Результат интеллектуальной деятельности: КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ CRY1Ca И CRY1Fa ДЛЯ БОРЬБЫ С РЕЗИСТЕНТНОСТЬЮ У НАСЕКОМЫХ
Вид РИД
Изобретение
Предшествующий уровень техники
Человек выращивает кукурузу для употребления в пищу и для энергетических целей. Человек также выращивает множество других культур, включая сою и хлопок. Насекомые поедают и повреждают растения, и тем самым наносят ущерб деятельности человека. Для борьбы с насекомыми-вредителями ежегодно тратятся миллиарды долларов, и еще миллиарды уходят на возмещение ущерба, наносимого этими вредителями. Инсектициды, синтезированные методами органической химии, являются главным инструментом, используемым для борьбы с насекомыми-вредителями, но в некоторых регионах, в борьбе с насекомыми-вредителями важную роль играют биологические инсектициды, такие как инсектицидные белки, происходящие от Bacillus thuringiensis (Bt). Возможность культивировать резистентные к насекомым растения посредством трансформации этих растений генами инсектицидных белков Bt явилась революцией в современном сельском хозяйстве и доказала важность и ценность инсектицидных белков и их генов.
Некоторые белки Bt были использованы для создания резистентных к насекомым трансгенных растений, которые успешно прошли испытания, и в настоящее время их производят в промышленном масштабе. Такими белками являются Cry1Ab, Cry1Ac, Cry1F и Cry3Bb, вводимые в кукурузу, Cry1Ac и Cry2Ab, вводимые в хлопок, и Cry3A, вводимый в картофель.
Коммерчески доступные продукты, экспрессирующие эти белки, экспрессируют только один из этих белков, за исключением случаев, когда желательно получить комбинированный инсектицидный спектр из 2 белков (например, в кукурузе, Cry1Ab и Cry3Bb объединены для вырабатывания резистентности к чешуекрылым вредителям и корневым личинкам, соответственно), или случаев, когда независимое действие этих белков делает их ценным инструментом для замедления развития резистентности к инсектицидам у восприимчивых популяций насекомых (например, Cry1Ac и Cry2Ab в хлопчатнике объединяют в целях вырабатывания у растений резистентности к табачной листовертке).
То есть, некоторые сорта резистентных к насекомым трансгенных растений, которые быстро и повсеместно адаптируются к этой технологии, также имеют тот недостаток, что популяции вредителей вырабатывают резистентность к инсектицидным белкам, продуцируемым этими растениями. Было предложено несколько стратегий для сохранения ценных признаков резистентности к Bt-насекомым, где указанные стратегии включают использование в высоких доз белков в комбинации с сохранением площадей «убежищ» нетрансгенных растений, и чередования или совместного использования различных токсинов (McGaughey et al.(1998), «Bt-Resistance Management» Nature Biotechnol. 16: 144-146).
Необходимо, чтобы белки, отобранные для использования в IRM-кластерах, обладали независимым инсектицидным действием, при котором резистентность, вырабатываемая к одному белку, не распространялась на другой белок (то есть, не наблюдалась перекрестная резистентность к белкам). Так, например, если популяция насекомых, выбранных на резистентность к «белку А», является восприимчивой к «белку В», то можно сделать вывод, что в данном случае перекрестная резистентность отсутствует, и комбинация «белок А и белок В» будет эффективной для замедления вырабатывания резистентности к одному белку А.
В случае отсутствия популяции резистентных насекомых, оценка может быть сделана исходя из других характеристик, которые, как предполагается, относятся к механизму действия и возможной перекрестной резистентности. Было высказано предположение, что применение опосредуемого рецептором связывания при идентификации инсектицидных белков, очевидно, не будет приводить к вырабатыванию перекрестной резистентности. (van Mellaert et al. 1999). Ключевым прогностическим фактором отсутствия перекрестной резистентности, появляющейся при таком подходе, является то, что инсектицидные белки не конкурируют за связывание с рецепторами у восприимчивых видов насекомых.
В случае, когда два токсина Bt конкурируют за связывание с одним и тем же рецептором, и если этот рецептор мутирует у насекомого так, что один из токсинов больше не связывается с этим рецептором, а поэтому не обладает инсектицидным действием против насекомого, то такое насекомое может приобретать резистентность ко второму токсину (который конкурентно связывается с тем же рецептором). То есть, можно сказать, что такое насекомое будет обладать перекрестной резистентностью к обоим токсинам Bt. Однако если два токсина связываются с двумя различными рецепторами, то это означает, что такое насекомое не обладает одновременной резистентностью к этим двум токсинам.
Cry1Fa может быть использован для борьбы с чешуекрылыми вредителями многих видов, включая европейского кукурузного пилильщика (ECB; Ostrinia nubilalis (Hubner)) и совку травяную (FAW; Spodoptera frugiperda), и обладает активностью против свекловичного пилильщика (SCB; Diatraea saccharalis).
Белок Cry1Fa, продуцирующийся в растениях кукурузы, содержащих модификацию TC1507, ответственен за вырабатывание признака резистентности к насекомым, и этот белок применяется в ведущих областях промышленности для борьбы с совкой травяной (FAW). Cry1Fa также используется в продуктах Herculex® , SmartStax™ и WideStrike™.
Возможность проводить исследование на связывание с рецептором (конкурентное или гомологичное) с использованием белка Cry1Fa имеет определенные ограничения, поскольку при применении большинства методов мечения белков для детектирования в анализах на связывание с рецептором происходит инактивация инсектицидной активности белка Cry1Fa.
Дополнительные токсины Cry перечислены на web-сайте офиса Комитета по номенклатуре B.t. (Crickmore et al; lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/). См. Приложение А в данной заявке. В настоящее время известно примерно 60 основных групп токсинов «Cry» (Cry1-Cry59), и кроме того, существуют другие токсины, такие как токсины Cyt и токсины VIP и т.п. Многие токсины из каждой пронумерованной группы имеют подгруппы, обозначенные прописными буквами, а подгруппы, обозначенные прописными буквами, в свою очередь, подразделяются на подгруппы (суб-подгруппы), обозначенные строчными буквами (например, Cry1 имеет подгруппу A-L, а Cry1A имеет подгруппу a-i).
Краткое описание сущности изобретения
Настоящее изобретение частично основано на неожиданном обнаружении того факта, что популяция травяной совки (Spodoptera frugiperda; FAW), отобранная на резистентность к инсектицидной активности белка Cry1Fa, не является резистентной к инсектицидной активности белка Cry1Ca. Для специалиста в данной области очевидно, что преимущество такого открытия состоит в том, что растения, экспрессирующие эти два инсектицидных белка или их инсектицидные части, могут быть использованы для замедления или предупреждения развития резистентности к любому из этих отдельно взятых инсектицидных белков.
Настоящее изобретение также подтверждается обнаружением того факта, что Cry1Fa и Cry1Ca не конкурируют друг с другом за связывание с кишечными рецепторами совки травяной (FAW) (или Diatraea saccharalis (свекловичного пилильщика; SCB)).
Настоящее изобретение также относится, в частности, к трехкомпонентным кластерам или «пирамидам» из трех (или более) токсинов, где токсины Cry1Fa и Cry1C представляет собой базовую пару. Одна из предпочтительных пирамид включает по меньшей мере два белка, не обладающих перекрестной резистентностью к двум вредителям - к совке травяной и к ECB (Европейскому кукурузному пилильщику; Ostrinia nubilalis); Cry1Fa плюс Cry1Ca плюс один или несколько анти-ECB токсинов, таких как Cry1Ab. В некоторых предпочтительных вариантах пирамиды, выбранные токсины обладают тремя различными механизмами действия против FAW. Предпочтительными пирамидными комбинациями являются Cry1Fa плюс Cry1Ca плюс другой токсин/ген, выбранный из группы, состоящей из Vip3Ab, Cry1D, Cry1Be и Cry1E. Растения (и посевные площади, засеянные такими растениями), которые продуцируют эти три токсина, входят в объем настоящего изобретения. Могут быть также добавлены дополнительные токсины/гены, но эти конкретные трехкомпонентные кластеры, в соответствии с настоящим изобретением, будут, как неожиданно было обнаружено, преимущественно действовать против FAW по трем механизмам. Это поможет снизить или вообще избежать потребности в площадях-«убежищах» нетрансгенных культур. В общих чертах, настоящее изобретение также относится к применению трех инсектицидных белков (в некоторых предпочтительных вариантах, белков Cry), которые не конкурируют друг с другом против одного вредителя-мишени.
Краткое описание графического материала
Фигура 1: Оценка степени повреждения кукурузы, зараженных in vitro только что вылупившимися личинками совки травяной (FAW) и совки травяной, резистентной к Cry1F (rFAW). Трансгенные растения T0, отобранные после трансформации плазмидой pDAS5162, были разделены на две группы посредством иммуноблот-скрининга с использованием антитела DIG152RPC1: на растения, которые не продуцируют DIG-109 (на фигуре, с левой стороны), и на растения, которые не имеют детектируемые уровни DIG-109 (в центре фигуры). DIG-109-позитивные растения ранжировали по уровню экспрессии (слева направо; от самого меньшего к самому большему). HP-анализ на DSM2 осуществляли на 36 pDAS5162-трансформированных линиях. В 95% образцов было детектировано простое событие интеграции, определенное как 1-2 копии этого гена. Результаты биологического анализа растений, используемых в качестве позитивного и негативного контроля, представлены на правой стороне фигуры.
Фигура 2: Конкурирование за связывание BBMV Spodoptera frugiperda с коровым токсином Cry1Fa, с коровым токсином Cry1Ca и с 125I-меченным коровым токсином Cry1Ca.
Краткое описание последовательностей
SEQ ID NO:1 - Химерный белок «коровый Cry1Ca/протоксин Cry1Ab», состоящий из 1164 аминокислот (DIG- 152) (вариант pMYC2547)
SEQ ID NO:2 - Второй химерный белок ««коровый Cry1Ca /протоксин Cry1Ab», состоящий из 1164 аминокислот (DIG-109) (сорт кукурузы)
SEQ ID NO:3 - Оптимизированная для кукурузы последовательность CDS, кодирующая 3492 п.н. DIG-109
SEQ ID NO:4 - коровый токсин Cry1Fa.
SEQ ID NO:5 - коровый токсин Cry1Ca.
Подробное описание изобретения
Как сообщается в настоящей заявке, токсин Cry1Ca, продуцируемый в трансгенной кукурузе и в других растениях (например, в хлопчатнике и сое), обнаруживает высокую эффективность в борьбе против совки травяной (FAW; Spodoptera frugiperda), у которой вырабатывается резистентность к активности Cry1Fa. Таким образом, настоящее изобретение частично основано на неожиданном обнаружении того факта, что совка травяная, резистентная к действию Cry1Fa, является восприимчивой (то есть, не обладает перекрестной резистентностью) к действию Cry1Ca.
Настоящее изобретение также частично основано на неожиданном обнаружении того факта, что токсин Cry1Ca является эффективным для защиты растений (таких как растения кукурузы) от поражения Cry1Fa-резистентной совки травяной. Обсуждение этого вредителя приводится, например, в публикации Tabashnik, PNAS (2008), vol. 105 no. 49, 19029-19030.
Настоящее изобретение включает применение токсина Cry1Ca для защиты кукурузы и других экономически ценных видов растений от поражения вредителями и снижения урожайности, вызываемого поеданием этих растений совкой травяной или популяциями совки травяной, у которых вырабатывается резистентность к Cry1Fa.
Настоящее изобретение также относится к кластеру IRM, используемому для предупреждения или замедления развития резистентности совки травяной к Cry1Fa и/или Cry1Ca.
Настоящее изобретение также относится к композициям, используемым для борьбы с чешуекрылыми вредителями, в клетках которых продуцируется белок, содержащий коровый токсин Cry1Fa, и белок, содержащий коровый токсин Cry1Ca.
Настоящее изобретение также относится к хозяину, трансформированному так, чтобы он продуцировал белок, содержащий коровый токсин Cry1Fa, и белок, содержащий коровый токсин Cry1Ca, где указанным хозяином является клетка микроорганизма или растения. Рассматриваемый(ые) полинуклеотид(ы) предпочтительно присутствует(ют) в генетической конструкции под контролем промотора, не являющегося промотором Bacillus thuringiensis (функционально присоединен(ы) к этому промотору/содержит(ат) этот промотор). Рассматриваемые полинуклеотиды могут содержать обычно встречающиеся в этом растении кодоны, способствующие повышению уровня экспрессии в растении.
Настоящее изобретение также относится к способу борьбы с чешуекрылыми вредителями, включающему контактирование указанных вредителей или среды их обитания с эффективным количеством композиции, содержащей белок, включающий коровый токсин Cry1Fa, а также белок, включающий коровый токсин Cry1Ca.
В одном из своих вариантов, настоящее изобретение относится к растению кукурузы (и к другим растениям, например, к хлопчатнику и сое), включающему экспрессируемый в этом растении ген, который кодирует белок, содержащий коровый токсин Cry1Ca, и экспрессируемый в этом растении ген, который кодирует белок, содержащий коровый токсин Cry1Fa, и к семенам такого растения.
В другом своем варианте, настоящее изобретение относится к растению кукурузы (и к другим растениям, например, к хлопчатнику и сое), где экспрессируемый в этом растении ген, который кодирует белок, содержащий коровый токсин Cry1Ca, и экспрессируемый в этом растении ген, который кодирует белок, содержащий коровый токсин Cry1Fa, были введены в указанные растения кукурузы и в семена таких растений путем интрогрессии.
(Описание Cry1C, используемого в растениях в качестве потенциального биоинсектицида, приводится в публикации Avisar et al. 2009). Avisar D, Eilenberg H, Keller M, Reznik N, Segal M, Sneh B, Zilberstein A (2009) The Bacillus thuringiensis delta-endotoxin Cry1C as a potential bioinsecticide in plants. Plant Science 176:315-324).
Рецепторы насекомых. Как описано в примерах, исследование на конкурентное связывание с рецептором, проводимое с использованием радиоактивно меченного белка корового токсина Cry1Ca, показало, что этот белок корового токсина Cry1Fa не конкурирует за высокоаффинное связывание с сайтом, присутствующим в тканях насекомого FAW, с которым связывается Cry1Ca. Эти результаты показали, что комбинация белков Cry1Fa и Cry1Ca представляет собой эффективное средство для ослабления вырабатывания резистентности у популяции FAW к Cry1Fa (и аналогично, для ослабления вырабатывания резистентности к Cry1Ca), а также для повышения уровня резистентности растений кукурузы, экспрессирующих оба эти белка, к указанному вредителю.
Таким образом, частично на основе данных, описанных выше, и данных, представленных в литературе, можно сделать вывод, что совместное продуцирование (в виде кластера) белков Cry1Ca и Cry1Fa может быть применено для получения высокой дозы IRM-кластера, используемого для борьбы с FAW. В эту комбинацию могут быть добавлены и другие белки для расширения спектра уничтожаемых насекомых-вредителей. Так, например, для кукурузы, добавление Cry1Ab позволит создать IRM-пирамиду, используемую для борьбы с европейским кукурузным пилильщиком.
В другом своем варианте, настоящее изобретение относится к применению одного или двух белков Cry1Fa и Cry1Ca в комбинации с белком Cry1Ab для ослабления вырабатывания резистентности у насекомых. Таким образом, в другом своем варианте, настоящее изобретение относится к применению одного или двух белков Cry1Fa и Cry1Ca в сельскохозяйственных регионах, в которых применение белка Cry1Ab становится неэффективным для борьбы со свекловичным пилильщиком, что обусловлено развитием резистентности у таких популяций вредителей. В соответствии с этим, предпочтительным «трехкомпонентным кластером» или «пирамидной» комбинацией, применяемой в соответствии с настоящим изобретением, являются Cry1F плюс Cry1C плюс Cry1Ab.
Настоящее изобретение также относится, в частности, к трехкомпонентным кластерам или «пирамидам» из трех (или более) токсинов, где токсины Cry1Fa и Cry1C представляет собой базовую пару. Одна из предпочтительных пирамид включает по меньшей мере два белка, не обладающих перекрестной резистентностью к двум вредителям - к FAW и к ECB (Европейскому кукурузному пилильщику; Ostrinia nubilalis); Cry1Fa плюс Cry1Ca плюс один или несколько анти-ECB токсинов, таких как Cry1Ab (см. заявку США 20080311096), поскольку Cry1F обладает активностью против обоих насекомых. Другими токсинами против ECB являются Cry1Be (см. заявку США рег.№ 61/284290, поданную 16 декабря, 2009), Cry II (см. заявку США рег.№ 61/284278, поданную 16 декабря, 2009), Cry2Aa (см. заявку США рег.№ 61/284278, поданную 16 декабря 2009) и DIG-3 (см. заявку США 201000269223). В некоторых предпочтительных вариантах «пирамиды», выбранные токсины обладают тремя различными механизмами действия против FAW, и такими предпочтительными пирамидными комбинациями являются Cry1Fa плюс Cry1Ca плюс другой токсин/ген, выбранный из группы, состоящей из Vip3Ab, Cry1D (см. заявку США рег.№ 61/284252, поданную 16 декабря, 2009), Cry1Be и Cry1E (см. заявку США рег.№ 61/284278, поданную 16 декабря 2009). Растения (и посевные площади, засеянные такими растениями), которые продуцируют эти три токсина, входят в объем настоящего изобретения. Могут быть также добавлены дополнительные токсины/гены, и эти конкретные трехкомпонентные кластеры, в соответствии с настоящим изобретением, будут, как неожиданно было обнаружено, преимущественно действовать против FAW по трем механизмам. Это позволяет снизить или избежать потребности в площадях-«убежищах» нетрансгенных культур. Таким образом, в настоящем изобретении рассматривается посевная площадь свыше 10 акров.
Другие токсины, например, Vip3, перечислены в Приложении A. Эти номера GENBANK могут быть также использованы для идентификации последовательностей любых описанных и упомянутых здесь генов и белков.
В патенте США No. 5188960 и в патенте США No. 5827514 описаны белки, которые содержат коровый токсин Cry1Fa, и которые могут быть использованы для осуществления настоящего изобретения. В патенте США № 6218188 описаны оптимизированные для растения последовательности ДНК, кодирующие белки, которые содержат коровый токсин Cry1Fa, и которые могут быть использованы в настоящем изобретении.
Комбинации токсинов, описанных в настоящем изобретении, могут быть использованы для борьбы с чешуекрылыми вредителями. Взрослые чешуекрылые, например, бабочки и моли, питаются, главным образом, нектаром и играют значительную роль в опылении. Почти все личинки чешуекрылых, то есть, гусеницы, поедают растения, и многие из них являются опасными вредителями. Гусеницы живут на листьях или поедают внутреннюю часть листьев, либо они повреждают корни или стебли растения, что приводит к истощению питательных веществ у растения, и в большинстве случаев, к разрушению основной физической структуры растения. Кроме того, гусеницы повреждают плоды, ткани и хранящееся зерно и муку, в результате чего продукты либо вообще становятся непригодными для продажи, либо их коммерческая ценность значительно снижается. Используемый здесь термин «чешуекрылые вредители» также относится к различным стадиям жизненного цикла вредителя, включая стадии развития личинок.
Некоторые химерные токсины согласно изобретению содержат полноразмерную часть N-концевого корового токсина Bt, и в определенном положении, расположенном за пределами части корового токсина, этот белок переходит в гетерологичную последовательность протоксина. N-концевая, инсектицидно активная часть токсина Bt называется «коровым токсином». Переход от корового сегмента токсина в гетерологичный сегмент протоксина может происходить приблизительно в области стыка токсин/протоксин, или альтернативно, часть нативного протоксина (простирающаяся за пределы коровой части токсина) может сохраняться, причем, переход в гетерологичную часть протоксина может происходить ниже.
Одним из примеров может служить один химерный токсин согласно изобретению, который представляет собой полноразмерную часть корового токсина Cry1Fa (аминокислоты 1-601) и гетерологичный протоксин (аминокислоты от положения 602 до С-конца). В одном предпочтительном варианте изобретения, часть химерного токсина, содержащая протоксин, происходит от токсина белка Cry1Ab. Вторым примером может служить второй химерный токсин согласно изобретению, который описан в SEQ ID NO:l и представляет собой часть полноразмерного корового токсина Cry1Ca (аминокислоты 1-619) и гетерологичный протоксин (аминокислоты от положения 620 до С-конца). В предпочтительном варианте изобретения, часть химерного токсина, содержащая протоксин, происходит от токсина белка Cry1Ab.
Для специалистов в данной области очевидно, что токсины Bt, даже токсины, принадлежащие к определенному классу, такому как Cry1F, могут до некоторой степени варьироваться по своей длине и точной локализации перехода от части корового токсина в часть протоксина. Обычно, токсины Cry1Ca и Cry1Fa имеют длину примерно от 1150 до 1200 аминокислот. Переход от части корового токсина в часть протоксина обычно происходит на участке между частями, составляющими примерно от 50% и примерно до 60% от всей длины токсина. Химерный токсин согласно изобретению включает полноразмерную область этой N-концевой части корового токсина. Таким образом, химерный токсин содержит по меньшей мере примерно 50% полноразмерного белка Cry1Fa токсина Bt или по меньшей мере примерно 50% полноразмерного белка токсина Cry1Ca. Этот белок имеет длину, обычно составляющую по меньшей мере примерно 590 аминокислот. Что касается части протоксина, то полноразмерная область части протоксина Cry1Ab простирается от конца части корового токсина до С-конца молекулы.
Гены и токсины. Гены и токсины, используемые в настоящем изобретении, включают не только описанные здесь полноразмерные последовательности, но также и фрагменты этих последовательностей, варианты, мутанты и гибридные белки, которые сохраняют характерную пестицидную активность токсинов, конкретно описанных в настоящей заявке. Используемые здесь термины «варианты» или «модификации» генов означают нуклеотидные последовательности, которые кодируют те же самые токсины или токсины, эквивалентные токсинам, обладающим пестицидной активностью. Используемый здесь термин «эквивалентные токсины» означает токсины, обладающие такой же или, по существу, такой же биологической активностью против вредителей-мишеней, как и заявленные токсины.
Используемые здесь пределы идентичности составляют приблизительно 95% (Cry1Fa и Cry1Ca), 78% (Cry1F и Cry1C) и 45% (Cry1) в соответствии с «изменениями номенклатуры для пестицидных кристаллических белков Bacillus thuringiensis» («Revision of the Nomenclature for the Bacillus thuringiensis Pesticidal Crystal Proteins», N. Crickmore, D.R. Zeigler, J. Feitelson, E. Schnepf, J. Van Rie, D. Lereclus, J. Baum, and D.H. Dean. Microbiology and Molecular Biology Reviews (1998) Vol 62: 807-813). Такие пределы могут быть также применены только для коровых токсинов (для токсинов Cry1F и Cry1C).
Для специалистов в данной области очевидно, что гены, кодирующие активные токсины, могут быть идентифицированы и получены несколькими способами. Специфические гены или части генов, описанные в настоящей заявке, могут быть получены из изолятов, депонированных в депозитариях культур. Эти гены или их части или варианты могут быть также сконструированы путем синтеза, например, на синтезаторе генов. Варианты генов могут быть легко сконструированы стандартными методами получения точковых мутаций. Кроме того, фрагменты этих генов могут быть получены с использованием коммерчески доступных экзонуклеаз или эндонуклеаз в соответствии со стандартными процедурами. Так, например, для систематического отщепления нуклеотидов от концов этих генов могут быть использованы ферменты, такие как Bal31, либо может быть применен сайт-направленный мутагенез. Гены, кодирующие активные фрагменты, могут быть также получены с использованием различных рестриктирующих ферментов. Для непосредственного получения активных фрагментов этих белков-токсинов могут быть использованы протеазы.
Фрагменты и эквиваленты, которые сохраняют пестицидную активность описанных здесь токсинов, входят в объем настоящего изобретения. Кроме того, вследствие избыточности генетического кода, ряд различных последовательностей ДНК может кодировать описанные здесь аминокислотные последовательности. Специалист в данной области может легко получить такие альтернативные последовательности ДНК, кодирующие те же самые или, по существу, те же самые токсины. Такие варианты последовательностей ДНК входят в объем настоящего изобретения. Используемый здесь термин «по существу, те же самые» последовательности означает последовательности, которые имеют аминокислотные замены, делеции, добавления или инсерции, которые фактически не оказывают влияния на пестицидную активность. В это определение также входят фрагменты генов, кодирующих белки, сохраняющие пестицидную активность.
Другим методом идентификации генов, кодирующих токсины и части генов, используемых в настоящем изобретении, является применение олигонуклеотидных зондов. Такими зондами являются детектируемые нуклеотидные последовательности. Такие последовательности могут быть детектированы с помощью соответствующей метки, либо они могут быть изначально сделаны флуоресцентными, как описано в Международной заявке No. WO93/16094. Как хорошо известно специалистам, если молекула-зонд и образец нуклеиновой кислоты гибридизуются посредством образования прочной связи между двумя молекулами, то разумно предположить, что такой зонд и образец будут обладать значительной гомологией. Гибридизацию, предпочтительно, проводят в жестких условиях с применением методов, хорошо известных специалистам, например, описанных Keller, G. Н., M. M. Manak (1987) DNA Probes, Stockton Press, New York, N.Y., pp. 169-170. Ниже приводятся некоторые примеры комбинаций концентраций соли и температур (в порядке возрастания жесткости): 2 х SSPE или SSC при комнатной температуре; 1 х SSPE или SSC при 42°C; 0,1 х SSPE или SSC при 42°C; 0,1 х SSPE или SSC при 65°C. Детектирование зонда представляет собой известный метод, применяемый для того, чтобы определить, происходит гибридизация или нет. Такой анализ с использованием зонда представляет собой быстрый метод идентификации токсин-кодирующих генов согласно изобретению. Нуклеотидные сегменты, используемые в качестве зондов согласно изобретению, могут быть синтезированы на синтезаторе ДНК в соответствии со стандартными процедурами. Эти нуклеотидные последовательности могут быть также использованы в качестве ПЦР-праймеров для амплификации генов согласно изобретению.
Варианты токсинов. Некоторые токсины согласно изобретению конкретно описаны в настоящей заявке. Поскольку эти токсины приводятся здесь просто в качестве примеров токсинов согласно изобретению, то следует отметить, что настоящее изобретение включает варианты токсинов или эквивалентные токсины (и нуклеотидные последовательности, кодирующие эквивалентные токсины), обладающие такой же пестицидной активностью, как и представленный здесь токсин, или аналогичной активностью. Эквивалентные токсины имеют аминокислотную последовательность, гомологичную аминокислотной последовательности представленного здесь токсина. Такая гомология аминокислотных последовательностей обычно составляет более чем 75%, предпочтительно, более чем 90%, а наиболее предпочтительно, более, чем 95%. Гомология аминокислотных последовательностей является наивысшей в критических областях токсина, ответственных за биологическую активность или определяющих трехмерную конфигурацию, которая, в конечном счете, ответственна за биологическую активность. В соответствии с этим, некоторые аминокислотные замены являются допустимыми и могут присутствовать в тех областях, которые не играют важной роли в сообщении активности, или являются консервативными аминокислотными заменами, которые не влияют на трехмерную конфигурацию молекулы. Так, например, аминокислоты могут быть подразделены на следующие классы: неполярные, незаряженные полярные, основные и кислотные. Таким образом, при консервативных заменах, аминокислоту одного класса заменяют другой аминокислотой того же типа, и такая замена входит в объем настоящего изобретения, при условии, что она, фактически, не будет влиять на биологическую активность соединения. В таблице 1 представлен список примеров аминокислот, принадлежащих к каждому классу.
|
В некоторых случаях могут быть также сделаны неконсервативные замены. Важным фактором является то, что такие замены не должны значительно снижать биологическую активность токсина.
Рекомбинантные хозяева. Гены, кодирующие токсины согласно изобретению, могут быть введены микробным или растительным хозяевам широкого ряда. Экспрессия гена токсина приводит, прямо или опосредованно, к продуцированию пестицида внутри клеток и его сохранению в этих клетках. Для получения штамма Bt, экспрессирующего оба токсина согласно изобретению, может быть применен конъюгативный и рекомбинантный перенос. Другие организмы-хозяева могут быть также трансформированы одним или обоими генами токсинов, используемыми для достижения синергического эффекта. С использованием подходящих микробов-хозяев, например, Pseudomonas, эти микробы могут быть внесены в места обитания вредителей, где они могут размножаться и поедать эти микробы. Это будет приводить к уничтожению вредителей. Альтернативно, микроб, содержащий ген токсина, может быть обработан в условиях, способствующих пролонгированию активности токсина и стабилизации клетки. Обработанная клетка, которая сохраняет токсическую активность, может быть затем внесена в среду обитания вредителей-мишеней.
Если ген токсина Bt вводят микробу-хозяину посредством подходящего вектора, и если указанный хозяин вносят в среду обитания в живом виде, то важно, чтобы были использованы определенные микробы-хозяева. При этом, выбирают такие микроорганизмы-хозяева, которые, как известно, занимают определенную «фитосферу» (филлоплан, филлосферу, ризосферу и/или ризоплан) одного или нескольких представляющих интерес культур. Эти микроорганизмы выбирают так, чтобы они обладали способностью успешно конкурировать в конкретных условиях (в культуре и в другой среде обитания насекомых) с микроорганизмами дикого типа, обеспечивали стабильное сохранение и экспрессию генов, кодирующих полипептид-пестицид, и желательно, обеспечивали лучшую защиту пестицида от разрушения и инактивации в условиях окружающей среды.
Известно, что большое число микроорганизмов обитает на филлоплане (на поверхности листьев растений) и/или на ризосфере (в почве, окружающей корни растений) ценных сельскохозяйственных культур широкого ряда. Такими микроорганизмами являются бактерии, водоросли и грибы. Особый интерес представляют такие микроорганизмы, как бактерии, например, бактерии рода Pseudomonas, Erwinia, Serratia, Klebsiella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylophilius, Agrobactenum, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc и Alcaligenes; грибы, а в частности, дрожжи, например, дрожжи рода Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotorula и Aureobasidium. Особый интерес представляют такие виды бактерий фитосфер, как Pseudomonas syringae, Pseudomonas fluorescens, Serratia marcescens, Acetobacter xylinum, Agrobactenium tumefaciens, Rhodopseudomonas spheroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes entrophus и Azotobacter vinlandii; и дрожжей-фитосфер, таких как Rhodotorula rubra, R. glutinis, R. marina, R. aurantiaca, Cryptococcus albidus, C. diffluens, C. laurentii, Saccharomyces rosei, S. pretoriensis, S. cerevisiae, Sporobolomyces roseus, S. odorus, Kluyveromyces veronae и Aureobasidium pollulans. Особый интерес представляют пигментированные микроорганизмы.
Для введения гена Bt, кодирующего токсин, микроорганизму-хозяину в условиях, обеспечивающих стабильное сохранение гена и стабильную экспрессию гена, могут быть применены методы широкого ряда. Такие методы хорошо известны специалистам в данной области и описаны, например, в патенте США No. 5135867, который вводится в настоящее описание посредством ссылки.
Обработка клеток. Bacillus thuringiensis или рекомбинантные клетки, экспрессирующие токсины Bt, могут быть обработаны в целях пролонгирования активности токсина и стабилизации клеток. Образующаяся пестицидная микрокапсула содержит токсин или токсины Bt в клеточной структуре, которая стабилизирует и защищает токсин в случае, когда эту микрокапсулу вносят в среду обитания вредителя-мишени. Подходящими клетками-хозяевами могут быть прокариоты или эукариоты, и такими клетками обычно являются, но не ограничиваются ими, клетки, которые не продуцируют вещества, являющиеся токсичными для высших организмов, таких как млекопитающие. Однако могут быть также использованы и микроорганизмы, которые продуцируют вещества, токсичные для высших организмов, но, при этом, эти токсические вещества являются нестабильными, или уровень их введения является достаточно низким, что исключает возможность какого-либо токсического воздействия на млекопитающего-хозяина. В качестве хозяев, особый интерес представляют прокариоты и низшие эукариоты, такие как грибы.
Обрабатываемые клетки обычно являются интактными и, по существу, находятся в пролиферативной форме, а не в форме спор, хотя, в некоторых случаях могут использоваться и споры.
Обработка микробных клеток, например, микробов, содержащих ген или гены токсина Bt, может быть осуществлена химическими и/или физическими методами, или комбинацией химических и физических методов, при условии, что такой метод не будет оказывать негативного влияния на свойства токсина и не будет приводить к снижению способности клеток защищать токсин. Примерами химических реагентов являются галогенирующие агенты, а в частности, галогены с атомными номерами 17-80. Более конкретно, может быть использован йод в мягких реакционных условиях в течение определенного периода времени, достаточного для достижения желаемых результатов. Другими подходящими методами являются обработка альдегидами, такими как глутаральдегид; противоинфекционными агентами, такими как хлорид зефирана и хлорид цетилпиридиния; спиртами, такими как изопропиловый спирт и этанол; различными гистологическими фиксаторами, такими как йод Люголя, фиксатор Боуина; различные кислоты и фиксатор Хелли (см: Humason, Gretchen L., Animal Tissue Techniques, W. H. Freeman and Company, 1967); или комбинацией физического метода (нагревания) и химических агентов, которые сохраняют и пролонгируют активность токсина, продуцируемого в клетках, введенных в среду обитания хозяина. Примерами физических методов являются коротковолновое излучение, такое как гамма-излучение и рентгеновское излучение, замораживание, УФ-облучение, лиофилизация и т.п. Методы обработки микробных клеток описаны в патентах США №№ 4695455 и 4695462, которые вводятся в настоящее описание посредством ссылки.
Эти клетки обычно имеют повышенную структурную стабильность, что приводит к увеличению резистентности к условиям окружающей среды. Если пестицид присутствует в форме предшественника, то способ обработки клеток должен быть выбран так, чтобы он не приводил к ингибированию процессинга предшественника с образованием зрелой формы пестицида под действием патогена вредителя-мишени. Так, например, формальдегид будет обеспечивать перекрестное сшивание с белками, и тем самым ингибировать процессинг предшественника полипептидного пестицида. Способ обработки должен сохранять по меньшей мере значительную степень биологической доступности или биологической активности токсина.
Свойствами, представляющими особый интерес для продуцирования при выборе клетки-хозяина, являются простота введения гена или генов Bt хозяину, доступность экспрессионных систем, эффективность экспрессии, стабильность пестицида в хозяине и наличие дополнительных генетических признаков. Представляющими интерес технологическими свойствами пестицидных микрокапсул являются их защитная способность в отношении пестицида, например, толщина клеточных стенок, пигментация и внутриклеточная упаковка или способность образовывать тельца включения; выживаемость в водной среде; отсутствие токсичности для млекопитающих; привлекательность с точки зрения поедания вредителями; простота утилизации; фиксация без повреждения токсина и т.п. Другими рассматриваемыми свойствами являются простота приготовления препарата и его транспортировки, материальные затраты, стабильность при хранении и т.п.
Культивирование клеток. Клетки-хозяева, содержащие инсектицидный ген или гены B.t., могут быть выращены в любой подходящей питательной среде, в которой ДНК-конструкция будет обеспечивать селективное преимущество, то есть, обеспечивать селективную среду, в которой все или почти все клетки будут сохранять ген B.t. Затем эти клетки могут быть собраны обычными способами. Альтернативно, эти клетки могут быть обработаны до их сбора.
Клетки B.t., продуцирующие токсины согласно изобретению, могут быть культивированы с использованием стандартных сред и методом ферментации. После завершения цикла ферментации, бактерии могут быть собраны сначала путем отделения спор и кристаллов B.t. от сбраживаемого бульона стандартными методами. Выделенные споры и кристаллы B.t. могут быть приготовлены в виде смачиваемого порошка, жидкого концентрата, гранул или других препаратов, полученных путем добавления поверхностно-активных веществ, диспергирующих веществ, инертных носителей и других компонентов, облегчающих транспортировку и обработку ими конкретных вредителей-мишеней. Такие процедуры приготовления и применения хорошо известны специалистам.
Препараты. Приготовленные гранулы-приманки, содержащие аттрактант и споры, кристаллы и токсины изолятов B.t., или рекомбинантные микробы, содержащие гены, полученные из описанных здесь изолятов B.t., могут быть внесены в почву. Приготовленный продукт может быть применен в виде покрытия, наносимого на семена, или препарата для обработки корней или всего растения на последних стадиях цикла выращивания сельскохозяйственной культуры. Для обработки растений и почвы, клетки B.t. могут быть приготовлены в виде смачиваемых порошков, гранул или дустов, путем смешивания с различными инертными материалами, такими как неорганические минеральные вещества (филлосиликаты, карбонаты, сульфаты, фосфаты и т.п.) или растительные материалы (измельченные в порошок початки кукурузы, рисовая шелуха, скорлупа грецкого ореха и т.п.). Такие препараты могут включать адъюванты типа «распылителей-связующих веществ», стабилизирующие агенты, другие пестицидные добавки или поверхностно-активные вещества. Жидкие препараты могут быть водными или безводными, и могут быть использованы в виде пен, гелей, суспензий, эмульгируемых концентратов или т.п. Ингредиентами могут быть реологические агенты, поверхностно-активные вещества, эмульгаторы, диспергирующие вещества или полимеры.
Как известно специалистам в данной области, концентрация пестицида может значительно варьироваться в зависимости от природы конкретного препарата, а в частности, в зависимости от того, используется ли он в виде концентрата или в чистом виде. Пестицид составляет по меньшей мере 1% по массе, а может составлять 100% по массе. Сухие препараты могут составлять примерно 1-95% по массе пестицида, а жидкие препараты обычно составляют примерно 1-60% по массе твердых веществ в жидкой фазе. Эти препараты обычно содержат примерно от 102 до 104 клеток/мг. Эти препараты могут быть введены в количестве примерно от 50 мг (в жидком или в сухом виде) до 1 кг или более на гектар.
Препараты могут быть внесены в среду обитания чешуекрылых вредителей, например, нанесены на листья или почву путем распыления, опыливания, орошения или т.п.
Трансформация растений. Предпочтительными рекомбинантными хозяевами, которые могут быть использованы для продуцирования инсектицидных белков согласно изобретению, являются трансформированные растения. Гены, кодирующие описанные здесь белки токсинов Bt, могут быть введены в растительные клетки с применением различных методов, хорошо известных специалистам. Так, например, существует большое число клонирующих векторов, содержащих систему репликации в Escherichia coli, и маркер, позволяющий проводить отбор трансформированных клеток, и эти векторы могут быть использованы в целях получения препарата для инсерции чужеродных генов в высшие растения. Векторы содержат, например, inter alia pBR322, серии pUC, серии M13mp, pACYC184. В соответствии с этим, ДНК-фрагмент, имеющий последовательность, кодирующую белок токсина Bt, может быть встроен в вектор в подходящий рестрикционный сайт. Полученную плазмиду используют для трансформации E. coli. Клетки E. coli культивируют в подходящей питательной среде, а затем собирают и подвергают лизису. Затем эту плазмиду выделяют. Методами анализа обычно являются анализ последовательности, рестрикционный анализ, электрофорез и другие методы, применяемые в биохимии и молекулярной биологии. После каждой манипуляции, используемая последовательность ДНК может быть расщеплена и присоединена к следующей последовательности ДНК. Каждая последовательность плазмиды может быть клонирована в одной и той же или в других плазмидах. В зависимости от метода встраивания нужных генов в растение, могут оказаться необходимыми и другие последовательности ДНК. Так, например, если плазмиду Ti или Ri используют для трансформации клеток растения, то по меньшей мере правую границу, а в большинстве случаев, правую и левую границы Т-ДНК-плазмиды Ti или Ri присоединяют в качестве фланкирующей области встраиваемых генов. Использование T-ДНК для трансформации клеток растений было тщательно исследовано и подробно описано в EP 120516, Lee and Gelvin (2008), Hoekema (1985), Fraley et al., (1986), и An et al., (1985), и хорошо известно специалистам.
После интеграции встроенной ДНК в геном растения, такая ДНК становится относительно стабильной. Трансформирующий вектор обычно содержит селективный маркер, который сообщает трансформированным клеткам растения резистентность к биоциду или антибиотику, таким как биалафос, канамицин, G418, блеомицин или гигромицин, inter alia. С использованием отдельно взятого маркера можно, соответственно, осуществлять отбор трансформированных клеток, а не клеток, которые не содержат встроенную ДНК.
Существует много методов, подходящих для встраивания ДНК в клетки растений-хозяев. Такими методами являются трансформация молекулой T-ДНК с использованием Agrobacterium tumefaciens или Agrobacterium rhizogenes в качестве трансформирующих агентов, слияние, инжекция, биобаллистические методы (бомбардировка микрочастицами) или электропорация, а также другие возможные методы. Если для трансформации используются агробактерии, то встраиваемую ДНК клонируют в конкретные плазмиды, а именно, в промежуточный вектор или в бинарный вектор. Промежуточные векторы могут быть интегрированы в плазмиду Ti или Ri посредством гомологичной рекомбинации с использованием последовательностей, гомологичных последовательностям, присутствующим в T-ДНК. Плазмида Ti или Ri также содержит область vir, необходимую для переноса T-ДНК. Промежуточные векторы не могут сами реплицироваться в агробактериях. Промежуточный вектор может быть перенесен в Agrobacterium tumefaciens с помощью хелперной плазмиды (конъюгирование). Бинарные векторы могут сами реплицироваться как в E. coli, так и в агробактериях. Они содержат селективный маркерный ген и линкер или полилинкер, которые замыкают правую и левую пограничные области T-ДНК. Они могут быть трансформированы непосредственно в агробактерии (Holsters et al., 1978). Агробактерия, используемая в качестве клетки-хозяина, содержит плазмиду, несущую область vir. Область vir необходима для переноса T-ДНК в клетку растения. Может также присутствовать и дополнительная T-ДНК. Трансформированную таким образом бактерию используют для трансформации клеток растений. Растительные эксплантаты могут быть преимущественно культивированы с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в клетку растения. Затем целые растения могут быть выращены из инфицированного растительного материала (например, из кусочков листьев, сегментов стеблей, корней, а также протопластов или клеток, культивированных суспензионным методом) в подходящей среде, которая может содержать антибиотики или биоциды для отбора. Затем выращенные таким образом растения могут быть протестированы на присутствие встроенной ДНК. В случае инжекции и электропорации, каких-либо специальных требований к получению плазмид не предъявляется. При этом, могут быть использованы стандартные плазмиды, такие как, например, производные pUC.
Трансформированные клетки развиваются в растении как обычно. Они могут образовывать зародышевые клетки и передавать трансформированный(ые) признак(и) потомству. Такие растения могут быть выращены обычным способом и скрещены с растениями, имеющими трансформированные наследуемые факторы или другие наследуемые факторы. Полученные гибридные растения имеют соответствующие фенотипические свойства.
В предпочтительном варианте изобретения, растения трансформируют генами, в которых встречаемость кодонов оптимизирована для растений. См, например, патент США No. 5380831, который вводится в настоящее описание посредством ссылки. Хотя в настоящей заявке описаны некоторые усеченные токсины, однако, специалистам по Bt хорошо известно, что токсины, принадлежащие к типу токсинов длиной 130 кДа (полноразмерные), имеют N-концевую половину, которая представляет собой коровый токсин, и С-концевую половину, которая представляет собой протоксиновый «хвост». Таким образом, соответствующие «хвосты» могут быть использованы вместе с усеченными/коровыми токсинами согласно изобретению. См., например, патент США № 6218188 и патент США № 6673990. Кроме того, методы создания синтетических генов Bt для их использования в растениях известны специалистам (Stewart and Burgin, 2007). Одним из неограничивающих примеров предпочтительного трансформированного растения является фертильное растение кукурузы, содержащее экспрессируемый в растении ген, кодирующий белок Cry1Fa, а также второй экспрессируемый в растении ген, кодирующий белок Cry1Ca.
Перенос (или интрогрессия) Cry1Fa- и Cry1Ca-детерминированного(ых) признака(ов) в инбредные линии кукурузы может быть достигнут путем рекуррентного селективного скрещивания, например, возвратного скрещивания. В этом случае, нужное рекуррентное растение сначала скрещивают с инбредным донором (нерекуррентным родителем), который несет соответствующий(ие) ген(ы), сообщающий(е) Cry1F- и Cry1C-детерминированные признаки. Затем потомство этого кросса подвергают возвратному скрещиванию с рекуррентным растением с последующим отбором полученного потомства на нужный(ые) признак(и), перенесенный(е) от нерекуррентного родителя. Через три, предпочтительно, четыре, а еще более предпочтительно, через пять или более поколений «бэккроссов» с рекуррентным родителем с отбором на нужный(е) признак(и), потомство будет гетерозиготным по локусам, контролирующим перенесенный(е) признак(и), но оно будет аналогично рекуррентному родителю по большинству или почти по всем другим генам (см., например, Poehlman & Sleper (1995) Breeding Field Crops, 4th Ed., 172-175; Fehr (1987) Principles of Cultivar Development, Vol. 1: Theory and Technique, 360-376).
Стратегии выращивания культур, резистентных к насекомым (IRM). Например, Roush и сотрудниками были описаны стратегии с использованием двух токсинов, также называемые созданием «пирамид» или «кластеров» для выращивания трансгенных культур, обладающих инсектицидными свойствами. (The Royal Society. Phil. Trans. R. Soc. Lond. B. (1998) 353, 1777- 1786).
На web-сайте Агенства США по защите окружающей среды (epa.gov/oppbppdl/biopesticides/pips/bt_corn_refuge_2006. htm) опубликованы следующие требования по обеспечению площадей-убежищ с нетрансгенными культурами (то есть, не-B.t.) культурами (земельного участка или блока с сельскохозяйственными не-Bt-культурами/кукурузой) для их использования под трансгенные культуры, продуцирующие один белок Bt, обладающий активностью против вредителей-мишеней.
«Конкретными структурными требованиями к продуктам из Bt-кукурузы (Cry1Ab или Cry1F), защищенной от кукурузного пилильщика, являются:
Структурные площади-«убежища»: 20% площади-убежища под Bt-кукурузу, не защищенную от Чешуекрылых в Кукурузном поясе;
50% площади-убежища под Bt-хлопчатник, не защищенный от Чешуекрылых в Хлопковом поясе;
Блоки:
Внутренние (то есть, в полях с Bt)
Внешние (то есть, отдельные поля в пределах ½ мили (по возможности ¼ мили) от Bt-поля для максимизации свободного скрещивания)
Полоски регулярно обрабатываемых сельскохозяйственных земель:
Эти полоски должны иметь ширину по меньшей мере в 4 ряда (предпочтительно, 6 рядов) для снижения числа случаев миграции личинок».
Кроме того, Национальная ассоциация производителей кукурузы, на своем web-сайте (ncga.com/insect-resistance-management-fact-sheet-bt-corn) также опубликовала аналогичное руководство по требованиям для площадей-убежищ. Так, например:
«Требования к IRM в случае кукурузного пилильщика:
- Засевать по меньшей мере 20% акров кукурузой для сохранения нетрансгенных гибридов
- В регионах выращивания хлопчатника, должно оставаться 50% площади-убежища
- Должно быть засеяно 1/2 мили нетрансгенными гибридами
- Площади-убежища могут быть засеяны полосами на Bt-поля; площади-убежища должны быть засеяны в виде полос, которые должны иметь ширину по меньшей мере в 4 ряда
- Площади-убежища могут быть обработаны стандартными пестицидами только, если достигаются экономические пороги для насекомых-мишеней
- Распыляемые инсектициды на основе Bt не могут быть использованы на площадях-убежищах под кукурузу
- Соответствующее убежище должно быть засеяно Bt-кукурузой на каждой ферме»
Как указывали Roush и сотрудники (например, на страницах 1780 и 1784 в правой колонке), кластеры или пирамиды из двух различных белков, каждый из которых является эффективным против вредителей-мишеней с минимальной перекрестной резистентностью или с отсутствием такой резистентности, могут быть использованы на более мелких «убежищах» нетрансгенных растений. Roush высказал предположение, что для успешного использования кластеров, площадь-убежище, размер которого составляет менее, чем 10%, может быть возделана культурой с резистентностью, сравнимой с резистентностью культур, возделываемой примерно на 50% площади-убежища для одного (не-пирамидного) токсина. Что касается доступных в настоящее время продуктов из кукурузы, содержащих «пирамидные» Bt, то Агенство США по защите окружающей среды требует, чтобы значительно меньшая (обычно 5%) площадь структурного убежища была засеяна не-Bt кукурузой, а не культурой с одним токсином (обычно 20%).
Существует различные пути обеспечения IRM-эффектов использования площадей-убежищ, включая различные геометрические схемы засева на поля (как упоминалось выше) и смеси семян в одном пакете, что также обсуждается Roush и др. (см. выше), и в патенте США № 6551962.
Вышеуказанные проценты или аналогичные соотношения площадей-убежищ могут быть использованы для рассматриваемых двухкомпонентных или трехкомпонентных кластеров или пирамид. Для трехкомпонентных кластеров с тремя механизмами действия против одного вредителя-мишени, убежища вообще быть не должно (или например, площадь-убежище должна быть менее 5%). Это особенно справедливо для площадей под коммерческие культуры - например, свыше 10 акров.
Все патенты, патентные заявки, предварительные заявки и публикации или цитируемые в них работы во всей своей полноте вводятся в настоящее описание посредством ссылки в той степени, в которой они соответствуют детальному описанию настоящей заявки.
Ниже приводятся примеры, которые иллюстрируют способы практического осуществления настоящего изобретения.
Эти примеры не должны рассматриваться как ограничение объема изобретения. Все проценты даны по массе, а все соотношения смесей растворителей даны по объему, если это не оговорено особо. Все температуры даны в градусах Цельсия.
Если это не указано или не подразумевается конкретно, то используемые здесь артикли «a», «an» и «the» означают «по меньшей мере один».
Пример 1
Конструирование химерных белков, содержащих коровые токсины Cry1Ca и протоксины Cry1Ab
Химерные токсины. Химерные белки, содержащие домен корового токсина одного Cry, присоединенного к протоксиновому сегменту другого токсина Cry, были уже описаны, например, в патенте США № 5593881 и в патенте США № 5932209. Последовательность белка эндотоксина Cry1Ca3-дельта депонирована в GenBank под регистрационным номером AAA22343 под устаревшим обозначением Cry1C(b).
Вариантами химерных белков Cry1Ca согласно изобретению являются химерные токсины, содержащие N-концевой сегмент корового токсина, происходящий от инсектицидного токсина Cry1Ca3, присоединенного к гетерологичному протоксиновому сегменту эндотоксина дельта в определенном положении, расположенном за пределами сегмента корового токсина. Переход от корового токсина в гетерологичный сегмент протоксина может происходить приблизительно в природной области стыка токсин/протоксин, или альтернативно, часть нативного протоксина (простирающаяся за пределы сегмента корового токсина) может сохраняться, причем, переход в гетерологичный протоксин может происходить ниже. В одном из вариантов, сегменты корового токсина и протоксина могут включать точно такую же аминокислотную последовательность нативных токсинов, от которых они происходят, либо они могут включать аминокислотные добавления, делеции или замены, которые не ухудшают, а могут даже улучшать биологическую функцию сегментов, связанных друг с другом.
Так, например, химерный токсин согласно изобретению содержит сегмент корового токсина, происходящий от Cry1Ca3, и гетерологичный протоксин. В предпочтительном варианте изобретения, сегмент корового токсина, происходящий от Cry1Ca3 (619 аминокислот), присоединен к гетерологичному сегменту, содержащему сегмент протоксина, происходящий от дельта-эндотоксина Cry1Ab (545 аминокислот). Последовательность химерного белка из 1164 аминокислот, обозначенного здесь DIG-152, представлена как SEQ ID NO:1. Второй предпочтительный вариант изобретения включает химерный белок, в котором сегмент корового токсина Cry1Ca (619 аминокислот) присоединен ко второму сегменту протоксина из 545 аминокислот, происходящему от Cry1Ab. Последовательность второго химерного белка из 1164 аминокислот, обозначенного здесь DIG-109, представлена как SEQ ID NO:2 (вариант, оптимизированный для кукурузы). Следует отметить, что в объем настоящего изобретения входят и другие химрные гибриды, содержащие варианты корового токсина Cry1Ca и протоксины, происходящие от Cry1Ab.
Следует отметить, что химерные белки DIG-152 и DIG-109, по существу, функционально эквивалентны друг другу, и отличаются только в одном положении последовательности (в положении аминокислоты 620, которая является положением присоединения сегмента корового токсина Cry1Ca к сегменту протоксина Cry1Ab).
Пример 2
Конструирование экспрессионных плазмид, кодирующих химерные белки «коровый токсин Cry1Ca/протоксин Cry1Ab», и их экспрессия в Pseudomonas
Для создания экспрессионной конструкции pMYC2547 Pseudomonas fluorescens (Pf), полученной в целях продуцирования полноразмерного химерного белка DIG-152 (представленного в SEQ ID NO:1) были применены стандартные методы клонирования (описанные, например, в руководстве Sambrook et al, (1989) и Ausubel et al, (1995), и в более поздних изданиях). Продуцирование белка осуществляли в штамме MB214 Pseudomonas fluorescens (производном штамма MB101; P. fluorescens, биовар 1), имеющем инсерцию модифицированного оперона lac, как описано в патенте США № 5169760. Основная стратегия клонирования включает субклонирование фрагмента ДНК, кодирующего DIG-152, в плазмидные векторы, в результате чего этот фрагмент будет помещен под экспрессионный контроль промотора Ptac и терминатора rrnBTlT2, происходящего от плазмиды pKK223-3 (PL Pharmacia, Milwaukee, WI). Одна из таких плазмид обозначается pMYC2547, а изолят MB214, содержащий эту плазмиду, обозначается Dpfl08.
Анализ на рост и экспрессию в шейкерных колбах
Продуцирование белка DIG-152 для характеризации и биологического анализа на его действие против насекомых осуществляли с использованием штамма Dpf108 P. fluorescens, выращенного в шейкерных колбах. Продуцирование белка DIG-152, находящегося под контролем промотора Ptac, осуществляли, как описано ранее в патенте США No. 5527883. Подробное описание микробиологических манипуляций приводится в публикации Squires et al., (2004), в заявке на патент США 20060008877, в заявке на патент США 20080193974 и в заявке на патент США 20080058262, которые вводятся в настоящее описание посредством ссылки. Экспрессия была индуцирована путем добавления изопропил-β-D-1-тиогалактопиранозида (IPTG) после первого инкубирования в течение 24 часов при 30°С со встряхиванием. Образцы культур брали во время индуцирования и в различные периоды времени после индуцирования. Клеточную плотность измеряли по оптической плотности на 600 нм (OD600).
Фракционирование клеток и анализ образцов в шейкерных колбах, проводимый с помощью электрофореза в ПААГ с ДСН. При каждом взятии образца, клеточную плотность образцов доводили до OD600=20, и 1 мл-аликвоты центрифугировали при 14000 x g в течение пяти минут. Клеточный осадок замораживали при -80°С. Растворимые и нерастворимые фракции, взятые из замороженных образцов клеточного осадка в шейкерных колбах, получали с использованием раствора для экстракции бактериального белка EasyLyse™ (EPICENTRE® Biotechnologies, Madison, WI). Каждый клеточный осадок ресуспендировали в 1 мл раствора EasyLyse™, а затем разводили 1:4 в буфере для лизиса и инкубировали со встряхиванием при комнатной температуре в течение 30 минут. Лизат центрифугировали при 14000 об/мин в течение 20 минут при 40C, и супернатант выделяли в виде растворимой фракции. Затем осадок (нерастворимую фракцию) ресуспендировали в равном объеме забуференного фосфатом физиологического раствора (PBS; 11,9 мМ Na2HPO4, 137 мМ NaCl, 2,7 мМ KCl, pH 7,4).
Образцы смешивали в отношении 1:1 с 2 х буфером для образцов Лэммли, содержащем β-меркаптоэтанол (Sambrook et al, см. выше), и кипятили в течение 5 минут, а затем загружали на 12% бис-трис-гель Criterion XT (Bio-Rad Inc., Hercules, CA). Электрофорез осуществляли в XT-буфере MOPS, рекомендованном производителем. Гели окрашивали кумасси синим Bio-Safe в соответствии с протоколом, рекомендованным производителем (Bio-Rad), и визуализировали с использованием визуализирующей системы Alpha Innotech (San Leandro, CA).
Получение телец включения. Получение телец включения белка DIG-152 (IB) осуществляли в клетках, полученных путем реакции ферментации P. fluorescens, которые продуцировали нерастворимый инсектицидный Bt-белок, как показал анализ, проводимый с помощью электрофореза в ПААГ с ДСН и MALDI-MS (матричная лазерная десорбция/ионизирующая масс-спектрометрия). Гранулы, полученные путем ферментации P. fluorescens, оттаивали на водяной бане при 37°С. Клетки ресуспендировали до 25% масс/об в буфере для лизиса [50 мМ триса, pH 7,5, 200 мМ NaCl, 20 мМ EDTA-динатриевой соли (этилендиаминтетрауксусной кислоты), 1% тритона X-100 и 5 мМ дитиотреитола (DTT); 5 мл/л «коктейля» ингибиторов бактериальной протеазы (Catalog # P8465; Sigma-Aldrich, St. Louis, MO), который добавляли непосредственно перед применением]. Клетки суспендировали на гомогенизаторе с ручным управлением с установкой шкалы наименьшего значения (Tissue Tearor, BioSpec Products, Inc., Bartlesville, OK). Лизоцим (25 мг Sigma L7651, выделенного из белка куриных яиц) добавляли к клеточной суспензии путем смешивания металлическим шпателем, и суспензию инкубировали при комнатной температуре в течение одного часа. Суспензию охлаждали на льду в течение 15 минут, а затем обрабатывали ультразвуком на ультразвуковом генераторе Branson Sonifier 250 (два раунда по 1 минуте, 50% дежурный цикл, 30% выход). Лизис клеток подтверждали с помощью микроскопии. При необходимости добавляли 25 мг лизоцима, а затем инкубирование и обработку ультразвуком повторяли. После подтверждения лизиса клеток под микроскопом, лизат центрифугировали при 11500 x g в течение 25 минут (4°С) с получением осадка телец включения (IB), и супернатант отбрасывали. Осадок IB ресуспендировали со 100 мл буфера для лизиса, гомогенизировали в миксере с ручным управлением и центрифугировали как описано выше. Осадок IB повторно промывали путем ресуспендирования (в 50 мл буфера для лизиса), гомогенизации, обработки ультразвуком и центрифугирования до тех пор, пока супернатант не становился бесцветным, а осадок IB твердым и не совсем белым. Для конечной промывки, осадок IB ресуспендировали в стерильно отфильтрованной (на фильтре 0,22 мкм) дистиллированной воде, содержащей 2 мМ EDTA, и центрифугировали. Конечный осадок ресуспендировали в стерильно отфильтрованной дистиллированной воде, содержащей 2 мМ EDTA, и хранили в виде 1 мл-аликвот при -80°С.
Анализ, проводимый посредством электрофореза в ПААГ с ДСН, и количественную оценку белка в препаратах IB осуществляли путем оттаивания 1 мл-аликвоты осадка IB и разведения 1:20 стерильно отфильтрованной дистиллированной водой. Затем разведенный образец кипятили с 4 х восстанавливающим буфером для образцов [250 мМ трис, pH 6,8, 40% глицерина (об/об), 0,4% бромфенолового синего (масс/об), 8% ДСН (масс/об) и 8% β-меркаптоэтанола (об/об)] и загружали на 4-20% трис-глицин Novex® в геле в лунках 12+2 (Invitrogen), обработанном 1 х трисом/глицином/ДСН-буфером (BioRad). Гель подвергали электрофорезу в течение 60 минут при 200 вольт, а затем окрашивали кумасси синим (50% G-250/50% R-250 в 45% металоле, 10% уксусной кислоте), и обесцвечивали 7% уксусной кислотой, 5% метанолом в дистиллированной воде. Количественную оценку полос-мишеней осуществляли путем сравнения денситометирических величин для полос со стандартными образцами альбумина бычьей сыворотки (BSA), проанализированных на том же геле для построения стандартной кривой.
Солюбилизация телец включения. 6 мл суспензии телец включения DIG-152 от клона Pf, DPfl08, центрифугировали на микроцентрифуге Эппендорфа модели 5415C, установленной на наивысшее значение (приблизительно 14000 x g), в результате чего получали осадок телец включения. Супернатант буфера для хранения удаляли и заменяли 25 мл 100 мМ натрийкарбонатного буфера, pH 11, в конической 50 мл-пробирке. Тельца включения ресуспендировали пипеткой и подвергали вихревому перемешиванию до получения однородной смеси. Пробирку помещали на медленно вращающуюся платформу и оставляли на ночь при 4°С для экстракции белка-мишени. Экстракт центрифугировали при 30000 x g в течение 30 минут при 4°С, и полученный супернатант 5-кратно концентрировали на центрифужном фильтрующем устройстве с регенерированной целлюлозой Amicon Ultra-15 (с отсечкой молекулярной массы 30000; Millipore). Буфер для образцов заменяли 10 мМ CAPS [3-(циклогексамино)-1-пропансульфоновой кислотой], pH 10, с использованием одноразовых колонок PD-10 (GE Healthcare, Piscataway, NJ).
Солюбилизация белка телец включения и их активация трипсином. В некоторых случаях, суспензию DIG-152 телец включения от клона Pf, DPfl08, центрифугировали на микроцентрифуге Эппендорфа модели 5415C, установленной на наивысшее значение (приблизительно 14000 x g), в результате чего получали осадок телец включения. Супернатант буфера для хранения удаляли и заменяли 100 мМ CAPS, pH 11, с получением белка, концентрация которого составляла примерно 50 мг/мл. Пробирку вращали при комнатной температуре в течение трех часов до полной солюбилизации белка. Затем добавляли трипсин в равных количествах до 5% - 10% (масс.%, по исходной массе порошка IB) и осуществляли гидролиз путем инкубирования при вращении в течение ночи при 4°С или вращении в течение 90-120 минут при комнатной температуре. Нерастворимый материал удаляли путем центрифугирования при 10000 x g в течение 15 минут, и супернатант наносили на анионообменную колонку MonoQ (10 мм х 10 см). Активированный белок DIG-152 элюировали (как было определено с помощью электрофореза в ПААГ с ДСН) 25 колоночными объемами 0% - 100% 1М градиента NaCl. Фракции, содержащие активированный белок, объединяли, и при необходимости, концентрировали до менее чем 10 мл на центрифужном фильтрующем устройстве с регенерированной целлюлозой Amicon Ultra-15 как описано выше. Затем материал пропускали через колонку с Superdex 200 (16 мм х 60 см) в буфере, содержащем 100 мМ NaCl, 10% глицерина, 0,5% твина-20 и 1 мМ EDTA. Анализ, проводимый с помощью электрофореза в ПААГ с ДСН, показал, что активированный (ферментативно усеченный) белок элюируется при 65-70 мл. Фракции, содержащие активированный белок, собирали и концентрировали на центрифужном концентраторе как описано выше.
Гель-электрофорез. Препараты концентрированного белка получали для проведения электрофореза путем разведения 1:50 в буфере для образцов LDS NuPAGE® (Invitrogen), содержащем 5 мМ DTT в качестве восстановителя, и нагревали при 95°С в течение 4 минут. Образец загружали на дорожки-дубликаты с 4-12% гелем NuPAGE® вместе с пятью BSA-стандартами в количестве от 0,2 мкг до 2 мкг/дорожку (для построения стандартной кривой). Затем подавали напряжение в 200 В с использованием буфера MOPS для электрофороза с ДНС (Invitrogen) до тех пор, пока краситель-«свидетель» не достигал основания геля. Гель окрашивали 0,2% кумасси синим G-250 в 45% метаноле, 10% уксусной кислотой, и обесцвечивали сначала путем быстрой обработки 45% металолом, 10% уксусной кислотой, а затем путем длительной обработки 7% уксусной кислотой, 5% метанолом до появления прозрачного фона. После обесцвечивания, гель сканировали на мультивизуализаторе BioRad Fluor-S. Для получения объемов окрашенных полос белка с вычитанием фона и для построения стандартной кривой BSA, которая была использована для вычисления концентрации химерного белка DIG-152 в маточном растворе, использовали компьютерную программу Quantity One Software v.4.5.2.
Пример 3
Инсектицидная активность белка DIG-152, продуцируемого в Pseudomonas fluorescens
Инсектицидная активность белка DIG-152 была продемонстрирована на личинках совки травяной (FAW, Spodoptera frugiperda (J.E. Smith)) и на личинках Cry1F-резистентной FAW (rFAW).
Приготовление и биоанализы образцов. Препараты телец включения (нативного полноразмерного белка или белка, активированного трипсином) переносили в 10 мМ CAPS-буфер, pH10, методами замены буфера, такими как диализ, или на колонках PD-10. Затем образцы соответствующим образом разводили в 10 мМ CAPS, pH 10, и все биоаналитические препараты содержали контроль, состоящий из этого буфера, который служил в качестве фона для подтверждения гибели насекомых или ингибирования их развития.
Концентрации белков в буфере для биоанализа оценивали с помощью гель-электрофореза с использованием BSA в целях построения стандартной денситометрической кривой для геля, и этот анализ проводили с использованием системы визуализации BioRad, как описано выше. Белки в гелевой матрице окрашивали красителем на основе кумасси синего, и перед считыванием обесцвечивали.
Очищенные белки тестировали на инсектицидную активность с помощью биоанализов, проводимых с использованием только что вылупившихся личинок чешуекрылых, которым давали искусственный корм для насекомых. Личинки FAW вылуплялись из яиц, полученных из колоний, хранящихся в коммерчески доступном инсектарии (Benzon Research Inc., Carlisle, PA). Личинки rFAW вылуплялись из яиц, полученных из не-промышленных колоний (Dow AgroSciences, Indianapolis, IN).
Биоанализы проводили в 128-луночных пластиковых планшетах, специально изготовленных для биоанализов насекомых (C-D International, Pitman, NJ). Каждая лунка содержала 1,0 мл корма для чешуекрылых многих видов (Southland Products, Lake Village, AR). 40 аликвот образца белка наносили пипеткой на поверхность в 1,5 см2 каждой лунки с кормом (то есть 26 мкл/см2). Концентрацию корма вычисляли как количество (нг) белка DIG-152 на квадратный сантиметр площади поверхности в лунке. Обработанные лунки выдерживали в вытяжном шкафу до тех пор, пока жидкость на поверхности корма не испарялась или не абсорбировалась в этом корме.
В течение нескольких часов размножения, отдельные личинки собирали увлажненной щеткой из верблюжьей шерсти и помещали на обработанный корм, одну личинку на лунку. Затем зараженные лунки герметично закрывали клейкими прозрачными пластиковыми пластинами, и снабжали отверстием для газообмена (C-D International). Планшеты для биоанализа выдерживали в регулируемых условиях окружающей среды [28°, относительная влажность приблизительно 40% (ОВ), 16 ч:8 ч (день:ночь)] в течение 5 дней, после чего регистрировали общее число насекомых, обработанных каждым образцом белка, число погибших насекомых и массу выживших насекомых. Процент погибших насекомых и процент ингибирования их роста вычисляли для каждой обработки. На каждой стадии обработки вычисляли процент погибших насекомых и процент ингибирования их роста. Процент ингибирования роста (GI) вычисляли следующим образом:
%GI = [1 - (TWIT/TNIT)/(TWIBC/TNIBC)] x 100,
где TWIT означает общую массу насекомых для данной обработки (Total Weight of Insects in the Treatment),
TNIT означает общее число насекомых для данной обработки (Total Number of Insects in the Treatment),
TWIBC означает общую массу насекомых в фоновом контроле (Total Weight of Insects in the Background Check) (буфер-контроль), и
TNIBC означает общее число насекомых в фоновом контроле (Total Number of Insects in the Background Check) (буфер-контроль).
GI50 определяли как концентрацию химерного белка DIG- 152 в корме, где величина %GI равна 50. LC50 (50% летальную концентрацию) регистрировали как концентрацию белка DIG-152 в корме, при которой погибало 50% тестируемых насекомых. Статистический анализ (односторонний ANOVA) проводили с использованием компьютерной программы JMP (SAS, Cary, NC).
В таблице 2 представлены результаты биоанализов белка DIG- 152 на его поедание личинками насекомого совки травяной.
Таблица 2. Величины GI50 и LC50 (в нг/см2), вычисленные для корма насекомых, на верхнюю часть которого был нанесен белок DIG-152.
|
Отличительным признаком белка DIG-152 согласно изобретению является то, что рост только что вылупившихся личинок совки травяной (Spodoptera frugiperda) ингибируется после поедания или белка DIG-152. Кроме того, личинки совки травяной, которые являются резистентными к интоксикации белком Cry1Fa, остаются восприимчивыми к активности DIG-152, как и личинки совки травяной дикого типа. Значимость восприимчивости этих Cry1Fa-резистентных насекомых к Cry1Ca более подробно обсуждалась выше.
Пример 4
Конструирование последовательности, оптимизированной по кодонам кукурузы и кодирующей белок DIG-109
Специалисту в области молекулярной биологии растений известно, что множество последовательностей ДНК может быть сконструировано так, чтобы они кодировали одну аминокислотную последовательность. Общеизвестный способ повышения уровня экспрессии области, кодирующей представляющей интерес белок, заключается в создании такой кодирующей области, в которой состав кодонов аналогичен общему составу кодонов хозяина, в котором должна быть достигнута экспрессия генов. Руководство по конструированию и продуцированию синтетических генов можно найти, например, в заявке WO 1997/13402 и в патенте США № 5380831.
Последовательность ДНК, имеющую смещение кодонов кукурузы, конструировали и синтезировали так, чтобы она продуцировала химерный инсектицидный белок DIG-109 в трансгенных однодольных растениях. Таблицу встречаемости кодонов в кукурузе (Zea mays L.) составляли исходя из 706 белок-кодирующих последовательностей, выведенных из последовательностей, депонированных в GenBank (world wide web: ncbi.nlm.nih.gov). Средневесовой набор кукурузных кодонов вычисляли путем исключения любого избыточного кодона, количество которого составляет примерно менее чем 10% от общей встречаемости кодонов для данной аминокислоты. Средневесовое представление для каждого кодона вычисляли по формуле:
Средневесовой % C1 = 1/(%C1+%C2+%C3+т.п.)x%C1x100, где C1 представляет собой рассматриваемый кодон, а %C2, %C3 и т.п. означает средний % встречаемости остальных синонимичных кодонов.
Для получения последовательности ДНК, оптимизированной по кодонам кукурузы и кодирующей белок DIG-109 SEQ ID NO:2, состоящий из 1164 аминокислот, были сделаны замены кодонов в нативной последовательности ДНК crylCa, кодирующей сегмент корового токсина Cry1Ca, в результате чего была получена последовательность ДНК, имеющая общий состав оптимизированных для кукурузы кодонов, представленный в таблице смещения кодонов. Аналогичным образом, замены кодонов в нативной последовательности ДНК crylAb, кодирующей сегмент протоксина Cry1Ab были сделаны так, чтобы полученная последовательность ДНК имела общий состав оптимизированных для кукурузы кодонов, представленный в таблице смещения кодонов. Дополнительные уточнения последовательностей были сделаны для устранения нежелательных сайтов распознавания рестриктирующих ферментов, потенциальных сайтов сплайсинга интронов растений, длинных участков остатков A/T или C/G, и других мотивов, которые могут негативно влиять на стабильность РНК, транскрипцию или трансляцию кодирующей области в клетках растений. Для введения нужных сайтов распознавания рестриктирующих ферментов и для удаления длинных внутренних открытых рамок считывания (кроме рамок +1) были сделаны нужные замены. Все эти замены были сделаны на участках, в пределах которых предположительно оставалось смещение кодонов по сравнению с составом кодонов кукурузы. Полная оптимизированная по кодонам кукурузы последовательность (кодирующая белок DIG-109) представлена в SEQ ID NO:3. Синтез фрагмента ДНК осуществляли в соответствии с инструкциями поставщика (DNA2.0, Menlo Park, CA).
Пример 5
Конструирование растительных трансформирующих векторов, содержащих экспрессируемые в растении гены, кодирующие белки DIG-109
Для трансформации однодольных растений-хозяев обычно используется супербинарная система Agrobacterium (Japan Tobacco, Tokyo, JP). В такой супербинарной системе используется челночный плазмидный вектор pSBl1, содержащий последовательности повторов правой границы T-ДНК (RB) и повторов левой границы T-ДНК (LB), разделенные сайтами множественного клонирования. Производное pSBl1 (обозначаемое pDAB7691) получали стандартными методами клонирования ДНК. Плазмида pDAB7691 содержит оптимизированную для кукурузы DIG- 109-кодирующую последовательность (CDS; т.е., SEQ ID NO:3), находящуюся под транскрипционным контролем промотора убихитина 1 кукурузы с присоединенным интроном 1 (патент США № 5510474) и с присоединенной 3'-нетранслируемой областью Per5 кукурузы (3' UTR) (патент США № 7179902). Кроме того, pDAB7691 содержит растительный селективный маркерный ген, включающий последовательность CDS DSM2 Dow AgroSciences (WO 2008/070845 A2), находящуюся под транскрипционным контролем промотора актина 1 риса с присоединенным интроном 1 (патент США № 5641876), и с 3'UTR липазы кукурузы (патент США № 7179902). Физическое расположение компонентов Т-области pDAB7691 можно представить как:
RB> промотор Ubil кукурузы:CDS DIG-109:3'UTR Per5 кукурузы > промотор Act1 риса:CDS DSM2:3'UTR липазы кукурузы >LB.
Второе производное pSB11 (обозначаемое pDAB100276) получали стандартными методами клонирования ДНК. Плазмида pDAB100276 содержит оптимизированную по кодонам кукурузы DIG-109-кодирующую последовательность (CDS; то есть, SEQ ID NO:3), находящуюся под транскрипционным контролем промотора убихитина 1 кукурузы с присоединенными интроном 1 и 3'UTR Per5 кукурузы. Кроме того, плазмида pDAB100276 содержит растительный селективный маркерный ген, включающий последовательность CDS AAD1 Dow AgroSciences (заявка на патент США № 20090093366), находящуюся под транскрипционным контролем промотора убихитина 1 кукурузы с присоединенными интроном 1 и 3'UTR липазы кукурузы. Физическое расположение компонентов Т-области pDAB100276 можно представить как:
RB>промотор Ubil кукурузы:CDS DIG-109:3'UTR Per5 кукурузы>промотор Ubil кукурузы:CDS AAD-1:3'UTR липазы кукурузы >LB.
Для трансформации Agrobacterium, клетки клонирующего штамма DH5α Escherichia coli, содержащего плазмиду pDAB7691 или плазмиду pDAB100276, культивировали при 37°С в течение ночи на среде с агаром LB (г/л: триптон Bacto, 10; дрожжевой экстракт Bacto, 5; NaCl, 10; агар, 15), содержали спектиномицин (100 мкг/мл). Клетки штамма DH5a, содержащие конъюгативную мобилизирующую плазмиду pRK2013, культивировали на агаре LB, содержащем канамицин (50 мкг/мл). После инкубирования, планшеты помещали при 4°С для обеспечения доступности штамма LBA4404 Agrobacterium tumefaciens, содержащего плазмиду pSBl.
Пример 6
Трансформация Agrobacterium для получения супербинарных векторов
Супербинарная система Agrobacterium, в которой используется штамм LBA4404 Agrobacterium tumefaciens, содержащий плазмиду pSB1, обычно применяется для трансформации однодольных растений-хозяев. Методика конструирования и подтверждения создания супербинарных векторов хорошо описана в Техническом руководстве для pSB1 (Japan Tobacco). Для генерирования и подтверждения создания супербинарной плазмиды pDAS5162, которая представляет собой ко-интегрированную плазмиду, содержащую плазмиды pSBl и pDAB7691, и супербинарной плазмиды pDAS5848, которая представляет собой ко-интегрированную плазмиду, содержащую плазмиды pSBl и pDAB100276, применяли стандартные методы микробиологии молекулярной биологии.
Пример 7
Продуцирование белка DIG-109 в растенях кукурузы
Agrobacterium -опосредуемая трансформация кукурузы. Семена от кроссов F1 Hi-II (Armstrong et al., 1991) высевали в 5-галлонные горшки, содержащие смесь 95% культуральной среды Metro-Mix 360 без почвы (Sun Gro Horticulture, Bellevue, WA) и 5% почвы с глиной/известью. Растения выращивали в теплице с использованием комбинации натриевых и галогеновых ламп высокого давления в режиме освещения 16 часов день : 8 часов ночь. Для получения незрелых эмбрионов F2, используемых для трансформации, осуществляли контролируемое опыление сибсов. Початки кукурузы собирали приблизительно через 8-10 дней после опыления, когда незрелые эмбрионы имели размер от 1,0 мм до 2,0 мм.
Инфицирование и совместное культивирование. С початков кукурузы снимали листья, и поверхность стерилизовали путем очистки жидким мылом, пропитанным в 20% коммерчески доступном отбеливателе (содержащем 5% гипохлорит натрия), оставляли приблизительно на 20 минут, а затем три раза промывали стерильной водой. Суспензию клеток Agrobacterium tumefaciens, содержащих pDAS5162, супербинарный вектор, включающий ген, кодирующий белок DIG-109 и растительный селективный маркерный ген DSM2, получали путем переноса 1 или 2 петель бактерий [культивированных в течение 2-3 дней при 28°С на твердой среде YEP (г/л: дрожжевой экстракт Bacto, 10; пептон Bacto, 10; NaCl, 5; агар, 15), содержащей 100 мг/л спектиномицина, 10 мг/л тетрациклина и 250 мг/л стрептомицина] в 5 мл жидкой инфекционной среды [базальной среды LS (Linsmaier and Skoog, 1965), витамины N6 (Chu et al., 1975), 1,5 мг/л 2,4-дихлорфеноксиуксусной кислоты (2,4-D), 68,5 г/л сахарозы, 36,0 г/л глюкозы, 6 мМ L-пролина, pH 5.2], содержащей 100 мкΜ ацетосирингона.
Альтернативно, суспензию клеток Agrobacterium tumefaciens, содержащих pDAS5848, супербинарный вектор, включающий ген, кодирующй белок DIG-109 и растительный селективный маркерный ген AAD-1, получали путем переноса 1 или 2 петель бактерий, культивированных как описано выше, в 5 мл жидкой инфекционной среды, содержащей 100-200 мкМ ацетосирингона.
В обоих случаях, раствор встряхивали до получения однородной суспензии, и ее концентрацию доводили до конечной плотности 200 единиц Клетта с помощью колориметра Клетта-Саммерсона с использованием пурпурного фильтра (для трансформации pDAS5162), или до оптической плотности 1,2 на 550 нм (для трансформации pDAS5848). Незрелые эмбрионы выделяли непосредственно в микроцентрифужную пробирку, содержащую 2 мл инфекционной среды. Затем среду удаляли и заменяли 1 мл раствора Agrobacterium, после чего раствор Agrobacterium/эмбриона инкубировали в течение 5 - 10 минут при комнатной температуре. Затем эмбрионы переносили в среду для совместного культивирования [базальная среда LS, витамины N6, 1,5 мг/л 2,4-D, 30,0 г/л сахарозы, 6 мМ L-пролина, 0,85 мг/л AgNO3, 2,8 г/л геллановой камеди (PhytoTechnology Laboratories, Lenexa, KS), pH 5,8], содержащую 100 мкΜ ацетосирингона (для трансформантов pDAS5162) или 100-200 мкΜ ацетосирингона (для трансформантов pDAS5848), и совместно культивировали в течение 3-4 дней при 20°С в темноте.
После совместного культивирования, эмбрионы переносили в регенерирующую среду, содержащую соли MS и витамины, 6 мМ L-пролина, 100 мг/г мио-инозита, 500 мг/г MES, 30 г/л сахарозы, 1,5 мг/г 2,4-D, 0,85 мг/г AgNO3 , 250 мг/г цефотаксима, 2,8 г/л геллановой камеди, pH 5,8. Приблизительно через 7 дней, эмбрионы переносили в ту же самую среду, в которую было добавлено 3 мг/г биалофоса (для трансформантов pDAS5162) или 100 нМ галоксифопа (для трансформантов pDAS5848) (селективная среда). Трансформированные изоляты идентифицировали примерно через 8 недель и объединяли в общую массу путем переноса на свежую селективную среду с 2-недельными интервалами для регенерации и анализа.
Регенерация и продуцирование семян. Для регенерации, культуры переносили в индукционную среду «28» (соли MS и витамины, 30 г/л сахарозы, 5 мг/г бензиламинопурина, 1,5 мг/г 2,4-D, 250 мг/г цефотаксима, 2,8 г/л геллановой камеди, pH 5,7), в которую было добавлено 3 мг/г биалофоса (для трансформантов pDAS5162) или 100 нМ галоксифопа (для трансформантов pDAS5848). Инкубирование проводили в течение 1 недели в условиях слабого освещения (14 мкΕм-2с-1), а затем в течение 1 недели в условиях сильного освещения (приблизительно 89 мкΕм-2с-1). Затем ткани переносили в среду для регенерации «36» (среду, аналогичную индукционной среде, за исключением того, что она не содержала регуляторов роста растения). Когда ростки достигали 3-5 см в длину, растения переносили в стеклянные пробирки для культивирования, содержащие среду SHGA [(Schenk and Hildebrandt (1972), salts and vitamins; PhytoTechnologies Labr.), содержащую 1,0 г/л мио-инозита, 10 г/л сахарозы и 2,0 г/л геллановой камеди, pH 5,8] для последующего культивирования и развития побегов и корней. Растения пересаживали в ту же самую смесь почвы, описанную ранее, и культивировали в теплице до начала цветения. Для получения семян проводили искусственное опыление.
Для специалиста в области трансформации кукурузы очевидно, что для трансформации кукурузы и для отбора трансформированных растений могут быть применены и другие методы в случае использования других экспрессируемых в растении селективных маркерных генов (например, генов толерантности к гербицидам).
Пример 8
Биохимический анализ и биоанализы растений кукурузы, продуцирующих белок DIG-109, на токсичность для насекомых
Продуцирование белка DIG-109 в трансгенных растениях кукурузы оценивали по белкам, экстрагированным из листьев молодых растений (генерация T0). Два диска листьев кукурузы диаметром 6 мм помещали в пробирку для образцов, взятую из глубокого 96-луночного планшета с набором пробирок (Costar Cat# 3957), и замораживали до -80° и хранили до начала анализа. В день анализа, два 4,5-миллилитровых покрытых цинком BB Daisy™ добавляли в каждую (замороженную) пробирку вместе с 200 мкл буфера для экстракции, состоящего из PBS (забуференного фосфатом раствора; Fisher Cat# BP665-1) и 0,05% твина 20. Каждую пробирку закрывали крышкой, и планшет помещали в шаровую мельницу (Kleco™ 4-96 Pulverizer; Garcia Manufacturing, Visalia, CA) максимум на три минуты. Измельченные в порошок образцы центрифугировали в течение 5 минут при 2500 x g, и супернатант, содержащий растворимые белки, использовали в иммуноанализах.
Иммуноблот-анализ белков, экстрагированных из листьев кукурузы, показал, что поликлональное антитело против DIG152RPC1 (полученное у кроликов, иммунизованных активированным трипсином белком, содержащим коровый токсин Cry1Ca) не вступает в перекрестную реакцию с белками, экстрагированными из листьев нетрансгенных растений. В экстрактах растений, трансформированных pDAS5162, белки некоторых видов были детектированы с использованием антитела против DIG152PRC1.
Очевидно, что несмотря на то, что трансген, введенный в кукурузу путем трансформации pDAS5162, будет кодировать полноразмерный белок DIG-109, однако, протеолитическая активность клеток кукурузы будет приводить к росту цепи белка с образованием избыточного количества стабильных молекул с меньшей молекулярной массой.
Листья, собранные от независимо полученных трансгенных растений кукурузы, трансформированных конструкцией pDAS5162, тестировали in vitro на токсичность для насекомых с использованием только что вылупившихся личинок совки травяной (FAW, Spodoptera frugiperda (J.E. Smith)) и Cry1F-резистентной FAW (rFAW). Яйца FAW были получены из коммерчески доступного инсектария (Benzob), а яйца rFAW были получены от не-промышленных популяций (Dow AgroSciences). Образцы сегментов листьев были взяты для биоанализов выращенных в теплице растений Т0 на токсичность для насекомых, и приблизительно через две недели, растения переносили из лаборатории в теплицу. Для кусочка листьев от каждого растения (приблизительно 1 кв.дюйм каждого) помещали в отдельные лунки 32-луночного планшета (CD International), поверхность которого была покрыта 3 мл отвержденного 2% агара. Из яиц вылуплялись личинки, которым давали корм, подходящий для чешуекрылых многих видов (Southland Products), и менее чем через 24 часа личинки отбирали. Приблизительно 10 личинок на сегмент листьев осторожно помещали в каждую лунку щеткой из верблюжьей шерсти. Инфицированные планшеты герметично закрывали перфорированными крышками, поставляемыми вместе с планшетами, а затем хранили при 28°С, при относительной влажности (ОВ) 40%, в режиме освещения 16 ч : 8 ч (день:ночь)] в течение трех дней. Процент поражения (% DAM) каждого кусочка листа регистрировали по окончании теста. Степень поражения усредняли и использовали для того, чтобы определить, какие растения имели наименьшее поражение тестируемыми насекомыми каждого типа. Тесты повторяли несколько раз для всех насекомых.
Данные анализировали с использованием статистической компьютерной программы JMP (SAS, Cary, NC) путем усреднения величин % DAM для каждого растения и для каждого типа насекомого. Для одностороннего анализа ANOVA использовали модель «Fit Y х X». Если это необходимо, то для оценки значимого различия средних величин %DAM для каждой обработки применяли метод разделения средних Тьюки-Крамера. Сравнение проводили для величин %DAM, полученных от контрольных растения одного и того же возраста. Растения позитивного контроля выращивали из семян коммерчески доступного гибрида Herculex I™, продуцирующего токсин Cry1Fa B.t. Негативный контроль (то есть, нетрансформированные растения) был представлен линиями Hi II и B104, и изолинией Herculex I™ (родитель-гибрид Herculex I™, не содержащий Cry).
На фигуре 1 систематизированы результаты, полученные в таких тестах с помощью биоанализа насекомых. Была неожиданно обнаружена позитивная корреляция между продуцированием DIG-109 в листьях трансгенных растений и %DAM. Для FAW, F=35,3; d.f. (степень свободы) =1,33; P<0,0001; r2=0,52, а для rFAW, F=25,3; d.f.=1,33; P<0,0001; r2=0,43. Было также неожиданно и впервые обнаружено, что личинки совки травяной, которые являются резистентными к интоксикации токсином Cry1Fa B.t., уже погибали после поедания ими листьев с токсином DIG-109 B.t..
Следует отметить, что другие насекомые-вредители кукурузы могут быть протестированы аналогичным образом. Такими вредителями являются, но не ограничиваются ими: Agromyza parvicornis (кукурузная минирующая муха), Agrotis ipsilon (совка-ипсилон), Anticarsia gemmatalis (совка фасолевая), Diatraea grandiosella (американский кукурузный пилильщик), Diatraea saccharalis (пилильщик свекловичный), Elasmopalpus lignosellus (точильщик зерновой), Helicoverpa zea (совка кукурузная), Heliothis virescens (совка табачная), Ostrinia nubilalis (европейский кукурузный пилильщик), Cry1F-резистентные O. nubilalis, Plutella xylostella (моль капустная), Cry1-резистентная P. xylostella, Spodoptera exigua (совка свекловичная) и Trichoplusia ni (пяденица капустная).
Трансгенные растения кукурузы, трансформированные pDAS5848 (генерация T0), также подвергали биоанализу на резистентность к насекомым и иммуноанализам. Количество белка DIG-109 в экстрактах листьев количественно оценивали с использованием коммерчески доступного набора для детектирования Cry1C с помощью ELISA-анализа (Envirologix™, Portland, MA; Cat# AP007), и уровень детектированного белка DIG-109 выражали в миллионных долях (м.д.; 1 м.д. означает 1 нг белка DIG-109 на мг общего растворимого белка в экстракте).
Степень поражения вредителями FAW и rFAW оценивали по следующим баллам: 0 = отсутствие поражения или несколько заметных дырок, 1= 25-50% поражение листьев, и 2 = съеден почти весь лист или съедена левая часть листа. Защищенным от вредителей растением является растение, степень поражения которого составляет 0,67 или ниже.
Данные, представленные в таблице 3, указывали на позитивную корреляцию между присутствием белка DIG-109, детектированного с помощью ELISA в растениях T0, и снижением уровня поражения личинкой совки травяной в in vitro биоанализах. Растения с самым высоким детектированным уровнем белка DIG-109 (растение 5848-005.4) имели самую низшую оценку степени поедания листьев. Листья от растений с более низкими уровнями детектированного белка DIG-109 в пределах концентраций 190-230 м.д. были также меньше поражены, чем листья растений негативного контроля (то есть, нетрансформированного контроля B104 и Hi II), которые имели средние оценки поражения 1,7 и 1,8. Во всех оцененных листьях с pDAS5848 преобладал детектированный белок типа DIG-109, который содержал дублет из пептидов размером приблизительно 60 кДа и 55 кДа.
Таблица 3. Уровни белка DIG-109 в экстрактах pDAS5848-трансформированных листьев трансгенной кукурузы и снижение степени поражения личинками совки травяной
|
|
Таким образом, отличительным признаком настоящего изобретения является то, что белок DIG-109, при его продуцировании в растениях кукурузы, сообщает растениям резистентность к поражению личинками совки травяной и личинками Cry1F-резистентной совки травяной.
Пример 9
Эксперименты по конкурентому связыванию белков, содержащих коровые токсины Cry1Fa и Cry1Ca, с выделенными мембранными везикулами щеточной каемки Spodoptera frugiperda
В нижеследующих примерах описана оценка конкурентного связывания белков, содержащих коровый токсин Cry1, с предполагаемыми рецепторами в ткани кишечника насекомого. Было показано, что 125I-меченный белок, содержащий коровый токсин Cry1Ca, связывается с высокой аффинностью с мембранными везикулами щеточной каемки (BBMV), полученными от Spodoptera frugiperda (совки травяной), и что белок, содержащий коровый токсин Cry1Fa, не конкурирует за связывание с этими везикулами.
Очистка белков Cry. Ген, кодирующий химерный белок, содержащий коровый токсин Cry1Ca и протоксин Cry1Ab, экспрессировали в экспрессионном штамме Pseudomonas fluorescens как описано в примере 2. Аналогичным образом, ген, кодирующий химерный белок, содержащий коровый токсин Cry1Fa (603 аминокислоты) и протоксин Cry1Ab (545 аминокислот), экспрессировали в системе Pf. Белки очищали методами, описанными в примере 2, и осуществляли гидролиз трипсином с получением активированных коровых токсинов из полноразмерных белков, а затем эти продукты очищали методами, описанными в примере 2. Препараты обработанных трипсином белков (содержащих активированный коровый токсин) имели чистоту >95%, а их молекулярная масса составляла приблизительно 65 кДа как было экспериментально определено с помощью электрофореза в ДСН-ПААГ.
Стандартные методы количественной оценки белка и электрофореза в полиакриламидном геле с ДСН проводили как описано, например, в руководстве Sambrook et al. (1989) и Ausubel et al. (1995), и в более поздних изданиях.
Получение и фракицонирование солюбилизированных BBMV. Личинки Spodoptera frugiperda в последней возрастной стадии выдерживали в условиях голодания в течение ночи, а затем, после охлаждения на льду в течение 15 минут, вскрывали. Ткань средней части кишечника удаляли из полости тела, а заднюю часть кишечника оставляли присоединенной к покровному слою. Среднюю часть кишечника помещали в 9 х объем охлажденного льдом гомогенизирующего буфера (300 мМ маннит, 5 мМ EGTA, 17 мМ основания трис, pH 7,5), в который была добавлена смесь ингибиторов протеазы (Sigma-Aldrich P-2714), разведенная в соответствии с рекомендации поставщиков. Ткань гомогенизировали 15-ю импульсами, подаваемыми стеклянным гомогенизатором ткани. BBMV получали методом MgCl2-преципитации, описанным Wolfersberger (1993). Вкратце, равный объем 24 мМ раствора MgCl2 в 300 мМ маннита смешивали с гомогенатом, выделенным из средней части кишки, перемешивали в течение 5 минут и оставляли на льду на 15 минут. Раствор центрифугировали при 2500 x g в течение 15 минут при 40C. Супернатант сохраняли, и осадок суспендировали в исходном объеме 0,5 х разведенного гомогенизирующего буфера, а затем снова центрифугировали. Два супернатанта объединяли и центрифугировали при 27000 x g в течение 30 минут при 4°С с получением фракции BBMV. Осадок суспендировали в буфере для хранения BBMV (10 мМ HEPES, 130 мМ KCl, 10% глицерин, pH 7,4) до получения концентрации белка примерно 3 мг/мл. Концентрацию белка определяли с использованием альбумина бычьей сыворотки (BSA) в качестве стандарта. Определение уровня щелочной фосфатазы (фермента-маркера для фракции BBMV) проводили до замораживания образцов с использованием набора для анализа щелочной фосфатазы QuantiChrom™ DALP-250 (Gentaur Molecular Products, Kampenhout, BE) в соответствии с инструкциями производителей. Удельная активность этого фермента обычно возрастала в 7 раз по сравнению с активностью, обнаруживаемой в исходной фракции гомогената средней части кишки. BBMV разделяли на образцы-аликвоты по 250 мкл, быстро замораживали в жидком азоте и хранили при -80°С.
Электрофорез. Анализ белков с помощью электрофореза в ДСН-ПААГ проводили в восстанавливающих условиях (то есть, в 5% β-меркаптоэтаноле, BME) и в денатурирующих условиях (то есть, при нагревании в течение 5 минут, при 90°С в присутствии 2% ДСН). Белки загружали на лунки с 4-20% трис-глициновым полиакриламидным гелем (BioRad; Hercules, CA) и разделяли под напряжением 200 вольт в течение 60 минут. Полосы белка детектировали путем окрашивания кумасси бриллиантовым голубым R-250 (BioRad) в течение одного часа, и обесцвечивали раствором 5% метанола в 7% уксусной кислоте. Гели визуализировали и анализировали на визуализаторе BioRad Fluro-S Multi Imager™. Относительные молекулярные массы полос белка определяли путем сравнения с подвижностью белков с известной молекулярной массой, наблюдаемых в образце лэддера белка BenchMark™ (Life Technologies, Rockville, MD), загруженного на одну лунку геля.
Иодирование белка, содержащего коровый токсин Cry1Ca. Очищенный белок, содержащий коровый токсин Cry1Ca, подвергали реакции иодирования с использованием иодирующих сфер Pierce Iodination Beads (Thermo Fisher Scientific, Rockford, IL). Вкратце, две иодирующих сферы два раза промывали 500 мкл PBS (20 мМ фосфат натрия, 0,15 M NaCl, pH7,5), и помещали в центрифужную 1,5 мл-пробирку, содержащую 100 мкл PBS. Затем добавляли 0,5 мКи 125I-меченного иодида натрия, после чего компоненты оставляли для реакции на 5 минут при комнатной температуре, а затем к раствору добавляли 1 мкг белка, содержащего коровый токсин Cry1Ca, и смесь оставляли для реакции еще на 3-5 минут. Реакцию завершали путем пипетирования раствора из иодирующих сфер и наносили на центрифужную колонку Zeba™ (Invitrogen), уравновешенную в 50 мМ CAPS, pH 10,0, 1 мМ DTT (дитиотреитол), 1 мМ EDTA и 5% глицерине. Иодирующие сферы два раза промывали 10 мкл PBS и промывочный раствор также наносили на обессоливающую колонку Zeba™. Радиоактивный раствор элюировали с центрифужной колонки путем центрифугирования при 1000 x g в течение 2 минут. Затем, белок, содержащий коровый токсин Cry1Ca и меченный радиоактивным 125I, диализовали против 50 мМ CAPS, pH 10,0, 1 мМ DTT, 1 мМ EDTA и 5% глицерина.
Визуализация. Нерадиоактивный иодированный белок, содержащий коровый токсин Cry1Ca, определяли с помощью электрофореза в ДСН-ПААГ и визуализации флуоресцентным методом. Вкратце, ДСН-ПААГ-гели сушили с использованием аппарата для сушки геля BioRad в соответствии с инструкциями производителей. Осушенные гели визуализировали путем их заворачивания в пленку Mylar (толщиной в 12 мкм) и экспонирования под флуоресцирующим экраном с накоплением Molecular Dynamics (35 см x 43 см) в течение 1 часа. Планшеты проявляли с помощью флуоресцентного визуализатора Molecular Dynamics Storm 820, и изображение анализировали с помощью компьютерной программы ImageQuant™.
Пример 10
Связывание 125 I-меченного белка, содержащего коровый токсин Cry1, с BBMV от Spodoptera frugiperda
В целях определения оптимального количества белка BBMV для его использования в анализах на связывание с белками, содержащими коровые токсины Cry1Ca и Cry1Fa, строили кривую насыщения с использованием 125I-меченного белка, содержащего коровый токсин Cry1Ac. 0,5 нМ 125I-меченного белка, содержащего коровый токсин Cry1Ac, инкубировали в течение 1 часа при 28°С в буфере для связывания (8 мМ NaHPO4, 2 мМ KH2PO4, 150 мМ NaCl, 0,1% BSA, pH7,4) с белком BBMV в количестве, составляющем от 0 мкг/мл до 500 мкг/мл (общий объем 0,5 мл). 125I-меченный белок, содержащий коровый токсин Cry1Ac и связанный с белками BBMV, отделяли от несвязанной фракции путем взятия образцов 150 мкл реакционной смеси с тремя повторностями, которые помещали в отдельные центрифужные 1,5 мл-пробирки и центрифугировали при 14000 x g в течение 8 минут при комнатной температуре. Супернатант осторожно удаляли, и осадок три раза промывали охлажденным льдом буфером для связывания. Дно центрифужной пробирки, содержащей осадок, отрезали, помещали в стеклянную пробирку для культивирования размером 13 x 75 мм, и каждый образец подсчитывали в течение 5 минут в гамма-счетчике. Затем строили график зависимости CPM (число импульсов в минуту) минус фоновый CPM (в отсутствии реакции с белком BBMV) от концентрации белка BBMV. Кроме того, в соответствии с результатами, полученными в другом анализе, оптимальная концентрация белка BBMV, используемого в анализах на связывание, составляла 150 мкг/мл.
Пример 11
Анализы на конкурентное связывание BBMV от S. frugiperda с белками, содержащими коровые токсины Cry1Ca и Cry1Fa
Анализы на гомологичное и гетерологичное конкурентное связывание проводили с использованием 150 мкг/мл белка BBMV и 0,5 нМ 125I-меченного белка, содержащего коровый токсин Cry1Ca. К реакционной смеси добавляли конкурентный нерадиоактивный белок, содержащий коровый токсин Cry1Fa в концентрациях от 0,045 нМ до 1000 нМ и одновременно добавляли радиоактивный белок, содержащий коровый токсин Cry1Ca, для гарантии истинного конкурентного связывания. Инкубирование проводили в течение 1 часа при 28°С и измеряли количество 125I-меченного белка, содержащего коровый токсин Cry1Ca и связанного с BBMV (специфически связанного) как описано выше. Неспецифическое связывание выражали как величину, полученную в присутствии 1000 нМ нерадиоактивного белка, содержащего коровый токсин Cry1Ca. 100%-ное общее связывание рассматривается как количество связывания в отсутствии какого-либо конкурентно связанного белка, содержащего коровый токсин Cry1Fa.
В анализах на связывание с рецептором, проводимых с использованием 125I-меченного белка, содержащего коровый токсин Cry1Ca, определяли способность белка, содержащего коровый токсин Cry1Fa, вытеснять этот радиоактивно меченный лиганд из его сайта связывания с BBMV от S. frugiperda. Результаты (фигура 2) показали, что белок, содержащий коровый токсин Cry1Fa, не вытеснял связанный 125I-меченный белок, содержащий коровый токсин Cry1Ca, из его рецепторного(ых) белка(ов) при концентрациях до 300 нМ (при концентрации, которая в 600 раз превышала концентрацию радиоактивного связывающего лиганда). Как и предполагалось, немеченный белок, содержащий коровый токсин Cry1Ca, обладал способностью вытеснять радиоактивно меченный белок, содержащий коровый токсин Cry1Ca, из его связывающегося(ихся) белка(ов), на что указывала сигмоидальная кривая доза-ответ, где 50%-ное вытеснение происходило при 5 нМ.
Таким образом, было показано, что белок, содержащий коровый токсин Cry1Ca взаимодействует с сайтом связывания BBMV S. frugiperda, который не связывается с белком, содержащим коровый токсин Cry1Fa.
См. фигуру 2: Конкуренция корового токсина Cry1Fa, корового токсина Cry1Ca и 125I-меченного корового токсина Cry1Ca за связывание с BBMV Spodoptera frugiperda.
Пример 12
Конкурентное связывание Белка, содержащего коровый токсин Cry1Ca, и меченного биотином белка, содержащего коровый токсин Cry1Fa, с BBMV Diatraea saccharalis
В нижеследующих примерах описана оценка конкурентного связывания белков, содержащих коровый токсин Cry1, с предполагаемыми рецепторами в ткани кишечника насекомого. Было показано, что меченный биотином белок, содержащий коровый токсин Cry1Fa связывается с высокой аффинностью с мембранными везикулами щеточной каемки (BBMV), полученными от Diatraea saccharalis (свекловичного пилильщика), и что белок, содержащий коровый токсин Cry1Ca не конкурирует за связывание с этими везикулами.
Получение и фракционирование солюбилизированных BBMV. Личинки D. saccharalis в последней возрастной стадии выдерживали в условиях голодания в течение ночи, а затем, после охлаждения на льду в течение 15 минут, вскрывали. Препараты BBMV получали методами, описанными в примере 10.
Результаты, полученные в анализе на конкурентное связывание, проводимом с использованием 125I-меченного белка, содержащего коровый токсин Cry1Fa, могут иметь ограниченную биологическую значимость, поскольку иодирование белка Cry1Fa делает этот белок неактивным в биологических анализах на поедание такого белка насекомыми. В противоположность этому, было обнаружено, что биотинилированный белок, содержащий коровый токсин Cry1Fa, сохранял свою токсичность по отношению к насекомым. Кроме того, можно измерить уровень взаимодействия этого (биотинилированого) белка с рецепторами в присутствии не-биотинилированных (конкурирующих) белков, содержащих коровый токсин Cry. Такой эксперимент по конкурентному связыванию позволяет детектировать связывание меченного биотином белка, содержащего коровый токсин Cry1Fa с рецепторами в BBMV D. saccharalis после электрофореза белков BBMV и переноса всего образца в ПВДФ-мембрану (поливинилиденфторидную мембрану). Авидин, конъюгированный с пероксидазой хрена, в комбинации с реагентом, усиливающим химическую люминесценцию, использовали для визуализации меченного биотином белка, содержащего коровый токсин Cry1Fa.
Белок, содержащий коровый токсин Cry1Fa метили биотином с использованием набора для биотинилирования сульфо-NHS-LC Pierce EZ-Link® (Thermo Fisher Scientific). Вкратце, 40 мкл сульфо-NHS-LC-биотина (10 мг/мл в диметилсульфоксиде) добавляли к 500 мкл белка, содержащего коровый токсин Cry1Fa (2,0 мг/мл), в 0,1M натрийфосфатного буфера, pH7,2. Реакционную смесь инкубировали при 4°С в течение ночи, а затем непрореагировавший сульфо-NHS-LC-биотин удаляли на обессоливающей колонке Zeba™. Уровень включения биотина в белок, содержащий коровый токсин Cry1Fa, измеряли с помощью анализа на вытеснение НАВА-авидина Pierce (Thermo Fisher Scientific) как описано производителем.
Меченный биотином белок, содержащий коровый токсин Cry1Fa (2,5 нМ), инкубировали в течение 1 часа при 28°С с 0,2 мг BBMV, полученного от D. saccharalis [в общем объеме 1,0 мл] в присутствии или в отсутствии 500-кратного избытка немеченного белка, содержащего коровый токсин Cry1Fa или Cry1Ca. Несвязанный меченный биотином белок, содержащий коровый токсин Cry1Fa удаляли путем центрифугирования при 16000 x g в течение 10 минут, и полученный осадок 3 раза промывали охлажденным льдом связывающим буфером. Осадок суспендировали в 15 мкл 4 х буфера для образцов Лэммли, встряхивали и обрабатывали ультразвуком для гарантии полной солюбилизации, а затем нагревали до 90°С в течение 3 минут. Весь образец загружали на 4%-20% трис-глициновый гель, разделяли с помощью электрофореза в ДСН-ПААГ, и подвергали электропереносу на ПВДФ-мембрану в соответствии с инструкциями поставщиков (BioRad). 1000 нг белка, содержащего коровый токсин Cry1Fa и коровый токсин Cry1Ca, также подвергали электрофорезу на геле и использовали в качестве негативного контроля. Меченный биотином белок, содержащий коровый токсин Cry1Fa, визуализировали с использованием конъюгированной с авидином пероксидазы хрена при разведении 1:15000 с активацией хемилюминисценции с использованием 1:1-смеси пероксидного раствора SuperSignal® West Pico с люминальным раствором-усилителем (Thermo Fisher Scientific, номера по каталогу 1859674 и 1859675). Полосы регистрировали на мультивизуализаторе Biorad Fluor-S с помощью компьютерной программы Quantity One v.4.5.2.
Результаты указывали на детектирование меченного биотином белка, содержащего коровый токсин Cry1Fa и связанного с рецепторами в BBMV D. saccharalis, а также показали, что небиотинилированный белок, содержащий коровый токсин Cry1Fa, при 500-кратной избыточной концентрации полностью вытеснял связанный биотинилированный белок, содержащий коровый токсин Cry1Fa, из его рецептора(ов). В противоположность этому, белок, содержащий коровый токсин Cry1Ca при 500-кратной избыточной концентрации не обладал способностью вытеснять связанный биотинилированный белок, содержащий коровый токсин Cry1Fa, что указывает на то, что белок, содержащий коровый токсин Cry1Ca ,не конкурирует за сайт(ы) связывания белка, содержащего коровый токсин Cry1Fa, в BBMV D. saccharalis. Также было показано, что в соответствии с результатами, полученными для BBMV S. frugiperda, белок, содержащий коровый токсин Cry1Fa, и белок, содержащий коровый токсин Cry1Ca, связывались с отдельными сайтами связывания BBMV D. saccharalis.
Библиография
Finney, D.J. 1971. Probit analysis. Cambridge University Press, England.
Hua, G., L. Masson, J. L. Jurat-Fuentes, G. Schwab, and M. J. Adang. Binding analyses of Bacillus thuringiensis Cry d-endotoxins using brush border membrane vesicles of Ostrinia nubilalis. Applied and Environmental Microbiology 67[2], 872-879. 2001.
LeOra Software. 1987. POLO-PC. A user's guide to probit and logit analysis. Berkeley, CA.
McGaughey, W. H. , F. Gould, and W. Gelernter. Bt resistance management. Nature Biotechnology 16[2], 144-146. 1998
Marcon, P.R.G.C., L.J. Young, K. Steffey, and B.D. Siegfried. 1999. Baseline susceptibility of the European corn borer, Ostrinia nubilalis (Hubner) (Lepidoptera: Pyralidae) to Bacillus thuringiensis toxins. J. Econ. Entomol. 92 (2): 280-285.
Robertson, L.J. and H.K. Preisler. 1992. Pesticide bioassays with arthropods. CRC Press, Boca Ranton, FL.
SAS Institute Inc. 1988. SAS procedures guide, Release 6.03 edition. SAS Institute Inc, Cary, NC.
Stone, B.F. 1968. A formula for determining degree of dominance in cases of monofactorial inheritance of resistance to chemicals. Bull. WHO 38:325-329.
Van Mellaert, H., J. Botterman, J. Van Rie, and H. Joos. Transgenic plants for the prevention of development of insects resistant to Bacillus thuringiensis toxins. (Plant Genetic Systems N.V., Belg. 89-401499[400246], 57-19901205. EP. 5-31-1989.
Приложение А
Список дельта-эндотоксинов - Crickmore et al. web-сайт (указанный в заявке)
Регистрационный номер для доступа в NCBI


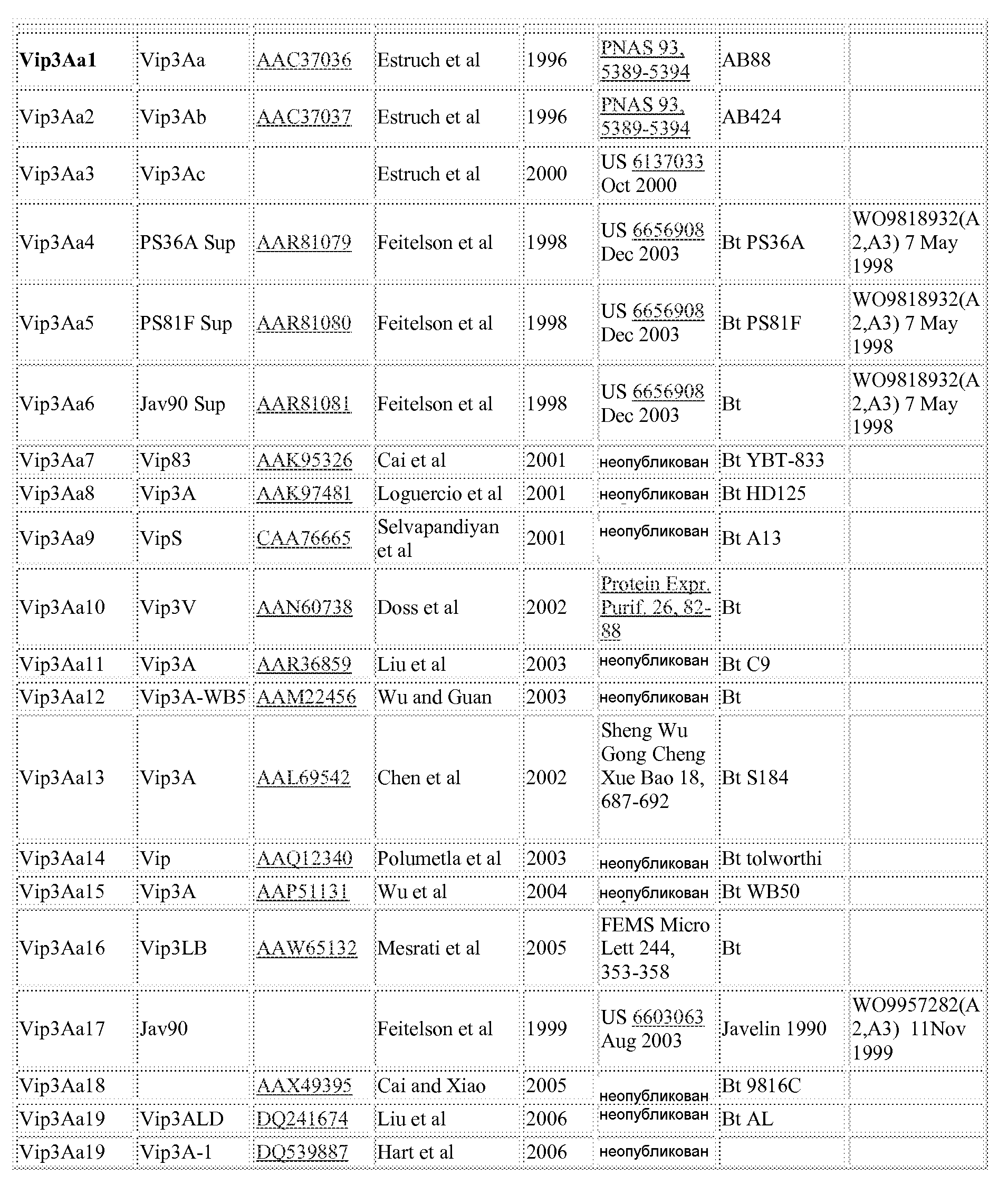















Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
5-фторпиримидиновые производные в качестве фунгицидов
Пестицидные композиции
Высококонцентрированные препараты 1-метилгептилового эфира флуроксипира, стабильные при низких температурах
Пестициды, пестицидная композиция и способ контроля вредителей
Синергическая гербицидная композиция, содержащая хлорацетанилиды и пиколиновые кислоты
Подсолнечник с низким содержанием насыщенных жиров и соответствующие способы
Способы переноса молекулярных веществ в клетки растений
Стабилизированные эмульсии масло-в-воде, содержащие агрономически активные ингредиенты, и способы их применения
Пестицидные композиции
Защита от повреждения гербицидом 6-(трехзамещенный фенил)-4-амино-2-пиридинкарбоксилата посеянного семенами и рассадного риса-сырца
Стабильная пестицидная композиция на основе сульфоксимина и способ борьбы с насекомыми
5-фторпиримидиновые производные в качестве фунгицидов

