Результат интеллектуальной деятельности: СПОСОБ ОЦЕНКИ ЭФФЕКТИВНОСТИ ЗАЩИТЫ ЛИМФОЦИТОВ ОТ АПОПТОЗА
Вид РИД
Изобретение
Известны способы оценки эффективности защиты клеток от апоптоза при активации антиоксидантной активности путем определения роста общего содержания SH-групп белков [Microtubule dynamics and glutathione metabolism in phagocytizing human polymorphonuclear leukocytes / B.R. Burchill, J.M. Oliver, С.B. Pearson et al. // J. of Cell Biology. - 1978. - Vol. 76, №2. - P. 439-447], но данная методика не позволяет оценить уровень восстановленного глутатиона, а следовательно, оценить эффективность защиты клеток от апоптоза. Известен также способ косвенного определения содержания восстановленного глутатиона по активности глутатионпероксидазы [Медицинские лабораторные технологии: В 2-х томах. / Под ред. А.И. Карпищенко - Т. 2 / А.И. Карпищенко. - СПб.: Интермедика. - 1999. - 656 с.], однако, активность этого фермента зависит от конформации активного центра фермента. При изменении конформации активного центра фермента его активность меняется, что не позволяет с высокой точностью оценить уровень восстановленного глутатиона, а, следовательно, оценить эффективность защиты клеток от апоптоза. Известен также способ косвенного определения концентрации восстановленного глутатиона по активности глутатионредуктазы [Медицинские лабораторные технологии: В 2-х томах. / Под ред. А.И. Карпищенко - Т. 2 / А.И. Карпищенко. - СПб.: Интермедика. - 1999. - 656 с], однако активность этого фермента зависит от конформации активного центра фермента. При изменении конформации активного центра фермента его активность меняется, что не позволяет достоверно в любой ситуации оценить с высокой точностью уровень восстановленного глутатиона, а следовательно, оценить эффективность защиты клеток от апоптоза.
Известен также способ определения антиоксидантной активности как защиты клеток от апоптоза по содержанию восстановленного глутатиона, предложенный М.Е. Anderson (1985) в модификации S. Kojima et al. (2004) [Low dose gamma-rays activate immune functions via induction of glutathione and delay tumor growth / S. Kojima, K. Nakayama, H. Ishida // J. Radiat. Res. - 2004. - Vol. 45, №1. - P. 33-39], основаный на взаимодействии восстановленного глутатиона (GSH) с 5,5′-дитио-бис(2-нитробензойной) кислотой (ДТНБ). При этом образуется окисленный глутатион (GSSG), который затем восстанавливается и вновь взаимодействует с ДТНБ. Данный способ является наиболее близким к предлагаемому по технической сущности и достигаемому результату, и выбран в качестве прототипа.
Целью предлагаемого изобретения является повышение эффективности и точности способа.
Указанная цель достигается тем, что дополнительно в инкубационную среду, содержащую лимфоциты, вводится 1,4-дитиоэритритол и аскорбиновая кислота в конечной концентрации 3,0 ммоль и 0,1 ммоль соответственно без которых невозможна последующая защита клеток от апоптоза
Следовательно, только комплексная модернизация способа-прототипа позволяет получить желаемый результат. Только комплексное внесение в инкубационную смесь 1,4-дитиоэритритола и аскорбиновой кислоты в минимальной концентрации 3.0 мМ и 0,1 мМ соответственно позволяет повысить оценку эффективности защиты лимфоцитов от апоптоза, а именно при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту низкоэффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту высокоэффективной.
Антиоксидантная система направлена на эффективную нейтрализацию гидроксирадикалов и снижение токсичной для организма гидроперекиси. Гидроксильный радикал (•OH) участвует в микробицидном и цитотоксическом действии нейтрофилов, моноцитов и Т-лимфоцитов [Окислительный стресс. Прооксиданты и антиоксиданты / Е.Б. Меньщикова, В.З. Панкин, Н.К. Зенков и соавт. - М.: Слово. - 2006. - 556 с.]. Есть два основных механизма синтеза •OH нейтрофилами: первый - образование из пероксида водорода в присутствии металлов переменной валентности в так называемой «реакции Фентона»: Fe2++Н2О2→Fe3++•ОН+ОН- [Лущак В.И. Окислительный стресс и механизмы защиты от него у бактерий / В.И. Лущак // Биохимия. - 2001. - Т. 66, вып. 5. - С. 592-609], второй - в ходе ряда реакций с участием гипогалоидов 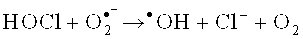 или взаимодействие гипохлорита с ионами Fe2+ [Spin trapping evidence for myeloperoxidase-dependent hydroxyl radical formation by human neutrophils and monocytes // C.L. Ramos, S. Pou, В.E. Britigan et al. // J. Biol. Chem - 1992. - Vol. 267. - P. 8307-8312; Candeias, L.P. Formation of hydroxyl radicals on reaction of hypochlorous acid with ferrocyanide, a model iron (II) complex / L.P. Candeias, M.R.L. Stratford, P. Wardman // Free Radical Res. - 1994. - Vol. 20. - P. 241-249.]. Следовательно, если в клетке нет металлов переменной валентности в свободном состоянии, для нее реакция Фентона и подобные реакции не будут опасными [Hampton М.В. Inside the Neutrophil Phagosome: Oxidants, Myeloperoxidase, and Bacterial Killing / M.B. Hampton, A.J. Kettle, С.C. Winterbourn // Blood. - 1998. - Vol. 92, №9. - P. 3007-3017].
или взаимодействие гипохлорита с ионами Fe2+ [Spin trapping evidence for myeloperoxidase-dependent hydroxyl radical formation by human neutrophils and monocytes // C.L. Ramos, S. Pou, В.E. Britigan et al. // J. Biol. Chem - 1992. - Vol. 267. - P. 8307-8312; Candeias, L.P. Formation of hydroxyl radicals on reaction of hypochlorous acid with ferrocyanide, a model iron (II) complex / L.P. Candeias, M.R.L. Stratford, P. Wardman // Free Radical Res. - 1994. - Vol. 20. - P. 241-249.]. Следовательно, если в клетке нет металлов переменной валентности в свободном состоянии, для нее реакция Фентона и подобные реакции не будут опасными [Hampton М.В. Inside the Neutrophil Phagosome: Oxidants, Myeloperoxidase, and Bacterial Killing / M.B. Hampton, A.J. Kettle, С.C. Winterbourn // Blood. - 1998. - Vol. 92, №9. - P. 3007-3017].
В клетках предусмотрен ряд защитных реакций по блокированию свободных ионов Fe2+ и Cu+. Например, активированные клетки синтезируют лактоферрин, связывающий свободное железо и переводящий его в каталитически неактивную форму, а также продуцируют высокие концентрации таурина, конъюгирующего с гипохлоритом и защищающего клетку от его токсичных эффектов [Taurine chloramines, a product of activated netrophils, inhibits in vitro the genetation of nitric oxide and other macrofage inflammatory mediators / J. Marcinkiewiez, A. Grabowska, J. Bereta et al. // J. Leukocyte Biol. - 1995. - Vol. 58. - P. 667-674].
Образование •ОН показано в ходе микросомального окисления, окисления арахидоновой кислоты, в реакциях с флавиновыми ферментами, убихиноном, пероксинитритом. В ряде исследований клеток in vitro получены свидетельства продукции •ОН данными клетками [Hydroxylation of salicylate by activated neutrophils / W.B. Davis, B.S. Mohammed, D.C. Mays et al. // Biochem Pharmacol. - 1989. - Vol. 38. - P. 4013-4019]. Однако изучение этих реакций зачастую основывалось на использовании ингибиторов и измерении уровня вторичных продуктов. Следовательно, реакции, приписанные •ОН, могли быть вызваны другими оксидантами, в частности 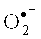 или гипохлорной кислотой [Do human neutrophils form hydroxyl radical. Evaluation of an unresolved controversy / M.S. Cohen, В.E. Britigan, D.J. Hassett et al. // Free Radic Biol Med. - 1988. - Vol. 5. - P. 81-90]. Ряд авторов считает, что in vitro за счет реакции Фентона клетки производят незначительные количества •ОН [Rosen, G.М. Free radicals and phagocytic cells / G.M. Rosen, S. Pou, C.L. Ramos // FASEB J. - 1995. - Vol. 9. - P. 200-211].
или гипохлорной кислотой [Do human neutrophils form hydroxyl radical. Evaluation of an unresolved controversy / M.S. Cohen, В.E. Britigan, D.J. Hassett et al. // Free Radic Biol Med. - 1988. - Vol. 5. - P. 81-90]. Ряд авторов считает, что in vitro за счет реакции Фентона клетки производят незначительные количества •ОН [Rosen, G.М. Free radicals and phagocytic cells / G.M. Rosen, S. Pou, C.L. Ramos // FASEB J. - 1995. - Vol. 9. - P. 200-211].
Генерация •ОН стимулированными клетками в очаге воспаления может существенно лимитироваться отсутствием в среде ионов железа. Исследование реакции Фентона в клетках выявило, что блокирование ионов железа лактоферрином ингибирует непосредственно саму реакцию [Rosen, G.М. Free radicals and phagocytic cells / G.M. Rosen, S. Pou, C.L. Ramos // FASEB J. - 1995. - Vol. 9. - P. 200-211], а утилизация H2O2 миелопероксидазой ограничивает реакцию, даже если железо доступно [Winterbourn, С.С. Myeloperoxidase as an effective inhibitor of hydroxyl radical production: Implications for the oxidative reactions of neutrophils / С.C. Winterbourn // J. Clin. Invest. - 1986. - Vol. 78. - P. 545-557]. Хотя большинство биологических форм железа каталитически неактивно, показана способность клеток к продукции •ОН в присутствии трансферрина, подверженного протеолитической деградации [Phagocyte-derived free radicals stimulated by ingestion of ironrich Staphylococcus aureus: Aspin-trapping study / M.S. Cohen, В.E. Britigan, Y.S. Chai et al. // J. Infect Dis. - 1991. - Vol. 163. - 819-826], или железа в составе Pseudomonas aeruginosa, содержащего сидерофор пиохелин [Possible role of bacterial siderophores in inflammation-Iron bound to the pseudomonas siderophore pyochelin can function as a hydroxyl radical catalyst / T.J. Coffman, C.D. Cox, B.L. Edeker et al. / J. Clin. Invest. - 1990. - Vol. 86. - P. 1030-1038]. Однако M.S. Cohen и соавторы обнаружили, что внутриклеточное железо не всегда доступно: в их экспериментах повышенного образования радикала •ОН не отмечалось, даже если клетки поглощали Staphylococcus aureus, который был преинкубирован с Fe2+ [Phagocyte-derived free radicals stimulated by ingestion of ironrich Staphylococcus aureus: Aspin-trapping study / M.S. Cohen, В.E. Britigan, Y.S. Chai et al. // J. Infect Dis. - 1991. - Vol. 163. - 819-826].
С помощью чувствительных спиновых меток обнаружена наработка гидроксил-радикала клетки in vitro в результате реакции HOCl и
, причем преобразованию в •ОН подверглась очень небольшая часть использованного клетками кислорода [Free hydroxyl radicals are formed on reaction between the neutrophilderived species superoxide and hypochlorous acid / L.P. Candeias, K.B. Patel, M.R.L. Stratford et al. // FEBS Lett. - 1993. - Vol. 333. - P. 151-159]. Вопрос о том, достаточно ли такого количества •ОН, чтобы играть существенную роль в цитотоксичности, до сих пор остается открытым. Здесь необходимо учитывать, что гораздо большей бактерицидной способностью •ОН обладает в присутствии Cl- [Radiation induced generation of chlorine derivatives in N2O-saturated phosphate buffered saline: Toxic effects on Escherichia coli cells / G. Czapski, S. Goldstein, N. Andorn et al. // Free Radic. Biol. Med. - 1992. - Vol. 12. - P. 353-361.], вероятно, вследствие реакции между ними с образованием гипохлорита [Bactericidal potency of hydroxyl radical in physiological environments / R.G. Wolcott, B.S. Franks, D.M. Hannum et al. // J. Biol. Chem. - 1994. - Vol. 269. - P. 9721-9734].
Гидроксильный радикал представляет собой один из наиболее реакционно-способных окислителей и может взаимодействовать почти с любой молекулой клетки. Он модифицирует дезоксирибозу и азотистые основания ДНК, окисляет молекулы белков, углеводов и липидов. Особенно активно •ОН в ходе реакций перекисного окисления липидов атакует фосфолипиды, содержащие в жирнокислотных радикалах ненасыщенные связи, что ведет к образованию гидроперекисей [Дубинина Е.Е. Продукты метаболизма кислорода в функциональной активности клеток (жизнь и смерть, созидание и разрушение). Физиологические и клинико-биохимические аспекты / Е.Е. Дубинина. - СПб.: Медицинская пресса, 2006. - 400 с.; Окислительный стресс. Прооксиданты и антиоксиданты / Е.Б. Меньшикова, В.З. Ланкин, Н.К. Зенков и соавт. - М.: Слово. - 2006. - 556 с.]. Основным компонентом антиокидантной системы является восстановленная форма глутатиона.
Глутатион - трипептид (L-γ-глутамил-L-цистеилглицин) с молекулярной массой 307 Da занимает особое место среди SH-содержащих соединений. Наличие γ-глутамильной связи защищает трипептид от ферментативной деградации. В организме глутатион присутствует в двух формах: окисленной - GSSG и восстановленной - GSH, причем содержание GSH в клетках на несколько порядков выше, чем GSSG [Колесниченко Л.С., 1989; Wu G. et al., 2004; Смирнова Г.В., Октябрьский О.Н., 2005; Марри Р. и соавт., 2009]. По данным P. Pietarinen-Runtti et al. (2000), концентрация GSH в нейтрофилах составляет около 5 нмоль/мг белка. Содержание глутатиона в сыворотке крови здоровых людей незначительно, поэтому клетки основную потребность в GSH обеспечивают путем нематричного синтеза [Wu G. et al., 2004] в ходе двух последовательных реакций, катализируемых γ-глутамилцистеин-синтетазой (КФ 6.3.2.2) и глутатион-синтетазой (КФ 6.3.2.3) [Кулинский В.И., 1990; Смирнова Г.В., Октябрьский О.Н., 2005; Марри Р. и соавт., 2009]. Лимитирующим звеном синтеза является образование γ-глутамилцистеина, зависящее от наличия L-цистеина и его способности окисляться в L-цистин [Зенков Н.К. и соавт., 2001]. В то же время недостаточность глутатион-синтетазы способствует развитию окислительных повреждений в нейтрофилах [Spielberg S.P. et al., 1979].
Глутатион при физиологических значениях рН имеет две анионные карбокси-группы, положительно заряженную аминогруппу и SH-группу цистеинового остатка, которая придает GSH свойства восстановителя и способность быстро обезвреживать свободные радикалы и АФК [Day R.M., 2005; Zhu Y., 2007; Circu C.L. et al., 2009]. Глутатин является типичным тиолом и, участвуя в одноэлектронных восстановительных реакциях, становится GS•, который димеризуется до GSSG, легко реагирующего со свободными SH-группами. Второй тип окислительно-восстановительных превращений с участием GSH - это реакции тиолдисульфидного обмена, которые известны как основной путь образования смешанных дисульфидов глутатиона с белками (белок-SSG) и играют роль в регуляции биологических процессов [Chai Y.C. et al., 1994]. В реакциях третьего типа происходит двухэлектронное окисление глутатиона с образованием интермедиата, который реагирует со второй молекулой GSH (получение GSSG) или иной молекулой (синтез смешанного дисульфида) [Смирнова Г.В., Октябрьский О.Н., 2005].
GSH является стабилизатором мембран [Биленко М.В, 1989; Udupi V., 1992; Trudel S. et al., 2009]. Он защищает клеточные структуры клеток от высокотоксичного OCI - [Carr А.С., Winterbourn С.С., 1997], при этом GSH превращается в глутатион-сульфонамид и дегидроглутатион [Harwood D.T. et al., 2006]. Связывая NO, глутатион образует токсичные для клетки нитрозильные комплексы. Мононитрозоглутатион может активировать апоптоз [Turpaev K.Т. et al., 1997].
Не всегда восстановительного потенциала GSH достаточно для полной нейтрализации прооксидантов. Существует мнение, что взаимодействие GSH с органическими радикалами эффективно только в условиях удаления
, поэтому глутатион образует с супероксиддисмутазой своеобразную антиоксидантную систему, ибо в противном случае развиваются реакции образования Н2О2 и GS• [Ланкин В.З. и соавт., 1997; Меньшикова Е.Б. и соавт., 2006]. В сочетании с витамином В12 глутатион, а также N-ацетилцистеин, могут потенцировать прооксидантное и цитотоксическое действие на клетку [Соловьева М.Е. и соавт., 2007].
Основной антиоксидантный эффект GSH реализует посредством участия в работе ферментов. Глутатион выступает донором водорода при восстановлении Н2О2 и перекисей липидов глутатион-пероксидазами и глутатион-S-трансферазами (ГТ) [Hirayama K., 1989; Sies Н. et al., 1997; Кулинский В.И., 1990; Hayes J.D. et al., 2005; Зенков H.К.. и соавт., 2009; Liu G. et al., 2010]. Высокая активность глутатион-редуктазы и накопление GSH оказывает протекторный эффект в отношении альвеолярных макрофагов, инкубируемых с прооксидантами in vitro [Pietarinen Р.K., 1995].
С изменением окислительно-восстановительного баланса сопряжено большое количество реакций, поэтому поддержание оптимального редокс-состояния цитозоля выступает важным условием нормальной жизнедеятельности клеток. Высокая концентрация глутатиона в цитоплазме, его редокс-активность и возможность поддержания в восстановленном состоянии делают систему GSH/GSSG важнейшим внутриклеточным редокс-буфером [Reed М.С. et al., 2008]. Концентрация GSH в клетке в 500-1000 раз превышает уровень НАДФН и других внутриклеточных редокс-систем, поэтому изменения соотношения GSH/GSSG прямо отражают изменения редокс-статуса клетки [Кулинский В.И., 2007; Aslan М., Canatan D., 2008; Reed М.С, 2008]. Считают, что буферная емкость системы глутатиона защищает репликативную систему клетки, а дефицит GSH приводит к снижению синтеза ДНК и белков [Poot М., 1991; Ланкин В.З., 1997; Day R.M., Suzuki Y.J., 2005; Liu G. et al., 2010], а затем и к апоптозу.
К природным антиоксидантам относят также аскорбиновую кислоту, которая играет важную роль в развитии окислительного стресса в организме.
Аскорбиновая кислота реализует свое антиоксидантное действие в плазме, межклеточной жидкости и на внеклеточном уровне. В организме человека аскорбиновая кислота преимущественно представлена в L-форме. Стрессовые ситуации увеличивают количество метаболитов витамина С в виде дегидроаскорбиновой кислоты.
Аскорбиновая кислота и дегидроаскорбиновая кислота играют активную роль в нескольких процессах, включая защиту от инфекции, повышении иммунности, в процессах заживления ран, а также принимая участие в образовании антистрессовых гормонов. Аскорбат является кофактором дофамин-β-гидроксилазы, которая катализирует синтез норадреналина и других катехоламинов. Аскорбиновая кислота является восстановителем для L-пролингидроксилазы, которая необходима для синтеза коллагена и соединительной ткани в целом. В организме с участием аскорбиновой кислоты происходит регенерация α-токоферола из токофероксильного радикала. Окислительный стресс коррелирует с ухудшением секреции инсулина, а терапия аскорбиновой кислотой прерывает повреждающее действие свободных радикалов, уменьшает степень проявления инсулиновой резистентности [М.И. Балаболкин и соавт., 2003]. Ионы аскорбата являются одним из активных элементов системы антиоксидантной защиты, предохраняя липиды от окисления их пероксидными радикалами. Антиоксидантный эффект аскорбата проявляется при достаточном количестве других антиоксидантов, таких как α-токоферол и глутатион. Глутатион восстанавливает дегидроаскорбиновую кислоту прямым и неферментативным путем до аскорбиновой кислоты. Эта реакция является одним из основных механизмов антиоксидантной системы, часто описываемых как восстановительные циклы - глутатион/глутатиондисульфид и аскорбиновая/дегидроаскорбиновая кислота. При этом клетки периферических тканей поглощают экзогенную дегидроаскорбиновую кислоту и в присутствии глутатиона конвертируют ее в цитоплазме в аскорбиновую кислоту. Восстановление глутатиондисульфида в глутатион катализируется глутатион редуктазой и требует участия NADPH в качестве кофактора. Недостаточность глутатиона снижает содержание аскорбиновой кислоты в тканях и одновременно повышает концентрацию дегидроаскорбиновой кислоты.
При недостатке α-токоферола и глутатиона может превалировать прооксидантный эффект аскорбата и его метаболитов. Прооксидантный эффект аскорбиновой кислоты может наблюдаться не только при недостатке α-токоферола и глутатиона, но и при применении высоких доз аскорбиновой кислоты. Избежать прооксидантного эффекта аскорбиновой кислоты можно в случае создания адекватного внутриклеточного уровня восстановленного глутатиона.
Исходя из вышесказанного, целесообразно использовать в эксперименте комплексное применение аскорбата с протектором SH-групп - а именно 1,4-дитиоэритритолом. Для проникновения внутрь клетки пассивным транспортом происходит превращение аскорбиновой кислоты в дегидроаскорбиновую кислоту, затем последняя подвергается обратимому превращению в аскорбиновую кислоту при участии восстановленного глутатиона. Следовательно, комплексная модернизация способа-прототипа позволяет повысить точность оценки эффективности защиты лимфоцитов от апоптоза.
Каждый вновь введенный в формулу изобретения признак выполняет функцию повышения точности и эффективности способа: дополнительное добавление в инкубационную среду 1,4-дитиоэритритола и аскорбиновой кислоты в низкой концентрации для последующего определения белково-связанного и восстановленного глутатиона. Установлено, что при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту низкоэффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту высокоэффективной.
Роль антиоксидантной системы клетки заключается в снижении токсического эффекта свободных радикалов, в том числе и гидроперекисей липидов.
Антиоксидантную защиту обеспечивает широкий круг веществ, различных по происхождению, физико-химической природе и механизмам действия. Общим их свойством, по определению J.M. Gutteridge (1992), является способность, присутствуя в низких по сравнению с окисляемым субстратом концентрациях, существенно задерживать или ингибировать его окисление. Постоянное образование прооксидантов должно быть уравновешено их инактивацией, поэтому для поддержания гомеостаза необходима адекватная ситуации непрерывная регенерация антиоксидантной способности клеток [Зенков Н.К. и соавт., 2001; Blokhina О. et al., 2003].
Общепринятой номенклатуры антиоксидантов в настоящее время не существует, хотя ряд авторов [Dimascio Р., 1990; Kalra V. et al., 2001; Зайцев В.Г. и соавт., 2003] выделяет два класса: превентивные, снижающие скорость инициации цепной реакции окисления, и гасящие (прерывающие цепь), препятствующие развитию цепной реакции. К превентивным относят каталазу и пероксидазы, разрушающие ROOH, а также агенты, образующие хелатные комплексы с металлами переменной валентности, к прерывающим цепь - фенолы, ароматические амины. В условиях in vivo главными гасящими антиоксидантами являются: витамин Е, нейтрализующий ROO• в липидной фазе мембран [Jore D. et al., 1990; Hong J.H. et al., 2004], фермент СОД, улавливающий
в водной фазе клетки [Fridovich I., 1989; Dimascio P., 1990; Ciurea D., 1992], и церулоплазмин - белок острой фазы, выполняющий антирадикальную функцию в крови [Marklund S.L., 1987; Atanasiu R.L. et al., 1998].
Более известно деление антиоксидантов на ферменты и соединения неферментативной природы. Последние в определенных концентрациях всегда присутствуют в липидной фазе мембран и водных средах организма и расходуются первыми при устранении проявлений окислительного стресса [Droge W., 2002; Blokhina О., 2003]. Ферменты наиболее активно присоединяется к антиоксидантной защите (АОЗ) после включения механизмов индукции [Лущак В.И., 2001]. При возникновении окислительного стресса (ОС) расход антиоксидантов возрастает и меняется экспрессия генов, кодирующих белковые компоненты АОЗ [Дубинина Е.Е., 2006]. Между ферментами и неферментативными элементами АОЗ существует равновесие, причем последние при ряде патологических состояний организма могут выступать в качестве прооксидантов [Зенков Н.К и др., 2001].
Главную роль среди неферментавных антиоксидантных систем защиты отводят глутатиону.
Функционирование клеток связано с уровнем белково-связанного глутатиона. Определение белково-связанного глутатиона основано на способности боргидрата натрия (NaBH4) высвобождать из связи с белками глутатион, который при взаимодействии с ДТНБ образует окрашенное соединение, а именно тио-2-нитробензойную кислоту, водный раствор которой имеет максимум поглощения при длине волны 412 нм [Burchill, B.R. Microtubule dynamics and glutathione metabolism in phagocytizing human polymorphonuclear leukocytes [Text] / B.R. Burchill, J.M. Oliver, C.B. Pearson et al. // J. of Cell Biology. - 1978. - Vol. 76, №2. - P. 439-447].
В настоящее время крайне важно определить уровень белково-связанного глутатиона для защиты клеток от апоптоза.
В настоящее время в лабораторной практике наиболее распространен способ оценки эффективности защиты лимфоцитов от апоптоза и переокисления с помощью определения концентрации белково-связанного и восстановленного глутатиона.
Концентрацию гидроксирадикалов определяют методом, предложенным [Thorn S.R, Elbuken M.E., 1991]. Метод основан на разрушении модельного субстрата 2-дезокси-D-рибозы гидроксильным радикалом, образуемым опсонизированными лимфоцитами.
Содержание восстановленного глутатиона определяют методом, предложенным М.Е. Anderson (1985) в модификации S. Kojima et al. (2004) [Kojima, S. Low dose gamma-rays activate immune functions via induction of glutathione and delay tumor growth / S. Kojima, K. Nakayama, H. Ishida // J. Radiat. Res. - 2004. - Vol. 45, №1. - P. 33-39]. Принцип метода основан на взаимодействии GSH с 5,5′-дитио-бис(2-нитробензойной) кислотой (ДТНБ) с образованием тио-2-нитробензойной кислоты, водный раствор которой имеет максимум поглощения при длине волны 412 нм. При этом образуется GSSG, который восстанавливается глутатионредуктазой до GSH и вновь взаимодействует с ДТНБ. Скорость образования окрашенного продукта пропорциональна содержанию общего глутатиона. Для определения содержания GSSG пробы прединкубируются с блокатором SH-групп 2-винилпиридином («Wako», Япония), который необратимо связывает GSH и, следовательно, скорость образования окрашенного продукта пропорциональна содержанию GSSG.
Лизат клеток готовят на 5% сульфосалициловой кислоте, которая осаждает белки, но не ингибирует активность глутатионредуктазы.
Концентрацию белка в клетках определяют методом [A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding / M.M. Bradford // Analyt. Biochem. - 1976. - Vol. 7, №1, 2. - P. 248-254], основанным на взаимодействии Кумасси голубого G-250 с остатками аргинина и лизина в белках. Свободный краситель красного цвета (максимум поглощения - 495 нм) при образовании комплекса с белком переходит в синюю форму (максимум поглощения - 595 нм).
К 0,1 мл лизата клеток добавляют 1,0 мл раствора Кумасси голубого (100 мг красителя, 50 мл 96° этанола, 100 мл 85% Н3РО4, Н2О до 1,0 л), перемешивают, инкубируют 3 мин при комнатной температуре и измеряют оптическую плотность проб (длина волны 595 нм) против контроля, содержащего 0,1 мл воды и 1,0 мл раствора Кумасси голубого. Содержание белка рассчитывают по калибровочной кривой, построенной по разведениям стандартного раствора альбумина (1,0 мг/мл) и выражают в мг/мл.
В настоящее время крайне важно для защиты клеток от апоптоза и переокисления оценить концентрацию различных форм глутатиона после стимуляции антиоксидантной активности низкими концентрациями 1,4-дитиоэритритола и аскорбиновой кислоты. Для решения этой задачи предложен новый способ защиты клеток от апоптоза после дополнительного добавления в инкубационную среду 1,4-дитиоэритритола и аскорбиновой кислоты в конечной концентрации 3.0 мМ и 0,1 мМ соответственно.
Все сказанное свидетельствует о крайней важности разработки способа защиты клеток от апоптоза, а также для защиты клеток от токсического действия активных форм кислорода.
Популярность указанного выше способа обоснована его высокой чувствительностью, простотой осуществления и достаточной адекватностью получаемых результатов, лежащих в основе определения концентрации гидроксильных радикалов в среде инкубации лимфоцитов.
Существенные признаки, характеризующие изобретение, проявили в заявленной совокупности новые свойства, явным образом не вытекающие из уровня техники в данной области, и не являются очевидными для специалиста.
Идентичной совокупности признаков не обнаружено при изучении патентной и научной медицинской литературы. Данное изобретение может быть использовано в медицинской практике для повышения точности защиты клеток от апоптоза при различных заболеваниях. Таким образом, следует считать предлагаемое изобретение соответствующим условиям патентоспособности: «новизна», «изобретательский уровень», «промышленная применимость».
Метод основан на определении концентрации белок-связанного и восстановленного глутатиона.
Способ осуществляется следующим образом поэтапно:
1. Выделение культуры клеток линии Jurkat.
2. Количественное определение численности жизнеспособных клеток с помощью окраски трипановым синим микроскопическим методом.
Клетки ресуспендируют в 1 мл клеточной взвеси. Отбирают 100 мкл ресуспендированной клеточной суспензии и добавляют 100 мкл 0,1% раствора трипанового синего на физ. растворе, хорошо перемешивают и заполняют камеру Горяева. Предварительно к камере притирают покровное стекло так, чтобы появлялись радужные, ньютоновые кольца (только при этих условиях соблюдался правильный объем камеры). Каплю клеточной взвеси с красителем вносят под притертое покровное стекло. Подсчет клеток производят в 5-ти больших квадратах по диагонали камеры Горяеева. Расчет клеточности лимфоцитов крови производят по формуле
А×106=(число клеток)/4
где А - клеточность лимфоцитов крови.
3. Внесение в инкубационную смесь заявленных добавок.
В культуральную смесь добавляют соединения: 1,4-дитиоэритритол в концентрации 3.0 мМ и аскорбиновую кислоту в конечной концентрации 0,1 мМ.
4. Биохимическое исследование белково-связанного, восстановленного и окисленного глутатиона.
Лизат лимфоцитов готовят на 5% сульфосалициловой кислоте, которая осаждает белки, но не ингибирует активность глутатионредуктазы. Количество общего глутатиона (GSH и GSSG) определяют в пробе содержащей 0,1 М Na-фосфатный буфер (рН=7,5) с 1 мМ ЭДТА, 0,4 мМ НАДФН2, 0,3 мМ ДТНБ и 1 U/мл глутатионредуктазы («Wako», Япония). Окисленный глутатион определяют аналогичным способом в клеточном лизате после предварительной инкубации пробы в течение 30 мин с 10 мМ 2-винилпиридином. Расчет содержания общего и окисленного глутатиона производят с помощью калибровочных графиков, для построения которых используют растворы GSH и GSSG («МР», США) в концентрации от 3 до 100 мкМ, обработанные аналогично опытным пробам. Концентрацию GSH рассчитывают как разницу между концентрацией общего глутатиона и GSSG, выражая результат в нмоль/мг белка.
Определение белково-связанного глутатиона.
После инкубации в экспериментальных условиях (5% и 20% кислорода) в присутствии или отсутствии NEM, DTE, NAC клетки центрифугируют 5 минут при 4°С и 1500 об/мин для их осаждения. Удаляют супернатант. Добавляют 1 мл охлажденного PBS (рН 7,4). Ресуспендируют на вортексе. Центрифугируют 5 минут при 4°С и 1500 об/мин. Удаляют супернатант. Осадок клеток ресуспендируют в 1 мл 5% сульфосалициловой кислоты для получения клеточного лизата. Центрифугируют при 3000 об/мин 10 мин. 1,0 мл осадка белка инкубируют 1 ч при 50°С с 1,0 мл 1% NaBH4». Далее оставшийся белок осаждают добавлением 0,4 мл 30% ТХУ. Пробу инкубируют 15 мин при 50°С. Затем пробу охлаждают 5 мин (0°С). Центрифугируют 10 мин при 3000 об/мин. Супернатант смешивают с 2,5 мл PBS (рН 7,4) и добавляют 2,0 мл ацетона для полного окисления NaBH4. Перемешивают. Центрифугировают 10 мин при 3000 об/мин. Удаляют верхнюю фазу. К нижней фазе добавляют равный объем диэтилового эфира (для удаления ТХУ). Перемешивают. Центрифугировают 10 мин при 3000 об/мин. Затем снова удаляют верхнюю фазу. Далее процедуру отмывки пробы от ТХУ с помощью диэтилового эфира производят 4-х кратно. Затем отбирают 0,1 мл жидкости (из нижней фазы) и смешивают с 0,4 мл 0,01 М фосфатного буфера (рН=7,0). В пробу добавляют 0,1 мл 0,4 мг/мл ДТНБ. Пробу спектрофотометрируют при 412 нм против контроля, содержащего воду вместо раствора осажденного белка. Расчет производят с учетом коэффициента молярной экстинкции 13·103 М-1см-1. Результаты определения концентрации белково-связанного глутатиона выражают в нмоль/мг белка.
Оценка способа оценки эффективности защиты лимфоцитов от апоптоза по способу-прототипу и предлагаемому способу выполнялось 40 раз. Результаты исследования обработаны статистически с использованием пакета программ Stat Soft Statistica 6.0.
При проведении исследования по способу-прототипу уровень восстановленного глутатиона в среде инкубации клеток в норме составил 2.15±0.18 нмоль/мг белка и белково-связанного глутатиона 2.9±0.25 нмоль/мг белка, а при оценке по предлагаемому способу при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту лимфоцитов от апоптоза низко эффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту лимфоцитов от апоптоза высоко эффективной (Табл. 1).
Полученные результаты уровня глутатиона в среде инкубации клеток соответствуют данным литературы [Смирнова Г.В., Октябрьский О.Н., 2005]. Итак, при применении способа-прототипа был получен недостаточно точный результат, не позволивший оценить эффективность защиты лимфоцитов от апоптоза, что связано с отсутствием биохимической стимуляции процесса, а наиболее эффективным и точным был предлагаемый способ. При этом предлагаемый способ прост в исполнении и интерпретации полученных результатов.
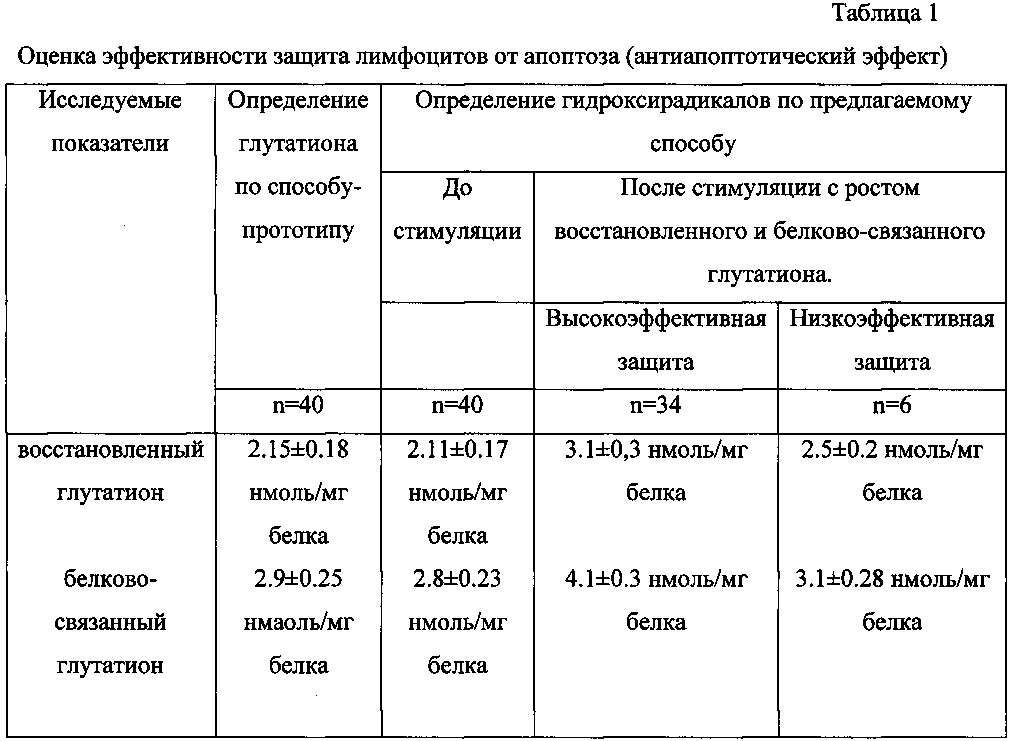
То есть при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту лимфоцитов от апоптоза низкоэффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту лимфоцитов от апоптоза высокоэффективной.
Полученные результаты уровня глутатиона в среде инкубации лимфоцитов соответствуют данным литературы [Смирнова Г.В., Октябрьский О.Н., 2005].
Итак, при применении способа-прототипа был получен недостаточно точный результат, не позволивший оценить эффективность защиты лимфоцитов от апоптоза, что связано с отсутствием биохимической стимуляции процесса, а наиболее эффективным и точным был предлагаемый способ.
При этом предлагаемый способ прост в исполнении и интерпретации полученных результатов.
Способ оценки эффективности защиты лимфоцитов от апоптоза, заключающийся в выделении лимфоцитов, инкубации в течение 48 ч при температуре 37°С и 5% содержании СО, в количественном определении жизнеспособности лимфоцитов по включению трипанового синего и биохимическом определении концентрации белково-связанного и восстановленного глутатиона в лизате лимфоцитов после предварительной инкубации в течение 30 минут с 10 ммоль 2-винилпиридином, отличающийся тем, что дополнительно в инкубационную среду, содержащую лимфоциты, вводят 1,4-дитиоэритритол и аскорбиновую кислоту в конечной концентрации 3,0 ммоль и 0,1 ммоль соответственно, и при росте окисленного глутатиона на 18-35% и восстановленного глутатиона на 25-41% оценивают защиту лимфоцитов от апоптоза как низкоэффективную, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более оценивают защиту лимфоцитов от апоптоза как высокоэффективную.Способ оценки эффективности лечения ишемической болезни сердца
Способ диагностики липидемии
Способ прогнозирования течения ишемической болезни сердца
Средство и способ индукции апоптоза опухолевых клеток
Способ определения раковых клеток в серозных жидкостях у больных с подозрением на злокачественные новообразования
Способ клинической оценки активности воспаления при хроническом калькулезном холецистите
Кальций-фосфатное биологически активное покрытие на имплантате
Способ прогнозирования течения ишемической болезни сердца
Способ определения липидов
Способ оценки эффективности стимуляции антиоксидантной активности
Средство и способ индукции апоптоза опухолевых клеток
Способ клинической оценки активности воспаления при хроническом калькулезном холецистите
Кальций-фосфатное биологически активное покрытие на имплантате
Способ оценки эффективности стимуляции антиоксидантной активности
Способ прогнозирования течения липидемии
Способ оценки прогрессирования атерогенности при ишемической болезни сердца
Способ прогнозирования эффективности защиты лимфоцитов от переокисления
Способ диагностики острого токсического повреждения печени
Способ прогнозирования ранней стадии апоптоза
Способ создания модели перекисного окисления лимфоцитов

