Результат интеллектуальной деятельности: САЙТЫ АНТИТЕЛ, СПЕЦИФИЧНО СВЯЗЫВАЮЩИЕ EGFRvIII
Вид РИД
Изобретение
Настоящее изобретение относится к связывающим белкам, которые специфически связываются с EGFRvIII и к мультиспецифичным связывающим белкам, которые специфически связываются с EGFRvIII и CD3. Также изобретение относится к мультиспецифичному тандемному диателу, которое связывается с EGFRvIII и CD3. В частности, настоящее изобретение относится к биспецифическому тандемному диателу (TandAb®) EGFRvIII/CD3 с высокой цитотоксичностью для рекрутинга Т-лимфоцитов для специфичного и мощного уничтожения солидных раковых опухолей нескольких типов.
Была показана взаимосвязь между нарушением регуляции EGFR (рецептора эпидермального фактора роста) и целым рядом типов рака, и малые молекулы и направленные на EGFR антитела успешно дошли до стадии применения в клинических условиях. Однако антитела, одобренные для клинического применения, и антитела, находящиеся в разработке, обладают общим профилем тяжелых побочных эффектов вследствие широкой распространенности экспрессии EGFR в здоровых тканях. Образующийся путем делеции вариант III EGFR (EGFRvIII), с процессированным внеклеточным доменом, и, как следствие, лиганд-независимой конститутивной активностью, является наиболее распространенной мутантной формой, ассоциированной с онкогенной трансформацией. EGFRvIII экспрессируется исключительно в раковых тканях и ассоциирован с солидными опухолями разных типов. Ограниченная экспрессия EGFRvIII в раковых клетках дает возможность разрабатывать цитотоксические антитела, которые направлены исключительно на раковые ткани, и не затрагивают здоровые ткани, и существенно уменьшает побочные эффекты, ассоциированные с терапией EGFR.
Согласно первому аспекту настоящего изобретения, в настоящей заявке предложены полностью человеческие высокоспецифичные EGFRvIII-связывающие белки с высоким сродством. Описываемые EGFRvIII-связывающие белки обладают следующими преимуществами:
Они способствуют отсутствию или очень низкой степени интернализации EGFRvIII с поверхности клеток, позитивных по наличию мишени, что благоприятно для рекрутинга цитотоксических Т-лимфоцитов или естественных киллеров (NK). Результаты, представленные на Фигуре 9, показывают, что по сравнению с необработанными клетками молекула рецептора EGFRvIII все еще остается на поверхности клетки после воздействия на клетки EGFRvIII-связывающих белков согласно настоящему изобретению, при всех оцениваемых условиях. Указанные результаты свидетельствуют о том, что EGFRvIII/CD3-связывающие белки согласно настоящему изобретению неожиданно не демонстрируют никакой тенденции к интернализации (Фигура 9). Вместо индукции интернализации связывание EGFRvIII-связывающих белков согласно настоящему изобретению может либо подавлять интернализацию, либо способствовать повышенной экспрессии EGFRvIII, приводящей к увеличению его плотности на поверхности клетки.
У указанных белков с высоким сродством их сродство может быть еще больше увеличено с применением технологий «созревания аффинности» с получением связывающих белков, специфичных в отношении EGFRvIII с KD в диапазоне 100 пМ или ниже. Удивительным оказалось то, что указанные связывающие белки демонстрируют тонкую специфичность связывания с EGFRvIII без перекрестной реактивности с природным EGFR (EGFR дикого типа или wtEGFR). Новый эпитоп в EGFRvIII по сравнению с EGFRwt образуется в результате делеции внутри рамки считывания 269 аминокислот из внеклеточного домена EGFR (экзоны 2-7), и предполагается, что он довольно маленький; он заключается в новом смежном положении аминокислот (аминокислота 5 соединяется с аминокислотой 274) и появлении одной новой аминокислоты GLY в сайте делеции.
Процедура скрининга с «созреванием аффинности» была направлена на повышение сродства связывания путем облегчения селекции связывающих белков со сниженной скоростью диссоциации (kOFF). Однако неожиданно, связывающие белки, подвергшиеся «созреванию аффинности» проявили существенно повышенную скорость ассоциации (kON), тогда как уменьшение kOFF внесло лишь незначительный вклад в достигающее 100 раз повышение связывания связывающих белков, и некоторые вариабельные домены антител демонстрировали KD ниже 100 пМ.
Указанные уникальные свойства делают вариабельные домены антител, специфичные в отношение EGFRvIII, описываемые в настоящей заявке, подходящими для разработки мультиспецифичных, например, биспецифических, поливалентных, рекрутирующих иммунные эффекторные клетки, направленных на опухоли лекарственных препаратов на основе антител, например, таких как биспецифическое тандемное диатело к EGFRvIII/CD3 (TandAb®).
Согласно дополнительному аспекту настоящего изобретения предложены поливалентные EGFRvIII-связывающие антитела, содержащие два сайта связывания для EGFRvIII или, в случае поливалентных биспецифических антител, содержащее дополнительно к двум сайтам связывания EGFRvIII сайты связывания антигена Т-лимфоцитов CD3 иди CD 16А, экспрессируемых специфически на поверхности естественных киллеров (NK).
Согласно одному примеру применялись тандемные диатела к EGFRvIII/CD3, содержащие два сайта связывания для EGFRvIII и дополнительно содержащие два сайта связывания для CD3, и исследовалось их связывание с рекомбинантным гибридными антигенами EGFRvIII- или EGFRv-Fc. Гибридные антигены EGFRvIII- или EGFR-Fc разделяли посредством SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) либо при невосстановительных условиях (молекулярные структуры остаются частично сохраненными благодаря интактным дисульфидным связям), или при восстановительных условиях (структура белка полностью денатурирована), переносили на мембрану из поливинилиденфторида (ПВДФ) посредством вестерн-блоттинга и оценивали на предмет связывания с применением разных тандемных диател к EGFRvIII/CD3 или EGFR-связывающего IgG цетуксимаба для проявления блоттинга (Фигура 4). Данный пример показывает, что и исходные, и подвергшиеся «созреванию аффинности» EGFRvIII-связывающие домены сохраняют свою селективную специфичность в отношении EGFRvIII также в форме поливалентного антитела, содержащей два сайта связывания для EGFRvIII. Все тандемные диатела EGFRvIII/CD3 распознают EGFRvIII/CD3 как при восстановительных, так и при невостановительных условиях, но не способны распознавать полноразмерный EGFR дикого типа при любых из указанных условий. Кроме того, данный пример демонстрирует, что EGFRvIII-специфичные связывающие домены, описываемые в настоящей заявке, распознают линейный эпитоп, и им не требуются интактные трехмерные структуры для их активности. Это отличает их от EGFR-связывающего антитела IgG - цетуксимаба (эрбитукса), которое распознает и EGRF дикого типа, и мутированные EGFRvIII, но ему явно требуются интактные дисульфидные связи для его активности с конформационным эпитопом (Фигура 4).
Согласно другому примеру применялись тандемные диатела к EGFRvIII/CD3, содержащие исходные или подвергшиеся «созреванию аффинности» EGFRvIII-специфичные домены, комбинированные с разными CD3-связывающими доменами и/или обладающие разным порядком отдельных связывающих доменов в молекулах тандемных диател. Тандемные диатела с порядком доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3 содержат двухвалентные сайты связывания EGFRvIII в центре молекул тандемного диатела, тогда как тандемные диатела с порядком доменов VHEGFRvIII-VLCD3-VHCD3-VLEGFRvIII содержат два сайта связывания EGFRvIII в краевых положениях, а два сайта связывания CD3 в центре. Варианты тандемных диател, содержащие разные EGFRvIII-домены, и с разным порядком доменов анализировали, чтобы определить концентрационную зависимость связывания с антигеном EGFRvIII, антигеном EGFR дикого типа, а также связывания с антигеном CD3 посредством ELISA (твердофазный иммуноферментный анализ) (Фигура 6). Все тандемные антитела, содержащие EGFRvIII-связывающие домены, подвергшиеся «созреванию аффинности», демонстрировали значительное усиление связывания с EGFRvIII, отчетливо проявляющееся как сдвиг соответствующих кривых связывания в сторону более низкой концентрации более чем на два порядка (Фигура 6). Связывание с антигеном EGFR дикого типа не наблюдалось ни у одного из указанных тандемных антител.
Селекцию и идентификацию антител к EGFRvIII, например, антиген-связывающих белков, которые селективно связываются с мутированной, но не нативной формой EGFR, можно проводить при помощи библиотек фагового дисплея. Т-лимфоциты являются мощными уничтожающими опухоли эффекторными клетками, которые не могут быть рекрутированы нативными антителами. Следовательно, согласно дополнительному аспекту настоящего изобретения предложен набор мультиспецифичных EGFRvIII/CD3-антиген-связывающих белков, в частности тандемных диател, способных рекрутировать Т-лимфоциты, с широким спектром связывающих и цитотоксических свойств. Связывающие свойства, опосредуемую Т-лимфоцитами цитотоксическую активность и мишень-опосредованную активацию Т-лимфоцитов антигенсвязывающих белков согласно настоящему изобретению характеризуют в наборе исследований in vitro. Антигенсвязывающие белки согласно настоящему изобретению проявляют тонкую специфичность в отношении антигена EGFRvIII в исследованиях Biacor, ELISA и с EGFRvIII-позитивными клетками в исследованиях FACS (сортировка клеток с активированной флуоресценцией). Ни с одним из связывающих белков не отмечалось заметного связывания на антигене EGFR или на клетках, экспрессирующих EGFRwt во всем исследуемом диапазоне концентраций. Большинство мощных тандемных диател с высоким сродством проявляли цитотоксичность в отношении EGFRvIII-экспрессирующих клеток глиомы F98 и клеток СНО (яичника китайского хомячка) с ЕС50 в диапазоне 1 пМ - 10 пМ; остальные тандемные диатела проявляли цитотоксичность с ЕС50 до приблизительно 10000 пМ. Также оценивалась цитотоксичность указанных тандемных диател в отношении клеток EGFRwt+, как более чувствительного зонда на остаточное связывание с нативной формой. Никакой цитотоксичности не наблюдалось ни в отношении EGFRwt+-клеток, ни в отношении других EGFRvIII-негативных клеток вплоть до максимальной оцениваемой концентрации тандемных диател 0,5 мкМ. Высокоаффинное связывание с CD3 важно для эффективного рекрутинга Т-лимфоцитов и, кроме того, в отсутствии EGFRvIII+клеток-мишеней in vitro, тандемные диатела согласно настоящему изобретению не вызывают активации Т-лимфоцитов, которую измеряют по отсутствию их пролиферации, что способствует хорошему профилю безопасности в доклинических исследованиях (Фигура 10). В заключении следует отметить, что тандемное диатело EGFRvIII/CD3 обусловливает строгую специфичность и высокую активность противоопухолевой цитотоксичности.
Были сгенерированы тандемные диатела с KD связывания с CD3 в диапазоне от 1,1 нМ до приблизительно 500 нМ, но относительно постоянной KD связывания с EGFRvIII (в диапазоне от 2,0 нМ до 6,7 нМ), и была продемонстрирована хорошая корреляция между силой связывания с CD3 и цитотоксической активностью в отношении клеток, экспрессирующих EGFRvIII, со значениями EC50 от 25 пМ до приблизительно 10000 пМ (Фигура 11).
Хотя сродство к EGFRvIII антигенсвязывающих белков согласно настоящему изобретению, было значительно увеличено, потери специфичности не происходило.
TandAb® является торговой маркой компании «Affimed Therapeutics», применяемой для обозначения тандемного диатела. (Kipriyanov et al., 1999, J. Mol. Biol, 293:41-56; Cochlovius et al., 2000, Cancer Res., 60:4336-4341; Reusch et al., 2004, Int. J. Cancer, 112:509-518, Kipriyanov, 2009, Methods Mol Biol, 562:177-93; McAleese and Eser, 2012, Future Oncol. 8:687-95). В контексте настоящего изобретения TandAb и тандемные диатела применяются как синонимы.
Последовательности гена и белка EGFR дикого типа известны. EGFRvIII является результатом делеции внутри рамки считывания экзонов 2-7 (801 по) в гене EGFR дикого типа, которая приводит к делеции 267 аминокислот во внеклеточном домене рецептора, и генерированию нового остатка глицина в месте соединения экзонов 1 и 8. Такое новое соседство аминокислот во внеклеточном домене EGFR создает опухоль-специфичный иммуногенный эпитоп.
Согласно первому аспекту настоящего изобретения описан связывающий белок, обладающий специфичностью по меньшей мере в отношении EGFRvIII и не обладающий перекрестной реактивностью с EGFR дикого типа. Указанный связывающий белок предпочтительно содержит CDR1, CDR2, CDR3 вариабельной области тяжелой цепи антитела и CDR1, CDR2 и CDR3 вариабельной области легкой цепи антитела, выбранных из CDR, описанных в Таблице 1А ниже.
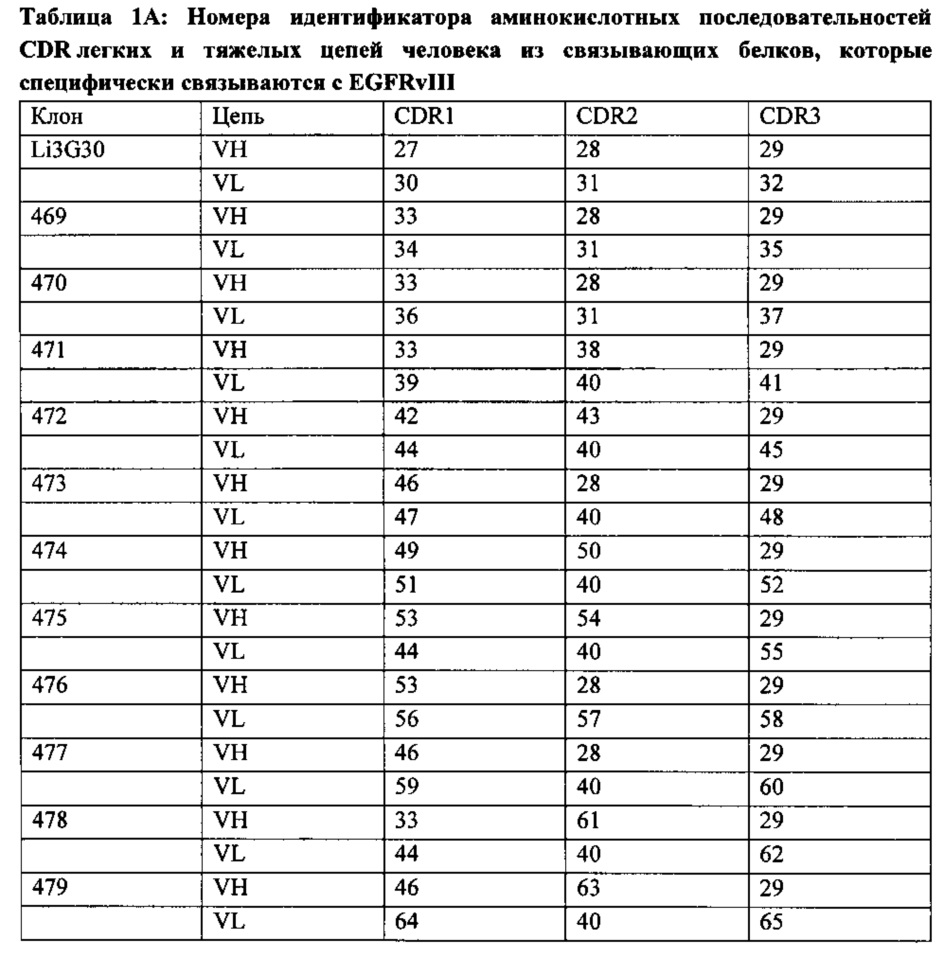
Следовательно, одним вариантом реализации настоящего изобретения является EGFRvIII-связывающий белок по меньшей мере с одним сайтом связывания, содержащим вариабельный домен тяжелой цепи антитела, содержащий CDR1, выбранный из группы, состоящей из SEQ ID №: 27, 33, 42, 46, 49, 53; CDR2, выбранный из группы, состоящей из SEQ ID №: 28, 38, 43, 50, 54, 61, 63; и CDR3 с последовательностью SEQ ID №: 29, и/или вариабельный домен легкой цепи антитела, содержащий CDR1, выбранный из группы, состоящей из SEQ ID №:30, 34, 36, 39, 44, 47, 51, 56, 59, 64; CDR2, выбранный из группы, состоящей из SEQ ID №: 31, 40, 57, и CDR3, выбранный из группы, состоящей из SEQ ID №: 32, 35, 37, 41, 45, 48, 52, 55, 58, 60, 62 и 65.
Согласно дополнительному варианту реализации указанный связывающий белок содержит по меньшей мере один сайт связывания EGFRvIII, содержащий вариабельный домен тяжелой цепи антитела, выбранный из группы, состоящей из SEQ ID №: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21 и 25, как показано в Таблице 1В, и/или вариабельный домен легкой цепи антитела, выбранный из группы, состоящей из SEQ ID №: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 и 26, как показано в Таблице 1В. Аминокислотные последовательности трех указанных вариабельных CDR легких цепей и трех указанных вариабельных CDR тяжелых цепей в Таблице 1 выделены жирным шрифтом и подчеркнуты. Указанные сайты связывания демонстрируют повышенное сродство к EGFRvIII без потери специфичности. Указанные антигенсвязывающие белки не обладают перекрестной реактивностью с антигеном EGFR дикого типа.
Также антигенсвязывающие белки согласно настоящему изобретению не демонстрируют никакой интернализации или лишь минимальную интернализацию EGFRvIII при его связывании. Согласно настоящему изобретению более чем 80%, предпочтительно более чем 90%, наиболее предпочтительно более чем 95% молекул рецептора EGFRvIII остается на поверхности клетки после воздействия на клетки EGFRvIII-связывающих антител, при условиях, исследуемых в Примере 9. Вместо индукции интернализации связывание EGFRvIII-специфичных связывающих белков согласно настоящему изобретению может либо подавлять интернализацию, либо способствовать усилению экспрессии EGFRvIII, что приводит к повышению его плотности на поверхности клетки.
Термин «связывающий белок» относится к производному иммуноглобулина с антигенсвязывающими свойствами, т.е. иммуноглобулины-полипептиды или их фрагменты, которые содержат антигенсвязывающий сайт. Связывающий белок содержит вариабельные домены антитела или их фрагменты. Каждый антигенсвязывающий домен образован вариабельным доменом тяжелой цепи (VH) антитела, т.е. иммуноглобулина, и вариабельным доменом легкой цепи (VL) антитела, связывающимися с одним и тем же эпитопом. Вариабельный домен легкой цепи или вариабельный домен тяжелой цепи согласно настоящему изобретению представляет собой полипептид, содержащий CDR1, CDR2 и CDR3. Предпочтительно, связывающий белок согласно настоящему изобретению лишен константных доменов иммуноглобулина. Также термин «связывающий белок» относится к фрагментам или производным антител, включая Fab, Fab', F(ab')2, фрагменты Fv, одноцепочечные Fv, диатело, тандемное диатело (TandAb®), флекситело (WO 03/025018), тандемный одноцепочечный Fv ((scFv)2).



Согласно предпочтительному варианту реализации связывающий белок, придающий специфичность к EGFRvIII, содержит антигенсвязывающий сайт из одного из следующих сочетаний вариабельного домена тяжелой цепи и вариабельного домена легкой цепи, показанных в Таблице 1:
(i) SEQ ID №:1 и SEQ ID №:2,
(ii) SEQ ID №:3 и SEQ ID №:4,
(iii) SEQ ID №:5 и SEQ ID №:6,
(iv) SEQ ID №:7 и SEQ ID №:8,
(v) SEQ ID №:9 и SEQ ID №:10,
(vi) SEQ ID №:11 и SEQ ID №:12,
(vii) SEQ ID №:13 и SEQ ID №:14,
(viii) SEQ ID №:15 и SEQ ID №:16,
(ix) SEQ ID №:17 и SEQ ID №:18,
(x) SEQ ID №:19 и SEQ ID №:20,
(xi) SEQ ID №:21 и SEQ ID №:22, или
(xii) SEQ ID №:25 и SEQ ID №:26
Также согласно настоящему изобретению предложены связывающие белки, которые не только обладают специфичностью в отношении EGFRvIII, но которые также содержат по меньшей мере один дополнительный функциональный домен. Согласно дополнительному варианту реализации указанным по меньшей мере одним функциональным доменом является эффекторный домен. «Эффекторный домен» представляет собой сайт связывания, специфичный в отношении эффекторной клетки, который может стимулировать или запускать цитотоксичность, фагоцитоз, представление антигена, выделение цитокинов. Такие эффекторные клетки включают, например, без ограничений, Т-лимфоциты или NK. В частности, дополнительный эффекторный домен содержит по меньшей мере один вариабельный домен тяжелой цепи антитела и по меньшей мере один вариабельный домен легкой цепи, образующие антигенсвязывающий сайт для CD3, предпочтительно, CD3 человека.
Таким образом, EGFRvIII-связывающий белок согласно настоящему изобретению может быть мультиспецифичным. В настоящей заявке термин «мультиспецифичный» означает, что связывающий белок согласно настоящему изобретению содержит по меньшей мере два антигенсвязывающих сайта для разных эпитопов, причем по меньшей мере один специфичен для EGFRvIII. Например, связывающий белок может быть триспецифическим и содержать сайты связывания с двумя разными эпитопами и/или антигенами на клетке опухоли и по меньшей мере один сайт связывания для эпитопа или антигена на эффекторной клетке, например, такой как CD3. Связывающий белок может содержать сайты связывания с разными эпитопами на одном и том же антигене и/или эпитопами на разных антигенах. Такие мультиспецифичные связывающие белки включают одноцепочечные диатела, (scFv)2, тандемные диатела (TandAb®) и флекситела (см. Le Gall et al. 1999 FEBS Letts 453:164-168 и WO 03/025018). Согласно конкретному варианту реализации настоящего изобретения связывающий белок является биспецифическим в отношении EGFRvIII и CD3.
Сайт связывания CD3 мультиспецифичного связывающего белка согласно настоящему изобретению может состоять из вариабельного домена тяжелой цепи (SEQ ID №: 23) и вариабельного домена легкой цепи (SEQ ID №: 24), которые показаны в Таблице 2, или близкого гомолога данной последовательности, отличающегося только на 2 аминокислоты в последовательностях CDR или вариабельных доменов любого другого полностью человеческого, гуманизированного или нечеловеческого антитела, специфичных в отношении CD3 (именуемых CD3), таких как, например, вариабельные домены антитела или вариабельные домены любого другого полностью человеческого, гуманизированного или нечеловеческого антитела, специфичные в отношении CD3, в частности CD3ε, такие как, например, вариабельные домены антитела UCHT1 или ОКТ3.
Таблица 2: Аминокислотные последовательности вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VK) полностью человеческого анти-CD3 антитела (аминокислотные последовательности CDR1, CDR2 и CDR3 показаны жирным шрифтом и подчеркнуты)

Кроме того, EGFRvIII-связывающий белок согласно настоящему изобретению может быть поливалентным. В настоящей заявке термин «поливалентный» означает, что связывающий белок согласно настоящему изобретению содержит по меньшей мере два антигенсвязывающих сайта. Указанные антигенсвязывающие сайты могут обладать одинаковой или разной специфичностью. Согласно одному варианту реализации связывающий белок связывается по меньшей мере при помощи двух сайтов связывания, т.е. двухвалентен, с одним и тем же эпитопом. Примером двухвалентного связывающего белка является диатело, а примером по меньшей мере трехвалентного связывающего белка является тандемное диатело.
Согласно дополнительному аспекту настоящего изобретения описываемые EGFRvIII-связывающие белки, а также биспецифические сайты связывания EGFRvIII и CD3 являются гуманизированными или полностью человеческими, т.е. происходят из антител человека. Согласно дополнительному аспекту настоящего изобретения EGFRvIII-связывающий белок или мультиспецифичный EGFRvIII- и CD3-связывающий белок является димером, т.е., содержит два полипептида с антигенсвязывающими сайтами для EGFRvIII и CD3.
Согласно настоящему изобретению предложен димерный и биспецифический EGFRvIII-и CD3-связывающий белок в форме тандемного диатела (TandAb®). Такие тандемные диатела конструируют путем соединения четырех вариабельных связывающих доменов антител (вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL) в одной генной конструкции) (см. McAleese and Eser, 2012, Future Oncol. 8:687-95), что делает возможной гомодимеризацию. В таких тандемных диателах длина линкера такова, что она препятствует межмолекулярному спариванию вариабельных доменов, так что молекула не может конъюгировать в себе, образуя одноцепочечное диатело, а вынуждено спариваться с комплементарными доменами другой цепи. Указанные домены также расположены так, чтобы соответствующие домены VH и VL спаривались во время такой димеризации. После экспрессии с конструкции из одиночного гена две идентичные полипептидные цепи сворачиваются в ориентации «голова-к-хвосту», образуя нековалентный гомодимер массой приблизительно 110 кДА (McAleese and Eser, 2012, Future Oncol. 8:687-95). Несмотря на отсутствие межмолекулярных ковалентных связей указанный гомодимер очень стабилен, когда он образовался, остается интактным и не переходит обратно в мономерную форму.
Согласно одному варианту реализации мультиспецифичного тандемного диатела предложено гуманизированное биспецифическое тетравалентное антитело (TandAb®) с двумя сайтами связывания и для CD3, и для EGFRvIII (EGFRvIII/CD3 RECRUIT-TandAb®), чтобы связать цитотоксическую способность Т-клеток с лечением глиобластомы (GB), рака предстательной железы, головы и шеи и других типов рака, позитивного по мутанту рецептора эпидермального фактора роста vIII (EGFRvIII+).
Тандемные диатела обладают целым рядом свойств, которые обеспечивают преимущества над традиционными моноклональными антителами, а также менее крупными мультиспецифичными молекулами. Тандемные диатела полностью функциональны в отсутствии гликозилирования. Тандемные диатела содержат только вариабельные домены антител и, следовательно, лишены побочных эффектов, которые могут быть ассоциированы с фрагментом Fc. Поскольку биспецифические тандемные диатела допускают двухвалентное связывание и с EGFRvIII, и с CD3, авидность у них такая же, как и у IgG. Размер тандемного диатела, приблизительно 110 кДа, меньше чем у IgG, что может обеспечивать лучшее проникновение в опухоль. Однако данный размер значительно превышает порог выведения для пресистемного клиренса, что обеспечивает фармакокинетическое преимущество по сравнению с менее крупными биспецифическими формами на основе антитело-связывающих доменов или структур, основанных не на антителах. Генерирование и производство таких тандемных диател описано, например, у Kipriyanov SM: Methods Mol. Biol. 2009;562:177-93 Kipriyanov SM: Methods Mol Biol 2003;207:323-33 или McAleese and Eser, 2012, Future Oncol. 8:687-95.
Тандемные диатела хорошо экспрессируются в клетках СНО. Может быть введен надежный восходящий и нисходящий технологический процесс.
EGFRvIII- и CD3-биспецифическое тандемное диатело согласно настоящему изобретению разработаны для получения специфичного направленного действия на EGFRvIII+-клетками опухоли посредством рекрутинга цитотоксических Т-лимфоцитов. Антитела не способны напрямую рекрутировать цитотоксические Т-лимфоциты. В отличие от них тандемные диатела посредством привлечения молекул CD3 могут обеспечивать перекрестное взаимодействие цитотоксических Т-клеток с EGFRvIII+-клетками опухоли с высокой специфичностью, тем самым значительно повышая цитотоксический потенциал таких молекул. Тандемные диатела проявляют сильную, специфическую и эффективную цитотоксичность. Существуют весомые факты в пользу того, что Т-лимфоциты могут играть определенную роль в контролировании роста опухолей. Например, было показано, что присутствие цитотксических Т-клеток в опухолях толстой и ободочной кишки, а также в лимфатических узлах от пациентов с неходжкинской лимфомой коррелируют с более хорошим клиническим исходом. Кроме того, потенциал лекарственных средств, предназначенных для индукции Т-клеточного ответа, был продемонстрирован для вакцин от меланомы, а также для антитела, направленного на CTLA-4 - отрицательный регулятор активации Т-лимфоцитов. Тандемное диатело согласно настоящему изобретению привлекает цитотоксические Т-лимфоциты посредством связывания с поверхностно-экспрессируемым CD3, предпочтительно CD3ε, который образует часть рецептора Т-лимфоцитов. Одновременное связывание данного тандемного диатела с CD3 и с EGFRvIII, экспрессируемого на поверхности клеток конкретных опухолей вызывает активацию Т-клеток и опосредует последующий лизис клетки опухоли.
Термин «димер» относится к комплексу из двух пептидов. Предпочтительно, указанные два полипептида нековалентно связаны друг с другом, в частности, при условии, что ковалентной связи между двумя указанными полипептидами нет. Предпочтительно указанный биспецифический димер является гомодимером, т.е. содержит два идентичных полипептида. Однако триспецифические или мультиспецифичные димеры могут быть гетеродимерными и содержать два разных полипептида, например, в которых по меньшей мере один сайт связывания вариабельного домена легкой цепи расположен в одном полипептиде, а вариабельный домен расположен в другом домене. Термин «полипептид» относится к полимеру из аминокислотных остатков, связанных амидными связями. Указанный полипептид предпочтительно является одноцепочечным гибридным белком, который не разветвлен. В указанном полипептиде вариабельные домены антитела связаны друг с другом. Указанный полипептид может содержать смежные аминокислотные остатки наряду с N-концевыми и/или С-концевыми остатками вариабельного домена. Например, такие смежные аминокислотные остатки могут включать Tag-последовательность, предпочтительно на С-конце, который может быть полезен для очистки полипептида.
Каждый полипептид указанного биспецифического тандемного диатела содержит по меньшей мере четыре вариабельных домена, вариабельный домен легкой цепи (VL) и вариабельный домен тяжелой цепи (VH) CD3-связывающего белка, а также вариабельный домен легкой цепи (VL) и вариабельный домен тяжелой цепи (VH) EGFRvIII-связывающего белка. Указанные четыре вариабельных домена связаны пептидными линкерами L1, L2 и L3, и могут располагаться в направлении N-конец - С-конец следующим образом:
(i) VL (CD3)-L1-VH (EGFRvIII)-L2-VL(EGFRvIII)-L3-VH(CD3); или
(ii) VH (CD3)-L1-VL(EGFRvIII)-L2-VH(EGFRvIII)-L3-VL(CD3); или
(iii) VL(EGFRvIII)-L1-VH(CD3)-L2-VL(CD3)-L3-VH(EGFRvIII); или
(iv) VH(EGFRvIII)-L1-VL(CD3)-L2-VH(CD3)-L3-VL(EGFRvIII).
Согласно одному варианту реализации настоящего изобретения указанные четыре вариабельных домена расположены в порядке VL (CD3)-L1-VH (EGFRvIII)-L2-VL(EGFRvIII)-L3-VH(CD3). Тандемные диатела с порядком доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3 и содержащие разные EGFRvIII-связывающие последовательности, хотя и обладают разным сродством к EGFRvIII, все демонстрируют весьма сходное связывание со второй специфической мишенью - CD3, что удается показать посредством ELISA на антигене CD3γε (Фигура 5А). В другом тандемном диателе с порядком расположения доменов (VHEGFRvIII-VLCD3-VHCD3-VLEGFRvIII), содержащем EGFRvIII-связывающие домены в наружном положении и два сайта связывания CD3 в центре, аналогично проявляется существенное улучшение связывания с EGFRvIII при подвергшихся «созреванию аффинности» EGFRvIII-связывающих доменах, тогда как связывание с CD3-доменом в центре тандамных диател несколько слабее и варьирует в большей степени между отдельными тандамными диателами, чем при первом порядке расположения доменов (Фигура 5В). Аналогичные наблюдения были сделаны, когда для конструирования диател, содержащих разные EGFRvIII-связывающие домены, применялся другой CD3-домен (SEQ ID №:23 и 24) (Фигура 5С).
Длина линкеров влияет на гибкость антигенсвязывающего тандемного диатела. Влияние длины линкера на образование димерных антигенсвязывающих белков описано, например, Todorovska et al., 2001 Journal of Immunological Methods 248:47-66; Perisic et al., 1994 Structure 2:1217-1226; Le Gall et al., 2004, Protein Engineering 17:357-366 и WO 94/13804.
Согласно настоящему изобретению предпочтительно, чтобы длина пептидных линкеров L1, L2 и L3 была такой, чтобы домены одного полипептида могли образовать межмолекулярные связи с доменами другого полипептида с образованием димерного антигенсвязывающего тандемного диатела. Такие линкеры являются «короткими», т.е. состоят из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 аминокислотных остатков. Таким образом, указанные линкеры состоят из приблизительно 12 или менее аминокислотных остатков. В случае 0 аминокислотных остатков ликер представляет собой пептидную связь. Такие короткие линкеры благоприятствуют межмолекулярной димеризации двух полипептидов за счет связывания и образования корректных антигенсвязывающий сайтов между вариабельными доменами легкой цепи антитела и вариабельными доменами тяжелых цепей антитела разных полипептидов. Короткий линкер из приблизительно 12 аминокислотных остатков или менее, как правило, препятствует тому, чтобы соседние домены одной и той же полипептидной цепи образовывали межмолекулярное взаимодействие друг с другом. Согласно одному варианту реализации настоящего изобретения указанные линкеры содержат приблизительно от 3 до приблизительно 10, например, 4, 5 или 6 соседних аминокислотных остатков.
Что касается аминокислотного состава линкеров, выбирают такие пептиды, которые не препятствуют димеризации двух указанных полипептидов. Например, линкеры, содержащие остатки глицина и серина, как правило, обеспечивают устойчивость к протеазам. Аминокислотную последовательность линкеров можно оптимизировать, например, при помощи способов фагового дисплея, чтобы улучшить связывание антигена и выработку димерного антиген-связывающего полипептида. Примеры пептидных линкеров, подходящих для тандемного диатела согласно настоящему изобретению, включают GGSGGS (SEQ ID №:66), GGSG (SEQ ID №:67) или GGSGG (SEQ ID №:68).
EGFRvIII-связывающий белок и мультиспецифичное тандемное диатело, описываемые в настоящей заявке, можно получить путем экспрессии полинуклеотидов, кодирующих полипептид тандемного диатела, который ассоциирован с другим идентичным полипептидом и образует вместе с ним антигенсвязывающее тандемное диатело. Следовательно, дополнительным вариантом реализации настоящего изобретения является полинуклеотид, например, ДНК или РНК, кодирующий полипептид антигенсвязывающего тандемного антитела, описываемого в настоящей заявке.
Указанный полинуклеотид можно сконструировать при помощи способов, известных специалистам в данной области техники, например, посредством сочетания генов, кодирующих по меньшей мере четыре вариабельных домена антитела, либо разделенные последовательностями, кодирующими пептидные линкеры, или напрямую соединенные пептидной связью, в единой генетической конструкции, функционально связанной с соответствующим промотором, и необязательно с белком-меткой для детекции и очистки, а также соответствующим терминатором транскрипции, и ее экспрессии в бактерии или другой подходящей системе экспрессии, такой как, например, клетки СНО. В зависимости от применяемой системы векторов и хозяина можно применять любое количество соответствующих элементов транскрипции и трансляции, включая конститутивные и индуцибельные промоторы. Промотор выбирают таким образом, чтобы он запускал экспрессию полинуклеотида в соответствующей клетке-хозяине.
Полинуклеотид можно вводить в вектор, предпочтительно вектор экспрессии, который представляет собой дополнительный вариант реализации настоящего изобретения. Данный рекомбинантный вектор можно сконструировать в соответствии со способами, хорошо известными специалистам в данной области техники.
Можно применять самые разнообразные системы вектор/хозяин, которые бы содержали и экспрессировали полинуклеотид, кодирующий полипептид согласно настоящему изобретению. Примеры векторов экспрессии, действующих в клетках E.coli включают pSKK (Le Gall et al., J Immunol Methods. (2004) 285(1):111-27) или pcDNA5 («Invitrogen») для экспрессии в клетках млекопитающих.
Таким образом, антигенсвязывающее тандемное диатело, описываемое в настоящей заявке, можно получить путем введения вектора, кодирующего полипептид, описанный выше, в клетку-хозяина, и культивирования указанной клетки при условиях, при которых полипептидные цепи экспрессируются, их можно выделить из питательной среды и, необязательно, подвергнуть дальнейшей очистке.
Согласно дополнительному варианту реализации настоящего изобретения предложена фармацевтическая композиция, содержащая EGFRvIII-связывающий белок, антигенсвязывающее тандемное диатело, вектор, содержащий полинуклеотид, кодирующий полипептид указанного антиген-связывающего тандемного диатела, или клетку-хозяина, трансформированную указанным вектором, и по меньшей мере один фармацевтически приемлемый носитель. Подразумевается, что термин «терапевтически приемлемый носитель» включает любой носитель, который не снижает эффективности биологической активности ингредиентов и который не токсичен для пациента, которому его вводят. Примеры подходящих фармацевтических носителей хорошо известны в технике и включают физиологические растворы, фосфатно-солевые буферы, воду, эмульсии, такие как эмульсии масло/вода, разные типы смачивающих агентов, стерильные растворы и др. Такие носители могут быть составлены при помощи традиционных способов, и их можно вводить субъекту в соответствующей дозе. Предпочтительно, указанные композиции стерильны. Указанные композиции также могут содержать вспомогательные вещества, такие как консерванты, эмульгаторы и диспергирующие агенты. Предупреждение действия микроорганизмов можно обеспечить путем включения разных антибактериальных и противогрибковых агентов.
Специалист в данной области техники без особых затруднений может сконструировать и получить антигенсвязывающие белки, такие как тандемные диатела, описываемые в настоящей заявке, применяя общепринятые технологии и стандартные способы, известные в технике, см., например, Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y.; The Protein Protocols Handbook, edited by John M. Walker, Humana Press Inc. (2002); или Antibody engineering: methods and protocols / edited by Benny K.C. Lo; Benny K.C. II Series: Methods in molecular biology (Totowa, N.J.)).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 иллюстрирует положения вариабельных последовательностей в EGFRvIII-специфичных связывающих доменов с высоким сродством, полученных после "созревания аффинности». Положения CDR вариабельных областей тяжелых и легких цепей, которые отличаются от исходной последовательности, показаны звездочками, а соответствующие аминокислоты - курсивом и подчеркиванием. Положения полезных мутаций выделены жирным шрифтом.
На Фигуре 2 показаны результаты мультициклических измерений кинетики связывания разных антител scFv с рекомбинантным антигеном EGFRvIII-Fc или с антигеном EGFR-Fc дикого типа в анализе Biacore Т200. Сенсограмма показывает кинетику единиц ответа (ЕО) поверхностного плазмонного резнонанса, измеренную во времени при 4 разных концентрациях антитела cFv (86 нМ, 17,2 нМ, 3,4 нМ, 0,69 нМ). Ассоциацию измеряют в течение 180 секунд, диссоциацию измеряют после. Данные анализируют на основании модели связывания 1:1.
На Фигуре 3 показаны результаты проточно-цитометрического измерения связывания антител scFv с (А) клетками глиомы крысы F98 с гиперэкспрессией (F98EGFRvIII), (В) клетками F98 с гиперэкспрессией нативного EGFR (F98EGFR), (С) нетрансфецированными клетками F98 (А98), а также связывания тех же антител scFv с (D) клетками СНО с гиперэкспрессией EGFRvIII (CHOEGFRvIII), (Е) клетками СНО с гиперэкспрессией нативного EGFR (CHOEGFR) и (F) нетрансфецированными клетками СНО (СНО).
Фигура 4 иллюстрирует связывание тандемных диател, содержащих разные EGFRvIII-связывающие домены, с гибридными антигенами, включающими очищенный EGFRvIII или нативный EGFR и Fc (невосстановленный белок или белок, восстановленный посредством DTT), которые разделяли посредством SDS-PAGE и переносили на мембрану из PVDF посредством вестерн-блоттинга. Fc-гибридные белки димеризуются за счет дисульфидных связей в Fc-части, но являются мономерными после восстановления под действием DTT. EGFRvIII-специфичные антитела связываются с EGFRvIII-гибридным антигеном в восстановленной и невосстановленной конформации, но не связываются с нативным EGFR. Анти-EGFR антитело цетуксимаб применялось в качестве контроля и распознавало как невосстановленный EGFRvIII-Fc, так и невосстановленный EGFR-Fc антиген, но не обработанные DTT антигены.
На Фигуре 5 показано зависимое от концентрации связывание разных EGFRvIII/CD3-специфичных тандемных диател с рекомбинантными антигенами EGFRvIII-Fc, EGFR-Fc и CD3γε, анализируемое посредством ELISA. Сигналы связывания показаны на (А) для EGFRvIII/CD3-специфических тандемных диател с порядком расположения доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3, на (В) для EGFRvIII/CD3-специфических тандемных диател с порядком расположения доменов VHEGFRvIII-VLCD3-VHCD3-VLEGFRvIII, и на (С) для EGFRvIII/CD3-специфичных тандемных диател, содержащих домен CD3 с последовательностью SEQ ID №: 23, 24 и обладающих порядком расположения доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3. Улучшение связывания с EGFRvIII тандемных диател, содержащих EGFRvIII-связывающие домены, подвергшиеся «созреванию аффинности», четко выявляется по сдвигу соответствующих кривых связывания в сторону более низких концентраций при всех формах/порядке доменов.
На Фигуре 6 показаны результаты мультициклических измерений кинетики связывания EGFRvIII/CD3-специфичных тандемных диател, содержащих разные двухвалентные EGFRvIII-связывающие домены при порядке расположения доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3 с рекомбинантным антигеном EGFRvIII-Fc в анализе Biacore X100. Сенсограммы показывают кинетику единиц ответа (ЕО) поверхностного плазмонного резнонанса, измеренную во времени при разных концентрациях тандемных диател. Ассоциацию измеряют в течение 180 секунд, диссоциацию измеряют после. Данные анализируют на основании модели связывания 1:1.
На Фигуре 7 показаны результаты иммуногистохимического окрашивания (ИГХ) срезов тканей от (А) трех отдельных пациентов с раком головы и шеи (Г и Ш), (В) трех отдельных пациентов с глиобластомой, и (С) репрезентативные результаты для рака предстательной железы, рака молочной железы (Her2 neg) и немелкоклеточного рака легких (НМРЛ). Срезы были окрашены двумя разными концентрациями EGFRvIII-специфического диатела, содержащего EGFRvIII-специфичный связывающий домен Li3G30 в двухвалентной конформации, аналогичной расположению в EGFRvIII-направленных тандемных диател (например, (VLCD3-)VHEGFRvIII-VLEGFRvIII(-VHCD3), но с отсутствующими CD3-связывающими частями. В качестве контроля срезы окрашивали EGFR-специфичным IgG-антителом цетуксимабом, который распознает EGFR (присутствует в здоровых, а также раковых тканях), а также EGFRvIII. Специфичность окрашивания подтверждали посредством контрольных срезов, инкубируемых со вторичными реагентами в отсутствии EGFR- или EGFRvIII-специфичных первичных антител.
Фигура 8 иллюстрирует улучшение цитотоксической активности, эффективности и мишень-зависимой специфичности EGFRvIII/CD3-тандемных диател, содержащих разные подвергшиеся «созреванию аффинности» EGFRvIII-специфичные связывающие домены, в сравнении с соответствующими тандемными диателами, содержащими исходный EGFRvIII/CD3-связывающий домен (Li3G30) перед «созреванием аффинности» на EGFRvIII-позитивных клетках: (А) ABC 470, содержащие EGFRvIII/CD3-тандемные диатела, (В) ABC 471, содержащие EGFRvIII/CD3-тандемные диатела, (C) ABC 472, содержащие EGFRvIII/CD3-тандемные диатела. Зависимую от концентрации антител цитотоксичность измеряли посредством проточной цитометрии на основании анализа, проводимого с применением EGFRvIII-экспрессирующих клеток СНО (CHOEGFRvIII), EGFRvIII-негативных EGFR-экспрессирующих клеток СНО (CHOEGFR) или нетрансфецированных клеток СНО в качестве клеток-мишеней и мононуклеарных клеток периферической крови (МКПК) в качестве эффекторных клеток. На каждой панели (А-С) тандемные диатела, содержащие отдельные EGFRvIII-связывающие домены и CD3-домены в порядке VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3 показаны слева, а тандемные диатела, содержащие отдельные EGFRvIII-домены в сочетании с разным CD3-доменом в порядке VHEGFRvIII-VLCD3-VHCD3-VLEGFRvIII - справа. На Фигуре 9 показаны результаты оценки интернализации рецепторов EGFRvIII с поверхности клетки при связывании с EGFRvIII-специфичными антителами. EGFRvIII-экспрессирующие клетки инкубировали с разными концентрациями трех разных тандемных диател к EGFRvIII/CD3 с порядком расположения доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3 (i) или VHEGFRvIII-VLCD3-VHCD3-VLEGFRvIII (iv), и/или содержащих разные CD3-связывающие домены при 37°С в течение 24 часов. После модуляции соответствующие тандемные диатела применяли при насыщающей концентрации 10 мкг/мл для окрашивания всех оставшихся молекул рецептора EGFRvIII на поверхности клетки.
На Фигуре 10 показаны результаты оценки пролиферации (включения BrdU (5-бромдезоксиуридина)) в культурах МКПК человека в присутствии EGFRvIII/CD3 (или EGFRvIII/CD16A) тандемных диател. МКПК пролиферируют в присутствии CD3-связывающего антитела ОКТ3 или фитогемагглютинина (ФГА) растительного лектина, которые применяются в качестве положительного контроля. Инкубация МКПК с разными тандемными диателами EGFRvIII/CD3 в отсутствии EGFRvIII-позитивных клеток-мишеней не вызывает активацию и пролиферацию МКПК.
Фигура 11 иллюстрирует положительную корреляцию сродства связывания с CD3 тандемных диател EGFRvIII/CD3 с их цитотоксической активностью. Более 20 тандемных диател EGFRvIII/CD3, содержащих идентичный EGFRvIII-связывающий домен (Li3G30), сочетаемый в порядке расположения доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3 с разными CD3-связывающими доменами, обладающих разным сродством к CD3, были проанализированы в отношении связывания CD3 в клетках CD3+-Jurkat, в отношении связывания EGFRvIII в клетках CHOEGFRvIII, а также в отношении цитотоксической активности. Значения KD для связывания CD3 варьировали от 1 нМ до приблизительно 500 нМ. Значения KD для связывания EGFRvIII и значения EC50 для цитотоксичности откладывали на графике относительно значений KD для связывания CD3 для каждого из анализируемых тандемных диател. Хотя тандемные диатела демонстрируют только небольшую вариабельность в значениях KD для связывания CD3, EC50 цитотксичности демонстрируют почти линейное увеличение при увеличении KD для связывания CD3 указанных тандемных диател.
На Фигуре 12 показаны результаты исследования обоснованности концепции in vivo. Дозозависимое подавляющее действие на рост подкожных ксенотрансплантатных опухолей F98EGFRvIII анализировали для двух тандемных диател EGFRvIII/CD3, содержащих один и тот же EGFRvIII-связывающий домен (Li3G30), но разные CD3-связывающие домены или порядок расположения доменов и сравнивали с действием цетуксимаба, который вводили при значительно более высоких концентрациях. Дозозависимое подавление роста опухоли EGFRvIII/CD3v6-тандемными диателами с порядком расположения доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3; (В) дозозависимое подавление роста опухоли EGFRvIII/CD3-тандемными диателами, содержащими CD3-разные связывающие домены с высоким сродством и с порядком расположения доменов VHEGFRvIII-VLCD3-VHCD3-VLEGFRvIII.
Изобретение дополнительно проиллюстрировано, но без ограничения, следующими примерами:
ПРИМЕР 1: Открытие и «созревание аффинности" EGFRvIII-специфичных связывающий белков: открытие Li3G30
Библиотеку последовательностей scFv человека, полученную при помощи фагового дисплея, подвергали двум-трем циклам пэннинга для обогащения связывающих белков, которые специфичны к EGFRvIII. Указанную библиотеку предварительно инкубировали на EGFR-Fc перед каждым циклом пэннинга, чтобы истощить запас белков, связывающихся с формой EGFR дикого типа или с частью Fc указанного гибридного белка. Селекцию проводили параллельно на твердой фазе, покрытой EGFRvIII, и в растворе. После двух и трех циклов пэннинга, отбирали единичные колонии, индуцировали экспрессию растворимого scFc и проводили скрининг экстрактов на связывание с EGFRvIII-Fc и EGFR-Fc.
Согласно настоящему изобретению описаны домены полностью человеческого высоко специфичного антитела, связывающиеся с EGFRvIII, - мутант, полученный в результате делеции в EGFR, который в мутантной форме экспрессируется исключительно в опухолях, но не в здоровых тканях. EGFRvIII-специфичные домены полностью человеческого антитела, описываемые в настоящей заявке, сначала изыскивались в ходе скрининга посредством фагового диспеля с применением библиотеки полностью человеческих scFv, происходящих из IgM. Неожиданно, несмотря на применение процедуры истощения на антигене EGFR дикого типа для двух или трех циклов пэннинга, которые должны были обогатить смесь связывающими белками, которые специфичны к EGFRvIII, большинство полученного scFv связывалось с обоими белками (EGFRvIII и EGFR дикого типа) при ELISA. Была идентифицирована одна последовательность с высокой специфичностью к EGFRvIII под названием Li3G30. За исключением нескольких несоответствий на N-конце VH и VL, в каркасных областях, а также в CD1 и 2, тяжелая и легкая цепь полностью идентичны зародышевым кодируемым последовательностям VH5-51 и VL3-25, соответственно. Li3G30 была идентифицирована из третьего жидкофазного цикла пэннинга.
«Созревание аффинности»
Библиотеку для пары каркасных участков антител VH5-51/VL3-25 разрабатывали по алгоритму, заказанному у компании «DistributedBio». Были идентифицированы остатки Li3G30, определяющие специфичность, и они были включены в дизайн библиотеки. Указанный дизайн включал 52 рандомизированных положения CDR, каждое с индивидуально определенным распределением аминокислот. В CDR3 VL были встроены петли трех разных длин. Фрагменты генов, кодирующие рандомизированные положения VH и VL, были заказаны у компаний «Geneart»/«Lifetechnologies» и синтезированы компанией «TRIM-technology». Фрагменты клонировали в вектор фагового дисплея рЕХНАМ (Schwarz et al. 2004), и размер итоговой библиотеки достигал 3,7Е+8 трансформированных клеток E.coli. Указанную библиотеку упаковывали в частицы фагов и подвергали процедуре пэннинга и скрининга с целью выделения вариантов с повышенным сродством и сохраненной специфичностью к EGFRvIII. Пэннинг проводили с антигеном EGFRvIII-Fc, который был иммобилизован на пластиковой поверхности, связывающей белки. Процедура пэннинга была разработана так, чтобы она благоприятствовала связыванию EGFRvIII с scFvs с низкими скоростями диссоциации (kOFF) за счет проведения следующих этапов: процедура отмывки включала несколько этапов инкубации в буфере отмывки длительностью до 30 минут, чтобы создать благоприятные условия для селекции scFvs с медленной диссоциацией. Другая процедура была основана на конкурентном инкубировании в течение ночи с EGFRvIII. Чтобы убедиться, что в процессе «созревания аффинности» были выбраны антитела с перекрестной реактивностью к EGFR-EGFRvIII или антитела, связывающиеся с FC, перед первым этапом пэннинга библиотеку истощали по возможным белкам, связывающимся с EGFR дикого типа посредством предварительной инкубации на EGFR-Fc дикого типа. Кроме того, протокол скрининга создавал неблагоприятные условия для связывающих веществ с перекрестной реактивностью из-за наличия растворимого EGFR в ходе процедуры пэннинга на иммобилизованном EGFRvIII.
Также уникальные EGFRvIII-специфичные scFv дополнительно проверяли на связывание с EGFRvIII-экспрессирующими трансфецированными клетками СНО и F98 путем проточной цитометрии. ScFv, которые демонстрировали связывание с экспрессирующими EGFR дикого типа и/или нетрансфецированными клетками, исключали из анализа. Для идентификации scFv с повышенным сродством к EGFRvIII по сравнению с исходным клоном Li3G30 применяли ранжирование начальной KD в анализе Biacore X100. Наилучшие варианты были охарактеризованы путем подробных измерений SPR и основанных на флуоресценции оценок стабильности.
Начальная библиотека для «созревания аффинности» включала петли трех разных длин для CDR3 VL, про все из которых было известно, что они совместимы с каркасами VH5-51/VL3-25. Неожиданно петля средней длины оказалось явно более благоприятной во время селекции. Как следствие CDR3 VL укорочен на одну аминокислоту по сравнению с исходным анти-EGFRvIII-антителом для всех вариантов, подвергнутых «созреванию аффинности» (Таблица 1, Фигура 1). Последовательности отобранных scFv сравнивали с входной библиотекой, и анализ указывает на три полезных в CDR3 VL. CD1 и 2 тяжелой и легкой цепи исходного анти-EGFRvIII-антитела идентичны последовательности, кодируемой соответствующими зародышевыми последовательностями VH5-51 и VL3-25. Соответствующий кодируемый зародышевой последовательностью остаток был восстановлен или явно был благоприятным в 75% положений. Одна благоприятная мутация предполагается в CDR1 VL (Фигура 1). Распределение аминокислот в остальных рандомизированных положениях было очень близко к распределению в библиотеке перед селекцией.
ПРИМЕР 2: Измерение связывания scFv с EGFRvIII и EGFR дикого типа в анализе Biacore:
Получение чипа СМ5 на основе IgG к Fc человека
Чип СМ5 на основе IgG к Fc человека получали посредством ковалентного присоединения аминов при помощи набора Amine-Coupling-Kit («GE») в соответствии с инструкциями производителя. IgG разводили в прилагаемом буфере для иммобилизации (10 мМ ацетат натрия, рН 5,0) до концентрации 25 мкг/мл. Для процедуры соединения применяли предустановленный способ «амин» в управляющей программе Biacore Т200. Для всех 4 проточных ячейках достигали целевого уровня от 5000 до 10000 УЕ. Перед введением растворов рекомбинантного белка поверхность чипа активировали дихлорэтаном/N-гидроксисукцинимидом. Свободные сайты связывания на поверхности блокировали этаноламином. 1×HBS-P+ был выбран в качестве подвижного буфера для всей процедуры соединения.
Диапазон концентраций одноцепочечных Fv (scFv), применяемых при измерении поверхностного плазмонного резонанса:
Концентрации ScFv проверяли перед измерением поверхностного плазмонного резонанса путем определения А280 с применением Nandrop. Концентрацию в матричном растворе (с) рассчитывали по формуле:
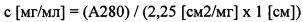
Разные коэффициенты экстинкции рассчитывали при помощи инструмента для прогнозирования «Expasy Protparam» (http://web.expasy.org/protparam/). Для расчета концентрации в молях антител scFv с меткой His, полагали молекулярную массу равной приблизительно 28 кДа. scFv корректировали на конечные концентрации, упоминаемые ниже, путем последовательного разведения в 1х HBS-P+буфере. Готовили следующие концентрации: 430 нМ, 86 нМ, 17,2 нМ, 3,44 нМ, 0,688 нМ и 0 нМ.
Условия измерения для scFvs:
Гибридные белки EGFRvIII-Fc и EGFR дикого типа-Fc разводили в 1×HBS-P+ до концентрации 6,25 нМ. Оба антигена вводили при скорости потока 10 мкл/мин в течение 75 сек для EGFRvIII-Fc и 90 сек для EGFR дикого типа-Fc. EGFRvIII-Fc вводили в проточную ячейку 2. A EGFR дикого типа-Fc вводили в проточную ячейку 4. Проточные ячейки 1 и 3 служили пустыми пробами.
1× HBS-P+ применяли в качестве подвижного буфера на протяжении всей процедуры. Антитела scFv вводили при скорости потока 30 мкл/мин в течение 180 сек во все четыре проточные ячейки.
Сигналы, измеренные в проточной ячейке 1 без захваченного антигена, вычитали из сигналов проточной ячейке 2, чтобы скорректировать кривые связывания на фоновое связывание. После диссоциации в течение 540 сек чип регенерировали, как описано ниже.
Чип на основе IgG к Fc человека регенерировали путем введения буфера с высоким содержанием соли 3,0 М MgCl2 (30 сек, 10 мкл/мин). Перед каждым измерением проводили четыре «пусковых периода» без scFv. Измерения в растворах scFv проводили от наименьшей до наибольшей концентрации. Включали один цикл с чистым подвижным буфером (0 мкМ).
Кинетику связывания антител scFv определяли на основании мультициклических измерений кинетики (МЦК). Для оценки кажущегося сродства связывания оценивали кривые связывания при помощи способа «Кинетика», включенного в программу для оценки Biacore. Посредством данного способа кривые связывания в целом аппроксимируют при помощи «модели взаимодействия Ленгмюра 1:1». Результаты приведены в Таблице 3.
Примененная процедура скрининга на основе «созревания аффинности» была направлена на повышение сродства связывания с EGFRvIII для EGFRvIII-специфичных связывающих доменов путем облегчения селекции связывающих белков со сниженной скоростью диссоциации (kOFF). Однако неожиданно полученные при «созревании аффинности» связывающие белки (Таблица 3) проявили существенное увеличение скорости ассоциации (kON), тогда как уменьшение kOFF вносило лишь незначительный вклад в достигающее 100 раз усиление связывания scFv, причем некоторые связывающие белки проявляли KD менее 100 пМ (Таблица 3). Особой неожиданностью явилось то, что уменьшение KD достигалось преимущественно за счет увеличение kON, хотя процедура скрининга благоприятствовала селекции scFv со сниженной kOFF. Также это было неожиданным, поскольку «созревание аффинности» антител обычно связано со снижением kOFF (Schier et al., 1996, Pini et al., 1998, Boder et al., 2000).
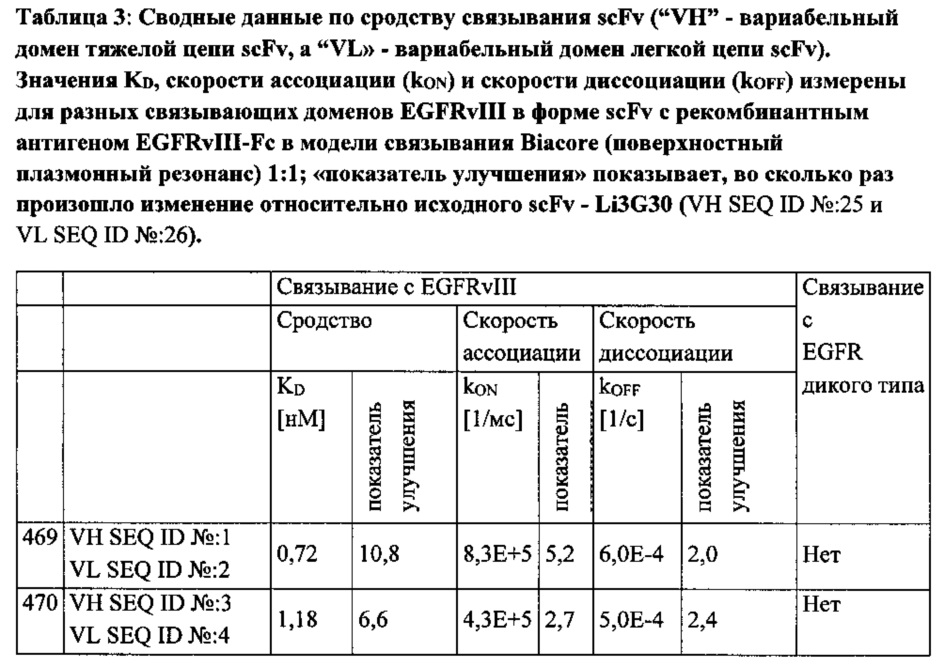
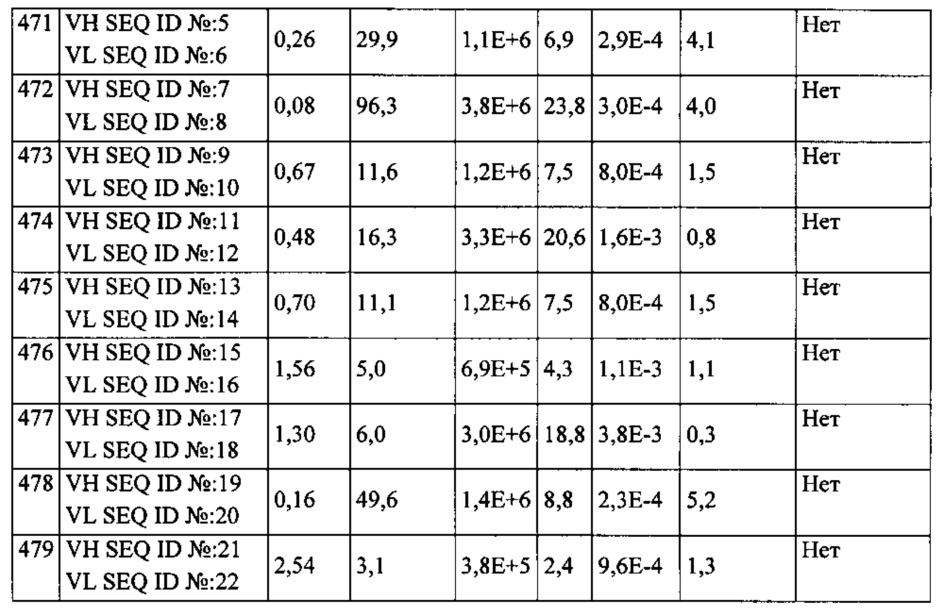
Восемь из 11 scFv, подвергшихся «созреванию аффинности», демонстрировали по меньшей мере 5-кратное усиление kON по сравнению с исходным scFv; три из которых демонстрировали 20-кратное увеличение kON. По сравнению с увеличением kON, эффекты "созревания аффинности" были весьма незначительными. Только три scFv, подвергшихся «созреванию аффинности», демонстрировали 4-5-кратное увеличение kOFF. Интересно, что два из них были выделены на основе селекции с конкуренцией, которая могла быть более эффективным способом селекции для scFv со сниженной kOFF по сравнению с длительной инкубацией в буфере отмывки.
Температуры плавления избранных кандидатов, подвергшихся «созреванию аффинности», имеют среднее значение 62,2°С, и которые, будучи определенными посредством дифференциальной сканирующей флуорометрии (ДСФ), варьировали от 53°С до 67,2°С.
Сохранение высокой специфичности к EGFRvIII по сравнению с EGFR дикого типа во время "созревания аффинности»
«Созревание аффинности», как правило, сопровождается небольшими изменениями эпитопа связывания, что может влиять на специфичность/перекрестную реактивность (Barbas et al., 1994, Parsons et al., 1996, Winkler et al., 2000). Новый эпитоп EGFRvIII no сравнению с EGFR дикого типа образуется в результате делеции внутри рамки считывания 269 аминокислот из внеклеточного домена EGFR (экзоны 2-7), и ожидается, что он довольно маленький; он заключается в новом смежном положении аминокислот (аминокислота 5 соединяется с аминокислотой 274) и одной новой аминокислоты GLY в сайте делеции. Предполагается, что даже небольшие изменения эпитопа связывания в данном случае быстро нарушают тонкую специфичность к EDFRvIII и могут приводить к образованию связывающих белков с перекрестной реактивностью, которые также распознают нативный EGFR.
Неожиданно «созревание аффинности», описываемое в настоящей заявке, привело к увеличению связывания анти-EGFRvIII антител до 100 раз без какой-либо потери исключительной специфичности к EGFRvIII (Таблица 3, Фигура 2).
Во время селекции подвергшихся «созреванию аффинности» связывающих белков уникальные EGFRvIII-специфичные scFv оценивали на предмет связывания с EGFRvIII-экспрессирующими клетками СНО (CHOEGFRvIII) и F98 (F98EGFRvIII) посредством проточной цитометрии, а также с экспрессирующими человеческий EGFR дикого типа клетками СНО (CHOEGFR) и F98 (F98EGFR), и с нетрансфецироваными клетками СНО и F98.
11 высоко специфичных анти-EGFRvIII scFv, не демонстрирующих измеримого фонового связывания на EGFR-позитивных клетках (CHOEGFR) или нетрансфецированных клетках СНО, были охарактеризованы более подробно. В лучшем случае определялось сродство к EGFRvIII 80 пМ, что соответствует показателю улучшения 100 раз по сравнению с исходным scFv (Таблица 3). В то же время ни один из исследуемых анти-EGFRvIII scFv не проявлял никакой перекрестной реактивности с антигеном EGFR дикого типа (Фигура 2).
Насколько нам известно, такое высокое сродство к EGFRvIII у домена антитела человека в сочетании с исключительной специфичностью к EGFRvIII и без определимой перекрестной реактивности с EGFR дикого типа ранее никогда не наблюдалось. Safdari с соавторами недавно описали результаты гуманизации EGFRvIII-связывающего домена антитела мыши - MR1 (Safdari et al., 2014), который в своей мышиной форме впервые был описан Lorimer с соавторами (Lorimer et al., 1996, Beers et al., 2000, Kuan et al., 2000). В ходе процесса гуманизации они достигли повышения сродства для humMR1, достигающего сходных пикомолярных значений KD, которые описаны в настоящей заявке, однако в отличие от нашего изобретения они не смогли полностью упразднить перекрестную реактивность указанного домена антитела с EGFR дикого типа, которую можно четко видеть на фигуре 4 в их публикации (Safdari et al., 2014). Следовательно, humMR1 не является специфичным в отношении EGFRvIII (145кДа), но также распознает EGFR (170кДа) после SDS PAGE и вестерн-блоттинга (Safdari et al., 2014). Мы всесторонне проверили специфичность и перекрестную реактивность наших EGFRvIII-связывающих доменов как до, так и после «созревания аффинности» и неожиданно не обнаружили никаких признаков перекрестной реактивности с EGFR дикого типа ни с очищенным рекомбинантным антигеном EGFR дикого типа, проанализированным посредством Biacore (Фигура 2), ни в SDS-PAGE и вестерн-блоттинге (Фигура 4), ни при проведении ELISA (Фигура 5). Наши EGFRvIII-связывающие домены также не проявляют никакой перекрестной реактивности с EGFR дикого типа, гиперэксепрссируемым на поверхности трансфецированных клеток СНО (CHOEGFR) или клеток глиомы F98 (F98EGFR) (Фигура 3).
ПРИМЕР 3: Проточно-цитометрическая оценка EGFRvIII-специфичных антител scFv с высокой специфичностью и высоким средством с клетками глиомы крысы F98, гиперэкспрессирующими EGFRvIII (F98EGFRvIII), или со стабильно трансфецированными клетками СНО, гиперэкспрессирующими EGFRvIII (CHOEGFRvIII); отсутствие связывания с клетками F98, гиперэкспрессирующими нативный EGFR (F98EGFR), или нетрансфецированными клетками F98 (F98), а также отсутствие связывания с клетками СНО, гиперэкспрессирующими нативный EGFR (CHOEGFR), или нетрансфецированными клетками СНО (СНО)
Материалы (среда, буфер и реагенты)
Анти-His IgG 13/45/31 («Dianova»), FCS (фетальная телячья сыворотка) («Invitrogen»), Anti-His IgG 13/45/31 (Dianova), FCS (Invitrogen), фикол-пак PLUS («GE Healthcare»), FITC-конъюгированный антимышиный IgG козы, мин X («Dianova»), L-глутамин («Invitrogen»), NaN3 («Carl Roth»), ФСБ («Invitrogen»), пенициллин/стрептомицин («Invitrogen»), пропидий иодид («Sigma»), RPMI-1640 («Invitrogen»).
Клетки и линии клеток
Клетки глиомы крысы F98, клетки F98, гиперэкспрессирующие EGFR (F98EGFR), а также клетки F98, гиперэкспрессирующие EGFRvIII (F98EGFRvIII), приобретали в Американской коллекции типовых культур (АТСС) и культивировали в соответствии с рекомендуемыми протоколами.
|
Стабильно трансфецированные клетки СНО, экспрессирующие EGFR или EGFRvIII, генерировали в компании «Affimae» в соответствии со следующим протоколом:
Гены, кодирующие EGFR дикого типа или мутант, полученный в результате делеции, EGFRvIII синтезировала компания «Life Technologies»/«GeneArt» (Регенсбург, Германия) и субклонировали в вектор экспрессии для млекопитающих pcDNA5/FRT. Стабильные линии клеток СНО, экспрессирующие рекомбинантный EGFR дикого типа или EGFRvIII, генерировали на основе линии клеток-хозяев Flp-In СНО («Life Technologies»), предварительно адаптированной к росту в суспензии. За один день до трансфекции клетки Flp-In СНО пересевали в среды HyClone CDM4 СНО с добавлением L-глутамина, добавки НТ и пенициллина/стрептомицина (без зеоцина). Линии клеток со стабильной экспрессией генерировали посредством совместной трансфекции линии клеток Flp-In СНО продуктом экспрессии на основе pcDNA5/FRT и плазмидой интеграции, а также плазмидой экспрессии рекомбиназы pOG44 («Life Technologies») с применением реагента трансфекции - полиэтиленимина (ПЭИ). 2,5 мкг тотальной ДНК разводили в 50 мкл среды OptiMEMI и комбинировали с 7,5 мкг ПЭИ, разбавленного в 50 мкл среды OptiMEMI. Смесь инкубировали в течение 10 минут, а затем добавляли к 2×106 клеток Flp-In СНО, суспендированных в 1 мл среды CHO-S-SFMII. Через один день после трансфекции клетки разбавляли до плотности 1×105 жизнеспособных клеток/мл в среде CHO-S-SFMII с добавлением 500 мкг/мл гигромицина В и засевали в колбы для культур Т75. Рекомбиназа Flp опосредует вставку конструкции экспрессии Flp-In в геном в интегрированном сайте FRT посредством сайт-специфичной рекомбинации ДНК. Во время фазы селекции плотность жизнеспособных клеток измеряли один раз в две недели, и клетки центрифугировали и повторно суспендировали в свежей среде для селекции при максимальной плотности 1×105 клеток/мл. Клетки, стабильно экспрессирующие EGFR или EGFRvIII, восстанавливались приблизительно после 2-3 недель селекции с применением гигромицина В, и затем их переносили в суспензионную питательную среду HyClone CDM4 СНО. Стабильно экспрессирующие EGFR дикого типа или EGFRvIII клетки замораживали в среде, содержащей 50% ProFreeze («Lonza») и 7,5% DMSO (диметилсульфоксид).
Для определения плотности поверхностных антигенов EGFR дикого типа или EGFRvIII на стабильно трансфецированных линиях клеток, применяли проточно-цитометрическую косвенную иммунофлуоресцентную оценку (QIFIKIT, Dako) в соответствии с инструкциями производителя, и были продемонстрированы почти равные плотности EGFR дикого типа на стабильно трансфецированных клетках CHOEGFR и EGFRvIII на стабильно трансфецированных клетках CHOEGFRvIII, тогда как на нетрансфецированных клетках СНО не удавалось определить сайтов связывания EGFR дикого типа или EGFRvIII.
Определение связывания антитела и сродства посредством проточной цитометрии
Клетки инкубировали со 100 мкл последовательных разведений указанных антител scFv, начиная с 100 мкг/мл (~3300 нМ) в буфере для FACS в течение 45 мин на льду. После трехкратной отмывки буфером для FACS клетки инкубировали с 0,1 мл 10 мкг/мл моноклонального анти-His антитела мыши клона 13/45/31 в том же буфере в течение 45 мин на льду. После второго цикла отмывки клетки инкубировали с 0,1 мл 15 мкг/мл FITC-конъюгированного антимышиного антитела IgG козы при тех же условиях, что и ранее. В качестве контроля клетки инкубировали с анти-His IgG 13/45/31, а затем с FITC-конъюгированными антимышиными антителами IgG козы без scFv. Затем клетки снова отмывали и повторно суспендировали в 0,2 мл буфера для FACS, содержащего 2 мкг/мл пропидия иодида, чтобы исключить погибшие клетки. Флуоресценцию 1×104 живых клеток измеряли при помощи проточного цитометра Beckman-Coulter FC500 MPL с применением программы МХР («Beckman-Coulter", Крефельд, Германия) или проточного цитометра Millipore Guava EasyCyte с применением программы Incyte («Merck Millipore», Швальбах, Германия). Средние интенсивности флуоресценции проб клеток рассчитывали при помощи программы СХР («Beckman-Coulter", Крефельд, Германия) или Incyte («Merck Millipore», Швальбах, Германия).
В случае анализа при помощи проточного цитометра Beckman-Coulter FC500 MPL применяли 0,5×106 клеток на окрашивание, а в случае применения проточного цитометра Millipore Guava EasyCyte - 0,25×106 клеток/окрашивание.
После вычитания значений интенсивности флуоресценции клеток, окрашенных только вторичными и третичными реагентами, рассчитывали значения KD по формуле для связывания в одном сайте (гипербола) в программе GraphPad Prism (GraphPad Prism версия 6.00 для Windows, программа GraphPad, Ла-Хойя, Калифорния, США). Результаты показаны на Фигуре 3.
ПРИМЕР 4: SDS PAGE и вестерн-блоттинг для анализа специфичности связывания EGFRvIII-связывающих антител
Для проверки специфичности связывания EGFRvIII/CD3-тандемных диател, содержащих разные связывающие домены для антигена EGFRvIII, и для демонстрации отсутствия связывания с антигеном EGFRvIII дикого типа проводили электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и вестерн-блоттинг. Последовательности, кодирующие внеклеточный домен EGFR дикого типа или процессированный EGFRvIII соединяли посредством технологий рекомбинантных ДНК с частью Fc антитела IgG человека. ДНК-конструкции трансфецировали в клетки СНО, которые секретировали рекомбинантный EGFR-Fc дикого типа или EGFRvIII-Fc в виде растворимых белков в надосадочную жидкость культуры клеток, из которой его очищают при помощи хроматографии с белком А. За счет образования межмолекулярной дисульфидной связи в части Fc гибридные антигены EGFR дикого типа-Fc (или EGFRvIII-Fc) образуют димеры из двух идентичных цепей. EGFRvIII-Fc в результате EGFRvIII-специфичной делеции 267 аминокислот обладает массой приблизительно на 25 кДа меньше, чем EGFR-Fc, что ведет к отличию размера димерных форм белков с Fc-частью в 50 кДа.
Равные количества очищенного EGFR-Fc дикого типа (и EGFRvIII-Fc) смешивают с невосстановительным буфером для проб SDS PAGEx2 или восстановительным буфером для проб SDS PAGEx2, содержащим дитиотриетол (ДТТ) в качестве восстановителя. Пробы с ДТТ нагревают при 95°С в течение 5 мин перед загрузкой на 4-20% гели для SDS PAGE Criterion TGX Precast («Biorad»). Применяют 1 мкг пробы белка на дорожку. Для разделения белков в геле SDS-PAGE запускают в 1х Трис/глицин/SDS-буфере ("Biorad») при 300 В приблизительно в течение 25 мин. Общий белок визуализируют в геле при помощи системы молекулярной визуализации Criterion Stain-free («Biorad»). В качестве маркера молекулярных масс применяют Page Ruler Unstained Protein («Thermo Scientific»). Затем гель подвергают блоттингу на мембране из ПВДФ с применением системы полусухого блоттинга Fastblot от компании «Biorad». Мембрану блокируют в 3% растворе (масса/объем) сухого обезжиренного молока в 1×TBS в течение 30 мин при комнатной температуре. Мембраны разрезают на куски, каждый из которых содержит аналогичные пробы белка, подвергнутые блоттингу. Разные тандемные диатела разводят до концентрации 2 мкг/мл в 3% растворе (масса/объем) сухого обезжиренного молока в 1 × TBS и инкубируют с отдельными фрагментами мембраны в течение 1 часа на качающейся платформе. В качестве контроля применяют анти-EGFR антитело цетуксимаб в концентрации 10 мкг/мл. Мембраны промывают TBST (TBS+0,1% (по объему) Tween 20) три раза по 10 мин каждую и однократно промывают ТВS перед инкубацией со вторичными конъюгированными с пероксидазой хрена (HRP) антителами детекции, Penta HIS-HRP (QIAGEN) с целью детекции тандемных диател или с белком L-HRP («Thermo Scientific») для определения цетуксимаба, разведенных 1:5000 в 3% растворе сухого обезжиренного молока в TBS в течение 1 часа на качающейся платформе. Мембраны промывают TBST (TBS+0,1% (по объему) Tween 20) три раза по 10 мин каждую и однократно промывают TBS. Обусловленное HRP развитие окраски на мембране инициируют путем добавления 0,06% диаминобензидина + 0,02% CoCl2+ 0,015% H2O2 в TBS. Реакцию останавливают путем добавления воды. Мембраны высушивают и сканируют. Результаты показаны на Фигуре 4.
ПРИМЕР 5: Анализ связывания при помощи твердофазного иммуноферментного анализа (ELISA) для EGFRvIII/CD3-тандемных диател, содержащих разные EGFRvIII-связывающие домены к антигену EGFRvIII, и связывания с CD3, а также специфичность их связывания с EGFRvIII в отсутствии связывания антигена EGFR дикого типа
Процедура ELISA
В планшеты для ELISA на 96 лунок (Immuno MaxiSorp; «Nunc») вносили EGFRvIII-Fc, EGFR дикого типа -Fc или рекомбинантный антиген в 100 мМ карбонат-бикарбонатного буфера и оставляли на ночь при 4°С. Чтобы получить приблизительно равную мольную плотность покрытия антигенами EGFRvIII-Fc вносили в концентрации 4 кг/мл, EGFR-Fc в концентрации 6 мкг/мл, а антиген CD3γε вносили в концентрации 1,5 мкг/мл. После этапа блокирования 3% раствором (масса/объем) сухого обезжиренного молока («Merck») в ФСБ, 11 последовательных разведений разных EGFRvIII/CD3-тандемных диател в концентрации от 200 мкг/мл до 6,5×106нг/мл в ФСБ, содержащих 3% раствор (масса/объем) сухого обезжиренного молока («Merck») в ФСБ, инкубировали на планшетах в течение 1,5 ч при 25°С. После инкубации планшеты отмывали трижды 300 мкл на лунку ФСБ, содержащего 0,1% (по объему) Tween 20 («Serva»). Добавляли 50 нг/мл белка L-HRP и инкубировали в планшетах в течение 1 ч при 25°С. После трехкратной отмывки 300 мкл на лунку ФСБ, содержащего 0,1% (по объему) Tween 20, для детекции добавляли субстрат тетраметилбензидин (ТМВ) («Seramun»). Реакцию останавливали приблизительно через 2 минуты путем добавления 100 мкл на лунку 0,5 M H2SO4. Поглощение в лунках измеряли на длине волны 450 нм на планшетном ридере для многолуночных планшетов (Victor, «Perkin Elmer»).
Значения поглощения откладывали на графике в форме диаграммы при помощи программы GraphPad Prism («GraphPad Software», Сан-Диего, Калифорния).
Результаты показаны на Фигуре 5.
ПРИМЕР 6: Измерение связывания тандемных диател с EGFRvIII посредством поверхностного пзамонного резнонанса
Получение чипа СМ5 на основе IgG к Fc человека
Чип СМ5 на основе IgG к Fc человека получали, как было описано выше (Пример 2).
Диапазон концентраций тандемных диател, применяемых при измерении поверхностного пзамонного резнонанса:
Концентрации тандемных диател проверяли до измерения поверхностного пзамонного резнонанса путем определения А280 при помощи Nandrop. Концентрацию в матричном растворе (с) рассчитывали по формуле:
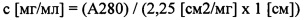
Разные коэффициенты экстинкции рассчитывали при помощи инструмента для прогнозирования «Expasy Protparam» (http://web.expasy.org/protparamA). Для расчета концентрации в молях, полагали молекулярную массу равной приблизительно 105 кДа для всех тандемных диател. Концентрации тандемных диател корректировали на конечные разведения, упоминаемые ниже, путем последовательного разведения в 1×HBS-P+буфере. Готовили следующие концентрации: 150 нМ, 30 нМ, 6 нМ, 1,2 нМ, 0,24 нМ, 0,048 нМ и 0 нМ, или 90 нМ, 30 нМ, 10 нМ, 3,33 нМ, 1,11 нМ, 0,37 нМ, 0,123 нМ, 0,041 нМ и 0 нМ, или как указано.
Условия анализа связывания для измерения KD тандемных диател
Гибридный белок EGFRvIII-Fc разводили в 1×HBS-P+ до концентраций 6,25 нМ. Раствор антигена вводили со скоростью потока 10 мкл/мин в течение 40 сек для EGFRvIII-Fc. Антиген вводили в проточную ячейку 2. Проточная ячейка 1 выступала в качестве ячейки сравнения.
В качестве подвижного буфера на протяжении всей процедуры применяли 1x HBS-P+. Разведения тандемных диател вводили со скоростью потока 30 мкл/мин в течение 180 сек во все проточные ячейки. Сигналы, измеренные в проточной ячейке 1 без захваченного антигена, вычитали из сигналов в проточной ячейке 2, чтобы скорректировать кривые связывания на фоновое связывание. После периода диссоциации 540 сек чип регенерировали, как описано ниже.
Чипа на основе IgG к Fc человека регенерировали путем введения буфера с высоким содержанием соли 3,0 M MgCl2 (3×15 сек, 10 мкл/мин). Перед каждым измерением проводили три пусковых периода без тандемных диател. Измерения в растворах тандемных диател проводили от наименьшей к наибольше концентрации. Включали один цикл с чистым подвижных буфером (0 мкМ).
Параметры связывания тандемных диател определяли путем мультицикилических измерений кинетики (МЦК). Для оценки кажущегося сродства связывания кривые связывания оценивали при помощи способа «Кинетика», включенного в программу оценки Biacore X100. При помощи указанного способа кривые связывания целиком и локально аппроксимировали при помощи программы с применением «модели взаимодействия Ленгмюра 1:1».
Параметры связывания тандемных диател
Чтобы проанализировать влияние сочетания двух EGFRvIII-связывающих доменов в двухвалентной или поливалентной, или мультиспецифичной молекуле, такой как EGFRvIII/CD3 тандемное антитело, кажущееся сродство EGFRvIII/CD3 тандемных диател к антигену EGFRvIII измеряли посредством поверхностного плазмонного резонанса с применением Biacore X100 (Фигура 6).
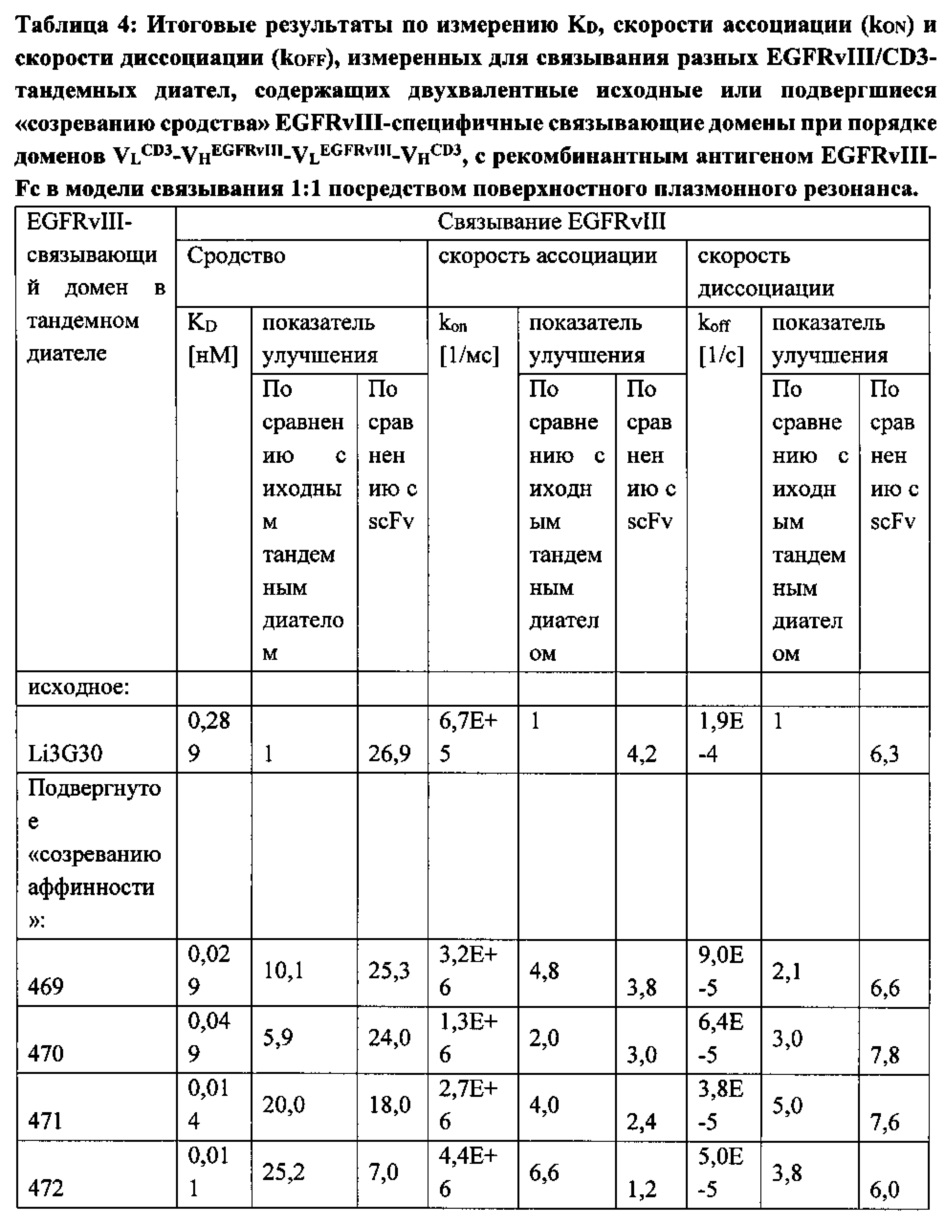
Кажущееся сродство EGFRvIII-специфичных связывающих доменов в EGFRvIII/тандемных диателах было повышенным во всех исследованных молекулах, и KD для самого лучшего тандемного диатела достигало 11 пМ. Уменьшение KD тандемных диател, содержащих домены, подвергшиеся «созреванию аффинности», по сравнению с соответствующим тандемным диателом, содержащим исходный EGFRvIII-связывающий домен, достигало 25 раз.
Повышение KD относительно KD, измеренного для одновалентных связывающих scFv, также достигало 25 раз. Интересно, что в отличие от повышения, достигаемого посредством «созревания аффинности», которое было преимущественно обусловлено повышением kON, дальнейшее увеличение сродства связывания у двухвалентной формы тандемного диатела достигалось главным образом за счет более медленной kOFF (Таблица 4).
Следовательно, форма тандемного диатела обеспечивает путь для дальнейшего увеличения сродства связывания при сохранении специфичности связывающего домена антитела.

ПРИМЕР 7: Иммуногистохимическое (ИГХ) окрашивание срезов ткани солидных опухолей анти- EGFRvIII биспецифическим диателом или цетуксимабом
В отличие от EGFR дикого типа, который широко экспрессируется в здоровых тканях, экспрессия мутированного рецептора EGFRvIII в высокой степени специфична для опухолей. Хотя высокая частота экспрессии EGFRvIII в глиобластома систематически описана в литературе, распространенность экспрессии EGFRvIII и однородность экспрессии в других опухолях остается противоречивой.
Было инициировано небольшое иммуногистохимическое (ИГХ) исследование, и срезы тканей от 3 разных пациентов с раком - с глиобластомой (ГБ), раком головы и шеи (РГШ), раком предстательной железы, Her2-негативным раком молочной железы, а также немелкоклеточным раком легких (NSCLC) анализировали на реактивность с применением EGFRvIII-специфичного связывающего домена согласно настоящему изобретению в форме двухвалентного диатела (Фигура 7).
Иммуногистохимическое исследование проводили, чтобы оценить экспрессию EGFRvIII в разных опухолях, а также специфичность EGFRvIII-связывающих антител в отношении опухолей. Связывание EGFRvIII EGFRvIII-специфичными вариабельными доменами исследовали в форме двухвалентного диатела, содержащего EGFRvIII-связывающий домен Li3G30 в расположении VHEGFRvIII-VLEGFRvIII. Вторичную детекцию проводили при помощи метки-His на белке диатела. В качестве контроля применяли EGFR-специфичное антитело IgG цетуксимаб (Эрбитукс).
Все срезы тканей человека получали из «BioChain Institute, Inc.». и со срезами работали в соответствии с инструкцией поставщика. Срезы тканей адаптировали к комнатной температуре и инкубировали с пероксидазным блоком DAKO после блокирования 10% сывороткой козы. EGFRvIII-специфичное диатело инкубировали при двух концентрациях 0,5 мкг/мл и 0,1 мкг/мл в течение одного часа, а затем с антителом анти-His («Dianova»), и определяли при помощи системы Envision+ HRP-DAB для антител мыши (DAKO). Окрашивание срезов тканей контрольным веществом эрбитукс («Merck KGaA») проводили при концентрации 10 мкг/мл с применением системы детекции Klear Human HRP-Polymer DAB («GBI Labs») в соответствии с инструкциями производителя. В конце срезы окрашивали гематоксилином и заливали в среду для заключения препаратов (Shandon Consul Mount™, «Thermo Scientific»).
Результаты показаны на Фигуре 7.
Два из трех образов глиобластомы демонстрировали явную и специфичную реактивность при применении EGFRvIII-специфичного диатела. Неожиданно, все три образца рака головы и шеи продемонстрировали сильную и явную позитивность в отношении EGFRvIII, а также в раке предстательной железы, раке молочной железы и НКРЛ было получено специфическое окрашивание ткани опухоли при применении EGFRvIII-специфичного диатела, описываемого в настоящей заявке (фигура 7).
На основании указанных свойств EGFRvIII-связывающие домены, описываемые в настоящей заявке, прекрасно подходят для разработки мультиспецифичных, поливалентных, вовлекающих иммуно-эффекторные клетки, лекарственных средств, направленных на опухоли, например, таких как, например, EGFRvIII/CD3-биспецифичные тандемные диатела.
ПРИМЕР 8: Оценка цитотоксической активности, опосредуемой EGFRvIII/CD3 - тандемными диателами в основанном на FACS анализе цитотоксичности
EGFRvIII/CD3-тандемные диатела, содержащие исходные или подвергшиеся «созреванию аффинности» EGFRvIII-специфичные домены в сочетании с разными CD3+-связывающими доменами и/или содержащими разный порядок отдельных связывающих доменов в молекулах тандемного диатела, анализировали в ходе исследования цитоткосичности на основе клеток (Фигура 8).
Мишень-опосредуемую зависимость, специфичность тандемных диател и Т-лимфоцит-опосредуемое уничтожение анализировали путем применения экспрессирующих EGFRvIII клеток СНО (CHOEGFRvIII), клеток СНО, экспрессирующих EGFR дикого типа (CHOEGFRI), и нетрансфецированных клеток СНО в качестве клеток-мишеней. Материал (среды, буфер и реагенты):
DMSO («Sigma»), EasySep™ Human Τ Cell Enrichment Kit («Stem Cell Technologies»), набор для обогащения Т-лимфоцитами CD4+ человека EasySep™ («Stem Cell Technologies»), набор для обогащения Т-лимфоцитами CD8+ человека EasySep™ («Stem Cell Technologies»), магнит Big Easy EasySep™ («Stem Cell Technologies»), FCS (Invitrogen), Lymphoprep («Stem Cell»), L-глутамин («Invitrogen»), mlgGl FITC («ADG»), CD16-FITC (MEM-154) («Thermo Fisher Scientific»), mIgG1-FITC / mIgG1-PE / mIgG1-ECD («Beckman Coulter»), mIgG1-PE («Beckman Coulter»), mIgG1-PC5 («Beckman Coulter»), mIgG1-PC7 («Beckman Coulter»), CD8-FITC / CD4-PE / CD3-ECD («Beckman Coulter»), CD16-PC5 («Beckman Coulter»), CD19-PC7 («Beckman Coulter»), CD16-FITC / CD56-PE / CD3-ECD («Beckman Coulter»), CD14-PC7 («Beckman Coulter»), CD33-PE («MACS Miltenyi Biotech»), NaN3 («Roth»), ФСБ («Invitrogen»), пенициллин/стрептомицин («Invitrogen»), набор PKH67 Green Fluorescent Cell Linker Midi Kit («Sigma»), IgG человека Gammanorm («Octapharma»), пропидий иодид («Sigma»), RPMI-1640 («Invitrogen»).
Выделение МКПК из лейкоцитарной пленки и обогащение Т-лимфоцитов
Лейкоцитарную пленку приобретали у Deutsches Rotes Kreuz, DRK-Blutspendedienst  (Мангейм, Германия) в день отбора крови; препараты лейкоцитарной пленки хранили в течение ночи при комнатной температуре, перед тем как выделить МКПК посредством центрифугирования в градиенте плотностей. Пробы лейкоцитарной пленки разводили двух-трехкратным объемом ФСБ, наслаивали на основание из Lymphoprep и центрифугировали при 800×g в течение 25 мин при комнатной температуре без разбивания комков. МКПК, расположенные на поверхности, отбирали и промывали трехкратно ФСБ, перед тем как их применяли для обогащения Т-лимфоцитов или проточно-цитометрического анализа. Обогащение Т-лимфоцитов из популяции МКПК проводили при помощи набора EasySep™ Human Т Cell Enrichment Kit для иммуномагнитного выделения интактных Т-лимфоцитов человека и магнита Big Easy EasySep™ Magnet в соответствии с инструкциями производителя.
(Мангейм, Германия) в день отбора крови; препараты лейкоцитарной пленки хранили в течение ночи при комнатной температуре, перед тем как выделить МКПК посредством центрифугирования в градиенте плотностей. Пробы лейкоцитарной пленки разводили двух-трехкратным объемом ФСБ, наслаивали на основание из Lymphoprep и центрифугировали при 800×g в течение 25 мин при комнатной температуре без разбивания комков. МКПК, расположенные на поверхности, отбирали и промывали трехкратно ФСБ, перед тем как их применяли для обогащения Т-лимфоцитов или проточно-цитометрического анализа. Обогащение Т-лимфоцитов из популяции МКПК проводили при помощи набора EasySep™ Human Т Cell Enrichment Kit для иммуномагнитного выделения интактных Т-лимфоцитов человека и магнита Big Easy EasySep™ Magnet в соответствии с инструкциями производителя.
Изучение свойств эффекторных клеток посредством проточно-цитометрического анализа
Для оценки распределения субпопуляции МКПК и чистоты подгрупп обогащенных Т-лимфоцитов проводили окрашивание МКПК.
Для проточно-цитометрического анализа выделенные МКПК или обогащенные субпопуляции МКПК повторно суспендировали в ФСБ с добавлением 2% термоинактивированной FCS и 0,1% азида натрия (называется буфером для FACS) и 1 мг/мл поликлонального IgG человека с получением плотности 0,5-2×106/мл. Аликвоты суспензии клеток по 0,5 мл затем инкубировали в течение 15 мин в темноте с антителами по следующей схеме, как рекомендует производитель.
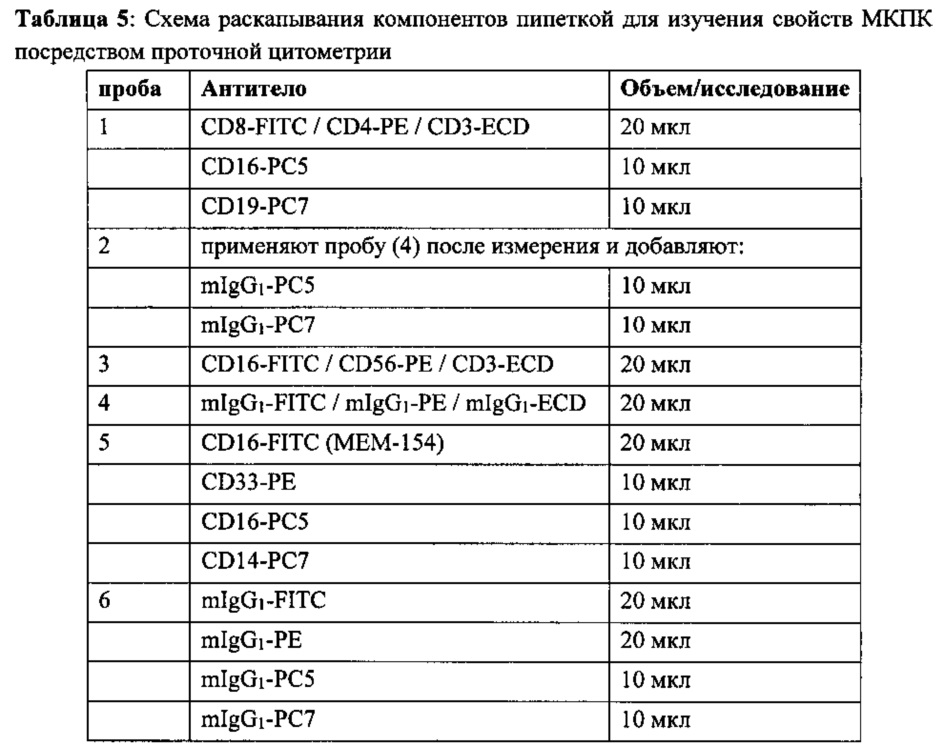
После инкубации пробы измеряли в порядке: 3. 4. 5. 6. 1. 2 на проточном цитометре Beckman-Coulter FC500 MPL («Весkman-Coulter", Крефельд, Германия) с применением протоколов, утвержденных для МКПК и многоцветового окрашивания (FITC РЕ ECD РС5 РС7; FITC РЕ ECD; FITC РЕ РС5 РС7) в программе захвата МХР («Beckman-Coulter»). Для анализа и построения графиков данных применяли программу для анализа СХР («Beckman-Coulter»).
Анализ цитотоксичности:
Т-лимфоциты, которые применяли в качестве эффекторных клеток, характеризовали посредством проточной цитометрии, как было описано. Клетками-мишенями, применяемыми при анализе цитотоксичности, были клетки СНО, стабильно трансфецированные для гиперэкспрессии EGFRvIII (CHOEGFRvIII), клетки СНО, стабильно трансфецированные для экспрессии EGFR дикого типа (CHOEGFR), или нетрансфецированные клетки СНО (СНО). Указанные линии клеток генерировали, как было описано ранее, и культивировали при стандартных условиях, как описано. Для анализа цитотоксичности клетки-мишени отбирали, дважды промывали средой RPMI 1640 без FCS и повторно суспендировали в растворителе С, поставляемом в составе набора РКН67 Green Fluorescent Cell Linker Midi Kit, чтобы получить плотность клеток 2×107/мл. Затем суспензию клеток смешивали с равным объемом концентрированного в 2 раза РКН67-меченного раствора (например, 1 мкл в 250 мкл растворителя С) и инкубировали в течение 2-5 мин при КТ и периодическом перемешивании в соответствии с инструкциями производителя. Реакцию окрашивания прекращали путем добавления равного объема FCS и инкубировали в течение 1 мин. После отмывки меченных клеток-мишеней полной средой RPMI, клетки подсчитывали и повторно суспендировали до плотности 2×105/мл в полной среде RPMI. Затем 2×104 клеток-мишеней засевали вместе с Т-лимфоцитами в соотношении Е:Т 5:1 и указанными антителами в отдельные лунки микропланшетов на 96 лунок с круглым дном в общем объеме 200 мкл/лунку. Обычно исследовали 9 последовательных разведений 1:5, начиная с 30 мкг/мл. Спонтанная гибель клеток и уничтожение мишеней эффекторами в отсутствии антител определяли по меньшей мере в трех повторах в каждом планшете. Опосредуемое тандемными диателами уничтожение обычно определяли в двух повторах.
После центрифугирования в течение 2 мин при 200 g при КТ планшеты для анализа инкубировали в течение 40 ч - 48 ч при 37°С в увлажненной атмосфере с 5% СО2. После инкубации культуры промывали однократно буфером для FACS, а затем повторно суспендировали в 150 мкл буфера для FACS с добавлением 2 мкг/мл PI (ингибитор протеазы). Абсолютное количество живых клеток-мишеней, которые характеризовались по положительной зеленой окраске на РКН67, но негативной на PI, измеряли при помощи проточного цитометра Millipore Guava EasyCyte («Merck Millipore»).
На основании измеренного количества оставшихся живых клеток-мишеней процент специфичнеского лизиса клеток рассчитывали по следующей формуле: [1-(количество живых мишеней (проба) / (количество живых мишеней (спонтанное))]×100%. S-образные кривые зависимости ответа от дозы и значения ЕС50 рассчитывали по нелинейной регрессии/4-параметрической линейной аппроксимации с применением программы GraphPad Prism (GraphPad Prism, версия 6.00 для Windows, «GraphPad Software», Ла-Хойя, Калифорния, США).
Статистический анализ
Значения лизиса, полученные для конкретной концентрации антител, определяли в двух повторах и анализировали по S-образной кривой доза-ответ/4-параметрической логистической аппроксимации при помощи программы GraphPad Prism (GraphPad Prism, версия 6.00 для Windows, «GraphPad Software», Ла-Хойя, Калифорния, США) и применяли для расчета средних значений ЕС50 и СО для повторов процентов лизиса. Результаты показаны на Фигуре 8 и в Таблице 6.
Все исследуемые тандемные диатела проявляли концентрационную зависимость, высокую специфичность и цитотоксичность, а также высокую эффективность в отношении EGFRvIII-экспрессирующих клеток, тогда как выживание EGFRvIII-негативных клеток не нарушалось. Во всем диапазоне концентраций любого из исследуемых тандемных диател не наблюдалось никакой определимой цитотоксичности на EGFRwt-экспрессирующие или нетрансфецированные клетки. Цитотоксичность, опосредуемая EGFRvIII/CD3-тандемными диателами, следовательно, строго зависит от мишени. Тандемное диатело, содержащее исходный EGFRvIII-связывающий домен перед «созреванием аффинности» и с порядком расположения доменов VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3, в анализе цитотоксичности демонстрировало ЕС50 25 пМ (Фигура 8). Соответствующие тандемные диатела, содержащие EGFRvIII-связывающие домены с улучшенным сродством при том же порядке расположения доменов (VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3), систематически демонстрируют усиленную активность цитотоксичности, достигающую ЕС50 1,5 пМ в самом лучшем случае (Таблица 6). Относительное повышение цитотоксической активности было более заметным в тандемных диателах, содержащих EGFRvIII-связывающие домены во внешнем положении (порядок доменов VHEGFRvIII-VLCD3-VHCD3-VLEGFRvIII) (Таблица 6). Удается достигнуть 70-кратного повышения цитотоксической активности у данных тандемных диател путем замены EGFRvIII-связывающего домена с низким сродством Li3G30 на домены с улучшенным сродством (Фигура 8, Таблица 6).
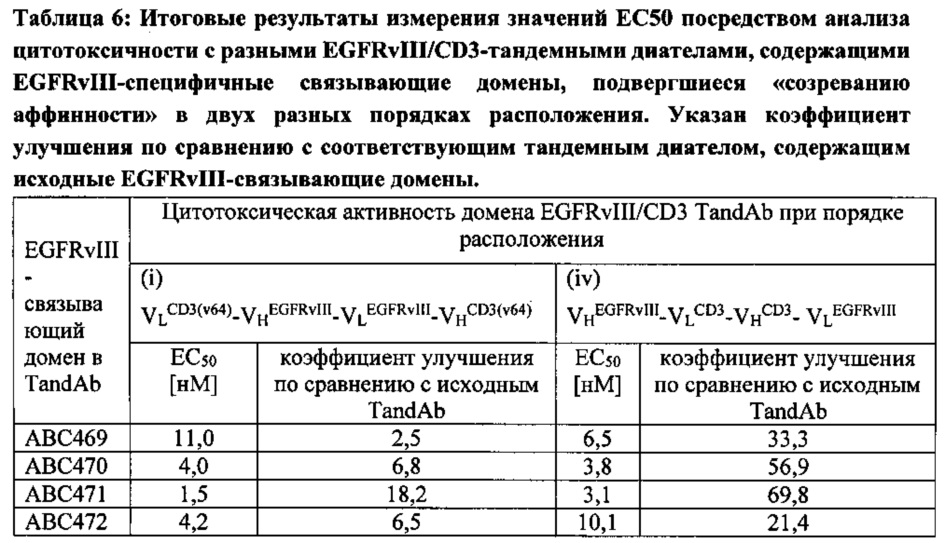
Choi et al. (Proc Natl Acad Sci USA. 2013 Jan 2; 110(1):270-5; WO2013/7185010) описали препарат сравнения для EGFRvIII/CD3-биспецифического активатора Т-лимфоцитов (BiTE), содержащий один сайт связывания для EGFRvIII (на основе домена MR1 мыши) и один сайт связывания для CD3 (на основе анти-CD3 антитела ОКТ3). При анализе цитотоксичности на основе клеток данное антитело достигало 50% от максимально достижимого специфичного лизиса при концентрации приблизительно 10-2 мкг/мл, который соответствует мольной концентрации ЕС50 приблизительно 200 пМ для BiTE 52 кДа (Фигура 4С в Choi et al., 2013). Сравнение показывает, что даже описываемые в настоящей заявке тандемные диатела, содержащие исходный, не подвергшийся «созреванию аффинности» EGFRvIII-связывающий домен, обладают приблизительно в 10 раз более высокой цитотоксичностью, чем EGFRvIII/CD3 BiTE. Кроме того, тандемные диатела согласно настоящему изобретению, содержащие подвергшиеся «созреванию аффинности» EGFRvIII-связывающие домены, проявляют даже 100-краткное увеличение цитотоксичности по сравнению с цитотоксичностью EGFRvIII/CD3 BiTE.
ПРИМЕР 9: интернализация рецепторов EGFRvIII под действием EGFRvIII/CD3-тандемных диател на клетках СНО, экспрессирующих рекомбинантный EGFRvIII (CHOEGFRvIII), или клетках F98, экспрессирующих рекомбинантный EGFRvIII (F98EGFRvIII)
Для оценки модуляции EGFRvIII EGFRvIII/CD3-тандемными диателами клетки СНО, экспрессирующие рекомбинантный EGFRvIII (CHOEGFRvIII), или клетки F98, экспрессирующие рекомбинантный EGFRvIII (F98EGFRvIII), инкубировали с аликвотами 2,5×105 в отдельных лунках с круглым дном микропланшета на 96 лунок вместе с возрастающими концентрациями EGFRvIII/CD3-тандемных диател в среде RPMI 1640 с добавлением 10% FCS, 2 мМ L-глутамина, 100 МЕ/мл натриевой соли пенициллина G, 100 мкг/мл стрептомицина сульфата, 100 мкг/мл пирувата натрия (в настоящей заявке называется полной средой RPMI; все компоненты от компании "Invitrogen") в течение 24 часов в влажном инкубаторе с 5% СО2 при 37°С. Несколько аликвот клеток культивировали в отсутствии антитела, и они служили контрольными пробами для определения максимального уровня EGFRvIII на поверхности клетки, и отрицательным контролем для окрашивания одними только вторичными реагентами. После инкубации клетки дважды отмывали ледяным фосфатно-солевым буфером (ФСБ, "Invitrogen"), с добавлением 2% термоинактивированного FCS ("Invitrogen") и 0,1% азида натрия («Roth», Карлсруэ, Германия) (в настоящей заявке называется буфер для FACS), а затем окрашивали насыщающими концентрациями (10 мкг/мл) идентичного антитела, с которым их инкубировали во время модуляции культуры для оценки максимального связывания антител, соответствующего оставшимся антигенам EGFRvIII на поверхности клетки. Аликвоты клеток, которые культивировали в отсутствии антител, окрашивали идентичным антителом при насыщающих концентрациях, чтобы определить максимальную способность связывания антитела, соответствующую максимальному измеримому уровню EGFRvIII на поверхности клетки. После трехкратной отмывки буфером для FACS клетки, которые были обработаны тандемными диателами и окрашены 10 мкг/мл анти-His mAb (Dia910, «Dianova»), затем обрабатывали 15 мкг/мл FITC-конъюгированным антимышиным IgG козы («Dianova»). После повторной отмывки буфером для FACS клетки повторно суспендировали в буфере для FACS, содержащем 2 мкг/мл пропидия иодида ("Sigma»), чтобы исключить мертвые клетки. Из каждой пробы ~5×103 пропидий иодид-негативных клеток анализировали при помощи программы МХР на проточном питометре FC500 MPL («Beckman-Coulter", Крефельд, Германия), и средние интенсивности флуоресценции (MFI) измеряемых проб определяли при помощи программы СХР («Beckman-Coulter"), или клетки анализировали на проточном цитометре Millipore Guava EasyCyte при помощи программы Incyte («Merck Millipore», Швальбах, Германия).
После вычитания сигнала, полученного при окрашивании клеток только вторичными реагентами, средние интенсивности флуоресценции измеряемых проб откладывали на графике-диаграмме при помощи программы GraphPad Prism («GraphPad Software», Сан-Диего, Калифорния), и относительное количество оставшегося на поверхности клетки EGFRvIII рассчитывали при помощи флуоресцентного сигнала, полученного путем окрашивания при 37°С в отсутствии EGFRvIII/CD3-тандемных диател проб необработанных клеток, которые принимали за 100% EGFRvIII на поверхности клетки. Результаты показаны на Фигуре 9.
В нескольких литературных сообщениях описано, что мутант, полученный путем делеции, - EGFRvIII демонстрирует нарушенную интернализацию по сравнению с нативным EGFR (Fenstermaker et al. 2000, Han et al. 2006, Grandal et al. 2007, Gan et al. 2009). Несмотря на это, несколько опубликованных сообщений о EGFRvIII-специфичных антителах или антителах, направленных на EGFR дикого типа, которые также связываются EGFRvIII (например, цетуксимаб), свидетельствуют о быстрой интернализации и деградации EGFRvIII в EGFRvIII-экспрессирующих клетках после связывания соответствующих антител (Reist et al. 1995, Foulon et al. 2000, Kuan et al. 2000, Patel et al. 2007, Jutten et al. 2009, Dreier et al. 2012).
Для анализа интернализации применяли трансфецированные клетки CHOEGFRvIII, которые экспрессируют EGFRvIII. Клетки, экспрессирующие EGFRvIII, и инкубировали их с разными концентрациями EGFRvIII/CD3-тандемных диател при 37°С в течение 24 часов, при условиях, которые, как описано, в контексте других EGFRvIII-связывающих антител вызывают сильную интернализацию. После такой модуляции, во время которой комплексы антитело-рецептор интернализуются или нет, соответствующие антитела применяли в насыщающей концентрации 10 мкг/мл для окрашивания всех оставшихся молекул рецептора EGFRvIII на поверхности клетки (Фигура 9). Результаты, представленные на Фигуре 9, показывают, что при всех исследуемых условиях более 80% и до 140% молекул рецептора EGFRvIII, присутствующих на необработанных клетках, все еще доступны после воздействия на данные клетки EGFRvIII-связывающих тандемных диател (140% или в более общем виде >100% означает, что клетки, обработанные тандемными диателами, в отличие от (за счет интернализации) уменьшения количества рецепторов на поверхности клетки, демонстрируют увеличение количества рецепторов на поверхности клетки. Нормировка с приведением к 100% получается из сравнения с клетками, которые не были обработаны.)
Данные результаты показывают, что EGFRvIII/CD3-тандемные диатела согласно настоящему изобретению неожиданно не демонстрируют никакой тенденции к интернализации (Фигура 9). Вместо индукции интернализации связывание EGFRvIII-связывающих доменов согласно настоящему изобретению может даже подавлять интернализацию и приводить к увеличению уровня рецептора EGFRvIII на поверхности клетки. Результаты, представленные на Фигуре 9, показывают, что относительно необработанных клеток по меньшей мере более чем 80% все еще остается на поверхности клетки после обработки клеток EGFRvIII-связывающими антителами, при всех исследуемых условиях. Указанные результаты указывают на то, что EGFRvIII/CD3-антитела согласно настоящему изобретению, в частности EGFRvIII/CD3-тандемные диатела, неожиданно не демонстрируют тенденции к интернализации (Фигура 9). Вместо индукции интернализации связывание EGFRvIII-связывающих доменов согласно настоящему изобретению может даже подавлять интернализацию или облегчать экспрессию EGFRvIII, что приводит к увеличению его плотности на поверхности клетки.
ПРИМЕР 10: Оценка пролиферации клеток в культурах МКПК в присутствии EGFRvIII/CD3 (или EGFRvIII/CD16A)-тандемных диател
Чтобы оценить, индуцируют ли EGFRvIII/CD3 (или EGFRvIII/CD16A)-тандемные диатела активацию Т-лимфоцитов и последующую пролиферацию в отсутствии EGFRvIII+-клеток-мишеней, МКПК человека культивировали в присутствии возрастающих концентраций EGFRvIII/CD3-тандемных диател. Пролиферацию оценивали в анализе включение BrdU после 5-дневной инкубации.
Материалы (среда, буфер и реагенты)
FCS («Invitrogen»), сыворотка человека («Sigma»), Lymphoprep («Stemcell Technologies»), L-глутамин («Invitrogen»), OKT3 (Biolegend), ФСБ («Invitrogen»), бета-меркаптоэтанол («Invitrogen»), пенициллин/стрептомицин («Invitrogen»), RPMI-1640 («Invitrogen»), пируват натрия («Invitrogen»), ELISA на пролиферацию клеток с BrdU («Roche»).
Клетки
Лейкоцитарную пленку приобретали у Deutsches Rotes Kreuz, DRK-Blutspendedienst (Мангейм, Германия) в день отбора крови; препараты лейкоцитарной пленки хранили в течение ночи при комнатной температуре перед тем как выделить МКПК.
Выделение МКПК из лейкоцитарных пленок и обогащение Т-лимфоцитов
МКПК выделяли из лейкоцитарной пленки посредством центрифугирования в градиенте плотности. Пробу лейкоцитарной пленки разводили двух-трехкратным объемом ФСБ, наслаивали на основание из Lymphoprep и центрифугировали при 800×g в течение 25 мин при комнатной температуре без разбивания комков. МКПК, расположенные на поверхности, отбирали и промывали трехкратно ФСБ перед тем, как их применяли для обогащения Т-лимфоцитов или проточно-цитометрического анализа.
Анализ пролиферации
Все планшеты для анализа блокировали средой RPMI 1640 с добавлением 10% термоинактивированной сыворотки человека, 4 мМ L-глутамина, 100 Е/мл натриевой соли пенициллина G, 100 мкг/мл стрептомицина сульфата, 1 мМ пирувата натрия (в настоящей заявке называется полной средой RPMI) в течение 2 часов при КТ для предотвращения неспецифического связывания антител с пластиковой поверхностью. После удаления блокирующей среды 4×105 МКПК засевали в полную среду RPMI в отдельные лунки микропланшетов на 96 лунок с плоским дном вместе с указанными концентрациями исследуемых и контрольных компонентов в общем объеме 200 мкл/лунку в трех повторах. В качестве положительного контроля применяли IgG анти-CD3 клон ОКТ3. Чтобы оценить спонтанную пролиферацию, клетки культивировали в отсутствии антител в 6 повторах. Затем планшеты инкубировали в течение 4 дней при 37°С и 5% CO2 в увлажненной атмосфере. За 18 часов до конца инкубации в культуры клеток импульсно вводили 100 мкМ BrdU. Включенный BrdU количественно оценивали при помощи набора BrdU Proliferation ELISA в соответствии с инструкциями производителя. После остановки развития окраски путем добавления 100 мМ H2SO4 поглощение измеряли на длине волны 450 нм при помощи планшетного ридера на много лунок (Victor 3, «Perkin Elmer»). Значения поглощения анализировали, и средние значения вместе с СО (стандартным отклонением) откладывали на графике при помощи программы GraphPad Prism (GraphPad Prism версия 6.00 для Windows, программа GraphPad, Ла-Хойя, Калифорния, США).
Результаты показаны на Фигуре 10.
ПРИМЕР 11: Корреляция аффинности связывания CD3 и цитотоксической активности EGFRvIII/CD3-тандемных диател
EGFRvIII/CD3-тандемные диатела, содержащие одинаковый EGFRvIII-связывающий домен (Li3G30) в сочетании с разными CD3-связывающими доменами, с определенным спектром сродства к антигену CD3 Т-лимфоцитов конструировали, экспрессировали и определяли их связывание с клетками CD3+ Jurkat и EGFRvIII+ CHOEGFRvIII; кроме того, проводили анализ цитотксичности с применением клеток CHOEGFRvIII в качестве клеток-мишеней и МКПК в качестве эффекторных клеток. Все исследуемые тандемные диатела в данном примере обладают одинаковым порядком расположения доменов VH и VL (тяжелая и легкая цепь): VLCD3-VHEGFRvIII-VLEGFRvIII-VHCD3.
Проточно-цитометрическую оценку и связывания антител с клетками EGFRvIII+ CHOEGFRvIII, и цитотоксической активности, опосредуемой EGFRvIII/CD3-тандемными диателами, проводили, как было описано в предыдущих примерах.
Кажущееся сродство EGFRvIII/CD3-тандемных диател к CD3+-клеткам определяли посредством проточной цитометрии после окрашивания клеток CD3+ Jurkat возрастающими концентрациями тандемных диател. Значения KD рассчитывали на основании нелинейной регрессии/гиперболы с применением средних значений флуоресценции, определенных посредством проточной цитометрии для отдельных концентраций тандемных диател.
Материалы (среда, буфер и реагенты)
Анти-His IgG 13/45/31 («Dianova»), FCS (фетальная телячья сыворотка) («Invitrogen»), FITC-конъюгированный антимышиный IgG козы, мин X («Dianova»), L-глутамин («Invitrogen»), NaN3 («Carl Roth»), ФСБ («Invitrogen»), пенициллин/стрептомицин («Invitrogen»), пропидиум иодид («Sigma»), RPMI-1640 («Invitrogen»).
Клетки
Клетки Jurkat (любезно предоставленные д-ром G. Moldenhauer, DKFZ, Гейдельберг, Германия) культивировали в среде RPMI 1640 с добавлением 2 мМ L-глутамина и 100 МЕ/мл натриевой соли пенициллина G, и 100 мкг/мл стрептомицина сульфата (все компоненты от компании "Invitrogen», в настоящей заявке называется средой RPMI 1640).
Определение сродства антител к CD3+-клеткам посредством проточной цитометрии: Клетки инкубировали с 100 мкл последовательных разведений указанных антител, начиная со 100 мкг/мл (~1000 нМ) в ледяном ФСБ с добавлением 2% термоинактивированной FCS и 0,1% азида натрия (называется буфером для FACS) в течение 45 мин на льду. После трехкратной отмывки буфером для FACS клетки инкубировали с 0,1 мл 10 мкг/мл моноклонального анти-His антитела мыши клона 13/45/31 в том же буфере в течение 45 мин на льду. После второго цикла отмывки клетки инкубировали с 0,1 мл 15 мкг/мл FITC-конъюгированного антимышиного антитела IgG козы при тех же условиях, что и ранее. В качестве контроля клетки инкубировали с анти-His IgG13/45/31, а затем с FITC-конъюгированными антимышиными антителами IgG козы без первичного антитела. Затем клетки снова отмывали и повторно суспендировали в 0,2 мл буфера для FACS, содержащего 2 мкг/мл пропидия иодида, чтобы исключить погибшие клетки. Флуоресценцию 1×104 живых клеток измеряли при помощи проточного цитометра Beckman-Coulter FC500 MPL с применением программы МХР («Beckman-Coulter", Крефельд, Германия) или проточного цитометра Millipore Guava EasyCyte с применением программы Incyte («Merck Millipore», Швальбах, Германия). Средние интенсивности флуоресценции проб клеток рассчитывали при помощи программы СХР («Beckman-Coulter", Крефельд, Германия) или Incyte («Merck Millipore», Швальбах, Германия). В случае анализа при помощи проточного цитометра Beckman-Coulter FC500 MPL применяли 0,5×106 клеток на окрашивание, а в случае применения проточного цитометра Millipore Guava EasyCyte - 0,25×106 клеток/окрашивание.
После вычитания значений интенсивности флуоресценции клеток, окрашенных только вторичными и третичными реагентами, рассчитывали значения KD по формуле для связывания в одном сайте (гипербола) в программе GraphPad Prism (GraphPad Prism версия 6.00 для Windows, программа GraphPad, Ла-Хойя, Калифорния, США). Значения KD для связывания EGFRvIII и ЕС50 для цитотоксичности откладывали на графике как функция значения KD для связывания CD3 для каждого из анализируемых тандемных диател. Хотя тандемные диатела показывают только небольшую вариабельность значений KD для связывания EGFRvIII, значения ЕС50 для цитотоксичности показывают почти линейное увеличение при росте значений KD для связывания CD3 указанных тандемных диател (Фигура 11).
ПРИМЕР 12: Подавление ксенотрансплантата опухоли F98EGFRvIII EGFRvIII/CD3-тандемными диателами (исследования обоснованности концепции in vivo)
Девяти группам мышей NOD/scid с иммунодефицитом (NOD/MrkBomTac-Prkdcscid, Taconic, Дания) прививали ксенотрнасплантат путем подкожной (п/к) инъекции суспензии 4×106 клеток F98npEGFRvIII, смешанных с 1×107 очищенных МКПК человека от здоровых доноров в день 0 (д0). Чтобы учесть потенциальную вариабельность МКПК доноров, каждую экспериментальную группу разделяли на две когорты, получавших МКПК только от одного отдельного донора. Животным вводили препараты в хвостовую вену (в/в) через 2 ч после прививки клеток опухоли и в дальнейшем через 24 ч, 48 ч, 72 ч и 96 ч со 100 мкг, 10 мкг или 1 мкг соответствующего исследуемого тандемного диатела. Контрольная группа получала только основу. Дополнительным контролем были животные, которым в/б два раза в неделю вводили 1 мг цетуксимаба (Эрбитукса), начиная с д0. В таблице 6 обобщены группы доз и распределение доноров. Объем опухоли отслеживали 3 раза в неделю с 3-го дня по 52-й день путем измерения большого диаметра и малого диаметра по следующей формуле: Объем = (малый диаметр)2 × большой диаметр × 0,5.
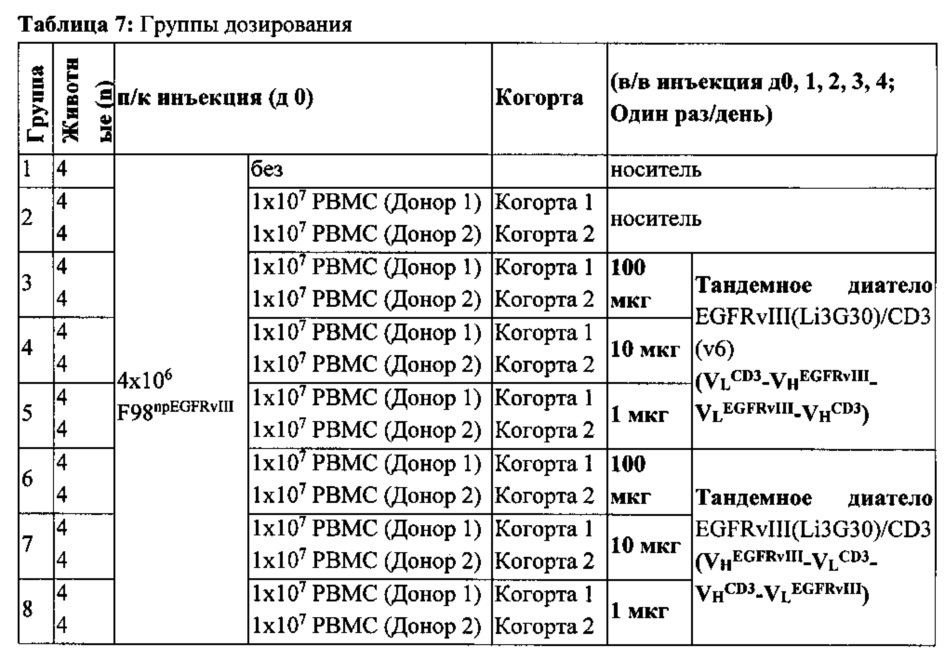
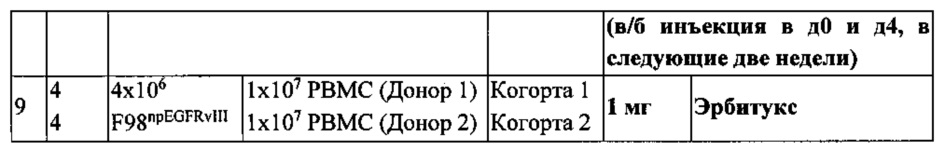
Ссылки:
Schwarz М,  Р, Takada Y, Le Gall F, Knackmuss S, Bassler N,
Р, Takada Y, Le Gall F, Knackmuss S, Bassler N,  C, Little M, Bode C, Peter K. Single-chain antibodies for the conformation-specific blockade of activated platelet integrin alphaIIbbeta3 designed by subtractive selection from naive human phage libraries. FASEB J. 2004 Nov; 18(14):1704-6.
C, Little M, Bode C, Peter K. Single-chain antibodies for the conformation-specific blockade of activated platelet integrin alphaIIbbeta3 designed by subtractive selection from naive human phage libraries. FASEB J. 2004 Nov; 18(14):1704-6.
Schier R, McCall A, Adams GP, Marshall KW, Merritt H, Yim M, Crawford RS, Weiner LM, Marks C, Marks JD. Isolation of picomolar affinity anti-c-erbB-2 single-chain Fv by molecular evolution of the complementarity determining regions in the center of the antibody binding site. J Mol Biol. 1996
Pini A, Viti F, Santucci A, Carnemolla B, Zardi L, Neri P, Neri D. Design and use of a phage display library. Human antibodies with subnanomolar affinity against a marker of angiogenesis eluted from a two-dimensional gel. J Biol Chem. 1998
Boder ET, Midelfort KS, Wittrup KD. Directed evolution of antibody fragments with monovalent femtomolar antigen-binding affinity. Proc Natl Acad Sci USA. 2000 Sep 26; 97(20):10701-5.
Barbas CF 3rd, Hu D, Dunlop N, Sawyer L, Cababa D, Hendry RM, Nara PL, Burton DR. In vitro evolution of a neutralizing human antibody to human immunodeficiency virus type 1 to enhance affinity and broaden strain cross-reactivity. Proc Natl Acad Sci USA. 1994 Apr 26; 91(9):3809-13.
Parsons HL, Earnshaw JC, Wilton J, Johnson KS, Schueler PA, Mahoney W, McCafferty J. Directing phage selections towards specific epitopes. Protein Eng. 1996 Nov; 9(11): 1043-9.
Winkler K, Kramer A,  G, Seifert M, Scholz C, Wessner H, Schneider-Mergener J,
G, Seifert M, Scholz C, Wessner H, Schneider-Mergener J,  W. Changing the antigen binding specificity by single point mutations of an anti-p24 (HIV-1) antibody. J Immunol. 2000 Oct 15; 165(8):4505-14.
W. Changing the antigen binding specificity by single point mutations of an anti-p24 (HIV-1) antibody. J Immunol. 2000 Oct 15; 165(8):4505-14.
Safdari Y, Farajnia S, Asgharzadeh M, Omidfar K, Khalili M. humMRl, a highly specific humanized single chain antibody for targeting EGFRvIII. Int Immunopharmacol. 2014 Feb; 18(2):304-10.
Lorimer IA, Keppler-Hafkemeyer A, Beers RA, Pegram CN, Bigner DD, Pastan I. Recombinant immunotoxins specific for a mutant epidermal growth factor receptor: targeting with a single chain antibody variable domain isolated by phage display. Proc Natl Acad Sci USA. 1996 Dec 10; 93(25): 14815-20.
Beers R, Chowdhury P, Bigner D, Pastan I. Immunotoxins with increased activity against epidermal growth factor receptor vIII-expressing cells produced by antibody phage display. Clin Cancer Res. 2000 Jul; 6(7):2835-43.
Kuan CT, Wikstrand CJ, Archer G, Beers R, Pastan I, Zalutsky MR, Bigner DD. Increased binding affinity enhances targeting of glioma xenografts by EGFRvIII-specific scFv. Int J Cancer. 2000 Dec 15; 88(6):962-9.
Choi BD, Kuan CT, Cai M, Archer GE, Mitchell DA, Gedeon PC, Sanchez-Perez L, Pastan I, Bigner DD, Sampson JH. Systemic administration of a bispecific antibody targeting EGFRvIII successfully treats intracerebral glioma. Proc Natl Acad Sci USA. 2013 Jan 2; 110(1):270-5. Fenstermaker RA, Ciesielski MJ. Deletion and tandem duplication of exons 2-7 in the epidermal growth factor receptor gene of a human malignant glioma. Oncogene. 2000 Sep 14; 19(39):4542-8.
Han W, Zhang T, Yu H, Foulke JG, Tang CK. Hypophosphorylation of residue Y1045 leads to defective downregulation of EGFRvIII. Cancer Biol Ther. 2006 Oct; 5(10): 1361-8.
Grandal MV, Zandi R, Pedersen MW, Willumsen BM, van Deurs B, Poulsen HS. EGFRvIII escapes down-regulation due to impaired internalization and sorting to lysosomes. Carcinogenesis. 2007 Jul; 28(7): 1408-17.
Gan HK, Kaye AH, Luwor RB. The EGFRvIII variant in glioblastoma multiforme. J Clin Neurosci. 2009 Jun; 16(6):748-54.
Reist CJ, Archer GE, Kurpad SN, Wikstrand CJ, Vaidyanathan G, Willingham MC, Moscatello DK, Wong AJ, Bigner DD, Zalutsky MR. Tumor-specific anti-epidermal growth factor receptor variant III monoclonal antibodies: use of the tyramine-cellobiose radioiodination method enhances cellular retention and uptake in tumor xenografts. Cancer Res. 1995 Oct 1; 55(19):4375-82.
Foulon CF, Reist CJ, Bigner DD, Zalutsky MR. Radioiodination via D-amino acid peptide enhances cellular retention and tumor xenograft targeting of an internalizing anti-epidermal growth factor receptor variant III monoclonal antibody. Cancer Res. 2000 Aug 15; 60(16):4453-60.
Kuan CT, Wikstrand CJ, Archer G, Beers R, Pastan I, Zalutsky MR, Bigner DD. Increased binding affinity enhances targeting of glioma xenografts by EGFRvIII-specific scFv. Int J Cancer. 2000 Dec 15; 88(6):962-9.
Patel D, Lahiji A, Patel S, Franklin M, Jimenez X, Hicklin DJ, Kang X. Monoclonal antibody cetuximab binds to and down-regulates constitutively activated epidermal growth factor receptor vIII on the cell surface. Anticancer Res. 2007 Sep-Oct; 27(5A):3355-66.
Jutten B, Dubois L, Li Y, Aerts H, Wouters BG, Lambin Ρ, Theys J, Lammering G. Binding of cetuximab to the EGFRvIII deletion mutant and its biological consequences in malignant glioma cells. Radiother Oncol. 2009 Sep; 92(3):393-8.
Dreier A, Barth S, Goswami A, Weis J. Cetuximab induces mitochondrial translocalization of EGFRvIII, but not EGFR: involvement of mitochondria in tumor drug resistance? Tumour Biol. 2012 Feb; 33(1):85-94.

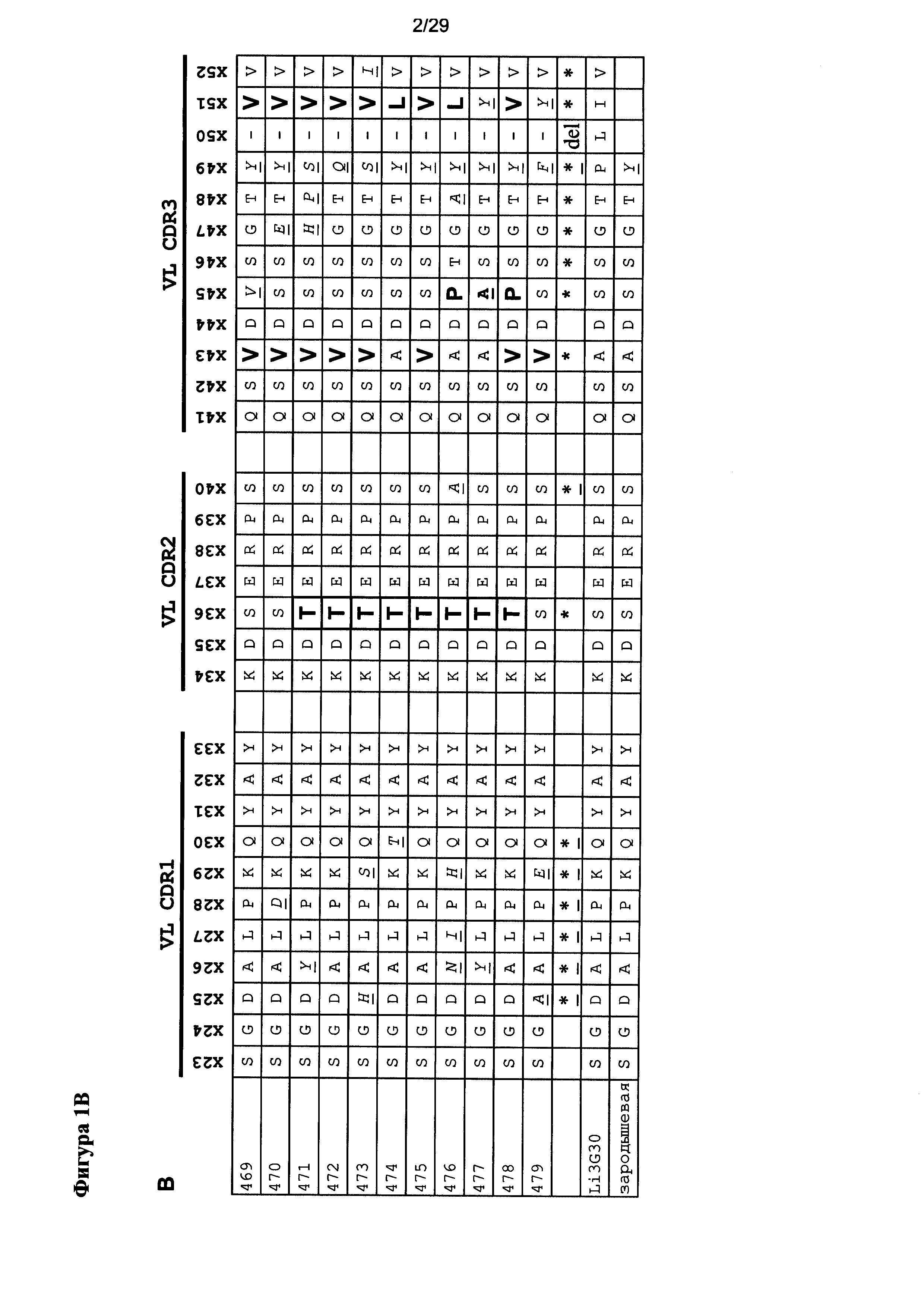
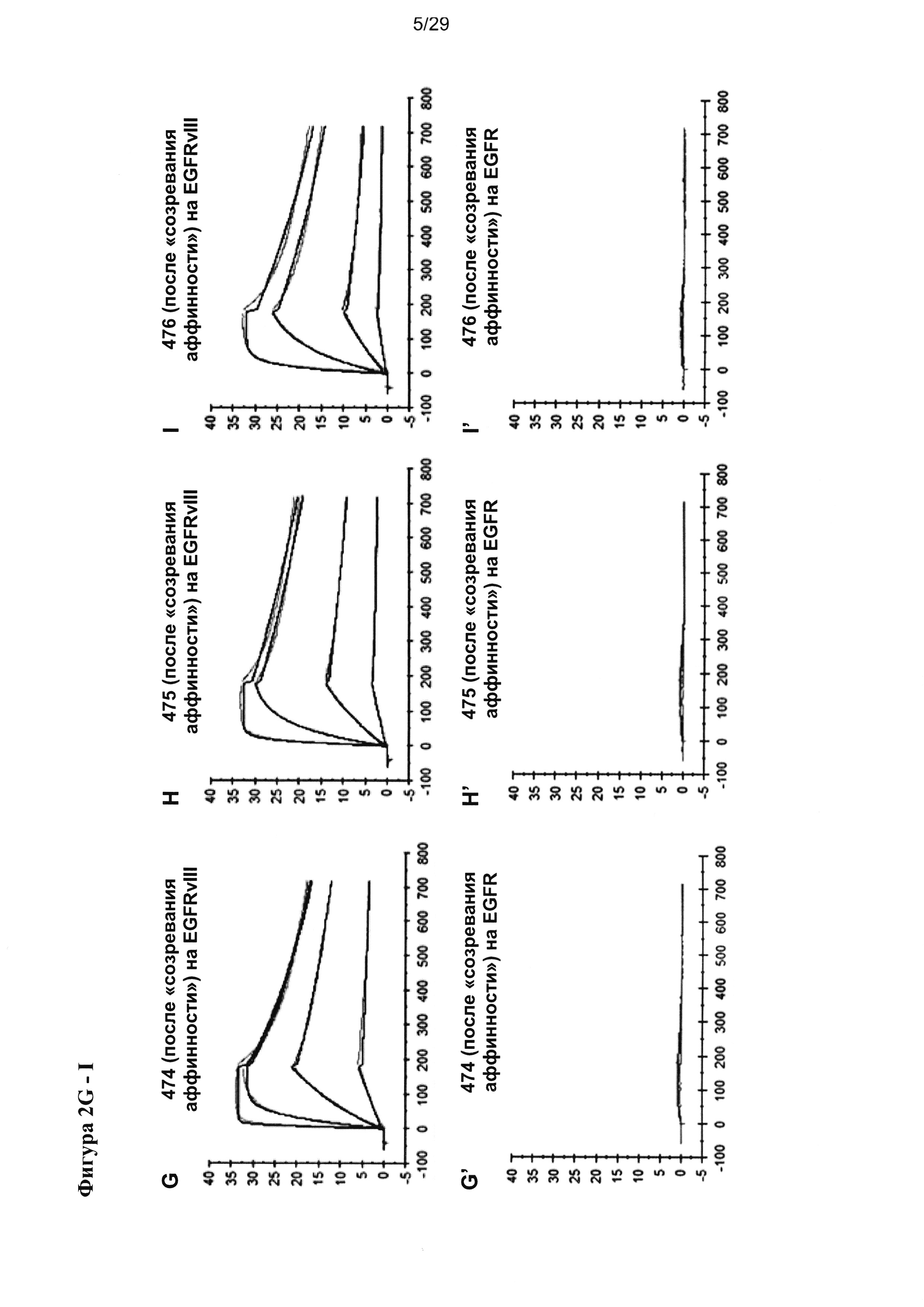
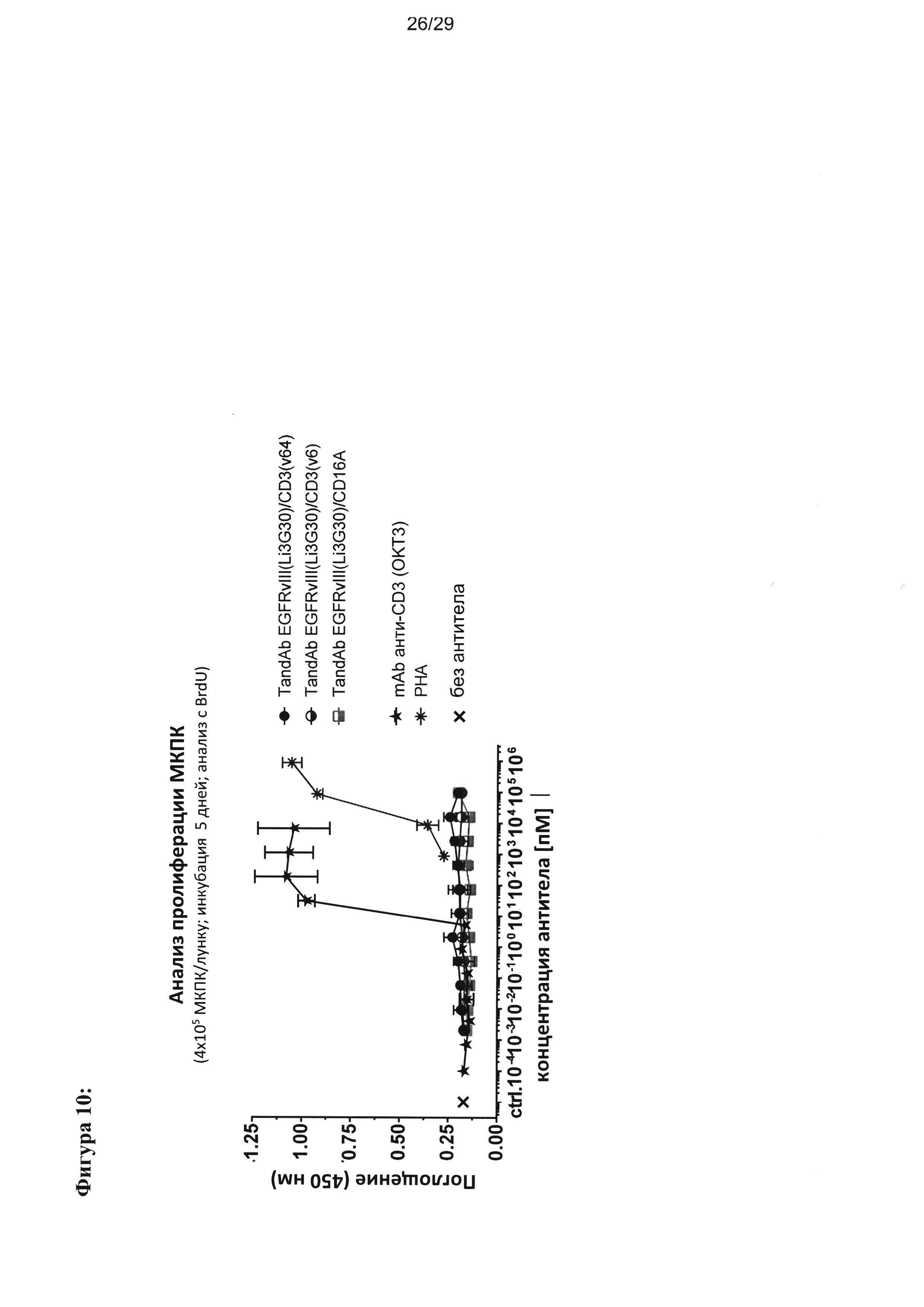
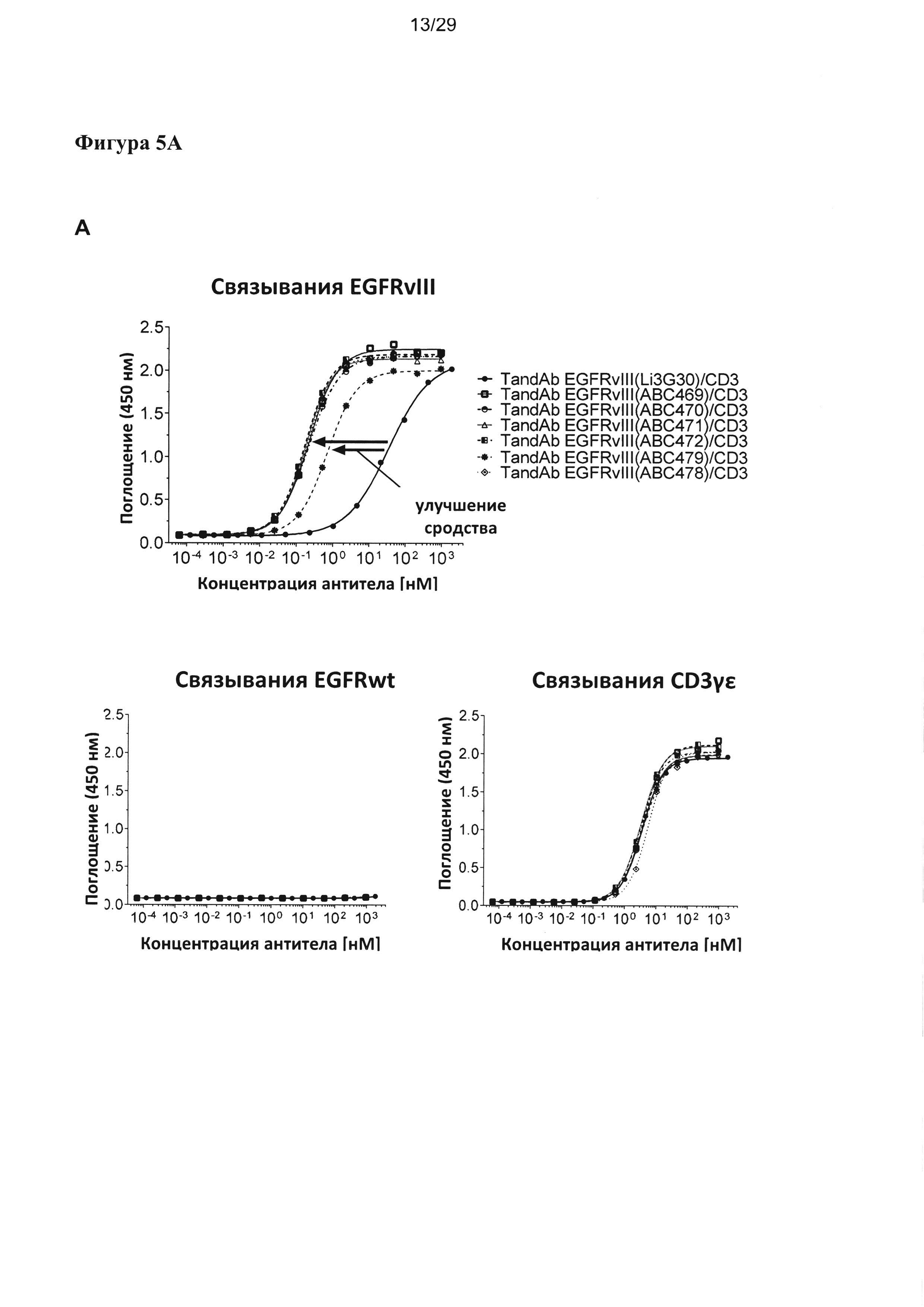
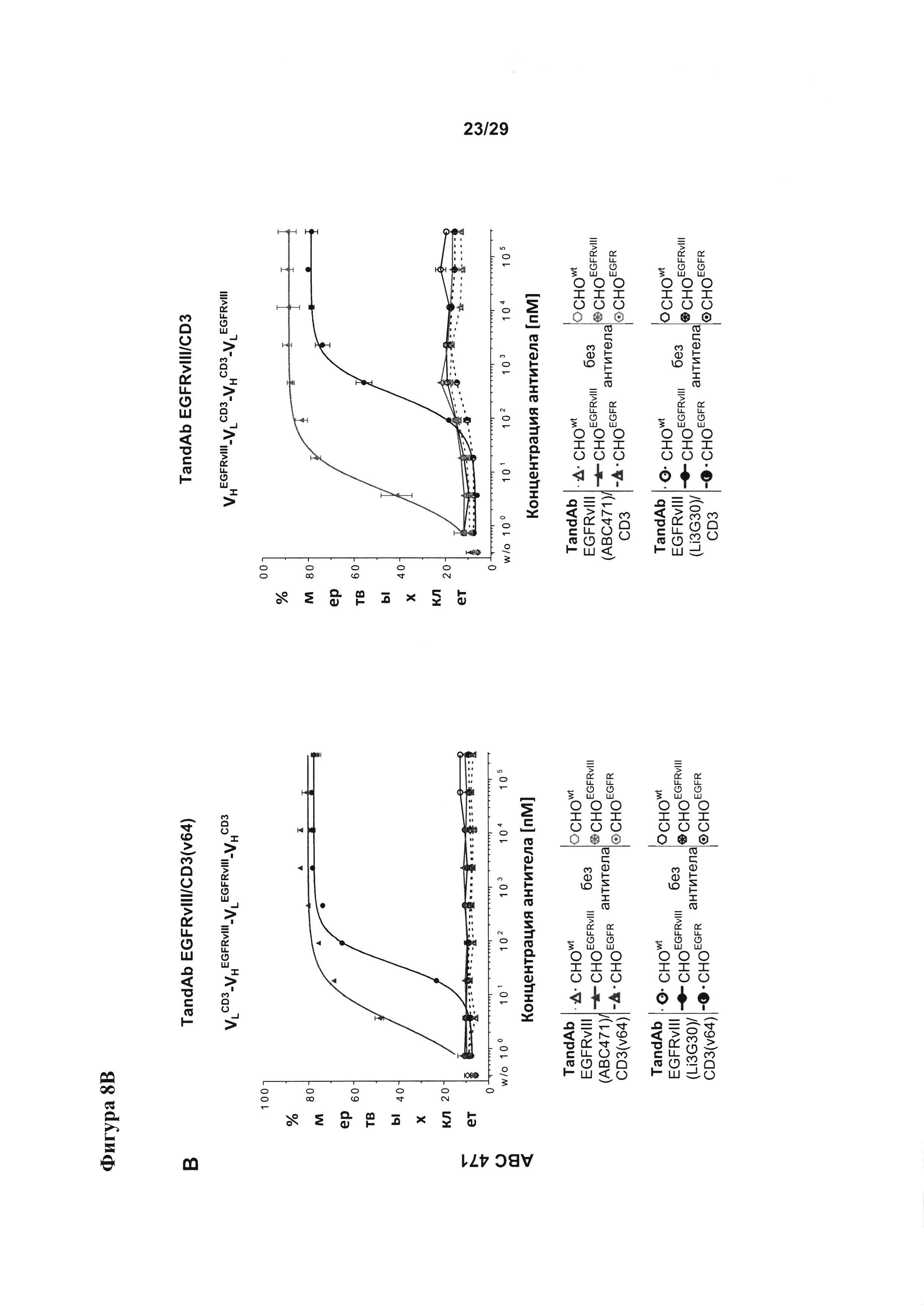
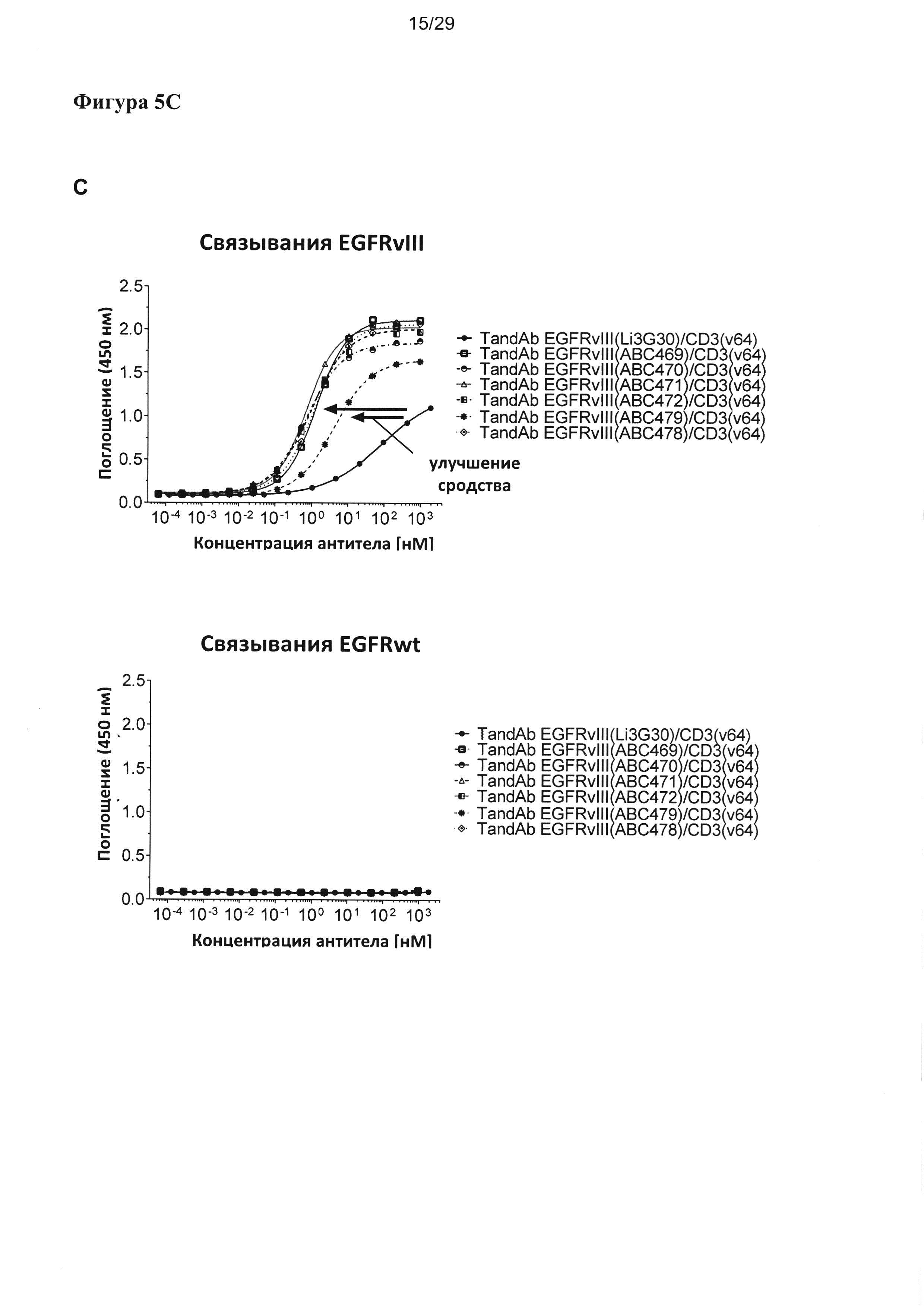
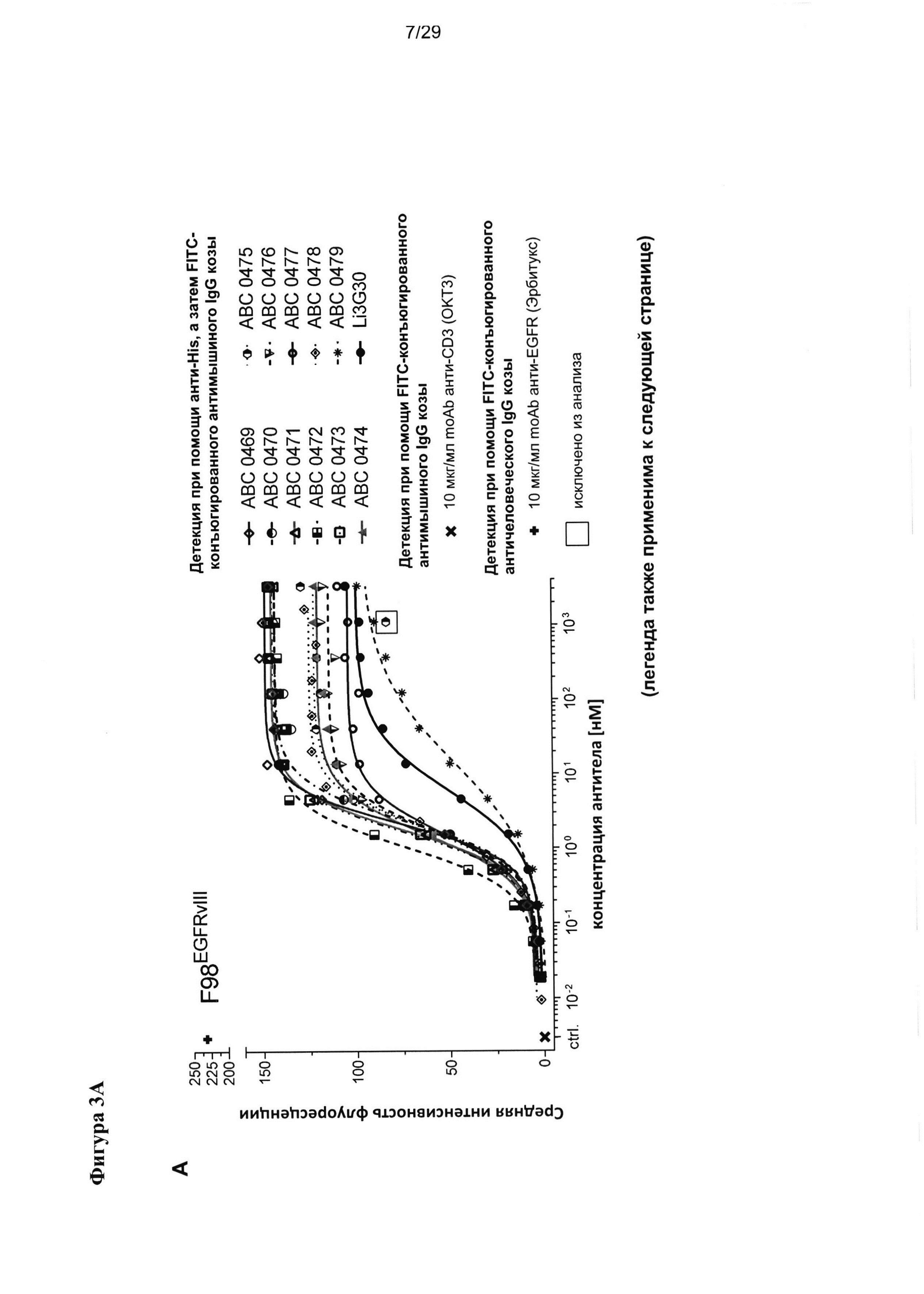
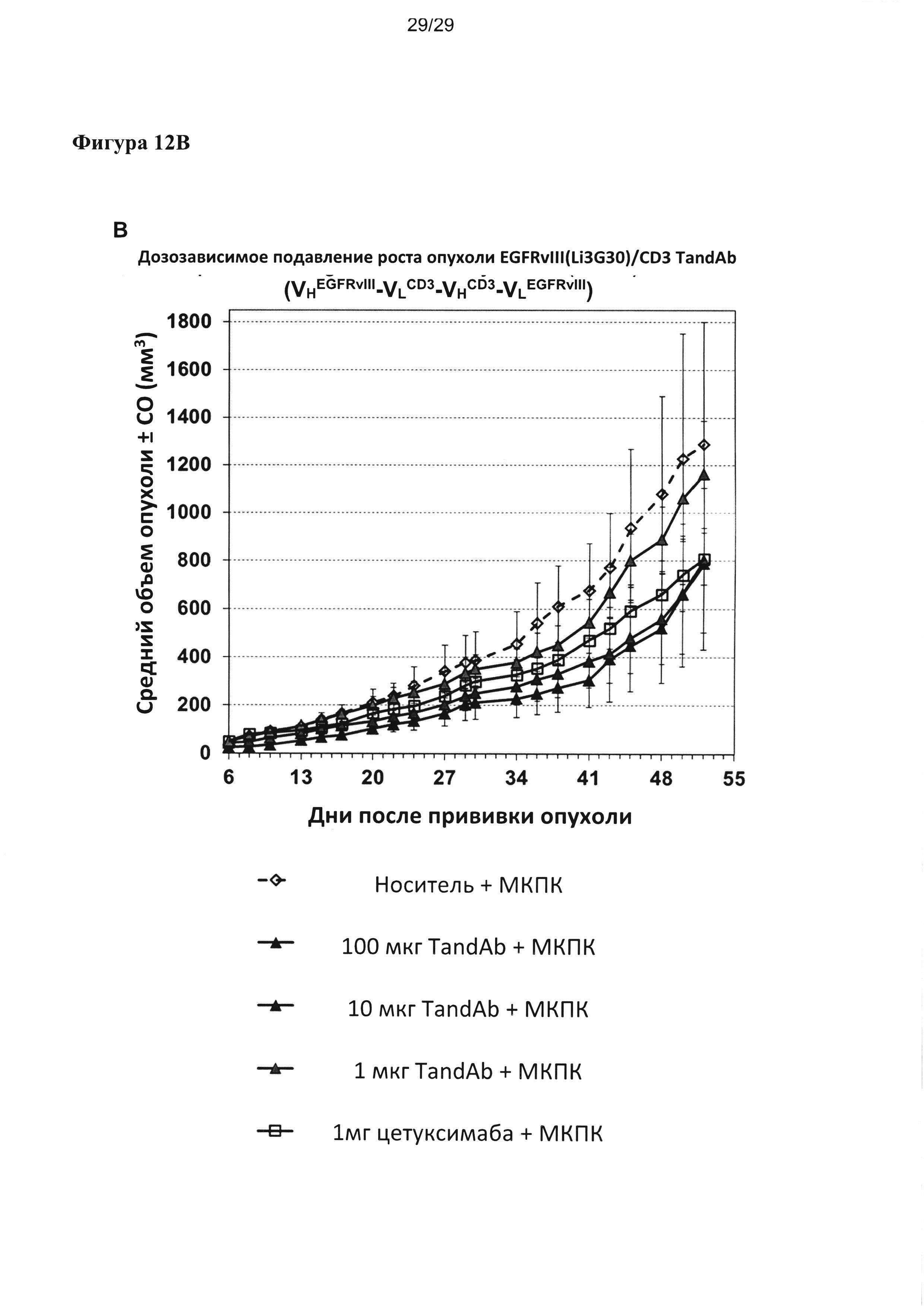

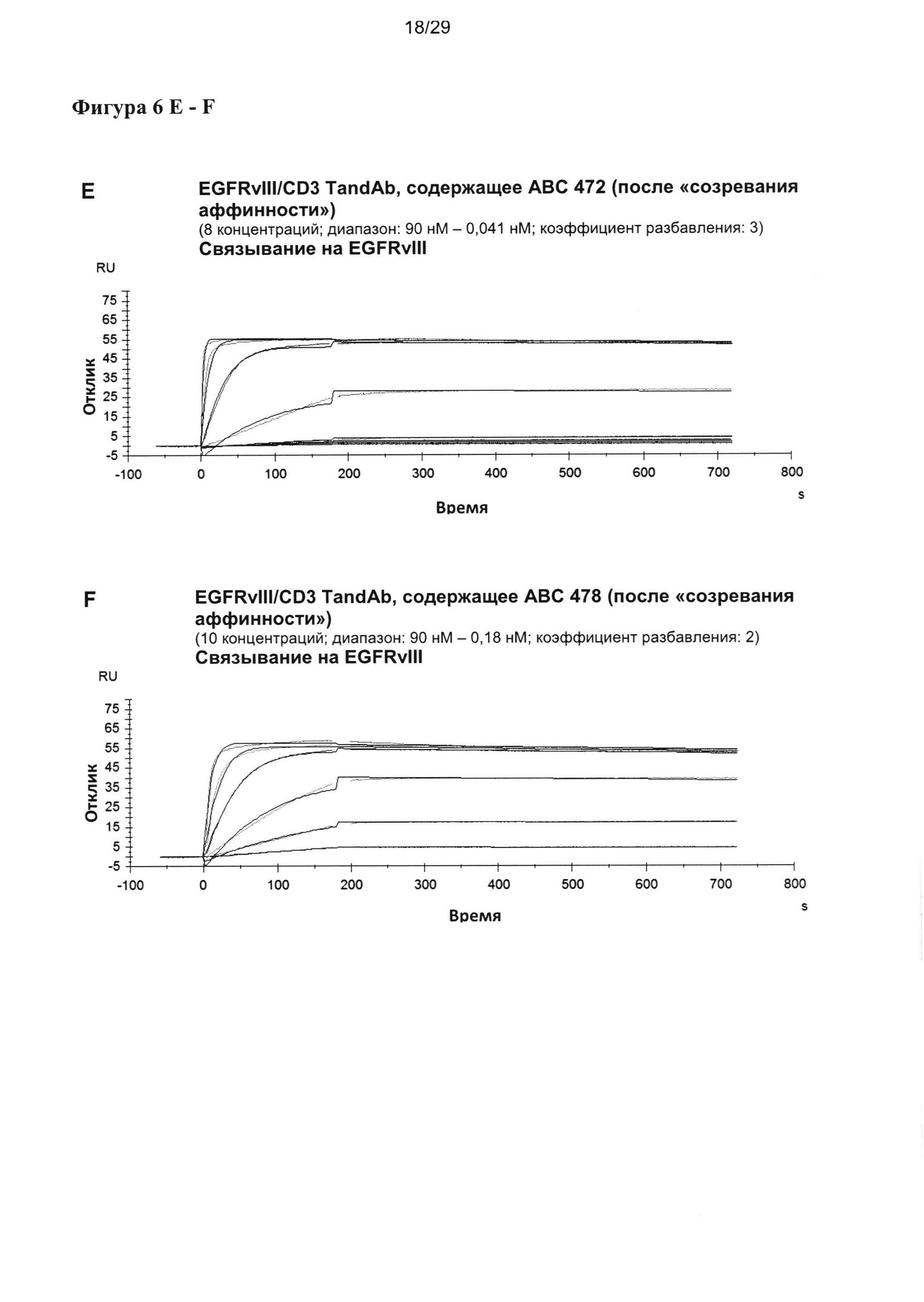

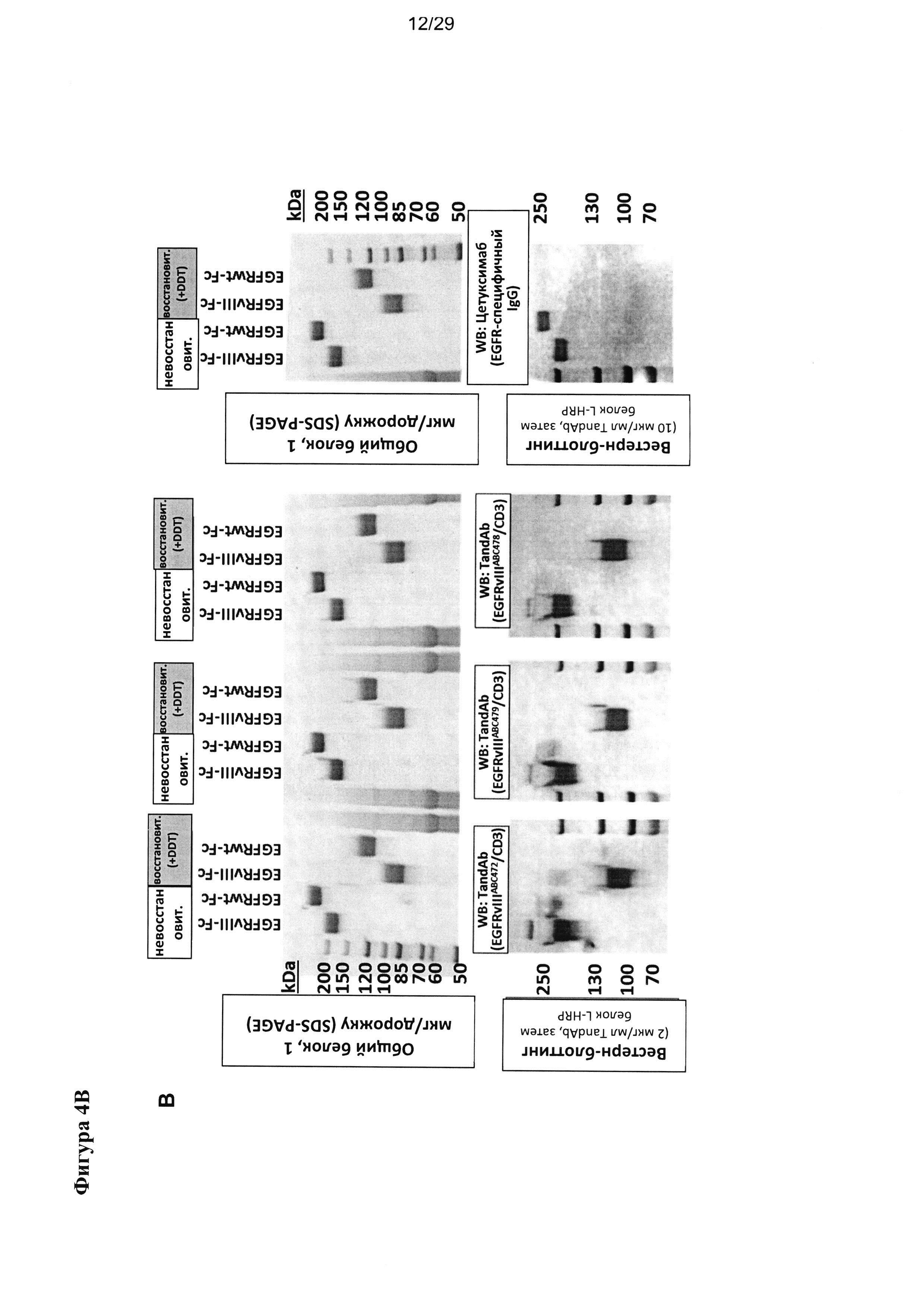
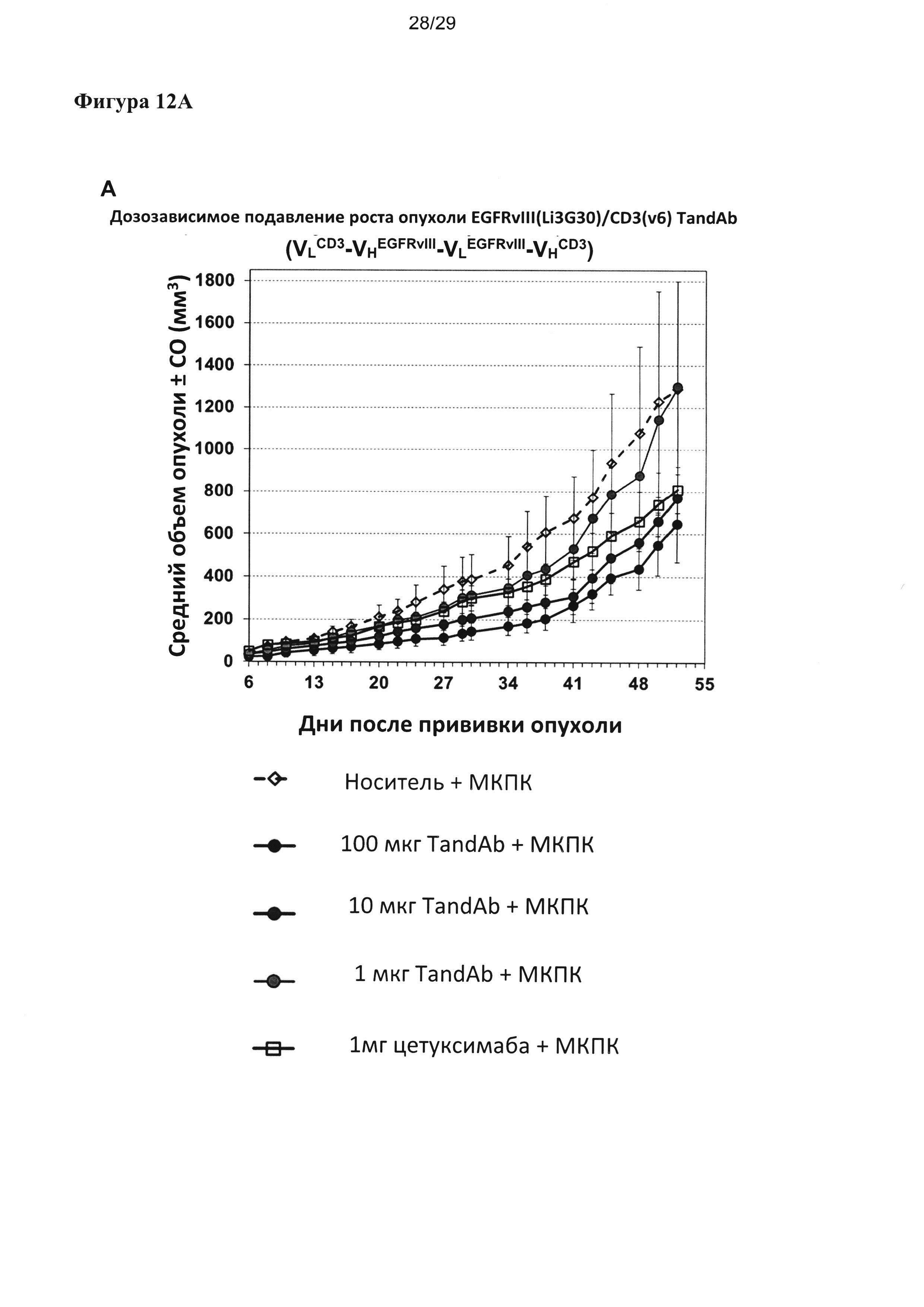


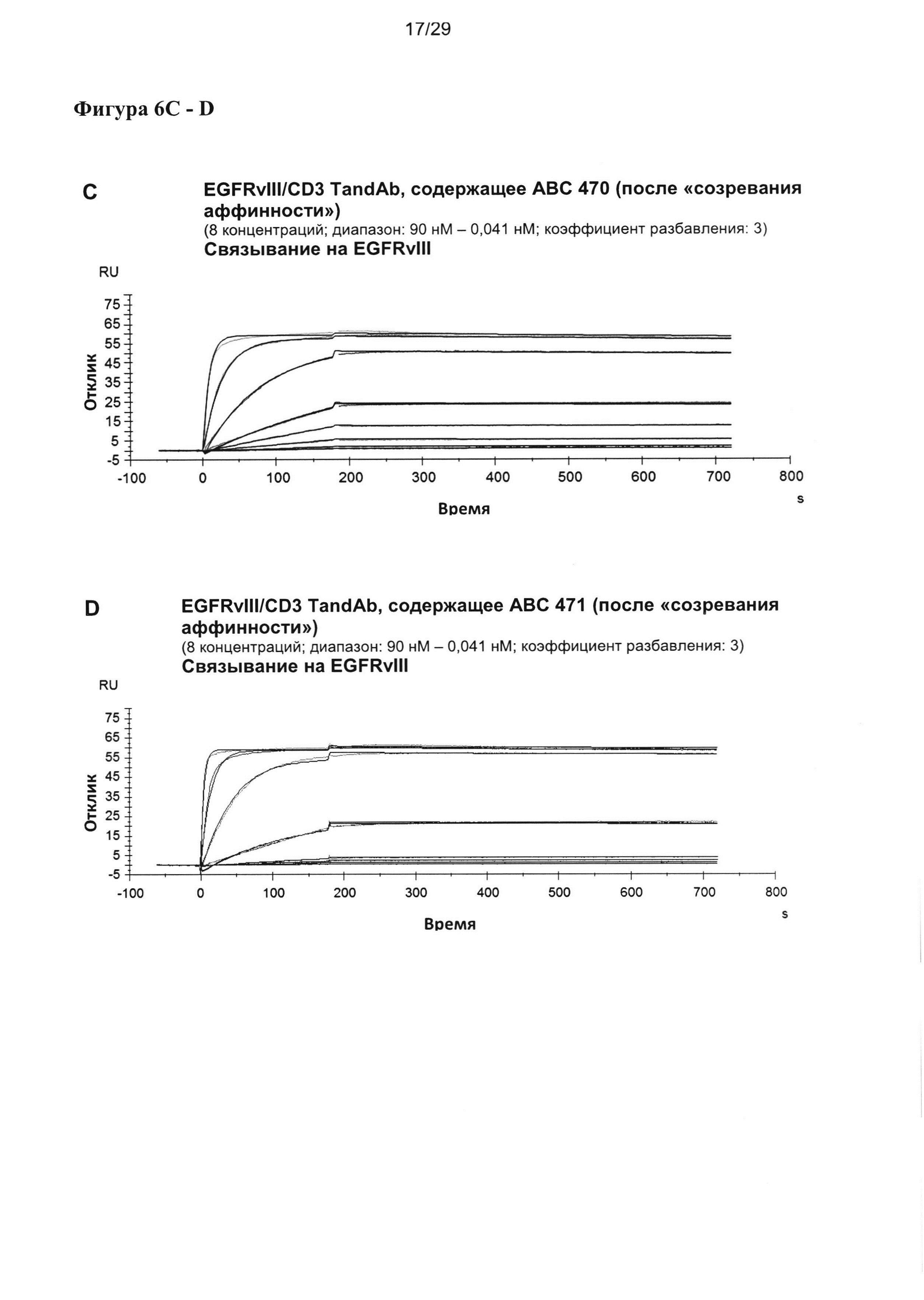


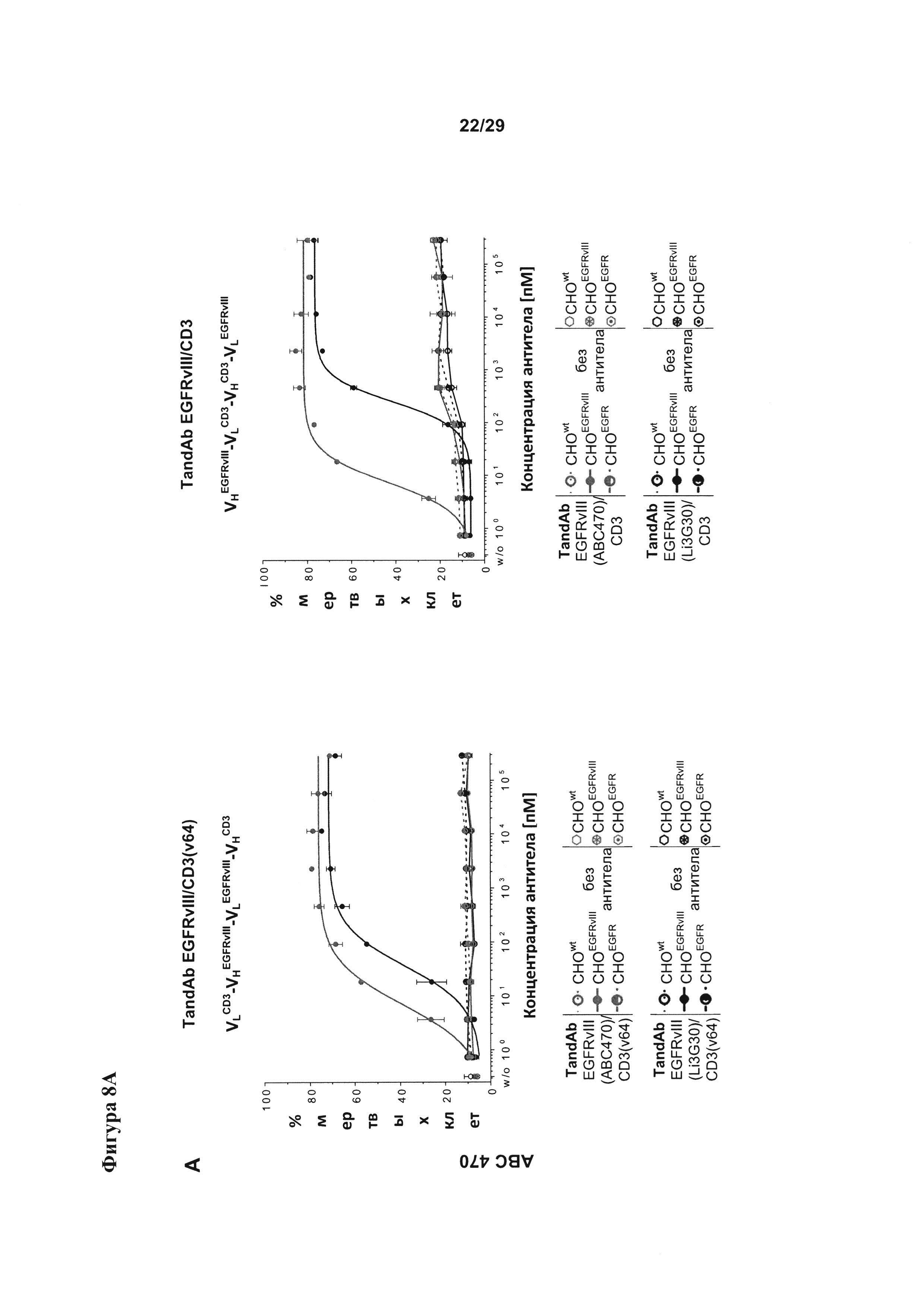


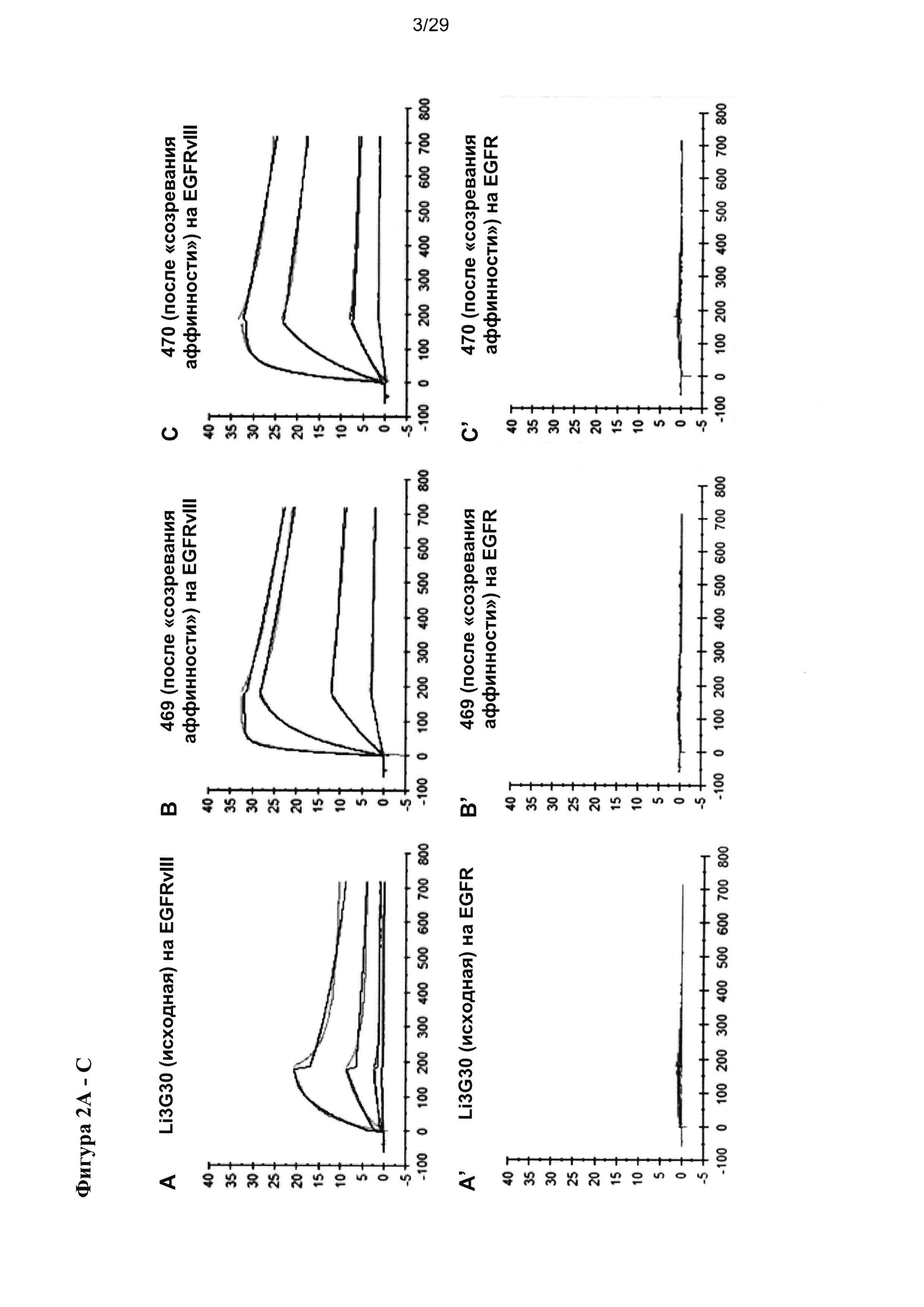

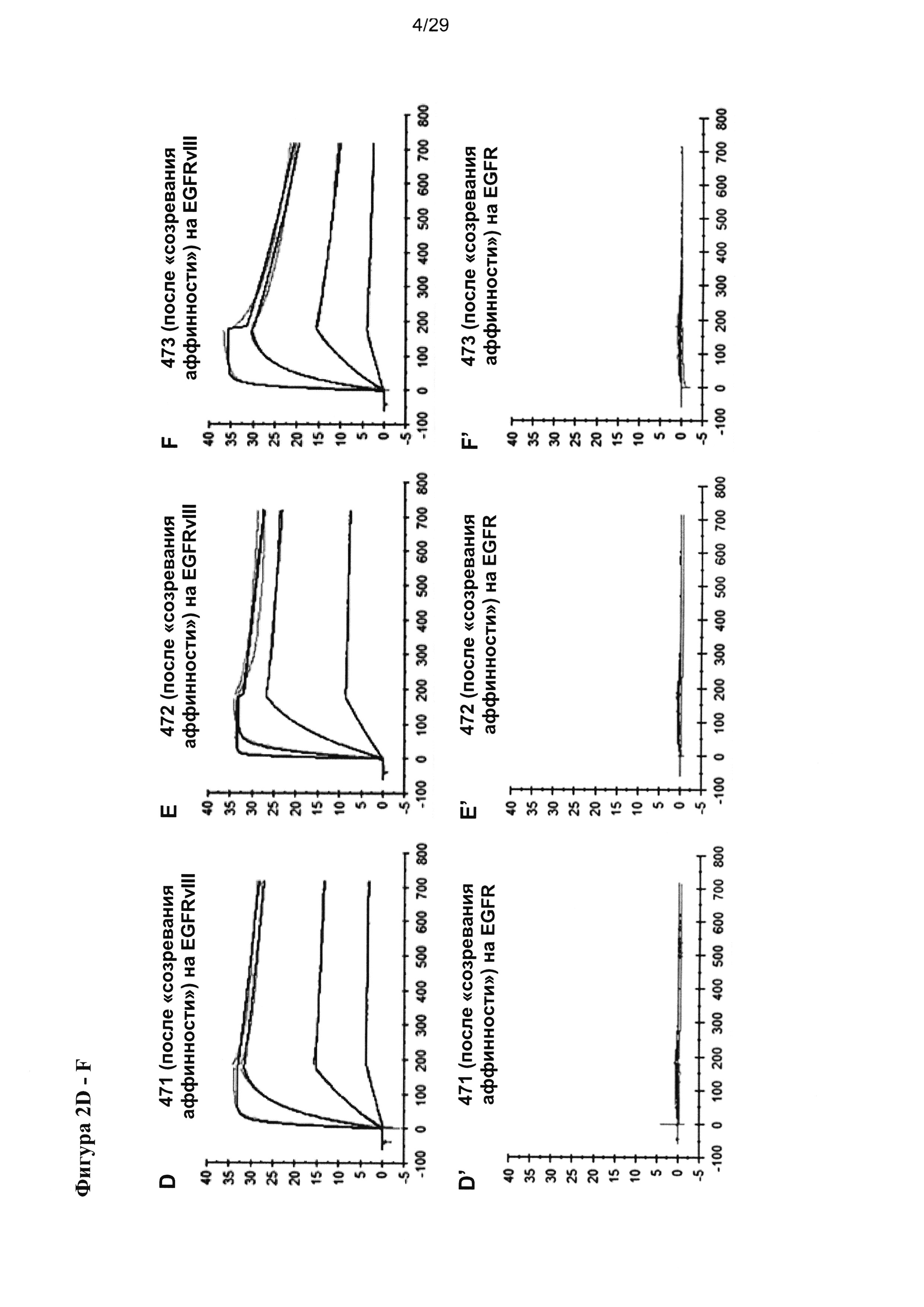


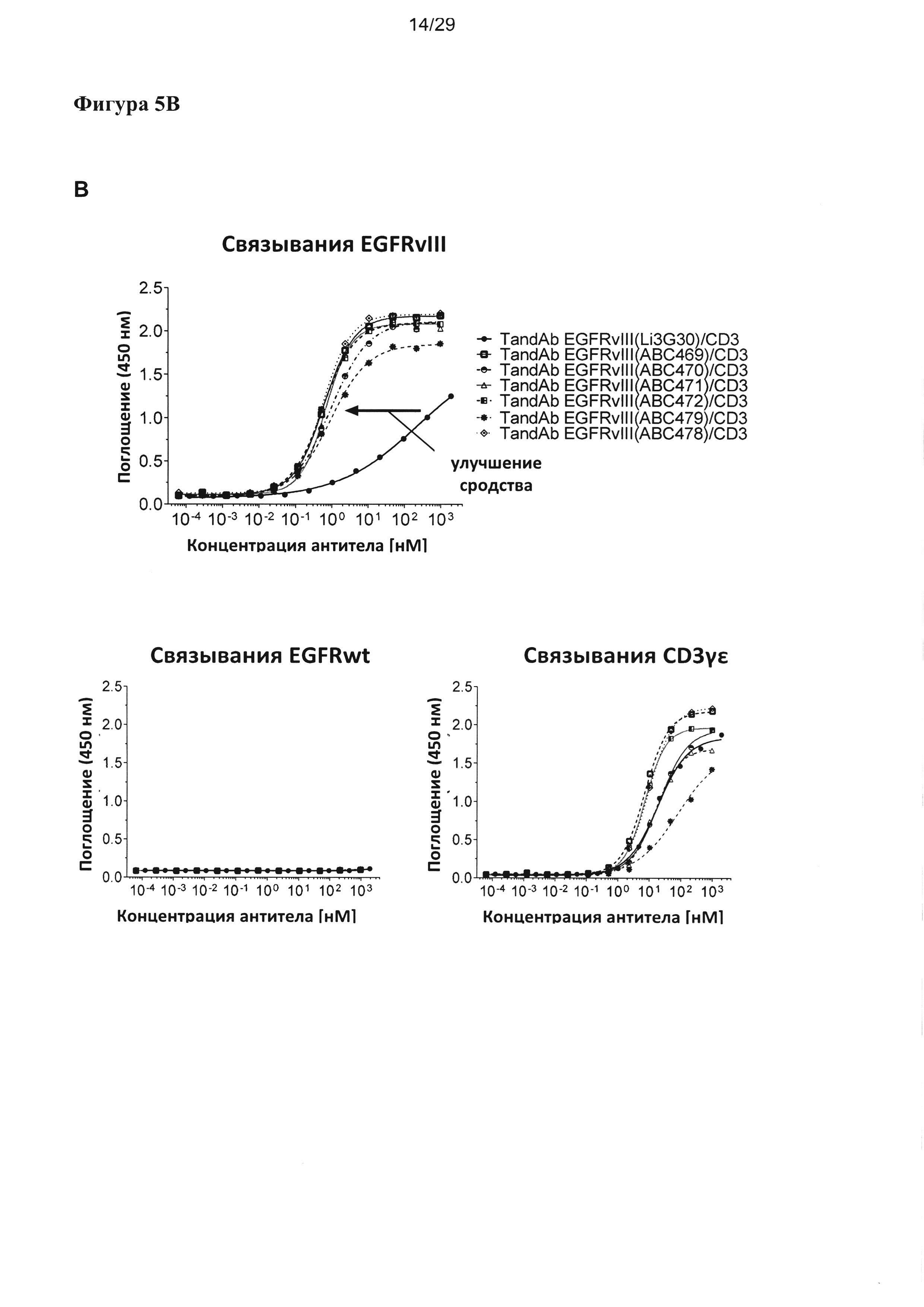


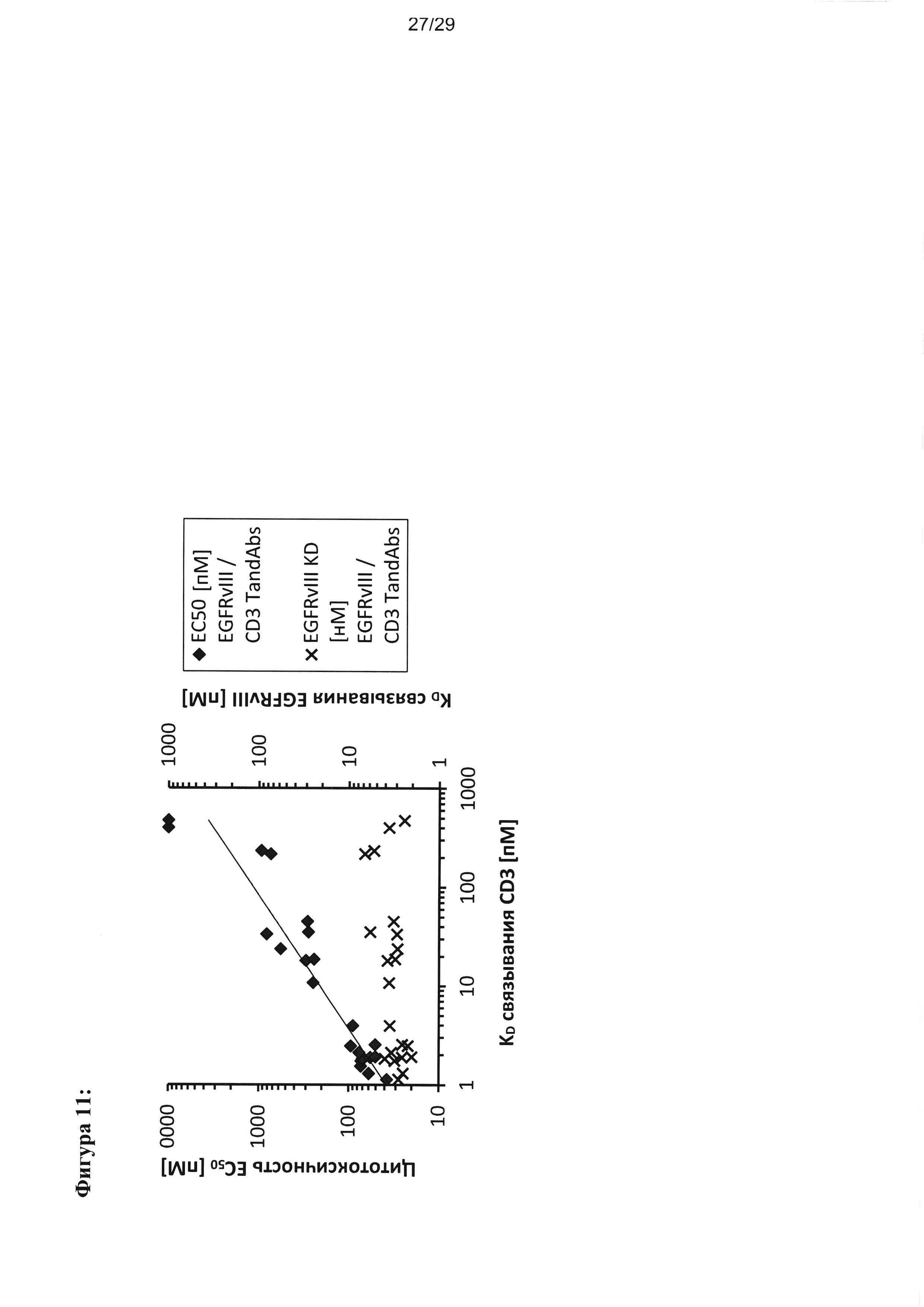
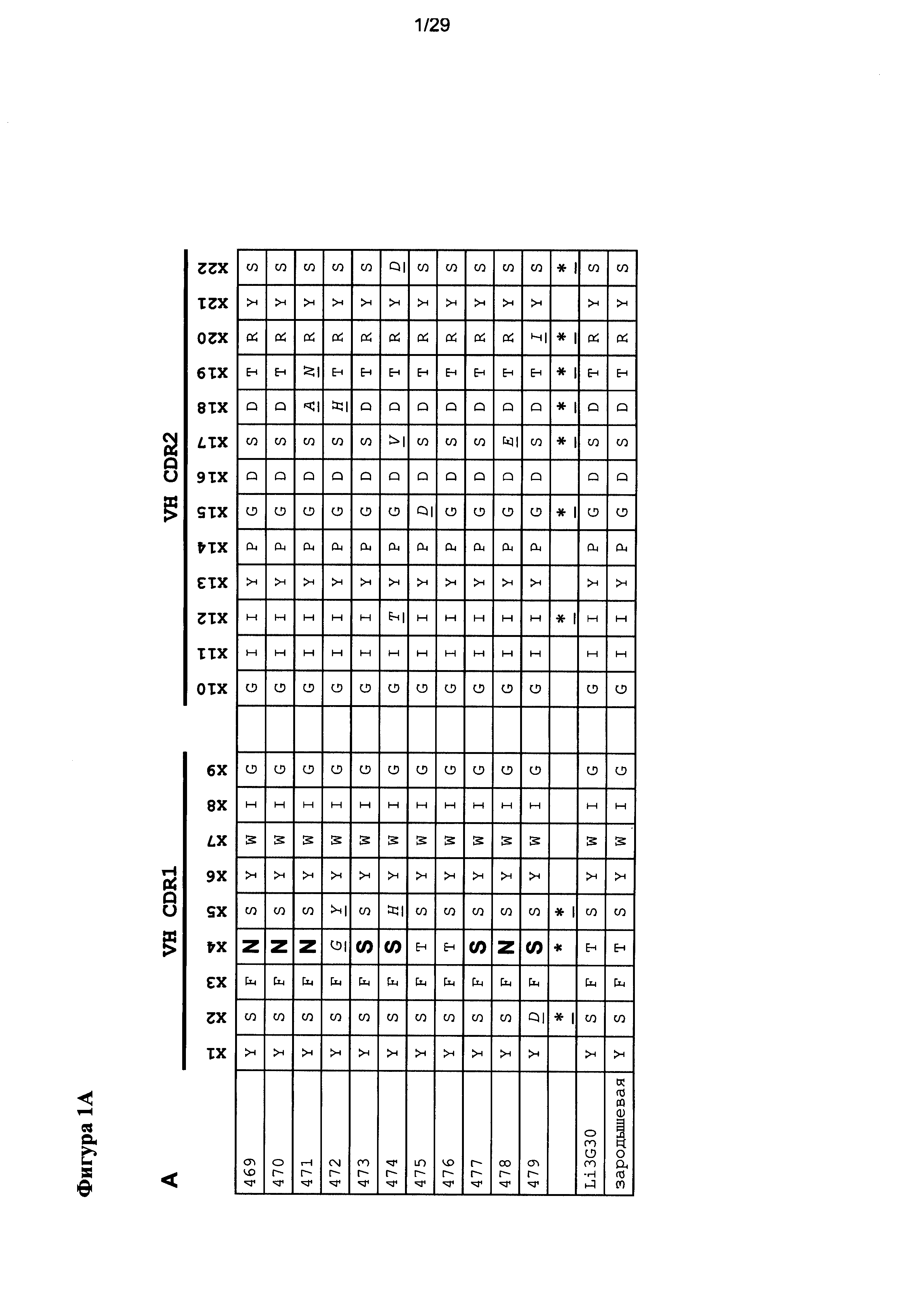

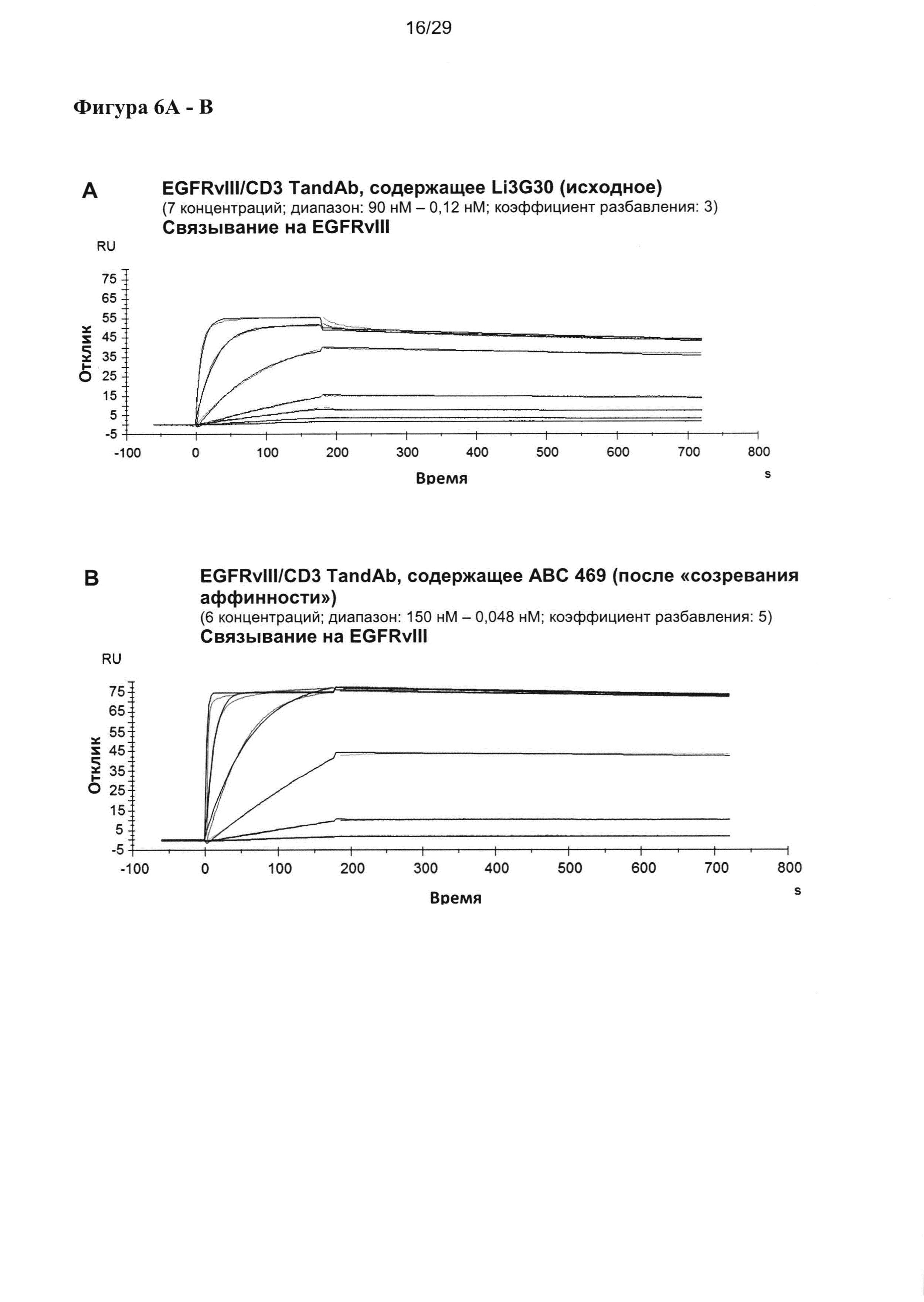


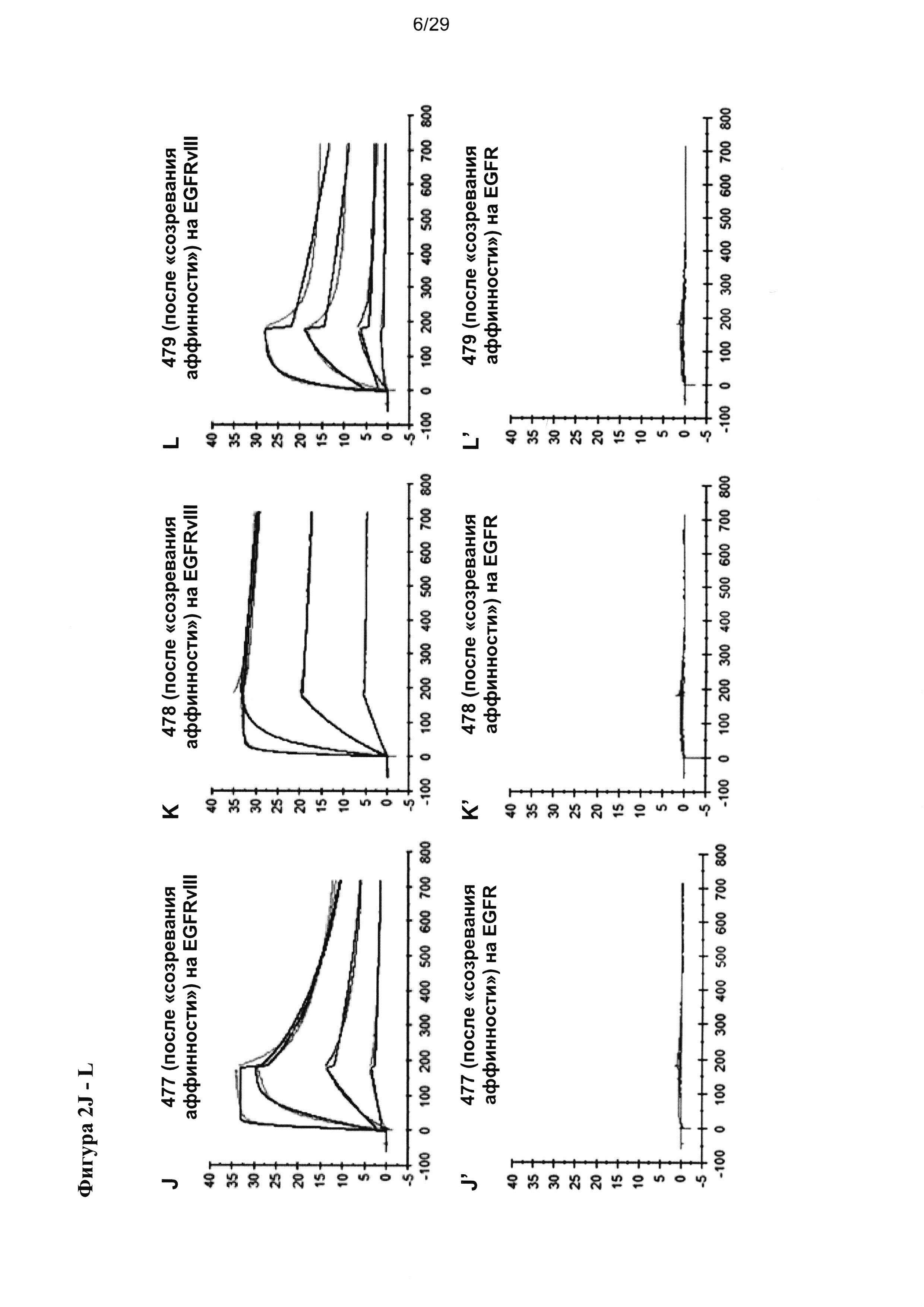
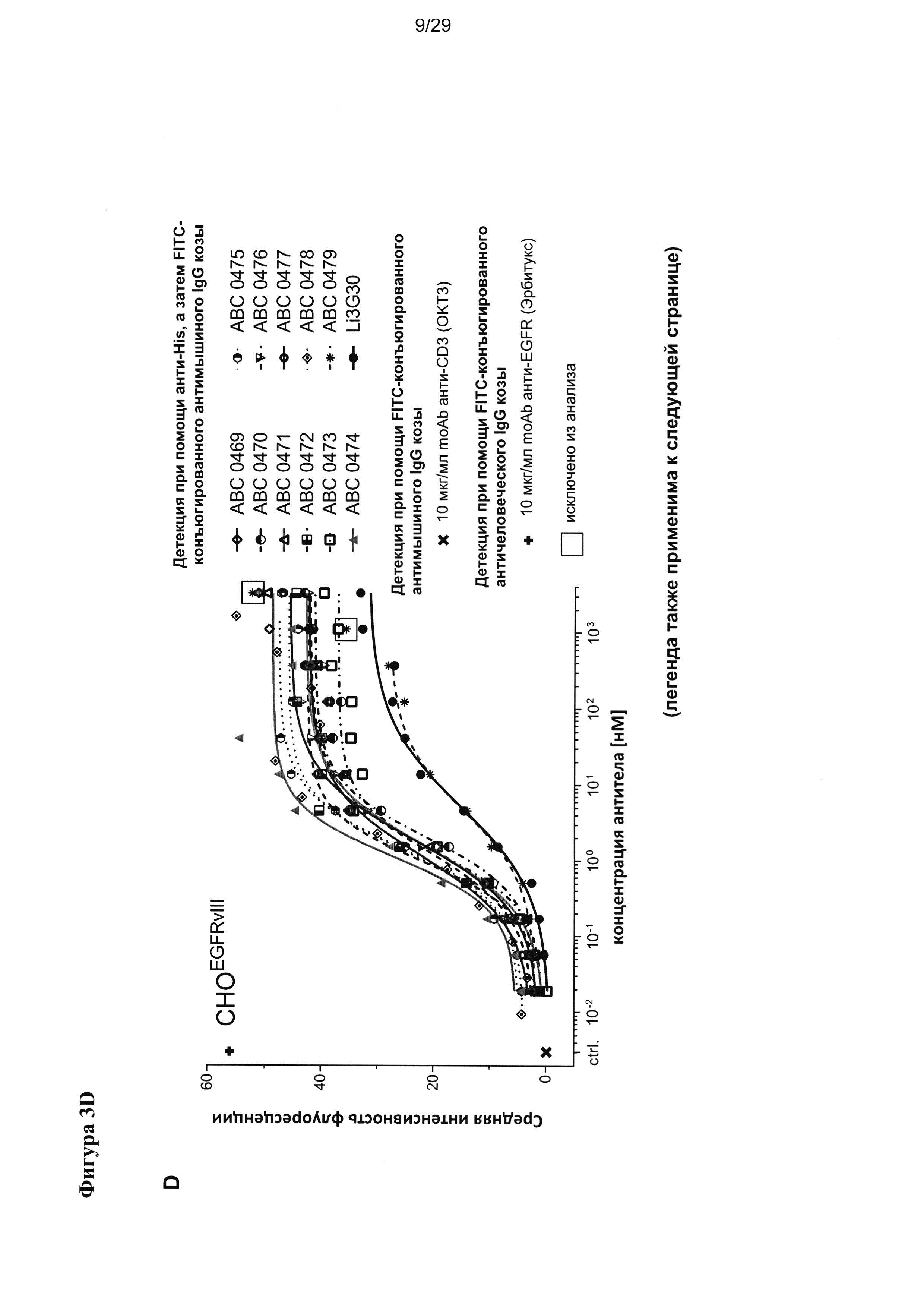


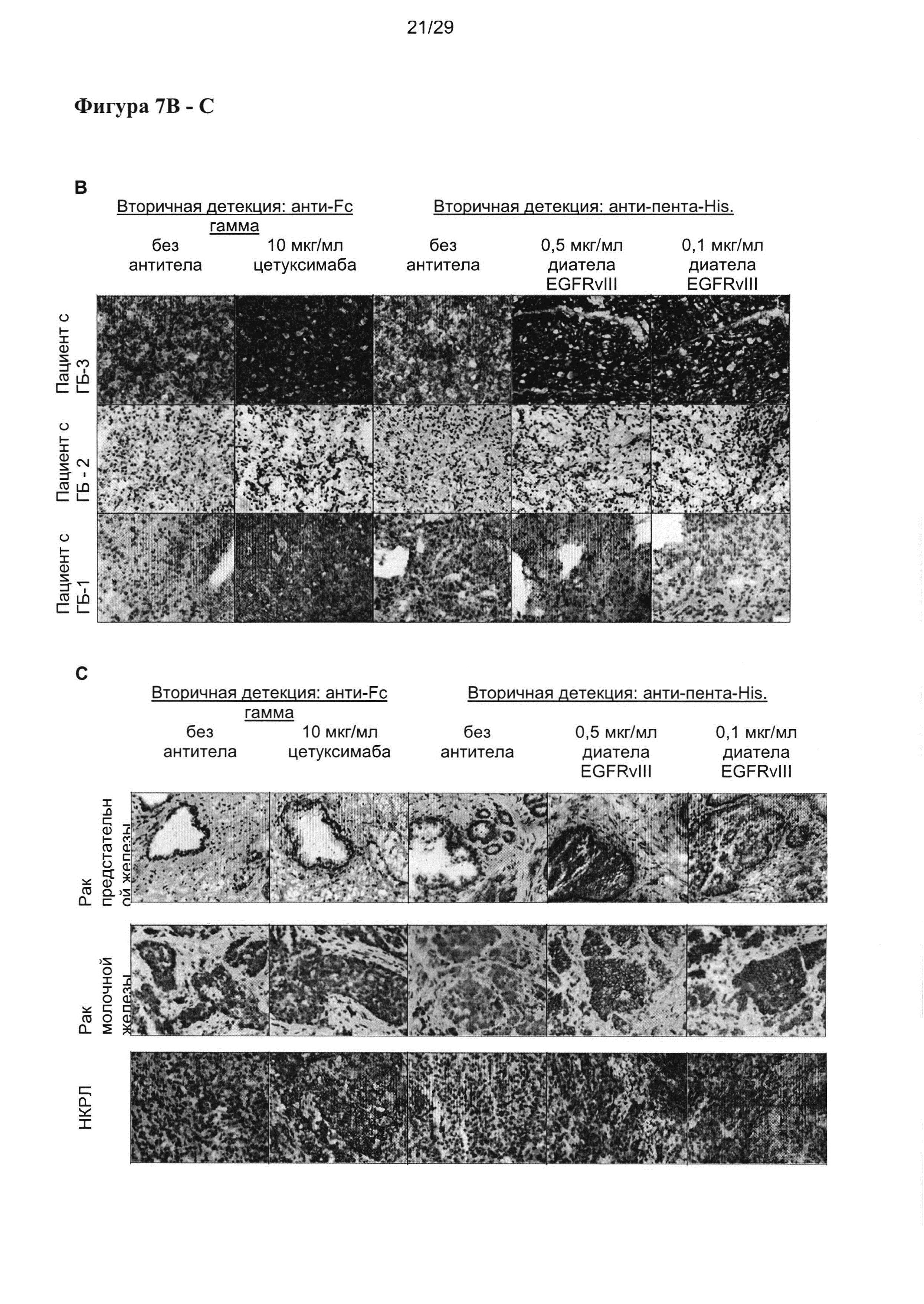
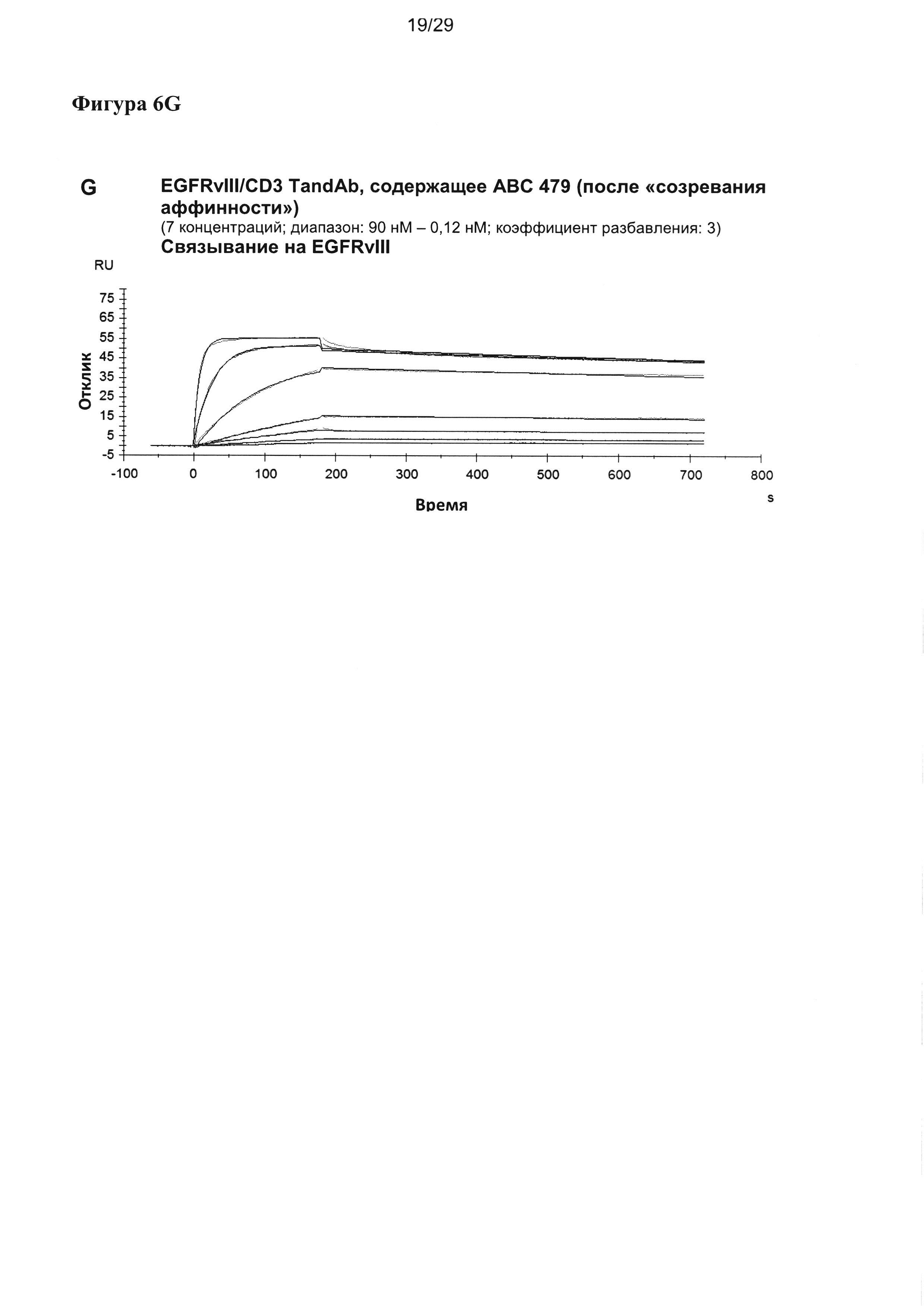
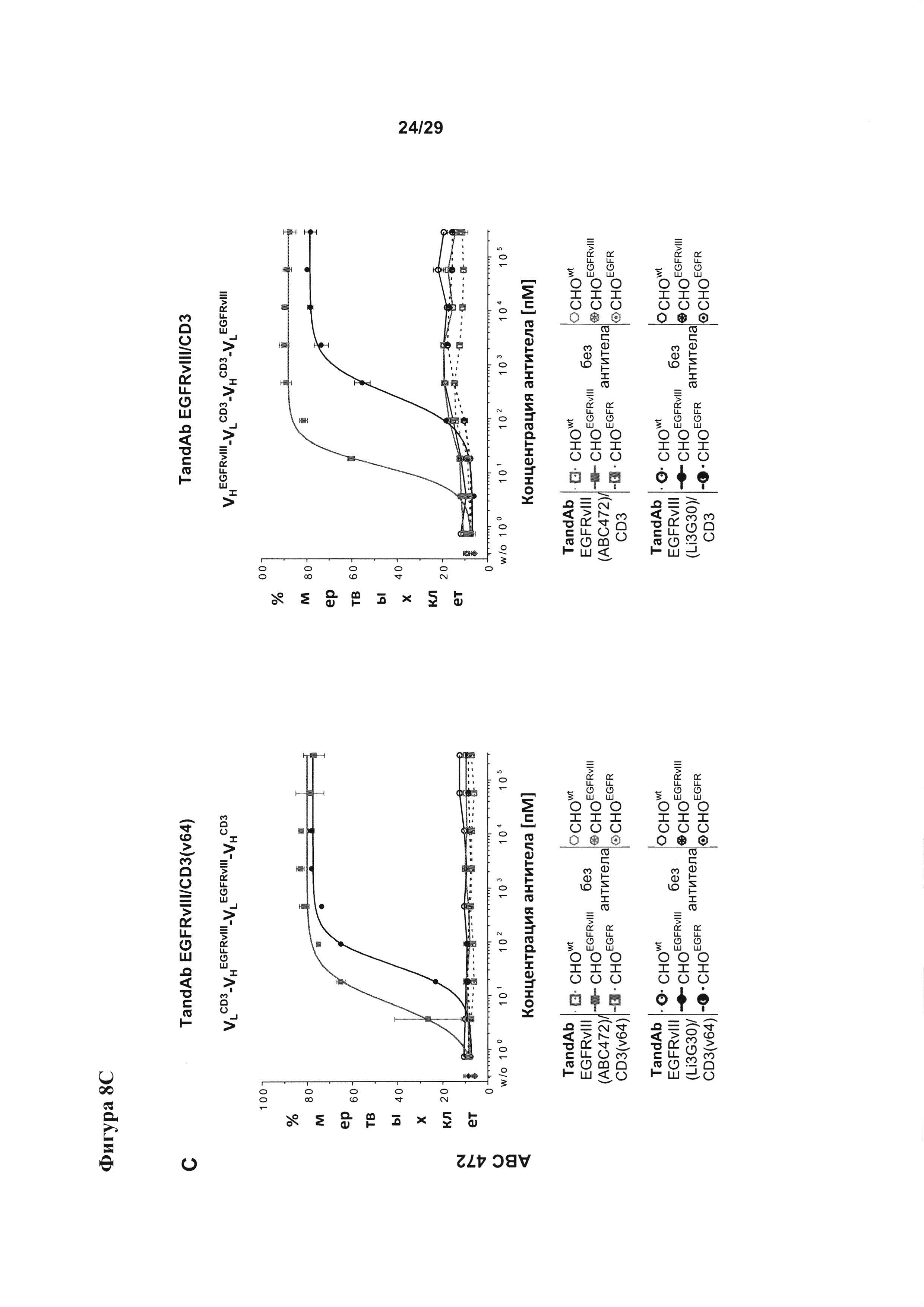


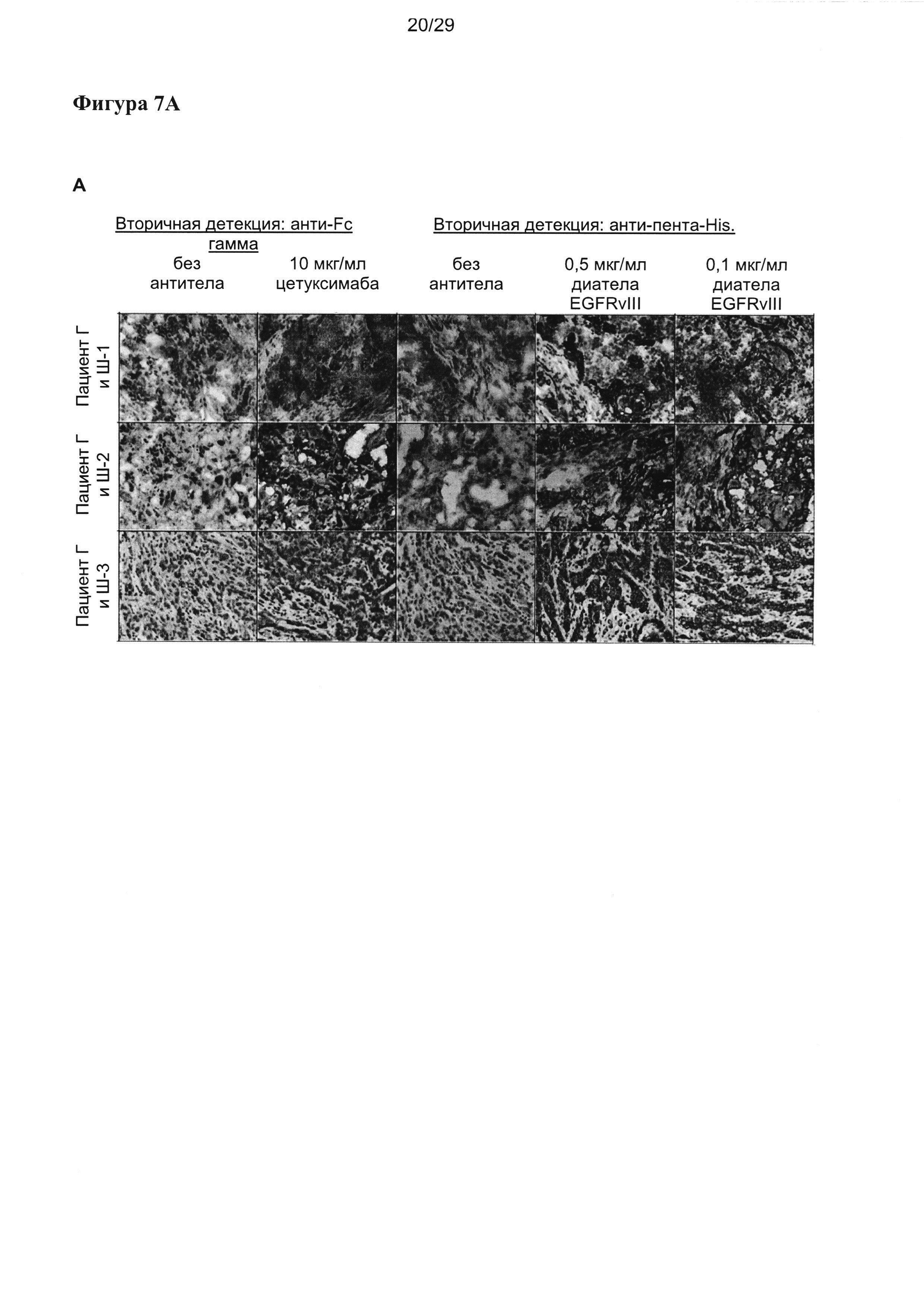


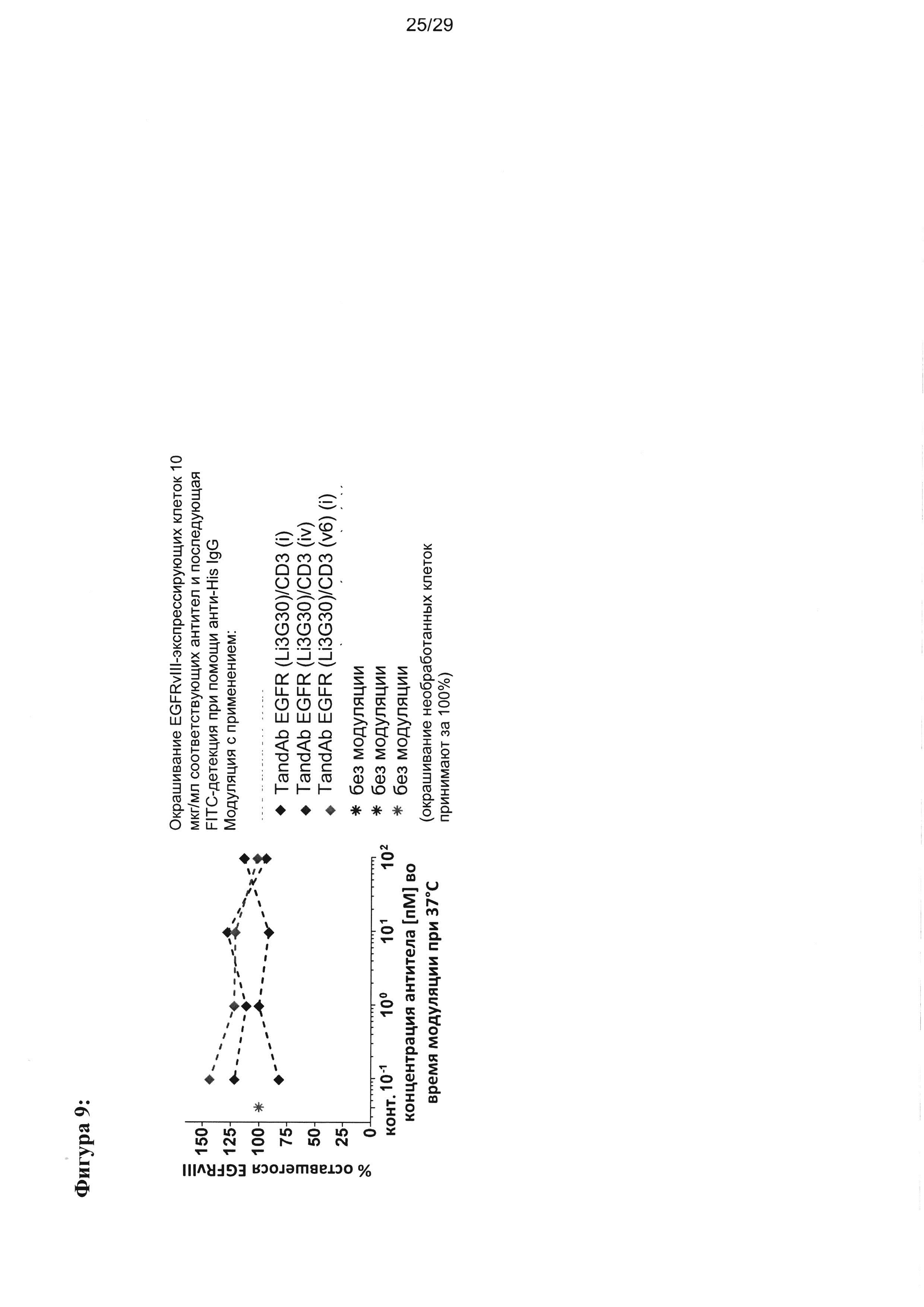




Поливалентная антиген-связывающая fv-молекула
Cd3-связывающий домен
Cd3-связывающий домен
Cd3-связывающий домен
Анти-cd 16 связывающие молекулы
Cd3-связывающий домен
Cd3-связывающий домен
Cd3-связывающий домен

