Результат интеллектуальной деятельности: Способ эффективного биосинтеза рекомбинантных белков в двудольных растениях с использованием промотора гена pro-SmAMP1 из растения Stellaria media
Вид РИД
Изобретение
Область техники, к которой относится изобретение
Изобретение относится к области биологии, биотехнологии и генетической инженерии растений, а именно, к способу достижения высокой экспрессии целевых генов и повышенного накопления целевого белка в клетках растений. Настоящее изобретение может применяться для наработки рекомбинантных белков в растениях методом транзиентной экспрессии с целью эффективного производства белковых препаратов для медицинской, фармацевтической, ветеринарной и пищевой промышленности. Настоящее изобретение может использоваться в фундаментальных научных исследованиях.
Уровень техники
В настоящее время из широкого круга организмов выделены промоторы многих генов, которые используются в растительных генно-инженерных системах. Промоторы для биотехнологии растений базируются на схожих последовательностях нуклеотидов, которые, как правило, включают сайт инициации транскрипции и TATA-бокс в качестве специфических цис-активных мотивов, связывающих факторы транскрипции.
В последние годы стали доступны для экспрессии гетерологичных генов в растениях несколько хорошо охарактеризованных промоторов. В генетической инженерии растений наиболее широко применяется сильный промотор CaMV35S, созданный на основе промоторной области 35S РНК вируса мозаики цветной капусты CaMV (Potenza et al. 2004). Благодаря широкому использованию в растительных векторах CaMV35S обычно все новые промоторы сопоставляют с ним по своей эффективности. Полноразмерный регуляторный участок для CaMV35S по размеру приближается к 3 тыс. п.н. (Odell et al. 1985). Однако, промоторной активностью обладают и меньшие по размеру участки: типичным промотором CaMV35S, используемым в экспрессионных векторах для трансформации растений, является делеционный вариант длиной в 352 п.н. (Fang et al. 1989). В большинстве случаев промотор CaMV35S обеспечивает высокий уровень экспрессии гетерологичных генов в растениях, тем не менее, он имеет ряд существенных недостатков. Из-за вирусного происхождения CaMV35S инфицирование трансгенных растений вирусом CaMV может приводить к сайленсингу гетерологичного гена под контролем CaMV35S (Al-Kaff et al. 2000).
Для однодольных растений известны сильные промоторы генов растений, которые при транзиентной экспрессии по активности достоверно превосходят CaMV35S. Промотор Actin1 из риса (Oryza sativa) запускает транзиентную экспрессию репортерного гена gusA в клетках риса и кукурузы в двадцать раз сильнее, чем CaMV35S (McElroy et al. 1991). Промотор гена ACT1 из банана (Musa spp.) более чем в два раза превосходил по активности CaMV35S при транзиентной экспрессии в клетках банана (Hermann et al. 2001).
Для генетической инженерии двудольных растений также были найдены сильные и конститутивные растительные промоторы, но превосходство их над промотором CaMV35S было не так существенно, как у однодольных. Промотор гена актина ACT2 из Arabidopsis thaliana был в два раза сильнее при транзиентной экспрессии в клетках арабидопсиса, чем CaMV35S (An et al. 2010). Промоторы UBQ1 и UBQ6 из A. thaliana активны во всех тканях табака (Nicotiana tabacum) на уровне вирусного промотора CaMV35S (Callis et al. 1990).
Несмотря на обилие литературных данных по активности промоторов многих генов, в настоящее время имеется дефицит эффективных промоторов для высокоуровневой транзиентной экспрессии целевых генов.
В наших исследованиях мы обратили внимание на группу генов растений, которые могут стать источником новых эффективных промоторов для транзиентной экспрессии. Ранее было установлено, что экспрессия генов антимикробных пептидов сорного растения Stellaria media увеличивается при взаимодействии растения с фитопатогенными грибами и при обработке растений метилжасмонатом (Shukurov et al. 2012). Экспрессия гена антимикробных пептидов pro-SmAMP1 увеличивалась от 10 до 100 раз, при этом достигая уровня экспрессии гена «домашнего хозяйства» β-актина. Экспрессия гена pro-SmAMP2 более подходила под определение конститутивной, изменялась не так сильно под воздействием вышеперечисленных факторов (в 2-5 раз), но при этом её уровень сопоставим или превосходил уровень экспрессии β-актина.
С целью поиска нового растительного промотора для биотехнологии растений была секвенирована и клонирована нуклеотидная последовательность промоторной области гена pro-SmAMP2 из S. media. Для изучения ее эффективности в качестве промотора при экспрессии целевых генов в гетерологичных растениях методом транзиентной экспрессии, были созданы два 5'-делеционных варианта -2160 и -862 п.н., включающие в себя также 5'-нетранслируюмую область pro-SmAMP2. Эти делеционные варианты были слиты с последовательностью репортерного гена gusА в растительном экспрессионном векторе pCambia1381Z и изучены методом транзиентной экспрессии в клетках растений Nicotiana benthamiana. Измерения активности GUS показали, что делеционный вариант -2160 п.н. pro-SmAMP2 обладает низкой эффективностью, а делеционный вариант -862 п.н. сопоставим по эффективности с вирусным промотором CaMV35S (Стрельникова и др. 2014).
Также была определена нуклеотидная последовательность фрагмента промоторной области гена pro-SmAMP1 длиной 1257 п.н. С учетом распределения цис-элементов было создано пять 5'-делеционных вариантов -1235, -771, -714, -603 и -481 п.н. промотора рro-SmAMP1, которые были слиты с кодирующей областью репортерного гена gusA в растительном экспрессионном векторе pCambia1381Z. Эффективность всех делеционных вариантов промотора pro-SmAMP1 определяли методом транзиентной экспрессии в растениях N. benthamiana. Уровни активности репортерного белка GUS в экстрактах из агроинфильтрированных растений при использовании всех делеционных вариантов промотора рro-SmAMP1 были существенно выше, чем при применении вирусного промотора CaMV35S (Высоцкий и др. 2016).
Для сравнительной оценки эффективности в растениях N. benthamiana использовали делеционный вариант -481 п.н. промотора pro-SmAMP1, делеционный вариант -495 п.н. промотора pro-SmAMP2 (Komakhin et al. 2016) и в качестве контроля вирусный промотор CaMV35S. С использованием агробактериальной инфильтрации установлено, что средние значения активности репортерного белка GUS составили при использовании промотора pro-SmAMP1 - 41600±3400 пмоль/мг/мин, промотора pro-SmAMP2 - 28400±2000 пмоль/мг/мин и вирусного промотора CaMV35S - 12000±900 пмоль/мг/мин. Сравнительные результаты транзиентной экспрессии репортерного гена gusА под контролем промоторов pro-SmAMP1, pro-SmAMP2 и CaMV35S в растениях N. benthamiana свидетельствуют, что уровни абсолютной активности GUS при использовании всех промоторов были примерно в 1.5-4.6 раз ниже, чем в наших более ранних исследованиях (Стрельникова и др., 2014; Высоцкий и др., 2016). Различия в уровнях активности при использовании одного и того же варианта промотора могут быть объяснены разным физиологическим состоянием растений или агробактерий в момент проведения экспериментов. Ранее в наших исследованиях транзиентная экспрессия gusА под контролем разных промоторов в N. benthamiana была выполнена в конце лета и начале осени (Стрельникова и др., 2014; Высоцкий и др., 2016), в то время как в данных исследованиях эксперименты проводились в конце зимы и весной. Косвенным подтверждением разного физиологического состояния растений может служить то, что весной они зацветали на 1-1.5 недели раньше, чем осенью. В тоже время во всех независимых повторениях соотношение уровней активности GUS при использовании разных промоторов оставалось постоянной величиной: самым эффективным оставался делеционный варианта -481 п.н. промотора pro-SmAMP1, существенно менее эффективным был делеционный вариант -495 п.н. промотора pro-SmAMP2 и наименее эффективным оказывался вирусный промотор CaMV35S (Маджарова и др. 2018).
Транзиентную экспрессию генов часто используют для получения каких-либо рекомбинантных белков с использованием растений N. benthamiana. Такой способ не приводит к созданию трансгенных растений, трансген не может передаваться в поколениях растений и попасть в окружающую среду через перекрестное опыление между близкими видами; при этом работа таким способом не создает проблем с сертификацией трансгенных растений. Поэтому метод транзиентной экспрессии имеет потенциал для масштабного получения различных целевых рекомбинантных белков. Однако необходимым условием для эффективной транзиентной экспрессии является создание генетических конструкций с промоторами генов растений, обеспечивающими высокий выход целевого белкового продукта.
Раскрытие сущности изобретения
Предлагаемый способ основан на инфильтрации растений с использованием клеток бактерий Agrobacterium tumefaciens, содержащих генетические конструкции с новыми делеционными -388, -364, -328, -276, -226 и -158 п.н. вариантами промотора pro-SmAMP1 из растения мокрица (Stellaria media), контролирующими целевой ген, и nos терминатор транскрипции. Способ транзиентной экспрессии целевого гена с использованием новых делеционных вариантов промотора pro-SmAMP1 обеспечивает статистически более высокий уровень накопления в инфильтрированных растениях кодируемого целевого белка, чем ранее использованные делеционные варианты промотора pro-SmAMP1 (Высоцкий и др. 2016) из мокрицы, промотора pro-SmAMP2 из мокрицы (Komakhin et al. 2016; Маджарова и др. 2018) и вирусного промотора CaMV35S. Нуклеотидные последовательности новых делеционных вариантов промоторной области гена pro-SmAMP1, использованные в данном способе транзиентной экспрессии целевых генов, представлены как -388 п.н. (SEC ID NO 1), -364 п.н. (SEC ID NO 2), -328 п.н. (SEC ID NO 3), -276 п.н. (SEC ID NO 4), -226 п.н. (SEC ID NO 5) и -158 п.н. (SEC ID NO 6). Цифровое обозначение -388, -364, -328, -276, -226 и -158 указывает количество пар нуклеотидов делеционного варианта промотора относительно сайта инициации трансляции (инициирующего кодона ATG, в котором А представлен в позиции +1) и включает 5'-нетранслируемую область гена pro-SmAMP1 размером 39 п.н.
Объектом настоящего изобретения является последовательность ДНК длиной -388 п.н. (SEC ID NO 1), представляющая собой вариант промоторной области гена pro-SmAMP1 из Stellaria media и включающая его 5'-нетраслируемую область. Данный вариант промотора pro-SmAMP1 при транзиетной экспрессии в растениях обеспечивает с вероятностью 95% значительно более высокий уровень экспрессии целевого гена, чем созданный ранее его более протяженный вариант -481 п.н. (Высоцкий и др. 2016) и вирусный промотор CaMV35S. Кроме этого, нуклеотидная последовательность -388 п.н. (SEC ID NO 1) промотора pro-SmAMP1 на 93 п.н. короче, чем нуклеотидная последовательность ранее созданного делеционного варианта -481 п.н. промотора pro-SmAMP1, что важно при создании генетических конструкций с минимизированным размером экспрессионных кассет.
Вариантами настоящего изобретения являются также последовательности ДНК, соответствующие делеционным вариантам промоторной области гена pro-SmAMP1 длиной -364 п.н. (SEC ID NO 2), -328 п.н. (SEC ID NO 3), -276 п.н. (SEC ID NO 4), -226 п.н. (SEC ID NO 5) и -158 п.н. (SEC ID NO 6), обеспечивающие с вероятностью 95% более высокий уровень экспрессии целевого гена в растениях, чем делеционный вариант -481 п.н. pro-SmAMP1 и вирусный промотор CaMV35S. Делеционные варианты -364 п.н. (SEC ID NO 2), -328 п.н. (SEC ID NO 3), -276 п.н. (SEC ID NO 4), -226 п.н. (SEC ID NO 5) и -158 п.н. (SEC ID NO 6) промотора pro-SmAMP1 короче ранее изученного варианта -481 п.н. промотора pro-SmAMP1 на 117-323 п.н., что важно при создании генетических конструкций с минимизированным размером экспрессионных кассет. В целом участок ДНК, представляющий собой вариант промоторной области гена pro-SmAMP1 S. media, соединенный с его 5'-нетранслируемой областью, где указанный фрагмент ДНК имеет нуклеотидную последовательность SEC ID NO 1.
Объектами настоящего изобретения также являются генетические конструкции для экспрессии в растениях целевых генов, находящихся под контролем указанных выше делеционных вариантов промотора pro-SmAMP1.
Настоящим изобретением является способ (процесс) наработки целевых белков в тканях растений при транзиентной экспрессии целевых генов под контролем любой из нуклеотидных последовательностей -388 п.н. (SEC ID NO 1), -364 п.н. (SEC ID NO 2), -328 п.н. (SEC ID NO 3), -276 п.н. (SEC ID NO 4), -226 п.н. (SEC ID NO 5) и -158 п.н. (SEC ID NO 6), находящихся в составе генетических конструкций.
Основной технический результат изобретения заключается в расширении арсенала новых эффективных промоторов для генетической инженерии растений, с использованием которых происходит достоверно более высокий уровень накопления целевых белков при транзиентной экспрессии целевых генов.
Краткое описание чертежей
Фиг. 1. Нуклеотидная последовательность 5'-фланкирующей промоторной области гена pro-SmAMP1 размером -481 п.н. (относительно сайта инициации трансляции ATG, в котором А представлен в позиции +1) и распределение цис-действующих элементов, выделенных цветом и обозначенных надписями. 5'-нетранслируемая область расположена между сайтами инициации транскрипции (TSS) и инициации трансляции (ATG). Сайт начала трансляции выделен красным курсивом. Вертикальными красными стрелками и цифрами отмечены начальные точки нуклеотидных последовательностей новых 5'-делеционных вариантов промотора. Вертикальной черной стрелкой и цифрами «-481» отмечена начальная точка нуклеотидной последовательности созданного ранее 5'-делеционного варианта -481 п.н. промотора (Высоцкий и др. 2016).
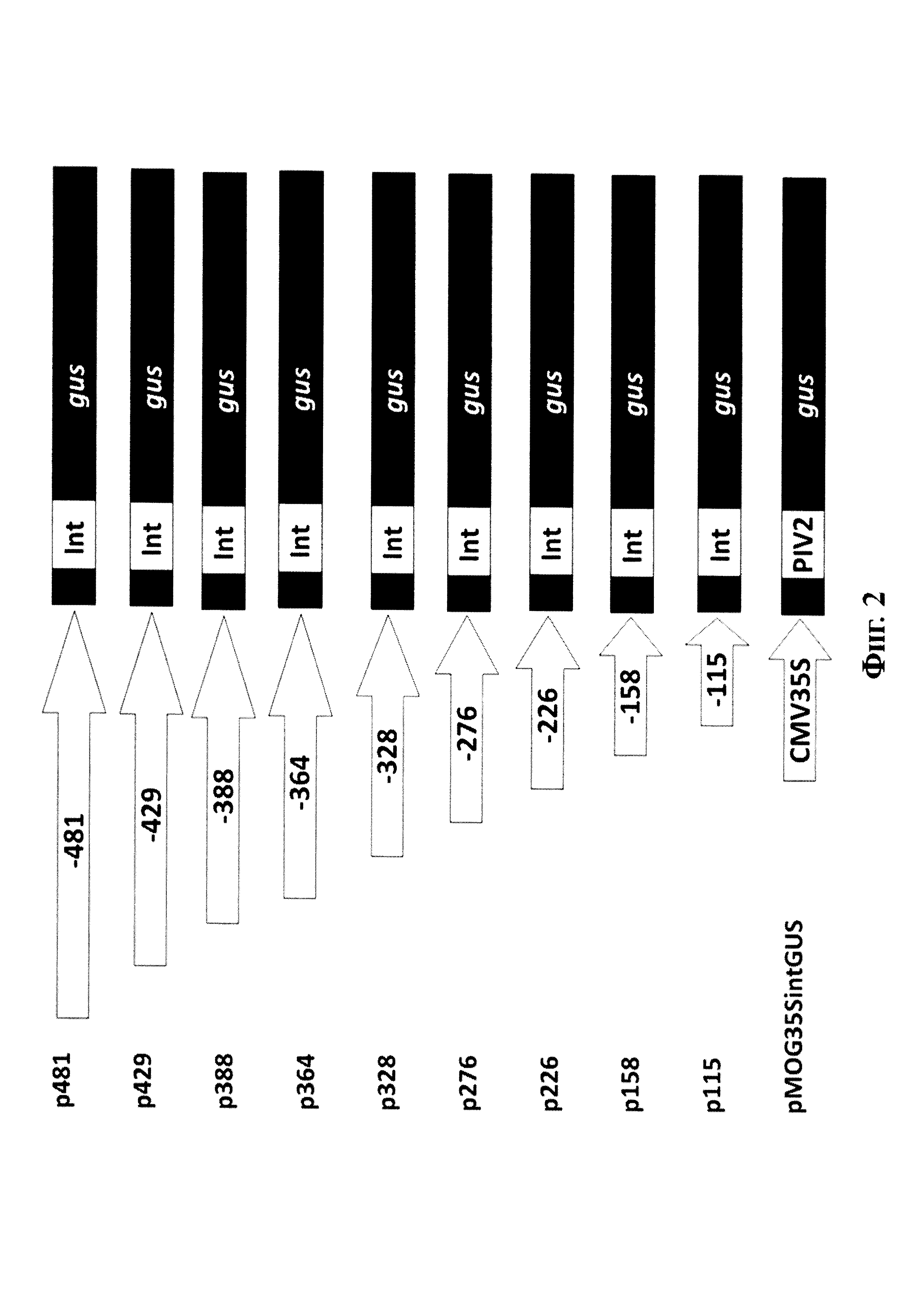
Фиг. 2. Схема генетических конструкций на основе растительного экспрессионного вектора pCAMBIA 1381Z в которых репортерный ген gusА (gus) находится под контролем новых делеционных вариантов промотора pro-SmAMP1. Генетическая конструкция p481 создана ранее (Высоцкий и др. 2016) и использована в качестве сравнительного контроля. Растительный экспрессионный вектор pMOG35SintGUS содержит ген gusА под контролем вирусного промотора CaMV35S и использован в качестве контроля. Черным показана транслируемая область гена gusА; Int - модифицированный интрон каталазы клещевины внутри транслируемой области гена gus; PIV2 - модифицированный интрон гена ST-LS1 картофеля внутри транслируемой области гена gusА. Промоторы изображены в виде стрелок с соответствующими подписями.
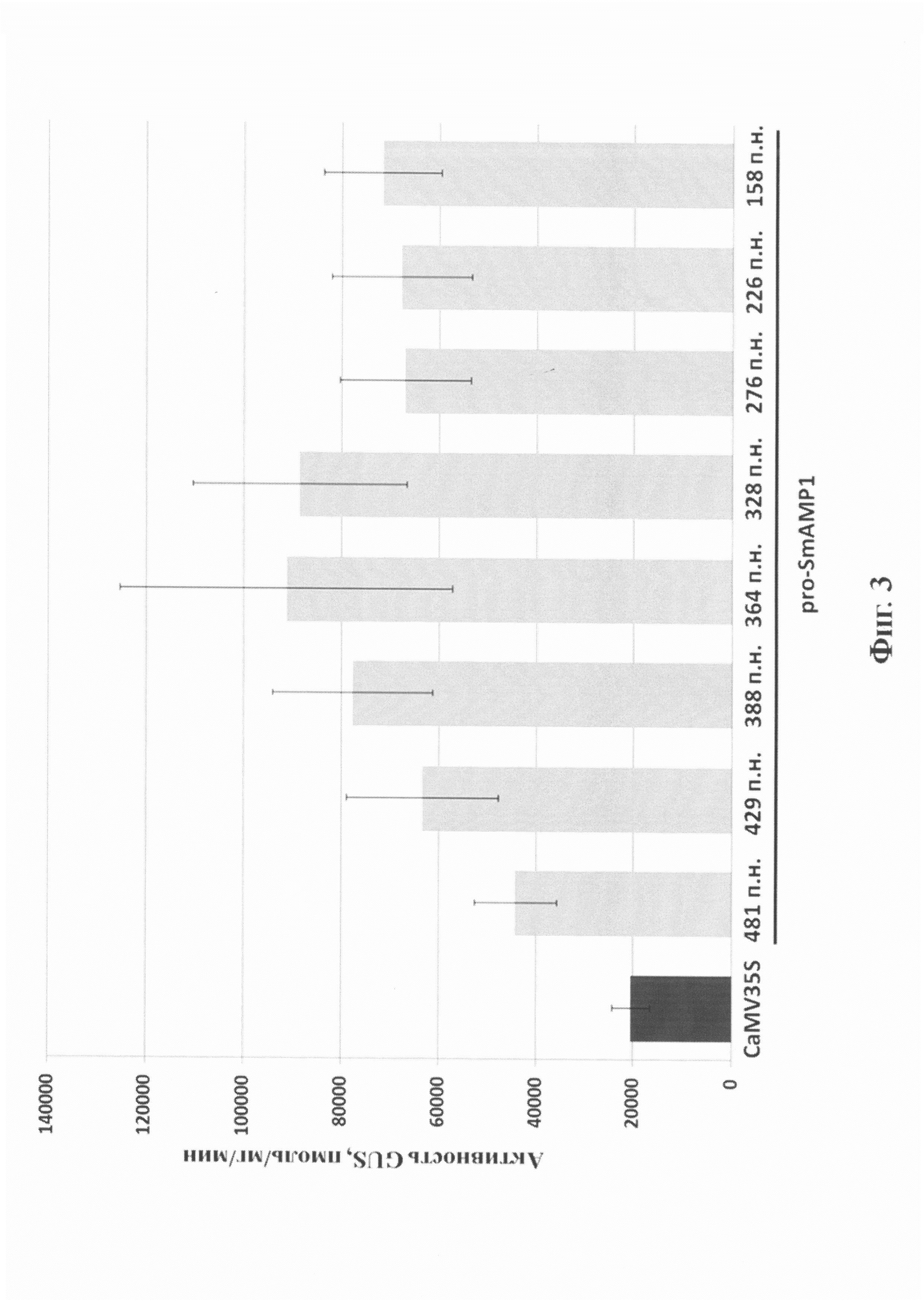
Фиг. 3. Активность репортерного белка бета-глюкуронидазы (GUS) в листьях растений N. benthamiana при транзиентной экспрессии гена gusА под контролем различных делеционных вариантов промотора pro-SmAMP1 и вирусного промотора CaMV35S. Горизонтальной линией отмечены делеционные варианты на основе промотора pro-SmAMP1 из растения мокрицы. Вертикальными линиями обозначен доверительный интервал на 5% уровне значимости. Цифрами ниже оси абсцисс показаны индивидуальные делеционные варианты промоторов, рассчитанные относительно старта инициации транскрипции. Активность бета-глюкуронидазы измерена флуориметрическим способом, используя 4-метилумбеллиферил-D-глюкуронид в качестве субстрата.
Осуществление изобретения
Приведенное выше раскрытие описывает изобретение в общих чертах. Более полное понимание может быть достигнуто за счет приведения соответствующих примеров. Представленные здесь примеры приводятся исключительно с целью иллюстрации и никоим образом не претендуют на то, чтобы сузить область данного изобретения в рамках этих примеров. Изменение формы или замещение эквивалентов допускается на основании его целесообразности в соответствующих условиях. Хотя здесь и будут применяться специфические термины, они используются исключительно с описательной целью.
Пример 1. Выбор новых делеционных вариантов промоторной области гена pro-SmAMP1
Этот пример раскрывает особенности создания новых делеционных вариантов промотора pro-SmAMP1, размер нуклеотидных последовательностей которых значительно меньше, чем у самого короткого из ранее полученных вариантов.
Ранее в растении мокрица (S. media) была определена наименьшая нуклеотидная последовательность фрагмента промоторной области гена антигрибных пептидов pro-SmAMP1 длиной -481 п.н. (Высоцкий и др. 2016). Биоинформационный анализ в программах PLACE (http://www.dna.affrc.go.jp/PLACE/signalscan.html) и PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) показал, что данный делеционный вариант промотора pro-SmAMP1 содержит консервативную последовательность TATA-бокса, CAAT-мотив и последовательность CAN(A/C)(A/C)(C/A)C(C/A)N2A(C/A), характеризующую сайт инициации транскрипции TSS (на фиг. 1). Сочетание этих мотивов характерно для промоторов, обеспечивающих высокий уровень экспрессии генов в растениях (Sawant et al. 1999). Кроме того, выявлен целый ряд цис-элементов и регуляторных мотивов, которые могут обуславливать определенный характер экспрессии генов. Так, были обнаружены регуляторные элементы, отвечающие за реакцию на свет, такие как TGA-элемент, I-бокс, G-бокс и LAMP-элемент. Кроме того, обнаружены мотивы, ответственные за реакцию на действие фитопатогенов (S-бокс, W-бокс, TGACG-мотив и др.), абиотические воздействия (ARE, ABRE), а также определяющие локальный характер экспрессии (GCN4, Skn-I). Необходимо отметить, что положение и количество обнаруженных регуляторных элементов отличает промотор pro-SmAMP1 от известного промотора pro-SmAMP2 (Komakhin et al. 2016). К примеру, TGA-элемент, ответственный за реакцию на свет, у промотора pro-SmAMP1 находится в другой позиции, чем у pro-SmAMP2. LAMP-элемент есть только в промоторе pro-SmAMP1.
Эффективность делеционного варианта -481 п.н. промотора pro-SmAMP1 при транзиентной экспрессии в растениях N. benthamiana была достоверно выше, чем при применении вирусного промотора CaMV35S (Высоцкий и др. 2016). В тоже время осталась неясна эффективность делеционных вариантов промотора pro-SmAMP1, размер нуклеотидной последовательности которых был меньше -481 п.н. относительно сайта инициации трансляции (инициирующего кодона ATG). Для обнаружения минимального активного делеционного варианта промотора pro-SmAMP1, содержащего минимальное количество цис-действующих элементов выполнено последовательное усечение созданного ранее делеционного варианта -481 п.н. с 5'-конца с шагом до 70 п.н., но с таким расчетом, чтобы ни вносить разрывы в цис-действующие элементы, обнаруженные компьютерными программами (на фиг. 1). Всего было выбрано восемь новых делеционных вариантов промотора pro-SmAMP1.
Пример 2. Создание генетических конструкций с новыми делеционными вариантами промотора pro-SmAMP1
На основе проведенного анализа последовательности делеционного варианта -481 п.н. pro-SmAMP1 были подобраны праймеры для создания новых более коротких делеционных вариантов промотора с целью последующей оценки их эффективности таким образом, чтобы прямые праймеры в сочетании с обратным последовательно ограничивали цис-действующие элементы. Было подобрано восемь прямых праймеров и один обратный, ограничивающие области промотора, обозначенные как -429, -388, -364, -328, -276, -226, -158 и -115 п.н. Обратный праймер был подобран непосредственно слева от сайта инициации трансляции, начиная с нуклеотидного остатка в положении -1. Последовательности праймеров приведены ниже:
-429f - 5'- GAATTCTAGAGCATCGTCAATAAA -3'
-388f - 5'- GAATTCCAATCTCGATAATACATTTT -3'
-364f - 5'- GAATTCAAATCACCCGATAACACT -3'
-328f - 5'- GAATTCTATATAGCCTTTATCTTTATCTCG -3'
-276f - 5'- GAATTCAGTCTATCCGTATAGACCCT -3'
-226f - 5'- GAATTCCAAGATATTAAAGTGTGTGT -3'
-158f - 5'- GAATTCGGTTATCATCAAGCATTT -3'
-115f - 5'- GAATTCTGCAAACGGCAAACC -3'
rev - 5'- CCATGGTTTCACTTGATTTTTTTG -3'
Все прямые праймеры содержали сайт рестрикции EcoRI, а обратный - NcoI, необходимые для клонирования в вектор pCAMBIA 1381Z.
В качестве матрицы для амплификации новых делеционных вариантов промотора pro-SmAMP1 использовали созданную ранее плазмидную генетическую конструкцию p481 (Высоцкий и др. 2016).
С использованием разработанных праймеров были амплифицированы и клонированы в экспрессионный вектор pCAMBIA 1381Z восемь новых делеционных вариантов промотора pro-SmAMP1. Схема полученных конструкций, обозначенных, соответственно, р429, р388, р364, р328, р276, р226, р158 и р115 отображена на фиг. 2.
Созданные генетические конструкции предназначены для транзиентной экспрессии целевого гена в растениях N. benthamiana, находящегося под контролем заявленных вариантов промотора pro-SmAMP1. В качестве целевого гена использован репортерный ген gusA, позволяющий оценить эффективность транзиентной экспрессии по активности его белкового продукта - фермента β-глюкуронидазы (GUS).
Пример 3. Агроинфильтрация растений N. benthamiana
Инфильтрацию листьев растений проводили десятью вариантами A. tumefacien штамма GV3101 с плазмидными генетическими конструкциями p429, р388, p364, р328, р276, р226, р158 и р115, а также с pMOG35SintGUS и с р481 (Высоцкий и др., 2016) в качестве сравнительных контролей. Генетическая конструкция pMOG35SintGUS содержит репортерный ген gusA под контролем вирусного промотора CaMV35S. Конструкция p481 содержит репортерный ген gusA под контролем изученного ранее делеционного варианта -481 п.н. промотора pro-SmAMP1 (Высоцкий и др. 2016). Полученные агробактерий штаммы выращивали на агаризованной среде LB с антибиотиками 100 мг/л канамицина, 100 мг/мл рифампицина и 25 мг/л гентамицина. Для проведения агроинфильтрации растений штаммы выращивали в жидкой среде LB в течение суток с добавлением антибиотиков 100 мг/л канамицина, 100 мг/мл рифампицина и 25 мг/л гентамицина при температуре 28°С и 160 об/мин. Штамм A. tumifaciens GV2260/C58C1 pLH7000 р19 выращивали в течении суток на среде LB с антибиотиками 100 мг/л рифампицина, 50 мг/л спектиномицина, 50мг/л стрептомицина, при температуре 28°С и 160 об/мин. и использовали для подавления РНК интерференции в клетках растений.
Агробактерии разводили до оптической плотности 0.6 при длине волны 600 нм. Полученные суспензии GV3101 pMOG35SintGUS, GV3101 р481, GV3101 p429, GV3101 р388, GV3101 p364, GV3101 р328, GV3101 р276, GV3101 р226, GV3101 р158 и GV3101 р115 смешивали в соотношение 1:1 с суспензией агробактерий GV2260/C58C1 pLH7000 р19. Готовую смесь агробактерий вводили в листья растений N. benthamiana шприцом без иглы с внутренней стороны листа. После инфильтрации растения содержали при температуре 26°C и 16-ти часовом искусственном освещении. На 7 день из места инокуляции брали высечки массой 10 мг и замороживали при -70°С.
Из одной высечки с каждого растения получали белковые экстракты, в которых измеряли активность фермента β-глюкуронидазы. Во всех проанализированных растениях, кроме инфильтрированных вариантом GV3101 р115, обнаружена флуоресценция продуктов гидролиза MUG (4-methylumbelliferyl-β-D-glucuronide Trihydrate, PhytoTechnology Laboratories) белком β-глюкуронидазой.
Таким образом, получены растения N. benthamiana, инфильтрированные агробактериальными штаммами GV3101 pMOG35SintGUS, GV3101 р481, GV3101 p429, GV3101 р388, GV3101 p364, GV3101 р328, GV3101 р276, GV3101 р226, GV3101 р158, продуцирующие β-глюкуронидазу.
Пример 4. Оценка эффективности промотора pro-SmAMP1 по активности репортерного белка GUS при транзиентной экспрессии
Уровни активности репортерного белка GUS в листьях агроинфильтрированных растений N. benthamiana сравнивали при использовании различных делеционных вариантов промотора pro-SmAMP1 и промотора CaMV35S. Первоначально из каждой высечки экстрагировали фракцию водорастворимых белков, которую использовали для определения уровня флуоресценции продуктов гидролиза субстрата 4MUG, полученных в результате активности репортерного белка GUS, затем в ней определяли концентрацию водорастворимых белков. Полученные результаты по флуоресценции продуктов гидролиза 4MUG и содержанию белка использовали для расчета уровня активности GUS.
С использованием полученных результатов по каждой высечке рассчитывали уровень активности GUS в пмоль4MU/мг/мин (или ед.а.) для каждого отдельного варианта генетической конструкции. Всего было проанализировано 180 образцов, в том числе при использовании GV3101 pMOG-35SintGus - 20 шт., GV3101 p481 - 20 шт., GV3101 p429 - 20 шт., GV3101 p388 - 20 шт., GV3101 p364 - 20 шт., GV3101 p328 - 20 шт., GV3101 p276 - 20 шт., GV3101 p226 - 20 шт. и GV3101 p158 - 20 шт. Средний уровень активности GUS при использовании каждого делеционного вариантов промотора pro-SmAMP1 и вирусного промотора CaMV35S представлен на фиг. 3.
Как следует из фиг. 3, средний уровень активности GUS в растениях, агроинфильтрированных генетическими конструкциями с различными делеционными вариантами промотора pro-SmAMP1, составлял от 44000 до 91000 ед.а. В частности, делеционный вариант -481 п.н. позволял достигать 44297±8431 ед.а., вариант -429 п.н. - 63433±15521 ед.а., вариант -388 п.н. - 77774±16380 ед.а., вариант -364 п.н. - 91273±34068 ед.а., вариант -328 п.н. - 88625±21868 ед.а., вариант -276 п.н. - 67034±13495 ед.а., вариант -226 п.н. - 67798±14436 ед.а. и вариант -158 п.н. - 71674±12089 ед.а. При этом с использование вирусного промотора CaMV35S удалось достичь только 20520±3937 ед.а. После знака «±» для каждого варианта представлен доверительный интервал значений активности при p=05.
Как следует из приведенных результатов, значения уровней активности GUS при использовании всех делеционных вариантов промотора pro-SmAMP1 с вероятностью 95% более чем два раза превышали уровни активности GUS с применением вирусного промотора CaMV35S. Сравнительный анализ показал, что наименее эффективным среди делеционных вариантов pro-SmAMP1 является созданный ранее вариант -481 п.н. (Высоцкий и др. 2016) и статистически достоверно не отличается от него новый делеционный вариант -429 п.н. Остальные новые делеционные варианты -388, -364, -328, -276, -226 и -158 п.н. промотора pro-SmAMP1 достоверно превосходят вариант -481 п.н., но между собой по эффективности не различаются (на фиг. 3).
Ранее нами был клонирован промотор гена pro-SmAMP2 из генома двудольного растения S. media. Он так же, как промотор гена pro-SmAMP1, является представителем регуляторных элементов, контролирующих экспрессию генов антимикробных пептидов. Уровень экспрессии репортерного гена gus в гетерологичных растениях N. benthamiana при использовании делеционного варианта -862 п.н. промотора proSmAMP2 был выше, чем при использовании промотора CaMV35S, но эти различия были не статистически значимыми при p=05 (Стрельникова и др. 2014).
При транзиентной экспрессии репортерного гена gus в агроинфильтрированных растениях N. benthamiana все делеционные варианты от -1235 до -481 п.н. (Высоцкий и др. 2016) и от -388 до -158 п.н. промотора pro-SmAMP1 из мокрицы были существенно более эффективными, чем вирусный промотор CaMV35S. Каждый из новых делеционных вариантов -388, -364, -328, -276, -226 и -158 п.н. промотора pro-SmAMP1 может быть использован для транзиентной экспрессии генов и предпочтителен для создания генетических конструкций. Все новые делеционные варианты обладают достоверно более высокой эффективностью, чем ранее изученный делеционный вариант -481 п.н., а длина их нуклеотидных последовательностей короче на 93-323 п.н. Для инфильтрации N. benthamiana предпочтительнее использовать генетические конструкции с делеционными вариантами от -388 до -158 п.н., чем -481 п.н. промотора pro-SmAMP1. Применение делеционных вариантов промотора pro-SmAMP1 для биотехнологии растений приоритетно с точки зрения экологической безопасности, поскольку они могут заменить в генетических конструкциях для трансформации растений промоторы вирусов, бактерий или животных.
Список литературы
1. Al-Kaff N. S., Kreike M. M., Covey S. N., Pitcher R., Page A. M., Dale P. J. (2000) Plants rendered herbicide-susceptible by cauliflower mosaic virus-elicited
suppression of a 35S promoter-regulated transgene // Nat Biotechnol. 18(9). P. 995-999.
2. An Y.-Q.C., Meagher R.B. (2010) Strong expression and conserved regulation of ACT2 in Arfbidopsis thaliana and Physcomitrella patens // Plant molecular biology reporter. Voi 28. №3. P. 484-490
3. Fang R.X., Nagy F., Sivasubramaniam S., Chua N.H. (1989) Multiple cis regylatory elements for maximal expression of the cauliflower mosaic virus 35S promoter in transgenic plants // Plant Cell. 1(1). P. 141-151.
4. Callis J., Raasch J.A., Vierstra R.D. (1990) Ubiquitin extension proteins of Arabidopsis thaliana. Structure, localization, and expression of their promoters in transgenic tobacco. // Journal of Biological Chemistry. Vol. 265. №21. P. 12486-12493.
5. Komakhin R.A., Vysotskii D.A., Shukurov R.R., Voblikova V.D., Komakhina V.V., Strelnikova S.R., Vetchinkina E.M., Babakov A.V. (2016) Novel strong promoter of antimicrobial peptides gene pro-SmAMP2 from chickweed (Stellaria media). // BMC Biotechnol. 18; 16(1):43. doi: 10.1186/s12896-016-0273-x.
6. McElroy D., Blowers A.D., Jenes B., Wu R. (1991) Construction of expression vectors based on the rice actin 1 (Act1) 50 region for use in monocot transformation. Mol Gen Genet 231:150-160.
7. Hermann S., Harding R., Dale J. (2001) The banana actin 1 promoter drives near-constitutive transgene expression in vegetative tissues of banana (Musa spp.) // Plant Cell Reports. Vol. 20. №6. P. 525-530.
8. Potenza C., Aleman L., Sengupta-Gopalan C. (2004) Targeting transgene expression in research, agricultural, and environmental pplication: Promoters used in plant transformation // In Vitro CellDevBiol-Plant. 40(1). P. 1-22.
9. Odell J.T., Nagy F., Chua N.H. (1985) Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter // Nature. 313 (6005). P. 810-812.
10. Sawant S., Singh P., Gupta S., Madnala R., Tuli R. (1999) Conserved nucleotide sequences in highly expressed genes in plants. Journal of Genetics 78:123-131 doi:10.1007/bf02924562.
11. Высоцкий Д.А., Стрельникова С.Р., Ефремова Л.Н., Ветчинкина Е.М., Бабаков А.В., Комахин Р.А. (2016) Структурно-функциональный анализ нового растительного промотора pro-SmAMP1 из Stellaria media // Физиология растений. Т. 63. №5. с. 705-715.
12. Стрельникова С.Р., Вобликова В.Д., Шукуров Р.Р., Бабаков А.В., Комахин Р.А. (2014) Изучение нового растительного промотора гена proSmAMP2 из Stellaria media методом агробактериальной инфильтрацией растений. Биотехнология: 8-17.







Стимулятор роста растений
Генетическая конструкция на основе системы редактирования генома crispr/cas9, кодирующая нуклеазу cas9, для геномного редактирования однодольных зерновых культур на основе двойного отбора растений
Сильный растительный промотор pro-smamp2 из сорного растения stellaria media
Способ получения трансформированных растительных клеток, содержащих рекомбинантную щелочную фосфатазу человека, и применение трансформированных растительных клеток, содержащих рекомбинантную щелочную фосфатазу человека

