Результат интеллектуальной деятельности: МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
Вид РИД
Изобретение
Область изобретения
Данное изобретение относится к антителам и гибридным полипептидам с Fc-областью, которые асимметрично модифицированы в отношении их взаимодействия с Fc-рецептором, особенно с FcRn, а также к способам их применения.
Уровень техники
Почти все Fc-рецепторы связываются с симметричной Fc-областью антител асимметрично.
Например, человеческий Fcγ-рецептор IIIA взаимодействует с различными аминокислотами остатками на двух полипептидных цепях Fc-области. Таким образом, асимметрично введенные мутации (например, в нижней шарнирной области на остатках с 233 по 238) могут быть использованы для увеличения или уменьшения взаимодействия антитела с человеческим Fcγ-рецептором IIIA.
Однако взаимодействие между человеческим неонатальным Fc-рецептором FcRn является симметричным: две FcRn-молекулы могут связываться с одним IgG со стехиометрией 2:1 (см., например, Huber, A.W., et al., J. Mol. Biol. 230 (1993) 1077-1083). Таким образом, асимметрично введенные мутации снижают связывание с одним FcRn, но не с обоими.
Примеры асимметричных IgG-подобных молекул включают, но не ограничиваясь ими, такие, которые получены с использованием следующих технологий или с использованием следующих форматов: Triomab/Quadroma, "ключ в замке", Crosstab, электростатически подобранные антитела, LUZ-Y, SEEDbody (Strand Exchange Engineered Domain body), Biclonic и DuoBody.
В WO 2012/125850 сообщается об Fc-содержащих белках, имеющих асимметричные замены в их Fc-областях и демонстрирующих повышенное связывание с человеческим Fcγ-рецептором IIIA и повышенную ADCC-активность.
В WO 2012/58768 сообщается об изолированных гетеромногомерах, содержащих гетеродимерную Fc-область, где эта гетеродимерная Fc-область содержит вариантный СН3-домен, содержащий аминокислотные мутации для облегчения формирования гетеродимера с повышенной стабильностью, и где гетеродимерная Fc-область также содержит вариантный СН2-домен, содержащий асимметричные аминокислотные модификации для облегчения селективного связывания с рецептором Fc-гамма.
В WO 2011/131746 сообщается, что путем введения асимметричных мутаций в СН3-области двух моноспецифических исходных белков можно усилить реакцию обмена Fab-ветвями, что она становится направленной и, таким образом, дает гетеродимерные белки с высокой стабильностью.
Kim et al. (Kim, H., et al., Invest. Ophthalmol. Vis. Sci. 49 (2008) 2025-2029) сообщают, что в ткани глаза, за исключением ретинального пигментного эпителия и ткани хориоидеи, показано наличие транскрипта FcRn предсказанного размера, в том числе в цилиарном теле и радужке, сетчатке, конъюнктиве, роговице, хрусталике и пучке зрительного нерва. Показана экспрессия рецептора FcRn в гематоофтальмическом барьере, что указывает на то, что транспорт IgG из тканей глаза в кровеносную систему может использовать этот рецептор. Поскольку внутренние глазные ткани, такие как сетчатка, отделены от системы крови гематоофтальмическим барьером, обнаружение полноразмерного антитела в системе крови в течение короткого времени после интравитреальной инъекции не ожидалось. Тем не менее, недавние фармакокинетические данные, полученные на обезьянах и людях, показали, что интравитреально введенный бевацизумаб появляется в крови в течение нескольких часов после интравитреальной инъекции. Таким образом, может оказаться, что функция рецептора FcRn в конъюнктивальных лимфатических сосудах заключается в действии его в качестве рецептора эффлюкса (оттока) для эффективного устранения комплексов "антиген-антитело IgG" из конъюнктивального пространства. Несмотря на схожий молекулярный вес IgG (150 кДа) был обнаружен в водянистой влаге, тем не менее, IgA (160 кДа) обнаружен не был. Различие в проникновении IgG и IgA из сыворотки в водянистую влагу можно объяснить присутствием FcRn-рецепторов, которые являются селективными для IgG.
Kim et al. также сообщали (Kim, Н., et al., Mol. Vis. 15 (2009) 2803-2812), что прямая интравитреальная инъекция стала стандартным подходом для доставки терапевтических антител в задний сегмент глаза при нарушениях сетчатки. Интравитреально введенный бевацизумаб (IgG) и куриный IgY преодолевали барьер внутренней пограничной мембраны и диффундировали в более глубокие структуры сетчатки. После диффузии через сетчатку бевацизумаб пересекал барьер "кровь - сетчатка" и проходил в большой круг кровообращения. Интраретинальный куриный IgY был локализован только вдоль аблюминальной стороны барьера "кровь - сетчатка". Кроме того, кровеносные сосуды хориоидеи были отрицательными на наличие куриного IgY. После интраретинального введения бевацизумаба были обнаружены его физиологически релевантные сывороточные уровни, представляющие до 30% от введенной дозы. Это говорит о более высоком риске системных побочных эффектов, чем считалось ранее. Гематоофтальмический барьер обнаруживает специфический механизм транспортировки и выведения полноразмерных IgG в системный кровоток. Данное исследование Kim подтверждает гипотезу о том, что этот механизм является неонатальным Fc-рецептором.
В US 2011/054151 сообщается о композициях и способах одновременного двухвалентного и одновалентного совместного взаимодействия антигенов.
В US 2011/236388 сообщается о биспецифических двухвалентных анти-VEGF/анти-ANG-2-антителах.
В WO 2010/121766 сообщается о гибридных белках-антителах с измененными сайтами связывания FcRn.
Kim, J.K., et al. сообщают о картировании на человеческом IgG сайта связывания рецептора, связанного с МНС класса I, FcRn (Eur. J. Immunol. 29 (1999) 2819-2825).
Qiao, S.-W., et al. сообщают о зависимости опосредованной антителом презентации антигена на FcRn (Proc. Natl. Acad. Sci. USA 105 (2008) 9337-9342).
Kuo, T.T., et al. сообщают о неонатальном Fc-рецепторе: от иммунитета к лечению (J. Clin. Immunol. 30 (2010) 777-789).
Firan, М., et al. сообщают, что рецептор, связанный с МНС класса I, FcRn, играет существенную роль в передаче у людей гамма-глобулина от матери плоду (Int. Immunol. 13 (2001) 993-1002).
Vidarsson, G., et al. сообщают, что FcRn является рецептором IgG на фагоцитах с новой ролью в фагоцитозе (Blood 108 (2006) 3573-3579).
Gillies, S.D., et al. сообщают об улучшении эффективности гибридных белков "антитело - интерлейкин 2" путем снижения их взаимодействия с Fc-рецепторами (Cancer Res. 59 (1999) 2159-2166).
В WO 2013/060867 сообщается о продукции гетеродимерных белков.
Сущность изобретения
Обнаружили, что FcRn-связывание антитела или гибридного полипептида Fc-области может быть модифицировано путем изменения аминокислотных остатков в несоответствующих позициях в отдельных полипептидах Fc-области, поскольку эти изменения действуют вместе, модифицируя FcRn-связывание. Антитела и гибридные полипептиды Fc-области, описанные в данном документе, могут быть использованы, например, для лечения заболеваний, при которых требуется индивидуальное время системного удерживания.
Одним из аспектов, описанных в данном документе, является вариантная (человеческая) Fc-область класса IgG, содержащая первый полипептид Fc-области и второй полипептид Fc-области,
где
a) первый полипептид Fc-области и второй полипептид Fc-области получены из одного и того же родительского (человеческого) полипептида Fc-области класса IgG, и
b) первый полипептид Fc-области имеет аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat,
где вариантная (человеческая) Fc-область класса IgG имеет аффинность к человеческому Fc-рецептору, отличающуюся от (человеческой) Fc-области класса IgG, которая имеет те же аминокислотные остатки (как в (родительском) человеческом полипептиде Fc-области из а)) в соответствующих позициях в соответствии с индексом ЕС по системе нумерации Kabat в первом полипептиде Fc-области и втором полипептиде Fc-области.
Одним из аспектов, описанных в данном документе, является вариантная (человеческая) Fc-область класса IgG, содержащая первый полипептид Fc-области и второй полипептид Fc-области,
где
а) первый полипептид Fc-области имеет аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat,
где вариантная (человеческая) Fc-область класса IgG имеет аффинность к человеческому Fc-рецептору, отличающуюся от Fc-области класса IgG, которая имеет те же аминокислотные остатки (как в соответствующей человеческой Fc-области) в первом и втором полипептиде Fc-области в соответствующих позициях.
Одним из аспектов, описанных в данном документе, является вариантная (человеческая) Fc-область класса IgG, содержащая первый полипептид Fc-области и второй полипептид Fc-области,
где
a) аминокислотная последовательность первого полипептида Fc-области отличается от аминокислотной последовательности первого родительского полипептида Fc-области по меньшей мере одним или более чем одним аминокислотным остатком,
и
аминокислотная последовательность второго полипептида Fc-области отличается от аминокислотной последовательности второго родительского полипептида Fc-области по меньшей мере одним или более чем одним аминокислотным остатком, и
b) первый полипептид Fc-области имеет аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat,
где вариантная (человеческая) Fc-область класса IgG имеет аффинность к человеческому Fc-рецептору, отличающуюся от родительской Fc-области класса IgG, содержащей первый и второй родительский полипептид Fc-области класса IgG из а).
Одним из аспектов, описанных в данном документе, является вариантная (человеческая) Fc-область класса IgG, содержащая первый полипептид Fc-области и второй полипептид Fc-области,
где
a) аминокислотная полипептида Fc последовательность первого полипептида Fc-области получена из первого родительского полипептида Fc-области, а аминокислотная последовательность второго полипептида Fc-области получена из второго родительского полипептида Fc-области, и
b) в первом полипептиде Fc-области и/или во втором полипептиде Fc-области одна или более чем одна мутация введены так, что первый полипептид Fc-области имеет аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat,
где вариантная (человеческая) Fc-область класса IgG имеет аффинность к человеческому Fc-рецептору, отличающуюся от Fc-области класса IgG, содержащей первый и второй родительские полипептиды Fc-области класса IgG из а).
В одном воплощении всех аспектов вариантная (человеческая) Fc-область класса IgG представляет собой вариантную (человеческую) гетеродимерную Fc-область класса IgG.
В одном воплощении всех аспектов первый родительский полипептид Fc-области класса IgG и второй родительский полипептид Fc-области класса IgG являются нечеловеческими полипептидами Fc-области класса IgG.
В одном воплощении всех аспектов первый родительский полипептид Fc-области класса IgG и второй родительский полипептид Fc-области класса IgG являются одним и тем те же полипептидом Fc-области класса IgG.
В одном воплощении всех аспектов спаривание первого полипептида Fc-области и второго полипептида Fc-области с образованием димерной (функциональной) Fc-области приводит к формированию гетеродимера.
В одном воплощении всех аспектов первый и второй полипептиды Fc-области отличаются независимо друг от друга по меньшей мере на один аминокислотный остаток от соответствующего родительского полипептида Fc-области класса IgG.
В одном воплощении всех аспектов класс IgG выбран среди подклассов IgG1, IgG2, IgG3n IgG4.
В одном воплощении всех аспектов человеческий Fc-рецептор выбран среди человеческого неонатального Fc-рецептора и человеческого Fcy-рецептора.
В одном воплощении всех аспектов первый полипептид Fc-области отличается от второго полипептида Fc-области на 1, или 2, или 3, или 4, или 5, или 6, или 7, или 8, или 9, или 10, или 11, или 12 аминокислотных остатков в соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat.
В одном воплощении вариантная (человеческая) Fc-область класса IgG имеет сниженное связывание со стафилококковым белком А, чем соответствующая родительская человеческая Fc-область класса IgG.
В одном воплощении вариантная (человеческая) Fc-область класса IgG имеет такое же связывание со стафилококковым белком А, как соответствующая родительская человеческая Fc-область класса IgG.
В одном воплощении всех аспектов вариантная (человеческая) Fc-область класса IgG содержит первый и второй полипептид Fc-области как человеческого IgG1, так и человеческого IgG4 (человеческого происхождения), которые содержат одну или две мутации, выбранные из I) группы I253A, Н310А и Н435А, или ii) группы Н310А, Н433А и Y436A, или iii) группы L251D, L314D и L432D, или
iv) группы L251S, L314S и L432S (нумерация согласно индексу ЕС по системе нумерации Kabat) в первом полипептиде Fc-области, и одну или две мутации, выбранные из группы, включающей мутации L251D, L251S, I253A, Н310А, L314D, L314S, L432D, L432S, Н433А, Н435А и Y436A (нумерация согласно индексу ЕС по системе нумерации Kabat) во втором полипептиде Fc-области, так что все мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A, или iii) L251D, L314D и L432D, или iv) L251S, L314S и L432S содержатся в вариантной (человеческой) Fc-области класса IgG.
В одном воплощении всех аспектов вариантная (человеческая) Fc-область класса IgG содержит первый и второй полипептид Fc-области как человеческого IgG1, так и человеческого IgG4 (человеческого происхождения), которые содержат мутации I253A/H310A/H435A, или H310A/H433A/Y436A, или L251D/L314D/L432D, или L251S/L314S/L432S, или их комбинации в Fc-области (нумерация согласно индексу ЕС по системе нумерации Kabat), где i) все мутации находятся в первом или втором полипептиде Fc-области, или ii) одна или две мутации находятся в первом полипептиде Fc-области, и одна или две мутации находятся во втором полипептиде Fc-области, так что все мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A, или iii) L251D, L314D и L432D, или iv) L251S, L314S и L432S находятся в Fc-области.
В одном воплощении всех аспектов вариантная (человеческая) Fc-область класса IgG содержит первый и второй полипептид Fc-области человеческого подкласса IgG1, где
a) первый и второй полипептиды Fc-области также содержат мутации L234A и L235A (нумерация согласно индексу ЕС по системе нумерации Kabat), или
b) первый и второй полипептиды Fc-области также содержат мутацию P329G (нумерация согласно индексу ЕС по системе нумерации Kabat), или
c) первый и второй полипептиды Fc-области также содержат мутации L234A и L235A и P329G (нумерация согласно индексу ЕС по системе нумерации Kabat), или
d) первый и второй полипептиды Fc-области также содержат мутации L234A и L235A (нумерация согласно индексу ЕС по системе нумерации Kabat), и первый полипептид Fc-области также содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области также содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V, или
e) первый и второй полипептиды Fc-области также содержат мутации L234A и L235A и P329G (нумерация согласно индексу ЕС по системе нумерации Kabat), и первый полипептид Fc-области также содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области также содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V.
В одном воплощении всех аспектов вариантная (человеческая) Fc-область класса IgG содержит первый и второй полипептид Fc-области человеческого подкласса IgG4, где
a) первый и второй полипептиды Fc-области также содержат мутации S228P и L235E (нумерация согласно индексу ЕС по системе нумерации Kabat), или
b) первый и второй полипептиды Fc-области также содержат мутацию P329G (нумерация согласно индексу ЕС по системе нумерации Kabat), или
c) первый и второй полипептиды Fc-области также содержат мутации S228P и L235A и P329G (нумерация согласно индексу ЕС по системе нумерации Kabat), или
d) первый и второй полипептиды Fc-области также содержат мутации S228P и L235A (нумерация согласно индексу ЕС по системе нумерации Kabat), и первый полипептид Fc-области также содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области также содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V, или
e) первый и второй полипептиды Fc-области также содержат мутации S228P и L235A и P329G (нумерация согласно индексу ЕС по системе нумерации Kabat), и первый полипептид Fc-области также содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области также содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V.
Один из аспектов, описанных в данном документе, представляет собой антитело или гибридный полипептид Fc-области, содержащий вариантную (человеческую) Fc-область класса IgG, описанную в данном документе.
В одном воплощении антитело представляет собой моноклональное антитело.
В одном воплощении антитело является человеческим, гуманизированным или химерным антителом.
Один из аспектов, описанных в данном документе, представляет собой нуклеиновую кислоту, кодирующую вариантную (человеческую) Fc-область класса IgG, описанную в данном документе.
Один из аспектов, описанных в данном документе, представляет собой нуклеиновую кислоту, кодирующую антитело, описанное в данном документе.
Один из аспектов, описанных в данном документе, представляет собой нуклеиновую кислоту, кодирующую гибридный полипептид Fc-области, описанный в данном документе.
Один из аспектов, описанных в данном документе, представляет собой клетку-хозяина, содержащую нуклеиновую кислоту, описанную в данном документе.
Один из аспектов, описанных в данном документе, представляет собой способ получения вариантной (человеческой) Fc-области IgG класса, о которой сообщается в данном документе, включающий культивирование клетки-хозяина, о которой сообщается в данном документе, так что вариантная (человеческая) Fc-область класса IgG продуцируется.
Один из аспектов, описанных в данном документе, представляет собой способ получения антитела, описанного в данном документе, включающий культивирование клетки-хозяина, описанной в данном документе, так что это антитело продуцируется.
Один из аспектов, описанных в данном документе, представляет собой способ получения гибридного полипептида Fc-области, описанного в данном документе, включающий культивирование клетки-хозяина, описанной в данном документе, так что гибридный полипептид Fc-области продуцируется.
Один из аспектов, описанных в данном документе, представляет собой фармацевтический состав, содержащий вариантную (человеческую) Fc-область класса IgG, описанную в данном документе, или антитело, описанное в данном документе, или гибридный полипептид Fc-области, описанный в данном документе.
Один из аспектов, описанных в данном документе, представляет собой вариантную (человеческую) Fc-область класса IgG, описанную в данном документе, или антитело, описанное в данном документе, или гибридный полипептид Fc-области, описанный в данном документе, для применения в качестве лекарственного средства.
Один из аспектов, описанных в данном документе, представляет собой применение вариантной (человеческой) Fc-области класса IgG, описанной в данном документе, или антитела, описанного в данном документе, или гибридного полипептида Fc-области, описанного в данном документе, в производстве лекарственного средства.
Антитела, описанные в данном документе, могут быть использованы, например, в качестве молекул, привлекающих Т-клетки, в качестве молекул, связывающих Fc-гамма-рецептор, с высокой биологической активностью (силой) и быстрым выведением из кровотока (сыворотки крови), в качестве конъюгатов "антитело - лекарственный агент" - с быстрым выведением в целях сокращения системных побочных эффектов, или в качестве предварительно нацеленных антител.
Краткое описание графических материалов
Фиг. 1 Схема концепции и преимущества антител <VEGF-ANG-2> IgG1 или IgG4 с мутацией IHH-AAA (= комбинация мутаций I253A, Н310А и Н435А (нумерация согласно индексу ЕС по Kabat)).
Фиг. 2 Маломасштабное измерение вязкости на основе DLS:
Экстраполированная вязкость при 150 мг/мл в 200 мМ растворе аргинина/сукцината, рН 5,5 (сравнение <VEGF-ANG-2> антител VEGFang2-0016 (с мутацией IHH-AAA) с референсным антителом VEGFang2-0015 (без таких мутаций IHH-AAA)).
Фиг. 3 Агрегация DLS в зависимости от температуры (в том числе температура начала агрегации DLS) в 20 мМ гистидиновом буфере, 140 мМ NaCl, рН 6,0 (сравнение описанных в данном документе <VEGF-ANG-2> антител VEGFang2-0016 (с мутацией IHH-AAA) с референсным антителом VEGFang2-0015 (без такой мутации IHH-AAA)).
Фиг. 4 Семидневное хранение при 40°C при концентрации 100 мг/мл (снижение основного пика и увеличение высокомолекулярной массы (HMW)) (сравнение описанных в данном документе <VEGF-ANG-2> антител VEGFang2-0016 (с мутацией IHH-AAA), которые продемонстрировали более низкую агрегацию, с референсным антителом VEGFang2-0015 (без такой мутации IHH-AAA)).
Фиг. 5А и В Состояние устойчивой аффинности к FcRn в A: VEGFang2-0015 (без мутации IHH-AAA) и в В: VEGFang2-0016 (с мутацией IHH-AAA).
Фиг. 6 Измерение взаимодействия с Fc-гамма RIIIa антител VEGFang2-0015 без мутации IHH-AAA и VEGFang2-0016 с мутацией IHH-AAA (оба относятся к подклассу IgG1 с мутациями P329G LALA; в качестве контролей использовали антидигоксигениновое антитело (анти-Dig) подкласса IgG1 и антитело на основе IgG4.
Фиг. 7А Схематический принцип фармакокинетического (PK) анализа ELISA для определения концентраций биспецифических <VEGF-ANG-2> антител в сыворотке крови и цельных глазных лизатах.
Фиг. 7В Сывороточная концентрация после внутривенного (iv) применения: сравнение VEGFang2-0015 без мутации IHH-AAA и VEGFang2-0016 с мутацией IHH-AAA.
Фиг. 7С Сывороточная концентрация после интравитреального применения: сравнение VEGFang2-0015 без мутации IHH-AAA и VEGFang2-0016 с мутацией IHH-AAA.
Фиг. 7D Концентрация VEGFang2-0016 (с мутацией IHH-AAA) в лизатах правого и левого глаз (после интравитреального введения только в правый глаз в сравнении с внутривенным введением): после интравитреального введения значимые концентрации могут быть обнаружены только в правом глазу; после внутривенного введения концентрация в глазных лизатах не могли не быть обнаружены из-за короткого периода полужизни VEGFang2-0016 (с мутацией IHH-AAA) в сыворотке.
Фиг. 7Е Концентрация VEGFang2-0015 (без мутации IHH-AAA) в лизатах правого и левого глаз (после интравитреального введения только в правом глазу в сравнении с внутривенным введением): в правом глазу (и в какой-то степени в левом глазу) после интравитреального введения концентрации VEGFang2-0015 могут быть обнаружены; это указывает на диффузию из правого глаза в сыворотку крови и оттуда в левый глаз, что может быть объяснено длительным периодом полужизни VEGFang2-0015 (без мутации IHH-AAA); после внутривенного введения также значительные концентрации могут быть обнаружены в лизатах обоих глаз в результате диффузии в глаза VEGFang2-0015 (без мутации IHH-AAA), который стабилен в сыворотке.
Фиг. 8 Антитела, разработанные в отношении их способности связывать FcRn, демонстрируют удлиненный (мутация YTE) или укороченный (мутация IHH-AAA) период полужизни in vivo, усиленное (мутация YTE) или уменьшенное (мутация IHH-AAA) связывание по сравнению с референсным антителом дикого типа (wild-type, wt) в анализе SPR, а также повышенное или сниженное время удерживания в FcRn-колоночной хроматографии; а) PK-данные после одного внутривенного болюсного введения 10 мг/кг самцам huFcRn-трансгенных мышей C57BL/6J +/- 276: данные AUC для IgG дикого типа, а также YTE и IHH-AAA Fc-модифицированные IgG; b) сенсограмма BIAcore; с) элюирование с колонки с FcRn-аффинностью; анти-IGF-1R-антитело дикого типа (референсное), YTE-мутант анти-IGF-IR-антитела, IHH-AAA-мутант анти-IGF-IR-антитела.
Фиг. 9 Изменение времени удерживания в FcRn-аффинной хроматографии в зависимости от числа мутаций, введенных в Fc-область.
Фиг. 10 Изменение FcRn-связывания в зависимости от асимметричного распределения мутаций, введенных в Fc-область.
Фиг. 11 Элютивная хроматограмма биспецифического <VEGF-ANG-2> антитела (VEGF/ANG2-0) с мутациями Н310А, Н433А и Y436A в обеих тяжелых цепях из двух последовательных колонок для аффинной хроматографии с белком А.
Фиг. 12 Элютивная хроматограмма анти-IGF-1R-антитела (IGF-1R-0045) с мутациями Н310А, Н433А и Y436A в обеих тяжелых цепях из колонки для аффинной хроматографии с белком А.
Фиг. 13 Связывание <VEGF-ANG-2> антител, имеющих модифицированную Fc-область IgG, с иммобилизованным белком А на чипе CIV15.
Фиг. 14 Элютивная хроматограмма различных <VEGF-ANG-2> антител на FcRn-аффинной колонке.
Фиг. 15 Связывание различных гибридных полипептидов со стафилококковым белком A (SPR).
Фиг. 16 Связывание различных мутантов <VEGF-ANG-2> антитела и анти-IGF-IR-антитела с иммобилизованным белком A (SPR).
Подробное описание воплощений изобретения
I. Определения
Термин "примерно" обозначает диапазон +/- 20% от следующего далее числового значения. В одном воплощении термин "примерно" обозначает диапазон +/- 10% от следующего далее числового значения. В одном воплощении термин "примерно" обозначает диапазон +/- 5% от следующего далее числового значения.
"Акцепторная человеческая каркасная область" в контексте данного изобретения является каркасной областью, включающей аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), полученную из каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, как определено ниже. Акцепторная человеческая каркасная область, "полученная из" каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, может содержать одну и ту же аминокислотную последовательность, или она может содержать изменения в аминокислотной последовательности. В некоторых воплощениях число аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или меньше, 3 или менее, или 2 или менее. В некоторых воплощениях акцепторная человеческая каркасная область VL по своей последовательности идентична последовательности VL каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной последовательности.
Понятие антитела "со зрелой аффинностью" относится к антителу с одним или более чем одним изменением в одной или более чем одной гипервариабельной области (hypervariable region, HVR) по сравнению с родительским антителом, которое не обладает такими изменениями, причем такие изменения приводят к повышению аффинности антитела к антигену.
Термин "изменение" обозначает мутацию (замещение), вставку (добавление) или делецию одного или более чем одного аминокислотного остатка в родительском антителе или гибридном полипептиде, например, гибридном полипептиде, содержащем по меньшей мере FcRn-связывающую часть Fc-области, чтобы получить модифицированное антитело или гибридный полипептид. Термин "мутация" означает, что указанный аминокислотный остаток замещен на другой аминокислотный остаток. Например, мутация L234A означает, что аминокислотный остаток лизин в позиции 234 в Fc-области антитела (полипептиде) замещен аминокислотным остатком аланином (замещение лизина аланином) (нумерация в соответствии с индексом ЕС).
Термин "аминокислотная мутация" обозначает замещение по меньшей мере одного существующего аминокислотного остатка другим отличающимся аминокислотным остатком (= замена аминокислотного остатка). Замененный аминокислотный остаток может быть "природным аминокислотным остатком" и может быть выбран из группы, состоящей из аланина (трехбуквенный код: Ala, один буквенный код: А), аргинина (Arg, R), аспарагина (Asn, N), аспарагиновой кислоты (ASP, D), цистеина (Cys, С), глутамина (Gin, Q), глутаминовой кислоты (Glu, Е), глицина (Gly, G), гистидина (His, Н), изолейцина (Не, I), лейцина (Leu, L), лизина (Lys, K), метионина (Met, М), фенилаланина (Phe, F), пролина (Pro, Р), серина (Ser, S), треонина (Thr, Т), триптофана (Trp, W), тирозина (Tyr, Y) и валина (Val, V). Замененный аминокислотный остаток может быть "неприродным аминокислотным остатком", см., например, US 6586207, WO 98/48032, WO 03/073238, US 2004/0214988, WO 2005/35727, WO 2005/74524, Chin, J.W., et al., J. Am. Chem. Soc. 124 (2002) 9026-9027; Chin, J.W. and Schultz, P.G., ChemBioChem 11 (2002) 1135-1137; Chin, J.W., et al., PICAS United States of America 99 (2002) 11020-11024; и Wang, L. and Schultz, P.O., Chem. (2002) 1-10 (все полностью включены в данный документ посредством ссылки).
Термин "аминокислотная вставка" обозначает (дополнительное) встраивание по меньшей мере одного аминокислотного остатка в заданной позиции в аминокислотной последовательности. В одном воплощении вставка будет представлять собой введение одного или двух аминокислотных остатков. Вставленный аминокислотный остаток (остатки) может быть любым природным или неприродным аминокислотным остатком.
Термин "аминокислотная делеция" означает удаление по меньшей мере одного аминокислотного остатка в заданной позиции в аминокислотной последовательности.
Термин "ANG-2", используемый в данном документе, относится к человеческому ангиопоэтину-2 (ANG-2) (альтернативно сокращается как ANGPT2 или ANG2) (SEQ ID №31), который описан, например, в Ivlaisonpierre, Р.С., et al, Science 277 (1997) 55-60, и в Cheung, А Н., et al., Genomics 48 (1998) 389-91. Ангиопоэтины 1 (SEQ ID №32) и 2 были открыты в качестве лигандов для Tie, семейства тирозинкиназ, которые селективно экспрессируются в сосудистом эндотелии (Yancopoulos, G.D., et al., Nature 407 (2000) 242-248). В настоящее время есть четыре определенных члена семейства ангиопоэтина. Ангиопоэтин 3 и 4 (ANG-3 и ANG-4) могут представлять собой широко разошедшиеся аналоги одного и того же генного локуса у мыши и человека (Kim, I., et al., FEBS Let, 443 (1999) 353-356; Kim, I., et al., J. Biol. Chem. 274 (1999) 26523-26528). ANG-1 и ANG-2 были первоначально идентифицирован в экспериментах на культурах тканей в качестве агониста и антагониста, соответственно (про ANG-1 см.: Davis, S., et al., Cell 87 (1996) 1161-1169; про ANG-2 см.: Ivlaisonpierre, P.C, et al., Science 277 (1997) 55-60). Все известные ангиопоэтины связывают, прежде всего, Tie2 (SEQ ID №33), и оба ANG-1 и -2 связывают Tie2 с аффинностью 3 нМ (Kd) (Ivlaisonpierre, Р.С., et al., Science 277 (1997) 55-60).
Термин "антитело" используется в данном документе в широком смысле и охватывает различные структуры антител, включая, но не ограничиваясь ими, моноклональные антитела, мультиспецифические антитела (например, биспецифические антитела, триспецифические антитела), а также фрагменты антител до тех пор, пока они проявляют нужную активность связывания с антигеном и/или белком А и/или FcRn.
Понятие "антитела, которое связывается с тем же эпитопом", что и референсное антитело, относится к антителу, которое блокирует связывание референсного антитела с его антигеном в конкурентном анализе на 50% или более, и, наоборот, референсное антитело блокирует связывание данного антитела с его антигеном в конкурентном анализе на 50% или более. В данном документе предложен иллюстративный конкурентный анализ.
Термин "асимметричная Fc-область" обозначает пару полипептидов Fc-области, которые имеют различные аминокислотные остатки в соответствующих позициях в соответствии с индексом ЕС по системе нумерации Kabat.
Термин "асимметричная Fc-область в отношении FcRn-связывания" обозначает Fc-область, которая состоит из двух полипептидных цепей, которые имеют различные аминокислотные остатки в соответствующих позициях, где эти позиции определяются в соответствии с индексом ЕС по системе нумерации Kabat, где различные позиции влияют на связывание Fc-области с человеческим неонатальным Fc-рецептором (FcRn). В данном описании различия между двумя полипептидными цепями Fc-области в "асимметричной Fc-области в отношении FcRn-связывания" не включают различия, которые были введены для облегчения формирования гетеродимерных Fc-областей, например, для получения биспецифических антител. Эти различия также могут быть асимметричными, т.е. две цепи имеют различия на несоответствующих аминокислотных остатках в соответствии с индексом ЕС по системе нумерации Kabat. Эти различия облегчают гетеродимеризацию и уменьшают гомодимеризацию. Примерами таких различий являются так называемые замены типа "ключ в замке" (см., например, US 7695936 и US 2003/0078385). Были обнаружены следующие замещения типа "ключ в замке" в отдельных полипептидных цепях Fc-области антитела IgG подкласса IgG1, которые повышали образование гетеродимера: 1) Y407T в одной цепи и T366Y в другой цепи; 2) Y407A в одной цепи и T366W в другой цепи; 3) F405A в одной цепи и T394W в другой цепи; 4) F405W в одной цепи и T394S в другой цепи; 5) Y407T в одной цепи и T366Y в другой цепи; 6) T366Y и F405A в одной цепи и T394W и Y407T в другой цепи; 7) T366W и F405W в одной цепи и T394S и Y407A в другой цепи; 8) F405W и Y407A в одной цепи и T366W и T394S в другой цепи; и 9) T366W в одной цепи и T366S, L368A и Y407V в другой цепи, где последние перечисленные являются особенно подходящими. Кроме того, изменения, создающие новые дисульфидные мостики между двумя полипептидными цепями Fc-области, облегчают формирование гетеродимеров (см., например, US 2003/0078385). Были найдены следующие замены, образующие надлежащим образом разнесенные остатки цистеина для образования новых внутрицепочечных дисульфидных связей в отдельных полипептидных цепях Fc-области антитела IgG подкласса IgG1, которые увеличивали формирование гетеродимера: Y349C в одной цепи и S354C в другой; Y349C в одной цепи и Е356С в другой; Y349C в одной цепи и Е357С в другой; L351C в одной цепи и S354C в другой; Т394С в одной цепи и Е397С в другой; или D399C в одной цепи и K392C в другой. Другими примерами аминокислотных замен, облегчающих гетеродимеризацию, являются так называемые "парные замещения с изменением заряда" (см., например, WO 2009/089004). Были найдены следующие парные замещения с изменением заряда в отдельных полипептидных цепях в Fc-области антитела IgG подкласса IgG1, которые увеличивали формирование гетеродимера: 1) K409D или K409E в одной цепи и D399K или D399R в другой цепи; 2) K392D или K392E в одной цепи и D399K или D399R в другой цепи; 3) K439D или K439E в одной цепи и E356K или E356R в другой цепи; 4) K370D или K370E в одной цепи и E357K или E357R в другой цепи; 5) K409D и K360D в одной цепи плюс D399K и E356K в другой цепи; 6) K409D и K370D в одной цепи плюс D399K и E357K в другой цепи; 7) K409D и K392D в одной цепи плюс D399K, E356K и E357K в другой цепи; 8) K409D и K392D в одной цепи и D399K в другой цепи; 9) K409D и K392D в одной цепи и D399K и E356K в другой цепи; 10) K409D и K392D в одной цепи и D399K и D357K в другой цепи; 11) K409D и K370D в одной цепи и D399K и D357K в другой цепи; 12) D399K в одной цепи и K409D и K360D в другой цепи; и 13) K409D и K439D в одной цепи и D399K и E356K в другой.
Термин "связывающееся (с антигеном)" обозначает связывание антитела с его антигеном в анализе in vitro, в одном воплощении с помощью анализа связывания, в котором антитело связано с поверхностью, и связывание антигена с антителом измеряется с помощью поверхностного плазмонного резонанса (Surface Plasmon Resonance, SPR). Связывание означает аффинность связывания (KD) 10-8 M или менее, в некоторых воплощениях от 10-13 до 10-8 M, в некоторых воплощениях от 10-13 до 10-9 М.
Связывание может быть исследовано с помощью анализа BIAcore (GE Healthcare Biosensor АВ, Уппсала, Швеция). Аффинность связывания определяется терминами ka (константа скорости ассоциации антитела из комплекса антитело/антиген), kd (константа диссоциации) и KD(kd/ka).
Термин "химерное" антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или вида, в то время как остальная часть тяжелой и/или легкой цепи получена из другого источника или вида.
Термин "СН2-домен" обозначает часть полипептида тяжелой цепи антитела, которая простирается примерно от позиции ЕС 231 до позиции ЕС 340 (в соответствии с индексом ЕС по системе нумерации Kabat). В одном воплощении СН2-домен имеет аминокислотную последовательность SEQ ID № 09: APELLGG PSVFLFPPKP KDTLMISRTP EVTCVWDVS HEDPEVKFNW YVDGVEVHNA KTKPREEQ E STYRWSVLT VLHQDWLNGK EYKCKVSNKA LPAPIEKTIS КАК.
Термин "СН3-домен" обозначает часть полипептида тяжелой цепи антитела, которая простирается примерно от позиции ЕС 341 до позиции ЕС 446. В одном воплощении СН3-домен имеет аминокислотную последовательность SEQ ID №10: GQPREPQ VYTLPPSRDE LTKNQVSLTC LVKGFYPSDI AVEWESNGQP ENNYKTTPPV LDSDGSFFLY SKLTVDKSRW QQGNVFSCSV MHEALHNHYT QKSLSLSPG.
Понятие "класса" антитела относится к типу константного домена или константной области, принадлежащей его тяжелой цепи. Есть пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
Термин "сравнимая длина" обозначает, что два полипептида содержат одинаковое число аминокислотных остатков или могут отличаться по длине на один или более и до 10 аминокислотных остатков максимум. В одном воплощении полипептиды Fc-области содержат одинаковое число аминокислотных остатков или отличаются друг от друга на 1-10 аминокислотных остатков. В одном воплощении полипептиды Fc-области содержат одинаковое число аминокислотных остатков или отличаются друг от друга на 1-5 аминокислотных остатков. В одном воплощении полипептиды Fc-области содержат одинаковое число аминокислотных остатков или отличаются друг от друга на 1-3 аминокислотных остатка.
Термин "получена из" означает, что аминокислотная последовательность получена из родительской аминокислотной последовательности путем введения изменений по меньшей мере в одной позиции. Таким образом, полученная аминокислотная последовательность отличается от соответствующей родительской аминокислотной последовательности по меньшей мере в одной соответствующей позиции (в соответствии с индексом ЕС по системе нумерации Kabat для Fc-областей антител). В одном воплощении аминокислотная последовательность, полученная из родительской аминокислотной последовательности, отличается на число аминокислотных остатков от одного до пятнадцати в соответствующих позициях. В одном воплощении аминокислотная последовательность, полученная из родительской аминокислотной последовательности, отличается на число аминокислотных остатков от одного до десяти в соответствующих позициях. В одном воплощении аминокислотная последовательность, полученная из родительской аминокислотной последовательности, отличается на число аминокислотных остатков от одного до шести в соответствующих позициях. Аналогичным образом, полученная аминокислотная последовательность имеет высокую идентичность аминокислотной последовательности со своей родительской аминокислотной последовательностью. В одном воплощении аминокислотная последовательность получена из последовательности аминокислот родительского имеет 80% или более идентичности аминокислотной последовательности. В одном воплощении аминокислотная последовательность, полученная из родительской аминокислотной последовательности, имеет идентичность аминокислотной последовательности 90% или более. В одном воплощении аминокислотная последовательность, полученная из родительской аминокислотной последовательности, имеет идентичность аминокислотной последовательности 95% или более.
"Эффекторные функции" относятся к таким биологическим активностям, которые приписываются Fc-области антитела, которая варьирует в зависимости от класса антитела. Примеры эффекторных функций антител включают: C1q-связывание и комплементзависимую цитотоксичность (complement dependent cytotoxicity, CDC); Fc-рецепторное связывание; антителозависимую клеточную цитотоксичность (antibody-dependent cell-mediated cytotoxicity, ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора); и В-клеточную активацию.
Понятие "эффективного количества" агента, например, фармацевтического препарата, относится к количеству, эффективному в дозах и в течение периода времени, необходимого для достижения желаемого терапевтического или профилактического результата.
Термин "Fc-гибридный полипептид" обозначает гибрид связывающего домена (например, антигенсвязывающего домена, такого как одноцепочечное антитело, или полипептида, такого как лиганд рецептора), и Fc-области антитела, который обладает нужной активностью связывания мишени и/или белка А и/или FcRn.
Термин "Fc-область человеческого происхождения" обозначает С-концевую область тяжелой цепи иммуноглобулина человеческого происхождения, которая содержит по меньшей мере часть шарнирной области, СН2-домен и СН3-домен. В одном воплощении Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Рго230 до карбоксильного конца тяжелой цепи. В одном воплощении Fc-область имеет аминокислотную последовательность SEQ ID №60. Тем не менее, С-концевой лизин (Lys447) в Fc-области может присутствовать или не присутствовать. Если не указано иное, то нумерация аминокислотных остатков в Fc-области или константной области осуществляется в соответствии с системой нумерации ЕС, также называемой индексом ЕС, как описано в Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91 3242. Fc-область состоит из двух полипептидов Fc-области тяжелых цепей, которые могут быть ковалентно связаны друг с другом через остатки цистеина шарнирной области, формирующие межполипептидные дисульфидные связи.
Термин "FcRn" обозначает человеческий неонатальный Fc-рецептор. Функция FcRn заключается в спасении IgG от деградации по лизосомальному пути, что приводит к снижению его выведения и увеличению времени полужизни. FcRn является гетеродимерным белком, состоящим из двух полипептидов: подобный белку главного комплекса гистосовместимости I класс размером 50 кДа (α-FcRn) и β2-микроглобулин размером 15 кДа (β2m). FcRn с высокой аффинностью связывается с СН2-СН3-частью Fc-области IgG. Взаимодействие между IgG и FcRn строго зависит от рН и происходит в стехиометрии 1:2, где одна молекула IgG связывается с двумя молекулами FcRn через две тяжелые цепи (Huber, А Н., et al., J. Mol. Biol. 230 (1993) 1077-1083). Связывание FcRn происходит в эндосоме в кислых условиях (рН<6,5), и IgG высвобождается на нейтральной клеточной поверхности (рН 7,4). рН-чувствительный характер взаимодействия облегчает FcRn-опосредованную защиту IgG, пиноциированных в клетки после внутриклеточной деградации, путем связывания с рецептором в кислой среде эндосом. Затем FcRn облегчает возвращение IgG к клеточной поверхности и последующее высвобождение в кровоток при воздействии на комплекс FcRn-IgG нейтральной рН-среды вне клетки.
Термин "FcRn-связывающая часть Fc-области" обозначает часть полипептида тяжелой цепи антитела, которая простирается примерно от позиции ЕС 243 до позиции ЕС 261, и примерно от позиции ЕС 275 до позиции ЕС 293, и примерно от позиции ЕС 302 до позиции ЕС 319, и примерно от позиции ЕС 336 до позиции ЕС 348, и примерно от позиции ЕС 367 до позиции ЕС 393 и позиции ЕС 408, и примерно от позиции ЕС 424 до позиции ЕС 440. В одном воплощении один или более из следующих аминокислотных остатков в соответствии с индексом ЕС согласно нумерации Kabat изменены: F243, Р244, Р245 Р, K246, Р247, K248, D249, Т250, L251, M252, I253, S254, R255, Т256, Р257, Е258, V259, Т260, С261, F275, N276, W277, Y278, V279, D280, V282, Е283, V284, Н285, N286, А287, K288, Т289, K290, Р291, R292, Е293, V302, V303, S304, V305, L306, Т307, V308, L309, Н310, Q311, D312, W313, L314, N315, G316, K317, Е318, Y319, I336, S337, K338, А339, K340, G341, Q342, Р343, R344, Е345, Р346, Q347, V348, С367, V369, F372, Y373, Р374, S375, D376, I377, А378, V379, Е380, W381, Е382, S383, N384, G385, Q386, Р387, Е388, N389, Y391, Т393, S408, S424, С425, S426, V427, М428, Н429, Е430, А431, L432, Н433, N434, Н435, Y436, Т437, Q438, K439 и S440 (нумерация ЕС).
"Каркасная область", или "FR", относится к остаткам вариабельных доменов за исключением остатков гипервариабельных областей (hypervariable region, HVR). FR вариабельного домена обычно состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно идут в следующей последовательности в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термин "полноразмерное антитело" обозначает антитело, имеющее структуру, по существу аналогичную структуре нативного антитела, или имеющее тяжелые цепи, которые содержат Fc-область, определенную в данном документе. Полноразмерное антитело может содержать другие домены, такие как, например scFv или scFab, конъюгированные с одной или более чем одной цепью полноразмерного антитела. Эти конъюгаты также охватываются термином полноразмерного антитела.
Термины "гетеродимер" или "гетеродимерный" обозначают молекулу, которая содержит две полипептидные цепи (например, сопоставимой длины), где две полипептидные цепи имеют аминокислотную последовательность, которая содержит по меньшей мере один отличающийся аминокислотный остаток в соответствующей позиции, где соответствующая позиция определяется в соответствии с индексом ЕС по Kabat.
Термины "гомодимер" и "гомодимерный" обозначают молекулу, которая содержит две полипептидные цепи сравнимой длины, где две полипептидные цепи содержат аминокислотную последовательность, которая идентична в соответствующих позициях, где соответствующие позиции определяются в соответствии с индексом ЕС по Kabat.
Антитело или гибридный полипептид Fc-области, описанный в данном документе, может быть гомодимерным или гетеродимерным в отношении его Fc-области, что определяется относительно мутаций или свойств, находящихся в фокусе. Например, в отношении связывания FcRn и/или белка А (т.е. фокус на свойствах) Fc-область (антитело) является гомодимерной (т.е. оба полипептида Fc-области тяжелой цепи содержат эти мутации) в отношении мутаций Н310А, Н433А и Y436A (эти мутации находятся в фокусе в отношении свойств связывания FcRn и/или белка А для гибридного полипептида Fc-области или антитела), но в то же время гетеродимерной в отношении мутаций Y349C, T366S, L368A и Y407V (эти мутации не в фокусе, поскольку эти мутации направлены на гетеродимеризацию тяжелых цепей, а не на свойства связывания FcRn и/или белка А), а также мутаций S354C и T366W, соответственно (первый перечень содержится только в первом полипептиде Fc-области, в то время как второй перечень содержится только во втором полипептиде Fc-области). Кроме того, например, гибридный полипептид Fc-области или антитело, описанное в данном документе, может быть гетеродимерным в отношении мутаций I253A, Н310А, Н433А, Н435А и Y436A (т.е. все эти мутации направлены на свойства димерного полипептида связывать FcRn и/или белок А), т.е. один полипептид Fc-области содержит мутации I253A, Н310А и Н435А, в то время как другой полипептид Fc-области содержит мутации Н310А, Н433А и Y436A.
Термины "клетка-хозяин", "клеточная линия-хозяин" и "клеточная культура-хозяин" используются взаимозаменяемо и относятся к клеткам, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают "трансформанты" и "трансформированные клетки", которые включают первичные трансформированные клетки и потомство, полученное из них, независимо от числа пассажей. Потомство может не быть полностью идентичным родительской клетке по содержанию нуклеиновых кислот, а может содержать мутации. Мутантное потомство, которое имеет такую же функцию или биологическую активность, как при скрининге или селекции первоначально трансформированной клетки, включено в данный документ.
"Человеческое антитело" является таким антителом, которое обладает аминокислотной последовательностью, которая соответствует антителу, продуцируемому человеком и/или человеческой клеткой или получено из нечеловеческого источника, который использует репертуар человеческих антител или других человеческих последовательностей, кодирующих антитела. Это определение человеческого антитела, в частности, исключает гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
"Человеческая консенсусная каркасная область" является каркасной областью, которая представляет наиболее часто встречающиеся аминокислотные остатки в выборе каркасных областей последовательностей VL или VH человеческого иммуноглобулина. Как правило, выбор VL- или VH-последовательностей человеческого иммуноглобулина осуществляется из подгруппы последовательностей вариабельного домена. Как правило, подгруппа последовательностей является такой подгруппой, как описано в Kabat, Е.А. et al., Sequences of Proteins of Immunological Interest, 5th ed., Bethesda MD (1991), NIH Publication 91-3242, Vols. 1-3. В одном воплощении для VL подгруппа является подгруппой каппа I, как описано в Kabat et al., см. выше. В одном воплощении для VH подгруппа является подгруппой III, как описано в Kabat et al., см. выше.
Термин "полипептид человеческой Fc-области" обозначает последовательность аминокислот, которая идентична "нативному" полипептиду или полипептиду "дикого типа" человеческой Fc-области. Термин "полипептид вариантной (человеческой) Fc-области" обозначает последовательность аминокислот, которая получена из "нативного" полипептида или полипептида "дикого типа" человеческой Fc-области путем по меньшей мере одного "аминокислотного изменения". "Человеческая Fc-область" состоит из двух полипептидов человеческой Fc-области. "Вариантная (человеческая) Fc-область" состоит из двух полипептидов Fc-области, причем оба могут быть полипептидами вариантной (человеческой) Fc-области, либо один представляет собой полипептид человеческой Fc-области, а другой представляет собой полипептид вариантной (человеческой) Fc-области.
В одном воплощении полипептид человеческой Fc-области имеет аминокислотную последовательность полипептида Fc-области человеческого IgG1 SEQ ID №60, или полипептида Fc-области человеческого IgG 2 SEQ ID №
61, или человеческого полипептид Fc-области IgG3 SEQ ID №62, или полипептида Fc-области человеческого IgG4 SEQ ID №63. В одном воплощении полипептид вариантной (человеческой) Fc-области получен из Fc-области полипептида SEQ ID №60, или 61, или 62, или 63, и имеет по меньшей мере одну аминокислотную мутацию по сравнению с полипептидом человеческой Fc-области SEQ ID №60, или 61, или 62, или 63. В одном воплощении полипептид вариантной (человеческой) Fc-области содержит/имеет от одной до примерно двенадцати аминокислотных мутаций, и в одном воплощении от примерно одной до примерно восьми аминокислотных мутаций. В одном воплощении полипептид вариантной (человеческой) Fc-области имеет по меньшей мере примерно 80% гомологию с полипептидом человеческой Fc-области SEQ ID №60, или 61, или 62, или 63. В одном воплощении полипептид вариантной (человеческой) Fc-области имеет по меньшей мере примерно 90% гомологию с полипептидом человеческой Fc-области SEQ ID №60, или 61, или 62, или 63. В одном воплощении полипептид вариантной (человеческой) Fc-области имеет по меньшей мере 95% гомологию с полипептидом человеческой Fc-области SEQ ID №60, или 61, или 62, или 63.
Полипептид вариантной (человеческой) Fc-области, полученный из полипептида человеческой Fc-области SEQ ID №60, или 61, или 62, или 63 определяется содержащимися аминокислотными изменениями. Так, например, термин "P329G" обозначает полипептид вариантной (человеческой) Fc-области, полученный из полипептида человеческой Fc-области, с заменой пролина на глицин в аминокислотной позиции 329 относительно полипептида человеческой Fc-области SEQ ID №60 или 61, или 62, или 63.
Используемые в данном документе аминокислотные позиции всех константных областей и доменов тяжелой и легкой цепи пронумерованы в соответствии с системой нумерации Kabat, описанной в Kabat, et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), которая называется "нумерацией согласно Kabat" в данном документе. В частности, система нумерации Kabat (см. страницы 647-660) из Kabat, et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991) используется для константного домена легкой цепи CL изотипов каппа и лямбда, и индекс ЕС по системе нумерации Kabat (см страницы 661-723) используется для константных доменов тяжелой цепи (СН1, шарнирный, СН2 и СН3).
Полипептид Fc-области человеческого IgG1 имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALP APIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №60).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями L234A, L235A имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALP APIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №64).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями Y349C, T366S, L368A и Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALP APIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №65).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALP APIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №66).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями L234A, L235A и мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALP APIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №67).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями L234A, L235A и мутациями S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALP APIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №68).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутацией P329G имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALG APIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №69).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями L234A, L235A и мутацией P329G имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALG APIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №70).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутацией P239G и мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALG APIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №71).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутацией P329G и мутацией S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALG APIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №72).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями L234A, L235A, P329G и мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALG APIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №73).
Полипептид Fc-области, полученный из Fc-области человеческого IgG1, с мутациями L234A, L235A, P329G и мутациями S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSHEDPEV KFNWYVDGVEVHNAKTKPREEQYNSTYRWSVLTVLHQDWLNGKEYKCKVSNKALG APIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQP ENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLS PGK (SEQ ID №74).
Полипептид Fc-области человеческого IgG4 имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSL SLSLGK (SEQ ID №63).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями S228P и L235E имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSL SLSLGK (SEQ ID №75).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями S228P, L235E и мутацией P329G имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LGSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSL SLSLGK (SEQ ID №76).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LPSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESN GQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKS LSLSLGK (SEQ ID №77).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LPSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSL SLSLGK (SEQ ID №78).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями S228P, L235E и S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LPSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESN GQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID №79).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями S228P, L235E и Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LPSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID №80).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутацией P329G имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LGSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID №81).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями P239G и Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LGSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID №82).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями P329G и S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LGSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESN GQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKS LSLSLGK (SEQ ID №83).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями S228P, L235E, P329G и Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCWVDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LGSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNG QPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSL SLSLGK (SEQ ID №84).
Полипептид Fc-области, полученный из Fc-области человеческого IgG4, с мутациями S228P, L235E, P329G и S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVWDVSQEDP EVQFNWYVDGVEVHNAKTKPREEQFNSTYRWSVLTVLHQDWLNGKEYKCKVSNKG LGSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESN GQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKS LSLSLGK (SEQ ID №85).
Ниже показано выравнивание различных человеческих Fc-областей (нумерация ЕС):


"Гуманизированное" антитело относится к химерному антителу, содержащему аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В некоторых воплощениях гуманизированное антитело будет содержать по существу все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или по существу все HVR (например, CDR) соответствуют таковым в нечеловеческом антителе, и все или по существу все FR соответствуют таковым в человеческом антителе. Гуманизированное антитело, возможно, может содержать по меньшей мере часть константной области антитела, полученной из человеческого антитела. "Гуманизированная форма" антитела, например, нечеловеческого антитела, относится к антителу, которое подверглось гуманизации.
Термин "гипервариабельная область", или "HVR", используемый в данном документе, относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными в последовательности ("области, определяющие комплементарность", или "CDR") и формируют структурно определенные петли ("гипервариабельные петли") и/или содержат остатки, контактирующие с антигеном ("контакты антигена"). Как правило, антитела содержат шесть HVR; три в VH (Н1, Н2, Н3), и три в VL (L1, L2, L3). HVR, указанные в данном документе, включают
(a) гипервариабельные петли, возникающие на аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3) (Chothia, С. and Lesk, A.M., J. Mol. Biol. 196 (1987) 901-917);
(b) CDR, возникающие на аминокислотных остатках 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (Н1), 50-65 (Н2) и 95-102 (Н3) (Kabat, Е.А. et al., Sequences of Proteins of Immunological Interest, 5th ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242.);
(c) контакты антигена, возникающие на аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (Н1), 47-58 (Н2) и 93-101 (Н3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)); и
(d) комбинации (a), (b) и/или (с), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (Н1), 26-35b (Н1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
В одном воплощении HVR-остатки содержат такие, которые определены в других местах в данном описании.
Если не указано иное, HVR-остатки и другие остатки в вариабельном домене (например, FR-остатки) нумеруются в данном документе в соответствии с индексом ЕС по системе нумерации Kabat (Kabat et al., см. выше).
Термин "IGF-1R", используемый в данном документе, относится к любому нативному IGF-1R от любого позвоночного животного, включая млекопитающих, таких как приматы (например, человек) и грызуны (например, мыши и крысы), если не указано иное. Термин охватывает "полноразмерный" непроцессированный IGF-1R, а также любую форму IGF-1R, которая образовалась в результате процессинга в клетке. Термин также охватывает встречающиеся в природе варианты IGF-1R, например, варианты сплайсинга или аллельные варианты. Аминокислотная последовательность человеческого IGF-1R показана в SEQ ID №11.
"Индивидуум" или "субъект" является млекопитающим. Млекопитающие включают, но не ограничиваются ими, домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В некоторых воплощениях индивидуум или субъект является человеком.
"Изолированное" антитело является таким антителом, которое было отделено от компонентов его природной среды. В некоторых воплощениях антитело очищают до чистоты более 95% или 99%, которую определяют, например, с помощью электрофореза (например, SDS-PAGE, изоэлектрофокусировки (IEF), капиллярного электрофореза) или хроматографии (например, эксклюзионной хроматографии или ионнообменной или обращено-фазовой HPLC). Обзор способов оценки чистоты антител см., например, в Flatman, S. et al., J. Chromatogr. В 848 (2007) 79-87.
Понятие "изолированной" нуклеиновой кислоты относится к нуклеиновокислотной молекуле, которая была отделена от компонентов ее природной среды. Изолированная нуклеиновая кислота включает нуклеиновокислотную молекулу, содержащуюся в клетках, которые обычно содержат нуклеиновокислотные молекулы, но нуклеиновокислотная молекула находится вне хромосомы или в том месте хромосомы, которое отличается от ее природной хромосомной локализации.
"Изолированная нуклеиновая кислота, кодирующая антитело против IGF-1R" относится к одной или более чем одной нуклеиновокислотной молекуле, кодирующей тяжелую и легкую цепи антитела (или их фрагменты), включая такие нуклеиновокислотные молекулы в одном векторе или в отдельных векторах, и такие нуклеиновокислотные молекулы присутствуют в одном или более чем одном месте в клетке-хозяине.
Термин "моноклональное антитело", используемый в данном документе, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. популяции, содержащей отдельные антитела, которые являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантных антител, например, содержащих встречающиеся в природе мутации или образованных во время производства препарата моноклонального антитела, где такие могут присутствовать в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклонального антитела направлено против одной детерминанты на антигене. Таким образом, указатель "моноклональное" указывает на характер антитела, которое было получено из популяции по существу гомогенных антител, и не должен быть истолкован как требующий получения антитела каким-либо конкретным способом. Например, моноклональные антитела, используемые в соответствии с данным изобретением, могут быть получены с помощью различных методик, включая, но не ограничиваясь ими, гибридомную методику, рекомбинантную ДНК-методику, методики фагового дисплея, а также методики с использованием трансгенных животных, содержащих все или часть человеческих иммуноглобулиновых локусов, и такие методики и другие иллюстративные методики получения моноклональных антител описаны в данном документе.
Понятие "нативных антител" относится к молекулам иммуноглобулина природного происхождения с различными структурами. Например, нативные антитела IgG являются гетеротетрамерными гликопротеинами размером примерно 150000 Да, состоящими из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидными мостиками. От N- к С-концу каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, а затем три константных домена (СН1, СН2 и СН3). Аналогично, от N- к С-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, с последующим константным легким доменом (CL). Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (к) и лямбда (А), на основании аминокислотной последовательности его константного домена.
Термин "вкладыш в упаковку" используется для обозначения инструкций, обычно включаемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях, касающихся применения таких терапевтических продуктов.
"Процент (%) идентичности аминокислотной последовательности" по отношению к референсной полипептидной последовательности определяется как процент аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам в референсной полипептидной последовательности после выравнивания последовательностей и введения при необходимости пробелов для достижения максимального процента идентичности последовательностей, и без учета любых консервативных замен как части идентичности последовательности. Выравнивание с целью определения процента идентичности аминокислотных последовательностей может быть достигнуто различными способами, которые известны специалистам в данной области, например, с использованием общедоступного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для выравнивания последовательностей, в том числе любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Тем не менее, для целей данного изобретения значения % идентичности аминокислотных последовательностей получают с использованием компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана Genentech, Inc, и исходный код был подан с пользовательской документацией в бюро авторских прав США, Вашингтон, округ Колумбия, 20559, где он был зарегистрирован под номером TXU510087. Программа ALIGN-2 находится в открытом доступе от Genentech, Inc., Саут-Сан-Франциско, штат Калифорния, или может быть составлен из исходного кода. Программа ALIGN-2 должна быть скомпилирована для применения на операционной системе UNIX, включая цифровую UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не меняются.
В ситуациях, когда для сравнения аминокислотных последовательностей используется ALIGN-2, процент идентичности аминокислотной последовательности данной аминокислотной последовательности А с данной аминокислотной последовательностью В (альтернативно можно сказать, что данная аминокислотная последовательность А имеет или содержит определенный процент идентичности аминокислотной последовательности с данной аминокислотной последовательностью В) вычисляется следующим образом:
100 × доля X/Y,
где X обозначает число аминокислотных остатков, оцененных как идентичные совпадения с помощью программы для выравнивания последовательностей ALIGN-2 при выравнивании в этой программе А и В, и где Y обозначает общее число аминокислотных остатков в В. Следует иметь в виду, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то % идентичности аминокислотной последовательности А с В не будет равен % идентичности аминокислотной последовательности В с А. Если специально не указано иное, то все значения % идентичности аминокислотной последовательности, используемые в данном описании, получены так, как описано в предыдущем абзаце, с использованием компьютерной программы ALIGN-2.
Термин "фармацевтический препарат" относится к препарату, который находится в такой форме, чтобы обеспечить биологическую активность активного ингредиента, содержащегося в нем, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому будет осуществляться введение препарата.
Понятие "фармацевтически приемлемого носителя" относится к ингредиенту в фармацевтическом препарате, помимо активного ингредиента, который является нетоксичным для субъекта. Фармацевтически приемлемые носители включают, но не ограничиваясь ими, буфер, эксципиент, стабилизатор или консервант.
Термин "пептидный линкер", используемый в данном документе, обозначает пептид с аминокислотной последовательностью, которая в одном воплощении имеет синтетическое происхождение. Пептидный линкер в одном воплощении является пептидом с аминокислотной последовательностью длиной по меньшей мере 30 аминокислот, в одном воплощении с длиной от 32 до 50 аминокислот. В одном воплощении пептидный линкер представляет собой пептид с аминокислотной последовательностью длиной от 32 до 40 аминокислот. В одном воплощении пептидный линкер представляет собой (G×S)n, где G = глицин, S = серии, (х=3, n=8, 9 или 10) или (х=4 и n=6, 7 или 8), в одном воплощении х=4, n=6 или 7, в одном воплощении х=4, n=7. В одном воплощении пептидный линкер представляет собой (G4S)6G2.
Термин "рекомбинантное антитело" обозначает все антитела (химерные, гуманизированные и человеческие), которые получены, экспрессированы, созданы или выделены с помощью рекомбинантных средств. Они включают антитела, выделенные из клетки-хозяина, такой как клетка NS0 или СНО, или из животного (например, мыши), которое является трансгенным по генам иммуноглобулина человека, или антитела, экспрессированные с помощью рекомбинантного экспрессионного вектора, которым трансфицировали клетку-хозяина. Такие рекомбинантные антитела имеют вариабельные и константные области в перегруппированной форме. Рекомбинантные антитела, описанные в данном документе, могут быть подвергнуты соматическому гипермутированию in vivo. Таким образом, аминокислотные последовательности VH- и VL-областей рекомбинантных антител представляют собой последовательности, которые, притом что получены из и связаны с человеческими зародышевыми последовательностями VH и VL, могут не существовать в природном репертуаре человеческого зародышевого антитела in vivo.
Термин "лечение" (и его грамматические варианты, такие как "лечить") относится к клиническому вмешательству в попытке изменить естественный ход заболевания у индивидуума, которого лечат, и которое может быть выполнено либо для профилактики, либо в ходе клинической патологии. Желательные эффекты лечения включают, но не ограничиваясь ими, предотвращение возникновения или рецидива заболевания, ослабление симптомов, уменьшение любого прямого или косвенного патологического последствия заболевания, предотвращение метастазирования, уменьшение скорости прогрессирования заболевания, улучшение или временное облегчение болезненного состояния, достижение ремиссии или улучшение прогноза. В некоторых воплощениях антитела или гибридные полипептиды Fc-области, описанные в данном документе, используются для задержки развития заболевания или для замедления прогрессии заболевания.
Термин "валентность", используемый в данной заявке, означает наличие определенного числа сайтов связывания в молекуле (антитела). Таким образом, термины "двухвалентный", "четырехвалентный" и "шестивалентный" обозначают наличие двух сайтов связывания, четырех сайтов связывания и шести сайтов связывания, соответственно, в молекуле (антитела). Биспецифические антитела, описанные в данном документе, в одном предпочтительном воплощении являются "двухвалентными".
Термин "вариабельная область" или "вариабельный домен" относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с его антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) антитела обычно имеют аналогичную структуру, при этом каждый домен содержит четыре консервативные каркасные области (FR, framework region) и три гипервариабельные области (HVR, hypervariable region) (см., например, Kindt, T.J. et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., N.Y. (2007), page 91). Один VH- или VL-домен может быть достаточным для придания антигенсвязывающей специфичности. Кроме того, антитела, которые связывают конкретный антиген, могут быть выделены с помощью VH- или VL-домена из антитела, которое связывается с этим антигеном, для скрининга библиотеки комплементарных VL- или VH-доменов, соответственно. См., например, Portolano et al., J. Immunol. 150: 880-887 (1993); Clarkson et al., Nature 352: 624-628(1991). 624-628).
Термин "глазное сосудистое заболевание" включает, но не ограничиваясь ими, внутриглазные неоваскулярные синдромы, такие как диабетическая ретинопатия, диабетический макулярный отек, ретинопатия недоношенных, неоваскулярная глаукома, окклюзии вен сетчатки, окклюзии центральной вены сетчатки, дегенерация желтого пятна, возрастная дегенерация желтого пятна, пигментный ретинит, ретинальная ангиоматозная пролиферация, макулярные телеангиэктазии, ишемическая ретинопатия, неоваскуляризация радужной оболочки, внутриглазная неоваскуляризация, неоваскуляризация роговицы, неоваскуляризация сетчатки, неоваскуляризация сосудистой оболочки и дегенерация сетчатки (см., например, Garner, A., Vascular diseases, In: Pathobiology of ocular disease, A dynamic approach, Garner, A., and Klintworth, G.K., (eds.), 2nd edition, Marcel Dekker, New York (1994), pp.1625-1710).
Термин "вектор", используемый в данном документе, относится к нуклеиновокислотной молекуле, способной размножать другую нуклеиновую кислоту, с которой она связана. Этот термин включает вектор как самореплицирующуюся нуклеиновокислотную структуру, а также как вектор, включенный в геном клетки-хозяина, в которую он был введен. Некоторые векторы способны направлять экспрессию нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называют в данном документе "экспрессионными векторами".
Термин "VEGF", используемый в данном документе, относится к человеческому сосудистому эндотелиальному фактору роста (VEGF/VEGF-A), 165-аминокислотному человеческому сосудистому эндотелиальному фактору роста (аминокислоты 27-191 из последовательности предшественника человеческого VEGF165: SEQ ID №30; аминокислоты 1-26 представляют сигнальный пептид) и к родственным изоформам сосудистого эндотелиального фактора роста 121, 189 и 206, которые описаны в Leung, D.W., et al., Science 246 (1989) 1306-1309; Houck et al., Mol. Endocrin. 5 (1991) 1806-1814; Keck, P.J., et al., Science 246 (1989) 1309-1312, и в Connolly, D.T., et al., J. Biol. Chem. 264 (1989) 20017-20024; вместе с природными аллельными и процессированными формами этих факторов роста. VEGF участвует в регуляции нормального и аномального ангиогенеза и неоваскуляризации, связанной с опухолями и внутриглазными нарушениями (Ferrara, N., et al., Endocrin. Rev. 18 (1997) 4-25; Berkman, R.A., et al., J. Clin. Invest. 91 (1993) 153-159; Brown, L.F., et al., Human Pathol. 26 (1995) 86-91; Brown, L.F., et al., Cancer Res. 53 (1993) 4727-4735; Mattern, J., et al., Brit. J. Cancer. 73 (1996) 931-934; и в Dvorak, H.F., et al., Am. J. Pathol. 146 (1995) 1029-1039). VEGF является гомодимерным гликопротеином, который был выделен из нескольких источников и включает несколько изоформ. VEGF демонстрирует высокую специфическую митогенную активность в отношении эндотелиальных клеток.
Термин "с мутацией IHH-AAA", используемый в данном документе, относится к комбинации мутаций I253A (Ile253Ala), Н310А (His310Ala) и Н435А (His435Ala); термин "с мутацией HHY-AAA", используемый в данном документе, относится к комбинации мутаций Н310А (His310Ala), Н433А (His433Ala) и Y436A (Tyr436Ala), а термин "с мутацией YTE", используемый в данном документе, относится к комбинации мутаций M252Y (Met252Tyr), S254T (Ser254Thr) и Т256Е (Thr256Glu) в константной области тяжелой цепи подкласса IgG1 или IgG4, где нумерация осуществляется согласно индексу ЕС по Kabat.
Термин "с мутациями P329G LALA", используемый в данном документе, относится к комбинации мутаций L234A (Leu234Ala), L235A (Leu235Ala) и P329G (Pro329Gly) в константной области тяжелой цепи подкласса IgG1, где нумерация осуществляется согласно индексу ЕС по Kabat. Термин "с мутацией SPLE", используемый в данном документе, относится к комбинации мутаций S228P (Ser228Pro) и L235E (Leu235Glu) в константной области тяжелой цепи подкласса IgG4, где нумерация осуществляется согласно индексу ЕС по Kabat. Термин "с мутациями SPLE и P239G", используемый в данном документе, относится к комбинации мутаций S228P (Ser228Pro), L235E (Leu235Glu) и P329G (Pro329Gly) в константной области тяжелой цепи подкласса IgG4, где нумерация осуществляется согласно индексу ЕС по Kabat.
II. Данное изобретение
Изобретение основано, по меньшей мере частично, на обнаружении того факта, что FcRn-связывание антитела или гибридного полипептида Fc-области может быть модифицировано путем изменения аминокислотных остатков в несоответствующих позициях в отдельных полипептидах Fc-области, потому что эти изменения действуют совместно, изменяя FcRn-связывание. Антитела и гибридные полипептиды Fc-области, описанные в данном документе, могут быть использованы, например, для лечения заболеваний, при которых требуется индивидуальное системное время удерживания.
А. Неонатальный Fc-рецептор (FcRn)
Неонатальный Fc-рецептор (FcRn) имеет важное значение для метаболического пути антител класса IgG in vivo. Функции FcRn заключаются в спасении IgG дикого типа от деградации по лизосомальному пути, что приводит к снижению его выведения и увеличению времени полужизни. FcRn является гетеродимерным белком, состоящим из двух полипептидов: подобный белку главного комплекса гистосовместимости I класса размером 50 кДа (α-FcRn) и 62-микроглобулин размером 15 кДа (β2m). FcRn с высокой аффинностью связывается с СН2-СН3-частью Fc-области антитела класса IgG. Взаимодействие между антителом класса IgG и FcRn зависит от рН и происходит в стехиометрии 1:2, где одна молекула IgG связывается с двумя молекулами FcRn через два полипептида Fc-области тяжелой цепи (Huber, А Н., et al., J. Mol. Biol. 230 (1993) 1077-1083).
Таким образом, FcRn-связывающие свойства/характеристики IgG in vitro свидетельствуют о его фармакокинетических свойствах in vivo в кровотоке.
Во взаимодействии между FcRn и Fc-областью антитела класса IgG участвуют различные аминокислотные остатки СН2- и СН3-домена тяжелой цепи. Аминокислотные остатки, взаимодействующие с FcRn, находятся примерно между позицией ЕС 243 и позицией ЕС 261, примерно между позицией ЕС 275 и позицией ЕС 293, примерно между позицией ЕС 302 и позицией ЕС 319, примерно между позицией ЕС 336 и позицией ЕС 348, примерно между позицией ЕС 367 и позицией ЕС 393, в позиции ЕС 408 и примерно между позицией ЕС 424 и позицией ЕС 440. Более конкретно, во взаимодействии между Fc-областью и FcRn участвуют следующие аминокислотные остатки в соответствии с нумерацией ЕС по Kabat: F243, Р244, Р245Р, K246, Р247, K248, D249, Т250, L251, M252, 1253, S254, R255, Т256, Р257, Е258, V259, Т260, С261, F275, N276, W277, Y278, V279, D280, V282, Е283, V284, Н285, N286, А287, K288, Т289, K290, Р291, R292, Е293, V302, V303, S304, V305, L306, Т307, V308, L309, Н310, Q311, D312, W313, L314, N315, G316, K317, Е318, Y319, I336, S337, K338, А339, K340, G341, Q342, Р343, R344, Е345, Р346, Q347, V348, С367, V369, F372, Y373, Р374, S375, D376, I377, А378, V379, Е380, W381, Е382, S383, N384, G385, Q386, Р387, Е388, N389, Y391, Т393, S408, S424, С425, S426, V427, М428, Н429, Е430, А431, L432, Н433, N434, Н435, Y436, Т437, Q438, K439 и S440.
Исследования путем сайт-направленного мутагенеза доказали, что критическими сайтами связывания в Fc-области IgG для FcRn являются гистидин 310, гистидин 435 и изолейцин 253, и в меньшей степени гистидин 433 и тирозин 436 (см., например,. Kim, J.K., et al., Eur. J. Immunol. 29 (1999) 2819-2825; Raghavan, M., et al., Biochem. 34 (1995) 14649-14657; Medesan, C, et al., J. Immunol. 158(1997) 2211-2217).
Способы увеличения связывания IgG с FcRn были выполнены путем мутирования IgG на различных аминокислотных остатках: треонин 250, метионин 252, серии 254, треонин 256, треонин 307, глутаминовая кислота 380, метионин 428, гистидин 433 и аспарагин 434 (см. Kuo, Т.Т., et al., J. Clin. Immunol. 30 (2010) 777-789).
В некоторых случаях желательными являются антитела с уменьшенным периодом полужизни в кровотоке. Например, препараты для интравитреального введения должны иметь длинный период полужизни в глазе и короткий период полужизни в кровотоке пациента. Такие антитела также имеют преимущество, которое заключается в повышенной экспозиции на месте заболевания, например, в глазу.
Известны различные мутации, которые влияют на связывание FcRn и тем самым на период полужизни в кровотоке. Остатки Fc-области, критичные для взаимодействия мышиного Fc и мышиного FcRn, были определены с помощью сайт-направленного мутагенеза (см., например, Dall'Acqua, W.F., et al. J. Immunol 169 (2002) 5171-5180). Остатки I253, H310, H433, N434 и Н435 (нумерация ЕС согласно Kabat) участвуют в этом взаимодействии (Medesan, С, et al., Eur. J. Immunol. 26 (1996) 2533-2536; Firan, M., et al., Int. Immunol. 13 (2001) 993-1002; Kim, J.K., et al., Eur. J. Immunol. 24 (1994) 542-548). Остатки I253, H310 и H435 оказались критическими для взаимодействия человеческого Fc с мышиным FcRn (Kim, U.K., et al., Eur. J. Immunol. 29 (1999) 2819-2825). Dall'Acqua et al. описали, что остатки M252Y, S254T, T256E повышают FcRn-связывание в исследованиях белок-белковых взаимодействий (Dall'Acqua, W.F., et al. J. Biol. Chem. 281 (2006) 23514-23524). Исследования комплекса человеческого Fc с человеческим FcRn показали, что остатки I253, S254, Н435 и Y436 имеют решающее значение для этого взаимодействия (Firan, M., et al., Int. Immunol. 13 (2001) 993-1002; Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604). In Yeung, Y.A., et al. (J. Immunol. 182 (2009) 7667-7671). Были зарегистрированы и рассмотрены различные мутанты остатков с 248 по 259, с 301 по 317, с 376 по 382 и с 424 по 437. Иллюстративные мутации и их влияние на связывание FcRn перечислены в следующей таблице.
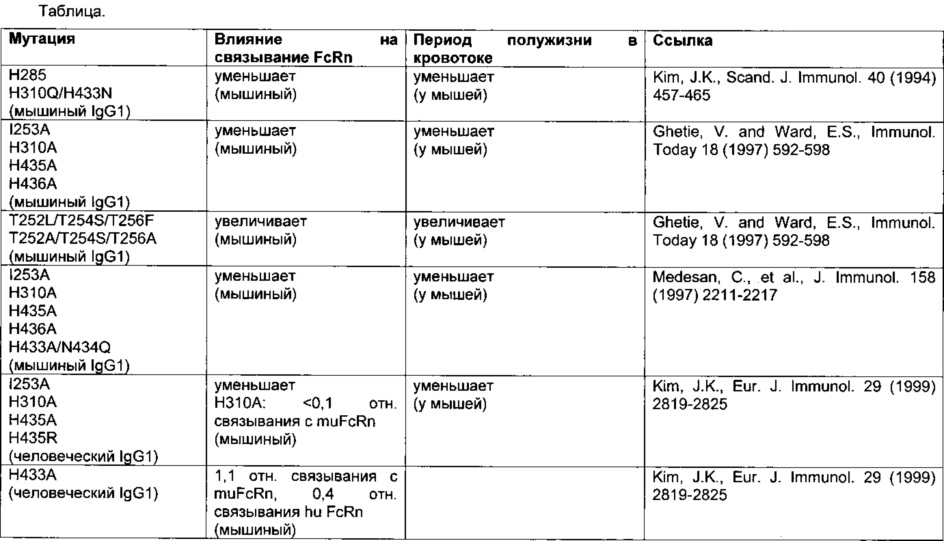
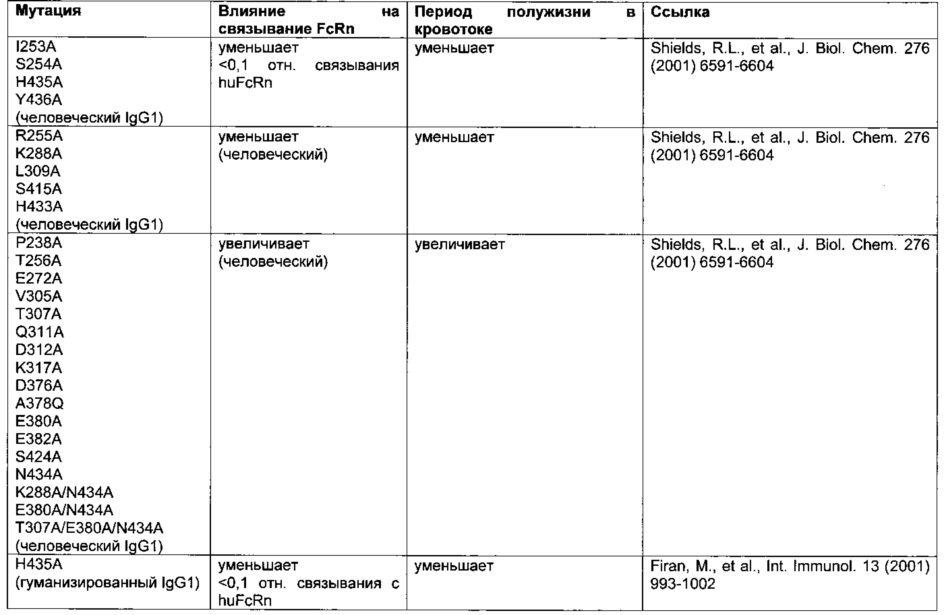
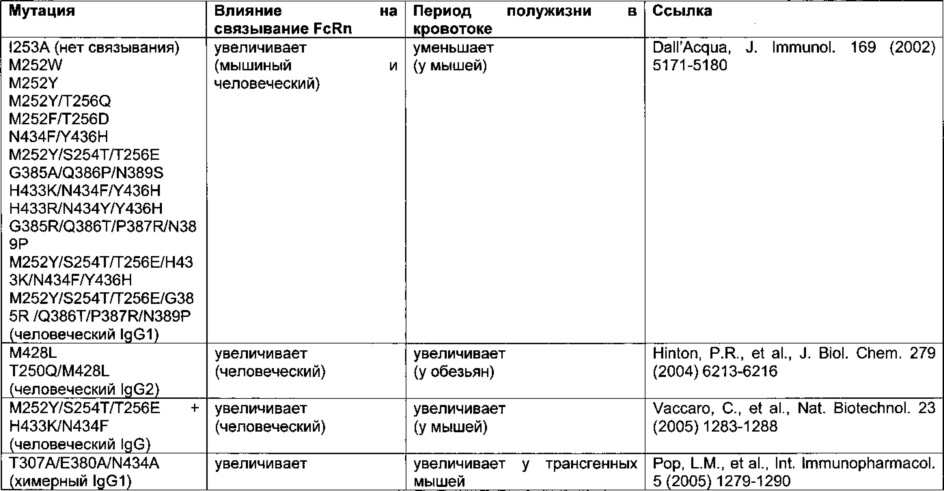
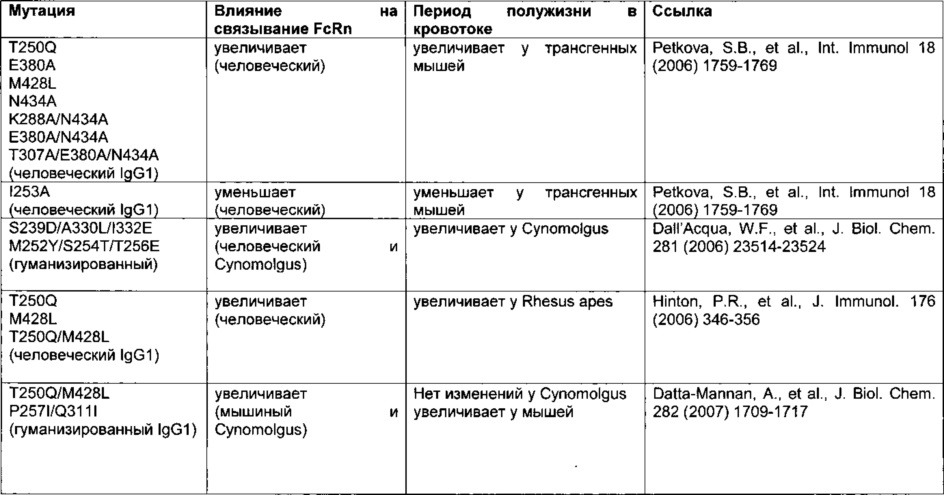
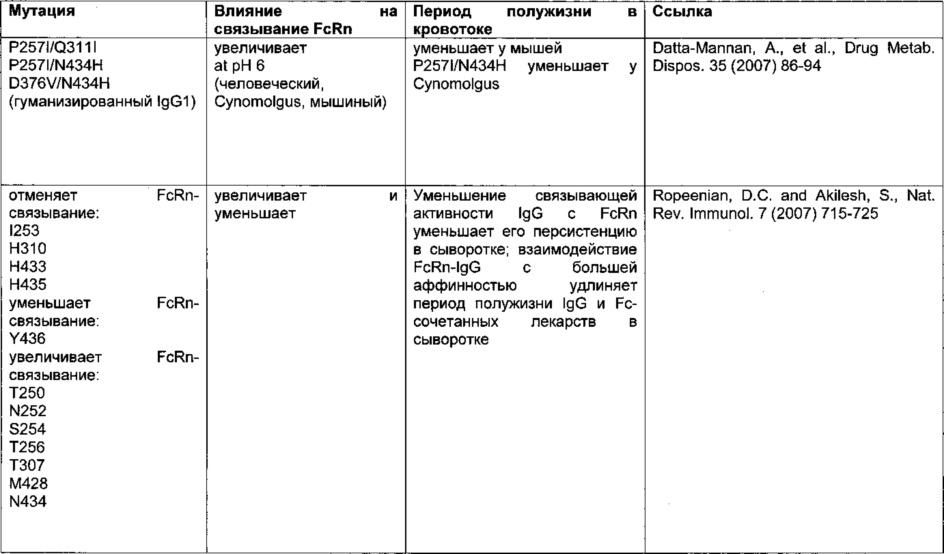
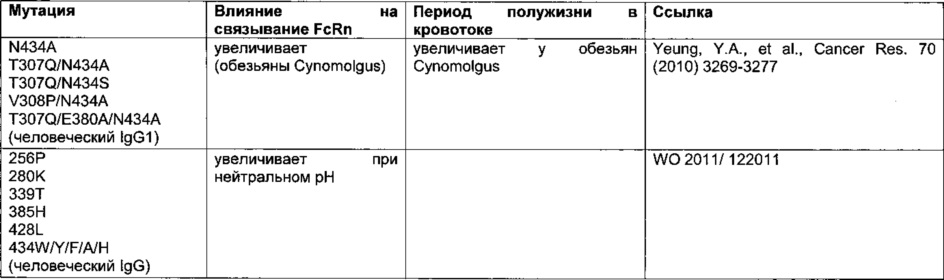
Было обнаружено, что одной мутации, односторонней в одном полипептиде Fc-области, достаточно для значительного ослабления связывания с рецептором Fc. Чем больше мутаций введено в Fc-область, тем более слабым становится связывание с FcRn. Но односторонние асимметричные мутации не являются достаточными, чтобы полностью ингибировать связывание FcRn. Для полного ингибирования связывания FcRn необходимы мутации с обеих сторон.
Таким образом, вариантная (человеческая) Fc-область класса IgG является гетеродимером, и спаривание первого полипептида Fc-области (тяжелой цепи) и второго полипептида Fc-области (тяжелой цепи) для формирования функциональной Fc-области приводит к образованию гетеродимера.
Результаты симметрично спроектированной Fc-области IgG1 для влияния на связывание FcRn показаны в следующей таблице (выравнивание мутаций и время удерживания на FcRn-аффинной хроматографической колонке).
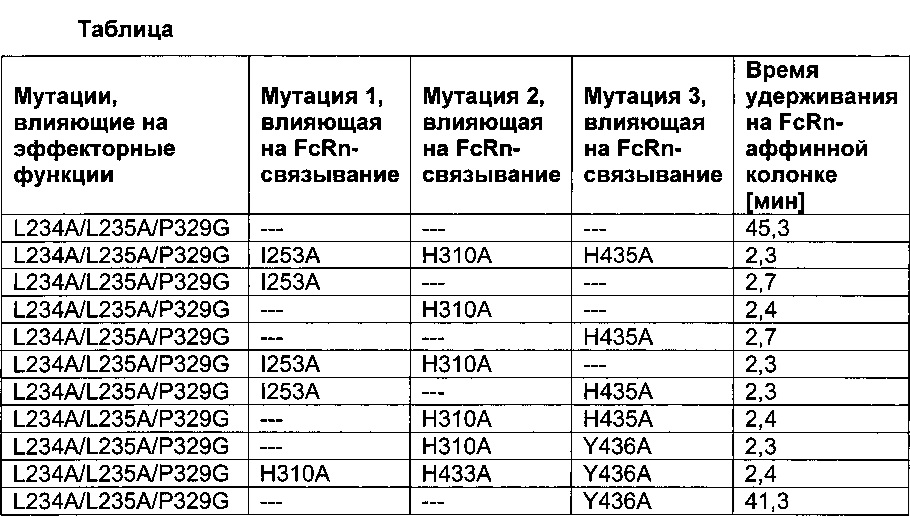
Время удерживания менее 3 минут соответствует отсутствию связывания, поскольку вещество проходит через колонку (отсутствие пика).
Одиночная мутация Н310А является наиболее молчащей симметричной мутацией, прекращающей любое FcRn-связывание.
Симметричная одиночная мутация I253A и Н435А приводит к относительному сдвигу времени удерживания 0,3-0,4 мин. Как правило, это может рассматриваться как не поддающееся обнаружению связывание.
Одиночная мутация Y436A приводит к обнаруживаемой силе взаимодействия с FcRn-аффинной колонкой. Без привязки к данной теории, эта мутация может иметь FcRn-опосредованный период полужизни, который можно отличить от нулевого взаимодействия, например, комбинации мутаций I253A, Н310А и Н435А (мутация IHH-AAA).
Результаты, полученные с симметрично модифицированным антителом к HER2, представлены в следующей таблице (для справки см. WO 2006/031370).
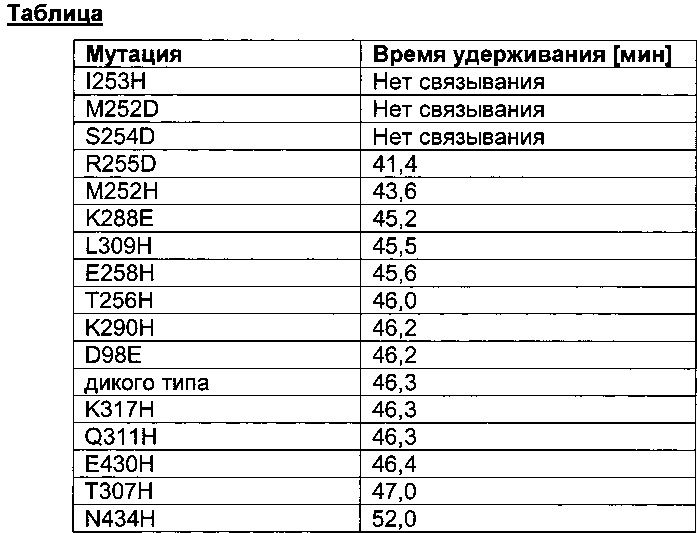
Эффект от введения в Fc-область асимметричных мутаций, влияющих на FcRn-связывание, был проиллюстрирован биспецифическим анти-VEGF/ANG2-антителом (см. ниже), собранным с использованием технологии "ключ в замке" (см., например, US 7695936, US 2003/0078385; мутации "цепи-ключа": S354C/T366W, мутации "цепи-замка": Y349C/T366S/L368A/Y407V). Влияние асимметрично вводимых мутаций на связывание с FcRn может быть легко определено с использованием методики FcRn-аффинной хроматографии (см. фиг. 9 и следующую таблицу). Антитела, которые имеют более позднюю элюцию с аффинной колонки FcRn, т.е. имеют более длительное время удерживания на FcRn-аффинной колонке, имеют длительный период полужизни in vivo, и наоборот.

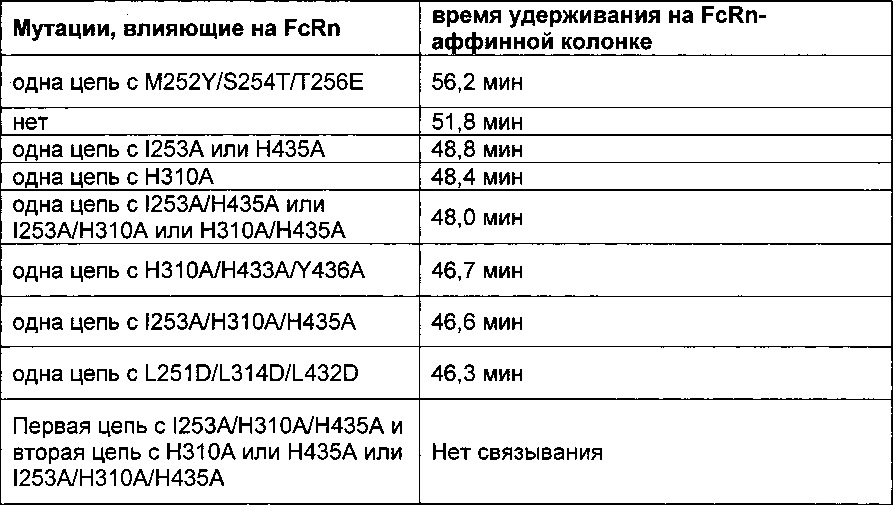
Асимметричные мутации IHH-AAA- и LLL-DDD (LLL-DDD-мутация = L251D, L314D и L432D) демонстрируют более слабое связывание, чем соответствующее родительское антитело или антитело дикого типа.
Симметричная мутация HHY-AAA (= комбинация мутаций Н310А, Н433А и Y436A) приводит к Fc-области, которая более не связывается с человеческим FcRn, в то время как связывание с белком А сохраняется (см. фиг.11, 12, 13 и 14).
Эффект от введения в Fc-область асимметричных мутаций, влияющих на FcRn-связывание, также было проиллюстрировано биспецифическим анти-VEGF/АМС2-антителом (VEGF/ANG2), моноспецифическим анти-IGF-IR-антителом (IGF-1R) и полноразмерным антителом со слиянием с С-концом обеих тяжелых цепей (гибрид), собранным с использованием технологии "ключ в замке" для того, чтобы позволить введение асимметричных мутаций (см., например, US 7695936, US 2003/0078385; мутации "цепи-ключа": S354C/T366W, мутации "цепи-замка": Y349C/T366S/L368A/Y407V). Влияние асимметрично введенных мутаций на связывание с FcRn и белком А может быть легко определено с использованием методики FcRn-аффинной хроматографии, методики аффинной хроматографии на белке А и методик на основании SPR (см. следующую таблицу).
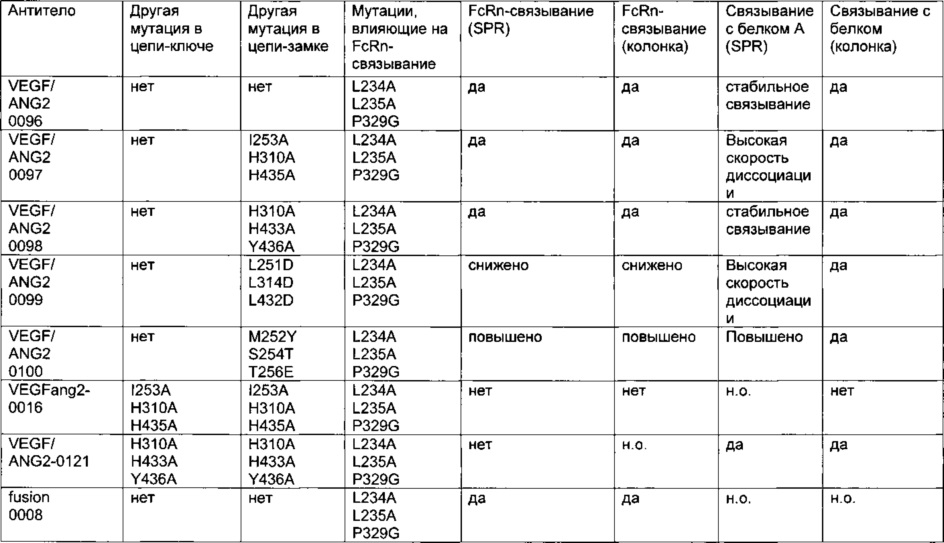
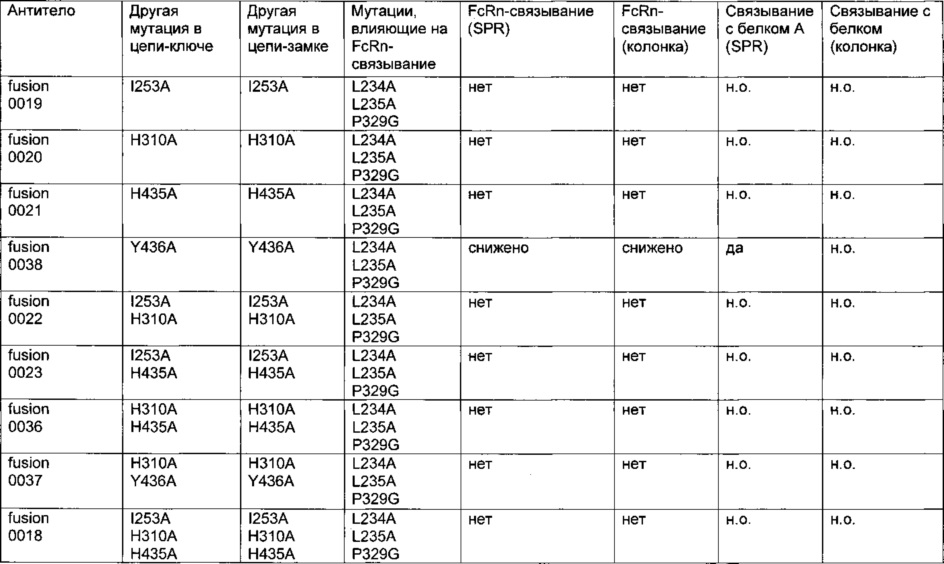
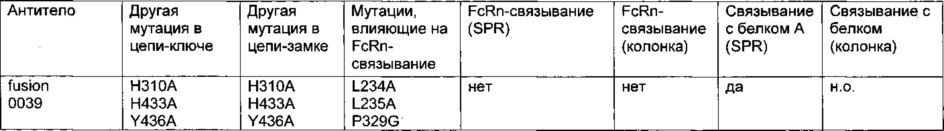
Сочетание мутаций I253A, Н310А, Н435А, или L251D, L314D, L432D, или L251S, L314S, L432S приводит к потере связывания с белком А, в то время как сочетание мутаций I253A, Н310А, Н435А, или Н310А, Н433А, Y436A, или L251D, L314D, L432D приводит к потере связывания с человеческим неонатальным Fc-рецептором.
Один из аспектов, описанных в данном документе, представляет собой антитело или гибридный полипептид Fc-области, содержащий вариантную человеческую Fc-область класса IgG, описанную в данном документе.
Fc-область в гибридном полипептиде Fc-области имеет описанные выше характеристики ее партнера, с которым она слита. Партнер по слиянию может быть любой молекулой, имеющей биологическую активность, период полужизни которой in vivo должен быть уменьшен или увеличен, то есть период полужизни которой должен быть четко определен и сделан индивидуальным для его предполагаемого применения.
Гибридные полипептиды Fc-области могут содержать, например, вариантную (человеческую) Fc-область класса IgG, описанную в данном документе, и рецепторный белок, который связывается с мишенью, включающей лиганд, такой как, например, гибридный полипептид TNFR-Fc-область (TNFR = человеческий рецептор фактора некроза опухоли), или гибридный полипептид IL-IR-Fc-область (IL-1R = человеческий рецептор интерлейкина-1), или гибридный полипептид VEGFR-Fc-область (VEGFR = человеческий рецептор фактора роста эндотелия сосудов), или гибридный полипептид ANG2R-Fc-область (ANG2R = человеческий рецептор ангиопоэтина 2).
Гибридные полипептиды Fc-области могут содержать, например, вариантную (человеческую) Fc-область класса IgG, описанную в данном документе, и фрагмент антитела, который связывается с мишенью, в том числе такой как, например, Fab-фрагмент антитела, scFv (см., например, Nat. Biotechnol. 23 (2005) 1126-1136), или доменные антитела (dAb) (см., например, WO 2004/058821, WO 2003/002609).
Гибридные полипептиды Fc-области могут содержать, например, вариантную (человеческую) Fc-область класса IgG, описанную в данном документе, и рецепторный лиганд (природный или искусственный).
В. Примеры антител
В одном аспекте данное изобретение предусматривает изолированные антитела, которые обладают модифицированным FcRn-связыванием, т.е. эти антитела связываются с человеческим FcRn с аффинностью, более высокой или более низкой, чем антитело, не имеющее мутации, затрагивающей FcRn-связывание.
В одном воплощении антитело включает Fc-область, содержащую первый полипептид Fc-области и второй полипептид Fc-области, где
a) первый полипептид Fc-области и второй полипептид Fc-области получены из одного того же полипептида человеческой Fc-области, и
b) первый полипептид Fc-области был модифицирован так, что его аминокислотная последовательность отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat, а второй полипептид Fc-области был модифицирован так, что его аминокислотная последовательность отличается от аминокислотной последовательности первого полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat, где модифицированная позиция в первом полипептиде Fc-области и модифицированная позиция во втором полипептиде Fc-области являются разными, и
c) Fc-область имеет аффинность к человеческому Fc-рецептору, отличающуюся от Fc-области, которая содержит в качестве первого и второго полипептида Fc-области полипептид человеческой Fc-области а) (т.е. имеет те же аминокислотные остатки, что и полипептид человеческой Fc-области а) в соответствующих позициях в соответствии с индексом ЕС по системе нумерации Kabat).
Одним из примеров антител, описанных в данном документе, а также одним из аспектов данного изобретения является биспецифическое, двухвалентное антитело с отмененным FcRn-связыванием, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2,
где
i) первый антигенсвязывающий сайт, специфически связывающийся с VEGF, содержит в вариабельном домене тяжелой цепи CDR3H-область с SEQ ID №14, CDR2H-область с SEQ ID №15 и CDR1H-область с SEQ ID №16, а в вариабельном домене легкой цепи CDR3L-область с SEQ ID №17, CDR2L-область с SEQ ID №18 и CDR1L-область с SEQ ID №19, и
ii) второй антигенсвязывающий сайт, специфически связывающийся с ANG-2, содержит в вариабельном домене тяжелой цепи CDR3H-область с SEQ ID №22, CDR2H-область с SEQ ID №23 и CDR1H-область с SEQ ID №24, а в вариабельном домене легкой цепи CDR3L-область с SEQ ID №25, CDR2L-область с SEQ ID №26 и CDRIL-область с SEQ ID №27, и
iii) биспецифическое антитело включает Fc-область, содержащую первый полипептид Fc-области и второй полипептид Fc-области,
где
a) первый полипептид Fc-области и второй полипептид Fc-области получены из одного и того же полипептида человеческой Fc-области, и
b) первый полипептид Fc-области был модифицирован так, что его аминокислотная последовательность отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat, а второй полипептид Fc-области был модифицирован так, что его аминокислотная последовательность отличается от аминокислотной последовательности первого полипептида Fc-области по меньшей мере в одной соответствующей позиции в соответствии с индексом ЕС по системе нумерации Kabat, где модифицированная позиция в первом полипептиде Fc-области и модифицированная позиция во втором полипептиде Fc-области являются различными, и
c) Fc-область имеет аффинность к человеческому Fc-рецептору, которая отличается от Fc-области, которая содержит в качестве первого и второго полипептида Fc-области полипептид человеческой Fc-области а) (т.е. имеет те же аминокислотные остатки, что и полипептид человеческой Fc-области а) в соответствующих позициях в соответствии с индексом ЕС по системе нумерации Kabat).
В одном воплощении всех аспектов Fc-область представляет собой вариантную (человеческую) Fc-область класса IgG. В одном воплощении вариантная (человеческая) Fc-область класса IgG представляет собой гетеродимер Fc-области класса IgG.
В одном воплощении всех аспектов спаривание первого полипептида Fc-области и второго полипептида Fc-области для образования (функциональной) Fc-области приводит к образованию гетеродимера.
В одном воплощении полипептид человеческой Fc-области представляет собой полипептид человеческой Fc-области подкласса IgG1 или подкласса IgG4.
В одном воплощении полипептид человеческой Fc-области представляет собой полипептид человеческой Fc-области подкласса IgG1, который также содержит мутации L234A, L235A и P329G.
В одном воплощении полипептид человеческой Fc-области представляет собой полипептид человеческой Fc-области подкласса IgG4, который также содержит мутации S228P и L235E.
В одном воплощении первый полипептид Fc-области также содержит мутации S354C и T366W, а второй полипептид Fc-области также содержит мутации Y349C, T366S, L368A и Y407V.
В одном воплощении биспецифическое антитело характеризуется тем, что
i) первый антигенсвязывающий сайт, специфически связывающийся с VEGF, включает в качестве вариабельного домена тяжелой цепи VH аминокислотную последовательность SEQ ID №20, а в качестве вариабельного домена легкой цепи VL аминокислотную последовательность SEQ ID №21, и
ii) второй антигенсвязывающий сайт, специфически связывающийся с ANG-2, включает в качестве вариабельного домена тяжелой цепи VH аминокислотную последовательность SEQ ID №28, а в качестве вариабельного домена легкой цепи VL аминокислотную последовательность SEQ ID №29.
В одном воплощении биспецифическое антитело характеризуется тем, что Fc-область iii) относится к человеческому подклассу IgG1. В одном воплощении биспецифическое антитело характеризуется тем, что Fc-область человеческого подкласса IgG1 также содержит мутации L234A, L235A и P329G (нумерация согласно индексу ЕС по Kabat).
В одном воплощении биспецифическое антитело характеризуется тем, что Fc-область iii) относится к человеческому подклассу IgG4. В одном воплощении биспецифическое антитело характеризуется тем, что Fc-область человеческого подкласса IgG4 также содержит мутации S228P и L235E (нумерация согласно индексу ЕС по Kabat). В одном воплощении биспецифическое антитело характеризуется тем, что Fc-область человеческого подкласса IgG4 также содержит мутации S228P, L235A и P329G (нумерация согласно индексу ЕС по Kabat).
В одном воплощении биспецифическое антитело содержит Fc-область, содержащую первый и второй полипептид Fc-области, оба человеческого подкласса IgG1 или IgG4 (т.е. человеческого происхождения), которая включает одну или две мутации, выбранные из i) группы I253A, Н310А, Н435А, или ii) группы Н310А, Н433А, Y436A, или iii) группы L251D, L314D, L432D, или iv) группы L251S, L314S, L432S (в соответствии с индексом ЕС по системе нумерации Kabat) в первом полипептиде Fc-области, и одну или две мутации, выбранные из группы, включающей мутации L251D, L251S, I253A, Н310А, L314D, L314S, L432D, L432S, Н433А, Н435А и Y436A (в соответствии с индексом ЕС по системе нумерации Kabat) во втором полипептиде Fc-области, так что все мутации i) I253A, Н310А, Н435А, или ii) Н310А, Н433А, Y436A, или iii) L251D, L314D, L432D, или iv) L251S, L314S, L432S содержатся в Fc-области.
В одном воплощении биспецифическое антитело включает Fc-область, содержащую первый и второй полипептид Fc-области, оба человеческого IgG1 подкласса или IgG4 (т.е. человеческого происхождения), которые содержат мутации I253A/H310A/H435A, или H310A/H433A/Y436A, или L251D/L314D/L432D, или L251S/L314S/L432S, или их комбинации в Fc-области (в соответствии с индексом ЕС по системе нумерации Kabat), где либо все мутации находятся в первом или втором полипептиде Fc-области, либо одна или две мутации находятся в первом полипептиде Fc-области, либо одна или две мутации находятся во втором полипептиде Fc-области, так что все мутации i) I253A, Н310А, Н435А, или ii) Н310А, Н433А, Y436A, или iii) L251D, L314D, L432D, или iv) L251S, L314S, L432S содержатся в Fc-области.
Кроме того, другими аспектами, описанными в данном документе, являются фармацевтический препарат, содержащий биспецифическое антитело, фармацевтический препарат для применения в лечении глазных сосудистых заболеваний, применение биспецифических антител для изготовления лекарственного средства для лечения глазных сосудистых заболеваний, способ лечения пациентов, страдающих от глазных сосудистых заболеваний, путем введения биспецифического антитела пациенту, нуждающемуся в таком лечении. В одном воплощении биспецифическое антитело или фармацевтический препарат, содержащий биспецифическое антитело, вводят интравитреально.
Еще один аспект согласно данному изобретению представляет собой нуклеиновокислотную молекулу, кодирующую тяжелую и/или легкую цепь биспецифического антитела, описанного в данном документе.
Данное изобретение также предусматривает экспрессионные векторы, содержащие нуклеиновую кислоту, описанную в данном документе, способные экспрессировать нуклеиновую кислоту в прокариотической или эукариотической клетке-хозяине, и клетки-хозяева, содержащие такие векторы, для рекомбинантной продукции биспецифического антитела, описанного в данном документе.
Данное изобретение также включает прокариотическую или эукариотическую клетку-хозяина, содержащую вектор, описанный в данном документе.
Данное изобретение также включает способ получения биспецифического антитела, описанного в данном документе, который характеризуется экспрессией нуклеиновой кислоты, описанной в данном документе, в прокариотической или эукариотической клетке-хозяине и выделение биспецифического антитела из клетки или супернатанта клеточной культуры. Одно воплощение представляет собой способ получения биспецифического антитела, описанного в данном документе, включающий этапы
a) трансформации клетки-хозяина векторами, содержащими нуклеиновокислотные молекулы, кодирующие антитело;
b) культивирования клетки-хозяина в условиях, позволяющих синтез антитела; и
c) выделения антитела из культуры.
Изобретение также включает антитело, полученное таким способом, для продукции биспецифического антитела.
Антитела, описанные в данном документе, имеют высокоценные свойства благодаря их специфическим модификациям в Fc-области, дающим пользу для пациента, страдающего от глазных сосудистых заболеваний. Они демонстрируют высокую стабильность в среде стекловидного тела и медленную диффузию из глаза (по сравнению с фрагментами антител меньшего размера без константной области тяжелой цепи), где заболевание фактически расположено и подвергается лечению (так схема лечения может быть потенциально улучшена по сравнению с не-IgG-подобными антителами, такими как, например, фрагменты Fab и (Fab)2). С другой стороны, антитела, описанные в данном документе, будут довольно быстро выводиться из сыворотки (что очень желательно для уменьшения потенциальных побочных эффектов, возникающих из-за системного воздействия). Удивительно, но они также демонстрируют более низкую вязкость (см. фиг. 2) (по сравнению с версиями без комбинации мутаций I253A, Н310А и Н435А в константной области) и, следовательно, особенно полезны для интравитреального применения через тонкие иглы при лечении глазных заболеваний (для такого применения обычно используются тонкие иглы, и высокая вязкость делает соответствующее применение довольно трудным). Более низкая вязкость также позволяет делать препараты с более высокой концентрацией.
Также удивительно, что антитела, описанные в данном документе, демонстрируют более низкую тенденцию к агрегации (см. фиг. 4) в процессе хранения (по сравнению с версиями без комбинации мутаций I253A, Н310А и Н435А в Fc-области), что является критическим для интравитреального введения в глаз (поскольку агрегация в глазе может привести к осложнениям во время такого лечения).
Биспецифические антитела, описанные в данном документе, демонстрируют хорошую эффективность в ингибировании сосудистых заболеваний.
В некоторых воплощениях биспецифические антитела, описанные в данном документе, из-за их специфических модификаций в константной области (например, P329G LALA) демонстрируют ценные свойства, такие как отсутствие связывания с Fc-гамма-рецепторами, что снижает риск побочных эффектов, таких как тромбоз и/или нежелательная клеточная гибель (например, в результате ADCC).
В одном воплощении, описанном в данном документе, биспецифическое антитело, описанное в данном документе, является двухвалентным.
В одном из аспектов в соответствии с данным изобретением биспецифическое двухвалентное антитело, описанное в данном документе, характеризуется тем, что содержит
a) тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и
b) модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, где константные домены CL и СН1 заменены друг на друга.
Этот биспецифический двухвалентный формат биспецифического антитела, специфически связывающегося с человеческим фактором роста сосудистого эндотелия (VEGF) и человеческим ангиопоэтином-2 (ANG-2), описан в заявке WO 2009/080253 (включая СН3-домены, модифицированные по типу "ключ в замке"). Антитела, основанные на этом биспецифическом двухвалентном формате антитела, называются CrossMab.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №38, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №40, и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №39, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №41.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №34, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №36, и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №35, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №37.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №42, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №44, и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №43, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №45.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №90, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №40, и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №91, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №41.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №88, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №36, и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №89, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №37.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №92, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №44, и
b) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №93, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №45.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №38, SEQ ID №39, SEQ ID №40 и SEQ ID №41.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №34, SEQ ID №35, SEQ ID №36 и SEQ ID №37.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №42, SEQ ID №43, SEQ ID №44 и SEQ ID №45.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №90, SEQ ID N2 91, SEQ ID №40 и SEQ ID №41.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №88, SEQ ID №89, SEQ ID №36 и SEQ ID №37.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №92, SEQ ID №93, SEQ ID №44 и SEQ ID №45.
В другом аспекте, описанном в данном документе, биспецифическое антитело, описанное в данном документе, характеризуется тем, что содержит
a) тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и
b) тяжелую цепь и легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, где N-конец тяжелой цепи соединен с С-концом легкой цепи с помощью пептидного линкера.
Этот биспецифический двухвалентный формат антитела для этого биспецифического антитела, специфически связывающегося с человеческим фактором роста сосудистого эндотелиального (VEGF) и человеческим ангиопоэтином-2 (ANG-2), описан в заявке WO 2011/117330 (включая СН3-домены, модифицированные по типу "ключ в замке"). Антитела, основанные на этом биспецифическом двухвалентном формате антитела, называются одноцепочечными Fab-фрагментами с одной ветвью (OAscFabs).
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №46, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №48, и
b) в качестве тяжелой цепи второго полноразмерного антитела, соединенной через пептидный линкер с легкой цепью второго полноразмерного антитела, аминокислотную последовательность SEQ ID №47.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №49, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №51, и
b) в качестве тяжелой цепи второго полноразмерного антитела, соединенной через пептидный линкер с легкой цепью второго полноразмерного антитела, аминокислотную последовательность SEQ ID №50.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №94, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №48, и
b) в качестве тяжелой цепи второго полноразмерного антитела, соединенной через пептидный линкер с легкой цепью второго полноразмерного антитела, аминокислотную последовательность SEQ ID №95.
В одном воплощении такое биспецифическое двухвалентное антитело характеризуется тем, что содержит
a) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №96, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №51, и
b) в качестве тяжелой цепи второго полноразмерного антитела, соединенной через пептидный линкер с легкой цепью второю полноразмерного антитела, аминокислотную последовательность SEQ ID №97.
В одном воплощении вариабельный домен тяжелой цепи антитела (VH) и вариабельный домен легкой цепи антитела (VL) тяжелой и легкой цепей второго полноразмерного антитела дополнительно стабилизированы введением дисульфидного мостика между следующими позициями: позиция 44 вариабельного домена тяжелой цепи и позиция 100 вариабельного домена легкой цепи (нумерация согласно Kabat (Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Методики введения дисульфидных мостиков для стабилизации описаны, например, в WO 94/029350, Rajagopal, V., et al, Prot. Eng. 10 (1997) 1453-1459, Kobayashi et al., Nuclear Medicine & Biology 25 (1998) 387-393, и в Schmidt, M., etal., Oncogene 18(1999) 1711-1721.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №46, SEQ ID №47 и SEQ ID №48.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №49, SEQ ID №50 и SEQ ID №51.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №94, SEQ ID №95 и SEQ ID №48.
Соответственно, один аспект, описанный в данном документе, представляет собой биспецифическое двухвалентное антитело, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, которое характеризуется тем, что включает аминокислотные последовательности SEQ ID №96, SEQ ID №97 и SEQ ID №51.
В одном воплощении СН3-домены биспецифического двухвалентного антитела, описанного в данном документе, изменены с помощью технологии "ключ в замке", которая подробно описана на нескольких примерах, например в WO 96/027011, Ridgway J.B., et al., Protein Eng. 9 (1996) 617-621, и в Merchant, A.M., et al., Nat. Biotechnol. 16 (1998) 677-681. В этом способе поверхности взаимодействия двух СН3-доменов изменены так, чтобы увеличить гетеродимеризацию обеих тяжелых цепей, содержащих эти два СН3-домена. Каждый из двух СН3-доменов (двух тяжелых цепей) может быть "ключом", а другой "замком". Введение дисульфидного мостика также стабилизирует гетеродимеры (Merchant, А.М, et al., Nature Biotech. 16 (1998) 677-681, Atwell, S., et al. J. Mol. Biol. 270 (1997) 26-35) и повышает выход.
В одном предпочтительном воплощении всех аспектов, описанном в данном документе, биспецифические антитела характеризуются тем, что
СН3-домен одной тяжелой цепи и СН3-домен другой тяжелой цепи встречаются на контактной поверхности, которая включает исходную контактную поверхность между СН3-доменами антитела;
где указанная контактная поверхность изменена таким образом, чтобы способствовать формированию биспецифического антитела, где изменение характеризуется тем, что
a) СН3-домен одной тяжелой цепи изменен так,
что в исходной контактной поверхности СН3-домена одной тяжелой цепи, которая встречается с исходной контактной поверхностью СН3-домена другой тяжелой цепи в биспецифическом антителе,
аминокислотный остаток заменен аминокислотным остатком, имеющим больший объем боковой цепи, тем самым формируя выступ в контактной поверхности СН3-домена одной тяжелой цепи, который может быть расположен в полости на контактной поверхности СН3-домена другой тяжелой цепи
и
b) СН3-домен другой тяжелой цепи изменен так,
что в исходной контактной поверхности второго СН3-домена, которая встречается с исходной контактной поверхностью первого СН3-домена в биспецифическом антителе,
аминокислотный остаток заменен на аминокислотный остаток, имеющий меньший объем боковой цепи, тем самым формируя полость в контактной поверхности второго СН3-домена, в которой может быть расположен выступ на контактной поверхности первого СН3-домена.
Таким образом, в одном предпочтительном воплощении антитело согласно изобретению характеризуется тем, что
СН3-домен тяжелой цепи полноразмерного антитела а) и СН3-домен тяжелой цепи полноразмерного антитела b) встречаются на контактной поверхности, которая включает изменение в исходной контактной поверхности между СН3-доменами антитела;
где
i) в СН3-домене одной тяжелой цепи
аминокислотный остаток заменен аминокислотным остатком, имеющим больший объем боковой цепи, тем самым формируя выступ в контактной поверхности СН3-домена одной тяжелой цепи, который может быть расположен в полости на контактной поверхности СН3-домена другой тяжелой цепи,
и
ii) в СН3-домене другой тяжелой цепи
аминокислотный остаток заменен на аминокислотный остаток, имеющий меньший объем боковой цепи, тем самым формируя полость в контактной поверхности второго СН3-домена, в которой может быть расположен выступ на контактной поверхности первого СН3-домена.
В одном предпочтительном воплощении указанный аминокислотный остаток, имеющий боковую цепь большего объема, выбран из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y), триптофана (W).
В одном предпочтительном воплощении указанный аминокислотный остаток, имеющий боковую цепь меньшего объема, выбран из группы, состоящей из аланина (А), серина (S), треонина (Т), валина (V).
В одном воплощении оба СН3-домена также изменены путем введения остатка цистеина (С) в соответствующих позициях каждого СН3-домена, так что между двумя СН3-доменами может быть сформирован дисульфидный мостик.
В одном воплощении биспецифическое антитело содержит мутацию T366W в СН3-домене "цепи-ключа" и мутации T366S, L368A, Y407V в СН3-домене "цепи-замка". Также может быть использован дополнительный межцепочечный дисульфидный мостик между СН3-доменами (Merchant, A.M, et al., Nature Biotech 16 (1998) 677-681), например, путем введения мутации Y349C или S354C в СН3-домен "цепи-ключа" и мутации Y439C или Е356С или S354C в СН3-домен "цепи-замка".
В одном воплощении указанное биспецифическое антитело, описанное в данном документе, содержит мутацию Y349C или S354C и мутацию T366W в одном из двух СН3-доменов и мутацию S354C или Е356С или Y439C и мутации T366S L368A и Y407V в другом из двух СН3-доменов. В одном предпочтительном воплощении указанное биспецифическое антитело содержит мутации Y349C, T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A, Y407V в другом из двух СН3-доменов (дополнительная мутация Y349C в одном СН3-домене и дополнительная мутация S354C в другом СН3-домене для формирования межцепочечного дисульфидного мостика) (нумерация согласно индексу ЕС по Kabat (Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). В одном предпочтительном воплощении указанное биспецифическое антитело содержит мутации S354C, T366W в одном из двух СН3-доменов и мутации Y439C, T366S, L368A, Y407V в другом из двух СН3-доменов (дополнительная мутация Y349C в одном СН3-домене и дополнительная мутация S354C в другом СН3-домене для формирования межцепочечного дисульфидного мостика) (нумерация согласно индексу ЕС по Kabat (Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)).
Альтернативно или дополнительно также могут быть использованы и другие технологии типа "ключ-замок", такие как описанные в ЕР 1870459 А1. Таким образом, другим примером указанного биспецифического антитела являются мутации R409D и K370E в СН3-домене "цепи-ключа" и мутации D399K и E357K в СН3-домене "цепи-замка" (нумерация согласно индексу ЕС по Kabat (Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)).
В другом воплощении указанное биспецифическое антитело содержит мутацию T366W в СН3-домене "цепи-ключа" и мутации T366S, L368A и Y407V в СН3-домене "цепи-замка", и дополнительно мутации R409D, K370E в СН3-домене "цепи-ключа" и мутации D399K, E357K в СН3-домене "цепи-замка".
В одном воплощении указанное биспецифическое антитело содержит мутации Y349C, T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A и Y407V в другом из двух СН3-доменов, или указанное биспецифическое антитело содержит мутации Y349C, T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A и Y407V в другом из двух СН3-доменов, и дополнительно мутации R409D, K370E в СН3-домене "цепи-ключа" и мутации D399K, E357K в СН3-домене "цепи-замка".
В одном воплощении указанное биспецифическое антитело содержит мутации S354C, T366W в одном из двух СН3-доменов и мутации Y349C, T366S, L368A, Y407V в другом из двух СН3-доменов, или указанное биспецифическое антитело содержит мутации S354C, T366W в одном из двух СН3-доменов и мутации Y349C, T366S, L368A и Y407V в другом из двух СН3-доменов, и дополнительно мутации R409D, K370E в СН3-домене "цепи-ключа" и мутации D399K, E357K в СН3-домене "цепи-замка".
В одном воплощении биспецифическое антитело, описанное в данном документе, характеризуется тем, что имеет одно или более чем одно из следующих свойств:
- демонстрирует более низкую сывороточную концентрацию по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (через 96 часов после интравитреального введения мышам, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn) (определяли с помощью анализов, описанных в примере 6),
- демонстрирует аналогичную (коэффициент от 0,8 до 1,2) концентрацию в цельных лизатах правого глаза по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (у мышей, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn, через 96 часов после интравитреального введения в правый глаз) (определяли с помощью анализов, описанных в примере 6),
- не демонстрирует связывания с человеческим неонатальным Fc-рецептором.
В одном воплощении биспецифическое двухвалентное антитело характеризуется тем, что содержит первый полипептид Fc-области и второй полипептид Fc-области, где
a) первый и второй полипептид Fc-области содержит мутацию Y436A, или
b) первый и второй полипептид Fc-области содержит мутации I253A, Н310А и H435A, или
c) первый и второй полипептид Fc-области содержит мутации H310A, Н433А и Y436A, или
d) первый и второй полипептид Fc-области содержит мутации L251D, L314D и L432D, или
e) первый и второй полипептид Fc-области содержит мутации L251S, L314S и L432S, или
f) первый полипептид Fc-области содержит мутацию Y436A, а второй полипептид Fc-области содержит
- мутации I253A, Н310А и Н435А, или
- мутации Н310А, Н433А и Y436A, или
- мутации L251D, L314D и L432D, или
- мутации L251S, L314S и L432S, или
g) первый полипептид Fc-области содержит мутации I253A, Н310А и Н435А, а второй полипептид Fc-области содержит
- мутации Н310А, Н433А и Y436A, или
- мутации L251D, L314D и L432D, или
- мутации L251S, L314S и L432S, или
h) первый полипептид Fc-области содержит мутации Н310А, Н433А и Y436A, а второй полипептид Fc-области содержит
а) мутации L251D, L314D и L432D, или
b) мутации L251S, L314S и L432S,
или
i) первый полипептид Fc-области содержит мутации L251D, L314D и L432D, а второй полипептид Fc-области содержит а) мутации L251S, L314S и L432S.
В одном воплощении биспецифическое антитело содержит Fc-область, содержащую первый и второй полипептид Fc-области человеческого IgG1 или человеческого IgG4 (человеческого происхождения), которые содержат одну или две мутации, выбранные из i) группы I253A, Н310А и Н435А, или ii) группы Н310А, Н433А и Y436A, или iii) группы L251D, L314D и L432D, или iv) группы L251S, L314S и L432S (нумерация согласно индексу ЕС по системе нумерации Kabat) в первом полипептиде Fc-области, и одну или две мутации, выбранные из группы, включающей мутации L251D, L251S, I253A, Н310А, L314D, L314S, L432D, L432S, Н433А, Н435А и Y436A (нумерация согласно индексу ЕС по системе нумерации Kabat) во втором полипептиде Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A, или iii) L251D, L314D и L432D, или iv) L251S, L314S и L432S содержатся в Fc-области.
В одном воплощении биспецифическое антитело содержит Fc-область, содержащую первый и второй полипептид Fc-области человеческого IgG1 или человеческого IgG4 (т.е. человеческого происхождения), которая содержит мутации I253A/H310A/H435A, или H310A/H433A/Y436A, или L251D/L314D/L432D, или L251S/L314S/L432S, или их комбинации в Fc-области (нумерация согласно индексу ЕС по системе нумерации Kabat), где все мутации находятся в первом или втором полипептиде Fc-области, или одна или две мутации находятся в первом полипептиде Fc-области, и одна или две мутации находятся во втором полипептиде Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A, или iii) L251D, L314D и L432D, или iv) L251S, L314S и L432S находятся в Fc-области.
В одном воплощении биспецифическое двухвалентное антитело характеризуется тем, что содержит
первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, и характеризуется тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №20, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №21, и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №28, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №29, и
iii) биспецифическое антитело содержит Fc-область, содержащую первый и второй полипептид Fc-области как человеческого IgG1, так и человеческого IgG4 (т.е. человеческого происхождения), которая включает одну или две мутации, выбранные из i) группы I253A, Н310А, Н435А, или ii) группы Н310А, Н433А, Y436A, или iii) группы L251D, L314D, L432D, или iv) группы L251S, L314S, L432S (в соответствии с индексом ЕС по системе нумерации Kabat) в первом полипептиде Fc-области, и одну или две мутаций, выбранные из группы, включающей мутации L251D, L251S, I253A, Н310А, L314D, L314S, L432D, L432S, Н433А, Н435А и Y436A (в соответствии с индексом ЕС по системе нумерации Kabat) во втором полипептиде Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А, Н435А, или ii) Н310А, Н433А, Y436A, или iii) L251D, L314D, L432D, или iv) L251S, L314S, L432S содержатся в Fc-области.
и имеет одно или более чем одно из следующих свойств:
- демонстрирует более низкую сывороточную концентрацию по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (через 96 часов после интравитреального введения мышам, которые являются FcRn-дефицитными, но гемизиготно трансгенны по человеческому FcRn) (определяли с помощью анализов, описанных в примере 6),
- демонстрирует аналогичную (коэффициент от 0,8 до 1,2) концентрацию в цельных лизатах правого глаза по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (у мышей, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn, через 96 часов после интравитреального введения в правый глаз) (определяли с помощью анализов, описанных в примере 6),
- не демонстрирует связывания с человеческим неонатальным Fc-рецептором.
В одном воплощении биспецифическое двухвалентное антитело характеризуется тем, что содержит
первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, и характеризуется тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №20, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №21, и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №28, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №29, и
iii) биспецифическое антитело содержит первый полипептид Fc-области и второй полипептид Fc-области, где
a) первый и второй полипептид Fc-области содержит мутацию Y436A,
или
b) первый и второй полипептид Fc-области содержит мутации I253A, H310A и Н435А, или
c) первый и второй полипептид Fc-области содержит мутации H310A, H433A и Y436A, или
d) первый и второй полипептид Fc-области содержит мутации L251D, L314D и L432D, или
e) первый и второй полипептид Fc-области содержит мутации L251S, L314S и L432S, или
f) первый полипептид Fc-области содержит мутацию Y436A, а второй полипептид Fc-области содержит
- мутации I253A, Н310А и Н435А, или
- мутации Н310А, Н433А и Y436A, или
- мутации L251D, L314D и L432D, или
- мутации L251S, L314S и L432S, или
g) первый полипептид Fc-области содержит мутации I253A, Н310А и Н435А, а второй полипептид Fc-области содержит
- мутации Н310А, Н433А и Y436A, или
- мутации L251D, L314D и L432D, или
- мутации L251S, L314S и L432S, или
h) первый полипептид Fc-области содержит мутации Н310А, Н433А и Y436A, а второй полипептид Fc-области содержит
a) мутации L251D, L314D и L432D, или
b) мутации L251S, L314S и L432S,
или
i) первый полипептид Fc-области содержит мутации L251D, L314D и L432D, а второй полипептид Fc-области содержит
а) мутации L251S,L314S и L432S.
и имеет одно или более чем одно из следующих свойств:
- демонстрирует более низкую сывороточную концентрацию по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (через 96 часов после интравитреального введения мышам, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn) (определяли с помощью анализов, описанных в примере 6),
- демонстрирует аналогичную (коэффициент от 0,8 до 1,2) концентрацию в цельных лизатах правого глаза по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (у мышей, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn, через 96 часов после интравитреального введения в правый глаз) (определяли с помощью анализов, описанных в примере 6),
- не демонстрирует связывания с человеческим неонатальным Fc-рецептором.
В одном воплощении биспецифическое антитело характеризуется тем, что содержит первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, и характеризуется тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №20 с 1, 2 или 3 аминокислотными заменами, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №21 с 1, 2 или 3 аминокислотными заменами, и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №28 с 1, 2 или 3 аминокислотными заменами, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №29 с 1, 2 или 3 аминокислотными заменами, и
iii) биспецифическое антитело содержит Fc-область, содержащую первый и второй полипептид Fc-области как человеческого IgG1, так и человеческого IgG4 (т.е. человеческого происхождения), которая включает одну или две мутации, выбранные из i) группы I253A, Н310А, Н435А, или ii) группы Н310А, Н433А, Y436A, или iii) группы L251D, L314D, L432D, или iv) группы L251S, L314S, L432S (в соответствии с индексом ЕС по системе нумерации Kabat) в первом полипептиде Fc-области, и одну или две мутаций, выбранные из группы, включающей мутации L251D, L251S, I253A, Н310А, L314D, L314S, L432D, L432S, Н433А, Н435А и Y436A (в соответствии с индексом ЕС по системе нумерации Kabat) во втором полипептиде Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А, Н435А, или ii) Н310А, Н433А, Y436A, или iii) L251D, L314D, L432D, или iv) L251S, L314S, L432S содержатся в Fc-области.
и имеет одно или более чем одно из следующих свойств:
- демонстрирует более низкую сывороточную концентрацию по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (через 96 часов после интравитреального введения мышам, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn) (определяли с помощью анализов, описанных в примере 6),
- демонстрирует аналогичную (коэффициент от 0,8 до 1,2) концентрацию в цельных лизатах правого глаза по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (у мышей, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn, через 96 часов после интравитреального введения в правый глаз) (определяли с помощью анализов, описанных в примере 6),
- не демонстрирует связывания с человеческим неонатальным Fc-рецептором.
В одном воплощении биспецифическое антитело характеризуется тем, что содержит первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2, и характеризуется тем, что
i) первый антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №20 с 1, 2 или 3 аминокислотными заменами, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №21 с 1, 2 или 3 аминокислотными заменами, и
ii) второй антигенсвязывающий сайт содержит в качестве вариабельного домена тяжелой цепи (VH) аминокислотную последовательность SEQ ID №28 с 1, 2 или 3 аминокислотными заменами, а в качестве вариабельного домена легкой цепи (VL) аминокислотную последовательность SEQ ID №29 с 1, 2 или 3 аминокислотными заменами, и
iii) биспецифическое антитело содержит первый полипептид Fc-области и второй полипептид Fc-области, где
a) первый и второй полипептид Fc-области содержит мутацию Y436A, или
b) первый и второй полипептид Fc-области содержит мутации I253A, H310A и Н435А, или
c) первый и второй полипептид Fc-области содержит мутации Н310А, Н433А и Y436A, или
d) первый и второй полипептид Fc-области содержит мутации L251D, L314D и L432D, или
e) первый и второй полипептид Fc-области содержит мутации L251S, L314S и L432S,или
f) первый полипептид Fc-области содержит мутацию Y436A, а второй полипептид Fc-области содержит
- мутации I253A, Н310А и Н435А, или
- мутации Н310А, Н433А и Y436A, или
- мутации L251D, L314D и L432D, или
- мутации L251S,L314S и L432S, или
g) первый полипептид Fc-области содержит мутации I253A, Н310А и Н435А, а второй полипептид Fc-области содержит
- мутации Н310А, Н433А и Y436A, или
- мутации L251D, L314D и L432D, или
- мутации L251S, L314S и L432S, или
h) первый полипептид Fc-области содержит мутации Н310А, Н433А и Y436A, а второй полипептид Fc-области содержит
a) мутации L251D, L314D и L432D, или
b) мутации L251S, L314S и L432S,
или
i) первый полипептид Fc-области содержит мутации L251D, L314D и L432D, а второй полипептид Fc-области содержит
а) мутации L251S, L314S и L432S. и имеет одно или более чем одно из следующих свойств:
- демонстрирует более низкую сывороточную концентрацию по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (через 96 часов после интравитреального введения мышам, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn) (определяли с помощью анализов, описанных в примере 6),
- демонстрирует аналогичную (коэффициент от 0,8 до 1,2) концентрацию в цельных лизатах правого глаза по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (у мышей, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn, через 96 часов после интравитреального введения в правый глаз) (определяли с помощью анализов, описанных в примере 6),
- не демонстрирует связывания с человеческим неонатальным Fc-рецептором.
Антигенсвязывающий сайт биспецифического антитела, описанного в данном документе, содержит шесть областей, определяющих комплементарность (CDR), которые способствуют в той или иной степени аффинности сайта связывания с его антигеном. Существует три CDR вариабельного домена тяжелой цепи (CDRH1, CDRH2 и CDRH3) и три CDR вариабельного домена легкой цепи (CDRL1, CDRL2 и CDRL3). Протяженность CDR и каркасных областей (FR) определяют путем сравнения с базой данных аминокислотных последовательностей, в которых эти области были определены в соответствии с вариабельностью среди последовательностей.
В одном воплощении всех аспектов антитело не связывается специфически с человеческим FcRn. В одном воплощении всех аспектов антитело, кроме того, специфически связывается со стафилококковым белком А.
В одном воплощении всех аспектов антитело не связывается специфически с человеческим FcRn. В одном воплощении всех аспектов антитело, кроме того, не связывается специфически со стафилококковым белком А.
В одном воплощении всех аспектов первый полипептид также содержит мутации Y349C, T366S, L368A и Y407V ("замок"), а второй полипептид содержит мутации S354C и T366W ("ключ").
В одном воплощении всех аспектов первый полипептид также содержит мутации S354C, T366S, L368A и Y407V ("замок"), а второй полипептид содержит мутации Y349C и T366W ("ключ").
В одном воплощении всех аспектов полипептиды Fc-области относятся к человеческому подклассу IgG1. В одном воплощении первый полипептид Fc-области и второй полипептид Fc-области также содержат мутации L234A и L235A. В одном воплощении первый полипептид Fc-области и второй полипептид Fc-области также содержат мутацию P329G.
В одном воплощении всех аспектов полипептиды Fc-области относятся к человеческому подклассу IgG4. В одном воплощении первый полипептид Fc-области и второй полипептид Fc-области также содержат мутации S228P и L235E. В одном воплощении первый полипептид Fc-области и второй полипептид Fc-области также содержат мутацию P329G.
Антитело, описанное в данном документе, получают с помощью рекомбинантных средств. Таким образом, один аспект, описанный в данном документе, представляет собой нуклеиновую кислоту, кодирующую антитело, описанное в данном документе, а другой аспект представляет собой клетку, содержащую указанную нуклеиновую кислоту, кодирующую антитело, описанное в данном документе. Способы рекомбинантной продукции широко известны в данной области и включают экспрессию белка в прокариотических и эукариотических клетках с последующим выделением антитела и, как правило, очисткой до фармацевтически приемлемой чистоты. Для экспрессии вышеупомянутых антител в клетке-хозяине нуклеиновые кислоты, кодирующие соответствующие (модифицированные) легкие и тяжелые цепи, вставляют в соответствующие экспрессионные векторы с помощью стандартных способов. Экспрессию осуществляют в подходящих прокариотических или эукариотических клетках-хозяевах, таких как клетки СНО, клетки NS0, клетки SP2/0, клетки HEK293, клетки COS, клетки PER.C6, дрожжевые клетки или клетки Е.coli, и выделяют антитело из клеток (из культурального супернатанта или из клеток после лизиса). Основные способы рекомбинантной продукции антител широко известны в данной области и описаны, например, в статьях Makrides, S C., Protein Expr. Purif. 17 (1999) 183-202; Geisse, S., et al., Protein Expr. Purif. 8 (1996) 271-282; Kaufman, R.J., Mol. Biotechnol. 16 (2000) 151-161; Werner, R.G., Drug Res. 48 (1998) 870-880.
Соответственно, один аспект, описанный в данном документе, относится к способу получения биспецифического антитела, описанного в данном документе, включающему этапы
a) трансформации клетки-хозяина векторами, содержащими нуклеиновокислотные молекулы, кодирующие антитело;
b) культивирования клетки-хозяина в условиях, обеспечивающих синтез антитела; и
c) выделения антитела из указанной культуры.
В одном воплощении этап выделения с) включает применение реагента, специфически захватывающего константный домен легкой цепи (который, например, специфичен для константного домена легкой цепи каппа или лямбда в зависимости от того, содержится ли в биспецифическом антителе легкая цепь каппа или лямбда). В одном воплощении этот реагент, специфически захватывающий легкую цепь, используется в режиме "связать и элюировать". Примерами таких реагентов, специфически захватывающих константный домен легкой цепи легкой цепи, являются, например, KappaSelect™ и LambdaFabSelect™ (доступны от GE Healthcare/BAC), которые основаны на матрице с высоко жесткой агарозной основой, что позволяет использовать высокие скорости потока и низкое давление при больших масштабах. Эти материалы содержат лиганд, который связывается с константной областью легкой цепи каппа или лямбда, соответственно (т.е. фрагменты, не имеющие константной области легкой цепи, не будут связываться; фиг. 1). Следовательно, они обе способны связываться с другими молекулами-мишенями, содержащими константную область легкой цепи, например, IgG, IgA и IgM. Лиганды прикреплены к матрице с помощью длинного гидрофильного спейсера, что делает их легко доступными для связывания с молекулой-мишенью. Они основаны на одноцепочечном фрагменте антитела, что позволяет проводить скрининг на человеческий Ig каппа или лямбда.
В одном воплощении этап выделения с) включает применение реагента, специфически захватывающего Fc-область. В одном воплощении реагент, специфически захватывающий Fc-область, используется в режиме "связать и элюировать". Примерами таких реагентов, специфически захватывающих Fc-область, являются, например, материалы для аффинной хроматографии на основе стафилококкового белка А.
Биспецифические антитела соответствующим образом выделяют из культуральной среды с помощью обычных процедур очистки иммуноглобулинов, таких как, например, аффинная хроматография (белок А-сефароза или KappaSelect™, LambdaFabSelect™), гидроксиапатитная хроматография, гель-электрофорез или диализ.
ДНК и РНК, кодирующие моноклональные антитела, можно легко выделить и секвенировать с помощью обычных процедур. В-клетки или гибридомные клетки могут служить источником таких ДНК и РНК. После выделения ДНК может быть вставлена в экспрессионные векторы, которыми затем трансфицируют клетки-хозяева, например, клетки НЕК 293, клетки СНО или клетки миеломы (которые в противном случае не продуцируют иммуноглобулиновый белок), чтобы получить синтез рекомбинантных моноклональных антител в клетках-хозяевах.
Некоторые из молекул, описанных в данном документе, обеспечивают простоту выделения/очистки благодаря содержанию Fc-областей, которые дифференциально модифицированы по меньшей мере одной из модификаций, которые приводят к i) дифференциальной аффинности молекулы к (стафилококковому) белку А и ii) дифференциальной аффинности молекулы к человеческому FcRn, и молекула выделяется из разрушенной клетки, из среды или из смеси молекул на основе ее аффинности к белку А.
Очистку антител проводят с целью устранения клеточных компонентов или других загрязняющих веществ, например, других клеточных нуклеиновых кислот или белков, с помощью стандартных методик, включая щелочную/SDS-обработку, мечение CsCl, колоночную хроматографию, электрофорез в агарозном геле и другие хорошо известные в данной области методики (см., например, Ausubel, F., et al., ed. Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)). Для очистки белков приняты и широко применяются различные способы, такие как, например, аффинная хроматография с микробными белками (например, аффинная хроматография с белком А или белком G), ионообменная хроматография (например, катионообменная (с карбоксиметиловыми смолами), анионообменная (с аминоэтиловыми смолами) и со смешанным режимом обмена), тиофильная адсорбционная хроматография (например, с бета-меркаптоэтанолом и другими SH-лигандами), хроматография гидрофобных взаимодействий или адсорбционная хроматография с ароматическими группами (например, с фенил-сефарозой, аза-аренофильными смолами или м-аминофенилбороновой кислотой), металл-хелатная аффинная хроматография (например, с носителем с аффинностью к Ni(II) и Cu(II)), эксклюзионная хроматография и электрофоретические способы (такие как гель-электрофорез, капиллярный электрофорез) (Vijayalakshmi, M.A., Appl. Biochem. Biotech. 75(1998) 93-102).
Двухвалентное биспецифическое антитело против человеческого VEGF и человеческого ANG-2, описанное в данном документе, может иметь ценный профиль эффективности/безопасности и может обеспечить преимущества для пациента, который нуждается в анти-VEGF- и анти-ANG-2-терапии.
Один из аспектов, описанных в данном документе, представляет собой фармацевтический препарат, содержащий антитело, описанное в данном документе. Другим аспектом изобретения, описанным в данном документе, является применение антитела, описанного в данном документе, для изготовления фармацевтического препарата. Еще один аспект, описанный в данном документе, представляет собой способ изготовления фармацевтического препарата, содержащего антитело, описанное в данном документе. В другом аспекте предусмотрен препарат, например, фармацевтический препарат, содержащий антитело, описанное в данном документе, собранное в препарат вместе с фармацевтическим носителем.
Препарат согласно данному изобретению может быть введен различными способами, известными в данной области. Как будет понятно специалисту, путь и/или режим введения будет варьировать в зависимости от нужного результата. Для введения определенным путем соединения, описанного в данном документе, может потребоваться покрытие соединения или одновременное введение соединения с материалом, предотвращающим его инактивацию. Например, соединение может быть введено субъекту в соответствующем носителе, например, в липосомах, или в разбавителе. Фармацевтически приемлемые разбавители включают физиологический раствор и водные буферные растворы. Фармацевтические носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления стерильных инъекционных растворов или дисперсий непосредственно перед применением. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области.
Могут быть использованы многие возможные режимы доставки, включая, но не ограничиваясь ими, внутриглазное введение или местное применение. В одном воплощении введение является внутриглазным и включает, но не ограничиваясь ими, субконъюнктивальную инъекцию, внутрикамерную инъекцию, инъекцию в переднюю камеру через темпоральную сторону лимба, внутристромальную инъекцию, внутрироговичную инъекцию, субретинальную инъекцию, инъекцию в водянисту влагу, субтеноновую инъекцию или устройство для замедленной доставки, интравитреальную инъекцию (например, переднюю, среднюю или заднюю витреальную инъекцию). В одном воплощении введение является местным и включает, но не ограничиваясь этим, глазные капли на роговицу.
В одном воплощении биспецифическое антитело или фармацевтический препарат, описанный в данном документе, вводят интравитреальным путем, например, с помощью интравитреальной инъекции. Она может быть выполнена в соответствии со стандартными процедурами, известными в данной области (см., например, Ritter et al., J. Clin. Invest. 116 (2006) 3266-3276, Russelakis-Carneiro et al., Neuropathol. Appl. Neurobiol. 25 (1999) 196-206, и Wray et al., Arch. Neurol. 33 (1976) 183-185).
В некоторых воплощении терапевтические наборы, описанные в данном документе, могут содержать одну или более чем одну дозу (биспецифического) антитела, присутствующего в фармацевтическом препарате, описанном в данном документе, подходящее устройство для интравитреальной инъекции фармацевтического препарата и инструкцию с подробным описанием подходящих субъектов и протоколами проведения инъекции. В этих воплощениях препараты, как правило, вводят субъекту, нуждающемуся в лечении, с помощью интравитреальной инъекции. Она может быть выполнена в соответствии со стандартными процедурами, известными в данной области (см., например, Ritter et al., J. Clin. Invest. 116 (2006) 3266-3276, Russelakis-Carneiro et al., Neuropathol. Appl. Neurobiol. 25 (1999) 196-206, и Wray et al., Arch. Neurol. 33 (1976) 183-185).
Эти препараты также могут содержать вспомогательные вещества, такие как консерванты, смачивающие агенты, эмульгаторы и диспергирующие агенты. Предотвращение наличия микроорганизмов может быть обеспечено как за счет процедур стерилизации, как сказано выше, так и за счет включения различных антибактериальных и противогрибковых агентов, например, парабена, хлорбутанола, фенола, сорбиновой кислоты и т.п. Также может быть целесообразным включение в препараты изотонических агентов, таких как сахара, хлорид натрия и т.п. Кроме того, пролонгированную абсорбцию инъекционной фармацевтической формы можно получить путем включения агентов, которые задерживают абсорбцию, таких как моностеарат алюминия и желатин.
Независимо от выбранного пути введения соединения, описанные в данном документе, которые могут быть использованы в подходящей гидратированной форме, и/или фармацевтические препараты, описанные в данном документе, собирают в фармацевтически приемлемые лекарственные формы с помощью традиционных способов, известных специалистам в данной области.
Фактические уровни доз активных ингредиентов в фармацевтических препаратах, описанных в данном документе, можно изменить таким образом, чтобы получить количество активного ингредиента, эффективное для достижения желаемого терапевтического эффекта для конкретного пациента, композиции и способа введения, но не токсичное для пациента. Выбранный уровень доз будет зависеть от ряда фармакокинетических факторов, включая активность конкретных используемых препаратов, путь введения, время введения, скорость выведения конкретного используемого соединения, от продолжительности лечения, других лекарственных препаратов, соединений и/или материалов, используемых в сочетании с конкретными используемыми препаратами, от возраста, пола, веса, условий, общего состояния здоровья и истории болезни пациента, подлежащего лечению, и т.п. факторов, хорошо известных в медицине.
Препарат должен быть стерильным и жидким, если он доставляется с помощью шприца. Кроме воды, предпочтительным носителем является изотонический буферный солевой раствор.
Надлежащая текучесть может быть обеспечена, например, путем использования покрытий, таких как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Во многих случаях в препарат предпочтительно включать изотонические агенты, например, сахара, многоатомные спирты, такие как маннит или сорбит, и хлорид натрия.
Препарат может содержать офтальмологический препарат-депо, содержащий активный агент для введения субконъюнктивального введения. Офтальмологический препарат-депо включает микрочастицы практически чистого активного агента, например, биспецифического антитела, описанного в данном документе. Микрочастицы, содержащие биспецифическое антитело, описанное в данном документе, могут быть встроены в биосовместимый фармацевтически приемлемый полимер или липидный инкапсулирующий агент. Препараты-депо могут быть приспособлены к высвобождению всего или практически всего активного материала в течение длительного периода времени. Полимер или липидная матрица, если они присутствуют, могут быть адаптированы для деградации, достаточной для того, чтобы быть перемещенными из места введения после высвобождения всего или практически всего активного агента. Препарат-депо может быть жидким препаратом, включающим фармацевтически приемлемый полимер и растворенный или диспергированный активный агент. После инъекции полимер образует депо в месте инъекции, например, путем гелеобразования или осаждения.
Другой аспект, описанный в данном документе, представляет собой биспецифическое антитело, описанное в данном документе, для применения в лечении глазных сосудистых заболеваний.
Другой аспект, описанный в данном документе, представляет собой фармацевтический препарат, описанный в данном документе, для применения в лечении глазных сосудистых заболеваний.
Другой аспект, описанный в данном документе, представляет собой применение антитела, описанного в данном документе, для изготовления лекарственного средства для лечения глазных сосудистых заболеваний.
Другой аспект, описанный в данном документе, представляет собой способ лечения пациента, страдающего от глазных сосудистых заболеваний, путем введения пациенту антитела, описанного в данном документе, в случае необходимости такого лечения.
Здесь явно указано, что термин "содержащий", используемый в данном документе, включает термин "состоящий из". Таким образом, все аспекты и воплощения, которые включают термин "содержащий", аналогично раскрываются термином "состоящий из".
Модификации
В другом аспекте антитело согласно любому из вышеописанных воплощений может включать любой из признаков, по отдельности или в комбинации, как описано в разделах 1-6 ниже:
1. Аффинность антитела
В некоторых воплощениях антитело, предложенное в данном документе, имеет константу диссоциации (Kd) ≤100 нМ, ≤10 нМ (например, 10-7 М или менее, например, от 10-7 М до 10-13 М, например, от 10-8 М до 10-13 M).
В одном воплощении Kd измеряли в анализе поверхностного плазмонного резонанса BIACORE®. Например, анализ проводили с использованием BIACORE®-2000 или BIACORE®-3000 (GE Healthcare Inc., Пискатавэй, Нью-Джерси) при 25°C с СМ5-чипами с иммобилизованным антигеном при примерно 10 единицах ответа (RU). В одном воплощении биосенсорный чип с карбоксиметилированным декстраном (CM5, GE Healthcare Inc.) активировали N-этил-N'-(3-диметиламинопропил)карбодиимида гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводили в 10 мМ растворе ацетата натрия, рН 4,8, до концентрации 5 мкг/мл (примерно 0,2 мкМ) и вводили со скоростью потока 5 мкл/мин до достижения примерно 10 единиц ответа (RU) связанного белка. После введения антигена вводили 1 M этаноламина для блокировки непрореагировавших групп. Для измерения кинетики двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) вводили в PBS с 0,05% поверхностно-активного вещества полисорбата 20 (TWEEN-20™) (PBST) при 25°C со скоростью потока примерно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывали с помощью простой модели связывания Ленгмюра 1:1 (программное обеспечение BIACORE® Evaluation Software версии 3.2), одновременно соединяя сенсограммы ассоциации и диссоциации. Константу равновесия диссоциации (Kd) рассчитывали как отношение Wkon (см., например, Chen, Y. et al., J. Mol. Biol. 293 (1999) 865-881). Если в анализе поверхностного плазмонного резонанса, описанном выше, оп-скорость превышает 106 М-1 с-1 то она может быть определена с помощью методики тушения флуоресценции, при которой измеряется увеличение или уменьшение интенсивности флуоресцентного излучения (возбуждение=295 нм; излучение = 340 нм; 16 нм полосовой фильтр) при 25°C с 20 нМ антитела против антигена (Fab-форма) в PBS, рН 7,2, в присутствии возрастающих концентраций антигена, измеряемых на спектрометре, таком как спектрофотометр, оборудованный системой быстрого смешивания с последующей остановкой (Aviv Instruments) или спектрофотометр SLM-AMINCO™ серии 8000 (ThermoSpectronic) со встряхивающейся кюветой.
2. Химерные и гуманизированные антитела
В некоторых воплощениях антитело, предложенное в данном документе, является химерным антителом. Некоторые химерные антитела описаны, например, в US 4816567 и в Morrison, S.L. et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855). В одном примере химерное антитело содержит нечеловеческую вариабельную область (например, вариабельную область, полученную от мыши, крысы, хомяка, кролика или примата, например, обезьяны) и человеческую константную область. В другом примере химерное антитело представляет собой антитело "с переключением класса", в котором класс или подкласс был изменен по сравнению с родительским антителом. Химерные антитела включают их антигенсвязывающие фрагменты.
В некоторых воплощениях химерное антитело представляет собой гуманизированное антитело. Как правило, нечеловеческое антитело гуманизируют, чтобы уменьшить иммуногенность для людей, сохраняя при этом специфичность и аффинность родительского нечеловеческого антитела. Как правило, гуманизированное антитело содержит один или более чем один вариабельный домен, в котором HVR, например, CDR (или их части) получены из последовательностей нечеловеческого антитела, a FR (или их части) получены из последовательностей человеческого антитела. Гуманизированное антитело, возможно, также будет содержать по меньшей мере часть человеческой константной области. В некоторых воплощениях некоторые FR-остатки в гуманизированном антителе замещены соответствующими остатками из нечеловеческого антитела (например, антитела, из которого получены HVR-остатки), например, для восстановления или повышения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения приведены, например, в Almagro, J.С.and Fransson, J., Front. Biosci. 13 (2008) 1619-1633, а также описаны, например, в Riechmann, I., et al., Nature 332 (1988) 323-329; Queen, C, et al., Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033; US 5,821,337, US 7,527,791, US 6,982,321 и US 7,087,409; Kashmiri, S.V., et al., Methods 36 (2005) 25-34 (описывает прививание SDR (a-CDR)); Padlan, E.A., Mol. Immunol. 28 (1991) 489-498 (описывает "перекладку"); Dall'Acqua, W.F. et al., Methods 36 (2005) 43-60 (описывает "перетасовку FR"); Osbourn, J. et al., Methods 36 (2005) 61-68; и Klimka, A. et al., Br. J. Cancer 83 (2000) 252-260 (описывают подход "направляемой селекции" к перетасовке FR).
Человеческие каркасные области, которые могут быть использованы для гуманизации, включают, но не ограничиваясь ими: каркасные области, выбранные с помощью методики "наилучшего соответствия" (см., например, Sims, M.J., et al., J. Immunol. 151 (1993) 2296-2308); каркасные области, полученные из консенсусной последовательности человеческих антител конкретной подгруппы вариабельных областей легкой или тяжелой цепей (см., например, Carter, P., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289; и Presta, L.G., et al., J. Immunol. 151 (1993) 2623-2632); человеческие зрелые (соматически мутантные) каркасные области или человеческие зародышевые каркасные области (см., например, Almagro, J.С.and Fransson, J., Front. Biosci. 13 (2008) 1619-1633); и каркасные области, полученные при скрининге FR-библиотек (см., например, Baca, M. et al., J. Biol. Chem. 272 (1997) 10678-10684, и Rosok, M.J. et al., J. Biol. Chem. 271 (19969 22611-22618).
3. Человеческие антитела
В некоторых воплощениях антитело, предложенное в данном документе, является человеческим антителом. Человеческие антитела могут быть получены с помощью различных методик, известных в данной области. Человеческие антитела в целом описаны в van Dijk, М.А. and van de Winkel, J.G., Curr. Opin. Pharmacol. 5 (2001) 368-374, и в Lonberg, N., Curr. Opin. Immunol. 20 (2008) 450-459.
Человеческие антитела могут быть получены путем введения иммуногена трансгенному животному, которое было модифицировано таким образом, чтобы производить интактные человеческие антитела или интактные антитела с человеческими вариабельными областями в ответ на антигенную стимуляцию. Такие животные, как правило, содержат все или часть локусов человеческого иммуноглобулина, которые заменяют локусы эндогенного иммуноглобулина или которые присутствуют вне хромосом или интегрированы случайным образом в хромосомы животного. У таких трансгенных мышей эндогенные локусы иммуноглобулина, как правило, инактивированы. Обзор способов получения человеческих антител с помощью трансгенных животных см. в Lonberg, N., Nat. Biotech. 23 (2005) 1117-1125. См. также, например, US 6075181 и US 6150584, описывающие методику XENOMOUSE™; патент США №5770429, описывающий методику HUMAB®; US 7041870, описывающий методику K-M MOUSE®, и US 2007/0061900, описывающий методику VELOCiMOUSE®). Человеческие вариабельные области из интактных антител, полученные с помощью таких животных, могут быть дополнительно модифицированы, например, путем объединения с другой человеческой константной областью.
Человеческие антитела также могут быть получены с помощью методик на основе гибридом. Были описаны линии клеток человеческой миеломы и мышиной/человеческой гетеромиеломы для получения человеческих моноклональных антител (см., например, Kozbor, D., J. Immunol. 133 (1984) 3001-3005; Brodeur, B.R., et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York (1987), pp.51-63; b Boerner, P., et al., J. Immunol. 147 (1991) 86-95). Человеческие антитела, полученные с помощью человеческой В-клеточной гибридомной методики, также описаны в Li, J. et al., Proc. Natl. Acad. Sci. USA 103 (2006) 3557-3562. Дополнительные методики включают те, которые описаны, например, в US 7189826 (описывается получение моноклональных человеческих антител IgM с помощью гибридомных клеточных линий) и Ni, J., Xiandai Mianyixue 26 (2006) 265-268 (описываются человек/человеческие гибридомы). Методика человеческих гибридом (методика Trioma) также описана в Vollmers, Н.Р. and Brandlein, S., Histology and Histopathology 20 (2005) 927-937 и Vollmers, H.P. and Brandlein, S., Methods and Findings in Experimental and Clinical Pharmacology 27 (2005) 185-191.
Человеческие антитела также могут быть получены путем выделения Fv-клональных последовательностей вариабельных доменов, выбранных из библиотек фагового дисплея человеческою происхождения. Такие последовательности вариабельного домена затем могут быть объединены с нужным человеческим константным доменом. Методики отбора человеческих антител из библиотек антител описаны ниже.
4. Антитела, полученные из библиотек
Антитела, описанные в данном документе, могут быть выделены путем скрининга комбинаторных библиотек на антитела с нужной активностью или активностями. Например, в данной области известны различные способы создания библиотек фагового дисплея и скрининга таких библиотек на антитела, обладающие нужными характеристиками связывания. Такие способы приведены, например, в Hoogenboom, H.R. et al., Methods in Molecular Biology 178 (2001) 1-37, a также описаны, например, в the McCafferty, J. et al., Nature348 (1990) 552-554; Clackson, T. et al., Nature 352 (1991) 624-628; Marks, J.D. et al., J. Mol. Biol. 222 (1992) 581-597; Marks, J.D. and Bradbury, A., Methods in Molecular Biology 248 (2003) 161-175; Sidhu, S.S. et al., J. Mol. Biol. 338 (2004) 299-310; Lee, C.V. et al., J. Mol. Biol. 340 (2004) 1073-1093; Fellouse, F.A., Proc. Natl. Acad. Sci. USA 101 (2004) 12467-12472; и Lee, C.V. et al., J. Immunol. Methods 284 (2004) 119-132.
В некоторых способах фагового дисплея репертуар VH- и VL-генов отдельно клонировали с помощью полимеразной цепной реакции (ПЦР) и рекомбинировали в случайном порядке в фаговые библиотеки, которые затем могли быть подвергнуты скринингу на антигенсвязывающий фаг, как описано в Winter, G., et al., Ann. Rev. Immunol. 12 (1994) 433-455. Фаг, как правило, выставляет фрагменты антитела либо как одноцепочечные Fv-фрагменты (scFv), либо как Fab-фрагменты. Библиотеки из иммунизированных источников предлагают высокоаффинные антитела к иммуногену без необходимости конструирования гибридом. Альтернативно, может быть клонирован наивный репертуар (например, от человека), чтобы обеспечить единый источник антител к широкому спектру чужих, а также собственных антигенов без какой-либо иммунизации, как описано в Griffiths, A.D., et al., EMBO J. 12 (1993) 725-734. Наконец, наивные библиотеки также могут быть получены синтетически путем клонирования неперестроенных сегментов V-гена из стволовых клеток и с использованием ПЦР-праймеров, содержащих случайную последовательность для кодирования высоко вариабельных CDR3-областей и для выполнения перестройки /л v/fro, как описано в Hoogenboom, H.R. and Winter, G., J. Mol. Biol. 227 (1992) 381-388. Патентные публикации, описывающие фаговые библиотеки человеческого антитела, включают, например US 5750373 и US 2005/0079574, US 2005/0119455, US 2005/0266000, US 2007/0117126, US 2007/0160598, US 2007/0237764, US 2007/0292936 и US 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, считаются в данном документе человеческими антителами или фрагментами человеческих антител.
5. Мультиспецифические антитела
В некоторых воплощениях антитело, предложенное в данном документе, является мультиспецифическим антителом, например, биспецифическим антителом. Мультиспецифические антитела представляют собой моноклональные антитела, которые обладают специфичностью связывания по меньшей мере с двумя различными сайтами. Биспецифические антитела также могут быть использованы для локализации цитотоксических агентов на клетках, которые экспрессируют один или более чем один антиген-мишень. Биспецифические антитела могут быть получены в виде полноразмерных антител или фрагментов антител.
Методики получения мультиспецифических антител включают, но не ограничиваясь ими, рекомбинантную коэкспрессию двух пар тяжелых цепей и легких цепей иммуноглобулина, имеющих различные специфичности (см. Milstein, С. and Cuello, А.С., Nature 305 (1983) 537-540, WO 93/08829, и Traunecker, A. et al., EMBO J. 10 (1991) 3655-3659), и инженерию типа "ключ в замке" (см., например, US 5731168). Мультиспецифические антитела также могут быть получены за счет использования инженерных электростатических перемешивающих влияний для создания Fc-гетеродимерных молекул антитела (WO 2009/089004); поперечного сшивания двух или более антител или фрагментов (см., например, US 4676980 и Brennan, М. et al., Science 229 (1985) 81-83); использования лейциновых застежек для получения биспецифических антител (см., например, Kostelny, S.A. et al., J. Immunol. 148 (1992) 1547-1553); использования методики "димерного антитела" для получения биспецифических фрагментов антител (см., например, Holliger, P. et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448); и использования одноцепочечных Fv-димеров (sFv) (см., например, Gruber, М et al., J. Immunol. 152 (1994) 5368-5374); и получения триспецифических антител, как описано, например, в Tutt, A. et al., J. Immunol. 147(1991)60-69).
Инженерные антитела с тремя или более функциональными антигенсвязывающими сайтами, в том числе "антитела-осьминоги", также включены в данный документ (см., например, US 2006/0025576).
Антитело или фрагмент в данном документе также включает "Fab двойного действия" или "DAF", включающий антигенсвязывающий сайт, который связывается с первым антигеном, а также с другим, отличающимся, антигеном (см., например, US 2008/0069820).
Антитело или фрагмент в данном документе также включают мультиспецифические антитела, описанные в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, WO 2010/112193, WO 2010/115589, WO 2010/136172, WO 2010/145792 и WO 2010/145793.
6. Варианты антител
В некоторых воплощениях рассматриваются варианты аминокислотных последовательностей антител, предложенных в данном документе. Например, может быть желательным улучшение аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотной последовательности антитела могут быть получены путем введения соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем пептидного синтеза. Такие модификации включают, например, делеции и/или вставки и/или замены остатков в аминокислотных последовательностях антитела. Можно использовать любую комбинацию делеций, инсерций и замен, чтобы получить конечную конструкцию, при условии, что конечная конструкция обладает желаемыми характеристиками, например, связыванием антигена.
а) Варианты с заменой, вставкой и делецией
В некоторых воплощениях предложены варианты антител, имеющие одну или более чем одну аминокислотную замену. Сайты для мутагенеза путем замен, представляющие интерес, включают HVR и FR. Консервативные замены приведены в таблице 1 под заголовком "предпочтительные замены". Более существенные изменения приведены в таблице 1 под заголовком "иллюстративные замены" и дополнительно описаны ниже со ссылкой на классы аминокислот по боковой цепи. Аминокислотные замены могут быть введены в антитело, представляющее интерес, и продукты могут быть проверены на нужную активность, например, сохранившееся/улучшенное связывание антигена, снижение иммуногенности или улучшенная ADCC или CDC.
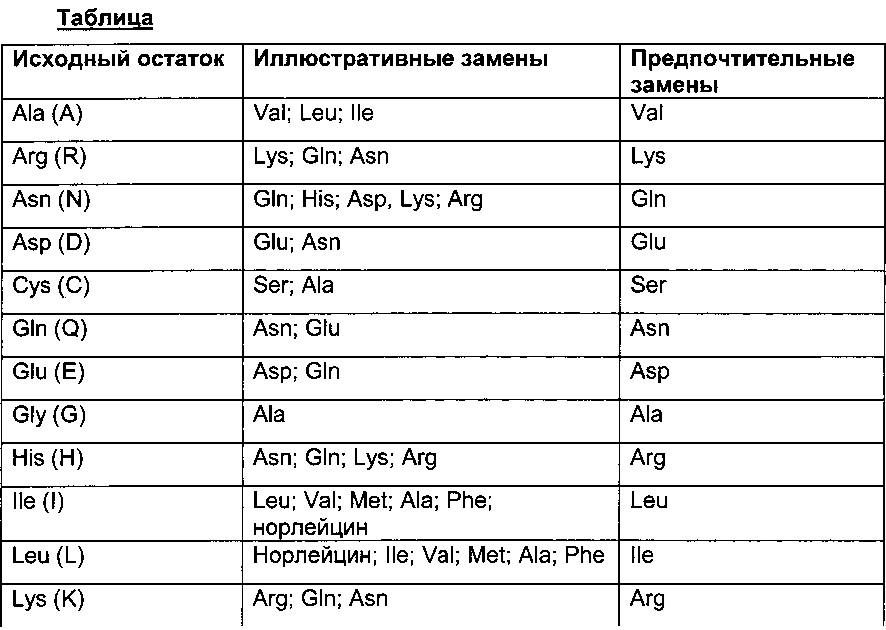
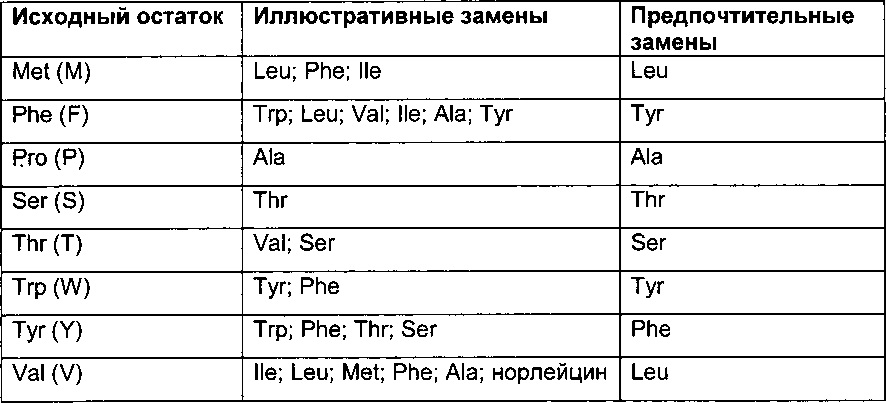
Аминокислоты могут быть сгруппированы в соответствии с общими свойствами боковой цепи:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Не;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gin;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены влекут за собой замену члена одного из этих классов на член другого класса.
Один тип варианта с заменой предполагает замену одного или более чем одного остатка гипервариабельной области родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный вариант(ы), выбранный для дальнейшего изучения, будет иметь модификации (например, улучшения) определенных биологических свойств (например, повышенную аффинность, сниженную иммуногенность) относительно родительского антитела и/или будет иметь практически сохранившиеся определенные биологические свойства родительского антитела. Иллюстративным вариантом с заменой является антитело со зрелой аффинностью, которое можно легко получить, например, с помощью методик созревания аффинности на основе фагового дисплея, таких как описанные в данном документе. Вкратце, один или более чем один остаток HVR подвергается мутированию, и вариантные антитела выставляются на фаге и проверяются на конкретную биологическую активность (например, аффинность связывания).
Изменения (например, замены) могут быть сделаны в HVR, например, для улучшения аффинности антитела. Такие изменения могут быть сделаны в "горячих точках" HVR, т.е. остатках, кодируемых кодонами, которые подвергаются мутации с высокой частотой в ходе процесса соматического созревания (см., например, Chowdhury, P.S., Methods Mol. Biol. 207 (2008) 179-196), и/или в остатках, которые контактируют с антигеном, при этом образующиеся вариантные VH или VL исследуются на аффинность связывания. Созревание аффинности путем конструирования и повторного выбора из вторичных библиотек было описано, например, в Hoogenboom, H.R. et al. in Methods in Molecular Biology 178 (2002) 1-37. В некоторых воплощениях созревания аффинности разнообразие вводится в вариабельные гены, выбранные для созревания, с помощью любой из разнообразных методик (например, ПЦР с внесением ошибок, перестановка цепей или олигонуклеотид-направленный мутагенез). Затем создается вторичная библиотека. Затем эту библиотеку скринируют для выявления любых вариантов антител с нужной аффинностью. Другой способ внесения разнообразия включает HVR-направленные подходы, в которых рандомизируют несколько остатков HVR (например, 4-6 остатков за один раз). HVR-остатки, участвующие в связывании антигена, могут быть конкретно выявлены, например, с помощью сканирующего аланином мутагенеза или моделирования. CDR-H3 и CDR-L3, в частности, часто становятся мишенями.
В некоторых воплощениях в одном или более чем одном HVR могут возникать замены, вставки или делеции до тех пор, пока такие изменения не вызывают существенного снижения способности антитела связывать антиген. Например, в HVR могут быть сделаны консервативные изменения (например, консервативные замены, предложенные в данном документе), которые существенно не снижают аффинность связывания. Такие изменения могут находиться вне контактирующих с антигеном остатков в HVR. В некоторых воплощениях вариантных VH- и VL-последовательностей, предложенных выше, каждый HVR либо не меняется, либо содержит не более чем одну, две или три аминокислотные замены.
Способ, который может быть использован для выявления остатков или областей антитела, которые могут стать мишенями мутагенеза, называется "сканирующим аланином мутагенезом", который описан в Cunningham, B.C. and Wells, J.A., Science 244 (1989) 1081-1085. В этом способе остаток или группу остатков-мишеней (например, заряженные остатки, такие как Arg, Asp, His, Lys и Glu) выявляют и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином), чтобы определить, влияют ли они на взаимодействие антитела с антигеном. Другие замены могут быть введены в аминокислотных позициях, демонстрирующих функциональную чувствительность к первоначальным заменам. Альтернативно или дополнительно может быть использована кристаллическая структура комплекса "антиген-антитело" для выявления точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки могут стать мишенями или могут быть устранены в качестве кандидатов на замену. Варианты можно подвергнуть скринингу, чтобы определить, содержат ли они нужные свойства.
Вставки в аминокислотную последовательность включают амино- и/или карбоксиконцевые слияния длиной от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или множества аминокислотных остатков внутрь последовательности. Примеры концевых вставок включают антитела с N-концевым остатком метионина. Другие варианты молекул антитела со вставками включают слияние N- или С-конца антитела с ферментом (например, ADEPT) или полипептидом, который увеличивает время полужизни антитела в сыворотке.
b) Варианты гликозилирования
В некоторых воплощениях антитело, предложенное в данном документе, изменено для увеличения или уменьшения степени, в которой антитело гликозилировано. Добавление или удаление к антителу сайтов гликозилирования может быть удобно проведено путем изменения аминокислотной последовательности, так что создается или удаляется один или более чем один сайт гликозилирования.
Если антитело содержит Fc-область, то углевод, прикрепленный к ней, может быть изменен. Нативные антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленный двухантенный олигосахарид, который, как правило, прикреплен с помощью N-связи к Asn297 СН2-домена Fc-области. См., например, Wright, A. and Morrison, S.L., TIBTECH 15 (1997) 26-32. Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, прикрепленную к GlcNAc в "стволе" двухантенной олигосахаридной структуры. В некоторых воплощениях модификации олигосахаридов в антителе изобретения могут быть сделаны для того, чтобы получить варианты антитела с определенными улучшенными свойствами.
В одном воплощении предложены варианты антитела, имеющее углеводную структуру, в которой отсутствует фукоза, прикрепленная (прямо или косвенно) к Fc-области. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяется путем вычисления среднего количества фукозы в сахарной цепи на Asn297 по отношению к сумме всех гликоструктур, прикрепленных к Asn 297 (например, сложных, гибридных и высокоманнозных структур), измеренного путем MALDI-TOF-масс-спектрометрии, как описано, например, в WO 2008/077546. Asn297 относится к остатку аспарагина, расположенному примерно в позиции 297 в Fc-области (нумерация остатков Fc-области согласно ЕС); тем не менее, Asn297 также может быть расположен в позиции примерно ±3 аминокислоты до или после позиции 297, т.е. между позициями 294 и 300, в связи с минорными изменениями последовательности в антителах. Такие варианты фукозилирования могут иметь улучшенные ADCC-функции. См., например, US 2003/0157108; US 2004/0093621. Примеры публикаций, связанных с "дефукозилированными" или "фукозодефицитными" вариантами антител, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140, US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2005/031140; Okazaki, A. et al., J. Mol. Biol. 336 (2004) 1239-1249; Yamane-Ohnuki, N. et al., Biotech. Bioeng. 87 (2004) 614-622. Примеры клеточных линий, способных продуцировать дефукозилированные антитела, включают СНО-клетки Lec13, дефицитные по фукозилированию белка (Ripka, J. et al., Arch. Biochem. Biophys. 249 (1986) 533-545; US 2003/0157108; и WO 2004/056312, особенно пример 11), и нокаутные клеточные линии, такие как СНО-клетки с нокаутом гена альфа-1,6-фукозилтрансферазы, FL/T8 (см., например, Yamane-Ohnuki, N. et al., Biotech. Bioeng. 87 (2004) 614-622; Kanda, Y. et al., Biotechnol. Bioeng. 94 (2006) 680-688; и WO 2003/085107).
Также предложены варианты антител с олигосахаридами с симметричным разветвлением, например, в которых двухантенный олигосахарид, присоединенный к Fc-области антитела, разделен пополам посредством GlcNAc. Такие варианты антител могут обладать уменьшенным фукозилированием и/или улучшенной ADCC-функцией. Примеры таких вариантов антител описаны, например, в WO 2003/011878; US 6602684 и US 2005/0123546. Также предусмотрены варианты антител по меньшей мере с одним остатком галактозы в олигосахариде, прикрепленном к Fc-области. Такие варианты антител могут обладать улучшенной CDC-функцией. Такие варианты антител описаны, например, в WO 1997/30087, WO 1998/58964 и WO 1999/22764.
с) варианты Fc-области
В некоторых воплощениях в Fc-область антитела, предложенного в данном документе, может быть введена одна или более чем одна дополнительная аминокислотная модификация, образующая тем самым вариант Fc-области. Вариант Fc-области может содержать последовательность человеческой Fc-области (например, Fc-области человеческого IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену/мутацию) в одной или более чем одной аминокислотной позиции.
В некоторых воплощениях изобретение предусматривает вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, которые делают его желательным кандидатом для применений, в которых время полужизни антитела in vivo является важным, а еще некоторые эффекторные функции (такие как CDC и ADCC) являются не необходимыми или вредными. Можно провести анализ цитотоксичности in vitro и/или in vivo, чтобы подтвердить уменьшение/истощение CDC- и/или ADCC-активности. Например, можно провести анализы Fc-рецепторного (FcR) связывания для того, чтобы убедиться, что антитело не связывается с FcγR (и, следовательно, скорее всего не обладает ADCC-активностью), но сохраняет FcRn-связывающую способность. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гемопоэтических клетках суммирована в таблице 3 на странице 464 в Ravetch, J.V. and Kinet, J.P., Annu. Rev. Immunol. 9 (1991) 457-492. Неограничивающие примеры анализов in vitro для оценки ADCC-активности молекулы, представляющей интерес, описываются в US 5500362 (см., например, Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 83 (1986) 7059-7063; и Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 82 (1985) 1499-1502); US 5821337 (см. Bruggemann, M. et al., J. Exp.Med. 166 (1987) 1351-1361). Альтернативно, могут быть использованы нерадиоактивные способы анализа (см., например, нерадиоактивный анализ цитотоксичности ACTI™ для проточной цитометрии (CellTechnology Inc., Маунтин-Вью, Калифорния; и нерадиоактивный анализ цитотоксичности CytoTox96® (Promega, Мэдисон, Висконсин). Используемые эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (РВМС) и натуральные киллеры (NK-клетки). Альтернативно или дополнительно ADCC-активность молекулы, представляющей интерес, может быть оценена in vivo, например, на животной модели, такой как описана в Clynes, R. et al., Proc. Natl. Acad. Sci. USA 95 (1998) 652-656. Также можно выполнить анализы Clq-связывания, чтобы подтвердить, что антитело не может связываться с C1q и, следовательно, не обладает CDC-активностью. См., например, C1q- и С3с-связывающий анализ ELISA в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента может быть выполнен анализ CDC (см., например, Gazzano-Santoro, Н. et al., J. Immunol. Methods 202 (1996) 163-171; Cragg, M.S. et al., Blood 101 (2003) 1045-1052; и Cragg, M.S. and M.J. Glennie, Blood 103 (2004) 2738-2743). Определение FcRn-связывания и периода выведения/полужизни in vivo также можно проводить с использованием способов, известных в данной области (см., например, Petkova, S.B. et al., Int. Immunol. 18 (2006) 1759-1769).
Антитела со сниженной эффекторной функцией включают таковые с заменой одного или более чем одного из остатков Fc-области 238, 265, 269, 270, 297, 327 и 329 (US 6737056). Такие варианты Fc-области включают Fc-области с заменами в двух или более чем двух из аминокислотных позиций 265, 269, 270, 297 и 327, включая так называемый "DANA"-мутант Fc-области с заменами остатков 265 и 297 на аланин (US 7332581).
Описаны некоторые варианты антител с улучшенным или уменьшенным связыванием с FcR (см., например, US 6737056; WO 2004/ 056312, и Shields, R.L. et al., J. Biol. Chem. 276 (2001) 6591-6604).
В некоторых воплощениях вариант антитела содержит Fc-область с одной или более чем одной аминокислотной заменой, которая улучшает ADCC, например, с заменами в позициях 298, 333 и/или 334 в Fc-области (нумерация остатков согласно ЕС).
В некоторых воплощениях в Fc-области сделаны изменения, которые приводят к измененному (т.е. повышенному или пониженному) C1q-связыванию и/или комплемент-зависимой цитотоксичности (CDC), например, как описано в US 6194551, WO 99/51642 и Idusogie, Е Е. et al., J. Immunol. 164 (2000) 4178-4184.
Антитела с удлиненным периодом полужизни и повышенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG к плоду (Guyer, R.L. et al., J. Immunol. 117 (1976) 587-593, и Kim, J.K. et al., J. Immunol. 24 (1994) 2429-2434), описаны в US 2005/0014934. Эти антитела содержат Fc-область с одной или более чем одной заменой в ней, что улучшает связывание Fc-области с FcRn. Такие варианты Fc-области включают варианты с заменами в одном или более чем одном из остатков Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, с заменой в Fc-области остатка 434 (US 7371826).
См. также Duncan, A.R. and Winter, G., Nature 322 (1988) 738-740; US 5648260; US 5624821 и WO 94/29351, касающиеся других примеров вариантов Fc-области.
d) Цистеин-инженерные варианты антител
В некоторых воплощениях может быть желательным создание цистеин-инженерных антител, например, "тиоМКА" ("thioMAb"), в которых один или более чем один остаток антитела замещен на остаток цистеина. В конкретных воплощениях замещенные остатки находятся в доступных сайтах антитела. При замещении этих остатков цистеином реактивные тиольные группы, таким образом, располагаются в доступных участках антитела и могут быть использованы для конъюгирования антитела с другими группировками, такими как лекарственные группировки или линкер-лекарственные группировки, для образования иммуноконъюгата, описанного далее. В некоторых воплощениях один или более чем один из следующих остатков может быть замещен цистеином: V205 (нумерация Кабата) легкой цепи; А118 (нумерация ЕС) тяжелой цепи; и S400 (нумерация ЕС) Fc-области тяжелой цепи. Цистеин-инженерные антитела могут быть получены, как описано, например, в US 7521541.
e) Производные антител
В некоторых воплощениях антитело, предложенное в данном документе, может быть дополнительно модифицировано так, чтобы содержать дополнительные небелковые группировки, которые известны в данной области и легко доступны. Группировки, пригодные для получения производных антитела, включают, но не ограничиваясь ими, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают, но не ограничиваясь ими, полиэтиленгликоль (PEG), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимеры этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или рандомные сополимеры), и декстран или поли(п-винилпирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры оксида полипропиленгликоля/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликоля пропиональдегид может давать преимущества в производстве благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединенных к антителу, может варьировать, и если присоединено больше одного полимера, то они могут быть одинаковыми или различными молекулами. Как правило, число и/или тип полимеров, используемых для получения производных, может быть определен на основании соображений, включая, но не ограничиваясь ими, конкретные свойства или функции антитела, которые будут улучшены, будет ли использоваться производное антитела в терапии в определенных условиях и т.д.
В другом воплощении предложены конъюгаты антитела и небелковой группировки, которая может селективно нагреваться при воздействии радиации. В одном воплощении небелковая группировка представляет собой углеродную нанотрубку (Kam, N.W. et al., Proc. Natl. Acad. Sci. USA 102 (2005) 11600-11605). Излучение может иметь любую длину волны, включая, но не ограничиваясь ими, такие длины волн, которые не повреждают обычные клетки, но нагревают небелковую группировку до температуры, при которой клетки, ближайшие к небелковой группировке антитела, погибают.
f) Гетеродимеризация
Существует несколько подходов к СН3-модификации, обеспечивающих гетеродимеризацию, которые хорошо описаны, например, в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012058768, WO 2013157954, WO 2013096291. Как правило, во всех таких подходах первый СН3-домен и второй СН3-домены разработаны на основе комплементарности, так что каждый СН3-домен (или тяжелая цепь, содержащая его) не может больше гомодимеризоваться сам с собой, но вынужден гетеродимеризоваться с комплементарно разработанным другим СН3-доменом (так что первый и второй СН3-домен гетеродимеризуются, и между двумя первыми или двумя вторыми СН3-доменами гомодимеры не формируются). Эти различные подходы к улучшению гетеродимеризации тяжелой цепи рассматриваются как различные альтернативы в сочетании с модификациями тяжелой - легкой цепи (обмен/замена VH и VL в одной связывающей ветви и введение замен заряженных аминокислот с противоположными зарядами в контактную поверхность CH1/CL) в мультиспецифические антитела согласно изобретению, которые снижают число побочных продуктов типа Бенс-Джонса с ошибочным спариванием легкой цепи.
В одном предпочтительном воплощении данного изобретения (в случае, когда мультиспецифическое антитело содержит СН3-домены в тяжелых цепях) СН3-домены указанного мультиспецифического антитела согласно изобретению могут быть изменены с помощью технологии "ключ в замке", которая подробно описана на нескольких примерах, например в WO 96/027011, Ridgway J.В., et al., Protein Eng. 9 (1996) 617-621, и Merchant, A.M., et al., Nat. Biotechnol. 16 (1998) 677-681; WO 98/ 050431. В этом способе поверхности взаимодействия двух СН3-доменов изменены так, чтобы увеличить гетеродимеризацию обеих тяжелых цепей, содержащих эти два СН3-домена. Каждый из двух СН3-доменов (двух тяжелых цепей) может быть "ключом", а другой - "замком". Введение дисульфидного мостика дополнительно стабилизирует гетеродимеры (Merchant, А.М, et al., Nature Biotech. 16 (1998) 677-681, Atwell, S., et al. J. Mol. Biol. 270 (1997) 26-35) и повышает выход.
Так, в одном предпочтительном воплощении изобретения указанное мультиспецифическое антитело (содержит СН3-домен в каждой тяжелой цепи и) характеризуется тем, что
первый СН3-домен первой тяжелой цепи антитела в а) и второй СН3-домен второй тяжелой цепи антитела в b) встречаются на контактной поверхности, которая включает исходную контактную поверхность между СН3-доменами антитела;
где указанная контактная поверхность изменена таким образом, чтобы способствовать формированию мультиспецифического антитела, где изменение характеризуется тем, что
i) СН3-домен одной тяжелой цепи изменен так,
что в исходной контактной поверхности СН3-домена одной тяжелой цепи, которая встречается с исходной контактной поверхностью СН3-домена другой тяжелой цепи в мультиспецифическом антителе,
аминокислотный остаток заменен аминокислотным остатком, имеющим больший объем боковой цепи, тем самым формируя выступ в контактной поверхности СН3-домена одной тяжелой цепи, который может быть расположен в полости на контактной поверхности СН3-домена другой тяжелой цепи
и
ii) СН3-домен другой тяжелой цепи изменен так,
что в исходной контактной поверхности второго СН3-домена, которая встречается с исходной контактной поверхностью первого СН3-домена в мультиспецифическом антителе,
аминокислотный остаток заменен на аминокислотный остаток, имеющий меньший объем боковой цепи, тем самым формируя полость в контактной поверхности второго СН3-домена, в которой может быть расположен выступ на контактной поверхности первого СН3-домена.
Предпочтительно, указанный аминокислотный остаток, имеющий боковую цепь большего объема, выбран из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y), триптофана (W)
Предпочтительно указанный аминокислотный остаток, имеющий боковую цепь меньшего объема, выбран из группы, состоящей из аланина (А), серина (S), треонина (Т), валина (V).
В одном аспекте изобретения оба СН3-домена также изменены путем введения цистеина (С) в качестве аминокислоты в соответствующих позициях каждого СН3-домена, так что может быть сформирован дисульфидный мостик между двумя СН3-доменами.
В одном предпочтительном воплощении указанное мультиспецифическое антитело содержит аминокислотную мутацию T366W в первом СН3-домене "цепи-ключа" и аминокислотные мутации T366S, L368A, Y407V во втором СН3-домене "цепи-замка". Также может быть использован дополнительный межцепочечный дисульфидный мостик между СН3-доменами (Merchant, А.М, et al., Nature Biotech 16 (1998) 677-681), например, путем введения аминокислотной мутации Y349C в СН3-домен "цепи-ключа" и аминокислотной мутации Е356С или аминокислотной мутации S354C в СН3-домен "цепи-замка".
В одном предпочтительном воплощении указанное мультиспецифическое антитело (которое содержит СН3-домен в каждой тяжелой цепи) содержит аминокислотные мутации S354C, T366W в одном из двух СН3-доменов и аминокислотные мутации Y439C, T366S, L368A, Y407V в другом из двух СН3-доменов (дополнительная аминокислотная мутация S354C в одном СН3-домене и дополнительная аминокислотная мутация Y349C в другом СН3-домене для формирования межцепочечного дисульфидного мостика) (нумерация согласно индексу ЕС по Kabat).
Другие методики СН3-модификаций, обеспечивающие гетеродимеризацию, предусмотрены в качестве альтернатив и описаны, например, в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012058768, WO 2013157954, WO 2013096291.
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в ЕР 1870459 А1. Этот подход основан на введении замен/мутаций заряженных аминокислот с противоположным зарядом в определенных аминокислотных позициях на контактной поверхности СН3/СН3-доменов между обеими тяжелыми цепями. Одним из предпочтительных воплощений для указанного мультиспецифического антитела являются аминокислотные мутации R409D; K370E в первом СН3-домене (мультиспецифического антитела) и аминокислотные мутации D399K; E357K во втором СН3-домене мультиспецифического антитела (нумерация согласно Kabat).
В другом воплощении указанное мультиспецифическое антитело содержит аминокислотную мутацию T366W в СН3-домене "цепи-ключа" и аминокислотные мутации T366S, L368A, Y407V в СН3-домене "цепи-замка" и дополнительные аминокислотные мутации R409D; K370E в СН3-домене "цепи-ключа" и аминокислотные мутации D399K; E357K в СН3-домене "цепи-замка".
В другом воплощении указанное мультиспецифическое антитело содержит аминокислотные мутации S354C, T366W в одном из двух СН3-доменов и аминокислотные мутации Y349C, T366S, L368A, Y407V в другом из двух СН3-доменов, или указанное мультиспецифическое антитело содержит аминокислотные мутации Y349C, T366W в одном из двух СН3-доменов и аминокислотные мутации S354C, T366S, L368A, Y407V в другом из двух СН3-доменов и дополнительные аминокислотные мутации R409D; K370E в СН3-домене "цепи-ключа" и аминокислотные мутации D399K; E357K в СН3-домене "цепи-замка".
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2013/157953. В одном воплощении первый СН3-домен содержит аминокислотную мутацию T366K, а второй полипептид СН3-домена содержит аминокислотную мутацию L351D. В другом воплощении первый СН3-домен содержит дополнительную аминокислотную мутацию L351K. В другом воплощении второй СН3-домен содержит дополнительную аминокислотную мутацию, выбранную среди Y349E, Y349D и L368E (предпочтительно L368E).
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2012/058768. В одном воплощении первый СН3-домен содержит аминокислотные мутации L351Y, Y407A, а второй СН3-домен содержит аминокислотные мутации Т366А, K409F. В другом воплощении второй СН3-домен содержит дополнительную аминокислотную мутацию в позиции Т411, D399, S400, F405, N390 или K392, например, выбранную среди а) T411N, T411R, T411Q, T411K, T411D, Т411Е или T411W, b) D399R, D399W, D399Y или D399K, с) S400E, S400D, S400R или S400K, F405I, F405M, F405T, F405S, F405V или F405W, N390R, N390K или N390D, K392V, K392M, K392R, K392L, K392F или K392E. В другом воплощении первый СН3-домен содержит аминокислотные мутации L351Y, Y407A, а второй СН3-домен содержит аминокислотные мутации T366V, K409F. В другом воплощении первый СН3-домен содержит аминокислотные мутации Y407A, а второй СН3-домен содержит аминокислотные мутации Т366А, K409F. В другом воплощении второй СН3-домен содержит дополнительные аминокислотные мутации K392E, Т411Е, D399R и S400R.
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2011/143545, например, с модификацией аминокислоты в позиции, выбранной из группы, состоящей из 368 и 409.
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2011/090762, который также использует технологию типа "ключ в замке". В одном воплощении первый СН3-домен содержит аминокислотную мутацию T366W, а второй СН3-домен содержит аминокислотную мутацию Y407A. В одном воплощении первый СН3-домен содержит аминокислотную мутацию T366Y, а второй СН3-домен содержит аминокислотную мутацию Y407T.
В одном воплощении мультиспецифическое антитело относится к изотипу IgG2, и в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2010/129304.
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2009/089004. В одном воплощении первый СН3-домен содержит аминокислотную замену K392 или N392 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D), предпочтительно K392D или N392D), а второй СН3-домен содержит аминокислотную замену D399, Е356, D356 или Е357 положительно заряженной аминокислотой (например, лизином (К) или аргинином (R)), предпочтительно D399K, E356K, D356K или E357K, и более предпочтительно D399K и E356K. В другом воплощении первый СН3-домен также содержит аминокислотную замену K409 или R409 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D), предпочтительно K409D или R409D). В другом воплощении первый СН3-домен дополнительно или альтернативно содержит аминокислотную замену K439 и/или К370 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D)).
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2007/147901. В одном воплощении первый СН3-домен содержит аминокислотные мутации K253E, D282K и K322D, а второй СН3-домен содержит аминокислотные мутации D239K, E240K и K292D.
В одном воплощении в качестве альтернативы может быть использован подход к гетеродимеризации, описанный в WO 2007/110205.
Рекомбинантные способы и препараты
Антитела могут быть получены с использованием рекомбинантных способов и препаратов, например, описанных в US 4816567. В одном воплощении предложена изолированная (выделенная) нуклеиновая кислота(ы), кодирующая антитело, описанное в данном документе. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В другом воплощении предложен один или более чем один вектор (например, экспрессионный вектор), содержащий такую нуклеиновую кислоту. В другом воплощении предложена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном таком воплощении клетка-хозяин содержит (например, была трансформирована ими): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном воплощении клетка-хозяин является эукариотической, например, клеткой яичника китайского хомячка (СНО) или лимфоидной клеткой (например, Y0, NSO, Sp2/0). В одном воплощении предложен способ получения антитела, описанного в данном документе, который включает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, описанное выше, в условиях, подходящих для экспрессии антитела, и, возможно, восстановление антитела из клетки-хозяина (или культуральной среды клетки-хозяина).
Для рекомбинантной продукции вариантной Fc-области нуклеиновую кислоту, кодирующую вариантную Fc-область, например, описанную выше, выделяют и встраивают в один или более чем один вектор для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такая нуклеиновая кислота может быть легко выделена и секвенирована с использованием обычных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими полипептиды вариантных Fc-областей или тяжелые и легкие цепи антитела).
Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитела, включают прокариотические или эукариотические клетки, описанные в данном документе. Например, антитела могут быть получены в бактериях, в частности, когда не нужны гликозилирование и Fc-эффекторные функции. Про экспрессию фрагментов антитела и полипептидов в бактериях см., например, в US 5648237, US 5789199 и US 5840523 (см. также Charlton, K.A., In: Methods in Molecular Biology, Vol.248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2003), pp.245-254, где описывается экспрессия фрагментов антител в Е. coli). После экспрессии антитело может быть выделено из массы бактериальных клеток в растворимой фракции и может быть далее очищено.
Помимо прокариотов для векторов, кодирующих антитела, подходящими клонирующими или экспрессирующими хозяевами являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, включая штаммы грибов и дрожжей, у которых пути гликозилирования являются "гуманизированными", что приводит к продукции антитела с частично или полностью человеческим характером гликозилирования. См. Gerngross, T.U., Nat. Biotech. 22 (2004) 1409-1414; и Li, Н. et al., Nat. Biotech. 24 (2006) 210-215.
Подходящие клетки-хозяева для экспрессии гликозилированного антитела также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые могут быть использованы в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев также могут быть использованы культуры растительных клеток. См., например, US 5959177, US 6040498, US 6420548, US 7125978 и US 6417429 (описывающие технологию PLANTIBODIES для получения антител в трансгенных растениях)).
В качестве хозяев также могут быть использованы клетки позвоночных животных. Например, могут быть использованы клеточные линии млекопитающих, приспособленные к росту в суспензии. Другими примерами используемых клеточных линий-хозяев от млекопитающих являются линия почки обезьяны CV1, трансформированная SV40 (COS-7); человеческая эмбриональная почечная линия (HEK293 или клетки 293, описанные, например, в Graham, F.L. et al., J. Gen Virol. 36 (1977) 59-74); клетки почек детенышей хомяка (ВНК); мышиные клетки Сертоли (TM4-клетки, описанные, например, в Mather, J.P., Biol. Reprod. 23 (1980) 243-252); клетки почки обезьяны (CV1); клетки почки африканской зеленой обезьяны (VERO-76); клетки человеческой карциномы шейки матки (HELA); клетки почки собаки (MDCK); клетки печени крысы buffalo (BRL 3А); клетки легких человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); TRI-клетки, описанные, например, в Mather, J.P. et al., Annals N Y. Acad. Sci. 383 (1982) 44-68; клетки MRC 5 и клетки FS4. Другие используемые линии клеток-хозяев от млекопитающих включают клетки яичника китайского хомячка (СНО), в том числе DHFR-CHO-клетки (Urlaub, G. et al., Proc. Natl. Acad. Sci. USA 77 (1980) 4216-4220); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор некоторых линий клеток-хозяев от млекопитающих, подходящих для продукции антител, см., например, в Yazaki, P. and Wu, A.M., Methods in Molecular Biology, Vol.248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2004), pp.255-268.
Анализы
Антитела, предусмотренные в данном документе, могут быть идентифицированы, подвергнуты скринингу или охарактеризованы по их физическим/химическим свойствам и/или биологической активности с помощью различных анализов, известных в данной области.
В одном аспекте антитело, описанное в данном документе, анализируют на активность связывания с антигеном, например, с помощью известных способов, таких как ELISA, вестерн-блот и т.п.
Иммуноконъюгаты
Изобретение также предусматривает иммуноконъюгаты, содержащие антитело, описанное в данном документе, которое конъюгировано с одним или более чем одним цитотоксическим агентом, таким как химиотерапевтические агенты или лекарства, ингибирующие рост агенты, токсины (например, белковые токсины, ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения или их фрагменты) или радиоактивные изотопы.
В одном воплощении иммуноконъюгат является конъюгатом "антитело-лекарственное средство" (antibody-drug conjugate, ADC), в котором антитело конъюгировано с одним или более чем одним лекарственным средством, включая, но не ограничиваясь ими, мейтансиноид (см. US 5208020, US 5416064 и ЕР 0425235 В1); ауристатин, такой как монометилауристатиновые лекарственные группировки DE и DF (ММАЕ и MMAF) (см. US 5635483, US 5780588 и US 7498298); доластатин; калихеамицин или его производное (см. US 5712374, US 5714586, US 5739116, US 5767285, US 5770701, US 5770710, US 5773001 и US 5877296; Hinman, L.M. et al., Cancer Res. 53 (1993) 3336-3342; и Lode, H.N. et al., Cancer Res. 58 (1998) 2925-2928); антрациклины, такие как дауномицин или доксорубицин (см. Kratz, F. et al., Curr. Med. Chem. 13 (2006) 477-523; Jeffrey, S.C. et al., Bioorg. Med. Chem. Lett. 16 (2006) 358-362; Torgov, M.Y. et al., Bioconjug. Chem. 16 (2005) 717-721; Nagy, A. et al., Proc. Natl. Acad. Sci. USA 97 (2000) 829-834; Dubowchik, G.M. et al., Bioorg.& Med. Chem. Letters 12 (2002) 1529-1532; King, H.D. et al., J. Med. Chem. 45 (2002) 4336-4343; и US 6630579); метотрексат; виндезин; таксаны, такие как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен; и СС1065.
В другом воплощении иммуноконъюгат содержит антитело, описанное в данном документе, конъюгированное с ферментативно активным токсином или его фрагментом, включая, но не ограничиваясь ими, дифтерийную А-цепь, несвязывающиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модецина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytoiacca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Saponaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены.
В другом воплощении иммуноконъюгат содержит антитело, описанное в данном документе, конъюгированное с радиоактивным атомом с образованием радиоактивного конъюгата. Множество радиоактивных изотопов доступно для получения радиоактивных конъюгатов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Когда для обнаружения используется радиоактивный конъюгат, он может содержать радиоактивный атом для сцинтиграфических исследований, например, TC99m или I123, или спиновую метку для визуализации путем ядерного магнитного резонанса (NMR) (также известного как магнитно-резонансная томография, MRI), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента могут быть получены с использованием множества бифункциональных связывающих белки агентов, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(параазидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин может быть получен, как описано в Vitetta, E.S. et al., Science 238 (1987) 1098-1104. Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен-триамин-пентауксксная кислота (MX-DTPA) является примером хелатирующего агента для конъюгации радионуклида с антителом. См. WO 94/11026. Линкер может представлять собой "расщепляемый линкер", облегчающий высвобождение цитотоксического лекарственного агента в клетке. Например, может быть использован кислотно-лабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, линкер, содержащий диметил или дисульфид (Chari, R.V. et al., Cancer Res. 52 (1992) 127-131; US 5208020).
Иммуноконъюгаты или ADC в данном документе рассмотрены особо, но не ограничиваются такими конъюгатами, полученными с помощью сшивающих реагентов, включая, но не ограничиваясь ими, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые коммерчески доступны (например, Pierce Biotechnology, Inc., Рокфорд, Иллинойс, США).
Способы и препараты для диагностики и обнаружения
В некоторых воплощениях любые антитела, предусмотренные в данном документе, используются для обнаружения их родственного антигена(ов) в биологическом образце. Термин "обнаружение", используемый в данном документе, включает количественное или качественное обнаружение. В некоторых воплощениях биологический образец содержит клетку или ткань.
В одном воплощении предусмотрено антитело, описанное в данном документе, для применения в способах диагносики или обнаржуения.
В некоторых воплощениях предусмотрены меченые антитела, описанные в данном документе. Метки включают, но не ограничиваясь ими, метки или группировки, которые обнаруживаются непосредственно (такие как флуоресцентные, хромофорные, электронно-плотные, хемилюминесцентные и радиоактивные метки), а также группировки, такие как ферменты или лиганды, которые обнаруживаются косвенно, например, через ферментативную реакцию или молекулярное взаимодействие. Примеры меток включают, но не ограничиваясь ими, радиоизотопы 32Р, 14С, 125I, 3H и 131I, флуорофоры, такие как хелаты редкоземельных металлов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например люциферазу светлячка и бактериальную люциферазу (US 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, оксидазы сахаридов, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфатдегидрогеназу, гетероциклические оксидазы, например, уриказу и ксантиноксидазу, в сочетании с ферментом, который использует пероксид водорода для окисления предшественника метки, таким как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и т.п.
Фармацевтические препараты
Фармацевтические препараты антитела, описанного в данном документе, получают путем смешивания такого антитела, имеющего требуемую степень чистоты, с одним или более чем одним возможным фармацевтически приемлемым носителем (Remington's Pharmaceutical Sciences, 16th edition, Osol, A. (ed.) (1980)), в виде лиофилизированных препаратов или водных растворов. Фармацевтически приемлемые носители, как правило, нетоксичны для реципиентов в используемых дозах и концентрациях и включают, но не ограничиваясь ими: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензетониума хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (PEG). Иллюстративные фармацевтически приемлемые носители в данном документе также включают агенты для диспергирования лекарственного средства в интерстициальном пространстве, такие как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, человеческие растворимые гликопротеины гиалуронидазы РН-20, такие как rhuPH20 (HYLENEX®, Baxter International, Inc.). Некоторые типичные sHASEGP и способы их применения, в том числе rhuPH20, описаны в US 2005/0260186 и US 2006/0104968. В одном аспекте sHASEGP комбинирован с одной или более чем одной дополнительной гликозамингликаназой, такой как хондроитиназа.
Иллюстративные лиофилизированные препараты антител описаны в US 6267958. Водные препараты антител включают те, которые описаны в US 6171586 и WO 2006/044908, последние препараты включают гистидин-ацетатный буфер.
Препарат в данном документе также может содержать более чем один активный ингредиент, что нужно при лечении по конкретным показаниям, предпочтительно с дополняющими активностями, которые не влияют друг на друга. Такие активные ингредиенты предпочтительно присутствуют в комбинации в количествах, которые эффективны для данной цели.
Активные ингредиенты могут быть заключены в микрокапсулы, полученные, например, с помощью методик коацервации или путем межфазной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметакрилатные) микрокапсулы, соответственно, в коллоидных системах доставки лекарственных средств (таких как липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие методики раскрыты в Remington's Pharmaceutical Sciences, 16th edition, Osol, A. (ed.)(1980).
Могут быть изготовлены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, при этом матрицы находятся в виде формованных изделий, например, пленок, или в виде микрокапсул.
Препараты, используемые для введения in vivo, как правило, являются стерильными. Стерильность может быть легко достигнута, например, путем фильтрации через стерилизующие фильтрационные мембраны.
Терапевтические способы и препараты
Любое из антител, предусмотренных в данном документе, может быть использовано в терапевтических способах.
В одном аспекте предусмотрено антитело, описанное в данном документе, для применения в качестве лекарственного средства.
В некоторых воплощениях предусмотрено антитело для применения в способе лечения. В одном таком воплощении указанный способ также включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, как описано ниже. "Индивидуум" в соответствии с любым из вышеописанных воплощений в предпочтительном воплощении является человеком.
В другом аспекте изобретение предусматривает применение антитела в изготовлении или получении лекарственного средства. "Индивидуум" в соответствии с любым из вышеописанных воплощений может быть человеком.
В другом аспекте изобретение предусматривает фармацевтические препараты, содержащие любое из антител, предусмотренных в данном документе, например, для применения в любом из указанных выше терапевтических способов. В одном воплощении фармацевтический препарат содержит любое из антител, предусмотренных в данном документе, и фармацевтически приемлемый носитель. В другом воплощении фармацевтический препарат содержит любое из антител, предусмотренных в данном документе, и по меньшей мере один дополнительный терапевтический агент, например, как описано ниже.
Антитела, описанные в данном документе, могут быть использованы в терапии по отдельности или в комбинации с другими агентами. Например, антитело, описанное в данном документе, может быть введено совместно по меньшей мере с одним дополнительным терапевтическим агентом.
Антитело, описанное в данном документе (и любой дополнительный терапевтический агент), может быть введено с помощью любых подходящих средств, в том числе путем парентерального, внутрилегочного и интраназального, и, если нужно местное лечение, внутриочагового введения. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Введение дозы может быть осуществлено любым подходящим способом, например, с помощью инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или хроническим. В данном документе рассматриваются различные схемы дозировки, включая, но не ограничиваясь ими, одно или множество введений в различные временные точки, болюсное введение и импульсную инфузию.
Антитела, описанные в данном документе, будут собраны в препарат, дозированы и введены в соответствии с надлежащей медицинской практикой. Факторы, подлежащие рассмотрению в этом контексте, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину нарушения, место доставки агента, способ введения, схему введения и другие факторы, известные практикующим врачам. Антитело не обязательно, но может быть собрано в препарат с одним или более чем одним агентом, используемым в настоящее время для профилактики или лечения рассматриваемого нарушения. Эффективное количество таких других агентов зависит от количества антитела, присутствующего в препарате, от типа заболевания или лечения и от других факторов, описанных выше. Как правило, они используются в тех же дозировках и с теми же путями введения, которые описаны в данном документе, или в дозировках примерно от 1 до 99% от описанных в данном документе, или в любой дозировке и любым путем, который является эмпирически/клинически подходящим.
Для профилактики или лечения заболевания подходящая доза антитела, описанного в данном документе (при использовании его отдельно или в сочетании с одним или более чем одним другим дополнительным терапевтическим агентом), будет зависеть от типа заболевания, подлежащего лечению, от типа антитела, от тяжести и течения заболевания, от того, вводят ли антитело с профилактической или терапевтической целью, от предыдущей терапии, истории болезни пациента и ответа на антитело и по усмотрению лечащего врача. Антитело вводят пациенту подходящим образом одномоментно или серийно. В зависимости от типа и тяжести заболевания изначальной кандидатной дозировкой антитела для введения пациенту может быть дозировка от примерно 1 мкг/кг до 15 мг/кг (например, от 0,1 мг/кг до 10 мг/кг) независимо от того, например, вводится ли оно в ходе одного или более отдельных введений или путем непрерывной инфузии. Одна из типичных суточных доз может варьировать от примерно 1 мкг/кг до 100 мг/кг или более в зависимости от указанных выше факторов. В случае повторных введений в течение нескольких дней или дольше в зависимости от состояния лечение, как правило, может быть продолжено до требуемого подавления симптомов заболевания. Один из примеров дозировки антитела будет находиться в диапазоне от примерно 0,05 мг/кг до примерно 10 мг/кг. Таким образом, пациенту может быть введена одна или более чем одна доза примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любая их комбинация). Такие дозы можно вводить периодически, например, каждую неделю или каждые три недели (например, так, чтобы пациент получал от примерно двух до примерно двадцати доз антитела или, например, примерно шесть доз антитела). Может быть введена изначальная более высокая нагрузочная доза с последующей одной или более чем одной более низкой дозой. Прогресс этой терапии легко контролировать с помощью обычных методик и анализов.
Понятно, что любой из вышеуказанных препаратов или терапевтических способов может быть осуществлен с использованием иммуноконъюгата, описанного в данном документе, вместо или в дополнение к антителу, описанному в данном документе.
Продукты производства
В другом аспекте, описанном в данном документе, предусмотрен продукт производства, содержащий материалы, используемые для лечения, профилактики и/или диагностики нарушений, описанных выше. Продукт производства содержит контейнер и этикетку или листок-вкладыш, прикрепленный или связанный с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для внутривенно вводимых растворов и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит препарат сам по себе или в сочетании с другим препаратом, эффективным для лечения, профилактики и/или диагностики состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенно вводимого раствора или флакон с пробкой, прокалываемой иглой для подкожной инъекции). По меньшей мере один активный агент в препарате представляет собой антитело, описанное в данном документе. На этикетке или вкладыше в упаковку указывается, что препарат используется для лечения выбранного состояния. Кроме того, продукт производства может включать: (а) первый контейнер с препаратом, содержащимся в нем, где препарат содержит антитело, описанное в данном документе; и (b) второй контейнер с препаратом, содержащимся в нем, где препарат также содержит цитотоксический или другой терапевтический агент.Продукт производства в этом воплощении изобретения также может содержать вкладыш в упаковку, указывающий, что препараты могут быть использованы для лечения конкретною состояния. Альтернативно или дополнительно, изделие также может включать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, например, бактериостатическую воду для инъекций (bacteriostatic water for injection, BWFI), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Он также может включать другие материалы, желательные с коммерческой и потребительской точки зрения, в том числе другие буферы, разбавители, фильтры, иглы и шприцы.
Понятно, что любой из указанных выше продуктов производства может включать иммуноконъюгат, описанный в данном документе, вместо или в дополнение к антителу, описанному в данном документе.
III. Конкретные воплощения
1. Fc-область класса IgG, содержащая первый полипептид вариантной Fc-области и второй полипептид вариантной Fc-области, где
a) первый полипептид вариантной Fc-области получен из первого родительскою полипептида Fc-области класса IgG, а второй полипептид вариантной Fc-области получен из второго родительскою полипептида Fc-области класса IgG, где первый родительский полипептид Fc-области класса IgG идентичен или отличается от второго родительского полипептида Fc-области класса IgG, и
b) первый полипептид вариантной Fc-области отличается от второго полипептида вариантной Fc-области в одном или более чем одном аминокислотном остатке, отличающемся от тех аминокислотных остатков, в которых первый родительский полипептид Fc-области класса IgG отличается от второго родительского полипептида Fc-области класса IgG, и
с) Fc-область класса IgG, содержащая первый полипептид вариантной Fc-области и второй полипептид вариантной Fc-области, обладает аффинностью к человеческому Fc-рецептору, который отличается тем, что Fc-область класса IgG содержит первый родительский полипептид Fc-области класса IgG из а) и второй родительский полипептид Fc-области класса IgG из а).
2. Fc-область класса IgG согласно воплощению 1, где человеческий Fc-рецептор представляет собой человеческий неонатальный Fc-рецептор (FcRn) или человеческий Fc-гамма-III-рецептор (FcyRIII).
3. Fc-область класса IgG согласно любому из воплощений 1-2, где человеческий Fc-рецептор представляет собой человеческий неонатальный Fc-рецептор.
4. Fc-область класса IgG согласно любому из воплощений 1-3, где аффинность Fc-области класса IgG, содержащей первый полипептид вариантной Fc-области и второй полипептид вариантной Fc-области к человеческому Fc-рецептору увеличивается или уменьшается на 10% или более, что определяется в анализе поверхностного плазмонного резонанса (SPR), по сравнению с Fc-областью класса IgG, содержащей первый родительский полипептид Fc-области класса IgG из а) и второй родительский полипептид Fc-области класса IgG а).
5. Fc-область класса IgG согласно любому из воплощений 1-4, где по меньшей мере некоторые из тех аминокислотных остатков, в которых первый родительский полипептид Fc-области класса IgG отличается от второго родительского полипептида Fc-области класса IgG, содействуют формированию гетеродимерной Fc-области класса IgG.
6. Fc-область класса IgG согласно любому из воплощений 1-5, где
I) первый родительский полипептид Fc-области класса IgG выбран из группы, включающей
- полипептид Fc-области человеческого IgG1,
- полипептид Fc-области человеческого IgG2,
- полипептид Fc-области человеческого IgG3,
- полипептид Fc-области человеческого IgG4,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A,
- полипептид Fc-области человеческого IgG1 с мутациями Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутацией P329G,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G,
- полипептид Fc-области человеческого IgG4 с мутациями Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутацией P329G,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, S354C, T366S, L368A, Y407V,
- человеческий IgG1, IgG2 или IgG4 с мутацией K392D, и
- человеческий IgG3 с мутацией N392D,
и
ii) второй родительский полипептид Fc-области класса IgG выбран из группы, содержащей
- полипептид Fc-области человеческого IgG1,
- полипептид Fc-области человеческого IgG2,
- полипептид Fc-области человеческого IgG3,
- полипептид Fc-области человеческого IgG4,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A,
- полипептид Fc-области человеческого IgG1 с мутациями S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями Y349C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, Y349C, T366W,
- полипептид Fc-области человеческого IgG1 с мутацией P329G,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, Y349C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G,
- полипептид Fc-области человеческого IgG4 с мутациями S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутацией P329G,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, Y349C, T366W,
- человеческий IgG1 с мутациями D399K, D356K и/или E357K, и
- человеческий IgG2, IgG3 или IgG4 с мутациями D399K, E356K и/или E357K.
7. Fc-область класса IgG согласно любому из воплощений 1-6, где
I) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность, выбранную из группы, содержащей SEQ ID №60, 61, 62, 63, 64, 65, 67, 69, 70, 71,73, 75, 76, 78, 80, 81, 82 и 84,
и
ii) второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность, выбранную из группы, содержащей SEQ ID №60, 61, 62, 63, 64, 66, 68, 69, 70, 72, 74, 75, 76, 77, 79, 81, 83 и 85.
8. Fc-область класса IgG согласно любому из воплощений 1-7, где
i) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1, или
ii) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, или
iii) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, или
iv) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, S354C, T366W, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, Y349C, T366S, L368A, Y407V, или
v) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, S354C, T366W, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области человеческою IgG1 с мутациями L234A, L235A, P329G, Y349C, T366S, L368A, Y407V, или
vi) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4, или
vii) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, или
viii) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, P329G, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, P329G, или
ix) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, S354C, T366W, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, Y349C, T366S, L368A, Y407V, или
x) первый родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, P329G, S354C, T366W, а второй родительский полипептид Fc-области класса IgG представляет собой полипептид Fc-области IgG4 с мутациями S228P, L235E, P329G, Y349C, T366S, L368A, Y407V.
9. Fc-область класса IgG согласно любому из воплощений 1-5 и 7, где I) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №60, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №60, или
ii) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №64, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №64, или
iii) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №70, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №70, или
iv) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №68, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №67, или
v) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №74, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №73, или
vi) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №63, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №63, или
vii) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №75, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №75, или
viii) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №76, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №76, или
ix) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №79, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №80, или
x) первый родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №85, а второй родительский полипептид Fc-области класса IgG имеет аминокислотную последовательность SEQ ID №84.
10. Fc-область класса IgG согласно любому из воплощений 1-9, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в 1-8 аминокислотных остатках, отличных от тех аминокислотных остатков, в которых первый родительский полипептид Fc-области класса IgG отличается от второго родительского полипетида Fc-области класса IgG.
11. Fc-область класса IgG согласно любому из воплощений 1-10, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в 1-6 аминокислотных остатках, отличных от тех аминокислотных остатков, в которых первый родительский полипептид Fc-области класса IgG отличается от второго родительского полипетида Fc-области класса IgG.
12. Fc-область класса IgG согласно любому из воплощений 1-11, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в 1-3 аминокислотных остатках, отличных от тех аминокислотных остатков, в которых первый родительский полипептид Fc-области класса IgG отличается от второго родительского полипетида Fc-области класса IgG.
13. Fc-область класса IgG согласно любому из воплощений 1-12, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в одном или более чем одном из аминокислотных остатков в позициях 228, 234, 235, 236, 237, 238, 239, 248, 250, 251, 252, 253, 254, 255, 256, 257, 265, 266, 267, 268, 269, 270, 272, 285, 288, 290, 291, 297, 298, 299, 307, 308, 309, 310, 311, 314, 327, 328, 329, 330, 331, 332, 385, 387, 428, 433, 434, 435 и 436 (нумерация в соответствии с индексом ЕС по системе нумерации Kabat).
14. Fc-область класса IgG согласно любому из воплощений 1-2 и 4-13, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в одном или более чем одном из аминокислотных остатков в позициях 228, 234, 235, 236, 237, 238, 239, 253, 254, 265, 266, 267, 268, 269, 270, 288, 297, 298, 299, 307, 311, 327, 328, 329, 330, 331, 332, 434 и 435 (нумерация в соответствии с индексом ЕС по системе нумерации Kabat).
15. Fc-область класса IgG согласно любому из воплощений 1-2 и 4-14, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в одном или более чем одном из аминокислотных остатков в позициях 233, 236, 265, 297, 329 и 331.
16. Fc-область класса IgG согласно воплощению 15, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области одним или более чем одним аминокислотным изменением Е233Р, AG236, D265A, N297A, N297D, Р329А и P331S.
17. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-13, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в одном или более чем одном из аминокислотных остатков в позициях 248, 250, 251, 252, 253, 254, 255, 256, 257, 272, 285, 288, 290, 291, 308, 309, 310, 311,314, 385, 386, 387, 428, 432, 433,434, 435 и 436.
18. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-17, где первый полипетид вариантной Fc-области отличается от второго полипетида вариантной Fc-области в одном или более чем одном из аминокислотных остатков в позициях 251, 253, 310, 314, 432, 433, 435 и 436.
19. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-18, где первый полипетид Fc-области отличается одной или двумя мутациями, выбранными из i) группы I253A, Н310А и Н435А, или ii) группы Н310А, Н433А и Y436A, или iii) группы L251D, L314D и L432D, или iv) группы L251S, L314S и L432S (нумерация в соответствии с индексом ЕС по системе нумерации Kabat), от второго полипетида Fc-области, и второй полипетид Fc-области отличается одной или двумя мутациями, выбранными из группы, включающей мутации L251D, L251S, I253A, Н310А, L314D, L314S, L432D, L432S, Н433А, Н435А и Y436A (нумерация в соответствии с индексом ЕС по системе нумерации Kabat), от первого полипетида Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A, или iii) L251D, L314D и L432D, или iv) L251S, L314S и L432S содержатся в Fc-области.
20. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-18, где первый полипетид Fc-области отличается одной или двумя мутациями, выбранными из i) группы I253A, Н310А и Н435А, или ii) группы Н310А, Н433А и Y436A (нумерация в соответствии с индексом ЕС по системе нумерации Kabat), от второго полипетида Fc-области, и второй полипетид Fc-области отличается одной или двумя мутациями, выбранными из группы, включающей мутации I253A, Н310А, Н433А, Н435А и Y436A (нумерация в соответствии с индексом ЕС по системе нумерации Kabat), от первого полипетида Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A содержатся в Fc-области.
21. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-18, содержащая мутации I253A/H310A/H435A, или H310A/H433A/Y436A, или L251D/L314D/L432D, или L251S/L314S/L432S, или их комбинацию в Fc-области (нумерация в соответствии с индексом ЕС по системе нумерации Kabat), где I) все мутации находятся в первом или втором полипептиде Fc-области, или ii) одна или две мутации находятся в первом полипептиде Fc-области, и одна или две мутации находятся во втором полипептиде Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A, или iii) L251D, L314D и L432D, или iv) L251S, L314S и L432S содержатся в Fc-области.
22. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-18, содержащая мутации I253A/H310A/H435A или H310A/H433A/Y436A или их комбинацию в Fc-области (нумерация в соответствии с индексом ЕС по системе нумерации Kabat), где i) все мутации находятся в первом или втором полипептиде Fc-области, или ii) одна или две мутации находятся в первом полипептиде Fc-области, и одна или две мутации находятся во втором полипептиде Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации i) I253A, Н310А и Н435А, или ii) Н310А, Н433А и Y436A содержатся в Fc-области.
23. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-22, где Fc-область класса IgG демонстрирует сниженное связывание со стафилококковым белком А по сравнению с Fc-областью класса IgG, содержащей первый родительский полипептид Fc-области класса IgG из а) и второй родительский полипептид Fc-области класса IgG из а).
24. Fc-область класса IgG согласно любому из воплощений 1-3 и 5-17, содержащая мутации M252Y/S254T/T256E в Fc-области (нумерация в соответствии
- с индексом ЕС по системе нумерации Kabat), где i) все мутации находятся в первом или втором полипептиде Fc-области, или ii) одна или две мутации находятся в первом полипептиде Fc-области, и одна или две мутации находятся во втором полипептиде Fc-области, так что все мутации в первом и втором полипептиде Fc-области, взятые вместе, приводят к тому, что мутации M252Y/S254T/T256E содержатся в Fc-области класса IgG.
25. Антитело, содержащее Fc-область класса IgG согласно любому из воплощений 1-24.
26. Антитело согласно воплощению 25, которое представляет собой моноклональное антитело.
27. Антитело согласно любому из воплощений 25-26, которое представляет собой человеческое, гуманизированное или химерное антитело.
28. Антитело согласно любому из воплощений 25-27, которое представляет собой биспецифическое антитело.
29. Антитело согласно любому из воплощений 25-28, которое представляет собой двухвалентное антитело.
30. Антитело согласно любому из воплощений 25-29, которое представляет собой биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2.
31. Биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее первый антигенсвязывающий сайт, который специфически связывается с человеческим VEGF, и второй антигенсвязывающий сайт, который специфически связывается с человеческим ANG-2,
где
α) первый антигенсвязывающий сайт, специфически связывающийся с VEGF, содержит в вариабельном домене тяжелой цепи CDRSH-область с SEQ ID №14, CDR2H-область с SEQ ID №15 и CDR1H-область с SEQ ID №16, и в вариабельном домене легкой цепи CDR3L-область с SEQ ID №17, CDR2L-область с SEQ ID №18 и CDR1L-область с SEQ ID №19, и
β) второй антигенсвязывающий сайт, специфически связывающийся с ANG-2, содержит в вариабельном домене тяжелой цепи CDR3H-область с SEQ ID №22, CDR2H-область с SEQ ID №23 и CDR1H-область с SEQ ID №24, и в вариабельном домене легкой цепи CDR3L-область с SEQ ID №25, CDR2L-область с SEQ ID №26 и CDR1L-область с SEQ ID №27, и
γ) биспецифическое антитело содержит Fc-область класса IgG согласно любому из воплощений 1-24.
32. Биспецифическое антитело согласно воплощению 31, где
α) первый антигенсвязывающий сайт, специфически связывающийся с VEGF, содержит в качестве вариабельного домена тяжелой цепи VH аминокислотную последовательность SEQ ID №20, а в качестве вариабельного домена легкой цепи VL аминокислотную последовательность SEQ ID №21, и
β) второй антигенсвязывающий сайт, специфически связывающийся с ANG-2, содержит в качестве вариабельного домена тяжелой цепи VH аминокислотную последовательность SEQ ID №28, а в качестве вариабельного домена легкой цепи VL аминокислотную последовательность SEQ ID №29, и
γ) биспецифическое антитело содержит Fc-область класса IgG согласно любому из воплощений 1-24.
33. Биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, в котором константные домены CL и СН1 заменены друг на друга, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №38, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №40, и
β) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №39, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №41.
34. Биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, в котором константные домены CL и СН1 заменены друг на друга, содержащее
а) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №34, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №36, и
В) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №35, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №37.
35. Биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, в котором константные домены CL и СН1 заменены друг на друга, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №42, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №44, и
β) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №43, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №45.
36. Биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, в котором константные домены CL и СН1 заменены друг на друга, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №90, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №40, и
β) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №91, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №41.
37. Биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, в котором константные домены CL и СН1 заменены друг на друга, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №88, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №36, и
β) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №89, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №37.
38. Биспецифическое двухвалентное антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и модифицированную тяжелую цепь и модифицированную легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, в котором константные домены CL и СН1 заменены друг на друга, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №92, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №44, и
β) в качестве модифицированной тяжелой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №93, а в качестве модифицированной легкой цепи второго полноразмерного антитела аминокислотную последовательность SEQ ID №45.
39. Биспецифическое антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и тяжелую цепь и легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, где N-конец тяжелой цепи связан с С-концом легкой цепи через пептидный линкер, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №46, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №48, и
β) в качестве тяжелой цепи второго полноразмерного антитела, связанной через пептидный линкер с легкой цепью второго полноразмерного антитела, аминокислотную последовательность SEQ ID №47.
40. Биспецифическое антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и тяжелую цепь и легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, где N-конец тяжелой цепи связан с С-концом легкой цепи через пептидный линкер, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №49, и в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №51, и
β) в качестве тяжелой цепи второго полноразмерного антитела, связанной через пептидный линкер с легкой цепью второго полноразмерного антитела, аминокислотную последовательность SEQ ID №50.
41. Биспецифическое антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и тяжелую цепь и легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, где N-конец тяжелой цепи связан с С-концом легкой цепи через пептидный линкер, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №94, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №48, и
β) в качестве тяжелой цепи второго полноразмерного антитела, связанной через пептидный линкер с легкой цепью второго полноразмерного антитела, аминокислотную последовательность SEQ ID №95.
42. Биспецифическое антитело с отмененным FcRn-связыванием, содержащее тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с VEGF, и тяжелую цепь и легкую цепь второго полноразмерного антитела, которое специфически связывается с ANG-2, где N-конец тяжелой цепи связан с С-концом легкой цепи через пептидный линкер, содержащее
α) в качестве тяжелой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №96, а в качестве легкой цепи первого полноразмерного антитела аминокислотную последовательность SEQ ID №51, и
β) в качестве тяжелой цепи второго полноразмерного антитела, связанной через пептидный линкер с легкой цепью второю полноразмерного антитела, аминокислотную последовательность SEQ ID №97.
43. Биспецифическое антитело согласно любому из воплощений 39-42, где вариабельный домен тяжелой цепи антитела (VH) и вариабельный домен легкой цепи антитела (VL) тяжелой и легкой цепи второго полноразмерного антитела стабилизированы путем введения дисульфидного мостика между позицией 44 в вариабельном домене тяжелой цепи и позицией 100 в вариабельном домене легкой цепи (нумерация согласно Kabat).
44. Биспецифическое антитело согласно любому из воплощений 33-43, где биспецифическое антитело содержит мутации S354C, T366W в одном из двух СН3-доменов и мутации Y349C, T366S, L368A, Y407V в другом из двух СН3-доменов, или где биспецифическое антитело содержит мутации Y349C, T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A, Y407V в другом из двух СН3-доменов.
45. Биспецифическое антитело согласно любому из воплощений 33-43, где биспецифическое антитело содержит мутации S354C, T366W в одном из двух СН3-доменов и мутации Y349C, T366S, L368A, Y407V в другом из двух СН3-доменов, а также мутации R409D, К370Е в СН3-домене в дополнение к мутациям Y349C, T366S, L368A, Y407V и мутации D399K, E357K в СН3-домене в дополнение к мутациям S354C, T366W, или где биспецифическое антитело содержит мутации Y349C, T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A, Y407V в другом из двух СН3-доменов, а также мутации R409D, К370Е в СН3-домене в дополнение к мутациям S354C, T366S, L368A, Y407V и мутации D399K, E357K в СН3-домене в дополнение к мутациям Y349C, T366W.
46. Биспецифическое антитело согласно любому из воплощений 25-45, где антитело имеет одно или более чем одно из следующих свойств:
- демонстрирует более низкую сывороточную концентрацию по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (через 96 часов после интравитреального введения мышам, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn), и/или
- демонстрирует аналогичную (коэффициент от 0,8 до 1,2) концентрацию в цельных лизатах правого глаза по сравнению с соответствующим биспецифическим антителом без мутаций, описанных в iii) (у мышей, которые являются FcRn-дефицитными, но гемизиготно трансгенными по человеческому FcRn, через 96 часов после интравитреального введения в правый глаз), и/или
- не демонстрирует связывания с человеческим неонатальным Fc-рецептором, и/или
- не демонстрирует связывания со стафилококковым белком А, и/или
- демонстрирует связывание со стафилококковым белком А.
47. Гибридный полипептид Fc-области, содержащий Fc-область класса IgG согласно любому из воплощений 1-24.
48. Фармацевтический препарат, содержащий антитело согласно любому из воплощений 25-46 или гибридный полипептид Fc-области согласно воплощению 47.
49. Фармацевтический препарат согласно воплощению 48, который предназначен для применения в лечении глазных сосудистых заболеваний.
50. Антитело согласно любому из воплощений 25-46 или гибридный полипептид Fc-области согласно воплощению 47 для применения в качестве лекарственного средства.
51. Применение согласно воплощению 50, где применение заключается в лечении глазных сосудистых заболеваний.
52. Применение антитела согласно любому из воплощений 25-46 или гибридного полипептида Fc-области согласно воплощению 47 в производстве лекарственного средства.
53. Применение согласно воплощению 52, где применение заключается в изготовлении лекарственного средства для лечения глазного сосудистого заболевания.
54. Антитело согласно любому из воплощений 25-46 или гибридный полипептид Fc-области согласно воплощению 47 для применения в лечении глазного сосудистого заболевания.
55. Способ лечения пациента, страдающего от глазных сосудистых заболеваний, путем введения антитела согласно любому из воплощений 25-46 или гибридного полипептида Fc-области согласно воплощению 47 пациенту в случае необходимости такого лечения.
IV. Примеры
Ниже приведены примерь; способов и препаратов, описанных в данном документе. Понятно, что могут быть осуществлены различные другие воплощения, учитывая общее описание, данное выше.
Хотя данное изобретение было подробно описано посредством иллюстраций и примеров для ясности понимания, эти описания и примеры не должны быть истолкованы как ограничивающие объем данного изобретения. Раскрытие всей патентной и научной литературы, цитируемой в данном документе, ясно включено во всей ее полноте посредством ссылки.
Методы
Масс-спектоометоия с ионизацией электрораспылением (ESI-MC)
Аликвоты белка (50 мкг) дегликозилировали добавлением 0,5 мкл реагента N-Glycanase plus (Roche) и натрий-фосфатного буфера (0,1 М, рН 7,1) для получения конечного объема образца 115 мкл. Смесь инкубировали при 37°С в течение 18 ч. Затем для восстановления и денатурации добавляли 60 мкл 0,5 М ТСЕР (Pierce) в 4 М растворе гуанидина*HCl (Pierce) и 50 мкл 8 M раствора гуанидина*HCl. Смесь инкубировали при 37°C в течение 30 мин. Образцы обессоливали путем эксклюзионной хроматографии (Sepharose G-25, изократическая, 40% ацетонитрила с 2% муравьиной кислотой). ESI-масс-спектры (положительные) записывали на приборе Q-TOF (maXis, Bruker), оснащенном источником ионов nano-ESI (TriVersa NanoMate, Advion). Настройки MS-параметров были следующими: перенос: RF воронки, 400 Vpp; энергия ISCID, 0 эВ; мультиполярный RF, 400 Vpp; квадруполь: энергия иона, 4,0 эВ; нижний предел, 600 m/z; источник: сухой газ, 8 л/мин; температура сухого газа, 160°C; столкновительная ячейка: энергия столкновения, 10 эВ; RF столкновения: 2000 Vpp; охлаждение ионов: RF охлаждения ионов, 300 Vpp; время переноса: 120 мкс; время перед импульсом, 10 мкс; диапазон сканирования m/z от 600 до 2000. Для оценки данных использовали самостоятельно разработанное программное обеспечение (MassAnalyzer).
Поверхностный плазмонный резонанс (SPR) FcRn
Связывающие свойства антитела дикого типа и мутантов с FcRn определяли путем методики поверхностного плазмонного резонанса (SPR) с помощью инструмента BIAcore Т100 (BIAcore АВ, Упсала, Швеция). Эта система хорошо известна для изучения молекулярных взаимодействий. Она позволяет осуществлять в режиме реального времени непрерывный мониторинг связываний лиганда/анализируемого вещества и, таким образом, определять кинетические параметры в различных условиях анализа. SPR-технология основана на измерении показателя преломления вблизи поверхности покрытого золотом биосенсорного чипа. Изменения показателя преломления отражают изменения массы на поверхности, вызванные взаимодействием иммобилизованного лиганда с анализируемым веществом, введенным в растворе. Если молекулы связывают иммобилизованный лиганд на поверхности, то масса увеличивается, в случае диссоциации масса уменьшается. Для данного анализа рецептор FcRn иммобилизировали на биосенсорном чипе BIAcore CM5 (GE Healthcare Bioscience, Упсала, Швеция) путем аминного связывания на уровне 400 единиц ответа (RU). Анализ проводили при комнатной температуре, используя в качестве рабочего буфера и буфера для разведения PBS, 0,05 Tween-20TM, рН 6,0 (GE Healthcare Bioscience). 200 нМ образцов антител вводили со скоростью потока 50 мкл/мин при комнатной температуре. Время ассоциации составляло 180 секунд, фаза диссоциации занимала 360 секунд. Регенерации поверхности чипа достигали путем короткого введения HBS-P, рН 8,0. Оценку SPR-данных проводили путем сравнения высоты сигнала биологического ответа на 180 секунде после введения и на 300 секунде после введения. Соответствующие параметры представляли максимальный уровень RU (180 секунд после введения) и позднюю стабильность (300 секунд после окончания введения).
Поверхностный плазменный резонанс (SPR) белка А
Анализ основан на спектроскопии поверхностного плазмонного резонанса. Белок А иммобилизовали на поверхности SPR-биосенсора. При введении образца в проточные ячейки SPR-спектрометра он образует комплекс с иммобилизованным белком А, что приводит к увеличению массы на поверхности сенсорного чипа и, следовательно, к более высокому ответу (1 RU определяется как 1 пг/мм2). После этого сенсорный чип подвергают регенерации путем растворения комплекса "образец - белок А". Затем полученные ответы оценивают по максимальной высоте сигнала в единицах ответа (RU) и поведению при диссоциации.
Примерно 3500 единиц ответа (RU) белка А (20 мкг/мл) было связано на чипе CM5 (GE Healthcare) при рН 4,0 с помощью набора для аминного связывания от GE Healthcare.
Буфер для образцов и системный буфер представлял собой HBS-P+(0,01 M HEPES, 0,15 M NaCI, 0,005% поверхностно-активного вещества Р20, стерильно профильтрованный, рН 7,4). Температура проточной ячейки была установлена на 25°C, а температуре отсека для образца на 12°C. Вначале в систему вводили рабочий буфер. Затем вводили 5 нМ растворы образцов конструкций в течение 120 секунд со скоростью потока 30 мкл/мин, а затем следовала фаза диссоциации длиной 300 секунд. Затем поверхность сенсорного чипа подвергали регенерации с помощью двух введений глицина-HCl с рН 1,5 в течение 30 секунд со скоростью потока 30 мкл/мин. Каждый образец измеряли трижды.
Биспецифические антитела и их соответствующие последовательности


Термин "с мутацией IHH-AAA", используемый в данном документе, относится к комбинации мутаций I253A (Не253А1а), Н310А (His310Ala) и Н435А (His435Ala) в константной области тяжелой цепи подклассов IgG1 или IgG4 (нумерация согласно индексу ЕС по системе нумерации Kabat); термин "с мутацией HHY-AAA", используемый в данном документе, относится к комбинации мутаций Н310А (His310Ala), Н433А (His433Ala) и Y436A (Tyr436Ala) в константной области тяжелой цепи подклассов IgG1 или IgG4 (нумерация согласно индексу ЕС по системе нумерации Kabat), термин "с мутацией P329G LALA", используемый в данном документе, относится к комбинации мутаций L234A (Leu234Ala), L235A (Leu235Ala) и P329G (Pro329Gly) в константной области тяжелой цепи подкласса IgG1 (нумерация согласно индексу ЕС по системе нумерации Kabat), а термин "с мутацией SPLE", используемый в данном документе, относится к комбинации мутаций S228P (Ser228Pro) и L235E (Leu235Glu) в константной области тяжелой цепи подкласса IgG4 (нумерация согласно индексу ЕС по системе нумерации Kabat).
Общая информация
Общая информация о нуклеотидных последовательностях легких и тяжелых цепей человеческих иммуноглобулинов приведена в: Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991). Аминокислотные остатки цепей антител пронумерованы согласно нумерации ЕС (Edelman, G.M., et al., Proc. Natl. Acad. Sci. USA 63 (1969) 78-85; Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)).
Технологии рекомбинантных ДНК
Для манипуляций с ДНК были использованы стандартные способы, описанные в Sambrook, J. et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989). Молекулярно-биологические реагенты использовали в соответствии с инструкциями изготовителей.
Синтез генов
Необходимые сегменты генов были заказаны в соответствии с заданными требованиями у Geneart (Регенсбург, Германия).
Определение последовательности ДНК
Последовательности ДНК были определены путем секвенирования двухцепочечной ДНК, выполненного в MediGenomix GmbH (Мартиншрид, Германия) или SequiServe GmbH (Фатерштеттен, Германия).
Анализ последовательности ДНК и белка и управление данными о последовательности
Пакет программного обеспечения от GCG (Genetics Computer Group, Мэдисон, Висконсин) версии 10.2 и Vector NT1 Advance предварительной версии 8.0 от Infomax использовали для создания, картирования, анализа, пояснения и иллюстрации последовательности.
Экспрессионные векторы
Для экспрессии описанных антител использовали экспрессионные плазмиды для временной экспрессии (например, в клетках HEK293-F) на основании кДНК-организации с промотором интрона А из CMV или без него, либо на основании геномной организации с промотором из CMV.
Транскрипционная единица гена антитела состоит из следующих элементов:
- уникального сайта (сайтов) рестрикции на 5-конце,
- немедленного раннего энхансера и промотора цитомегаловируса человека,
- последовательности интрона А в случае кДНК-организации,
5- '-нетранслируемой области человеческого гена иммуноглобулина,
- нуклеиновой кислоты, кодирующей сигнальную последовательность тяжелой цепи иммуноглобулина,
- нуклеиновой кислоты, кодирующей цепь человеческого антитела (дикого типа или с обменом доменов) либо в виде кДНК, либо в геномной организации с экзон-интронной организацией иммуноглобулина,
- 3'-нетранслируемой области с сигнальной последовательностью полиаденилирования,и
- уникального сайта (сайтов) рестрикции на 3'-конце.
Кроме кассеты для экспрессии антитела данные плазмиды содержали:
- сайт начала репликации, который обеспечивает репликацию этой плазмиды в Е. coli,
- ген бета-лактамазы, который придает устойчивость Е. coli к ампициллину, и
- ген дигидрофолатредуктазы от Mus musculus в качестве селективного маркера в эукариотических клетках.
Нуклеиновые кислоты, кодирующие цепи антитела, были получены с помощью ПЦР и/или генного синтеза и собраны известными рекомбинантными способами и методиками соединения соответствующих нуклеиновокислотных сегментов, например, с помощью уникальных сайтов рестрикции в соответствующих векторах. Субклонированные нуклеиновокислотные последовательности были проверены путем секвенирования ДНК. Для временных трансфекций большие количества плазмид получали путем создания культур Е. coli, трансформированных плазмидой (Nucleobond АХ, Macherey-Nagel).
Методики клеточного культивирования
Стандартные методики клеточного культивирования использовали в соответствии с описанием в Current Protocols in Cell Biology (2000), Bonifacino, J.S., Dasso, M., Harford, J.В., Lippincott-Schwartz, J. and Yamada, K.M. (eds.), John Wiley & Sons, Inc.
Биспецифические антитела экспрессировали путем временной котрансфекции соответствующими экспрессионными плазмидами клеток HEK293-F, растущих в суспензии, как описано ниже.
Пример 1
Экспрессия и очистка
Временные трансфекций в системах HEK293-F
Моноспецифические и биспецифические антитела получали путем временной трансфекций соответствующими плазмидами (например, кодирующими тяжелую и модифицированную тяжелую цепи, а также соответствующую легкую и модифицированную легкую цепи) с помощью системы HEK293-F (Invitrogen) в соответствии с инструкцией производителя. Вкратце, клетки HEK293-F, растущие в суспензии во встряхиваемой колбе или в перемешивающем ферментере в бессывороточной среде для экспрессии Freestyle™ 293 (Invitrogen), трансфицировали смесью соответствующих плазмид и 293 fectin™ или fectin (Invitrogen). В 2 л встряхиваемой колбе (Corning) клетки HEK293-F высевали с плотностью 1*106 клеток/мл в 600 мл и инкубировали на скорости 120 об/мин, 8% СО2. На следующий день клетки трансфицировали при плотности клеток примерно 1,5*106 клеток/мл с использованием примерно 42 мл смеси А) 20 мл Opti-MEM (Invitrogen) с 600 мкг тотальной плазмидной ДНК (1 мкг/мл), кодирующей тяжелую или модифицированную тяжелую цепь, соответственно, и соответствующую легкую цепь в эквимолярном соотношении и В) 20 мл Opti-MEM с 1,2 мл 293 fectin или fectin (2 мкл/мл). Согласно потреблению глюкозы в ходе ферментации добавляли раствор глюкозы. Супернатант, содержащий секретированное антитело, собирали через 5-10 дней, и антитела либо непосредственно очищали из супернатанта, либо супернатант замораживали и хранили.
Очистка
Биспецифические антитела очищали из клеточных культуральных супернатантов с помощью аффинной хроматографии с использованием белка MabSelectSure-Sepharose™ (для не-IHH-ААА-мутантов) (GE Healthcare, Швеция) и KappaSelect-Agarose (для IHH-AAA-мутантов) (GE Healthcare, Швеция), хроматографии гидрофобных взаимодействий с использованием бутил-сефарозы (GE Healthcare, Швеция) и эксклюзионной хроматографии на Superdex200 (GE Healthcare, Швеция).
Вкратце, стерильные профильтрованные клеточные культуральные супернатанты очищали на смоле MabSelectSuRe, уравновешенной (не-IHH-ААА-мутанты и антитела дикого типа) буфером PBS (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, рН 7,4), промывали уравновешивающим буфером и элюировали 25 мМ раствором цитрата натрия при рН 3,0. Мутанты IHH-AAA захватывали на смоле KappaSelect, уравновешенной 25 мМ Tris, 50 мМ NaCl, рН 7,2, промывали уравновешивающим буфером и элюировали 25 мМ раствором цитрата натрия с рН 2,9. Элюированные фракции антител объединяли и нейтрализовали с помощью 2 M Tris, рН 9,0. Объединенные антитела готовили для хроматографии гидрофобных взаимодействий, добавляя 1,6 М раствор сульфата аммония до конечной концентрации 0,8 М сульфата аммония и доводили рН до 5,0 с помощью уксусной кислоты. После уравновешивания бутил-сефарозой смолы с помощью 35 мМ ацетата натрия, 0,8 М сульфата аммония, рН 5,0, антитела наносили на смолу, промывали уравновешивающим буфером и элюировали линейным градиентом до 35 мМ ацетата натрия, рН 5,0. Фракции, содержащие антитела (моноспецифические или биспецифические), объединяли и далее очищали с помощью эксклюзионной хроматографии с использованием колонки Superdex 200 26/60 GL (GE Healthcare, Швеция), уравновешенной 20 мМ гистидином, 140 мМ NaCl, рН 6,0. Фракции, содержащие антитела (моноспецифические или биспецифические), объединяли, концентрировали до необходимой концентрации с использованием устройств для ультрафильтрации Vivaspin (Sartorius Stedim Biotech S.A., Франция) и хранили при -80°C.

Чистоту и целостность антитела анализировали после каждого этапа очистки с использованием CE-SDS по микрожидкостной методике Labchip (Caliper Life Science, США). 5 мкл раствора белка готовили для анализа CE-SDS с помощью набора реагентов НТ Protein Express согласно инструкции производителя и анализировали на системе LabChip GXII с помощью НТ Protein Express Chip.Данные анализировали с помощью программного обеспечения LabChip GX.
Таблица: Удаление типичных побочных продуктов различными последовательными этапами очистки, определяемое по CE-SDS
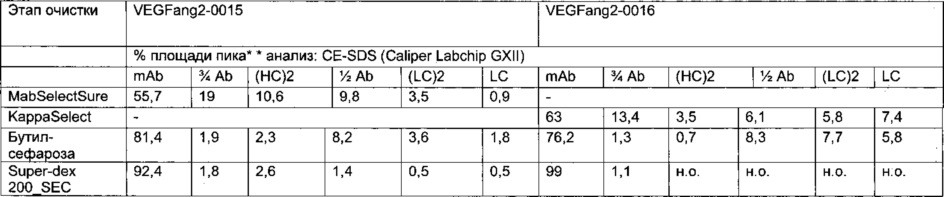
Содержание агрегатов в образцах антител анализировали с помощью высокоэффективной SEC с использованием аналитической эксклюзионной колонки Superdex 200 (GE Healthcare, Швеция) в рабочем буфере 2×PBS (20 мМ Na2HPO4, 2 мМ KH2PO4, 274 мМ NaCl и 5,4 мМ KCl, рН 7,4) при 25°C. 25 мкг белка вводили в колонку со скоростью потока 0,75 мл/мин и элюировали изократически в течение 50 минут.
Аналогичным образом получали и очищали биспецифические <VEGF-ANG-2> антитела VEGFang2-0012 и VEGFang2-0201, получая следующие выходы:

Кроме того, биспецифические <VEGF-ANG-2> антитела <VEGF-ANG-2> CrossMAb IgG4 с мутацией IHH-AAA и с мутацией SPLE (SEQ ID №42, SEQ ID №43, SEQ ID №44, SEQ ID №45), <VEGF-ANG-2> OAscFab IgG1 с мутацией IHH-AAA (SEQ ID №46, SEQ ID №47, SEQ ID №48), <VEGF-ANG-2> OAscFab IgG4 с мутацией IHH-AAA и с мутацией SPLE (SEQ ID №49, SEQ ID №50, SEQ ID №51), <VEGF-ANG-2> CrossMab IgG1 с мутацией HHY-AAA и мутацией P329G LALA (SEQ ID №90, SEQ ID №91, SEQ ID №40, SEQ ID №41), <VEGF-ANG-2>Crosstab IgG4 с мутацией с HHY-AAA и мутацией SPLE (SEQ ID №92, SEQ ID №93, SEQ ID №44, SEQ ID №45), <VEGF-ANG-2> OAscFab IgG1 с мутацией HHY-AAA (SEQ ID №94, SEQ ID №95, SEQ ID №48) и <VEGF-ANG-2> OAscFab IgG4 с мутацией HHY-AAA и мутацией SPLE (SEQ ID №96, SEQ ID №97, SEQ ID №51) могут быть получены и очищены аналогичным образом.
Пример 2
Аналитика и проявление
Измерение вязкости на основе DLS малого масштаба Измерение вязкости по существу проводили так, как описано в Не, F. et al., Analytical Biochemistry 399 (2009) 141-143. Вкратце, образцы концентрировали до различных концентраций белка в 200 мМ сукцинате аргинина, рН 5,5, а затем добавляли полистироловые латексные шарики (диаметром 300 нм) и полисорбат 20 (0,02% объем/объем). Образцы переносили в 384-луночные планшеты с оптическим дном путем центрифугирования через фильтровальный планшет с размером пор 0,4 мкм и покрывали парафиновым маслом. Кажущийся диаметр латексных шариков определяли методом динамического светорассеяния при 25°C. Вязкость раствора можно рассчитать как η=η0(rh/rh,0) (η: вязкость; η0: вязкость воды; rh: кажущийся гидродинамический радиус латексных шариков; rh,0: гидродинамический радиус латексных шариков в воде).
Чтобы можно было сравнивать различные образцы при одинаковой концентрации, данные "вязкость-концентрация" подгоняли под уравнение Муни (уравнение 1) (Mooney, M., Colloid. Sci., 6 (1951) 162-170; Monkos, К., Biochem. Biophys. Acta 304 (1997) 1339) и соответствующим образом интерполировали данные. 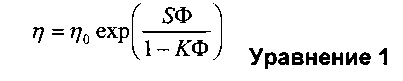
(S: параметр гидродинамического взаимодействия белка; К: фактор самовытеснения; Ф: объемная доля растворенного белка)
Результаты показаны на фиг. 2: VEGFang2-0016 с мутацией IHH-AAA в Fc-области демонстрирует более низкую вязкость при всех измеренных температурах по сравнению с VEGFang2-0015 без мутации IHH-AAA в Fc-области.
Температура начала агрегации DLS
Образцы готовили в концентрации 1 мг/мл в 20 мМ гистидине/гидрохлориде гистидина, 140 мМ NaCl, рН 6,0, переносили в 384-луночные планшеты с оптическим дном путем центрифугирования через фильтровальный планшет с размером пор 0,4 мкм и покрывали парафиновым маслом. Гидродинамический радиус повторно измеряли методом динамического светорассеяния, в то время как образцы нагревали со скоростью 0,05°С/мин от 25°С до 80°С. Температуру начала агрегации определяли как температуру, при которой гидродинамический радиус начинал возрастать. Результаты показаны на фиг. 3. На фиг. 3 показана агрегация VEGFang2-0015 без мутации IHH-AAA по сравнению с VEGFang2-0016 с мутацией IHH-AAA в Fc-области. VEGFang2-0016 демонстрировал температуру начала агрегации 61°C, тогда как VEGFang2-0015 без мутации IHH-AAA демонстрировал температуру начала агрегации 60°C.
DLS с течением времени
Образцы готовили в концентрации 1 мг/мл в 20 мМ гистидине/гидрохлориде гистидина, 140 мМ NaCl, рН 6,0, переносили в 384-луночные планшеты с оптическим дном путем центрифугирования через фильтровальный планшет с размером пор 0,4 мкм и покрывали парафиновым маслом. Гидродинамический радиус повторно измеряли методом динамического светорассеяния, а образцы инкубировали при постоянной температуре 50°C в течение периода до 145 часов. В этом эксперименте тенденция к агрегации нативного, нефолдированного белка при повышенной температуре может привести к увеличению среднего диаметра частиц с течением времени. Этот метод на основе DLS очень чувствителен к агрегатам, так как это способствует большей пропорциональности интенсивности рассеянного света. Даже после 145 часов при 50°C (температура, близкая к температуре начала агрегации, см. выше) средний диаметр частиц увеличивался менее чем на 0,5 нм как для VEGFang2-0015, так и для VEGFang2-0016.
Семидневное хранение при 40°С в дозе 100 мг/мл
Образцы концентрировали до конечной концентрации 100 мг/мл в 200 мМ сукцинате аргинина, рН 5,5, стерильно фильтровали и хранили в покое при 40°C в течение семи дней. До и после хранения определяли содержание высоко- и низкомолекулярных видов (HMW и LMW, соответственно) путем эксклюзионной хроматографии. Разница в содержании HMW и LMW между хранимым образцом и образцом, измеренным сразу после приготовления, сообщается как "увеличение HMW" и "увеличение LMW", соответственно. Результаты, приведенные в расположенной далее таблице и на фиг.4, показывают, что VEGFang2-0015 (без мутации IHH-AAA) демонстрирует более сильное снижение основного пика и более сильный рост HMW по сравнению с VEGFang2-0016 (с мутацией IHH-AAA). Неожиданно, VEGFang2-0016 (с мутацией IHH-AAA) демонстрирует более низкую тенденцию к агрегации по сравнению с VEGFang2-0015 (без мутации IHH-AAA).
Таблица: Изменение главного пика, HMW- и LMW-пиков через семь дней при 40°C
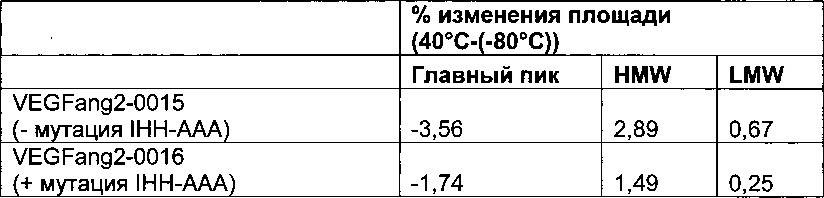
Функциональный анализ биспецифических <VEGF-ANG-2> антител проводили путем анализа поверхностного плазмонного резонанса (SPR) с помощью инструмента BIAcore® Т100 или Т200 (GE Healthcare) при 25°C. Система BIAcore® хорошо известна для изучения молекулярных взаимодействий. SPR-технология основана на измерении показателя преломления вблизи поверхности покрытого золотом биосенсорного чипа. Изменения показателя преломления отражают изменения массы на поверхности, вызванные взаимодействием иммобилизованного лиганда с анализируемым веществом, введенным в растворе. Если молекулы связывают иммобилизованные лиганды на поверхности, то масса увеличивается, и наоборот, в случае диссоциации анализируемого вещества с иммобилизованного лиганда масса уменьшается (отражая диссоциацию комплекса). SPR позволяет осуществлять в реальном времени непрерывный мониторинг связываний лиганда/анализируемого вещества и, следовательно, определять константу скорости ассоциации (ка), константу скорости диссоциации (kd) и константу равновесия (KD).
Пример 3
Связывание с VEGF, ANG-2, Fc-raiviMaR и FcRn
Кинетическая аффинность изоформ VEGF, включающая оценку межвидовой перекрестной реактивности
Примерно 12000 единиц ответа (RU) системы захвата (10 мкг/мл козлиного противочеловеческого F(Ab)'2; код заказа: 28958325; GE Healthcare Bio-Sciences АВ, Швеция) связывали с чипом CM5 (GE Healthcare BR-1005-30) при рН 5,0 с помощью набора для аминного связывания, поставляемого GE Healthcare. Буфер для образцов и системный буфер представлял собой PBS-T (10 мМ забуференный фосфатом физиологический раствор с 0,05% Tween-20TM), рН 7,4. Температуру проточной ячейки устанавливали на 25°C, а температуру отсека для образца на 12°C, и вначале дважды вводили рабочий буфер. Биспецифическое антитело было захвачено при введении 50 нМ раствора в течение 30 секунд при скорости потока 5 мкл/мин. Ассоциацию измеряли при введении человеческого hVEGF121, мышиного mVEGF120 или крысиного rVEGF164 в различных концентрациях в растворе в течение 300 секунд со скоростью потока 30 мкл/мин, начиная с 300 нМ в разведениях 1:3. Фазу диссоциации наблюдали в течение 1200 секунд и запускали при переключении с раствора образца на рабочий буфер. Поверхность подвергали регенерации путем промывания в течение 60 секунд раствором глицина с рН 2,1 со скоростью потока 30 мкл/мин. Различия в объемных показателях преломления корректировали путем вычитания ответа, полученного с поверхности с козлиным противочеловеческим F(Ab)'2. Пустые (контрольные) введения также вычитали (= двойной референс). Для расчета кажущейся KD и других кинетических параметров использовали модель Ленгмюра 1:1. Результаты показаны ниже.
Аффинность раствора ANG-2, включая оценку межвидовой перекрестной реактивности
Аффинность раствора измеряли как аффинность взаимодействия путем определения концентрации свободных партнеров взаимодействия в равновесной смеси. Анализ аффинности раствора включает смешивание биспецифического <VEGF-ANG-2> антитела, с сохранением его в постоянной концентрации, с лигандом (=ANG-2) в различных концентрациях. Максимально возможное число единиц ответа (например, 17000 единиц ответа (RU)) антитела иммобилизовали на поверхности чипа CM5 (GE Healthcare BR-1005-30) при рН 5,0 с использованием набора для связывания амина, поставляемого компанией GE Healthcare. Буфер для образцов и системный буфер представлял собой HBS-P с рН 7,4. Температуру проточной ячейки устанавливали на 25°C, а температуру отсека для образца на 12°C, и вначале дважды вводили рабочий буфер. Для создания калибровочной кривой в проточную камеру BIAcore, содержащую иммобилизованное биспецифическое <VEGF-ANG-2> антитело, вводили возрастающие концентрации ANG-2. Количество связанного ANG-2 определяли как единицы ответа (RU) и наносили на график в зависимости от концентрации. Растворы каждого лиганда (11 концентраций от 0 до 200 нМ для биспецифического <VEGF-ANG-2> антитела) инкубировали с 10 нМ ANG-2 и дожидались равновесия при комнатной температуре. Концентрации свободного ANG-2 определяли из калибровочной кривой, созданной до и после измерения ответа растворов с известными количествами ANG-2. 4-параметрическую подгонку осуществляли с XLfit4 (IDBS Software) с использованием модели 201, используя концентрацию свободного ANG-2 как у-ось и используемые концентрации антитела для ингибирования как х-ось. Аффинность рассчитывали путем определения точки перегиба этой кривой.
Поверхность подвергали регенерации путем однократного промывания в течение 30 секунд 0,85% раствором Н3РО4 со скоростью потока 30 мкл/мин. Различия в объемных показателях преломления корректировали путем вычитания ответа, полученного с поверхности, связанной с отрицательным (пустым) контролем. Результаты приведены ниже.
Состояние устойчивой аффинности к FcRn
Для измерения FcRn использовали состояние устойчивой аффинности, чтобы сравнить биспецифические антитела друг с другом. Человеческий FcRn разводили в буфере для связывания (10 мкг/мл, Na-ацетат, рН 5,0) и иммобилизовали на чипе C1-Chip (GE Healthcare BR-1005-35) согласно процедуре ориентированной иммобилизации с использованием программы BIAcore до конечного ответа 200 RU. Температуру проточной ячейки устанавливали на 25°С, а температуру отсека для образца на 12°С, и вначале дважды вводили рабочий буфер. Буфер для образцов и системный буфер представлял собой HBS-P (10 мМ забуференный фосфатом физиологический раствор с 0,05% Tween-20TM) с рН 6,0. Для оценки различных концентраций IgG для каждого антитела готовили концентрации 62,5 нМ, 125 нМ, 250 нм и 500 нМ. Скорость потока устанавливали на 30 мкл/мин и на поверхность чипа последовательно вводили различные образцы, выбирая время ассоциации 180 секунд. Поверхность подвергали регенерации путем введения PBS-T с рН 8 в течение 60 секунд со скоростью потока 30 мкл/мин. Различия в объемных показателях преломления корректировали путем вычитания ответа, полученного с поверхности, связанной с отрицательным (пустым) контролем. Введения буфера также вычитали (= двойной референс). Для расчета состояния устойчивой аффинности использовали способ из программного обеспечения BIA-Evaluation. Вкратце, значения RU наносили на график против анализируемых концентраций, получая кривую "доза-ответ". На основании 2-параметрического соответствия рассчитывали верхнюю асимптоту, что позволяло определять полумаксимальное значение RU и, следовательно, аффинность. Результаты показаны на фиг.5 и ниже в таблице. Аналогичным образом может быть определена аффинность к FcRn Cynomolgus, мышей и кроликов.
Измерение Fc-raMMa-RIIIa
Для измерения аффинности Fc-raMMa-RIIIa использовали прямой анализ связывания. Примерно 3000 единиц ответа (RU) системы захвата (1 мкг/мл Penta-His; Qiagen) связывали с чипом СМ5 (GE Healthcare BR-1005-30) при рН 5,0 с помощью набора для аминного связывания, поставляемого GE Healthcare. Буфер для образцов и системный буфер представлял собой HBS-P+ с рН 7,4. Температуру проточной ячейки устанавливали на 25°C, а температуру отсека для образца на 12°C, и вначале дважды вводили рабочий буфер. Fc-гамма-RIIIa-His-рецептор были захвачен при введении 100 нМ раствора в течение 60 секунд при скорости потока 5 мкл/мин. Связывание измеряли при введении 100 нМ биспецифического антитела или моноспецифических контрольных антител (антидигоксигениновые антитела (анти-Dig) для антител подкласса IgG1 и подкласса IgG4) в течение 180 секунд со скоростью потока 30 мкл/мин. Поверхность подвергали регенерации путем промывания в течение 120 секунд раствором глицина с рН 2,5 со скоростью потока 30 мкл/мин. Поскольку связывание Fc-гамма-PIIIa отличается от модели Ленгмюра 1:1, в этом анализе оценивали только наличие/отсутствие связывания. Аналогичным образом может быть определено связывание Fc-гамма-RIa и Fc-гамма-RIIa. Результаты показаны на фиг. 6, из которых следует, что при введении мутаций P329G LALA связывание с Fc-гамма-RIIIa не может быть обнаружено.
Оценка независимого VEGF- и ANG-2-связывания с биспецифическими <VEGF-ANG-2> антителами
Примерно 3500 единиц ответа (RU) системы захвата (10 мкг/мл козлиного противочеловеческого IgG; GE Healthcare Bio-Sciences AB, Швеция) связывали на чипе CM4 (GE Healthcare BR-1005-34) при рН 5,0 с помощью набора для аминного связывания, поставляемого GE Healthcare. Буфер для образцов и системный буфер представлял собой PBS-T (10 мМ забуференный фосфатом физиологический раствор с 0,05% Tween-20TM) с рН 7,4. Температуру проточной ячейки устанавливали на 25°С, а температуру отсека для образца на 12°С. Перед захватом в проточную ячейку дважды вводили рабочий буфер.
Биспецифическое антитело были захвачено при введении 10 нМ раствора в течение 60 секунд со скоростью потока 5 мкл/мин. Независимое связывание каждого лиганда с биспецифическим антителом анализировали, определяя активную связывающую способность для каждого лиганда, добавляя последовательно или одновременно (скорость потока 30 мкл/мин):
1. Введение человеческого VEGF с концентрацией 200 нМ в течение 180 секунд (идентифицирует одиночное связывание антигена).
2. Введение человеческого ANG-2 с концентрацией 100 нМ в течение 180 секунд (идентифицирует одиночное связывание антигена).
3. Введение человеческого VEGF с концентрацией 200 нМ в течение 180 секунд с последующим дополнительным введением человеческого ANG-2 с концентрацией 100 нМ в течение 180 секунд (идентифицирует связывание ANG-2 в присутствии VEGF).
4. Введение человеческого ANG-2 с концентрацией 100 нМ в течение 180 секунд с последующим дополнительным введением человеческого VEGF с концентрацией 200 нМ (идентифицирует связывания VEGF в присутствии ANG-2).
5. Совместное введение человеческого VEGF с концентрацией 200 нМ и человеческого ANG-2 с концентрацией 100 нМ в течение 180 секунд (идентифицирует связывание VEGF и ANG-2 в одно и то же время).
Поверхность подвергали регенерации путем промывания 3 M раствором MgCl2 в течение 60 секунд со скоростью потока 30 мкл/мин. Различия в объемных показателях преломления корректировали путем вычитания ответа, полученного с поверхности, связанной с козлиным противочеловеческим IgG.
Биспецифическое антитело было способно связываться с обоими антигенами взаимонезависимо, если полученный конечный сигнал из подходов 3, 4 и 5 был равен или близок сумме отдельных конечных сигналов из подходов 1 и 2. Результаты приведены в таблице ниже, где показано, что оба антитела, VEGFang2-0016 и VEGFang2-0012, способны связываться взаимонезависимо с VEGF и ANG-2.
Оценка одновременного VEGF- и ANG-2-связывания с биспецифическими <VEGF-ANG-2> антителами
Во-первых, примерно 1600 единиц ответа (RU) системы захвата VEGF (20 мкг/мл) связывали с чипом CM4 (GE Healthcare BR-1005-34) при рН 5,0 с помощью набора для аминного связывания, поставляемого GE Healthcare. Буфер для образцов и системный буфер представлял собой PBS-T (10 мМ забуференный фосфатом физиологический раствор с 0,05% Tween-20TM) с рН 7,4. Температуру проточной ячейки устанавливали на 25°C, а температуру отсека для образца на 12°C, и вначале в проточную ячейку дважды вводили рабочий буфер. Во-вторых, 50 нМ раствор биспецифического антитела вводили в течение 180 секунд со скоростью потока 30 мкл/мин. В-третьих, hANG-2 вводили в течение 180 секунд со скоростью потока 30 мкл/мин. Ответ связывания hANG-2 зависел от количества биспецифического антитела, связанного с VEGF, и показывал одновременное связывание. Поверхность подвергали регенерации путем промывания 0,85% раствором Н3РО4 в течение 60 секунд со скоростью потока 30 мкл/мин.
Одновременное связывание показано дополнительным сигналом специфического связывания hANG-2 с биспецифическими <VEGF-ANG-2> антителами, ранее связанными с VEGF. Для обоих биспецифических антител VEGFang2-0015 и VEGFang2-0016 может быть обнаружено одновременное VEGF- и ANG-2-связывание с биспецифическими <VEGF-ANG-2> антителами (данные не показаны).
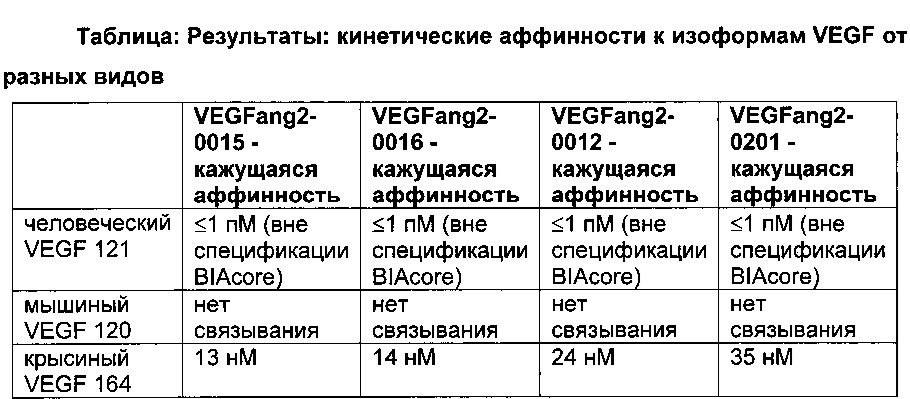
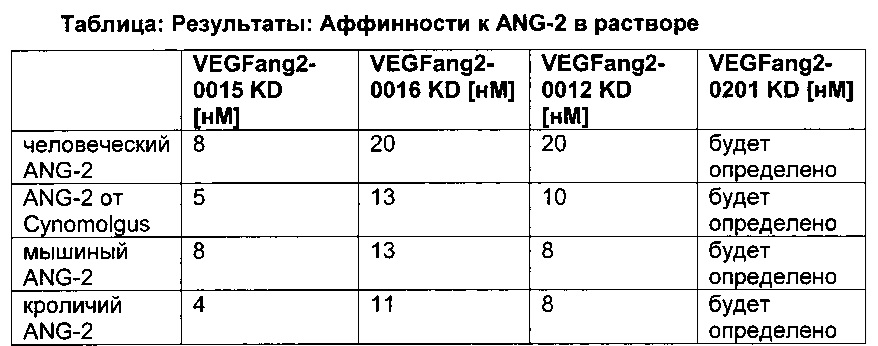



Пример 4
Масс-спектрометрия
В этом разделе описывается характеризация биспецифических <VEGF-ANG-2> антител с акцентом на правильную сборку. Ожидаемые первичные структуры были подтверждены путем масс-спектрометрии с ионизацией электрораспылением (ESI-MS) дегликозилированных и интактных или IdeS-расщепленных (IgG-расщепляющим ферментом от S. pyogenes) биспецифических <VEGF-ANG-2> антител. IdeS-расщепление проводили с 100 мкг очищенного антитела, инкубированного с 2 мкг ldeS-протеазы (Fabricator) в 100 ммоль/л NaH2PO4/Na2HPO4, рН 7,1 при 37°C в течение 5 ч. Затем антитела дегликозилировали с помощью N-гликозидазы F, нейраминидазы и О-гликозидазы (Roche) в 100 ммоль/л NaH2PO4/Na2HPO4, рН 7,1, при 37°С в течение 16 часов при концентрации белка 1 мг/мл, и затем обессоливали с помощью HPLC на колонке Sephadex G25 (GE Healthcare). Общую массу определяли с помощью ESI-MS в системе maXis 4G UHR-QTOF MS (Bruker Daltonik), снабженной источником TriVersa NanoMate (Advion).
Массы, полученные для IdeS-расщепленных, дегликозилированных (в таблице ниже), или интактных, дегликозилированных (в таблице ниже) молекул соответствуют предсказанным массам, выведенным из аминокислотных последовательностей для биспецифических <VEGF-ANG-2> антител, состоящих из двух различных легких цепей LCANG-2 и LCLucentis и двух различных тяжелых цепей HCANG-2 И HCLucentis.
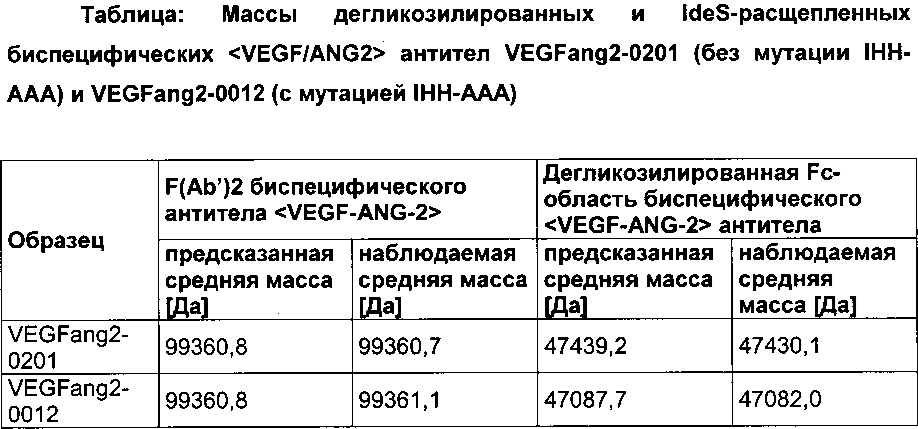
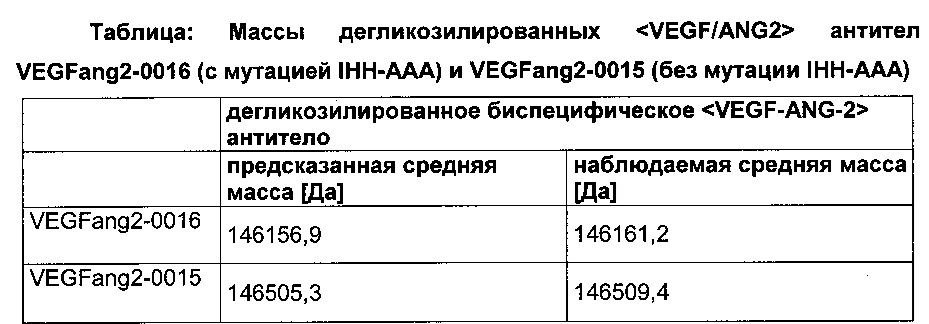
Пример 5
FcRn-хроматография
Соединение со стрептавидин-сефарозой:
Один грамм стрептавидин-сефарозы (GE Healthcare) добавляли к биотинилированному и диализованному рецептору и инкубировали в течение двух часов при встряхивании. Рецепторное производное сефарозы помещали в 1 мл колонку ХК (GE Healthcare).
Хроматография с использованием FcRn-аффинной колонки:
Условия:
|
Хроматография на колонке с аффинностью к человеческому FcRn
В следующий таблице приведено время удерживания биспецифических <VEGF-ANG-2> антител на аффинных колонках, содержащих человеческий FcRn.
Данные получены при условиях, описанных выше.
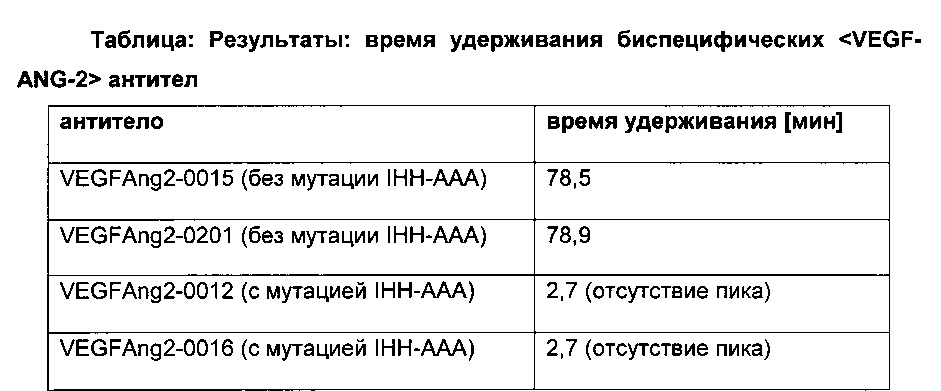
Пример 6
Фармакокинетические (PK) свойства антител с мутацией IHH-AAA
Данные PK на FcRn мышах, трансгенных по человеческому FcRn
Прижизненная фаза:
В исследование были включены самки мышей C57BL/6J (фон); мыши, дефицитные по FcRn, но гемизиготно трансгенные по человеческому FcRn (huFcRn, линия 276 -/tg)
Часть 1:
Всем мышам вводили однократно интравитреально в правый глаз 2 мкл/животное соответствующего раствора (т.е. 21 мкг соединения/животное VEGFAng2-0015 (без мутации IHH-AAA)) или 23,6 мкг соединения/животное VEGFAng2-0016 (с мутацией IHH-AAA)).
Мышей делили на 2 группы по 6 животных в каждой. Образцы крови брали в группе 1 через 2, 24 и 96 часов, а в группе 2 через 7, 48 и 168 часов после введения дозы.
Инъекцию в стекловидное тело правого глаза мыши выполняли с использованием системы NanoFil Microsyringe для нанолитровых инъекций от World Precision Instruments, Inc., Берлин, Германия. Мышей анестезировали с помощью 2,5% изофлурана и для визуализации глаза мыши использовали микроскоп Leica MZFL 3 с 40-кратным увеличением и кольцевым осветителем Leica KL 2500 LCD. Затем вводили 2 мкл соединения с помощью иглы размера 35.
У каждого животного собирали кровь из ретробульбарного венозного сплетения контралатерального глаза для определения уровней соединений в сыворотке крови.
После инкубации при комнатной температуре в течение 1 часа из крови получали образцы сыворотки объемом по меньшей мере 50 мкл путем центрифугирования (9300 д) при 4°C в течение 3 мин. Образцы сыворотки замораживали сразу после центрифугирования и хранили в замороженном виде при -80°C до анализа. Глаза, в которые вводили соединение, у животных из группы 1 выделяли через 96 часов после введения, а у животных из группы 2 через 168 ч после введения. Образцы хранили в замороженном виде при -80°C до анализа.
Часть 2:
Всем мышам однократно внутривенно (через хвостовую вену) вводили 200 мкл/животное соответствующего раствора (т.е. 21 мкг соединения/животное (VEGFAng2-0015 (без мутации IHH-AAA)) или 23,6 мкг соединения/животное (VEGFAng2-0016 (с мутацией IHH-AAA)).
Мышей делили на 2 группы по 5 животных в каждой. Образцы крови брали в группе 1 через 1, 24 и 96 часов, а в группе 2 через 2, 48 и 168 часов после введения дозы. У каждого животного собирали кровь из ретробульбарного венозного сплетения для определения уровней соединения в сыворотке крови.
После инкубации при комнатной температуре в течение 1 часа из крови получали образцы сыворотки объемом по меньшей мере 50 мкл путем центрифугирования (9300 д) при 4°C в течение 3 мин. Образцы сыворотки замораживали сразу после центрифугирования и хранили в замороженном виде при -80°C до анализа.
Получение цельных лизатов глаз (мышиных)
Глазные лизаты получали путем физико-химического разрушения цельных глаз лабораторных животных. Для механического разрушения каждый глаз переносили в микропробирку объемом 1,5 мл с коническим дном. После замораживания и размораживания глаза промывали один раз 1 мл промывочного буфера (Bio-Rad, Bio-Plex Cell Lysis Kit, кат.№171-304011). На следующем этапе добавляли 500 мкл свежеприготовленного буфера для лизиса клеток и измельчали глаза с помощью пестика для измельчения тканей в микропробирках объемом 1,5 мл (Kimble Chase, 1.5 mL pestle, арт. №749521-1500). Затем смесь замораживали и размораживали пять раз и снова измельчали. Чтобы отделить лизат от остальных тканей, образцы центрифугировали в течение 4 мин при 4500 д. После центрифугирования супернатант собирали и хранили при -20°C до дальнейшего анализа путем количественной ELISA.
Анализ
Концентрации <VEGF-ANG-2> антител в сыворотках и в глазных лизатах мышей определяли путем твердофазного иммуноферментного анализа (ELISA).
Для количественной оценки <VEGF-ANG-2> антител в образцах сыворотки и глазных лизатах мышей проводили стандартный твердофазный серийный иммуноферментный анализ типа "сэндвич" с биотинилированными и дигоксигенилированными моноклональными антителами, используемыми в качестве антител захвата и обнаружения. При проверке сохранности биспецифичности анализируемого антитела биотинилированное антитело захвата распознает сайт связывания VEGF, тогда как дигоксигенилированное антитело обнаружения связывается с сайтом связывания ANG-2 анализируемого антитела. Затем проводят обнаружение связанного иммунного комплекса антитела захвата, анализируемого антитела и антитела обнаружения на твердой фазе микротитровального планшета, покрытого стрептавидином (SA-MTP), с помощью пероксидазы хрена, соединенной с антидигоксигениновым антителом. После отмывания несвязанного материала из SA-MTP и добавления ABTS-субстрата полученный сигнал пропорционален количеству анализируемого антитела, связанного с твердой фазой SA-MTP. Затем проводят количественную оценку путем преобразования измеренных сигналов от образцов в концентрации согласно стандартам (калибраторам), анализируемым параллельно.
На первом этапе в SA-MTP вносили раствор биотинилированного антитела захвата (антиидиотипическое антитело mAb<Id<VEGF>>M-2.45.51-IgG-Bi(DDS)) в объеме 100 мкл/лунка с концентрацией 1 мкг/мл на один час на MTP-шейкере со скоростью 500 об/мин. В это время готовили стандарты, QC-образцы (образцы контроля качества) и анализируемые образцы. Стандарты и ОС-образцы разводили до 2% сывороточной матрицы; анализируемые образцы разводили таким образом, чтобы сигналы находились в пределах линейного диапазона стандартов.
После нанесения на SA-MTP антитела захвата планшет промывали три раза промывочным буфером в объеме 300 мкл/лунка. Затем в SA-MTP пипеткой вносили по 100 мкл/лунка стандартов, QC-образцов и анализируемых образцов и снова инкубировали в течение одного часа при 500 об/мин. Теперь анализируемое антитело прикреплялось его анти-VEGF-сайтом связывания к твердой фазе SA-MTP через антитело захвата. После инкубации и удаления несвязанного анализируемого антитела путем промывания планшета в SA-MTP добавляли 100 мкл/лунка первого антитела обнаружения (антиидиотипическое антитело mAb<Id-<ANG-2>>M-2.6.81-IgG-Dig(XOSu)) в концентрации 250 нг/мл. Планшет снова инкубировали в течение одного часа на шейкере при 500 об/мин. После промывания в лунки SA-MTP вносили по 100 мкл/лунка второго антитела обнаружения (pAb<Digoxigenin>S-Fab-POD (poly)) в концентрации 50 мЕд/мл, и планшет снова инкубировали в течение одного часа при 500 об/мин. После финального этапа промывания, чтобы удалить избыток антитела обнаружения, вносили по 100 мкл/лунка субстрата (ABTS). Конъюгат "антитело-фермент" катализировал цветовую реакцию субстрата ABTS®. Затем сигнал измеряли с помощью ELISA-ридера на длине волны 405 нм (референсная длина волны: 490 нм ([405/490] нм)).
Фармакокинетическая оценка
Фармакокинетические параметры рассчитывали путем некомпартментного анализа, используя программу для фармакокинетической оценки WinNonlinTM (Pharsight) версии 5.2.1.
Результаты:
А) Концентрации в сыворотке
Результаты для концентраций в сыворотке крови приведены в следующих таблицах и на фиг. 7В-7С.
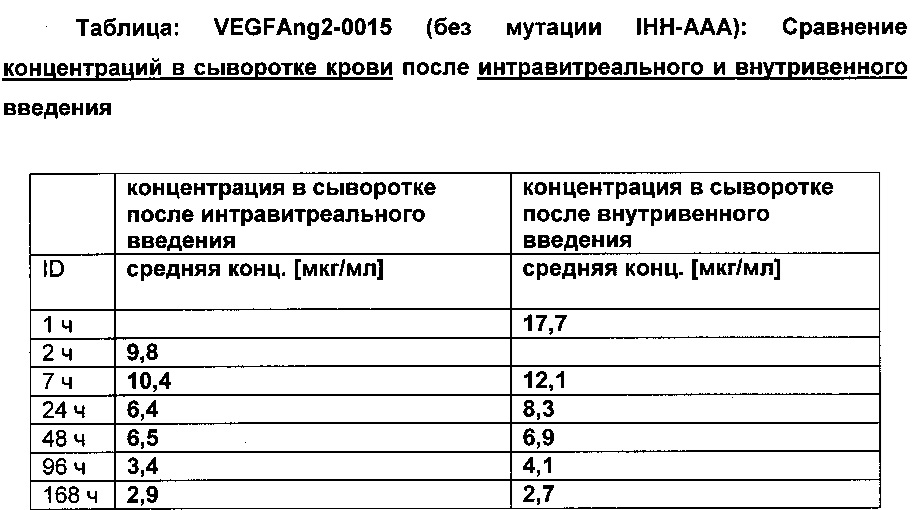
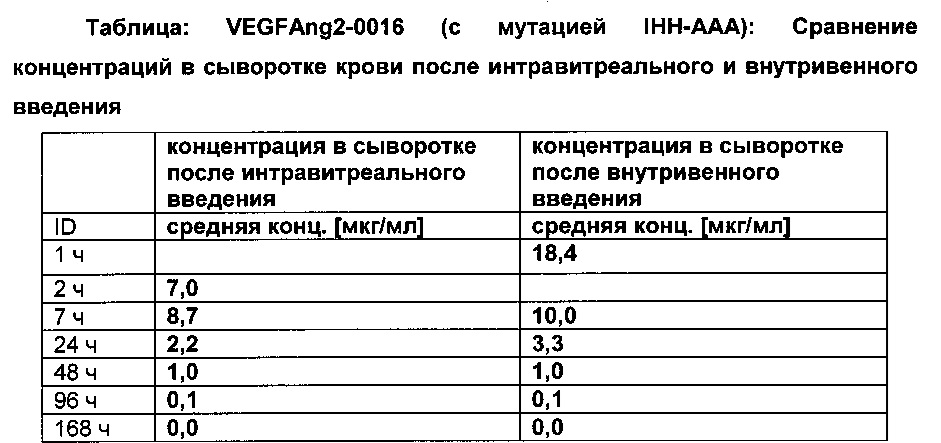
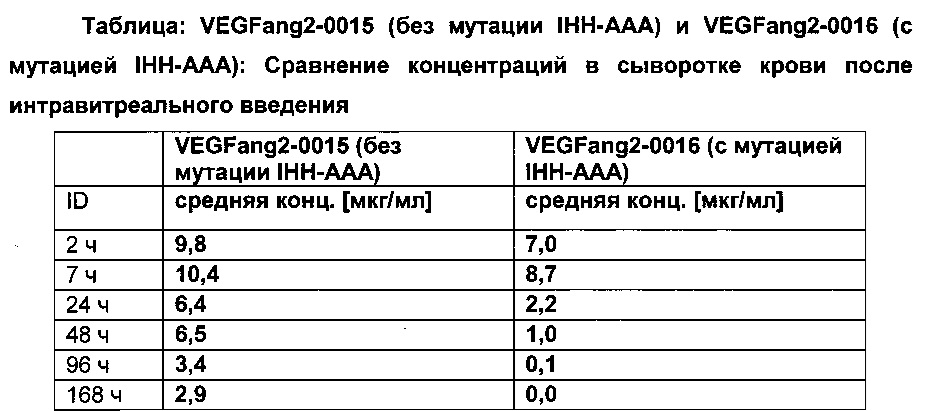
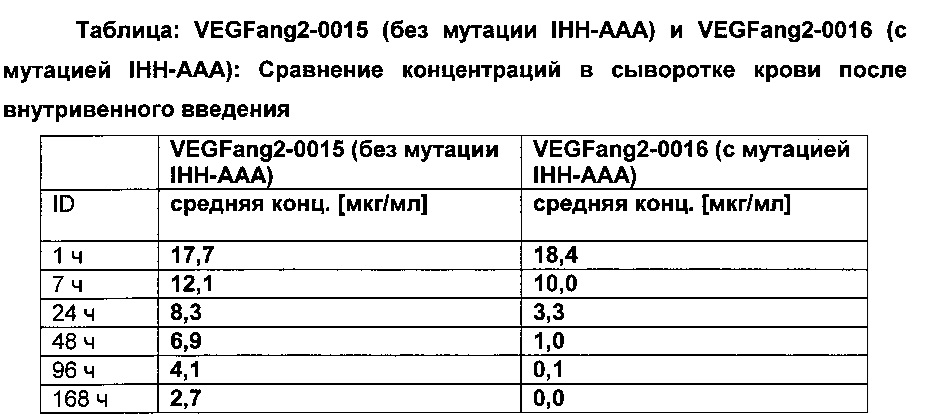
Результаты:
В) Концентрации в глазных лизатах левого и правого глаз Результаты для концентраций в глазных лизатах приведены в следующих таблицах и на фиг. 7D-7E.




Краткое изложение результатов:
После интравитреального применения описанное в данном документе биспецифическое <VEGF-ANG-2> антитело VEGFang2-0016 (с мутацией IHH-AAA) демонстрирует аналогичные концентрации (через 96 и 168 часов) в глазных лизатах при сравнении с биспецифическим <VEGF-ANG-2> антителом без мутации IHH-AAA VEGFang2-0015.
Кроме того, после интравитреального применения описанное в данном документе биспецифическое <VEGF-ANG-2> антитело VEGFang2-0016 (с мутацией IHH-AAA) демонстрирует более быстрое выведение и более короткий период полужизни в сыворотке крови по сравнению с биспецифическим <<VEGF-ANG-2> антителом VEGFang2-0015 без мутации IHH-AAA.
Пример 7
Микрокарманный анализ ангиогенеза на роговице мышей
Чтобы проверить антиангиогенное влияние биспецифического <VEGF-ANG-2> антитела с соответствующими VEGF-связывающими VH и VL с SEQ ID №20 и 21 и ANG-2-связывающими VH и VL с SEQ ID №28 и 29 на VEGF-индуцированный ангиогенез in vivo, проводили микрокарманный анализ ангиогенеза на роговице мышей. В этом анализе диск Nylaflo, пропитанный VEGF, имплантировали в карман в бессосудистой роговице на фиксированном расстоянии от лимбальных сосудов. Сосуды немедленно росли в роговицу в направлении увеличивающегося градиента VEGF. Самок мышей Balb/c в возрасте от 8 до 10 недель приобретали у Charles River, Зульцфельд, Германия. Протокол модифицировали в соответствии со способом, описанным в Rogers, M.S., et al., Nat. Protoc. 2 (2007) 2545-2550. Вкратце, у анестезированных мышей под микроскопом делали микрокарманы с шириной примерно 500 мкм на расстоянии примерно 1 мм от лимба к верхней части роговицы, используя хирургический скальпель и острые пинцеты. Имплантировали диск (Nylaflo®, Pall Corporation, Мичиган) с диаметром 0,6 мм и поверхность области имплантации сглаживали. Диски инкубировали в соответствующем факторе роста или в носителе в течение по меньшей мере 30 мин. Через 3, 5 и 7 дней (или, альтернативно, только через 3, 5 или 7 дней) глаза фотографировали и измеряли сосудистую реакцию. Анализ количественно оценивали путем подсчета процента площади новых сосудов от общей площади роговицы.
В диски загружали 300 нг VEGF или PBS в качестве контроля и имплантировали на 7 дней. Разрастание сосудов от лимба к диску контролировали с течением времени на 3, 5 и/или 7 день. За день до имплантации диска внутривенно вводили антитела в дозе 10 мг/кг (в связи с внутривенным применением стабильное в сыворотке антитело VEGFang2-0015 (без мутации IHH-AAA), которое отличается от антитела VEGFang2-0016 только мутацией IHH-AAA и имеет те же анти-VEGF- и анти-ANG-2-VH и -VL для опосредования эффективности, использовано в качестве заместителя) для проверки антиангиогенного влияния на VEGF-индуцированный ангиогенез in vivo. Животные в контрольной группе получали носитель. Вводимый объем составлял 10 мл/кг.
Пример 8
Фармакокинетические (РК) свойства антител с мутацией HHY-AAA
Данные РК на FcRn мышах, трансгенных по человеческому FcRn Прижизненная фаза:
В исследование были включены самки мышей C57BL/6J (фон); мыши, дефицитные по FcRn, но гемизиготно трансгенные по человеческому FcRn (huFcRn, линия 276 -/tg)
Часть 1:
Всем мышам вводили однократно интравитреально в правый глаз 2 мкл/животное соответствующего раствора VEGF/ANG2-0016, VEGF/ANG2-0096, VEGF/ANG2-0098, VEGF/ANG2-0121.
Мышей делили на 2 группы по 6 животных в каждой. Образцы крови брали в группе 1 через 2, 24 и 96 часов, а в группе 2 через 7, 48 и 168 часов после введения дозы.
Инъекцию в стекловидное тело правого глаза мыши выполняли с использованием системы NanoFil Microsyringe для нанолитровых инъекций от World Precision Instruments, Inc., Берлин, Германия. Мышей анестезировали с помощью 2,5% изофлурана и для визуализации глаза мыши использовали микроскоп Leica MZFL 3 с 40-кратным увеличением и кольцевым осветителем Leica KL 2500 LCD. Затем вводили 2 мкл соединения с помощью иглы размера 35.
У каждого животного собирали кровь из ретробульбарного венозного сплетения контралатерального таза для определения уровней соединений в сыворотке крови.
После инкубации при комнатной температуре в течение 1 часа из крови получали образцы сыворотки объемом по меньшей мере 50 мкл путем центрифугирования (9300 д) при 4°C в течение 3 мин. Образцы сыворотки замораживали сразу после центрифугирования и хранили в замороженном виде при -80°С до анализа. Глаза, в которые вводили соединение, у животных из группы 1 выделяли через 96 часов после введения, а у животных из группы 2 через 168 ч после введения. Образцы хранили в замороженном виде при -80°C до анализа.
Часть 2:
Все мышам однократно внутривенно (через хвостовую вену) вводили 200 мкл/животное соответствующего VEGF/ANG2-0096, VEGF/ANG2-0098 или VEGF/ANG2-0121.
Мышей делили на 2 группы по 5 животных в каждой. Образцы крови брали в группе 1 через 1, 24 и 96 часов, а в группе 2 через 7, 48 и 168 часов после введения дозы. У каждого животного собирали кровь из ретробульбарного венозного сплетения для определения уровней соединения в сыворотке крови.
После инкубации при комнатной температуре в течение 1 часа из крови получали образцы сыворотки объемом по меньшей мере 50 мкл путем центрифугирования (9300 д) при 4°C в течение 3 мин. Образцы сыворотки замораживали сразу после центрифугирования и хранили в замороженном виде при -80°C до анализа.
Получение цельных лизатов глаз (мышиных)
Глазные лизаты получали путем физико-химического разрушения цельных глаз. Для механического разрушения каждый глаз переносили в микропробирку объемом 1,5 мл с коническим дном. После замораживания и размораживания глаза промывали один раз 1 мл промывочного буфера (Bio-Rad, Bio-Plex Cell Lysis Kit, кат. №171-304011). На следующем этапе добавляли 500 мкл свежеприготовленного буфера для лизиса клеток и измельчали глаза с помощью пестика для измельчения тканей в микропробирках объемом 1,5 мл (Kimble Chase, 1.5 mL pestle, арт. №749521-1500). Затем смесь замораживали и размораживали пять раз и снова измельчали. Чтобы отделить лизат от остальных тканей, образцы центрифугировали в течение 4 мин при 4500 g. После центрифугирования супернатант собирали и хранили при -20°C до дальнейшего анализа путем количественной ELISA.
Анализ
Концентрации антител в сыворотках и в глазных лизатах мышей определяли путем твердофазного иммуноферментного анализа (ELISA).
Для количественной оценки антител в образцах сыворотки и глазных лизатах мышей проводили стандартный твердофазный серийный иммуноферментный анализ типа "сэндвич" с биотинилированными и дигоксигенилированными моноклональными антителами, используемыми в качестве антител захвата и обнаружения. При проверке сохранности биспецифичности анализируемого антитела биотинилированное антитело захвата распознает сайт связывания VEGF, тогда как дигоксигенилированное антитело обнаружения связывается с сайтом связывания ANG-2 анализируемого антитела. Затем проводят обнаружение связанного иммунного комплекса антитела захвата, анализируемого антитела и антитела обнаружения на твердой фазе микротитровального планшета, покрытого стрептавидином (SA-MTP), с помощью пероксидазы хрена, соединенной с антидигоксигениновым антителом. После отмывания несвязанного материала из SA-MTP и добавления ABTS-субстрата полученный сигнал пропорционален количеству анализируемого антитела, связанного с твердой фазой SA-MTP. Затем проводят количественную оценку путем преобразования измеренных сигналов от образцов в концентрации согласно стандартам (калибраторам), анализируемым параллельно.
На первом этапе в SA-MTP вносили раствор биотинилированного антитела захвата (антиидиотипическое антитело, например, mAb<Id<VEGF>>M-2.45.51-IgG-Bi(DDS)) в объеме 100 мкл/лунка с концентрацией 1 мкг/мл на один час на MTP-шейкере со скоростью 500 об/мин. В это время готовили стандарты, QC-образцы и анализируемые образцы. Стандарты и QC-образцы разводили до 2% сывороточной матрицы; анализируемые образцы разводили таким образом, чтобы сигналы находились в пределах линейного диапазона стандартов.
После нанесения на SA-MTP антитела захвата планшет промывали три раза промывочным буфером в объеме 300 мкл/лунка. Затем в SA-MTP пипеткой вносили по 100 мкл/лунка стандартов, QC-образцов и анализируемых образцов, соответственно, и снова инкубировали в течение одного часа при 500 об/мин. Теперь анализируемое антитело прикреплялось одним из его сайтов связывания к твердой фазе SA-MTP через антитело захвата. После инкубации и удаления несвязанного анализируемого антитела путем промывания планшета в SA-MTP вносили 100 мкл/лунка первого антитела обнаружения (антиидиотипическое антитело, например, mAb<Id-<ANG-2>>M-2.6.81-IgG-Dig(XOSu)) в концентрации 250 нг/мл. Планшет снова инкубировали в течение одного часа на шейкере при 500 об/мин. После промывания в лунки SA-MTP вносили по 100 мкл/лунка второго антитела обнаружения (например, pAb<Digoxigenin>S-Fab-POD (poly)) в концентрации 50 мЕд/мл, и планшет снова инкубировали в течение одного часа при 500 об/мин. После финального этапа промывания, чтобы удалить избыток антитела обнаружения, вносили по 100 мкл/лунка субстрата (ABTS®). Конъюгат "антитело-фермент" катализировал цветовую реакцию субстрата ABTS®. Затем сигнал измеряли с помощью ELISA-ридера на длине волны 405 нм (референсная длина волны. 490 нм ([405/490] нм)).
Фармакокинетическая оценка
Фармакокинетические параметры рассчитывали путем некомпартментного анализа, используя программу для фармакокинетической оценки WinNonlin™ (Pharsight) версии 5.2.1.
Хотя данное изобретение было подробно описано посредством иллюстраций и примеров для ясности понимания, эти описания и примеры не должны быть истолкованы как ограничивающие объем данного изобретения. Раскрытие всей патентной и научной литературы, цитируемой в данном документе, ясно включено во всей ее полноте посредством ссылки.
















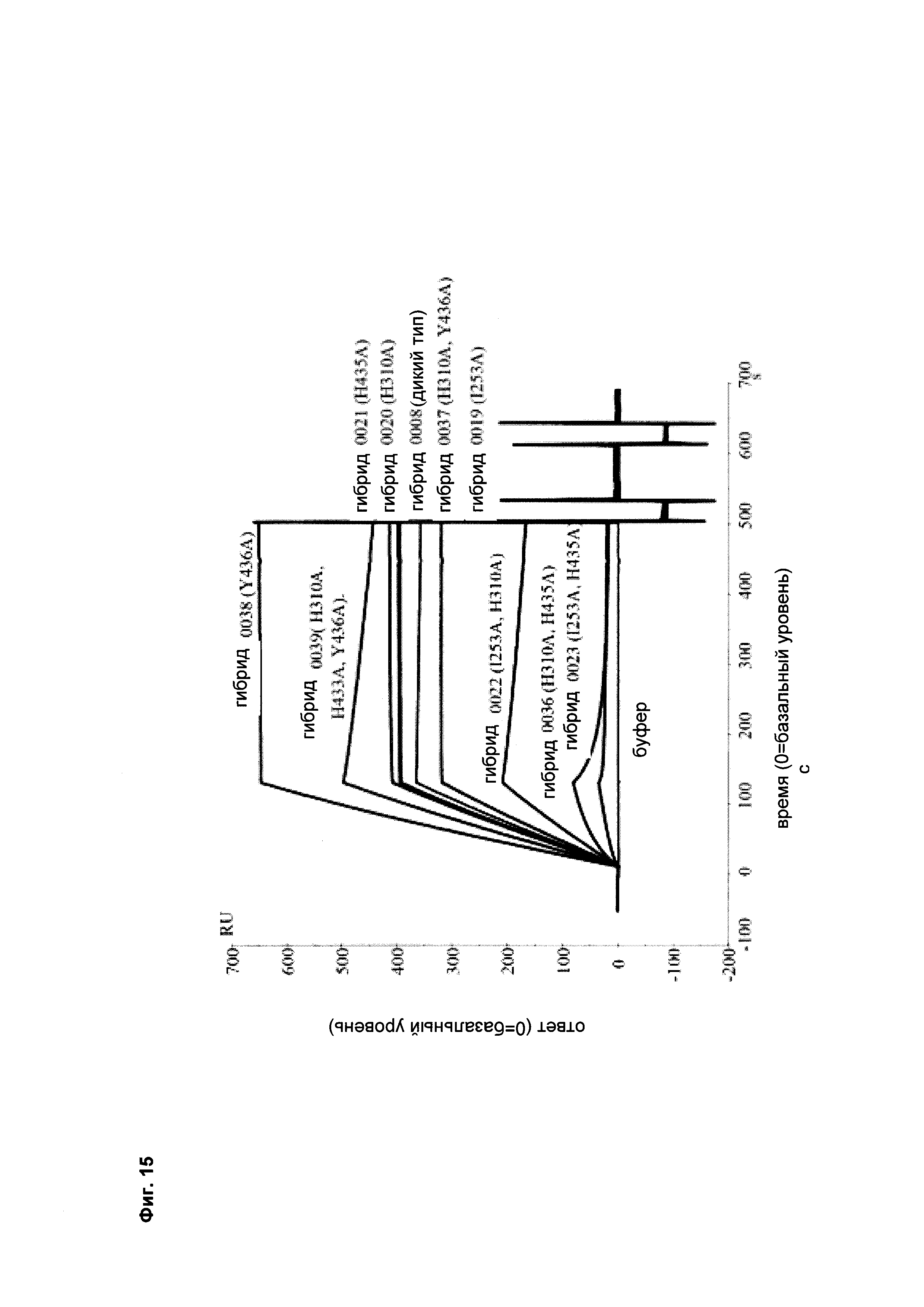




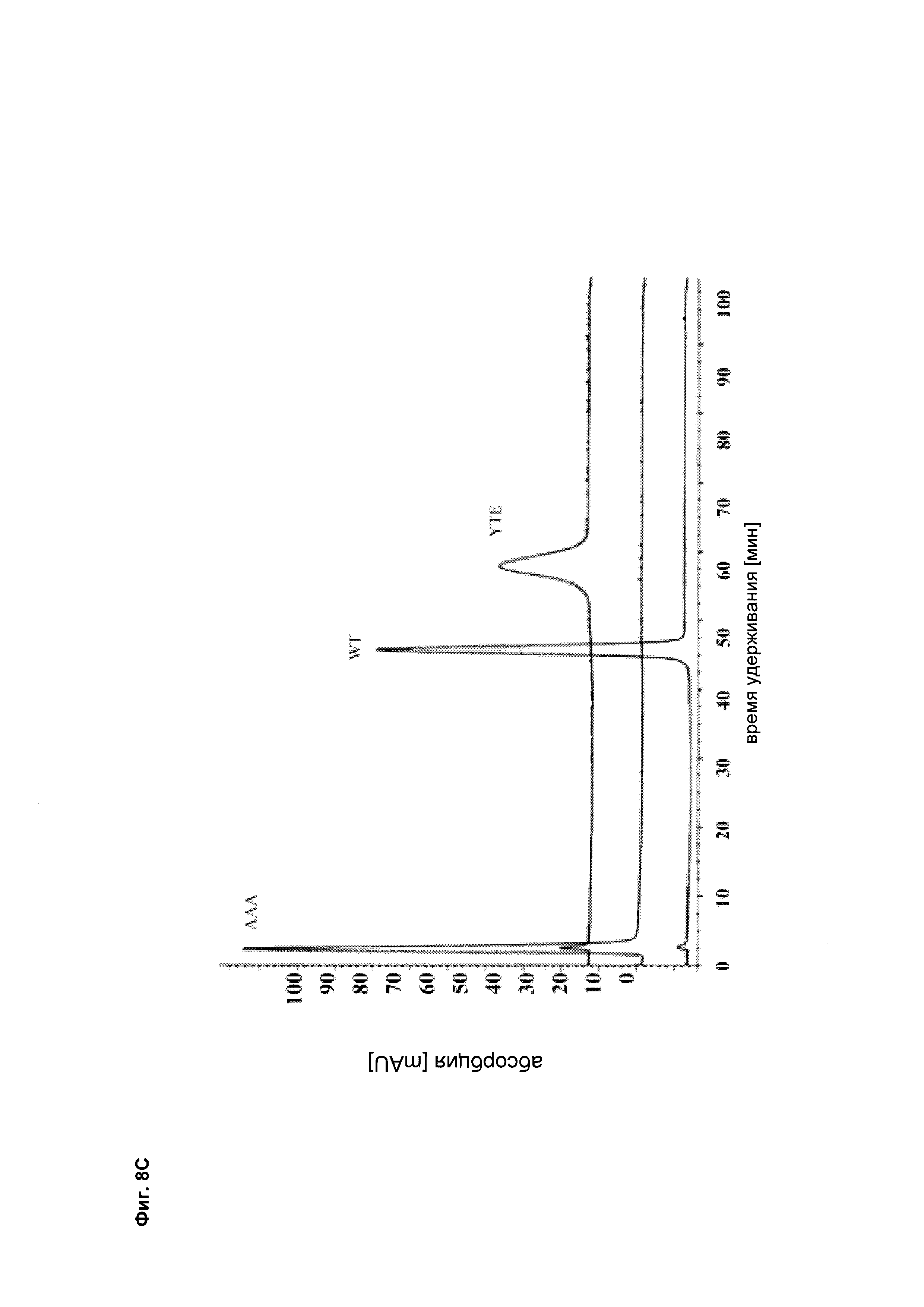













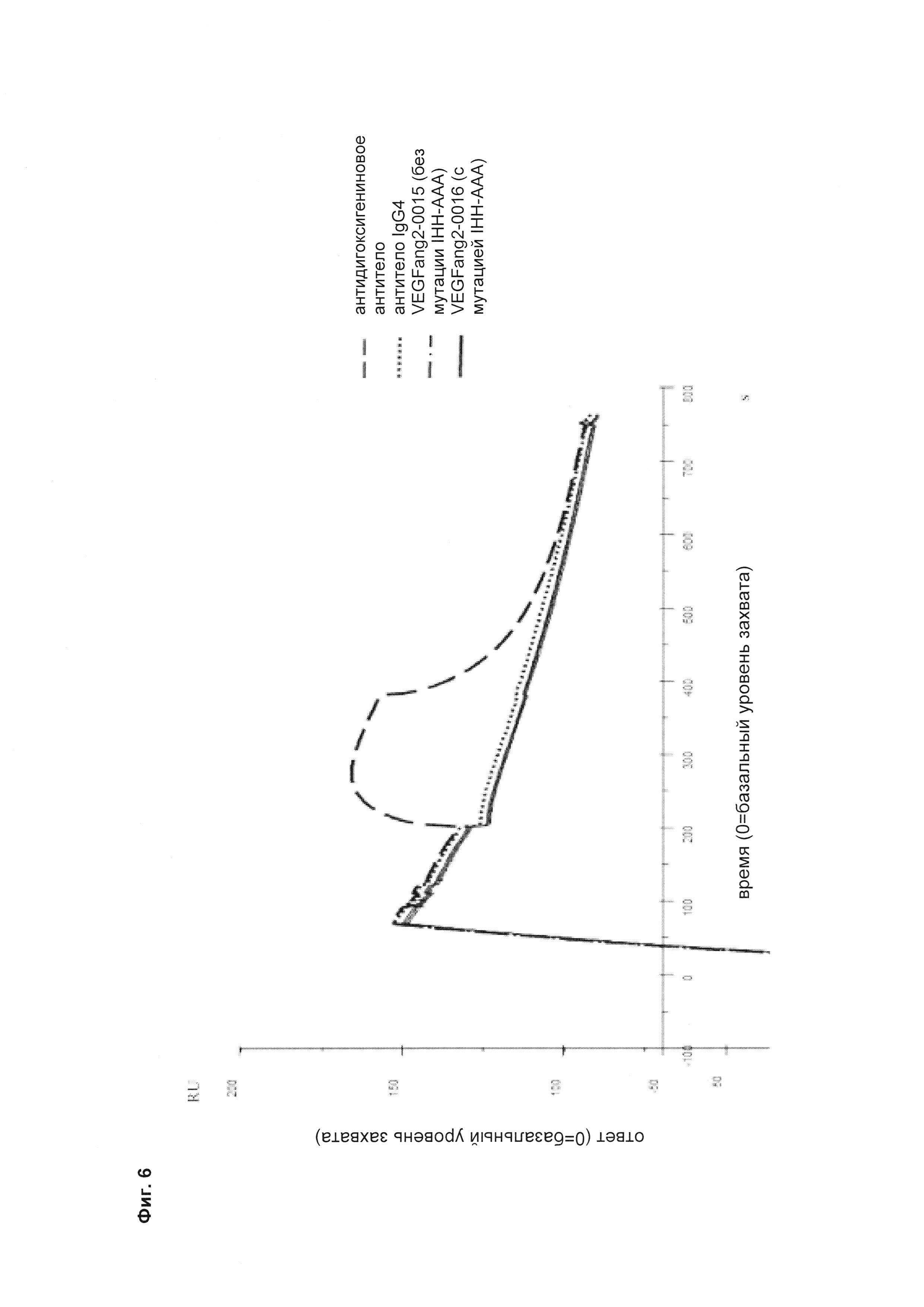





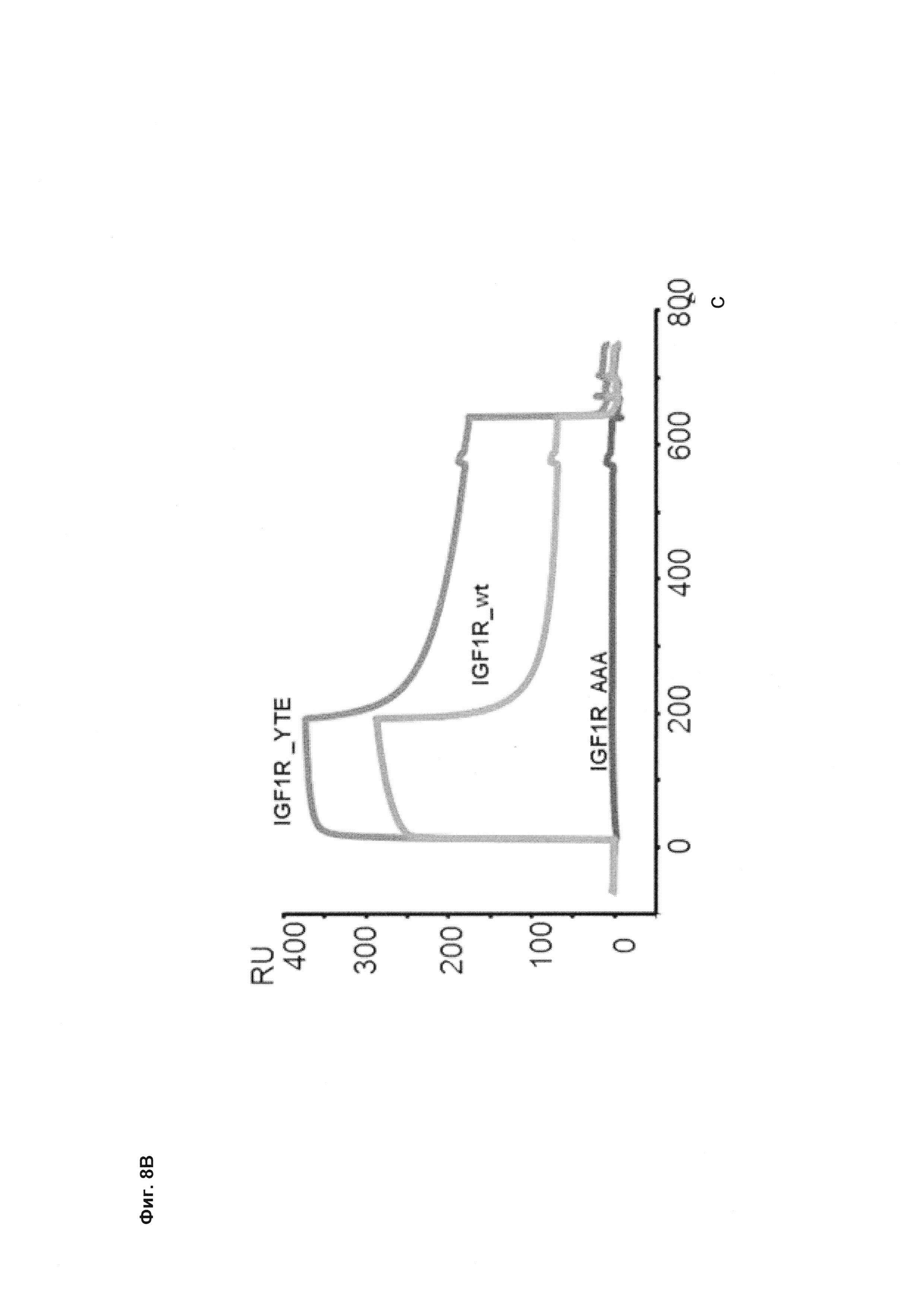


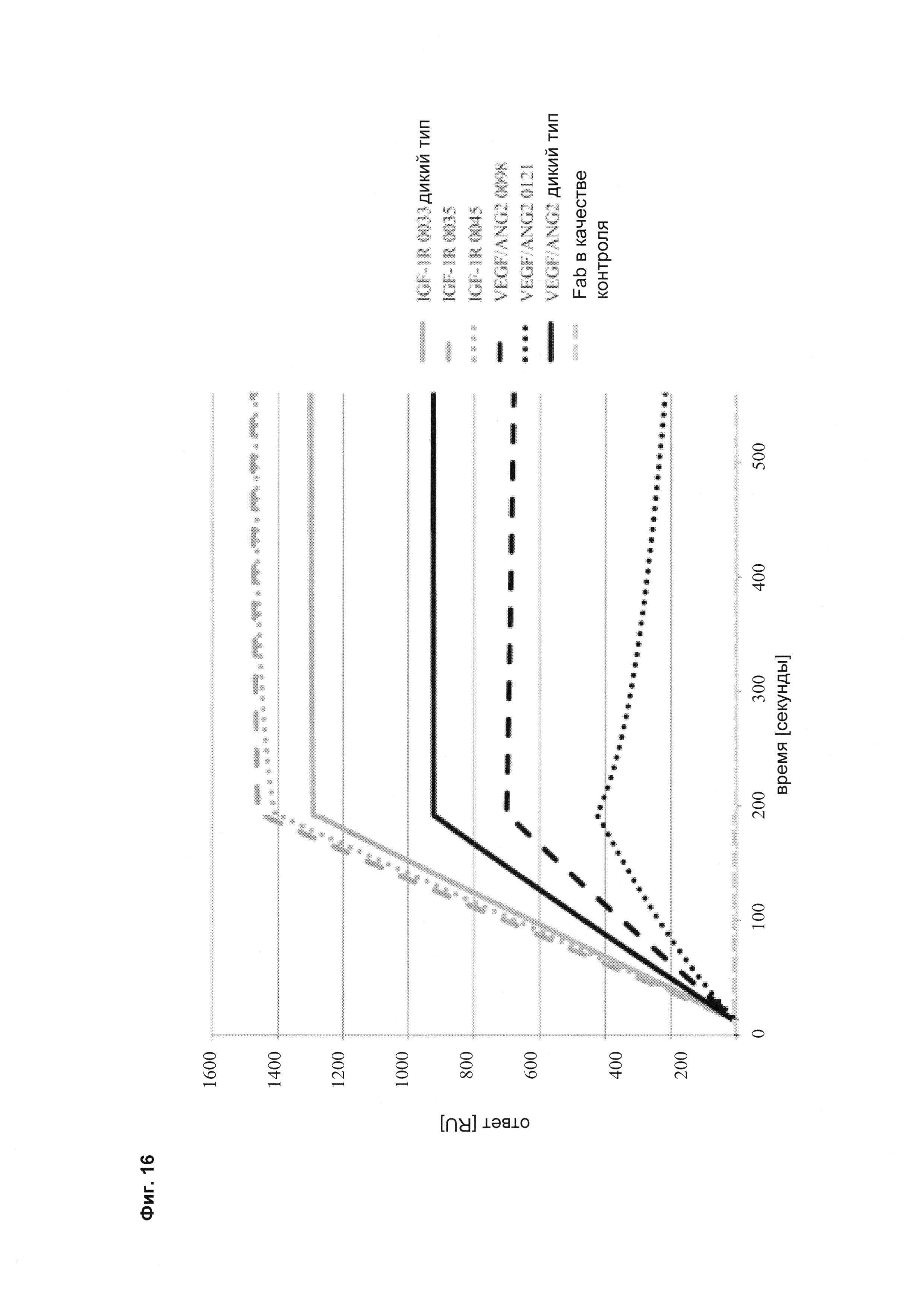
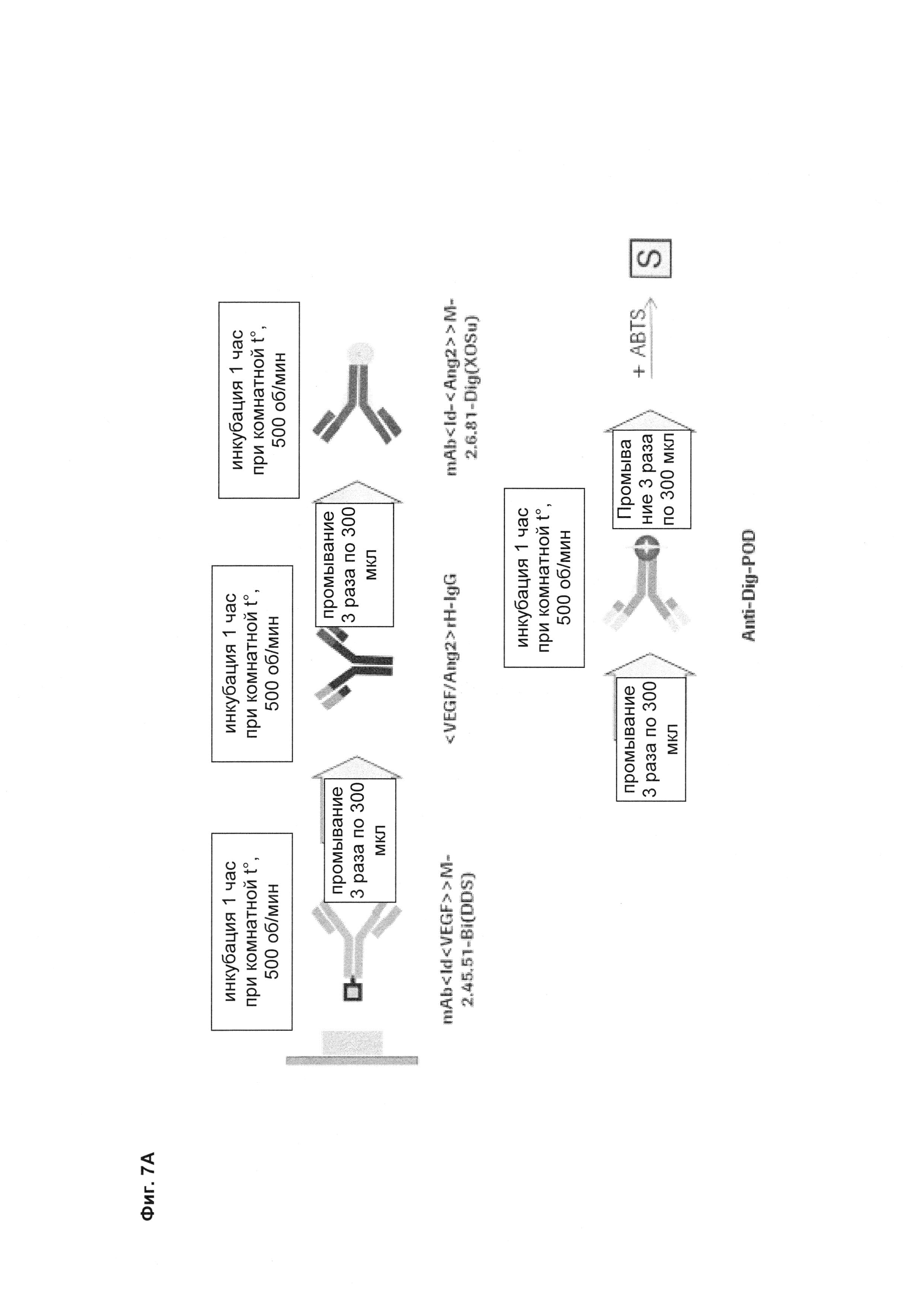

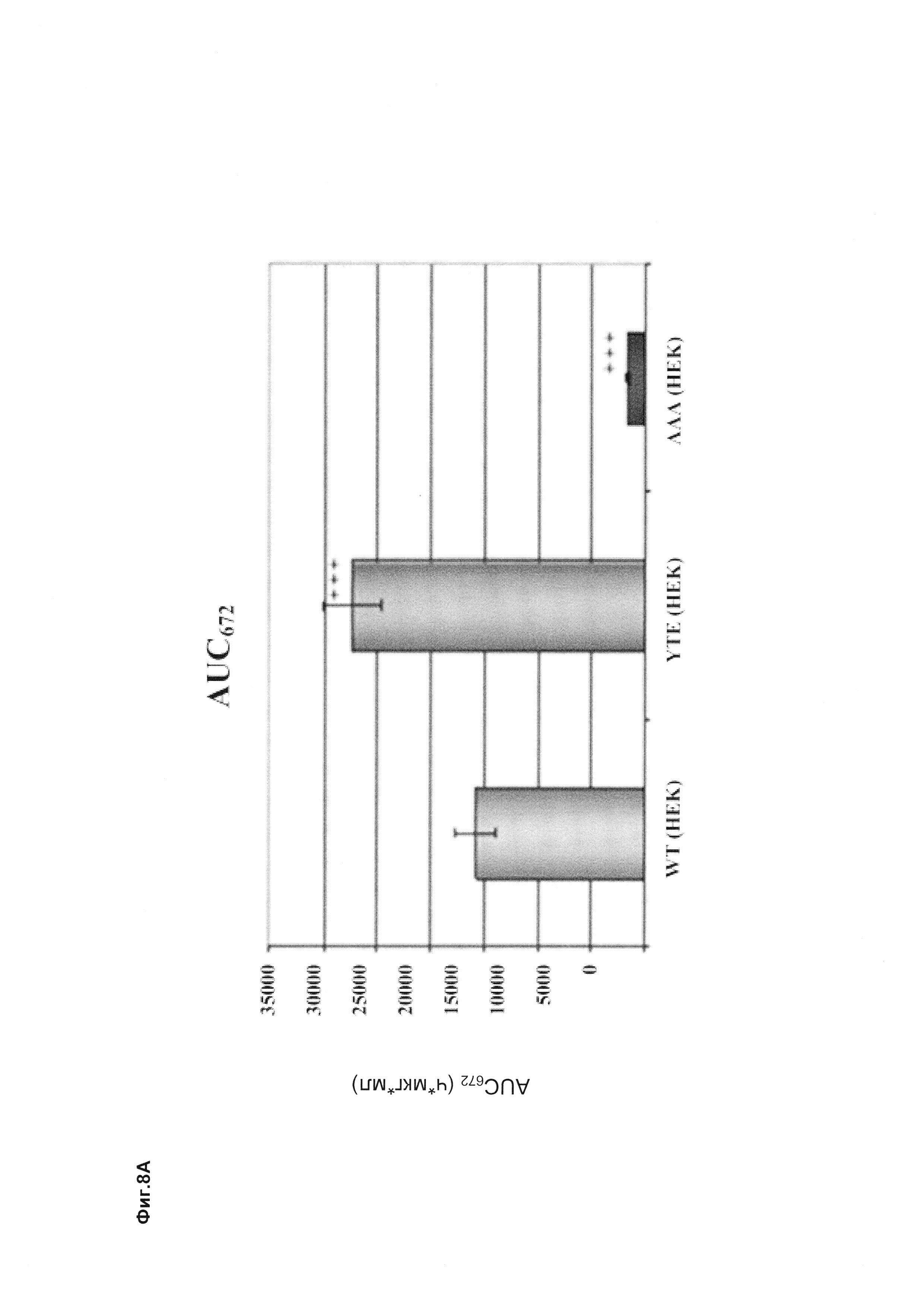


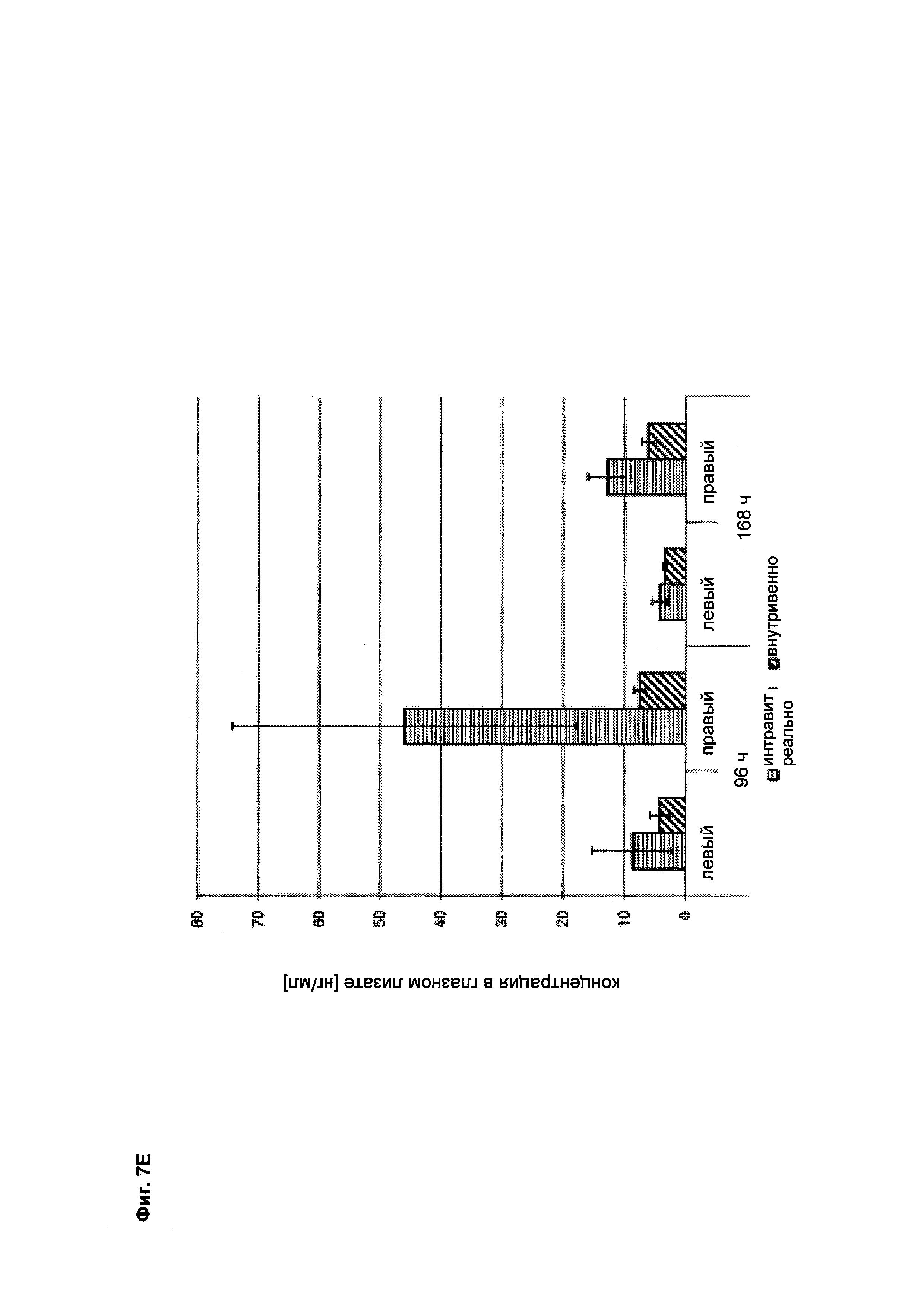





















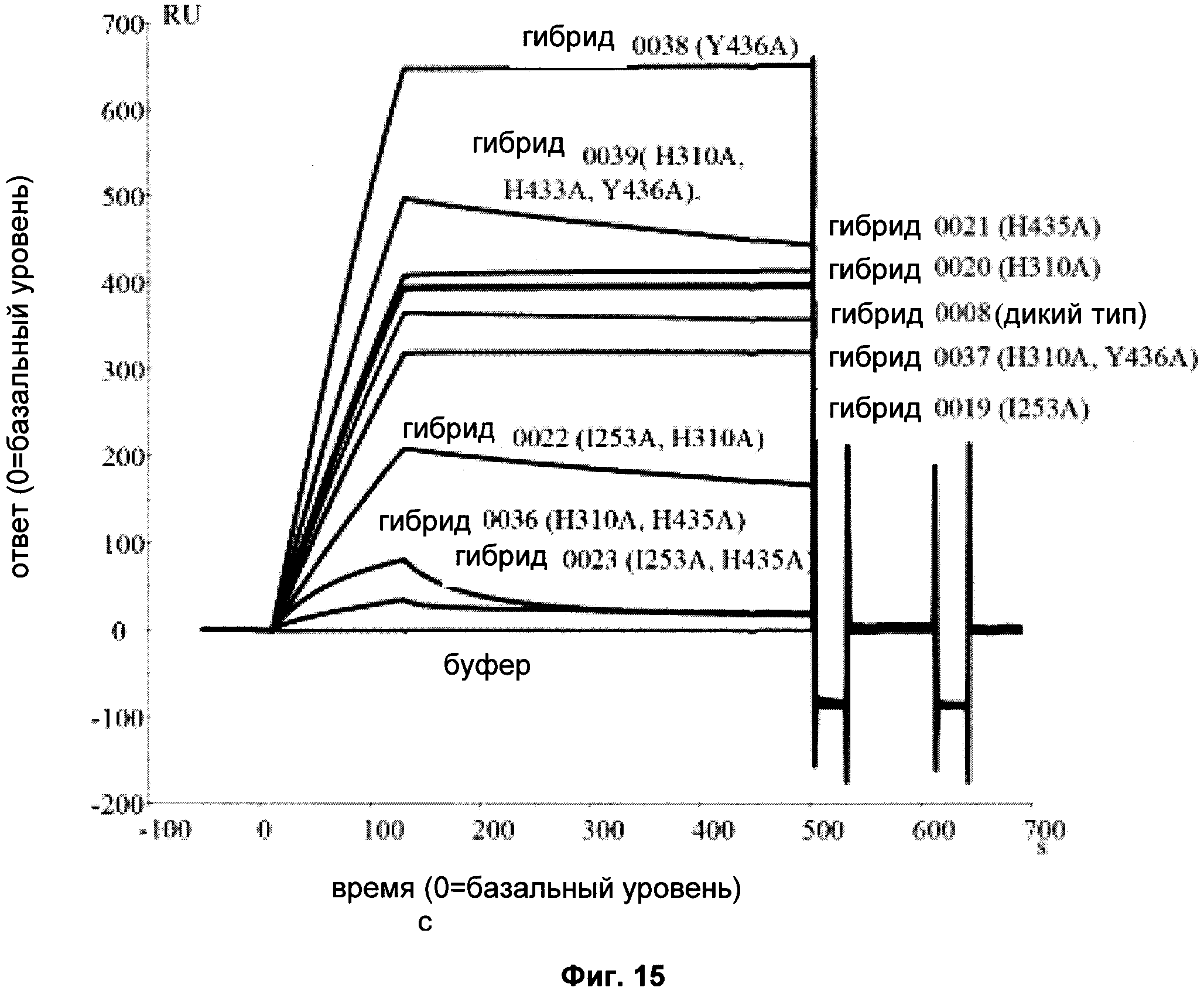
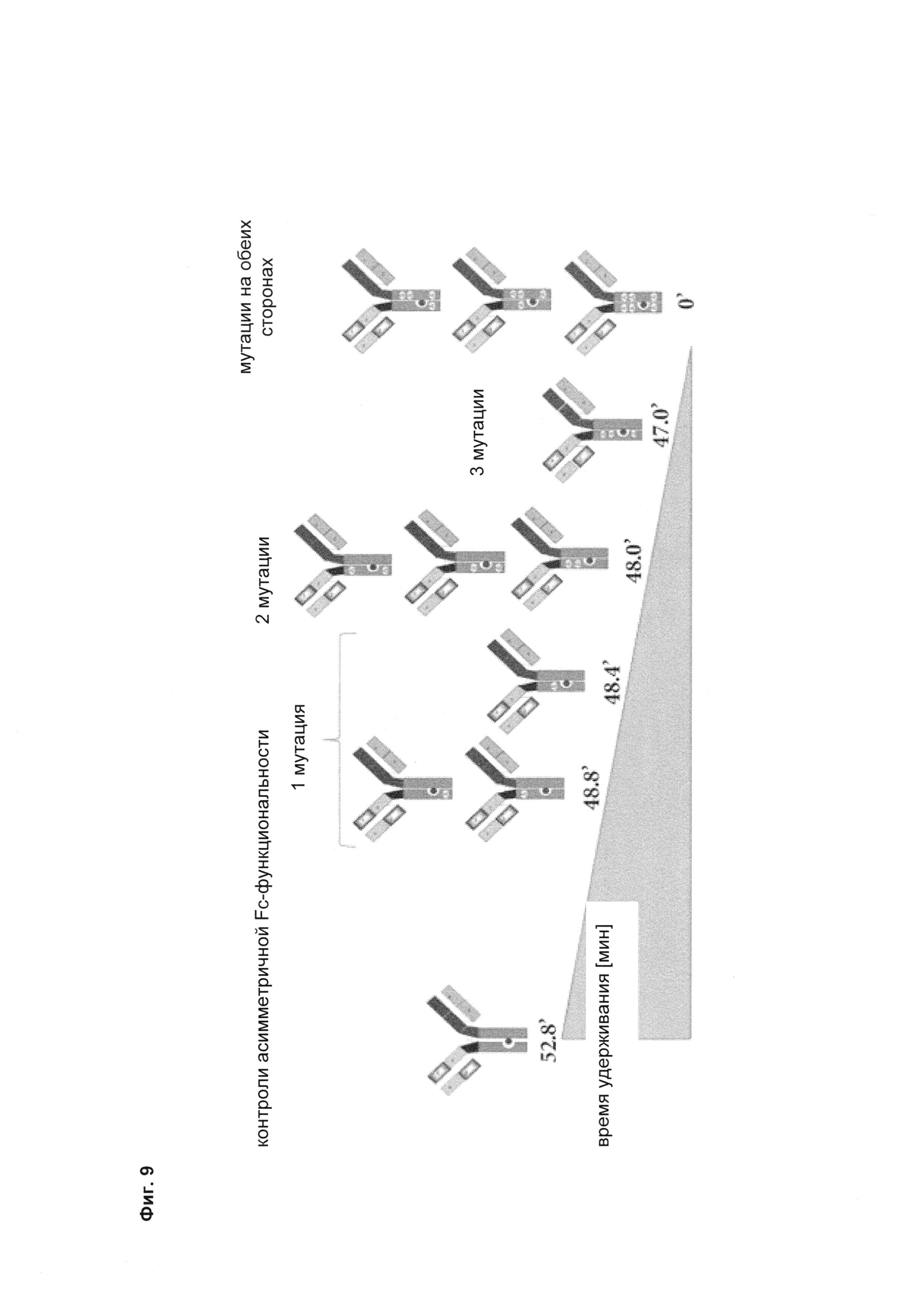

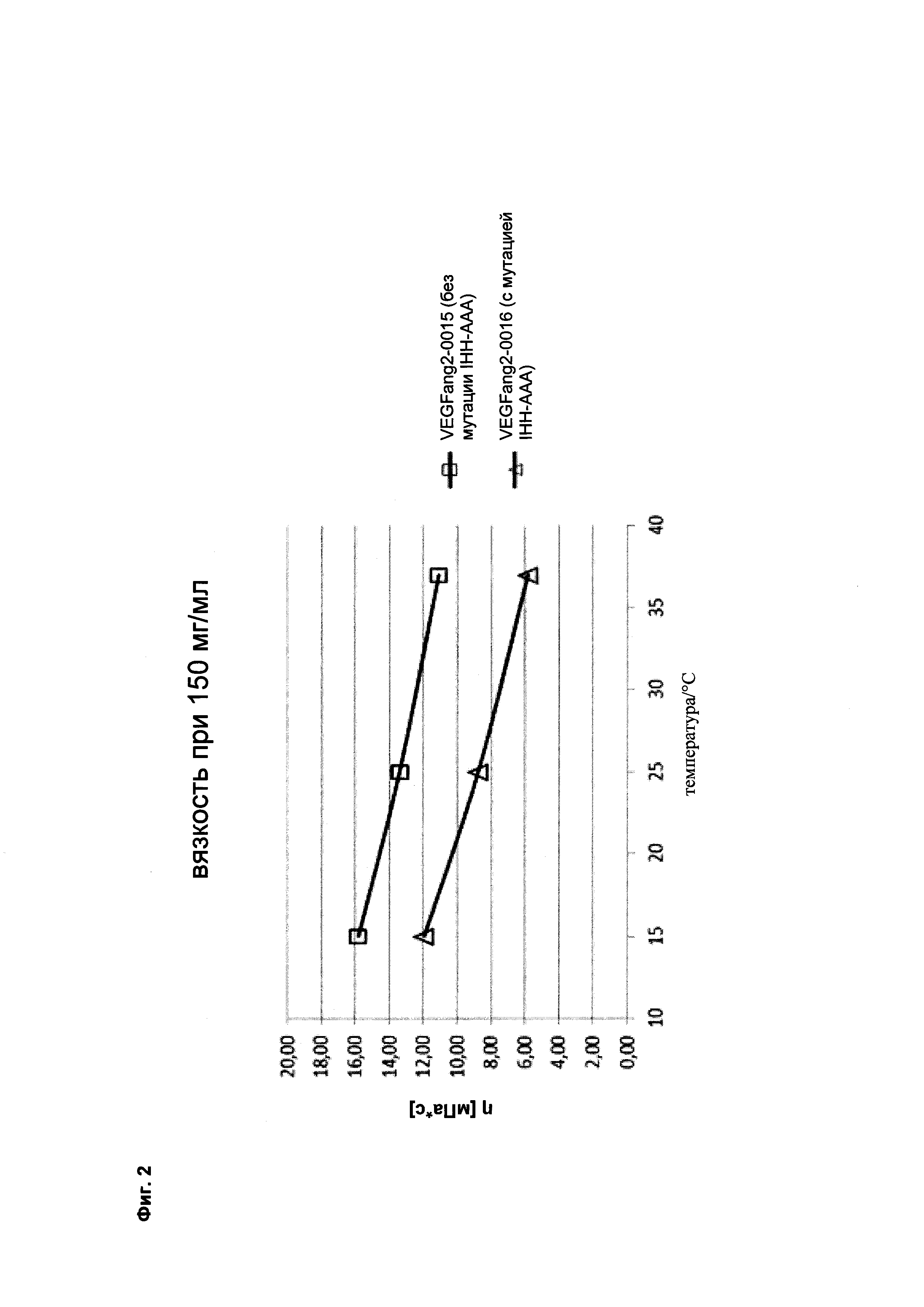





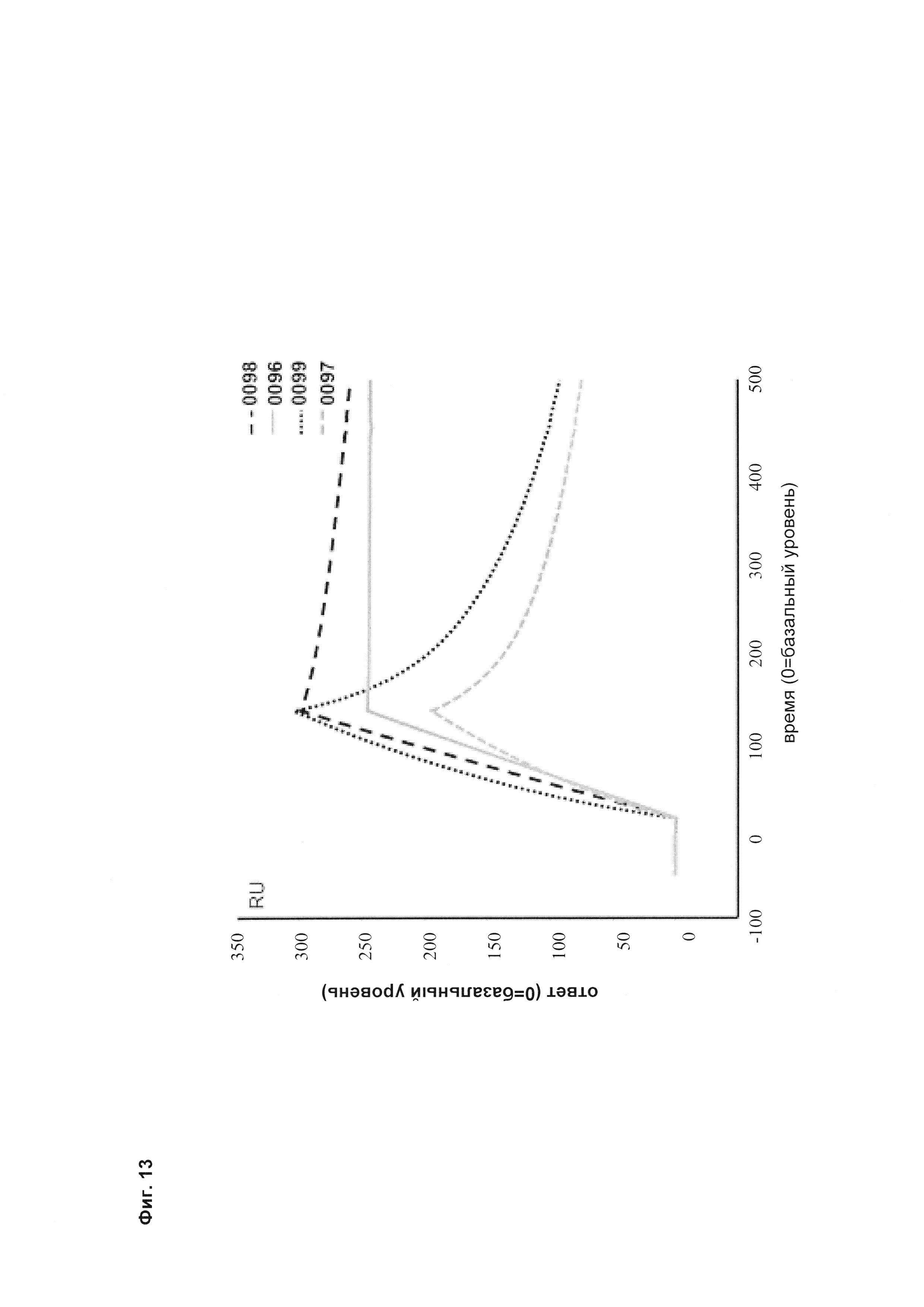
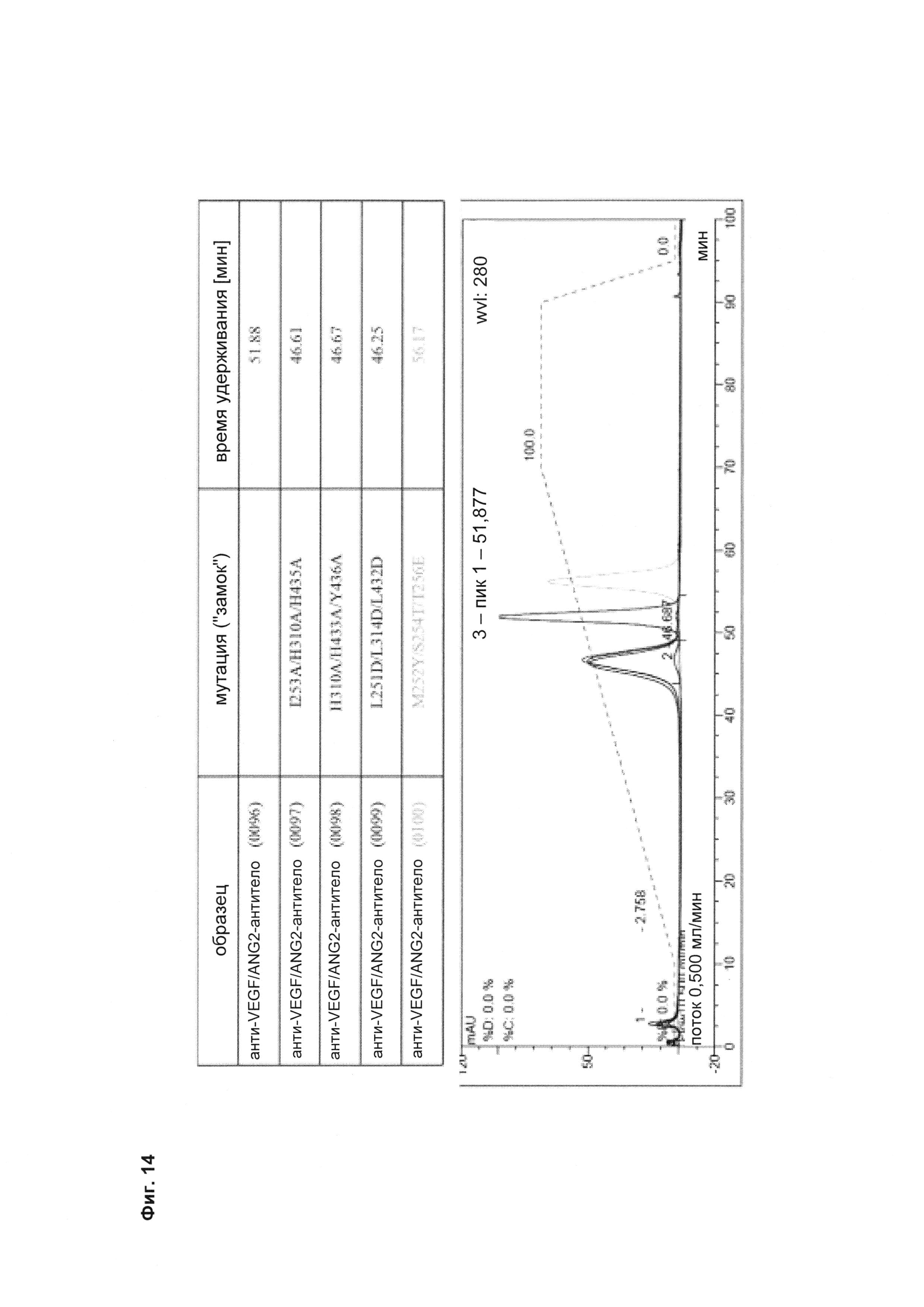

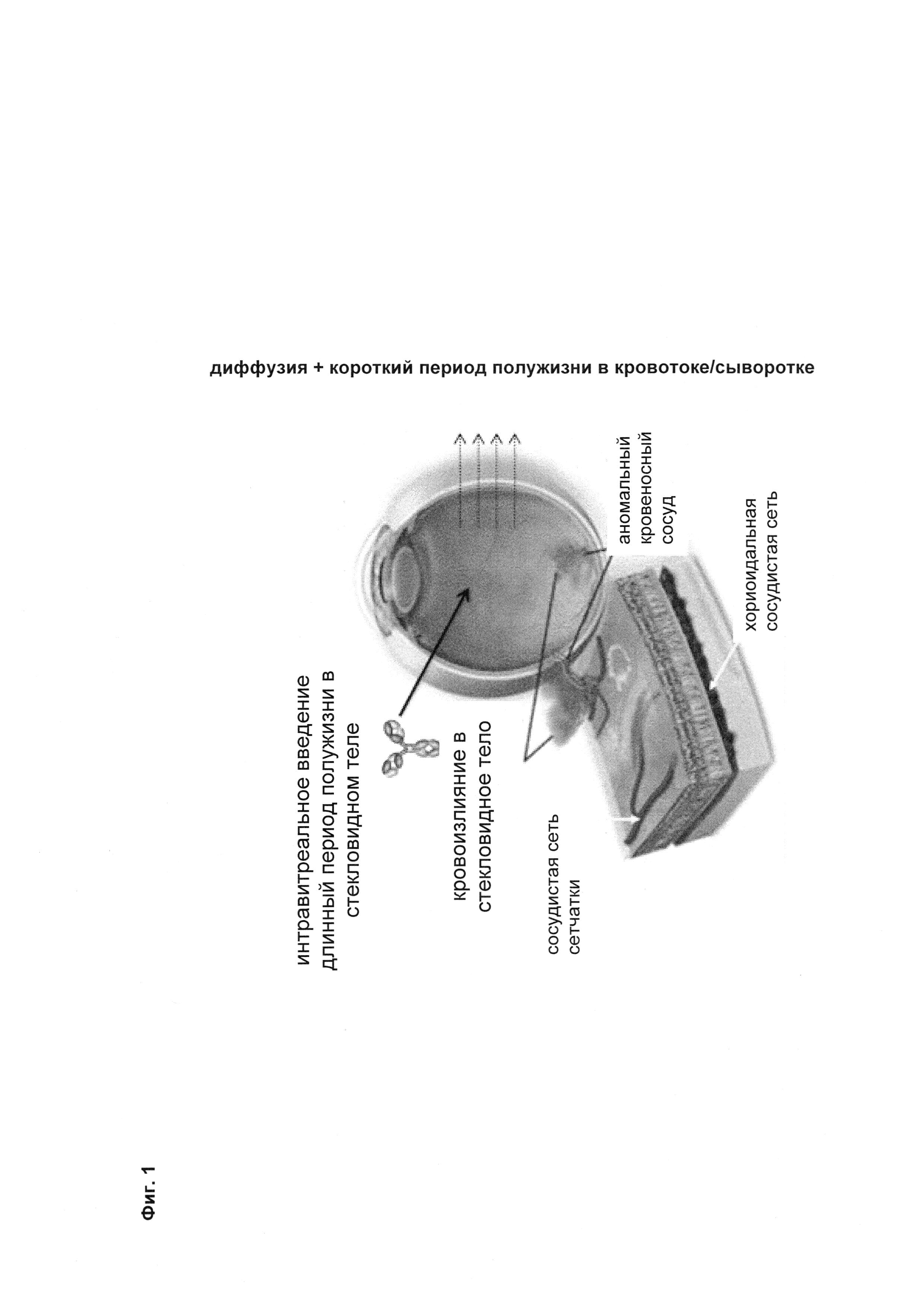




















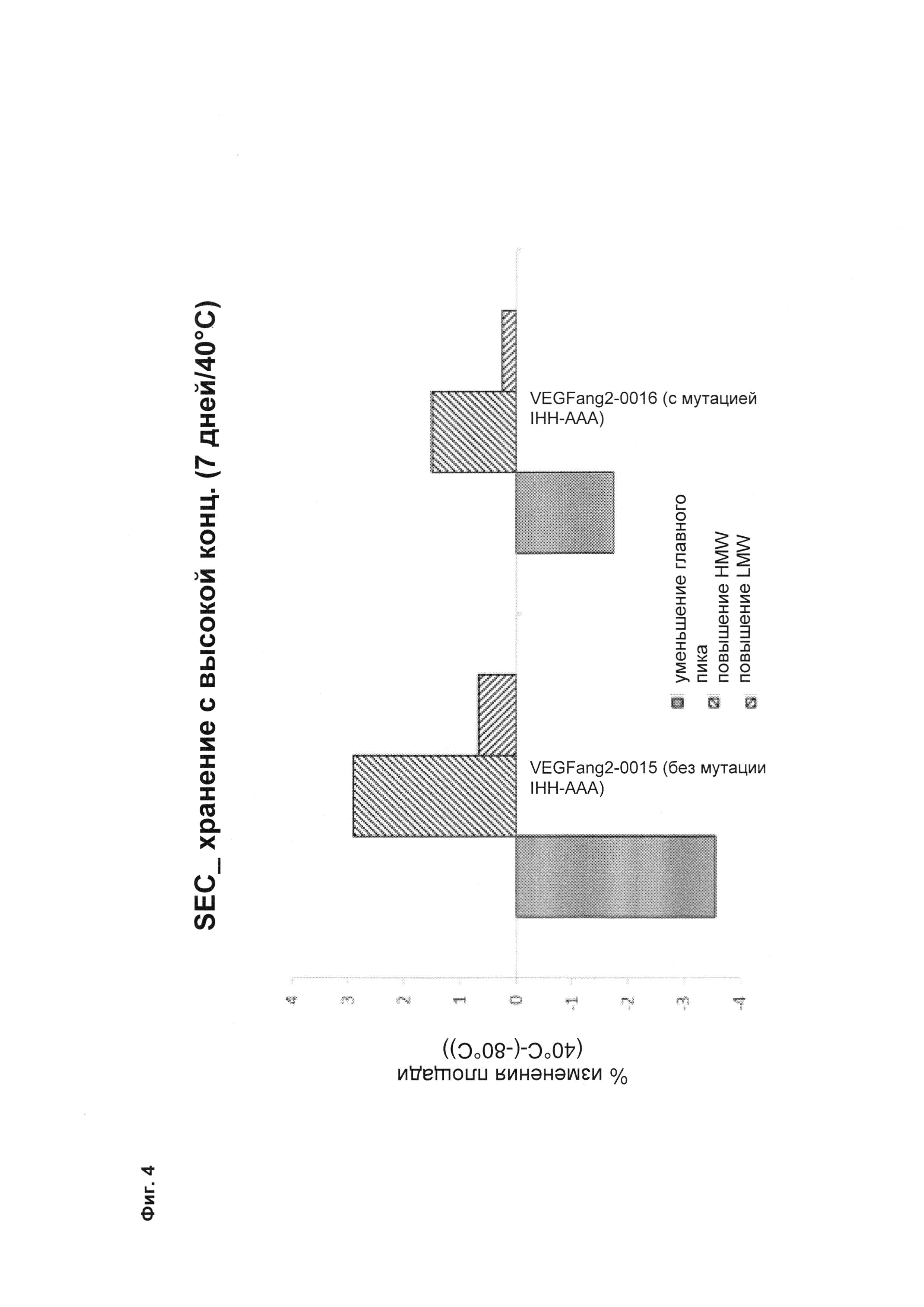


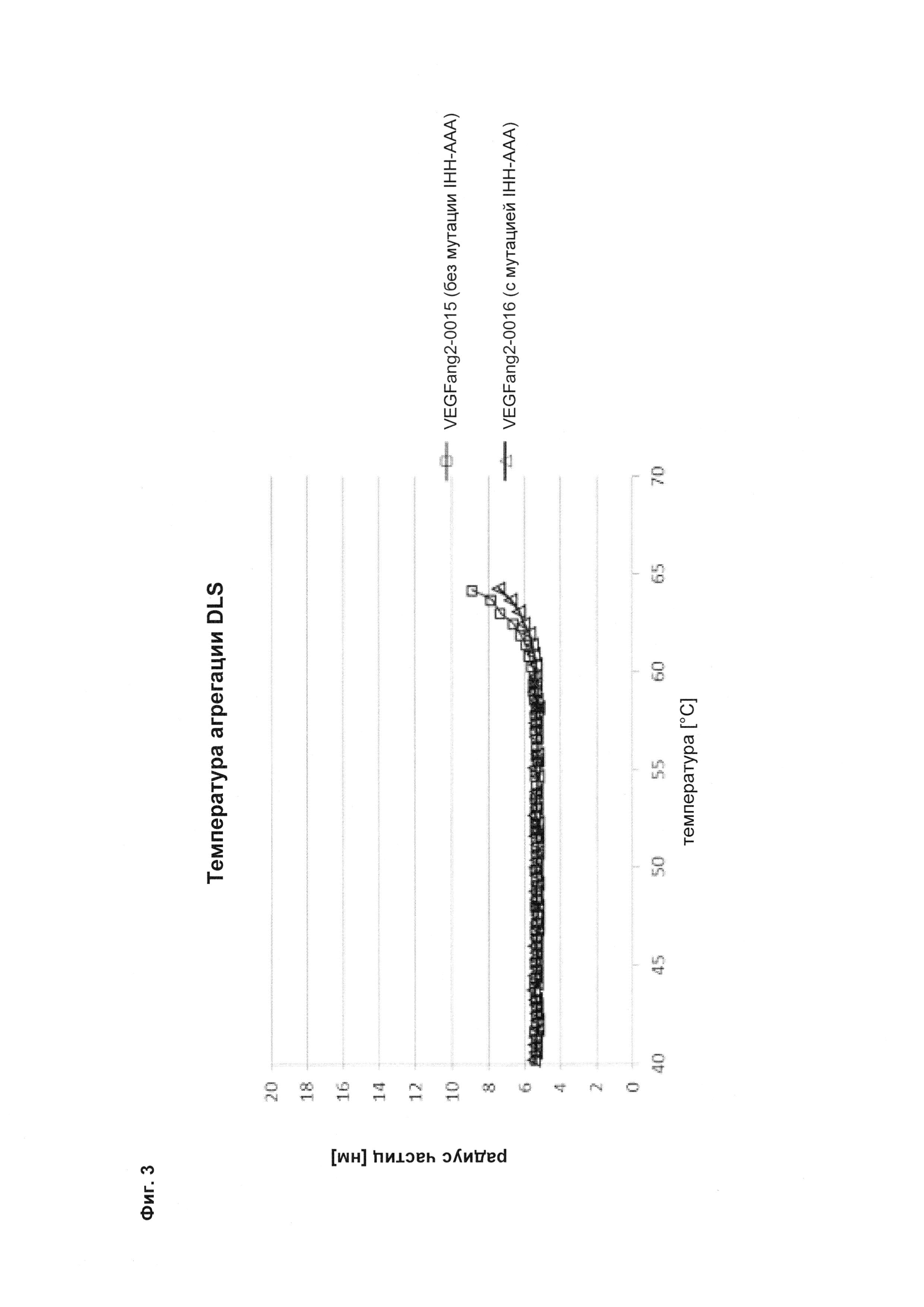


























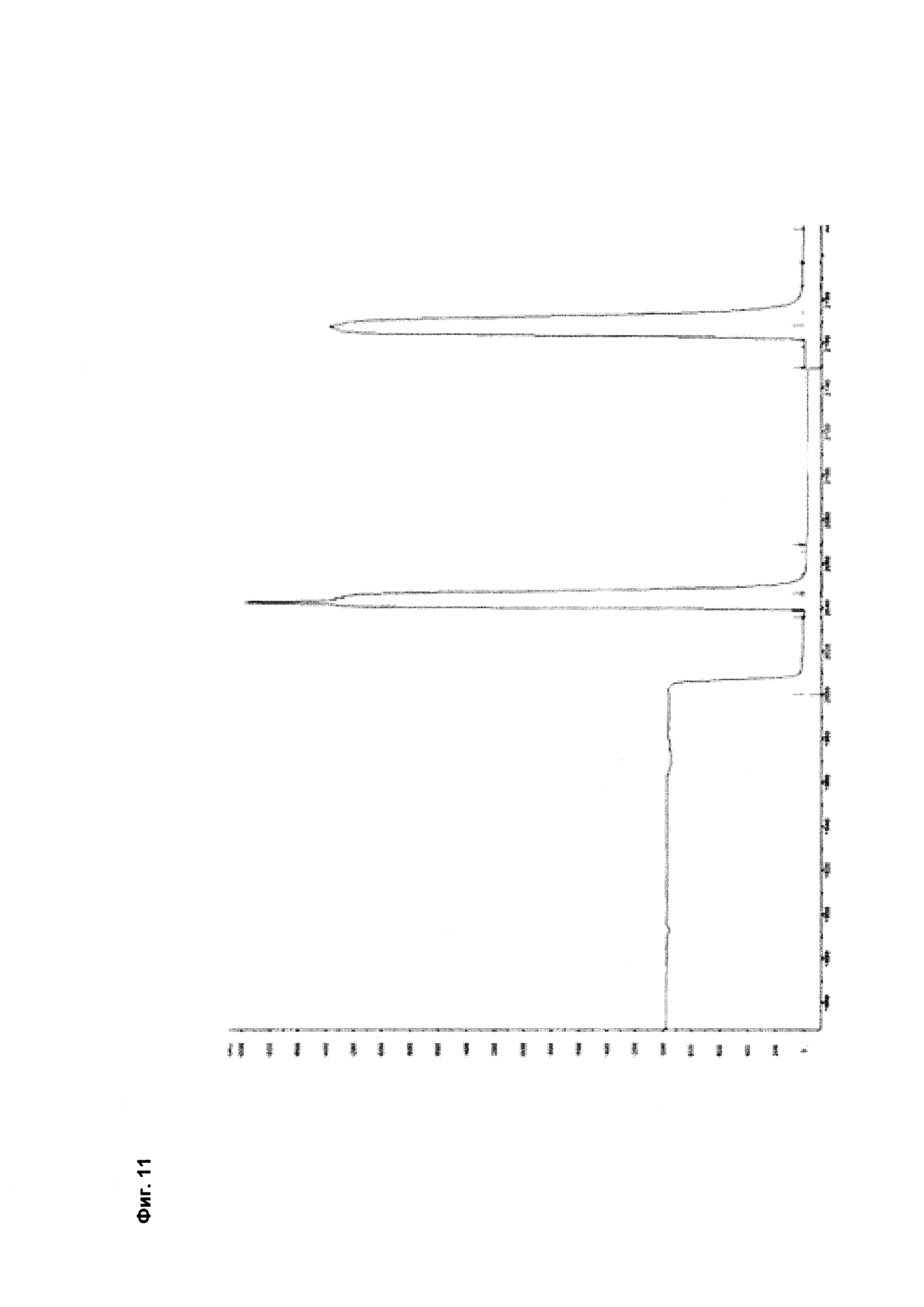




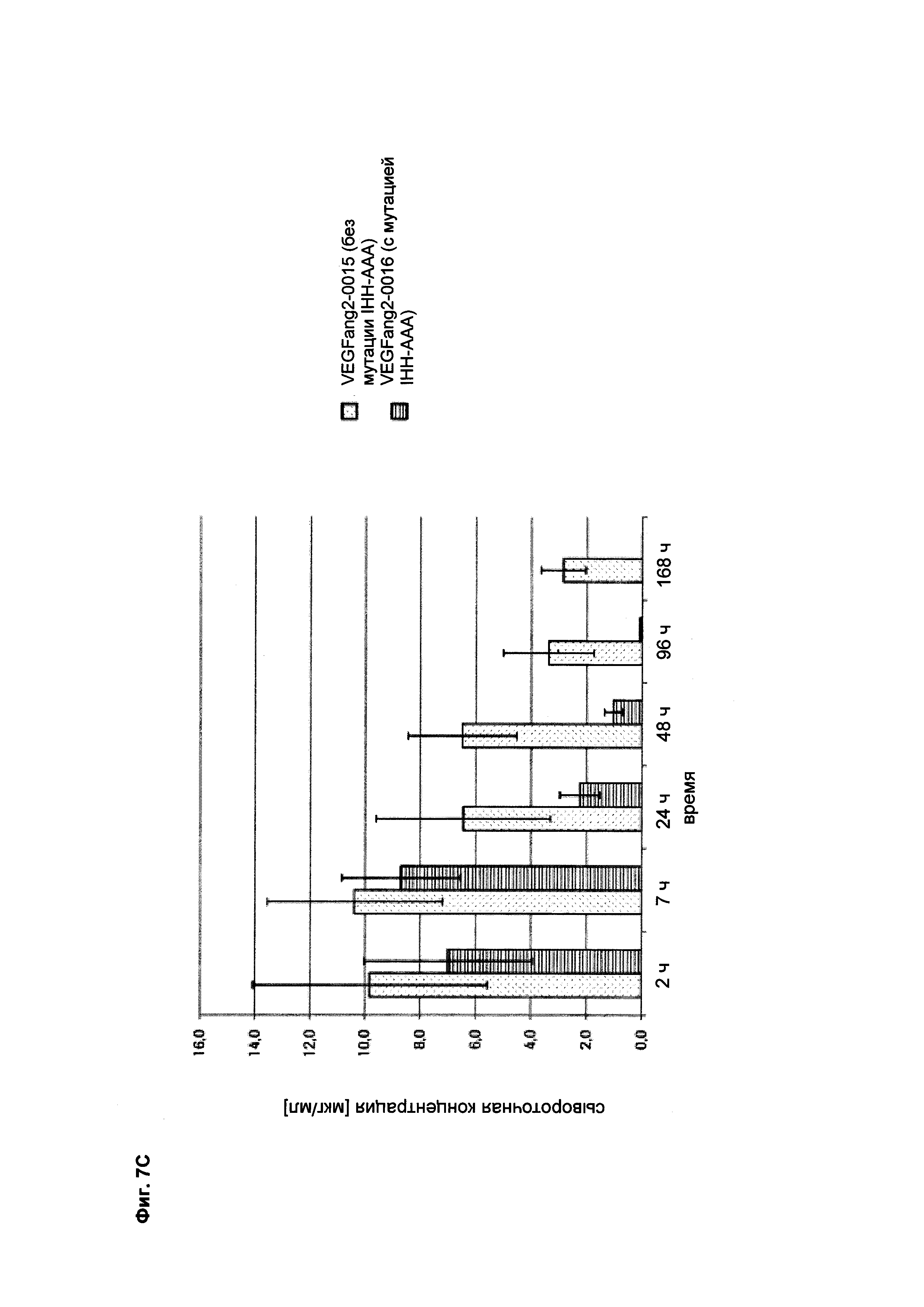














Препарат антитела
Производные гетероарилпирролидинил- и пиперидинилкетона
Производные изоксазоло-пиридина
Производные имидазопиридина или имидазопиримидина в качестве ингибиторов фосфодиэстеразы 10а
Ингибиторы jnk
Бензопирановые и бензоксепиновые ингибиторы рi3k и их применение
Арилциклогексилэфиры дигидротетраазабензоазуленов для применения в качестве антагонистов рецептора вазопрессина v1a
Пуриновые соединения, ингибирующие рi3к, и способы применения
Алкилциклогексиловые эфиры дигидротетраазабензоазуленов
Способ синтеза производных амино-метилтетралина
Антитела против ангиопоэтина-2 человека
Биспецифические двухвалентные антитела анти-vegf/анти-ang-2
Fc варианты антитела
Антигенсвязывающие белки
Антитела, не содержащие fc-фрагмента, включающие два fab-фрагмента, и способы их применения
Аффинная хроматография с применением fc-рецепторов
Биспецифические анти-vegf/анти-ang-2 антитела
Способ получения растворимого fcr в виде fc-гибрида с инертной fc-областью иммуноглобулина и их применения
Биспецифические антигенсвязывающие молекулы
In vitro прогнозирование времени полужизни антител in vivo

