Результат интеллектуальной деятельности: ТРАНСГЕННОЕ РАСТЕНИЕ БЕРЕЗЫ С ПОВЫШЕННОЙ ПРОДУКТИВНОСТЬЮ
Вид РИД
Изобретение
Область техники, к которой относится изобретение
Настоящее изобретение относится к области генной инженерии растений и связано с получением трансгенных растений березы, обладающих повышенной продуктивностью. Результаты данного изобретения могут быть использованы в лесном хозяйстве, ландшафтном озеленении, строительстве, целлюлозно-бумажной промышленности, мебельном производстве и многих других отраслях народного хозяйства, использующих березу для выращивания или в качестве сырья.
Уровень техники
Береза очень широко распространена в России, занимая до 2/3 площади всех лиственных лесов страны. Древесина березы находит самое разнообразное применение (производство фанеры, стройматериалов, мебели, паркета, высококачественной целлюлозы, бочек для пищевых продуктов и др.). Древесина карельской березы из-за оригинального рисунка очень высоко ценится и используется как декоративный материал. Известно также большое число декоративных форм березы, отличающихся окраской листьев и коры, формой листьев и кроны, которые широко используются в озеленении. Однако береза не отличается высокой скоростью роста, и повышение продуктивности ее насаждений, особенно карельской березы, отличающейся медленным ростом, может иметь очень большое значение. Наиболее перспективным способом решения этой проблем является применение методов биотехнологии, а именно генной инженерии, позволяющих путем встраивания соответствующих генов повысить продуктивность этой ценной породы деревьев.
Для повышения скорости роста древесных растений использовали ряд генно-инженерных подходов, направленных на модификацию различных сфер метаболизма растений. Положительный эффект был достигнут при суперэкспрессии гена ксилоглюканазы из Aspergillus aculeatus в растениях тополя белого (Park et al., 2004, FEBS Lett., 564:183-187), гена пероксидазы хрена prxC1a в растениях осины (Kawaoka et al., 2003, Plant Physiol., 132:1177-1185), гена гемоглобина vhb из Vitreoscilla spp. в растениях тополя белого (Zelasco et al., 2006, Mol. Breed., 17:201-216), а также при суперэкспрессии ключевых генов биосинтеза гиббереллинов в растениях гибридной осины (Eriksson et al., 2000, Nat. Biotechnol., 18:784-788). Однако наиболее детальное освещение в научной литературе получил способ повышения скорости роста путем модификации метаболизма азота за счет суперэкспрессии гена глутаминсинтетазы (GS). Связано это с тем, что доступность неорганического азота в почве зачастую является лимитирующим фактором в росте и развитии растения. Фермент глутаминсинтетаза играет центральную роль в метаболизме азота у растений, катализируя превращение аммония в глутамин, который является аминокислотным предшественником глутамата и всех азотсодержащих компонентов, необходимых для роста растения. Растительная GS - это голофермент, состоящий из 8 идентичных полипептидных субъединиц и включающий два основных изофермента, GS1 и GS2, которые локализуются в различных внутриклеточных компартментах, цитозоле и хлоропластах соответственно (Kirby et al., 2006, Silvae Genet., 55:278-284). Генетические и молекулярные исследования показали, что GS может играть ключевую роль в эффективном использовании азота растениями (Hirel et al., 1992, Plant Mol. Biol., 20:207-218; Obara et al., 2001, J. Exp.Bot., 52:1209-1217). Продукты GS - глутамат и глутамин - являются предшественниками всех азотных соединений растения (Ireland and Lea, 1999, in: Plant Amino Acids: Biochemistry and Biotechnology, pp. 49-109, edited by В.K. Singh, Marcel Dekker Inc., New York). Поэтому генетическая модификация процессов, катализируемых ферментами GS1 и GS2, может быть очень результативным подходом повышения продуктивности лесных древесных растений, особенно в случаях их выращивания на бедных по азоту почвах. В работе Gallardo et al. (1999, Planta, 210:19-26) было продемонстрировано положительное влияние встраивания гена глутамин синтетазы на скорость роста трансгенных растений гибрида осины и тополя (Populus tremula × P. alba) в теплице. Трехлетние полевые испытания этих растений также подтвердили их ускоренный рост (Jing et. al., 2004, New Phytol., 164:137-145). Таким образом, встраивание гена глутаминсинтетазы в растения является перспективным способом повышения их продуктивности.
Все известные к настоящему времени работы по ускорению роста древесных растений с помощью генетической трансформации проводились на растениях из рода Populus: тополе, осине и их гибридах. Насколько нам известно, каких-либо исследований, направленных на повышение продуктивности растений березы методами генной инженерии, в том числе и с использованием гена глутаминсинтетазы, ранее не проводилось.
Раскрытие изобретения.
Задачей настоящего изобретения было получение трансгенной березы, отличающейся повышенной продуктивностью на основе элитного генотипа B. pubescens бп3ф1.
Используемый в настоящем описании термин «растение» охватывает целые растения, предшественники и потомство растений и части растений, включая семена, побеги, стебли, листья, корни (включая клубни), цветки и ткани и органы, причем все они содержат нуклеотидную последовательность с SEQ ID NO: 1. Термин «растение» также охватывает клетки растений, суспензионные культуры, каллусную ткань, зародыши, меристемы, гаметофиты, спорофиты, пыльцу и микроспоры, причем все они, опять же, содержат нуклеотидную последовательность с SEQ ID NO: 1.
Учитывая то, что большинство древесных растений, в том числе и береза, являются труднотрансформируемыми и труднорегенерируемыми, а также то, что даже при условии осуществления акта трансформации вероятность регенерации из трансформированной клетки экспланта целого трансгенного растения, в котором уровень экспрессии встроенного гена достаточен для проявления соответствующего фенотипического признака, а экспрессия эндогенных генов растения не нарушена, весьма мала, получение положительного результата при решении поставленной задачи не являлось очевидным фактом.
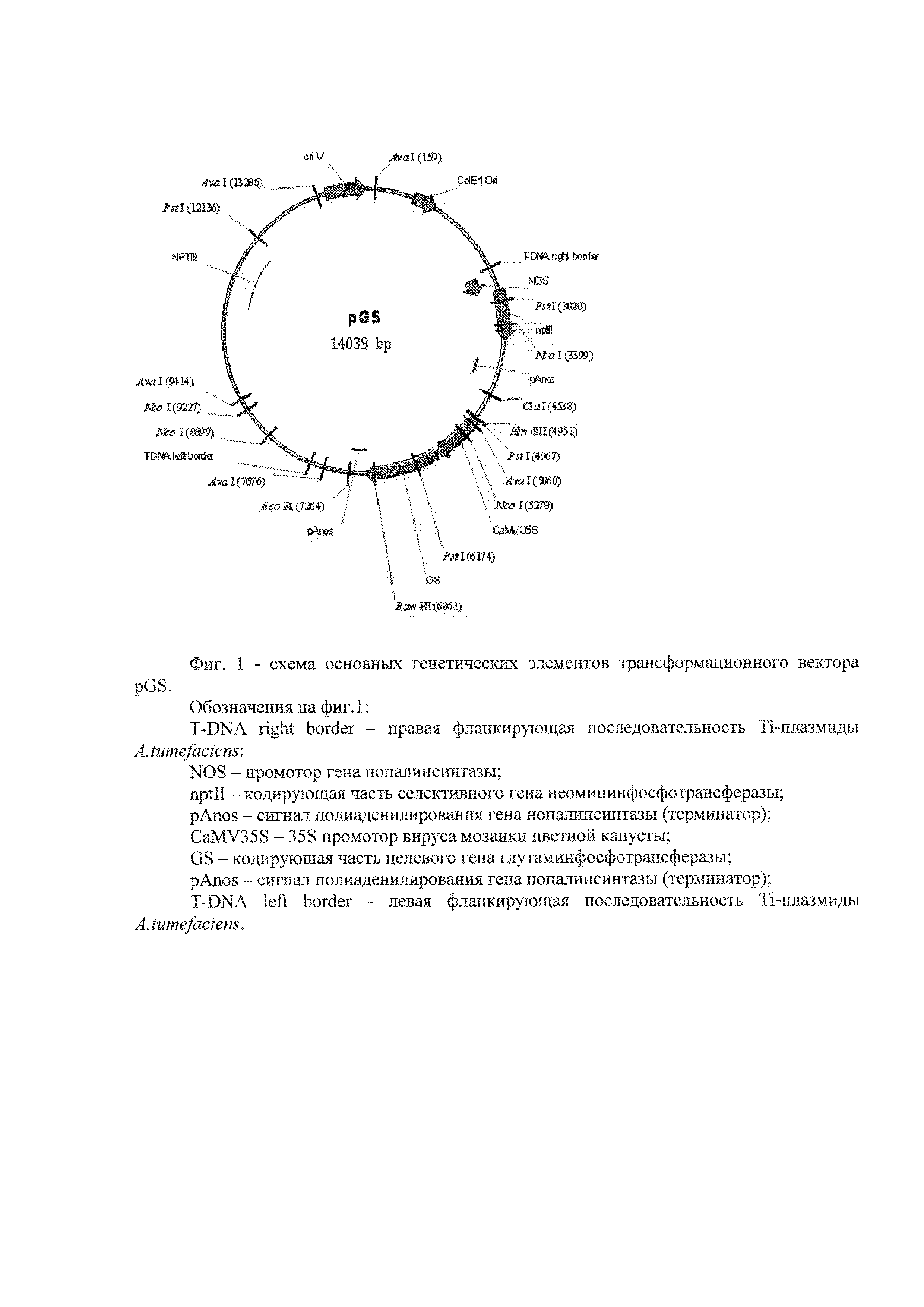
В качестве трансформирующего агента при получении трансгенной березы по изобретению использовали бинарный вектор pGS, включающий нуклеотидную последовательность глутаминсинтетазы. Структура вектора представлена на Фиг. 1.
В первом аспекте настоящее изобретение относится к растению березы с повышенной продуктивностью, где указанное растение березы трасформировано молекулой нуклеиновой кислоты, которая кодирует глутаминсинтетазу.
Молекула нуклеиновой кислоты может представлять собой кДНК, РНК или их гибридную молекулу. Предпочтительно молекулой нуклеиновой кислоты является молекула кДНК, кодирующая глутаминсинтетазу. Наиболее предпочтительно молекула кДНК имеет нуклеотидную последовательность, представленную в SEQ ID NO: 1.
Молекулу нуклеиновой кислоты глутаминсинтетазы можно выделить из любого вида растений. Предпочтительно растение представляет собой Pinus sylvestris.
Растение березы, трансформированное молекулой нуклеиновой кислоты глутаминсинтетазы, может представлять любой вид березы из рода Betula. Предпочтительно растение березы выбрано из группы, состоящей из B. pubescens, B. pendula, B. pendula var. carelica, В. nigra и B. schmidtii.
Во втором аспекте настоящее изобретение относится к трансгенному растению березы, содержащему молекулу нуклеиновой кислоты, кодирующей глутаминсинтетазу, где экспрессия данной молекулы приводит к получению трансгенного растения, которое демонстрирует следующие признаки повышения продуктивности: увеличенный выход биомассы, листья большего размера, увеличенная активность глутаминсинтетазы, повышенная эффективность использования азота по сравнению с аналогом дикого типа.
В третьем аспекте настоящее изобретение относится к способу получения трансгенного растения березы с повышенной продуктивностью, где способ включает стадии:
а) агробактериальной трансформации эксплантов березы in vitro молекулой нуклеиновой кислоты, кодирующей глутаминсинтетазу;
б) регенерации целых растений из трансформированных клеток;
в) идентификации молекулы нуклеиновой кислоты, кодирующей глутаминсинтетазу, в регенерированных растениях;
в) отбора растений березы с повышенной продуктивностью по сравнению с растениями того же генотипа, не содержащими молекулу нуклеиновой кислоты, кодирующей глутаминсинтетазу.
В еще одном варианте осуществления способ дополнительно включает стадию трансформации растения березы нуклеотидной последовательностью, кодирующей селективный или репортерный ген, который функционально связан с молекулой нуклеиновой кислоты, кодирующей глутаминсинтетазу, посредством чего облегчается отбор трансгенного растения березы среди регенерированных растений.
Краткое описание чертежей
Фиг. 1 - схема основных генетических элементов трансформационного вектора pGS.
Фиг. 2 - результат проведения рестрикционной проверки вектора pGS рестриктазами XbaI и SacI.
Фиг. 3 - результат проведения рестрикционной проверки вектора pGS рестриктазой BglII.
Фиг. 4 - результат регенерации побегов березы после трансформации вектором pGS.
Фиг. 5 - результат проведения ПЦР на фрагмент гена глутаминсинтетазы на ДНК трансформированных линий березы.
Осуществление изобретения
Пример 1. Ген глутаминсинтетазы
Целью выделения являлась нуклеотидная последовательность мРНК глутаминсинтетазы из P. sylvestris (GenBank Х69822), представленная в SEQ ID NO: 1. Тотальную РНК выделяли из 7-10-дневных проростков сосны по методике Gehrig et. al. (2000, Plant Mol. Biol. Rep., 18:369-376) с использованием готовых наборов экстракции фирмы BioRad (США). Электрофорез выделенной РНК проводили в специальном буфере (10× MOPS) в электрофорезной камере фирмы Amersham (США). Препарат РНК перед нанесением денатурировали путем приготовления следующей смеси: 2 мкл препарата тотальной РНК, 0,9 мкл 10× MOPS, 1,7 мкл формалина (37% раствор формальдегида в воде, рН не выше 4), 4,5 мкл формамида. После этого смесь инкубировали при 65°С 15 мин и наносили в лунки агарозного геля в сочетании со стандартным буфером. Оценку количества полученного препарата проводили стандартным спектрофометрическим методом. Для клонирования одного гена необходимо экстрагировать не менее 10 мкг тотальной РНК.
Для получения кДНК копии гена глутаминсинтетазы методом реакции обратной транскрипции использовали фермент обратную транскриптазу фирмы Fermentas (США), олиго-dT (Синтол, Россия) и комплект буферов и нуклеотидов Fermentas. Для синтеза кДНК гена глутаминсинтетазы брали 2-5 мкг РНК, добавляли 1-2 мкМ олигосdT и доводили объем до 10 мкл водой, обработанной ДЭПК. После этого смесь инкубировали при 70°С в течение 10 минут с последующим переносом на лед. По окончанию инкубации добавляли ревертазную смесь общим объемом 20 мкл, содержащую 10% DTT, 5 единиц ревертазы, буфер для ревертазы, 0,5 мМ нуклеотидов и 2-5 единиц ингибитора РНК-азы. Смесь инкубировали в течение часа при 42°С, после чего прогревали при 70°С 15 минут и далее разбавляли реакционную смесь бидистиллированной водой до 100 мкл.
Для проведения амплификации целевого фрагмента с кДНК копии использовали термоциклер (амплификатор) типа Mastercycler фирмы Eppendorf (Германия), полимеразу Pfu с соответствующим буфером и набор нуклеотидов от фирмы Fermentas. Условия ПЦР на ген глутаминсинтетазы были следующие: 5 минут при 93°С (горячий старт), 45 секунд при 93°С (денатурация), 30 секунд при 63°С (отжиг), 30 секунд при 72°С (элонгация), циклов амплификации 30. Использовали праймеры GS1-UTR-1 (SEQ ID NO: 2) и GS1-UTR-2 (SEQ ID NO: 3). Ожидаемый размер фрагмента 1443 п.о. Электрофорез сплайсированных фрагментов ДНК проводили в 1% агарозном геле в электрофорезной камере фирмы Amersham (США).
Пример 2. Клонирование гена глутаминсинтетазы в вектор pGEM-T Easy
5′- и 3′-концы фрагмента ДНК из примера 1, кодирующего ген глутаминсинтетазы с сайтами рестрикции XbaI и SacI, подвергали обработке соответствующими рестриктазами для образования липких кончиков. Далее для очистки полученных препаратов от примесей ферментов и нежелательных фрагментов ДНК образцы разделяли путем электрофореза в агарозе. Для выделения фрагментов ДНК из агарозного геля использовали фирменный кит MBI Fermentas (#К0513).
В качестве промежуточного вектора использовали вектор pGEM-T Easy (Promega), который также обрабатывали рестриктазами XbaI и SacI. Перед лигированием для удаления фосфата на 5′ конце вектор обрабатывали щелочной фосфатазой из кишечника теленка MBI Fermentas. Реакцию лигирования проводили Т4 ДНК лигазой MBI Fermentas в течение 12 часов при 12°С. Фермент инактивировали 10 мин при 65°С, а затем лигазной смесью трансформировали E. coli.
Компетентные клетки E. coli трансформировали лигазной смесью кальциевым методом и высевали на агар LB, содержащий канамицин для отбора клеток E. coli, трансформированных вектором. Количество колоний-трансформантов оценивали после 16-18 часов инкубации при 37°С. Трансформированные колонии E. coli тестировали на присутствие гена глутаминсинтетазы в векторе амплификацией ПЦР фрагмента гена глутаминсинтетазы после предварительной обработки рестриктазами SacI и XbaI. У целевых клонов после рестрикции с данными рестриктазами на агарозном гельэлектрофорезе видна вставка соответствующего размера.
Пример 3. Конструирование вектора pGS, содержащего ген глутаминсинтетазы
Отобранные в примере 2 колонии использовали для инокуляции среды LB и получения ночной культуры бактерий, из которой далее выделяли плазмидную ДНК щелочным методом (Маниатис и др., Методы генетической инженерии. Молекулярное клонирование, 1984). Секвенирование клонов со вставками целевых фрагментов проводили в фирме ЗАО «Евроген» (Россия).
В качестве вектора использовали бинарный вектор рВI121 размером 14758 п.о., из которого по сайтам XbaI и SacI путем обработки соответствующими рестриктазами удаляли фрагмент длиной 1900 п. о. (ген β-глюкуронидазы). Базовую часть вектора длиной 12858 п.о. (фрагмент pBI-XbaI-SacI) использовали для дальнейшего клонирования. Вставку гена глутаминсинтетазы получали путем обработки плазмидной ДНК рестриктазами XbaI и SacI. 5′- и 3′- концы фрагмента ДНК, кодирующего глутаминсинтетазу, обрабатывали щелочной фосфатазой. Лигирование фрагментов рВI-XbaI-SacI и ДНК, кодирующей глутаминсинтетазу, проводили при температуре 4°С в присутствии Т4 ДНК лигазы (Fermentas) с образованием вектора pGS. Перенос вектора в клетки E. coli, выделение плазмидной ДНК и проверку вставки гена глутаминсинтетазы выполняли, как описано в примере 2.
Рестрикционную проверку полученного вектора pGS проводили путем обработки рестриктазами XbaI и SacI (должны быть получены полосы 12,7 т.п.о. и 1,3 т.п.о) и BglII (должны быть получены полосы 8,2 т.п.о., 2,5 т.п.о. и 3,6 т.п.о.). Результаты проверки показаны на Фиг. 2 и Фиг. 3, соответственно.
Перенос бинарного вектора pGS в агробактериальный штамм СВЕ21 проводили по стандартной методике. Селекцию колоний осуществляли на среде, содержащей 50 мг/л канамицина.
Пример 4. Подготовка штамма бактерий A. tumefaciens СВЕ21 (рСВЕ21, pGS) для трансформации растений
Для трансформации растений используют ночную культуру бактерий A. tumefaciens. Для этого 100 мкл суспензии клеток бактерий A. tumefaciens СВЕ21 (рСВЕ21, pGS) добавляют к 50 мл жидкой среды LB, содержащей 50 мг/л канамицина и инкубируют в течение ночи на термостатируемом орбитальном шейкере при 28°С и 120-150 об/мин, после чего центрифугируют полученную суспензию 5 минут при 4000 об/мин, осадок промывают жидкой средой MS и повторяют центрифугирование и промывание. После осаждения клеток их заливают 50 мл жидкой среды MS и ресуспендируют.
Пример 5. Подготовка растительного материала березы in vitro для трансформации.
Для трансформации растений березы используют листовые экспланты с растений in vitro. Размножение культуры березы проводят на питательной среде WPM, содержащей 0,6 мг/л БАП, 0,1 мг/л ИМК, 20 г/л сахарозы и 7 г/л агара. Растения выращивают при фотопериоде 16/8 часов, температуре 22-24°С и освещенности 3000-3500 люкс.
Пример 6. Трансформация растений березы клетками бактерий A. tumefaciens СВЕ21 (pCBE21, pGS).
Для трансформации используют листья с растений in vitro возрастом 1 месяц. У листьев удаляют черешки и верхушки (у крупных листьев - также и боковые стороны) и наносят несколько надрезов перпендикулярно центральной жилке, не доводя их до краев листа. Подготовленные таким образом экспланты помещают на 40-50 минут в суспензию агробактерий, после чего осушают стерильными фильтрами и размещают на фильтрах, расположенных в чашках Петри на поверхности среды для кокультивации, содержащей минеральные соли MS, 5 мг/л зеатина, 5 мг/л БАП, 0,2 мг/л ИМК, 30 г/л сахарозы и 7 г/л агара. В каждую чашку помещают по 10-15 эксплантов. Кокультивацию проводят в течение 3 суток.
Пример 7. Регенерация растений березы из трансформированных эксплантов.
После периода кокультивации экспланты промывают в дистиллированной воде с добавлением 1 г/л цефотаксима в течение 20-30 минут и затем дважды в воде без цефотаксима. Отмытые экспланты подсушивают на фильтрах и переносят на среду для регенерации и селекции трансформантов того же состава, как и среда для кокультивации, содержащую дополнительно 50 мг/л канамицина и 500 мг/л цефотаксима. На этой среде экспланты выдерживают в условиях 16-часового светового дня при 22-23°С с пересадкой каждые 4 недели. Результаты регенерации показаны на Фиг. 4. Регенерированные побеги пересаживают на среду для размножения, содержащую 50 мг/л канамицина и 250 мг/л цефотаксима.
Пример 8. Идентификация фрагмента последовательности гена глутамин синтетазы в регенерантах березы методом ПЦР
Присутствие гена GS в трансгенных растениях березы подтверждают методом ПЦР с праймерами GS1 (SEQ ID NO: 4) и GS2 (SEQ ID NO: 5), специфичными для кодирующей области трансгенной конструкции.
Геномную ДНК из растений березы выделяют по методу Rogers and Bendich (1994, in: Gelvin SB, Schilperoort RA (eds) Plant Molecular Biology Manual. Boston, MA: Kluwer Academic Publishers, D1: 1-8). Для выделения используют листья растений in vitro (около 100 мг). Полученную растительную ДНК используют в качестве матрицы в ПЦР-анализах. Реакционная смесь содержит 67 мМ Tris-HCI, рН 9.0, 16 мМ (NH4)2SO4, 2 мМ MgCI2, 0,01% желатина, по 0,2 мМ каждого dNTP, 0,6 мкМ конечной концентрации каждого праймера и 0,2 единицы/мкл Taq полимеразы. Реакцию проводят в объеме 25 мкл при следующих условиях: 92°С - 3 мин; 35 циклов: 92°С - 20 сек, 62°С - 10 сек, 72°С - 1 мин, затем 72°С - 5 мин.
Продукты ПЦР анализируют в 1,8% агарозном геле с добавлением бромистого этидия. Гель фотографируют в ультрафиолете при длине волны 260-280 нм. Появление продукта ПЦР (ДНК размером 1190 н.п.) при использовании указанных праймеров, а также при условии отсутствия его в реакциях, поставленных на контрольной ДНК, свидетельствует о присутствии искомого гена в ДНК исследуемых растений (Фиг. 5).
Пример 9. Отбор трансгенных растений березы с повышенной продуктивностью
Укорененные в условиях in vitro растения березы (линии, по итогам ПЦР содержащие ген глутаминсинтетазы и нетрансформированный контроль) высаживали в теплицу в пластиковые кассеты. В качестве субстрата использовали пропаренную смесь торфа и песка (3:1), обогащенную 20 г комплексного минерального удобрения "Кристаллин" на 1 кг смеси. На период адаптации растений накрывали полиэтиленовой пленкой, которую снимали через один месяц. Оценку продуктивности проводили путем измерения биомассы растения, площади листьев, активность глутаминсинтетазы, эффективности использования азота.
Биомассу растений определяли в конце первого сезона вегетации. Корневую систему отмывали от субстрата, растение высушивали при 105°С в течение 24 ч и взвешивали на весах с точностью до 0,1 г.
Площадь листьев определяли в период вегетации. Листья, срезанные из средней части побега, сканировали с разрешением 300 dpi и площадь определяли с помощью программы ImageJ.
Активность глутаминсинтетазы определяли в листовой ткани в период вегетации по методу Cai et. al. (2009, Plant Cell Rep, 28:527-537).
Эффективность использования азота определяли в конце первого сезона вегетации по методу Finzi et. al. (2007, Proc Natl Acad Sci USA, 104:14014-14019).
Результаты показаны в таблице 1.

В таблице 1 показано превышение различных показателей продуктивности растений березы с геном глутаминсинтетазы по сравнению с нетрансформированным контролем на 9-84%.
Перечень последовательностей
SEQ ID NO: 1 - нуклеотидная последовательность мРНК глутаминсинтетазы из P. sylvestris (GenBank Х69822):

SEQ ID NO: 2 - олигонуклеотидный праймер GS1-UTR-1:
5′-AGCTTCTAGATTCCCTTCCTCTGGTTTGTT-3′
SEQ ID NO: 3 - олигонуклеотидный праймер GS1-UTR-2:
5′-AGCTGAGCTCCACAAAGAATATATCATATT-3′
SEQ ID NO: 4 - олигонуклеотидный праймер GS1:
5′-TTATGAGCTCTTAGCTTATCAGTGACGCTGG-3′
SEQ ID NO: 5 - олигонуклеотидный праймер GS2:
5′-CTCATCTAGACTGGTTTGTTTTGGAGAGTGGC-3′.


Клеточная линия hufshik, секретирующая рекомбинантный человеческий фсг
Способ длительного хранения in vitro растений осины
Способ криоконсервации пазушных почек in vitro растений осины
Способ подготовки микропобегов in vitro ясеня, осины, ивы для последующего укоренения в условиях ex vitro
Трансгенное растение березы с устойчивостью к гербицидам
Способ получения генетически модифицированных древесных растений
Трансгенное растение осины с повышенной продуктивностью и модифицированной древесиной
Способ сохранения качественных характеристик культуры in vitro некоторых древесных видов растений (лимонник китайский, рододендрон, сирень, береза повислая)
Трансгенное растение осины с повышенным содержанием целлюлозы в древесине
Трансгенное растение осины с пониженной скоростью разложения древесины
Клеточная линия hufshik, секретирующая рекомбинантный человеческий фсг
Способ длительного хранения in vitro растений осины
Способ криоконсервации пазушных почек in vitro растений осины
Способ подготовки микропобегов in vitro ясеня, осины, ивы для последующего укоренения в условиях ex vitro
Трансгенное растение березы с устойчивостью к гербицидам
Способ получения генетически модифицированных древесных растений
Трансгенное растение осины с повышенной продуктивностью и модифицированной древесиной
Способ сохранения качественных характеристик культуры in vitro некоторых древесных видов растений (лимонник китайский, рододендрон, сирень, береза повислая)
Трансгенное растение осины с повышенным содержанием целлюлозы в древесине
Трансгенное растение осины с пониженной скоростью разложения древесины

