Результат интеллектуальной деятельности: Генетическая конструкция на основе двух индуцибельных экспрессионных векторов для экспрессии тиазол/оксазол модифицированного пептида в клетках бактерий E. coli; способ получения рекомбинантного штамма бактерий E. coli и способ получения модифицированного пептида на его основе
Вид РИД
Изобретение
Тиазол/оксазол-модифицированные микроцины (ТОММ) представляют собой подмножество более крупного класса натуральных продуктов - рибосомально синтезируемых и посттрансляционно модифицируемых пептидов (РиПП) [1, Paul G. Arnison et al., 2013]. Биосинтез РиПП начинается с трансляции рибосомой небольшого пептида-предшественника (20-110 остатков), состоящего из N-концевого отрезаемого лидерного пептида и С-концевого модифицируемого пептида, который подвергается последующим посттрансляционным модификациям специальными модифицирующими ферментами.
ТОММ образуют структурно разнообразное семейство натуральных продуктов, характеризующееся наличием посттрансляционно установленных тиазольных и оксазольных гетероциклов, формируемых цистеиновыми, сериновыми и треониновыми остатками в составе пептида-предшественника [2, Melby JO et al., 2011]. Кроме того, к этим основным модификациям многие ТОММ содержат множество других посттрансляционных модификаций [2, Melby JO et al., 2011; 3, McIntosh JA et al., 2011; 4, Lee J et al., 2013; 5, McIntosh JA et al., 2009]. ТОММ характеризуются разнообразными биологическими активностями, включая ингибиторы ДНК-гиразы, ингибиторы трансляции и гемолитические токсины [1, Paul G. Arnison et al., 2013; 6, Molloy EM et al., 2011; 7, Lee SW et al., 2008; 8,  JL et al., 1991; 9, Bagley MC et al., 2005].
JL et al., 1991; 9, Bagley MC et al., 2005].
Тиазолы и оксазолы устанавливаются путем последовательного действия АТФ-зависимой циклодегидратазы (С- и D-белка) и FMN-зависимой дегидрогеназы (В-белка), которые ответственны за образование и за окисление азолина соответственно [10, Koehnke J et al., 2013; 11, McIntosh JA, Schmidt EW 2010; 12, Dunbar KL et al., 2012; 13, Milne JC et al., 1998].
Технический результат заключается в разработке такой генетической конструкции, которая позволяет получать пептид из класса тиазол-оксазол модифицированных микроцинов с нетипичной для пептидов модификацией, то есть расширить арсенал структурных мотивов тиазол-оксазол модифицированных пептидов.
Задача состоит в разработке способа получения тиазол-оксазол модифицированного пептида, названного авторами клебсазолицин, имеющего аминокислотную последовательность SerGlnSerProGly-AsnCysAlaSerCysSerAsnSerAlaSerAlaAsnCysThrGlyGlyLeuGly, в модельном гетерологическом организме Е. coli.
Для получения указанного технического результата предложено создание рекомбинантного штамма бактерии Е. coli, продуцирующего модифицированный пептид клебсазолицин в культуральную среду. Предложено разработать генетическую конструкцию, обеспечивающую биосинтез целевого пептида в гетерологическом организме - клетках бактерий Е. coli, и его экстракцию в культуральную среду.
Кроме того, для достижения указанного технического результата предложено разработать способ получения тиазол-оксазол модифицированного пептида клебсазолицина путем выделения из культуральной жидкости бактерий, полученной при культивировании микроорганизма, созданного указанным выше способом.
Заявляемый технический результат достигается тем, что, разработана генетическая конструкция для обеспечения гетерологической экспрессии тиазол-оксазол модифицированного пептида клебсазолицина в клетках бактерий Е. coli, сконструированная на основе двух совместимых индуцибельных векторных плазмид pBAD His/B и рЕТ28а. Конструкция содержит фрагменты геномной ДНК микроорганизма Klebsiella pneumoniae subsp.ozaenae (АТСС 11296), составляющие генетический кластер биосинтеза клебсазолицина, а именно, ген пептида-предшественника klpA, гены белков процессинга klpB, klpC, klpD и ген белка-транспортера klpE.
При этом плазмида pBAD His/B по сайтам рестрикции SacI и PstI содержит фрагмент, включающий гены klpB, klpC, klpD и klpE, который получают путем амплификации геномной ДНК Klebsiella pneumoniae subsp. ozaenae АТСС 11296 с использованием олигонулеотида klpCF с последовательностью 5'-ATTATGAGCTCaaataacatttataaggccgcag-3' и олигонулеотида klpER с последовательностью ATTATCTGCAGCttaaaaattatttaattcca-ttacaacat-3', а плазмида рЕТ28а по сайтам рестрикции NcoI и XhoI содержит фрагмент, включающий ген klpA, который получают путем амплификации геномной ДНК Klebsiella pneumoniae subsp.ozaenae АТСС 11296 с использованием олигонулеотида klpAF с последовательностью 5'-ATAATCCatggctaaaatcaagaatcgttttgg-3' и олигонуклеотида klpAR с последовательностью 5'-ATTATCTCGAGttaacctaagccacctgtacaatta-3'.
Заявляемый технический результат достигается также тем, что предложен способ получения на основе бактерии рекомбинантного штамма бактерии E. coli штамма продуцента клебсазолицина продуцирующего модифицированный пептид клебсазолицин, предусматривающий трансформацию клеток бактерии Е. coli BL21(DE3) генетической конструкцией, описанной выше.
Заявляемый технический результат достигается также тем, что предложен способ получения тиазол-оксазол модифицированного пептида клебсазолицина, путем выделения из культуральной среды, получаемой при культивировании описанного выше микроорганизма.
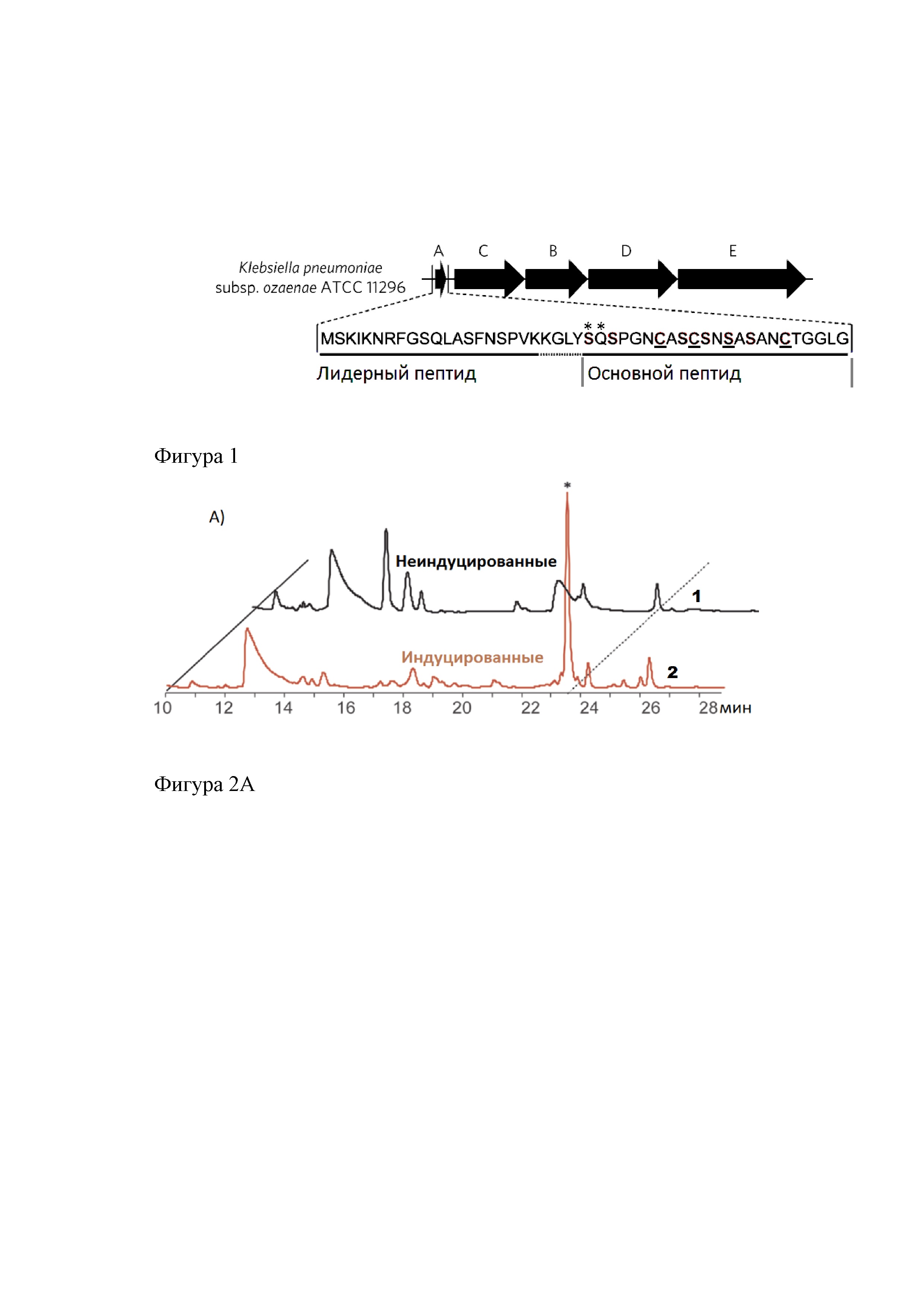
На фиг. 1 представлена организация кластера биосинтеза клебсазолицина. Обозначены: ген klpA, кодирующий пептид-предшественник; гены klpB, klpC, klpD, кодирующие BCD-синтетазу, и ген klpE, кодирующий ABC-транспортер. Также приведена аминокислотная последовательность пептида-предшественника.
Аминокислотные остатки, образующие азолы, подчеркнуты; вовлеченные в формирование амидинового кольца - обозначены звездочкой.
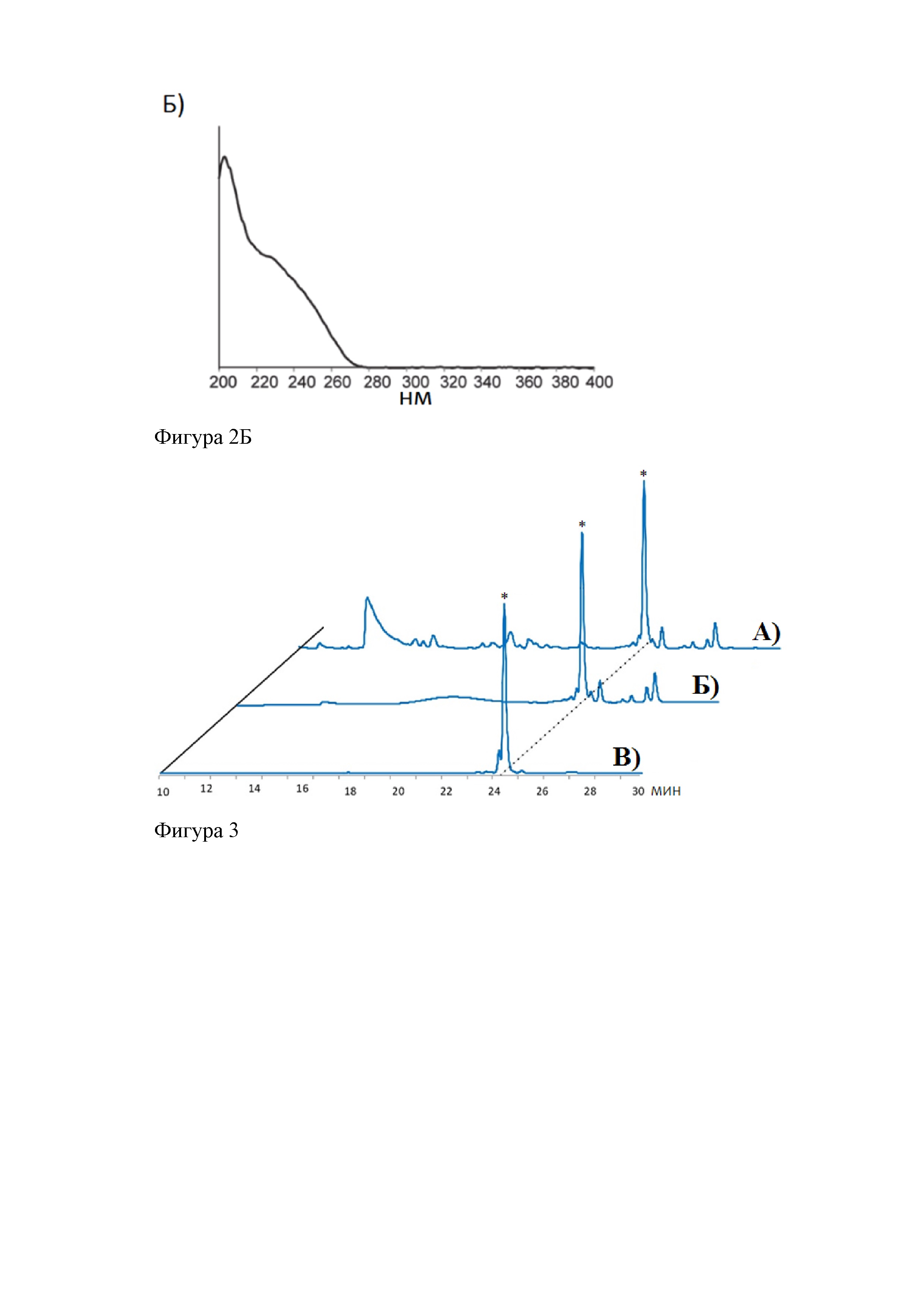
На фиг. 2 представлено А) Сравнение УФ-поглощения при ВЭЖХ супернатантов неиндуцированных (1) и индуцированных (2) культур клеток Е. coli, несущих кластер генов klpABCDE. Б) Спектр поглощения ультрафиолетового излучения клебсазолицина.
На фиг. 3 представлено сравнение УФ-поглощения при ВЭЖХ образцов культуральной среды индуцированных клеток Е. coli BL21(DE3), содержащих кластера биосинтеза клебсазолицина, при последовательной очистке посредством твердофазной экстракции на картридже С18 и последующей ВЭЖХ с обращенной фазой: А) неочищенной, Б) очищенной посредством твердофазной экстракции на картридже С18 с В) последующей очисткой ВЭЖХ с обращенной фазой, клебсазолицин-содержащая фракция помечена звездочкой.
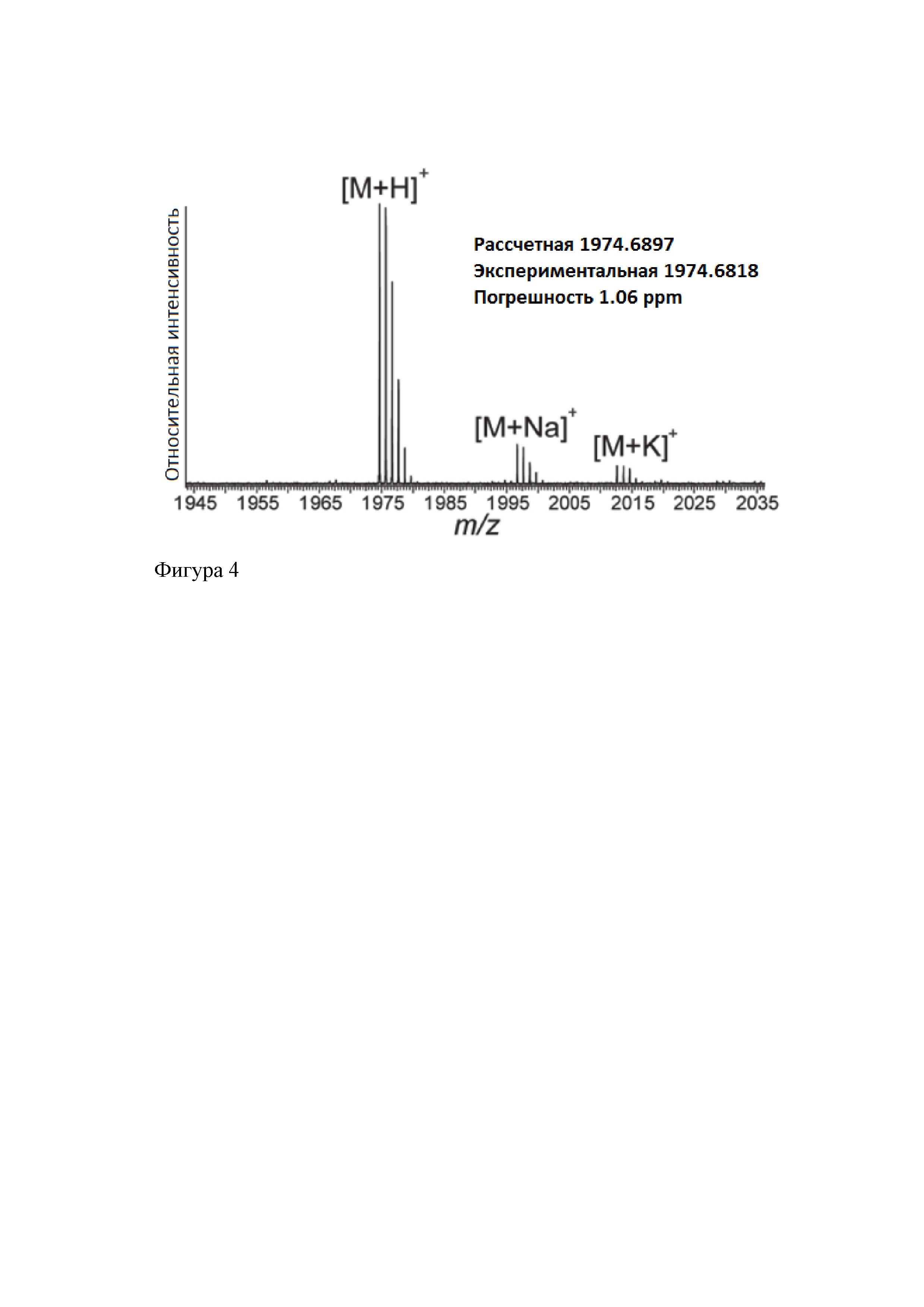
На фиг. 4 представлен ФП-МС спектр ВЭЖХ очищенного клебсазолицина. Показаны значения m/z основного пика [М+Н]+, а также [M+Na]+ и [М+K]+. Представлены экспериментально измеренное и рассчитанное значения m/z [М+Н]+.
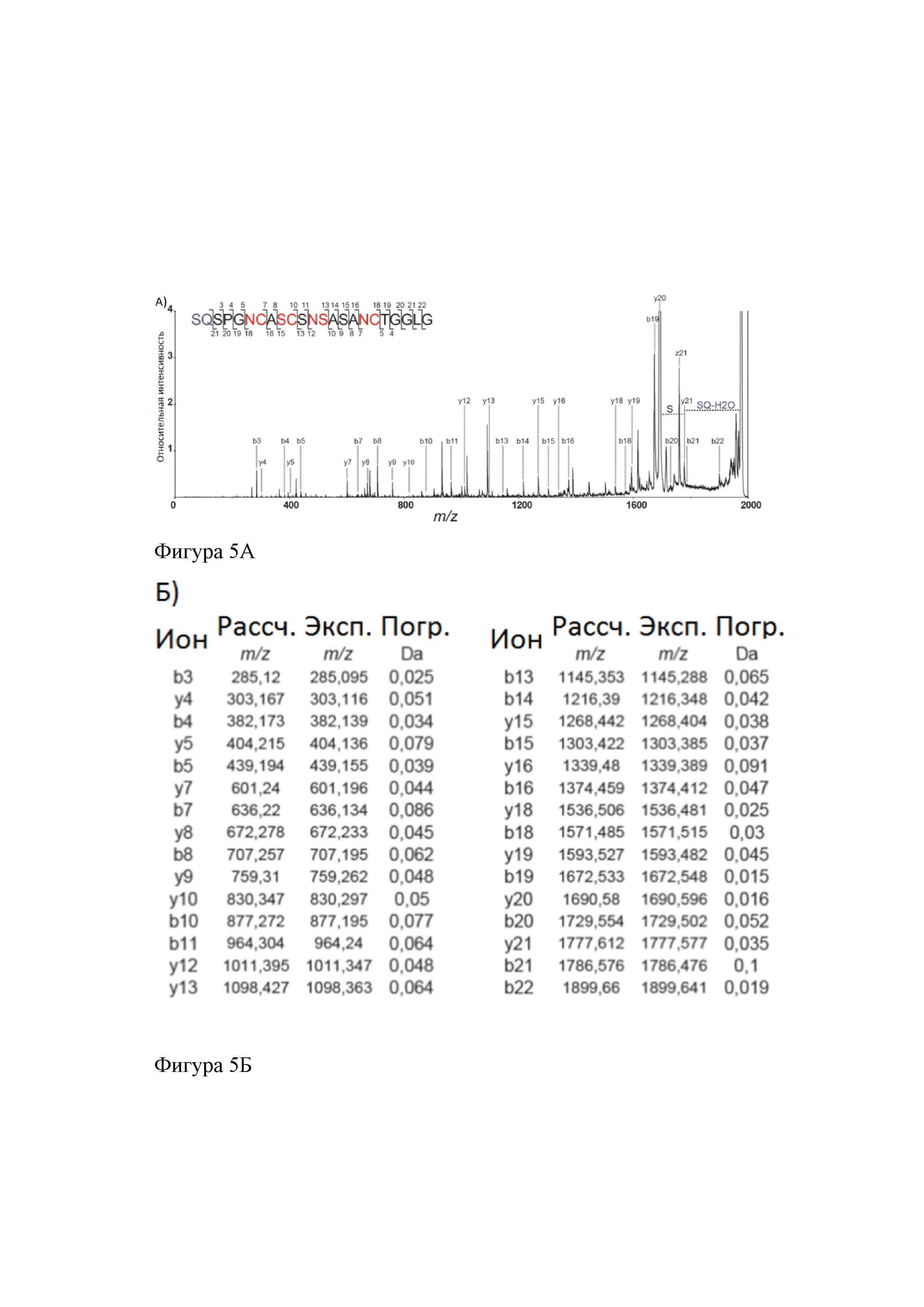
На фиг. 5 представлены: А) МС-МС спектрограмма клебсазолицина с наблюдаемыми b- и y-ионами; Б) значения m/z присутствующих в спектре b- и y-ионов: рассчитанные, экспериментальные, погрешности.
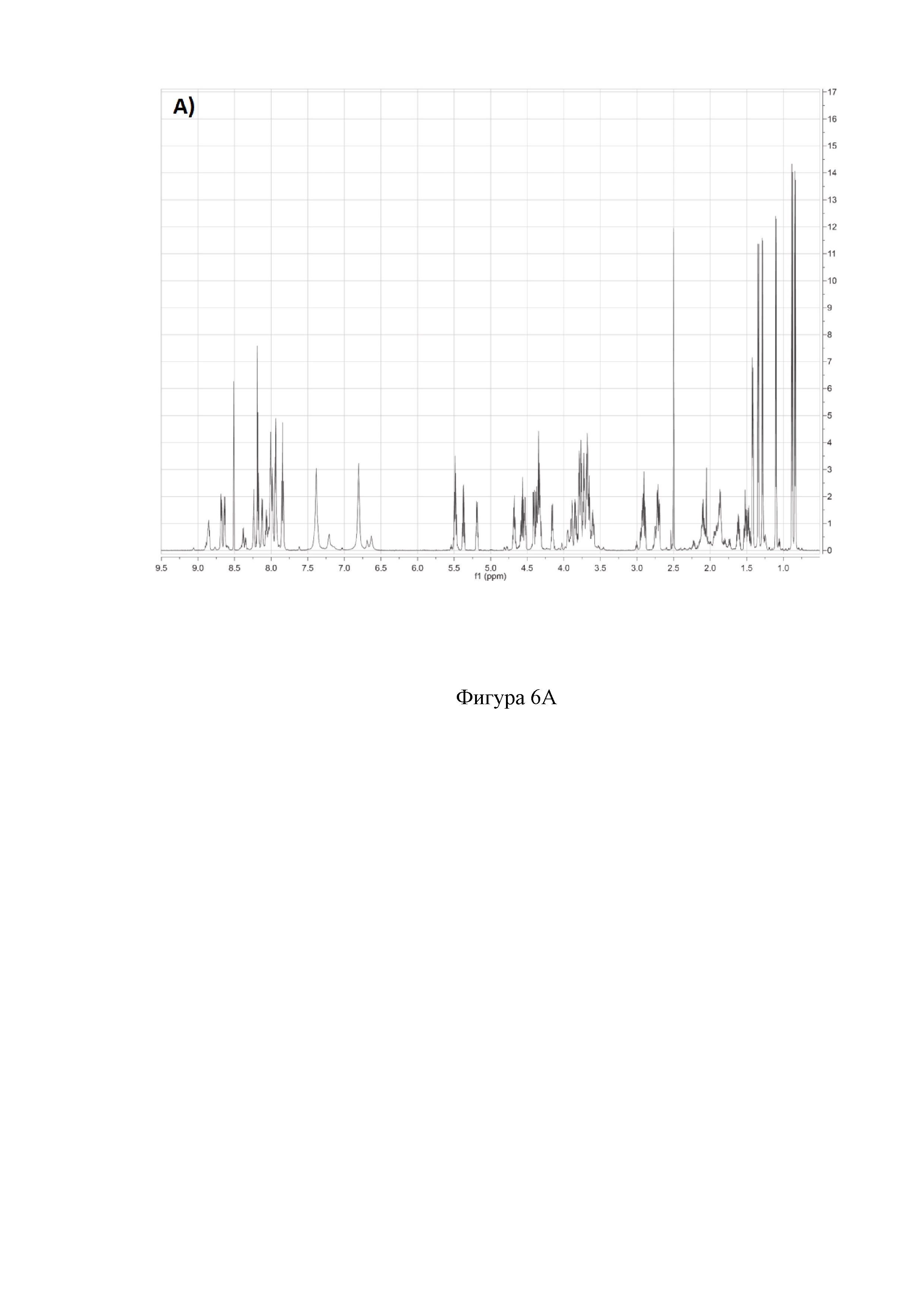
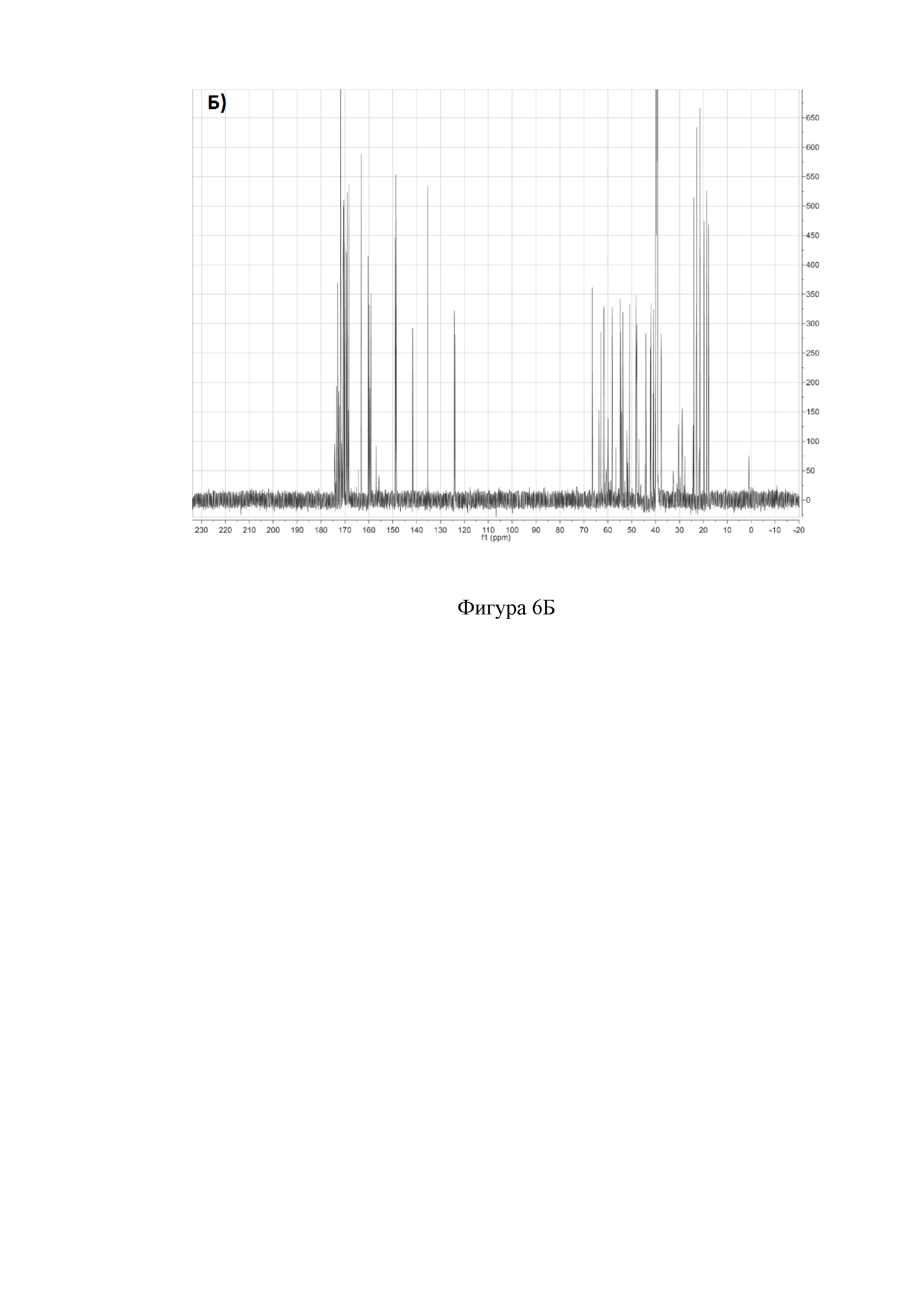
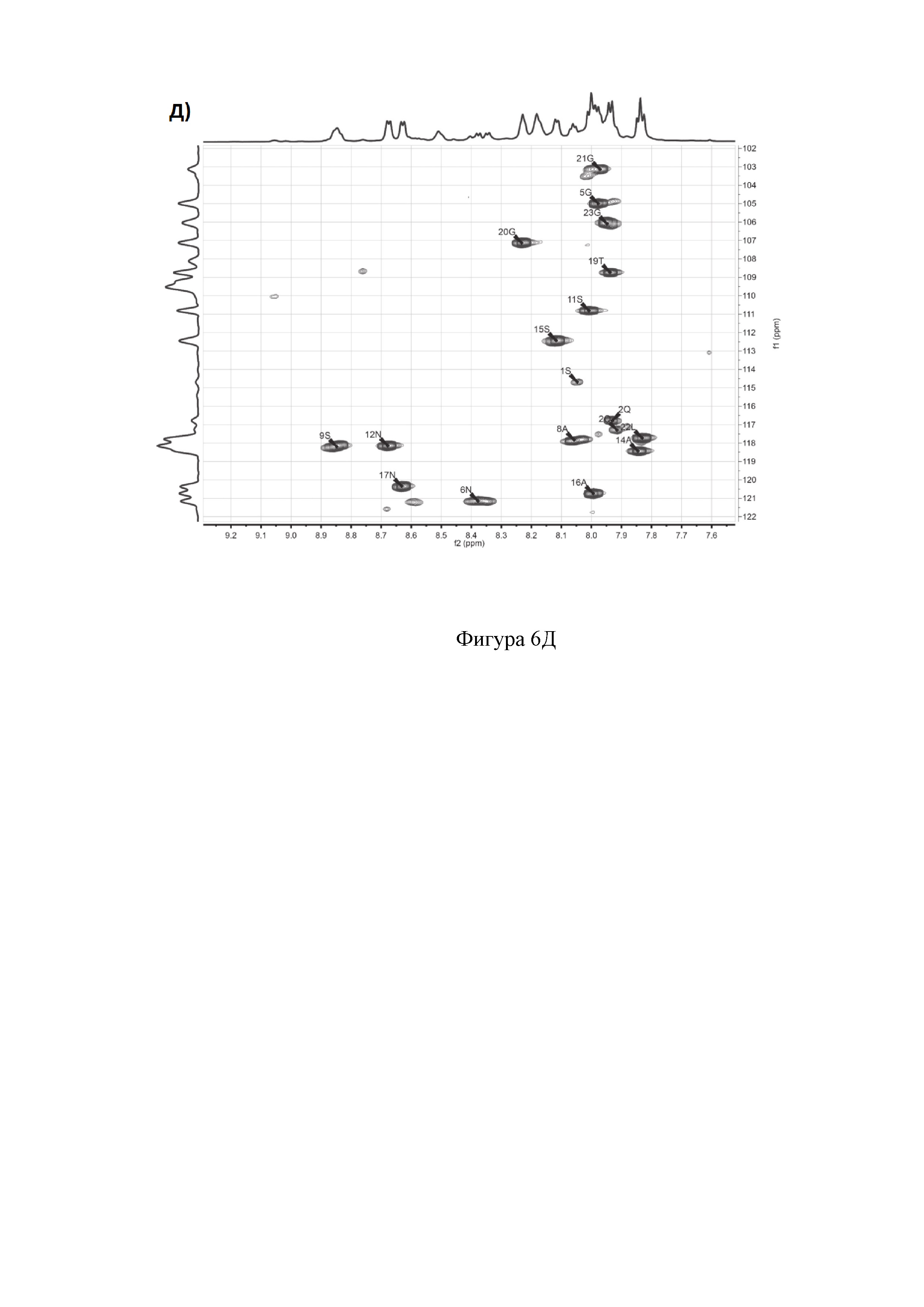
На фиг. 6 представлены двумерные ЯМР-спектры клебсазолицина:
A) 1Н спектр клебазолицина в растворителе DMSO-d6.
Б) 13С спектр клебсазолицина в растворителе DMSO-d6.
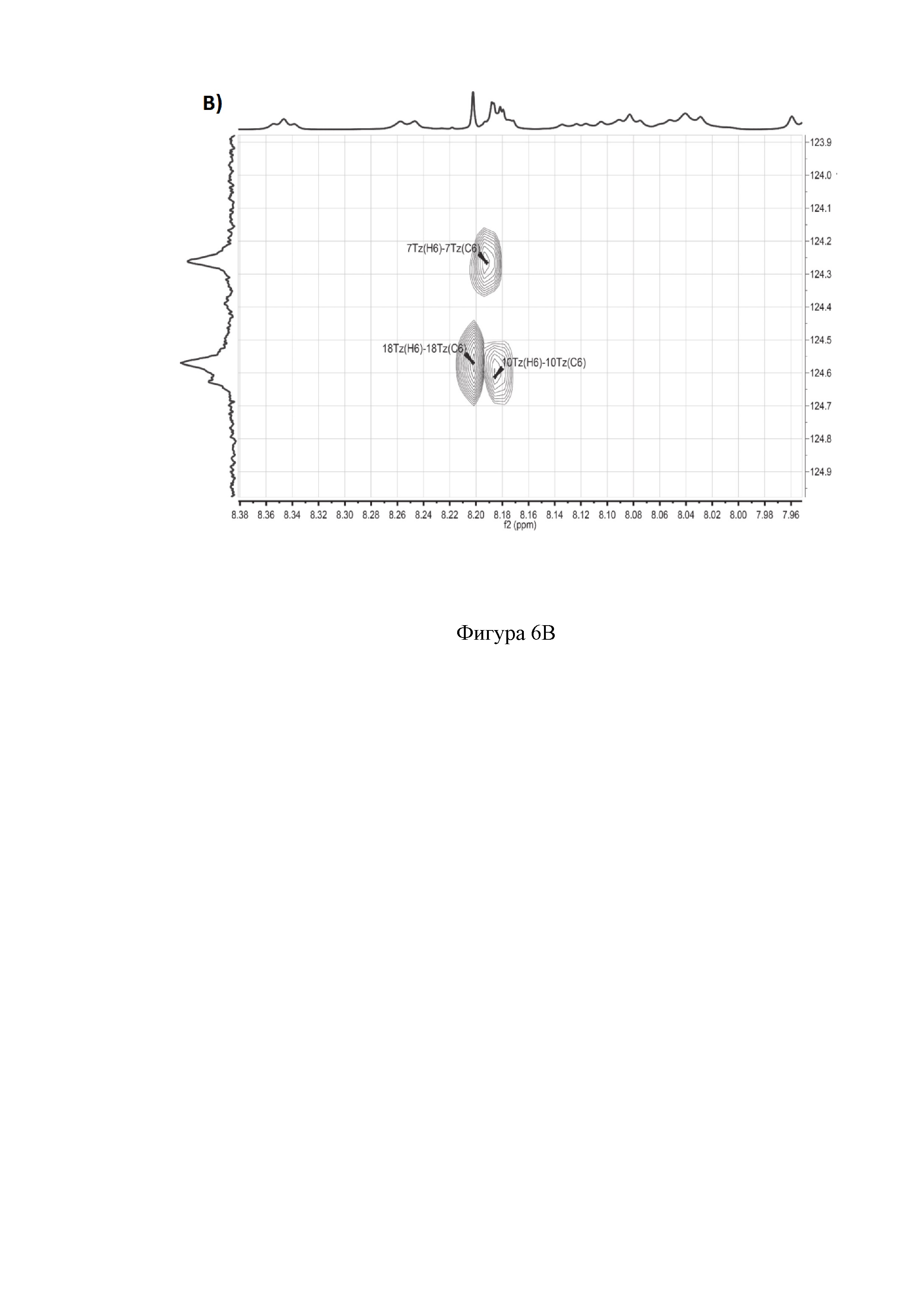
B) Фрагменты 13C-HSQC спектра с определением тиазолов.
Г) Фрагменты 13C-COZY спектра с определением оксазола.
Д) Амидновый регион 1H-15N HSQC- спектра клебсазолицина.
Е) Фрагменты 13С-НМВС-спектра.
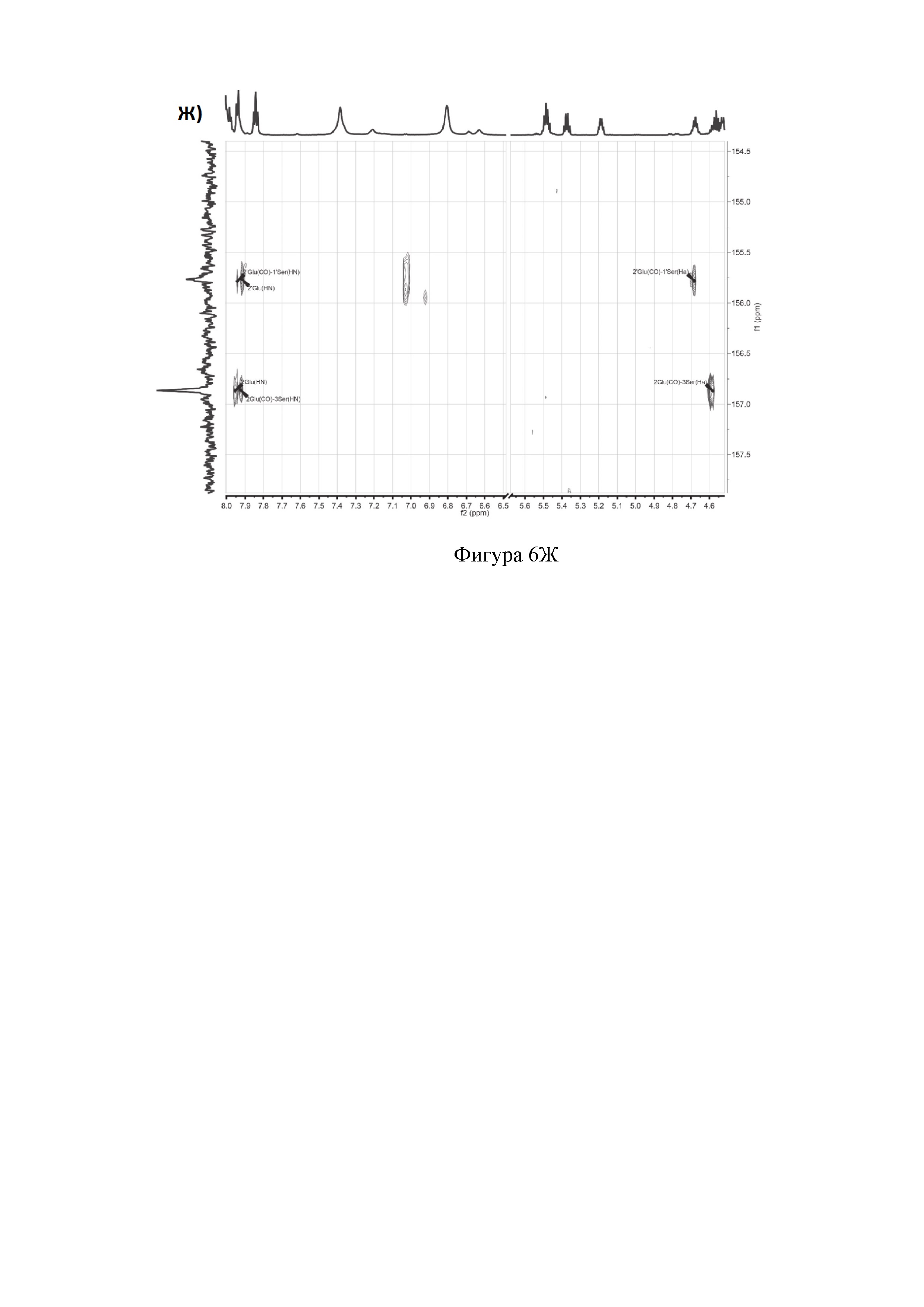
Ж) Фрагменты 13С-НМВС-спектра (Ser1, Glu2 и Ser3).
Описание изобретения
В этом изобретении мы сообщаем о способе получения структурно уникального ТОММ, который является первым членом нового класса ингибиторов синтеза белка.
В приложении 1 представлен полный генетический кластер синтеза клебсазолицина, обнаруженный в геноме Klebsiella pneumoniae subsp. ozaenae (АТСС 11296), включает ген пептида-предшественника klpA, гены белков процессинга klpB, klpC, klpD и ген белка-транспортера klpE, ответственного за экспорт из клеток зрелого клебсазолицина из клеток (фигура 1).
Модифицируемая часть пептида-предшественника, образуемая после отщепления лидерного пептида, состоит из 23-х остатков аминокислот: SerGlnSerProGlyAsnCysAlaSerCysSerAsnSerAlaSerAlaAsnCysThrGlyGlyLeu Gly. Подвергнутый процессингу этот пептид имеет молекулярную массу 1973, 684 Да и содержит модификации аминокислотных остатков в положениях 7, 10, 13, 18 с образованием азольных колец и ранее не показанную для пептидов модификацию первых двух N-концевых остатков с образованием 6-ти членного азолинового цикла.
Схема способа получения клебсазолицина состоит из трех типовых этапов, описанных ниже. Применение описанного способа позволяет достичь выхода клебсазолицина 15-40 мг на литр культуральной среды.
Этап 1. Создание генетической конструкци для обеспечения гетерологичной экспрессии клебсазолицина в клетках бактерий Е. coli.
На этом этапе осуществляется получение фрагментов геномной ДНК Klebsiella pneumoniae subsp.ozaenae АТСС 112 96, содержащих гены кластера биосинтеза клебсазолицина и искусственные сайты рестрикции, посредством ПЦР амплификации. Полученные фрагменты клонируют в стандартные плазмидные векторы pBAD His/B и рЕТ28а, несущие в качестве селективных маркеров гены устойчивости к антибиотикам ампициллину и канамицину соответственно.
Фрагмент 1 (162 bp), включающий ген пептида-предшественника, klpA, и сайты для рестриктаз NcoI (на 5'-конце) и XhoI (на 3'-конце), амплифицируют с использованием пары олигонулеотидов klpAF и klpAR и геномной ДНК Klebsiella pneumoniae subsp.ozaenae в качестве матрицы:
klpAF: 5'-ATAATCCatggctaaaatcaagaatcgttttgg-3'
klpAR: 5'-ATTATCTCGAGttaacctaagccacctgtacaatta-3'
Фрагмент 2 (4495 bp), содержащий участок кластера биосинтеза клебсазолицина с генами белков процессинга и геном белка-транспортера, klpBCDE, фланкированный сайтами для рестриктаз SacI (на 5'-конце) и PstI (на 3'-конце), амплифицируют с использованием олигонулеотидов klpCF и klpER и геномной ДНК Klebsiella pneumoniae subsp. ozaenae в качестве матрицы:
klpCF: 5'-ATTATGAGCTCaaataacatttataaggccgcag-3'
klpER: 5'-ATTATCTGCAGCttaaaaattatttaattccattacaacat-3'
Внедрение полученных ДНК фрагментов в векторные плазмиды осуществляют по следующей схеме: Фрагмент 1, так же как и плазмидный вектор рЕТ28а, обрабатывают парой эндонуклеаз рестрикции NcoI и XhoI, что приводит к образованию попарно комплементарных одноцепочечных участков на фрагменте и на векторе. Полученные на этом шаге рестриктный фрагмент и линеаризованный вектор лигируют друг с другом, в результате чего образуется плазмидный вектор, названный нами pET28-klpA.
Аналогичным образом осуществляют клонирование Фрагмента 2 в плазмидный вектор pBAD His/B с применением пары эндонуклеаз рестрикции SacI и PstI. Сконструированный таким способом вектор мы назвали pBAD-klpBCDE.
Результатом описанных манипуляций является получение двух совместимых индуцибельных экспрессионных векторов pET28-klpA и pBAD-klpBCDE, несущих суммарно полный генетический кластер биосинтеза клебсазолицина с транспортером под контролем Т7-промоторов.
Этап 2. Получение рекомбинантного штамма бактерии Е. coli, продуцирующего модифицированный пептид клебсазолицин.
Рекомбинантный штамм-продуцент клебсазолицина создают путем котрансформации реципиентного штамма Е. coli BL21(DE3) двумя плазмидными векторами, полученными на Этапе 1. Отбор колоний, несущих одновременно оба плазмидных вектора, осуществляют при помощи культивирования трансформированных клеток Е. coli на агарной питательной среде LB в присутствии двух селективных антибиотиков: ампициллина (100 мкг/мл) и канамицина (50 мкг/мл).
Этап 3. Получение модифицированного пептида клебсазолицина, путем культивирования рекомбинантного микроорганизма, созданного на Этапе 2.
Штамм Е. coli BL21(DE3), трансформированный плазмидами pBAD-klpBCDE и рЕТ2 8-KlpA, культивируют в жидкой питательной среде LB, дополненной селективными антибиотиками ампициллином (100 мкг/мл) и канамицином (50 мкг/мл). При достижении бактериальной культурой оптической плотности OD600 = 0,6, клетки осаждают центрифугированием в течение 10 мин с ускорением 4000G при комнатной температуре, промывают раствором фосфатного буфера, осаждают повторно при тех же условиях и переносят в индуцирующую минимальную питательную среду М9 (1 л содержит 12.8 г Na2HPO4⋅7H2O, 3 г KH2PO4, 0.5 г NaCl, 1 г NH4Cl, 1 мл 2М раствора MgSO4, 100 мкл 1М раствора CaCl2), дополненную 1% глицерола (объем/объем), 10 мкг/мл тиамина, 100 мкг/мл ампициллина, 50 мкг/мл канамицина и двумя индукторами - 10 мкМ арабинозы, 0.1 мкМ IPTG. Индуцированные клетки инкубируют в течение 24 ч при 30°С, после чего осаждают путем центрифугирования в течение 30 минут с ускорением 12000G при комнатной температуре и отбирают культуральный супернатант, содержащий целевой пептид.
На фигуре 2.А приведены данные сравнения УФ-поглощений при ОФ ВЭЖХ-анализе культуральных супернатантов индуцированных и неиндецированных клеток Е. coli BL21(DE3), трансформированных плазмидами pBAD-klpBCDE и рЕТ28-KlpA. На представленных хроматограммах видно, что после индукции генетического кластера klpABCDE появляется продукция соединения со временем задерживания 23-24 минуты, и его спектр УФ-поглощения характерен для азолсодержащих пептидов (фигура 2.Б).
Очистку целевого пептида из культурального супернатанта осуществляют в две стадии: с помощью твердофазной экстракции на картриджах и при помощи обращенно-фазовой высокоэффективной жидкостной хроматографии (ОФ-ВЭЖХ) (фигура 3).
На первой стадии культуральный супернатант индуцированных клеток пропускают через картридж Waters Sep-Pak 12-сс Vac С18 (размер частиц 55-105 мкм), позволяя целевому пептиду связаться с сорбентом, после чего картридж интенсивно промывают 9% водным раствором ацетонитрила. Элюирование целевого пептида осуществляют 15%-ным водным раствором ацетонитрила и собирают в несколько фракций с последующим их ФП-МС-анализом. Фракции элюата, содержащие по данным ФП-МС-анализа клебсазолицин, подвергают дальнейшей очистке посредством ОФ-ВЭЖХ.
Очистку ОФ-ВЭЖХ проводят на полупрепаративной колонке Jupiter С18 HPLC column (300 ; 250×4.6; размер пор 5-μm), предварительно уравновешенной 0,1% водным раствором трифторуксусной кислоты (trifluoroacetic acid, TFA). Связавшийся с колонкой материал элюируют линейным градиентом ацетонитрила в воде при 0,1% TFA (от 0 до 30% ацетонитрила в течение 30 мин). Отобранные на этом этапе фракции оценивают посредством масс-спектрометрического анализа с преобразованием Фурье (ФП-МС). Показано, что вещество со временем задерживания 23-24 минуты имеет молекулярную массу 1 973,684 Да (фигура 4), что соответствует рассчитанной для клебсазолицина массе.
; 250×4.6; размер пор 5-μm), предварительно уравновешенной 0,1% водным раствором трифторуксусной кислоты (trifluoroacetic acid, TFA). Связавшийся с колонкой материал элюируют линейным градиентом ацетонитрила в воде при 0,1% TFA (от 0 до 30% ацетонитрила в течение 30 мин). Отобранные на этом этапе фракции оценивают посредством масс-спектрометрического анализа с преобразованием Фурье (ФП-МС). Показано, что вещество со временем задерживания 23-24 минуты имеет молекулярную массу 1 973,684 Да (фигура 4), что соответствует рассчитанной для клебсазолицина массе.
Образцы, содержащие чистый клебсазолицин, лиофилизируют, растворяют в ДМСО и храняти при -80°С.
Доказательство структуры
Использование тандемного масс-спектрометрического (МС-МС) анализа позволило установить аминокислотную последовательность данного соединения, а также дало информацию об остатках, участвующих в образовании азолов: Cys7, Cys10, Ser13 и Cys18 (фигура 5). Однако, модификация, приводящая к потере молекулы воды (-18 Da), вовлекающая первые две N-концевые аминокислоты (Ser1 и Gln2), не могла быть установлена с помощью масс-спектрометрии из-за отсутствия МС-МС фрагментации в этой области. Поэтому, для установления химической структуры клебсазолицина, мы использовали двумерный ядерный магнитный резонанс (ЯМР).
ЯМР-спектры регистрировали на растворенном в DMSO-D6 образце 5 мМ клебсазолицина, обогащенного изотопами 13С и 15N, находящегося в трубке Шигеми при 55°С. Все спектры ЯМР регистрировались на спектрометре Varian DirectDrive NMR System 700 МГц, снабженном 5-миллиметровым инверсным солеустойчивым датчиком тройного резонанса 1H/13C/15N. Данные ЯМР обрабатывали и анализировали с помощью Mestrelab Mnova версии 10.
Полное определение отдельных аминокислот, включая один пролин, четыре глицина, три аспарагина, три аланина, один лизин, пять серинов и один неканонический глутамин, а также три тиазола и один оксазол, получили из спектров 1Н, 13С, 1H-TOCSY, 1H-COZY, 13С-HSQC, 13С-НМВС, 13C-COZY, 13C-HSQCTOXY, 15N-HSQC, 15N-HSQCTOXY и HNCO (фигура 6).
Аланины, глицины, аспарагины, треонин и лейцин были определены из 1Н, 1H-TOCSY, 1H-COZY спектров 1Н-сигнала (фигура 6. А) и из 13С-сигнала спектра 13C-HSQC (фигура 6. Б). Определение серинов, пролина и глутамина сделали на основе 13C-HSQCTOXY спектра. Боковые цепи аспарагинов и глутамин были определены из 13С-НМВС спектра. Тиазолы и оксазол были установлены из 13C-HSQC, 13C-HSQCTOXY и 13С-НМВС спектров. Корреляции С5-Н5 были установлены из 13C-HSQC спектра (фигура 6. В). Сигналы С4, С6 и Oxz13 были определены из 13C-COZY (фигура 6. Г). Сигналы карбонильных атомов углерода установили из 13С-НМВС, 13C-COZY и Н-С проекций HNCO спектра. NH-группы были определены из спектров 15N-HSQC (фигура 6. Д) и 15N-HSQCTOXY спектров. Последовательность связей между различными аминокислотами была установлена на основе анализа наблюдаемых корреляций НМВС 1Н-13С сигналов HSQC (фигура 6. Е). Связь Pro4-Gly5 была определена из НМВС- корреляций Рго4-СО с Gly5-HN и Pro4-СО с Gly5-Hα. Связи Gly5-Asn6, Ala8-Ser9, Ser11-Asn12 и Thr19-Gly20 были установлены аналогичным образом. Связи между Ser15-Ala16 и Ala16-Asn17 была установлена через НМВС-корреляции Ser15-CO с Ala16-Нα и Ala16-СО с Asn17-Hα соответственно. Связь между Gly20 и Gly21 была установлена через НМВС-корреляции Gly20-CO с Gly21-HN. Связи Gly21-Leu22 и Leu22-Gly23 были определены аналогичным образом. Связть Asn6-Thz7 очевидна из НМВС- корреляций Thz7-C2 и Thz7-C4 с Asn6-Hα и Asn6-Hβ соответственно. Аналогично обнаружено соединение Ser9-Thz10 и Asn17-Thz18. Связь Oxz13-Ala14 очевидна из НМВС-корреляций Oxz13-С6 с Ala14-Нα и Ala14-HN. Аналогичным образом были установлены связи Ala8-Thz7, Thz10-Ser11 и Thz18-Thr19. Связь Ala12-Oxz13 очевидна из НМВС-корреляций Oxz13-C2 с Ala12-Нα и Ala12-Нβ. Связывание Ser1'-Glu2 и Glu2-Ser3 очевидно из НМВС- корреляций Glu2-CO с Ser1'-Нα, Ser3-Hα и Ser1'-HN, Ser3-HN (фигура 6. Ж).
Наблюдаемые две группы сигналов от Ser1, Gln2, Ser3 и Asn6, где совокупность состояний, связанных как 5/3, указывают на наличие таутомерного равновесия между двумя протонированными формами амидного фрагмента. Эти данные доказывают, что N-концевые аминокислотные остатки Ser1 и Gln2 образуют шестичленный гетероцикл через амидиновую связь.
Таким образом, показано, что экспрессия генетического кластера klpABCDE в клетках бактерий Е. coli BL21(DE3) приводит к продукции тиазол-оксазол модифицированного пептида, названного авторами клебсазолицин, и его химическая структура была полностью подтверждена, что означает что генетическая конструкция разработана верно и способ получения клебсазолицина обеспечивает возможность расширить структурный арсенала тиазол-оксазол модифицированных пептидов.


Способ получения порошкового композита на основе меди с улучшенными прочностными характеристиками
Преселективная коробка передач "ромашка-3ф" для трансмиссий транспортных средств.
Полуавтоматическое огнестрельное оружие "редуктор"
Способ формирования идентификационного кода информационно-защитной этикетки с заданным уровнем уникальности
Электромеханический двухпоточный привод транспортной машины с бортовым способом поворота
Способ фильтрации тока намагничивания и воспроизведения первичного напряжения измерительных двухобмоточных трансформаторов напряжения
Интеллектуальная система и способ измерения расхода двухфазного потока нефтяных скважин
Способ изготовления спеченных трубчатых изделий с буртом из порошка
Термомеханический силовой привод
Вибропоглощающий слоистый композитный металл-полимерный материал с использованием термопластичного эластомера на основе сополиуретанимида ган-р
Состав материала для изготовления электродов генераторов низкотемпературной плазмы
Семейство пептидов - ингибиторов активности белка reca, блокирующих sos-ответ у бактерий
Тиазол-оксазол-модифицированные пептиды, обладающие способностью ингибировать бактериальную рибосому
Способ получения оптического излучения
Нуклеаза pacas9
Средство разрезания днк на основе cas9 белка из defluviimonas sp.
Средство разрезания днк на основе cas9 белка из биотехнологически значимой бактерии clostridium cellulolyticum
Средство разрезания днк на основе cas9 белка из бактерии demequina sediminicola
Средство разрезания днк на основе cas9 белка из бактерии pasteurella pneumotropica
Применение cas9 белка из бактерии pasteurella pneumotropica для модификации геномной днк в клетках

