Результат интеллектуальной деятельности: ИСПОЛЬЗОВАНИЕ ГЛУБИННОЙ МОРСКОЙ ВОДЫ ИЗ СЕРОВОДОРОДНОЙ ЗОНЫ ЧЕРНОГО МОРЯ В КАЧЕСТВЕ СРЕДЫ КУЛЬТИВИРОВАНИЯ МОРСКИХ ВОДОРОСЛЕЙ
Вид РИД
Изобретение
Изобретение относится к марикультуре и предназначено для культивирования морских водорослей в лабораторных или промышленных условиях.
Развитие марикультуры, как и развитие земледелия, ставит задачу получения урожая на одних и тех же плантациях с его последующим изъятием и постоянное возобновление запасов питательных веществ (биогенов и микроэлементов) на плантациях для сохранения необходимой продуктивности угодий. Как показал опыт земледелия, внесение извне удобрений промышленного или естественного происхождения наряду с функцией восполнения запаса питательных веществ, имеет и отрицательное влияние в виде загрязнения плантаций и окружающей среды токсичными примесями, ухудшения качества продукции, нарушения природных потоков элементов не только на плантациях, но и далеко за их пределами, в том числе и в водных экосистемах. Это искажает природный баланс элементного состава вод, что при достижении критических количественных показателей ведет к ухудшению биологического качества водной среды, в том числе и в морских экосистемах. Эффективным направлением решения этих вопросов при развитии марикультуры в морских акваториях является использование природных обогащенных вод этих же водоемов без внесения извне дополнительного антропогенного потока веществ. В связи с этим, применение природных вод в качестве плодородной среды культивирования водорослей в лабораторных или производственных условиях является актуальной задачей.
Существуют рекомендации (см. А.С. №1618896, СССР, МКИ А01K61/00) по использованию глубинной сероводородной воды Черного моря для культивирования гидробионтов. Глубинную воду сероводородной зоны Черного моря поднимают на поверхность и освобождают от смеси сероводорода и других горючих газов. После подогрева глубинная морская вода направляется в пруды марикультурального предприятия.
В основу изобретения поставлена задача применения глубинной воды Черного моря, содержащей сероводород, а также глубинной окисленной воды в качестве плодородной среды культивирования водорослей.
Для достижения поставленной задачи было проведено комплексное исследование экологического качества как природной, так и окисленной вод, из сероводородной зоны Черного моря и дана оценка экологического качества этих вод для черноморских гидробионтов на примере ряда видов одноклеточных водорослей и многоклеточной бентосной зеленой водоросли.
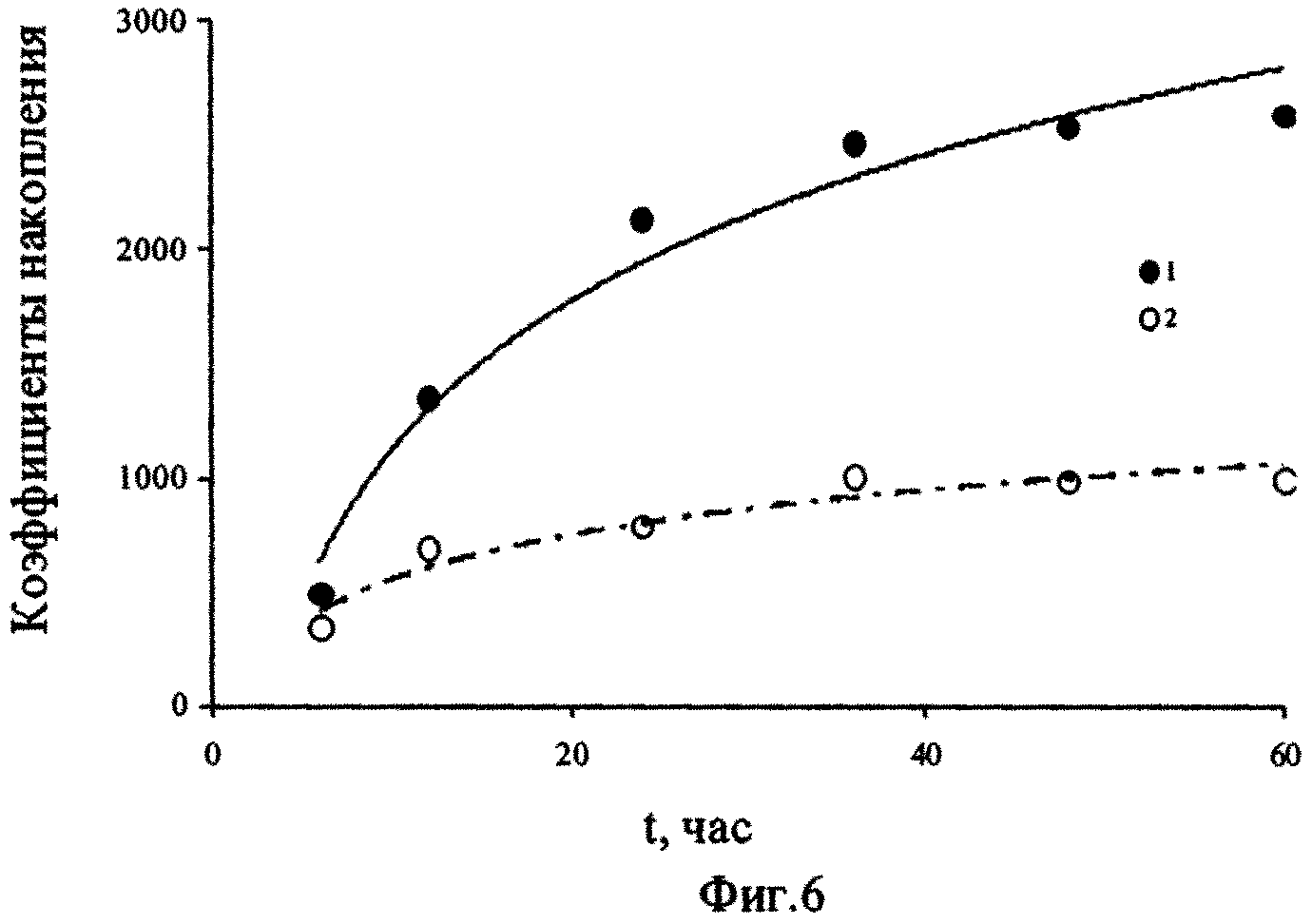
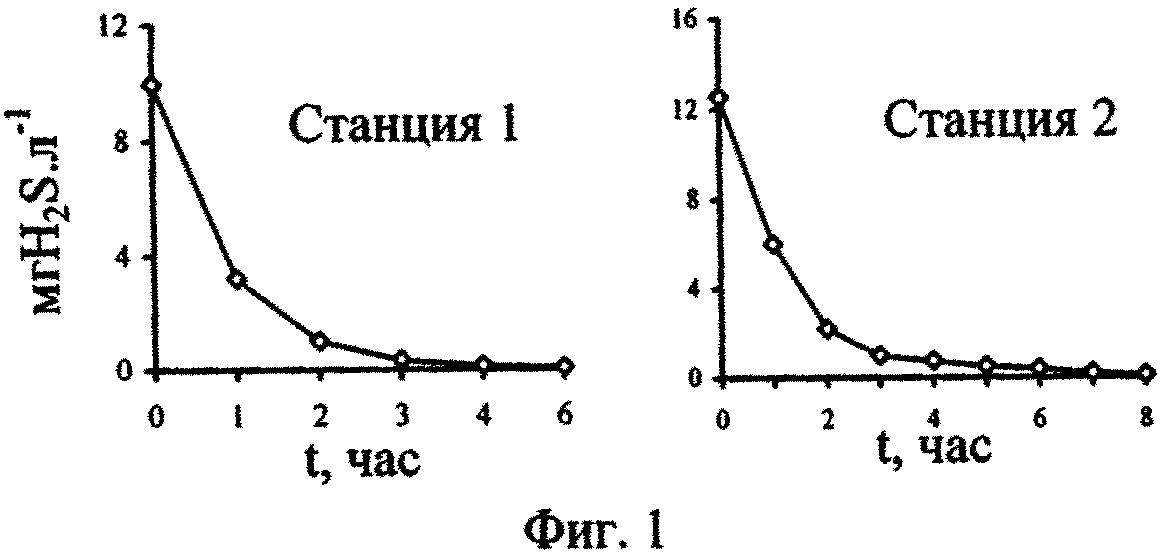
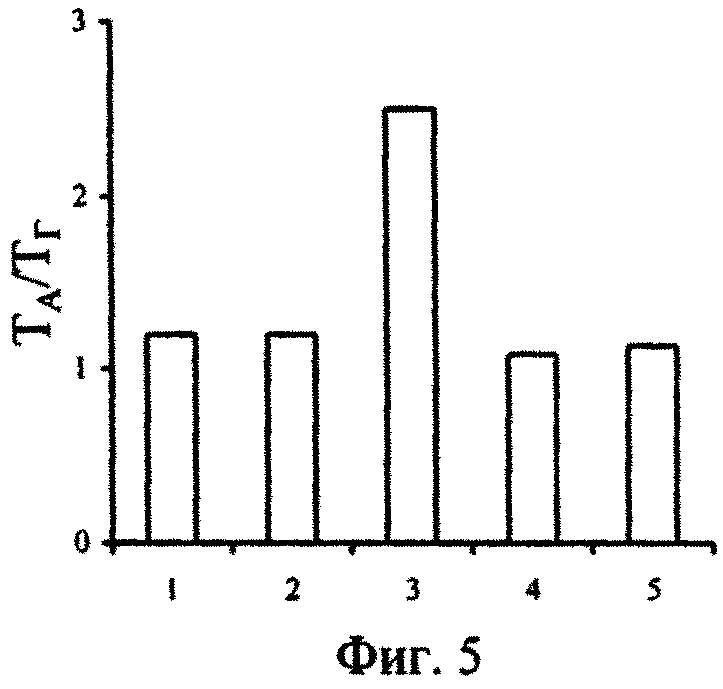
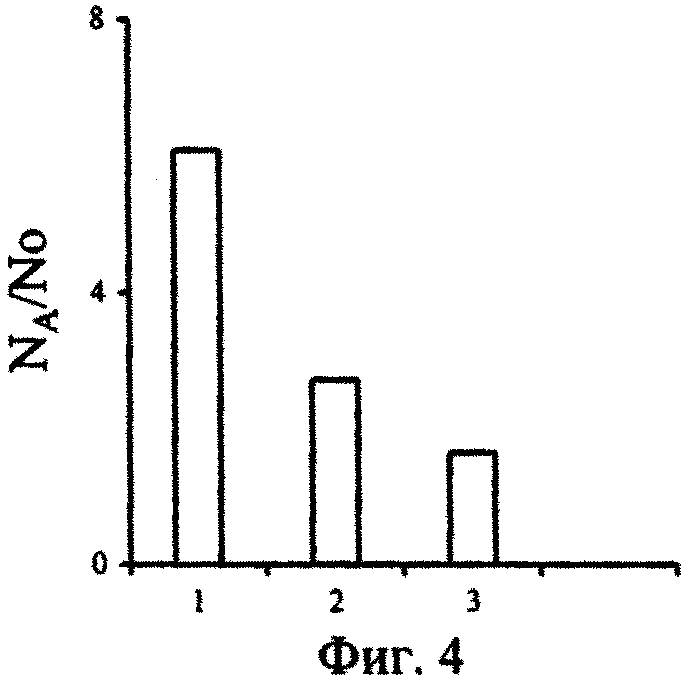
На фиг.1 - Изменение концентраций сероводорода в воде, поднятой с глубины 2000 м, при ее активной аэрации; фиг. 2 - Изменение численности P. micans, в поверхностной (1) и глубинной воде с разными исходными концентрациями сероводорода (2-5) при содержании в ней фосфатов 246 мкгР·л-1 (серия а) и 264 мкгР·л-1 (серия б); фиг. 3 - Изменение численности G. kovalevskii (а) и D. brightwellii (б) в зависимости от времени их культивирования в поверхностной (1) и глубинной воде (2) Черного моря; фиг. 4 - Отношение численности черноморских динофлагеллят в глубинной воде (NA) с начальной концентрацией сероводорода, равной 12,2 мгН2S·л-1, и поверхностной воде (No) через 17 сут от начала экспериментов; фиг. 5 - Отношение темпов деления черноморских одноклеточных водорослей в аэрированной воде, отобранной с глубины 2000 м (ТА), и среде Гольдберга (Тг); фиг. 6 - Изменение во времени Кн 32Р в ульве в глубинной (1) и поверхностной (2) воде; фиг.7 - Зависимость скорости поглощения (V) 32Р ульвой от концентрации фосфатов в воде, после предварительной адаптации ульвы в течение 72 часов в глубинной аэрированной (1) и в поверхностной воде (2); фиг. 8 - Зависимость Кн 32Р от времени для водоросли, культивируемой в глубинной воде (1), поверхностной воде (2) и поверхностной воде с добавкой минерального фосфора (3); фиг. 9 - Выведение Ρ из ульвы в среды сравнения: глубинную воду (1), поверхностную воду (2) и поверхностную черноморскую воду с добавкой минерального фосфора (3).
Выбор объектов культивирования связан с их ролью в экосистеме Черного моря. Так одноклеточные водоросли были выбраны как первичные продуценты органического вещества в море. Они являются звеном пищевой цепи для других объектов марикультуры и от их реакции на качество среды зависит функционирование более высоких уровней трофических цепей в экосистеме, а также они сами могут использоваться как ценные продукты марикультуры.
Выбор макроводоросли ульвы (Ulva rigida Ag.) для исследования глубинной воды обусловлен принадлежностью данного макрофита к мезосапробным гидробионтам, широко распространенных в прибрежных районах моря, рекомендованных для культивирования.
При исследовании реакции черноморских одноклеточных водорослей опыты проводили с альгологически чистыми культурами динофлагеллят (Prorocentrum micans Ehr., Peridinium trochoideum (Stein) Lammermann, Gymnodinium kovalevskii Pitz., Gymnodinium sp.) и диатомовых (Ditylum brightwellii (West), Coscinodiscus granii, Sceletonema costatum). Главным показателем их состояния были темпы деления и численность клеток. Испытуемой средой для них служила черноморская вода, отобранная в море с глубин 1000-2000 м. Средами сравнения были та же вода, активно аэрированная в течение 6-8 час до полного окисления в ней сероводорода, поверхностная вода, взятая на той же станции отбора, что и глубинная, а также среда Гольдберга, традиционно используемая в лабораторных условиях для культивирования одноклеточных водорослей.
Эксперименты с одноклеточными водорослями проводили в трех повторностях. В каждую склянку вносили равное исходное количество клеток исследуемых водорослей. Через промежутки времени 1-17 суток от начала экспериментов из каждой склянки отбирали по 0,1 мл суспензии клеток, в которой определяли их количество.
Для оценки возможного влияния глубинной окисленной воды на процессы жизнедеятельности макрофитов исследовали кинетику обмена фосфатов между ульвой и водной средой с помощью метода меченых атомов с использованием 32Р. Определяемыми параметрами кинетики фосфорного обмена ульвы со средой были: коэффициент накопления (Кн) 32Р, скорость поглощения минерального фосфора ульвой из среды (V, мегР·л-1·час-1), доля 32Р, выведенного из водоросли в водную среду.
Во всех опытах с ульвой использовали воду, поднятую с глубины 1000 м, которую перед началом экспериментов активно аэрировали в течение 6 час. Контроль состоял из двух вариантов, средой сравнения в которых была поверхностная вода с природным содержанием фосфатов, (вариант 1), и поверхностная вода с добавкой фосфатов в таком количестве, что в сумме концентрация фосфатов в среде была равной таковой в глубинной воде (вариант 2).
Выбор двойного контроля в опытах был обусловлен тем, что параметры кинетики обмена 32Р ульвой зависят от концентрации растворенного минерального фосфора в водной среде. Поэтому наличие второго контроля - поверхностной воды с добавкой фосфатов - позволяло исключить эту зависимость и выявить возможное влияние других гидрохимических компонентов окисленной глубинной воды на значение параметров кинетики фосфорного обмена между водорослью и водной средой.
Эксперименты с ульвой проводили на фрагментах, вырезанных из центральной части ее таллома, диаметр которых составил 15 мм, а исходная биомасса - 10 - 12 мг в расчете на сырую массу. В каждый из стеклянных сосудов емкостью 1-2 л помещали 10 таких фрагментов. Температура воды в аквариумах составляла 14 - 15°С.
Погрешность радиометрических измерений не превышала 5%. В каждой временной точке отбирали 8-10 фрагментов и 3-5 проб воды. Для каждой выборки в одной временной точке рассчитывали среднее значение и определяли доверительный интервал среднего значения с 95% надежностью. Во всех выборках ширина доверительного интервала не превышала 10% от значения среднего. Все эксперименты выполнены в трех повторностях.
Для определения концентраций сероводорода и фосфатов в поверхностной и глубинной воде использовали стандартные гидрохимические методики.
Результаты экспериментов.
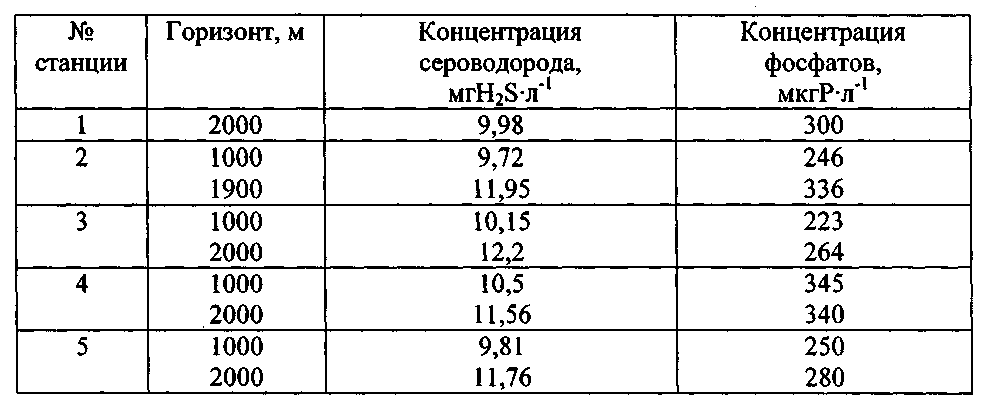
Таблица 1
Исходные концентрации сероводорода и фосфатов в черноморской воде, отобранной из восстановительной зоны Черного моря для экспериментов с водорослями
Различные начальные концентрации сероводорода в испытуемых средах получали, аэрируя глубинную воду в течение разного периода времени. Кривые, отражающие изменение концентраций сероводорода в воде, поднятой на разных станциях с горизонтов 2000 м, в зависимости от времени аэрации, представлены на фиг. 1.
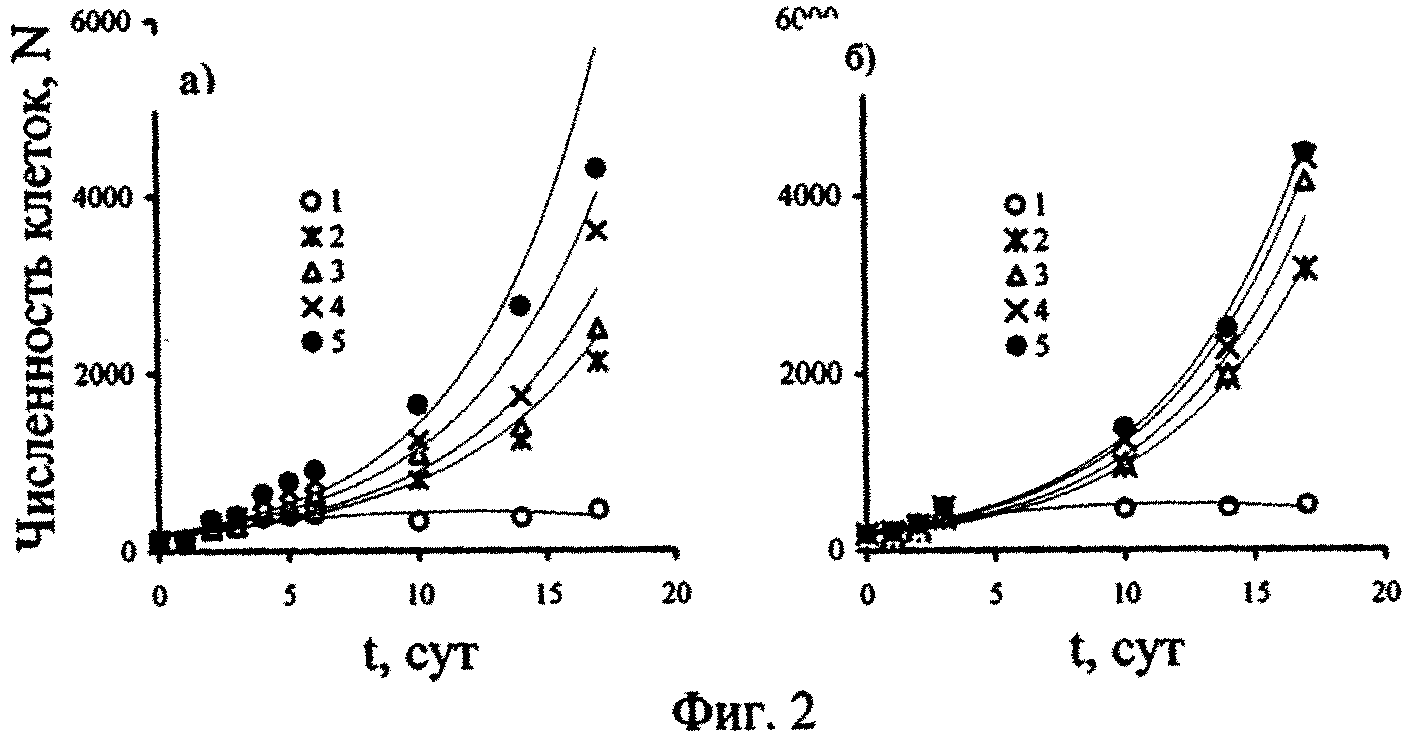
При изучении влияния черноморской воды, поднятой с глубин 1000-2000 м на исследуемые одноклеточные водоросли авторами установлено, что они адаптируются к действию изначально высоких исходных, а затем снижающихся концентраций сероводорода до его полного окисления в первые 3-6 дней от начала эксперимента. Вначале наблюдается затормаживающее действие этого ксенобиотика на жизнедеятельность исследованных одноклеточных водорослей, которое затем нивелируется достаточно быстрым восстановлением их клеточного деления, что продемонстрировано на примере данных, полученных для P. micans (фиг. 2). Эксперименты с P. micans состояли из двух серий, отличительной чертой которых были разные исходные концентрации минерального фосфора и сероводорода в глубинной воде, а контролем служила поверхностная вода с разной естественной концентрацией минерального фосфора (табл. 2).
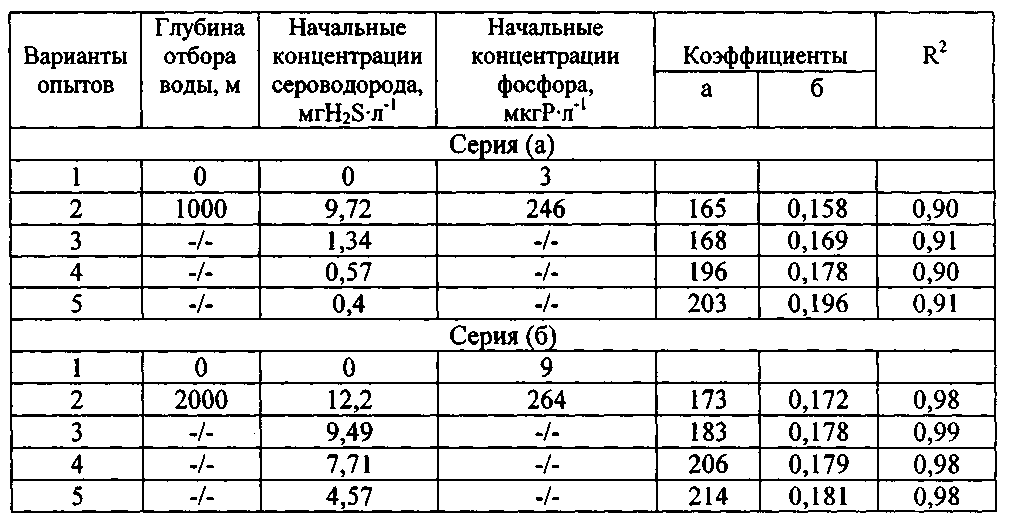
Таблица 2
Начальные концентрации сероводорода, минерального фосфора в испытуемой воде, а также численное значение коэффициентов в уравнении и достоверность аппроксимации
R2 экспоненциальных кривых на фиг. 2
Временные зависимости изменения численности клеток водоросли P. micans, представленные на фиг. 2, имеют экспоненциальный вид и описываются уравнением: N=a·ebt (1), где N - численность клеток P. micans, t - время культивирования водоросли в данной среде, а и b - коэффициенты.
Как видно из табл. 2, величина достоверности аппроксимации (R2) экспоненциальных кривых 2-5, построенных на фиг. 2, достаточно высока, изменяясь от 0,9 до 0,98. Из данных, приведенных на фиг. 2 и в табл. 2, можно заключить, что величина «а» в уравнении (1) соответствует исходному количеству клеток водоросли, которое необходимо ее популяции в начале эксперимента для достижения сопоставимого уровня выживания при разных начальных концентрациях сероводорода в среде.
Величина «b», определяя наклон кривых по отношению к оси ординат (численности клеток N), может быть оценена как средний темп деления клеток (средний уровень изменения численности клеток за единицу времени) при экспоненциальной временной зависимости этого показателя функционирования популяции. Этот коэффициент, скорее всего, отражает влияние разных исходных концентраций сероводорода в среде на темп деления клеток P. micans. Так, в опытах серии (а) этот показатель увеличивался от 0,158 (при исходной концентрации сероводорода в среде, равной 9,72 мгН2S·л-1) до 0,196 (при 0,4 мгН2S·л-1), т.е. почти на 24%. Для опытов серии (б) изменение коэффициента «b» было значительно меньше и составляло 4%. Такое различие может быть обусловлено, прежде всего, разницей в начальных концентрациях сероводорода в средах сравнения, варьируя от 9,72 до 0,4 мгН2S·л-1 в опытах серии (а) и от 12,2 до 4,57 мгН2S·л-1 в серии (б).
С другой стороны, для глубинной воды, испытуемой в обеих сериях экспериментов с P. micans, отмечена разница в содержании минерального фосфора, равная 18 мкгР·л-1 (табл. 2, фиг. 2). Поэтому, возможно, количество минерального фосфора в среде является вторым фактором, влияющим на численное значение коэффициента 'b».
Дополнительным, непосредственно неучтенным, в наших опытах фактором среды, влияние которого на популяцию исследуемой планктонной водоросли также отражается на коэффициенте «b», может быть содержание в испытуемой воде других микро- и макроэлементов или органических соединений, необходимых для оптимального развития популяции P. micans.
Кривые 1 (фиг. 2) а), б), полученные по результатам опытов с поверхностной водой, представляют собой полиномиальные линии, соответствующие зависимостям типа: N=-сх2+dx+к, где N - численность клеток в популяции, с и d - коэффициенты, определяющие наклон кривых по отношению к оси ординат, а величина k, по-видимому, соответствует тому исходному количеству клеток, которое необходимо учитывать в конкретных сравнительных экспериментах.
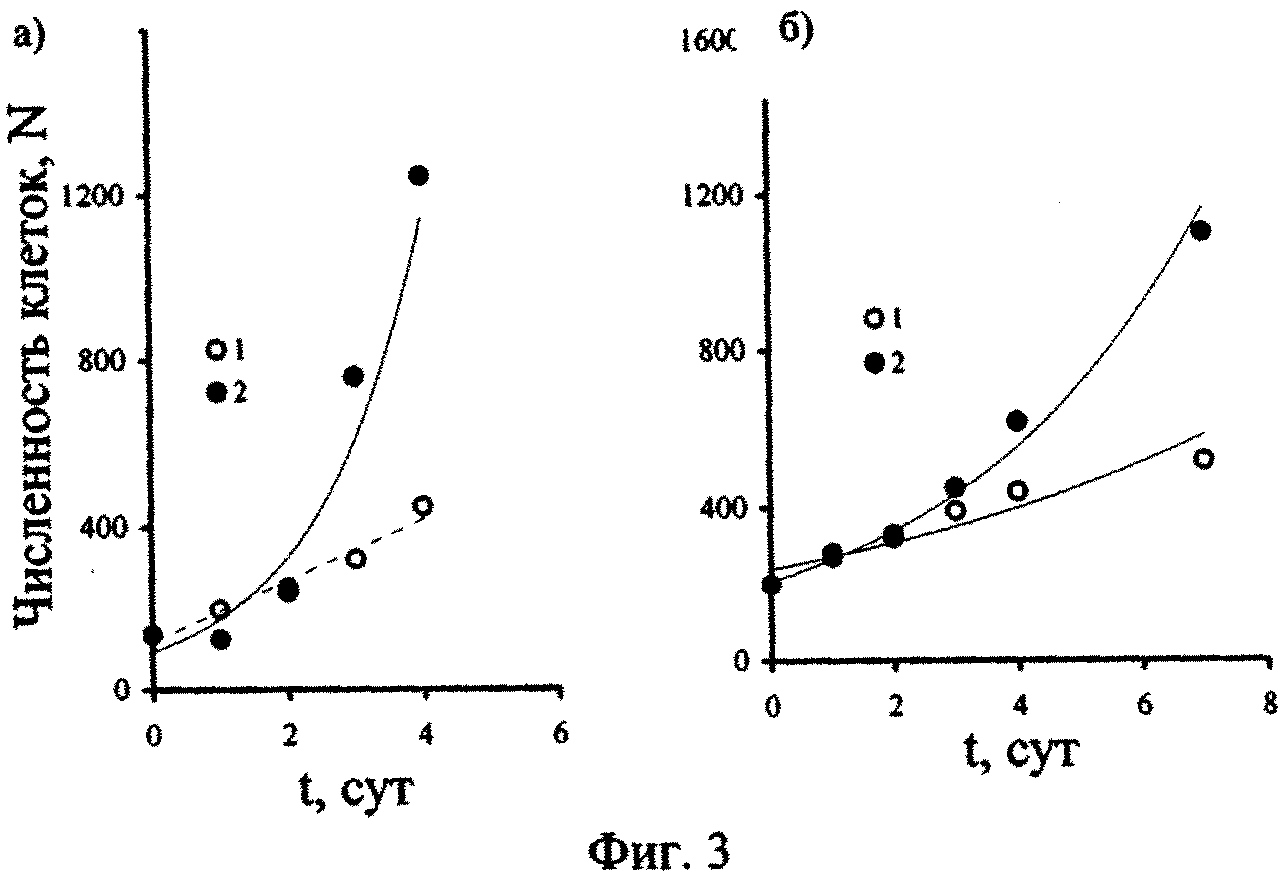
Результаты изучения отклика других видов черноморских планктонных водорослей, в частности, G. kovalevskii и D. brightwellii, на состав воды из восстановительной зоны моря представлены на фиг. 3, из которого следует, что уравнения, характеризующие изменения численности клеток в популяциях водорослей во времени, имеют экспоненциальный вид.
В опыте с глубинной водой при величине достоверности аппроксимации кривой R2=0,9, уравнение, описывающее изменение численности клеток G. kovalevskii со временем, имело вид: N=94e0,624t. Численность клеток этой же водоросли в поверхностной воде изменялась со временем также по экспоненциальному закону и выражена уравнением N=140e0,29t при величине R2=0,98.
Аналогично для D. brightwellii уравнения имели вид: N=207e0,247t при R2=0,98 (для варианта опыта с глубинной водой) и N=239e0,129t при R2=0,88 (для поверхностной воды).
Как видно, коэффициенты «а» и «b» в уравнениях, описывающих закономерности изменения численности популяций G. kovalevskii и D.brightwellii в зависимости от времени экспериментов, несут такую же смысловую нагрузку, как и в уравнениях для Р. micans.
Из представленных на фиг. 2, 3 данных видно, что адаптационный период для исследованных черноморских одноклеточных водорослей зависит от их видовой принадлежности, исходной концентрации сероводорода в среде и времени, в течение которого происходит естественное окисление ксенобиотика в опытных сосудах.
Отношение темпов деления черноморских динофлагеллят в глубинной и поверхностной воде (фиг. 4) свидетельствует о том, что, несмотря на высокие исходные концентрации сероводорода, глубинная вода не оказывает токсического действия на исследованные виды водорослей. Напротив, по мере окисления сероводорода в среде из восстановительной зоны Черного моря, численность и темпы деления клеток в ней выше, чем в поверхностной воде моря (фиг. 2-4).
Известно, что поверхностная вода Черного моря не содержит биогенные элементы в количествах, достаточных для длительного и полноценного культивирования одноклеточных водорослей. В первую очередь это относится к фосфору, концентрации которого в пробах воды, взятых в разных районах Черного моря для экспериментов с одноклеточными водорослями, были равны 3-9 мкгР·л-1. В то же время в воде с глубин 1000-2000 м, содержание фосфатов было выше в 30-130 раз.
Кроме фосфора, на численность одноклеточных водорослей в среде культивирования влияют и другие элементы. Поэтому в другой серии экспериментов качество окисленной глубинной воды из восстановительной зоны Черного моря сравнивали с, традиционно используемой в лабораторном культивировании микроводорослей, средой Гольдберга, основой приготовления которой служит поверхностная вода открытой части моря. Содержание основных элементов в приготовленной питательной среде Гольдберга выше, чем в глубинной воде. Исключение составил лишь марганец (табл. 3).
Таблица 3.
Концентрации важнейших химических элементов (мг-атом·л-1) в глубинной воде
Черного моря и среде Гольдберга.

Наши эксперименты показали, что по сравнению со средой Гольдберга, темпы деления и численность популяций черноморских микроводорослей в глубинной воде после полного окисления в ней сероводорода достоверно выше в воде из восстановительной зоны Черного моря (фиг. 5). Это обусловлено особенностями солевого, в том числе микроэлементного состава, характерного для воды из восстановительной зоны Черного моря, и ее благоприятное действие на микрофиты вызвано не только более широким спектром разнообразия химических элементов, но и природной сбалансированностью состава химических элементов и форм их физико-химического состояния, соотношением основных биогенных макроэлементов, а также наличием других биологически активных веществ, более доступных для потребления их водорослями из глубинной воды, которые положительно влияют на темп деления клеток.
Следовательно, вода из восстановительной зоны Черного моря даже при самых высоких природных начальных концентрациях в ней сероводорода не оказывает губительного действия на популяции черноморских одноклеточных водорослей. Более того, после полного окисления сероводорода, биологическое качество воды из восстановительной зоны Черного моря превышает таковое не только в поверхностной воде, но даже в культуральной питательной среде Гольдберга.
При исследовании кинетики накопления 32Р многоклеточной зеленой черноморской водорослью U. rigida Ag. в глубинной аэрированной воде, установлено, что, по сравнению с поверхностной водой (с концентрацией фосфатов, равной 3 мкгР·л-1 аккумулирование 32Р фрагментами таллома ульвы происходит более интенсивно в глубинной воде, содержание фосфатов в которой составляло 230 мкгР·л-1 (Фиг. 6). Изменение Кн 32Р ульвой во времени в течение первых 2,5 суток эксперимента описывается уравнением вида
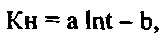
где а и b - коэффициенты.
В частности, для варианта в глубинной воде при величине достоверности аппроксимации, равной 0,96 (кривая 1) уравнение принимает вид:

а для варианта в поверхностной воде, при R=0,93, кривая 2 описывается уравнением
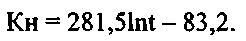
Как видно, численные значения коэффициентов «а» и «b» в уравнениях для кривых 1 и 2 (фиг.6), различались значительно. Так, для коэффициента «а» эта разница составляла 3,3 раза, а для коэффициента «b» - 12,5 раза.
К концу эксперимента, результаты которого представлены на фиг. 6, коэффициенты накопления меченого фосфора водорослью в глубинной воде почти в 6 раза выше, чем в поверхностной воде, что может быть обусловлено, как одной из причин, разным содержанием фосфатов в средах сравнения.
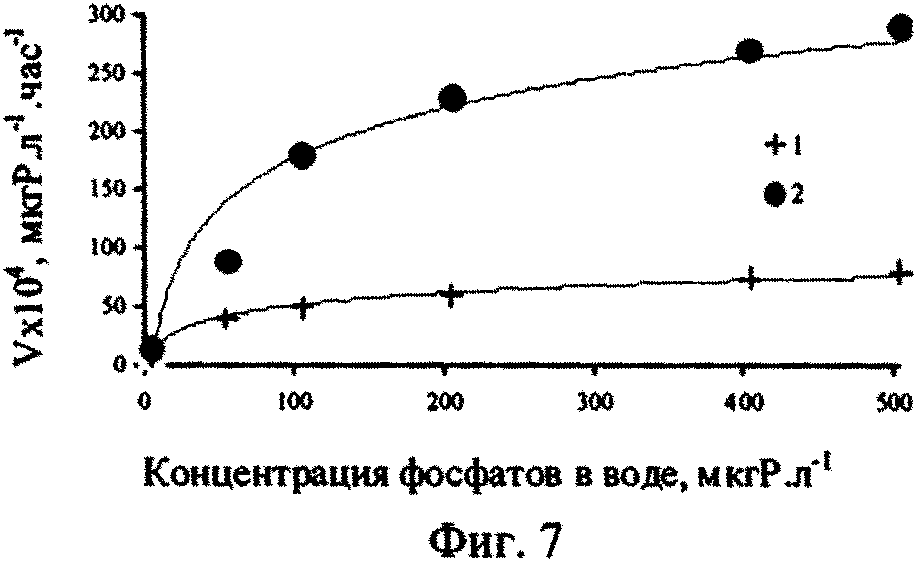
Для уточнения наличия возможных факторов, регулирующих фосфорный обмен ульвы, нами были проведены эксперименты по изучению влияния разных добавок минерального фосфора в поверхностной воде на скорость поглощения 32Р ульвой (фиг. 7). Перед началом опытов по поглощению Ρ водоросли выдерживали в сравниваемых средах (глубинная аэрированная и поверхностная вода) 72 часа с целью возможной оценки влияния преадаптации водорослей к разному количеству фосфатов в среде и формирования разного уровня внутриклеточного содержания фосфора в опытных образцах ульвы.
Из представленных на фиг. 7 результатов видно, что при добавках минерального фосфора к поверхностной воде, примерно до 10-20 мкгР·л-1, скорость поглощения 32Р ульвой практически не зависит от условий предварительной адаптации водоросли. При дальнейшем увеличении концентрации минерального фосфора, вносимого в черноморскую поверхностную воду в виде добавок фосфатов, скорость поглощения 32Р ульвой почти в 3,6 раза ниже для водорослей, проходивших преадаптацию в глубинной аэрированной воде (кривая 1), содержащей 230 мкгР·л-1, чем для водорослей, культивировавшихся перед началом эксперимента в поверхностной воде (кривая 2). Для варианта опыта, в котором в период адаптации водоросли культивировали в глубинной аэрированной воде (Фиг. 7, кривая 1), при величине достоверности аппроксимации, равной 0,94, зависимость скорости поглощения (V) ульвой 32Р от концентрации минерального фосфора в воде (С) имеет логарифмический вид:
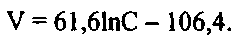
При величине достоверности аппроксимации, равной 0,98, зависимость скорости поглощения меченого фосфора водорослью от концентрации минерального фосфора в водной среде, для варианта адаптации в поверхностной воде (фиг. 7, кривая 2), может быть описана также уравнением аналогичного вида:
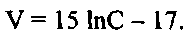
Как видно из фиг. 7, на процесс аккумулирования 32Р ульвой влияет количество минерального фосфора в водной среде и сформировавшаяся внутриклеточная концентрация фосфора в ульве.
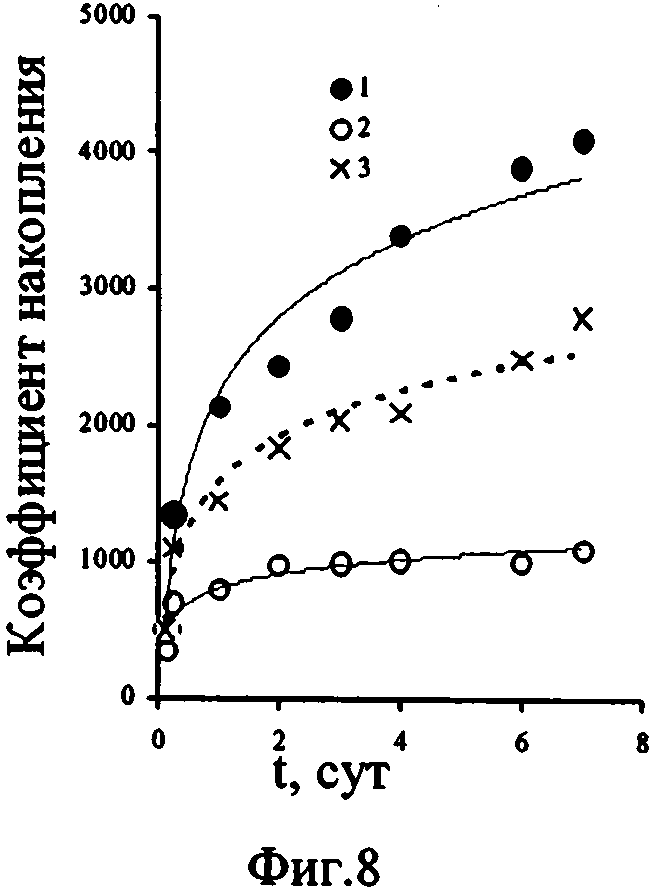
Кинетические зависимости Кн 32Р ульвой, полученные авторами в экспериментах с разными концентрациями минерального фосфора в воде, были предпосылкой для проведения сравнительного анализа кривых, описывающих накопление меченого фосфора в условиях, когда средами сравнения были глубинная аэрированная вода с концентрацией фосфатов, равной 230 мкгР·л-1, поверхностная вода, в которой фосфаты были определены на уровне 3 мкгР·л-1 и эта же поверхностная вода, в которую минеральный фосфор внесли в количестве, равном 227 мкгР·л-1, т.е. в сумме в данной среде концентрация фосфатов соответствовала таковой в глубинной воде (фиг. 8). Через семь суток после начала эксперимента Кн меченого фосфора составили следующие величины: в глубинной аэрированной воде - 4000, в поверхностной - 1080, в поверхностной с добавкой фосфатов - 2900 единиц. Следовательно, результаты, представленные на Фиг.8, свидетельствуют, что количество минерального фосфора в среде может выступать в роли фактора, регулирующего процесс аккумулирования меченого фосфора, но не является единственным параметром глубинной воды, определяющим интенсивность этого процесса.
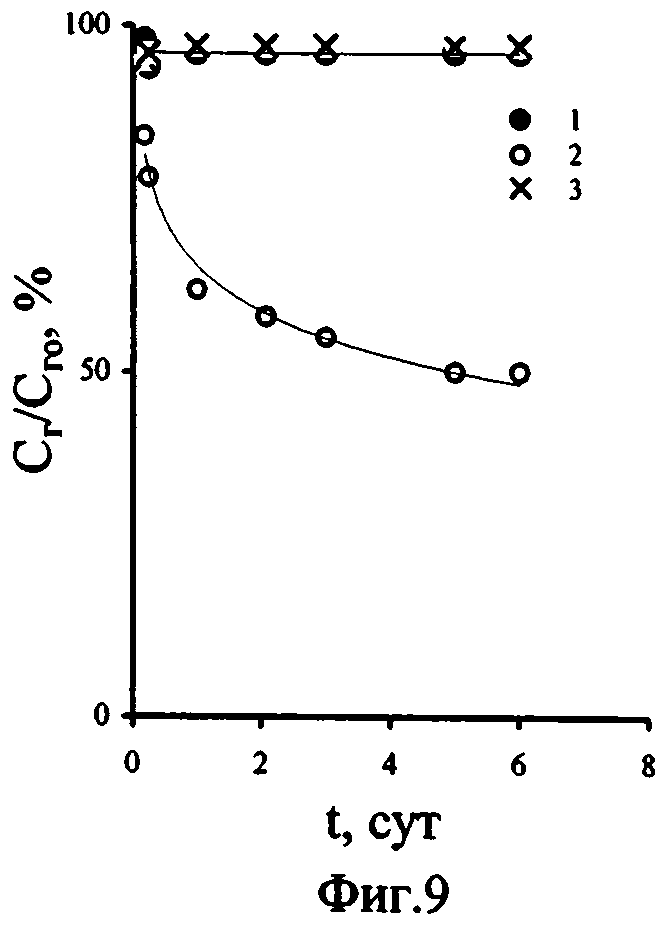
Влияние содержания фосфатов в воде на процессы фосфорного обмена ульвы подтверждают и опыты по изучению выведения 32Р водорослями. Анализируя кривые выведения 32Ρ ульвой (фиг.9), можно сделать вывод о том, что меченый фосфор, накопленный ульвой в средах с концентрацией минерального фосфора в воде, равной 230 мкгР·л-1 (кривые 1, 3), отражает преимущественное поступление меченного фосфора в необменные фонды фосфора, используемые ульвой на рост и продукцию именно в условиях более высоких концентраций фосфатов в среде, как в поверхностной, с добавкой фосфатов, так и в глубинной аэрированной воде. При этом кинетика выведения фосфатов в естественной поверхностной (без добавок фосфора - кривая 2) и глубинной черноморской воде (кривая 3) достоверно различается. Как видно из представленных графиков (фиг.9), в поверхностной воде доля выводимого на третьи сутки 32Ρ достигает 50% и в дальнейшем увеличивается, в то время, как в глубинной воде за это же время доля выведенного фосфора составляет лишь 6 % от исходного количества аккумулированного водорослями меченого фосфора и с течением времени остается на этом же уровне. Выведение 32Р в глубинной и поверхностной воде с добавкой фосфатов, в которых концентрации фосфора уравнены, не различается. Таким образом, для процессов выведения меченного фосфора ульвой концентрация фосфатов в водной среде играет ведущую роль.
Величины же Кн 32Р достоверно различаются в средах сравнения (фиг. 8), несмотря на одинаковые условия по содержанию фосфатов в воде (кривые 1, 3). И хотя в поверхностной воде с добавкой фосфатов Кн 32Р водорослями выше (кривая 3), чем в поверхностной воде с природным содержанием фосфатов (кривая 2), самые высокие Кн 32Р макрофитами наблюдали в глубинной воде. Вероятно, величина Кн 32Р ульвой обусловлена не только повышенным содержанием фосфора в воде, но и более интенсивным поступлением фосфора в необменные фонды водоросли, что, в свою очередь, может быть вызвано благоприятным влиянием на фосфорный обмен ульвы других гидрохимических компонентов глубинных вод. Это может быть как их повышенное содержание по сравнению с поверхностными водами, так и различие их форм нахождения в глубинной и поверхностной воде. В роли таких компонентов могут выступать микроэлементы, играющие важную роль в метаболизме водорослей (например, марганец) и биогенные макроэлементы (в частности, например, азот), находящиеся в формах доступных для усвоения водорослями.
Поэтому, хотя количество неорганического фосфора в воде является одним из основных факторов, влияющих на процессы обмена фосфора между ульвой и водной средой., исходный уровень содержания фосфора в ульве и другие гидрохимические компоненты водной среды из восстановительной зоны моря могут вносить свой вклад в регуляцию этих процессов.
Результаты исследования глубинной воды Черного моря показывают, что на процессы жизнедеятельности черноморской многоклеточной водоросли U. xigida Ag. естественная глубинная вода, после ее аэрации, не оказывает угнетающего действия.
Кроме того, можно сделать вывод о том, что повышенное количество фосфатов, присутствующих в глубинной воде, в значительной степени определяя кинетику обмена фосфора между ульвой и средой, не является единственным благоприятным фактором, влияющим на процесс аккумулирования минерального 32Р зеленой водорослью U. rigida Ag.
Результаты исследования биогенных свойств водной среды из восстановительной зоны Черного моря показали, что глубинная вода не оказывает губительного действия на черноморские планктонные водоросли в присутствии высоких исходных концентраций сероводорода. После полного окисления этого ксенобиотика черноморская глубинная вода может применяться в качестве питательной плодородной среды культивировании одноклеточных планктонных и многоклеточных бентосных водорослей в лабораторных или промышленных условиях и в марикультурных хозяйствах.
Способ мелиорации прибрежных экосистем
Устройство для культивирования макрофитов
Способ культивирования одноклеточной зеленой микроводоросли dunaliella salina для получения биомассы
Способ определения объемов и площадей поверхностей клеток диатомовых водорослей
Способ определения концентрации тория-234 в морской воде
Способ определения концентрации тория-234 в морских донных отложениях
Способ биологической доочистки сточных вод
Способ длительного хранения микроводорослей
Устройство для сбора количественных проб зоо- и ихтиопланктона
Питательная среда для культивирования штамма возбудителя рожи свиней erysipelothrix rhuisipathie
Способ мелиорации прибрежных экосистем
Устройство для культивирования макрофитов
Способ культивирования одноклеточной зеленой микроводоросли dunaliella salina для получения биомассы
Способ определения объемов и площадей поверхностей клеток диатомовых водорослей
Способ определения концентрации тория-234 в морской воде
Способ определения концентрации тория-234 в морских донных отложениях
Способ биологической доочистки сточных вод
Способ длительного хранения микроводорослей
Устройство для сбора количественных проб зоо- и ихтиопланктона
Питательная среда для культивирования штамма возбудителя рожи свиней erysipelothrix rhuisipathie

