Результат интеллектуальной деятельности: НОВЫЕ БЕЛКОВЫЕ КОНСТРУКЦИИ ДЛЯ ДИАГНОСТИКИ И ТЕРАПИИ GD2-ПОЗИТИВНЫХ ЗАБОЛЕВАНИЙ
Вид РИД
Изобретение
Область техники
Изобретение относится к биотехнологии и может быть использовано для получения конструкций на основе антител к опухолеассоциированному ганглиозиду GD2. Данные антитела и их фрагменты могут найти применение в медицине, иммунологии и биотехнологии для приготовления диагностических реагентов и терапевтических препаратов для лечения GD2-позитивных онкологических заболеваний.
Уровень техники
Ганглиозиды - универсальные компоненты мембран эукариотических клеток, известные как важные регуляторы и медиаторы межклеточных взаимодействий и иммунного ответа. В последнее время активно изучается их роль в опухолевом процессе. В трансформированных клетках появляются ганглиозиды, которые в нормальных клетках либо отсутствуют, либо экспрессированы в ограниченном количестве [Hakomori S. ,. Adv Cancer Res., 52, р. 257-331, 1986]. Наиболее ярким опухолеспецифическим ганглиозидом является GD2, маркер опухолей различного происхождения. Дисиалоганглиозид GD2 - это гликосфинголипид, содержащий два остатка сиаловой кислоты, экспрессируемый преимущественно на клеточной поверхности опухолевых клеток. Экспрессия GD2 в нормальных фетальных и взрослых тканях преимущественно ограничивается центральной нервной системой, периферическими нервами и меланоцитами кожи [Zhang S., et al., Int. Selection of tumor antigens as targets for immune attack using immunohistochemistry: I. Focus on gangliosides. J. Cancer, Sep 26;73(1):42-9., 1997]. В злокачественных новообразованиях описана повышенная экспрессия GD2 на клетках нейробластомы, глиомы, меланомы, мелкоклеточного рака легких, различных сарком и ряде других опухолей, с трудом поддающихся классическим методам лечения [F. Navid, V.M. Santana and R.C. Barfield., Anti-GD2 antibody therapy for GD2-expressing tumors. Current Cancer Drug Targets, Mar; 10(2):200-9, 2010]. Функциональные свойства ганглиозида GD2 изучены не до конца, однако точно известно, что он играет важную роль в адгезии опухолевых клеток на внеклеточном матриксе, а также усиливает пролиферацию и инвазивность опухолевых клеток [Shibuya, H., et al., Enhancement of malignant properties of human osteosarcoma cells with disialyl gangliosides GD2/GD3. Cancer Sci, 2012. 103(9): p. 1656-64.]. Кроме того, одним из важных механизмов иммуносупрессии у онкологических больных являются процессы, вызываемые свободными ганглиозидами, которые сбрасываются в кровь. Шеддинг опухолевых ганглиозидов с клеточной поверхности показан для большого числа опухолевых клеток, включая клетки нейробластомы, лимфомы, меланомы, лейкемии, опухолевые клетки мозга также сбрасывают ганглиозиды [Lauc G, Heffer-Lauc M., Shedding and uptake of gangliosides and glycosylphosphatidylinositol-anchored proteins. Biochim Biophys Acta., 1760(4):584-602. Epub 2005 Dec 20. 2006]. Биологическая роль шеддинга опухолевых ганглиозидов достаточно подробно изучена. Опухолевые ганглиозиды, свободно циркулирующие в крови, ингибируют некоторые иммунные ответы как in vivo, так и in vitro [Li R., Villacreses N., Ladisch S., Human tumor gangliosides inhibit murine immune responses in vivo., Cancer Res., 55(2):211-4., 1995]. Их сброс с поверхности трансформированных клеток стимулирует развитие опухолей у мышей и у человека. Благодаря его избирательной экспрессии на поверхности опухолевых клеток GD2 является привлекательной мишенью для таргетной иммунотерапии с применением моноклональных GD2-специфичных антител, кроме того они могут быть использованы для диагностического определения ганглиозида GD2 в сыворотке крови онкологических больных.
Основной проблемой в создании эффективных препаратов для иммунотерапии GD2-позитивных опухолей является получение антител, обладающих высокой аффинностью и специфичностью к ганглиозиду GD2. Известно, что иммунизация чистыми ганглиозидами индуцирует очень низкий титр ганглиозид-специфичных антител с очень коротким периодом полужизни. Как правило специфичность получаемых GD2-специфичных антител является низкой, поскольку имеется сильная структурная гомология между различными ганглиозидами, вследствие чего наблюдается значительная кроссреактивность разрабатываемых GD2-специфичных антител.
Большинство GD2-позитивных новообразований представляют собой солидные опухоли, для которых характерна невысокая противоопухолевая эффективность моноклональных антител, обусловленная большими размерами этих молекул. Кроме того, нежелательные побочные эффекты терапии зачастую обусловлены наличием Fc- опосредованной цитотоксичности. Поэтому, в связи с общим ростом распространенности различных типов онкологических заболеваний в России и в мире, и возникновении резистентности к имеющимся лекарствам, имеется существенная потребность в создании новых лекарств, и лекарств нового формата, на основе антител к GD2. Данное изобретение расширяет круг имеющихся кандидатов для лечения GD2-позитивных онкологических заболеваний.
Сущность изобретения
Задачей настоящего изобретения является создание терапевтического агента на основе антител к GD2 и разработка нового эффективного и безопасного способа лечения пациентов с онкологическими заболеваниями, характеризующимися присутствием ганглиозида GD2 на поверхности клеток опухоли. Также задачей настоящего изобретения является расширение арсенала средств для подавления пролиферации и индукции гибели О02-позитивных опухолевых клеток, а также для диагностики ганглиозида GD2 в сыворотке крови пациентов с С02-позитивными опухолями.
Указанные задачи решаются путем создания нового терапевтического агента для лечения онкологических заболеваний, представляющего собой антитело или его антигенсвязывающий фрагмент, специфически связывающее ганглиозид GD2, включающее тяжёлую цепь, содержащую H-CDR1 с последовательностью SEQ ID N0:1, H-CDR2 с последовательностью SEQ ID N0:2, и H-CDR3 с SEQ ID N0:3, и лёгкую цепь, содержащую L-CDR1 с последовательностью SEQ ID N0:4, L-CDR2 с последовательностью SEQ ID N0:5, и L-CDR3 с SEQ ID N0:6.
В некоторых вариантах изобретения данное собой антитело или его антигенсвязывающий фрагмент характеризуется тем, что тяжёлая цепь содержит вариабельный домен с последовательностью, гомологичной по меньшей мере на 95% последовательности SEQ ID N0:7. В некоторых вариантах изобретения данное антитело или его антигенсвязывающий фрагмент характеризуется тем, что лёгкая цепь содержит вариабельный домен с последовательностью, гомологичной по меньшей мере на 95% последовательности SEQ ID N0:8. В некоторых вариантах изобретения данное антитело представляет собой химерное антитело.
Указанная задача также решается путем создания мультивалентной конструкции, специфически связывающей ганглиозид GD2, и содержащей два, три или четыре описанных выше функциональных фрагмента антител, специфически связывающих ганглиозид GD2 и соединенных между собой при помощи ковалентной конъюгации с молекулой полиэтиленгликоля. В некоторых вариантах изобретения данная мультивалентная конструкция характеризуется тем, что содержит антигенсвязывающий фрагмент антитела с последовательностью, гомологичной по меньшей мере на 95% последовательности SEQ ID N0:9 или последовательности SEQ ID N0:10.
Указанные антитела или их функциональные фрагменты, а также указанные мультивалентные конструкции, содержащие такие антитела или их функциональные фрагменты, можно применять для лечения или диагностики С02-специфичных опухолей, таких как, например, нейробластома. Другими примерами онкологических заболеваний, характеризующихся увеличенной экспрессией ганглиозида GD2 на поверхности опухолевых клеток, являются мелкоклеточный рак легкого, саркома, глиома, ретинобластома и меланома. Диагностику подобных опухолей можно проводить как in vivo, как и по срезам тканей (ex vivo), а также с помощью других, известных специалистам методов.
При осуществлении изобретения достигаются следующие технические результаты:
- создан новый вариант терапевтического агента на основе фрагментов антител к GD2 для лечения онкологических заболеваний, характеризующихся присутствием ганглиозида GD2 на поверхности клеток опухоли;
- разработан новый эффективный способ уничтожения клетки в организме субъекта, на поверхности которой находится ганглиозид GD2;
- разработан новый способ диагностики онкологических заболеваний, характеризующихся присутствием ганглиозида GD2 на поверхности клеток опухоли.
Краткое описание рисунков
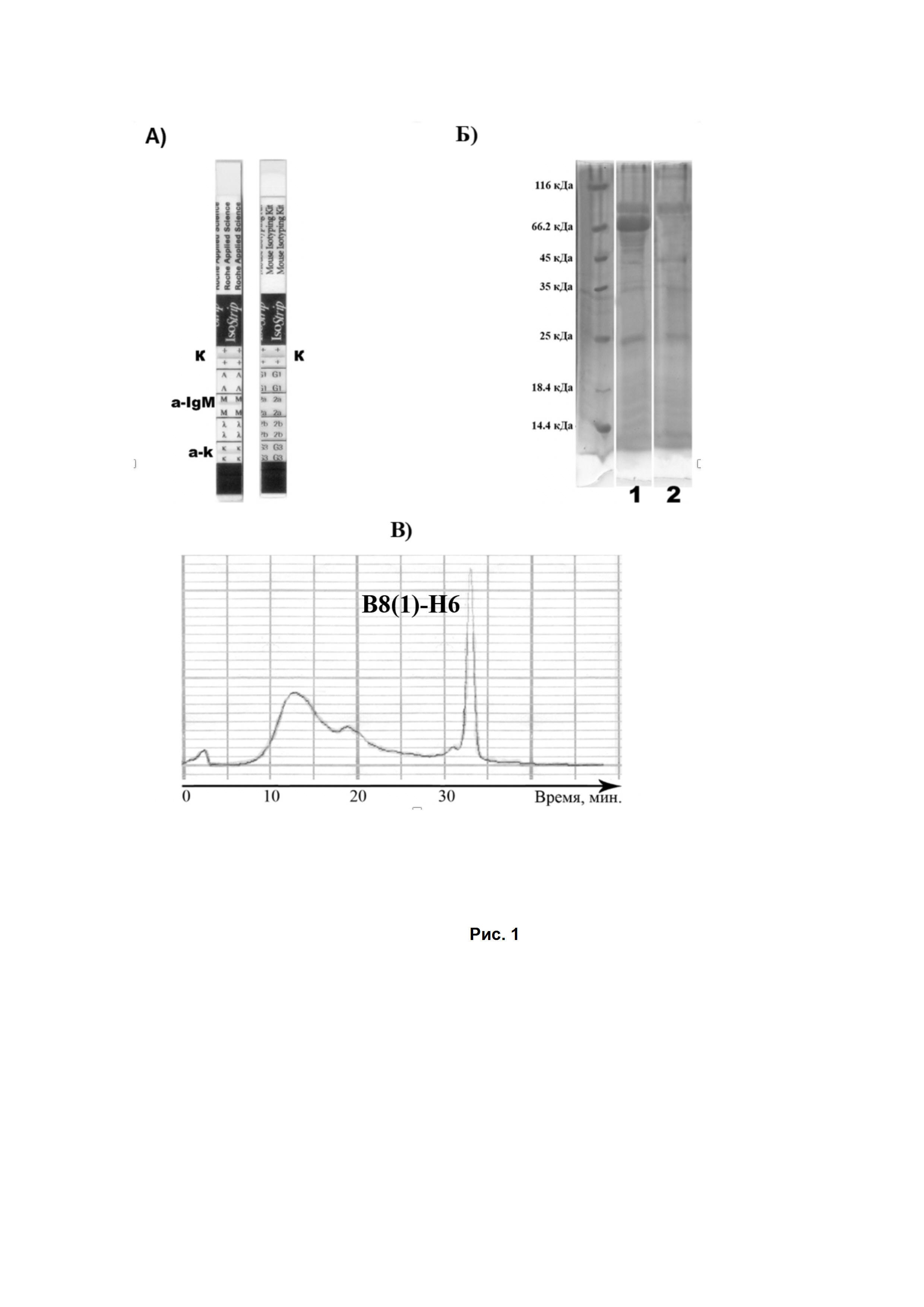
Рис. 1. (А). Результаты изотипирования при помощи IsoStrip Isotyping Kit. (Б). ПААГ-электрофорез в 10% геле антител B8(1)-H6 в восстанавливающих условиях: 1) Фракция антител B8(1)-H6 после очистки на Protein A колонке. 2) Фракция антител B8(1)-H6 после последующей очистки на HPLC. (В). Аналитическая гель-фильтрационная хроматография антител B8(1)-H6.
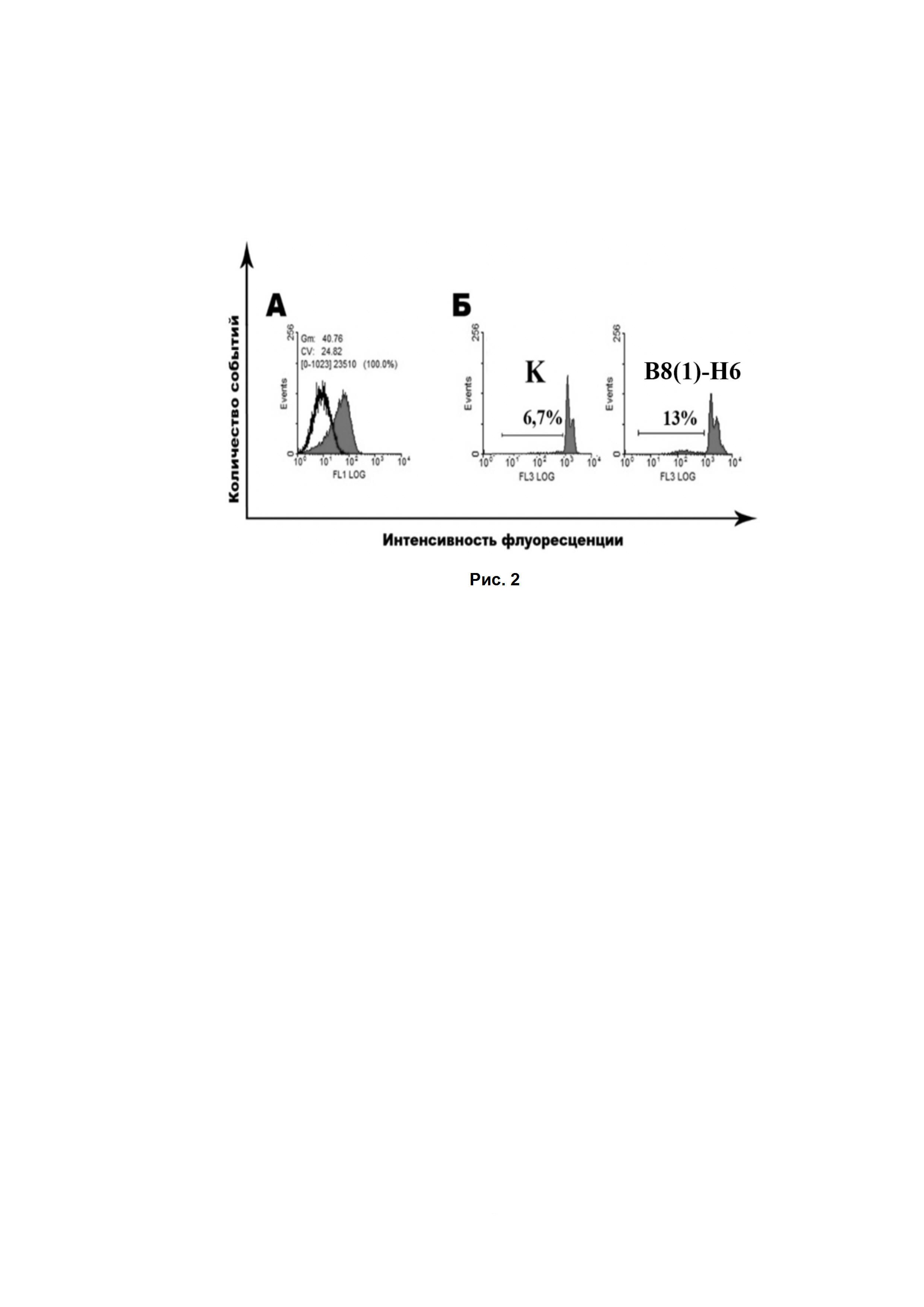
Рис. 2. (А) Цитометрические измерения клеток EL-4, окрашенных GD2-специфичными антителами B8(1)-H6, вторые антитела anti-mouse fab-specific-FITC (Титр 1:2000). Пик, обозначенный цветом, отражает флуоресценцию клеток, окрашенных GD2-мАт, контур - пик контрольных клеток, окрашенных вторичными антивидовыми антителами. Представлены результаты репрезентативного эксперимента. Было проведено как минимум 3 эксперимента. (Б) PI-тест. Анализ уровня фрагментации ДНК клеток GD2-позитивной клеточной линий EL-4 после инкубации в течение 24 ч с GD2-специфичными антителами B8(1)-H6 (500 тыс. клеток /образец, 20 мкг/мл). Маркером обозначена доля клеток с фрагментированной ДНК. Представлены результаты репрезентативного эксперимента. Было проведено как минимум 3 эксперимента.
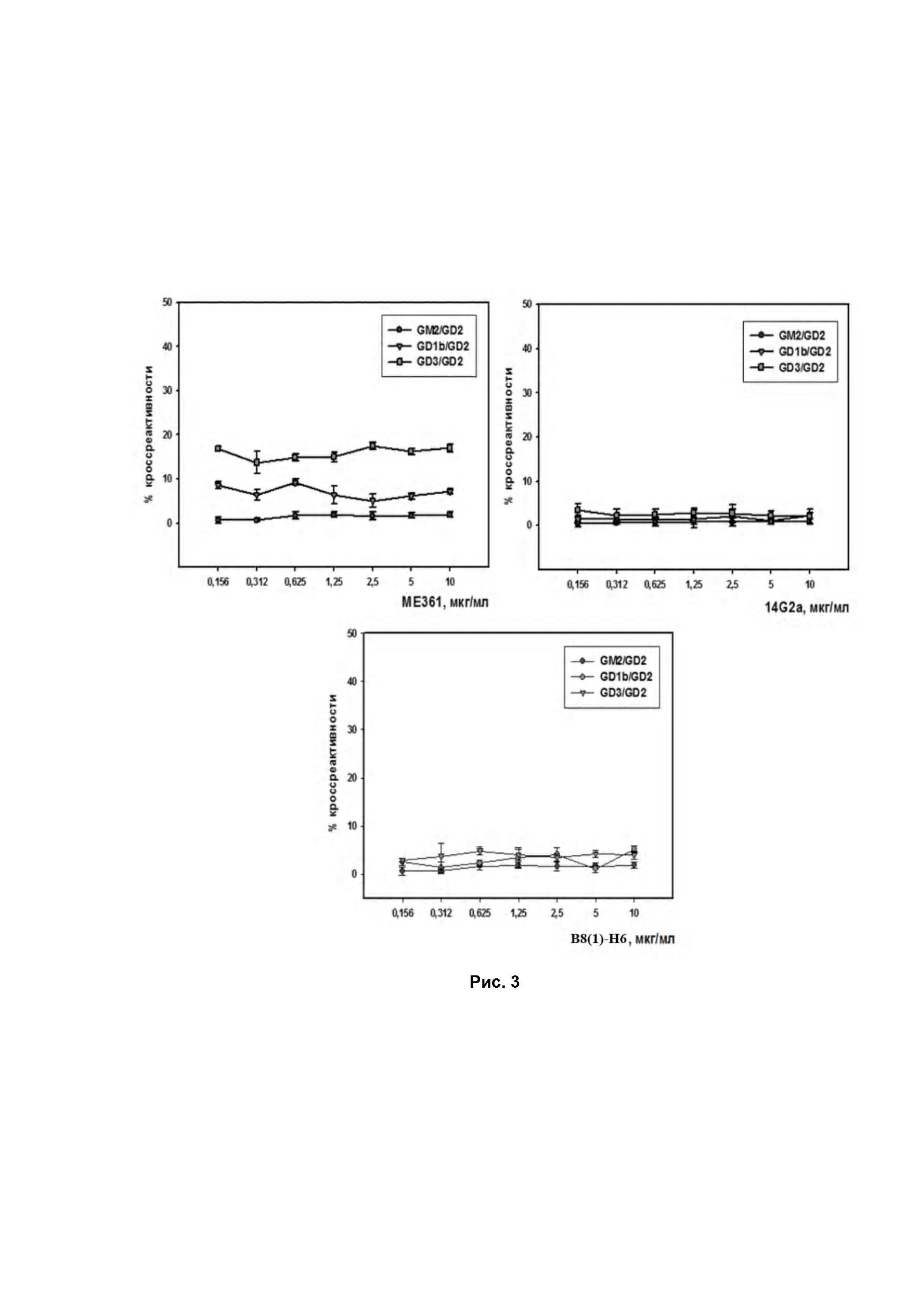
Рис. 3. Оценка кросс-реактивности различных GD2-специфичных антител с ганглиозидами GM2, GD1b и GD3 по результатам ИФА. Наносили различные разведения антител 14G2a, ME361 и B8(1)-H6. Антивидовые HRP-меченые анти-Fab-специфичные антитела (1:6000). На подложке 250 нг/лунка различных ганглиозидов. % кросс-реактивности рассчитывался по отношению OD раствора пероксидазной реакции после связывания антител с ганглиозидами GM2, GD1b или GD3 к OD раствора после связывания антител с ганглиозидом GD2.
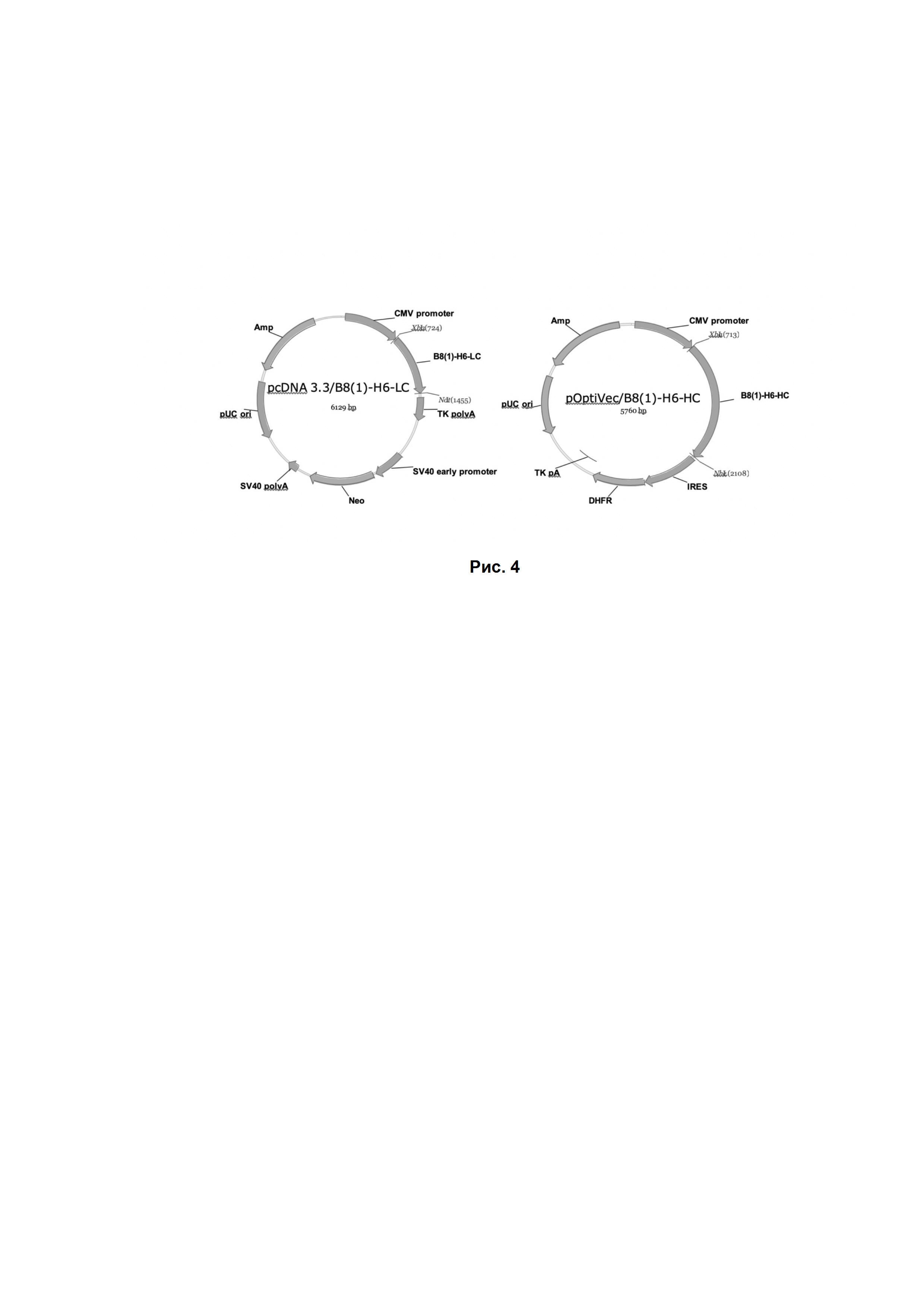
Рис. 4. Генетические конструкции, использованные при экспрессии химерных антител. Химерная легкая и тяжелая цепи антитела B8(1)-H6 находятся под контролем промотера цитомегаловируса (CMV promoter) и снабжены сигналами полиаденилирования (TK polyA). В качестве маркеров для селекции используются ген устойчивости к неомицину и дигидрофолатредуктаза, соответственно.
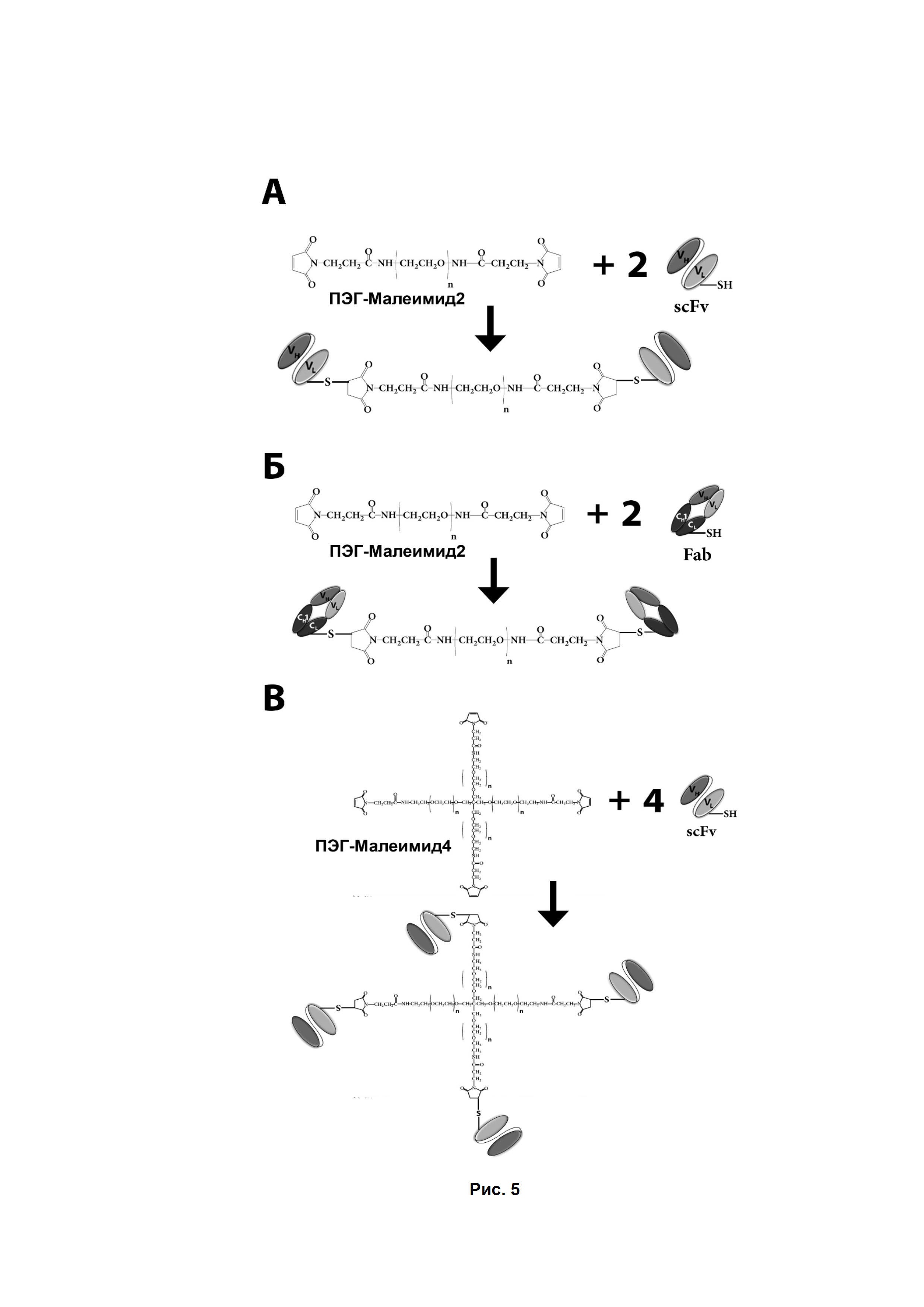
Рис. 5. Схема получения мультимерных фрагментов GD2-специфичных антител.
А - получение димеров scFv-фрагментов с использованием ПЭГ-Малеимид2.
Б - получение димеров Fab-фрагментов с использованием ПЭГ-Малеимид2.
В - получение тетрамеров scFv-фрагментов с использованием ПЭГ-Малеимид4.
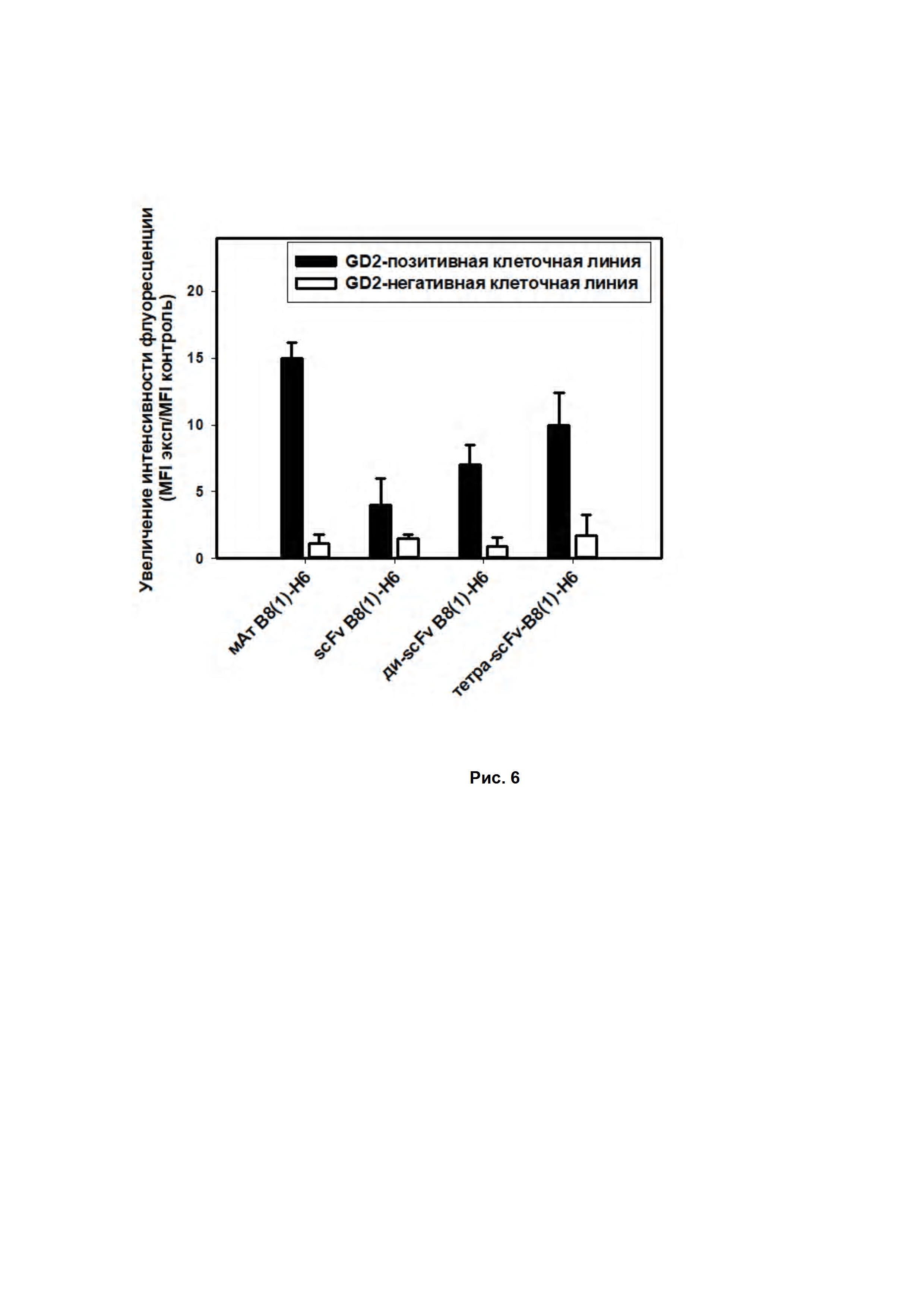
Рис. 6. Анализ антиген-связывающей способности молекул на основе антител B8(1)-H6 методом проточной цитофлуориметрии. GD2-позитивная клеточная линия - клетки мышиной лимфомы EL-4, GD2-негативная клеточная линия - клетки человеческой нейробластомы NGP-127.
Рис. 7. Анализ антиген-связывающей способности молекул на основе антител B8(1)-H6 методом ИФА с использованием ганглиозидов GD2, GD3 и GD1b, сорбированных на подложке планшета.
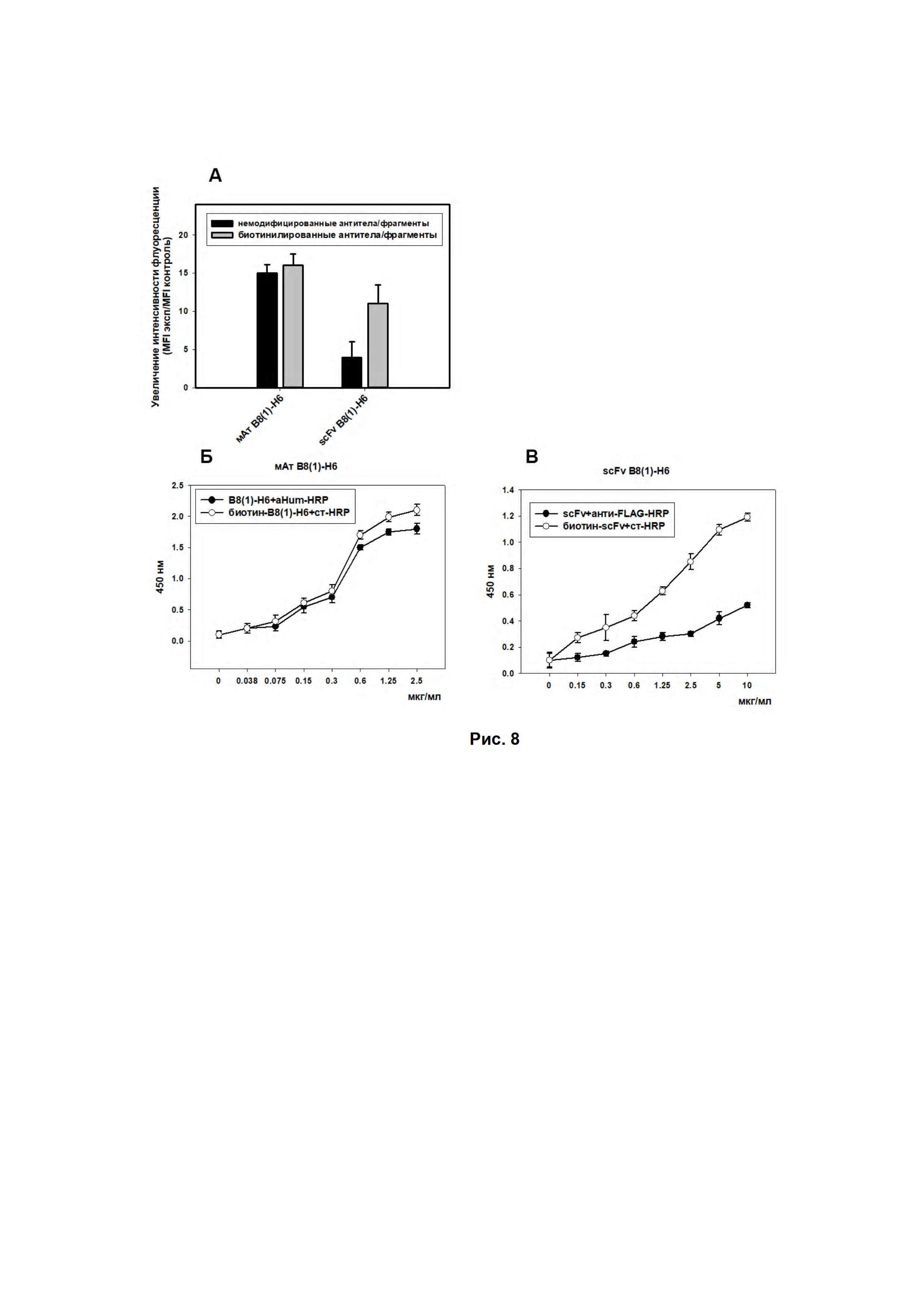
Рис. 8. Оценка связывания биотинилированных антител/фрагментов антител B8(1)-H6 с ганглиозидом GD2. (А) - Цитометрические измерения клеток EL-4, окрашенных антителами/фрагментами B8(1)-H6 или их биотинилированными производными, вторичные антитела anti-human Fc-specific-FITC в случае антител или anti-FLAG-FITC в случае scFv-фрагментов, детекцию биотинилированных антител/фрагментов проводили с использованием стрептавидин-FITC. Представлены результаты репрезентативного эксперимента. Было проведено как минимум 3 эксперимента. (Б) и (В) - Оценка связывания антител/фрагментов антител B8(1)-H6 и их биотинилированных производных с ганглиозидом GD2 в прямом ИФА. Различные концентрации GD2-связывающих молекул, вторичные антитела anti-human Fc-specific-HRP в случае антител или anti-FLAG-HRP в случае scFv-фрагментов, детекцию биотинилированных антител/фрагментов проводили с использованием стрептавидин-HRP, титр вторых антител и стрептавидин-HRP 1:4000. На подложке ганглиозид GD2 (250 нг/лунка). Каждая точка в трех повторах.
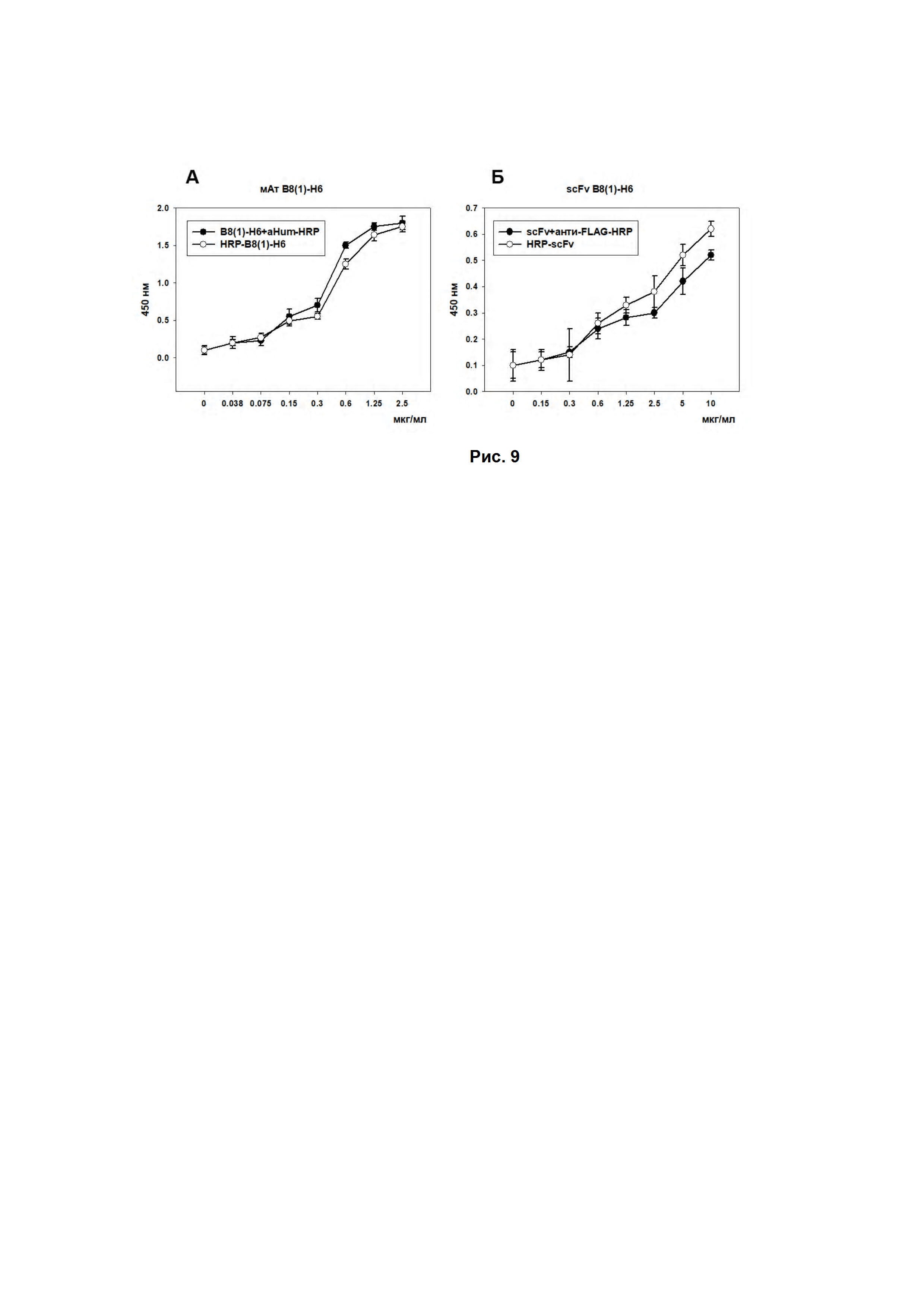
Рис. 9. (А) - Оценка связывания антител B8(1)-H6 и антител B8(1)-H6, меченных пероксидазой хрена, с ганглиозидом GD2 в прямом ИФА. Различные концентрации GD2-специфичных антител, вторичные антитела anti-human Fc-specific-HRP для немеченых антител B8(1)-H6, титр вторых антител 1:4000. На подложке ганглиозид GD2 (250 нг/лунка). Каждая точка в трех повторах. (Б) - Оценка связывания scFv-фрагментов антител B8(1)-H6 и scFv-фрагментов антител B8(1)-H6, меченных пероксидазой хрена, с ганглиозидом GD2 в прямом ИФА. Различные концентрации фрагментов GD2-специфичных антител, вторичные антитела anti-FLAG-HRP для немеченных фрагментов антител B8(1)-H6, титр вторых антител 1:4000. На подложке ганглиозид GD2 (250 нг/лунка). Каждая точка в трех повторах.
Рис. 10. Дот-блот анализ ганглиозида GD2 в сыворотках крови, сорбированных на нитроцеллюлозной мембране. (А). Инкубация с немечеными антителами B8(1)-H6 с последующей инкубацией с антителами anti-human Fc-specific-HRP (титр 1:6000); (Б). Инкубация с биотинилированными антителами B8(1)-H6 с последующей инкубацией с стрептавидин-HRP; (В). Инкубация с антителами B8(1)-H6, меченными пероксидазой хрена. 1 - контрольная сыворотка здорового донора. 2 - экспериментальная сыворотка с экзогенно добавленным ганглиозидом GD2. 3 - сыворотка крови пациента с нейробластомой.
Рис. 11. ИФА анализ ганглиозида GD2 в сыворотках крови. А. Инкубация с немечеными антителами B8(1)-H6 с последующей инкубацией с антителами anti-human Fc-specific-HRP (титр 1:6000); Б. Инкубация с биотинилированными антителами B8(1)-H6 с последующей инкубацией с стрептавидин-HRP; В. Инкубация с антителами B8(1)-H6, меченными пероксидазой хрена.
Рис. 12. Оценка клеточной гибели под действием GD2-специфичных антител B8(1)-H6, scFv- и ди-scFv-фрагментов антител B8(1)-H6. (А) - МТТ-тест, 72 ч инкубации, (Б) - PI-тест и 7AAD-тест, 24 ч. инкубации. К - интактные клетки.
Рис. 13. Оценка антитело-зависимой клеточно-опосредованной цитотоксичности антител B8(1)-H6 в клетках линии EL-4.
Подробное раскрытие изобретения
Определения и термины
Для лучшего понимания настоящего изобретения ниже приведены некоторые термины, использованные в настоящем описании изобретения.
В описании данного изобретения термины «включает» и «включающий» интерпретируются как означающие «включает, помимо всего прочего». Указанные термины не предназначены для того, чтобы их истолковывали как «состоит только из».
Под «субъектом» следует понимать человека или другое млекопитающее.
Ганглиозидом GD2 в настоящем описании называется дисиалоганглиозид, экспрессированный на поверхности опухолевых клеток нейроэктодермального происхождения со следующей IUPAC формулой: (2R,4R,5S,6S)-2-[3-[(2S,3S,4R,6S)-6-[(2S,3R,4R,5S,6R)-5-[(2S,3R,4R,5R,6R)-3-acetamido-4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-2-[(2R,3S,4R,5R,6R)-4,5-dihydroxy-2-(hydroxymethyl)-6-[(E)-3-hydroxy-2-(octadecanoylamino)octadec-4-enoxy]oxan-3-yl]oxy-3-hydroxy-6-(hydroxymethyl)oxan-4-yl]oxy-3-amino-6-carboxy-4-hydroxyoxan-2-yl]-2,3-dihydroxypropoxy]-5-amino-4-hydroxy-6-(1,2,3-trihydroxypropyl)oxane-2-carboxylic acid.
Термин «функциональный фрагмент антитела» в использованном здесь значении означает один или несколько фрагментов антитела, которые сохраняют способность специфически связываться с антигеном (например, ганглиозидом GD2). Примеры функциональных фрагментов включают (i) Fab-фрагмент, одновалентный фрагмент, состоящий из VL, VH, CL и СН1 доменов; (ii) F(ab')2-фрагмент, двухвалентный фрагмент, включающий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (iii) Fv-фрагмент, состоящий из VL и VH доменов одного плеча антитела; (iv) два домена Fv-фрагмента, VL и VH, объединенных при помощи синтетического линкера в одну белковую цепь, в которой VL и VH области спариваются с образованием одновалентной конструкции (известна как одноцепочечный Fv-фрагмент (scFv); см., например, Bird et al., Science, 1988, 242:423-426). Указанные фрагменты антител получают обычными методами, известными специалистам в данной области. В предпочтительных вариантах настоящего изобретения функциональные фрагменты антител не обладают или обладают сниженными эффекторными функциями; в частности, функциональные фрагменты антител могут не содержать Fc-регион.
Некоторые мультивалентные конструкции по настоящему изобретению могут содержать от двух до нескольких функциональных фрагментов антител, каждый из которых способен специфически связываться с ганглиозидом GD2; таким образом, содержать два или несколько антиген-связывающих участков. Функциональные фрагменты антител в мультивалентной конструкции могут включать вариабельные области, а также, иногда, константные области, выделенные из последовательностей иммуноглобулинов, специфически связывающих GD2. Предпочтительно, но без обязательных ограничений, функциональные фрагменты в составе мультивалентной конструкции по настоящему изобретению не обладают или обладают сниженными эффекторными функциями. Мультивалентность, т.е. способность связывать сразу несколько молекул антигена, обеспечивает конструкции по настоящему изобретению высокую авидность (стабильность взаимодействия между конструкцией и клеткой, экспрессирующей антигены на поверхности), что в свою очередь обеспечивает улучшенные функциональные свойства конструкции.
Используемые функциональные фрагменты антител по настоящему изобретению могут представлять собой «химерные» или «гуманизированные» фрагменты антител. Химерные фрагменты означают фрагменты антител, которые включают последовательности вариабельной области тяжелой и/или легкой цепей из одного вида и последовательности константной области из другого вида. Гуманизированные фрагменты означают фрагменты антител, которые включают последовательности фрагментов человеческих антител, в которых некоторые остатки CDR и, возможно, некоторые остатки каркасной области (FR) заменены остатками аналогичных сайтов из антител грызунов.
В некоторых вариантах изобретения функциональные фрагменты антител согласно изобретению содержат области тяжелых и/или легких цепей, включающие в себя аминокислотные последовательности, гомологичные аминокислотным последовательностям, описанным в Таблице 1 предпочтительных фрагментов антител, где указанные функциональные фрагменты антител сохраняют нужные функциональные свойства согласно изобретению (связывание с ганглиозидом GD2 и индукция клеточной гибели). Гомологичные фрагменты антител могут быть получены путем мутагенеза (например, сайт-направленного или ПЦР-опосредуемого мутагенеза) соответствующих молекул нуклеиновых кислот с последующим тестированием кодируемого модифицированного фрагмента антител на сохранение его функций в соответствии с описанными здесь функциональными анализами.
Используемый здесь термин «процент гомологии двух последовательностей» эквивалентен термину «процент идентичности двух последовательностей». Процент идентичности двух последовательностей определяется числом положений идентичных аминокислот в этих двух последовательностях с учетом числа пробелов и длины каждого пробела, которые необходимо ввести для оптимального сопоставления двух последовательностей путем выравнивания. Процент идентичности равен числу идентичных аминокислот в данных положениях с учетом выравнивания последовательностей, разделенному на общее число положений и умноженному на 100. Процент идентичности двух аминокислотных последовательностей может быть определен с помощью программы NCBI Protein BLAST (https://blast.ncbi.nlm.nih.gov/).
Создание гибридомы, продуцирующей GD2-специфичные моноклональные антитела B8(1)-H6.
Подбор и синтез GD2-мимикрирующих пептидов. В качестве иммуногена были выбраны ганглиозид-мимикрирующие пептиды, полученные с использованием GD2-специфичных антител 14G2a или МЕ61, наиболее схожие по своей пространственной структуре с углеводными детерминантами ганглиозида GD2, а также их KLH-модифицированные аналоги. Были отобраны следующие ганглиозид GD2-мимикрирующие пептиды: 14G2a-94L (RCNPNMEPPRCWAAEGD) и 361-P10S (WRYTAPVHLGDG), а так же их KLH-модифицированные варианты: 14G2a-94L-KLH (RCNPNMEPPRCWAAEGD-KLH) и 361-P10S-KLH (WRYTAPVHLGDG-KLH), синтезированные методом твердофазного синтеза. Пришивка KLH осуществлялась с N-конца пептидов (Peptide 2.0 inc, США). Структура данных пептидов основана на результатах рентгеноструктурного анализа комплексов GD2-антитело. Основным преимуществом пептидов по сравнению с ганглиозидом GD2 является повышенная иммуногенность. Кроме того, известно, что при образовании коньюгатов с опухолевыми антигенами, гемоцианин улитки (KLH) существенно повышает их иммуногенность.
В качестве объекта для иммунизации были выбраны лабораторные мыши линии BALB/с. Была проведена четырехкратная иммунизация лабораторных мышей с последующим слиянием выделенных спленоцитов и клеток миеломы X63 в полиэтиленгликоле с получением клеток гибридомы. Самки мышей линии BALB/c возрастом 10 недель были четырехкратно иммунизированы 40 мкг GD2-мимикрирующих пептидов (чистота более 95%), растворенных в стерильном PBS в объеме 250 мкл, смешанными с равным объемом полного адъюванта Фрейнда до образования устойчивой эмульсии. Иммунизация проводилась внутрибрюшинно с интервалами раз в две недели. Эффективность иммунизации и оценку продукции антител в сыворотке крови животных проводили методом иммуноферментного анализа сыворотки животных, согласно методике, описанной выше. Через 3 дня после последней иммунизации, выделенные мышиные спленоциты были слиты с клетками миеломы X63 в полиэтиленгликоле согласно общепринятой методике с использованием ПЭГ 4000.
Отбор и анализ моноклонов. Поиск и отбор клонов, продуцирующих GD2-специфичные моноклональные антитела, осуществлялся методом лимитирующих разведений. Контроль эффективности и специфичности связывания полученных антител с ганглиозидом GD2 осуществлялся методом иммуноферментного анализа.
Исходя из сравнения аффинности полученных антител, продуцированных различными клонами, при помощи иммуноферментного анализа, были выбраны наиболее оптимальные по ряду характеристик гибридомы, продуцирующие антитела, связывающие ганглиозид GD2 (антитела, продуцирующиеся в выбранных клетках гибридомы, были обозначены как B8(1)-H6). Изотипирование этих антител показало, что они относятся к IgM классу. Изотипирование полученных антител проводилось при помощи IsoStrip Mouse Monoclonal Antibody Isotyping Kit (Santa Cruz, США) согласно методике производителя. Наработка и выделение выбранных антител осуществлялась с использованием асцитов и Protein A. Антитела B8(1)-H6 проявили способность связывания с Protein A, однако для получения чистой фракции антител использовали дополнительную хроматографическую очистку с использованием гель-фильтрации. Дополнительная очистка полученных на Protein A-колонке антител B8(1)-H6 проводилась на колонке TSKgel 2000pw (7.5 мм × 300 мм, Tosoh Corporation, Japan) с использованием ВЭЖХ-системы Beckman System gold в PBS. Скорость потока 1 мл/мин. Белковые фракции собирали с использованием детектора на 280 нм. Очищенную фракцию антител концентрировали на фильтрах Amicon Ultra (максимальная пропускающая способность 100 кДа) при 1000 g в течение 20 мин. Использование хроматографии позволило получить фракции антител B8(1)-H6 с чистотой порядка 80% (Рис. 1), что было показано при помощи ПААГ-электрофореза.
Оценка кросс-реактивности полученных GD2-специфичных антител класса IgM с ганглиозидами GM1, GD1b и GD3 проводилась согласно стандартной методике ИФА.
Полученные антитела B8(1)-H6 обладают хорошей GD2-связывающей способностью, сравнимой с контрольными моноклональными антителами МЕ361, что было показано в ходе прямого иммуноферментного анализа с ганглиозидом GD2. Также полученные антитела способны эффективно связываться с ганглиозидом GD2 опухолевых клеток линии мышиной лимфомы EL-4. Более того, после 24-х часов инкубации клеток мышиной лимфомы EL-4 с антителами B8(1)-H6 в концентрации 20 мкг/мл процент клеток с фрагментированной ДНК увеличивался в 1.9+0.4 по сравнению с базовым уровнем (Рис. 2), что показывает индукцию гибели GD2-позитивных опухолевых клеток под действием GD2-специфичных антител.
Далее проводили анализ кроссреактивности B8(1)-H6 с другими ганглиозидами. Контрольные антитела ME361 проявляют кросс-реактивность к ганглиозиду GD3, который является очень близким к ганглиозиду GD2 по структуре, но при этом его экспрессия отмечается на клетках нормальных тканей и органов, что существенно ограничивает возможности использования полноразмерных антител этого типа. Для изучения кроссреактивности полученных антител B8(1)-H6 был использован метод иммуноферментного анализа. Как и ожидалось, антитела МЕ361 обладают значимой кросс-реактивностью с ганглиозидом GD3 составляющей около 15% от связывания с GD2 (Рис. 3). Для антител 14G2a и B8(1)-H6 параметры кросс-реактивности для всех исследованных ганглиозидов не превышали 5%, что говорит о строгой специфичности этих антител к ганглиозиду GD2 (Рис. 3).
Для подтверждения результатов также было проведено цитометрическое окрашивание клеток различными GD2-специфичными антителами. Исходя из анализа ганглиозидного состава клеточных линий, а также результатов окрашивания клеток анти-GD2-мАт 14G2a и анти-GD3-мАт R24 были отобраны GD2+GD3--, GD2+GD3+-, GD2-GD3+- и GD2-GD3--клеточные линии. Результаты поверхностного окрашивания этих линий подтверждают результаты об отсутствии кросс-реактивности антител B8(1)-H6 с ганглиозидом GD3, поскольку они не показали связывания с GD3-позитивными клетками линии SKOV-3.
Определение вариабельных областей антитела B8(1)-H6.
Полученная гибридома продуцировала антитела класса IgM, что не является оптимальным в силу большой молекулярной массы антител (700-1100 кДа). Поскольку антитела B8(1)-H6 обладали хорошими характеристиками связывания и специфичностью, их использовали для получения фрагментов антител и различных рекомбинантных вариантов антител, специфичных к GD2, и имеющих большие перспективы применения на практике.
Секвенирование вариабельных доменов антител. Для определения последовательностей антитела B8(1)-H6 были выделены фрагменты ДНК, кодирующие вариабельные домены легкой и тяжелой цепей этих антител. Для этого из клеток гибридомы выделяли тотальную РНК набором RNeasy mini kit (Qiagen). Качество препаратов РНК контролировали с помощью агарозного электрофореза с последующим окрашиванием гелей бромистым этидием.
Получение библиотеки кДНК проводили набором RevertAid H Minus First strand cDNA Synthesis kit (Thermo Scientific) c использованием oligo dT20 праймера. ПЦР с препаратов кДНК проводили с панелями вырожденных праймеров, комплементарных сигнальным пептидам иммуноглобулинов (Chardes et al., 1999). В качестве обратных праймеров использовали последовательности, комплементарные константным доменам тяжелых и легких цепей. Обратные праймеры также содержали рестриктный сайт SalI для последующего клонирования в плазмидный вектор. Положительным контролем для ПЦР служили прямые олигонуклеотиды, комплементарные консервативным FR1 районам вариабельных доменов иммуноглобулинов.
Реакцию ПЦР проводили с использованием High Fidelity Enzyme mix (Thermo Scientific), с использованием минимального числа циклов. Продукты реакции анализировали на агарозном электрофорезе; идентифицировали комбинации вырожденных олигонуклеотидов, давших ПЦР продукт. Подходящие комбинации использовали для препаративной ПЦР, продукты очищали с помощью электрофореза, расщепляли рестриктазой SalI и клонировали в вектор pBluescript KS+ по сайтам EcoRV и SalI. Бактериальные клоны выращивали в жидкой культуре, содержащей селективный антибиотик; плазмидную ДНК выделяли с помощью набора QIAprep Spin Miniprep Kit (Qiagen), и вставки секвенировали.
Сравнение полученных последовательностей с последовательностями ближайших гомологов по базам данных подтвердило принадлежность последовательности антител B8(1)-H6 к генам иммуноглобулинов класса IgM. В Таблице 1 представлены последовательности вариабельных областей тяжелых и легких цепей антител.
Таблица 1.
|
Нижеследующие примеры приведены в целях раскрытия характеристик настоящего изобретения и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения.
Примеры создания молекул на основе GD2-специфичного антитела B8(1)-H6 для терапии онкологических заболеваний.
Пример 1. Получение химерных моноклональных антител.
Последовательности химерных антител создавались на основе синтезированных вариабельных доменов антител B8(1)-H6. Одновременно с синтезом последовательностей проводили оптимизацию кодонов для экспрессии в клетках млекопитающих. Для получения тяжелых и легких цепей вариабельные домены сшивали с константными доменами тяжелых цепей (ТЦ) и легких цепей (ЛЦ) изотипа IgG1 человека. Данные последовательности были также оптимизированы по кодонам для экспрессии в клетках млекопитающих. Для получения химерных ТЦ и ЛЦ использовали стратегию overlap extension, позволяющую сшить два фрагмента ДНК, используя реакцию ПЦР. Реакцию ПЦР проводили с использованием полимеразы High Fidelity PCR mix (Thermo Scientific) c минимальным числом циклов. Кроме полноразмерных антител, аналогично были сконструированы функциональные фрагменты антител (см. ниже).
Продукты ПЦР клонировали в вектор pcDNA3.4 (Thermo Scientific). Для снижения фона при лигировании вектора со вставками вектор подвергали дефосфорилированию с использованием фосфатазы FastAP (Thermo Scientific). Бактериальные колонии скринировали с помощью ПЦР. Плазмиды выделяли набором Wizard DNA purification system kit (Promega), после чего проверяли рестрикцией. Плазмиды проверяли секвенированием, и в дальнейшей работе использовали конструкции, не содержащие ни единой нуклеотидной замены. Получение ДНК, пригодной для трансфекции, проводили с использованием набора Qiagen plasmid maxi kit c небольшими модификациями. Концентрацию и чистоту ДНК определяли по оптическому поглощению при 260 и 280 нм. Препараты стерилизовали фильтрованием через фильтры с размером пор 0.22 мкм.
Для постоянной экспрессии химерных антител на основе B8(1)-H6 был создан клон-продуцент на основе клеток CHO. Получение кодирующей последовательности легкой цепи ДНК проводили методом ПЦР, с использованием праймеров, содержащих сайты клонирования в экспрессионный вектор pcDNA3.3 с измененным полилинкером (Thermo Fisher Scientific). Клонирование проводили по сайтам XbaI и NotI.
Клонирование тяжелой цепи осуществляли в вектор pOptiVec (Thermo Fisher Scientific) с использованием рестриктных сайтов XbaI и NheI. Генетические карты полученных конструкций показаны на Рис. 4. Препараты ДНК очищали с помощью ионообменной хроматографии и использовали для трансфекции клеток DXB-11.
Введение экспрессионных конструкций в клетки осуществляли с помощью электропорации на приборе Gene Pulser Xcell System (BioRad), с использованием смеси плазмид, несущих тяжелую и легкую цепи антитела. Спустя 48 часов после начала трансфекции проводили смену среды культивирования на бессывороточную среду, содержащую 200 мкг/мкл G418 и лишенную гипоксантина и тимидина. Культивирование продолжали в течение 20 дней, в присутствии антибиотика G418 в среде, не содержащей гипоксантина и тимидина; в течение этого периода проводили регулярное измерение клеточной плотности и жизнеспособности.
После достижения устойчивых показателей роста (жизнеспособность выше 95% и время удвоения - не более 30 часов) стабильный клеточный пул клонировали лимитирующими разведениями (1 клетка на лунку 96 луночного планшета). В среду для клонирования клеток добавляли низкие концентрации метотрексата (25нМ). Спустя еще 14 дней, позитивные по росту клеток лунки анализировали с помощью ИФА на предмет секреции рекомбинантных антител. Из 225 проанализированных колоний детектируемой экспрессией обладали 25 первичных клона. Колонии с максимальными значениями продукции масштабировали до лунок 24 луночного планшета и подвергали второму раунду амплификации с использованием 100 нМ метотрексата. Клоны с максимальными значениями продуктивности дополнительно амплифицировали при 500 нМ метотрексата, после чего повторно клонировали методом лимитирующих разведений. Полученные финальные клоны с максимальными значениями продуктивности и хорошими показателями роста масштабировали и далее культивировали в суспензионной культуре для получения препаративных количеств химерных антител.
Было выбрано два финальных клона (№6 и №74), обладавших хорошими значениями удельной продуктивности и приемлемыми ростовыми характеристиками. Клоны выращивали в суспензионном состоянии, в объемах 30 мл в среде Dynamis (ThermoFisher Scientific) с использованием двух комбинаций коммерчески доступных подпиток - Cell Boost 7a и 7b (Hyclone) или Efficient Feed C+ (ThermoFisher Scientific). Ежедневно корректировали содержание глюкозы в среде и оценивали клеточную плотность и жизнеспособность. Культивирование продолжали в течение 13-14 дней и заканчивали после снижения жизнеспособности культуры ниже 60%. Кондиционированную среду осветляли центрифугированием и наносили на SDS электрофорез. Продукция антител B8(1)-H6 клоном №74 была на достаточно высоком уровне (около 30 мг/л), что позволило выделить достаточные количества антител для проведения последующих экспериментов.
Пример 2. Получение scFv- и Fab-фрагментов антитела B8(1)-H6.
Для экспрессии в клетках линии СНО scFv- и Fab-фрагментов GD2-специфичных антител соответствующие кДНК были встроены в векторы для стабильной экспрессии в клетках CHO линии DXB-11, дефицитной по гену дигидрофолатредуктазы (DHFR). Для Fab-фрагмента получение кодирующей последовательности легкой цепи ДНК проводили методом ПЦР; далее использовали клонирование по сайтам XbaI и NotI в экспрессионный вектор pcDNA3.3 с измененным полилинкером (Thermo Fisher Scientific). Клонирование части тяжелой цепи осуществляли в вектор pOptiVec (Thermo Fisher Scientific) с использованием рестриктных сайтов XbaI и NheI. Для гена scFv-фрагмента использовали клонирование по сайтам XbaI и NotI в экспрессионный вектор pcDNA3.3. Полученные конструкции проверяли секвенированием. Препараты ДНК очищали с помощью ионообменной хроматографии и использовали для трансфекции клеток DXB-11. Введение экспрессионных конструкций в клетки осуществляли с помощью электропорации на приборе Gene Pulser Xcell System (BioRad), с использованием смеси плазмид, несущих тяжелую и легкую цепи антитела. Спустя 48 часов после начала трансфекции проводили смену среды культивирования на бессывороточную среду, содержащую 200 мкг/мкл G418 и лишенную гипоксантина и тимидина. Культивирование продолжали 19 дней, в течение которых проводили регулярное измерение клеточной плотности и жизнеспособности. После достижения устойчивых показателей роста (жизнеспособность выше 95% и время удвоения - не более 30 часов) стабильный клеточный пул клонировали лимитирующими разведениями (1 клетка на лунку 96 луночного планшета). В среду для клонирования клеток добавляли низкие концентрации метотрексата (25нМ). Спустя еще 14 дней позитивные по росту клеток лунки анализировали с помощью иммуноферментного анализа (ИФА) на предмет секреции фрагментов антител. Колонии с максимальными значения продукции масштабировали до 24 луночных планшетов и подвергали второму раунду амплификации с использованием 100 нМ метотрексата. Клоны с максимальными значениями продуктивности дополнительно амплифицировали при 250 и 500 нМ метотрексата, после чего повторно клонировали методом лимитирующих разведений, как описано выше.
Полученные финальные клоны с максимальными значениями продуктивности и хорошими показателями роста масштабировали и далее культивировали в суспензионной культуре для получения препаративных количеств фрагментов антител.
Для экспрессии в E.coli scFv- и Fab-фрагментов GD2-специфичных антител соответствующие кДНК были встроены в вектор pET22b+ по рестриктным сайтам NdeI и XhoI. Идентичность конструкций ожидаемым была подтверждена секвенированием. Для экспрессии конструкции pET22b+/scFv и pET22b+/Fab трансформировали в штаммы E. coli Rosetta2(DE3)pLysS или HMS174(DE3)pLysS, и проводили индукцию экспрессии по стандартному протоколу, известному специалистам, в течение 4 ч при 25°С или 22 ч при 20°С. Количество, а также локализацию белка оценивали при помощи BugBuster Protein extraction reagent (Novagen, USA) в соответствии с инструкцией производителя. Для выделения тотальной фракции белка клетки E. coli, осажденные из 0.5 л культуры, тщательно ресуспендировали в 5 мл холодного лизирующего буфера (50 мМ NaH2PO4, 300 мМ NaCl, 10 мМ имидазол, 0.5 мМ фенилметилсульфонил фторид (PMSF), pH 7.2) и дополнительно охлаждали в течение 15 мин на льду. Затем суспензию обрабатывали ультразвуком до полного разрушения клеток. Полученный лизат центрифугировали при 10 000 g в течение 20 мин при 4°С. Осадок, содержащий клеточный дебрис, удаляли. Перевод белка в конечный буфер (фосфатный буфер (PBS), дополненный 0.02% NaN3) осуществляли центрифугированием на фильтрах Amicon Ultra-4 с диаметром пор 10 кДа.
Для экспрессии в Pichia pastoris scFv- и Fab-фрагментов GD2-специфичных антител соответствующие кДНК были встроены в вектор pPICZalphaB по сайтам рестрикции XhoI/NotI. При этом пропептид для секреции (альфа-фактор из Saccharomyces cerevisiae) находился в одной рамке считывания с фрагментами антител. Для отщепления альфа-фактора были предусмотрены сайты действия протеаз Kex2 (EKRE) и Ste13 (EA EA). Клонирование проводили в стандартном штамме E. coli XL1-blue, селекцию осуществляли при посеве на низко-солевую среду LB c 25 мкг/мл зеоцина. Методом электропорации трансфицировали клетки Pichia pastoris штаммов GS115 и smd1168h согласно инструкции к EasySelect Pichia Expression Kit (Invitrogen). Трансформанты сеяли на среду YPDS-агар с разной концентрацией зеоцина (500-2000 мкг/мл), трансформанты вырастали на 4-5 сутки. Полученные колонии рассевали для получения отдельных клонов, которые высевали на среду BMGH в 24-луночные планшеты и инкубировали при 30°C и 200 об/мин трое суток. Затем клетки собирали центрифугированием при 1500×g и комнатной температуре, и высевали на среду BMMH в 24-луночные планшеты. После чего инкубировали при 30°C и 200 об/мин в течение 4 суток. Каждые сутки добавляли 0,5% метанола. После окончания инкубации клетки отделяли центрифугированием при 5000×g, супернатанты оставляли для анализа.
Пример 3. Выделение и очистка фрагментов антител на примере экспрессии в E.coli.
Для очистки scFv-фрагментов к полученным лизатам добавляли 50% суспензии Ni-NTA-агарозы и инкубировали 1 ч при 4°С при легком перемешивании. Затем суспензию переносили в колонку и дважды промывали буфером, содержащим 50 мМ NaH2PO4, 300 мМ NaCl, 20 мМ имидазол (pH 7.2). Элюцию проводили буфером, содержащим 50 мМ NaH2PO4, 300 мМ NaCl, 250 мМ имидазол (pH 7.2).
Для выделения и очистки Fab-фрагментов из клеточных лизатов использовали колонку с Protein L агарозой из набора реактивов HiTrap Protein L (GE Healthcare, США). После промывки и активации колонки связывающим буфером на нее наносили лизат клеток E.coli. Колонку промывали от несвязавшихся белков, затем элюировали Fab-фрагменты, используя элюирующий буфер. Очищенные фрагменты собирали в 15 мл пробирку, содержащую 750 мкл нейтрализующего буфера. Нейтрализованный раствор Fab-фрагментов переводили в фосфатный буфер (PBS), содержащий азид натрия (0.02%), центрифугированием через фильтры Amicon Ultra (максимальная пропускающая способность 100 кДа) при 1000 g в течение 20 мин.
Чистота полученных Fab- и scFv-фрагментов по белковому электрофорезу составила >95%. Полученные препараты были переведены в PBS, pH 7,4, и затем стерилизованы фильтрацией через мембрану с размером пор 0,22 мкм. Концентрацию белка определяли спектрофотометрически при длине волны 280 нм.
Пример 4. Мультимеризация scFv- и Fab-фрагментов GD2-специфичных антител через пегилирование.
Мультимеризация является одной из стратегий улучшения фармакокинетических и фармакодинамических свойств фрагментов антител. Больший молекулярный размер мультимеров на основе фрагментов антител способствует увеличению периода полужизни в сыворотке, а также более быстрому поглощению опухолью за счет EPR-эффекта, что приводит к их лучшему накоплению в опухоли по сравнению с исходными IgG антителами или мономерами фрагментов. Кроме того, мультимеризация, как правило, способствует мультивалентности против антигена-мишени, которая приводит не только к более высокой аффинности благодаря эффекту авидности, но также может способствовать улучшению функциональных свойств и компенсировать отсутствие индукции вторичных иммунных функций ADCC и CDC. В случае моноклональных антител к ганглиозиду GD2 Fc-фрагмент полноразмерных антител во многом определяет побочные эффекты, и иммунные механизмы ADCC и CDC имеют ограниченный положительный терапевтический эффект. В связи с этим, создание мультимерных фрагментов GD2-специфичных антител для терапии онкологических заболеваний имеет значительный потенциал.
В то же время мультимеры фрагментов антител, полученные стандартными способами, такими как использование стерической агрегации, формирования цистеиновых мостиков или кроссшивающих реагентов, часто характеризуются крайне низкой стабильностью и проблемами с масштабированием, что ограничивает перспективы их использования.
Одной из стратегий увеличения времени полужизни терапевтических молекул в кровотоке является увеличение гидродинамического объема молекулы путем связывания с инертными полимерами, такими как полиэтиленгликоль или другими гидрофильными полимерами. Причем только полиэтиленгликоль (ПЭГ)-производные белков одобрены FDA и другими регуляторными ведомствами для использования в медицине. Действительно, слияние или конъюгация различных пептидных молекул с полимерами существенно повышает их время полужизни, что было подтверждено экспериментально и в клинической практике.
Для усиления терапевтического потенциала фрагментов GD2-специфичных антител и улучшения их фармакокинетических свойств авторами было использовано сайт-направленное пегилирование, целью которого было не только увеличение времени циркуляции фрагментов в крови, но и усиление цитотоксических свойств за счет получения стабильных мультимеров фрагментов GD2-специфичных антител. Для получения ди-, три- и тетрамеров фрагментов антител использовали стандартные реагенты ПЭГ-малеимид-2 и ПЭГ-малеимид-4 (JenKem Technology USA, США https://www.jenkemusa.com). Использованные производные ПЭГ позволяют присоединять к одной молекуле ПЭГ несколько (от 1 до 4) молекул фрагментов. Схемы реакций пегилирования представлены на Рис. 5.
Для проведения реакции пегилирования scFv- и Fab-фрагментов моноклональных антител к раствору фрагментов (2.5 мг/мл) в буфере для пегилирования (20 mM фосфатный буфер, с добавлением 50 mM NaCl, 10 mM EDTA, pH 7.5) добавляли стоковый раствор 15 mM TCEP до достижения конечной концентрации TCEP 0.1 mM. Инкубацию проводили при комнатной температуре в течение 90 мин при аккуратном перемешивании. От восстановителя избавлялись центрифугированием через фильтры Amicon Ultra-4 (максимальная пропускающая способность 10 кДа), либо используя колонки для высаливания Zeba Desalting Columns (TermoFisher, США). Затем к раствору фрагментов антител добавляли соответствующие растворы малеимид-ПЭГ, в различных соотношениях. Инкубация длилась 16-18 ч при комнатной температуре с аккуратным перемешиванием на шейкере. Эффективность пегилирования оценивали с помощью ПААГ-электрофореза. После подбора оптимальных условий проведения пегилирования выход реакций составлял не менее 90%.
Для разделения полученных пегилированных фрагментов и получения чистых фракций, содержащих индивидуальные моно- и мультимеры фрагментов антител, проводили хроматографическую очистку с использованием анионообменной хроматографии или гель-фильтрации.
В одном из вариантов, для разделения полученных продуктов реакций пегилирования фрагментов антител проводили катионообменную хроматографию на колонке TSK-GEL SP-5PW с использованием ВЭЖХ-системы Beckman System gold, подвижная фаза: элюирующий буфер (25 мМ CH3COOH, pH 4.0), линейный градиент 0 mM - 250 mM NaCl (в течение 90 мин при комнатной температуре, скорость потока 1 мл/мин).
Альтернативно, для разделения полученных продуктов реакций пегилирования использовали гель-фильтрацию (эксклюзионную хроматографию) - колонку Superdex 200 10/30 GL и элюент PBS, пропущенный через фильтр 0.22 мкм. Скорость потока 0.6 мл/мин.
Пример 5. Определение антиген-связывающих свойств GD2-специфичных антител B8(1)-H6, scFv-фрагментов антител B8(1)-H6 и мультимерных фрагментов антител B8(1)-H6.
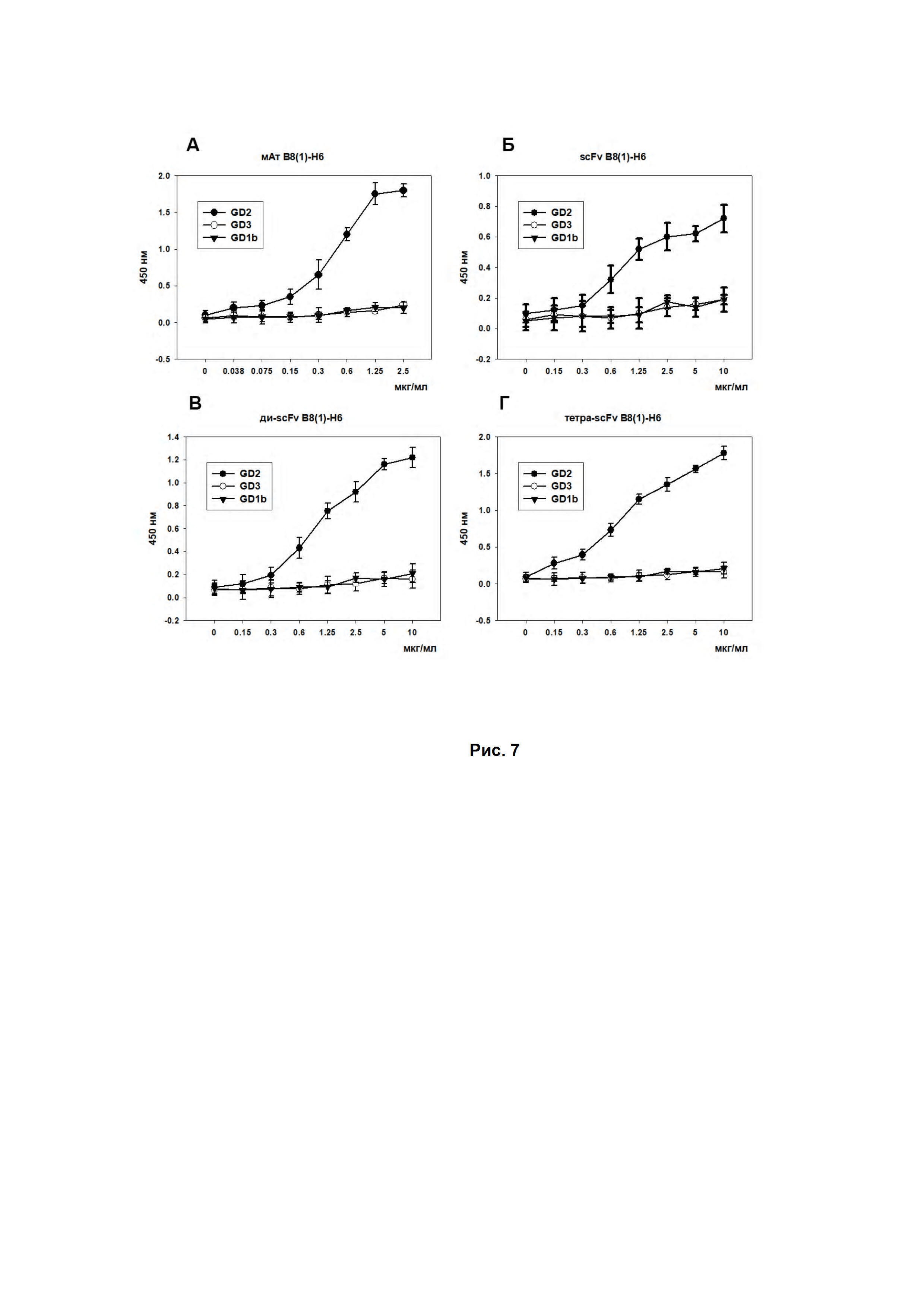
Изучение антиген-связывающих свойств полученных GD2-специфичных молекул на основе антител B8(1)-H6 проводили методами проточной цитофлуориметрии (Рис. 6) и иммуноферментного анализа (ИФА) (Рис. 7).
Рекомбинантные GD2-специфичные антитела B8(1)-H6, а также scFv-фрагменты антител B8(1)-H6 и их модифицированные производные проявили способность специфически взаимодействовать с ганглиозидом GD2. Так, при окрашивании GD2-позитивной линии EL-4 антителами B8(1)-H6 и фрагментами антител наблюдалось значимое увеличение интенсивности флуоресценции клеток по сравнению с контрольными клетками, к которым были добавлены только вторичные антивидовые антитела. GD2-негативные клетки линии NGP не связывались с молекулами на основе GD2-специфичного антитела B8(1)-H6 (Рис. 6).
Аналогично, в ИФА было показано дозо-зависимое увеличение интенсивности развития пероксидазной реакции при добавлении последовательных разведений GD2-специфичных антител B8(1)-H6 и scFv- фрагментов антител B8(1)-H6 к сорбированному на подложке ганглиозиду GD2. Было показано отсутствие кросс-реактивности использованных соединений к ганглиозидам GD3 и GD1b, которые являются структурно схожими с ганглиозидом GD2 (Рис. 7).
Также оценка связывания различных фрагментов GD2-специфичных антител показала, что пегилирование с образованием мультимерных фрагментов GD2-специфичных антител показывало значимое увеличение связывания с антигеном (почти в 2 раза по сравнению с мономерным scFv-фрагментом), что подтверждалось как методом ИФА (рис. 2 В, Г), так и окрашиванием GD2-позитивных опухолевых линий с последующей оценкой на проточном цитофлуориметре (Рис. 6).
Пример 6. Получение конъюгатов GD2-специфичных антител B8(1)-H6 и scFv- фрагментов антител B8(1)-H6 с биотином и пероксидазой хрена (HRP).
Полученные GD2-специфичные антитела можно использовать для определения ганглиозида GD2, локализованного на поверхности опухолевых клеток, а также в сыворотке крови больных с GD2-позитивными онкологическими заболеваниями. Для сокращения стадий детектирования ганглиозида GD2, повышения чувствительности метода, а также снижения неспецифического связывания антител была проведена модификация антител и фрагментов для получения биотин- и HRP-меченных антител B8(1)-H6 и их scFv-фрагментов.
Биотинилирование антител B8(1)-H6 проводили с использованием биотин-сукцинимида (Termo Scientific, США). К раствору антител с концентрацией 2 мг/мл в боратном буфере, рН 9.0, добавляли биотин-сукцинимид в двух весовых соотношениях антитело:биотин - 1:0.1 и 1:0.05. После инкубации в течение 4 ч добавляли NH4Cl (10 мкл 1М NH4Cl на 100 мкг эфира) и проводили диализ против PBS в течение 24 ч. Для оценки эффективности прохождения реакции биотинилирования антител B8(1)-H6 использовали методы проточной цитофлуориметрии и ИФА (Рис. 8).
Биотинилирование scFv-фрагментов антител B8(1)-H6 проводили с использованием биотин-малеимида (Termo Scientific, США). К раствору фрагментов антител с концентрацией 2 мг/мл в фосфатном буфере, рН 6.0, содержащем 100 мМ EDTA, добавляли восстановитель TCEP до конечной концентрации 0.5 мМ. После инкубации в течение 1.5 ч от восстановителя избавлялись центрифугированием через фильтры Amicon Ultra-4, либо используя колонки для высаливания. Затем к раствору фрагментов антител добавляли биотин-малеимид в 4-х кратном молярном избытке. Инкубация длилась 18 ч при 4°С с аккуратным перемешиванием на шейкере, после чего от не связавшегося биотин-малеимид избавлялись центрифугированием через фильтры Amicon Ultra-4. Для оценки эффективности прохождения реакции биотинилирования scFv-фрагментов антител B8(1)-H6 использовали методы проточной цитофлуориметрии и ИФА (Рис. 8).
Для оценки свойств биотинилированных антител B8(1)-H6 и scFv-фрагментов антител B8(1)-H6 использовали GD2-позитивные клетки мышиной лимфомы EL-4. Клетки инкубировали с биотинилированными и немечеными антителами и фрагментами в течение 1 ч, а затем с антивидовыми FITC-меченными антителами или стрептавидин-FITC в случае полноразмерных антител, либо с FITC-меченными анти-FLAG антителами или стрептавидин-FITC в случае фрагментов антител в течение 1 часа. Измерения проводили на проточном цитофлуриметре (EpicsCoulter XL). Результаты эксперимента представлены на Рис. 8.
В случае полноразмерных антител B8(1)-H6 биотинилирование привело к сравнимому сигналу флуоресценции клеток инкубированных с немечеными и биотинилированными антителами с последующим окрашиванием клеток антивидовыми FITC-меченными антителами или стрептавидин-FITC, соответственно. Данные результаты говорят об эффективном прохождении реакции биотинилирования антител B8(1)-H6.
При использовании биотинилированных scFv-фрагментов антител B8(1)-H6 и FITC-меченного стрептавидина было зарегистрировано значительное усиление сигнала флуоресценции клеток по сравнению с окрашиванием клеток немодифицированными scFv-фрагментами и анти-FLAG антителами (Рис. 8А).
Также эффективность реакции биотинилирования оценивалась методом ИФА. Проведение анализа было аналогично вышеописанной методике. Использование пары биотин-меченные антитела B8(1)-H6 и стрептавидин-HRP также позволило детектировать минимальные значения сорбированного ганглиозида GD2 (Рис. 8 Б), а в случае определения ганглиозида GD2 scFv-фрагментами битинилирование позволило существенно усилить детекцию сигнала (Рис. 8В).
Мечение антител B8(1)-H6 и scFv-фрагментов антител B8(1)-H6 пероксидазой хрена.
Конъюгацию антител B8(1)-H6 проводили с использованием набора реагентов EZ-link Plus activated peroxidase kit (Thermo Scientific, США), содержащих аминореактивную пероксидазу хрена, согласно инструкции производителя. Для конъюгации 1 мг антител B8(1)-H6 переводили в бикарбонатный буфер, pH 9.4, гельфильтрацией, используя колонку Zeba (Thermo Scientific, США). Растворяли 1 мг лиофилизированной EZ-link Plus активированной пероксидазы в 100 мкл ddH2O и добавляли к раствору антител. После инкубации в течение 1 ч при комнатной температуре реакцию останавливали добавлением цианоборгидрида натрия. Затем проводили диализ против PBS в течение 24 ч.
Активность полученных HRP-меченных антител B8(1)-H6 определяли, используя метод ИФА, по методике, описанной выше. В качестве контроля использовали немеченые антитела B8(1)-H6 с последующим добавлением вторичных антител anti-human Fc-specific-HRP. Результаты представлены на Рис. 9.
Конъюгацию scFv-фрагментов антител B8(1)-H6 проводили с использованием набора реагентов EZ-Link™ Maleimide Activated Horseradish Peroxidase Kit (Thermo Scientific, США), содержащих пероксидазу хрена, модифицированную малеимидными группами (HRP-малеимид), согласно инструкции производителя. К раствору фрагментов антител с концентрацией 2 мг/мл в фосфатном буфере, рН 6.0, содержащем 100 мМ EDTA, добавляли восстановитель TCEP до конечной концентрации 0.5 мМ. После инкубации в течение 1.5 ч от восстановителя избавлялись центрифугированием через фильтры Amicon Ultra-4, либо используя колонки для высаливания. Затем к раствору фрагментов антител добавляли HRP-малеимид в 4-х кратном молярном избытке. Инкубация длилась 18 ч при 4°С с аккуратным перемешиванием на шейкере, после чего от не связавшегося HRP-малеимид избавлялись центрифугированием через фильтры Amicon Ultra-4.
Активность полученных HRP-меченных scFv-фрагментов антител B8(1)-H6 определяли, используя метод ИФА, по методике описанной выше. В качестве контроля использовали немеченые scFv-фрагменты антител B8(1)-H6 с последующим добавлением вторичных антител anti-FLAG-HRP. Результаты представлены на Рис. 9Б.
Из Рис. 9 видно, что мечение антител B8(1)-H6 и scFv-фрагментов антител B8(1)-H6 пероксидазой хрена не привело к потере аффинности связывания с антигеном и при этом позволило сократить количество стадий метода ИФА по определению ганглиозида GD2.
Пример 7. Определение ганглиозида GD2 в сыворотке крови пациентов с нейробластомой с использованием антител B8(1)-H6.
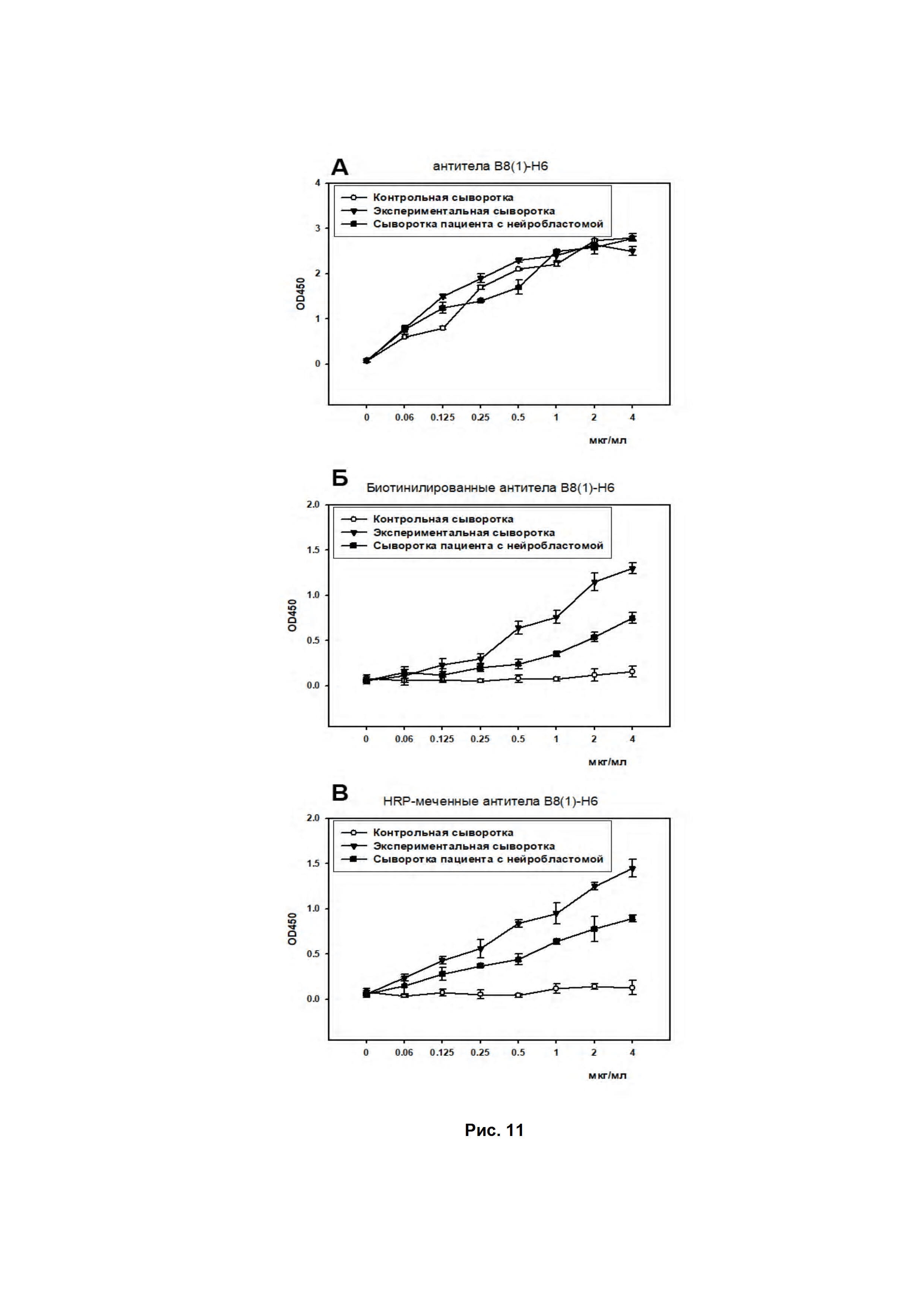
Наличие ганглиозида GD2 в сыворотке крови с использованием немеченых, биотинилированных и HRP-меченных GD2-специфичных антител B8(1)-H6 было проведено методами дот-блота и ИФА. В экспериментальной сыворотке с экзогенно добавленным ганглиозидом и в сыворотке крови пациента с нейробластомой было зафиксировано наличие GD2, в отличие от контрольной сыворотки здорового донора при использовании биотинилированных и HRP-меченных GD2-специфичных антител B8(1)-H6. Окрашивание мембраны при использовании немеченых антител B8(1)-H6 с последующей инкубацией с антителами anti-human Fc-specific-HRP привело к развитию пероксидазной реакции во всех сыворотках, что было обусловлено связыванием вторичных anti-human Fc-specific-HRP антител с различными антителами, содержащимися в сыворотках крови (Рис. 10).
Аналогичная картина наблюдалась в ИФА (Рис. 11). После сорбции трех вариантов сывороток, в различных разведениях в течение 2 ч проводилась стандартная методика ИФА с использованием немеченых, биотинилированных и HRP-меченных GD2-специфичных антител B8(1)-H6.
Из представленных данных видно, что биотинилированные и HRP-меченные антитела B8(1)-H6 можно использовать для определения ганглиозида GD2 в сыворотках крови пациентов с GD2-позитивными онкологическими заболеваниями, в частности с нейробластомой.
Пример 8. Иммуногистохимия биопсийного материала пациентов.
Из биопсийного материала нейробластомы, саркомы и рака эндометрия, замороженного в жидком азоте, получали срезы толщиной 5 мкм на криотоме HM550 MVP, Thermo. Стекла со срезами держали при комнатной температуре 30 мин, затем фиксировали холодным ацетоном и окрашивали с помощью непрямого иммунопероксидазного метода. Для блокирования внутренней пероксидазной активности срезы инкубировали в течение 30 мин в 3% перекиси водорода. После отмывок в PBS, содержащем 0,1% Tween-20, блокировали неспецифическое связывание с антигеном, инкубируя срезы в PBS, содержащем 0,1% Tween-20 и 5% человеческой сыворотки, а затем инкубировали с биотинилированными GD2-специфическими антителами 14G2a и B8(1)-Н6, после чего проводили инкубацию с HRP-меченным стрептавидином. Реакцию проявляли реактивом TMB 1-Step Solution (Thermo). Срезы контрастировали гематоксилином и эозином, после чего их покрывали мовиолом (Mowiol), накрывали покровным стеклом и запаивали. Визуализацию изображений проводили на микроскопе Zeiss Axiovert 40. Результаты иммуногистохимических экспериментов представлены в Таблице 2.
Таблица 2.
|
Обозначения в Таблице 2: +++ - сильное положительное окрашивание, ++ - положительное окрашивание, + слабое положительное окрашивание, - отсутствие окрашивания.
Использованные антитела B8(1)-Н6 показали избирательное окрашивание биопсийного материала опухолей различного происхождения. В случае срезов нейробластомы антитела показали сильное положительное или положительное окрашивание, срезы саркомы характеризовались слабым развитием реакции, а срезы рака эндометрия не окрашивались антителами B8(1)-Н6. Аналогичная картина была характерна для GD2-специфичных антител 14G2a, которые широко используются для определения ганглиозида GD2 методом иммуногистохимии. Исходя из этих данных можно заключить, что антитела B8(1)-Н6 могут применяться для определения GD2 в биопсийных образцах онкологических больных.
Пример 9. Прямая цитотоксическая активность GD2-специфичных антител B8(1)-H6, scFv-фрагментов антител B8(1)-H6 и мультимерных фрагментов антител B8(1)-H6.
Сравнительная оценка цитотоксических эффектов, индуцируемых антителами и фрагментами GD2-специфичных антител, проводилась с использованием нескольких классических подходов. В качестве клеточной модели исследования была использована линия мышиной лимфомы EL-4, которая характеризуется гиперэкспрессией ганглиозида GD2.
МТТ тест проводили с использованием стандартной колориметрической процедуры. При этом клетки GD2-позитивных опухолевых линий инкубировали в 96-луночных планшетах с последовательными разведениями GD2-специфических антител и их фрагментов в течение 72 ч, затем центрифугировали (8 мин, 300 g), осадок суспендировали и добавляли по 30 мкл/лунка раствора MTT (5 мг/мл в PBS). После выпадения кристаллов формазана (2-4 ч) его растворяли добавлением 100 мкл DMSO. Оптическое поглощение измеряли на спектрофотометре для планшетов Multiscan FC (Thermo Scientific, США) при длине волны 540 нм.
Оценка клеточной гибели в PI-тесте
Окрашивание пропидиум иодидом (PI) проводили по общепринятой методике. Вкратце, после окончания инкубации с GD2-специфичными антителами и их фрагментами в течение 24 ч, клетки EL-4 осаждали, фиксировали ледяным 70% EtOH в течение 1 ч при 4°С, дважды отмывали в PBS (300 g, 10 мин, 4°C). Затем клетки ресуспендировали в растворе для окрашивания ДНК (PBS, содержащий 20 мкг/мл PI и 10 мкг/мл РНКазы). Измерения проводили на проточном цитофлуориметре EPICS ELITE (Coulter, США). В каждом образце регистрировали не менее 1×104 клеток. На гистограммах клетки с фрагментированной ДНК, дифференцировали от нормальных клеток по более низкой интенсивности флуоресценции. Обработку результатов проводили с использованием программы WinMDI.
Оценку проницаемости плазматической мембраны клеток проводили с помощью флуоресцентного ДНК-красителя 7-AAD. Перед окрашиванием клетки однократно отмывали в PBS и к осадку добавляли 0.5 мл раствора 7-AAD (2 мкг/мл). Флуоресценцию клеток регистрировали цитофлуориметрически. В каждом образце регистрировали не менее 1×104 клеток. Обработку результатов проводили с использованием программы WinMDI.
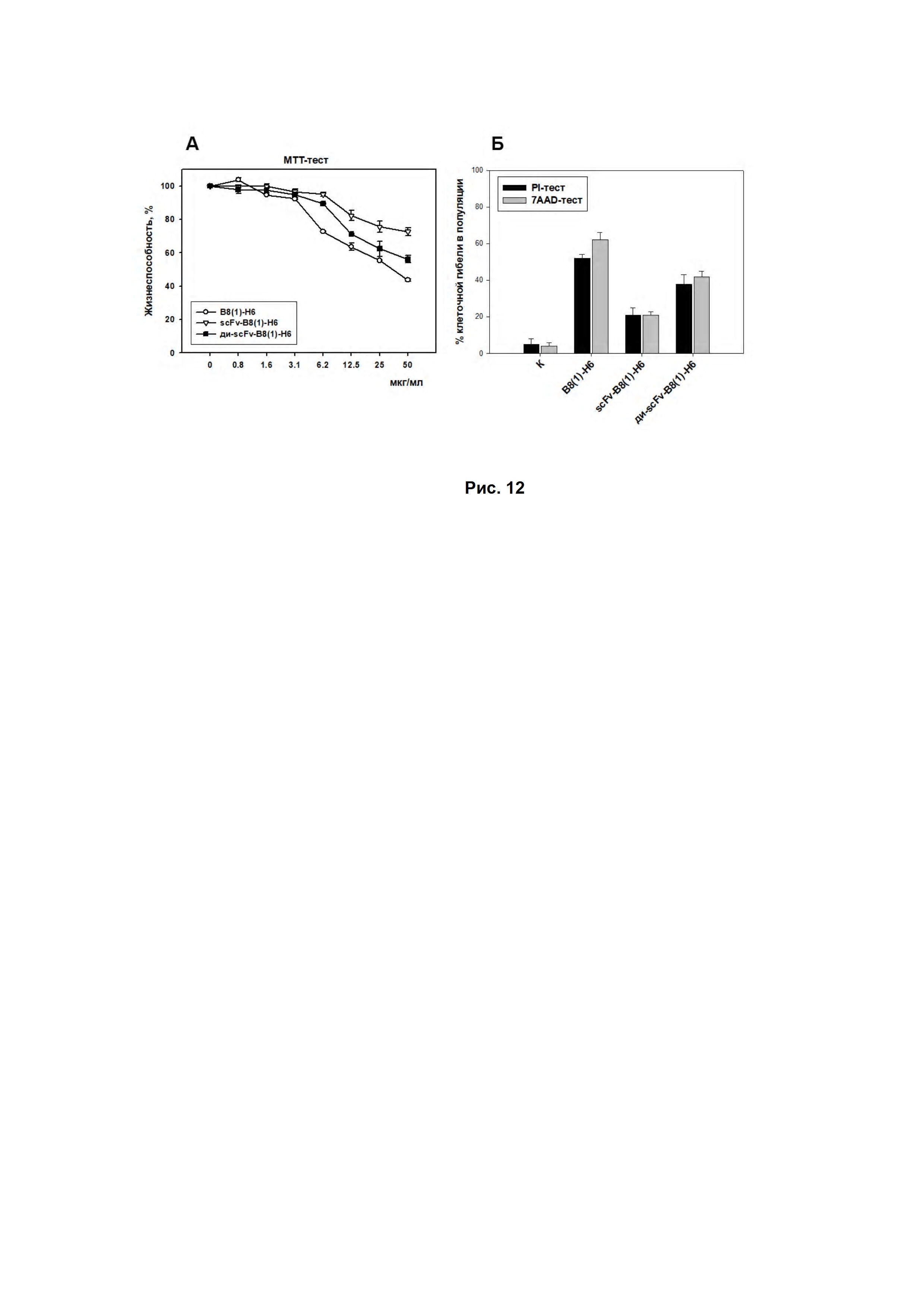
На Рис. 12 представлены данные цитотоксической активности GD2-специфичных антител B8(1)-H6, scFv- и ди-scFv-фрагментов антител B8(1)-H6.
Как видно из. Рис. 12А в МТТ-тесте жизнеспособность клеток EL-4 под действием GD2-связывающих молекул падала в дозозависимой манере. Наибольшей цитотоксической активностью обладали полноразмерные GD2-специфические антитела, так при максимальной концентрации антител жизнеспособность опухолевых клеток падала на 60%. Воздействие scFv-фрагментов оказывало меньший эффект, снижая жизнеспособность клеток только на 25%, а пегилированные димеры scFv-фрагментов обладали значительно большим цитотоксическим эффектом, который приближался к действию полноразмерных антител B8(1)-H6 (Рис. 12Б).
Методом проточной цитофлуориметрии в PI- и 7AAD-тестах было также показано, что инкубация клеток с scFv-фрагментами приводила к незначительному увеличению количества клеток с фрагментированной ДНК и клеток с нарушением целостности плазматической мембраны относительно интактных клеток, в то время как пегилированные димеры scFv-фрагментов характеризовались большими цитотоксическими эффектами по отношению к GD2-позитивным опухолевым клеткам линии EL-4.
Полученные данные говорят о том, что GD2-специфичные антитела B8(1)-H6, а также их scFv-фрагменты обладают прямой цитотоксической активностью по отношению к GD2-позитивным опухолевым клеткам, что может служить существенным вкладом в противоопухолевые эффекты при использовании данных соединений в иммунотерапии GD2-позитивных заболеваний.
Пример 10. Оценка антитело-зависимой клеточно-опосредованной цитотоксичности антител B8(1)-H6.
В качестве клеток мишеней использовали GD2-позитивную клеточную линию EL-4. До проведения теста клетки линии EL-4 окрашивали флуоресцентным красителем calcein AM Molecular Probes (Eugene, USA). Для этого клетки EL-4 переводили в полную ростовую среду DMEM с концентрацией 4 млн/мл и добавляли calcein AM до концентрации 10 мкМ. После инкубации в течение 30 мин при 37°С клетки три раза отмывали и ресуспендировали в полной среде в концентрации необходимой для проведения теста. В качестве эффекторных клеток использовали NK-клетки человека. Клетки периферических мононуклеаров выделяли из крови здоровых доноров по стандартной методике на градиенте фиколла. Фракцию периферических мононуклеаров дважды отмывали и ресуспендировали в буфере для сепарации. Выделения NK-клеток из периферических мононуклеаров проводили при помощи метода отрицательной магнитной сепарации с использованием набора для изоляции NK-клеток (Miltenyi Biotec, Германия). Сепарацию осуществляли при помощи колонки и магнита (MACS Columns and MACS Separator, Miltenyi Biotech, Германия). Чистота выделения NK-клеток составляла не менее 97%.
Оценку клеточно-опосредованной цитотоксичности антител B8(1)-H6 на клетках EL-4 проводили с использованием модифицированного метода высвобождения кальцеина [Lichtenfels et al., 1994; Neri et al., 2001]. Мембранно-проницаемый краситель calcein-AM при проникновении в клетку за счет активности внутриклеточных эстераз расщепляется до зеленого флуоресцентного красителя кальцеина. В результате лизиса клеток мишеней флуоресцентный краситель высвобождается в среду культивирования и интенсивность ее флуоресценции отражает уровень антитело-зависимой клеточно-опосредованной цитотоксичности.
При проведении теста calcein-AM-меченные клетки EL-4 рассевали в лунки 96-луночного планшета и добавляли серийные разведения антител B8(1)-H6. После инкубации с антителами к клеткам мишеням добавляли эффекторные клетки и продолжали инкубацию в течение 3-х часов. Соотношение клеток-мишеней и эффекторных клеток составляло 1:2. После инкубации клетки осаждали центрифугированием, собирали супернатант и измеряли интенсивность флуоресценции на планшетном спектрофлуориметре Glomax (Promega, США) при λвозб. = 490 нм и λ исп. = 510-570 нм.
Эффективность клеточно-опосредованной цитотоксичности антител определялась по формуле: % ADCC = 100*(MFI образца - MFI AICC)/(MFI максимального высвобождения красителя - MFI спонтанного высвобождения красителя).
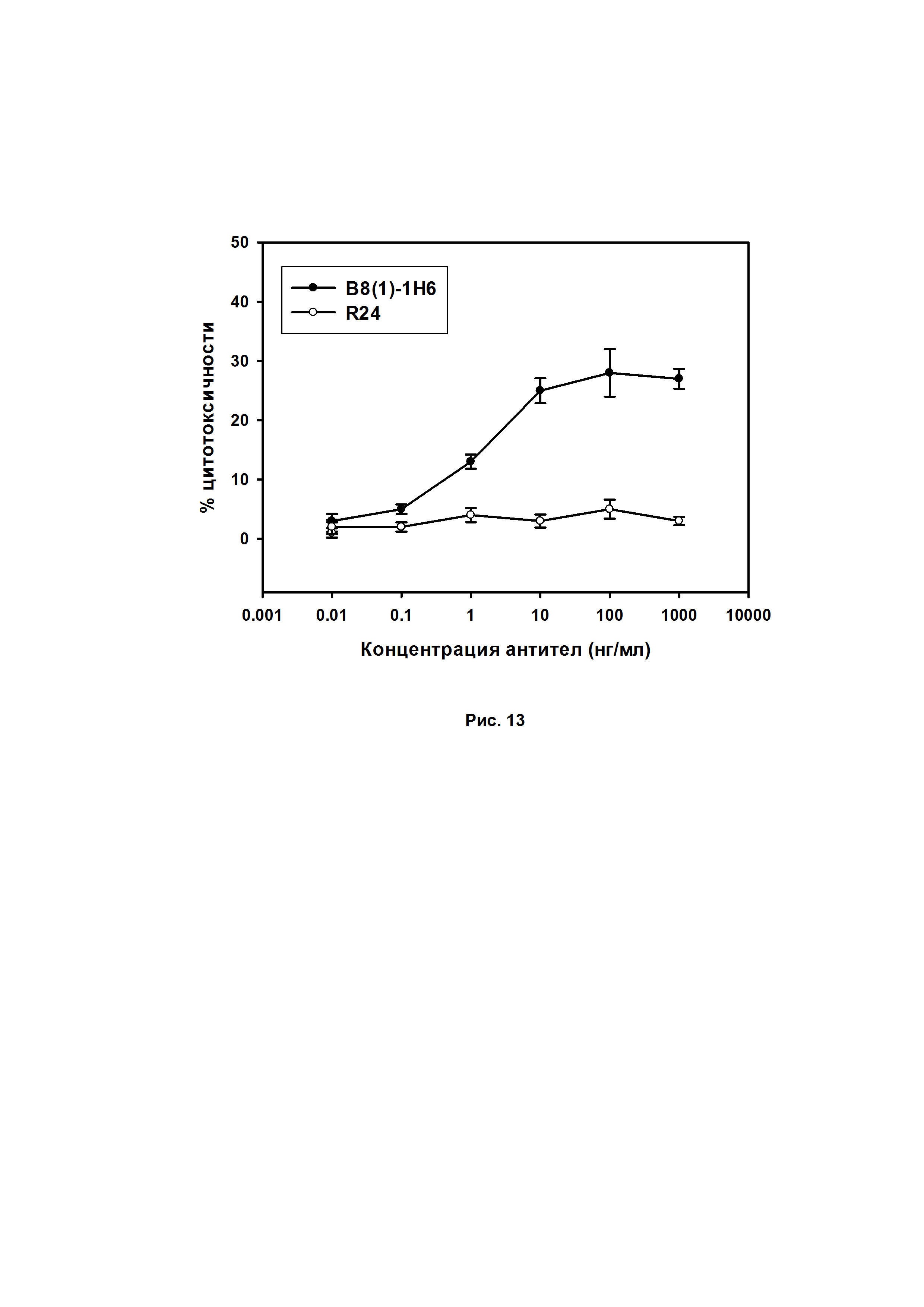
Где MFI - средняя интенсивность флуоресценции, AICC - антитело независимое высвобождение образца, MFI AICC определяется по измерению флуоресценции образца, в котором инкубировались клетки-мишени и эффекторные клетки без антител, MFI максимального высвобождения красителя определяется по измерению флуоресценции образца, в котором инкубировались клетки-мишени с добавлением 1% детергента Triton X-100, MFI спонтанного высвобождения красителя определяется по измерению флуоресценции образца, в котором инкубировались клетки-мишени без эффекторных клеток и антител. В качестве отрицательного контроля использовали антитела R24, специфичные к ганглиозиду GD3, не содержащегося на поверхности клеток EL-4.
Как видно из Рис. 13, антитела B8(1)-H6 опосредуют значимую цитотоксичность NK-клеток в GD2-позитивных клетках EL-4 в отличие от GD3-специфичных антител R24, которые не активировали ADCC даже при высоких концентрациях.
Несмотря на то, что изобретение описано со ссылкой на раскрываемые варианты воплощения, для специалистов в данной области должно быть очевидно, что конкретные подробно описанные эксперименты приведены лишь в целях иллюстрирования настоящего изобретения, и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения. Должно быть понятно, что возможно осуществление различных модификаций без отступления от сути настоящего изобретения.





Получение пегилированных фрагментов gd2-специфичных антител, индуцирующих прямую клеточную гибель gd2-позитивных опухолевых клеток, и их применение в терапии gd2-позитивных опухолей
Рекомбинантные однодоменные антитела, специфически связывающие белок f4/80, способ их получения и использования для детекции этого белка
Получение пегилированных фрагментов gd2-специфичных антител, индуцирующих прямую клеточную гибель gd2-позитивных опухолевых клеток, и их применение в терапии gd2-позитивных опухолей

