Результат интеллектуальной деятельности: СПОСОБ ПОЛУЧЕНИЯ САЙТ-НАПРАВЛЕННЫХ ТРАНСЛОКАЦИЙ В КЛЕТКАХ SACCHAROMYCES CEREVISIAE
Вид РИД
Изобретение
Область техники
Изобретение относится к области генетики и молекулярной биологии и касается способа получения транслокантов между любыми локусами, принадлежащими различным хромосомам, в клетках почкующихся дрожжей Saccharomyces cerevisiae. Предложенный способ может быть использован для моделирования хромосомных транслокаций в клетках эукариот в исследовательских и биотехнологических целях.
Уровень техники
В последнее время широкие возможности для редактирования и манипулирования геномами различных организмов связаны с РНК-зависимыми эндонуклеазами семейства CRISPR/Cas (Clustered Regularly Interspaced Short Palindromic Repeats). Эти белки являются сиквенс-специфичными защитными механизмами архей и бактерий против чужеродных плазмид и вирусов (Cell, 2016, 164: 29-44). Они встречаются у 50% бактерий и 90% архей. CRISPR представляют собой генетические локусы, включающие серии прямых повторов длиной 20-50 п.н. и разделяющие их уникальные спейсеры того же размера. Как оказалось, спейсеры происходят из вирусных геномов и коньюгативных плазмид, остающихся от соответствующих инфекций. Области CRISPR фланкированы cas генами. Cas-белки играют важную роль в работе указанных защитных систем. CRISPR/Cas системы можно разделить на шесть основных типов, причем наиболее важными являются первые три (Cell, 2016, 164: 29-44). Характерным маркером систем типа I являются Cas3 белки, несущие домены хеликазы и нуклеазы. Они разрезают чужеродную ДНК, в распознавании которой участвует мультибелковый комплекс Cascade (CRISPR-associated complex for antiviral defense). У систем типа II в разрушении чужеродной ДНК принимает участие единственный белок Cas9. Для систем типа III характерен Cas 10 белок, образующий Cascade-подобный комплекс для поиска и деградации чужеродных ДНК.
CRISPR РНК (crRNA) вместе с транс-активирующими crRNA (tracrRNA), образуют комплекс с Cas-белками для определения специфичности разрезания через Уотсон-Криковские взаимодействия между нуклеиновыми кислотами. Для наиболее простой системы типа II при формировании эффективного нуклеазного комплекса необходимы три компонента: Cas9 белок, зрелая crRNA и tracrRNA. Более того, было установлено, что crRNA и tracrRNA могут быть объеденены в единую gRNA (guide RNA) молекулу, и, модифицируя ее последоваетельность в определенном месте, можно разрезать новые сайты ДНК (Mol. Ther., 2016, doi: 10.1038/mt. 2016.10). Данная упрощенная CRISPR/Cas система получила широкое распространение для технологий редактирования генома через удаление/модификацию не желательных последовательностей ДНК (Mol. Ther., 2016, doi: 10.1038/mt.2016.10). На ее основе также были запатентованы методы получения транслокантов в геномах эукариот. Суть методов заключается в ведении gRNA-зависимого двуцепочечного разрыва в гены эукариот. Так как репарация разрыва осуществляется путем гомологичной рекомбинации (HR) или соединением не гомологичных концов (NHEJ) в результате могут образовываться транслокантные хромосомы, которые затем отбираются и подтверждаются методом ПЦР с последующим секвенированием. Недостатком этих методов является непредсказуемость выхода получаемых транслокантов, так как разрезанные CRISPR/Cas системами хромосомы могут репарироваться и рекомбинировать многими способами, выход продуктов репарации гетерогенен, и, таким образом, трудно получить сайт-специфичные транслокации. Кроме того, подготовка gRNA для разрезания новых генов достаточно трудоемка и включает мутагенез, этапы клонирования и секвенирования.
Этого недостатка лишены подходы, в которых транслокации происходят между короткими гомологичными последовательностями, расположенными в различных участках генома (Nature, 2009, 460(7258):984-9; PloS Genetics, 2011, 7(5): е1002089). На основе предложенных систем была установлена роль многих клеточных факторов, включая, например, SGS1, ТОР3, RMI1, SRS2, RAD6, SLX1, SLX4, SLX5, MSH2, MSH6, RAD10, MRC1 и TOF1 в поддержании стабильности генома. Однако в этом случае сайты транслокаций ограничены расположением гомологичных последовательностей.
Известна методика мост-индуцируемых транслокаций (Chromosoma, 2005, 114(1): 15-27; PNAS, 2008, 105(28): 2703-8; Genetics, 2010, 186(3): 775-90), заключающаяся в интеграции гена устойчивости к канамицину в локусы, принадлежащие разным хромосомам в клетках почкующихся дрожжей S. cerevisiae с образованием транслокантной хромосомы. По данной методике можно получать транслоканты между любыми участками генома, которые определяются 5'- и 3'-концами используемых кассет для интеграции. Суть метода заключается в проведении литиево-ацетатной трансформации клеток S. cerevisiae кассетой, несущей ген устойчивости к канамицину, фланкированный короткими (30-65 п.н.) участками гомологии, принадлежащими разным хромосомам; отбора транслокантов по устойчивости к канамицину и проверки правильности интеграции кассеты с помощью ПЦР и секвенирования полученного ПЦР-продукта. Минусами данного метода является не возможность отрицательной селекции, что затрудняет отслеживание дальнейшей судьбы транслокантной хромосомы и последующие генетические манипуляции с маркером устойчивости к канамицину; а также не возможность повторного использования этого маркера (гена устойчивости к канамицину) для генетических манипуляций с клетками транслокантов. Следует учесть, что перечень используемых селективных маркеров для клеток S. cerevisiae очень ограничен. Эти недостатки преодолевает предлагаемый новый подход, сохраняя основное преимущество метода мост-индуцируемых транслокаций - возможность осуществления транслокаций между любыми локусами генома по желанию.
Раскрытие изобретения
Задачей настоящего изобретения является создание способа для осуществления транслокаций между любыми двумя геномными локусами в клетках почкующихся дрожжей S. cerevisiae с последующей возможностью как положительной, так и отрицательной селекции транслокантных хромосом.
Остается актуальной задача получения генетического маркера транслокантной хромосомы, позволяющего проводить как положительную (выживают клетки, несущие этот маркер), так и отрицательную селекцию (клетки, несущие этот маркер, погибают); а также различных генетических манипуляций с ним с отбором продуктов манипуляций с помощью отрицательной селекции; в клетках транслокантов должна сохраняться возможность использования маркера устойчивости к канамицину для генетических манипуляций, в том числе новых мост-индуцируемых транслокаций.
Поставленная задача решается способом получения сайт-направленных транслокаций в клетках Saccharomyces cerevisiae, заключающимся в трансформации клеток ura3Δ0/ura3Δ0 штамма почкующихся дрожжей S.cerevisiae (Yeast, 1998, 14(2): 115-32) кассетой для транслокации, содержащей ген URA3, фланкированный последовательностями ДНК длиной более 30 п.н. с гомологией к произвольным локусам разных хромосом. Далее высевают клетки на селективные среды без урацила, выращивают в течение ночи, с последующим отбором выросших колоний и проведением ПЦР-реакции для подтверждения правильности транслокации. Предпочтительно трансформацию клеток проводить литиево-ацетатным методом.
Техническим результатом изобретения является получение заявляемым способом транслокантной хромосомы, состоящих из двух частей, принадлежащих до транслокации различным хромосомам, которая пригодна для повторного использования, а также позволяет проводить как положительную, так и отрицательную селекцию. Также полученная транслокантная хромосома позволяет проводить различные генетические манипуляции с отбором продуктов манипуляций с помощью отрицательной селекции. В клетках транслокантов сохраняется возможность использования маркера устойчивости к канамицину для генетических манипуляций, в том числе новых мост-индуцируемых траснлокаций между другими хромосомами, которых в диплоидных клетках S. cerevisiae 16 пар.
По сравнению с начальной методикой предлагаемый способ обладает следующими преимуществами:
1) Ген URA3 позволяет проводить как положительную, так и отрицательную селекцию: в присутствии 5-флуорооротатной кислоты клетки с этим геном погибают, что позволяет отслеживать судьбу транслокантной хромосомы;
2) Предложенное изобретение позволяет использовать ген устойчивости к канамицину для дополнительных манипуляций с генами и хромосомами, включая траснлокаций;
3) Присутствие уникального гена URA3 с возможностью отрицательной селекции позволяет интегрировать в транслокантную хромосому дополнительные гены и/или генетические элементы и модифицировать место транслокации.
Краткое описание чертежей
Изобретение поясняется чертежами.
На фиг. 1 показана принципиальная схема сайт-направленных транслокаций. Клетки ura3Δ0/ura3Δ0 S.cerevisiae (Yeast, 1998, 14(2): 115-32) трансформируются кассетой для транслокации. Области кассеты для транслокации, гомологичные двум разным хромосомам выделены пунктиром различной длины.
На фиг. 2 представлена фотография клеток транслокантов, окрашенных FITC-фаллоидином для визуализации цитоскелета.
Осуществление изобретения
Все используемые реагенты являются коммерчески доступными, все процедуры, если не оговорено особо, осуществляли при комнатной температуре или температуре окружающей среды, то есть в диапазоне от 18 до 25°C.
Для выращивания колоний ura3Δ0/ura3Δ0 штамма S.cerevisiae возможно использование любых питательных сред, жидких или агаризованных, поддерживающих рост этих клеток. При селекции транслокантных клеток, несущих интегрированный ген URA3, также возможно использование любых жидких или агаризованных питательных сред, принципиальным требованием к ним является отсутствие урацила.
Для получения кассеты для трансформации должны использоваться праймеры, обладающие гомологией не менее 30 п. н. к двум различным хромосомам и не менее 25 п.н - к гену URA3 (фиг. 1).
Для проведения отрицательной селекции необходимо использовать в селективной среде для отбора желаемых мутаций/делеций/инсерций 5-флуорооротатную кислоту: клетки с геном URA3 погибают в ее присутствии (Methods Enzymol, 1987, 154: 164-75), а выживут лишь колонии клеток, в которых этот ген был инактивирован в результате генетических манипуляций.
Приведенные примеры конкретного осуществления изобретения приведены для предоставления специалистам в данной области техники полного описания проведения и применения анализа по изобретению, и подразумевают, что приведенные примеры не ограничивают предполагаемый авторами изобретения объем изобретения.
1. Провести литиево-ацетатную трансформацию (PNAS, 2008, 105(28): 2703-8) ura3Δ0/ura3Δ0 штамма почкующихся дрожжей кассетой для транслокации, содержащей ген URA3, фланкированный последовательностями ДНК длиной более 30 п. н. с гомологией к произвольным локусам разных хромосом (фиг. 1).
2. После появления колоний провести проверку правильности интеграции кассеты с помощью ПЦР и секвенирования полученного ПЦР-продукта. Фотография клеток транслокантов приведена на фиг. 2.
Принципиально важной стадией изобретения является трансформация ura3Δ0/ura3Δ0 клеток S.cerevisiae кассетой для транслокации с геном URA3, обладающей областями гомологии более 30 п.н двум разным хромосомам. Районы гомологии определяют место интеграции кассеты (фиг. 1). Транслоканты, несущие ген URA3, способны расти на среде без урацила, что позволяет провести селективный отбор этих клеток. Однако в присутствии 5-флуорооротатной кислоты клетки с этим геном погибают, что позволяет проводить отрицательную селекцию. Последняя возможность может быть важна для осуществления генетических манипуляций в точке транслокации: инсерции/делеции последовательностей ДНК, а также ее точечного мутагенеза.
Модифицируемым параметром изобретения являются 5'- и 3'-концы последовательности кассеты для транслокации, которые могут быть гомологичными любым двум локусам в геноме, но принадлежать двум разным хромосомам. Диплоидные клетки S.cerevisiae обладают 16 парами хромосом.
В качестве примера осуществления изобретения были проведены следующие действия:
1. Инокулировали свежую колонию ura3Δ0/ura3Δ0 штамма S.cerevisiae в 2 мл питательной среды без урацила. Растили в течение ночи при 30°C.
2. Инокулировали 200 μl ночной культуры в 50 мл той же питательной среды. Растили культуру до OD600=0.7-0.8 (~7 ч). Собрали клетки центрифугированием при 2-3 g.
3. Ресуспендировали клетки в 5 мл раствора, содержащего 100 мМ трис-HCl, рН 7.5; 50 мМ ЭДТА, рН 8.0 и 100 мМ ацетата лития. Собрали клетки центрифугированием при 2-3g.
4. Ресуспендировали клетки в 0.5 мл того же раствора, перенесли в пластиковую пробирку с закрывающейся крышкой. Инкубировали 60 мин при комнатной температуре на шейкере с перемешиванием.
5. Смешали 10 μл 10 мг/мл ДНК из спермы лосося, ~5-10 μг в ~10 μл ДНК кассеты для транслокации и 200 μл клеток. Инкубировали в течение 30 мин при комнатной температуре на шейкере с перемешиванием. Кассету для транслокации получали заранее с помощью ПЦР, используя в качестве матрицы ДНК штаммов S.cerevisiae с геном URA3, как, например, S288C, и праймеры с последовательностями SEQ ID NO:1 и SEQ ID NO:2.
Реакция ПЦР включала следующие стадии:
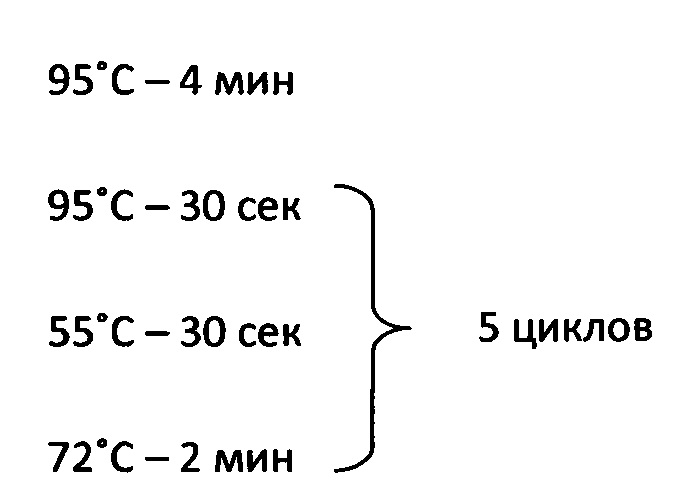
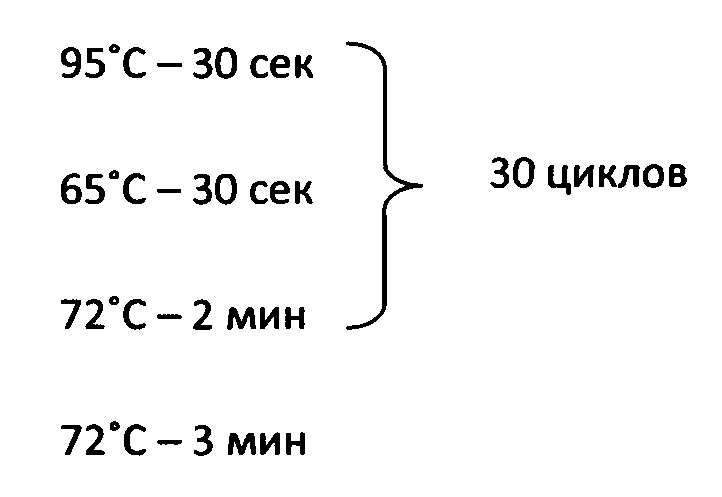
Последовательность используемой кассеты приведена в SEQ ID NO:3.
6. Добавили 1 мл раствора, содержащего 100 мМ трис-HCl, рН 7.5; 50 мМ ЭДТА, рН 8.0; 100 мМ ацетата лития и ПЭГ 3500. Инкубировали 10 мин при 42°C. Собрали клетки центрифугированием при 2-3 g.
7. Удалили все следы супенатанта с помощью пипетки. Ресуспендировали клетки в 200 ил раствора, содержащего 100 мМ трис-HCl, рН 7.5 и 50 мМ ЭДТА, рН 8.0, и высеяли клетки на селективную среду без урацила.
8. После появления колоний провели проверку правильности интеграции кассеты с помощью ПЦР и секвенирования полученных ПЦР-продуктов. Фотография клеток транслокантов приведена на фиг. 2.
В результате предлагаемого способа были получены транслоканты с интегрированным геном URA3.






