АНТИТЕЛА ИЛИ ИХ ФРАГМЕНТЫ, НАПРАВЛЕННЫЕ ПРОТИВ ЭПИТОПА СТАФИЛОКОККА ЗОЛОТИСТОГО (STAPHYLOCOCCUS AUREUS)
Вид РИД
Изобретение
Изобретение относится к антителам и их фрагментам, которые направлены против эпитопа стафилококка золотистого (Staphylococcus aureus) (=S. aureus), к набору, содержащему эти антитела или фрагменты, к использованию этих антител или фрагментов, к линии клеток гибридомы, которая продуцирует эти антитела, и к способу лечения.
Из журнала FEMS Imunol. Med. Microbial., октябрь 2000 г.; 29(2), страницы 145 -153 известны иммунодоминантные структуры, которые экспрессируются in vivo при сепсисе, вызванном метициллин-резистентным стафилококком золотистым (MRSA). Эти структуры представляют собой белок иммунодоминантного секреторного антигена A (IsaA) с молекулярной массой 29 кДа и белок иммунодоминантного секреторного антигена В (IsaB) с молекулярной массой 17 кДа. Заявляется, что эти белки могут служить потенциальной мишенью для разработки терапии против MRSA на основе антител.
Из реферата статьи «Разработка терапии на основе антител, нацеленной против иммунодоминантных антигенов стафилококка золотистого (Staphylococcus aureus)», Олсен К. и др., стр.128 сборника рефератов, опубликованного по результатам 59-й конференции Немецкого общества гигиены и микробиологии (Deutsche Gesellschaft fur Hygiene und Mikrobiologie e.V.) в сентябре 2007 г., и из статьи Лоренца У. и др. «Терапевтическая эффективность моноклональных антител к стафилококку золотистому (Staphylococcus aureus) на модели сепсиса и абсцесса у мышей», Хирургический форум 2008 г., Springer Berlin Heidelberg, 29 мая 2008 г., выпуск 17, стр.225-226, известно применение одного мышиного моноклонального антитела, нацеленного против иммунодоминантного антигена IsaA, в двух инфекционных моделях на животных. Исследование показало, что применение моноклональных антител к IsaA снижает инфекционную нагрузку в обеих инфекционных моделях.
Задачей настоящего изобретения является создание новых антител или их фрагментов, которые бы хорошо подходили для лечения инфекций, вызванных стафилококком золотистым, и для обнаружения S. aureus. Кроме того, могут быть предоставлены набор, содержащий эти антитела или их фрагменты, способ использования этих антител или фрагментов, линия клеток гибридомы, секретирующая эти антитела или фрагменты, и способ лечения.
Эту задачу решает объект, описанный в пунктах формулы изобретения 1, 8, 15, 16, 17 и 19. Варианты осуществления изобретения раскрываются в пунктах формулы изобретения 2-7, 9-14, 18 и 20-24.
Предлагается использовать антитела или их фрагменты, когда упомянутые антитела или фрагменты направлены против эпитопа стафилококка золотистого, который определяется моноклинальными новыми антителами, секретируемыми линией клеток гибридомы, хранящейся в «Deutsche Sammlung von Mikroorganismen und Zeilkulturen GmbH, Inhoffenstr. 76, D-38124 Braunschweig, Germany» (DSMZ) под инвентарным номером DSM ACC2987 или DSM ACC2988. Линия клеток гибридомы, хранящаяся в DSMZ под инвентарным номером DSM ACC2987, далее по тексту именуется «клеточная линия DSM ACC2987», а линия клеток гибридомы под инвентарным номером DSM ACC2988 далее по тексту именуется «клеточная линия DSM ACC2988». Моноклональные новые антитела, которые секретируются клеточными линиями DSM ACC2987 и DSM ACC2988, являются мышиными моноклональными антителами.
Предлагаемые антитела содержат полные молекулы иммуноглобулина, предпочтительно IgMs, IgDs, IgEs, IgAs или IgGs, более предпочтительно lgG1, lgG2a, lgG2b, lgG3 или lgG4, а фрагменты содержат часть таких молекул иммуноглобулинов, как например, Fab-фрагменты либо V-, VH- или CDR-области. Антитела также содержат модифицированные и/или измененные антитела, как например, химерные или гуманизированные антитела. Антитела также содержат модифицированные или измененные моноклональные или поликлональные антитела, а также созданные рекомбинантным или синтетическим образом или же синтезированные антитела. Фрагменты содержат фрагменты антител, а также их части, как например, отделенные легкие или тяжелые цепочки Fab, Fab/c, Fv, Fab', F(ab')2. Предлагаемые антитела также содержат производные антител, как например, биспецифические (бифункциональные) антитела и конструкты антител, как например, одноцепочечные Fv фрагменты (scFv, или оцFv), биспецифические оцру фрагменты или гибридные белки антител. Все производные антител демонстрируют связывающую специфичность антител, из которых они получены, т.е. они нацелены против эпитопа стафилококка золотистого, который определяется моноклональными новыми антителами, секретируемыми линиями клеток гибридомы DSM ACC2987 или DSM ACC2988.
Эпитоп, который определяется моноклональными новыми антителами, секретируемыми линией клеток гибридомы DSM ACC2987, находится на иммунодоминантном антигене S. aureus IsaA. Эпитоп, который определяется моноклональными новыми антителами, секретируемыми линией клеток гибридомы DSM ACC2988, находится на иммунодоминантном антигене S. aureus IsaB. Авторы настоящего изобретения обнаружили, что эпитопы, определяемые предлагаемыми антителами, особенно уязвимы у S.aureus. Антитела или их фрагменты, нацеленные против этих эпитопов, отлично подходят для обнаружения S.aureus и для лечения инфекции, вызванной S. aureus. По причине высокой изменчивости S. aureus, результатом которой является различный уровень экспрессии и мутации антигенов у различных штаммов, каждое антитело, которое определяет дополнительный эпитоп, не определяемый другими антителами, подходит для определения S. aureus, а также для лечения инфекции, вызванной S.aureus.
Антитела могут быть моноклональными или поликлональными антителами. В частности, антитела могут быть моноклональными новыми антителами, т.е. антителами, которые секретируются линией клеток гибридомы DSM ACC2987 или DSM ACC2988. Эти антитела очень подходят для обнаружения S.aureus, а также для лечения инфекции. Они демонстрируют очень высокую аффинность и специфичность. На высокую аффинность антител, которые секретируются линией клеток гибридомы DSM ACC2987, к эпитопу, определяемому этими антителами, указывает низкое значение Ко для связывания упомянутого антитела с упомянутым эпитопом. В зависимости от способа определения, для этого связывания были определены значения KD≤18 пМ и 1,7 нМ (см. Фиг.6 и 8 и соответствующий текст).
Моноклональные антитела или антитела, секретируемые линией клеток гибридомы DSM ACC2987 и/или DSM ACC2988, могут быть антителами типа IgG, в частности, типа lgG1 или типа lgG2b. Фрагменты могут представлять собой Fab-фрагменты, Fab/c-фрагменты, Fv-фрагменты, Fab'-фрагменты или F(ab')2-фрагменты. Эти фрагменты особенно подходят для обнаружения S.aureus, поскольку клеточная стенка S.aureus содержит белок А, который неспецифически связывается с иммуноглобулинами через их Fc-области.
Антитела могут представлять собой антитела животных, т.е. антитела, выработанные у животных, в частности, мышиные, бычьи или верблюжьи антитела, человеческие антитела, антитела, выработанные у растений, в яйце или у грибов, в частности у сахаромицетов, рекомбинантные антитела, выработанные в клетках клеточной линии, химерные антитела или гуманизированные антитела. Гуманизированное антитело может представлять собой моноклональное антитело, которое содержит связывающую часть моноклонального мышиного антитела, например, моноклональное антитело, секретируемое линией клеток гибридомы DSM ACC2987 или DSM ACC2988, и не связывающую часть человеческого антитела.
Каждое из антител может иметь тяжелую цепочку с первой вариабельной областью и легкую цепочку со второй вариабельной областью,
где линия клеток гибридомы - DSM ACC2987, и где первая вариабельная область содержит аминокислотную последовательность, которая, по меньшей мере, на 90% идентична, в частности, по меньшей мере, на 92,5% идентична, в частности, по меньшей мере, на 95% идентична, в частности, по меньшей мере, на 97,5% идентична, в частности, на 100% идентична последовательности №2 (SEQ ID NO: 2), и где вторая вариабельная область содержит последовательность, которая, по меньшей мере, на 90% идентична, в частности, по меньшей мере, на 92,5% идентична, в частности, по меньшей мере, на 95% идентична, в частности, по меньшей мере, на 97,5% идентична, в частности, на 100% идентична последовательности №4 (SEQ ID NO: 4)
или
где линия клеток гибридомы - DSM ACC2988, и где первая вариабельная область содержит аминокислотную последовательность, которая, по меньшей мере, на 90% идентична, в частности, по меньшей мере, на 92,5% идентична, в частности, по меньшей мере, на 95% идентична, в частности, по меньшей мере, на 97,5% идентична, в частности, на 100% идентична последовательности №6(SEQ ID NO: 6), и где вторая вариабельная область содержит последовательность, которая по меньшей мере, на 90% идентична, в частности, по меньшей мере, на 92,5% идентична, в частности, по меньшей мере, на 95% идентична, в частности, по меньшей мере, на 97,5% идентична, в частности, на 100% идентична последовательности №8 (SEQ ID NO: 8).
Первая вариабельная область и вторая вариабельная область, как охарактеризовано выше, вместе образуют участок связывания, обладающий высокой аффинностью и специфичностью к эпитопу.
Одной возможной последовательностью ДНК, кодирующей первую вариабельную область согласно последовательности №2 (SEQ ID NO: 2), является последовательность №1 (SEQ ID NO: 1). Последовательность №1 (SEQ ID NO: 1) является последовательностью, кодирующей первую вариабельную область антител, секретируемых клеточной линией DSM ACC2987. Вторая вариабельная область согласно последовательности №4 (SEQ ID NO: 4) может кодироваться последовательностью №3 (SEQ ID NO: 3). Последовательность №3 является последовательностью, кодирующей вторую вариабельную область антител, секретируемых клеточной линией DSM ACC2987.
Первая вариабельная область согласно последовательности №6 (SEQ ID NO: 6) может кодироваться последовательностью №5 (SEQ ID NO: 5), которая является последовательностью, кодирующей первую вариабельную область антител, секретируемых клеточной линией DSM ACC2988. Вторая вариабельная область согласно последовательности №8 (SEQ ID NO: 8) может кодироваться последовательностью №7 (SEQ ID NO: 9). Последовательность №7 (SEQ ID NO: 7) является последовательностью, кодирующей вторую вариабельную область антител, секретируемых клеточной линией DSM ACC2988.
Если линия клеток гибридомы - DSM ACC2987, т.е. если эпитоп находится на иммунодоминантном антигене S.aureus IsaA, эпитоп может содержать, по меньшей мере, одну аминокислотную последовательность, которая идентична, по меньшей мере, на 10, в частности, по меньшей мере, на 11, в частности, по меньшей мере, на 12, в частности, по меньшей мере, на 13, в частности, по меньшей мере, на 14, в частности, по меньшей мере, на 15 аминокислот с одной из последовательностей №15, 17-19, 21-26, 32-34 и 57 (SEQ ID NO: 15, 17-19, 21-26, 32-34 и 57) согласно списку последовательностей.
Картирование эпитопов показало, что последовательности №15, 17-19, 21-26, 32-34 и 57 участвуют в связывании антител или фрагментов с эпитопом. Эпитоп может содержать более одной аминокислотной последовательности, как указано выше. Эпитоп может быть даже образован двумя или более последовательностями, расположенными на расстоянии друг от друга в аминокислотной последовательности IsaA.
Предлагаемые в данном изобретении антитела или фрагменты могут использоваться в качестве лекарственного средства. В особенности, они могут использоваться в качестве лекарственного средства для лечения человека или животного, которое инфицировано S. aureus, особенно метициллин-резистентным или метициллин-чувствительным S.aureus, или у которого есть риск инфицирования. У человека или животного может быть мастит или сепсис, вызванный инфекцией. Мастит может представлять собой мастит крупного рогатого скота. Если корова заболела маститом крупного рогатого скота, корова не дает пригодного к использованию молока, а если корова получает лечение антибиотиками, как обычно, в этом случае молоко от этой коровы подлежит утилизации до тех пор, пока в молоке от этой коровы присутствуют антибиотики. Этого недостатка традиционного лечения можно избежать при использовании предлагаемых антител или фрагментов в качестве лекарственного средства для лечения мастита крупного рогатого скота.
Лекарственное средство может быть лекарственным средством для системного и/или местного применения. Авторы изобретения признают, что лечение тяжелой инфекции S.aureus предлагаемыми антителами или фрагментами приводит к существенному снижению уровня смертности и количества S.aureus в органах получавшего лечение человека или животного. Более того, авторы изобретения признают, что фагоцитарный лизис (цитолиз) бактерий S.aureus полиморфно-ядерными лейкоцитами существенно увеличивается, если предлагаемые антитела связаны с бактериями S.aureus, по сравнению с бактериями S.aureus без этих антител.
Антитела или фрагменты могут быть представлены в комбинации с другими антителами или фрагментами этих других антител, которые нацелены против, по меньшей мере, одного дополнительного эпитопа стафилококка золотистого. Этот дополнительный эпитоп может находиться на антигене, на котором находится эпитоп, т.е. IsaA или IsaB, либо на другом антигене. Использование такой комбинации в качестве лекарственного средства может быть более эффективным, чем использование лекарственного средства, которое содержит только предлагаемые антитела или фрагменты. Это может объясняться высокой изменчивостью S. aureus, которая приводит к различному уровню экспрессии антигенов у различных штаммов, поэтому при помощи комбинации антител или фрагментов можно распознать больше бактерий, чем при помощи одних лишь антител или фрагментов.
Антитела или фрагменты могут быть представлены в комбинации, по меньшей мере, с одним антибиотиком. У человека или животного, которому необходимо лечение лекарственным средством, помимо обычного S.aureus может присутствовать и мутировавший S.aureus. Мутировавший S.aureus может иметь мутировавший IsaA и/или IsaB, который не может быть определен предлагаемыми антителами или фрагментами. В подобном случае против мутировавшего S.aureus может оказаться эффективным антибиотик.
Предлагаемые антитела или фрагменты могут быть представлены в смеси с плазмой крови млекопитающего, в особенности, с плазмой крови человека. Авторы изобретения обнаружили, что предлагаемые антитела или фрагменты, смешанные с плазмой крови, могут оказаться намного эффективнее, чем предлагаемые антитела или фрагменты в физиологическом растворе.
Изобретение также относится к набору, содержащему предлагаемые антитела или фрагменты, для обнаружения S.aureus. Такой набор может использоваться в диагностических целях.
Изобретение также относится к использованию предлагаемых антител или фрагментов для обнаружения, в особенности высокоспецифического обнаружения, S.aureus.
Кроме того, изобретение относится к линии клеток гибридомы, которая продуцирует предлагаемые антитела. Линия клеток гибридомы может представлять собой клеточную линию, хранящуюся в DSMZ под инвентарным номером DSM ACC2987 или DSM ACC2988.
Изобретение также относится к способу лечения человека или животного, которое инфицировано стафилококком золотистым, в особенности метициллин-резистентным или метициллин-чувствительным стафилококком золотистым, или которое подвержено риску инфицирования, когда человеку или животному вводят предлагаемые антитела или фрагменты. Антитела или фрагменты вводятся в дозе, достаточной для снижения количества S.aureus или для удаления S.aureus из организма человека или животного. Антитела или фрагменты могут быть смешаны с подходящим носителем.
У человека или животного может быть мастит или сепсис, вызванный инфекцией. Антитела или фрагменты могут быть представлены в виде комбинации с другими антителами или фрагментами этих других антител, которые нацелены против, по меньшей мере, одного дополнительного эпитопа стафилококка золотистого. Кроме того, антитела или фрагменты могут быть смешаны с плазмой или кровью млекопитающего, в особенности, человека, перед их введением. Антитела или фрагменты могут вводиться системно, в частности, внутривенно, назально или сублингвально. Они также могут вводиться вместе, по меньшей мере, с одним антибиотиком.
ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
На фиг.1а, 1b, 1с показано иммунофлуоресцентное окрашивание штамма S.aureus МА12 (фиг.1а), штамма S.aureus MA12AisaA без IsaA (фиг.1b) и штамма S. aureus Cowan I ∆spa::Tcг без белка А (фиг.1с) с применением предлагаемых антител по данному изобретению.
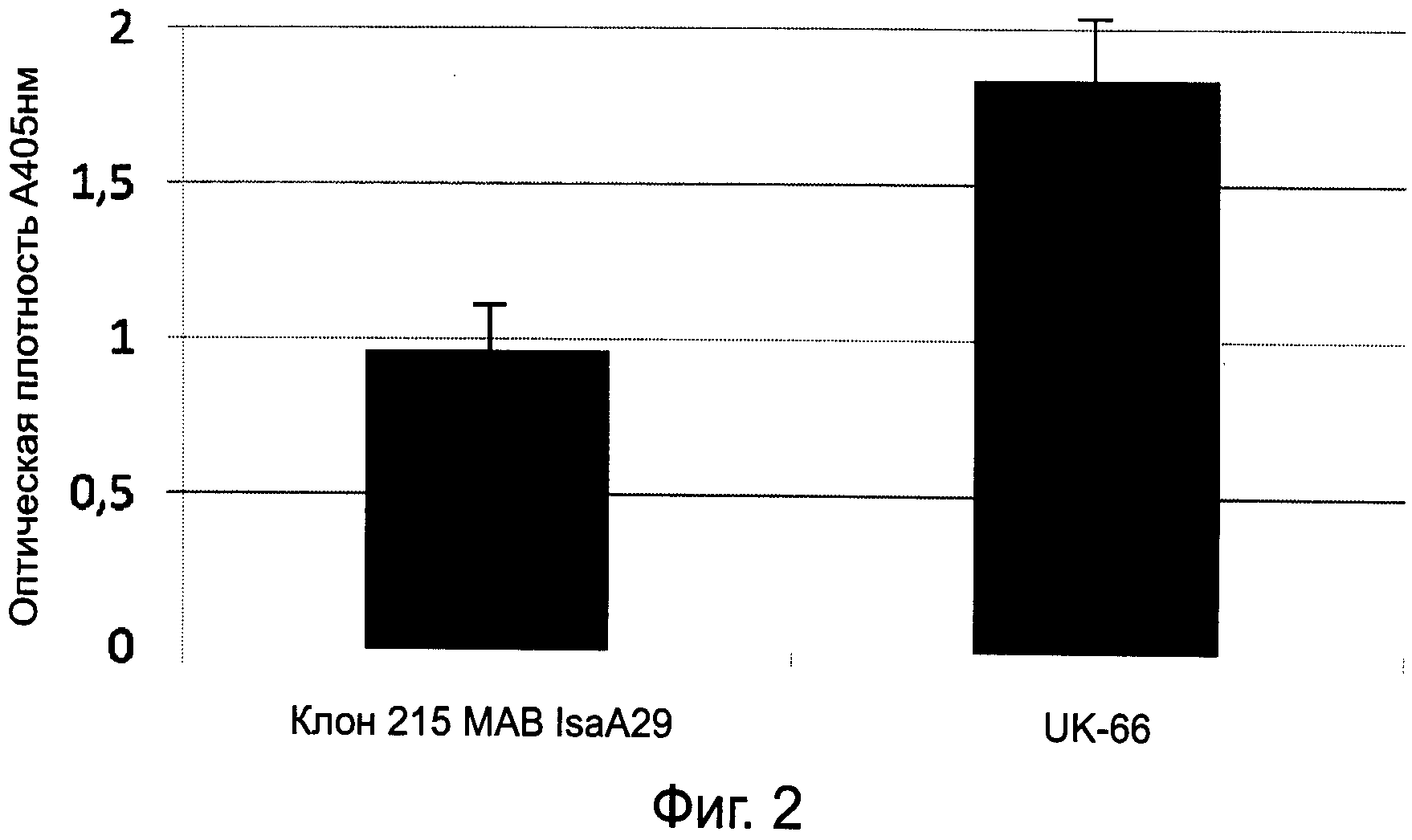
На фиг.2 показаны данные иммуноферментного анализа (ELISA) связывания IsaA с клоном 250 MAB-IsaA29 в сравнении с MAB-UK-66.
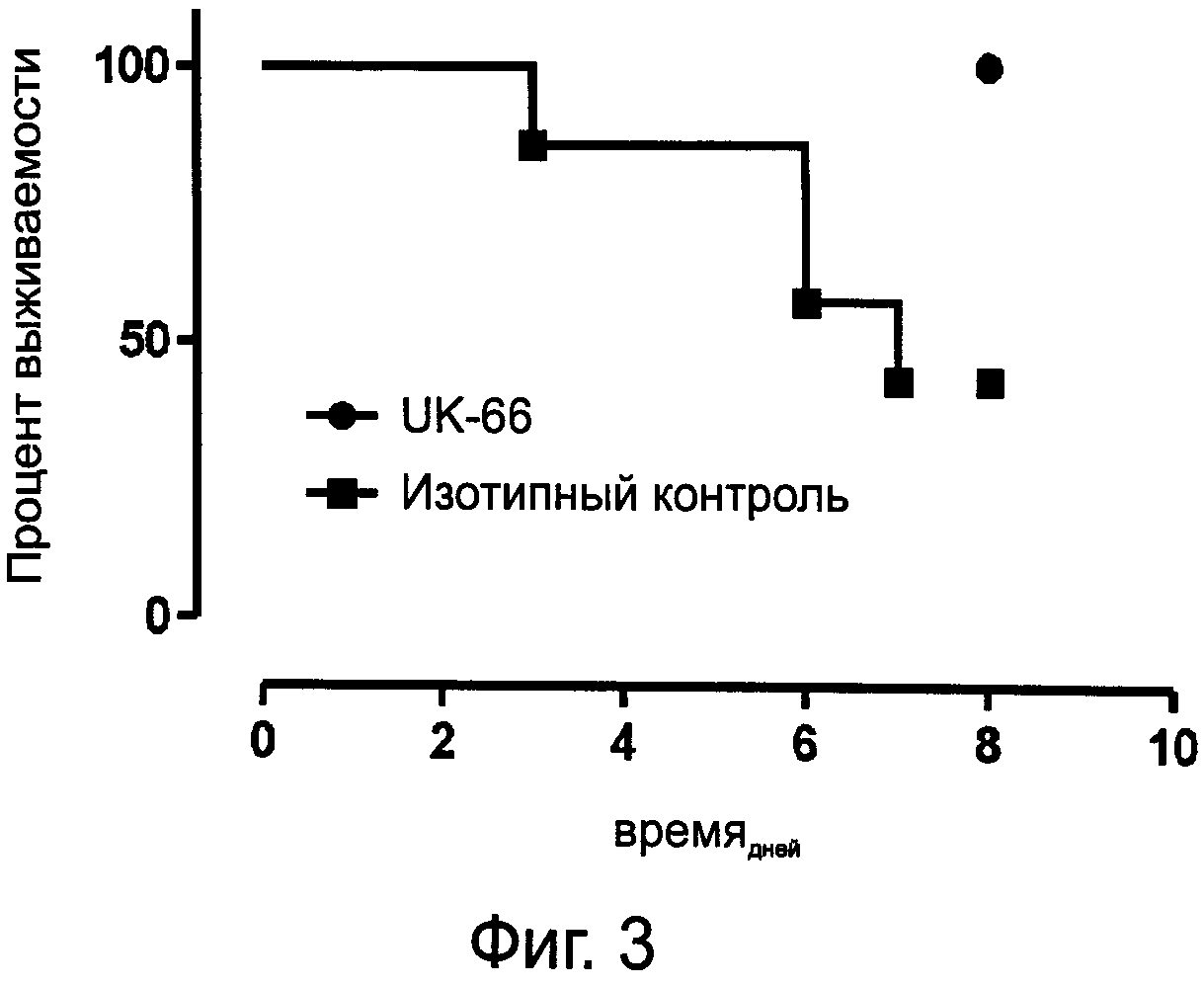
На фиг.3 показана выживаемость мышей после внутривенного введения штамма S.aureus USA300 и внутривенного введения предлагаемых моноклональных антител или изотипных контрольных антител.
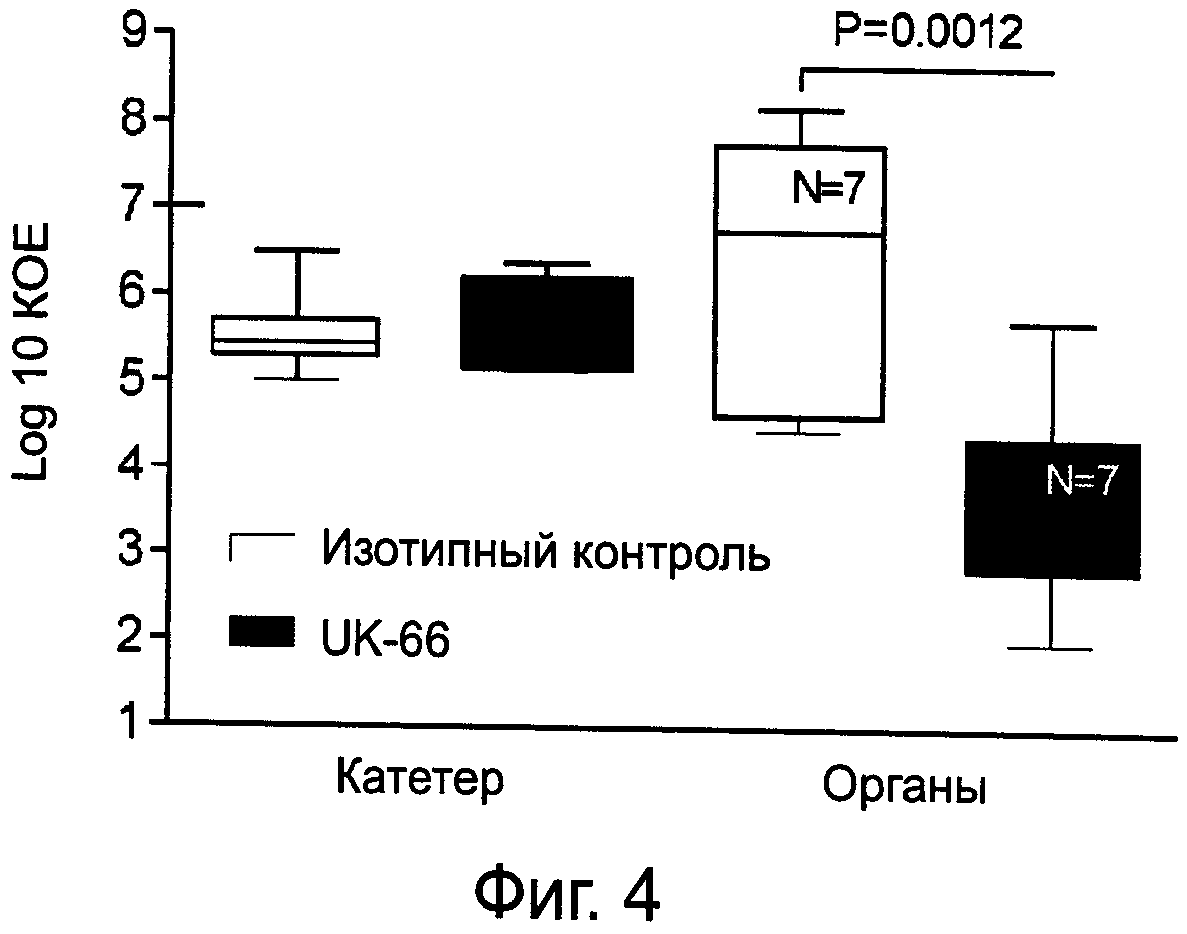
На фиг.4 показано выделение штамма S.aureus MA12 из центрального венозного катетера и органов мышей после введения S. aureus и предлагаемых моноклональных антител.
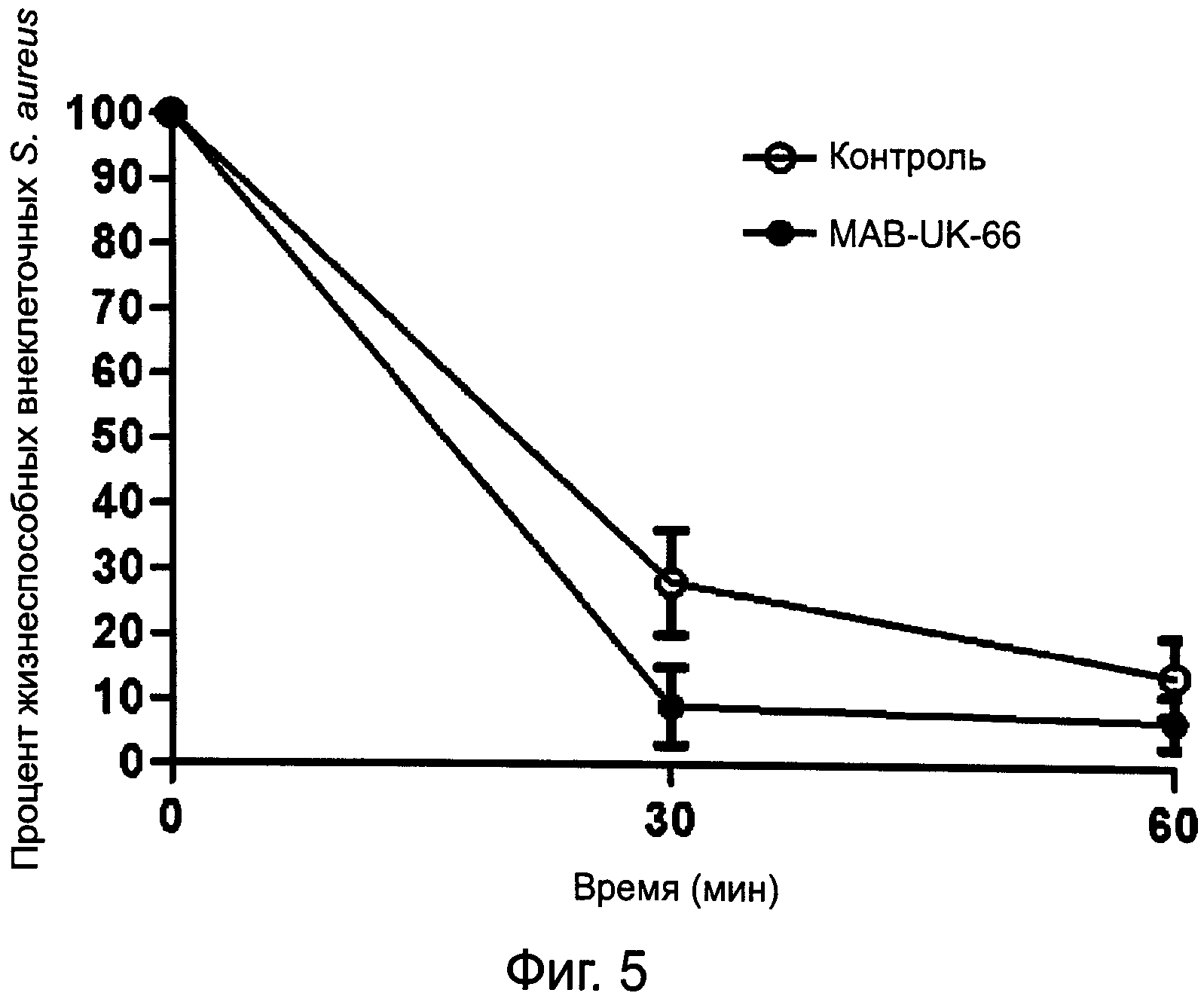
На фиг.5 показан фагоцитоз S.aureus полиморфно-ядерными лейкоцитами в присутствии и в отсутствие предлагаемых антител.
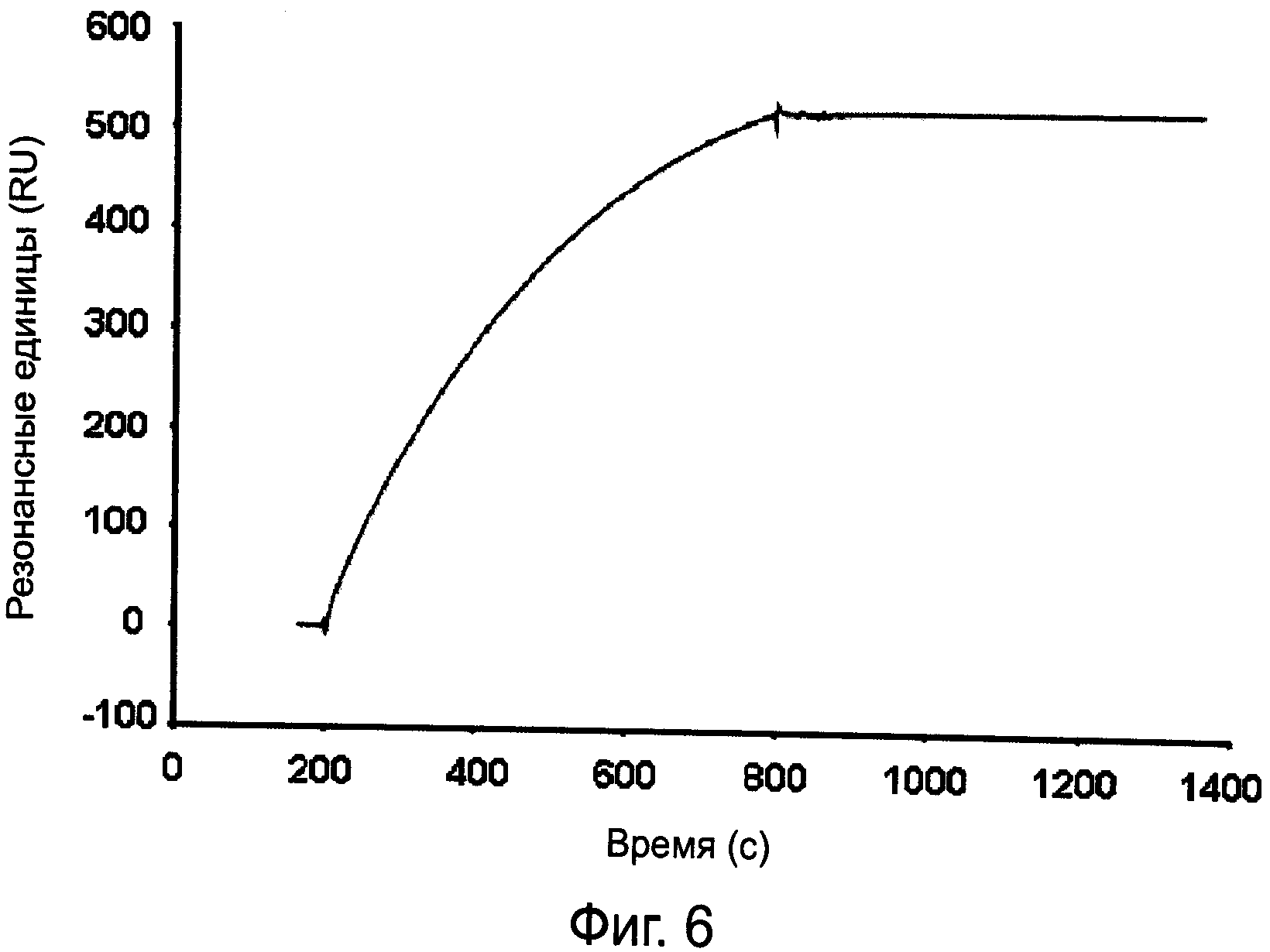
На фиг.6 показана кинетика связывания моноклональных антител MAB-UK-66 с иммобилизованным IsaA.
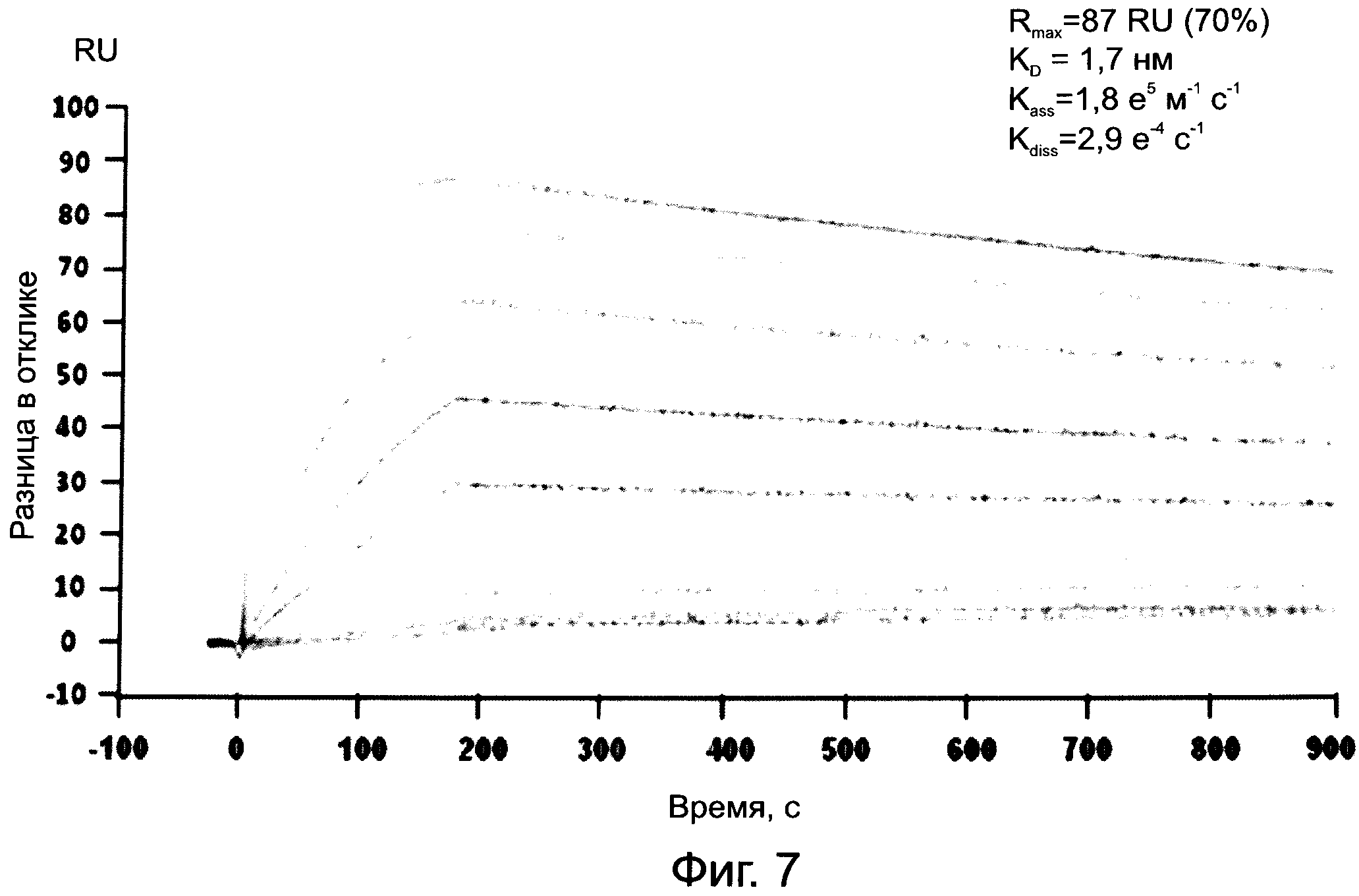
На фиг.7 показана кинетика связывания IsaA с иммобилизованными моноклональными антителами MAB-UK-66.
На фиг.1а-1с показан результат окрашивания с использованием моноклональных антител, направленных против эпитопов IsaA, в качестве первичных антител, которые были произведены линией клеток гибридомы DSM ACC2987 и обозначены MAB-UK-66. Конъюгированные с флуоресцинизотиоцианатом антитела, направленные против мышиного IgG, использовались в качестве вторичных антител.
На фигурах 1а и 1с показано положительное иммунофлуоресцентное окрашивание S.aureus - штаммы MA12 и Cowan I ∆spa::Tcг - при этом на фиг.1b показано отсутствие флуоресцентного окрашивания S.aureus, штамм MA12∆isaA. В отличие от исходных бактерий S.aureus, бактерии S.aureus штамма Cowan I ∆spa::Tcг не продуцируют белок А. Белок А обладает высокой аффинностью к Fc-части антител. Результатом присутствия белка А в бактерии будет сильное неспецифическое связывание первичных и вторичных антител с бактериями. Штамм Cowan I ∆spa::Tcг связывает первичные антитела, указывая на присутствие IsaA, но при этом не наблюдается перекрестной реактивности между антителом и белком А.
На фиг.2 показан результат иммуноферментного анализа (ИФА), который проводился для сравнения связывания с IsaA известного моноклонального антитела MAB-IsaA29, клон 215, и моноклонального антитела MAB-UK-66. Клон 215 MAB-IsaA29 представляет собой моноклональное антитело, описанное в рефератах «Разработка терапии на основе антител, нацеленной против иммунодоминантных антигенов стафилококка золотистого (Staphylococcus aureus)», Оулсен К. и др., страница 128 сборника рефератов, опубликованного по результатам 59-й конференции Немецкого общества гигиены и микробиологии (Deutsche Gesellschaft fur Hygiene und Mikrobiologie e.V.) в сентябре 2007 г., и из статьи Лоренца У. и др. «Терапевтическая эффективность моноклональных антител к стафилококку золотистому (Staphylococcus aureus) на модели сепсиса и абсцесса у мышей», Хирургический форум 2008 г., Springer Berlin Heidelberg, 29 мая 2008 г., выпуск 17, стр.225-226. ИФА (ELISA) проводился следующим образом:
После того, как на ночь каждую лунку планшета для микротитрации покрыли 100 мкл образца рекомбинантного белка IsaA (rIsaA) в концентрации 0,5 мкг/мл в фосфатно-буферном растворе (PBS, рН 7,4), лунки блокировали 1% бычьим сывороточным альбумином на 2 ч. Вышеупомянутые антитела к IsaA развели в пропорции 1 к 4000 и добавили в лунки. После инкубации в течение 1 часа добавили конъюгированный с пероксидазой хрена кроличий противомышиный IgG (DAKO, Глоструп, Дания) и инкубировали в течение 1 часа. Затем добавили субстрат ABTS [2,2'-азинобис(3-этилбензтиазолинсульфоновая KHcnoTa)](Sigma Chemical Co., Дайзенхофен, Германия) и инкубировали в течение 1 часа. Оптическую плотность определяли при длине волны 405 нм при помощи автоматического считывателя микропланшетов. На фиг.1 видно, что связывание MAB-UK-66 и rIsaA намного более интенсивное, чем связывание известного клона 215 антитела MAB-IsaA29 с rIsaA.
Эффективная терапия против S.aureus должна защищать мышей против летального инфицирования S.aureus. Для изучения эффективности предлагаемых антител in vivo была использована следующая модель выживаемости при вызванном S.aureus сепсисе на мышах:
Распределенным по возрастно-половым группам мышам NMRI (Charles River, Зульцфельд, Германия) в день 0 внутривенно ввели 5×108 колониеобразующих единиц (КОЕ) S.aureus USA300 (№АТСС ВАА-1556). Пролеченные мыши получали внутривенно MAB-UK-66 или соответствующее изотипное антитело в качестве контроля (двукратное введение: 15 мг/кг в 100 мкл PBS, рН 7,4, сразу и через 24 часа после введения бактерий). За животными велось наблюдение в течение 8 дней, летальные случаи заболевания документировались.
Значимость защиты измеряли при помощи логарифмического рангового критерия/критерия Кокса-Мантеля: Р=0,022. Результат показан на фиг.3.
Для дальнейшего изучения эффективности предлагаемых антител in vivo применялась следующая модель вызванного S.aureus сепсиса вследствие осложнений катетеризации на мышах:
В эксперименте использовались мыши NMRI (Charles River Wiga Deutschland GmbH, 97633 Зульцфельд, Германия), разделенные на группы по возрасту, полу и массе. Мышам делали интраперитонеальную анестезию ксилазином (8 мг/кг массы тела)/кетамином (100 мг/кг массы тела), после чего делали минимальный кожный разрез с левой стороны выбритой шеи. При помощи операционного микроскопа (Carl Zeiss Jena GmbH, 07745 Иена, Германия) при увеличении 10-16 X изолировали подчелюстную железу, чтобы получить доступ к бифуркации передней и задней лицевой вены. Выполняли венотомию между не затянутыми лигатурами на изолированной передней лицевой вене. Через разрез вставляли и продвигали до верхней полой вены стерильный одноканальный полиэтиленовый катетер (внутренний диаметр 0,28 мм×наружный диаметр 0,6 мм). Лигатуры затягивали, и катетер подкожно направляли и выводили через серединный разрез в лопаточной области. Проверяли проходимость; катетер наполняли раствором гепарина, закрывали заглушкой и оставляли в течение всего эксперимента. Через двадцать четыре часа после операции мышам через катетер вводили 100 мкл суспензии S.aureus, содержащей 1×107 КОЕ штамма МА12 бактерий S.aureus. MA12 представляет собой изолят со слизистой оболочки среднего медицинского персонала, описанный в статье Оулсена К., Зибура В., Коллера К.П., Хелла В., Вишельхауса Т.А. и Хакера Дж. «Влияние субингибиторной концентрации антибиотиков на экспрессию гена альфа-токсина (hia) метициллин-чувствительных и метициллин-резистентных изолятов стафилококка (Staphylococcus)», журнал Antimicrob. Agents Chemother. (1998), 42, стр.2817-2823. Бактериальную суспензию оставляли внутри канала катетера на 15 минут. Затем содержимое катетера смывали в мышей при помощи 0,2 мл физиологического раствора 0,9%. Обработанным мышам вводили антитела, произведенные линией клеток гибридомы DSM ACC2987 (двукратное введение: 15 мг/кг в объеме 100 мкл сразу же и через 24 часа после введения бактерий), внутривенно или физиологический раствор внутривенно (контрольная группа). Массу тела и внешний вид оценивали ежедневно на протяжении эксперимента. Через пять дней после инокуляции мыши подверглись эвтаназии при помощи ингаляции СО2.
У мышей, подвергнувшихся эвтаназии, в асептических условиях был произведен забор органов, которые затем гомогенизировали в 2 мл физиологического раствора. Кроме того, проверяли нахождение катетера в верхней полой вене; затем извлеченный катетер промывали 2 мл физиологического раствора, а промывочную жидкость собирали. Последовательные разведения гомогенатов органов и сборов жидкости из катетеров культивировали на планшетах с маннитоловым солевым агаром с феноловым красным по меньшей мере в течение 48 часов при температуре 37ºС. Производили подсчет колониеобразующих единиц как КОЕ/орган или КОЕ/катетер. Результаты показаны на фиг.4. Данные показывают, что лечение предлагаемыми антителами привело к существенному снижению бактериальной нагрузки на органы.
Для изучения влияния предлагаемых антител на фагоцитоз нейтрофилы человека изолировали при помощи Polymorphprep (Nycomed, Осло, Норвегия) в соответствии с инструкциями производителя. 1×107 КОЕ S.aureus MA12 в 1 мл сбалансированного солевого раствора Хенкса (HBSS) с добавлением 0,1% (масса/объем) желатина (HBSS-gel) и 15% (объем/объем) очищенных антител MAB-UK-66, произведенных линией клеток гибридомы DSM ACC2987, или PBS (контрольный) инкубировали в течение 30 минут при температуре 37ºС на водяной бане-шейкере с медленным взбалтыванием. Равные объемы 5×106 обработанных антителами и PBS S.aureus и 1×106 клеток ПМН/мл HBSS-gel в конечном объеме 1,5 мл инкубировали при температуре 37ºС и медленном взбалтывании. С интервалами от нуля до 60 минут образец этой суспензии отбирали, центрифугировали в течение 4 минут при 250×g и определяли количество бактерий в надосадочной жидкости посредством подсчета числа КОЕ. Фагоцитоз выражается как средние значения трехкратных определений ± СО процентного количества внеклеточных бактерий. Проводили статистический анализ с использованием непараметрического U-критерия Манна-Уитни. Для всех сравнений статистически значимым считалось значение Р<0,05. Значения выражались как средние значения ± СО (SD). Результат на Фиг.5 показывает, что S.aureus фагоцитируется полиморфно-ядерными лейкоцитами независимо от присутствия предлагаемых антител. Однако в присутствии предлагаемых антител процесс фагоцитоза существенно ускорялся по сравнению с контрольными образцами. Антитела действуют как опсонин для фагоцитоза S.aureus полиморфно-ядерными лейкоцитами.
Для определения аффинности моноклональных антител MAB-UK-66 к IsaA определяли кинетику связывания этих антител с иммобилизованным IsaA посредством безмаркерного измерения поверхностного плазменного резонанса с использованием системы BIACORE®2000 (GE Healthcare Europe GmbH, Мунцингер Штрассе 5, 79111 Фрайбург, Германия). Для иммобилизации антигена, IsaA подвергали обработке N-биотином посредством инкубации с эквимолярными концентрациями сульфо-NHS-LC-биотина (Thermo Fisher Scientific, p/a Perbio Science, Аденауэралле 113, 53113 Бонн, Германия). В этих условиях большинство молекул подвергались биотинилированию только в одном участке, и большинство эпитопов, определяемых моноклональными антителами MAB-UK-66, оставались нетронутыми. Иммобилизация антигена на покрытых стрептавидином матрицах чипов-биосенсоров СМ5 осуществлялась как описано в статье Никеля Дж., Котжа А., Себальда В., и Мюллера Т.Д. «Один остаток фактора роста и дифференциации GDF-5 определяет специфичность связывания с рецепторами морфогенетических белков кости типа IB» журнал J. Mol. Biol. (2005), 349, страницы 933-947. Количество иммобилизованного антигена соответствует примерно 100 резонансным единицам [RU], измеренным при помощи системы BIACORE®2000.
Анализы взаимодействия проводились с использованием буфера HBS150 (10 мМ 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота (HEPES) рН7,4, 150 мМ NaCl, 3,4 мМ ЭДТА). Сенсограммы регистрировались при скорости потока 10 мкл/мин при температуре 25ºС. Время ассоциации и диссоциации было установлено на 10 мин. После каждого цикла производили регенерацию чипов при помощи различных растворов (А: 1 мМ СН3СООН, 1 М NaCl, рН 3; В: 4 М MgCl2; С: 1 мМ СН3СООН, 1 М NaCl, 6 М мочевина, рН 3) в течение 2 мин. Кинетика связывания этих антител с иммобилизованным IsaA показана на фиг.6.
Все значения кажущейся аффинности связывания рассчитывались с использованием программного обеспечения Biaevaluation 2.2.4. Значения аффинности взаимодействия kon (<106 M-1s-1) и koff (<10-2 s-1) рассчитывались посредством подстановки данных по кинетике kon и koff в модель связывания Ленгмюра в масштабе 1:1. Таким образом была определена константа диссоциации KD. Константа диссоциации KD показывает аффинность между двумя взаимодействующими молекулами (как например, антитело и соответствующий антиген). Низкое значение KD указывает на высокую аффинность, а высокое значение KD указывает на низкую аффинность. Стандартное отклонение значений KD, определенных указанным образом, составляет меньше 50%. Таким образом, различия в значениях аффинности связывания более чем в два раза считаются значимыми.
В условиях измерений, описанных выше, антитела MAB-UK-66 взаимодействуют с антигенами IsaA массой 29 кДа необратимо ввиду не поддающихся оценке низких скоростей диссоциации. Поскольку оценка константы скорости ограничена 10-5 с-1, наблюдаемая скорость диссоциации должна быть меньше этого значения. Установив скорость диссоциации 10-5 с-1, можно определить, что скорость ассоциации составляет 5,6 105 М-1с-1, что дает значение константы диссоциации 1,8 10-11 М. Таким образом, значение Ко для этого взаимодействия составляет ≤1,8 10-11 М. Важно, что антитело после взаимодействия с антигеном нельзя удалить с поверхности чипа ни одним из вышеупомянутых растворов для регенерации. Это указывает на очень сильное и высокоспецифическое взаимодействие.
Для подтверждения высокой аффинности моноклонального антитела MAB-UK-66 к IsaA кинетика связывания IsaA с иммобилизованными антителами определялась посредством безмаркерного измерения поверхностного плазмонного резонанса с использованием системы BIACORE®2000 (GE Healthcare Europe GmbH, Мунцингер Штрассе 5, 79111 Фрайбург, Германия). Обратимая иммобилизация антитела MAB-UK-66 достигалась за счет использования противомышиного антитела Fc, ковалентно присоединенного с высокой плотностью (18700 резонансных единиц RU) к поверхности датчика СМ5 в соответствии с инструкциями производителя (Набор для захвата мышиных антител, GE Healthcare). Среднее количество захваченных антител MAB-UK-66 на поверхности противомышиного Fc соответствует примерно 640 RU. Чистая поверхность противомышиного Fc использовалась в качестве контрольной поверхности для мониторинга неспецифического связывания и для выполнения опорного вычитания. Анализы взаимодействия выполнялись с использованием буфера HBS-EP (10 мМ HEPES рН 7,4, 150 мМ NaCl, 3 мМ ЭДТА, 0,005% Твин 20). Сенсограммы регистрировались при скорости потока 30 мкл/мин и при температуре 25ºС. Скорость ассоциации и диссоциации была установлена на 3 и 15 мин, соответственно. После каждого цикла поверхности, захватывающие Fc, подвергались регенерации с использованием коротких импульсов глицина 10 мМ с рН 1,7. Кинетика связывания IsaA с иммобилизованными моноклональными антителами MAB-UK-66 показана на Фиг.7.
Значения аффинности и константы скорости ассоциации (kon) и диссоциации (koff) были рассчитаны с использованием программного обеспечения BIAevaluation 4.0.1 посредством подстановки полученных сенсограмм в модель связывания Ленгмюра в масштабе 1:1. Таким образом константа диссоциации KD 1.7 нМ была определена по двум независимым измерениям. Было определено, что константы скорости ассоциации и диссоциации для взаимодействия между MAB-UK-66 и IsaA равняются 1,8 105 М-1с-1 (kon) и 2,9 10-4 с-1 (koff), соответственно.
В описанных выше условиях измерений антитело MAB-UK-66 взаимодействует с антигеном IsaA массой 29 кДа с высокой аффинностью и низкой скоростью диссоциации, подтверждая сильное и высокоспецифическое взаимодействие, как уже было определено связыванием антител MAB-UK-66 с иммобилизованным IsaA.
Для характеристики эпитопа IsaA, который определяется предлагаемыми антителами или фрагментами, было выполнено картирование эпитопа. Для этого были синтезированы олигопептиды длиной 15 аминокислот. Последовательность каждого из олигонуклеотидов идентична последовательности 15 аминокислот IsaA. У каждого нуклеотида 11 аминокислот совпадает с олигонуклеотидом, представляющим идущую далее часть общей последовательности. Последовательности нуклеотидов - это последовательности №9-64 из списка последовательностей.
Каждый нуклеотид был иммобилизован на небольшом участке предметного стекла. Связывание моноклональных антител, секретируемых линией клеток гибридомы DSM ACC2987, и контрольных антител с этими участками было изучено во время инкубации с этими антителами посредством связывания меченных флуоресцентными маркерами вторичных антител и определения интенсивности флуоресценции. Результаты приведены в таблице ниже:
Таблица
|
|
Как видно из приведенной выше таблицы, последовательности №15, 17-19, 21-26, 32-34 и 57 (SEQ ID No. 15, 17-19, 21-26, 32-34 и 57) участвуют в связывании эпитопов с антителами, секретируемыми клеточной линией DSM ACC2987.