Результат интеллектуальной деятельности: СПОСОБ ИЗМЕРЕНИЯ ДИНАМИКИ РОСТА ЛИСТЬЕВ ДЕРЕВА В ЧИСТЫХ ЭКОЛОГИЧЕСКИХ УСЛОВИЯХ
Вид РИД
Изобретение
Изобретение относится к инженерной биологии и биоиндикации окружающей части кроны деревьев воздушной среды измерениями динамики, прежде всего во времени активной жизнедеятельности растений от начала сокодвижения и распускания почек до последних этапов онтогенеза, преимущественно в процессе жизнедеятельности ростовых органов различных видов древесных растений, например, в виде учетных листьев без их взятия у древесных растений с простой и небольшой листовой пластинкой: липы, клена полевого или американского, березы, тополя.
Предлагаемое изобретение может быть также использовано при экологическом и технологическом мониторинге за развитием и ростом молодых древесных растений, в частности за молодняками лесных деревьев, а также за качеством листвы деревьев в городских парках, аллеях, придорожных полосах и других древесных насаждений, естественного облесения пустырей, бывших строительных и иных площадок при рекультивации земель, естественного лесного возобновления на залежи земельных участков сельскохозяйственного назначения.
Известен способ измерения площади листьев у древесных растений (см. эту же книгу: Федорова А.И., Никольская А.Н. Практикум по экологии и охране окружающей среды: Учеб. пос. М.: Гуманит. изд. центр ВЛАДОС, 2001. 288 с.. С. 123-126), когда срезают по 20-25 листьев каждой древесной породы с деревьев, растущих в разных экологических условиях, складывают в пакеты, а затем засушивают между листами газетной бумаги в лабораторных условиях.
Недостатком является низкая чувствительность (точность) индикации и высокая трудоемкость практического применения способа из-за смешивания листьев у разных деревьев в одну общую пробу.
Известен также способ измерения площади листьев у древесных растений по патенту №2466351, включающий взятие листьев от учетных деревьев, растущих в разных экологических условиях, далее каждый измеряемый лист укладывается на подложку с белой поверхностью, а на лист сверху накладывается палетка для картографических измерений, причем продольная ось листа растения совмещается с одной из линий сетки палетки примерно в середине палетки, затем лист через прозрачную палетку с сеткой фотографируется, после фотография помещается в память компьютера, а измерения длины, ширины и площади листа выполняются по клеткам сетки палетки на увеличенном изображении листа растения.
Недостатком является взятие пробы листьев, что дает только однократные времени онтогенеза листьев измерения, отсутствие измерений с начала вегетационного периода по датам, что не позволяет изучать динамику поведения учетных листьев в ходе их развития и роста. При этом не выделяется локальная зона кроны дерева, что не позволяет изучать влияние окружающей части кроны деревьев воздушной среды измерениями динамики параметров листьев от начал распускания почек до разных этапов онтогенеза листьев.
Технический результат - расширение функциональных возможностей изучения динамики по суткам среднестатистического поведения конкретной группы учетных листьев, выбранных без их срезания из локальной зоны на поверхности кроны дерева с примерно одинаковым освещением, причем изменением во времени на разных этапах вегетации по четырем параметрам (длины и ширины листа, периметра и площади листа), а также повышение точности измерений динамики этих параметров учетных листьев во времени, начиная от начала распускания почек.
Этот технический результат достигается тем, что способ измерения динамики роста листьев дерева, включающий укладку подложки с белой поверхностью снизу на измеряемый лист, а на лист сверху накладывают прозрачную палетку для картографических измерений, причем продольную ось листа растения совмещают с одной из линий сетки палетки, затем лист через прозрачную палетку с сеткой фотографируют, после фотография помещается в память компьютера, а измерения длины и ширины листа выполняют по клеткам сетки палетки на увеличенном изображении листа растения, отличающийся тем, что на боковой поверхности кроны дерева выделяют локальную зону с примерно одинаковым освещением, на которой выбирают группу учетных листьев с метками, измерения параметров учетных листьев проводят за вегетационный период по суткам от начала распускания почек листьев с фотографированием каждого учетного листа, причем первое фотографирование выполняют не менее чем через неделю и не позже чем три недели после распускания почек листьев дерева, в течение времени вегетации проводят измерения параметров учтенных листьев, а затем в группе по всем учетным листьям, принятым на одной локальной зоне поверхности кроны дерева, статистическим моделированием выявляют среднестатистические закономерности динамики роста учетных листьев по отдельным параметрам учетных листьев, произрастающих на одной локальной зоне поверхности кроны дерева.
В кроне дерева на удобной для измерений высоте на боковой поверхности кроны дерева выделяют локальную зону с примерно одинаковым освещением, на которой выбирают группу из не менее 10 учетных листьев, каждый из которых отмечают меткой в виде куска белой нитки, привязанной к основанию листа на черешке, при этом локальную зону выбирают примерно в виде круга диаметром не более 0,5 м, а в качестве учетных принимают листья, расположенные на поверхности кроны с освещенной утренними солнечными лучами стороны кроны дерева.
Дополнительно к длине, ширине и площади учетных листьев измеряют периметр листа с учетом масштаба клеток прозрачной палетки по количеству клеток, через которые проходит ребристая линия края листа, при этом площадь листа определяют как сумму полных клеток внутри поверхности листа и неполных клеток вдоль периметра листа также с учетом масштаба клеток палетки.
Периметр листа измеряют по количеству клеток, через которые проходит ребристая линия края листа с учетом масштаба клеток палетки 2×2 мм, по формуле:
P=0,28284IP,
где P - периметр листа, см;
IP - количество по периферии листа неполных клеток, шт.
Площадь листа определяют как сумму полных клеток внутри поверхности листа и неполных клеток вдоль периметра листа также с учетом масштаба клеток палетки 2×2 мм, по формуле:
S=0,04IS+0,02IP,
где S - площадь листа, измеряемая количеством клеток размерами 2×2 мм внутри листа и количеством клеток периметра листа, см2;
IS - количество на изображении листа полных клеток, шт.;
IP - количество по периферии листа неполных клеток, шт.
Измерения параметров учетных листьев проводят за время вегетации по суткам от начала распускания почек листьев с фотографированием каждого учетного листа через прозрачную палетку с сеткой 2 мм, причем первое фотографирование выполняют не менее чем через неделю и не позже чем три недели после распускания почек листьев дерева, при этом по дате начала времени вегетации текущее время в сутках принимают за влияющую переменную, причем значение влияющей переменной в начале вегетации листьев принимают равным нулю, а визуально параметры учетных листьев по длине и ширине, периметру и площади учетных листьев в начале вегетации принимают также с нулевыми значениями, при этом фотографирование до конца вегетации выполняют не менее 10 раз.
В группе по всем не менее 10 учетным листьям, принятым на одной локальной зоне поверхности кроны дерева, статистическим моделированием выявляют среднестатистические закономерности динамики роста учетных листьев, произрастающих на одной локальной зоне кроны дерева, по четырем параметрам учетных листьев - длины и ширины листа, периметра и площади листа, а при необходимости выявляют биотехнические закономерности и по расчетным относительным параметрам учетных листьев.
Измерения и статистическое моделирование проводят от начала распускания почек до любого этапа онтогенеза учетных листьев для деревьев - также с учетом времени активной жизнедеятельности растений от начала распускания почек, при этом время исчисляется в сутках, то есть в циклах обращения Земли вокруг самого себя, а измерения с фотографированием учетных листьев через прозрачную палетку с делениями 2 мм проводят примерно в одно и то же время суток, например около 15 часов, для повышения точности выявления закономерностей динамики параметров учетных листьев статистическим моделированием измерения в условиях средней полосы европейской части Российской Федерации проводят не менее чем до конца августа.
Каждый учетный лист измеряется без срезки от растения и через прозрачную палетку для картографических измерений фотографируется, например, применением сотового телефона с памятью для хранения фотографий, как один цельный объект, а сами измерения параметров каждого учетного листа проводят на компьютере на увеличенном до формата A4 фотографии учетного листа по клеткам через 2 мм на сетке у прозрачной палетки, причем до фотографирования продольная ось листа растения совмещается с одной из линий сетки прозрачной палетки примерно в середине ширины палетки.
Для выявления биотехнической закономерности динамики длины и ширины листа во времени вегетации применяют обобщенную двухчленную формулу:
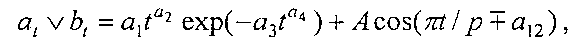
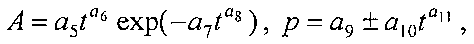
где at, bt - переменные во времени среднестатистические длина или ширина у популяции из не менее 10 учетных листьев, мм,
 - знак логических отношений «или»,
- знак логических отношений «или»,
t - время вегетации с начала распускания листьев, сутки,
A - амплитуда (половина) колебательного возмущения длины или ширины листьев в динамике роста популяции из не менее 10 листьев, мм,
p - полупериод колебания длины или ширины у не менее 10 учетных листьев, сутки,
a1…a12 - параметры модели, получающие числовые значения после статистического моделирования идентификацией модели по данным измерений длины или ширины у не менее 10 учетных листьев, при этом происходит чередование знаков:
+a10, -a12 - принимают для закономерности динамики среднестатистической длины at у популяции из не менее 10 учетных листьев;
-a10, +a12 - принимают для закономерности динамики среднестатистической ширины bt у популяции из не менее 10 учетных листьев.
Для выявления биотехнической закономерности динамики периметра и площади листа во времени вегетации рекомендуют обобщенную формулу:
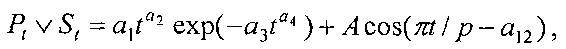
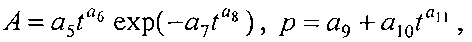
где Pt, St - переменные во времени среднестатистические периметр (см) или площадь (см2) у популяции из не менее 10 учетных листьев,
 - знак логических отношений «или»,
- знак логических отношений «или»,
t - время вегетации с начала распускания листьев, сутки,
A - амплитуда (половина) колебательного возмущения периметра (см) или площади (см2) в динамике среднестатистического роста популяции из не менее 10 учетных листьев,
p - полупериод колебательного возмущения при адаптации к внешним воздействиям периметра или площади учетных листьев, сутки,
a1…a12 - параметры модели, получающие числовые значения после статистического моделирования идентификацией модели по данным измерений периметра или площади у не менее 10 учетных листьев.
Сущность изобретения заключается в том, что в кроне дерева на удобной для измерений высоте на боковой поверхности кроны дерева выделяют локальную зону с однообразным внешним воздействием солнечного света, на которой выбирают не менее 10 учетных листьев, каждый из которых отмечают меткой в виде куска белой нитки, привязанной к основанию листа на черешке. При этом локальную зону выбирают примерно в виде круга диаметром не более 0,5 м, а в качестве учетных принимают листья, расположенные на поверхности кроны с освещенной утренними солнечными лучами стороны кроны дерева.
Сущность изобретения заключается также в том, что за вегетационный период по суткам от начала распускания почек листьев (по дате начала вегетационного периода время t принимается равным нулю) с фотографированием каждого учетного листа через прозрачную палетку с сеткой 2 мм. При этом фотографирование до конца вегетационного периода выполняют не менее 10 раз, начиная не менее чем через неделю после распускания почек листьев дерева.
Сущность изобретения заключается также и в том, что по всем не менее 10 учетным листьям, принятым на одной локальной зоне кроны дерева, статистическим моделированием выявляют среднестатистическую закономерность динамики роста учетных листьев, произрастающих на одной локальной зоне кроны дерева, по четырем параметрам учетных листьев: длины и ширины листа, периметра и площади листа. При необходимости могут быть выявлены закономерности и по расчетным относительным параметрам учетных листьев.
Сущность изобретения заключается также и в том, что измерения и статистическое моделирование проводят от начала вегетационного периода до любого этапа онтогенеза учетных листьев для деревьев - также с учетом времени активной жизнедеятельности растений от начала распускания почек. При этом время исчисляется в сутках, то есть в циклах обращения Земли вокруг самого себя. А измерения с фотографированием учетных листьев через прозрачную палетку с делениями 2 мм проводят примерно в одно и то же время суток, например около 15 часов. Для повышения точности выявления закономерностей динамики параметров учетных листьев статистическим моделированием измерения в условиях средней полосы европейской части Российской Федерации проводят не менее чем до конца августа.
Новизна технического решения заключается в том, что впервые рассматривается средняя динамика поведения не менее 10 учетных листьев в одной локальной зоне на поверхности кроны дерева. При этом среднестатистическая закономерность из 10 учетных листьев идентифицируется по данным измерений не менее 10 раз с начала распускания почек, причем не менее чем до конца августа.
При этом каждый учетный лист измеряется без срезки от растения и через прозрачную палетку для картографических измерений фотографируется, например, применением сотового телефона с памятью для хранения фотографий, как один цельный объект. А сами измерения на компьютере на увеличенном до формата А4 фотографии учетного листа выполняются по клеткам через 2 мм у палетки. Причем до фотографирования продольная ось листа растения совмещается с одной из линий примерно в середине ширины палетки.
Положительный эффект заключается в том, что на поверхности кроны дерева можно выделять разные локальные зоны с учетом освещенности кроны и учетом других влияющих на развитие и рост учетных листьев факторов. Принятие с одной локальной зоны не менее 10 учетных листьев достаточно для идентификации тренда с волновой составляющей у биотехнической закономерности по четырем первичным параметрам (длина, ширина, периметр, площадь) учетных листьев. Для более глубокого научного анализа возможен расчет и дополнительных вторичных параметров.
Положительный эффект также заключается в том, что впервые точно можно выявлять закономерности динамики поведения по измеренным 10 учетным листам в конкретной одной локальной зоне кроны дерева. Даже на такой локальной зоне каждый из 10 листьев липы оказываются очень разными по динамике роста, что свидетельствует о высокой разрешающей способности предлагаемого способа при учете разных факторов локальной зоны на поверхности кроны дерева на динамику роста каждого учетного листа. При этом появляется возможность, кроме абсолютных значений измеренных четырех параметров листа, дополнительно учитывать и расчетные параметры в виде скорости и ускорений роста листьев дерева от начала распускания почек до любого этапа онтогенеза учтенных листьев вплоть до конца вегетационного периода.
Таким образом, предлагаемое научно-техническое решение обладает существенными признаками, новизной и положительным эффектом. В научно-технической и патентной литературе информационных материалов, порочащих новизну предлагаемого изобретения, нами не обнаружено.
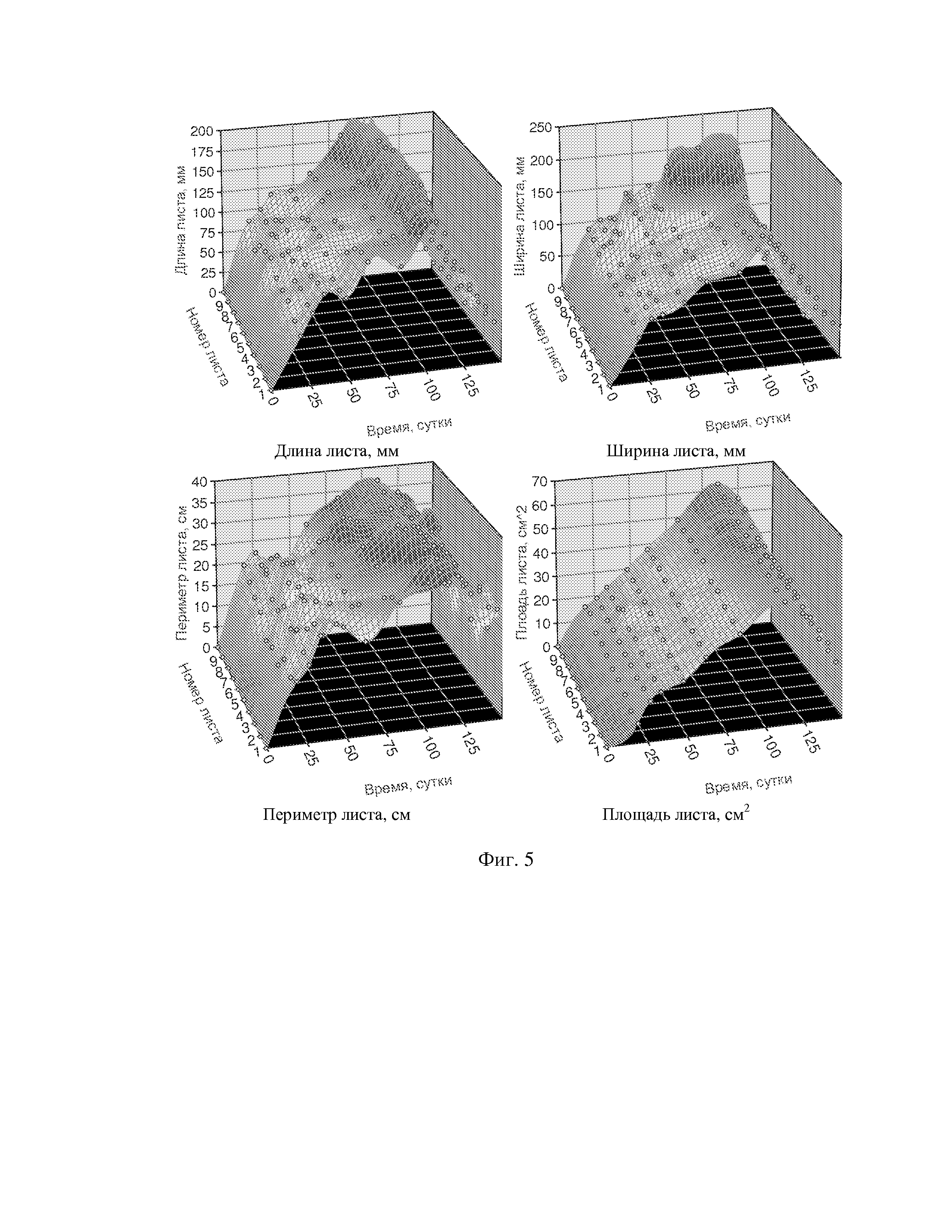
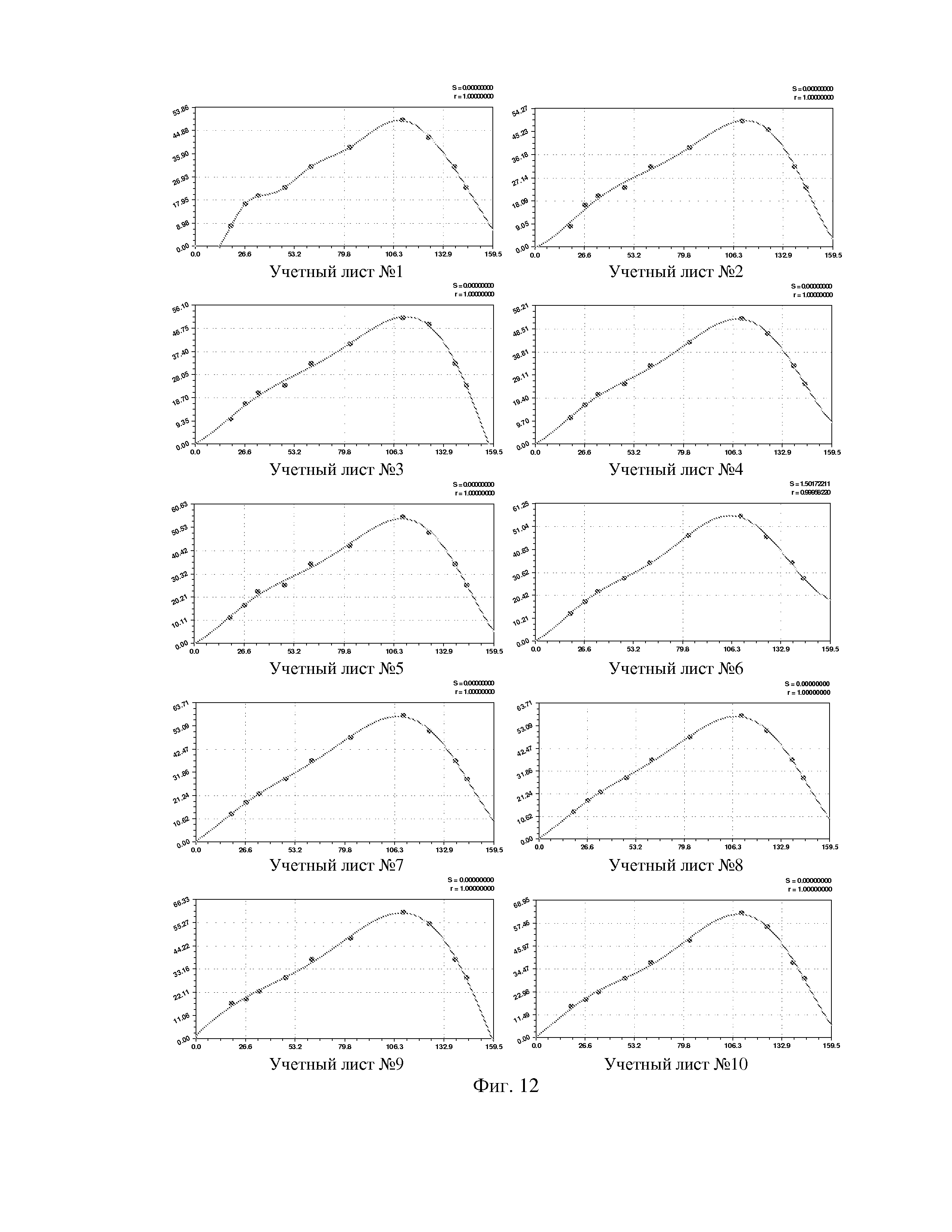
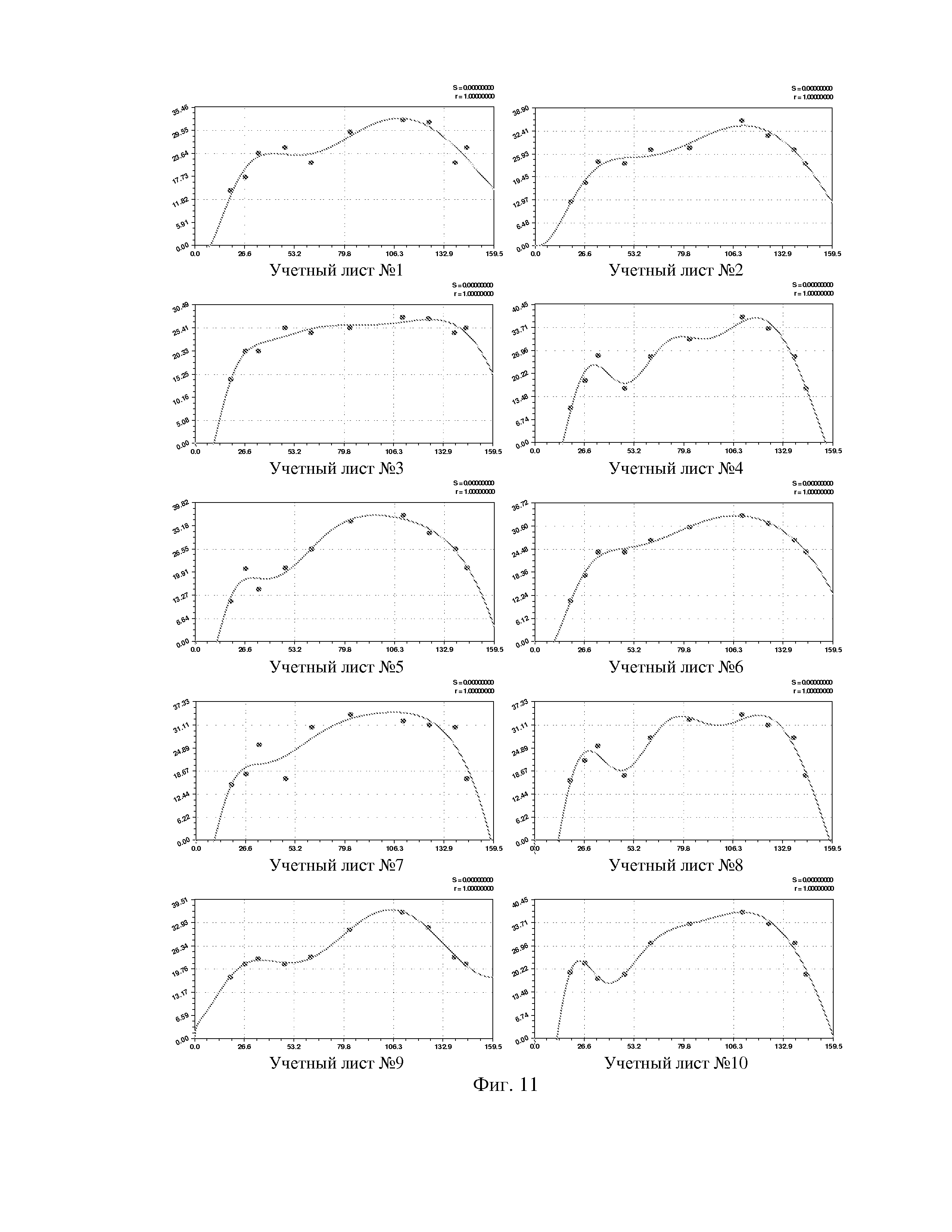
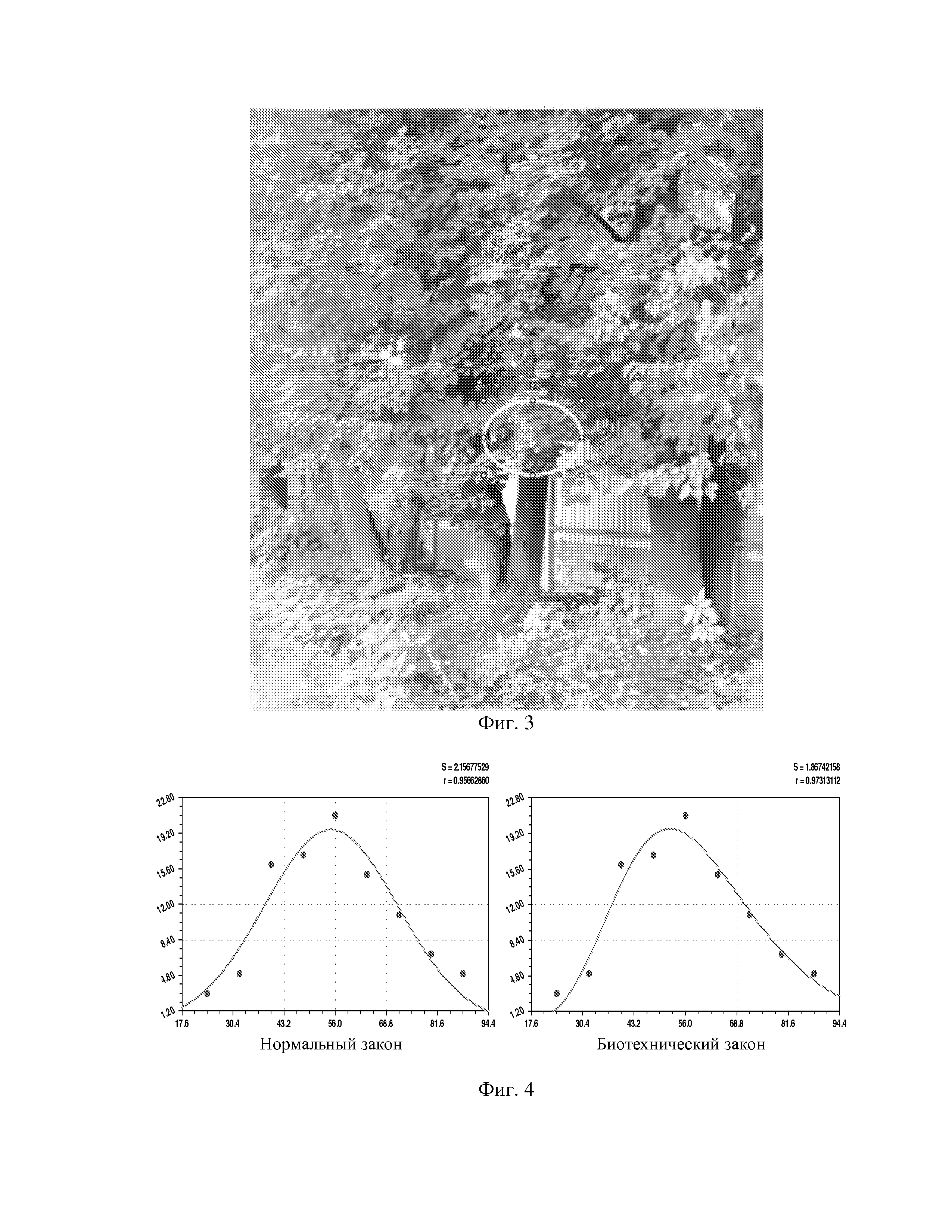
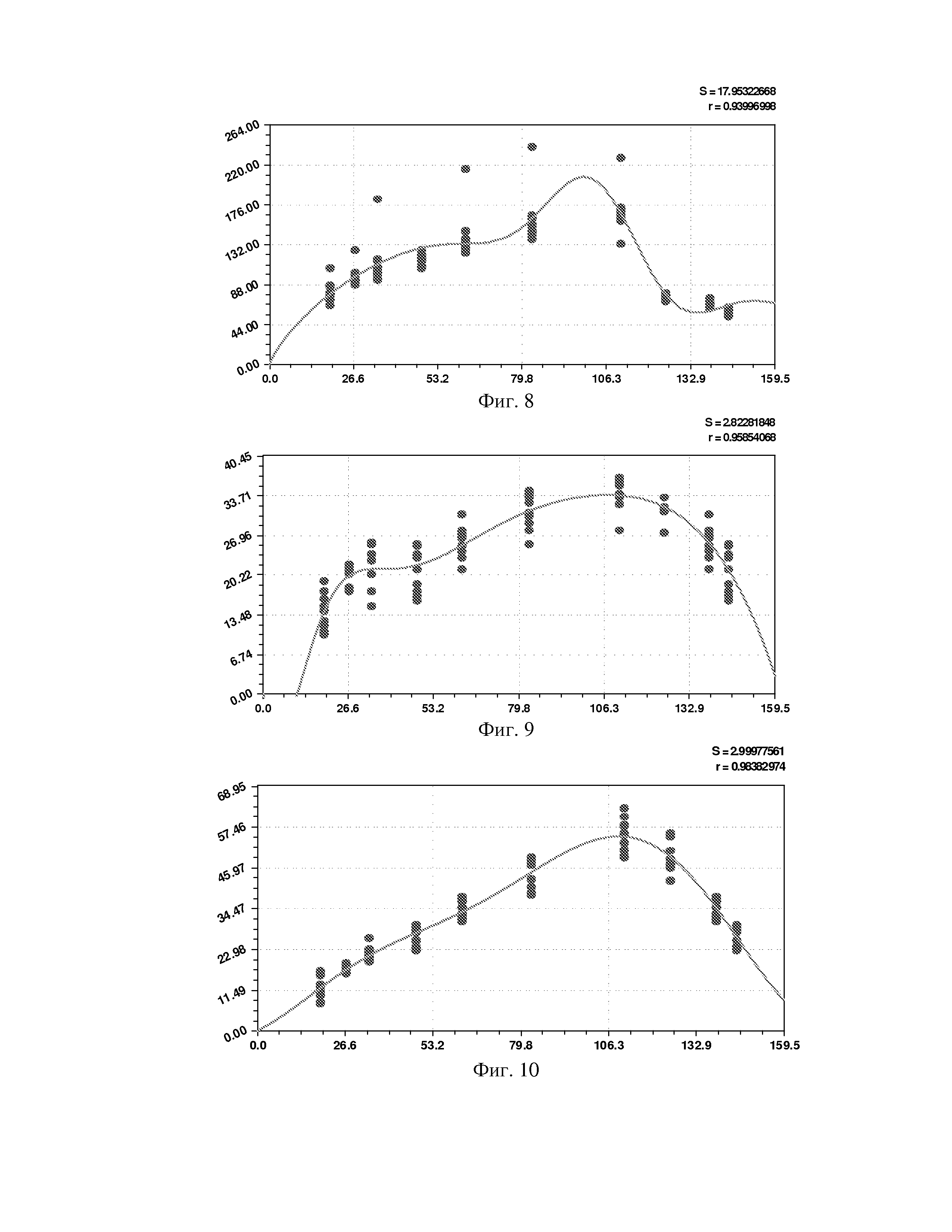
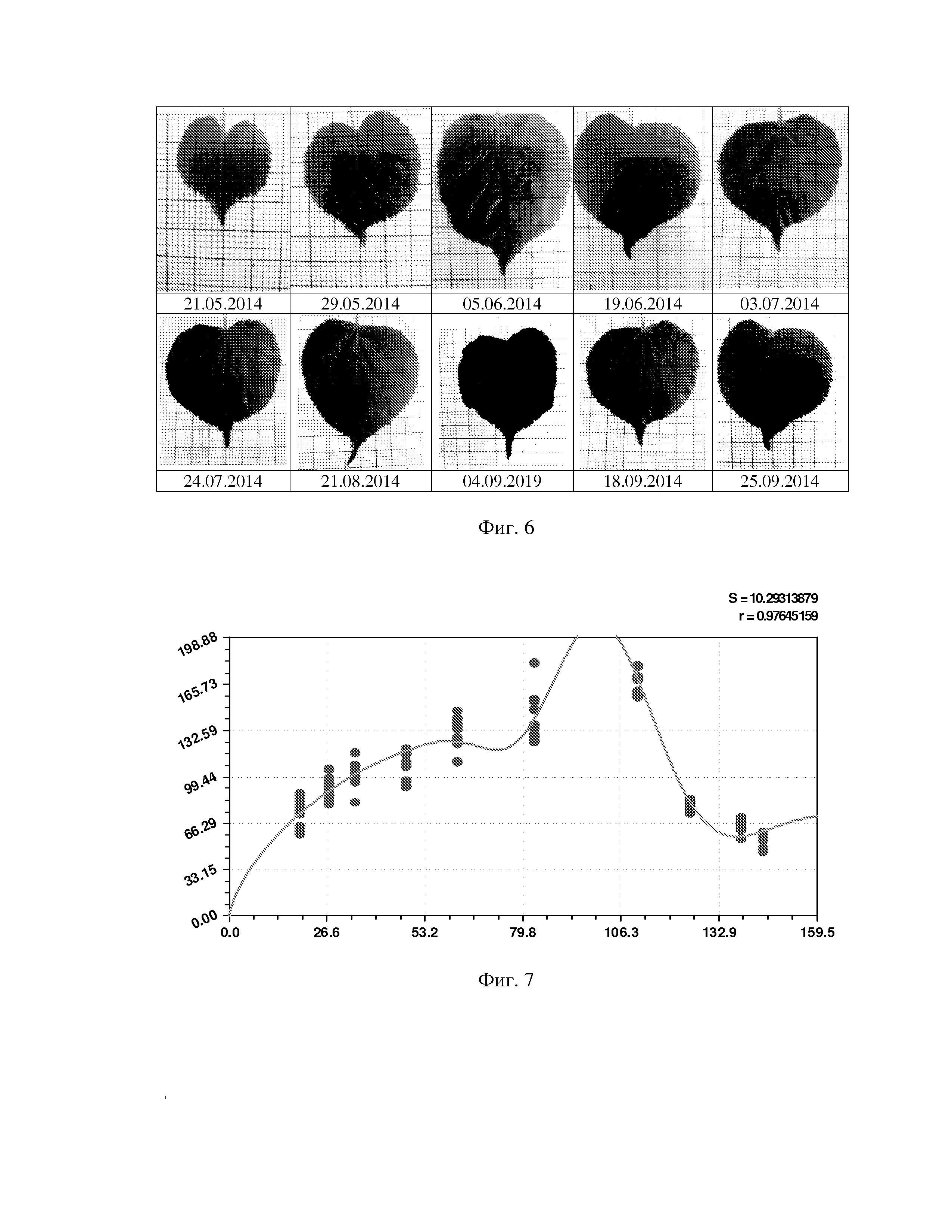
На фиг. 1 приведен спутниковый снимок части города Йошкар-Ола, на котором стрелкой показано дерево липы, причем вершина стрелки ориентирована на локальную зону кроны дерева, где выделены 10 учетных листьев; на фиг. 2 показан увеличенный спутниковый снимок дерева липы, причем локальная зона кроны липы расположена со стороны ботанического сада и она находится в чистой воздушной среде (от загрязнения с улицы и относительно малой интенсивности движения автомобилей защищает высокий забор и вся крона липы); на фиг. 3 приведена фотография трех лип, из которых на средней липе кружком выделена локальная зона кроны липы для выделения из листвы 10 учетных листьев; на фиг. 4 даны графики двух законов распределения всех 100 измерений (10 учетных листьев по 10 замеров у каждого листа за вегетационный период); на фиг. 5 приведены пространственные графики изменения четырех параметров учтенных листьев в зависимости от времени вегетационного периода и номера листа; на фиг. 6 как пример показаны фотографии листа №1 за вегетационный период (разница в фотографиях объясняется поворотом к фотоаппарату лицевой или тыльной стороной листа); на фиг. 7 показан график среднестатистической закономерности динамики длины листьев; на фиг. 8 - то же на фиг. 7 ширины листьев; на фиг. 9 - то же на фиг. 7 периметра листьев; на фиг. 10 - то же на фиг. 7 площади листьев; на фиг. 11 приведены графики динамики периметра каждого учетного листа из группы из 10 учетных листьев на одной локальной зоне поверхности кроны липы; на фиг. 12 - то же на фиг. 11 динамики площади каждого учетного листа.
Способ измерения динамики роста листьев дерева, например от начала распускания почек до любой стадии вегетационного периода не менее чем до конца августа, включает такие действия.
В отличие от прототипа выбранный учетный лист не срезают, а помечают меткой в виде бирки с номером учтенного листа на белой нитке, привязанной к основанию учтенного листа за черешок. Измерению подвергают каждый учетный лист. Поэтому способ измерения индивидуален, а выявление закономерностей - групповое по не менее 10 учетным листьям, выделенным на поверхности кроны дерева примерно с одной освещенностью в виде локальной зоны.
К каждому измеряемому листу без его повреждений подводится прозрачная палетка для картографических измерений с сеткой с делениями 2 мм, причем вручную берут пальцами левой руки за черешок учтенного листа, а палетку укладывают сверху на учетный лист. Затем палетку прижимают к листу и держат левой рукой так, чтобы не сорвать учетный лист. После этого в правую руку берут сотовый телефон, и лист через прозрачную палетку фотографируют. При этом продольная ось листа дерева совмещается с одной из линий сетки палетки примерно в середине палетки.
После проведения всех действий со всеми учетными листьями фотографии помещаются в память компьютера, а измерения длины, ширины, периметра и площади листа выполняются по клеткам сетки палетки на увеличенном до формата А4 изображении листа растения. После проведения измерений у необходимой группы листьев статистическим моделированием выявляют биотехнические закономерности изменения параметров листьев в зависимости от времени в сутках.
До проведения измерений у необходимой группы листьев у фотографического изображения каждого листа вначале обрезают края так, чтобы лист оказался в описанном прямоугольнике с сантиметровыми клетками, после этого рисунок листа увеличивают до формата А4, например, располагая поворотом рисунка измеряемого листа растения вдоль длинной стороны формата А4, причем четкость изображения здесь не имеет значения - лишь бы хорошо были видны малые клетки у сетки палетки.
На боковой поверхности кроны дерева выделяют локальную зону с примерно одинаковым освещением, на которой выбирают группу учетных листьев с метками. Измерения параметров учетных листьев проводят за вегетационный период по суткам от начала распускания почек листьев с фотографированием каждого учетного листа. Причем первое фотографирование выполняют не менее чем через неделю и не позже чем три недели после распускания почек листьев дерева. В течение времени вегетации проводят измерения параметров учтенных листьев, а затем в группе по всем учетным листьям, принятым на одной локальной зоне поверхности кроны дерева, статистическим моделированием выявляют среднестатистические закономерности динамики роста учетных листьев по отдельным параметрам учетных листьев, произрастающих на одной локальной зоне поверхности кроны дерева.
В кроне дерева на удобной для измерений высоте на боковой поверхности кроны дерева выделяют локальную зону с примерно с одинаковым освещением, на которой выбирают группу из не менее 10 учетных листьев, каждый из которых отмечают меткой в виде куска белой нитки, привязанной к основанию листа на черешке. При этом локальную зону выбирают примерно в виде круга диаметром не более 0,5 м, а в качестве учетных принимают листья, расположенные на поверхности кроны с освещенной утренними солнечными лучами стороны кроны дерева.
Дополнительно к длине, ширине и площади учетных листьев измеряют периметр листа с учетом масштаба клеток прозрачной палетки по количеству клеток, через которые проходит ребристая линия края листа. При этом площадь листа определяют как сумму полных клеток внутри поверхности листа и неполных клеток вдоль периметра листа также с учетом масштаба клеток палетки.
Периметр листа измеряют по количеству клеток, через которые проходит ребристая линия края листа с учетом масштаба клеток палетки 2×2 мм, по формуле:
P=0,28284IP,
где P - периметр листа, см;
IP - количество по периферии листа неполных клеток, шт.
Площадь листа определяют как сумму полных клеток внутри поверхности листа и неполных клеток вдоль периметра листа также с учетом масштаба клеток палетки 2×2 мм, по формуле:
S=0,04IS+0,02IP,
где S - площадь листа, измеряемая количеством клеток размерами 2×2 мм внутри листа и количеством клеток периметра листа, см2;
IS - количество на изображении листа полных клеток, шт.;
IP - количество по периферии листа неполных клеток, шт.
Измерения параметров учетных листьев проводят за время вегетации по суткам от начала распускания почек листьев с фотографированием каждого учетного листа через прозрачную палетку с сеткой 2 мм. Причем первое фотографирование выполняют не менее чем через неделю и не позже чем три недели после распускания почек листьев дерева. При этом по дате начала времени вегетации текущее время в сутках принимают за влияющую переменную. Причем значение влияющей переменной в начале вегетации листьев принимают равным нулю, а визуально параметры учетных листьев по длине и ширине, периметру и площади учетных листьев в начале вегетации с принимают также с нулевыми значениями, при этом фотографирование до конца вегетации выполняют не менее 10 раз.
В группе по всем не менее 10 учетным листьям, принятым на одной локальной зоне поверхности кроны дерева, статистическим моделированием выявляют среднестатистические закономерности динамики роста учетных листьев, произрастающих на одной локальной зоне кроны дерева, по четырем параметрам учетных листьев - длины и ширины листа, периметра и площади листа. А при необходимости выявляют биотехнические закономерности и по расчетным относительным параметрам учетных листьев.
Измерения и статистическое моделирование проводят от начала распускания почек до любого этапа онтогенеза учетных листьев для деревьев - также с учетом времени активной жизнедеятельности растений от начала распускания почек. При этом время исчисляется в сутках, то есть в циклах обращения Земли вокруг самого себя, а измерения с фотографированием учетных листьев через прозрачную палетку с делениями 2 мм проводят примерно в одно и то же время суток, например около 15 часов. Для повышения точности выявления закономерностей динамики параметров учетных листьев статистическим моделированием измерения в условиях средней полосы европейской части Российской Федерации проводят не менее чем до конца августа.
Каждый учетный лист измеряется без срезки от растения и через прозрачную палетку для картографических измерений фотографируется, например, применением сотового телефона с памятью для хранения фотографий, как один цельный объект. А сами измерения параметров каждого учетного листа проводят на компьютере на увеличенном до формата А4 фотографии учетного листа по клеткам через 2 мм на сетке у прозрачной палетки. Причем до фотографирования продольная ось листа растения совмещается с одной из линий сетки прозрачной палетки примерно в середине ширины палетки.
Для выявления биотехнической закономерности динамики длины и ширины листа во времени вегетации применяют обобщенную двухчленную формулу:
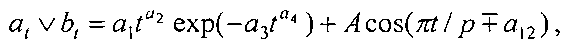
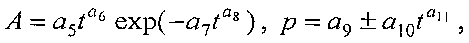
где at, bt - переменные во времени среднестатистические длина или ширина у популяции из не менее 10 учетных листьев, мм,
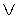 - знак логических отношений «или»,
- знак логических отношений «или»,
t - время вегетации с начала распускания листьев, сутки,
A - амплитуда (половина) колебательного возмущения длины или ширины листьев в динамике роста популяции из не менее 10 листьев, мм,
p - полупериод колебания длины или ширины у не менее 10 учетных листьев, сутки,
a1…a12 - параметры модели, получающие числовые значения после статистического моделирования идентификацией модели по данным измерений длины или ширины у не менее 10 учетных листьев, при этом происходит чередование знаков:
+a10, -a12 принимают для закономерности динамики среднестатистической длины at у популяции из не менее 10 учетных листьев;
-a10, +a12 принимают для закономерности динамики среднестатистической ширины bt у популяции из не менее 10 учетных листьев.
Для выявления биотехнической закономерности динамики периметра и площади листа во времени вегетации рекомендуют обобщенную формулу:
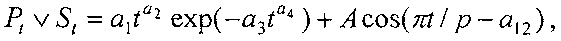
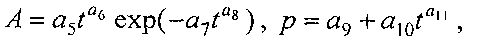
где Рt, St - переменные во времени среднестатистические периметр (см) или площадь (см2) у популяции из не менее 10 учетных листьев,
 - знак логических отношений «или»,
- знак логических отношений «или»,
t - время вегетации с начала распускания листьев, сутки,
A - амплитуда (половина) колебательного возмущения периметра (см) или площади (см2) в динамике среднестатистического роста популяции из не менее 10 учетных листьев,
p - полупериод колебательного возмущения при адаптации к внешним воздействиям периметра или площади учетных листьев, сутки,
a1…a12 - параметры модели, получающие числовые значения после статистического моделирования идентификацией модели по данным измерений периметра или площади у не менее 10 учетных листьев.
Пример. Опыты были проведены в городе Йошкар-Оле (фиг. 1) в вегетационный период 2014 года на одних и тех же 10 учетных листьях липы (фиг. 2), расположенной в ботаническом саду по улице Красноармейской. Учетные листья выбирали в одной локальной зоне (фиг. 3) на высоте примерно 1,6 м с восточной стороны освещения липы.
Каждый учетный лист помечали меткой в виде куска белой нитки с биркой с номером листа, привязанной к черешку у основания листа.
Способ измерения площади листьев липы включает такие действия: на лист сверху кладут прозрачную палетку с сеткой, например, с малыми клетками размерами 2×2 мм, так, чтобы средняя линия вдоль палетки совпала с осью продольной жилки листа. Затем лист с палеткой фотографируют, например, цифровым фотоаппаратом с функциями фотографирования и хранения множества фотоснимков в памяти.
У каждого листа взять его фото и подрезать на компьютере лишние кромки, а затем увеличить для удобства подсчетов клеток до формата А4.
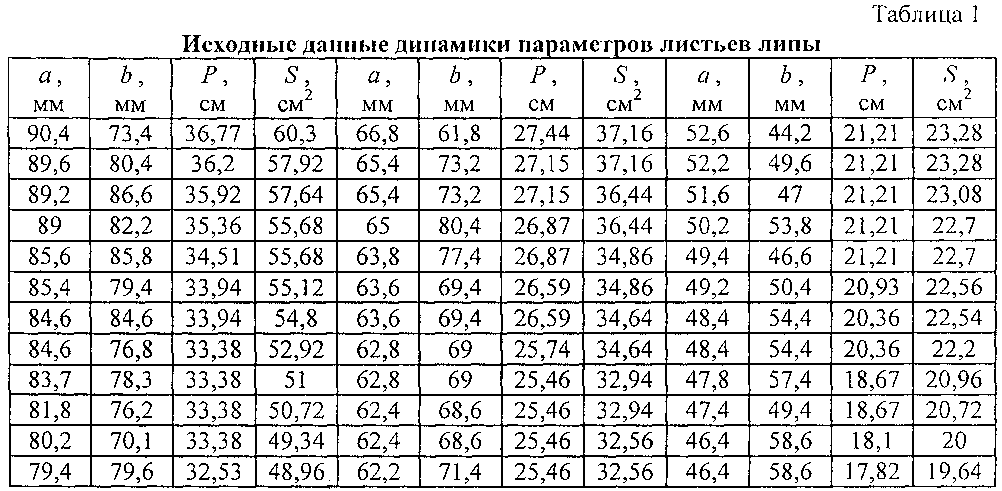
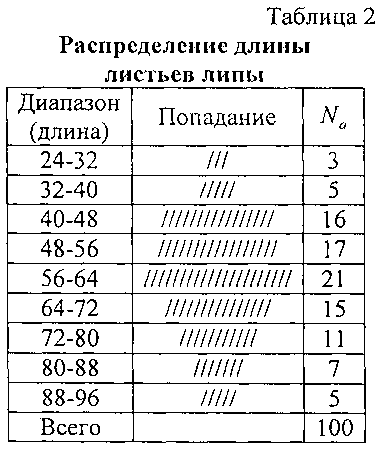
Исходные данные по замерам приведены в таблице 1, вначале без указания даты измерения. Поэтому вначале рассмотрим все 100 измерений как одну статистическую выборку для классического анализа.
=
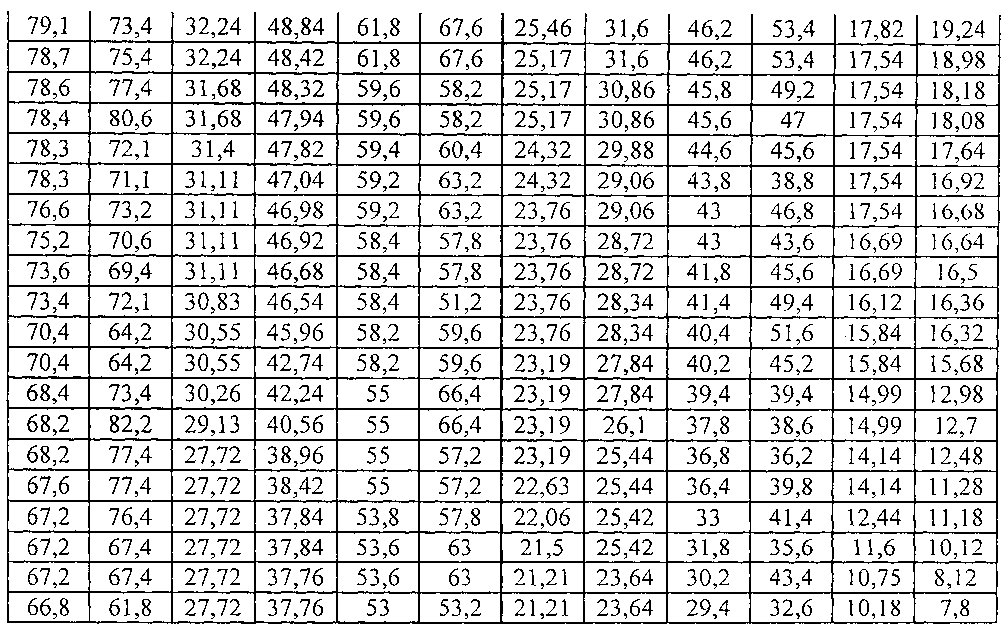
В таблице 1 даны следующие условные обозначения физическим параметрам листа:
a - длина листа вдоль главной жилки, измеряемая от места соединения черешка с листовой пластиной растения до конца вершинки листа, мм;
b - ширина листа по крайним точкам поперек листовой пластины или же общая ширина листа в наибольшем поперечном сечении листа, мм;
P - периметр листа, см;
S - площадь листа, измеряемая количеством клеток размерами 2×2 мм и количеством клеток периметра листа, см2.
Периметр и площадь учетного листа вычисляют по формулам:
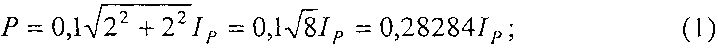

Для множества из 100 измерений длина листа изменяется в интервале от 29,4 мм до 92,2 мм. Ширина листа изменяется в пределах 32,6-80,2 мм. Периметр изменяется от 10,18 до 36,77 см. Площадь листа изменяется от 7,8 до 62,68 см2.
Для определения ширины разряда Δx используют приближенную формулу, предложенную Г.А. Стреджесом:

где xmax - xmin - размах вариации; N - число замеров.
В нашем случае N=100. Поэтому в знаменателе будет число 1+3,322×2=7,644. Значение xman - xmin по длине листа изменяется в интервале 62,8 мм, у ширины листа 47,60 мм, периметра 26,59 см, площади листа 54,88 см2.
Значение Δx будет равным: Длина листа 8,21 или примерно 8 мм: 96, 88, 80, 72, 64, 56, 48, 40, 32, 24.
Ширина листа 6,22 или около 6 мм: 84, 78, 72, 66, 60, 54, 48, 42, 36, 30. Периметр 3,47 или около 4 см: 40, 36, 32, 28, 24, 20, 16, 12, 8. Площадь листа 7,17 или около 7 см2: 63, 56, 49, 42, 35, 28, 21, 14, 7.
Нормальный закон распределения играет исключительно важную роль в существующей теории математической статистики и поэтому ныне занимает среди других законов распределения особое положение. Это - наиболее часто встречающийся на практике закон распределения.
В нашем случае кривая распределения по нормальному закону имеет симметричный холмообразный вид, который соответствует стандартному нормальному распределению.
А биотехнический закон оказывается точнее нормального из-за асимметричности расположения двух частей графика
На фигуре 4 даны графики законов распределения длины учетных листьев (по остальным параметрам учетных листьев получились аналогичные законы распределения) липы (табл. 2):
- нормальный
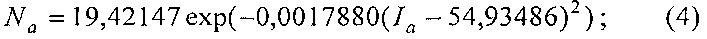
- биотехнический
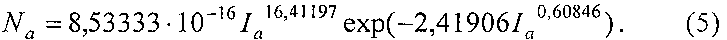
Коэффициент корреляции нормального закона равен 0,9566, а у биотехнического закона 0,9731.
Поэтому, при прочих равных условиях, биотехнический закон точнее отражает распределение фактических значений у всех четырех параметров листьев липы.
Из таблицы 2 и фигуры 4 видны остатки от формул (4) и (5), которые в дальнейшем можно идентифицировать волновыми функциями.
В таблице 3 приведены исходные данные для построения пространственных графиков динамики четырех основных параметров учетных листьев (фиг. 5) и последующего статистического моделирования динамики без учета номера учетных листьев.
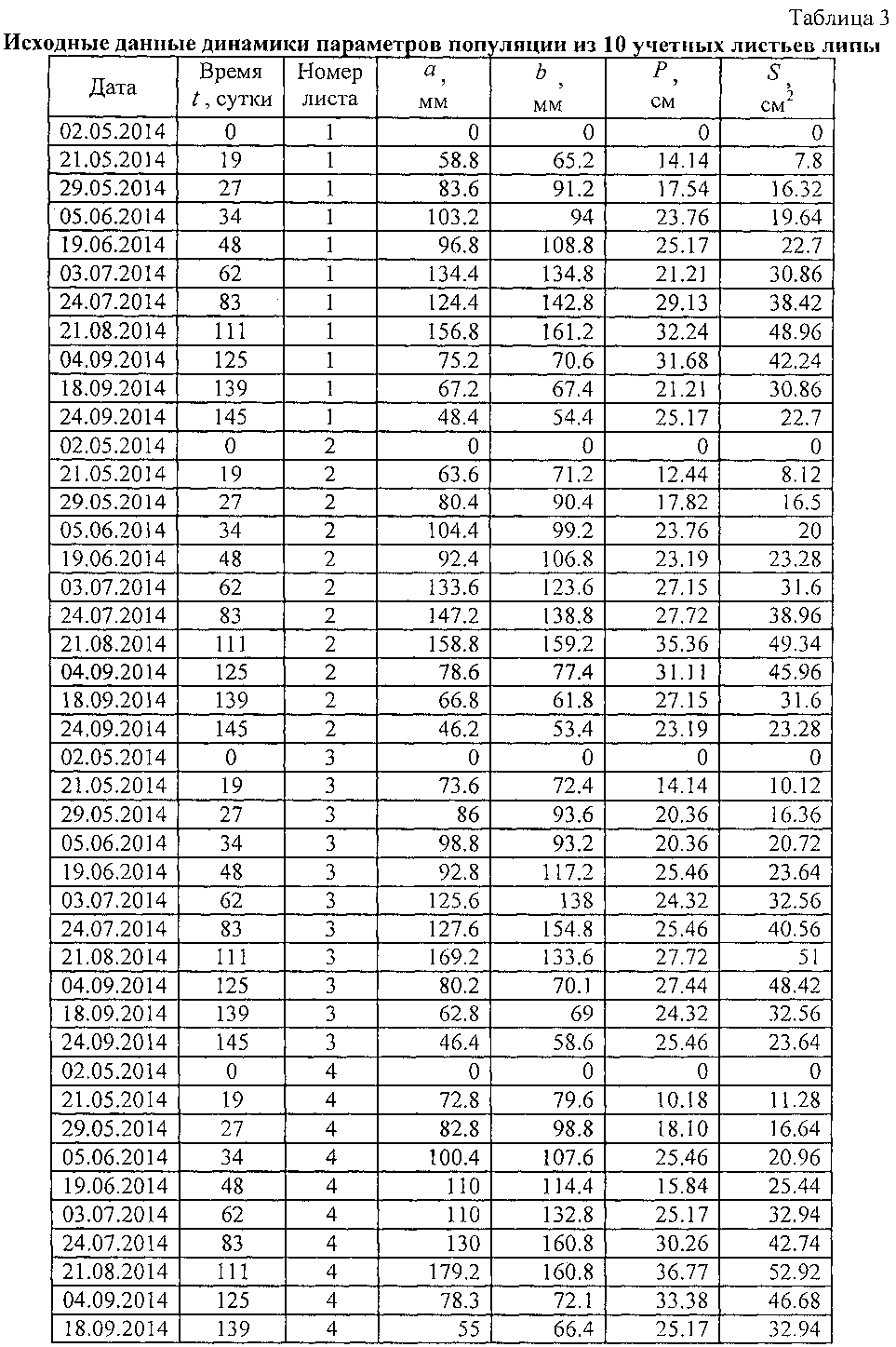
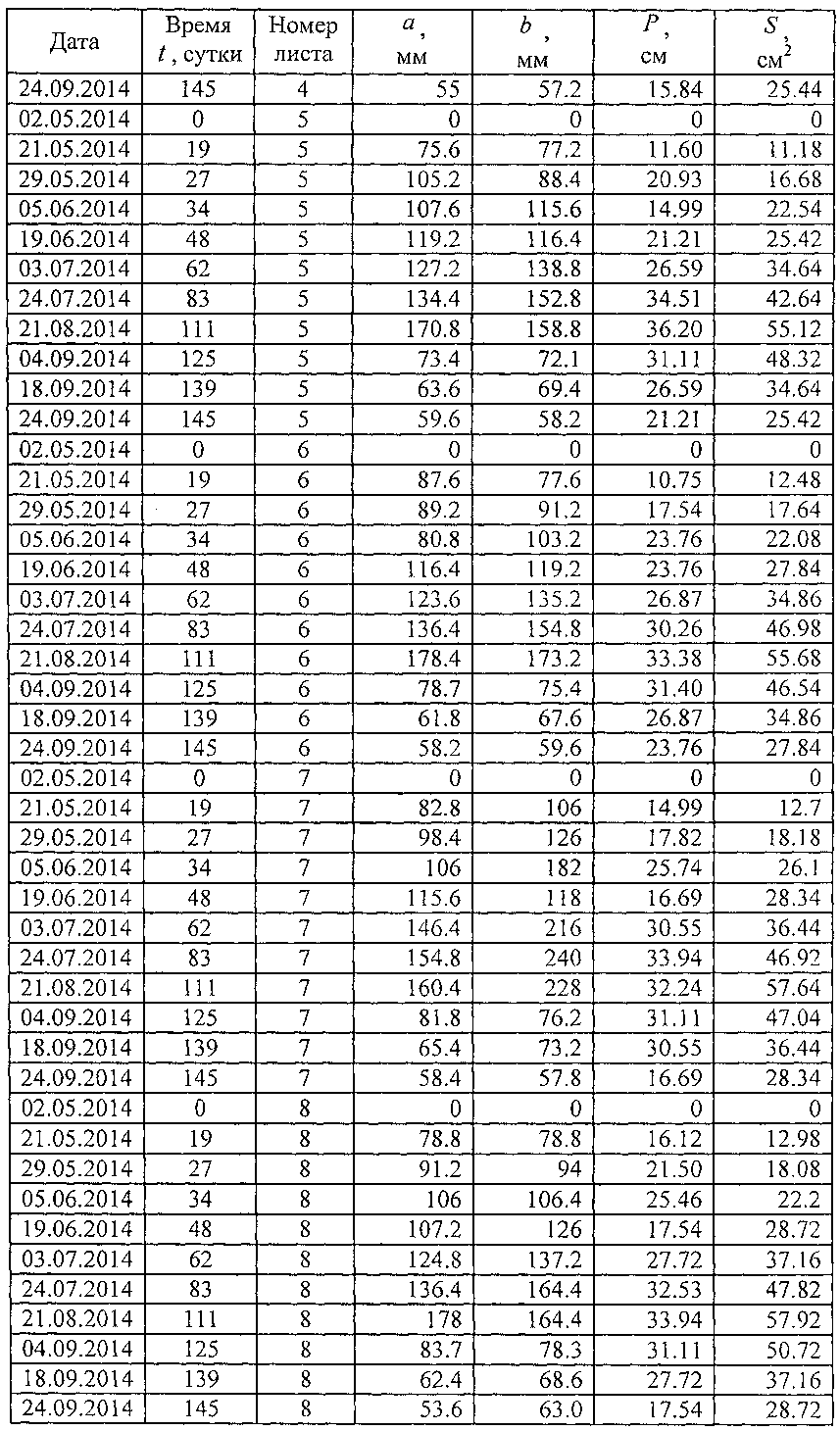
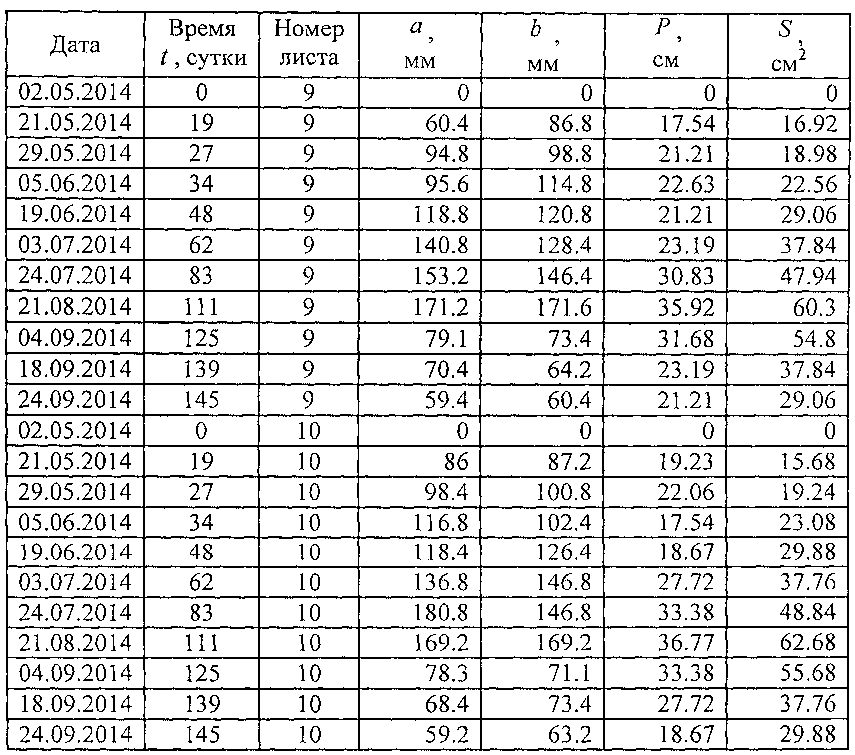
В сравнении с данными таблицы 1 в таблице 3 дополнены первые три столбца: дата, время t, сутки и номер листа. Время t становится объясняющей переменной, а параметры учетного листа - зависимыми параметрами (факторами) или показателями. Поэтому статистическим моделированием идентифицируются закономерности в виде структур a=f(t), b=f(t), P=f(t) и S=f(t).
Номер листа является только вспомогательной переменной, не имеющей физического (в нашем случае физиологического) смысла. Он нами применен только для построения пространственных графиков, показывающих различие в характере поведения каждого учетного листа. Таким образом, становится понятным, что каждый учетный лист испытывает различное внешнее воздействие. Это позволяет в дальнейших опытах учитывать те влияющие факторы среды, освещенности, расположения в кроне дерева и другие, которые отличают поведение в процессе жизнедеятельности каждого учетного листа от другого.
В таблице 3 за начало вегетационного периода принята дата 02.05.2014, то есть дата начала распускания почек или процесса видимого роста и развития учетных листьев. Для этой даты принимаются нулевые значения всех четырех изучаемых основных параметров учетных листьев.
По-видимому, принятие за начало вегетационного периода начала процесса сокодвижения (примерно 21.04.2014) в будущем позволит изучать и физиологические процессы, происходящие в листве дерева.
Из графиков на фигуре 5 заметно, что до периода роста учетных листьев в 30 суток все четыре показателя имеют в динамике гладкий характер. Здесь происходит быстрый рост без волновых возмущений.
Очевидно, что тогда существует оптимальное значение второго (первое измерение визуальное по дате 02.05.2014 с нулевыми значениями параметров) измерения в пределах от 7 до 21 суток от начала вегетационного периода по распусканию почек. Меньше недели после распускания почек листья еще очень малы и поэтому погрешность измерений параметров учетных листьев в сетке 2 мм прозрачной палетки значительно возрастает. Если же второе измерение проводят после трех недель, то существует опасность недостаточной фиксации начала колебательных возмущений в росте учтенных листьев. Из данных таблицы 3 видно, что в наших экспериментах втрое измерение проводилось через 19 суток, что находится в пределах допустимого интервала 7-21 суток.
Дальнейшее сравнение графиков на фигуре 5 показывает, что после 30 суток динамика поведения каждого учетного листа сильно меняется. Это связано с тем, что в популяционной динамике учетных листьев наблюдаются колебательные возмущения, различные для среды обитания каждого учетного листа. Примерно через месяц после распускания почек листья начинают самостоятельную жизнь, причем вплоть до конца своего существования. Эти процессы можно изучать через дополнительные параметры в виде скорости и ускорений роста листьев.
Сравнение графиков на фигуре 5 также показывает, что линейные параметры (длина, ширина и периметр) более чувствительны к динамике роста учетных листьев, а площадные параметры - менее чувствительны и на динамику площади учетных листьев среднестатистическим обоснованием индивидуальные влияния на каждый учтенный лист выравниваются. Поэтому следует ожидать, что при измерениях объемных параметров учетных листьев образуется еще большее нивелирование колебательных возмущений отдельных учетных листьев.
Но одно ясно, на что показывает динамический ряд на фигуре 6 изменений листа №1, что наибольшая перспектива по чувствительности имеются именно у линейных параметров листа. При этом изменяется также и форма листа во времени: одни части учтенного листа развиваются быстрее, чем другие части. Это - на будущее по поиску новых технических решений измерения динамики листьев как цельного растения, например, по динамике жилок и углов между ними.
Далее покажем закономерности среднестатистической динамики четырех основных параметров у 10 учетных листьев липы.
В таблице 4 показан фрагмент из данных таблицы 3 по листу №1.
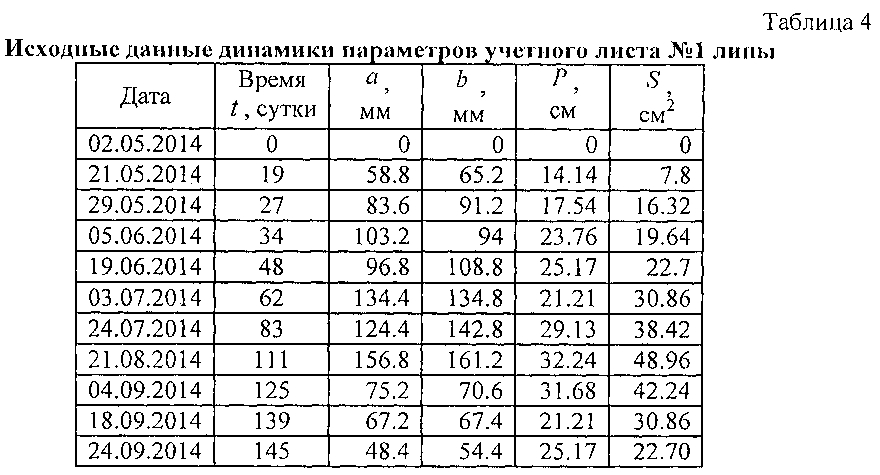
В таблице 4 удален столбец «Номер листа». Можно моделировать данные по каждому учетному листу. Однако мы не знаем влияющие факторы среди популяции из 10 листьев, выбранных как учетные в локальной зоне на поверхности кроны липы. Поэтому будет искать по всем 10 учетным листьям из данных по таблице 3 среднестатистическую линию поведения у всех 10 учетных листьев.
Длина листа изменяется (фиг. 7) по формуле
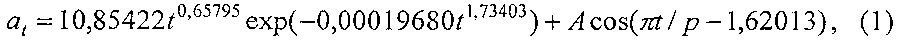
A=4,75240·10-33t20,44582exp(-0,063164t1,19166),
p=2,99774+0,10895t0,94969,
где at - переменная во времени среднестатистическая длина листьев, мм,
t - время вегетации с начала распускания листьев, сутки,
A - амплитуда (половина) колебательного возмущения длины листьев в динамике роста популяции из 10 листьев, мм,
p - полупериод колебания длины учетных листьев, сутки.
Первый член формулы (1) является биотехническим законом, а половина амплитуды у второго члена в виде волновой функции также является биотехническим законом. При этом второй член имеет перед собой положительный знак, что указывает на позитивное влияние волнового возмущения на рост длины листьев. Тем самым становится понятным, что волновая адаптация растений к окружающей сред является основным механизмом их роста и развития. При этом в начале вегетации при распускании почек на дату 02.05.2014 период колебания был равен 2×2,99774≈6 суток, а начало колебания произошло чуть позже даты 02.05.2014 (примерно 3-5 суток). Затем, по мере взросления организма листа, этот период возрастает, то есть колебательная адаптация успокаивается.
Адекватность формулы (1) по коэффициенту корреляции равен 0,9765 и этот уровень тесноты связи является сверхсильным (допустимый коэффициент корреляции более 0,95) факторным связям.
Ширина листа изменяется (фиг. 8) по аналогичной по конструкции двухчленной формуле
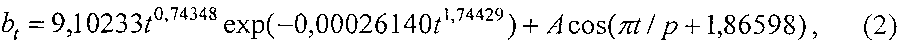
A=1,00341·10-22t13,54328exp(-0,0025789t1,72624),
p=192,24495-29,76665t0,30703.
Хотя конструкция формулы одинакова, но характер поведения ширины у популяции 10 учетных листьев иной.
Коэффициент корреляции 0,9400 меньше, чем у формулы динамики длины листа, и относит формулу (2) к сильнейшим (более 0,9) связям. При этом коренным образом иной полупериод колебания. При этом начало колебания ширины листа намного раньше срока распускания почек (по-видимому, рост ширины листа заложен генетически намного раньше даже начала сокодвижения). На дату 02.05.2014 период колебания равен 2×192,24495≈384 суток. Это больше годичного периода обращения Земли вокруг самого себя в 365 суток.
Здесь можно выдвинуть две гипотезы:
1) в древности более 100 млн. лет назад, липа была с круглогодичной листвой, а затем изменение глобального климата на планете вынудило это древесное растение перейти на годичные циклы распускания почек, роста листьев и их опадания;
2) на генетическом уровне липа заложила период вегетационного цикла в 384 суток для того, чтобы надежно превысить любую периодичность вращения Земли вокруг самого себя (тем самым липа переживет дальнейшее торможение вращения Земли вокруг Солнца еще много миллионов лет вперед на будущее).
Отрицательный знак в формуле полупериода показывает, что фенотипической волновой адаптацией листья липы быстро убавляют значение периода колебания. Это апостериорный факт показывает, что листья липы о ширине как бы быстро «приземляются» и, по-видимому, сброс листвы связан именно с динамикой ширины листьев липы.
Для длины и ширины листа рекомендуется обобщенная формула
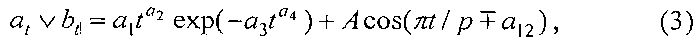
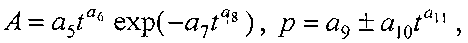
где at, bt - переменные во времени среднестатистические длина или ширина у популяции из не менее 10 учетных листьев, мм,
 - знак логических отношений «или»,
- знак логических отношений «или»,
t - время вегетации с начала распускания листьев, сутки,
A - амплитуда (половина) колебательного возмущения длины или ширины листьев в динамике роста популяции из не менее 10 листьев, мм,
p - полупериод колебания длины или ширины у не менее 10 учетных листьев, сутки,
a1…a12 - параметры модели (3), получающие числовые значения после статистического Моделирования идентификацией модели (3) по данным измерений длины или ширины у не менее 10 учетных листьев, при этом происходит чередование знаков:
+a10, -a12 принимают для закономерности динамики среднестатистической длины at у популяции из не менее 10 учетных листьев;
-a10, +a12 принимают для закономерности динамики среднестатистической ширины bt у популяции из не менее 10 учетных листьев.
Периметр листа Pt изменяется (фиг. 9) по формуле
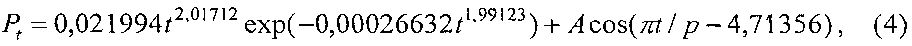
A=6,82928exp(0,00028005t1,76364), p=8,57053+0,49522t0,81824.
Адекватность формулы (4) по коэффициенту корреляции равен 0,9585 и этот уровень тесноты связи является сверхсильным (допустимый коэффициент корреляции более 0,95) факторным связям.
Начало волнения периметра происходит раньше 02.05.2014 (знак минус перед сдвигом волны). При этом амплитуда колебания изменяется со временем по закону экспоненциального роста, начиная от даты распускания почек при 2×6,82928≈13,7 см. Затем амплитуда колебания возрастает и такое возмущение, по-видимому, приводит к превышению допустимой амплитуды колебательного возмущения периметра и затем к отторжению листа от веточки.
В момент распускания почек период колебания равен 2×8,57053≈17,2 суток. Затем колебание успокаивается. Таким образом, для периметра характерно рост амплитуды и рост периода колебания.
Площадь листа St изменяется (фиг. 10) по формуле
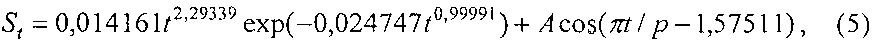
A=4,46628exp(0,00011616t1,88117), p=-9,37122+32,12674t0,10813.
Адекватность формулы (5) по коэффициенту корреляции равен 0,9838 и этот уровень тесноты связи является наибольшим из четырех параметров ученых листьев и при этом является сверхсильным (допустимый коэффициент корреляции более 0,95) факторным связям.
Амплитуда колебания на 02.05.2014 равна 2×4,46628≈8,93 см2, и это намного больше поперечного сечения самой почки. С дальнейшим ростом учтенные листья резко наращивают амплитуду колебания. По-видимому, наступает момент превышения амплитудой допустимого предела. При этом периметр и площадь действуют одинаковым образом.
Конструкция формулы (5) полностью аналогична закономерности (4). Однако начальное значение полупериода отрицательное и эту особенность биотехнической закономерности мы пока не можем объяснить. Но уже при t=1 сутки полупериод становится положительным и равным 22,8 суткам. Таким образом, в момент распускания почек учетный лист получает какие-то аномальные явления и процессы.
Для периметра и площади листа рекомендуется обобщенная формула
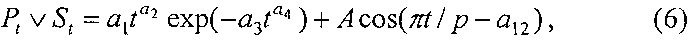
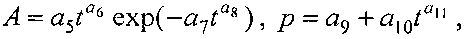
где Pt, St - переменные во времени среднестатистические периметр (см) или площадь (см2) у популяции из не менее 10 учетных листьев,
 - знак логических отношений «или»,
- знак логических отношений «или»,
t - время вегетации с начала распускания листьев, сутки,
A - амплитуда (половина) колебательного возмущения периметра (см) или площади (см2) в динамике среднестатистического роста популяции из не менее 10 учетных листьев,
p - полупериод колебательного возмущения при адаптации к внешним воздействиям периметра или площади учетных листьев, сутки,
a1…a12 - параметры модели (6), получающие числовые значения после статистического моделирования идентификацией модели (6) по данным измерений периметра или площади у не менее 10 учетных листьев.
По отношению к динамике параметров каждого из 10 листьев липы общая формула (3) оказалась неустойчивой, а обобщенная формула (6) для описания динамики периметра (табл. 5, фиг. 11) и площади (табл. 6, рис. 12) оказалась высоко устойчивой.
Запишем оба члена у каждой модели (4) и (5) в виде вейвлет-сигнала, исходящего от каждого учетного листа о своем поведении.
Волновое уравнение является асимметричным вейвлетом вида
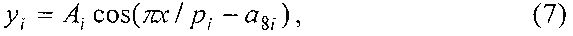
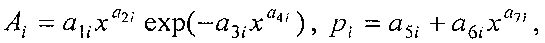
где Ai - амплитуда (половина) вейвлета, pi - полупериод колебания.
По формуле (7) c двумя фундаментальными физическими постоянными e (число Непера или число времени) и π (число Архимеда или число пространства) образуется изнутри изучаемого явления и/или процесса квантованный вейвлет-сигнал. Понятие вейвлет-сигнала позволяет абстрагироваться от физического смысла самих рядов (в общем случае не только динамических) и рассматривать их аддитивное разложение.
Для периметра у 10 учетных листьев параметры вейвлет-сигналов даны в таблице 5, а соответствующие графики приведены на фигуре 11. Во всех моделях коэффициент корреляции, из-за превышения количества параметров модели 13 над количеством наблюдений 11, равен 1,0000.
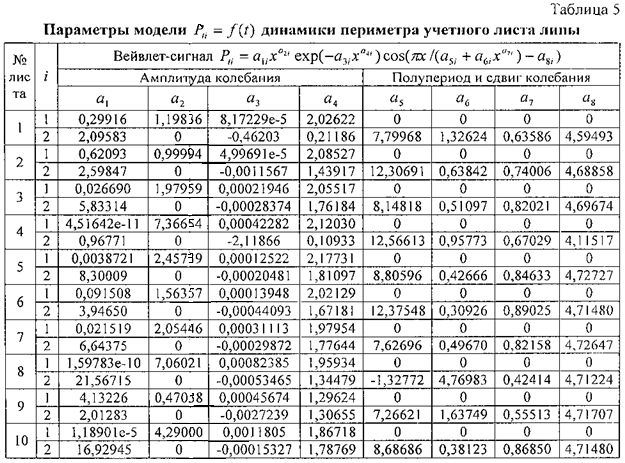
Все восемь параметров, кроме сдвига волны a8, имеют значительный интервал изменения своих значений. Этот факт указывает на высокую чувствительность предлагаемого способа и на практическую возможность изучения влияния разных факторов на динамику физических параметров у одного учетного листа липы и других деревьев.
Для площади у популяции из 10 учетных листьев параметры вейвлет-сигналов у каждого учетного листа даны в таблице 6, а соответствующие графики приведены на фигуре 12. Во всех моделях коэффициент корреляции, из-за превышения количества параметров модели 13 над количеством наблюдений 11, равен 1,0000.
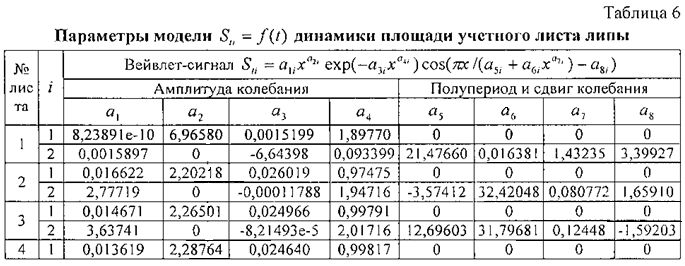
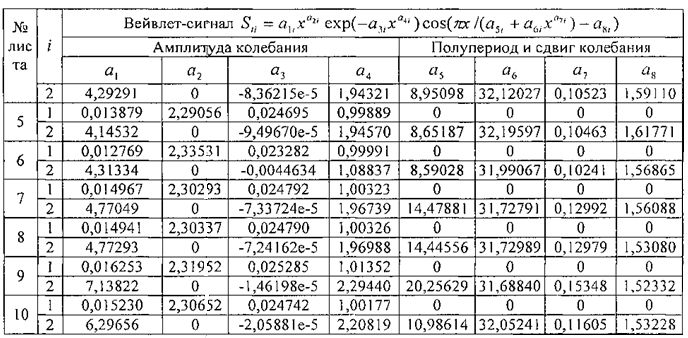
Из 10 учетных листьев только у листа №2 имеется отрицательный полупериод в начале распускания почек. Интервал изменения полупериода a5 находится в пределах от -3,57412 до 21,47660 суток.
По площади каждого учетного листа также можно выявлять закономерности влияния разных внешних и даже внутренних (например, физиологических) факторов. Поэтому предлагаемый способ имеет значимую перспективу методологического совершенствования.
Преимуществом предлагаемого способа является техническая простота исполнения, так как из оборудования требуется только изготовить даже самому прозрачную палетку, например из листового оргстекла, а сотовый телефон с функцией фотографирования ныне очень распространен и имеется почти у каждого студента или школьника. Кроме того, преимуществом является идентификация динамических рядов измерений среднестатистической динамики роста не менее 10 учетных листьев двумя обобщенными функциями, содержащими, по крайней мере, две составляющие. Первая составляющая является трендом в виде биотехнического закона, а вторая является положительно направленным на рост листьев волновым уравнением. При этом у второго члена, показывающего среднестатистическое колебательное возмущение учетных листьев, половина амплитуды для длины и ширины учтенных листьев является биотехническим законом, а для периметра и площади учетных листьев половина амплитуды изменяется по закону экспоненциального роста.
Изобретение может быть широко реализовано в школьных экологических кружках, пришкольных лесничествах, и даже в детских садах, а также в любых географических и иных экспедициях. Для использования предлагаемого способа нужно изготовить прозрачную палетку с сеткой клетками размером 2×2 мм, иметь сотовый телефон с функциями фотографирования, хранения фотографий в памяти и передачи их в персональный компьютер. В компьютере должна быть программная среда типа CurveExpert для выявления устойчивых закономерностей.
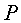
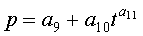
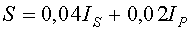
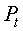
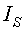

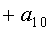
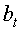
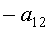
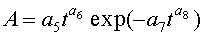
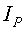
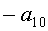
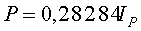
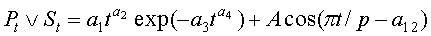
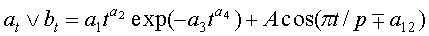
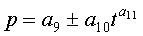
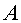
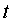
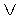
Способ отбора проб для анализа почвы луга
Способ измерения и анализа динамики объемного расхода водотока родника
Способ анализа кроны и места произрастания березы на склоне оврага
Способ динамического уравновешивания манипуляторной лесной машины и устройство для его осуществления
Способ ультразвукового испытания технической древесины
Способ анализа относительного сбега комбля березы на склоне оврага
Способ анализа комля и места произрастания березы на склоне оврага
Способ анализа формы комля дерева
Способ и устройство для изготовления образцов древесины
Способ испытания травяного покрова лесной прибрежной луговины
Способ сравнительной индикации загрязненности воздуха по флуктуирующей асимметрии листьев березы
Способ анализа видового состава луговой травы
Способ взятия пробы для агрохимического анализа по высоте ее расположения над урезом воды малой реки
Способ индикации загрязненности воздуха по флуктуирующей асимметрии листьев березы
Способ оценки устойчивости травяного покрова
Способ измерения высоты листа дерева от почвы
Способ анализа динамики роста в онтогенезе загрязненных листьев березы около автомобильной дороги
Способ оценки видового разнообразия травы по наличию отдельных видов растений на пробных площадках разных размеров
Способ факторного анализа онтогенеза учетных листьев дерева
Способ моделирования гипогонадизма, вызванного метаболическими нарушениями

