими из элементов последовательности с высокой повторяемостью.
Пример 17: Идентификация и характеристика нейтрального сайта интеграции в хромосоме Agrobacterium tumefaciens LBA4404
Индуцируемый в растении локус picA/pgl хромосомы штамма C58 A. tumefaciens (GENBANK номер регистрации AE0009243) был идентифицирован как несущественный ген, в который могут быть интегрированы фрагменты ДНК (Rong et al., (1990) J. Bacteriol. 172:5828-5836; Rong et al., (1991) J. Bacteriol. 173:5110-5120). О сходном нейтральном сайте интеграции в геноме штамма LBA4404 A. tumefaciens не сообщалось. В настоящем описании заявители описывают идентификацию и секвенирование геномной области LBA4404, которая включает последовательности с частичной гомологией локусу picA/pgl C58. Клетки LBA4404 (INVITROGEN) выращивали в среде YM (г/л: дрожжевой экстракт, 0,4; манит, 10; NaCl, 0,1; MgSO4 7H2O, 0,2; K2HPO4 3H2O, 0,5) при 30°С в течение ночи. Геномную ДНК получали из 1 мл культуры с помощью набора EASY DNA (INVITROGEN) в соответствии с протоколами производителя. Были сконструированы вырожденные праймеры на основе двух областей гомологии между последовательностью белка PicA C58 и гомологами, выявленными у Arabidopsis thaliana, Caldicellulosiruptor saccharolyticus, Alkaliphilus metalliredigenes и Clostridium acetobutylicum. Геномную ДНК LBA4404 использовали в качестве матрицы для полимеразной цепной реакции (ПЦР) с применением HERCULASETM MASTER MIX (STRATAGENE; San Diego, CA) и вырожденных праймеров AtnilA1Fa (5’-GACAGTCCNAATACSGAYGG-3’; SEQ ID NO:14; соответствующего аминокислотам 273-279 белка PicA C58) и AtnilA3R (5’-GTYTTSAGNCGSAGSCCSCGRTCSGT-3’; SEQ ID NO:15, соответствующего комплементарной цепи, кодирующей аминокислоты 364-369 белка PicA C58). Использовали следующие условия термоцикла: 1 цикл 94°С, 2 мин; 25 циклов [94°С, 30 сек; 55°С 30 сек; 72°С, 60 сек]; 1 цикл 72°С, 7 мин. Обозначения вырожденных нуклеотидов в последовательностях праймеров соответствуют нуклеотидам ДНК следующим образом: N = A, C, G или T; Y = T или C; R = A или G; и S = C или G. Был выделен продукт из 285 пар оснований (п.н.), клонирован в вектор pCR2.1-TOPO (INVITROGEN) в клетках Escherichia coli TOP 10 (INVITROGEN), и была определена последовательность ДНК. Было обнаружено, что последовательность гомологична, но не идентична области гена picA C58 (85% идентичность последовательности), и геномная область LBA4404, которую она представляет, обозначается в настоящем описании как фрагмент nilA.
Были сконструированы дополнительные праймеры, комплементарные фрагменту из 285 п.н. nilA LBA4404, для использования в качестве якорей при амплификации с помощью ПЦР геномных фрагментов, примыкающих с обеих сторон к последовательности из 285 п.н. В реакциях ПЦР их использовали в парах с праймерами, сконструированными на основе последовательностей фланкирующих областей гена picA C58. Были определены последовательности амплифицированных фрагментов, берущих начало от последовательности из 285 п.н. и расширенных в обе стороны, примыкающие к фрагменту nilA, и использованы для конструирования других праймеров для последующих реакций ПЦР. При использовании матрицы геномной ДНК LBA4404 с праймерами nilA2F (5’-CCATCCTCATAACACCAGCT-3’; SEQ ID NO:16) и nilA2R (5’-GCAGATCATCGATACGACCA-3’; SEQ ID NO:17), был получен фрагмент ПЦР из приблизительно 2 тысяч оснований (т.п.н.), который был клонирован в pCR®-BLUNT II/XL-TOPO® с использованием набора для клонирования TOPO TA (INVITROGEN) с получением плазмиды pDOW3719 (фиг.2). Анализ последовательности вставленного фрагмента pDOW3719 дал последовательность из 1796 п.н. (SEQ ID NO:18), которая включает наиболее длинную открытую рамку считывания (ORF), кодирующую предполагаемый белок из 531 аминокислот. Более короткая ORF в той же рамке считывания кодирует предполагаемый белок из 523 аминокислот. Предполагаемый белок из 523 аминокислот LBA4404 имеет 88% сходство, 85% идентичность с белком PicA C58. Кодирующие последовательности для предполагаемого белка из 523 аминокислот LBA4404 и белка PicA C58 имеют 81% идентичность. Таким образом, фрагмент nilA LBA4404 представляет собой геномный сегмент, который включает предполагаемый ген, существенно дивергирующий от гена picA C58. В настоящем раскрытии геномная последовательность из 1,8 т.п.н., представленная SEQ ID NO:18, обозначается как локус nilA.
Как ожидается, плазмида pDOW3719, содержащая точку начала репликации colE1, не реплицируется автономно в клетках A. tumefaciens. ДНК плазмиды pDOW3719 использовали для трансформации штамма LBA4404 A. tumefaciens путем электропорации. Отбор по устойчивости к канамицину (носимой pDOW3719) выявил трансформанты с pDOW3719, интегрированной в хромосому LBA4404 посредством рекомбинации, опосредованной областями гомологии из 1,8 т.п.н., имеющимися в хромосоме LBA4404 и pDOW3719. Такое явление интеграции ведет к созданию линейной копии последовательности плазмидного вектора pDOW3719, окруженной с обеих сторон отныне дуплицированной областью гомологии из 1,8 т.п.н. Трансформанты LBA4404 с устойчивостью к канамицину были выделены и подвергнуты скринингу на наличие вставки pDOW3719 с помощью анализа ПЦР. В качестве матрицы использовали препараты геномной ДНК трансформантов с 5 наборами праймеров: i) праймера M13F в паре с праймером M13R, который примыкает к вставке в pDOW3719, ii) праймера M13F в паре с праймером AS4R (включающим основания, комплементарные остаткам с 1041 по 1060 SEQ ID NO: 18), iii) праймера M13F в паре с праймером AS10R (включающим основания, комплементарные остаткам с 1320 по 1337 SEQ ID NO: 18), iv) праймера M13F в паре с праймером AS11R (включающим основания, комплементарные остаткам с 1391 по 1406 SEQ ID NO: 18), и v) праймера M13R в паре с праймером AS9F (включающим основания с 634 по 649 SEQ ID NO: 18). В контрольных реакциях все эти наборы праймеров амплифицировали фрагменты ожидаемого размера при использовании в качестве матрицы ДНК плазмиды pDOW3719. Однако, когда в качестве матрицы использовали геномную ДНК из устойчивого к канамицину трансформанта LBA4404, ПЦР с использованием пары праймеров M13F и M13R не дала амплифицированных продуктов, что указывает на то, что интактная (неинтегрированная) ДНК плазмиды pDOW3719 не очищалась вместе с геномной ДНК. Анализ ПЦР образцов геномной ДНК с четырьмя другими парами праймеров показал образование фрагментов ДНК ожидаемого размера. Эти результаты указывают на то, что устойчивость к канамицину трансформантов LBA4404 обеспечивается ДНК pDOW3719, которая была интегрирована в геном.
Один такой трансформант [LBA4404nilA-int1] был использован для проверки явления, заключающегося в том, что геномная вставка в локус nilA включает способность штамма трансформировать Arabidopsis thaliana. Бинарным вектором pDAB3779, который содержит экспрессируемый в растениях ген, кодирующий белок PAT (который придает устойчивость к гербициду BASTATM), трансформировали клетки штаммов LBA4404nilA-int1 и LBA4404 с отбором по устойчивости к спектиномицину. Эти штаммы затем использовали для проведения экспериментов с трансформацией Arabidopsis с использованием методов Weigel and Glazebrook {выше). Разницы в частоте трансформации, получаемой для двух штаммов, не наблюдалось. Таким образом, особенность вариантов осуществления и методов, описанных в настоящем описании, заключается в том, что вставка фрагмента чужеродной ДНК в хромосомный локус nilA штамма LBA4404 A. tumefaciens, который включает SEQ ID NO: 18, не оказывает влияния на рост растения или способность растения к трансформации таким сконструированным штаммом.
Пример 18: Конструирование суицидного производного pDOW3719 для интегрирования в локус nilA LBA4404
Вставка сайта множественного клонирования в локус nilA, клонированный в pDOW3719, была осуществлена с помощью ПЦР с удлинением посредством сплайсингового перекрытия (SOE) (Horton et al., (1990) BioTechniques 8:528-535). Реакции ПЦР SOE проводили с использованием HERCULASETM master mix в соответствии с протоколами производителя. Часть локуса nilA амплифицировали с использованием ДНК pDOW3719 в качестве матрицы с праймером nilA5’ (5’-CCGGCTCTTCCAGCTCCTCATGCACGAACAACGAGAAACGAGC-3’; SEQ ID NO: 19) в паре с праймером nilA_MCS_SOEF (5’-GAATGGTGAAACCTCTAGATTAATTAAGGATCCCCGGGTACCGAAAAGCCCGACATTGC-3’; SEQ ID NO:20) с получением фрагмента из приблизительно 800 п.н. Вторую часть локуса nilA амплифицировали с использованием ДНК pDOW3719 в качестве матрицы и праймера nilA_MCS_SOEF (5’-GCAATGTCGGGCTTTTCGGTACCCGGGGATCCTTAATTAATCTAGAGGTTTCACCATTC-3’; SEQ ID NO:21) в паре с праймером nilA3’ (5’-GGAATTCTCAGTGGCTTTCATGGGTTTTCTCG-3’; SEQ ID NO:22) с получением фрагмента из приблизительно 900 п.н. Полученные фрагменты затем очищали в геле (NUCLEOSPINTM, CLONTECH; Mountain View, CA) и использовали в качестве матрицы для амплификации с праймерами nilA5’ и nilA3’ с получением фрагмента из 1,6 т.п.н., последовательность которого раскрыта как SEQ ID NO:23 (nilA MCS). Полученный фрагмент гидролизовали с помощью Pvu I и Sap I (NEB) и лигировали с использованием ДНК-лигазы T4 (NEB) с ДНК pBCSK+sacBI (INVITROGEN), гидролизованной теми же самыми ферментами рестрикции. Клетки E. coli TOP10 трансформировали смесью лигирования, и трансформанты отбирали на агаре с соей LB TEKNOVA; Hollister, CA) с добавкой 30 мкг/мл хлорамфеникола. Клоны подвергали скринингу с помощью рестрикционного гидролиза с использованием Pvu I и Kpn I (NEB). Область локуса nilA позитивных клонов была подтверждена секвенированием, и полученная плазмида была названа pDOW3721. Особенность pDOW3721 заключается в том, что к сайту множественного клонирования (MCS), содержащему последовательности, узнаваемые ферментами рестрикции Sph I, Kpn I, Sma I, BamH I, Pac I, Ase I и Xba I, примыкает с одной стороны последовательность LBA4404 из 852 п.н., а с другой стороны последовательность LBA4404 из 745 п.н.
Таким образом, фрагмент чужеродной ДНК может быть клонирован в MCS pDOW3721 и, следовательно, интегрирован в хромосомный локус nilA LBA4404 путем гомологичной рекомбинации, опосредуемой фланкирующими последовательностями из LBA4404. Одиночные события кроссовера, посредством которых вся последовательность плазмиды pDOW3721 интегрируется в хромосому LBA4404, могут быть разделены на двойные события кроссовера с помощью обратной селекции на содержащих сахарозу средах. На таких средах сахароза превращается в токсичный продукт после ферментативного разложения под действием белка SacB, кодируемого геном sacB (Reid and Collmer, (1987) Gene 57:239-246; Quandt et al., (1993) Gene 127:15-21). Таким образом, трансформанты, способные выживать на содержащей сахарозу среде выращивания, должны быть подвергнуты второму событию кроссовера, которое удаляет основу векторной плазмиды pDOW3721 из хромосомы и оставляет разрушенный локус nilA, содержащий интегрированный фрагмент чужеродной ДНК. Во многих сообщениях за последние 10 лет было показано, что перенос последовательностей основы вектора является вполне обычным явлением. Доля растений с приобретенными последовательностями основы вектора в трансформантах обычно варьировала между 20% и 50% и иногда достигала 75% или более.
В качестве одного примера пригодности pDOW3721 был получен фрагмент 14,8 KpnI VirBCDG из плазмиды pSB1 и лигирован с помощью ДНК-лигазы T4 с ДНК pDOW3721, гидролизованной KpnI. Клетки E. coli TOP10 трансформировали смесью лигирования, и трансформанты отбирали на агаре с соей LB с добавкой 30 мкг/мл хлорамфеникола. Клоны были подвергнуты скринингу с помощью рестрикционного гидролиза EcoR I и Hind III. Полученная плазмида была названа pDOW3722.
Пример 19: Идентификация нуклеотидных последовательностей выше и ниже nilA
Штамм LBA4404nilA-int1 A. tumefaciens, содержащий интегрированную в геном плазмиду pDOW3719 (фиг.2 и пример 18), использовали для идентификации дополнительных последовательностей, расположенных выше и ниже геномной области nilA. pDOW3719 содержит ПЦР ампликон из 1796 п.н. локуса nilA штамма LBA4404 A. tumefaciens, клонированного в PCR-BLUNT II/XL-TOPO® (INVITROGEN). Эта плазмида была интегрирована в геном штамма LBA4404 A. tumefaciens посредством гомологичной рекомбинации. Колонии, которые содержали интегрированную плазмиду, были идентифицированы по устойчивости к канамицину (пример 17). Интегрированная плазмида и содержащиеся в ней элементы могут быть использованы в качестве инструментов для выделения и характеристики дополнительных нуклеотидных последовательностей методом "спасения плазмиды".
Геномную ДНК (гДНК) получали из клеток штамма LBA4404nilA-int1 в соответствии с протоколом выделения бактериальной геномной ДНК (Sambrook et al., выше). Один микрограмм гДНК раздельно гидролизовали следующими ферментами (все получены от NEB): Hind III, BamH I, Pst I, Asc I и Sac II. Эти ферменты рестрикции были выбраны специально для получения фрагментов гДНК, которые расположены выше и ниже локуса nilA. Ферменты рестрикции Hind III, BamH I и Pst I были выбраны потому, что узнаваемые ими сайты уникальны в последовательности pDOW3719. Более того, эти узнаваемые ферментами сайты расположены в областях соединения ампликонового фрагмента локуса nilA и вектора PCR-BLUNT II/XL-TOPO® (фиг.2). Расщепление гДНК этими ферментами и самолигирование полученных фрагментов приводит, таким образом, к спасению фрагмента плазмиды, который содержит неохарактеризованные геномные последовательности, лигированные рядом с сайтами связывания прямого универсального праймера M13 или обратного универсального праймера M13 плазмиды pDOW3917. Такие клоны выделяют путем трансформации смесью лигирования клеток E. coli с отбором по гену устойчивости к канамицину, несомому pDOW3917.
Кроме того, плазмида pDOW3719 не содержит сайты, узнаваемые ферментами рестрикции Asc I и Sac II. Поэтому фрагменты гДНК, создаваемые этими ферментами рестрикции, должны давать гибридный фрагмент ДНК, который охватывает всю длину последовательности интегрированной плазмиды pDOW3719 и включает области гДНК, которые примыкают с обеих сторон к интегрированной плазмиде pDOW3719.
Фрагменты гДНК, полученные в результате гидролиза ферментами рестрикции, как описано выше, самолигировали с помощью лигазы T4 (ROCHE APPLIED SCIENCES; Indianapolis, IN). Продуктами лигирования трансформировали клетки E. coli ONESHOT® TOP10 (INVITROGEN), и клетки высевали на среду LB, содержащую канамицин (50 мкг/мл). Отбирали индивидуальные колонии, и плазмидную ДНК выделяли и характеризовали по паттернам гидролиза ферментами рестрикции плазмиды. Клоны, которые содержали плазмиды, проявляющие согласующийся друг с другом паттерн полос после гидролиза ферментами рестрикции, отделяли для использования в реакциях секвенирования.
Нуклеотидные последовательности гДНК выше и ниже локуса nilA определяли с помощью метода "прогулки по геному". Были сконструированы секвенирующие праймеры, соответствующие известной последовательности гДНК (присутствующей в pDOW3719), и использованы с набором CEQTM DYE TERMINATOR CYCLE SEQUENCING в соответствии с рекомендациями производителя (BECKMAN COULTER; Fullerton, CA). На основе выявленной последовательности был сконструирован второй ряд праймеров, расположенных в ранее неизвестной геномной последовательности, и эти праймеры были использованы для получения дополнительных данных секвенирования. Этот процесс повторяли до тех пор, пока не была определена вся имеющаяся нуклеотидная последовательность гДНК. Этот метод дал 2936 п.н. последовательности выше локуса nilA и 4361 п.н. последовательности ниже локуса nilA. Вместе с 1796 п.н. ранее идентифицированного локуса nilA впервые идентифицированные последовательности фланкирующих сверху и снизу областей охватывают последовательность из 9093 п.н., включающую геномную область nilA (SEQ ID NO:24), которая распространяется в обоих направлениях от исходно идентифицированного локуса nilA.
Пример 20: Конструирование вектора для интегрирования в геномную область nilA LBA4404
Был сконструирован и создан интеграционный вектор для опосредуемой гомологией интеграции последовательностей чужеродной ДНК в геномную область nilA A. tumefaciens LBA4404. Фрагмент из 6,3 т.п.н. геномной области nilA, охватывающей локус nilA, амплифицировали с помощью ПЦР с использованием набора для ПЦР FAILSAFETM (EPICENTRE®, Madison, WI). Амплифицированный фрагмент лигировали в вектор PCR®8/GW/TOPO® (INVITROGEN), и позитивные клоны были подтверждены посредством гидролиза ферментами рестрикции и проверки последовательности ДНК. Полученный вектор, pDAB9615, был дополнительно модифицирован добавлением олигонуклеотидного фрагмента, содержащего множество уникальных сайтов узнавания ферментами рестрикции. Эти сайты рестрикции, окруженные областями из 3244 п.н. и 3128 п.н. геномной области nilA, служат в качестве сайтов клонирования для введения чужеродных нуклеотидных последовательностей. Полученный вектор был назван pDAB9618 (фиг.3).
Фрагмент из 15549 п.н., содержащий фрагмент 14,8 KpnI VirBCDG из pSB1 и бактериальный ген устойчивости к канамицину, получали путем гидролиза ДНК pDAB9292 с помощью Kpn I плюс Bst1107 I. Этот фрагмент затем лигировали с ДНК pDAB9618, которая была гидролизована Kpn I плюс Swa I, с получением вектора pDAB9621 (фиг.3). Затем использовали реакцию GATEWAY® для перемещения части pDAB9621, содержащей 14,8 KpnI VirBCDG и бактериальный ген устойчивости к канамицину и окруженной с каждой стороны последовательностями из 3 т.п.н. геномной области nilA LBA4404, в вектор GATEWAY® PDESTTM14 посредством реакции AN L-R CLONASE® (INVITROGEN). Полученная плазмида, pDAB9698 (фиг.3, SEQ ID NO:25), была подтверждена посредством гидролиза ферментами рестрикции и реакций секвенирования ДНК. pDAB9698 служила в качестве интеграционного вектора для интегрирования генов vir из pTiBo542 pSB1 (находящихся во фрагменте 14,8 KpnI VirBCDG) в хромосомную область nilA штамма LBA4404 A. tumefaciens.
Пример 21: Хромосомная интеграция фрагмента 14,8 KpnI VirBCDG посредством гомологичной рекомбинации
ДНК плазмиды pDAB9698 получали с использованием набора NUCLEOBOND® AX ANION EXCHANGE CHROMATOGRAPHY PLASMID DNA ISOLATION (MACHEREY-NAGEL). Очищенную плазмидную ДНК электропорировали в клетки LBA4404 A. tumefaciens (INVITROGEN). Вкратце, 500 нг плазмидной ДНК инкубировали с клетками при 4°С в течение 10 минут. Эту смесь переносили пипеткой в охлажденную на льду 0,2 см кювету GENE PULSER® (BIO-RAD) и электропорировали с помощью BIO-RAD GENE PULSER при следующих установках: выходная емкость 25 мкФ, расширитель емкости 960 мкФ, сопротивление 200 Ом и напряжение 2,5 кВ. Сразу после электропорации добавляли 950 мкл среды SOC (INVITROGEN), и смесь переносили в пробирку Falcon 2059 (BECTON DICKINSON AND CO.; Franklin Lakes, NJ). После этого трансформированные клетки инкубировали при 28°С в течение от 5 до 6 часов. После инкубации клетки высевали на отдельные чашки со средой YEP, содержащей канамицин (50 мкг/мл). Чашки выращивали в перевернутом состоянии при 28°С в течение от 36 до 48 часов. Одиночные колонии отбирали и размножали в 5 мл жидкой среды YEP, содержащей канамицин (50 мкг/мл), в течение приблизительно 36 часов при 28°С. Эти культуры использовали для получения партий культур в глицерине путем тщательного перемешивания с равным объемом 100% стерильного глицерина с последующим замораживанием и хранением при -80°С.
Пример 22: Фенотипическое и молекулярное подтверждение хромосомной интеграции фрагмента 14,8 KpnI VirBCDG
Как ожидается, pDAB9698, содержащая точку начала репликации colE1, не реплицируется автономно в клетках A. tumefaciens. После трансформации клеток LBA4404 ДНК pDAB9698 стабильная устойчивость к канамицину возникает в результате интеграции ДНК pDAB9698 в автономно реплицируемые генетические элементы Agrobacterium. Эти плазмидные интегранты обычно делятся на четыре класса, которые могут быть использованы в соответствии с разными вариантами осуществления штаммов Agrobacterium и методами их использования, как описано в настоящем описании. Первый класс включает клетки, в которых ДНК pDAB9698 интегрирована в сайт, отдаленный от геномной области nilA, посредством негомологичной рекомбинации. Эти клетки должны быть устойчивы к канамицину благодаря гену устойчивости к канамицину, расположенному рядом с фрагментом 14,8 KpnI VirBCDG, и дополнительно устойчивы к ампициллину благодаря гену устойчивости к ампициллину, содержащемуся в основе вектора pDAB9698 (pDESTTM14). Второй класс включает клетки, в которых ДНК pDAB9698 интегрирована в автономно реплицирующуюся вспомогательную плазмиду pAL4404 Ti (в естественных условиях являющуюся резидентной в LBA4404) посредством гомологичной рекомбинации, опосредуемой генами VirBCDG из pTiBo542, находящимися на pDAB9698, и генами VirBCDG из pTiACH5, находящимися на pAL4404. Эти клетки также должны быть устойчивы и к канамицину, и к ампициллину. Третий класс включает клетки, в которых ДНК pDAB9698 интегрирована в геномную область nilA LBA4404 посредством одиночного события гомологичной рекомбинации (кроссовера), опосредуемого либо последовательностями геномной области nilA из приблизительно 3 т.п.н., находящимися на pDAB9698 и примыкающими к фрагменту из 15549 п.н., содержащему фрагмент 14,8 KpnI VirBCDG из pSB1 и ген устойчивости к канамицину. Эти клетки также должны быть устойчивы и к канамицину, и к ампициллину. Четвертый класс включает клетки, в которых одиночное событие кроссовера клеток указанного выше класса 3 подвергается второму событию кроссовера, опосредуемого отныне дуплицированными последовательностями геномной области nilA из 3 т.п.н., которые возникли в результате одиночного события кроссовера. В зависимости от того, какая из последовательностей, примыкающих к геномной области nilA из 3 т.п.н., вызвала одиночное событие кроссовера, и какая из этих примыкающих последовательностей вызывает второе событие кроссовера, полученные клетки должны быть либо чувствительными к канамицину и устойчивыми к ампициллину, либо устойчивыми к канамицину и чувствительными к ампициллину. Предпочтительные клетки, описанные в настоящем описании, включают последний класс, то есть клетки, которые устойчивы к канамицину и чувствительны к ампициллину. Эти события двойного кроссовера, которые не содержат основу pDESTTM14 плазмиды, желательны, поскольку они не содержат сверхподвижных генетических элементов, таких как точка начала репликации colE1 и ген устойчивости к ампициллину.
Позитивные трансформанты, выделенные в примере 21, были подвергнуты скринингу на желаемое событие интеграции, опосредуемое двойной гомологичной рекомбинацией. Устойчивые к канамицину изоляты, содержащие фрагмент из 15549 п.н., включающий фрагмент 14,8 KpnI VirBCDG из pSB1 и ген устойчивости к канамицину (и не включающий основу вектора pDESTTM14), были идентифицированы по чувствительности к ампициллину. Позитивные трансформанты выращивали в 3 мл среды YEP, содержащей канамицин (50 мкг/мл), при 28°С в течение приблизительно 36 часов. Эти культуры затем высевали штрихом на твердые среды YEP, содержащие раздельно следующие различные антибиотики (концентрации в мкг/мл): рифампицин, 100; канамицин, 50; стрептомицин, 125; хлорамфеникол, 50; эритромицин, 200; тетрациклин, 12,5; и ампициллин, 100. Чашки инкубировали при 28°С в течение 48 часов, и выросшие колонии подсчитывали. Был идентифицирован штамм, который был устойчив к канамицину, рифампицину (хромосомный маркер) и стрептомицину (маркер pAL4404; Ooms et al., выше). Более того, штамм был чувствителен к хлорамфениколу, эритромицину и тетрациклину. Но самое важное, штамм был чувствителен к ампициллину. Этот лекарственный скрининговый фенотип указывает на желаемое событие двойной кроссоверной гомологичной рекомбинации, где фрагмент из 15549 п.н., содержащий фрагмент 14,8 KpnI VirBCDG из pSB1 и ген устойчивости к канамицину, интегрирован в хромосому LBA4404 A. tumefaciens. Этот штамм назван DAt16174.
Наличие генов VirBCDG из pTiBo542 в штамме DAt16174 было дополнительно подтверждено с помощью молекулярной характеристики. Геномная ДНК штамма DAt16174 была выделена с использованием протокола выделения бактериальной геномной ДНК (Sambrook et al., выше, и ее обновления). Были сконструированы ПЦР праймеры для амплификации перекрывающихся фрагментов интегрированных в хромосому генов VirBCDG. Реакции ПЦР с использованием праймеров, описанных в таблице 6, были проведены с помощью набора ПЦР FAILSAFETM (EPICENTRE®) в соответствии с указаниями производителя. Из-за большого суммарного молекулярного размера интегрированных генов VirBCDG были проведены реакции амплификации с получением пяти перекрывающихся фрагментов. Ампликоны ожидаемого размера очищали из агарозных гелей с помощью набора QIAEX II GEL EXTRACTION KIT (QIAGEN; Valencia, CA) в соответствии с протоколом производителя. Эти фрагменты были клонированы в вектор PCR2.1®-TOPO®TA с использованием набора PCR2.1®-TOPO®TA CLONING® KIT (INVITROGEN). Колонии бактерий с предполагаемым наличием клонов ампликонов ПЦР были подтверждены с помощью гидролиза ферментами рестрикции. Последовательности ДНК ампликоновых фрагментов были определены с помощью набора CEQTM DYE TERMINATOR CYCLE SEQUENCING KIT в соответствии с инструкциями производителя, и данные секвенирования были проанализированы с использованием программы SEQUENCHERTM, версия 4.1.4 (GENE CODES CORP.; Ann Arbor, MI). Полученные последовательности дали непрерывную последовательность из 22 т.п.н., которая охватывала весь фрагмент из 15549 п.н., содержащий фрагмент 14,8 KpnI VirBCDG из pSB1 и ген устойчивости к канамицину, плюс обе области из приблизительно 3 т.п.н., примыкающие к геномной области nilA и дополнительно расширенные вверх и вниз от геномных областей nilA (тем самым включая хромосомную последовательность LBA4404, которая исходно не содержалась в pDAB9698).
Принадлежность штамма DAt16174 к Agrobacterium tumefaciens была проверена с помощью кетолактозного теста. Предположительно трансформированные колонии высевали штрихом на агар с лактозой и инкубировали при 28°С в течение 48 часов. Чашки затем наполняли раствором Бенедикта и отслеживали при комнатной температуре. Изоляты, которые превращали раствор Бенедикта и подстилающий агар из синего в желтый, были таким образом подтверждены как являющиеся Agrobacterium.
Особенность штамма DAt16174 A. tumefaciens заключается в том, что его можно с выгодой использовать в качестве агента трансформации растений для переноса генов Т-ДНК с бинарных векторов, содержащих точки начала репликации, например, классов IncP, IncW или VS1. В широком плане введенный бинарный вектор может содержать точку начала репликации любого класса, способного к репликации в Agrobacterium при условии, что он совместим с точкой начала репликации pTi (и связанными функциями) резидентной плазмиды pAL4404 в DAt16174. Таким образом, область возможных применений штамма DAt16174 охватывает совместное присутствие в штамме DAt16174 более одной бинарной векторной плазмиды, если плазмиды содержат совместимые точки начала репликации (т.е. относятся к разным группам несовместимости). Селекция таких введенных бинарных векторов не должна быть связана с маркерами селекции, придающими устойчивость к канамицину, рифампицину или стрептомицину, поскольку штамм DAt16174 устойчив к этим трем антибиотикам.
Бинарные векторы могут реплицироваться автономно в клетках и E. coli, и Agrobacterium. Они включают последовательности, обрамленные правой и левой областями пограничных повторов Т-ДНК, которые могут включать маркерный ген селекции, пригодный для отбора трансформированных клеток растения, клонирующий линкер, клонирующий полилинкер или другую последовательность, которая может действовать в качестве сайта введения генов, предназначенных для трансформации растительной клетки. Ими можно прямо трансформировать клетки Agrobacterium путем электропорации, путем прямой опосредованной химически трансформации ДНК, введением с помощью бактериальной конъюгации или с помощью других методов. Используемая в качестве клетки-хозяина Agrobacterium содержит, по меньшей мере, одну плазмиду, несущую область vir. Область vir необходима для получения белков Vir, выполняющих все необходимые функции, принимающие участие в переносе Т-ДНК в растительную клетку. Плазмида, несущая область vir, обычно представляет собой мутантную плазмиду Ti или Ri (вспомогательную плазмиду), из которой была удалена область Т-ДНК, включая левый и правый пограничные повторы Т-ДНК. Примеры штаммов Agrobacterium, которые содержат вспомогательные плазмиды и пригодны для трансформации растений, включают, например, LBA4404, GV3101(pMP90), GV3101(pMP90RK), GV2260, GV3850, EHA101, EHA105 и AGL1. Многочисленные примеры бинарных векторных систем рассмотрены в обзоре Hellens et al., (2000, Trends Plant Sci. 5:446-451).
Кроме того, преимущества трансформации растений, придаваемые штамму LBA4404(pSB1) [используемому в супербинарной системе] опероном virB из pTiBo542 (который включает гены virB1, virB2, virB3, virB4, virB5, virB6, virB7, virB8, virB9, virB10 и virB11), геном virG, опероном virC (который включает гены virC1 и virC2) и частью оперона virD, включающей ген virD1, содержащимися в плазмиде pSB1, сохраняются в штамме DAt16174. Поскольку перечисленные выше супербинарные гены vir интегрированы в хромосому LBA4404, штамм DAt16174 обозначается как штамм SUPERCHROME. В отличие от супербинарной системы, для использования штамма DAt16174 не требуется образование нестабильных супербинарных плазмид посредством гомологичной рекомбинации между pSB1 и челночными векторами, такими как pSB11. Дополнительным преимуществом штамма SUPERCHROME является возможность введения в штамм стандартных бинарных векторов для трансформации растений.
Пример 23: Биохимическая и молекулярная характеристика тканей маиса, трансформированного разными штаммами Agrobacterium, несущими pDAB101556
Бинарный вектор для трансформации растений pDAB101556 (фиг.5) был сконструирован путем сочетания стандартных методов клонирования и технологии GATEWAYTM. Бинарный вектор pDAB101556 основывается на точке начала репликации типа IncP плазмиды RK2, и основа вектора содержит бактериальный ген, придающий устойчивость к спектиномицину в концентрации 100 мкг/мл. Пограничные повторы Т-ДНК берут начало от области TL pTi15955. Внутри правой границы (RB) и множественных левых границ (LB) области Т-ДНК плазмиды pDAB101556 расположены 2 экспрессируемые в растениях кодирующие белок последовательности (CDS). Первый ген (маркер селекции) включает кодирующую область для белка-маркера селекции AAD1 (SEQ ID NO: 13) (патент США № 7838733), которая находится под транскрипционным контролем промотора убиквитина 1 маиса из 1991 п.н. с ассоциированным интроном 1 (патент США № 5510474). Этот ген заканчивается 3’UTR липазы маиса (патент США № 7179902). Второй ген (маркер для скрининга) включает CDS для желтого флуоресцентного белка (YFP, в точности как раскрыто в патенте США № 7951923), транскрипция которой контролируется промотором убиквитина 1 маиса с ассоциированным интроном 1. Этот ген заканчивается 3’UTR Per5 маиса (патент США № 6384207).
Плазмида pDAB101556 была успешно введена путем электропорации в клетки штамма LBA4404 A. tumefaciens с получением штамма LBA4404(pDAB101556). Это сочетание штамм/плазмида, таким образом, включает стандартную бинарную систему трансформации растений. Трансформанты, отобранные на основе устойчивости к стрептомицину и спектиномицину, проверяли с помощью гидролиза плазмидной ДНК ферментами рестрикции перед получением замороженных в глицерине партий и их хранением при -80°С. Собирали все клетки штамма LBA4404(pDAB101556) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их для трансформаций маиса с помощью методов, раскрытых в примере 24.
Плазмида pDAB101556 была успешно введена путем электропорации в клетки штамма DAt13192 A. tumefaciens (см. пример 7) с получением штамма DAt13192(pDAB101556). Это сочетание штамм/плазмида, таким образом, включает тройную систему трансформации растений с недостаточностью рекомбинации. Трансформанты, отобранные на основе устойчивости к эритромицину, канамицину и спектиномицину, проверяли с помощью гидролиза плазмидной ДНК ферментами рестрикции перед получением замороженных в глицерине партий и их хранением при -80°С. Собирали все клетки штамма DAt13192(pDAB101556) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их для трансформаций маиса с помощью методов, раскрытых в примере 24.
Было предпринято несколько попыток введения ДНК плазмиды pDAB101556 в штамм DAt20711 (см. пример 9), тройную систему с эффективной рекомбинацией. Во всех случаях было обнаружено, что в этом штамме плазмида pDAB101556 нестабильна, и штамм Dat2071l(pDAB101556) не был сконструирован.
Плазмида pDAB101556 была успешно введена путем электропорации в клетки штамма DAt16174 A. tumefaciens (пример 22) с получением штамма DAt16174(pDAB101556). Это сочетание штамм/плазмида, таким образом, включает SUPERCHROME/бинарную систему трансформации растений. Трансформанты, отобранные на основе устойчивости к стрептомицину, канамицину и спектиномицину, проверяли с помощью гидролиза плазмидной ДНК ферментами рестрикции перед получением замороженных в глицерине партий и их хранением при -80°С. Собирали все клетки штамма DAt16174(pDAB101556) с агаровой чашки, инокулированной замороженной в глицерине партией, и использовали их для трансформаций маиса с помощью методов, раскрытых в примере 24.
Пример 24: Трансформация маиса штаммами Agrobacterium, несущими бинарный вектор pDAB101556
Получение незрелых зародышей. Семена сорта B104 были высажены в 3,5-дюймовые (88,9 мм) горшки SVD с METRO MIX 360 (SUN GRO HORTICULTURE Inc.; Bellevue, WA). Когда растения достигали стадии роста V4-V5, их переносили в 4-галлоновые (15,1 л) горшки, содержащие смесь 1:1 METRO MIX 360 и кальцинированную глину PROFILE GREENS GRADE (PROFILE PRODUCTS LLC; Buffalo Grove, IL) с 20 граммами OSMOCOTE 19-6-12 и 20 граммами IRONITETM в качестве добавок. Растения выращивали в оранжерее при использовании сочетания 1000 Вт HPS (натриевая лампа высокого давления) и 1000 Вт MH (металл-галоидных) ламп с установленным режимом освещения 16:8 освещение/темнота, если освещенность снаружи не превышала 450 Вт/м2. Для получения незрелых зародышей для трансформации проводили контролируемое сестринское опыление или самоопыление. Незрелые зародыши были получены через 10-13 дней после опыления, кода зародыши имели размер приблизительно от 1,6 до 2,0 мм.
Инфицирование и совместное культивирование. Поверхность колосьев маиса стерилизовали погружением в 20% имеющийся в продаже отбеливатель с поверхностно-активным веществом LIQUINOXTM (1 или 2 капли на 500 мл) в течение 20 минут и последующей трехкратной промывкой стерильной водой. Суспензию клеток Agrobacterium, содержащих бинарный вектор pDAB101556, получали из бактерий, выращенных на твердой среде AB при 20°C в течение от 2 до 3 дней, с последующим выращиванием на твердой среде YEP при 28°С в течение от 1 до 3 дней. Обе среды AB и YEP содержали добавки соответствующих антибиотиков, как описано в примере 23, для каждого штамма Agrobacterium, тестированного с бинарным вектором pDAB101556. Петли с клетками, соскобленными с чашки YEP, переносили в жидкую среду для инфицирования объемом от 10 до 15 мл, содержащую: соли MS (Frame et al., выше), модифицированные ISU витамины MS (Frame et al., выше), 3,3 мг/л дикамба-этанола, 68,4 г/л сахарозы, 36 г/л глюкозы, 700 мг/л L-пролина, pH 5,2, и содержащую 100-200 мкМ ацетосирингона. Раствор мягко отбирали и сливали пипеткой с использованием 5 мл стерильной пипетки или перемешивали в смесителе до получения однородной суспензии, и концентрацию доводили до оптической плотности приблизительно 1,0 при 600 нм (ОП600) с использованием спектрофотометра Hewlett-Packard P8452a.
Совместное культивирование. Незрелые зародыши выделяли прямо в микроцентрифужную пробирку, содержащую 2 мл среды для инфицирования. Среду удаляли и заменяли свежей средой для инфицирования объемом от 1 до 2 мл, после чего ее заменяли 1,5 мл раствора Agrobacterium. Раствор Agrobacterium и зародыша инкубировали в течение 5 минут при комнатной температуре и затем переносили в среду для совместного культивирования, которая содержала соли MS, модифицированные ISU витамины MS, 3,3 мг/л дикамба-этанола, 30 г/л сахарозы, 700 мг/л L-пролина, 100 мг/л мио-инозита, 100 мг/л ферментативного гидролизата казеина, 15 мг/л AgNO3, 100-200 мкМ ацетосирингона и 2,3 г/л GELRITETM (SIGMA-ALDRICH; St. Louis, MO), при pH 5,8. Инкубация совместного культивирования продолжалась в течение 3 дней в темноте при 20°С.
Восстановление и отбор. После совместного культивирования зародыши переносили в неселективную восстанавливающую среду на основе MS, содержащую соли MS, модифицированные ISU витамины MS, 3,3 мг/л дикамба-этанола, 30 г/л сахарозы, 700 мг/л L-пролина, 100 мг/л мио-инозита, 100 мг/л ферментативного гидролизата казеина, 15 мг/л AgNO3, 0,5 г/л MES, 250 мг/л цефотаксима и 2,3 г/л GELRITETM при pH 5,8. Инкубацию продолжали в течение 7 дней в темноте при 28°С. После 7 дней периода восстановления зародыши переносили в селективную среду. Для отбора тканей маиса, трансформированных супербинарной или бинарной плазмидой, содержащей экспрессируемый в растениях маркерный ген селекции aad-1, использовали восстанавливающую среду на основе MS (выше) с добавкой галоксифопа. Зародыши сначала переносили в селективную среду I, содержащую 100 нМ галоксифопа, и инкубировали в течение 2 недель, а затем переносили в селективную среду II с 500 нМ галоксифопа и инкубировали еще в течение 2 недель. Трансформированные изоляты были получены в течение периода приблизительно 5 недель при 28°С в темноте. При необходимости, выделенные изоляты объединяли путем переноса в свежую селективную среду II еще на 2 недели перед переносом в среды для регенерации.
Специалисты в области техники трансформации маиса должны понимать, что для отбора трансформированных растений пригодны другие методы при использовании других экспрессируемых маркерных генов селекции растений (например, генов устойчивости к гербицидам).
Регенерация I. После процесса отбора культуры переносили в среду для регенерации I на основе MS, содержащую соли MS, модифицированные ISU витамины MS, 60 г/л сахарозы, 350 мг/л L-пролина, 100 мг/л мио-инозита, 50 мг/л ферментативного гидролизата казеина, 1 мг/л AgNO3, 250 мг/л цефотаксима, 2,5 г/л GELRITETM и 500 нМ галоксифопа, при pH 5,8. Инкубацию продолжали в течение 2 недель при 28°С в темноте.
Регенерация II. Культуры переносили в среду для регенерации II на основе MS, содержащую соли MS, модифицированные ISU витамины MS, 30 г/л сахарозы, 100 мг/л мио-инозита, 250 мг/л цефотаксима, 2,5 г/л GELRITETM и 500 нМ галоксифопа, при pH 5,8. Через 3 недели при 28°С при режиме освещения 16/8 часов белым флуоресцентным светом (приблизительно 80 мкэйнштейн·м-2·сек-1) проростки изымали и переносили в среду для удлинения побегов/корней на основе MS (или на основе ½ MS), состоящую из солей MS (или солей ½ MS), модифицированных ISU витаминов MS, 0,5 г/л MES, 30 г/л сахарозы, 100 мг/л мио-инозита, 2,5 г/л GELRITETM, при pH 5,8. Когда проростки достигали длины от 4 до 6 см, их переносили в ростовую камеру и в конечном итоге в оранжерею.
Получение семян. Регенерированные растения переносили в 3,5-дюймовые (88,9 мм) горшки SVD с METRO MIX 360 и помещали в ростовую камеру для закаливания. Когда растения достигали стадии роста V3, их перемещали в оранжерею, а на стадии роста V4/V5 их переносили в 5-галонные (18,9 л) горшки, содержащие смесь 1:1 METRO MIX 360 и кальцинированную глину PROFILE GREENS GRADE с 20 граммами OSMOCOTE 19-6-12 и 20 граммами IRONITETM в качестве добавок. Растения выращивали в оранжерее при режиме освещения 16:8 освещение/темнота. Посредством контролируемого опыления (обратное скрещивание с B104) были получены семена T1. Семена собирали через 6 недель после опыления.
Были проведены многочисленные эксперименты по трансформации маиса сконструированными штаммами A. tumefaciens LBA4404(pDAB 101556), DAt13192(pDAB101556) и DAt16174(pDAB101556), и трансгенные клетки, отобранные при ингибирующих концентрациях галоксифопа, использовали для регенерации проростков и дальнейших исследований. В общей сложности при трансформациях LBA4404(pDAB101556) было зафиксировано 16 событий, при трансформациях DAt13192(pDAB101556) было зафиксировано 49 событий, а при трансформациях DAt16174(pDAB101556) было зафиксировано 60 событий (таблица 7).
Количество копий трансгена aad-1 в трансгенных растениях T0 было установлено с помощью тестов гидролиза зондов (Bubner and Baldwin, выше) с использованием специфичных для гена олигонуклеотидов. Анализы Саузерн-блотингом расщепленной NcoI ДНК, полученной из отобранных событий методом экстракции цетилтриметиламмония бромидом, были проведены с использованием амплифицированного ПЦР фрагмента гена aad-1 в виде меченного 32P зонда. Кроме того, с помощью тестов гидролиза зондов было выявлено наличие интегрированных последовательностей основы вектора, берущих начало от pDAB101556.
Таким образом, штамм DAt16174, SUPERCHROME штамм, включающий полный набор генов vir из pTiACH5, находящихся на pAL4404, и дополнительно включающий частичный набор генов vir из pTiBo542, интегрированных в локус nilA хромосомы LBA4404, способен эффективно продуцировать трансформированные растения маиса. Кроме того, хотя общая эффективность трансформации этим штаммом несколько ниже эффективности тройного штамма, качество вызываемых SUPERCHROME событий является превосходным, поскольку в 90% вызванных событий имеются вставки единственной копии при отсутствии выявляемых загрязнений основой.
Хотя настоящее изобретение было описано в виде конкретных иллюстративных вариантов осуществления, предназначенных для иллюстрации немногих аспектов изобретения, настоящее изобретение может быть дополнительно модифицировано в пределах сущности и объема настоящего раскрытия. Заявка, таким образом, предназначена для охвата любых вариаций, применений или адаптаций изобретения, использующих его общие принципы. Кроме того, настоящая заявка предназначена для охвата таких приложений настоящего раскрытия, которые относятся к известной или деловой практике в той области техники, к которой относится настоящее изобретение, и которые оказываются в границах прилагаемой формулы изобретения.
Все ссылки, включая публикации, патенты и патентные заявки, цитируемые в настоящем описании, тем самым включены в качестве ссылки в той же мере, как если бы для каждой ссылки было отдельно и специально указано, что она включена в качестве ссылки, и были представлены в своем полном объеме в настоящем описании. Обсуждавшиеся в настоящем описании ссылки представлены исключительно для их раскрытия до даты регистрации настоящей заявки. В настоящем описании ничто не следует толковать как допущение того, что изобретатели не уполномочены датировать такое раскрытие на основании предшествующего изобретения.
|
|
|
|
|
|
|


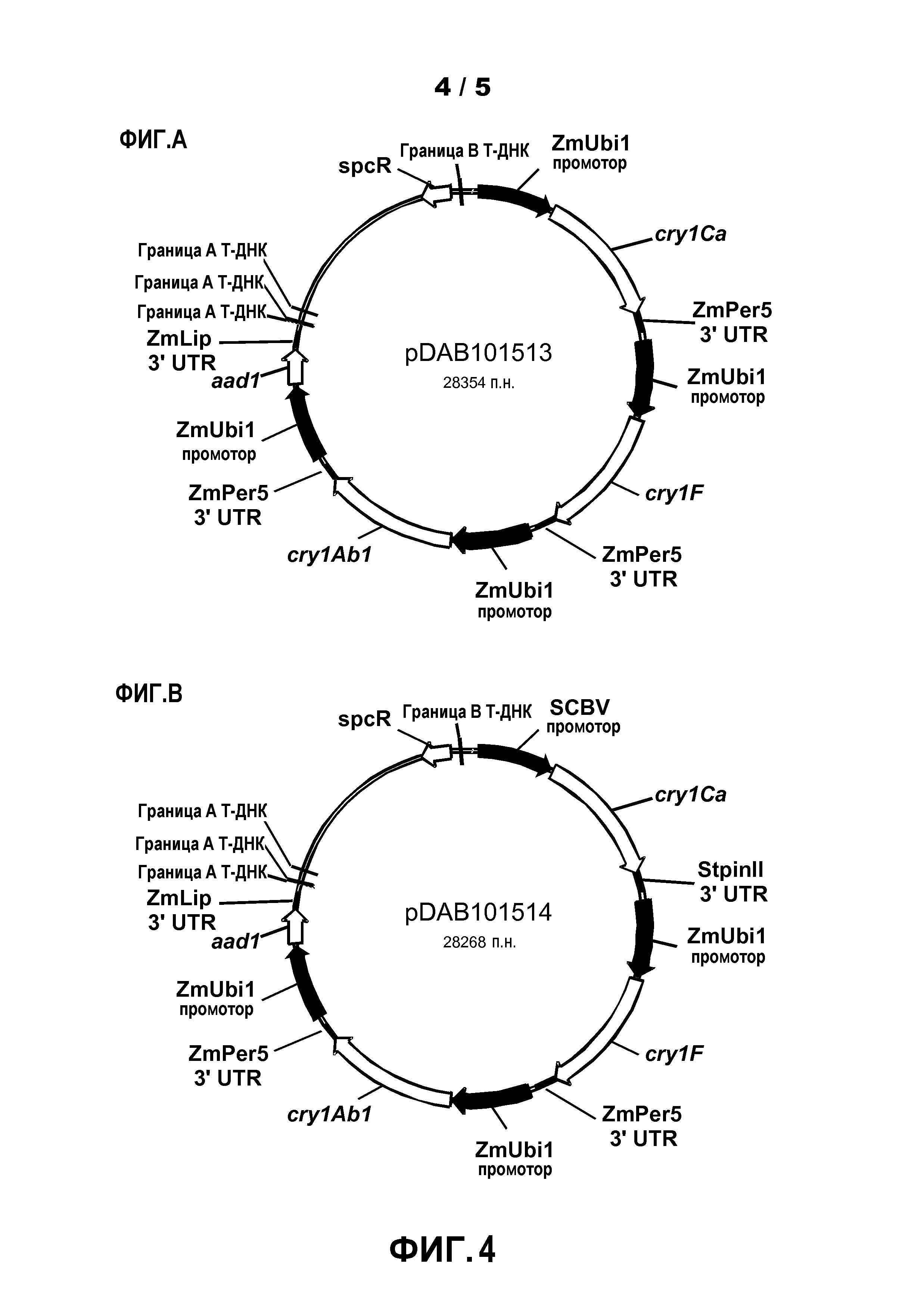

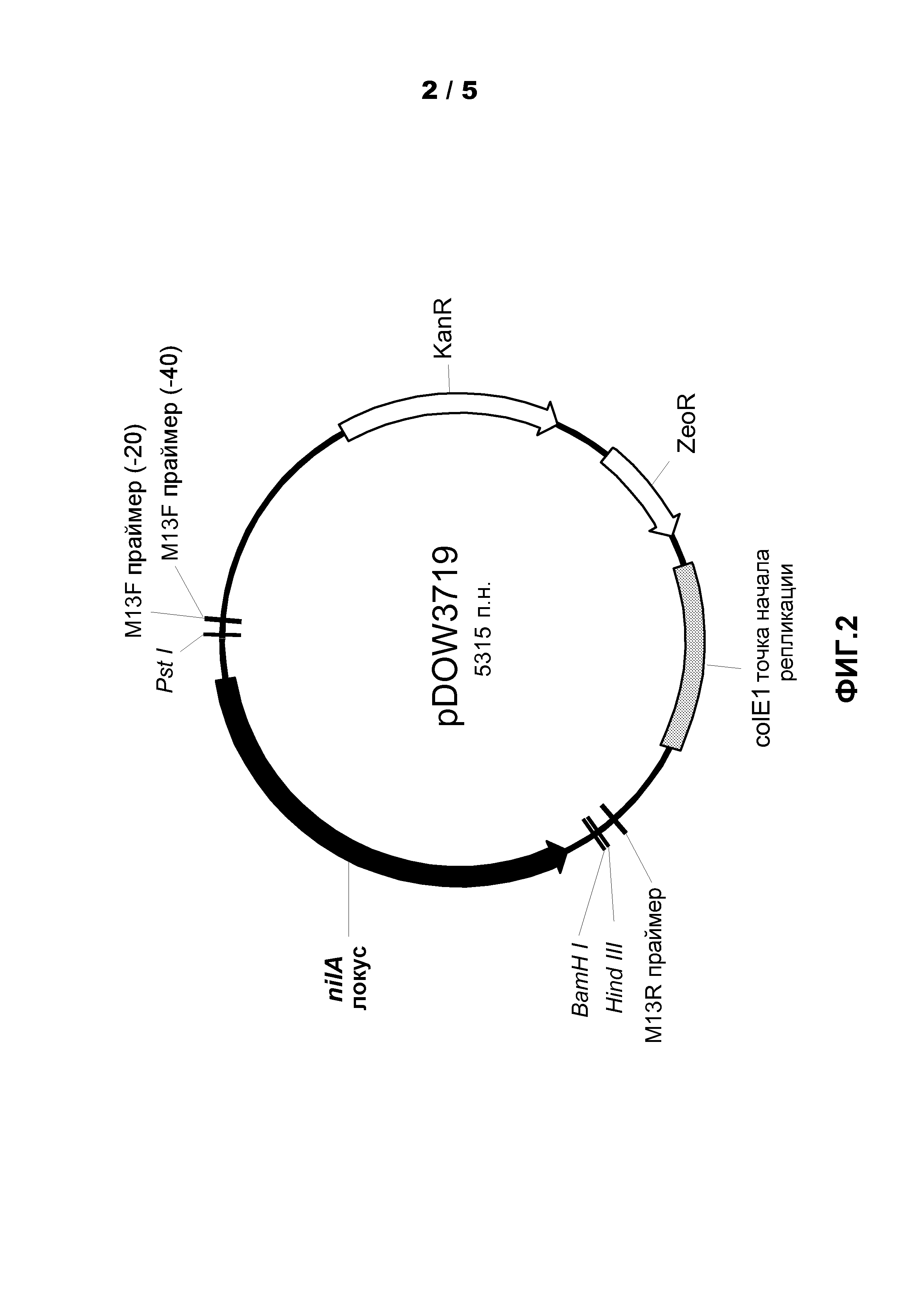







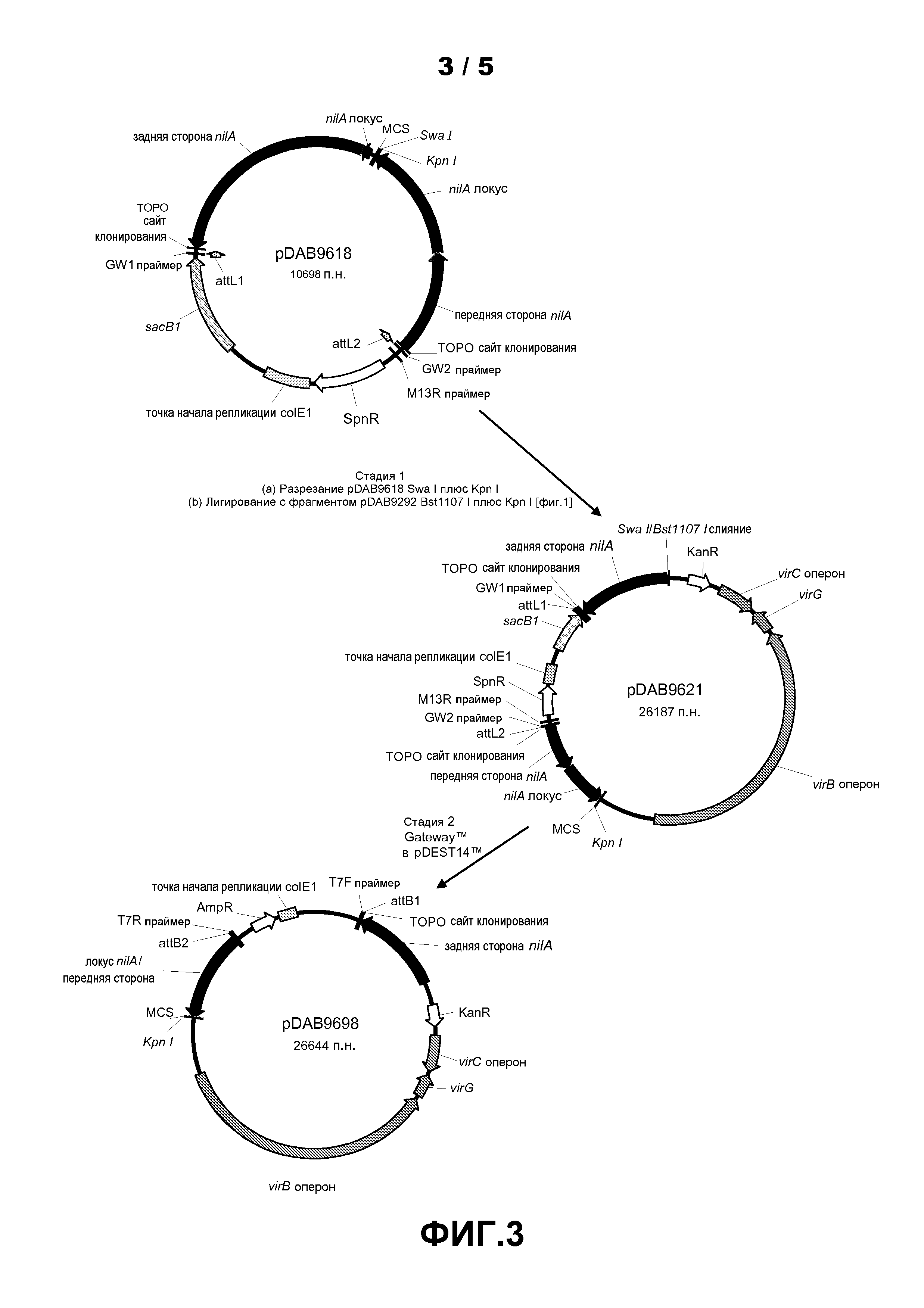





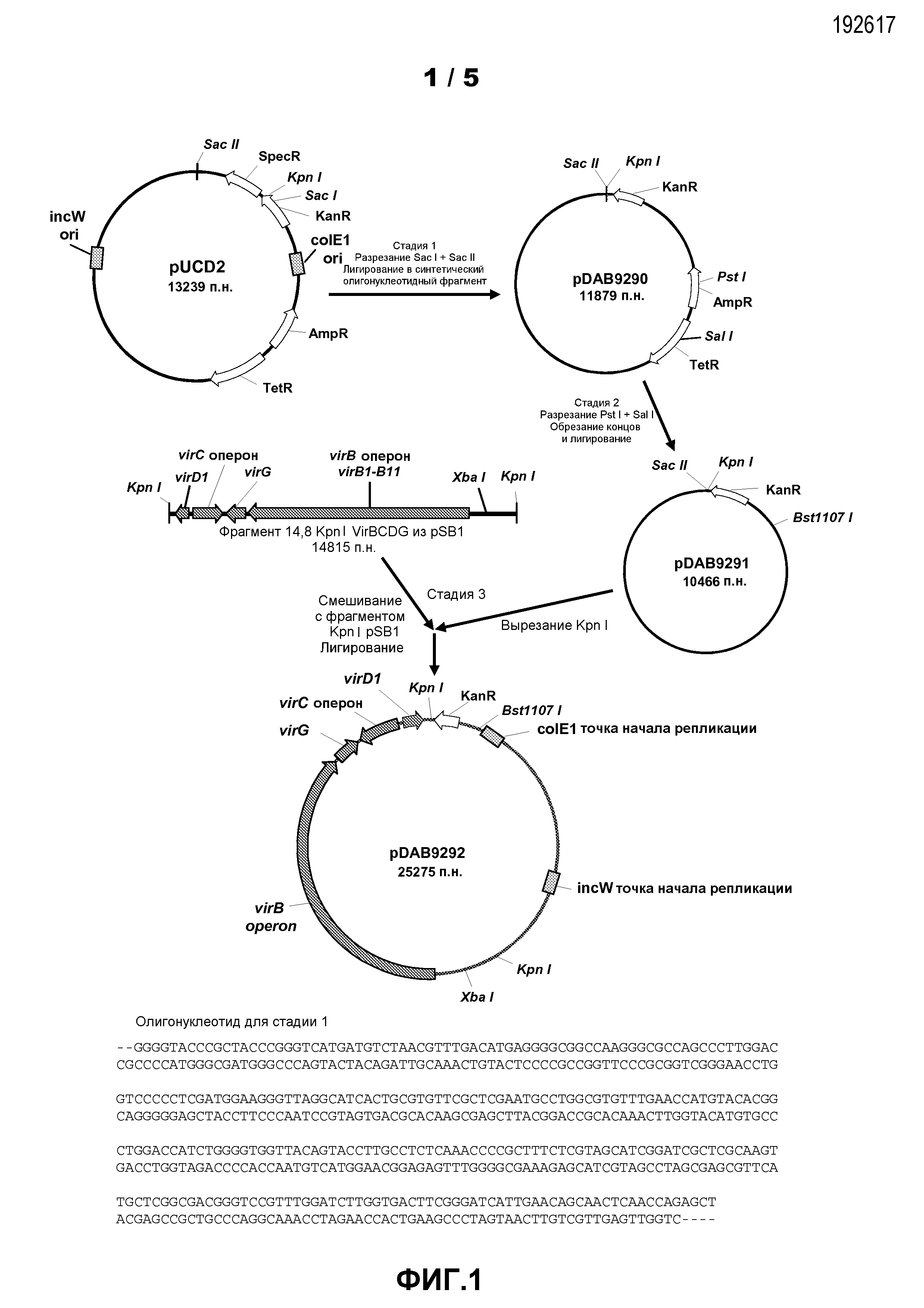






































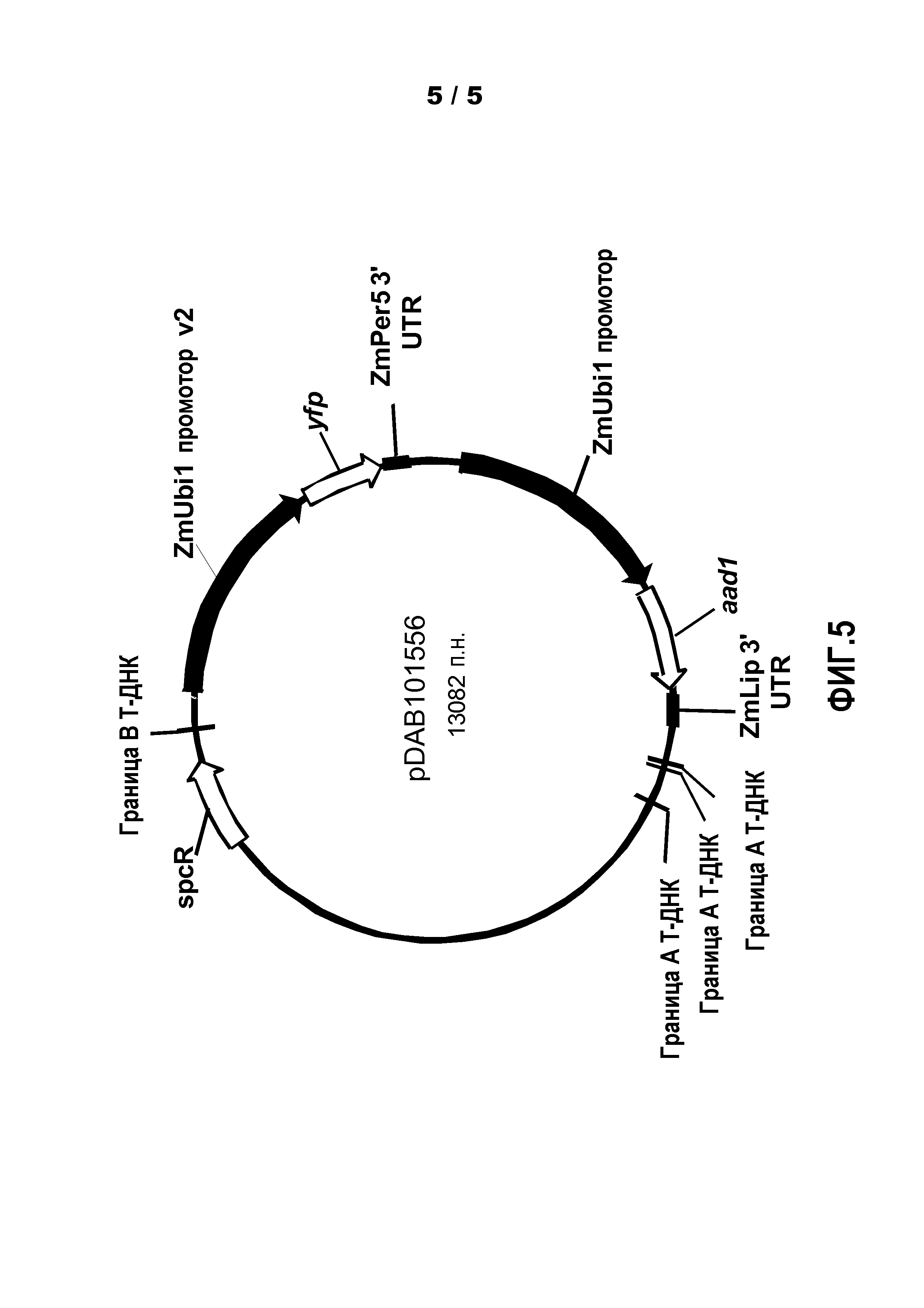





















Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
5-фторпиримидиновые производные в качестве фунгицидов
Пестицидные композиции
Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
Стабильная пестицидная композиция на основе сульфоксимина и способ борьбы с насекомыми
5-фторпиримидиновые производные в качестве фунгицидов

