Результат интеллектуальной деятельности: КОНСТРУКТ И СПОСОБ ДЛЯ ЭКСПРЕССИИ ТРАНСГЕНОВ С ИСПОЛЬЗОВАНИЕМ ДВУНАПРАВЛЕННОГО КОНСТИТУТИВНОГО ПРОМОТОРА BRASSICA
Вид РИД
Изобретение
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Эта заявка заявляет приоритет предварительной заявки США №61/656,634, поданной 7 июня 2012 года, описание которой включено здесь специально посредством ссылки в полном объеме.
ВКЛЮЧЕНИЕ ПОСРЕДСТВОМ ССЫЛКИ МАТЕРИАЛА, ПРЕДСТАВЛЕННОГО В ЭЛЕКТРОННОМ ВИДЕ
Посредством ссылки в полном объеме включен считываемый компьютером список последовательностей, представленный одновременно здесь и идентифицированный следующим образом: one 78,835 byte ASCII (text) file named «70136_ST25.txt», created on May 13, 2013.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение, в общем, относится к области молекулярной биологии и, более конкретно, к области стабильной экспрессии множественных генов в трансгенных растениях.
УРОВЕНЬ ТЕХНИКИ
Многие виды растений способны трансформироваться трансгенами из других видов для введения агрономически желательных признаков или характеристик, например, улучшения пищевой (питательной) ценности, увеличения урожая, придания устойчивости к насекомым-вредителям или устойчивости к болезням, увеличения толерантности к стрессу, вызванному засухой, и увеличения плодоовощных качеств (таких как пигментация и рост), передачи устойчивости к гербицидам, позволяя получать промышленным образом используемые соединения и/или материалы из этого растения и/или позволяя производить фармацевтические лекарственные средства. Введение трансгенов в клетки растений и последующее восстановление фертильных трансгенных растений, которые содержат стабильно интегрированную копию этого трансгена, может быть использовано для получения трансгенных растений, которые обладают желаемыми признаками.
Контроль и регуляция экспрессии генов могут осуществляться посредством многочисленных механизмов. Инициация транскрипции гена является доминирующим регулирующим механизмом экспрессии гена. Инициация транскрипции обычно регулируется полинуклеотидными последовательностями, локализованными в 5'-фланкирующей области или в области, расположенной выше транскрибирумого гена. Эти последовательности вместе называются промоторами и распределяются по категориям как регуляторный элемент гена. Промоторы в растениях, которые были клонированы и широко использовались как для научно-исследовательской работы, так и для биотехнологического применения, являются обычно однонаправленными, управляющими только одним геном, который был слит на его 3'-конце (т.е. ниже). См., например, Xie et al. (2001) Nat. Biotechnol. 19(7):677-9; США патент № 6,388,170.
Дополнительные регуляторные элементы гена включают в себя последовательности, которые взаимодействуют со специфическими ДНК-связывающими факторами. Эти мотивы последовательности называют иногда cis-элементами, и они обычно являются зависимыми от положения и зависимыми от ориентации, хотя они могут быть найдены 5'- или 3'- относительно кодирующей ген последовательности, или в интроне. Такие cis-элементы, с которыми связаны ткане-специфичные или развитие-специфичные факторы транскрипции, индивидуально или в комбинации, могут определять пространственно-временной паттерн промотора на транскрипционном уровне. Эти cis-элементы широко варьируются в типе контроля, который они оказывают на функционально связанные гены. Некоторые элементы действуют для увеличения транскрипции функционально-связанных генов в ответ на реакции окружающей среды (например, температуру, влажность и скарификацию). Другие cis-элементы могут реагировать на другие сигналы (например, прорастание, созревание семян и зацветание) или на пространственную информацию (например, специфичность ткани). См., например, Langridge et al. (1989) Proc. Natl. Acad. Sci. USA 86:3219-23.
Часто необходимым является введение множественных генов в растения для метаболической инженерии и стекинга генов («стеллажных» генов), причем эти гены часто регулируются идентичными или гомологичными промоторами. Однако сайленсинг генов на основе гомологии (HBGS) повышается, по-видимому, когда множественные введенные трансгены имеют гомологичные промоторы, запускаемые ими. См., например, Mol et al. (1989) Plant Mol. Biol. 13:287-94. Сообщалось, что HBGS встречаются широко в трансгенных растениях. См., например, Vaucheret and Fagard (2001) Trends Genet. 17:29-35. Предлагались несколько механизмов для объяснения феномена of HBGS, все из которых включают в себя признак, что гомология последовательности в этом промоторе запускает механизмы клеточного узнавания, которые приводят к сайленсингу повторяющихся генов. См., например, Matzke and Matzke (1995) Plant Physiol. 107:679-85; Meyer and Saedler (1996) Ann. Rev. Plant Physiol. Plant Mol. Biol. 47:23-48; Fire (1999) Trends Genet. 15:358-63; Hamilton and Baulcombe (1999) Science 286:950-2; и Steimer et al. (2000) Plant Cell 12:1165-78.
Стратегии для избегания HBGS в трансгенных растениях часто включают в себя развитие различных промоторов, которые являются функционально эквивалентными, но имеют минимальную гомологию последовательности. Таким образом, остается потребность в конструктах и способах для стабильной экспрессии множественных трансгенов эффективно с минимальным риском в отношении рекомбинации или потери трансгенов в результате бридинга или множественных генераций в трансгенных растениях.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Обеспечены конструкты и способы для экспрессии множественных генов в клетках растений и/или тканей растений с использованием раскрытого двунаправленного промотора из Brassica napus или двунаправленного конститутивного промотора Brassica (BBCP). Эти обеспеченные конструкты содержат по меньшей мере один такой двунаправленный промотор, связанный с экспрессионными кассетами множественных генов, где каждая из этих экспрессионных кассет генов содержит по меньшей мере один трансген. В некоторых вариантах осуществления, эти обеспеченные конструкты и способы делают возможной экспрессию от двух до двадцати генов.
В одном аспекте обеспечен конструкт нуклеиновой кислоты для экспрессии множественных генов в клетках и/или тканях растений. Указанный конструкт нуклеиновой кислоты содержит: (а) двунаправленный промотор, содержащий последовательность нуклеотидов, имеющую по меньшей мере 80%-ную идентичность SEQ ID NO: 1; и (b) две экспрессионные кассеты генов на противоположных концах этого двунаправленного промотора.
В одном варианте осуществления, указанный двунаправленный промотор содержит по меньшей мере один энхансер. В другом варианте осуществления, указанный двунаправленный промотор не содержит энхансера. В другом варианте осуществления, указанный конструкт нуклеиновой кислоты содержит бинарный вектор для трансформации растений. В другом варианте осуществления, указанный конструкт нуклеиновой кислоты содержит бинарный вектор для Agrobacterium-опосредованной трансформации. В другом варианте осуществления, указанный двунаправленный промотор содержит по меньшей мере один интрон. В другом варианте осуществления, указанный двунаправленный промотор содержит по меньшей мере одну 5’-нетранслируемую область. В одном варианте осуществления, указанный двунаправленный промотор содержит нуклеотидную последовательность, выбранную из SEQ ID NO: 2 или 3. В другом варианте осуществления, указанный двунаправленный промотор, содержит нуклеотидную последовательность, имеющую по меньшей мере 85%, 90%, 95% или 100% идентичность SEQ ID NO: 1. В другом варианте осуществления указанный двунаправленный промотор содержит нуклеотидную последовательность, выбранную из SEQ ID NO: 1, 22-25 или их комплементов. В дополнительном или альтернативном варианте осуществления, указанный двунаправленный промотор содержит нуклеотидную последовательность, выбранную из SEQ ID NO: 1, 22-24 или их комплементов. В одном дополнительном или альтернативном варианта осуществления, указанный двунаправленный промотор содержит нуклеотидную последовательность, выбранную из SEQ ID NO: 1, 22-23 или их комплементов. В одном дополнительном или альтернативном вариантах осуществления, указанный двунаправленный промотор содержит нуклеотидную последовательность, выбранную из SEQ ID NO: 1, 22 или их комплементов.
В одном варианте осуществления, по меньшей мере одна из экспрессионных кассет гена содержит два или более генов, связанных через ген-переключатель трансляции. В другом варианте осуществления, обе эти кассеты экспрессии генов содержат два или более генов, связанных через ген-переключатель трансляции. В дополнительном или альтернативном варианте осуществления, указанный ген-переключатель трансляции выбран из группы, состоящей из внутреннего сайта вхождения в рибосому (IRES), альтернативного сайта сплайсинга, сайта связывания рибозима, полинуклеотидной последовательности, кодирующей 2А-подобный пептид, полинуклеотидной последовательности, кодирующей интеин, полинуклеотидной последовательности, кодирующей сайт расщепления протеазы, и их комбинаций. В дополнительном или альтернативном варианте осуществления, указанный ген-переключатель содержит цис-действующий элемент гидролазы (CHYSEL). В одном дополнительном варианте осуществления, этим CHYSEL является последовательность пептида 2A или 2A-подобного пептида. В другом варианте осуществления, ген выше переключателя трансляции не содержит стоп-кодона трансляции.
В одном варианте осуществления, указанный конструкт нуклеиновой кислоты содержит по меньшей мере один трансген. В другом варианте осуществления, указанный конструкт нуклеиновой кислоты позволяет или допускает экспрессию по меньшей мере четырех генов. В дополнительном варианте осуществления все четыре гена являются трансгенами. В другом варианте осуществления, указанный конструкт нуклеиновой кислоты позволяет экспрессию от трех до двадцати генов. В другом варианте осуществления, указанный конструкт нуклеиновой кислоты позволяет экспрессию от четырех до восьми генов. В одном дополнительном или альтернативном варианте осуществления, указанные гены являются трансгенами. В другом варианте осуществления, по меньшей мере одна экспрессионная кассета генов содержит полинуклеотидную последовательность, кодирующую слитый белок. В другом варианте осуществления, указанный слитый белок содержит три - пять генов. В другом варианте осуществления, обе эти экспрессионные кассеты не содержат гена EPSPS или паралога.
В другом аспекте, обеспечен конструкт нуклеиновой кислоты, содержащий регуляторный элемент, применимый для терминации экспрессии одного или множественных генов в клетках и/или тканях растений. Этот регуляторный элемент содержит 3'-нетранслируемую область паралога А (UTR) или поли А-область, которые могут быть слиты с 3'-концом трансгена. В одном варианте осуществления, 3'UTR паралога A содержит функциональную последовательность полиаденилирования, которая применима для терминации и регуляции транскрипции и трансляции. В дополнительном или альтернативном варианте осуществления, этот регуляторный элемент содержит полинуклеотидную последовательность, имеющую по меньшей мере 80%-, 85%-, 90%-, 95%- или 100%-ную идентичность SEQ ID NO: 26 или ее комплемента. В одном дополнительном варианте осуществления, этот регуляторный элемент содержит полинуклеотидную последовательность SEQ ID NO: 26 или ее комплемент.
В другом аспекте, обеспечен способ генерирования трансгенного растения, предусматривающий трансформацию клетки растения конструктом нуклеиновой кислоты, обеспеченным здесь. В другом аспекте, обеспечен способ генерирования трансгенной клетки, предусматривающий трансформацию указанной клетки конструктом нуклеиновой кислоты, обеспеченной здесь. В другом аспекте, обеспечена клетка растения, содержащая конструкт нуклеиновой кислоты, обеспеченный здесь. В одном дополнительном или альтернативном варианте осуществления, указанный конструкт нуклеиновой кислоты стабильно трансформируют в указанную клетку растения. В другом аспекте, обеспечено трансгенное растение или семя, содержащие конструкт нуклеиновой кислоты, обеспеченный здесь. В одном дополнительном или альтернативном варианте осуществления, этот конструкт нуклеиновой кислоты стабильно трансформируют в клетки этого трансгенного растения или семени. В дополнительном варианте осуществления, указанное трансгенное растение является двудольным растением. В другом дополнительном варианте осуществления, указанное трансгенное растение является однодольным растением. В другом аспекте, обеспечен способ для экспрессии множественных генов в клетках и/или тканях растений, предусматривающий введение в эти клетки и/или ткани растения конструкта нуклеиновой кислоты, обеспеченного здесь. В одном дополнительном или альтернативном варианте осуществления, эти клетки и/или ткани растения стабильно трансформируют конструктом нуклеиновой кислоты, обеспеченным здесь. В другом аспекте, обеспечен бинарный вектор для Agrobacterium-опосредуемой трансформации. Этот бинарный вектор содержит конструкт нуклеиновой кислоты, обеспеченный здесь. В другом аспекте, обеспечено использование двунаправленного промотора, обеспеченного здесь, для экспрессии множественных трансгенов в растениях. В одном варианте осуществления, этот двунаправленный промотор, содержит нуклеотидную последовательность, имеющую по меньшей мере 80%-, 85%-, 90%-, 95%- или 100%-ную идентичность SEQ ID NO: 1. В другом аспекте, обеспечено использование двунаправленного промотора, обеспеченного здесь, в производстве трансгенных растений или семян. В одном варианте осуществления, этот двунаправленный промотор содержит нуклеотидную последовательность, имеющую по меньшей мере 80%-, 85%-, 90%-, 95%- или 100%-ную идентичность SEQ ID NO: 1.
В другом аспекте, обеспечен конструкт нуклеиновой кислоты, содержащий по меньшей мере одну последовательность интрона Brassica в трансгенных клетках и/или тканях растений. В одном варианте осуществления, последовательность интрона Brassica выбрана из SEQ ID NO: 27-33. В другом аспекте обеспечено использование по меньшей мере одной последовательности интрона Brassica в приготовлении трансгенных растений или семян. В одном варианте осуществления, последовательность интрона Brassica выбрана из SEQ ID NO: 27-33.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ И ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Фиг. 1А показывает последовательность 739 nt центрального двунаправленного промотора Brassica, двунаправленного конститутивного промотора (BBCP) (SEQ ID NO: 1).
Фиг. 1В показывает модифицированную последовательность 1226 nt BBCP (SEQ ID NO: 2), где нуклеотидная последовательность «gg» добавлена для введения сайта расщепления рестрикционного фермента (рестриктазы).
Фиг.1C показывает SEQ ID NO: 3, которая является обратным комплементом SEQ ID NO: 2, где добавленная последовательность «gg» SEQ ID NO: 2 показана как последовательность «cc» SEQ ID NO: 3.
Фиг. 2А показывает идентификацию BBCP из генома Brassica napus, где экспрессия гена паралога А запускается BBCP. На противоположном конце BBCP от гена паралога А EPSPS, также идентифицирован предложенный неизвестный ген.
Фиг. 2В показывает полинуклеотидную последовательность нативной геномной последовательности (SEQ ID NO: 4), содержащую (1) предположенный неизвестный ген, (2) BBCP и (3) ген паралога А EPSPS.
Фиг. 3А дополнительно показывает предсказанную последовательность белка этого предложенного неизвестного гена (SEQ ID NO: 5) и
Фиг. 3В показывает соответствующую кодирующую последовательность (SEQ ID NO: 6).
Фиг. 4А показывает частичную последовательность гена паралога А EPSPS (SEQ ID NO: 7), которая является такой же, как SEQ ID NO: 10 US 2009/0205083.
Фиг. 4В показывает полную последовательность гена паралога А EPSPS.
Фиг. 4С дополнительно показывает последовательность белка паралога А EPSPS (SEQ ID NO: 9), и
Фиг. 4D показывает соответствующую кодирующую последовательность паралога А EPSPS (SEQ ID NO: 10).
Фиг. 5А показывает последовательность гена паралога В EPSPS (SEQ ID NO: 11), которая является такой же, что и SEQ ID NO: 11 US 2009/0205083.
Фиг. 5В дополнительно показывает последовательность белка паралога В EPSPS (SEQ ID NO: 12), и
Фиг. 5С показывает соответствующую кодирующую последовательность паралога В EPSPS (SEQ ID NO: 13).
Фиг. 6А показывает последовательность гена паралога С EPSPS (SEQ ID NO: 14), которая является такой же, что и SEQ ID NO: 12 US 2009/0205083.
Фиг. 6В дополнительно показывает последовательность белка паралога С EPSPS (SEQ ID NO: 15), и
Фиг. 6С показывает соответствующую кодирующую последовательность паралога С EPSPS (SEQ ID NO: 16).
Фиг. 7А показывает последовательность гена паралога Е EPSPS (SEQ ID NO: 17), которая является такой же, что и SEQ ID NO: 14 US 2009/0205083.
Фиг. 7В дополнительно показывает последовательность белка паралога Е EPSPS (SEQ ID NO: 18), и Фиг. 7С показывает соответствующую кодирующую последовательность паралога С EPSPS (SEQ ID NO: 19).
Фиг. 8 показывает сопоставление примерных последовательностей среди белковых последовательностей паралога А EPSPS (SEQ ID NO: 9), паралога В EPSPS B (SEQ ID NO: 12), EPSPS паралога C (SEQ ID NO: 15) и паралога Е EPSPS (SEQ ID NO: 18).
Фиг. 9 показывает примерную последовательность из pDAB100331 (SEQ ID NO: 20), содержащую экспрессионные кассеты генов для GUS и GFP на противолежащих концах BBCP. Экспрессия как GUS, так и GFP запускается BBCP.
Фиг. 10 показывает примерную последовательность из pDAB100331 (SEQ ID NO: 21), содержащую экспрессионные кассеты генов для GUS и GFP на противолежащих концах BBCP. Экспрессия как GUS, так и GFP запускается BBCP.
Фиг. 11A и 11B показывают альтернативные последовательности BBCP, включающие в себя SEQ ID NO: 22-25.
Фиг. 12 показывает репрезентативные карты плазмид pDAB100331 и pDAB100333.
Фиг. 13 показывает репрезентативные карты плазмид pDAB108710 и pDAB108711.
Фиг. 14 показывает примерную последовательность гена 3’UTR паралога А EPSPS (SEQ ID NO: 26) и семь последовательностей интронов паралога А (SEQ ID No: 27-33).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Развитие трансгенных продуктов становится все более сложным, что требует стекинга множественных трансгенов в единый локус. Традиционно каждый трансген обычно требует уникального промотора для экспрессии, так что множественные промоторы требуются для экспрессии различных трансгенов в одной стопке генов. Кроме этого, увеличение размера стопки генов часто приводит к повторяемому использованию одного и того же промотора для получения сходных уровней паттернов экспрессии различных трансгенов для экспрессии единственного полигенного признака. Известно, что мультигенные конструкты, запускаемые одним и тем же промотором, вызывают сайленсинг генов, делая таким образом трансгенные продукты менее эффективными в данной области. Избыток сайтов связывания фактора транскрипции (TF) вследствие повторения промотора может вызывать истощение эндогенных TF, приводя к инактивации транскрипции. Этот сайленсинг трансгенов будет вероятно нежелательным образом влиять на производительность трансгенного растения, полученного для экспрессии трансгенов. Повторяющиеся последовательности в трансгене могут приводить к интра-локусной гомологичной рекомбинации, приводящей к реаранжировкам полинуклеотидов.
Обеспечены способы и конструкты, использующие двунаправленный конститутивный промотор Brassica (BBCP) для экспрессии трансгенов в растении. Также обеспечены способы и конструкты, объединяющие систему двунаправленного промотора с бицистронной организацией генов на каждом одном конце или на обоих концах промотора, например, с использованием последовательности 2A из вируса Thosea asigna. Белок 2A, который имеет в длину только 16-20 аминокислот, расщепляет этот полипротеин на его собственном карбоксил-конце. Это «саморасщепление» или свойство «пропуска (скачка) рибосомы» пептида 2A или 2A-подобного пептида могут быть использованы для процессирования искусственных полипротеинов, продуцируемых в трансгенных растениях. В одном варианте осуществления, гены Cry34 и Cry35 сливают в другую экспрессионную кассету генов, где гены GFP (или YFP или PhiYFP) и AAD1 слиты в другую экспрессионную кассету генов (с единственной открытой рамкой считывания (ORF) с копией гена белка 2А, расположенной между этими двумя генами в каждой комбинации). Например, каждая из этих экспрессионных кассет (или пар генов) могут быть помещены на любом конце двунаправленного промотора для запуска 4 трансгенов с использованием единственного промотора. Таким образом, конструкты и способы, обеспеченные здесь, применимы для избегания повторяемого использования одного и того же промотора и значимого уменьшения размера коммерческих конструктов. Кроме того, регуляция четырех или более генов одним промотором также обеспечивает способность ко-экспрессии генов, контролирующих единственный полигенный признак.
Некоторые раскрытые аббревиатуры перечислены в таблице 1.
|
Промоторы растений, используемые для научных исследований или биотехнологического применения, обычно являются однонаправленными, направленными только на один ген, который был слит на его 3'-конце (по ходу транскрипции). Часто является необходимым введение множественных генов в растения для метаболической инженерии и стекинга признака, и, следовательно, множественные промоторы обычно являются необходимыми в будущих трансгенных культурах для запуска экспрессии множественных генов. Желательной является разработка стратегий, которые могут экономить количество используемых промоторов и делать возможной одновременную совместно регулируемую экспрессию для укладки (стекинга) генов. В одном варианте осуществления, обеспеченные двунаправленные промоторы могут запускать транскрипцию множественных единиц транскрипции, включающих в себя последовательности RNAi, искусственные miRNA или имеющие «шпилечную петлю» последовательности РНК.
В данном контексте артикли, «a», «an» и «the» включают в себя множественные ссылки, если этот контекст не диктует ясно и недвусмысленно другое.
В данном контексте, фраза «обратное скрещивание» относится к процессу, в котором селекционер скрещивает гибридного потомка обратно с одним из родителей, например, гибрида первого поколения F1 с одним из родительских генотипов гибрида F1.
В данном контексте, фраза «интрон» относится к любой последовательности нуклеиновой кислоты, содержащейся в гене (или экспрессируемой представляющей интерес последовательности), которая транскрибируется, но не транслируется. Интроны включают в себя нетранслируемую последовательность нуклеиновой кислоты в экспрессируемой последовательности ДНК, а также соответствующую последовательность в молекулах РНК, транскрибируемых из них.
Этот обеспеченный конструкт может также содержать последовательности, которые усиливают трансляцию и/или стабильность мРНК, такие как интроны. Примером одного такого интрона является первый интрон гена II варианта гистона H3.III Arabidopsis thaliana или любую другую обычно известную интронную последовательность. Chaubet et al. Journal of Molecular Biology, 225:569-574 (1992). В данной области известно, что интроны могут быть использованы в комбинации с последовательностями промоторов для усиления трансляции и/или стабильности мРНК.
В данном контексте, фраза «5' нетранслируемая область» или «5’UTR» относится к нетранслируемому сегменту в 5' конце пре-мРНК или зрелых мРНК. Например, на зрелых мРНК, этот 5’UTR обычно захватывает на его 5' конце 7-метилгуанозиновый кэп и участвует во многих процессах, таких как сплайсинг, полиаденилирование, экспорт мРНК в направлении к цитоплазме, идентификация 5' конца этой мРНК посредством аппарата трансляции и защита мРНК против деградации.
В данном контексте, фраза «3' нетранслируемая область» или «3’UTR» относится к нетранслируемому сегменту в 3'-конце пре-мРНК или зрелых мРНК. Например, на зрелых мРНК эта область захватывает поли (A) хвост и, как известно, играет многие роли в стабильности мРНК, инициации трансляции, экспорте мРНК.
В данном контексте, фраза «сигнал полиаденилирования» относится к последовательности нуклеиновой кислоты, присутствующей в транскриптах мРНК, что позволяет этим транскриптам мРНК, в присутствии поли (А)-полимеразы, быть полиаденилированными на сайтах полиаденилирования, например, расположенных в 10-30 основаниях ниже поли (А)-сигнала. Многие сигналы полиаденилирования известны в данной области и применимы для данного изобретения. Примеры включают в себя сигнал полиаденилирования варианта гормона роста человека, поздний сигнал полиаденилирования SV40 (вируса обезьяны) и сигнал полиаденилирования бычьего гормона роста.
В данном контексте, фраза «выделенный» относится к биологическому компоненту (в том числе нуклеиновой кислоте или белку), который был по существу отделен, получен отдельно или очищен от других биологических компонентов в клетке организма, в котором этот компонент встречается в природе (т.е. других хромосомных и внехромосомных ДНК и РНК и белков), хотя и с вызыванием химического или функционального изменения в этом компоненте (то есть, нуклеиновая кислота может быть выделена из хромосомы разрушением химических связей, соединяющих эту нуклеиновую кислоту с остальной ДНК в хромосоме). Молекулы нуклеиновых кислот и белков, которые могут быть «изолированы», включают в себя молекулы нуклеиновых кислот и белков, очищенные стандартными способами очистки. Фраза «выделенные» включает в себя также нуклеиновые кислоты и белки, полученные рекомбинантной экспрессией в клетке-хозяине, а также химически синтезированные молекулы нуклеиновых кислот, белков и пептидов.
В данном контексте, фраза «экспрессия генов» относится к процессу, при помощи которого кодируемая информация транскрипционной единицы (в том числе, например, геномной ДНК) превращается в функциональную, нефункциональную или структурную часть клетки, часто с включением синтеза белка. На экспрессию генов могут влиять внешние факторы; например, подвергание клетки, ткани или организма действию агента, который увеличивает или уменьшает экспрессию гена. Экспрессия гена может также регулироваться где-нибудь в пути от ДНК до РНК к белку. Регуляция экспрессии гена осуществляется, например, через контроли, действующие на транскрипцию, трансляцию, транспорт и процессинг РНК, деградацию промежуточных молекул, таких как мРНК, или через активацию, инактивацию, компартментализацию или деградацию молекул специфического белка, после их получения, или их комбинаций. Экспрессия гена может быть измерена на уровне РНК или уровне белка любым способом, известным в данной области, включающим в себя, без ограничения, Нозерн-блоттинг, RT-PCR, Вестерн-блот, или анализом (анализами) активности белка in vitro, in situ или in vivo.
В данном контексте, фраза «сайленсинг гена на основе гомологии» (HBGS) относится к родовому термину, который включает в себя как транскрипционный сайленсинг гена, так и пост-транскрипционный сайленсинг гена. Сайленсинг локуса-мишени несвязанным локусом связывания может происходить из ингибирования транскрипции (транскрипционный сайленсинг гена; TGS) или деградации мРНК (пост-транскрипционный сайленсинг гена; PTGS), вследствие продуцирования двухцепочечной РНК (dsRNA), соответствующей промотору или транскрибируемым последовательностям, соответственно. Участие отдельных клеточных компонентов в каждом процессе предполагает, что dsRNA-индуцируемые TGS и PTGS, происходят, по-видимому, из диверсификации старого общего механизма. Однако строгое сравнение TGS и PTGS было трудно получить, так как оно обычно основано на анализе индивидуальных локусов сайленсинга. Было описано, что один локус трансгена запускает как TGS, так и PTGS, вследствие продуцирования dsRNA, соответствующей промотору и транскрибируемым последовательностям различных генов-мишеней. См., например, Mourrain et al. (2007) Planta 225:365-79. Вероятно, что siRNA являются фактическими молекулами, которые запускают TGS и PTGS на гомологичных последовательностях: siRNA могли бы в этой модели запускать сайленсинг и метилирование гомологичных последовательностей в cis и в trans через распространение метилирования трансгенных последовательностей в эндогенный промотор.
В данном контексте, фраза «молекула нуклеиновой кислоты» (или «нуклеиновая кислота» или «полинуклеотид») относится к полимерной форме нуклеотидов, которая может включать в себя как смысловые, так и антисмысловые цепи РНК, кДНК, геномной ДНК и синтетические формы и смешанные полимеры описанные выше. Один нуклеотид может быть рибонуклеотидом, дезоксирибонуклеотидом или модифицированной формой нуклеотида любого типа. «Молекула нуклеиновой кислоты» в данном контексте является синонимом «нуклеиновой кислоты» и «полинуклеотида». Молекула нуклеиновой кислоты обычно имеет длину по меньшей мере 10 оснований, если нет других указаний. Этот термин может также относиться к молекуле РНК или ДНК неопределенной длины. Этот термин включает в себя одноцепочечные и двухцепочечные формы ДНК. Молекула нуклеиновой кислоты может включать в себя каждый или оба природно-встречающихся и модифицированных нуклеотидов, связанных вместе природно-встречающимися и/или не встречающимися в природе нуклеотидными связями.
Молекулы нуклеиновых кислот могут быть модифицированы химически или биохимически или могут содержать неприродные или дериватизованные нуклеотидные основания, а также будут легко оцениваться квалифицированными в данной области специалистами. Такие модификации включают в себя, например, метки, метилирование, замену одного или нескольких природно-встречающихся нуклеотидов аналогом, межнуклеотидные модификации (например, незаряженные связи: например, метилфосфонаты, фосфотриэфиры, фосфорамидаты, карбаматы и т.д.; заряженные связи: например, фосфоротиоаты, фосфородитиоаты и т.д.; свисающие части: например, пептиды; интеркаляторы: например, акридин, псорален и т.д.; хелаторы; алкиляторы; и модифицированные связи; например, альфа-аномерные нуклеиновые кислоты и т.д.). Термин «молекула нуклеиновой кислоты» включает в себя также любую топологическую конформацию, в том числе одноцепочечную, двухцепочечную, частично дуплексную, триплексную, шпилечную, кольцевую и висячую конформации.
Транскрипция протекает от 5' к 3' вдоль цепи ДНК. Это означает, что РНК образуется последовательным добавлением рибонуклеотид-5'-трифосфатов к 3'-концу растущей цепи (с необходимым элиминированием пирофосфата). В каждой линейной или в кольцевой молекуле нуклеиновой кислоты, отдельные элементы (например, конкретные нуклеотидные последовательности) могут называться как находящиеся «выше» относительно дополнительного элемента, если они связаны или могли бы быть связаны с той же самой нуклеиновой кислотой в направлении 5' от этого элемента. Подобным образом, отдельные элементы могут находиться «ниже» относительно дополнительного элемента, если они связаны или могли бы быть связаны с той же самой нуклеиновой кислотой в направлении 3' от этого элемента.
В данном контексте, фраза «положение основания» относится к местоположению этого основания или нуклеотидного остатка в определенной нуклеиновой кислоте. Эта определенная нуклеиновая кислота может быть определена сопоставлением (см. ниже) со ссылочной нуклеиновой кислотой.
В данном контексте, фраза «гибридизация» относится к процессу, в котором олигонуклеотиды и их аналоги гибридизуются водородным связыванием, которое включает в себя связывание Уотсона-Крика, Хугстиновское спаривание или обращенное водородное Хугстиновское спаривание, между комплементарными основаниями. Обычно, молекулы нуклеиновых кислот состоят из азотистых оснований, которые являются либо пиримидинами (цитозин (C), урацил (U) и тимин (T)), либо пуринами (аденин (A) и гуанин (G)). Эти азотистые основания образуют водородные связи между пиримидином и пурином, и это связывание пиримидина с пурином называют «спариванием оснований». Более конкретно, А будет водородной связью с Т или U, а G будет связью с С. Термин «комплементарный» относится к спариванию оснований, которое осуществляется между двумя разными последовательностями нуклеиновых кислот или двумя разными областями одной и той же последовательности нуклеиновой кислоты.
В данном контексте, фразы «специфически гибридизируемые» и «специфически комплементарные» относятся к достаточной степени комплементарности, так что осуществляется стабильное и специфическое связывание между этим олигонуклеотидом и ДНК- или РНК-мишенью. Этот олигонуклеотид не должен быть на 100% комплементарным его последовательности-мишени, чтобы быть специфически гибридизируемым. Олигонуклеотид является специфически гибридизируемым, когда связывание этого олигонуклеотида с ДНК-мишенью или РНК мешает нормальной функции ДНК- или РНК-мишени, и имеется достаточная степень комплементарности для избегания неспецифического связывания этого олигонуклеотида с последовательностями, не являющимися мишенями, при условиях, в которых желаемым является специфическое связывание, например, при физиологических условиях в случае анализов или систем in vivo. Такое связывание называют специфической гибридизацией.
Условия гибридизации, приводящие к конкретным степеням строгости, будут варьироваться в зависимости от характера выбранного способа гибридизации и состава и длины гибридизующихся последовательностей нуклеиновых кислот. Обычно, температура гибридизации и ионная сила (особенно концентрация Na+ и/или Mg2+) буфера гибридизации будут способствовать строгости гибридизации, хотя периоды времени промывок также влияют на строгость. Расчеты в отношении условий гибридизации, требуемых для получения конкретных степеней строгости, обсуждаются в Sambrook et al. (ed.), Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, chs. 9 и 11.
В данном контексте, фраза «строгие условия» включает в себя условия, при которых гибридизация будет осуществляться только в случае, если имеется меньшее, чем 50%, ошибочное спаривание между молекулой гибридизации и ДНК-мишенью. «Строгие условия» включают в себя дополнительно конкретные уровни строгости. Таким образом, в данном контексте, условиями «средней строгости» являются условия, при которых молекулы с более чем 50%, ошибочного спаривания последовательности, не будут гибридизироваться; условиями «высокой строгости» являются условия, при которых последовательности с более, чем 20%, ошибочного спаривания не будут гибридизироваться; и условиями «очень высокой строгости» являются условия, при которых более, чем 10% ошибочного спаривания, не будут гибридизироваться.
В конкретных вариантах осуществления строгие условия могут включать в себя гибридизацию при 65°C, с последующими промываниями при 65°C с использованием 0,1× SSC/0,1% SDS в течение 40 минут.
Далее следуют репрезентативные, не ограничивающие гибридизацию условия:
Очень высокая строгость: Гибридизация в буфере 5× SSC при 65°C в течение 16 часов; дважды в буфере 2× SSC при комнатной температуре в течение 15 минут каждый раз; и промывание дважды в буфере 0,5× SSC при 65°C в течение 20 минут каждый раз.
Высокая строгость: Гибридизация в буфере 5-6× SSC при 65-70°C в течение 16-20 часов; промывание дважды в буфере 2× SSC при комнатной температуре в течение 5-20 минут каждый раз; и промывании дважды в буфере 1× SSC при 55-70°C в течение 30 минут каждый раз.
Средняя строгость: Гибридизация в буфере 6× SSC при комнатной температуре до 55°C в течение 16-20 часов; промывание дважды в буфере 2×-3× SSC при комнатной температуре до 55°C в течение 20-30 минут каждый раз.
В конкретных вариантах осуществления, специфически гибридизируемые молекулы нуклеиновых кислот могут оставаться связанными при условиях очень высокой строгости. В этих и дополнительных вариантах осуществления, специфически гибридизируемые молекулы нуклеиновых кислот могут оставаться связанными при условиях высокой строгости. В этих и дополнительных вариантах осуществления, специфически гибридизируемые молекулы нуклеиновых кислот могут оставаться связанными при условиях умеренной строгости.
В данном контексте, фраза «олигонуклеотид» относится к короткому полимеру нуклеиновой кислоты. Олигонуклеотиды могут быть образованы расщеплением более длинных сегментов нуклеиновых кислот или полимеризацией индивидуальных предшественников нуклеотидов. Автоматизированные синтезаторы делают возможным синтез олигонуклеотидов до нескольких сотен пар оснований в длину. Поскольку олигонуклеотиды могут связываться с комплементарной нуклеотидной последовательностью, они могут использоваться в качестве зондов для детектирования ДНК или РНК. Олигонуклеотиды, состоящие из ДНК (олигодезоксирибонуклеоиды), могут быть использованы в ПЦР, способе для амплификации малых ДНК-последовательностей. В ПЦР, этот олигонуклеотид обычно называется «праймером», который позволяет ДНК-полимеразе удлинять этот олигонуклеотид и реплицировать комплементарную цепь.
В данном контексте, фраза «идентичность последовательности» или «идентичность» относится к контексту, где две последовательности нуклеиновых кислот или полипептидные последовательности могут называться остатками в двух последовательностях, которые являются одними и теми же при сопоставлении для максимального соответствия на протяжении установленного окна сравнения.
В данном контексте, фраза «процент идентичности последовательности» относится к величине, определяемой сравнением двух оптимально сопоставленных последовательностей (например, последовательностей нуклеиновых кислот и аминокислотных последовательностей) на протяжении окна сравнения, где часть этой последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) при сравнении со ссылочной последовательностью (которая не содержит добавлений или делеций) для оптимального сопоставления этих двух последовательностей. Процент рассчитывают определением количества положений, в которых идентичный нуклеотидный остаток или аминокислотный остаток встречается в обеих последовательностях, для получения количества совместимых положений, делением этого количества совместимых положений на общее количество положений в окне сравнения, и умножением этого результата на 100 для получения процента идентичности последовательности положений.
Способы сопоставления последовательностей для сравнения хорошо известны в данной области. Например, различные программы и алгоритмы сопоставления описаны в Smith and Waterman (1981) Adv. Appl. Math. 2:482; Needleman and Wunsch (1970) J. Mol. Biol. 48:443; Pearson and Lipman (1988) Proc. Natl. Acad. Sci. U.S.A. 85:2444; Higgins and Sharp (1988) Gene 73:237-44; Higgins and Sharp (1989) CABIOS 5:151-3; Corpet et al. (1988) Nucleic Acids Res. 16:10881-90; Huang et al. (1992) Comp. Appl. Biosci. 8:155-65; Pearson et al. (1994) Methods Mol. Biol. 24:307-31; Tatiana et al. (1999) FEMS Microbiol. Lett. 174:247-50. Одно подробное рассмотрение способов сопоставления последовательностей и расчеты гомологии могут быть найдены, например, в Altschul et al. (1990) J. Mol. Biol. 215:403-10.
Национальный Центр для Информации Биотехнологии (The National Center for Biotechnology Information (NCBI) Basic Local Alignment Search Tool (BLAST™; Altschul et al. (1990)) является доступным из нескольких источников, в том числе из National Center for Biotechnology Information (Bethesda, MD), и в Интернете, для использования в связи с несколькими программами анализа последовательностей. Описание, как определить идентичность последовательности с использованием этой программы доступно в Интернете в разделе «help» для BLAST™. Для сравнений последовательностей нуклеиновых кислот, функция «Blast 2 sequences» программы BLAST™ (Blastn) может быть использована с использованием параметров по умолчанию. Последовательности нуклеиновых кислот даже с более высоким сходством со ссылочными последовательностями будут показывать увеличение процента идентичности при оценивании посредством этого способа.
В этом контексте, фраза «функционально связанные» относится к контексту, где первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, когда эта первая последовательность находится в функциональном отношении с этой второй последовательностью нуклеиновой кислоты. Например, промотор является функционально связанным с кодирующей последовательностью, когда этот промотор влияет на транскрипцию или экспрессию этой кодирующей последовательности. При рекомбинантном получении, функционально связанные последовательности нуклеиновых кислот являются обычно смежными и, при необходимости, соединяют две кодирующих белок области в одной и той же рамке считывания. Однако элементы не должны быть смежными, чтобы быть функционально связанными.
В данном контексте, термин «промотор» относится к области ДНК, которая обычно расположена выше (в направлении 5'-области гена), которая подлежит транскрипции. Промоторы могут позволять правильную активацию или репрессию гена, который они регулируют. Промотор может содержать специфические последовательности, которые распознаются факторами транскрипции. Эти факторы могут связываться с последовательностями ДНК промотора и приводить к рекрутингу РНК-полимеразы, фермента, который синтезирует РНК из кодирующей области этого гена.
В данном контексте, термин «трансформирует» или «трансдуцирует» относится к процессу, в котором вирус или вектор переносит молекулы нуклеиновых кислот в клетку. Клетка является «трансформированной» молекулой нуклеиновой кислоты, «трансдуцированной» в эту клетку, когда эта молекула нуклеиновой кислоты становится стабильно реплицированной этой клеткой, либо включением молекулы нуклеиновой кислоты в клеточный геном, либо эписомной репликацией. В данном контексте, термин «трансформация» включают в себя все способы, при помощи которых молекула нуклеиновой кислоты может быть введена в такую клетку. Примеры включают в себя, но не ограничиваются ими: трансфекцию вирусными векторами; трансформацию плазмидными векторами; электропорацию (Fromm et al. (1986) Nature 319:791-3); липофектин (Felgner et al. (1987) Proc. Natl. Acad. Sci. USA 84:7413-7); микроинъекцию (Mueller et al. (1978) Cell 15:579-85); Agrobacterium-опосредованный перенос (Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80:4803-7); прямое поглощение ДНК; whiskers-опосредованную трансформацию; и баллистическую трансформацию (бомбардировку микроснарядами) (Klein et al. (1987) Nature 327:70).
В данном контексте, термин «трансген» относится к экзогенной последовательности нуклеиновой кислоты. В одном примере, трансгеном является последовательность гена (например, гена устойчивости к гербицидам), гена, кодирующего промышленно или фармацевтически применимое соединение, или гена, кодирующего желательный сельскохозяйственный признак. В другом примере, трансгеном является антисмысловая последовательность нуклеиновой кислоты, где экспрессия последовательности этой антисмысловой нуклеиновой кислоты ингибирует экспрессию последовательности нуклеиновой кислоты-мишени. Один трансген может содержать регуляторные последовательности, функционально связанные с трансгеном (например, промотор). В некоторых вариантах осуществления, представляющая интерес последовательность нуклеиновой кислоты является трансгеном. Однако, в других вариантах осуществления, представляющая интерес последовательность нуклеиновой кислоты является эндогенной последовательностью нуклеиновой кислоты, где являются желательными дополнительные геномные копии этой эндогенной последовательности, или последовательностью нуклеиновой кислоты, которая находится в антисмысловой ориентации относительно последовательности молекулы нуклеиновой кислоты-мишени в организме хозяина.
В данном контексте, термин «вектор» относится к молекуле нуклеиновой кислоты, вводимой в клетку с образованием тем самым трансформированной клетки. Вектор может включать в себя последовательности нуклеиновых кислот, которые позволяют ему реплицироваться в клетку-хозяина, такие как сайт инициации репликации (ориджин репликации). Примеры включают в себя, но не ограничиваются ими, плазмиду, космиду, бактериофаг или вирус, который несет экзогенную ДНК в клетку. Вектор может также включать в себя один или несколько генов, антисмысловых молекул и/или селектируемых маркерных генов и других генетических элементов, известных в данной области. Вектор может трансдуцировать, трансформировать или инфицировать клетку, заставляя эту клетку посредством этого экспрессировать молекулы нуклеиновых кислот и/или белки, кодируемые этим вектором. Вектор может необязательно включать в себя вещества, помогающие вхождению молекулы нуклеиновой кислоты в эту клетку (например, липосому).
В данном контексте, термин «растение» включает в себя растения и части растений, включающие в себя, но не ограничивающиеся ими, клетки растений и ткани растений, такие как листья, стебли, корни, цветы, пыльцу и семена. Класс растений, которые могут быть использованы в данном изобретении, обычно является таким широким, как класс высших и низших растений, подверженный мутагенезу, включающий в себя покрытосемянные (однодольные и двудольные растения), голосемянные растения, папоротникообразные растения и многоклеточные водоросли. Таким образом, «растение» включает в себя двудольные растения и однодольные растения. Примеры двудольных растений включают в себя табак, Arabidopsis, сою, томаты, папайю (дынное дерево), канолу, подсолнечник, хлопчатник, люцерну, картофель, винные сорта винограда, голубиный горох (каянус), горох, Brassica, нут бараний, сахарную свеклу, семена рапса, арбуз, дыню, перец, арахис, тыкву, редьку, шпинат, тыкву крупноплодную, брокколи, капусту огородную, морковь, капусту цветную, сельдерей, капусту Китайскую, огурец, баклажан и салат-латук. Примеры однодольных растений включают в себя кукурузу, рис, пшеницу, сахарный тростник, ячмень, сорго, орхидеи, бамбук, бананы, рогозы, лилии, овес, лук, просо и тритикале.
В данном контексте, выражение «растительный материал» относится к листьям, стеблям, корням, цветкам или частям цветков, плодам, пыльце, яйцеклеткам, зиготам, семенам, черенкам, культурам клеток или ткани или любой другой части или продукта растения. В некоторых вариантах осуществления, растительный материал включает в себя семядолю и лист.
В данном контексте, фраза «переключатель трансляции» относится к механизму в конце гена, позволяющему трансляцию гена, находящегося непосредственно ниже. Этот механизм переключения трансляции может функционировать на уровне нуклеиновых кислот (например, с использованием вирусного или эукариотического внутреннего сайта связывания рибосомы (IRES), альтернативного сайта сплайсинга, сайта расщепления рибозима) или на уровне пептида/белка (например, пептида 2A, 2A-подобного пептида, пептида интеина или сайта расщепления протеазы).
Эти механизмы переключения трансляции на уровне нуклеиновых кислот или на уровне пептида/белка хорошо известны в данной области. См., например, li, Z., H.M. Schumacher, et al. (2010) J Biotechnol 145(1): 9-16; Chen, Y., K. Perumal, et al. (2000) Gene Expr 9(3): 133-143; Dinkova, T.D., H. Zepeda, et al. (2005) Plant J 41(5): 722-731; Dorokhov, Y.L., M.V. Skulachev, et al. (2002) Proc Natl Acad Sci U S A 99(8): 5301-5306; Fernandez-Miragall, O. and C. Hernandez (2011) PLoS One 6(7): e22617; Groppelli, E., G. J. Belsham, et al. (2007) J Gen Virol 88(Pt 5): 1583-1588; Ha, S.H., Y.S. Liang, et al. (2010) Plant Biotechnol J 8(8): 928-938; Karetnikov, A. and K. Lehto (2007) J Gen Virol 88(Pt 1): 286-297; Karetnikov, A. and K. Lehto (2008) Virology 371(2): 292-308; Khan, M.A., H. Yumak, et al. (2009) J Biol Chem 284(51): 35461-35470; and Koh, D.C., S.M. Wong, et al. (2003) J Biol Chem 278(23): 20565-20573, содержание которых включено здесь посредством ссылки в их полном объеме. Мультигенные конструкты экспрессии, содержащие модифицированные интеины, были описаны в патентах США №7,026,526 и 7,741,530, а также в заявке на патент США 2008/0115243, содержания которых включено здесь посредством ссылки в полном объеме.
В данном контексте, фраза «селектируемый маркер» или «ген селектируемого маркера» относится к гену, который необязательно используется в трансформации растений, например, для защиты клеток растения от селективного агента или обеспечения устойчивости/толерантности в отношении селективного агента. Только те клетки или растения, которые получают функциональный селектируемый маркер, способны делиться или расти при условиях, имеющих селективный агент. Примеры селективных агентов могут включать в себя, например, антибиотики, в том числе спектиномицин, неомицин, канамицин, паромомицин, гентамицин и гигромицин. Эти селектируемые маркеры включают в себя ген неомицинфосфотрансферазы (npt II), который экспрессирует фермент, придающий устойчивость к антибиотику канамицину, и гены для родственных антибиотиков неомицина, паромомицина, гентамицина, и G418, или ген для гигромицинфосфотрансферазы (hpt), который экспрессирует фермент, придающий устойчивость к гигромицину. Другие селектируемые маркерные гены могут включать в себя гены, кодирующие устойчивость к гербицидам, включающие в себя Bar (устойчивость против BASTA® (глюфосинат аммония), или фосфинoтрицин (PPT)), ацетолактатсинтазу (ALS, устойчивость против ингибиторов, таких как сульфонилмочевины (SU), имидазолиноны (IMI), триазолопиримидины (TP), пиримидинилоксибензоаты (POB) и сульфониламинокарбонилтриазолиноны, которые предотвращают первую стадию в синтезе аминокислот с разветвленной цепью), глифосат, 2,4-D, и устойчивость или чувствительность к металлам. Фраза «маркер-положительные» относятся к растениям, которые были трансформированы для включения селектируемого маркерного гена.
Различные селектируемые или детектируемые маркеры могут быть включены в выбранный экспрессирующий вектор для возможности идентификации и селекции трансформированных растений, или трансформантов. Многие способы являются доступными для подтверждения экспрессии селективных маркеров в трансформированных растениях, включающие в себя, например, секвенирование ДНК и ПЦР (полимеразную цепную реакцию), Саузерн-блоттинг, РНК-блоттинг, иммунологические способы для детектирования белка, экспрессированного из этого вектора, например, осажденного белка, который опосредует устойчивость к фосфинотрицину, или других белков, таких как репортерные гены β-глюкуронидаза (GUS), люцифераза, зеленый флуоресцентный белок (GFP), DsRed, β-галактозидаза, хлорамфениколацетилтрансфераза (CAT), щелочная фосфатаза и т.п. (См. обзор Sambrook, et al., Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Press, N.Y., 2001, содержание которого включено здесь посредством ссылки в полном объеме).
Селектируемые маркерные гены используются для селекции (отбора) трансформированных клеток и тканей. Селектируемые маркерные гены включают в себя гены, кодирующие устойчивость к антибиотикам, такие как гены, кодирующие неомицинфосфотрансферазу II (NEO) и гигромицинфосфотрансферазу (HPT), а также гены, придающие устойчивость к гербицидным соединениям. Гены устойчивости к гербицидам обычно кодируют модифицированный белок-мишень, нечувствительный к этому гербициду, или фермент, который деградирует или обезвреживает этот гербицид в растении до того, как он действует. Например, устойчивость к глифосату получали с использованием генов, кодирующих мутантные ферменты-мишени, 5-енолпирувилшикимат-3-фосфатсинтазу (EPSPS). Гены и мутанты для EPSPS были описаны в патентах США №4,940,835, 5,188,642, 5,310,667, 5,633,435, 5,633,448 и 6,566,587, содержания которых включены посредством ссылок в полном объеме. Устойчивость к глюфосинату аммония, бромоксинилу и 2,4-дихлорфеноксиацетату (2,4-D) получали с использованием бактериальных генов, кодирующих фосфинотрицинацетилтрансферазу, нитрилазу или 2,4-дихлорфеноксиацетатмонооксигеназу, которые обезвреживают соответствующие гербициды. Ферменты/гены для устойчивости/толерантности к глюфосинату были описаны в патентах США №5,273,894, 5,276,268, 5,550,318 и 5,561,236, содержания которых включены здесь посредством ссылки в их полном объеме. Ферменты/гены для устойчивости к 2,4-D были ранее описаны в патентах США №6,100,446 и 6,153,401, а также в заявках на патенты США 2009/0093366 и WO 2007/053482, содержания которых включены здесь посредством ссылки в их полном объеме. Ферменты/гены для нитрилазы были ранее описаны в патенте США №4,810,648, содержание которого включено посредством ссылки в его полном объеме.
Другие гербициды могут ингибировать точку роста или меристему, в том числе имидазолинон или сульфонилмочевина, и были описаны гены для устойчивости/толерантности ацетогидроксикислота-синтазы (AHAS) и ацетолактатсинтазы (ALS) для этих гербицидов. Гены и мутанты для AHAS и мутанты были описаны в патентах США №4,761,373, 5,304,732, 5,331,107, 5,853,973 и 5,928,937, содержания которых включены посредством ссылки в их полном объеме. Гены и мутанты для ALS были описаны в патентах США №5,013,659 и 5,141,870, содержания которых включены посредством ссылки в их полном объеме.
Гены устойчивости к глифосату включают в себя мутантные гены 5-енолпирувилшикимат-3-фосфатсинтазы (EPSPs) (через введение рекомбинантных нуклеиновых кислот и/или различных форм мутагенеза in vivo нативных генов EPSP), гены aroA и гены глифосатацетилтрансферазы (GAT), соответственно). Гены устойчивости для других фосфоно-соединений включают в себя гены глюфосинат (фосфинотрицинацетилтрансферазы (PAT) из видов Streptomyces, включающих в себя Streptomyces hygroscopicus и Streptomyces viridichromogenes), и гены пиридинокси- или феноксипропионовых кислот и циклогексонов (кодирующие ингибитор ACCase гены). Гены устойчивости/толерантности ацетилкоэнзим А-карбоксилазы (ACCase) были описаны в патентах США №5,162,602 и 5,498,544, содержания которых включены посредством ссылки в их полном объеме.
Молекула ДНК, кодирующая мутантный ген aroA, может быть получена под номером доступа ATCC 39256, и эта нуклеотидная последовательность этого мутантного гена описана в патенте США №4,769,061, выданном Comai, Заявке на Европейский патент №0333033, Kumada et al., и патенте США №4,975,374, выданном Goodman et al., с описанием нуклеотидных последовательностей генов глутаминсинтетазы, которые придают устойчивость к гербицидам, таким как L-фосфинотрицин. Нуклеотидная последовательность гена PAT обеспечена в Европейской заявке №0242246 Leemans et al. Кроме того, DeGreef et al., Bio/Technology 7:61 (1989), описывают получение трансгенных растений, которые экспрессируют химерные гены bar, кодирующие активность PAT. Примерами генов, придающих устойчивость к феноксипроприоновым кислотам и циклогексонам, включающим в себя сетоксидим и галоксифоп, являются гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные Marshall et al., Theon. Appl. Genet. 83:435 (1992). Гены GAT, способные придавать устойчивость к глифосату, описаны в WO 2005012515 Castle et al. Гены, придающие устойчивость к гербицидам 2,4-D, фоп и пиридилоксиауксину, описаны в WO 2005107437 и заявке на патент США Ser. №11/587,893.
Другие гербициды могут ингибировать фотосинтез, в том числе триазин (гены psbA и 1s+) или бензонитрил (ген нитрилазы). Przibila et al., Plant Cell 3:169 (1991), описывает трансформацию Chlamydomonas плазмидами, кодирующими мутантными генами psbA. Нуклеотидные последовательности для генов нитрилазы описаны в патенте США №4,810,648, Stalker, и молекулы DNA, содержащие эти гены, являются доступными под номерами доступа ATCC 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион S-трансферазу, описаны Hayes et al., Biochem. J. 285:173 (1992).
Для целей данного изобретения гены селектируемых маркеров включают в себя, но не ограничиваются ими, гены, кодирующие: неомицинфосфотрансферазу II (Fraley et al. (1986) CRC Critical Reviews in Plant Science, 4:1-25); цианамидгидратазу (Maier-Greiner et al. (1991) Proc. Natl. Acad. Sci. USA, 88:4250-4264); аспартаткиназу; дигидродипиколинатсинтазу (Perl et al. (1993) Bio/Technology, 11:715-718); триптофандекарбоксилазу (Goddijn et al. (1993) Plant Mol. Bio., 22:907-912); дигидродипиколинатсинтазу и десенсибилированную аспартаткиназу (Perl et al. (1993) Bio/Technology, 11:715-718); ген bar (Toki et al. (1992) Plant Physiol., 100:1503-1507 и Meagher et al. (1996) и Crop Sci., 36:1367); триптофандекарбоксилазу (Goddijn et al. (1993) Plant Mol. Biol., 22:907-912); неомицинфосфотрансферазу (NEO) (Southern et al. (1982) J. Mol. Appl. Gen., 1:327; гигромицинфосфотрансферазу (HPT или HYG) (Shimizu et al. (1986) Mol. Cell Biol., 6:1074); дигидрофолатредуктазу (DHFR) (Kwok et al. (1986) PNAS USA 4552); фосфинотрицинацетилтрансферазу (DeBlock et al. (1987) EMBO J., 6:2513); дегалогеназу 2,2-дихлорпропионовой кислоты (Buchanan-Wollatron et al. (1989) J. Cell. Biochem. 13D:330); ацетогидроксикислота-синтазу (Anderson et al., патент США №4,761,373; Haughn et al. (1988) Mol. Gen. Genet. 221:266); 5-енолпирувил-шикимат-фосфат-синтазу (aroA) (Comai et al. (1985) Nature 317:741); галогенарилнитрилазу (Stalker et al., published PCT application WO 87/04181); ацетил-коэнзим А-карбоксилазу (Parker et al. (1990) Plant Physiol. 92:1220); дигидроптероатсинтазу (sul I) (Guerineau et al. (1990) Plant Mol. Biol. 15:127); и полипептид 32 кД фотосистемы II (psbA) (Hirschberg et al. (1983) Science, 222:1346).
Включены также гены, кодирующие: хлорамфеникол (Herrera-Estrella et al. (1983) EMBO J., 2:987-992); метотрексат (Herrera-Estrella et al. (1983) Nature, 303:209-213; Meijer et al. (1991) Plant Mol Bio., 16:807-820 (1991); гигромицин (Waldron et al. (1985) Plant Mol. Biol., 5:103-108; Zhijian et al. (1995) Plant Science, 108:219-227 и Meijer et al. (1991) Plant Mol. Bio. 16:807-820); стрептомицин (Jones et al. (1987) Mol. Gen. Genet., 210:86-91); спектиномицин (Bretagne-Sagnard et al. (1996) Transgenic Res., 5:131-137); блеомицин (Hille et al. (1986) Plant Mol. Biol., 7:171-176); сульфонамид (Guerineau et al. (1990) Plant Mol. Bio., 15:127-136); бромоксинил (Stalker et al. (1988) Science, 242:419-423); 2,4-D (Streber et al. (1989) Bio/Technology, 7:811-816); глифосат (Shaw et al. (1986) Science, 233:478-481); и фосфинотрицин (DeBlock et al. (1987) EMBO J., 6:2513-2518). Все ссылки, приведенные в этом описании включены посредством ссылки в их полном объеме, если нет другого указания.
Приведенный выше перечень генов селектируемого маркера и репортера не должен считаться ограничивающим. Любой репортерный или селектируемый ген является включенным в данное изобретение. Если необходимо, такие гены могут быть секвенированы способами, известными в данной области.
Эти репортерные гены и гены селектируемых маркеров синтезируют для оптимальной экспрессии в растении. То есть, кодирующая последовательность этого гена может быть модифицирована для усиления экспрессии в растениях. Этот синтетический маркерный ген конструируют для экспрессии в растениях при более высоком уровне, приводящем к более высокой эффективности трансформации. Способы для синтетической оптимизации генов доступны в данной области. Действительно, несколько генов оптимизировали для увеличения экспрессии продукта этого гена в растениях.
Последовательность маркерного гена может быть оптимизирована для экспрессии в конкретном виде растения или альтернативно может быть модифицирована для оптимальной экспрессии в семействах растений. Предпочтительные кодоны растения могут быть определены из кодонов самой высокой частоты в белках, экспрессируемых в наибольшем количестве в конкретном представляющем интерес виде растения. См., например, EPA 0359472; EPA 0385962; WO 91/16432; Perlak et al. (1991) Proc. Natl. Acad. Sci. USA, 88:3324-3328; и Murray et al. (1989) Nucleic Acids Research, 17: 477-498; патент США №5,380,831; и патент США №5,436,391, включенные здесь посредством ссылки. Таким путем, эта нуклеотидная последовательность может быть оптимизирована для экспрессии в любом растении. Считается, что вся последовательность гена или любая часть последовательности гена может быть оптимизированной или синтетической. То есть, могут быть также использованы полностью оптимизированные или частично оптимизированные последовательности.
Гены, которые придают устойчивость к гербициду:
A. Резистентность/толерантность синтазы ацетогидроксикислоты (AHAS) и ацетолактатсинтазы (ALS) против гербицидов имидазолинона или сульфонилмочевины. Гены и мутанты были описаны в патентах США №4,761,373, 5,304,732, 5,331,107, 5,853,973 и 5,928,937. Гены и мутанты для ALS были описаны в патентах США №5,013,659 и 5,141, 870.
B. Гены устойчивости/толерантности карбоксилазы ацетилкофермента A (ACCase) против гербицидов циклогександионов и/или арилоксифеноксипропановой кислоты (в том числе Haloxyfop, Diclofop, Fenoxyprop, Fluazifop, Quizalofop) были описаны в патентах США №5,162,602 и 5,498,544.
C. Гены для устойчивости/толерантности к глифосату. Ген 5-енолпирувил-3-фосфошикиматсинтазы (ES3P-синтазы) был описан в патенте США №4,769,601. Гены 5-енолпирувилшикимат-3-фосфатсинтазы (EPSPS) и мутанты были описаны в патентах США №4,940,835, 5,188,642, 5,310,667, 5,633,435, 5,633,448 и 6,566,587.
D. Гены для устойчивости/толерантности к глюфосинату (биалафосу, фосфинотрицину (PPT)). Ген для фосфинотрицинацетилтрансферазы (Pat) был описан в патентах США №5,273,894, 5,276,268 и 5,550,318; и ген для устойчивости к биалафосу (Bar) был описан в патентах США №5,561,236 и 5,646,024, 5,648,477 и 7,112,665. Ген для глутаминсинтетазы (GS) был описан в патенте США №4,975,372 и Европейской заявке на патент EP 0333033 A1.
E. Гены устойчивости/толерантности гидроксифенилпируватдиоксигеназы (HPPD) против гербицидов изоксазола, дикетонитрилов и/или трикетонов, включающих в себя сулкотрион и мезотрион, описаны в патентах США №6,268,549 and 6,069,115.
F. Гены для 2,4-D устойчивости/толерантности. Ген 2,4-D-монооксигеназы был описан в патентах США №6,100,446 и 6,153,401. Дополнительные гены для 2,4-D-устойчивости/толерантности описаны в патенте США 2009/0093366 и WO 2007/053482.
G. Ген имидазолглицеринфосфатдегидратазы (IGPD) против гербицидов имидазола и/или триазола был описан в патенте США №5,541,310. Гены Dicamba, деградирующие ферменты (оксигеназу, ферредоксин и редуктазу) против гербицида Dicamba были описаны в патентах США №7,022,896 и 7,105,724.
H. Гены для гербицидов, которые ингибируют фотосинтез, включающие в себя триазин (гены psbA и 1s+) или бензонитрил (ген нитрилазы). См., например, Przibila et al., Plant Cell 3:169 (1991) описывающую трансформацию Chlamydomonas генами psbA. Нуклеотидные последовательности для генов нитрилазы описаны в патенте США № 4,810,648, и молекулы DNA, содержащие такие гены, являются доступными под номерами доступа ATTC 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион S-трансферазу, описаны Hayes et al., Biochem. J. 285:173 (1992).
Если нет других указаний, все технические и научные термины имеют значение, которое обычно понимается специалистами с ординарной квалификацией в области, к которой они относятся. Определения обычных терминов в молекулярной биологии могут быть найдены, например, в: Lewin, Genes V, Oxford University Press, 1994 (ISBN 0-19-854287-9); Kendrew et al. (eds.), The Encyclopedia of Molecular Biology, Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и Meyers (ed.), Molecular Biology and Biotechnology: A Comprehensive Desk Reference, VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Обеспечены конструкты и способы, относящиеся к трем паралогам гена EPSPS (A, B и C), и их генетическим компонентам, таким как 5’UTR, промоторы и транзитные пептиды, в Brassica napus. Описаны также трансгенные и нетрансгенные (в их природной среде) применения этих генов и их генетических элементов. В некоторых вариантах осуществления, трансгенное применение этих генов или элементов может придавать признаки толерантности к гербицидам (например, глифосату или 2,4-D) в растении. Паралоги А и В разделяют высокую гомологию или идентичность (~92%) и также действуют С и Е (~95%). Паралог A EPSPS является паралогом с наивысшей экспрессией во множественных типах тканей и в различных стадиях роста. Ген паралога А EPSPS экспрессируется конститутивно во всех испытанных тканях растений, например, листьях, корнях, стеблях, верхущечной меристеме, цветках, цветочных почках и т.д., на стадии 4-8 листьев. Кроме того, паралог А имеет уникальную последовательность транзитного пептида в сравнении с другими четырьмя паралогами. В некоторых вариантах осуществления, этот транзитный пептид паралога А EPSPS используется для обеспечения эффективной транслокации предшественников белка из цитоплазмы в пластиды. Далее, этот транзитный пептид паралогов B, C и E EPSPS может быть использован для обеспечения транслокации предшественников белка из цитоплазмы в пластиды. Эти ферменты EPSPS представляют шестой ключевой фермент шикиматного пути для синтеза ароматических аминокислот и ароматического метаболита в растениях, грибах и микроорганизмах. Таким образом, эти гены EPSPS могут повышающим образом или понижающим образом регулироваться в растениях любыми существующими или будущими технологиями, которые применяют для манипуляциии содержания аминокислот в растениях. См., например, WO 2009/042164, содержание которого включено здесь посредством ссылки в его полном объеме. Все из этих признаков, либо по отдельности, либо в комбинации, делают паралоги EPSPS важными для использования в трансгенных или не-трансгенных (природной среде генов) применениях для придания признака, такого как толерантность к гербициду и/или изменения в содержании аминокислот, углерода и азота в результате манипуляции шикиматного или ассоциированного путей. Особый интерес представляет трансгенная канола (Brassica napus) в соответствии с данным изобретением.
Обеспечена последовательность промотора паралога А EPSPS из сорта B. napus variety Nex710. Этот промотор паралога А EPSPS является двунаправленным на основе результатов, показанных в каллусной ткани B. napus и растениях, и, следовательно, названным двунаправленным конститутивным промотором Brassica (BBCP). Применение BBCP в трансгенных растениях может обеспечивать по меньшей мере одно из следующих преимуществ: (a) больше генов могут быть атакованы в одном раунде трансформации в геном растения; (b) трансгены могут быть конститутивно экспрессированы во всех тканях и частях растения; и (c) новые гены могут быть дополнительно добавлены или изменены в локусе-мишени с опосредованным «цинковыми пальцами» точным стекингом генов. Например, применение BBCP может позволить экспрессию селектируемого маркера/признака устойчивости к гербицидам в одном направлении и представляющего интерес гена (например, признака защиты культуры или увеличения урожая) в другом направлении. Дополнительно обеспечен уникальный транзитный пептид, содержащийся в последовательности гена паралога А, для создания возможности нацеливания на пластиду, например, хлоропласт.
B. napus является амфидиплоидным видом, происходящим из комбинации двух наборов хромосом B. rapa (2n=20, AA) и B. oleracea (2n=18, CC). Таким образом, обеспеченные множественные гены-паралоги EPSPS могли бы быть либо гомеологичными, либо паралогичными генами в зависимости от их происхождения либо из геномов А или С (гомеологичные), либо в качестве результата их удвоения в геноме после видообразования (паралогичные).
Обеспеченные способы и конструкты могут быть использованы для экспрессии любого признака в каноле или двудольных/однодольных растениях, такого как вводимый признак (например, признаки устойчивости к насекомым и толерантности к гербицидам), агрономические признаки (например, увеличение выхода), конечные признаки (например, целебное масло). Все способы, подходящие для конструирования специфических векторов с использованием BBCP, и его трансформация в канолу или другие растения также обеспечены.
Доставка и трансформация: Подходящие способы для трансформации растений включают в себя любой способ, посредством которого ДНК может быть введена в клетку, например, и без ограничения: электропорацию (см., например, патент США 5,384,253); бомбардировку микроснарядами (см., например, патенты США 5,015,580, 5,550,318, 5,538,880, 6,160,208, 6,399,861 и 6,403,865); Agrobacterium-опосредуемую трансформацию (см., например, патенты США 5,635,055, 5,824,877, 5,591,616; 5,981,840 и 6,384,301); и трансформацию протопластов (см., например, патент США 5,508,184). Посредством применения таких способов, как предыдущие, клетки в сущности любых видов растений могли быть стабильно трансформированы, и эти клетки могут развиваться в трансгенные растения способами, известными квалифицированному в данной области специалисту. Например, способы, которые могут быть особенно применимыми в контексте трансформации хлопчатника, описаны в патентах США 5,846,797, 5,159,135, 5,004,863 и 6,624,344; способы для трансформации растений Brassica в частности, описаны, например, в патенте США 5,750,871; способы для трансформации сои описаны, например, в патенте США 6,384,301; и способы для трансформации маиса описаны, например, в патентах 7,060,876 и 5,591,616 и Международной Публикации PCT Publication WO 95/06722.
После выполнения доставки экзогенной нуклеиновой кислоты в реципиентную клетку, эту трансформированную клетку идентифицируют для дополнительного культивирования и регенерации растений. Для улучшения способности идентификации трансформантов, может быть желательным применение селектируемого или сортируемого маркерного гена с трансформирующим вектором, используемым для генерирования трансформанта. В этом случае, эта потенциально трансформируемая популяция клеток может быть анализирована подверганием этих клеток действию селективного агента или агентов, или эти клетки могут быть подвергнуты скринингу на желаемый признак маркерного гена.
Клетки, которые выживают при подвергании действию селективного агента, или клетки, которые оценивались положительно в анализе скрининга, могут быть культивированы в средах, которые поддерживают регенерацию растений. В некоторых вариантах осуществления, любые подходящие для культуры ткани растений (например, среды MS и N6) могут быть модифицированы включением дополнительных веществ, таких как регуляторы роста. Ткань может поддерживаться на основных средах с регуляторами роста, пока достаточное количество ткани является доступным для начала попыток регенерации растений или после повторяемых раундов мануальной селекции, пока морфология этой ткани является пригодной для регенерации (например, по меньшей мере 2 недель), затем ее переносят в среды, благоприятные для образования побегов. Культуры переносят периодически, пока наблюдается достаточное образование побегов. После образования побегов, растения могут быть перенесены на почву для дальнейшего роста и созревания.
Для подтверждения присутствия желаемой молекулы нуклеиновой кислоты, содержащей конструкты, обеспеченные в регенерирующих растениях, может быть выполнено разнообразие анализов. Такие анализы могут включать в себя: молекулярные биологические анализы, такие как Саузерн- и Нозерн-блоттинг и ПЦР; биохимические анализы, такие как детектирование присутствия белкового продукта, например, иммунологическими способами (ELISA, Вестерн-блоты, и/или масс-спектрометрия, сопряженная с жидкостной хроматографией (LC-MS Масс-спектрофотометрия) или с использованием ферментативной функции; анализы частей растений, такие как анализы листьев или корней; и/или анализ фенотипа всего регенерированного растения.
События нацеленной интеграции могут быть подвергнуты скринингу, например, посредством ПЦР-амплификации с использованием, например, олигонуклеотидных праймеров, специфических в отношении представляющих интерес молекул нуклеиновых кислот. Понятно, что ПЦР-генотипирование включает в себя, но не ограничивается ими, амплификацию полимеразной цепной реакции (ПЦР) геномной ДНК, полученной из выделенной каллусной ткани растений-хозяев, в которой предполагается наличие представляющей интерес молекулы нуклеиновой кислоты, интегрированной в геном, с последующим стандартным клонированием и анализом последовательности продуктов амплификации ПЦР. Способы ПЦР-генотипирования были хорошо описаны (см., например, Rios et al. (2002) Plant J. 32:243-53), и могут быть использованы в отношении геномной ДНК, полученной из любого вида растения или типа ткани, в том числе культур клеток. Комбинации олигонуклеотидных праймеров, которые связываются как с последовательностью-мишенью, так и вводимой последовательностью, могут быть использованы последовательно или многократно в реакциях ПЦР-амплификации. Могут быть получены олигонуклеотидные праймеры, сконструированные для ренатурации (отжига) с сайтом-мишенью, вводимые последовательности нуклеиновых кислот и/или комбинации из этих двух. Таким образом, стратегии ПЦР-генотипирования могут включать в себя, например, и без ограничения: амплификацию специфических последовательностей в геноме этого растения; амплификацию множественных специфических последовательностей в геноме этого растения; амплификацию неспецифических последовательностей в геноме этого растения и комбинации любых из предыдущих. Квалифицированный в данной области специалист может придумать дополнительные комбинации праймеров и реакций амплификации для запрашивания этого генома. Например, может быть сконструирован набор прямых или обратных олигонуклеотидных праймеров для отжига с последовательностью (последовательностями), специфическими в отношении мишени вне границ введенной последовательности нуклеиновой кислоты.
Прямые и обратные олигонуклеотидные праймеры могут быть сконструированы для специфического отжига с введенной молекулой нуклеиновой кислоты, например, при последовательности, соответствующей кодирующей области в представляющей интерес нуклеотидной последовательности, содержащейся в ней, или для других частей этой молекулы нуклеиновой кислоты. Эти праймеры могут быть использованы вместе с описанными выше праймерами. Олигонуклеотидные праймеры могут быть синтезированы в соответствии с желаемой последовательностью и являются коммерчески доступными (например, из Integrated DNA Technologies, Inc., Coralville, IA). Амплификация может сопровождаться клонированием и секвенированием, или прямым анализом последовательности продуктов амплификации. Квалифицированный в данной области специалист может рассматривать альтернативные способы для анализа продуктов амплификации, генерированных во время ПЦР-генотипирования. В одном варианте осуществления, в ПЦР-амплификации используют олигонуклеотидные праймеры, специфические для гена-мишени.
Хотя это изобретение было описано со ссылкой на специфические способы и варианты, будет понятно, что различные модификации и изменения могут быть произведены без отклонения от этого изобретения. Все публикации, цитированные здесь, особо включены посредством ссылки для целей описания и раскрытия композиций и методологий, которые могут быть использованы в связи с этим изобретением. Все цитированные патенты, заявки на патент и информация последовательности в ссылочных web-сайтах и публичных базах данных также включены здесь посредством ссылки.
ПРИМЕРЫ
Пример 1
Идентификация последовательности промотора (BBCP) паралога А EPSPS
Пять последовательностей гена 5-енолпирувилшикимат-3-фосфатсинтазы (EPSPS) (паралоги или гомологи) в Brassica napus были описаны в US 2009/0205083 A1. Среди этих пяти генов, этот промотор паралога А EPSPS запускает самую сильную экспрессию в различных тканях растений. Для удлинения последовательности 1571 nt паралога А EPSPS (SEQ ID NO: 7), дополнительные последовательности гена паралога А EPSPS получают через геном с использованием универсального набора GenomeWalker™ (Clonetech Laboratories, Palo Alto, CA) для получения полной последовательности паралога А EPSPS (SEQ ID NO: 8), включающей в себя область его промотора и 3’-нетранслируемую область (например, SEQ ID NO: 26).
Для идентификации последовательности промотора гена паралога А EPSPS, осуществляли поиск полной последовательности с использованием инструмента Basic Local Alignment Search Tool (BLAST) против баз данных различных растений и Brassica. Идентифицировали шесть кДНК и/или мРНК последовательностей Brassica napus и Brassica rapa, которые сопоставляли в направлении экспрессии гена паралога А EPSPS. Номера идентификации (ID) GenBank для этих последовательностей являются ES937178, ES904055, CD825798, CD835768, CD837464 и EV121915. Эти последовательности кДНК и/или мРНК могут быть детектированы из библиотек листа, корня или зародыша видов-мишеней, но конкретная роль этих кДНК или мРНК не была аннотирована в GenBank. Интересно, что три кДНК и/или мРНК идентифицировали спариванием 5'-последовательности гена паралога А EPSPS в противоположном направлении относительно экспрессии гена А паралога EPSPS. ID GenBank этих последовательностей являются: CD836095, EV100366 и EE568337. кДНК и/или мРНК Brassica napus являются источниками этих последовательностей, опять без приписывания специфической функции им в GenBank.
Анализ последовательности этого примера показывает, что последовательность промотора гена паралога А EPSPS является двунаправленным промотором, который был назван двунаправленным конститутивным промотором Brassica (BBCP).
Пример 2
Дизайн и конструирование конструктов BBCP
Один отдельный бинарный вектор, названный как pDAB100333 (Фиг. 12) конструировали с использованием известных в данной области процедур. Бинарный pDAB100333 содержит два набора экспрессионных кассет генов или Транскрипционных Единиц Растений (PTU). Первый набор PTU состоит из двунаправленного промотора паралога А Brassica napus (BBCP), который запускает два репортерных гена. Один конец BBCP конструируют для запуска экспрессии репортерного гена β-глюкуронидазы (GUS) и терминируют 3'-нетранслируемой областью открытой рамки считывания Agrobacterium tumefaciens (AtuORF24 3'UTR). Противоположный конец BBCP конструируют для запуска репортерного гена зеленого флуоресцентного белка (GFP) и терминируют 3'-нетранслируемой областью нопалинсинтазы Agrobacterium tumefaciens (Atu Nos 3'UTR).
Второй набор PTU pDAB100333 включает в себя селектируемый маркер, клонируемый в кодирующей последовательности изопентенилтрансферазы (ipt CDS; Genbank Acc No. X00639.1), прерывая таким образом кодирующую последовательность jpt, где используют промотор убиквитина 10 Arabidopsis thaliana (промотор AtUbi10) для запуска кодирующей последовательности фосфинотрицинацетилтрансферазы (PAT), и этот PTU терминируют 3'-нетранслируемой областью открытой рамки считывания-1 A. tumefaciens (AtuORF1 3'UTR). Этот полученный бинарный вектор содержит два визуальных репортерных гена (GUS и GFP), запускаемые двунаправленным промотором, и селектируемый маркерный ген (PAT).
Этот бинарный вектор, pDAB100333, мобилизуют в Agrobacterium tumefaciens с использованием электропорации. Индивидуальные колонии идентифицируют на среде YEP, содержащей антибиотик спектиномицин. Отдельные колонии выделяют и присутствие этого бинарного вектора pDAB100333 может быть подтверждено посредством расщепления рестрикционными ферментами.
Другой бинарный вектор pDAB100331 (Фиг. 12) также конструируют с использованием известных процедур. Бинарный pDAB100331 конструируют таким образом, что он содержит BBCP в обратной ориентации относительно pDAB100333, но с теми же признаками, что и признаки pDAB100333. Таким образом, бинарный вектор pDAB100331 состоит из двух наборов экспрессионных кассет генов или Единиц Транскрипции Растений (PTU). Первый набор PTU состоит из бинаправленного промотора паралога А Brassica napus Paralog A (BBCP в обратной ориентации в сравнении с pDAB100333), который запускает два репортерных гена. Один конец BBCP конструируют таким образом, что он запускает репортерный ген зеленого флуоресцентного белка (GFP), и терминируют 3'-нетранслируемой областью нопалинсинтазы Agrobacterium tumefaciens (Atu Nos 3'UTR). Противоположный конец этого BBCP конструируют для запуска экспрессии репортерного гена β-глюкуронидазы (GUS) и терминируют 3'-нетранслируемой областью открытой рамки считывания-24 Agrobacterium tumefaciens (AtuORF24 3'UTR).
Второй набор pDAB100331 также включает в себя селектируемый маркер, клонированный в кодирующей последовательности изопентенилтрансферазы (ipt CDS; Genbank Acc No. X00639.1), прерывая таким образом кодирующую последовательность jpt, где используют промотор убиквитина 10 Arabidopsis thaliana (промотор AtUbi10) для запуска кодирующей последовательности фосфинотрицинацетилтрансферазы (PAT), и этот PTU терминируют 3'-нетранслируемой областью открытой рамки считывания-1 A. tumefaciens (AtuORF1 3'UTR). Этот полученный бинарный вектор содержит два визуальных репортерных гена (GUS и GFP), запускаемые двунаправленным промотором, и селектированный маркерный ген (PAT).
Подобным образом, бинарный вектор, pDAB100331, мобилизуют в Agrobacterium tumefaciens с использованием электропорации. Индивидуальные колонии идентифицируют на среде YEP, содержащей антибиотик спектиномицин. Отдельные колонии выделяют и присутствие этого бинарного вектора может быть подтверждено посредством расщепления рестрикционными ферментами.
Векторы для прямой доставки, которые клонируют в плазмиды на основе pUC с высокой копийностью, конструируют с использованием только первых PTU, содержащих промотор BBCP, который описан выше. Плазмиды pDAB108710 и pDAB108711 (Фиг. 14) конструируют таким образом, что они содержат BBCP в другой ориентации для запуска тех же самых признаков. Эта единственная PTU состоит из двунаправленного промотора паралога А Brassica napus, который запускает два репортерных гена. Один конец BBCP конструируют таким образом, что он запускает репортерный ген зеленого флуоресцентного белка (GFP), и терминируют 3'-нетранслируемой областью нопалинсинтазы Agrobacterium tumefaciens (AtuNos 3'UTR). Противоположный конец этого BBCP конструируют для запуска экспрессии репортерного гена β-глюкуронидазы (GUS) и терминируют 3'-нетранслируемой областью открытой рамки считывания-24 Agrobacterium tumefaciens (AtuORF24 3'UTR). Векторы прямой доставки ДНК используют для бомбардировки частицами тканей кукурузы.
Пример 3
Экспрессия конструкта BBCP в Brassica napus
Трансформация канолы - Получение сегмента гипокотиля и предобработка: Семена элитного генотипа канолы, Nex710, поверхностно стерилизуют 10% коммерческим отбеливателем Вт 10 минут и промывают 3 раза стерильной дистиллированной водой. Эти семена сушат посредством стерильного бумажного полотенца и затем помещают в Phyta-лоток, содержащий «среду для прорастания», состоящую из ½ концентрации базальной среды MS [Phytotech Cat# M 519 (PhytoTechnology Laboratories, Shawnee Mission, KS)], 20 г/л сахарозы и 8 г/л Агара TC, и поддерживают при режиме роста, установленном при 23°C с фотопериодом 16 часов свет/8 часов темнота.
В пятый день, проростки проверяют на стерильность и этот Phyta-лоток помещают внутрь ламинарного вытяжного шкафа (The Baker Company EdgeGARD) для поддержания стерильности. С использованием стерильного пинцета и ножниц для препарирования, растения вынимают из Phyta-лотка и надземную область (меристему и котиледон) и корни отделяют и выбрасывают. Гипокотили помещают в чашку Петри 100×25 мм, содержащую стерильную дистиллированную воду, которая требуется для предотвращения высушивания. Гипокотили разрезают поперечно на сегменты 2 мм и накладывают горизонтально на стерильной фильтровальной бумаге и наносят на «среду для индуцирования каллуса MSK1D1», состоящую из MS (Phytotech M519), 30 г/л сахарозы, 1 мг/л кинетина и 1 мг/л 2,4-D, отвержденной 7 г/л TC-Агара. Эти чашки помещают в прозрачную кадку Sterilite® и поддерживают при одном и том же режиме в течение 3 дней, в качестве предварительной обработки.
Приготовление Agrobacterium: За четыре дня перед инфицированием Agrobacterium, pDAB10333 и pDAB10331 в штамме Agrobacterium DA2552 (см., например, WO 2012/016222) высевают штрихом из исходного раствора глицерина на среду YEP (10 г/л Пептон, 10 г/л дрожжевой экстракт, 5 г/л NaCl, 10 г/л сахароза плюс 100 мг/л спектиномицин и 150 мг/л эритромицин и отверждают 15 г/л Бакто Агар) и выращивают в течение двух дней в термостате (Fisher Scientific Isotemp Incubator) при 28°C. Спустя два дня, малую петлю Agrobacterium помещают в 500 мл стерильную одноразовую колбу, содержащую 150 мл «жидкой среды для бактериального роста» (ту же самую среду, описанную выше, но минус отверждаемый агент). Эту культуру выращивают в течение шестнадцати часов при 28°С в темноте на закрытом шейкере (New Brunswick Scientific Innova 4330 охлаждаемый инкубаторный шейкер) при 200 об/мин. Спустя шестнадцать часов культуру Agrobacterium удаляют из шейкера и готовят аликвоты в центрифужных пробирках на 50 мл. Эти центрифужные пробирки помещают в центрифугу (Beckman Model J2-21) и центрифугируют при 6000 об/мин в течение 15 минут и затем ресуспендируют в «среде М для жидкой культуры», состоящей из LS-солей (Phytotech L689), 3% глюкозы, модифицированных витаминов Gamborg B5 (Phytotech G249), 215 мг/л кинетина и 221 мг/л 2,4-D при рН 5.
Введение инфекции и каллуса: В день инфицирования, сегменты гипокотилей канолы переносят в стерильную чашку Петри 100×25, содержащую 20 мл «среды для жидкой культуры», ожидая, когда Agrobacterium будет готовым. Затем эту «среду для жидкой культуры» удаляют из сегментов гипокотилей и 40 мл суспензии встряхивают кратковременно вихревым перемешиванием (на вортексе) и выливают в чашку Петри 100×25 мм, содержащую сегменты гипокотилей, в течение 30 минутной обработки. После этой 30-минутной обработки, все суспензии Agrobacterium удаляют с использованием двойной стекинг-пипетки. Эти обработанные гипокотили помещают обратно на слой фильтровальной бумаги на «среде для индуцирования каллуса MSK1D1». Эту культуру возвращают в Sterilite® tub, покрывают темной крышкой и возвращают в комнату для культивирования при режиме роста, описанном выше, в течение трех недель периода сокультивирования. После трех дней периода сокультивирования гипокотили помещают непосредственно на «среду для отбора 1 MSK1D1H1» (состоящую из «среды для индуцирования каллуса» плюс 1 мг/л Herbiace), помещают обратно в кадку с прозрачной крышкой и возвращают в комнату для культивирования с поддержанием того же режима роста, описанного выше. Затем, после одной недели, эти гипокотили переносят непосредственно в «среду 2 для селекции MSK1D1H3» (состоящую из «среды для индуцирования каллуса» плюс 3 мг/л Herbiace). После двух недель, эти гипокотили переносят непосредственно в «среду 3 для селекции MSK1D1H3» (состоящую из «среды для индуцирования каллуса» плюс 5 мг/л Herbiace). Эти сегменты гипокотилей продолжают переносить каждые две недели на свежую среду для селекции 3, пока не образуется достаточно каллуса на обоих концах гипокотилей. Затем эти каллусы анализируют на GUS.
Краситель GUS сегментов гипокотилей канолы: Процедура красителя GUS известна в данной области с небольшой модификацией раствора красителя GUS следующим образом: 0,1 M NaPO4-буфер при pH 8, 0,5 мМ K3(Fe(CN)6, 10 мМ Na2EDTA, 1 мг/мл X-Gluc и 0,06% Triton X-100. Хлорофилл из окрашенной ткани удаляют 70% этанолом. Анализ GUS выполняют в сегментах гипокотилей после нахождения на среде для селекции 3 в течение по меньшей мере двух недель. Эти каллусы погружают в окрашивающий GUS раствор и инкубируют в течение ночи в темноте при 37°C. Неинфицированную ткань и GUS-положительный контроль рутинным образом включают в этот анализ для отрицательного и положительного контролей.
Эти результаты показывают значимую экспрессию GUS в пробах трансгенного каллуса, полученных из трансформации как pDAB100331, так и pDAB100333. Синяя окраска не видна в пробах нетрансгенных контролей. Таким образом, эти трансгенные экспериментальные результаты подтверждают, что BBCP является двунаправленным промотором.
Пример 4
Экспрессия конструкта BBCP в сое
Штамм Agrobacterium tumefaciens EHA105 электропорируют бинарным вектором pDAB9381 (контрольным бинарным вектором, который не содержит двунаправленный промотор BBCP), pDAB100331 и pDAB100333 по отдельности. Выделенные колонии идентифицируют на средах YEP, содержащих антибиотик спектиномицин. Отдельные колонии выделяют и присутствие бинарного вектора pDAB9381, pDAB100331 и pDAB100333 может быть подтверждено с использованием расщепления рестрикционным ферментом (рестриктазой). Agrobacterium-опосредуемая трансформация сои (Glycine max c.v. Maverick) может выполняться согласно способам, хорошо известным в данной области.
После трансформации, как только развиваются корни, проростки с корнями фотографируют для наблюдения экспрессии GFP с фильтром 482 нм/502 нм GFP, охватывающим возбуждение/эмиссию (излучение). Из этих проростков берут пробы листьев для окрашивания GUS в соответствии с протоколом, адаптированным из Jefferson, R., (1987) «Histochemical localization of β-glucuronidase (GUS) reporter activity in plant tissues» Plant Mol. Biol. Reporter, 5: 387-405. Затем листья погружают в окрашивающий раствор, состоящий из: 2X Фосфатного буфера рН 7,0 (1× из 0,1 M NaH2PO4 и 0,1 M Na2HPO4), 0,5 мМ K3(Fe(CN)6, 10 мМ Na2EDTA, 1 мг/мл X-Gluc и 0,06% Тритон Х-100 и инкубируют в течение ночи при 37°C. После инкубирования, окрашивающий раствор удаляют и ткань промывают несколькими сменами 70% этанола и оставляют стоять в течение ночи перед получением фотографий.
Результаты четырех раундов экспериментов показаны в таблице 2 для испытанных эксплантатов и трансгенных проростков. Таблица 2 показывает число трансгенных побегов, которые, как подтверждено, содержат экспрессируемые белковые продукты репортерных генов GFP и GUS, запускаемые BBCP. Трансгенные растения сои, стабильно трансформируемые конструктами pDAB100331 и pDAB100333, регенерируют с частотой 3-14%. Приблизительно 81-99% регенерируемых побегов (проростков) экспрессируют GFP, тогда как 41% из них экспрессируют как GFP, так и GUS. В растениях, регенерируемых pDAB100333, экспрессия GUS является более однородной по всему листу и первично GUS не экспрессируется в ткани центральной жилки и в других жилках. В сравнении с этим в растениях, регенерируемых из pDAB100331, экспрессия GUS является более локализованной в центральной жилке и жилках листьев, как наблюдалось в исследованиях, выполненных на множественных листьях. Трансгенные растения, трансформированные контрольным конструктом, pDAB9381, не обнаруживают экспрессии ни GFP, ни GUS. Эти результаты подтверждают, что BBCP управляет экспрессией трансгена в обоих направлениях при приемлемом уровне, хотя могут быть некоторые минорные различия в направлении в картинах экспрессии в ткани листа.
|
Пример 5
Временная трансформация ткани листа кукурузы
Получение ткани: Выращенную в темноте ткань листа собирают за три-четыре часа перед бомбардировкой и помещают на среду получения бомбардировкой, описанную в таблице 3. Чашки обертывают и хранят при 28°C в темноте, пока они не становятся готовыми для бомбардировки.
Препарат микрочастиц (золото): 30 мг золота (1 мкм в размере, купленные из Bio-Rad, Hercules, CA) промывают в 500 мкл холодного этанола, обрабатывают ультразвуком в течение пятнадцати секунд и затем подвергают вихревому перемешиванию в течение пятнадцати секунд. Эти частицы центрифугируют в течение шестидесяти секунд при 3000 об/мин после осаждения в течение десяти минут. Затем супернатант выбрасывают и осадок промывают холодной водой (разрушают осадок кончиком пипетки и/или фингер-вортексом) с последующим центрифугированием в течение шестидесяти секунд при 3000 об/мин. Эту промывку и стадию центрифугирования можно повторять два раза или более. После конечного промывания, добавляют 250 мкл 50%-ного глицерина (конечная концентрация ~120 мг/мл). Эти пробы обрабатывают ультразвуком в течение пятнадцати секунд, вихревым перемешиванием в течение пятнадцати секунд и готовят аликвоты в пробирках Эппендорфа.
Приготовление микрочастиц: Для каждой чашки ткани, подлежащей бомбардировке, реакцию золото/ДНК получают следующим образом: 5-15 мкг ДНК (плазмидная ДНК pDAB108710, pDAB108711 и контрольная плазмидная ДНК), 6 мг золота, конечные концентрации 1 M CaCl2 и 16 мМ спермидин, в общем реакционном объеме 125 мкл. Немедленно после обработки ультразвуком и вихревым перемешиванием берут аликвоты в каждую реакционную пробирку. Эти реакционные пробирки обрабатывают вихревым перемешиванием перед добавлением 50 мкл предварительно охлажденного CaCl2, и затем опять обрабатывают вихревым перемешиванием перед добавлением 20 мкл 0,1 М спермидина. Затем реакционные пробирки обрабатывают вихревым перемешиванием в течение до десяти минут и оставляют стоять в течение десяти минут на лабораторном столе перед центрифугированием в течение десяти секунд при 5000 об/мин. Супернатант выбрасывают и осадки ресуспендируют в 150 мкл 70% этанола. Затем осадки разрушают пипеткой и/или фингер-вортексом перед центрифугированием в течение пятнадцати секунд при 5000 об/мин. Затем супернатант выбрасывают и осадки ресуспендируют в 150 мкл этанола. Затем осадки опять разрушают пипеткой и/или фингер-вортексом перед центрифугированием в течение пятнадцати секунд при 5000 об/мин. Наконец, осадки ресуспендировали в 36 мкл этанола (36 мкл на 6 мг золота) перед экспериментом с бомбардировкой. Перед помещением аликвот на микроносители, осадки обрабатывают ультразвуком в течение пятнадцати секунд и вихревым перемешиванием, пока золото не будет хорошо суспендированным. Аликвоту осаждают на три микроносителя с 10 мкл, каждая. Рекомендуются обработка ультразвуком и вихревое перемешивание между аликвотами.
|
Условия бомбардировки: Диски микроносителей на основе гелия и держатель для микроносителей готовят и автоклавируют перед использованием. ДНК-покрытые частицы золота (10 мкл, хорошо суспендированные) помещают на микроносители на пять минут до высушивания. Для сборки пусковой части для запуска микроносителя, в этот собранный узел помещают стоп-экран. Затем этот микроноситель помещают вверх дном на этот собранный узел и затем крышку собранного узла закрывают перед тем, как свежий и кратковременно сполоснутый (в 70% этаноле), разрываемый при давлении 1350 psi (фунт-сила на квадратный дюйм) диск помещают в удерживающую крышку и присоединяют к ускоряющей газ трубе. Затем этот собранный узел микроносителя немедленно помещают на щель верхнего уровня перед закрыванием двери камеры. Вакуум активируют в камере эвакуации перед бомбардировкой. Пробы бомбардируют с использованием давления и удерживанием пусковой кнопки, пока разрываемый при давлении 1350 psi (фунт-сила на квадратный дюйм) диск не взрывается и калибр давления гелия падает до нуля. Каждую чашку можно бомбардировать до трех раз одной и той же плазмидой, где эти чашки могут поворачиваться таким образом, что бомбардировка может иметь место в разном сайте/направлении на этой чашке. После бомбардировки чашки хранят в темноте при 28°С в течение двадцати четырех часов перед наблюдением окрашивания GFP и/или окрашивания GUS с использованием набора для окрашивания Sigma-Aldrich.
Результаты: экспрессию GUS наблюдают в тканях листьев, бомбардируемых плазмидами, содержащими ген GUS регулируемый BBCP. Ткани листьев кукурузы, бомбардируемые контрольной плазмидой, не обнаруживают никакой экспрессии GUS. Экспрессию GFP наблюдают в пробах листьев кукурузы, трансформированных плазмидами, имеющими ген GFP, запускаемый BBCP. Ткани листьев кукурузы, бомбардируемые контрольным конструктом, не обнаруживают никакой экспрессии GFP. Эти результаты подтверждают, что BBCP является двунаправленным промотором, регулирующим экспрессию репортерных генов, GUS и GFP, в обоих направлениях. В заключение автор делает вывод, что двунаправленная экспрессия от BBCP может наблюдаться как в двудольных (соя и B. napus), так и в однодольных (кукуруза) тканях растений.
Хотя ряд примерных аспектов и вариантов обсуждались выше, квалифицированные в данной области специалисты будут узнавать определенные модификации, пермутации, добавления и их субкомбинации. Таким образом, предполагается, что следующие прилагаемые пункты формулы изобретения и вводимые после этого пункты формулы изобретения, интерпретируются как включающие в себя все такие модификации, пермутации, добавления, и их субкомбинации, и находятся в пределах сущности и объема изобретения.





















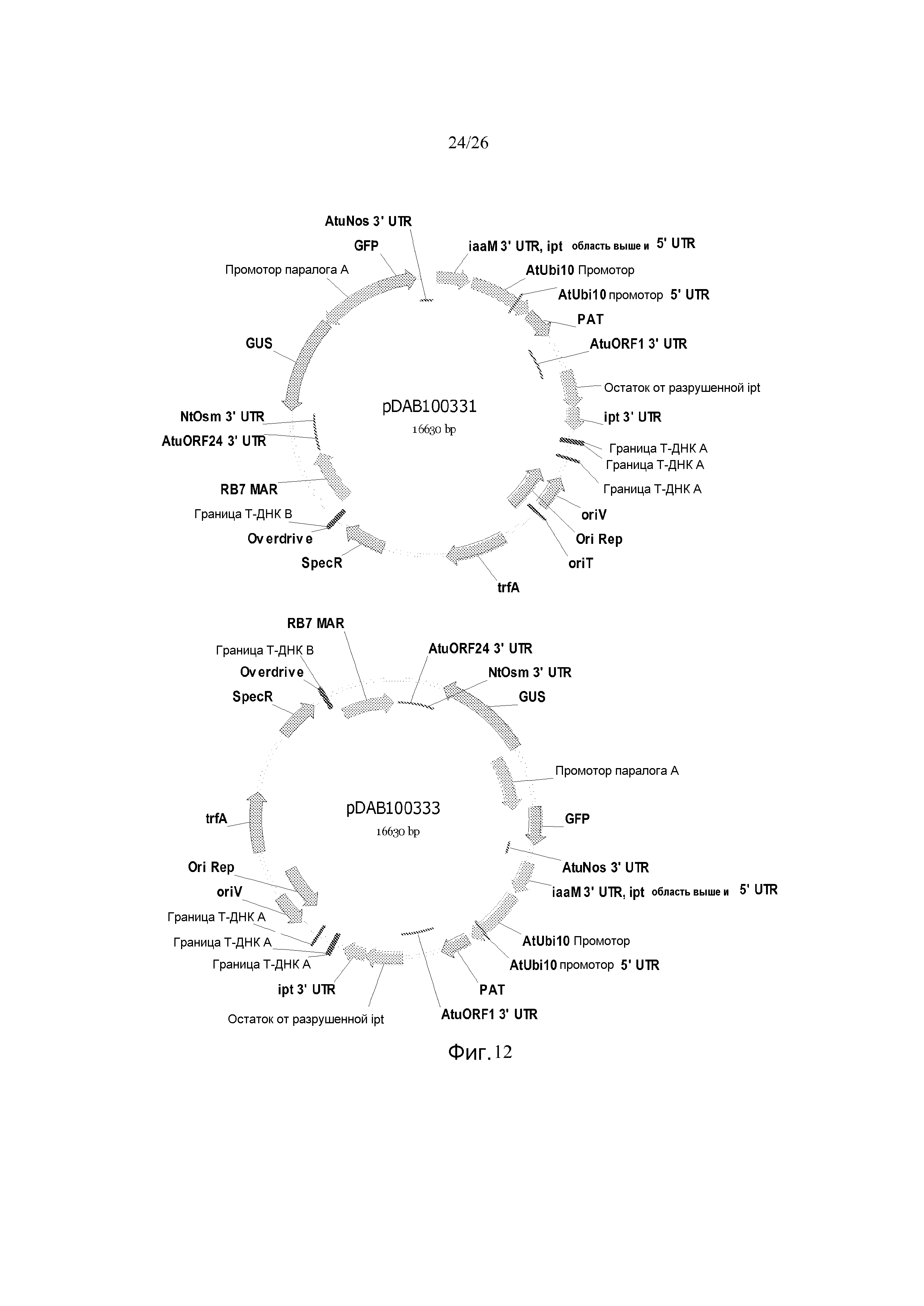















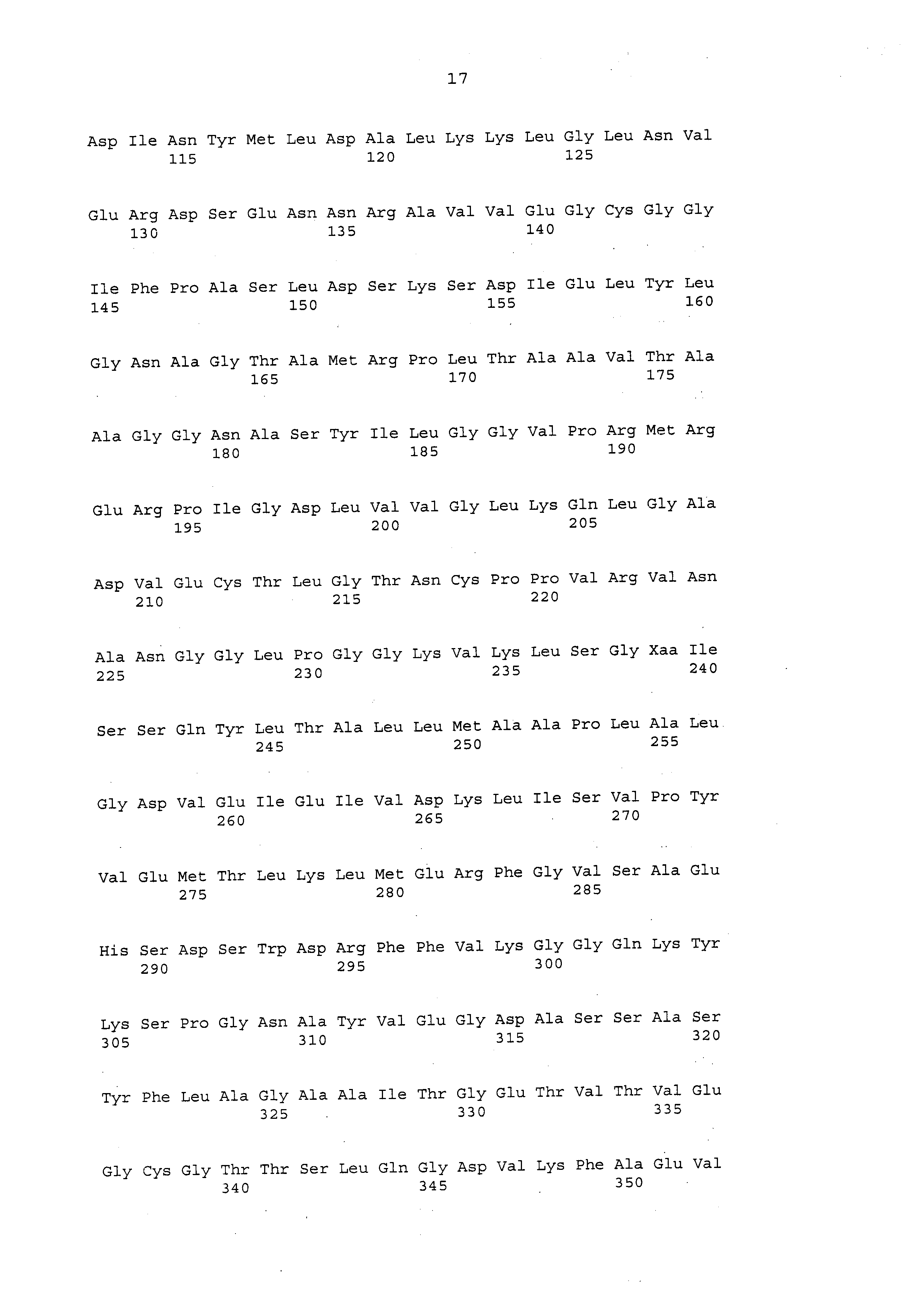























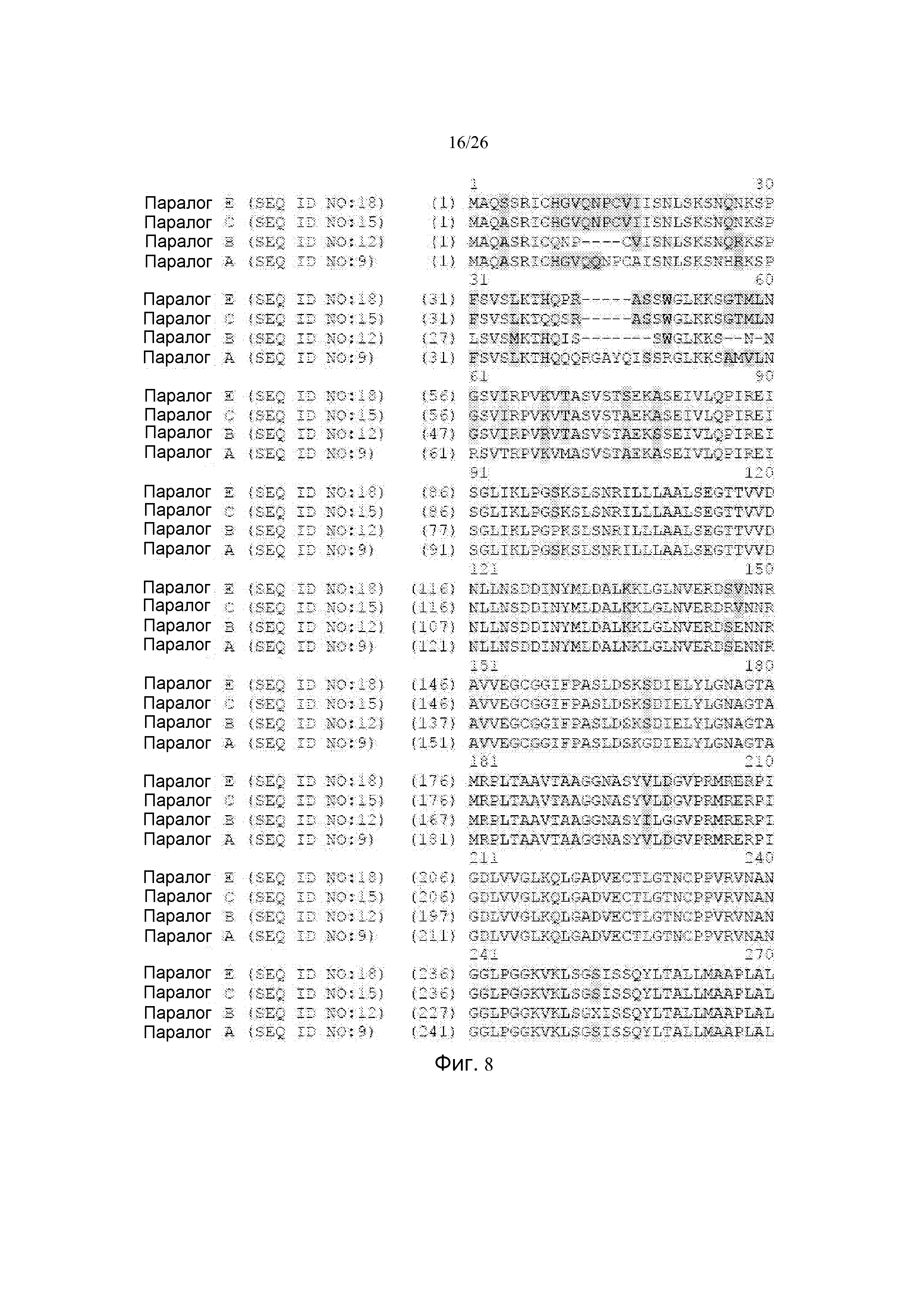

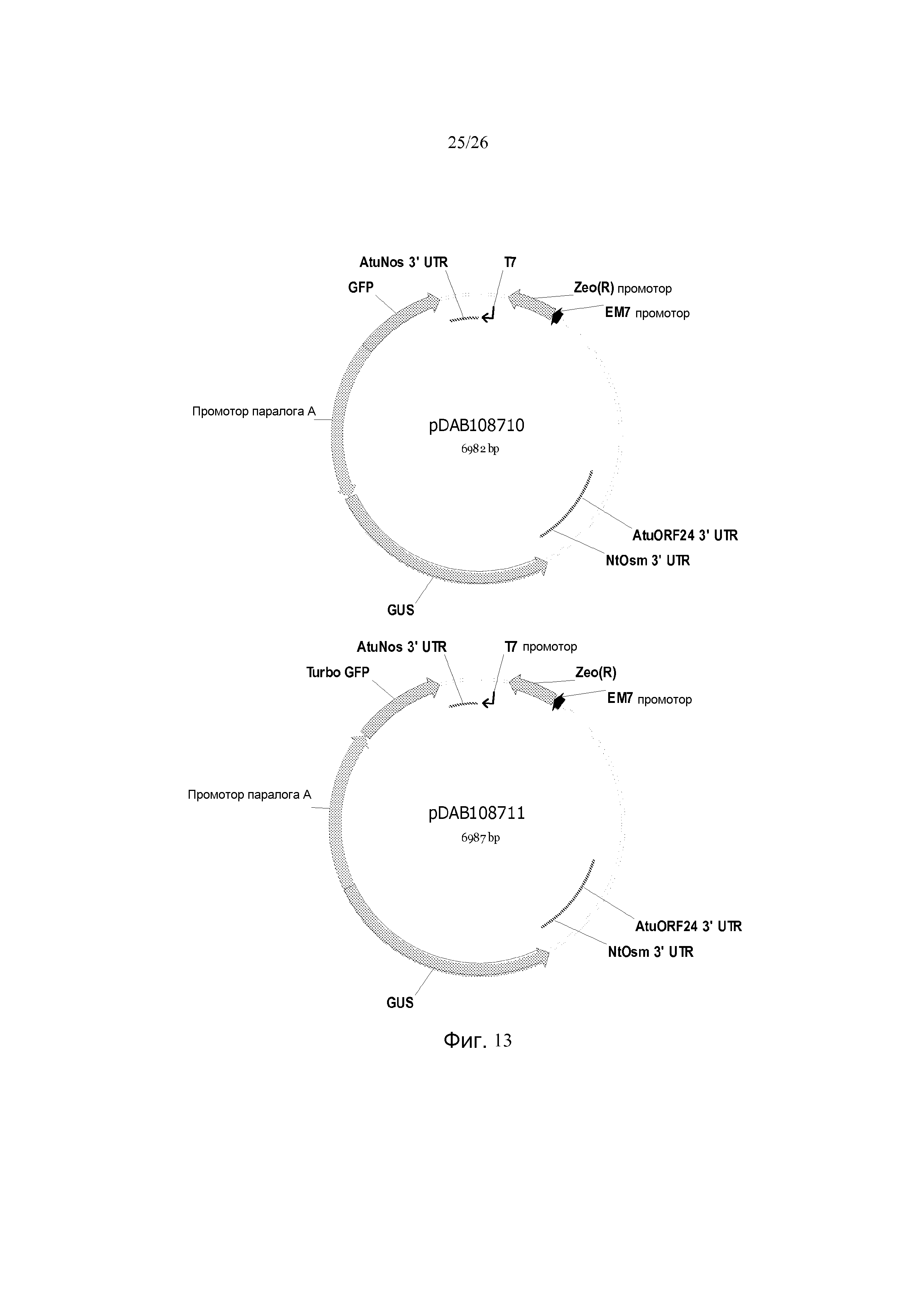

Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
5-фторпиримидиновые производные в качестве фунгицидов
Пестицидные композиции
Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
Стабильная пестицидная композиция на основе сульфоксимина и способ борьбы с насекомыми
5-фторпиримидиновые производные в качестве фунгицидов

