Результат интеллектуальной деятельности: СВЯЗЫВАЮЩАЯ МСР-1 НУКЛЕИНОВАЯ КИСЛОТА И ЕЕ ПРИМЕНЕНИЕ
Вид РИД
Изобретение
Настоящее изобретение относится к нуклеиновым кислотам, которые связываются с MCP-1, и к их применению для получения лекарственного средства и диагностического средства, соответственно.
MCP-1 (хемоаттрактантный белок моноцитов-1; альтернативные названия: MCAF [хемоаттрактантный и активирующий фактор моноцитов]; CCL2; SMC-CF [колониестимулирующий фактор гладкомышечных клеток]; HC-11; LDCF; GDCF; TSG-8; SCYA2; A2; код доступа SwissProt: P13500) человека был охарактеризован независимо тремя исследовательскими группами (Matsushima 1988; Rollins 1989; Yoshimura 1989). Он состоит из 76 аминокислот и, подобно всем хемокинам, характеризуется участком связывания гепарина. Две внутримолекулярных дисульфидных связи обеспечивают стабильную жесткую структуру молекулы. Более того, MCP-1 имеет пироглутамат на N-конце. В участке Thr 71 расположен вероятный участок O-связанного гликозилирования. Существуют другие члены семейства MCP, как у человека (MCP-2, -3, -4), так и у мышей (MCP-2, -3, -5). Белки человека приблизительно на 70% гомологичны MCP-1 человека.
Структура MCP-1 была выполнена с помощью ЯМР (Handel 1996) и рентгенограммы (Lubkowski 1997). Мономер MCP-1 имеет характерную для хемокинов укладку, при которой за N-концевыми остатками цистеина следует длинная петля, которая образует три встречно параллельные β-складчатые слои в повторе "Greek key". Белок заканчивается α-спиралью, которая расположена над тремя β-слоями (код доступа данных PDB: 1DOK).
Несмотря на то, что трехмерная структура форм MCP-1 различных видов млекопитающих в основном сохраняется, аминокислотная последовательность в ходе эволюции сохранилась в меньшей степени. При выравнивании последовательностей общее сходство последовательностей составляет 55%, между MCP-1 (также называемым JE) человека и мыши в участке первых 76 аминокислот. Кроме аминокислотной последовательности, MCP-1 мыши отличается от MCP-1 человека размером молекулы (125 аминокислоты) и степенью гликозилирования. MCP-1 мыши содержит С-концевой домен из 49 аминокислот, который отсутствует у MCP-1 человека, и не требуется для биологической активности in vitro. MCP-1 человека имеет следующий процент аминокислотной идентичности с MCP-1 следующих видов:
|
С учетом высокой степени дивергенции, для успешного проведения фармакологических испытаний на моделях грызунов может возникнуть необходимость в получении антагонистов MCP-1 грызунов.
MCP-1 является сильным аттрактантом моноцитов/макрофагов, базофилов, активированных T-клеток и NK-клеток. Широкое множество типов клеток, таких как эндотелиальные клетки, эпителиальные клетки, фибробласты, кератиноциты, синовиальные клетки, мезангиальные клетки, остеобласты, гладкомышечные клетки, а также множество опухолевых клеток, экспрессируют MCP-1 (Baggiolini 1994). Его экспрессию стимулирует несколько типов провоспалительных агентов, таких как IL-1β, TNF-α, IFN-γ, LPS (липополисахарид) и GM-CSF.
Довольно необычный в разнородной сети хемокинов, MCP-1 является высоко специфичным в отношении использования рецепторов, так как связывается с высокой аффинностью только с рецептором хемокинов CCR2. Подобно всем рецепторам хемокинов, CCR2 является сопряженным с G-белком рецептором (GPCR) (Dawson 2003). CCR2, по-видимому, экспрессируется в виде двух немного отличающихся формах, CCR2a и CCR2b, образующихся в результате альтернативного сплайсинга мРНК, кодирующей С-концевую область (Charo 1994). Эти рецепторы экспрессируются на моноцитах, миелоидных клетках-предшественниках и активированных T-клетках (Myers 1995; Qin 1996). Константа диссоциации MCP-1 и рецептора, трансфицированного в клетки HEK-293, составляет 260 пМ, что согласуется со значениями, измеренными на моноцитах (Myers 1995; Van Riper 1993). Активация CCR2b на трансфицированных клетках HEK-293 посредством MCP-1 ингибирует аденилилциклазу в концентрации 90 пМ, и мобилизует внутриклеточный кальций при несколько более высоких концентрациях, по-видимому, независимо от гидролиза фосфатидилинозитола. Эффекты аденилилциклазы и внутриклеточного высвобождения кальция жестко ингибируются токсином коклюша, что указывает на вовлечение в передачу сигнала гетеротримерных G-белков Gi-типа (Myers 1995).
MCP-1 связан с привлечением моноцитов в воспаленные ткани. Там, оседлые макрофаги высвобождают хемокины, такие как MCP-1 и другие хемокины, и цитокины, такие как TNF, IL-1β и другие цитокины, которые активируют экспрессию ряда молекул адгезии эндотелиальными клетками. В результате этого процесса, моноциты катятся по поверхности "липкого" эндотелия. В этот момент, моноциты встречаются с MCP-1, находящимся на поверхности эндотелия, и МСР-1 связывается с CCR2 моноцитов, активируя их. Такое связывание в конечном итоге приводит к полной остановке моноцитов, распространению их по эндотелию, и миграции в окружающую ткань, где моноциты дифференцируются до макрофагов и мигрируют в направлении области максимальной концентрации MCP-1.
MCP-1 является членом семейства хемокинов, которое представляет собой семейство низкомолекулярных (приблизительно 8-14 кДа) связывающих гепарин, в большинстве случаев основных и структурно родственных, молекул. Они образуются, главным образом, в воспаленных тканях, и регулируют привлечение, активацию и пролиферацию белых клеток крови (лейкоцитов) (Baggiolini 1994; Springer 1995; Schall 1994). Хемокины селективно индуцируют хемотаксис нейтрофилов, эозинофилов, базофилов, моноцитов, макрофагов, тучных клеток, T- и B-клеток. Кроме хемотактического эффекта, они могут селективно оказывать другие эффекты в отвечающих клетках, например, изменять форму клеток, временно повышать концентрацию свободных внутриклеточных ионов кальция, обеспечивать дегрануляцию, активацию интегринов, образование биологически активных липидов, таких как лейкотриены, простагландины, тромбоксаны, или дыхательный взрыв (высвобождение активных форм кислорода для разрушения патогенных организмов или опухолевых клеток). Таким образом, провоцируя высвобождение дополнительных провоспалительных медиаторов, хемотаксис и экстравазацию лейкоцитов в инфицированной области или в области воспаления, хемокины стимулируют усиление воспалительной реакции.
По расположению первых двух из четырех консервативных остатков цистеина, хемокины разделяют на четыре класса: CC или β-хемокины, в которых цистеины расположены тандемно, CXC или α-хемокины, где они разделены одним дополнительным аминокислотным остатком, XC или γ-хемокины, единственным представителем которых на сегодняшний день является лимфотактин, которые обладают лишь одним дисульфидным мостиком, и CX3C-хемокины, которые характеризуются тремя аминокислотными остатками между остатками цистеина, и единственным представителем которых на сегодняшний день является мембраносвязанный фракталкин (Bazan 1997).
CXC-хемокины, в частности, те CXC-хемокины, которые обладают аминокислотной последовательностью ELR на их N-конце, действуют, главным образом, на нейтрофилы. Примерами CXC-хемокинов, которые являются активными в отношении нейтрофилов, являются IL-8, GROα, -β и -γ, NAP-2, ENA-78 и GCP-2. CC-хемокины действуют на широкое множество лейкоцитов, такие как моноциты, макрофаги, эозинофилы, базофилы, а также T- и B-лимфоциты (Oppenheim 1991; Baggiolini 1994; Miller 1992; Jose 1994; Ponath 1996a). Их примерами являются I-309; MCP-1, -2, -3, -4, MIP-1α и -β, RANTES и эотаксин.
Хемокины действуют посредством рецепторов, которые относят к суперсемейству сопряженных с G-белком рецепторов с семью трансмембранными доменами (GPCR; Murphy 2000). В целом, взаимодействия хемокинов и рецепторов хемокинов обычно бывает разнородным в том смысле, что один хемокин может связывать множество рецепторов хемокинов и, наоборот, один рецептор хемокинов может взаимодействовать с несколькими хемокинами. Некоторые известные рецепторы CC-хемокинов включают CCR1, который связывает MIP-1α и RANTES (Neote 1993; Gao 1993); CCR2, который связывает хемокины, включая MCP-1, -2, -3 и -4 (Charo 1994; Myers 1995; Gong 1997; Garcia-Zepeda 1996); CCR3, который связывает хемокины, включая эотаксин, RANTES и MCP-3 (Ponath 1996b); CCR4, который, как было выявлено, передает сигнал в ответ на MCP-1, MIP-1α и RANTES (Power 1995); и CCR5, который, как было показано, передает сигнал в ответ на MIP-1α и -β и RANTES (Boring 1996; Raport 1996; Samson 1996).
Как указано выше, все четыре члена семейства MCP (1-4) связываются с CCR2, в то время как MCP-2, MCP-3 и MCP-4 также могут взаимодействовать с CCR1 и CCR3 (Gong 1997; Heath 1997; Uguccioni 1997) а, в случае MCP-2, с CCR5 (Ruffing 1998). Другим CC-хемокином, демонстрирующим высокую гомологию с семейством MCP, является эотаксин, который первоначально был выделен из жидкости бронхоальвеолярного лаважа, взятого у сенсибилизированных морских свинок, стимулированных аллергеном (Jose 1994). Было показано, что эотаксин также способен активировать CCR2 (Martinelli 2001).
Проблемой, с которой связано создание настоящего изобретения, является получение средства, которое специфично взаимодействует с MCP-1 и которое может использоваться для профилактики и/или лечения хронического заболевания и хронического нарушения, соответственно. Более конкретно, проблемой, с которой связано создание настоящего изобретения, является получение средств на основе нуклеиновой кислоты, которая специфично взаимодействует с MCP-1 и которая может использоваться для профилактики и/или лечения хронического заболевания и хронического нарушения, соответственно.
Еще одной проблемой, с которой связано создание настоящего изобретения, является разработка способа получения диагностического средства для лечения заболевания, которое представляет собой хроническое заболевание и хроническое нарушение, соответственно.
В связи с указными выше проблемами, хроническое заболевание и хроническое нарушение, соответственно, предпочтительно, представляет собой хроническое респираторное заболевание, хроническое заболевание почек и системную красную волчанку.
Эти и другие проблемы, с которыми связано создание настоящего изобретения, решают благодаря объекту изобретения, представленным в независимых пунктах формулы изобретения. Предпочтительные варианты осуществления представлены в зависимых пунктах формулы изобретения.
Проблема, с которой связано создание настоящего изобретения, решается благодаря объектам независимых пунктов формулы изобретения. Предпочтительные варианты осуществления представлены в зависимых пунктах формулы изобретения.
Более конкретно, в первом аспекте, который также является первым вариантом осуществления указанного аспекта, проблема, с которой связано создание настоящего изобретения, решается с помощью молекулы нуклеиновой кислоты, способной связываться с MCP-1, причем молекула нуклеиновой кислоты предназначена для использования в качестве лекарственного средства для лечения и/или профилактики хронического заболевания или хронического нарушения, предпочтительно, выбранного из группы, состоящей из хронического респираторного заболевания, хронического заболевания почек и системной красной волчанки.
Во втором аспекте, который также является первым вариантом осуществления указанного аспекта, проблема, с которой связано создание настоящего изобретения, решается с помощью молекулы нуклеиновой кислоты, способной связываться с MCP-1, причем молекула нуклеиновой кислоты предназначена для использования в качестве диагностического средства для диагностики хронического заболевания или хронического нарушения, предпочтительно, выбранного из группы, состоящей из хронического респираторного заболевания, хронического заболевания почек и системной красной волчанки.
Во втором варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого варианта осуществления первого аспекта и второго аспекта, хроническое респираторное заболевание выбрано из группы, состоящей из пневмонии, воспаления легких и плевры, плеврита, плеврального выпота, пневмонии при волчанке, хронической диффузной интерстициальной болезни легких, эмболии легких, кровоизлияния в легкие, синдрома сморщенного легкого, легочной гипертензии и хронической обструктивной болезни легких и их комбинаций.
В третьем варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого и второго варианта осуществления первого аспекта и второго аспекта, легочная гипертензия выбрана из группы, состоящей из легочной гипертензии, связанной с болезнью левого отдела сердца, легочной гипертензии, связанной с заболеваниями легких и/или гипоксемией, легочной гипертензии вследствие хронического тромботического и/или эмболического заболевания, гипертензии легочной артерии, предпочтительно, идиопатической гипертензии легочной артерии, гипертензии легочной артерии, связанной с коллагенозом, семейной гипертензии легочной артерии, гипертензии легочной артерии, связанной с другими заболеваниями, и гипертензии легочной артерии, связанной с заболеваниями вен и капилляров.
В четвертом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого, второго и третьего варианта осуществления первого аспекта и второго аспекта, хроническая обструктивная болезнь легких представляет собой хроническую обструктивную болезнь легких с вовлечением сосудов легких или без такого вовлечения.
В пятом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого, второго, третьего и четвертого варианта осуществления первого аспекта и второго аспекта, хроническая обструктивная болезнь легких выбрана из группы, состоящей из хронического бронхита и эмфиземы.
В шестом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого варианта осуществления первого аспекта и второго аспекта, хроническое заболевание почек выбрано из группы, состоящей из волчаночного нефрита, мембрано-пролиферативного гломерулонефрита, мембранозного гломерулонефрита, IgA-нефропатии, пост-стрептококкового гломерулонефрита, быстро прогрессирующего гломерулонефрита, нефритического синдрома, фокального сегментного гломерулосклероза, диабетической нефропатии, нефротического синдрома и нефротического синдрома, предпочтительно, волчаночного нефрита.
В седьмом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого, пятого и шестого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота выбрана из группы, содержащей нуклеиновые кислоты типа 1A, нуклеиновые кислоты типа 1B, нуклеиновые кислоты типа 2, нуклеиновые кислоты типа 3, нуклеиновые кислоты типа 4 и нуклеиновые кислоты, имеющие последовательность нуклеиновой кислоты согласно любой из SEQ ID NO:87-115.
В восьмом варианте осуществления первого и второго аспекта, который также является вариантом осуществления седьмого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота типа 2 содержит в направлении 5'->3' первый участок Бокс B1A, второй участок Бокс B2 и третий участок Бокс B1B, где
первый участок Бокс B1A и третий участок Бокс B1B необязательно гибридизуются друг с другом, причем при гибридизации образуется двухцепочечная структура,
первый участок Бокс B1A содержит нуклеотидную последовательность, выбранную из группы, содержащей ACGCA, CGCA и GCA,
второй участок Бокс B2 содержит нуклеотидную последовательность CSUCCCUCACCGGUGCAAGUGAAGCCGYGGCUC, и
третий участок Бокс B1B содержит нуклеотидную последовательность, выбранную из группы, содержащей UGCGU, UGCG и UGC.
В девятом варианте осуществления первого и второго аспекта, который также является вариантом осуществления восьмого варианта осуществления первого аспекта и второго аспекта,
второй участок Бокс B2 содержит нуклеотидную последовательность CGUCCCUCACCGGUGCAAGUGAAGCCGUGGCUC.
В десятом варианте осуществления первого и второго аспекта, который также является вариантом осуществления восьмого и девятого варианта осуществления первого аспекта и второго аспекта,
a) первый участок Бокс B1A содержит нуклеотидную последовательность ACGCA, и
третий участок Бокс B1B содержит нуклеотидную последовательность UGCGU; или
b) первый участок Бокс B1A содержит нуклеотидную последовательность CGCA, и
третий участок Бокс B1B содержит нуклеотидную последовательность UGCG; или
c) первый участок Бокс B1A содержит нуклеотидную последовательность GCA, и
третий участок Бокс B1B содержит нуклеотидную последовательность UGC или UGCG.
В одиннадцатом варианте осуществления первого и второго аспекта, который также является вариантом осуществления восьмого, девятого и десятого варианта осуществления первого аспекта и второго аспекта,
первый участок Бокс B1A содержит нуклеотидную последовательность GCA.
В двенадцатом варианте осуществления первого и второго аспекта, который также является вариантом осуществления восьмого, девятого, десятого и одиннадцатого варианта осуществления первого аспекта и второго аспекта, предпочтительно, одиннадцатого варианта осуществления первого и второго аспекта
третий участок Бокс B1B содержит нуклеотидную последовательность UGCG.
В 13-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления восьмого, девятого, десятого, одиннадцатого и двенадцатого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота содержит последовательность нуклеиновой кислоты согласно SEQ ID NO:37, SEQ ID NO:116, SEQ ID NO:117 и SEQ ID NO:278.
В 14-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого, пятого, шестого и седьмого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота типа 3 содержит в направлении 5'->3' первый участок Бокс B1A, второй участок Бокс B2A, третий участок Бокс B3, четвертый участок Бокс B2B, пятый участок Бокс B4, шестой участок Бокс B5A, седьмой участок Бокс B6, восьмой участок Бокс B5B и девятый участок Бокс B1B, где
первый участок Бокс B1A и девятый участок Бокс B1B необязательно гибридизуются друг с другом, причем при гибридизации образуется двухцепочечная структура,
второй участок Бокс B2A и четвертый участок Бокс B2B необязательно гибридизуются друг с другом, причем при гибридизации образуется двухцепочечная структура,
шестой участок Бокс B5A и восьмой участок Бокс B5B необязательно гибридизуются друг с другом, причем при гибридизации образуется двухцепочечная структура,
первый участок Бокс B1A содержит нуклеотидную последовательность, которая выбрана из группы, содержащей GURCUGC, GKSYGC, KBBSC и BNGC,
второй участок Бокс B2A содержит нуклеотидную последовательность GKMGU,
третий участок Бокс B3 содержит нуклеотидную последовательность KRRAR,
четвертый участок Бокс B2B содержит нуклеотидную последовательность ACKMC,
пятый участок Бокс B4 содержит нуклеотидную последовательность, выбранную из группы, содержащей CURYGA, CUWAUGA, CWRMGACW и UGCCAGUG,
шестой участок Бокс B5A содержит нуклеотидную последовательность, выбранную из группы, содержащей GGY и CWGC,
седьмой участок Бокс B6 содержит нуклеотидную последовательность, выбранную из группы, содержащей YAGA, CKAAU и CCUUUAU,
восьмой участок Бокс B5B содержит нуклеотидную последовательность, выбранную из группы, содержащей GCYR и GCWG, и
девятый участок Бокс B1B содержит нуклеотидную последовательность, выбранную из группы, содержащей GCAGCAC, GCRSMC, GSVVM и GCNV.
В 15-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого варианта осуществления первого аспекта и второго аспекта,
третий участок Бокс B3 содержит нуклеотидную последовательность GAGAA или UAAAA.
В 16-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого и 15-ого варианта осуществления первого аспекта и второго аспекта,
пятый участок Бокс B4 содержит нуклеотидную последовательность CAGCGACU или CAACGACU.
В 17-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого и 16-ого варианта осуществления первого аспекта и второго аспекта,
пятый участок Бокс B4 содержит нуклеотидную последовательность CAGCGACU, и Бокс B3 содержит нуклеотидную последовательность UAAAA.
В 18-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого и 16-ого варианта осуществления первого аспекта и второго аспекта,
пятый участок Бокс B4 содержит нуклеотидную последовательность CAACGACU, и третий участок Бокс B3 содержит нуклеотидную последовательность GAGAA.
В 19-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого, 16-ого, 17-ого и 18-ого варианта осуществления первого аспекта и второго аспекта,
седьмой участок Бокс B6 содержит нуклеотидную последовательность UAGA.
В 20-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого, 16-ого, 17-ого, 18-ого и 19-ого варианта осуществления первого аспекта и второго аспекта,
a) первый участок Бокс B1A содержит нуклеотидную последовательность GURCUGC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GCAGCAC; или
b) первый участок Бокс B1A содержит нуклеотидную последовательность GKSYGC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GCRSMC; или
c) первый участок Бокс B1A содержит нуклеотидную последовательность KBBSC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GSVVM; или
d) первый участок Бокс B1A содержит нуклеотидную последовательность BNGC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GCNV.
В 21-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 20-ого варианта осуществления первого аспекта и второго аспекта,
a) первый участок Бокс B1A содержит нуклеотидную последовательность GUGCUGC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GCAGCAC; или
b) первый участок Бокс B1A содержит нуклеотидную последовательность GUGCGC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GCGCAC; или
c) первый участок Бокс B1A содержит нуклеотидную последовательность KKSSC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GSSMM; или
d) первый участок Бокс B1A содержит нуклеотидную последовательность SNGC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GCNS.
В 22-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 21-ого варианта осуществления первого аспекта и второго аспекта,
первый участок Бокс B1A содержит нуклеотидную последовательность GGGC, и
девятый участок Бокс B1B содержит нуклеотидную последовательность GCCC.
В 23-ем варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого, 16-ого, 17-ого, 18-ого, 19-ого, 20-ого, 21-ого и 22-ого варианта осуществления первого аспекта и второго аспекта, второй участок Бокс B2A содержит нуклеотидную последовательность GKMGU, и четвертый участок Бокс B2B содержит нуклеотидную последовательность ACKMC.
В 24-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 23-ого варианта осуществления первого аспекта и второго аспекта, второй участок Бокс B2A содержит нуклеотидную последовательность GUAGU, и четвертый участок Бокс B2B содержит нуклеотидную последовательность ACUAC.
В 25-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого, 16-ого, 17-ого, 18-ого, 19-ого, 20-ого, 21-ого, 22-ого, 23-его и 24-ого варианта осуществления первого аспекта и второго аспекта,
a) шестой участок Бокс B5A содержит нуклеотидную последовательность GGY, и
восьмой участок Бокс B5B содержит нуклеотидную последовательность GCYR; или
b) шестой участок Бокс B5A содержит нуклеотидную последовательность CWGC, и
восьмой участок Бокс B5B содержит нуклеотидную последовательность GCWG.
В 26-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 25-ого варианта осуществления первого аспекта и второго аспекта,
шестой участок Бокс B5A содержит нуклеотидную последовательность GGC, и
восьмой участок Бокс B5B содержит нуклеотидную последовательность GCCG.
В 27-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого, 16-ого, 17-ого, 18-ого, 19-ого, 20-ого, 21-ого, 22-ого, 23-его, 24-ого, 25-ого и 26-ого варианта осуществления первого аспекта и второго аспекта, предпочтительно, 25-ого и 26-ого варианта осуществления первого и второго аспекта, шестой участок Бокс B5A гибридизуется с нуклеотидами GCY восьмого участка Бокс B5B.
В 28-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого, 16-ого, 17-ого и 19-ого, 20-ого, 21-ого, 22-ого, 23-его, 24-ого, 25-ого, 26-ого и 27-ого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота содержит последовательность нуклеиновой кислоты согласно SEQ ID NO:56.
В 29-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 14-ого, 15-ого, 16-ого и 18-ого, 19-ого, 20-ого, 21-ого, 22-ого, 23-его, 24-ого, 25-ого, 26-ого и 27-ого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота содержит последовательность нуклеиновой кислоты, выбранную из группы, содержащей последовательности нуклеиновых кислот согласно SEQ ID NO:57-61, SEQ ID NO:67-71 и SEQ ID NO:73.
В 30-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого, пятого, шестого и седьмого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота типа 4 содержит в направлении 5'->3' первый участок Бокс B1A, второй участок Бокс B2, третий участок Бокс B1B, где
первый участок Бокс B1A и третий участок Бокс B1B необязательно гибридизуются друг с другом, причем при гибридизации образуется двухцепочечная структура,
первый участок Бокс B1A содержит нуклеотидную последовательность, выбранную из группы, содержащей AGCGUGDU, GCGCGAG, CSKSUU, GUGUU и UGUU;
второй участок Бокс B2 содержит нуклеотидную последовательность, выбранную из группы, содержащей AGNDRDGBKGGURGYARGUAAAG, AGGUGGGUGGUAGUAAGUAAAG и CAGGUGGGUGGUAGAAUGUAAAGA, и
третий участок Бокс B1B содержит нуклеотидную последовательность, выбранную из группы, содержащей GNCASGCU, CUCGCGUC, GRSMSG, GRCAC и GGCA.
В 31-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 30-ого варианта осуществления первого аспекта и второго аспекта,
a) первый участок Бокс B1A содержит нуклеотидную последовательность GUGUU, и
третий участок Бокс B1B содержит нуклеотидную последовательность GRCAC;
b) первый участок Бокс B1A содержит нуклеотидную последовательность GCGCGAG, и
третий участок Бокс B1B содержит нуклеотидную последовательность CUCGCGUC; или
c) первый участок Бокс B1A содержит нуклеотидную последовательность CSKSUU, и
третий участок Бокс B1B содержит нуклеотидную последовательность GRSMSG, или
d) первый участок Бокс B1A содержит нуклеотидную последовательность UGUU, и
третий участок Бокс B1B содержит нуклеотидную последовательность GGCA, или
e) первый участок Бокс B1A содержит нуклеотидную последовательность AGCGUGDU, и
третий участок Бокс B1B содержит нуклеотидную последовательность GNCASGCU.
В 32-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 31-ого варианта осуществления первого аспекта и второго аспекта, первый участок Бокс B1A содержит нуклеотидную последовательность CSKSUU, и третий участок Бокс B1B содержит нуклеотидную последовательность GRSMSG.
В 33-ем варианте осуществления первого и второго аспекта, который также является вариантом осуществления 32-ого варианта осуществления первого аспекта и второго аспекта, первый участок Бокс B1A содержит нуклеотидную последовательность CCGCUU, и третий участок Бокс B1B содержит нуклеотидную последовательность GGGCGG.
В 34-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 30-ого, 31-ого, 32-ого и 33-его варианта осуществления первого аспекта и второго аспекта,
второй участок Бокс B2 содержит нуклеотидную последовательность AGGUGGGUGGUAGUAAGUAAAG.
В 35-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 30-ого, 31-ого, 32-ого, 33-его и 34-ого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота содержит последовательность нуклеиновой кислоты согласно SEQ ID NO:80 и SEQ ID NO:81.
В 36-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого, пятого, шестого и седьмого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота типа 1A содержит в направлении 5'->3' первый участок Бокс B1A, второй участок Бокс B2, третий участок Бокс B3, четвертый участок Бокс B4, пятый участок Бокс B5, шестой участок Бокс B6 и седьмой участок Бокс B1B, где
первый участок Бокс B1A и седьмой участок Бокс B1B необязательно гибридизуются друг с другом, причем при гибридизации образуется двухцепочечная структура,
первый участок Бокс B1A содержит нуклеотидную последовательность AGCRUG,
второй участок Бокс B2 содержит нуклеотидную последовательность CCCGGW,
третий участок Бокс B3 содержит нуклеотидную последовательность GUR,
четвертый участок Бокс B4 содержит нуклеотидную последовательность RYA,
пятый участок Бокс B5 содержит нуклеотидную последовательность GGGGGRCGCGAYC
шестой участок Бокс B6 содержит нуклеотидную последовательность UGCAAUAAUG или URYAWUUG, и
седьмой участок Бокс B1B содержит нуклеотидную последовательность CRYGCU.
В 37-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого варианта осуществления первого аспекта и второго аспекта,
первый участок Бокс B1A содержит нуклеотидную последовательность AGCGUG.
В 38-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого и 37-ого варианта осуществления первого аспекта и второго аспекта,
второй участок Бокс B2 содержит нуклеотидную последовательность CCCGGU.
В 39-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого, 37-ого и 38-ого варианта осуществления первого аспекта и второго аспекта,
третий участок Бокс B3 содержит нуклеотидную последовательность GUG.
В 40-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого, 37-ого, 38-ого и 39-ого варианта осуществления первого и второго аспекта
четвертый участок Бокс B4 содержит нуклеотидную последовательность GUA.
В 41-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого, 37-ого, 38-ого, 39-ого и 40-ого варианта осуществления первого аспекта и второго аспекта,
пятый участок Бокс B5 содержит нуклеотидную последовательность GGGGGGCGCGACC.
В 42-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого, 37-ого, 38-ого, 39-ого, 40-ого и 41-ого варианта осуществления первого аспекта и второго аспекта,
шестой участок Бокс B6 содержит нуклеотидную последовательность UACAUUUG.
В 43-ем варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого, 37-ого, 38-ого, 39-ого, 40-ого, 41-ого и 42-ого варианта осуществления первого аспекта и второго аспекта,
седьмой участок Бокс B1B содержит нуклеотидную последовательность CACGCU.
В 44-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 36-ого, 37-ого, 38-ого, 39-ого, 40-ого, 41-ого, 42-ого и 43-его варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота содержит последовательность нуклеиновой кислоты согласно SEQ ID NO:21.
В 45-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого, пятого, шестого и седьмого варианта осуществления первого аспекта и второго аспекта, нуклеиновая кислота типа 1B содержит в направлении 5'->3' первый участок Бокс B1A, второй участок Бокс B2, третий участок Бокс B3, четвертый участок Бокс B4, пятый участок Бокс B5, шестой участок Бокс B6 и седьмой участок Бокс B1B, где
первый участок Бокс B1A и седьмой участок Бокс B1B необязательно гибридизуются друг с другом, причем при гибридизации образуется двухцепочечная структура,
первый участок Бокс B1A содержит нуклеотидную последовательность AGYRUG,
второй участок Бокс B2 содержит нуклеотидную последовательность CCAGCU или CCAGY,
третий участок Бокс B3 содержит нуклеотидную последовательность GUG,
четвертый участок Бокс B4 содержит нуклеотидную последовательность AUG,
пятый участок Бокс B5 содержит нуклеотидную последовательность GGGGGGCGCGACC
шестой участок Бокс B6 содержит нуклеотидную последовательность CAUUUUA или CAUUUA, и
седьмой участок Бокс B1B содержит нуклеотидную последовательность CAYRCU.
В 46-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 45-ого варианта осуществления первого аспекта и второго аспекта,
первый участок Бокс B1A содержит нуклеотидную последовательность AGCGUG.
В 47-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 45-ого и 46-ого варианта осуществления первого аспекта и второго аспекта,
второй участок Бокс B2 содержит нуклеотидную последовательность CCAGU.
В 48-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 45-ого, 46-ого и 47-ого варианта осуществления первого аспекта и второго аспекта,
шестой участок Бокс B6 содержит нуклеотидную последовательность CAUUUUA.
В 49-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 45-го, 46-го, 47-го и 48-го варианта осуществления первого и второго аспекта,
седьмой участок Бокс B1B содержит нуклеотидную последовательность CACGCU.
В 50-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 45-ого, 46-ого, 47-ого, 48-ого и 49-ого варианта осуществления первого и второго аспекта, нуклеиновая кислота содержит последовательность нуклеиновой кислоты согласно SEQ ID NO:28 и SEQ ID NO:27.
В 51-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления любого из с первого по 50-ый вариантов осуществления первого и второго аспекта, MCP-1 выбран из группы, содержащей MCP-1 обезьяны, MCP-1 лошади, MCP-1 кролика, MCP-1 быка, MCP-1 собаки, MCP-1 свиньи и MCP-1 человека.
В 52-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления любого из с первого по 51-ый вариантов осуществления первого и второго аспекта, нуклеиновая кислота способна связывать MCP-1 человека.
В 53-ем варианте осуществления первого и второго аспекта, который также является вариантом осуществления любого из с первого по 52-ой вариантов осуществления первого и второго аспекта, предпочтительно, 52-ого варианта осуществления первого и второго аспекта, MCP-1 имеет аминокислотную последовательность согласно SEQ ID NO:1.
В 54-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления любого из с первого по 53-ий вариантов осуществления первого и второго аспекта, нуклеиновая кислота содержит модификацию, где модифицирующая группа предпочтительно представляет собой высокомолекулярную группу и/или где модифицирующая группа предпочтительно позволяет модифицировать характеристики нуклеиновой кислоты по любому из 1-54 с точки зрения времени нахождения в организме животного или человека, предпочтительно, в организме человека.
В 55-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 54-ого варианта осуществления первого и второго аспекта, модифицирующая группа выбрана из группы, включающей группу HES, группу PEG, биологически деградируемые модифицирующие группы и их комбинации.
В 56-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 55-ого варианта осуществления первого и второго аспекта, модифицирующая группа представляет собой группу PEG, состоящую из прямого или разветвленного PEG, где молекулярная масса группы PEG предпочтительно составляет приблизительно от 20000 до 120000 Да, более предпочтительно приблизительно от 30000 до 80000 Да и наиболее предпочтительно приблизительно 40000 Да.
В 57-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 55-ого варианта осуществления первого и второго аспекта, модифицирующая группа представляет собой группу HES, где предпочтительно молекулярная масса группы HES составляет приблизительно от 10000 до 200000 Да, более предпочтительно приблизительно от 30000 до 170000 Да и наиболее предпочтительно приблизительно 150000 Да.
В 58-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 54-ого, 55-ого, 56-ого и 57-ого варианта осуществления первого и второго аспекта, модифицирующая группа связана с нуклеиновой кислотой через линкер, где линкер представляет собой линкер или биологически деградируемый линкер.
В 59-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления 54-ого, 55-ого, 56-ого, 57-ого и 58-ого варианта осуществления первого и второго аспекта, модифицирующая группа связана с 5'-концевым нуклеотидом и/или 3'-концевым нуклеотидом нуклеиновой кислоты и/или с нуклеотидом нуклеиновой кислоты между 5'-концевым нуклеотидом и 3'-концевым нуклеотидом.
В 60-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления любого из с первого по 59-ый вариантов осуществления первого и второго аспекта, нуклеотиды нуклеиновой кислоты или нуклеотиды, образующие нуклеиновую кислоту, представляют собой L-нуклеотиды.
В 61-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления любого из с первого по 60-ый вариантов осуществления первого и второго аспекта, нуклеиновая кислота представляет собой L-нуклеиновую кислоту.
В 62-ом варианте осуществления первого и второго аспекта, который также является вариантом осуществления любого из с первого по 60-ый вариантов осуществления первого и второго аспекта, часть нуклеиновой кислоты, способная связывать MCP-1, состоит из L-нуклеотидов.
В третьем аспекте, который также является первым вариантом осуществления указанного третьего аспекта, проблема, с которой связано создание настоящего изобретения, решается с помощью фармацевтической композиции, содержащей молекулу нуклеиновой кислоты, как определено в любом из вариантов осуществления первого и второго аспекта, и, необязательно, дополнительный компонент, причем дополнительный компонент выбран из группы, содержащей фармацевтически приемлемые эксципиенты, фармацевтически приемлемые носители и фармацевтически активные средства, и где фармацевтическая композиция предназначена для лечения и/или профилактики хронического заболевания или хронического нарушения.
Во втором варианте осуществления третьего аспекта, который также является вариантом осуществления первого варианта осуществления третьего аспекта, фармацевтическая композиция содержит молекулу нуклеиновой кислоты, как определено в любом из вариантов осуществления первого и второго аспекта, и фармацевтически приемлемый носитель.
В третьем варианте осуществления третьего аспекта, который также является вариантом осуществления первого и второго варианта осуществления третьего аспекта, хроническое заболевание или хроническое нарушение является заболеванием или нарушением, как определено в любом из указанных выше пунктов.
В четвертом варианте осуществления третьего аспекта, который также является вариантом осуществления первого, второго и третьего варианта осуществления третьего аспекта, фармацевтическая композиция содержит второе фармацевтически активное средство, которое представляет собой иммунодепрессивное средство.
В пятом варианте осуществления третьего аспекта, который также является вариантом осуществления четвертого варианта осуществления третьего аспекта, иммунодепрессивное средство содержится в указанной фармацевтической композиции в качестве самостоятельной лекарственной формы.
В шестом варианте осуществления третьего аспекта, который также является вариантом осуществления четвертого и пятого варианта осуществления третьего аспекта, фармацевтическая композиция содержит меньшее количество иммунодепрессивного средства, чем фармацевтическая композиция, содержащая иммунодепрессивное средство для монотерапии.
В седьмом варианте осуществления третьего аспекта, который также является вариантом осуществления четвертого, пятого и шестого варианта осуществления третьего аспекта, лекарственная форма иммунодепрессивного средства содержит меньшее количество иммунодепрессивного средства, чем лекарственная форма в случае использования при монотерапии.
В восьмом варианте осуществления третьего аспекта, который также является вариантом осуществления шестого и седьмого варианта осуществления третьего аспекта, уменьшение количества иммунодепрессивного средства, которое является предметом шестого и седьмого варианта осуществления третьего аспекта, составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90%, предпочтительно, по меньшей мере 75%.
В девятом варианте осуществления третьего аспекта, который также является вариантом осуществления четвертого, пятого, шестого, седьмого и восьмого варианта осуществления третьего аспекта, иммунодепрессивное средство выбрано из группы, содержащей циклофосфамид и микофенолата мофетил.
В десятом варианте осуществления третьего аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого и девятого варианта осуществления третьего аспекта, более предпочтительно восьмого и девятого варианта осуществления третьего аспекта, хроническое заболевание представляет собой волчаночный нефрит и/или пневмонию.
В одиннадцатом варианте осуществления третьего аспекта, который также является вариантом осуществления первого, второго и третьего варианта осуществления третьего аспекта, фармацевтическая композиция содержит второе фармацевтически активное вещество, где такое второе фармацевтически активное вещество представляет собой противовоспалительное средство.
В двенадцатом варианте осуществления третьего аспекта, который также является вариантом осуществления одиннадцатого варианта осуществления третьего аспекта, противовоспалительное средство содержится в указанной фармацевтической композиции в качестве отдельной лекарственной формы.
В 13-ом варианте осуществления третьего аспекта, который также является вариантом осуществления одиннадцатого и двенадцатого варианта осуществления третьего аспекта, фармацевтическая композиция содержит меньшее количество противовоспалительного средства, чем фармацевтическая композиция, содержащая противовоспалительное средство для монотерапии.
В 14-ом варианте осуществления третьего аспекта, который также является вариантом осуществления одиннадцатого, двенадцатого и 13-ого варианта осуществления третьего аспекта, лекарственная форма противовоспалительного средства содержит меньшее количество иммунодепрессивного средства, чем лекарственная форма в случае применения при монотерапии.
В 15-ом варианте осуществления третьего аспекта, который также является вариантом осуществления 13-ого и 14-ого варианта осуществления третьего аспекта, уменьшение количества иммунодепрессивного средства, являющегося предметом 13-ого и 14-ого варианта осуществления третьего аспекта, составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90%, предпочтительно по меньшей мере 75%.
В 16-ом варианте осуществления третьего аспекта, который также является вариантом осуществления одиннадцатого, двенадцатого, 13-ого, 14-ого и 15-ого варианта осуществления третьего аспекта, противовоспалительное средство выбрано из группы, содержащей дексаметазон и рофлумиласт, предпочтительно, противовоспалительное средство представляет собой дексаметазон.
В 17-ом варианте осуществления третьего аспекта, который также является вариантом осуществления одиннадцатого, двенадцатого, 13-ого, 14-ого, 15-ого и 16-ого варианта осуществления третьего аспекта, предпочтительно, 15-ого и 16-ого варианта осуществления третьего аспекта, хроническое заболевание представляет собой хроническое респираторное заболевание и, более предпочтительно, COPD.
В четвертом аспекте, который также является первым вариантом осуществления указанного четвертого аспекта, проблема, с которой связано создание настоящего изобретения, решается с помощью молекулы нуклеиновой кислоты, как определено в любом из вариантов осуществления 1-62 первого и второго аспекта, для использования в способе лечения индивида, страдающего хроническим заболеванием или хроническим нарушением или имеющего риск его развития, где способ включает
- введение индивиду фармацевтически активного количества молекулы нуклеиновой кислоты.
Во втором варианте осуществления четвертого аспекта хроническое заболевание или хроническое нарушение является таким, как определено в любом из предшествующих пунктов.
В третьем варианте осуществления четвертого аспекта, который также является вариантом осуществления первого и второго варианта осуществления четвертого аспекта, способ дополнительно включает стадию
- введения индивиду иммунодепрессивного средства.
В четвертом варианте осуществления четвертого аспекта, который также является вариантом осуществления третьего варианта осуществления четвертого аспекта, количество иммунодепрессивного средства, вводимого в ходе лечения, меньше, чем количество иммунодепрессивного средства, которое вводили бы индивиду при монотерапии.
В пятом варианте осуществления четвертого аспекта, который также является вариантом осуществления четвертого варианта осуществления четвертого аспекта, количество иммунодепрессивного средства уменьшают по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90%, предпочтительно по меньшей мере на 75%.
В шестом варианте осуществления четвертого аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого и пятого варианта осуществления четвертого аспекта, иммунодепрессивное средство выбрано из группы, содержащей циклофосфамид и микофенолата мофетил.
В седьмом варианте осуществления четвертого аспекта, который также является вариантом осуществления первого, второго, третьего, четвертого, пятого и шестого варианта осуществления четвертого аспекта, и, более конкретно, пятого и шестого варианта осуществления четвертого аспекта, хроническое заболевание представляет собой хроническое заболевание почек, предпочтительно, волчаночный нефрит и/или пневмонию.
В восьмом варианте осуществления четвертого аспекта, который также является вариантом осуществления первого и второго варианта осуществления четвертого аспекта, способ дополнительно включает стадию
- введения индивиду противовоспалительного средства.
В девятом варианте осуществления четвертого аспекта, который также является вариантом осуществления восьмого варианта осуществления четвертого аспекта, количество противовоспалительного средства, вводимого в ходе лечения, является меньшим, чем количество иммунодепрессивного средства, которое вводили бы индивиду при монотерапии.
В десятом варианте осуществления четвертого аспекта, который также является вариантом осуществления девятого варианта осуществления четвертого аспекта, количество иммунодепрессивного средства уменьшают по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90%, предпочтительно по меньшей мере на 75%.
В одиннадцатом варианте осуществления четвертого аспекта, который также является вариантом осуществления восьмого, девятого и десятого варианта осуществления четвертого аспекта, противовоспалительное средство выбрано из группы, содержащей дексаметазон и рофлумиласт, предпочтительно, противовоспалительное средство представляет собой дексаметазон.
В двенадцатом варианте осуществления четвертого аспекта, который также является вариантом осуществления восьмого, девятого, десятого и одиннадцатого варианта осуществления четвертого аспекта, и, более конкретно, десятого и одиннадцатого варианта осуществления четвертого аспекта, хроническое заболевание представляет собой хроническое респираторное заболевание, предпочтительно COPD.
В пятом аспекте, который также является первым вариантом осуществления указанного пятого аспекта, проблема, с которой связано создание настоящего изобретения, решается с помощью применения молекулы нуклеиновой кислоты, как определено в любом из вариантов осуществления 1-62 первого и второго аспекта, для получения лекарственного средства для лечения и/или профилактики хронического заболевания или хронического нарушения.
Во втором варианте осуществления пятого аспекта, заболевание или нарушение представляет собой заболевание или нарушение, как определено в любом из вариантов осуществления 1-62 первого и второго аспекта.
В третьем варианте осуществления пятого аспекта, который также является вариантом осуществления первого и второго варианта осуществления пятого аспекта, лекарственное средство предназначено для использования в медицине человека или в ветеринарной медицине.
В шестом аспекте, который также является первым вариантом осуществления указанного шестого аспекта, проблема, с которой связано создание настоящего изобретения, решается с помощью способа диагностики хронического заболевания или хронического нарушения, включающего следующие стадии:
- приведение образца, взятого у индивида, который подлежит тестированию на наличие или отсутствие хронического заболевания или хронического нарушения или на наличие риска его развития, в контакт с молекулой нуклеиновой кислоты, как определено в любом из вариантов осуществления 1-62 первого и второго аспекта; и
- непосредственную или косвенную детекцию комплекса, содержащего MCP-1, и молекулы нуклеиновой кислоты.
Во втором варианте осуществления шестого аспекта, хроническое заболевание или хроническое нарушение представляет собой хроническое нарушение или хроническое заболевание, как определено в любом из вариантов осуществления 1-62 первого и второго аспекта.
В седьмом аспекте, который также является первым вариантом осуществления указанного седьмого аспекта, проблема, с которой связано создание настоящего изобретения, решается путем использования молекулы нуклеиновой кислоты, как определено в любом из вариантов осуществления 1-62 первого и второго аспекта, для получения диагностического средства для диагностики хронического заболевания или хронического нарушения, как определено в любом из вариантов осуществления 1-62 первого и второго аспекта.
Специалистам в данной области будет понятно, что также можно осуществлять последующие варианты осуществления и признаки, применительно к признакам и вариантам осуществления, описанным в настоящем документе, в частности, применительно к аспектам и вариантам осуществления, заявленных в качестве объекта изобретения в формуле изобретения.
Как используют в настоящем документе, термины "хроническое заболевание" и "хроническое нарушение" предпочтительно относятся к хроническому респираторному заболеванию, хроническому заболеванию почек и системной красной волчанке. Предпочтительно, термин "хроническое респираторное заболевание", как используют в настоящем документе, включает пневмонию, воспаление легких и плевры, плеврит, плевральный выпот, волчаночную пневмонию, хроническую диффузную интерстициальную болезнь легких, эмболию легких, кровоизлияние в легкие, синдром сморщенного легкого, легочную гипертензию и хроническую обструктивную болезнь легких и их комбинации. Более предпочтительно, термин "легочная гипертензия" включает легочную гипертензию, связанную с болезнью левого отдела сердца, легочную гипертензию, связанную с заболеваниями легких и/или гипоксемией, легочную гипертензию вследствие хронического тромботического и/или эмболического заболевания, гипертензию легочной артерии, предпочтительно, идиопатическую гипертензию легочной артерии, гипертензию легочной артерии, связанную с коллагенозом, семейную гипертензию легочной артерии, гипертензию легочной артерии, связанную с другими заболеваниями, и гипертензию легочной артерии, связанную с заболеваниями вен и капилляров. Более того, термин "хроническая обструктивная болезнь легких" предпочтительно включает хроническую обструктивную болезнь легких с вовлечением сосудов легких или без него. Наконец, термин "хроническая обструктивная болезнь легких", предпочтительно, включает хронический бронхит и эмфизему. Также, термин "хроническое заболевание почек", предпочтительно, включает волчаночный нефрит, мембрано-пролиферативный гломерулонефрит, мембранозный гломерулонефрит, IgA-нефропатию, пост-стрептококковый гломерулонефрит, быстро прогрессирующий гломерулонефрит, нефритический синдром, фокальный сегментный гломерулосклероз, диабетическую нефропатию, нефротический синдром, и нефротический синдром, предпочтительно, волчаночный нефрит.
Признаки нуклеиновой кислоты по настоящему изобретению, как раскрыто в настоящем описании, можно осуществлять в любом аспекте настоящего изобретения, если используют нуклеиновую кислоту, либо отдельно, либо в сочетании.
MCP-1 человека, а также мыши, представляют собой основные белки, имеющие аминокислотную последовательность согласно SEQ ID NO: 1 и 2, соответственно.
Обнаружение того факта, что можно идентифицировать короткие высоко аффинные нуклеиновые кислоты, связывающиеся с MCP-1, является в значительной мере удивительным, поскольку Eaton et al. (1997) выявили, что получение аптамеров, т.е. D-нуклеиновых кислот, связывающихся с молекулой-мишенью, направленных на основной белок, является, как правило, очень трудным, вследствие того, что этот тип мишеней дает высокое, но неспецифическое отношение сигнала к шуму. Это высокое отношение сигнала к шуму является следствием высокой неспецифической аффинности, которую демонстрируют нуклеиновые кислоты к основным мишеням, таким как MCP-1.
Как более подробно указано в формуле изобретения и примере 1, авторы настоящего изобретения, более неожиданно, идентифицировали ряд различных связывающих MCP-1 молекул нуклеиновых кислот, в которых большинство нуклеиновых кислот можно было охарактеризовать с точки зрения участков из нуклеотидов, которые называют в настоящем документе боксами. Различные связывающие MCP-1 молекулы нуклеиновых кислот могут быть подразделены на категории на основе указанных боксов и некоторых структурных признаков и элементов, соответственно. Различные категории, определенные таким образом, также называют в настоящем документе типами и, более конкретно, типом 1A, типом 1B, типом 2, типом 3 и типом 4.
Согласно настоящему изобретению, нуклеиновые кислоты по настоящему изобретению или их участки или их любая часть(и), могут, в принципе, гибридизоваться друг с другом. При такой гибридизации образуется двухцепочечная структура. Специалистам в данной области будет понятно, что такая гибридизация может происходить или может не происходить, в частности, в условиях in vitro и/или in vivo. Также, в случае такой гибридизации, не обязательно, чтобы гибридизация происходила на протяжении всей длины двух участков, где, по меньшей мере на основании правил спаривания оснований, такая гибридизация и, таким образом, образование двухцепочечной структуры, могут, в принципе, произойти. Как предпочтительно используют в настоящем документе, двухцепочечная структура является частью молекулы или структуры, образуемой двумя или более отдельными цепями или двумя пространственно разделенными участками одной цепи, где существует по меньшей мере одна, предпочтительно, две или более пар оснований, которые образуют пары оснований, предпочтительно, согласно правилам спаривания оснований по принципу Уотсона-Крика. Также специалистам в данной области будет понятно, что другой тип спаривания оснований, такой как спаривание оснований по принципу Хугстена, может существовать в двухцепочечной структуре или формировать ее.
В предпочтительном варианте осуществления расположение терминов, как используют в настоящем документе, означает порядок или последовательность структурных или функциональных признаков или элементов, описанных в настоящем документе, применительно к нуклеиновым кислотам, описанным в настоящем документе.
Согласно настоящему изобретению, нуклеиновая кислота по настоящему изобретению представляет собой молекулу нуклеиновой кислоты. Таким образом, термины "нуклеиновая кислота" и "молекула нуклеиновой кислоты" используют в настоящем документе взаимозаменяемо, если не указано иного. В одном из вариантов осуществления настоящей заявки нуклеиновая кислота и, таким образом, молекула нуклеиновой кислоты, содержит молекулу нуклеиновой кислоты, которая характеризуется тем, что все последовательно расположенные нуклеотиды, образующих молекулу нуклеиновой кислоты, связаны или соединены друг с другом одной или несколькими ковалентными связями. Более конкретно, каждый из таких нуклеотидов связан или соединен с другими нуклеотидами, предпочтительно, через фосфодиэфирные связи или другие связи, образующие участок последовательно расположенных нуклеотидов. Однако в такой конфигурации каждый из концевых нуклеотидов, т.е., предпочтительно нуклеотид на 5'-конце и на 3'-конце, связан только с одним нуклеотидом, при условии, что такая конфигурация является линейной, а не замкнутой конфигурацией, и, таким образом, линейной, а не замкнутой молекулой.
В другом варианте осуществления настоящей заявки нуклеиновая кислота и, таким образом, молекула нуклеиновой кислоты, содержит по меньшей мере две группы последовательно расположенных нуклеотидов, где в каждой группе последовательно расположенных нуклеотидов каждый нуклеотид связан или соединен с другими нуклеотидами, предпочтительно, через фосфодиэфирные связи или другие связи, образуя участок последовательно расположенных нуклеотидов. Однако в такой конфигурации каждый из двух концевых нуклеотидов, т.е. предпочтительно нуклеотид на 5'-конце и на 3'-конце, связан только с одним нуклеотидом. Однако в таком варианте осуществления, две группы последовательно расположенных нуклеотидов не связаны или не соединены друг с другом ковалентной связью, которая связывает один нуклеотид одной группы и один нуклеотид другой группы ковалентной связью, предпочтительно, ковалентной связью, образованной между группой сахара одного из указанных двух нуклеотидов и группой фосфора другого из указанных двух нуклеотидов или нуклеозидов. Однако в альтернативном варианте осуществления, две группы последовательно расположенных нуклеотидов, связаны или соединены друг с другом ковалентной связью, которая связывает один нуклеотид одной группы и один нуклеотид другой группы ковалентной связью, предпочтительно, ковалентной связью, образованной группой сахара одного из указанных двух нуклеотидов и группой фосфора другого из указанных двух нуклеотидов или нуклеозидов. Предпочтительно, по меньшей мере две группы последовательно расположенных нуклеотидов не связаны ковалентной связью. В другом предпочтительном варианте осуществления, по меньшей мере две группы связаны ковалентной связью, которая отличается от фосфодиэфирной связи. В другом варианте осуществления по меньшей мере две группы связаны ковалентной связью, которая представляет собой фосфодиэфирную связь.
Нуклеиновые кислоты по настоящему изобретению также включают нуклеиновые кислоты, которые по существу гомологичны конкретным последовательностям, описанным в настоящем документе. Под термином "по существу гомологичный" следует понимать, что гомология составляет по меньшей мере 75%, предпочтительно, 85%, более предпочтительно 90% и наиболее предпочтительно более 95%, 96%, 97%, 98% или 99%.
Действительный процент гомологичных нуклеотидов, присутствующих в нуклеиновой кислоте по настоящему изобретению может зависеть от общего числа нуклеотидов, находящихся в нуклеиновой кислоте. В основе изменения процента может лежать общее число нуклеотидов, присутствующих в нуклеиновой кислоте.
Гомологию можно определить, способами известными специалистам в данной области. Более конкретно, алгоритм сравнения последовательностей вычисляет процентную идентичность последовательностей для тестируемой последовательности(ей) относительно эталонной последовательности, исходя из определенных параметров программы. Тестируемая последовательность предпочтительно представляет собой последовательность или молекулу нуклеиновой кислоты, которую тестируют или которая подлежит тестированию в отношении гомологии, и если она гомологична, то в отношении степени гомологии, с другой молекулой нуклеиновой кислоты, где такую другую молекулу нуклеиновой кислоты также называют эталонной последовательностью. В одном из вариантов осуществления, контрольная последовательность представляет собой молекулу нуклеиновой кислоты, как описано в настоящем документе, более предпочтительно, молекулу нуклеиновой кислоты, имеющую последовательность согласно любой из SEQ ID NO:10-129, 132-256 и 278-282. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма локальной гомологии Smith & Waterman (Smith & Waterman, 1981) с помощью алгоритма выравнивания для гомологии Needleman & Wunsch (Needleman & Wunsch, 1970) способом поиска сходства Pearson & Lipman (Pearson & Lipman, 1988), с помощью компьютерных вариантов осуществления этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis.), или с помощью визуального исследования.
Одним из примеров алгоритма, который можно использовать для определения процента идентичности последовательностей, является алгоритм, используемый в основном инструменте поиска локальных блоков (далее в настоящем документе "BLAST"), см., например Altschul et al (Altschul et al. 1990 и Altschul et al, 1997). Программное обеспечение для проведения анализа BLAST является общедоступным от National Center for Biotechnology Information (далее в настоящем документе "NCBI"). Параметры по умолчанию, используемые в определении идентичности последовательностей с использованием программного обеспечения, доступного от NCBI, например, BLASTN (для нуклеотидных последовательностей) и BLASTP (для аминокислотных последовательностей) описаны в McGinnis et al (McGinnis et al, 2004).
Термин "нуклеиновая кислота по изобретению" также включает нуклеиновые кислоты, содержащие последовательности нуклеиновых кислот, описанные в настоящем документе, или их часть, предпочтительно, до такой степени, чтобы нуклеиновые кислоты или указанные части были вовлечены в связывание с MCP-1. Термин "нуклеиновая кислота по изобретению", как предпочтительно используют в настоящем документе, также включает, в одном из вариантов осуществления, нуклеиновую кислоту, которая может использоваться для связывания с любой молекулой, выбранной из группы, содержащей MCP-2, MCP-3, MCP-4 и эотаксин. Специалистам в данной области будет понятно, что отдельные нуклеиновые кислоты по настоящему изобретению будут связываться с одной или несколькими такими молекулами. Такая нуклеиновая кислота представляет собой, в одном из вариантов осуществления, одну из молекул нуклеиновых кислот, описанных в настоящем документе, или их производное и/или метаболит, где такое производное и/или метаболит, предпочтительно, представляет собой нуклеиновую кислоту, укороченную по сравнению с молекулами нуклеиновых кислот, описанными в настоящем документе. Укорочение может быть на любом или на обоих концах нуклеиновых кислот, как описано в настоящем документе. Также укорочение может быть во внутренней последовательности нуклеотидов нуклеиновой кислоты, т.е. оно может относиться к нуклеотиду(ам) между 5'- и 3'-концевыми нуклеотидами, соответственно. Более того, укорочение может включать делецию только одного нуклеотида из последовательности нуклеиновых кислот, описанных в настоящем документе. Укорочение может относиться к более чем одному участку нуклеиновой кислоты(кислот), где участок может иметь только один нуклеотид в длину. Связывание нуклеиновой кислоты по настоящему изобретению, предпочтительно, с молекулой, выбранной из группы, содержащей MCP-1, MCP-2, MCP-3, MCP-4 и эотаксин, может быть определено специалистами в данной области общепринятыми экспериментами или используя или внедрив способ, описанный в настоящем документе, предпочтительно, описаный в настоящем документе в разделе "Примеры". В одном из вариантов осуществления настоящего изобретения, если нет иных указаний, если в настоящем документе приведено указание на связывание нуклеиновых кислот по настоящему изобретению с MCP-1, то это также относится к связыванию нуклеиновых кислот по настоящему изобретению с любой молекулой, выбранной из группы, содержащей MCP-2, MCP-3, MCP-4 и эотаксин.
Нуклеиновые кислоты по настоящему изобретению могут представлять собой либо D-нуклеиновые кислоты, либо L-нуклеиновые кислоты. Предпочтительно, нуклеиновые кислоты по изобретению представляют собой L-нуклеиновые кислоты. Кроме того, возможен вариант, когда одна или несколько частей нуклеиновой кислоты присутствуют в качестве D-нуклеиновых кислот или по меньшей мере одна или несколько частей нуклеиновых кислот представляли собой L-нуклеиновые кислоты. Термин "часть" нуклеиновых кислот означает как минимум один нуклеотид. Такие нуклеиновые кислоты, как правило, называют в настоящем документе D- и L-нуклеиновыми кислотами, соответственно. Таким образом, в особенно предпочтительном варианте осуществления, нуклеиновые кислоты по настоящему изобретению состоят из L-нуклеотидов и содержат по меньшей мере один D-нуклеотид. Такой D-нуклеотид предпочтительно связан с частью, отличной от участков, определяющих нуклеиновые кислоты по настоящему изобретению, предпочтительно, с частями, где вовлечено взаимодействие с другими частями нуклеиновой кислоты. Предпочтительно, такой D-нуклеотид связан с концом любого из участков и любой из нуклеиновых кислот по настоящему изобретению, соответственно. В следующем предпочтительном варианте осуществления такие D-нуклеотиды могут действовать в качестве спейсера или линкера, предпочтительно, связывая модифицирующие группы, такие как PEG и HES, с нуклеиновыми кислотами по настоящему изобретению.
Также в одном из вариантов осуществления настоящего изобретения, каждая и любая из молекул нуклеиновых кислот, описанных в настоящем документе в полном объеме с точки зрения их последовательности(ей) нуклеиновой кислоты, ограничивается конкретной нуклеотидной последовательностью(ями). Иными словами, термины "включающий" или "включает(ют)" следует интерпретировать в таком варианте осуществления, как означающий "содержащий" или "состоящий из".
Также к настоящему изобретению относится то, что нуклеиновые кислоты по настоящему изобретению являются частью более длинной нуклеиновой кислоты, где эта более длинная нуклеиновая кислота содержит несколько частей, где по меньшей мере одна такая часть представляет собой нуклеиновую кислоту по настоящему изобретению, или ее часть. Другая часть(и) этих более длинных нуклеиновых кислот может представлять собой либо одну или несколько D-нуклеиновую кислоту(кислот), либо одну или несколько L-нуклеиновую кислоту(кислот). В контексте настоящего изобретения можно использовать любую комбинацию. Эта другая часть(и) более длинной нуклеиновой кислоты, либо отдельно или взятые вместе, либо в целом или в конкретной комбинации, может проявлять функцию, которая отличается от связывания, предпочтительно, от связывания с MCP-1. Одной из возможных функций является обеспечение взаимодействия с другими молекулами, где такие другие молекулы предпочтительно отличаются от MCP-1, например, для иммобилизации, перекрестного связывания, детекции или амплификации. В другом варианте осуществления настоящего изобретения нуклеиновые кислоты по изобретению содержат, в качестве самостоятельной группы или объединенных частей, несколько нуклеиновых кислот по настоящему изобретению. Такая нуклеиновая кислота, содержащая несколько нуклеиновых кислот по настоящему изобретению, также входит в состав термина "более длинная нуклеиновая кислота".
L-нуклеиновые кислоты, как используют в настоящем документе, представляют собой нуклеиновые кислоты, состоящие из L-нуклеотидов, предпочтительно, состоящие только из L-нуклеотидов.
D-нуклеиновые кислоты, как используют в настоящем документе, представляют собой нуклеиновые кислоты, состоящие из D-нуклеотидов, предпочтительно, состоящие только из D-нуклеотидов.
Термины "нуклеиновая кислота" и "молекула нуклеиновой кислоты" используют в настоящем документе взаимозаменяемо, если нет иных указаний.
Также, если нет иных указаний, любая нуклеотидная последовательность указана в настоящем документе в направлении 5' → 3'.
Независимо от того, состоит ли нуклеиновая кислота по изобретения из D-нуклеотидов, L-нуклеотидов или представляет собой их комбинацию, которая является, например, случайной комбинацией или определенной последовательностью участков, состоящей из по меньшей мере одного L-нуклеотида и по меньшей мере одной D-нуклеиновой кислоты, нуклеиновая кислота может состоять из дезоксирибонуклеотида(ов), рибонуклеотида(ов) или их комбинаций.
Конструирование нуклеиновых кислот по изобретению в качестве L-нуклеиновой кислоты является преимущественным по нескольким причинам. L-нуклеиновые кислоты представляют собой энантиомеры природных нуклеиновых кислот. Однако D-нуклеиновые кислоты не являются высокостабильными в водных растворах и, в частности, в биологических системах или биологических образцах из-за большого количества нуклеаз. Природные нуклеазы, в частности, нуклеазы, полученные из клеток животных, не способны деградировать L-нуклеиновые кислоты. Вследствие этого, биологическое время полужизни L-нуклеиновой кислоты в такой системе, включая организм животного и человека, значительно увеличивается. Из-за отсутствия способности к деградации L-нуклеиновой кислоты, не образуются продукты деградации нуклеазой, и, таким образом, не наблюдается побочных эффектов, связанных с ними. Этот аспект отличает L-нуклеиновую кислоту фактически от всех других соединений, которые используют в терапии заболеваний и/или нарушений, связанных с MCP-1. L-нуклеиновые кислоты, которые специфично связываются с молекулой-мишенью посредством механизма, отличного от спаривания оснований по принципу Уотсона-Крика, или аптамеры, которые состоят частично или полностью из L-нуклеотидов, в частности если эти части аптамера, вовлечены в связывание аптамера с молекулой-мишенью, также называют шпигельмерами.
Также настоящее изобретение относится к нуклеиновым кислотам, также называемым в настоящем документе нуклеиновыми кислотами по изобретению, независимо от того, находятся ли они в виде D-нуклеиновых кислот, L-нуклеиновых кислот или D,L-нуклеиновых кислот, или являются ли они ДНК или РНК, которые могут быть представлены в качестве одноцепочечных или двухцепочечных нуклеиновых кислот. Как правило, нуклеиновые кислоты по изобретению представляют собой одноцепочечные нуклеиновые кислоты, которые обладают определенными вторичными структурами благодаря первичной последовательности, и, таким образом, также могут образовывать третичные структуры. Однако нуклеиновые кислоты по изобретению также могут быть двухцепочечными в том смысле, что две их цепи, которые комплементарны или частично комплементарны друг другу, являются гибридизованными друг с другом. Это придает стабильность нуклеиновым кислотам, которая, в частности, может быть преимущественной, если нуклеиновая кислота находится в природной D-форме, а не в L-форме.
Нуклеиновые кислоты по изобретению могут быть модифицированными. Такие модификации могут относиться к одному нуклеотиду нуклеиновой кислоты, и они хорошо известны в данной области. Примеры такой модификации описаны, среди прочих, в Venkatesan (Venkatesan 2003); Kusser (Kusser 2000); Aurup (Aurup 1994); Cummins (Cummins 1995); Eaton et al. (Eaton 1995); Green et al. (Green 1995); Kawasaki et al. (Kawasaki 1993); Lesnik et al. (Lesnik 1993); и Miller & Kragel (Miller 1993). Такая модификация может представлять собой атом H, атом F или группу O-CH3 или группу NH2 во 2'-положении отдельных нуклеотидов, из которых состоит нуклеиновая кислота. Также нуклеиновая кислота по настоящему изобретению может содержать по меньшей мере один нуклеотид LNA. В одном из вариантов осуществления нуклеиновая кислота по настоящему изобретению состоит из нуклеотидов LNA.
В одном из вариантов осуществления, нуклеиновые кислоты по настоящему изобретению могут представлять собой нуклеиновую кислоту из нескольких частей. Нуклеиновая кислота из нескольких частей, как используют в настоящем документе, представляет собой нуклеиновую кислоту, которая состоит по меньшей мере из двух цепей нуклеиновых кислот. Эти по меньшей мере две цепи нуклеиновых кислот образуют функциональную единицу, где функциональная единица является лигандом для молекулы-мишени. По меньшей мере две цепи нуклеиновых кислот могут быть образованы из любой из нуклеиновых кислот по изобретению, либо путем расщепления нуклеиновой кислоты с получением двух цепей, либо путем синтеза одной нуклеиновой кислоты, соответствующей первой части всей нуклеиновой кислоты по изобретению, т.е. всей нуклеиновой кислоты, и другой нуклеиновой кислоты, соответствующей второй части всей нуклеиновой кислоты. Также будет понятно, что для получения нуклеиновой кислоты из нескольких частей, где имеется более двух цепей, как объяснено выше, можно использовать как расщепление, так и синтез. Иными словами, по меньшей мере две цепи нуклеиновой кислоты, как правило, отличаются от двух цепей, являющихся комплементарными и гибридизующимися друг с другом, хотя может существовать определенная степень комплементарности между различными частями нуклеиновой кислоты.
Наконец, также настоящее изобретение относится к тому, что является осуществимой полностью замкнутая, т.е. кольцевая структура нуклеиновых кислот по настоящему изобретению, т.е. что нуклеиновые кислоты по настоящему изобретению являются замкнутыми, предпочтительно, посредством ковалентной связи, где, более предпочтительно, такая ковалентная связь находится между 5'-концом и 3'-концом последовательностей нуклеиновых кислот, как описано в настоящем документе.
Авторы настоящего изобретения обнаружили, что нуклеиновые кислоты по настоящему изобретению проявляют высоко благоприятный диапазон значений KD.
Определение константы связывания можно проводить с использованием так называемого устройства “biacore”, которое также известно специалисту в данной области. Аффинность, как используют в настоящем документе, также измеряют с использованием анализа "опускания", как описано в разделе "Примеры". Соответствующим показателем для выражения интенсивности связывания между нуклеиновой кислотой по изобретению с мишенью, которая в данном случае представляет собой MCP-1, является так называемая величина KD, которая вместе со способом ее определения хорошо известна специалисту в данной области.
Нуклеиновые кислоты по настоящему изобретению характеризуются определенным значением KD. Предпочтительно, значение KD, которое демонстрируют нуклеиновые кислоты по настоящему изобретению, составляет менее 1 мкМ. Значение KD приблизительно 1 мкМ считают характерным для неспецифического связывания нуклеиновой кислоты с мишенью. Как будет понятно специалистам в данной области, значение KD для группы соединений, таких как нуклеиновые кислоты по настоящему изобретению, находится в определенном диапазоне. Указанное выше значение KD приблизительно 1 мкМ является предпочтительной верхней границей для значения KD. Предпочтительной нижней границей KD для связывающих мишень нуклеиновых кислот может быть приблизительно 10 пикомоль или более. Настоящее изобретение относится к варианту, когда значения KD отдельных нуклеиновых кислот, связывающихся с MCP-1, предпочтительно, находится в этом диапазоне. Предпочтительный диапазон можно определить путем выбора любого первого числа из этого диапазона и любого второго числа из этого диапазона. Предпочтительные верхние значения составляют 250 нМ и 100 нМ, предпочтительные нижние значения составляют 50 нМ, 10 нМ, 1 нМ, 100 пМ и 10 пМ.
Молекулы нуклеиновых кислот по настоящему изобретению могут иметь любую длину, при условии, что молекулы будут сохранять способность связываться с молекулой-мишенью. Специалисту в данной области будет понятно, что существует предпочтительная длина нуклеиновых кислот по настоящему изобретению. Как правило, длина составляет от 15 до 120 нуклеотидов. Специалистам в данной области будет понятно, что любое целое число от 15 до 120 является возможной длиной для нуклеиновых кислот по настоящему изобретению. Более предпочтительным диапазоном длины нуклеиновых кислот по настоящему изобретению является длина, составляющая приблизительно от 20 до 100 нуклеотидов, приблизительно от 20 до 80 нуклеотидов, приблизительно от 20 до 60 нуклеотидов, приблизительно от 20 до 50 нуклеотидов и приблизительно от 30 до 50 нуклеотидов.
Настоящее изобретение относится к нуклеиновым кислотам, описанным в настоящем документе, которые содержат группу, которая, предпочтительно, представляет собой высокомолекулярную группу и/или которая, предпочтительно, позволяет модифицировать характеристики нуклеиновой кислоты с точки зрения, среди прочих, времени нахождения в организме животного, предпочтительно, в организме человека. Особенно предпочтительным вариантом осуществления такой модификации является ПЭГ-илирование и HES-илирование нуклеиновых кислот по настоящему изобретению. Как используют в настоящем документе PEG означает поли(этиленгликоль), а HES означает гидроксиэтилкрахмал. ПЭГ-илирование, как предпочтительно используют в настоящем документе, представляет собой модификацию нуклеиновой кислоты по настоящему изобретению, где такая модификация состоит в присоединении группы PEG к нуклеиновой кислоте по настоящему изобретению. HES-илирование, как предпочтительно используют в настоящем документе, представляет собой модификацию нуклеиновой кислоты по настоящему изобретению, где такая модификация состоит в присоединении группы HES к нуклеиновой кислоте по настоящему изобретению. Эти модифицирующие группы, а также процесс модификации нуклеиновой кислоты с использованием таких модифицирующих групп, описаны в европейской патентной заявке EP 1306382 и в международной патентной заявке WO2005/074993, описания которых включены в настоящий документ в качестве ссылок в полном объеме.
Предпочтительно, молекулярная масса модифицирующей группы, состоящий из высокомолекулярной группы, или включающей ее, составляет приблизительно от 2000 до 250000 Да, предпочтительно, от 20000 до 200000 Да. Если такой высокомолекулярной группой является PEG, то молекулярная масса предпочтительно составляет от 20000 до 120000 Да, более предпочтительно, от 40000 до 80000 Да. Если такой высокомолекулярной группой является HES, то молекулярная масса составляет предпочтительно от 20000 до 200000 Да, более предпочтительно от 40000 до 150000 Да. Процесс модификации с помощью HES описан, например, в патентной заявке Германии DE 1 2004 006 249.8 и в международной патентной заявке WO2002/080979, описания которых включены в настоящий документ в качестве ссылки в полном объеме.
Настоящее изобретение относится к варианту, когда PEG, так и HES, можно использовать в качестве либо линейной, либо разветвленной формы, как дополнительно описано в патентных заявках WO2005/074993, WO2002/080979 и PCT/EP02/11950. Такую модификацию молекул нуклеиновых кислот по настоящему изобретению, в принципе, можно проводить в любом их положении. Предпочтительно, такую модификацию проводит либо на 5'-концевом нуклеотиде, либо на 3'-концевом нуклеотиде и/либо на любом нуклеотиде между 5'-нуклеотидом и 3'-нуклеотидом молекулы нуклеиновой кислоты.
Модифицирующую группу, и, предпочтительно, группу PEG и/или HES, можно присоединять к молекуле нуклеиновой кислоты по настоящему изобретению либо напрямую, либо через линкер. Также настоящее изобретение относится к молекуле нуклеиновой кислоты по настоящему изобретению, которая содержит одну или несколько модифицирующих групп, предпочтительно, одну или несколько групп PEG и/или HES. В одном из вариантов осуществления, отдельная линкерная молекула присоединяет более одной группы PEG или группы HES к молекуле нуклеиновой кислоты по настоящему изобретению. Линкер, используемый применительно к настоящему изобретению, сам может быть либо линейным, либо разветвленным. Этот тип линкеров известен специалистам в данной области и дополнительно описан в патентных заявках WO2005/074993 и PCT/EP02/11950.
В предпочтительном варианте осуществления линкер представляет собой биологически деградруемый линкер. Биологически деградируемый линкер позволяет модифицировать характеристики нуклеиновой кислоты по настоящему изобретению с точки зрения, среди прочих, времени нахождения в организме животного, предпочтительно, в организме человека, благодаря высвобождению модифицирующей группы из нуклеиновой кислоты по настоящему изобретению. Использование биологически деградируемого линкера может позволить лучший контроль времени нахождения нуклеиновой кислоты по настоящему изобретению. Предпочтительно вариантом осуществления такого биологически деградируемого линкера является биологически деградируемый линкер, как описано, но ими не ограничивающиеся, в международных патентных заявках WO2006/052790, WO2008/034122, WO2004/092191 и WO2005/099768, где в международных патентных заявках WO2004/092191 и WO2005/099768, линкер является частью полимерного олигонуклеотидного пролекарства, которое состоит из одной или двух модифицирующих групп, как описано в настоящем документе, молекулы нуклеиновой кислоты и биологически деградируемого линкера между ними.
Настоящее изобретение относится к тому, что модифицирующая группа является биологически деградируемой модифицирующей группой, где биологически деградируемая модифицирующая группа может быть присоединена к молекуле нуклеиновой кислоты по настоящему изобретению либо напрямую, либо через линкер. Биологически деградируемая модифицирующая группа позволяет модифицировать характеристики нуклеиновой кислоты по настоящему изобретению с точки зрения, среди прочих, времени нахождения в организме животного, предпочтительно, в организме человека, благодаря высвобождению модифицирующей группы из нуклеиновой кислоты по настоящему изобретению. Использование биологически деградруемой модифицирующей группы может обеспечить лучший контроль времени нахождения нуклеиновой кислоты по настоящему изобретению. Предпочтительно, вариантом осуществления такой биологически деградируемой модифицирующей группы являются биологически деградируемые полимеры, описанные, но ими не ограничивающиеся, в международных патентных заявках WO2002/065963, WO2003/070823, WO2004/113394 и WO2000/41647, в WO2000/41647, предпочтительно, на стр. 18, строках 4-24. Более предпочтительно, биологически деградируемый полимер представляет собой биологически деградируемый PEG или биологически деградируемую полигликолевую кислоту (сокращенно PLGA), как описано в международных патентных заявках WO2004/113394 и WO2000/41647, соответственно.
Не основываясь на какой-либо конкретной теории, полагают, что при модификации нуклеиновых кислот по настоящему изобретению высокомолекулярной группой, такой как полимер и, более конкретно, полимеры, описанные в настоящем документе, причем группа, предпочтительно, является физиологически приемлемой, изменяется кинетика выведения нуклеиновой кислоты. Более конкретно, полагают, что благодаря увеличению молекулярной массы таких модифицированных нуклеиновых кислот по изобретению и вследствие того, что нуклеиновые кислоты не подвергаются метаболизму, в частности, если они находятся в L-форме, выведение из организма животного, предпочтительно, из организма млекопитающего, и, более предпочтительно, из организма человека, снижается. Поскольку выведение, как правило, осуществляется через почки, авторы настоящего изобретения полагают, что скорость гломерулярной фильтрации модифицированной таким образом нуклеиновой кислоты значительно снижена по сравнению с нуклеиновыми кислотами, не имеющими этого типа высокомолекулярной модифицирующей группы, что приводит к повышению времени нахождения в организме. В связи с этим, особенно примечательно, что, несмотря на такую высокомолекулярную модифицирующую группу, на специфичность нуклеиновой кислоты по настоящему изобретению не оказывается неблагоприятного влияния. Таким образом, нуклеиновые кислоты по настоящему изобретению имеют неожиданные характеристики - которые обычно нельзя ожидать от фармацевтически активных соединений - состоящие в том, что для обеспечения замедленного высвобождения не обязательно требуется фармацевтический состав, предоставляемый для замедленного высвобождения. Вместо этого, нуклеиновые кислоты по настоящему изобретению в их модифицированной форме, содержащей высокомолекулярную группу, сами по себе уже можно использовать в качестве состава для замедленного высвобождения. Таким образом, модификация(и) молекул нуклеиновых кислот, как описано в настоящем документе, и, таким образом, модифицированные молекулы нуклеиновых кислот и любая композиция, содержащая их, могут обеспечить особую, предпочтительно, контролируемую их фармакокинетику и биораспределение. Также эти характеристики включают время нахождения в кровотоке и распределение в тканях. Такие модификации дополнительно описаны в патентной заявке PCT/EP02/11950.
Однако также настоящее изобретение относится к нуклеиновым кислотам, которые не содержат модификацию и, в частности, модификацию высокомолекулярной группой, такую как ПЭГ-илирование или HES-илирование. Такой вариант осуществления является особенно предпочтительным, если нуклеиновая кислота имеет предпочтительное распределение в какой-либо орган-мишень или ткань-мишень в организме, или когда требуется быстрое выведение нуклеиновых кислот из организма после введения. Нуклеиновые кислоты, описанные в настоящем документе, с предпочтительными характеристиками распределения в какой-либо орган-мишень или ткань-мишень в организме, могут обеспечить эффективные локальные концентрации в ткани-мишени при сохранении низкой системной концентрации нуклеиновых кислот. Это позволяет снизить применяемую дозу, что не только выгодно с экономической точки зрения, но также уменьшает ненужное воздействие средства на основе нуклеиновой кислоты на другие ткани, и таким образом, снижает вероятный риск развития побочных эффектов. Быстрое выведение нуклеиновых кислот, описанных в настоящем документе, из организма после введения может быть желательным в случае визуализации in vivo или специальных требований к дозированию терапевтического средства, в котором используются нуклеиновые кислоты, или лекарственных средств, содержащих их, по настоящему изобретению.
Нуклеиновые кислоты, которые также в настоящем документе называют нуклеиновыми кислотами по настоящему изобретению, и/или антагонистами по настоящему изобретению, можно использовать для получения лекарственного средства. Такое лекарственное средство или фармацевтическая композиция по настоящему изобретению содержит по меньшей мере одну нуклеиновую кислоту по изобретению, необязательно, вместе с дополнительными фармацевтически активными соединениями, при этом нуклеиновая кислота по изобретению предпочтительно действует в качестве самостоятельного фармацевтически активного соединения. Такие лекарственные средства содержат, в предпочтительных вариантах осуществления, по меньшей мере один фармацевтически приемлемый носитель. Такой носитель может представлять собой, например, воду, буфер, PBS, раствор глюкозы, предпочтительно, 5% сбалансированный солевой раствор глюкозы, крахмал, сахар, желатин или любой другой приемлемый носитель. Такие носители обычно известны специалистам в данной области. Специалисту в данной области будет понятно, что любые варианты осуществления, использование и аспекты лекарственного средства по настоящему изобретению, или варианты осуществления, использование и аспекты, связанные с ним, также могут использоваться для фармацевтической композиции по настоящему изобретению и наоборот.
Показания, заболевания и нарушения, для лечения и/или профилактики которых подходят нуклеиновые кислоты, фармацевтические композиции и лекарственные средства по настоящему изобретению или полученные по настоящему изобретению, являются следствием участия, либо непосредственного, либо косвенного, MCP-1 в соответствующем механизме патогенеза. Однако, лечение и профилактика этих показаний, заболеваний и нарушений также можно проводить, если в патогенный механизм напрямую или косвенно вовлечены MCP-2, MCP-3, MCP-4 и/или эотаксин. Для специалиста в данной области очевидно, что, в частности, можно использовать те нуклеиновые кислоты по настоящему изобретению, например, при заболеваниях, связанных в широком смысле с MCP-2, MCP-3, MCP-4 и эотаксином, которые взаимодействуют и связываются, соответственно, с MCP-2, MCP-3, MCP-4 и эотаксином, соответственно.
Более конкретно, такое применение является следствием, среди прочих, характера экспрессии MCP-1, который указывает на то, что он играет важную роль в развитии заболеваний человека, которые характеризуются инфильтрацией мононуклеарных клеток. Такая инфильтрация клеток происходит при большом числе воспалительных и аутоиммунных заболеваниях.
На моделях животных было показано, что MCP-1 экспрессируется в головном мозге после очаговой ишемии (Kim 1995; Wang 1995) и при экспериментальном аутоиммунном энцефаломиелите (Hulkower 1993; Ransohoff 1993; Banisor 2005). MCP-1 может быть важным хемокином, который нацелен на мононуклеарные клетки, при заболевании, моделируемом на этих животных, таком как инсульт и рассеянный склероз.
Большое количество данных свидетельствуют в пользу уникальной роли пути MCP-1/CCR2 в хемоатракции моноцитов и, таким образом, в хроническом воспалении: (i) дефицитные по MCP-1 или CCR2 мыши демонстрируют в значительной степени сниженный хемотактический ответ макрофагов, а в остальном выглядят нормальными (Kuziel 1997; Kurihara 1997; Boring 1997; Lu 1998). (ii) Несмотря на функциональное дублирование другими хемокинами in vitro, утрата эффекторной функции только MCP-1 является достаточной для нарушения миграции моноцитов в некоторых моделях воспаления (Lloyd 1997; Furuichi 2003; Egashira 2002; Galasso 2000; Ogata 1997; Kennedy 1998; Gonzalo 1998; Kitamoto 2003). (iii) Уровни MCP-1 являются повышенными при многих воспалительных заболеваниях. В действительности, полагают, что MCP-1 участвует в развитии большого числа заболеваний с очевидным воспалительным компонентом или без него, таких как ревматоидный артрит (Koch 1992; Hosaka 1994; Akahoshi 1993; Harigai 1993; Rollins 1996), заболевание почек (Wada 1996; Viedt 2002), рестеноз после ангиопластики (Economou 2001), аллергия и астма (Alam 1996; Holgate 1997; Gonzalo 1998), злокачественная опухоль (Salcedo 2000; Gordillo 2004), атеросклероз (Nelken 1991; Yla-Herttuala 1991; Schwartz 1993; Takeya 1993; Boring 1998), псориаз (Vestergaard 2004), воспаление нервной системы (Huang 2001), атопический дерматит (Kaburagi 2001), колит (Okuno 2002), эндометриоз (Jolicoeur 2001), увеит (Tuaillon 2002), нарушения сетчатки (Nakazawa2007), идиопатический фиброз и саркоидоз легких (Iyonaga 1994) и полимиозит/дерматомиозит (De Bleecker 2002).
Терапевтическое воздействие средствами против MCP-1 или антагонистов CCR2 - может влиять на избыточную воспалительную миграцию моноцитов, но может сохранить базовую миграцию фагоцитов, тем самым позволяя избежать общей иммуносупрессии и повышенного риска инфекций (Dawson 2003).
Кроме того, исходя из растущих знаний о молекулярных механизмах воспалительного процесса и взаимодействии локально секретируемых медиаторов воспаления, были идентифицированы новые мишени для терапии заболеваний почек (Holdsworth 2000; Segerer 2000). Одной из этих мишеней, для которых существуют надежные данные из исследований экспрессии и исследований со вмешательством с помощью конкретных антагонистов в соответствующих моделях на животных, является MCP-1. Этот белок является далеко не излишним в привлечении иммунных клеток в области воспаления почек. Полагают, что инфильтрация иммунных клеток в почку является основным механизмом структурного повреждения почки и снижения почечной функции при развитии различных форм болезни почек.
При стимуляции in vitro все типы клеток почек могут экспрессировать хемокины, включая MCP-1 (Segerer 2000); существует большой перечень стимулов, которые запускают экспрессию MCP-1 in vitro, включая цитокины, радикалы кислорода, иммунные комплексы и липидные медиаторы.
В здоровых почках крыс и мышей MCP-1 не экспрессируется, но легко активируется в ходе острого и хронического воспаления почек на моделях грызунов, включая иммунокомплексный гломерулонефрит, быстрый прогрессирующий гломерулонефрит, пролиферативный гломерулонефрит, диабетическую нефропатию, обструктивную нефропатию или острый тубулярный некроз (Segerer 2000; Anders 2003). Данные по экспрессии MCP-1 у грызунов высоко коррелируют с соответствующей экспрессией, выявленной в биоптатах почек человека (Rovin 1994; Cockwell 1998; Wada 1999). Более того, экспрессия в почках человека связана с активностью заболевания и снижается, если соответствующая терапия приводит к ремиссии заболевания (Amann 2003).
Инфильтрация гломерул мононуклеарными клетками связана с развитием диффузного гломерулосклероза у пациентов с диабетической нефропатией. MCP-1 играет важную роль в привлечении и накоплении моноцитов и лимфоцитов в гломерулах (Banba 2000; Morii 2003).
Локально продуцируемый MCP-1, по-видимому, особенно тесно связан с инициацией и прогрессированием тубулоинтерстициального повреждения, что показано в экспериментах с использованием трансгенных мышей с индуцированным нефротоксической сывороткой нефритом (NSN). В интерстициальных очагах повреждения MCP-1 был выявлен, главным образом, в эндотелиальных клетках сосудов, эпителиальных клетках канальцев и в инфильтрированных мононуклеарных клетках. Опосредуемая MCP-1 активация эпителиальных клеток канальцев согласуется с точкой зрения, что MCP-1 приводит к тубулоинтерстициальному воспалению, признаку прогрессирующего заболевания почек (Wada 2001; Viedt 2002).
Вследствие гомологии между MCP-1 с одной стороны и MCP-2, MCP-3, MCP-4 и эотаксином с другой стороны, нуклеиновые кислоты по настоящему изобретению, по меньшей мере те из них, которые взаимодействуют или связываются с MCP-2, MCP-3, MCP-4 и эотаксином, соответственно, можно, главным образом, использовать для лечения, профилактики и/или диагностики любого заболевания, в которое напрямую или косвенно вовлечены MCP-2, MCP-3, MCP-4 и эотаксин, соответственно. "Вовлеченный", как предпочтительно используют в настоящем документе, означает, что если предотвратить одну, несколько или все из функций, связанных с патогенным механизмом, лежащим в основе заболевания, соответствующей молекулы, которая вовлечена в заболевание, то заболевание будет вылечено или его степень будет снижена или будет предотвращена его вспышка; по меньшей мере симптомы или любой показатель такого заболевания будут снижены или улучшены, соответственно, так чтобы симптомы и показатель, соответственно, были идентичными или сходными с симптомами и показателем, наблюдаемыми у индивида, не страдающего заболеванием или не имеющего риска развития такого заболевания.
Безусловно, поскольку связывающие MCP-1 нуклеиновые кислоты по настоящему изобретению взаимодействуют или связываются с MCP-1 человека или мыши, специалисту в данной области, главным образом, будет понятно, что связывающие MCP-1 нуклеиновые кислоты по настоящему изобретению можно легко использовать для лечения, профилактики и/или диагностики любого заболевания, как описано в настоящем документе, человека и животных.
Эти члены семейства хемоаттрактантных белков моноцитов (MCP), т.е. MCP-2, MCP-3, MCP-4 и эотаксин, таким образом, обладают высокой степенью сходства последовательностей с MCP-1. Хотя не исключительно, эотаксин, MCP-2, -3 и -4 действуют через CCR3, характерный рецептор хемокинов на эозинофилах человека (Heath 1997). Рецептор CCR3 активируется при неопластических состояниях, таких как Т-клеточная лимфома кожи (Kleinhans 2003), глиобластома (Kouno 2004) или почечноклеточный рак (Johrer 2005).
Более конкретно, повышенные уровни эотаксина напрямую связаны с диагнозом астма и нарушенной функцией легких (Nakamura 1999). Повышенную экспрессию эотаксина в участках аллергического воспаления наблюдали как у атопических, так и у неатопических астматиков (Ying 1997; Ying 1999). Также, мРНК, кодирующие MCP-2 и -4, конститутивно экспрессируются в различных тканях; однако их физиологические функции в этом контексте неизвестны. Уровни MCP-2 в плазме повышены при сепсисе вместе с MCP-1 (Bossink 1995); MCP-3 экспрессируется у астматиков (Humbert 1997). Наконец, MCP-4 может быть обнаружен на обращенной к просвету поверхности атеросклеротических сосудов (Berkhout 1997).
Таким образом, заболевания и/или нарушения и/или болезненные состояния, для лечения и/или профилактики которых можно использовать лекарственное средство по настоящему изобретению, включают, но ими не ограничиваются, воспалительные заболевания, аутоиммунные заболевания, аутоиммунный энцефаломиелит, инсульт, острый и хронический рассеянный склероз, хроническое воспаление, ревматоидный артрит, заболевания почек, рестеноз, рестеноз после ангиопластики, острые и хронические аллергические реакции, первичные и вторичные иммунологические или аллергические реакции, астму, конъюнктивит, бронхит, злокачественную опухоль, атеросклероз, артериосклеротическую сердечно-сосудистую недостаточность или инсульт, псориаз, псориатический артрит, воспаление нервной системы, атопический дерматит, колит, эндометриоз, увеит, нарушения сетчатки, включая дегенерацию желтого пятна, отслоение сетчатки, диабетическую ретинопатию, ретролентальную фиброплазию, пигментный ретинит, пролиферативную витреоретинопатию, и центральную серозную хориоретинопатию; идиопатический фиброз легких, идиопатическую и/или связанную с коллагенозом гипертензию легочной артерии, саркоидоз, полимиозит, дерматомиозит, уклонение от иммуносупрессии, снижение риска инфекции, сепсис, воспаление почек, гломерулонефрит, быстрый прогрессирующий гломерулонефрит, пролиферативный гломерулонефрит, диабетическую нефропатию, обструктивную нефропатию, острый тубулярный некроз, и диффузный гломерулосклероз, системную красную волчанку, хронический бронхит, болезнь Бехчета, боковой амиотрофический склероз (ALS), преждевременный атеросклероз после болезни Кавасаки, инфаркт миокарда, ожирение, хроническую болезнь печени, болезнь Пейрони, острое повреждение спинного мозга, трансплантацию легкого или почки, миокардит, болезнь Альцгеймера и невропатию, карциному молочной железы, карциному желудка, рак мочевого пузыря, рак яичника, гемартому, карциному ободочной и прямой кишки, аденому толстой кишки, панкреатит, хроническую обструктивную болезнь легких (COPD) с вовлечением сосудов легких или без него, и воспалительные заболевания кишечника, такие как болезнь Крона или язвенный колит.
Особенно предпочтительно хроническое заболевание почек представляет собой волчаночный нефрит, предпочтительный для комбинированной терапии.
Волчаночный нефрит представляет собой воспаление почки, вызываемое системной красной волчанкой (сокращенно SLE), а также оно известно как волчаночный гломерулонефрит, тип или форма гломерулонефрита. Гломерулонефрит, также известный как гломерулярный нефрит, представляет собой заболевание почек, характеризующееся воспалением гломерул или небольших кровеносных сосудов в почках.
SLE, также известная как волчанка, представляет собой хроническое аутоиммунное заболевание, приводящее к воспалению и повреждению тканей. Помимо почек, SLE может поражать любую часть организма, но наиболее часто она повреждает сердце, суставы, кожу, легкие, кровеносные сосуды, печень и нервную систему. Повреждение легких проявляется респираторными заболеваниями, такими как пневмония, легочные проявления могут включать воспаление легких и плевры, которое может вызывать плеврит, плевральный выпот, волчаночную пневмонию, хроническую диффузную интерстициальную болезнь легких, легочную гипертензию, эмболию легких, кровоизлияние в легкие и синдром сморщенного легкого.
Диагноз волчаночного нефрита, как правило, основан на анализах крови, анализе мочи, рентгенограмме, ультразвуковом исследовании почки и/или биопсии почки.
Всемирная организация здравоохранения подразделила волчаночный нефрит на пять классов, на основании биопсии, все из которых охватываются термином волчаночный нефрит, как используют в настоящем документе.
Эта классификация была проведена в 1982 году и пересмотрена в 1995 году.
- Класс I представляет собой минимальный мезангиальный гломерулонефрит, который является гистологически нормальным при световой микроскопии, но имеет отложения в мезангии при электронной микроскопии.
- Класс II основан на обнаружении мезангиального пролиферативного волчаночного нефрита. Эта форма, как правило, полностью отвечает на лечение кортикостероидами.
- Класс III представляет собой очаговый пролиферативный нефрит и часто успешно отвечает на лечение высокими дозами кортикостероидов.
- Класс IV представляет собой диффузный пролиферативный нефрит. Эту форму, главным образом, лечат кортикостероидами и иммунодепрессивными лекарственными средствами.
- Класс V представляет собой мембранозный нефрит и он характеризуется крайней степенью отека и потери белка.
- Класс VI - гломерулосклероз.
Другими типами или формами гломерулонефрита при заболеваниях почек являются
- Мембрано-пролиферативный гломерулонефрит (сокращенно MPGN) - тип гломерулонефрита, вызываемый отложениями в гломерулярном мезангии почки и утолщением базальной мембраны (сокращенно GBM), активацией комплемента и повреждением гломерул;
- Мембранозный гломерулонефрит (сокращенно MGN), также известный как мембранозная нефропатия - медленно прогрессирующее заболевание почек;
- IgA-нефропатия, наиболее распространенный в мире гломерулонефрит; название заболевания произошло вследствие отложений иммуноглобулина A (сокращенно IgA) в виде пятен в мезангии, центральной части гломерул.
- Пост-стрептококковый гломерулонефрит, нарушение гломерул (гломерулонефрит) или небольших кровеносных сосудов в почках после стрептококковой инфекции;
- Быстро прогрессирующий гломерулонефрит (сокращенно RPGN), синдром почки, который, без лечения, быстро прогрессирует в острую почечную недостаточность и приводит к гибели в течение нескольких месяцев. В 50% случаев RPGN связан с лежащим в его основе заболеванием, таким как синдром Гудпасчера, системная красная волчанка или гранулематоз Вегенера; остальные случаи являются идиопатическими;
- Нефритический синдром (или острый нефритический синдром), который скорее означает набор признаков, связанных с нарушениями, поражающими почки, более конкретно, гломерулярными нарушениями;
- Фокальный сегментный гломерулосклероз, причина нефротического синдрома у детей и подростков, а также существенная причина почечной недостаточности у взрослых;
- Диабетическая нефропатия, также известная как синдром Киммелстила-Уилсона и интеркапиллярный гломерулонефрит, представляет собой прогрессирующее заболевание почки, вызываемое ангиопатией капилляров в гломерулах почек. Она характеризуется нефротическим синдромом и узловым гломерулосклерозом. Она является следствием продолжительного сахарного диабета, и представляет собой основную причину диализа во многих западных странах;
- Нефротический синдром, неспецифическое нарушение, при котором почки повреждаются, что вызывает выход через них больших количеств белка (> 3,5 грамм в сутки на площадь поверхности тела, составляющую 1,73 м2) в мочу;
- Интерстициальный нефрит (или тубуло-интерстициальный нефрит) - форма нефрита, поражающая интерстиций почек, окружающий канальцы. Это заболевание может быть как острым, так и хроническим.
Существуют убедительные данные, что MCP-1 и его соответствующий рецептор хемокинов CCR2 играют важную роль в аутоиммунном повреждении ткани, таком как клинические проявления SLE (Gerard & Rollins 2001). Например, мыши MRLlpr/lpr, дефицитные либо по гену MCP-1, либо по гену CCR2, защищены от подобного волчанке аутоиммунитета (Perez de Lema 2005, Tesch 1999). Таким образом, ось MCP-1/CCR2 может представлять собой перспективную терапевтическую мишень, например, от волчаночного нефрита.
Хронические респираторные заболевания, также известные как хронические легочные заболевания или хронические заболевания легких, представляют собой хронические заболевания дыхательных путей и других структур легкого, например, таких как сосуды легкого. Некоторые из наиболее распространенных хронических респираторных заболеваний представляют собой астму, хроническую обструктивную болезнь легких (сокращенно COPD), респираторную аллергию, профессиональные заболевания легких и легочную гипертензию.
Хроническая обструктивная болезнь легких (сокращенно COPD) представляет собой болезнь легких, которая характеризуется постоянной блокадой выхода струи воздуха из легких. Она представляет собой плохо диагностируемое угрожающее жизни заболевание легких, которое препятствует нормальному дыханию и не является полностью обратимым. COPD включает несколько заболеваний легких: наиболее распространенными являются хронический бронхит и эмфизема. Многие люди с COPD имеют оба из этих заболеваний. Эмфизема представляет собой повреждение альвеолярных мешочков на концах дыхательных путей, что затрудняет получение организмом требуемого количества кислорода. В ходе хронического бронхита воздухоносные пути являются раздраженными, красными и образуют слишком большое количество липкой слизи. Стенки воздухоносных путей набухают и практически блокируют прохождение воздуха через них.
Вовлечение MCP-1 в COPD и/или развитие COPD не было очевидным, пока De Boer и коллеги не выявили в полуколичественном анализе периферических тканей легкого курящего на тот момент времени или курящего в прошлом человека с COPD повышение уровней мРНК MCP-1 в 1,5 раза (De Boer 2000). На основании этих результатов, авторы предположили, что при COPD MCP-1 может быть вовлечен в привлечение макрофагов и тучных клеток в эпителий воздухоносных путей. Traves et al (Traves 2002) выявили повышенные уровни MCP-1 в мокроте, но не в жидкости бронхоальвеолярного лаважа (сокращенно BAL), и предположили, что MCP-1 вовлечен в миграцию моноцитов и нейтрофилов в дыхательные пути, приводящую к повышенной воспалительной нагрузке, связанной с COPD (Traves 2002). Однако в 2006 году Ko et al. (Ko 2006) исследовали конденсат в выдыхаемом воздухе у пациентов с COPD. Они не смогли выявить повышенных уровней MCP-1 у пациентов COPD (Ko 2006).
Несмотря на то, что MCP-1 вовлечен в процесс воспаления и привлечение моноцитов и/или нейтрофилов, которые вызывают воспаление, не было абсолютно очевидно, вовлечен ли MCP-1 в COPD и/или развитие COPD. Неожиданно, как показано в примере 11 по настоящему изобретению, в общепризнанной модели на животных, которую широко используют для скрининга веществ в отношении пригодности для лечения COPD, введение связывающего MCP-1 шпигельмера приводило к снижению клеточной инфильтрации в легкие. На основе данных, представленных в настоящей заявке, связывающие MCP-1 шпигельмеры пригодны для применения в терапии хронических респираторных заболеваний, предпочтительно, COPD, отдельно или в качестве одного из элементов комбинированной терапии, предпочтительно, комбинированной терапии стероидным лекарственным средством, предпочтительно, комбинированной терапией дексаметазоном, и связывающими MCP-1 шпигельмерами, причем дексаметазон или другие лекарственные средства имеют преимущество двух независимых способов действия для лечения респираторных заболеваний, таких как COPD.
Изменения в структуре и функции легочных сосудов в высокой степени распространены у пациентов с COPD (Peinado 2008), называемой в настоящем документе как COPD с вовлечением легочных сосудов. Аномалии сосудов нарушают газообмен и могут приводить к легочной гипертензии, которая является одним из основных факторов, связанных со сниженной выживаемостью пациентов с COPD (Peinado 2008). Изменение кровотока в легких идентифицируют на начальных стадиях заболевания, обеспечивая новое понимание его патогенеза. Повреждение эндотелиальных клеток и дисфункцию, вызываемую действием продуктами сигаретного дыма или воспалительными элементами, в настоящее время считают первичным изменением, которое запускает последовательность событий, приводящую к легочной гипертензии (Peinado 2008).
Легочная гипертензия (сокращенно PH) представляет собой повышение кровяного давления в легочной артерии, легочной вене или капиллярах легких, в совокупности известных как сосуды легких, приводящее к одышке, головокружению, потерям сознания и другим симптомам, все из которых усиливаются при напряжении. PH может быть тяжелым заболеванием со значительно сниженной переносимостью нагрузки и сердечной недостаточностью. С 1973 года было проведено разделение между первичной PH и вторичной PH (Hatano & Strasser 1975).
Первичная PH представляет собой синдром, характеризующийся хронически повышенной резистентностью сосудов легких в отсутствии известной причины, которая, без лечения, обычно приводит к смерти в течение четырех лет (Rubin 1997).
Вторичная PH является следствием длительного сужения сосудов и структурных изменений сосудистого ложа легких (Hopkins 2002). Основными стимулами, которые отвечают за эти изменения, являются хроническая альвеолярная гипоксия, хроническое воспаление и избыточное напряжение сдвига (Voelker 1995).
В 2003 году, в Венеции был созван 3-ий Всемирный симпозиум по гипертензии легочной артерии для изменения классификации на основе новых представлений о механизмах заболевания. Пересмотренная система, разработанная этой группой, обеспечивает современный каркас для понимания легочной гипертензии (Simonneau 2004):
- Группа I ВОЗ - Гипертензия легочной артерии (сокращенно PAH)
- Идиопатическая гипертензия легочной артерии (сокращенно IPAH)
- Семейная гипертензия легочной артерии (сокращенно FPAH)
- Гипертензия легочной артерии, связанная с другими заболеваниями (сокращенно APAH), где другими заболеваниями являются коллагеновая болезнь сосудов (например, склеродермия), врожденный шунт между системным и легочным кровотоком, портальная гипертензия, ВИЧ-инфекция, связанные с лекарственными средствами, токсинами или другие заболевания или нарушения
- Гипертензия легочной артерии, связанная с заболеванием вен или капилляров
- Группа II ВОЗ - Легочная гипертензия, связанная с болезнью левого отдела сердца
- Заболевание предсердий или желудочков
- Заболевание клапанов (например, стеноз митрального клапана)
- Группа III ВОЗ - Легочная гипертензия, связанная с заболеваниями легких и/или гипоксемией
- Хроническая обструктивная болезнь легких (сокращенно COPD), интерстициальное заболевание легких (сокращенно ILD)
- Нарушенное дыхание во сне, альвеолярная гиповентиляция
- Хроническое воздействие больших высот
- Связанные с развитием аномалии легких
- Группа IV ВОЗ - Легочная гипертензия вследствие хронического тромботического и/или эмболического заболевания
- Легочная эмболия в проксимальных или дистальных легочных артериях
- Эмболия другого типа, такая как эмболия клетками опухоли или паразитами
- Группа V ВОЗ - Прочее
Для первичной и вторичной PAH недавно был разработан ряд средств: производное простациклина, такое как эпопростенол, антагонист рецептора эндотелина, такой как босентан, и ингибитор фосфодиэстеразы 5 типа, такой как силденафил (Torres 2007).
У пациентов с идиопатической гипертензией легочной артерии или первичной легочной гипертензией уровни MCP-1 являются повышенными по сравнению со здоровыми контролями. Эти результаты подразумевают участие MCP-1 в развитии легочной гипертензии (Itoh 2006). Как показано в примере 12, связывающие MCP-1 шпигельмеры демонстрируют положительные эффекты при PH на модели животных, которую широко используют для скрининга веществ, пригодных для лечения легочной гипертензии. Таким образом, связывающие MCP-1 шпигельмеры пригодны в качестве средств для лечения PH.
В следующем варианте осуществления, лекарственное средство содержит дополнительное фармацевтически активное средство. Такие дополнительные фармацевтически активные соединения представляют собой, среди прочих, но не ограничиваясь ими, соединения, известные тем, что они контролируют кровяное давление и диабет, такие как ингибиторы ангиотензинпревращающего фермента (ACE) и блокаторы рецептора ангиотензина. Дополнительное фармацевтически активное соединение может представлять собой, в следующем варианте осуществления, также одно из соединений, которые снижают инфильтрацию иммунных клеток в области хронического воспаления или, в общем, подавляют избыточный иммунный ответ, который существует в условиях хронического воспаления и который приводит к повреждению тканей. Такие соединения могут представлять собой, но не ограничиваться ими, стероиды или иммунодепрессивные средства и предпочтительно их выбирают из группы, содержащей кортикостероиды, такие как преднизон, метилпреднизолон, гидрокортизон, дексаметазон и общие иммунодепрессанты, такие как циклофосфамид, циклоспорин, хлорамбуцил, азатиоприн, такролимус или микофенолата мофетил. Кроме того, в следующих вариантах осуществления пригодны более специфические блокаторы костимуляции T-клеток, например блокаторы CD154 или CD40, или CD28, или CD86, или CD80; или средства, устраняющие T- и/или B-клетки, такие как средство против CD20. Наконец, дополнительное фармацевтически активное средство может представлять собой модулятор активности любого другого хемокина, который может представлять собой агонист или антагонист хемокина или агонист или антагонист рецептора хемокина. Альтернативно или дополнительно, такое дополнительное фармацевтически активное средство представляет собой дополнительную нуклеиновую кислоту по настоящему изобретению. Альтернативно лекарственное средство содержит по меньшей мере одну дополнительную нуклеиновую кислоту, которая связывается с молекулой-мишенью, отличной от MCP-1, или проявляет функцию, отличную от функции одной из нуклеиновых кислот по настоящему изобретению.
Настоящее изобретение относится к тому, что лекарственное средство используют, альтернативно или дополнительно, в принципе, для профилактики любого из заболеваний, описанных применительно к применению лекарственного средства для лечения указанных заболеваний. Таким образом, соответствующие маркеры, т.е. для соответствующих заболеваний, известны специалистам в данной области. Предпочтительно, соответствующим маркером является MCP-1. Альтернативно и/или дополнительно, соответствующий маркер выбран из группы, содержащей MCP-2, MCP-3, MCP-4 и эотаксин. Следующая группа маркеров выбрана из группы, содержащей аутореактивные антитела в плазме, например, такие как антитела против дцДНК или ревматоидного фактора.
В одном из вариантов осуществления лекарственного средства по настоящему изобретению, такое лекарственное средство предназначено для применения в сочетании с другими способами лечения любых из заболеваний, описанных в настоящем документе, в частности, заболеваний, для которых намереваются использовать лекарственное средство по настоящему изобретению.
"Комбинированная терапия" (или "ко-терапия") включает введение лекарственного средства по изобретению и по меньшей мере второго средства в качестве части конкретного режима лечения, предназначенного для обеспечения благоприятного эффекта вследствие совместного действия этих лекарственных средств, т.е. лекарственного средства по настоящему изобретению и указанного второго средства. Благоприятный эффект комбинации включает, но не ограничивается ими, фармакокинетическое или фармакодинамическое совместное действие в результате комбинирования лекарственных средств. Введение этих лекарственных средств в комбинации, как правило, проводят в течение определенного периода (как правило, минут, часов, суток или недель, в зависимости от выбранной комбинации).
"Комбинированная терапия" может включать, но обычно не включает, введение двух или более из этих лекарственных средств в качестве частей отдельных схем монотерапии, которые случайным образом и произвольно приводят к комбинациям по настоящему изобретению. "Комбинированная терапия" включает введение этих лекарственных средств последовательно, т.е., где каждое лекарственное средство вводят в отдельное время, а также введение этих лекарственных средств, или по меньшей мере двух из этих лекарственных средств, по существу одновременно. По существу одновременное введение можно проводить, например, путем введения индивиду одной капсулы, имеющей фиксированную долю каждого лекарственного средства, или нескольких отдельных капсул для каждого из лекарственных средств.
В предпочтительном варианте осуществления, хроническое заболевание представляет собой либо хроническое заболевание почек, и более предпочтительно, волчаночный нефрит, либо хроническое заболевание легких, и более предпочтительно, пневмонию, которые подлежат лечению с использованием комбинированной терапии. В такой комбинированной терапии используется комбинация молекулы нуклеиновой кислоты, как описано в настоящем документе, и иммунодепрессивного средства. Предпочтительно, иммунодепрессивное средство выбрано из группы, содержащей циклофосфамид и микофенолата мофетил.
Циклофосфамид (непатентованное название для цитоксана, неосара, ревиммуна), также известный как цитофосфан, представляет собой алкилирующее средство на основе азотистого иприта, из группы оксазофоринов. Внутривенное и пероральное введение циклофосфамида является стандартом для лечения волчаночного гломерулонефрита (Steinberg 1991). Циклофосфамид представляет собой "пролекарство", которое превращается в печени в активные формы, которые имеют химиотерапевтическую активность. Действительно, применение циклофосфамида ограничивается потенциально тяжелыми токсическими эффектами, включающими подавление костного мозга, геморрагический цистит, оппортунистические инфекции, злокачественные заболевания и преждевременную недостаточность половых желез (Boumpas 1995). Клинические испытания лечения периодическим внутривенным циклофосфамидом в комбинации с кортикостероидами демонстрирует более продолжительную выживаемость почек, но не общую выживаемость, по сравнению с лечением кортикостероидами отдельно (Austin 1986; Valeri 1994; Lehman 1989; Boumpas 1992). Более того, неуспех в достижении ремиссии, который связан с увеличенной скоростью прогрессирования в почечную недостаточность, описан для 18-57 процентов пациентов, получавших циклофосфамид (Korbert 2000; Gourley 1996; Ionnidis 2000; Mok 2004).
Микофенолата мофетил представляет собой иммунодепрессивное средство, которое метаболизируется в печени до активной группы микофеноловой кислоты. Он ингибирует инозинмонофосфатдегидрогеназу, фермент, который контролирует скорость синтеза гуанинмонофосфата в каскаде синтеза пуринов de novo, используемом при пролиферации B- и T-лимфоцитов. Микофенолата мофетил, одобренный для профилактики отторжения трансплантата, использовали у пациентов с волчаночным нефритом, которые являются рефрактерными к циклофосфамиду, и у пациентов, которые не переносят циклофосфамид (Dooley 1990; Gaubitz 1999; Kingdon 2001; Karim 2002). В 4-недельном испытании, микофенолата мофетил был более эффективным, чем внутривенный циклофосфамид, в отношении индукции ремиссии волчаночного нефрита (Ginzler 2005).
Однако каждое из этих двух лекарственных средств связано со значительной заболеваемостью и смертностью. Например, в исследовании Aspreva Lupus Management Study (ALMS) испытуемый микофенолата мофетил вызывал побочные эффекты у 27,7% и связанную с лечением смерть в 4,9%, а циклофосфамид - у 22,8% и 2,8% подвергнутых лечению пациентов, соответственно (Appel 2007). Наиболее тяжелые побочные эффекты и смерти были связаны с инфекциями вследствие неспецифических иммуносупрессивных эффектов циклофосфамида и микофенолата мофетилса (Appel 2007). Новые лекарственные средства, специфично блокирующие аутоиммунное воспаление, могут позволить снизить токсичность современных протоколов лечения либо путем замены циклофосфамида и микофенолата мофетила, либо путем допущения существенных снижений доз при использовании в комбинации.
Применительно к такой комбинированной терапии, является возможным значительное снижение общего количества иммунодепрессивного средства, при достижении, тем не менее, терапевтического эффекта. Снижение общего количества иммунодепрессивного средства может быть осуществлено путем уменьшения количества иммунодепрессивного средства при каждом введении, или путем снижения частоты введения иммунодепрессивного средства при лечении заболевания. Независимо от того, какой из двух указанных вариантов используют, в любом случае общее количество иммунодепрессивного средства, которое вводят пациенту в ходе лечения, снижается по сравнению с общим количеством иммунодепрессивного средства, вводимого при лечении пациенту введением только иммунодепрессивного средства вместо комбинации иммунодепрессивного средства и молекулы нуклеиновой кислоты по настоящему изобретению. Такое введение иммунодепрессивного средства применительно к лечению указанного заболевания в качестве единственного фармацевтически активного средства также называют в настоящем документе монотерапией. Степень такого уменьшения зависит от конкретного иммунодепрессивного средства и конкретного заболевания, а также от индивидуальных характеристик пациента, подлежащего лечению. В любом случае, комбинированная терапия по настоящему изобретению протекает с меньшими побочными эффектами по сравнению с использованием соответствующего иммунодепрессивного средства при монотерапии.
Следующим предпочтительным вариантом осуществления комбинированной терапии с использованием в качестве одного фармацевтически активного средства молекулы нуклеиновой кислоты по настоящему изобретению, является комбинированная терапия применительно к лечению хронических респираторных заболеваний, где хроническое респираторное заболевание предпочтительно представляет собой COPD. Средство, подлежащее применению в указанной комбинированной терапии молекулой нуклеиновой кислоты по настоящему изобретению, представляет собой противовоспалительное средство. Предпочтительно, противовоспалительное средство выбрано из группы, содержащей дексаметазон и рофлумиласт; более предпочтительно, указанным противовоспалительным средством является дексаметазон.
Последовательное или по существу одновременное введение каждого лекарственного средства можно осуществлять любым пригодным способом, включая, но не ограничиваясь ими, местные способы, пероральные способы, внутривенные способы, внутримышечные способы и прямое всасывание через ткани слизистых оболочек. Лекарственные средства можно вводить одинаковыми или различными способами. Например, первое из выбранных лекарственных средств комбинации можно вводить путем инъекции, а другие лекарственные средства комбинации можно вводить местным способом.
Альтернативно, например, все лекарственные средства можно вводить местно или все лекарственные средства можно вводить путем инъекции. Последовательность, в которой вводят лекарственные средства, не является строго критичной, если нет иных указаний. "Комбинированная терапия" также может включать введение лекарственных средств, как описано выше, дополнительно в комбинации с другими биологически активными ингредиентами. Когда комбинированная терапия дополнительно включает немедикаментозное лечение, немедикаментозное лечение можно проводить в любое пригодное время, при условии достижения благоприятного эффекта совместного действия комбинации лекарственных средств и немедикаментозного лечения. Например, в соответствующих случаях, благоприятный эффект все еще достигается, когда немедикаментозное лечение удалено по времени от введения лекарственных средств, возможно на сутки или даже недели.
Как указано в общих терминах, выше, лекарственное средство по настоящему изобретению можно вводить, в принципе, в любой форме, известной специалистам в данной области. Предпочтительным способом введения является системное введение, более предпочтительно, парентеральное введение, предпочтительно, инъекция. Альтернативно лекарственное средство можно вводить местно. Другие способы введения включают внутримышечный, внутрибрюшинный и подкожный, пероральный, интраназальный, интратрахеальный или легочный, причем предпочтительным является способ введения, который является наименее инвазивным, но в то же время обеспечивающим эффективность.
Парентеральное введение, как правило, используют для подкожных, внутримышечных или внутривенных инъекций и инфузий. Кроме того, в одном подходе для парентерального введения используется имплантация систем с замедленным высвобождением или непрерывным высвобождением, которые обеспечивают поддержание дозирования на постоянном уровне и которые хорошо известны специалистам в данной области.
Более того, предпочтительные лекарственные средства по настоящему изобретению можно вводить в интраназальной форме посредством местного применения пригодных интраназальных носителей, ингаляторов, или трансдермальных способов с использованием форм трансдермальных накожных пластырей, хорошо известных специалистам в данной области. Для введения в форме трансдермальной системы для доставки, введение дозы, безусловно, будет непрерывным, а не прерывающимся, на протяжении режима дозирования. Другие предпочтительные местные препараты включают кремы, мази, лосьоны, аэрозольные спреи и гели, где концентрация активного ингредиента, как правило, может находиться в диапазоне от 0,01% до 15% масс./масс. или масс./об.
Лекарственное средство по настоящему изобретению, как правило, содержит эффективное количество активного компонента(ов) терапии, включая, но не ограничиваясь ими, молекулу нуклеиновой кислоты по настоящему изобретению, растворенную или диспергированную в фармацевтически приемлемой среде. Фармацевтически приемлемые среды или носители включают любые и все растворители, диспергирующие среды, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие всасывание средства и т.п. Применение таких сред и средств для фармацевтически активных веществ хорошо известно в данной области. Также в лекарственное средство по настоящему изобретению могут быть включены дополнительные активные ингредиенты.
В следующем аспекте настоящее изобретение относится к фармацевтической композиции. Такая фармацевтическая композиция содержит по меньшей мере одну из нуклеиновых кислот по настоящему изобретению и предпочтительно фармацевтически приемлемый носитель. Такой носитель может представлять собой любой носитель или любое связывающее вещество, используемое и/или известное в данной области. Более конкретно такое связующее вещество или носитель представляет собой любое связующее вещество или носитель, рассмотренный в связи с изготовлением лекарственного средства, описанного в настоящем документе. В следующем варианте осуществления, фармацевтическая композиция содержит дополнительное фармацевтически активное средство.
Изготовление лекарственного средства и фармацевтической композиции станет известным специалистам в данной области с учетом настоящего описания. Как правило, такие композиции можно получать в качестве инъецируемых средств, в форме либо жидких растворов, либо суспензий; твердых форм, пригодных для растворения или суспендирования в жидкости перед инъекцией; в качестве таблеток или других твердых веществ для перорального введения; в качестве капсул для замедленного высвобождения; или в любой другой форме, применяемой в настоящее время, включая глазные капли, кремы, лосьоны, бальзамы, ингалируемые средства и т.п. Также может быть особенно пригодным применение стерильных составов, таких как средства для промывания на основе солевого раствора, хирургами, терапевтами или работниками по медицинскому уходу для обработки конкретной области операционного поля. Также композиции можно доставлять с помощью микроустройства, микрочастицы или губки.
После изготовления лекарственное средство можно вводить способом, совместимым с дозированным составом, и в таком количестве, которое является фармакологически эффективным. Составы легко вводят в различных дозированных формах, таких как тип инъецируемых растворов, описанный выше, однако также можно использовать капсулы для высвобождения лекарственного средства.
В этом контексте, количество активного ингредиента и объем композиции, подлежащей введению, зависит от индивида или индивида, подлежащего лечению. Конкретные количества активного соединения, требуемые для введения, зависят от решения специалиста и являются индивидуальными для каждого индивида.
Как правило, используют минимальный объем лекарственного средства, требуемый для диспергирования активных соединений. Пригодные режимы для введения также варьируют, но в качестве примера может быть приведено исходное введение соединения и мониторинг результатов, а затем назначение дополнительных контролируемых доз через дополнительные промежутки времени.
Например, для перорального введения в форме таблетки или капсулы (например, желатиновой капсулы), активный лекарственный компонент, т.е. молекулу нуклеиновой кислоты по настоящему изобретению и/или любое другое фармацевтически активное средство, также называемое в настоящем документе лекарственным средством(ами) или активным соединением(ями), можно комбинировать с пероральным нетоксичным фармацевтически приемлемым инертным носителем, таким как этанол, глицерин, вода и т.п. Более того, когда желательно или необходимо, также в смесь можно включать пригодные связующие вещества, смазывающие вещества, дезинтегрирующие вещества и красители. Пригодные связующие вещества включают крахмал, алюмосиликат магния, крахмальную пасту, желатин, метилцеллюлозу, карбоксиметилцеллюлозу натрия и/или поливинилпирролидон, природные сахара, такие как глюкоза или бета-лактоза, кукурузные подсластители, природные и синтетические камеди, такие как гуммиарабик, трагакант или альгинат натрия, полиэтиленгликоль, воски и т.п. Смазывающие вещества, используемые в этих дозированных формах, включают олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия, диоксид кремния, тальк, стеариновую кислоту, ее соль с магнием или кальцием и/или полиэтиленгликоль, и т.п. Дезинтегрирующие вещества включают, но ими не ограничиваются, крахмал, метилцеллюлозу, агар, бентонит, крахмалы ксантановой смолы, агар, альгиновую кислоту или ее соль с натрием, или шипучие смеси и т.п. Разбавители включают, например, лактозу, декстрозу, сахарозу, маннит, сорбит, целлюлозу и/или глицин.
Лекарственное средство по изобретению также можно вводить в таких пероральных дозированных формах в качестве таблеток или капсул с замедленным или непрерывным высвобождением, пилюль, порошков, гранул, эликсиров, настоек, суспензий, сиропов и эмульсий. Суппозитории преимущественно получают из жировых эмульсий или суспензий.
Фармацевтическая композиция или лекарственное средство могут быть стерилизованными и/или могут содержать адъюванты, такие как консерванты, стабилизаторы, смачивающие средства или эмульгаторы, промоторы растворов, соли для регуляции осмотического давления и/или буферы. Кроме того, также они могут содержать другие терапевтически полезные вещества. Композиции получают в соответствии с общепринятыми способами смешивания, гранулирования или нанесения покрытия, и, как правило, они содержат приблизительно от 0,1% до 75%, предпочтительно, приблизительно от 1% до 50%, активного ингредиента.
Жидкие, в частности, инъецируемые композиции можно получать, например, путем растворения, диспергирования и т.д. Активное соединение растворяют в фармацевтически чистом растворителе, например, таком как вода, физиологический раствор, водный раствор декстрозы, глицерин, этанол и т.п., или смешивают с ним, с получением, таким образом, инъецируемого раствора или суспензии. Кроме того, можно изготавливать твердые формы, пригодные для растворения в жидкости перед инъекцией.
Для твердых композиций, эксципиенты включают фармацевтически пригодные маннит, лактозу, крахмал, стеарат магния, сахарин натрия, тальк, целлюлозу, глюкозу, сахарозу, карбонат магния и т.п. Активное соединение, определенное выше, также можно изготавливать в качестве суппозиториев, с использованием в качестве носителя, например, полиалкиленгликолей, например, пропиленгликоля. В некоторых вариантах осуществления, суппозитории преимущественно получают из жировых эмульсий или суспензий.
Лекарственные средства и молекулы нуклеиновых кислот, соответственно, по настоящему изобретению также можно вводить в форме систем для доставки липосом, таких как небольшие однослойные везикулы, крупные однослойные везикулы и многослойные везикулы. Липосомы можно получать из различных фосфолипидов, включающих холестерин, стеариламин или фосфатидилхолины. В некоторых вариантах осуществления, пленка липидных компонентов является гидратированной водным раствором лекарственного средства с образованием липидного слоя, инкапсулирующего лекарственное средство, как хорошо известно специалисту в данной области. Например, молекулы нуклеиновых кислот, описанные в настоящем документе, могут быть предоставлены в качестве комплекса с липофильным соединением или неиммуногенным высокомолекулярным соединением, сконструированного с использованием способов, известных в данной области. Кроме того, липосомы могут нести такие молекулы нуклеиновых кислот на их поверхности для нацеливания и переноса цитотоксических средств внутрь клетки для обеспечения ее уничтожения. Примеры связанных с нуклеиновой кислотой комплексов представлены в патенте США No. 6011020.
Лекарственные средства и молекулы нуклеиновых кислот, соответственно, по настоящему изобретению также можно связывать с растворимыми полимерами в качестве носителей для нацеливаемого лекарственного средства. Такие полимеры могут включать поливинилпирролидон, сополимер пирана, полигидроксипропил-метакриламид-фенол, полигидроксиэтиласпанамидфенол или полиэтиленоксидполилизин, замещенный пальмитоильными остатками. Более того, лекарственные средства и молекулы нуклеиновых кислот, соответственно, по настоящему изобретению можно связывать с классом биологически деградируемых полимеров, пригодным для обеспечения контролируемого высвобождения лекарственного средства, например, полимолочной кислотой, полиэпсилонкапролактоном, полигидроксимасляной кислотой, полиортоэфирами, полиацеталями, полидигидропиранами, полицианоакрилатами и поперечно-сшитыми или амфипатическими блок-сополимерами гидрогелей.
Если желательно, фармацевтическая композиция и лекарственное средство, соответственно, подлежащие введению, также могут содержать небольшие количества нетоксичных вспомогательных веществ, таких как смачивающие вещества или эмульгаторы, pH-буферные средства и другие вещества, например, такие как ацетат натрия и олеат триэтаноламина.
Режим дозирования с использованием молекул нуклеиновых кислот и лекарственных средств, соответственно, по настоящему изобретению выбирают в соответствии с различными факторами, включая тип, вид, возраст, массу тела, пол и медицинское состояние пациента; тяжесть состояния, подлежащего лечению; способ введения; функцию почек и печени пациента; и конкретного используемого аптамера или его соли. Терапевт или ветеринар в данной области может легко определить и назначить эффективное количество лекарственного средства, требуемое для предупреждения, борьбы или остановки прогрессирования состояния.
Эффективные уровни в плазме нуклеиновой кислоты по настоящему изобретению, предпочтительно, находятся в диапазоне от 500 фМ до 500 мкМ при лечении любых из заболеваний, описанных в настоящем документе.
Молекулы нуклеиновых кислот и лекарственные средства, соответственно, по настоящему изобретению, предпочтительно, можно вводить в единичной суточной дозе, раз в двое или трое суток, раз в неделю, раз в две недели, в однократной ежемесячной дозе или раз в три месяца.
Настоящее изобретение относится к тому, что лекарственное средство, как описано в настоящем документе, представляет собой фармацевтическую композицию, описанную в настоящем документе.
В следующем аспекте настоящее изобретение относится к способу лечения индивида, нуждающегося в таком лечении, где способ включает введение фармацевтически активного количества по меньшей мере одной из нуклеиновых кислот по настоящему изобретению. В одном из вариантов осуществления, индивид страдает заболеванием или имеет риск развития такого заболевания, где заболевание представляет собой любое из заболеваний, описанных в настоящем документе, в частности, любое из заболеваний, описанных применительно к применению любой из нуклеиновых кислот по настоящему изобретению для получения лекарственного средства.
Следует понимать, что нуклеиновую кислоту, а также антагонисты по настоящему изобретению, можно использовать не только в качестве лекарственного средства или для получения лекарственного средства, но также для косметических целей, в частности, в связи с вовлечением MCP-1 в воспаленные регионарные очаги повреждения кожи. Таким образом, следующим состоянием или заболеванием для лечения или профилактики которого можно использовать нуклеиновую кислоту, лекарственное средство и/или фармацевтическую композицию по настоящему изобретению, являются воспаленные регионарные очаги повреждения кожи.
Как предпочтительно используют в настоящем документе, диагностическое средство или диагностический способ пригодны для детекции, либо напрямую, либо косвенно, MCP-1, предпочтительно, MCP-1, как описано в настоящем документе, и более предпочтительно MCP-1, как описано в настоящем документе применительно к различным нарушениям и заболеваниям, описанным в настоящем документе. Однако когда молекулы нуклеиновых кислот по настоящему изобретению также связываются с любым, несколькими или всеми из MCP-2, MCP-3, MCP-4 и эотаксина, такие молекулы нуклеиновых кислот также можно использовать для диагностики заболеваний и нарушений, соответственно, патогенный механизм которых либо напрямую, либо косвенно связан или ассоциирован со сверхэкспрессией или избыточной активностью MCP-2, MCP-3, MCP-4 и/или эотаксина. Диагностическое средство пригодно для выявления и/или наблюдения нарушений и заболеваний, соответственно, описанных в настоящем документе. Такое выявление возможно посредством связывания нуклеиновых кислот по настоящему изобретению с MCP-1. Такое связывание можно выявлять либо напрямую, либо косвенно. Соответствующие способы и средства известны специалистам в данной области. Среди прочих, нуклеиновые кислоты по настоящему изобретению могут содержать метку, которая позволяет детекцию нуклеиновых кислот по настоящему изобретению, предпочтительно, нуклеиновой кислоты, связанной с MCP-1. Такую метку предпочтительно выбирают из группы, включающей радиоактивные, ферментные или и флуоресцентные метки. В принципе, любые известные анализы, разработанные для антител, могут быть переняты для нуклеиновых кислот по настоящему изобретению, где связывающее мишень антитело заменяют связывающей мишень нуклеиновой кислотой. В анализах антител с использованием немеченых связывающих мишень антител детекцию предпочтительно проводят с помощью вторичного антитела, модифицированного радиоактивными, ферментными и флуоресцентными метками и связывающегося со связывающим мишень антителом в его Fc-фрагменте. В случае нуклеиновой кислоты, предпочтительно, нуклеиновой кислоты по настоящему изобретению, нуклеиновую кислоту модифицируют такой меткой, где предпочтительно такая метка выбрана из группы, содержащей биотин, Cy-3 и Cy-5, и такую метку подвергают детекции с помощью антитела, направленного против такой метки, например антитела против биотина, антитела против Cy3 или антитела против Cy5, или - в случае, когда меткой является биотин - детекцию метки проводят с помощью стрептавидина или авидина, которые естественным образом связываются с биотином. Такое антитело, стрептавидин или авидин в свою очередь предпочтительно модифицируют соответствующей меткой, например радиоактивной, ферментной или флуоресцентной меткой (подобно вторичному антителу).
В следующем варианте осуществления детекцию или анализ молекул нуклеиновых кислот по изобретению проводят с помощью средств для вторичной детекции, где указанное средство для детекции представляет собой молекулярный маяк. Технология молекулярного маяка известна специалистам в данной области. В кратком изложении, зонды нуклеиновых кислот, которые также называют молекулярными маяками, являются обратными комплементарными последовательностями для образца нуклеиновых кислот, подлежащих детекции, и они вследствие этого гибридизуются с частью образца нуклеиновой кислоты, подлежащей детекции. При связывании с образцом нуклеиновой кислоты флуорофорные группы молекулярного маяка отделяются, что приводит к изменению флуоресцентного сигнала, предпочтительно, изменению интенсивности. Это изменение коррелирует с количеством нуклеиновых кислот, имеющимся в образце.
Будет понятно, что детекция MCP-1 с использованием нуклеиновых кислот по настоящему изобретению может, в частности, позволить детекцию MCP-1, как определено в настоящем документе.
Применительно к детекции MCP-1, предпочтительный способ включает следующие стадии:
(a) предоставление образца, подлежащего тестированию на наличие MCP-1,
(b) предоставление нуклеиновой кислоты по настоящему изобретению,
(с) реакцию образца с нуклеиновой кислотой, предпочтительно, в реакционной емкости
где стадию (a) можно проводить перед стадией (b), или стадию (b) можно проводить перед стадией (a).
В предпочтительном варианте осуществления предусмотрена дополнительная стадия d), которая состоит в детекции реакции образца с нуклеиновой кислотой. Предпочтительно, нуклеиновая кислота стадии b) иммобилизована на поверхности. Поверхность может представлять собой поверхность реакционной емкости, такой как реакционная пробирка, лунка планшета или поверхность приспособления, содержащегося в такой реакционной емкости, например, такого как гранула. Иммобилизацию нуклеиновой кислоты на поверхность можно проводить любым способом, известным специалистам в данной области, включая, но не ограничиваясь ими, нековалентное или ковалентное присоединение. Предпочтительно, связь образуют путем ковалентной химической связи между поверхностью и нуклеиновой кислотой. Однако также настоящее изобретение относится к тому, что нуклеиновая кислота косвенно иммобилизована на поверхности, где такая косвенная иммобилизация вовлекает применение дополнительного компонента или пары партнеров по взаимодействию. Такой дополнительный компонент предпочтительно представляет собой соединение, которое специфично взаимодействует с нуклеиновой кислотой, подлежащей иммобилизации, которую также называют партнером по взаимодействию, и, таким образом, он опосредует связывание нуклеиновой кислоты с поверхностью. Партнера по взаимодействию предпочтительно выбирают из группы, содержащей нуклеиновые кислоты, полипептиды, белки и антитела. Предпочтительно, партнер по взаимодействию представляет собой антитело, более предпочтительно моноклональные антитело. Альтернативно партнером по взаимодействию является нуклеиновая кислота, предпочтительно, функциональная нуклеиновая кислота. Более предпочтительно, такая функциональная нуклеиновая кислота выбрана из группы, содержащей аптамеры, шпигельмеры и нуклеиновые кислоты, которые по меньшей мере частично комплементарны нуклеиновой кислоте. В следующем альтернативном варианте осуществления, связывание нуклеиновой кислоты с поверхностью опосредуется партнером по взаимодействию из нескольких частиц. Таким партнером по взаимодействию из нескольких частиц предпочтительно является пара партнеров по взаимодействию или партнер по взаимодействию, состоящий из первого члена и второго члена, где первый член содержится в нуклеиновой кислоте, или связан с ней, а второй член связан с поверхностью или находится на ней. Партнер по взаимодействию предпочтительно выбирают из группы пар партнеров по взаимодействию, включающей биотин и авидин, биотин и стрептавидин, и биотин и нейтравидин. Предпочтительно, первым членом пары партнеров по взаимодействию является биотин.
Предпочтительным результатом такого способа является образование иммобилизованного комплекса MCP-1 и нуклеиновой кислоты, где более предпочтительно указанный комплекс подвергают детекции. Один вариант осуществления относится к тому, что в комплексе проводят детекцию MCP-1.
Соответствующим средством для детекции, которое согласуется с этим требованием является, например, любое средство для детекции, которое является специфичным для этой/этих части(ей) MCP-1. Особенно предпочтительным средством для детекции является средство для детекции, которое выбрано из группы, содержащей нуклеиновые кислоты, полипептиды, белки и антитела, получение которых известно специалистам в данной области.
Способ детекции MCP-1 также включает удаление образца из реакционной емкости, которую предпочтительно используют для проведения стадии c).
В следующем варианте осуществления способ также включает стадию иммобилизации партнера MCP-1 по взаимодействию на поверхности, предпочтительно, на поверхности, как определено выше, где партнер по взаимодействию является таким, как определено в настоящем документе, и предпочтительно как указано выше применительно к соответствующему способу, и более предпочтительно он включает нуклеиновые кислоты, полипептиды, белки и антитела в их различных вариантах осуществления. В этом варианте осуществления, особенно предпочтительным способом детекции является нуклеиновая кислота по настоящему изобретению, где такая нуклеиновая кислота предпочтительно может быть меченой или немеченой. В случае, когда такая нуклеиновая кислота является меченой, ее детекцию можно проводить напрямую или косвенно. Такая детекция также может вовлекать применение второго средства для детекции, которое, предпочтительно, также выбрано из группы, включающей нуклеиновые кислоты, полипептиды, белки и варианты осуществления в различных вариантах осуществления, описанных в настоящем документе. Такие средства для детекции предпочтительно являются специфичными к нуклеиновой кислоте по настоящему изобретению. В более предпочтительном варианте осуществления, вторым средством для детекции является молекулярный маяк. В предпочтительном варианте осуществления либо нуклеиновой кислота, либо второе средство для детекции, либо оба из них могут содержать метку для детекции. Метку для детекции предпочтительно выбирают из группы, включающей биотин, бром-дезоксиуридиновую метку, дигоксигениновую метку, флуоресцентную метку, УФ-метку, радиоактивную метку и хелатирующую молекулу. Альтернативно второе средство для детекции взаимодействует с поддающейся детекции меткой, которая предпочтительно находится в нуклеиновой кислоте, включена в нуклеиновую кислоту или связана с ней. В частности предпочтительными комбинациями являются следующие:
поддающейся детекции меткой является биотин, а вторым средством для детекции является антитело, направленное против биотина, или где
поддающейся детекции меткой является биотин, а вторым средством для детекции является авидин или несущая авидин молекула, или где
поддающейся детекции меткой является биотин, а вторым средством для детекции является стрептавидин или несущая стрептавидин молекула, или где
поддающейся детекции меткой является биотин, а вторым средством для детекции является нейтравидин или несущая нейтравидин молекула, или
где поддающейся детекции меткой является бром-дезоксиуридин, а вторым средством для детекции является антитело, направленное против бром-дезоксиуридина, или где
поддающейся детекции меткой является дигоксигенин и вторым средством для детекции является антитело, направленное против дигоксигенина, или где
поддающейся детекции меткой является хелатирующий агент, а вторым средством для детекции является радионуклид, где предпочтительно, чтобы метка для детекции была связана с нуклеиновой кислотой. Также понятно, что этот тип комбинации также применим к варианту осуществления, где нуклеиновая кислота связана с поверхностью. В таком варианте осуществления предпочтительно, чтобы поддающаяся детекции метка была связана с партнером по взаимодействию.
Наконец, также настоящее изобретение относится к тому, что детекцию второго средства для детекции проводят с использованием третьего средства для детекции, предпочтительно, третьим средством для детекции является фермент, более предпочтительно, демонстрирующий ферментативную реакцию при детекции второго средства для детекции, или третьим средством для детекции является средство для детекции излучения, более предпочтительно излучения, испускаемого радионуклидом. Предпочтительно, третье средство для детекции специфично детектируется вторым средством для детекции/или взаимодействует с ним.
Также в варианте осуществления с партнером MCP-1 по взаимодействию, иммобилизованным на поверхности, и нуклеиновой кислотой по настоящему изобретению, предпочтительно, добавленной к комплексу, образованному между партнером по взаимодействию и MCP-1, образец можно удалять из реакционной смеси, более предпочтительно из реакционной емкости, где проводят стадию c) и/или d).
В одном из вариантов осуществления нуклеиновая кислота по настоящему изобретению содержит флуоресцентную группу, где флуоресценция флуоресцентной группы отличается при образовании комплекса между нуклеиновой кислотой и MCP-1 и свободным MCP-1.
В следующем варианте осуществления нуклеиновая кислота представляет собой производное нуклеиновой кислоты по настоящему изобретению, где производное нуклеиновой кислоты содержит по меньшей мере одно флуоресцентное производное аденозина, заменяющее аденозин. В предпочтительном варианте осуществления флуоресцентным производным аденозина является этеноаденозин.
В следующем варианте осуществления детекцию комплекса, состоящего из производного нуклеиновой кислоты по настоящему изобретению и MCP-1, проводят с использованием флуоресценции.
В одном из вариантов осуществления способа, сигнал формируется на стадии (c) или стадии (d) и предпочтительно сигнал коррелирует с концентрацией MCP-1 в образце.
В предпочтительном аспекте, анализы можно проводить в 96-луночных планшетах, где компоненты иммобилизованы в реакционных сосудах, как описано выше и в лунах, выступающих в качестве реакционных сосудов.
Специалистам в данной области будет понятно, что все вышесказанное также применимо к MCP-2, MCP-3, MCP-4 и/или эотаксину, по меньшей мере в том, что нуклеиновые кислоты по настоящему изобретению также связываются с MCP-2, MCP-3, MCP-4 и/или эотаксином.
К настоящему изобретению относится то, что способ для детекции MCP-1 в образце, как описано в настоящем документе, также можно использовать в качестве способа диагностики заболевания, такого как хронические заболевания и хронические нарушения, как описано в настоящем документе более подробно.
Кроме того, нуклеиновую кислоту по изобретению можно использовать в качестве исходного материала для конструирования лекарственных средств. В принципе, существует два возможных подхода. Одним из подходов является скрининг библиотек соединений, где такие библиотеки соединений предпочтительно представляют собой библиотеки низкомолекулярных соединений. В одном из вариантов осуществления, скрининг представляет собой высокопроизводительный скрининг. Предпочтительно, высокопроизводительный скрининг представляет собой быструю эффективную и эмпирическую оценку соединений в анализе на основе мишени. В лучшем случае, анализ проводят путем колориметрического определения. Библиотеки, используемые для этого, хорошо известны специалисту в данной области.
Альтернативно нуклеиновую кислоту по настоящему изобретению можно использовать для рационального конструирования лекарственных средств. Предпочтительно, рациональное конструирование лекарственных средств представляет собой конструирование основной структуры фармацевтического средства. Начиная с 3-мерной структуры мишени, которую, как правило, идентифицируют такими способами, как рентгеновская кристаллография или ядерная магнитно-резонансная спектроскопия, используют компьютерные программы для поиска по базам данных, содержащим структуры множества различных химических соединений. Выбор проводят с помощью компьютера, идентифицированные соединения затем можно тестировать в лаборатории.
Рациональное конструирование лекарственных средств можно начинать с любой нуклеиновой кислоты по настоящему изобретению, и оно вовлекает структуру, предпочтительно, трехмерную структуру, которая сходна со структурой нуклеиновых кислот по изобретению или идентична опосредующим связывание частям структуры нуклеиновых кислот по изобретению. В любом случае такая структура, тем не менее, демонстрирует такие же или сходные характеристики связывания, что и нуклеиновые кислоты по изобретению. Предпочтительно либо на следующей стадии, либо в качестве альтернативной стадии рационального конструирования лекарственных средств, трехмерная структура этих частей нуклеиновых кислот, связывающихся с нейротрансмиттером, имитируется химическими группами, которые отличаются от нуклеотидов и нуклеиновых кислот. Путем этой имитации можно конструировать соединение, отличающееся от нуклеиновых кислот. Предпочтительно такое соединение представляет собой низкомолекулярное соединение или пептид.
В случае скрининга библиотек соединений, например, с использованием конкурентного анализа, который известен специалисту в данной области, могут быть найдены соответствующие аналоги MCP-1, агонисты MCP-1 или антагонисты MCP-1. Такие конкурентные анализы могут быть устроены следующим образом. Нуклеиновую кислоту, предпочтительно, шпигельмер, который представляет собой связывающую мишень L-нуклеиновую кислоту, связывают с твердой фазой. Для идентификации аналогов MCP-1 в анализ можно добавлять меченый MCP-1. Потенциальный аналог может конкурировать с молекулами MCP-1, связывающимися со шпигельмером, что сопровождается снижением сигнала, полученного с помощью соответствующей метки. Скрининг агонистов или антагонистов может вовлекать применение анализа клеточной культуры, как известно специалистам в данной области.
Набор по настоящему изобретению может содержать по меньшей мере одну или несколько из нуклеиновых кислот по изобретению. Кроме того, набор может содержать по меньшей мере один или несколько положительных или отрицательных контролей. Положительный контроль может представлять собой, например, MCP-1, в частности, против которого отбирают нуклеиновую кислоту по изобретению или с которым она связывается, предпочтительно, в жидкой форме. Отрицательный контроль может представлять собой, например, пептид, который определен с точки зрения биофизических свойств, сходных с MCP-1, но который не распознается нуклеиновыми кислотами по изобретению. Более того, указанный набор может содержать один или несколько буферов. Различные ингредиенты могут содержаться в наборе в сухой или лиофилизированной форме или они могут быть растворены в жидкости. Набор может включать один или несколько контейнеров, которые, в свою очередь, могут содержать один или несколько ингредиентов набора. В следующем варианте осуществления, набор содержит инструкцию или листок с инструкцией, на котором предоставлена информация для пользователя о применении набора и его различных ингредиентов. Понятно, что этот тип набора также особенно пригоден для диагностики и детекции хронического заболевания и хронического нарушения, как описано в настоящем документе.
Фармацевтическое и биоаналитическое определение нуклеиновой кислоты по настоящему изобретению для оценки ее фармакокинетического и биодинамического профиля в различных жидкостях, тканях и органах организма человека и не человека, является элементарным. Для такой цели можно использовать любой из способов детекции, описанных в настоящем документе или известных специалисту в данной области. В следующем аспекте настоящего изобретения для детекции нуклеиновой кислоты по настоящему изобретению предусмотрен сэндвич-анализ гибридизации. В анализе для детекции используют улавливающий зонд и зонд для детекции. Улавливающий зонд комплементарен первой части, а зонд для детекции комплементарен второй части нуклеиновой кислоты по настоящему изобретению. Как улавливающий зонд, так и зонд для детекции, могут быть образованы нуклеотидами ДНК, модифицированными нуклеотидами ДНК, модифицированными нуклеотидами РНК, нуклеотидами РНК, нуклеотидами LNA и/или нуклеотидами PNA.
Таким образом, улавливающий зонд содержит участок последовательности, комплементарный 5'-концу нуклеиновой кислоты по настоящему изобретению, а зонд для детекции содержит участок последовательности, комплементарный 3'-концу нуклеиновой кислоты по настоящему изобретению. В этом случае улавливающий зонд иммобилизуют на поверхности или матрице через его 5'-конец, где улавливающий зонд может быть иммобилизован непосредственного через его 5'-конец или через линкер между его 5'-концом и поверхностью или матрицей. Однако в принципе линкер может быть связан с каждым нуклеотидом улавливающего зонда. Линкер может быть образован гидрофильными линкерами, известными в данной области, или нуклеотидами D-ДНК, модифицированными нуклеотидами D-ДНК, нуклеотидами D-РНК, модифицированными нуклеотидами D-РНК, нуклеотидами D-LNA, нуклеотидами PNA, нуклеотидами L-РНК, нуклеотидами L-ДНК, модифицированными нуклеотидами L-РНК, модифицированными нуклеотидами L-ДНК и/или нуклеотидами L-LNA.
Альтернативно, улавливающий зонд содержит участок последовательности, комплементарный 3'-концу нуклеиновой кислоты по настоящему изобретению, а зонд для детекции содержит участок последовательности, комплементарный 5'-концу нуклеиновой кислоты по настоящему изобретению. В этом случае улавливающий зонд иммобилизуют на поверхности или матрице через его 3'-конец, где улавливающий зонд может быть иммобилизован непосредственного через его 3'-конец или через линкер между его 3'-концом и поверхностью или матрицей. Однако в принципе линкер может быть связан с каждым нуклеотидом участка последовательности, который комплементарен нуклеиновой кислоте по настоящему изобретению. Линкер может быть образован гидрофильными линкерами, известными в данной области, или нуклеотидами D-ДНК, модифицированными нуклеотидами D-ДНК, нуклеотидами D-РНК, модифицированными нуклеотидами D-РНК, нуклеотидами D-LNA, нуклеотидами PNA, нуклеотидами L-РНК, нуклеотидами L-ДНК, модифицированными нуклеотидами L-РНК, модифицированными нуклеотидами L-ДНК и/или нуклеотидами L-LNA.
Число нуклеотидов в улавливающем зонде и зонде для детекции, которые могут гибридизоваться с нуклеиновой кислотой по настоящему изобретению, варьирует и может зависеть от числа нуклеотидов в улавливающем зонде и/или зонде для детекции и/или самой нуклеиновой кислоте по настоящему изобретению. Общее число нуклеотидов в улавливающем зонде и зонде для детекции, которые могут гибридизоваться с нуклеиновой кислотой по настоящему изобретению, должно представлять собой максимальное количество нуклеотидов, которые содержатся в нуклеиновой кислоте по настоящему изобретению. Минимальное число нуклеотидов (от 2 до 10 нуклеотидов) в зонде для детекции и улавливающем зонде должно обеспечить гибридизацию с 5'-концом или 3'-концом, соответственно, нуклеиновой кислоты по настоящему изобретению. Для достижения высокой специфичности и селективности для нуклеиновой кислоты по настоящему изобретению относительно других нуклеиновых кислот, встречающих в анализируемых образцах, общее количество нуклеотидов в улавливающем зонде и зонде для детекции должно представлять собой максимальное число нуклеотидов, которые содержатся в нуклеиновой кислоте по настоящему изобретению.
Более того зонд для детекции предпочтительно содержит маркерную молекулу или метку, которую можно подвергать детекции, как описано в настоящем документе выше. Метка или маркерная молекула в принципе может быть связана с каждым нуклеотидом зонда для детекции. Предпочтительно, метка или маркер расположены на 5'-конце или 3'-конце зонда для детекции, где между нуклеотидами в зонде для детекции, которые комплементарны нуклеиновой кислоте по настоящему изобретению, и меткой может быть встроен линкер. Линкер может быть образован гидрофильными линкерами, известными в данной области, или нуклеотидами D-ДНК, модифицированными нуклеотидами D-ДНК, нуклеотидами D-РНК, модифицированными нуклеотидами D-РНК, нуклеотидами D-LNA, нуклеотидами PNA, нуклеотидами L-РНК, нуклеотидами L-ДНК, модифицированными нуклеотидами L-РНК, модифицированными нуклеотидами L-ДНК и/или нуклеотидами L-LNA.
Детекцию нуклеиновой кислоты по настоящему изобретению можно проводить следующим образом:
Нуклеиновая кислота по настоящему изобретению гибридизуется одним из ее концов с улавливающим зондом, а другим концом с зондом для детекции. После этого не связавшийся зонд для детекции удаляют, например, посредством одной или нескольких стадий промывания. Затем можно измерять количество связавшегося зонда для детекции, который предпочтительно содержит метку или маркерную молекулу, например, как более подробно описано в WO/2008/052774, которая включена в настоящий документ в качестве ссылки.
Как предпочтительно используют в настоящем документе, термин "лечение" в предпочтительном варианте осуществления включает, дополнительно или альтернативно, профилактику и/или наблюдение.
Как предпочтительно используют в настоящем документе, термины "заболевание и нарушение" используют взаимозаменяемо, если нет иных указаний.
Как используют в настоящем документе, термин "содержит" предпочтительно не предназначен для ограничения объекта, следующего после такого термина или описываемого им. Однако в альтернативном варианте осуществления термин "содержит" следует понимать в значении "состоит из" и, таким образом, как ограничивающий объект, следующий после такого термина или описываемый им.
Различные SEQ ID NO:, химическая природа молекул нуклеиновых кислот по настоящему изобретению и молекулы MCP-1-мишени как используют в настоящем документе, их фактическая последовательность и внутренний номер для ссылки обобщены в следующей таблице.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Настоящее изобретение дополнительно иллюстрируется фигурами, примерами и списком последовательностей, из которых могут быть установлены дополнительные признаки, варианты осуществления и преимущества, где
На фиг. 1 представлено выравнивание последовательностей родственных РНК-лигандов, связывающихся с MCP-1 человека, где указан мотив последовательности ("тип 1A"), который в предпочтительном варианте осуществления полностью необходим для связывания с MCP-1 человека;
На фиг. 2 представлено выравнивание последовательностей родственных РНК-лигандов, связывающихся с MCP-1 человека, где указан мотив последовательности ("тип 1B"), который в предпочтительном варианте осуществления полностью необходим для связывания с MCP-1 человека, и производных РНК-лигандов 180-D1-002;
На фиг. 3 представлено выравнивание последовательностей родственных РНК-лигандов, связывающихся с MCP-1 человека, где указан мотив последовательности ("тип 2"), который в предпочтительном варианте осуществления полностью необходим для связывания с MCP-1 человека;
На фиг. 4 представлено выравнивание последовательностей родственных РНК-лигандов, связывающихся с MCP-1 человека, где указан мотив последовательности ("тип 3"), который в предпочтительном варианте осуществления полностью необходим для связывания с MCP-1 человека;
На фиг. 5 представлены производные РНК-лигандов 178-D5 и 181-A2 (РНК-лиганды мотива последовательности "тип 3" для MCP-1 человека);
На фиг. 6 представлено выравнивание последовательностей родственных РНК-лигандов, связывающихся с MCP-1 человека, где указан мотив последовательности ("тип 4"), который в предпочтительном варианте осуществления полностью необходим для связывания с MCP-1 человека (другие последовательности);
На фиг. 7 представлена таблица последовательностей нескольких различных РНК-лигандов, связывающихся с MCP-1 человека, которые могут быть родственными связывающим MCP-1 мотивам последовательностей "тип 1A", "тип 1B"; "тип 2", "тип 3" или "тип 4";
На фиг. 8 представлено выравнивание производных РНК-лигандов 188-A3-001 и 189-G7-001, которые связываются с MCP-1 мыши;
На фиг. 9 представлен результат анализа связывания аптамера D-NOX-E36 с биотинилированным D-MCP-1 человека при комнатной температуре и 37°C, представленный в качестве связывания аптамера относительно концентрации биотинилированного D-MCP-1 человека;
На фиг. 10 представлен результат анализа связывания аптамера D-mNOX-E36 с биотинилированным D-MCP-1 мыши при 37°C, представленный в качестве связывания аптамера относительно концентрации биотинилированного D-MCP-1 мыши;
На фиг. 11 представлено индуцируемое MCP-1 высвобождение Ca++ в клетках THP-1, где была получена кривая "доза-эффект" для MCP-1 человека, показывающая половинную эффективную концентрацию (EC50), составляющую приблизительно 3 нМ, представленная как разница флуоресценции относительно пустого образца в зависимости от концентрации MCP-1 человека;
На фиг. 12 показана эффективность шпигельмера NOX-E36 в анализе высвобождения кальция; клетки стимулировали 3 нМ MCP-1 человека, предварительно инкубированным при 37°C с различными количествами шпигельмера NOX-E36, представленная в качестве процента от контроля относительно концентрации NOX-E36;
На фиг. 13 показана эффективность шпигельмера mNOX-E36 в анализе высвобождения кальция; клетки стимулировали 5 нМ MCP-1 мыши, предварительно инкубированным при 37°C с различными количествами шпигельмера mNOX-E36, представленная в качестве процента от контроля относительно концентрации mNOX-E36;
На фиг. 14 представлен индуцируемый MCP-1 человека хемотаксис клеток THP-1, где через 3 часа после миграции клеток THP-1 в направлении различных концентраций MCP-1 получали кривую "доза-эффект" для MCP-1, представленный в качестве повышения в X раз по сравнению с контролем относительно концентрации MCP-1 человека;
На фиг. 15 представлена эффективность шпигельмера NOX-E36 в анализе хемотаксиса; клеткам позволяли мигрировать в направлении 0,5 нМ MCP-1 человека, предварительно инкубированного при 37°C с различными количествами шпигельмера NOX-E36, представленная в качестве процента от контроля относительно концентрации шпигельмера NOX-E36;
На фиг. 16 представлена эффективность шпигельмера mNOX-E36 в анализе хемотаксиса; клеткам позволяли мигрировать в направлении 0,5 нМ MCP-1 мыши, предварительно инкубированного при 37°C с различными количествами шпигельмера NOX-E36, представленная в качестве процента от контроля относительно концентрации шпигельмера mNOX-E36;
На фиг. 17 представлена сенсограмма Biacore 2000, показывающая значение KD шпигельмера NOX-E-36, связывающегося с MCP-1 человека, который был иммобилизован на сенсорном чипе PioneerF1 способом присоединения аминов, представленная в качестве ответа (RU) с течением времени;
На фиг. 18 представлена сенсограмма Biacore 2000, показывающая связывание шпигельмера NOX-E36 с белками семейства MCP человека (huMCP-1, huMCP-2, huMCP-3) и эотаксином человека, которые были иммобилизованы с помощью способа присоединения аминов на сенсорном чипе PioneerF1 и CM4, соответственно, представленная в качестве ответа (RU) с течением времени;
На фиг. 19 представлена сенсограмма Biacore 2000, показывающая связывание шпигельмера NOX-E36 с MCP-1 различных видов (MCP-1 собаки, MCP-1 обезьяны, MCP-1 человека, MCP-1 свиньи, MCP-1 кролика, MCP-1 мыши, MCP-1 крысы), где различные формы MCP-1 были иммобилизованы с помощью способа присоединения аминов на сенсорных чипах PioneerF1 и CM4, соответственно, представленная в качестве ответа (RU) с течением времени;
На фиг. 20 представлена сенсограмма Biacore 2000, показывающая значение KD связывания шпигельмера 181-A2-018 с MCP-1 человека, который был иммобилизован на сенсорном чипе CM4 способом присоединения аминов, представленная в качестве ответа (RU) с течением времени;
На фиг. 21 представлена сенсограмма Biacore 2000, показывающая связывание шпигельмера 181-A2-018 с белками семейства MCP человека (huMCP-1, huMCP-2, huMCP-3) и эотаксином человека, которые были иммобилизованы с помощью способа присоединения аминов на сенсорном чипе PioneerF1 и CM4, соответственно, представленная в качестве ответа (RU) с течением времени;
На фиг. 22 представлена сенсограмма Biacore 2000, показывающая связывание шпигельмера 181-A2-018 с MCP-1 из различных видов (MCP-1 собаки, MCP-1 обезьяны, MCP-1 человека, MCP-1 свиньи, MCP-1 кролика, MCP-1 мыши, MCP-1 крысы), где различные формы MCP-1 были иммобилизованы с помощью способа присоединения аминов на сенсорных чипах PioneerF1 и CM4, соответственно, представленная в качестве ответа (RU) с течением времени;
На фиг. 23 представлено выравнивание с помощью Clustal W MCP-1 различных видов млекопитающих, а также MCP-2, MCP-3 и эотаксина человека (только положения 1-76);
На фиг. 24A представлена таблица, в которой обобщена специфичность связывания NOX-E36 и 181-A2-018 в отношении MCP-1 из различных видов млекопитающих, а также MCP-2, MCP-3 и эотаксина человека;
На фиг. 24B представлена таблица, в которой обобщена селективность NOX-E36 при определении анализом Biacore, где биотинилированный NOX-E36 был иммобилизован на поверхности сенсорного чипа, и проанализировано связывание панели различных CC- и CXC-хемокинов с NOX-E36;
На фиг. 24C представлен анализ кинетики взаимодействия NOX-E36 с хемокинами при определении анализом Biacore, где хемокины были ковалентно иммобилизованы на поверхности сенсорного чипа CM5 и были инъецированы различные концентрации NOX-E36, и проанализировано связывающее поведение NOX-E36 с использованием программного обеспечения BiaEvaluation;
На фиг. 24D представлена кривая "доза-эффект" для хемотаксиса при стимуляции клеток THP-1 посредством MIP-1α с половинной эффективной концентрацией, составляющей приблизительно 0,2 нМ;
На фиг. 24E представлено ингибирование индуцируемого MIP-1α хемотаксиса посредством NOX-E36. NOX-E36 не оказывал влияния на индуцируемый MIP1a хемотаксис клеток THP-1;
На фиг. 25 представлена эффективность шпигельмера NOX-E36-3'-PEG в анализе высвобождения кальция; клетки стимулировали 3 нМ MCP-1 человека, предварительно инкубированным при 37°C с различными количествами шпигельмера NOX-E36-3'-PEG, представленная в качестве процента от контроля относительно концентрации шпигельмера NOX-E36-3'-PEG;
На фиг. 26 представлена эффективность шпигельмера NOX-E36-3'-PEG в анализе хемотаксиса; клеткам позволяли мигрировать в направлении 0,5 нМ MCP-1 человека, предварительно инкубированного при 37°C с различными количествами шпигельмера NOX-E36-3'-PEG, представленная в качестве процента от контроля относительно концентрации NOX-E36-3'-PEG;
На фиг. 27A представлена эффективность шпигельмера NOX-E36-5'-PEG в анализе высвобождения кальция; клетки стимулировали 3 нМ MCP-1 человека, предварительно инкубированным при 37°C с различными количествами шпигельмера NOX-E36-5'-PEG, представленная в качестве процента от контроля относительно концентрации шпигельмера NOX-E36-5'-PEG;
На фиг. 27B представлена эффективность шпигельмера NOX-E36-5'-PEG в анализе хемотаксиса; клеткам позволяли мигрировать в направлении 0,5 нМ MCP-1 человека, предварительно инкубированного при 37°C с различными количествами шпигельмера NOX-E36-5'-PEG, представленная в качестве процента контроля относительно концентрации шпигельмера NOX-E36-5'-PEG;
На фиг. 28 представлено индуцируемое MCP-1 высвобождение Ca++ в клетках THP-1, где получали кривую доза-эффект для MCP-1 мыши, указывающую половинную эффективную концентрацию (EC50), составляющую приблизительно 5 нМ, представленное в качестве разницы флуоресценции относительно пустого образца в зависимости от концентрации MCP-1 мыши;
На фиг. 29 представлена эффективность шпигельмера mNOX-E36-3'-PEG против MCP-1 человека в анализе высвобождения кальция; клетки стимулировали 3 нМ MCP-1 мыши, предварительно инкубированным при 37°C с различными количествами шпигельмера mNOX-E36-3'-PEG, представленная в качестве процента от контроля относительно концентрации шпигельмера mNOX-E36-3'-PEG;
На фиг. 30 представлен индуцируемый MCP-1 мыши хемотаксис клеток THP-1, где после миграции клеток THP-1 в течение 3 часов в направлении различных концентраций mMCP-1 получали кривую "доза-эффект" для mMCP-1, представленный в качестве повышения в X раз по сравнению с контролем относительно концентрации MCP-1 мыши;
На фиг. 31 представлена эффективность шпигельмера mNOX-E36-3'-PEG против MCP-1 мыши в анализе хемотаксиса; клеткам позволяли мигрировать в направлении 0,5 нМ MCP-1 мыши, предварительно инкубированного при 37°C с различными количествами шпигельмера mNOX-E36-3'-PEG, представленная в качестве процента от контроля относительно концентрации противомышиного шпигельмера mNOX-E36-3'-PEG;
На фиг. 32 представлена сенсограмма Biacore 2000, показывающая значение KD связывания аптамера D-mNOX-E36 с D-MCP-1 мыши, который был иммобилизован на сенсорном чипе PioneerF1 способом присоединения аминов, представленная в качестве ответа (RU) с течением времени;
На фиг. 33 представлена сенсограмма Biacore 2000, показывающая связывание аптамера D-mNOX-E36 с D-MCP-1 человека и D-MCP-1 мыши, где две различных формы D-MCP-1 были иммобилизованы способом присоединения аминов на сенсорных чипах PioneerF1 и CM4, соответственно, представленная в качестве ответа (RU) с течением времени;
На фиг. 34 представлены срезы почек мышей MRLlpr/lpr в возрасте 24 недель, окрашенные перйодной кислотой по Шифу (PAS), антителами к Mac-2 (макрофаги) и CD3 (T-клетки) как указано; изображения соответствуют 7-12 мышам в каждой группе (исходное увеличение PAS: ×100, вставки PAS: ×400, Mac2: ×400, CD3: ×100;
На фиг. 35 представлена таблица, иллюстрирующая параметры функции почек и гистологические данные в различных группах мышей MRLlpr/lpr в возрасте 24 недели;
На фиг. 36 представлено количественное определение гистологических изменений посредством морфометрии, проведенное на окрашенных серебром срезах мышей из всех групп; A, индекс объема интерстиция; B, индекс расширения канальцев, и C, индекс повреждения клеток канальцев, вычисляли в качестве процента поля под большим увеличением и они выражены в качестве средних значений ± SEM;
На фиг. 37 представлено выживание мышей MRLlpr/lpr из различных групп введения при вычислении с помощью анализа Каплана-Мейера;
На фиг. 38 представлена экспрессия в почках мРНК для CC-хемокинов CCL2 и CCL5 при определении с помощью ОТ-ПЦР с использованием тотальной РНК почки, объединенной от 5 мышей из каждой группы, где уровни РНК для каждой группы мышей выражены посредством соответствующей экспрессии 18S рРНК;
На фиг. 39 представлено уменьшение патологии легких посредством введения mNOX-E36-3'PEG; ткань легких получали от всех четырех групп в возрасте 24 недели и подвергали полуколичественной оценке; введение mNOX-E36 и mNOX-E36-3'PEG снижало перибронхиолярное воспаление у мышей MRLlpr/lpr; изображения соответствуют 7-11 мышам в каждой группе; исходное увеличение ×100;
На фиг. 40 представлены кожные проявления волчанки у мышей MRLlpr/lpr в возрасте 24 недель, встречающиеся, как правило, на области морды или шеи (левая мышь), которые были менее распространены у мышей, которым вводили шпигельмер против mCCL2 (правая мышь);
На фиг. 41 представлены сывороточные и гистологические данные у мышей MRLlpr/lpr в возрасте 24 недель;
На фиг. 42 представлена фармакокинетика ПЭГ-илированных и неПЭГ-илированных шпигельмеров против mCCL2 в плазме в ходе исследования, представленная в качестве концентрации в плазме шпигельмера mNOX-E36 в зависимости от времени;
На фиг. 43 представлена проточная цитометрия для CCR2 на костном мозге и периферической крови у мышей MRLlpr/lpr, которым вводили носитель или mNOX-E36-3'PEG в возрасте 24 недель; данные представлены в качестве среднего процента положительных по CCR2 клеток ± SEM как в костном мозге, так и в периферической крови у 5 мышей в каждой группе;
На фиг. 44 представлены уровни CCL2 в сыворотки у мышей 1K db/db, которым вводили PoC-PEG- (белые столбцы) и mNOX-E36-3'PEG (mNOX-E36-P) (черные столбцы) при определении посредством ELISA в различные моменты времени, как указано; данные представляют собой средние значения ± SEM; *, p<0,05 mNOX-E36-3'PEG (mNOX-E36-P) против PoC-PEG;
На фиг. 45 представлено число инфильтрированных положительных по Mac-2 и Ki-67 клеток в гломерулах и интерстиции мышей db/db, которым не проводили введение или вводили POC-PEG или mNOX-E36-3'PEG;
На фиг. 46 представлен диабетический гломерулосклероз у мышей db/db в возрасте 6 месяцев; срезы почек мышей из различных групп окрашивали перйодной кислотой по Шифу и 15 гломерул из каждого среза почки оценивали в отношении степени гломерулосклероза; изображения показывают репрезентативные гломерулы, оцененные по соответствующим показателям, как указано, исходное увеличение 400 x; на графике представлен средний процент каждого показателя ± SEM для всех мышей в каждой группе (n=7-10); *, p<0,05 для мышей 1K db/db, которым вводили mNOX-E36-3'PEG (mNOX-E36-P) против PoC-PEG (PoC-P);
На фиг. 47 представлена скорость гломерулярной фильтрации (GFR) у мышей 1K db/db в возрасте 6 месяцев, которым вводили mNOX-E36-3'PEG (mNOX-E36-P) и PoC-PEG(PoC-P); GFR определяли по кинетике выведения FITC-инулина в группах мышей 1K db/db, которым вводили PoC-PEG и mNOX-E36-3'PEG, в конце исследования;
На фиг. 48 представлена атрофия канальцев и интерстициальный объем у мышей db/db в возрасте 6 месяцев; изображения окрашенных серебром срезов почки иллюстрируют соответствующие почки из соответствующих групп (исходное увеличение 100x); значения представляют собой средние значения ± SEM для соответствующего индекса морфометрического анализа для 7-10 мышей в каждой группе; *, p<0,05 2K db/db против мышей BKS дикого типа; #, p<0,05 мышей 1K против 2K db/db; †, p<0,05 у мышей 1K db/db, которым вводили mNOX-E36-3'PEG (mNOX-E36-PEG) против PoC-PEG;
На фиг. 49 представлена экспрессия мРНК CCL2 у мышей db/db при определении посредством ОТ-ПЦР в реальном времени с использованием общей РНК почки, объединенной от 6-10 мышей каждой группы; уровни мРНК для каждой группы мышей выражены посредством соответствующей экспрессии 18S рРНК;
На фиг. 50 представлена пространственная экспрессия CCL2 в почках мышей db/db при определении посредством иммунного окрашивания; изображения иллюстрируют соответствующие срезы почек от мышей в возрасте 6 месяцев в соответствующих группах, как указано (исходное увеличение, 200×);
На фиг. 51A-E представлены маркеры волчаночного нефрита у мышей MRLlpr/lpr после введения мышам MRLlpr/lpr носителя, revmNOX-E36-3'-PEG, mNOX-E36-3'-PEG, CYC на низком уровне, CYC на высоком уровне, CYC на низком уровне + mNOX-E36-3'-PEG или MMF, где индекс активности (фиг. 51A) и индекс хронического состояния (фиг. 51B) для DPLN определяли на окрашенных PAS срезах почек, как описано Austin et al (Austin et al. 1984); и где определяли среднее число гломерулярных макрофагов (фиг. 51C) в срезах почек мышей MRLlpr/lpr в возрасте 24 недель (клетки Mac2+ в 15 гломерулах на срез), количества интерстициальных макрофагов (фиг. 51D) или количества T-клеток (фиг. 51E) в срезах почек мышей MRLlpr/lpr в возрасте 24 недель, соответственно (клетки Mac2+ или CD3+ в 15 полях под большим увеличением на срез);
На фиг. 52 представлена полуколичественная оценка повреждения легкого из срезов легких, окрашенных перйодной кислотой по Шифу, мышей MRLlpr/lpr в возрасте 24 недель;
На фиг. 53A представлено общее число клеток в жидкости BAL через 24 ч после нагрузки LPS (×106/животное; среднее значение ± SEM; * p<0,05, ** p<0,01 против группы положительного контроля), где животным вводили носитель (положительный контроль), дексаметазон, рофлумиласт или связывающий MCP-1 шпигельмер mNOX-E36-3'-PEG перед нагрузкой LPS или носитель перед нагрузкой чистым воздухом (отрицательный контроль);
На фиг. 53B представлено абсолютное число нейтрофилов в жидкости BAL через 24 ч после нагрузки LPS (среднее значение ± SEM; ** p<0,01 против группы положительного контроля), где животным вводили носитель (положительный контроль), дексаметазон, рофлумиласт или связывающий MCP-1 шпигельмер mNOX-E36-3'-PEG перед нагрузкой LPS или носитель перед нагрузкой чистым воздухом (отрицательный контроль);
На фиг. 54A представлена гипертрофия правого отдела сердца у здоровых животных или животных после введения MCT/носителя или связывающего MCT/MCP-1 шпигельмера mNOX-E36-3'-PEG; где показателем было отношение массы правого желудочка к массе левого желудочка вместе с массой перегородки RV/(LV+S);
На фиг. 54B представлено систолическое давление правого желудочка (RSVP [мм. рт. ст.]) у здоровых животных или животных после введения MCT/носителя или MCT/связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG.
Пример 1: Нуклеиновые кислоты, которые связывают MCP-1 человека
С использованием биотинилированного D-MCP-1 человека в качестве мишени, можно получить несколько нуклеиновых кислот, которые связываются с MCP-1 человека, нуклеотидные последовательности которых представлены на фиг.1-7. Нуклеиновые кислоты охарактеризовывали на уровне аптамера, т.е. D-нуклеиновой кислоты, с использованием конкурентных анализов или прямых "pull-down"-анализов с биотинилированным D-MCP-1 человека (пример 4), или на уровне шпигельмера, т.е. L-нуклеиновой кислоты, с природной конфигурацией MCP-1 (L-MCP) измерением с помощью поверхностного плазмонного резонанса с использованием инструмента Biacore 2000 (пример 7), в анализе высвобождения Ca++ в культуре клеток in vitro (пример 5), или в анализе хемотаксиса in vitro (пример 6).
Молекулы нуклеиновых кислот, полученные таким образом, обладают различными мотивами последовательностей, и их четыре основных типа определены на фиг. 1 и 2 (тип 1A/1B), фиг. 3 (тип 2), фиг. 4 и 5 (тип 3) и фиг. 6 (тип 4). Дополнительные связывающие MCP-1 нуклеиновые кислоты, которые не могут быть родственными друг другу и другим мотивам последовательностей, описанным в настоящем документе, приведены на фиг. 7. Для определения мотивов нуклеотидной последовательности, используют сокращения IUPAC для неоднозначных нуклеотидов:
|
Если нет иных указаний, любая последовательность нуклеиновой кислоты или последовательность участков и боксов, соответственно, указаны в направлении 5' → 3'.
Связывающие MCP-1 нуклеиновые кислоты типа 1A (фиг. 1)
Как представлено на фиг. 1, все последовательности связывающих MCP-1 нуклеиновых кислот типа 1A содержат несколько последовательностей, участков или боксов, где боксы B1A и B1B представляют собой 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом. Однако такая гибридизация не обязательно существует в молекуле, в действительности находящейся в физиологических условиях. Боксы B2, B3, B4, B5 и бокс B6 фланкируются боксом B1A и боксом B1B.
Нуклеиновые кислоты охарактеризовывали на уровне аптамеров с использованием прямых и конкурентных "pull-down"-анализов с биотинилированным D-MCP-1 человека для ранжирования их в отношении их связывающего поведения (пример 4). Выбранные последовательности синтезировали в качестве шпигельмеров (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе высвобождения Ca++ в клеточной культуре in vitro (пример 5).
Последовательности в определенных боксах могут отличаться между связывающими MCP-1 нуклеиновыми кислотами типа 1A, что влияет на аффинность связывания с MCP-1. Исходя из анализа связывания различных связывающих MCP-1 нуклеиновых кислот, обобщенно представленных как связывающие MCP-1 нуклеиновые кислоты типа 1A, боксы B1A, B2, B3, B4, B5, B6 и B1B и их нуклеотидные последовательности, как описано ниже, индивидуально и более предпочтительно полностью необходимы для связывания с MCP-1:
- боксы B1A и B1B представляют собой 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом; где B1A представляет собой AGCRUG, предпочтительно, AGCGUG; и где B1B представляет собой CRYGCU, предпочтительно, CACGCU;
- бокс B2, который представляет собой CCCGGW, предпочтительно, CCCGGU;
- бокс B3, который представляет собой GUR, предпочтительно, GUG;
- бокс B4, который представляет собой RYA, предпочтительно, GUA;
- бокс B5, который представляет собой GGGGGRCGCGAYC, предпочтительно, GGGGGGCGCGACC;
- бокс B6, который представляет собой UGCAAUAAUG или URYAWUUG, предпочтительно, UACAUUUG
Как представлено на фиг. 1, молекула нуклеиновой кислоты, обозначаемая как 176-E10trc, имеет наилучшую аффинность связывания с MCP-1 (в качестве аптамера в "pull-down"-анализе с KD 5 нМ, а также в качестве шпигельмера с IC50 4-5 нМ в анализе высвобождения Ca++ в клеточной культуре in vitro) и, таким образом, она может представлять собой оптимальную последовательность и оптимальную комбинацию элементов последовательности B1A, B2, B3, B4, B5, B6 и B1B.
Связывающие MCP-1 нуклеиновые кислоты типа 1B (фиг. 2)
Как представлено на фиг. 2, все последовательности типа 1B содержат несколько последовательностей, участков или боксов, где боксы B1A и B1B представляют собой 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом, и боксы B2, B3, B4, B5 и бокс B6 фланкируются боксом B1A и боксом B1B. Однако такая гибридизация не обязательно существует в молекуле, в действительности находящейся в физиологических условиях.
Нуклеиновые кислоты охарактеризовывали на уровне аптамеров с использованием прямых и конкурентных "pull-down"-анализов с биотинилированным D-MCP-1 человека для ранжирования их в отношении их связывающего поведения (пример 4). Выбранные последовательности синтезировали в качестве шпигельмеров (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе высвобождения Ca++ в клеточной культуре in vitro (пример 5).
Последовательности в определенных боксах могут отличаться между связывающими MCP-1 нуклеиновыми кислотами типа 1B, что влияет на аффинность связывания с MCP-1. Исходя из анализа связывания различных связывающих MCP-1 нуклеиновых кислот, обобщенно представленных как связывающие MCP-1 нуклеиновые кислоты типа 1B, боксы B1A, B2, B3, B4, B5, B6 и B1B и их нуклеотидные последовательности, как описано ниже, индивидуально и более предпочтительно полностью необходимы для связывания с MCP-1:
- боксы B1A и B1B могут гибридизоваться друг с другом; где B1A представляет собой AGYRUG, предпочтительно, AGCGUG; и где B1B представляет собой CAYRCU, предпочтительно, CACGCU;
- бокс B2, который представляет собой CCAGCU или CCAGY, предпочтительно, CCAGU;
- бокс B3, который представляет собой GUG;
- бокс B4, который представляет собой AUG;
- бокс B5, который представляет собой GGGGGGCGCGACC;
- бокс B6, который представляет собой CAUUUUA или CAUUUA, предпочтительно, CAUUUUA.
Как представлено на фиг. 2, молекула нуклеиновой кислоты, обозначаемая как 176-C9trc, имеет наилучшую аффинность связывания с MCP-1 (в качестве аптамера в "pull-down"-анализе с KD 5 нМ, а также в качестве шпигельмера с IC50 4-5 нМ в анализе высвобождения Ca++ в клеточной культуре in vitro) и, таким образом, она может представлять собой оптимальную последовательность и оптимальную комбинацию элементов последовательности B1A, B2, B3, B4, B5, B6 и B1B
Связывающие MCP-1 нуклеиновые кислоты типа 2 (фиг. 3)
Как представлено на фиг. 3, все последовательности типа 2 содержат несколько последовательностей, участков или боксов, где боксы B1A и B1B представляют собой 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом, и бокс B2 является центральным элементом последовательности. Однако такая гибридизация не обязательно существует в молекуле, в действительности находящейся в физиологических условиях.
Нуклеиновые кислоты охарактеризовывали на уровне аптамеров с использованием прямых и конкурентных "pull-down"-анализов с биотинилированным D-MCP-1 человека для ранжирования их в отношении их связывающего поведения (пример 4). Выбранные последовательности синтезировали в качестве шпигельмеров (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе высвобождения Ca++ в клеточной культуре in vitro (пример 5) или в анализе хемотаксиса in vitro (пример 6).
Последовательности в определенных боксах могут отличаться между связывающими MCP-1 нуклеиновыми кислотами типа 3, что влияет на аффинность связывания с MCP-1. Исходя из анализа связывания различных связывающих MCP-1 нуклеиновых кислот, обобщенно представленных как связывающие MCP-1 нуклеиновые кислоты типа 2, боксы B1A, B2 и B1B и их нуклеотидные последовательности, как описано ниже, индивидуально и более предпочтительно полностью необходимы для связывания с MCP-1:
- боксы B1A и B1B, 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом; где B1A представляет собой ACGCA и B1B представляет собой UGCGU, или B1A представляет собой CGCA и B1B представляет собой UGCG, или B1A представляет собой GCA и B1B представляет собой UGCG или UGC; предпочтительно B1A представляет собой GCA и B1B представляет собой UGCG;
- бокс B2, CSUCCCUCACCGGUGCAAGUGAAGCCGYGGCUC, предпочтительно, CGUCCCUCACCGGUGCAAGUGAAGCCGUGGCUC
Как представлено на фиг. 3, нуклеиновая кислота, обозначаемая как 180-D1-002, а также производные 180-D1-002, такие как 180-D1-011, 180-D1-012, 180-D1-035, и 180-D1-036 (= NOX-E36) имеют наилучшую аффинность связывания с MCP-1 в качестве аптамера в "pull-down"-анализе или в конкурентном "pull-down"-анализе с KD < 1 нМ, и, таким образом, она может представлять собой оптимальную последовательность и оптимальную комбинацию элементов последовательности B1A, B2 и B1B
Для молекулы нуклеиновой кислоты D-NOX-E36 (D-180-D1-036; SEQ ID NO:159), была определена константа диссоциации (KD) 890±65 пМ при комнатной температуре (RT) и 146 ± 13 пМ при 37°C (пример 4; фиг. 9). Соответствующий шпигельмер NOX-E36 (180-D1-036; SEQ ID NO:37) обладал ингибиторной концентрацией (IC50) 3-4 нМ в анализе высвобождения Ca++ in vitro (пример 5; фиг. 12) и приблизительно 0,5 нМ в анализе хемотаксиса in vitro (пример 6; фиг. 15). Для ПЭГ-илированных производных NOX-E36, NOX-E36-3'PEG и NOX-E36-5'PEG, были определены IC50 приблизительно 3 нМ в анализе высвобождения Ca++ (пример 5, фиг. 25 и фиг.27A) и < 1 нМ в анализе хемотаксиса (пример 6; фиг. 26 и фиг. 27B).
Связывающие MCP-1 нуклеиновые кислоты типа 3 (фиг. 4+5)
Как представлено на фиг. 4 и 5, все последовательности типа 3 содержат несколько последовательностей, участков или боксов, где три пары боксов являются характерными для связывающих MCP-1 нуклеиновых кислот типа 3. Оба бокса B1A и B1B, а также боксы B2A и B2B и боксы B5A и B5B обладают способность гибридизоваться друг с другом. Однако такая гибридизация не обязательно существует в молекуле, в действительности находящейся в физиологических условиях. Между этими потенциально гибридизующимся элементами последовательности, расположены не гибридизующиеся нуклеотиды, определенные как бокс B3, бокс B4 и бокс B6.
Нуклеиновые кислоты охарактеризовывали на уровне аптамеров с использованием прямых и конкурентных "pull-down"-анализов с биотинилированным D-MCP-1 человека для ранжирования их в отношении их связывающего поведения (пример 4). Выбранные последовательности синтезировали в качестве шпигельмеров (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе высвобождения Ca++ в клеточной культуре in vitro (пример 5), в анализе хемотаксиса in vitro (пример 6), или посредством измерения с помощью Biacore (пример 7).
Последовательности в определенных боксах могут отличаться между связывающими MCP-1 нуклеиновыми кислотами типа 3, что влияет на аффинность связывания с MCP-1. Исходя из анализа связывания различных связывающих MCP-1 нуклеиновых кислот, обобщенно представленных как связывающие MCP-1 нуклеиновые кислоты типа 3, боксы B1A, B2A, B3, B2B, B4, B5A, B6, B5B, B1B и их нуклеотидные последовательности, как описано ниже, индивидуально и более предпочтительно полностью необходимы для связывания с MCP-1:
- боксы B1A и B1B, 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом; где B1A представляет собой GURCUGC и B1B представляет собой GCAGCAC; предпочтительно B1A представляет собой GUGCUGC и B1B представляет собой GCAGCAC;
или B1A представляет собой GKSYGC и B1B представляет собой GCRSMC; предпочтительно B1A представляет собой GUGCGC и B1B представляет собой GCGCAC;
или B1A представляет собой KBBSC и B1B представляет собой GSVVM; предпочтительно B1A представляет собой KKSSC и B1B представляет собой GSSMM;
или B1A представляет собой BNGC и B1B представляет собой GCNV; предпочтительно B1A представляет собой SNGC и B1B представляет собой GCNS; наиболее предпочтительно B1A представляет собой GGGC и B1B представляет собой GCCC;
- боксы B2A и B2B , участки, которые могут гибридизоваться друг с другом; где B2A представляет собой GKMGU и B2B представляет собой ACKMC; предпочтительно B2A представляет собой GUAGU и B2B представляет собой ACUAC;
- бокс B3, который представляет собой KRRAR, предпочтительно, UAAAA или GAGAA;
- бокс B4, который представляет собой CURYGA или CUWAUGA или CWRMGACW или UGCCAGUG, предпочтительно, CAGCGACU или CAACGACU;
- B5A и B5B, участки, которые могут гибридизоваться друг с другом; где B5A представляет собой GGY и B5B представляет собой GCYR, а GCY может гибридизоваться с нуклеотидами B5A; или B5A представляет собой CWGC и B5B представляет собой GCWG; предпочтительно B5A представляет собой GGC и B5B представляет собой GCCG;
- бокс B6, который представляет собой: YAGA или CKAAU или CCUUUAU, предпочтительно, UAGA.
Как представлено на фиг. 4 и 5, нуклеиновая кислота, обозначаемая как 178-D5, и ее производное 178-D5-030, а также 181-A2 с ее производными 181-A2-002, 181-A2-004, 181-A2-005, 181-A2-006, 181-A2-007, 181-A2-017, 181-A2-018, 181-A2-019, 181-A2-020, 181-A2-021 и 181-A2-023, имеют наилучшую аффинность связывания с MCP-1. 178-D5 и 178-D5-030 оценивали в качестве аптамеров в прямых и конкурентных "pull-down"-анализах (пример 4) с KD приблизительно 500 пМ. В той же экспериментальной схеме была определена KD для 181-A2 приблизительно 100 пМ. Посредством анализа Biacore (пример 7), было определено, что KD 181-A2 и ее производных в отношении MCP-1 составляет 200-300 пМ. В анализах высвобождения Ca++ и хемотаксиса с культивируемыми клетками (пример 5 и 6, соответственно), была измерена IC50 как для 178-D5, так и для 181-A2, составляющая приблизительно 500 пМ. Таким образом, 178-D5, а также 181-A2 и их производные могут представлять собой оптимальную последовательность и оптимальную комбинацию элементов последовательности B1A, B2A, B3, B2B, B4, B5A, B6, B5B и B1B.
Связывающие MCP-1 нуклеиновые кислоты типа 4 (фиг. 6)
Как представлено на фиг. 6, все последовательности типа 4 содержат несколько последовательностей, участков или боксов, где боксы B1A и B1B представляют собой 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом, и бокс B2 является центральным элементом последовательности.
Нуклеиновые кислоты охарактеризовывали на уровне аптамеров с использованием прямых и конкурентных "pull-down"-анализов с биотинилированным D-MCP-1 человека для ранжирования их в отношении их связывающего поведения (пример 4). Выбранные последовательности синтезировали в качестве шпигельмеров (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе высвобождения Ca++ в клеточной культуре in vitro (пример 5) или в анализе хемотаксиса in vitro (пример 6).
Последовательности в определенных боксах могут отличаться между связывающими MCP-1 нуклеиновыми кислотами типа 4, что влияет на аффинность связывания с MCP-1. Исходя из анализа связывания различных связывающих MCP-1 нуклеиновых кислот, обобщенно представленных как связывающие MCP-1 нуклеиновые кислоты типа 4, боксы B1A, B2 и B1B и их нуклеотидные последовательности, как описано ниже, индивидуально и более предпочтительно полностью необходимы для связывания с MCP-1:
- боксы B1A и B1B, 5'- и 3'-концевые участки, которые могут гибридизоваться друг с другом; где B1A представляет собой AGCGUGDU и B1B представляет собой GNCASGCU; или B1A представляет собой GCGCGAG и B1B представляет собой CUCGCGUC; или B1A представляет собой CSKSUU и B1B представляет собой GRSMSG; или B1A представляет собой GUGUU и B1B представляет собой GRCAC; или B1A представляет собой UGUU и B1B представляет собой GGCA; предпочтительно B1A представляет собой CSKSUU и B1B представляет собой GRSMSG; наиболее предпочтительно B1A представляет собой CCGCUU и B1B представляет собой GGGCGG; и
- бокс B2, который представляет собой AGNDRDGBKGGURGYARGUAAAG или AGGUGGGUGGUAGUAAGUAAAG или CAGGUGGGUGGUAGAAUGUAAAGA, предпочтительно, AGGUGGGUGGUAGUAAGUAAAG
Как представлено на фиг. 6, нуклеиновая кислота, обозначаемая как 174-D4-004 и 166-A4-002 имеет наилучшую аффинность связывания с MCP-1 (в качестве шпигельмера с IC50 2-5 нМ в анализе высвобождения Ca++ в клеточной культуре in vitro) и, таким образом, она может представлять собой оптимальную последовательность и оптимальную комбинацию элементов последовательности B1A, B2 и B1B.
Кроме того, было идентифицировано 29 других связывающих MCP-1 нуклеиновых кислот, которые нельзя описать как комбинацию элементов нуклеотидной последовательности, как показано для типов 1-4 связывающих MCP-1 нуклеиновых кислот. Эти последовательности приведены на фиг. 7.
Понятно, что любые из последовательностей, представленных на фиг.1-7, представляют собой нуклеиновые кислоты по настоящему изобретению, включая их укороченные формы, а также включая их удлиненные формы, однако при условии что укороченные или удлиненные таким образом, соответственно, молекулы нуклеиновых кислот все еще способны связываться с мишенью.
Пример 2: Нуклеиновые кислоты, которые связывают MCP-1 мыши
С использованием биотинилированного D-MCP-1 мыши в качестве мишени, можно было получить несколько молекул нуклеиновых кислот, связывающихся с ним. Результат анализа последовательности этих молекул нуклеиновых кислот можно видеть на фиг. 8.
Нуклеиновые кислоты охарактеризовывали на уровне аптамеров с использованием прямых и конкурентных "pull-down"-анализов с биотинилированным D-MCP-1 человека для ранжирования их в отношении их связывающего поведения (пример 4). Выбранные последовательности синтезировали в качестве шпигельмеров (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе высвобождения Ca++ в клеточной культуре in vitro (пример 5) или в анализе хемотаксиса in vitro (пример 6).
Как представлено на фиг. 8, D-188-A3-001 и D-189-G7-001 и их производные связывают D-MCP-1 в "pull-down"-анализе с субнаномолярной KD (фиг. 8).
Для D-mNOX-E36 (= D-188-A3-007; SEQ ID NO:244), была определена константа диссоциации (KD) 0,1-0,2 нМ при 37°C (пример 4; фиг. 10). Соответствующий шпигельмер mNOX-E36 (188-A3-007; SEQ ID NO:122) имел ингибиторную концентрацию (IC50) в анализе высвобождения Ca++ in vitro приблизительно 12 нМ (пример 5; фиг. 13) и приблизительно 7 нМ в анализе хемотаксиса in vitro (пример 6; фиг. 16). Для ПЭГ-илированного производного mNOX-E36, mNOX-E36-3'PEG (SEQ ID NO:254), была определена IC50 приблизительно 8 нМ в анализе высвобождения Ca++ (пример 5, фиг. 29) и приблизительно 3 нМ в анализе хемотаксиса (пример 6; фиг. 31).
Следует понимать, что любые из последовательностей, представленных на фиг.1-7, представляют собой нуклеиновые кислоты по настоящему изобретению, включая их укороченные формы, а также включая их удлиненные формы, однако при условии что укороченные или удлиненные таким образом, соответственно, молекулы нуклеиновых кислот все еще способны связываться с мишенью.
Пример 3: Синтез и преобразование аптамеров и шпигельмеров
Мелкомасштабный синтез
Аптамеры и шпигельмеры получали твердофазным синтезом с помощью устройства для синтеза ABI 394 (Applied Biosystems, Foster City, CA, USA) с использованием 2'TBDMS РНК-фосфорамидитного химического соединения (M.J. Damha, K.K. Ogilvie, Methods in Molecular Biology, Vol. 20 Protocols for oligonucleotides and analogous, ed. S. Agrawal, p. 81-114, Humana Press Inc. 1993). rA(N-Bz)-, rC(Ac)-, rG(N-ibu)- и rU-фосфорамидиты в D- и L-конфигурации приобретали от ChemGenes, Wilmington, MA. Аптамеры и шпигельмеры очищали гель-электрофорезом.
Крупномасштабный синтез с модификацией
Шпигельмер NOX-E36 получали твердофазным синтезом с помощью устройства для синтеза ÄktaPilot100 (Amersham Biosciences; General Electric Healthcare, Freiburg) с использованием 2'TBDMS РНК-фосфорамидитного химического соединения (M.J. Damha, K.K. Ogilvie, Methods in Molecular Biology, Vol. 20 Protocols for oligonucleotides and analogous, ed. S. Agrawal, p. 81-114, Humana Press Inc. 1993). L-rA(N-Bz)-, L-rC(Ac)-, L-rG(N-ibu)- и L-rU-фосфорамидиты приобретали от ChemGenes, Wilmington, MA. 5'-амино-модификатор приобретали от American International Chemicals Inc. (Framingham, MA, USA). Синтез немодифицированного шпигельмера начинали на L-riboG-модифицированном CPG с размером пор 1000 Å (Link Technology, Glasgow, UK); для 3'-NH2-модифицированного шпигельмера использовали 3'-аминомодификатор-CPG, 1000 Å (ChemGenes, Wilmington, MA). Для присоединения (15 мин на цикл), использовали 0,3 M бензилтиотетразол (CMS-Chemicals, Abingdon, UK) в ацетонитриле, и 3,5 эквивалентов соответствующего 0,1 M раствора фосфорамидита в ацетонитриле. Использовали цикл окисления-кэпирования. Дополнительные стандартные растворители и реагенты для олигонуклеотидного синтеза приобретали от Biosolve (Valkenswaard, NL). Синтезировали шпигельмер DMT-ON; после удаления защитной группы, его очищали посредством препаративной ОФ-ВЭЖХ (Wincott F. et al. (1995) Nucleic Acids Res 23:2677) с использованием среды Source15RPC (Amersham). 5'DMT-группу удаляли 80% уксусной кислотой (30 мин при комнатной температуре). Затем добавляли водный 2 M раствор NaOAc, и шпигельмер обессоливали тангенциальной проточной фильтрацией с использованием регенерированной целлюлозной мембраны 5 K (Millipore, Bedford, MA).
ПЭГ-илирование NOX-E36
Для продления времени нахождения шпигельмера в плазме in vivo, шпигельмер NOX-E36 ковалентно присоединяли к группе полиэтиленгликоля (PEG) размером 40 кДа на 3'-конце или 5'-конце.
3'-ПЭГ-илирование NOX-E36
Для ПЭГ-илирования (для технических деталей способа ПЭГ-илирования см. патентную заявку Европы EP 1306382), очищенный 3'-аминомодифицированный шпигельмер растворяли в смеси H2O (2,5 мл), DMF (5 мл) и буфера A (5 мл; полученной смешиванием лимонной кислоты • H2O [7 г], борной кислоты [3,54 г], фосфорной кислоты [2,26 мл] и 1 M NaOH [343 мл] и добавлением H2O до конечного объема 1 л; pH = 8,4, доводили с помощью 1 M HCl).
pH раствора шпигельмера доводили до 8,4 с помощью 1 M NaOH. Затем каждые 30 мин добавляли сложный эфир PEG-NHS массой 40 кДа (Nektar Therapeutics, Huntsville, AL) в четырех порциях по 0,6 эквивалентов при 37°C до достижения максимального выхода 75-85%. pH реакционной смеси в процессе добавления сложного эфира PEG-NHS держали при 8-8,5 с помощью 1 M NaOH.
Реакционную смесь смешивали с 4 мл раствора мочевины (8 M), 4 мл буфера A и 4 мл буфера B (0,1 M ацетат триэтиламмония в H2O) и нагревали до 95°C в течение 15 мин. Затем ПЭГ-илированный шпигельмер очищали посредством ОФ-ВЭЖХ со средой Source 15RPC (Amersham), с использованием градиента ацетонитрила (буфер B; буфер C: 0,1 M ацетат триэтиламмония в ацетонитриле). Избыток PEG элюировали при 5% буфере C, ПЭГ-илированный шпигельмер - при 10-15% буфере C. Фракции продукта с чистотой >95% (как оценивают по ВЭЖХ) комбинировали и смешивали с 40 мл 3 M NaOAC. ПЭГ-илированный шпигельмер обессоливали тангенциальной проточной фильтрацией (регенерированная целлюлозная мембрана 5 K, Millipore, Bedford MA).
5'-ПЭГ-илирование NOX-E36
Для ПЭГ-илирования (для технических деталей способа ПЭГ-илирования см патентную заявку Европы EP 1306382), очищенный 5'-аминомодифицированный шпигельмер растворяли в смеси H2O (2,5 мл), DMF (5 мл) и буфера A (5 мл; полученной смешиванием лимонной кислоты ·H2O [7 г], борной кислоты [3,54 г], фосфорной кислоты [2,26 мл] и 1 M NaOH [343 мл] и добавлением H2O до конечного объема 1 л; pH = 8,4, доводили с помощью 1 M HCl).
pH раствора шпигельмера доводили до 8,4 с помощью 1 M NaOH. Затем каждые 30 мин добавляли сложный эфир PEG-NHS массой 40 кДа (Nektar Therapeutics, Huntsville, AL) в шести порциях по 0,25 эквивалентов при 37°C до достижения максимального выхода 75-85%. pH реакционной смеси в процессе добавления сложного эфира PEG-NHS держали при 8-8,5 с помощью 1 M NaOH.
Реакционную смесь смешивали с 4 мл раствора мочевины (8 M), и 4 мл буфера B (0,1 M ацетат триэтиламмония в H2O) и нагревали до 95°C в течение 15 мин. Затем ПЭГ-илированный шпигельмер очищали посредством ОФ-ВЭЖХ со средой Source 15RPC (Amersham), с использованием градиента ацетонитрила (буфер B; буфер C: 0,1 M ацетат триэтиламмония в ацетонитриле). Избыток PEG элюировали при 5% буфере C, ПЭГ-илированный шпигельмер - при 10-15% буфере C. Фракции продукта с чистотой >95% (как оценивают по ВЭЖХ) комбинировали и смешивали с 40 мл 3 M NaOAC. ПЭГ-илированный шпигельмер обессоливали тангенциальной проточной фильтрацией (регенерированная целлюлозная мембрана 5 K, Millipore, Bedford MA).
Пример 4: Определение констант связывания ("pull-down"-анализ)
Прямой "pull-down"-анализ
Аффинность аптамеров к D-MCP-1 измеряли в "pull-down"-анализе при 20 или 37°C, соответственно. Аптамеры метили по 5'-фосфату T4-полинуклеотидкиназой (Invitrogen, Karlsruhe, Germany) с использованием [γ-32P]-меченного ATP (Hartmann Analytic, Braunschweig, Germany). Удельная радиоактивность меченных аптамеров составляла 200000-800000 cpm/пмоль. Аптамеры инкубировали после денатурации и ренатурации при концентрации 20 пМ при 37°в буфере для селекции (20 мМ Tris-HCl pH 7,4; 137 мМ NaCl; 5 мМ KCl; 1 мМ MgCl2; 1 мМ CaCl2; 0,1% [масс./об.] Tween-20) вместе с различными количествами биотинилированного D-MCP-1 в течение 4-12 часов для достижения равновесия при низких концентрациях. Буфер для селекции дополняли 10 мкг/мл сывороточного альбумина человека (Sigma-Aldrich, Steinheim, Germany), и 10 мкг/мл РНК дрожжей (Ambion, Austin, USA) для предотвращения адсорбции партнеров по связыванию на поверхностях используемого пластмассового изделия или матрицы для иммобилизации. Диапазон концентрации биотинилированного D-MCP-1 составлял от 8 пМ до 100 нМ; общий объем реакции составил 1 мл. Пептид и комплексы пептид-аптамер иммобилизовывали на 1,5-мкл частицах Streptavidin Ultralink Plus (Pierce Biotechnology, Rockford, USA), которые были предварительно уравновешены буфером для селекции и ресуспендированы в общем объеме 6 мкл. Частицы держали в суспензии в течение 30 мин при соответствующей температуре в термомиксере. Иммобилизованную радиоактивность количественно определяли в сцинтилляционном счетчике после удаления супернатанта и соответствующего промывания. Процент связывания наносили на график против концентрации биотинилированного D-MCP-1, и константы диссоциации получали с использованием алгоритмов программного обеспечения (GRAFIT; Erithacus Software; Surrey U.K.), исходя из стехиометрии 1:1.
Конкурентный "pull-down"-анализ
Для сравнения различных связывающих D-MCP-1 аптамеров проводили конкурентный анализ с ранжированием. Для этой цели наиболее аффинный доступный аптамер метили радиоактивной меткой (см. выше) и он служил в качестве эталона. После денатурации и ренатурации его инкубировали при 37°C с биотинилированным D-MCP-1 в 1 мл буфера для селекции в условиях, которые приводили приблизительно к 5-10% связыванию пептида после иммобилизации и промывания на агарозе NeutrAvidin или Streptavidin Ultralink Plus (оба от Pierce) без конкуренции. Добавляли избыток денатурированных и ренатурированных немеченых вариантов D-РНК-аптамеров с различными концентрациями (например 2, 10 и 50 нМ) меченного эталонного аптамера для параллельных реакций связывания. Аптамеры, подлежащие тестированию, конкурировали с эталонным аптамером за связывание мишени, таким образом, снижая сигнал связывания в зависимости от их связывающих характеристик. Аптамер, который был выявлен в качестве наиболее активного в этом анализе, затем мог служить в качестве нового эталона для сравнительных анализов других вариантов аптамеров.
Пример 5: Определение ингибиторной концентрации в анализе высвобождения Ca ++
Клетки THP-1 (DSMZ, Braunschweig) культивировали в течение ночи при плотности клеток 0,3×106/мл при 37°C и 5% CO2 в среде RPMI 1640 с GlutaMAX (Invitrogen), которая дополнительно содержала 10% эмбриональную телячью сыворотку, 50 единиц/мл пенициллина, 50 мкг/мл стрептомицина и 50 мкМ β-меркаптоэтанол.
Шпигельмеры инкубировали вместе с рекомбинантным MCP-1 человека (Bachem) в сбалансированном солевом растворе Хэнкса (HBSS), содержащем 1 мг/мл бычьего сывороточного альбумина, 5 мМ пробенецида и 20 мМ HEPES (HBSS+) в течение 15-60 мин при 37°C в 0,2-мл низкопрофильном 96-луночном планшете ("раствор для стимуляции").
В целях нагрузки индикаторным красителем для кальция, клетки центрифугировали при 300×g в течение 5 мин, ресуспендировали в 4 мл раствора индикаторного красителя (10 мкМ fluo-4 [Molecular Probes], 0,08% pluronic 127 [Molecular Probes] в HBSS+) и инкубировали в течение 60 мин при 37°C. Затем добавляли 11 мл HBSS+ и клетки центрифугировали, как описано выше, промывали один раз 15 мл HBSS+, а затем ресуспендировали в HBSS+ с получением плотности клеток 1,1×106/мл. В каждую лунку черного 96-луночно планшета добавляли по 90 мкл этой суспензии клеток.
Измерение флуоресцентных сигналов проводили при длине волны возбуждения 485 нм и длине волны испускания 520 нм в устройстве для считывания планшетов со множественной детекцией Fluostar Optima (BMG). Для параллельного измерения в нескольких образцах, лунки одного (перпендикулярного) ряда 96-луночного планшета регистрировали совместно. Для определения основной линии проводили первые три считывания данных с временем запаздывания 4 с. Затем регистрацию прерывали и планшет извлекали из устройства. С использованием многоканальной пипетки, в лунки добавляли 10 мкл раствора для стимуляции, затем планшет снова помещали в устройство и измерение продолжали. Всего проводили 20 регистраций с временными интервалами 4 секунды.
Для каждой лунки различие между максимальной флуоресценцией и величиной основной линии определяли и наносили на график против концентрации MCP1 или, в экспериментах по ингибированию высвобождения кальция шпигельмерами, против концентрации шпигельмера.
Определение половинной максимальной эффективной концентрации (EC 50 ) для MCP-1 человека
После стимуляции клеток THP-1 различными концентрациями hMCP-1 и нанесения на график различий между максимальным и исходным сигналами, получали кривую "доза-эффект" для MCP-1 человека, показывающую половинную эффективную концентрацию (EC50), составляющую приблизительно 2-4 нМ (фиг. 11). Эту концентрацию использовали для дальнейших экспериментов по ингибированию высвобождения Ca++ шпигельмерами.
Определение половинной максимальной эффективной концентрации (EC 50 ) для MCP-1 мыши
После стимуляции клеток THP-1 различными концентрациями mMCP-1 и нанесения на график различий между максимальным и исходным сигналами, получали кривую "доза-эффект" для MCP-1 мыши, показывающую половинную эффективную концентрацию (EC50), составляющую приблизительно 5 нМ (фиг. 28). Эту концентрацию использовали для дальнейших экспериментов по ингибированию высвобождения Ca++ шпигельмерами.
Пример 6: Определение ингибиторной концентрации в анализе хемотаксиса
Клетки THP-1, выращенные, как описано выше, центрифугировали, промывали один раз в HBH (HBSS, содержащая 1 мг/мл бычьего сывороточного альбумина и 20 мМ HEPES) и ресуспендировали в количестве 3×106 клеток/мл. 100 мкл этой суспензии добавляли к вставкам Transwell с порами 5 мкм (Corning, #3421). В более низких отделениях перед добавлением клеток MCP-1 предварительно инкубировали со шпигельмерами в различных концентрациях в 600 мкл HBH при 37°C в течение от 20 до 30 мин. Клеткам позволяли мигрировать при 37°C в течение 3 часов. Затем вставки удаляли и в нижние отделения добавляли 60 мкл 440 мкМ резазурина (Sigma) в фосфатно-солевом буфере. После инкубации при 37°C в течение 2,5 часов, измеряли флуоресценцию при длине волны возбуждения 544 нм и длине волны испускания 590 нм в устройстве для считывания с множественной детекцией Fluostar Optima (BMG).
Определение половинной максимальной эффективной концентрации (EC 50 ) для MCP-1 человека
После миграции в течение 3 часов клеток THP-1 в направлении различных концентраций MCP-1 человека, получили кривую "доза-эффект" для MCP-1 человека, показывающую максимальную эффективную концентрацию приблизительно 1 нМ и сниженную активацию при более высоких концентрациях (фиг. 14). Для дальнейших экспериментов по ингибированию хемотаксиса шпигельмерами использовали концентрацию MCP-1 0,5 нМ.
Определение половинной максимальной эффективной концентрации (EC 50 ) для MCP-1 мыши
После миграции в течение 3 часов клеток THP-1 в направлении различных концентраций MCP-1 человека, получили кривую "доза-эффект" для MCP-1 мыши, показывающую максимальную эффективную концентрацию приблизительно 1 нМ и сниженную активацию при более высоких концентрациях (фиг. 14). Для дальнейших экспериментов по ингибированию хемотаксиса шпигельмерами использовали концентрацию MCP-1 мыши 0,5 нМ.
Пример 7: Анализ связывания посредством измерения с помощью поверхностного плазмонного резонанса
7.1 Оценка специфичности NOX-E36, 181-A2-018 и mNOX-E36
Для анализа связывания нуклеиновых кислот с MCP-1 человека и родственными белками использовали устройство Biacore 2000 (Biacore AB, Uppsala, Sweden). Когда связывание намеревались обеспечить через аминогруппы, белки подвергали диализу против воды в течение 1-2 ч (смешанные сложные эфиры целлюлозы Millipore VSWP; размер пор 0,025 мкМ) для удаления мешающих аминов. Перед присоединением белка сенсорные чипы PioneerF1 или CM4 (Biacore AB) активировали путем инъекции 35 мкл разведения 1:1 0,4 M NHS и 0,1 M EDC при скорости потока 5 мкл/мин. Затем инъецировали хемокин в концентрациях 0,1-1,5 мкг/мл при скорости потока 2 мкл/мин до тех пор, пока ответ устройства не попадал в диапазон 1000-2000 RU (относительных единиц). Не вступившие в реакцию сложные эфиры NHS подвергали дезактивации инъекцией 35 мкл раствора гидрохлорида этаноламина (pH 8,5) со скоростью потока 5 мкл/мин. Сенсорный чип заливали два раза буфером для связывания и уравновешивали при 10 мкл/мин в течение 1-2 часов до тех пор, пока исходный уровень не становился стабильным. Для всех белков параметры кинетики и константы диссоциации оценивали с помощью серии инъекций шпигельмера в концентрациях 1000, 500, 250, 125, 62,5, 31,25 и 0 нМ в буфере для селекции (Tris-HCl, 20 мМ; NaCl, 137 мМ; KCl, 5 мМ; CaCl2, 1 мМ; MgCl2, 1 мМ; Tween20, 0,1% [об./масс.]; pH 7,4). Во всех экспериментах анализ проводили при 37°C с использованием команды Kinject, определяющей время ассоциации 180 секунд и время диссоциации 360 секунд при скорости потока 10 мкл/мин. Анализ данных и вычисление констант диссоциации (KD) проводили с помощью программного обеспечения BIAevaluation 3.0 (BIACORE AB, Uppsala, Sweden) с использованием алгоритма 1:1 стехиометрического приведения в соответствии Ленгмюра.
7.1.1 NOX-E36 и 181-A2-018 (специфичные к MCP-1 человека нуклеиновые кислоты)
Только для MCP-1 человека представлены все сенсограммы (фиг. 17 и 20, соответственно); в случае других белков, для ясности представлена только сенсограмма, полученная при концентрации шпигельмера 125 нМ (фиг. 18/19 и 21/22).
Анализ взаимодействия NOX-E36●hMCP-1: рекомбинантный MCP-1 человека иммобилизовывали на сенсорном чипе PioneerF1 в соответствии с рекомендациями изготовителя (способ присоединения аминов) до тех пор, пока не устанавливался ответ устройства 1381 RU (относительных единиц). Определенная константа диссоциации (KD) для NOX-E36, связывающихся с MCP-1 человека, составила приблизительно 890 пМ (фиг. 17).
Анализ взаимодействия 181-А2-018●hMCP-1: рекомбинантный MCP-1 человека иммобилизовывали на сенсорном чипе СМ4 в соответствии с рекомендациями изготовителя (способ присоединения аминов) до тех пор, пока не устанавливался ответ устройства 3111 RU (относительных единиц). Определенная константа диссоциации (KD) для 181-А2-018, связывающихся с MCP-1 человека, составила приблизительно 370 пМ (фиг. 20).
Для определения специфичности NOX-E36 и 181-A2-018, различные белки семейства MCP-1 человека, а также эотаксин человека иммобилизовывали на сенсорном чипе PioneerF1 и CM4 (hMCP-1, 1754 RU; hMCP-2, 1558 RU; hMCP-3, 1290 RU; эотаксин, 1523 RU). Анализ кинетики показал, что NOX-E36 связывается с эотаксином и hMCP-2 с константами диссоциации (KD) 5-10 нМ; hMCP-3 не распознавался (фиг. 18 и 24A). Напротив, 181-A2-018 связывает эотаксин, hMCP-2 и hMCP-3, но с немного меньшей аффинностью (10-20 нМ; фиг. 21 и 24A).
Межвидовую перекрестную реактивность NOX-E36 и 181-A2-018 оценивали с использованием иммобилизованного присоединения по аминогруппе MCP-1 из человека (1460 RU), обезьяны (1218 RU), свиньи (1428 RU), собаки (1224 RU), кролика (1244 RU), крысы (1267 RU) и мыши (1361 RU) на сенсорном чипе PioneerF1 и CM4. Анализ кинетики показал, что NOX-E36 связывается с MCP-1 человека, обезьяны, свиньи и собаки со сравнимыми константами диссоциации (KD) 0,89-1,2 нМ, в то время как MCP-1 из мыши, крысы и кролика не распознавались (фиг. 19 и 24A). 181-A2-018 связывается с MCP-1 человека и обезьяны со сравнимыми константами диссоциации (KD) 0,5-0,6 нМ, в то время как MCP-1 свиньи, кролика и собаки связываются со значительно меньшей аффинностью. MCP-1 крысы и мыши не распознавались NOX-A2-018 (фиг. 22 и 24A).
Последовательности, а также степень гомологии в аминокислотах с процентной идентичностью между белком MCP-1 из различных видов и близко родственными белками представлены на фиг. 23; вычисленные значения KD для NOX-E36 и 181-A2-018 представлены в виде таблицы на фиг. 24A.
7.1.2 mNOX-E36 (специфичная к MCP-1 мыши нуклеиновая кислота)
Для анализа связывающего поведения mNOX-E36, 3759 RU синтетического биотинилированного D-MCP-1 мыши (проточная ячейка 3) и 3326 RU биотинилированного D-MCP-1 человека (проточная ячейка 4) иммобилизовывали на сенсорном чипе с конъюгированным стрептавидином (Biacore AB, Freiburg, Germany), соответственно. 500, 250, 125, 62,5, 31,25 и 0 нМ растворы аптамера mNOX-E36 (D-РНК) инъецировали с использованием команды Kinject, определяющей время ассоциации 180 с и время диссоциации 360 с. Проточную ячейку 1 использовали в качестве буфера и контроля в виде декстрановой матрицы (поверхность Biacore SA-Chip), а на проточной ячейке 2 иммобилизовывали неспецифический D-пептид для определения неспецифического связывания аптамера. На фиг. 32 представлена сенсограмма кинетики D-NOX-E36 для связывания с D-MCP-1 мыши с вычисленной константой диссоциации (KD) 200-300 пМ. mNOX-E36 не связывает D-MCP-1 человека (фиг. 33); для ясности представлена только сенсограмма, полученная с 125 нМ шпигельмером.
7.2 Оценка селективности NOX-E36
Селективность NOX-E36 оценивали анализом посредством поверхностного плазмонного резонанса путем иммобилизации 5'-биотинилированного NOX-E36 на стрептавидине (SA-чип). 352 RU NOX-E36 иммобилизовывали на проточной ячейке (FC) 1, и равное количество 5'-концевого биотинилированного нефункционального контрольного шпигельмера (POC) иммобилизовывали на FC 2 посредством связывания стрептавидин/биотин. FC3 использовали в качестве контрольной поверхности для определения неспецифического связывания с сенсорной поверхностью декстран-SA.
100 нМ панели хемокинов человека из всех четырех подгрупп (CC, CXC, CX3C и XC) инъецировали в течение 360 с, и комплексам позволяли диссоциировать в течение 360 с при скорости потока 10 мкл/мин и 37°C. Единицы ответа после ассоциации (Resp.1; степень взаимодействия) и после диссоциации (Resp.2, аффинность взаимодействия) наносили на график. После каждой инъекции поверхность чипа регенерировали в течение 240 с 1 M хлоридом натрия с 0,1% Tween; затем иммобилизованным шпигельмерам позволяли повторно сворачиваться в течение 2 минут при физиологических условиях (буфер для анализа). Инъекцию каждого хемокина повторяли 3 раза. CXCL1, CXCL2, CXCL6 и CXCL9 показали неспецифическое связывание с рибонуклеиновыми кислотами и декстрановой поверхностью чипа. Специфическое высокоаффинное связывание с иммобилизованным NOX-E36 можно было выявить только для CCL2/MCP-1, CCL8/MCP-2, CCL11/эотаксина, CCL3/MIP1α и CXCL7/NAP-2 (фиг. 24B). Открытие, что MCP-2 и эотаксин связываются с NOX-E36 не удивительно, вследствие относительно высокой гомологии между этими хемокинами и MCP-1, составляющей 62 и 70%, для неожиданных положительных результатов CCL3/MIP-1α и CXCL7/NAP-2 проводили тесты на функциональное ингибирование in vitro, или они в настоящее время являются общепризнанными, соответственно.
Наконец, параметры кинетики взаимодействия между NOX-E36 и CCL2/MCP-1, CCL8/MCP-2, CCL11/эотаксином, CCL3/MIP1α, CXCL7/NAP-2, CCL7/MCP-3 и CCL13/MCP-4 определяли в "перевернутой" системе. Здесь, хемокины иммобилизовывали и инъецировали NOX-E36 (для подробного протокола, см. 7.1). Данные кинетики обобщены на фиг.24C.
7.3 Оценка функциональности против MIP-1α in vitro
Измерения Biacore показали перекрестную реактивность NOX-E36 с MIP-1α. С использованием функционального анализа in vitro на основе клеточной культуры, следовало проверить, переходит ли явное связывание в Biacore NOX-E36 с MIP-1α также в функциональность, например, антагонизм.
Для достижения этого проводили эксперименты по хемотаксису с клетками THP-1, которые могут стимулироваться MIP-1α. Клетки THP-1, выращенные, как описано выше, центрифугировали, промывали один раз в HBH (HBSS, содержащая 1 мг/мл бычий сывороточный альбумин и 20 мМ HEPES) и ресуспендировали в количестве 3×106 клеток/мл. 100 мкл этой суспензии добавляли к вставкам Transwell с порами размером 5 мкм (Corning, #3421). В нижних отделениях перед добавлением клеток MIP-1α предварительно инкубировали со шпигельмерами в различных концентрациях в 600 мкл HBH при 37°C в течение от 20 до 30 мин. Клеткам позволяли мигрировать при 37°C в течение 3 часов. Затем вставки удаляли и в нижние отделения добавляли 60 мкл 440 мкМ резазурина (Sigma) в фосфатно-солевом буфере. После инкубации при 37°C в течение 2,5 часов, измеряли флуоресценцию при длине волны возбуждения 544 нм и длине волны испускания 590 нм в устройстве для считывания планшетов со множественной детекцией Fluostar Optima (BMG).
После миграции в течение 3 часов клеток THP-1 в направлении различных концентраций MIP-1α человека, получили кривую "доза-эффект" для MIP-1α человека, указывающую на половинную максимальную эффективную концентрацию, составляющую приблизительно 1 нМ и сниженную активацию при более высоких концентрациях (фиг. 24D). Для дополнительных экспериментов по ингибированию хемотаксиса шпигельмерами использовали концентрацию MIP-1α, составляющую 0,5 нМ.
Эксперименты по определению ингибирования хемотаксиса посредством NOX-E36 проводили со стимулом, представляющим собой 0,5 нМ MIP-1α. Можно было отчетливо показать, что NOX-E36 не ингибирует индуцируемый MIP-1α хемотаксис вплоть до наиболее высокой тестируемой концентрации, составляющей 1 мкМ MIP-1α. В качестве положительного контроля параллельно проводили соответствующий эксперимент с MCP-1 в качестве стимула (фиг. 24E).
Пример 8: Лечение подобного волчанке заболевания у мышей MRL lpr/lpr шпигельмером против mMCP-1
Блокирование провоспалительных медиаторов стало успешным подходом для лечения хронического воспаления (Steinman 2004). В дополнение к TNF и интерлейкинам, CC-хемокины являются важными кандидатами для специфического антагонизма, поскольку CC-хемокины опосредуют привлечение лейкоцитов из внутрисосудистого пространства в области воспаления (Baggiolini 1998, Luster 2005). Существует убедительные доказательства, что MCP-1 (=CCL2) и его соответствующий рецептор хемокинов CCR2 играют важную роль в аутоиммунном воспалении тканей, таком как клинические проявления системной красной волчанки (Gerard & Rollins 2001). Например, мыши MRLlpr/lpr, дефицитные либо по гену Ccl2, либо по гену Ccr2, защищены от подобного волчанке аутоиммунитета (Perez de Lema 2005, Tesch 1999). Таким образом, ось CCL2/CCR2 может представлять собой перспективную терапевтическую мишень, например при волчаночном нефрите. В действительности, как замедленная генная терапия, так и перенос трансфицированных клеток, приводящие к продукции in situ NH2-укороченного MCP-1, значительно снижали аутоиммунное повреждение тканей у мышей MRLlpr/lpr. Однако такие экспериментальные подходы нельзя использовать у человека вследствие неподавляемой продукции антагониста и образования опухоли (Hasegawa 2003, Shimizu 2004). Таким образом, остается необходимость в разработке новых антагонистов CCL2 с благоприятными фармакокинетическими профилями in vivo. В этом примере показано, что блокада CCL2 мыши шпигельмером против mCCL2 mNOX-E36 или mNOX-E36-3'PEG может быть пригодной для лечения волчаночного нефрита и других проявлений системной красной волчанки. Терапия шпигельмером против mCCL2 с поздним началом приводит к эффективному улучшению волчаночного нефрита, аутоиммунного перибронхита, и подобного волчанке заболевания кожи у мышей MRLlpr/lpr, независимо от какой-либо предшествующей проблемы, связанной с терапевтической блокадой CCL2/CCR2.
Животные и протокол эксперимента
Самок мышей MRLlpr/lpr в возрасте десяти недель получали от Harlan Winkelmann (Borchen, Germany) и держали в нормальных условиях содержания при 12-часовом цикле света и темноты. Вода и стандартный корм (Ssniff, Soest, Germany) были доступны без ограничений. В возрасте 14 недель, группам из 12 мышей проводили подкожные инъекции шпигельмеров в 5% глюкозе (объем инъекции, 4 мл/кг) три раза в неделю следующим образом: mNOX-E36, 1,5 мкмоль/кг; mNOX-E36-3'PEG, 0,9 мкмоль/кг; нефункциональный контрольный шпигельмер PoC (5'-UAAGGAAACUCGGUCUGAUGCGGU AGCGCUGUGCAGAGCU-3'), 1,9 мкмоль/кг; PoC-PEG, 0,9 мкмоль/кг; носитель (5% глюкоза). Уровни mNOX-E36 и mNOX-E36-3'PEG в плазме определяли из образцов крови, которые получали каждую неделю из ретроорбитального синуса через 3 или 24 часов часа после инъекции, соответственно. Уровни шпигельмера в образцах плазмы определяли путем модификации способа "сэндвич"-гибридизации, как описано в примере 8. Мышей умерщвляли смещением шейных позвонков в конце 24 недели жизни.
Оценка системной волчанки
Очаги повреждения кожи регистрировали с помощью полуколичественной шкалы (Schwarting 2005). Отношение масс селезенки и совокупности брыжеечных лимфатических узлов к общей массе тела вычисляли в качестве маркеров ассоциированного с волчанкой лимфопролиферативного синдрома. Образцы крови и мочи собирали от каждого животного в конце периода исследования путем кровопускания из ретроорбитального венозного сплетения под общей анестезией ингалируемым эфиром. Образцы крови и мочи собирали от каждого животного в конце исследования и определяли отношение альбумин в моче/креатинин и сывороточные титры аутоантител изотипа IgG против дцДНК, как описано ранее (Pawar 2006). Скорость гломерулярной фильтрации (GFR) определяли через 24 недели по кинетике выведении FITC-инулина из плазмы (Sigma-Aldrich, Steinheim, Germany) через 5, 10, 15, 20, 35, 60 и 90 минут после однократной болюсной инъекции (Qi 2004). Флуоресценцию определяли при возбуждении 485 нм и считывали при испускании 535 нм. GFR вычисляли на основе двухкомпартментной модели с использованием программного обеспечения для подгонки кривой с нелинейной регрессией (GraphPad Prism, GraphPad Software Inc., San Diego, CA). Уровни цитокинов в сыворотке определяли с использованием коммерческих наборов ELISA для IL-6, IL-12p40 (OptEiA, BD Pharmingen), и IFN-α (PBL Biomedical Labs, USA). От всех мышей, почки и легкие фиксировали в 10% забуференном формалине, обрабатывали и погружали в парафин. Получали 5-мкм срезы для окраски серебром и перйодной кислотой по Шифу в соответствии с общепринятыми протоколами (Anders 2002). Тяжесть очагов повреждения в почке оценивали с использованием индексов активности и хронического состояния, как описано для волчаночного нефрита у человека (Austin 1984), и морфометрию интерстициального повреждения проводили, как описано ранее (Anders 2002). Тяжесть перибронхиального воспаления оценивали полуколичественно от 0 до 4. Для иммунного окрашивания, срезы фиксированных формалином и погруженных в парафин тканей депарафинировали и регидратировали. Эндогенную пероксидазу блокировали 3% пероксидом водорода и проводили извлечение антигена в растворе для извлечения антигена (Vector, Burlingame, CA) в автоклаве. Биотин блокировали с использованием набора для блокирования авидина/биотина (Vector). Предметные стекла инкубировали с первичными антителами в течение одного часа, а затем с биотинилированными вторичными антителами (противокрысиный IgG, Vector), и реагентом ABC (Vector). Предметные стекла промывали между стадиями инкубации в фосфатно-солевом буфере. В качестве системы для детекции использовали 3'3'-диаминобензидин (DAB, Sigma, Taufkirchen, Germany) с усилением металлом, что приводило к продукту черного цвета. В качестве контрастной краски использовали метиловый зеленый, предметные стекла дегидратировали и помещали в Histomount (Zymed Laboratories, San Francisco, CA). Использовали следующие первичные антитела: антитело крысы против Mac2 (макрофаги, Cederlane, Ontario, Canada, 1:50), антитело против CD3 мыши (1:100, клон 500A2, BD), антитело против IgG1 мыши (1:100, M32015, Caltag Laboratories, Burlingame, CA, USA), антитело против IgG2a мыши (1:100, M32215, Caltag), антитело против C3 мыши (1:200, GAM/C3c/FITC, Nordic Immunological Laboratories, Tilburg, Netherlands). Отрицательные контроли включали инкубацию с соответствующим изотипическим антителом. Для количественного анализа клетки подсчитывали в 15 гломерулах коры на срез. Отложения Ig и C3c в гломерулах оценивали от 0 до 3 на 15 срезах гломерул коры.
Получение РНК и количественная ОТ-ПЦР в реальном времени (TaqMan)
Ткань почки от каждой мыши быстро замораживали в жидком азоте и хранили при -80°C. Для каждого животного проводили получение тотальной РНК почки и обратную транскрипцию, как описано (Anders 2002). Праймеры и зонды были от PE Biosystems, Weiterstadt, Germany. Праймеры (300 нМ), использованные для детекции Ccl2, Ccl5 и 18S rRNA представляли собой предварительно разработанный реагент для анализа TaqMan от PE Biosystems.
Проточная цитометрия
От мышей всех групп в конце исследования получали общие образцы крови и костного мозга. Проточную цитометрию проводили с использованием устройства FACScalibur и ранее охарактеризованного антитела против mCCR2 MC21 (Mack 2001). Для детекции использовали биотинилированное антитело против IgG крысы (BD Biosciences). В качестве изотипического контроля использовали IgG2b крысы (BD Biosciences).
Статистический анализ
Данные выражали в качестве среднего значения ± стандартная ошибка среднего значения (SEM). Сравнение между группами проводили с использованием одномерного ANOVA. Для множественных сравнений использовали апостериорную коррекцию Бонферрони. Значение p < 0,05 считали указывающим на статистическую значимость.
Сэндвич-анализ гибридизации
Количество шпигельмера в образцах количественно определяли с помощью сэндвич-анализа гибридизации на основе анализа, описанного Drolet et al. 2000 (Pharm Res 17:1503). Образцы крови собирали параллельно для наблюдения выведения из плазмы NOX-E36. Выбранные ткани подготавливали для определения концентраций шпигельмера.
Подготовка планшета для гибридизации
Шпигельмер mNOX-E36 количественно определяли с использованием невалидированного сэндвич-анализа гибридизации. В кратком изложении, улавливающий зонд для mNOX-E36 (SEQ ID NO:281) иммобилизовывали на белых 96-луночных планшетах DNA-BIND (Corning Costar, Wiesbaden, Germany) в концентрации 0,75 мМ в 0,5 M фосфате натрия, 1 мМ ЭДТА, pH 8,5 в течение ночи при 4°C. Лунки промывали два раза и блокировали 0,5% масс./об. BSA в 0,25 M фосфате натрия, 1 мМ ЭДТА, pH 8,5 в течение 3 ч при 37°C, снова промывали и хранили при 4°C до применения. Перед гибридизацией лунки предварительно нагревали до 37°C и промывали два раза предварительно нагретым буфером для промывания (3×SSC, 0,5% [масс./об.] додецилсаркозинат натрия, pH 7,0; заранее получаемый 20x исходный раствор [3 M NaCl, 0,3 M Na3Цитрат] без лауросаркозина натрия и разбавленный соответствующим образом).
Получение образца
Все образцы анализировали в двух экземплярах. Образцы плазмы размораживали на льду, встряхивали и кратковременно осаждали в охлажденной настольной центрифуге. Гомогенаты тканей размораживали при RT и центрифугировали в течение 5 мин при максимальной скорости и RT. Только 5 мкл каждого образца удаляли для анализа, а затем возвращали в холодильник для хранения. Образцы разбавляли буфером для гибридизации (8 нМ зонд для детекции mNOX-E36 [SEQ ID NO:282] в буфере для промывания) при RT согласно следующей схеме:
|
Анализировали все разведения образца. Стандарт mNOX-E36 серийно разбавляли для построения калибровочной кривой из 8 точек, охватывающей диапазон 0-4 нМ. Не было получено и проанализировано образцов QC. Калибровочный стандарт был идентичным стандарту в исследуемых образцах.
Гибридизация и детекция
Образцы нагревали в течение 10 мин при 95°C и охлаждали до 37°C. Комплексы шпигельмер/зонд для детекции подвергали отжигу с иммобилизованными улавливающими зондами в течение 30 мин при 37°C. Не связавшиеся шпигельмеры удаляли путем промывания два раза в буфере для промывания и 1× TBST (20 мМ Tris-Cl, 137 мМ NaCl, 0,1% Tween 20, pH 7,5), соответственно. Детекцию гибридизованных комплексов проводили стрептавидин-щелочной фосфатазой, разбавленной 1:5000 в 1×TBST, в течение 1 ч при комнатной температуре. Для удаления не связавшегося конъюгата, лунки снова промывали 1× TBST и 20 мМ Tris-Cl, 1 мМ MgCl2, pH 9,8 (по два раза каждый). Лунки в конце заполняли 100 мл субстрата CSDP (Applied Biosystems, Darmstadt, Germany) и инкубировали в течение 45 мин при комнатной температуре. Измерение хемилюминесценции проводили на устройстве для считывания микропланшетов FLUOstar Optima (BMG Labtechnologies, Offenburg, Germany).
Анализ данных
Для количественного анализа данных использовали следующие разведения образцов:
|
Данные, полученные от группы носителя (без введения шпигельмера) вычитали в качестве фонового сигнала.
Анализ сэндвич-гибридизации, как описано в настоящем документе, также аналогично работает для шпигельмера NOX-36, NOX-E36-5'-PEG и NOX-E36-3'-PEG, где необходимо использовать соответствующий улавливающий зонд для NOX-E36 (SEQ ID NO:255) и соответствующий зонд для детекции NOX-E36 (SEQ ID NO:256) (данные не представлены).
Результаты
mNOX-E36-3'PEG повышает выживаемость и приводит к улучшению при заболевании почек у мышей MRL lpr/lpr
У самок мышей MRLlpr/lpr развивается пролиферативный иммунокомплексный гломерулонефрит, поразительно сходный с диффузным пролиферативным волчаночным нефритом у человека, и они впоследствии погибают от него. В этой терапевтической схеме исследования, подвергаемым лечению мышам MRLlpr/lpr вводили ПЭГ-илированный и неПЭГ-илированный шпигельмер против mCCL2, ПЭГ-илированный и неПЭГ-илированный контрольный ("PoC")-шпигельмер или носитель от 14 недели до 24 недели жизни. За это время мыши MRLlpr/lpr, которым вводили носитель, PoC или PoC-PEG показали диффузный пролиферативный гломерулонефрит, характеризующийся гломерулярной инфильтрацией макрофагов и смешанный перигломерулярный и интерстициальный инфильтрат воспалительных клеток, состоящий из гломерулярных и интерстициальных Mac2-положительных макрофагов и интерстициальных CD3-положительных лимфоцитов (фиг. 34 и 35). mNOX-E36-3'PEG улучшал индекс активности и хронического состояния для волчаночного нефрита, а также указанные выше маркеры воспаления почек (фиг. 35). НеПЭГ-илированая молекула mNOX-E36 была менее эффективной в отношении индекса хронического состояния и количества интерстициальных макрофагов и T-клеток (фиг. 35). Хроническое заболевание почек на развернутой стадии далее иллюстрировалось атрофией канальцев и сливающимися областями интерстициального фиброза у мышей, которым вводили носитель, PoC и PoC-PEG (фиг. 34). С использованием морфометрии для количественного определения этих изменений было выявлено, что ПЭГ-илированный и неПЭГ-илированный mNOX-E36 снижали интерстициальный объем, повреждение клеток канальцев и расширение канальцев, все из которых являются маркерами тяжести и прогноза хронического заболевания почек (фиг. 36). mNOX-E36-3'PEG, но не неПЭГ-илированный mNOX-E36 приводили к 50% снижению смертности (фиг. 37). Таким образом, mNOX-E36-3'PEG может снижать число макрофагов в почке и T-клеточные инфильтраты и приводить к улучшению при волчаночном нефрите и (почечной) выживаемости мышей MRLlpr/lpr. Для исследования того, влияет ли введение mNOX-E36 и mNOX-E36-3'PEG на внутрипочечное воспаление у мышей MRLlpr/lpr, проводили ОТ-ПЦР в реальном времени для оценки уровней экспрессии провоспалительных хемокинов CCL2 и CCL5, которые, как было показано ранее, прогрессирующим образом активируются в почках мышей MRLlpr/lpr в ходе развития заболевания почек (Perez de Lema 2001). Введение mNOX-E36 и mNOX-E36-3'PEG с 14 по 24 неделю жизни снижало экспрессию в почках мРНК CCL2 и CCL5 по сравнению с контролями, которым вводили носитель (фиг. 38).
Шпигельмеры против CCL2 снижают внепочечное аутоиммунное повреждение тканей у мышей MRL lpr/lpr
Кожа и легкие также обычно поражаются при аутоиммунном повреждении тканей у мышей MRLlpr/lpr. У мышей, которым вводили носитель, аутоиммунное заболевание легких характеризовалось умеренными перибронхиолярными и околососудистыми инфильтратами воспалительных клеток и повреждения кожи наблюдали у 60% мышей (фиг. 39, 40 и 35). Как mNOX-E36, так и mNOX-E36-3'PEG, снижали перибронхиальное воспаление и заболевание кожи по сравнению с мышами MRLlpr/lpr, которым вводили носитель, PoC и PoC-PEG, соответственно (фиг. 39, 40 и 35). Таким образом, эффекты CCL2-специфических шпигельмеров не ограничиваются волчаночным нефритом, а распространяются на другие проявления аутоиммунного повреждения тканей у мышей MRLlpr/lpr.
mNOX-E36 и лимфопролиферативный синдром, аутоантитела к дцДНК, и уровни цитокинов в сыворотке у мышей MRL lpr/lpr
У самок мышей MRLlpr/lpr развивается лимфопролиферативный синдром, характеризующийся массивной спленомегалией и увеличением шейных, подмышечных, паховых и мезентериальных лимфатических узлов. Как mNOX-E36, так и mNOX-E36-3'PEG не оказывали эффекта на массу селезенок и лимфатических узлов мышей MRLlpr/lpr (фиг. 41). Аутоиммунитет у мышей MRLlpr/lpr характеризуется продукцией аутоантител против множества ядерных антигенов, включая дцДНК. У мышей MRLlpr/lpr в возрасте 24 недель сывороточные аутоантитела против дцДНК IgG, IgG1, IgG2a, IgG2b присутствовали на высоких уровнях. Как mNOX-E36, так и mNOX-E36-3'PEG не оказывали эффекта ни на одно из этих аутоантител против ДНК (фиг. 41). Подобное волчанке заболевание у мышей MRLlpr/lpr, которым вводили носитель, характеризовалось повышенными уровнями в сыворотке IFN-α, IL-12p40 и IL-6. Как mNOX-E36, так и mNOX-E36-3'PEG не оказывали эффекта на какой-либо из этих медиаторов воспаления (фиг. 41). Таким образом, оба варианта mNOX-E36 не влияют на лимфопролиферацию, продукцию IgG против дцДНК и уровни цитокинов в сыворотке у мышей MRLlpr/lpr.
Уровни mNOX-E36 и mNOX-E36-3'PEG в плазме мышей MRL lpr/lpr
Уровни mNOX-E36 и mNOX-E36-3'PEG в плазме определяли с недельными интервалами для мониторинга экспозиции лекарственного средства в ходе прогрессирующего заболевания почек у мышей MRLlpr/lpr. Средние уровни mNOX-E36 в плазме через 3 ч после инъекции и mNOX-E36-3'PEG через 24 ч после инъекции составляли приблизительно 300 нМ и 1 мкМ на протяжении исследования, соответственно (фиг. 42). Таким образом, ПЭГ-илирование повышало уровни mNOX-E36 в плазме, и прогрессирующее заболевание почек у мышей MRLlpr/lpr не модулировало фармакокинетику обоих шпигельмеров.
mNOX-E36-3'PEG блокирует эмиграцию моноцитов из костного мозга
Было показано, что эмиграция моноцитов из костного мозга в ходе бактериальной инфекции вовлекает рецептор хемокинов CCR2 (Serbina 2006), но роль CCL2 в контексте аутоиммунитета остается гипотетической. Таким образом, исследовали популяцию CCR2-положительных моноцитов в периферической крови и костном мозге групп мышей MRLlpr/lpr, которым вводили mNOX-E36-3'PEG и носитель в возрасте 24 недель. Введение mNOX-E36-3'PEG повышало процент положительных по CCR2 клеток в костном мозге с 13% до 26%, в то время как оно снижало эту популяцию в периферической крови от 26% до 11% (фиг. 43). Эти данные подтверждают роль CCL2 в выходе положительных по CCR2 клеток из костного мозга в процессе аутоиммунного заболевания мышей MRLlpr/lpr.
Обобщение
С использованием технологии шпигельмеров, был создан новый и специфический антагонист mCCL2, который потенциально блокирует mCCL2 in vitro и in vivo. В действительности, лечение с поздним началом с помощью шпигельмера против CCL2 приводило к значительному улучшению при развернутом подобном волчанке аутоиммунном повреждении ткани у мышей MRLlpr/lpr. Эти данные подтверждают центральную роль CCL2 в хроническом воспалительном повреждении ткани и идентифицируют шпигельмеры CCL2 в качестве нового лекарственного средства от аутоиммунного повреждения ткани.
Пример 9: Лечение диабетической нефропатии у диабетических мышей с односторонней нефрэктомией шпигельмером против mMCP-1
Диабетическая нефропатия остается ведущей причиной заболевания почек на конечной стадии, поскольку нацеливание на ангиотензин-зависимые патологические механизмы не всегда предотвращают прогрессирование заболевания (Zimmet 2001; Ritz 1999; United States Renal Data System 2004; Svensson 2003). Таким образом, требуется добавление других стратегий лечения для терапевтического контроля диабетической нефропатии.
Данные недавних экспериментальных исследований связаны с прогрессированием диабетической нефропатии во внутрипочечное воспаление (Galkina 2006; Mora 2005; Meyer 2003; Tuttle 2005). Например, микофенолата мофетил, метотрексат или облучение снижают экскрецию альбумина с мочой и гломерулосклероз у крыс с индуцируемой стрептозотоцином диабетической нефропатией (Yozai 2005; Utimura 2003). В настоящее время, молекулярные и клеточные механизмы внутрипочечного воспаления при диабетической нефропатии остаются слабо охарактеризованными. Пациенты с диабетической нефропатией имеют повышенные уровни в сыворотке маркеров острой фазы воспаления, но это может не отражать внутрипочечное воспаление (Dalla Vestra 2005; Navarro 2003). У пациентов с диабетической нефропатией экскретируются высокие уровни CC-хемокинового хемоаттрактантного белка моноцитов 1 (MCP-1/CCL2) в моче, который может быть более специфичным для внутрипочечного воспаления (Morii 2003; Tashiro 2002; Takebayashi 2006). В действительности, MCP-1/CCL2 экспрессируется мезангиальными клетками человека, на которые воздействуют либо высокими концентрациями глюкозы, либо поздними продуктами гликирования (Ihm 1998; Yamagishi 2002). CCL2 вовлечен в комплексный многостадийный процесс привлечения лейкоцитов из внутрисосудистых компартментов во внесосудистые компартменты, т.е. гломерулы и интерстиций почки (Baggiolini 1998). В действительности, макрофагальные инфильтраты часто встречаются у человека и при экспериментальном диабетическом гломерулосклерозе и тубулоинтерстициальном повреждении (Bohle 1991; Furuta 1993; Chow 2007). Дефицитные по Ccl2 мыши с диабетом 1 типа или 2 типа имеют сниженное число гломерулярных макрофагов, которое связано с меньшим гломерулярным повреждением (Chow 2004; Chow 2006). В этих исследованиях также была показана функциональная роль CCL2 в гломерулярной патологии нефропатии при диабете 1 типа и 2 типа. Таким образом, CCL2 может представлять собой потенциальную терапевтическую мишень для диабетической нефропатии, и пригодные CCL2 антагонисты с благоприятными фармакокинетическими профилями должны быть проверены в этом контексте заболевания. В этом примере мы описываем эффекты ПЭГ-илированного шпигельмера против CCL2 mNOX-E36-3'PEG у мышей db/db с диабетом 2 типа с диабетической нефропатией на развернутой стадии. Мы показали, что шпигельмер против CCL2 может быть пригодным для лечения диабетической нефропатии.
Животные и протокол эксперимента
Самцов мышей C57BLKS db/db или C57BLKS дикого типа в возрасте 5 недель получали от Taconic (Ry, Denmark) и содержали в клетках с фильтрующей крышкой с 12-часовым циклом темнота/свет и неограниченным доступом к корму и воде на протяжении исследования. Перед использованием клетки, подстил, гнезда, корм и воду стерилизовали автоклавированием. В возрасте 6 недель проводили одностороннюю нефрэктомию (мыши "1K") или имитирующую хирургическую операцию (мыши "2K") посредством бокового разреза размером 1 см, как ранее описано для мышей db/db и мышей дикого типа (Bower 1980). У мышей в группе имитирующей операции почку оставляли in situ. Через 10 недель в возрасте 4 месяцев, мышей 1K db/db подразделяли на две группы, которым проводили три раза в неделю подкожные инъекции либо mNOX-E36-3'PEG, либо PoC-PEG, в 5% глюкозе (доза: 0,9 мкмоль/кг; объем инъекции: 1 мл/кг). Введение продолжали в течение 8 недель (до возраста 6 месяцев), когда животных умерщвляли и получали ткани для гистопатологической оценки. Все экспериментальные процедуры были одобрены органами местного самоуправления.
Оценка диабетической нефропатии
Все иммуногистохимические исследования проводили на погруженных в парафин срезах, как описано (Anders 2002). Следующие антитела использовали в качестве первичных антител: антитело крысы против Mac2 (гломерулярные макрофаги, Cederlane, Ontario, Canada, 1:50), антитело против Ki-67 (пролиферация клеток, Dianova, Hamburg, Germany, 1:25). Для гистопатологической оценки из каждой мыши части почек фиксировали в 10% формалине в фосфатно-солевом буфере и погружали в парафин. 3-мкм срезы окрашивали реагент перйодной кислоты по Шифу или серебром в соответствии с инструкциями поставщика (Bio-Optica, Milano, Italy). Гломерулярные склеротические очаги повреждения оценивались с использованием полуколичественной шкалы "слепым" исследователем следующим образом: 0 = нет очага повреждения, 1 = < 25% склеротических, 2 = 25-49% склеротических, 3 = 50-74% склеротических, 4 = 75-100% склеротических, соответственно. Анализировали 15 гломерул на срез. Индексы интерстициального объема и расширения канальцев определяли путем наложения решетки из 100 точек на 10 неперекрывающихся областей коры, как описано ранее (Anders 2002). Количества интерстициальных клеток определялись в 15 полях под большим увеличением (hpf, 400 x) "слепым" исследователем. Получение РНК и количественную ОТ-ПЦР в реальном времени (TaqMan) проводили в депарафинизированных гломерулах. После инкубации в лизирующем буфере (10 мМ Tris-HCl, 0,1 мМ ЭДТА, 2% SDS и 20 мкг/мл протеазы K) в течение 16 ч при 60°C, проводили экстракцию РНК на основе фенол-хлороформа. Гломерулярную РНК растворяли в 10 мкл не содержащей РНКазы воды. Обратную транскрипцию и ОТ-ПЦР в реальном времени из тотальной РНК органа и гломерулярной РНК проводили, как описано (Anders 2002, Cohen 2002). Контроли, состоящие из ddH2O были отрицательными в отношении генов-мишеней и генов домашнего хозяйства. Олигонуклеотидный праймер (300 нМ) и зонды (100 нМ) для mCcl2, Gapdh и 18 S rRNA представляли собой предварительно разработанные реагенты для анализа TaqMan от PE. Праймеры и зонды были от ABI Biosystems, Weiterstadt, Germany. Скорость гломерулярной фильтрации (GFR) определяли по кинетике выведения FITC-инулина из плазмы (Sigma-Aldrich, Steinheim, Germany) через 5, 10, 15, 20, 35, 60 и 90 минут после однократной болюсной инъекции (Qi 2004). Флуоресценцию определяли при возбуждении при 485 нм и считывании при испускании при 535 нм. GFR вычисляли на основе двухкомпонентной модели с использованием программного обеспечения для подгонки кривой с нелинейной регрессией (GraphPad Prism, GraphPad Software Inc., San Diego, CA). Все данные представлены в качестве среднего значения ± SEM. Сравнение между группами проводили с использованием ANOVA и для множественных сравнений использовали апостериорную коррекцию Бонферрони. Значение p<0,05 считали указывающим на статистическую значимость.
Результаты
mNOX-E36-3'PEG снижает число гломерулярных макрофагов и общий гломерулосклероз у мышей db/db с односторонней нефрэктомией
Если отсутствие функционального CCL2 связано со сниженным привлечением гломерулярных макрофагов у мышей db/db (Chow 2007) и mNOX-E36-3'PEG способен блокировать опосредуемое CCL2 привлечение макрофагов in vitro и in vivo, mNOX-E36-3'PEG должен снижать привлечение почечных макрофагов у мышей db/db с диабетической нефропатией при диабете 2 типа на развернутой стадии. Для проверки этой гипотезы, авторы настоящего изобретения проводили подкожные инъекции mNOX-E36-3'PEG или PoC-PEG в возрасте 4 месяцев у мышей db/db ("1K") с односторонней нефрэктомией. Введение продолжали в течение 8 недель, после чего ткани собирали для оценки диабетической нефропатии. В ходе этого периода введение mNOX-E36-3'PEG не оказывало значительного влияния на число лейкоцитов и тромбоцитов, уровни глюкозы в крови или массу тела, все из которых были значительно повышены во всех группах мышей db/db по сравнению с недиабетическими мышами BLKS (данные не представлены). Интересно, что mNOX-E36-3'PEG повышал у мышей 1K db/db уровни в сыворотке CCL2, указывая на то, что антагонист CCL2 сохраняет CCL2 в кровотоке (фиг. 44). В соответствии с гипотезой авторов настоящего изобретения mNOX-E36-3'PEG значительно снижал число гломерулярных макрофагов, на 40%, по сравнению с мышами db/db, которым вводили PoC-PEG или носитель, что было связано с более низкими числами положительных по Ki-67 пролиферирующих клеток в гломеруле у мышей db/db, которым вводили mNOX-E36-3'PEG (фиг. 45). Эти данные были связаны со значительным улучшением при общем диабетическом гломерулосклерозе у мышей 1K db/db (фиг. 46). Действительно, введение mNOX-E36-3'PEG уменьшало диабетический гломерулосклероз у мышей 1K db/db до степени гломерулосклероза, присутствующей у совпадающих по возрасту мышей ("2K") db/db без нефрэктомии (фиг. 46). Эти данные показывают, что замедленная блокада CCL2-зависимого привлечения гломерулярных макрофагов посредством mNOX-E36-3'PEG препятствует общему диабетическому гломерулосклерозу мышей db/db с диабетом 2 типа.
mNOX-E36-3'PEG приводит к улучшению GFR у мышей 1K db/db
Благоприятные эффекты введения mNOX-E36-3'PEG на диабетический гломерулосклероз у мышей 1K db/db должны быть связаны с лучшей GFR. Авторы настоящего изобретения проанализировали кинетику выведения FITC-инулина в качестве маркера GFR у мышей db/db (Qi 2004). По сравнению с нормальной GFR, составляющей приблизительно 250 мл/мин у мышей db/db (Qi 2004), авторы настоящего изобретения выявили сниженную GFR, составляющую 112±23 мл/мин, у мышей 1K db/db в возрасте 6 месяцев, которым инъецировали PoC-PEG (фиг. 47). Введение mNOX-E36-3'PEG значительно повышало GFR до 231±30 мл/мин у мышей 1K db/db (p<0,001) указывая на то, что блокирование CCL2-зависимого привлечении макрофагов в гломерулы также может улучшить функцию почек у мышей с диабетом 2 типа.
mNOX-E36-3'PEG уменьшает число интерстициальных макрофагов и тубулоинтерстициальное повреждение у мышей 1K db/db
Диабетическая нефропатия на развернутой стадии у человека связана со значительными количествами интерстициальных макрофагов и тубулоинтерстициальным повреждением (Bohle 1991). У мышей 2K db/db интерстициальные макрофагальные инфильтраты и значительное тубулоинтерстециальное повреждения не встречаются до возраста 8 месяцев (Chow 2007). Ранняя односторонняя нефрэктомия ускоряет развитие тубулоинтерстициальной патологии у мышей db/db (Ninichuk 2005), таким образом, авторы настоящего изобретения провели количественное определение интерстициальных макрофагов, расширения канальцев и интерстициальный объем в качестве маркеров тубулоинтерстициального повреждения у мышей во всех группах в возрасте 6 месяцев. В этот момент времени у мышей 1K db/db были выявлены повышенные количества интерстициальных макрофагов и значительное увеличение расширения канальцев и интерстициального объема по сравнению с мышами 2K db/db (фиг. 45, фиг. 48). Введение mNOX-E36-3'PEG снижало количества интерстициальных макрофагов на 53%, а также расширение канальцев и интерстициальный объем у мышей 1K db/db (фиг. 45, фиг. 48). Таким образом, блокирование CCL2-зависимого привлечения макрофагов в почку также препятствует тубулоинтерстициальному повреждению у мышей db/db с диабетом 2 типа.
mNOX-E36-3'PEG снижает экспрессию в почках Ccl2 у мышей 1K db/db
Макрофагальные инфильтраты усиливают воспалительные ответы в поврежденной ткани, например, локальную экспрессию CCL2. Таким образом, авторы настоящего изобретения предположили, что связанное с mNOX-E36-3'PEG уменьшение количества макрофагов в почке может быть связано с меньшей экспрессией CCL2 в почках. Авторы настоящего изобретения использовали ОТ-ПЦР в реальном времени для количественного определения экспрессии мРНК CCL2 у мышей db/db. mNOX-E36-3'PEG снижал уровни мРНК CCL2 в почках мышей 1K db/db в возрасте 6 месяцев по сравнению с совпадающими по возрасту мышами, которым вводили PoC-PEG (фиг. 49). Для дальнейшей оценки пространственной экспрессии CCL2 авторы настоящего изобретения провели иммунное окрашивание на белок CCL2 на срезах почки. У мышей 1K db/db экспрессия CCL2 была значительно усилена в гломерулах, канальцах и интерстициальных клетках по сравнению с мышами 2K db/db или мышами 2K дикого типа (фиг. 50). mNOX-E36-3'PEG значительно снижал окрашивание на CCL2 в этих отделах по сравнению с мышами 1K db/db, которым вводили носитель или PoC-PEG. Эти данные указывают на то, что блокирование CCL2-зависимого привлечения макрофагов в почки посредством mNOX-E36-3'PEG снижает локальную экспрессию CCL2 у мышей 1K db/db.
Обобщение
Идея о том, что воспаление приводит к прогрессированию диабетической нефропатии у человека, становится все более признанной (Tuttle 2005), сосредотачивая внимание на MCP-1/CCL2 в качестве потенциальной мишени для лечения этого заболевания. В этом примере, авторы настоящего изобретения показали, что введение диабетическим мышам с односторонней нефроэктомией mNOX-E36-3'PEG снижало количества гломерулярных (и интерстициальных) макрофагов в возрасте 6 месяцев, что было связано с меньшим количеством пролиферирующих гломерулярных клеток. Кроме того, экспрессия в почках/гломерулах мРНК CCL2 значительно снижалась при введении mNOX-E36-3'PEG. Более того, более низкие количества глоумерулярных макрофагов и гломерулярных пролиферирующих клеток в группе лечения были связаны с защитой от общего гломерулосклероза и со значительным улучшением скорости гломерулярной фильтрации. Благоприятные эффекты mNOX-E36-3'PEG на патологию гломерул и функцию почек у диабетических мышей согласуются с исследованиями, в которых использовались другие антагонисты CCL2 в других моделях гломерулярного повреждения (Lloyd 1997, Hasegawa 2003, Tang 1996, Wenzel 1997, Fujinaka 1997, Schneider 1999). Примечательно, что замедленное начало блокады CCL2 также снижало числа интерстициальных макрофагов, что было связано с меньшей тубулоинтерстициальной патологией у мышей 1K db/db.
Взятые вместе, эти данные подтверждают, что CCL2 является перспективной терапевтической мишенью для диабетической нефропатии и указывают на то, что начало блокады CCL2 шпигельмером - даже на развернутой стадии заболевания - тем не менее, может иметь защитный эффект.
Пример 10: mNOX-E36-3'-PEG позволяет снизить дозу циклофосфамида для контроля диффузного пролиферативного волчаночного нефрита и пневмонии при волчанке у мышей MRL lpr/lpr
Контроль диффузного пролиферативного волчаночного нефрита (сокращенно DPLN) у человека требует сильной иммуносупрессиии либо циклофосфамидом (сокращенно CYC), либо микофенолата мофетилом (сокращенно MMF). Каждое из двух лекарственных средств связано с существенной болезненностью и смертностью (Appel 2007). Большинство тяжелых неблагоприятных явлений и смертей были связаны с инфекциями вследствие неспецифических иммуносупрессивных эффектов CYC и MMF (Appel 2007). Новые лекарственные средства, специфично блокирующие аутоиммунное воспаление, могут обеспечить снижение токсичности современных протоколов лечения либо путем замены CYC и MMF, либо путем обеспечения значительного снижения дозы при применении в комбинации.
Экспериментальные исследования показали, что MCP-1 и его рецептор CCR2 имеют важные роли в аутоиммунном повреждении тканей, таком как проявления системной красной волчанки (сокращенно SLE) (Gerard 2001); например, было показано, что дефицитные по MCP-1 или CCR2 мыши MRLlpr/lpr с экспериментальной SLE защищены от DPLN (Perez 2005; Tesch 1999). Благоприятный эффект блокады MCP-1 шпигельмером против mMCP-1 mNOX-E36-3'-PEG при монотерапии уже был продемонстрирован in vivo на самках мышей MRLlpr/lpr: введение mNOX-E36-3'-PEG в течение 10 недель, начиная с возраста 14 недель, приводило к значительному улучшению DPLN, как показано в примере 9. Несмотря на то, что терапевтический эффект был очевиден, осталось неясным, каким образом эффективность mNOX-E36-3'-PEG можно сравнить с эффективностью CYC или MMF. Для оценки гипотезы о том, что терапевтические эффекты, эквивалентные полной дозе CYC - которая эффективно подавляет иммунную систему - также можно достигать комбинацией CYC в низкой дозе вместе с mNOX-E36-3'-PEG, проводили второе исследование in vivo.
Животные и протокол эксперимента
Самок мышей MRLlpr/lpr в возрасте семи недель получали от Harlan Winkelmann (Borchen, Germany) и держали в нормальных условиях содержания с 12-часовым циклом света и темноты. Вода и стандартный корм (Ssniff, Soest, Germany) были доступны без ограничений. С возраста 14 недель мышам проводили в течение 10 недель следующие инъекции: (A), 5% глюкоза подкожно (группа носителя); (B), 0,89 мкмоль/кг ПЭГ-илированного контрольного шпигельмера revmNOX-E36 подкожно; (C), 0,89 мкмоль/кг mNOX-E36-3'-PEG подкожно; (D), 30 мг/кг/4 недели CYC внутрибрюшинно (низкий CYC); (E), 30 мг/кг/неделя CYC внутрибрюшинно (высокий CYC); (F), 0,89 мкмоль/кг mNOX-E36-3'-PEG вместе с 30 мг/кг/4 недели CYC (комбинация) и (G), 100 мг/кг/сутки MMF перорально (Roche, Mannheim, Germany). Все инъекции носителя и шпигельмера проводили 3x/неделя. Мышей умерщвляли смещением шейных позвонков в конце 10 недели введения. Все экспериментальные процедуры проводили согласно законодательству по уходу за животными и этике Германии и они были одобрены местными органами самоуправления.
Оценка системной волчанки
В качестве маркеров ассоциированного с волчанкой лимфопролиферативного синдрома вычисляли отношение масс селезенки и совокупности брыжеечных лимфатических узлов к общей массе тела. Отношение альбумин/креатинин в моче определяли, как описано ранее (Pawar 2006). От всех мышей, почки и легкие фиксировали в 10% забуференном формалине, обрабатывали и погружали в парафин. Получали 5-мкм срезы для окраски серебром и перйодной кислотой по Шифу в соответствии с общепринятыми протоколами (Anders 2002). Тяжесть очагов повреждения в почке оценивали с использованием индексов активности и хронического состояния, как описано для волчаночного нефрита у человека (Austin 1984). Тяжесть перибронхиального воспаления оценивалась полуколичественно от 0 до 4 "слепым" исследователем. Иммунное окрашивание проводили, как описано ранее (Anders 2002). Использовали следующие первичные антитела: антитело крысы против Mac2 (макрофаги, Cederlane, Ontario, Canada, 1:50), антитело против CD3 мыши (1:100, клон 500A2, BD). Отрицательные контроли включали инкубацию с соответствующим изотипическим антителом. Положительные гломерулярные клетки подсчитывали в 15 гломерулах коры на срез. Интерстициальные клетки подсчитывали в поле под большим увеличением (сокращенно hpf).
Статистический анализ
Данные выражали в качестве среднего значения ± стандартная ошибка среднего значения (SEM). Сравнение между группами проводили с использованием одномерного ANOVA. Для множественных сравнений использовали апостериорную коррекцию Бонферрони. Значение p<0,05 считали указывающим на статистическую значимость.
Дополнение терапии mNOX-E36-3'-PEG приводит к улучшению эффектов ежемесячного введения CYC на заболевание почек у мышей MRL lpr/lpr .
У самок мышей MRLlpr/lpr развивается пролиферативный иммунокомплексный гломерулонефрит, сходный с DPLN у человека. Мышам MRLlpr/lpr вводили CYC, MMF, шпигельмер или носитель в возрасте 14-24 неделю. Это соответствует протоколу терапевтического лечения, поскольку в возрасте 14 недель мыши MRLlpr/lpr демонстрировали DPLN с индексом по шкале активности 4,1±1,1. В этом возрасте большинство нарушений тубулоинтерстициального отдела отсутствовали (не показано). После введения в течение 10 недель, у мышей MRLlpr/lpr, которым вводили носитель и контрольный шпигельмер, был выявлен DPLN, связанный с повышенным содержанием паренхиматозных клеток в гломерулах, экспансией гломерулярного матрикса, фокальным некрозом пучков, и смешанным перигломерулярным и интерстициальным инфильтратом воспалительных клеток. CYC раз в неделю и CYC раз в месяц вместе с mNOX-E36-3'-PEG были равно эффективными в отношении повышения индекса активности и хронического состояния для волчаночного нефрита (фиг. 51A и 51B). mNOX-E36 и CYC в низкой дозе отдельно, а также MMF, были менее эффективными, но, тем не менее, значительно улучшали индексы активности и хронического состояния для волчаночного нефрита. Таким образом, добавление mNOX-E36-3'-PEG к режиму на основе CYC раз в месяц является настолько же эффективным, как терапия CYC раз в неделю, при DPLN у мышей MRLlpr/lpr.
mNOX-E36 и CYC раз в месяц оказывают аддитивные эффекты на снижение инфильтратов иммунных клеток в почках мышей MRL lpr/lpr .
Инфильтраты иммунных клеток приводят к повреждению почек при волчаночном нефрите (Vielhauer 2006), и MCP-1 опосредует привлечение T-клеток и макрофагов у мышей MRLlpr/lpr (Tesch 1999). Таким образом, было предположено, что аддитивные эффекты комбинации mNOX-E36-3'-PEG/CYC раз в месяц могут быть связаны с нарушенным привлечением макрофагов и T-клеток у мышей MRLlpr/lpr. Оценка количества гломерулярных и интерстициальных макрофагов и интерстициальных T-клеток (Mac2+ макрофаги и CD3+ T-клетки) путем иммунного окрашивания показала, что CYC раз в неделю и CYC раз в месяц вместе с mNOX-E36 были равно эффективными в отношении снижения количеств гломерулярных, а также интерстициальных Mac2+ макрофагов в почках мышей MRLlpr/lpr (фиг 51C и 51D). mNOX-E36-3'-PEG и CYC раз в месяц отдельно, а также MMF были менее эффективными, однако, тем не менее, значительно снижали количество макрофагов в обоих отделах (фиг. 51C и 51D). Те же данные были получены для количеств интерстициальных CD3-положительных T-клеток (фиг. 3E). Таким образом, аддитивный эффект mNOX-E36-3'-PEG и CYC раз в месяц на патологию почек мышей MRLlpr/lpr связан со значительным снижением интерстициальных макрофагов и T-клеток, а также гломерулярых макрофагов, который был сходен с эффектом CYC раз в неделю.
mNOX-E36-3'-PEG и CYC раз в неделю имеют аддитивные эффекты на уменьшение повреждения легких у мышей MRL lpr/lpr .
Аутоиммунный перибронхит является другим проявлением подобного волчанке системного аутоиммунитета у мышей MRLlpr/lpr. CYC раз в неделю был более эффективен, чем CYC раз в месяц в отношении контроля повреждения легких у мышей MRLlpr/lpr. Однако CYC раз в месяц вместе с mNOX-E36 были настолько же эффективными, как и CYC раз в неделю (фиг 52). Удивительно, что MMF не оказывал эффекта на повреждение легких у мышей MRLlpr/lpr.
Обобщение
Данные демонстрируют, что комбинация введения mNOX-E36 и CYC в низкой дозе, начатая в возрасте 14 недель - момент времени, когда аутоиммунное повреждение ткани уже развито (Tesch 1999; Perez 2001) - является настолько же эффективной, как и CYC в высокой дозе, в отношении подавления DPLN и повреждения легких у мышей MRLlpr/lpr. В заключение, ингибирование MCP-1 в сочетании с CYC позволяет значительное снижение дозы CYC, которое позволяет избежать тяжелого иммунотоксического эффекта CYC, несмотря на равноэффективный контроль аутоиммунного повреждения тканей, такого как DPLN. Эта новая концепция позволяет снизить тяжелую и потенциально угрожающую жизни токсичность CYC у пациентов с DPLN и другие потенциально тяжелые проявления аутоиммунного заболевания, которые вовлекают зависимые от MCP-1 инфильтраты иммунных клеток.
Пример 11: скрининговое исследование COPD - Снижение клеточного инфильтрата в легкие путем введения связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG
Гетерогенная группа хронических респираторных заболеваний включает хронический бронхит, хроническую обструктивную болезнь легких (сокращенно COPD) и астму. Гистология легких пациентов, страдающих COPD и астмой, показывает выраженную инфильтрацию дыхательных путей макрофагами и гранулоцитами, главным образом, нейтрофилами при COPD и эозинофилами при астме. В клинических исследованиях было показано, что эти параметры воспаления коррелируют со снижением функции легких и повышенным сужением бронхов (гиперреактивность дыхательных путей [сокращенно AHR]) в ответ на неспецифические стимулы. Несколько моделей in vivo имитируют хроническое воспаление при COPD, позволяя исследование функции легких в течение многих дней и стимуляции гиперсекреции слизи, связанной с нейтрофилией и AHR. Было показано, что однократное воздействие на крыс/человека липополисахарида (сокращенно LPS) вызывает острую нейтрофилию легких и AHR. Ингаляция LPS вызывает дальнейшие признаки, аналогичные COPD, а именно, прогрессирующее снижение функции легких, персистирующую AHR, и популяцию нейтрофильных воспалительных клеток в бронхоальвеолярной жидкости, вместе со сверхпродукцией оксида азота. Полагают, что медиаторы, образующиеся вследствие активации и привлечения воспалительных клеток, и LPS индуцируют фенотип пролиферации, проницаемости эпителия и гиперсекреции слизи.
В исследовании, описанном в этом отчете, использовали модель с нагрузкой с использованием бактериального LPS для оценки терапевтического эффекта связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG в индуцируемой LPS модели воспаления легких у крыс. Всем животным давали нагрузку LPS для индукции острого респираторного воспаления. Проводили терапевтическое вмешательство с помощью связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG, дексаметазона (фармакологическое эталонное вещество 1), и рофлумиласта (фармакологическое эталонное вещество 2) в различных дозах.
Дексаметазон представляет собой сильнодействующий синтетический представитель стероидных гормонов класса глюкокортикоидов. Он действует в качестве противовоспалительного средства, а также иммунодепрессанта:
(I), противовоспалительное средство: глюкокортикоиды индуцируют синтез липокортина-1 (аннексина-1), который затем связывается с клеточными мембранами, препятствуя контактированию фосфолипазы A2 с ее субстратом, арахидоновой кислотой. Это приводит к сниженной продукции эйкозаноидов. Экспрессия циклооксигеназы (как COX-1, так и COX-2) также подавляется, усиливая этот эффект. Иными словами, два основных продукта при воспалении, простагландины и лейкотриены, инибируются под действием глюкокортикоидов. Глюкокортикоиды также стимулируют вхождение липокортина-1 во внеклеточное пространство, где он связывается с мембранными рецепторами лейкоцитов и ингибирует различные воспалительные явления: адгезию к эпителию, эмиграцию, хемотаксис, фагоцитоз, дыхательный взрыв и высвобождение различных медиаторов воспаления (лизосомальных ферментов, цитокинов, тканевого активатора плазминогена, хемокинов и т.д.) из нейтрофилов, макрофагов и тучных клеток.
(II), иммунодепрессант: глюкокортикоиды подавляют клеточно-опосредуемый иммунитет. Они действуют путем ингибирования генов многих цитокинов, наиболее важным из которых является ген IL-2, который впоследствии снижает пролиферацию T-клеток. В дополнение к препятствованию T-клеточной пролиферации, другим хорошо известным эффектом является индуцируемый глюкокортикоидами апоптоз. Эффект является более выраженным в незрелых T-клетках, которые все еще находятся в тимусе, а также он оказывает влияние на периферические T-клетки. Наконец, глюкокортикоиды подавляют гуморальный иммунитет, вызывая экспрессию в B-клетках меньших количеств IL-2 и рецепторов IL-2 рецепторы. Это снижает как клональную экспансию B-клеток, так и синтез антител. Сниженные количества IL-2 также приводят к активации меньшего количества T-клеток.
Рофлумиласт представляет собой лекарственное средство, которое действует в качестве селективного ингибитора длительного действия фермента фосфодиэстеразы PDE-4. Он обладает проивовоспалительными эффектами и находится на стадии разработки в качестве перорально вводимого лекарственного средства для лечения воспалительных состояний легких, таких как астма, хроническая обструктивная болезнь легких и эмфизема. Хотя в клинических испытаниях было выявлено, что рофлумиласт является эффективным, он вызывал некоторые ограничивающие дозу побочные эффекты, включая тошноту, диарею и головную боль, и его разработка продолжается в попытках минимизировать частоту встречаемости побочных эффектов при сохранении клинической эффективности.
Животные и их содержание
В этом исследовании использовали самцов крыс Sprague Dawley, а также общепризнанную модель индуцируемого LPS воспаления. Крыс приобретали в возрасте 5 недель (приблизительно 80-110 г) от Harlan Winkelmann, Borchen, Germany и в начале исследования из возраст составил 7 недель. Животных держали в клетках Makrolon® (поликарбонат) (две крысы на клетку) и содержали в общепринятых лабораторных условиях. Клетки и подстилку из мягкого дерева (Ssniff 3/4, Soest, Germany) заменяли два раза в неделю. Проводили электронный мониторинг температуры и относительной влажности комнаты с животными и их регистрировали постоянно. Пределы были установлены на 22±2°C для температуры и 55±15% для относительной влажности. Для освещения использовали 12-часовой цикл свет/темнота, контролируемый автоматическим таймером. В качестве рациона использовали коммерческий корм в гранулах (Ssniff R/M-H V1534, Ssniff-Spezialdiäten, Soest, Germany). Корм и питьевая вода (Stadtwerke Hannover) были доступны без ограничений.
Животным позволяли приспособиться к окружающей обстановке и аклиматизироваться в течение двух недель, а затем случайным образом разделяли на группы и проводили первую сенсибилизацию. Животные, используемые в исследовании, не продемонстрировали каких-либо признаков ухудшения их состояния здоровья. Всех животных наблюдали в их клетках каждые сутки.
Материалы
Связывающий MCP-1 шпигельмер mNOX-E36-3'PEG
Носитель для mNOX-E36-3'-PEG: 5% раствор глюкозы для инъекции
LPS: Липополисахарид из Escherichia coli 0111:B4 (Sigma/Aldrich, партия No. 76K4085). Свежий рабочий раствор получали в день применения.
Фармакологическое эталонное средство (1): Дексаметазона дигидрофосфат натрия, Ratiopharm, партия No. H22416, раствор 4 мг/мл. Исходный раствор хранили после открытия в холодильнике в течение 7 суток. Свежий рабочий раствор получали в день применения.
Фармакологическое эталонное средство (2): Рофлумиласт (селективный ингибитор PDE4, партия No. K429927). Свежий рабочий раствор получали в день применения.
Носитель для дексаметазона: Фосфатно-солевой буфер Дульбекко (сокращенно DPBS) - 0,0095 M (PO4) без Ca++ и Mg++
Проведение исследования
Всех животных взвешивали и случайным образом разделяли на группы перед первой сенсибилизацией: с учетом их массы их распределили равномерно в группы из десяти животных каждая. После распределения в группы проверяли средние значения и стандартное отклонение средних масс тела (± SD), и оно составило менее 20% каждой группе, а также между группами. Массы тел животных измеряли и регистрировали по отдельности.
На 1 сутки исследования проводили нагрузку LPS путем ингаляции, обеспечивающей введенную дозу приблизительно 2,93 мкг LPS. Всем животным групп положительного и отрицательного контроля вводили внутривенно носитель (5% глюкозу) за один час до нагрузки LPS (положительный контроль) или проводили имитирующую нагрузку чистым воздухом (отрицательный контроль). Животным в группе фармакологического контроля (1) вводили 2 мг/кг дексаметазона за 18 и 1 час до нагрузки LPS внутрибрюшинно; животным в группе фармакологического контроля (2) вводили 600 мкг рофлумиласта на животное внутрижелудочно. Введение с использованием связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG проводили в четырех различных дозах путем внутривенной инъекции за один час до нагрузки LPS (0,02 мг/кг; 0,2 мг/кг; 2 мг/кг; 20 мг/кг).
Через 24 часа после нагрузки, животных умерщвляли безболезненно смертельной дозой пентобарбитала натрия и собирали бронхоальвеолярный лаваж (сокращенно BAL). Лаваж легких животных проводили пять раз, каждый раз с помощью 5,0 мл ледяного 0,9% NaCl. Для оценки BAL, супернатант первого лаважа разделяли на аликвоты после осаждения клеток центрифугированием. После этого, клетки от всех лаважей объединяли и центрифугировали сразу после сбора (10 мин при 1200 Е/мин). Клетки ресуспендировали в 1 мл PBS и подсчитывали автоматически в цитометре CasyÒ. Клеточные пятна подготавливали и окрашивали в соответствии с Pappenheim для оценки количеств разных клеток. Анализировали воспалительное состояние в легких, включая числа макрофагов/моноцитов, нейтрофилов, эозинофилов и лимфоцитов путем подсчета всего 400 клеток на клеточное пятно.
Статистические способы
Для тестирования значимых отличий между группами использовали непараметрические тесты. Для множественного сравнения (> двух групп), проводили тест Anova и непараметрический тест Даннета. Значимыми считали отличия с p < 0,05.
Результаты
Общее число клеток в BAL было значимо сниженным в группе отрицательного контроля по сравнению с группой положительного контроля, как и ожидалось. Введение 20 мг/кг связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG привело к значительно сниженному общему числу клеток, на 41%, в группе положительного контроля в BAL. Введение с использованием дексаметазон индуцировало 71% снижение числа клеток, а рофлумиласт не показал какого-либо значимого эффекта (см. фиг. 53A).
Ингаляционная нагрузка LPS индуцировала воспаление в легком, отражающееся нейтрофилией, составляющей 71,8% нейтрофильных гранулоцитов в BAL. Жидкость лаважа легких в животных без введения в группе отрицательного контроля не содержала каких-либо нейтрофильных гранулоцитов. Введение дексаметазона и 2 или 20 мг/кг связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG привело к значимо сниженному числу нейтрофилов. Уже 2 мг/кг mNOX-E36-3'-PEG снижало число нейтрофилов приблизительно на 42% и 20 мг/кг mNOX-E36-3'-PEG привело к снижению нейтрофилов приблизительно на 48%. Введение рофлумиласта не влияло на абсолютное и относительное количество нейтрофилов в бронхоальвеолярном лаваже по сравнению с группой положительного контроля (см. фиг. 53B).
Заключение
Результаты для количеств различных клеток в BAL подтвердили индукцию острого индуцируемого LPS воспалительного ответа в легких. Терапевтическое введение крысам с нагрузкой LPS дексаметазона, как было показано, значимо предотвращает воспалительный ответ после воздействия LPS. Значимый терапевтический эффект также был получен для животных, которым вводили связывающий MCP-1 шпигельмер mNOX-E36-3'-PEG. На основании данных, представленных в настоящем документе, связывающие MCP-1 шпигельмеры имеют потенциал к применению в терапии хронических респираторных заболеваний, предпочтительно, COPD, отдельно или в комбинированной терапии, предпочтительно, в комбинированной терапии с дексаметазоном. Комбинированная терапия связывающими MCP-1 шпигельмерами с дексаметазоном имеет преимущество двух независимых способов действия для лечения хронических респираторных заболеваний, таких как COPD.
Пример 12: Эффекты связывающего MCP-1 шпигельмера mNOX-E36 при экспериментальной легочной гипертензии
Исследование, как описано в настоящем документе, проводили для определения эффектов связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG на гемодинамику и ремоделирование в общепризнанной модели индуцируемой монокроталином легочной гипертензии у крыс.
Гипертензию легочной артерии (сокращенно PAH) определяют путем оценки среднего давления в легочной артерии ≥20 мм.рт.ст. в покое, ремоделирования сосудов и гипертрофии правого желудочка. Идиопатическая PAH, также известная как первичная легочная гипертензия (сокращенно PPH), часто встречается у молодых женщин и без лечения приводит к смерти от недостаточности правого отдела сердца в пределах 3 лет. В основе тяжести заболевания лежит ремоделирование сосудов легких, характеризующееся пролиферацией и миграцией гладкомышечных клеток легочной артерии (сокращенно PASMC). Также на развернутых стадиях PAH можно наблюдать неоинтимальные очаги повреждения, как следствие пролиферации эндотелиальных клеток. Наблюдаемые патологические явления потенциально являются постоянными, с одновременным нарушением регуляции факторов роста и воспалительных медиаторов, участвующих в прогрессировании заболевания.
Монокроталин в качестве модели легочной гипертензии на грызунах
Эта модель успешно предсказала клиническую эффективность всех современных способов лечения клинической легочной гипертензии, включая простаноиды, ингибиторы фосфодиэстеразы и антагонисты рецептора эндотелина. Кроме того, она обеспечивает возможность исследования как профилактики, так и реверсии PAH. На модели с монокроталином, крысам проводят однократную подкожную инъекцию пирролизидинового алкалоидного токсина монокроталина. Токсин приводит к воспалительной легочной васкулопатии, вызывающей выраженную легочную гипертензию через 3-4 недели. Определяемые показатели для этой модели включают давление в правом желудочке, системное давление, отношение правый желудочек/левый желудочек + перегородка (RV/[LV + S]) и ремоделирование сосудов легких.
Животные
Взрослых самцов крыс Sprague Dawley (масса тела 300 - 350 г) получали от Charles River Laboratories (Sulzfeld, Germany). Эксперименты проводили согласно National Institutes of Health Guidelines on the Use of Laboratory Animals.
Протокол эксперимента
Прижизненная процедура: Монокроталин (сокращенно MCT; Sigma, Deishofen) растворяли в 1 M HCl, доводили до pH 7,4 посредством 1 M NaOH и вводили в качестве однократной подкожной инъекции в дозе 60 мг/кг массы тела, как описано. Контрольным крысам вводили равный объем изотонического солевого раствора.
Для исследований длительного вмешательства, крыс, которым вводили MCT, разделяли случайным образом на группы для введения либо 5% глюкозы в качестве плацебо (n = 10), либо связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG (n = 10 для обеих доз, 2 и 20 мг/кг, соответственно) путем подкожных инъекций 3 раза в неделю. Введение начинали через 3 недели после инъекции MCT - момент времени, когда, согласно ожиданиям, полностью устанавливалась легочная гипертензия. Животным проводили введение в течение последующих 2 недель, т.е. всего шести инъекций. На 35 сутки определяли гемодинамические параметры и получали ткань.
Гемодинамика: Для измерения гемодинамических параметров крыс подвергали анестезии. После этого крысам проводили внутримышечную инъекцию атропина (250 мкг/кг массы тела) для минимизации вазовагальных побочных эффектов в ходе препарирования. Крысам проводили трахеотомию и вентилировали с частотой 60/мин. Положительное давление в конце выдоха было установлено на 1 см H2O. Левую сонную артерию канюлировали для мониторинга артериального давления, и в правый отдел сердца помещали катетер через правую яремную вену для измерения давления в правом желудочке с помощью наполненных жидкостью датчиков силы.
Приготовление ткани: После обескровливания, легкие промывали изотоническим солевым раствором при константном давлении 22 см H2O через легочную артерию. Правое легкое лигировали в области ворот, быстро замораживали в жидком азоте, и хранили при -80°C; левую долю перфузировали в течение 5 минут с помощью фиксажа Замбони под давлением 22 см H2O через легочную артерию. Ткань фиксировали в формалине (4%) в течение 12 часов при 4°C, а затем переносили в 0,1 M фосфатный буфер.
Оценка гипертрофии правого отдела сердца: Для оценки гипертрофии правого желудочка, сердце удаляли и нарезали. Вычисляли отношение массы правого желудочка к массе левого желудочка и перегородки RV/(LV+S).
Статистический анализ
Все данные приведены в качестве среднего значения ± SEM. Отличия между группами оценивали посредством ANOVA и апостериорного теста Стьюдента-Ньюмана-Кейлса для множественных сравнений.
Результаты
Как и ожидалось, животные, которым вводили MCT/плацебо, показали резкое и статистически значимое увеличение гипертрофии правого отдела сердца по сравнению со здоровыми животными. В то время как животные, которым вводили MCT/плацебо, обладали RV/(LV+S) приблизительно 0,61, здоровые животные имели только приблизительно 0,23. Введение связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG вместо плацебо приводило у животных, которым вводили MCT, к значимо сниженной гипертрофии правого отдела сердца, составляющей приблизительно 0,39 для 2 мг/кг и приблизительно 0,45 для 20 мг/кг mNOX-E36 (см. фиг. 54A).
Наряду с гипертрофией правого отдела сердца, измеренное систолическое давление правого желудочка у крыс, которым вводили MCT/плацебо, было повышено до 69 мм. рт. ст. (здоровые животные, 29 мм. рт. ст.). Введение связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG вместо плацебо привело к значимому снижению систолического давления правого желудочка, составляющему приблизительно 46 мм. рт. ст. для 2 мг/кг и приблизительно 49 мм. рт. ст. для 20 мг/кг mNOX-E36-3'PEG (фиг. 54B).
Заключение
Результаты для гипертрофии правого отдела сердца и систолического давления правого желудочка у животных, которым вводили MCT/плацебо, подтвердили индукцию гипертензии легочной артерии посредством MCT. Введение связывающего MCP-1 шпигельмера mNOX-E36-3'-PEG крысам, которым вводили MCT, в значительной степени предотвращало гипертрофию правого отдела сердца и повышение систолического давления правого желудочка. Таким образом, связывающие MCP-1 шпигельмеры являются перспективными средствами для лечения легочной гипертензии.
Ссылки
Полные библиографические данные документов, приведенных в настоящем документе, содержание которых включено в настоящий в качестве ссылки, являются, если нет иных указаний, следующими.
Akahoshi T, Wada C, Endo H, Hirota K, Hosaka S, Takagishi K, Kondo H, Kashiwazaki S, Matsushima K (1993). Expression of monocyte chemotactic and activating factor in rheumatoid arthritis. Regulation of its production in synovial cells by interleukin-1 and tumor necrosis factor. Arthritis Rheum. 36:762
Alam R, York J, Moyars M, Stafford S, Grant JA, Lee J, Forsythe P, Sim T, Ida N (1996). Increased MCP-1, RANTES, and MIP-1α in bronchoalveolar lavage fluid of allergic asthmatic patients. Am. J. Respir. Crit. Care Med. 153:1398
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990), Basic local alignment search tool. J Mol Biol. 215(3):403-10.
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. Sep 1;25(17):3389-402.
Amann B, Tinzmann R, Angelkort B (2003). ACE inhibitors improve diabetic nephropathy through suppression of renal MCP-1. Diabetes Care 26:2421
Anders HJ, Vielhauer V, Frink M, Linde Y, Cohen CD, Blattner SM, Kretzler M, Strutz F, Mack M, Grone HJ, Onuffer J, Horuk R, Nelson PJ, Schlöndorff D (2002). A chemokine receptor CCR-1 antagonist reduces renal fibrosis after unilateral ureter ligation. J. Clin. Invest. 109:251
Anders HJ, Vielhauer V, Schlöndorff D (2003). Chemokines and chemokine receptors are involved in the resolution or progression of renal disease. Kidney Int. 63:401
Appel G, Dooley MA, Ginzler EM, Isenberg D, Jayne D, Solomons N, Wolfs D. (2007). Mycophenolate mofetil compared with intravenous cyclophosphamide as induction therapy for lupus nephritis: Aspreva Lupus Management Study (ALMS) results. J Am Soc Nephrol 18: SA-FC057 [abstract]
Aurup H Tuschl T, Benseler F, Ludwig J, Eckstein F. (1994). Oligonucleotide duplexes containing 2'-amino-2'-deoxycytidines: thermal stability and chemical reactivity. Nucleic Acids Res 22:20
Austin HA 3rd, Muenz LR, Joyce KM, Antonovych TT, Balow JE (1984). Diffuse proliferative lupus nephritis: identification of specific pathologic features affecting renal outcome. Kidney Int. 25:689
Austin HA III, Klippel JH, Balow JE, et al. Therapy of lupus nephritis: controlled trial of prednisone and cytotoxic drugs. (1986) N Engl J Med 314:614-619.
Baggiolini M (1998). Chemokines and leukocyte traffic. Nature 392:565
Baggiolini M, Dewald B, Moser B. (1994). Interleukin-8 and related chemotactic cytokines - CXC and CC chemokines. Adv. Immunol. 55:97
Banba N, Nakamura T, Matsumura M, Kuroda H, Hattori Y, Kasai K (2000). Possible relationship of monocyte chemoattractant protein-1 with diabetic nephropathy. Kidney Int. 58:684
Banisor I, Leist TP, Kalman B (2005). Involvement of β-chemokines in the development of inflammatory demyelination. J. Neuroinflammation 2:7
Bazan JF, Bacon KB, Hardiman G, Wang W, Soo K, Rossi D, Greaves DR, Zlotnik A, Schall TJ (1997). A new class of membrane-bound chemokine with a CX3C motif. Nature 385:640
Berkhout TA (1997). J Biol Chem 272:16404
Bohle A, Wehrmann M, Bogenschutz O, Batz C, Muller CA, Muller GA (1991). The pathogenesis of chronic renal failure in diabetic nephropathy. Investigation of 488 cases of diabetic glomerulosclerosis. Pathol. Res. Pract. 187:251
Boring L, Gosling J, Chensue SW, Kunkel SL, Farese RV Jr, Broxmeyer HE, Charo IF (1997). Impaired monocyte migration and reduced type 1 (Th1) cytokine responses in C-C chemokine receptor 2 knockout mice. J. Clin. Invest. 100:2552
Boring L, Gosling J, Cleary M, Charo IF (1998). Decreased lesion formation in CCR2-/- mice reveals a role for chemokines in the initiation of atherosclerosis. Nature 394:894
Boring L, Gosling J, Monteclaro FS, Lusis AJ, Tsou CL, Charo IF (1996). Molecular cloning and functional expression of murine JE (monocyte chemoattractant protein 1) and murine macrophage inflammatory protein 1alpha receptors: evidence for two closely linked C-C chemokine receptors on chromosome 9. J. Biol. Chem. 271:7551
Bossink AW, Paemen L, Jansen PM, Hack CE, Thijs LG, Van Damme J (1995). Plasma levels of the chemokines monocyte chemotactic proteins-1 and -2 are elevated in human sepsis. Blood 86:3841
Boumpas DT, Austin HA III, Fessler BJ, Balow JE, Klippel JH, Lockshin MD. Systemic lupus erythematosus: emerging concepts (1995). Part I. Renal, neuropsychiatric, cardiovascular, pulmonary, and hematologic disease. Ann Intern Med 122:940-950.
Boumpas DT, Austin HA III, Vaughn EM, et al. (1992) Controlled trial of pulse methylprednisolone versus two regimens of pulse cyclophosphamide in severe lupus nephritis. Lancet 340:741-745.
Bower G, Brown DM, Steffes MW, Vernier RL, Mauer SM (1980). Studies of the glomerular mesangium and the juxtaglomerular apparatus in the genetically diabetic mouse. Lab. Invest. 43:333
Charo IF, Myers SJ, Herman A, Franci C, Connolly AJ, Coughlin SR (1994). Molecular cloning and functional expression of two monocyte chemoattractant protein 1 receptors reveals alternative splicing of the carboxyl-terminal tails. Proc. Natl Acad. Sci. USA 91:2752
Chow F, Ozols E, Nikolic-Paterson DJ, Atkins RC, Tesch GH (2004). Macrophages in mouse type 2 diabetic nephropathy: Correlation with diabetic state and progressive renal injury. Kidney Int. 65:116
Chow FY, Nikolic-Paterson DJ, Ma FY, Ozols E, Rollins BJ, Tesch GH (2007). Monocyte chemoattractant protein-1-induced tissue inflammation is critical for the development of renal injury but not type 2 diabetes in obese db/db mice. Diabetologica 50:471
Chow FY, Nikolic-Paterson DJ, Ozols E, Atkins RC, Rollin BJ, Tesch GH (2006). Monocyte chemoattractant protein-1 promotes the development of diabetic renal injury in streptozotocin-treated mice. Kidney Int. 69:73
Cockwell P, Howie AJ, Adu D, Savage CO (1998). In situ analysis of C-C chemokine mRNA in human glomerulonephritis. Kidney Int. 54:827
Cohen CD, Gröne HJ, Gröne EF, Nelson PJ, Schlöndorff D, Kretzler M (2002). Laser microdissection and gene expression analysis on formaldehyde-fixed archival tissue. Kidney Int. 61:125
Cummins LL Owens SR, Risen LM, Lesnik EA, Freier SM, McGee D, Guinosso CJ, Cook PD. (1995). Characterization of fully 2'-modified oligoribonucleotide hetero- and homoduplex hybridization and nuclease sensitivity. Nucleic Acids Res 23:2019
Dalla Vestra M, Mussap M, Gallina P, Bruseghin M, Cernigoi AM, Saller A, Plebani M, Fioretto P (2005). Acute-phase markers of inflammation and glomerular structure in patients with type 2 diabetes. J. Am. Soc. Nephrol. 16 Suppl 1:S78
Dawson J, Miltz W, Mir AK, Wiessner C (2003). Targeting monocyte chemoattractant protein-1 signalling in disease. Expert Opin. Ther. Targets 7:35
De Bleecker JL, De Paepe B, Vanwalleghem IE, Schroder JM (2002). Differential expression of chemokines in inflammatory myopathies. Neurology 58:1779
De Boer WI, Sont JK, van Schadewijk A, Stolk J, van Krieken JH, Hiemstra PS. (2000) Monocyte chemoattractant protein 1, interleukin 8, and chronic airways inflammation in COPD (2000) J Pathol. 190(5):619-26
Dooley MA, Cosio FG, Nachman PH, et al. Mycophenolate mofetil therapy in lupus nephritis: clinical observations. (1999) J Am Soc Nephrol 10:833-839.
Drolet DW, Nelson J, Tucker CE, Zack PM, Nixon K, Bolin R, Judkins MB, Farmer JA, Wolf JL, Gill SC, Bendele RA (2000). Pharmacokinetics and safety of an anti-vascular endothelial growth factor aptamer (NX1838) following injection into the vitreous humor of rhesus monkeys. Pharm. Res. 17:1503
Eaton BE, Gold L, Hicke BJ, Janjic N, Jucker FM, Sebosta DP, Tarasow TM, Willis MC, Zichi DA (1997). Bioorg Med Chem 5:1087
Eaton BE, Gold L, Zichi DA. (1995). Let's get specific: the relationship between specificity and affinity. Chem Biol 2:633
Economou E, Tousoulis D, Katinioti A, Stefanadis C, Trikas A, Pitsavos C, Tentolouris C, Toutouza MG, Toutouzas P (2001). Chemokines in patients with ischaemic heart disease and the effect of coronary angioplasty. Int. J. Cardiol. 80:55
Egashira K, Zhao Q, Kataoka C, Ohtani K, Usui M, Charo IF, Nishida K, Inoue S, Katoh M, Ichiki T, Takeshita A (2002). Importance of monocyte chemoattractant protein-1 pathway in neointimal hyperplasia after periarterial injury in mice and monkeys. Circ. Res. 90:1167
Fujinaka H, Yamamoto T, Takeya M, Feng L, Kawasaki K, Yaoita E, Kondo D, Wilson CB, Uchiyama M, Kihara I (1997). Suppression of anti-glomerular basement membrane nephritis by administration of anti-monocyte chemoattractant protein-1 antibody in WKY rats. J. Am. Soc. Nephrol. 8:1174
Furuichi K, Wada T, Iwata Y, Kitagawa K, Kobayashi K-I, Hashimoto H, Ishiwata Y, Tomosugi N, Mukaida N, Matsushima K, Egashira K, Yokoyama H (2003). Gene therapy expressing amino-terminal truncated monocyte chemoattractant protein-1 prevents renal ischemia-reperfusion injury. J. Am. Soc. Nephrol. 14:1066
Furuta T, Saito T, Ootaka T, Soma J, Obara K, Abe K, Yoshinaga K (1993). The role of macrophages in diabetic glomerulosclerosis. Am. J. Kidney Dis. 21:480
Galasso JM, Liu Y, Szaflarski J, Warren JS, Silverstein FS (2000). Monocyte chemoattractant protein-1 is a mediator of acute excitotoxic injury in neonatal rat brain. Neuroscience 101:737
Galkina E, Ley K (2006). Leukocyte recruitment and vascular injury in diabetic nephropathy. J. Am. Soc. Nephrol. 17:368-377
Gao JL, Kuhns DB, Tiffany HL, McDermott D, Li X, Francke U, Murphy PM (1993). Structure and functional expression of the human macrophage inflammatory protein 1 alpha/RANTES receptor. J. Exp. Med. 177:1421
Garcia-Zepeda EA, Combadiere C, Rothenberg ME, Sarafi MN, Lavigne F, Hamid Q, Murphy PM, Luster AD (1996). Human monocyte chemoattractant protein (MCP)-4 is a novel CC chemokine with activities on monocytes, eosinophils, and basophils induced in allergic and nonallergic inflammation that signals through the CC chemokine receptors (CCR)-2 and -3. J. Immunol. 157:5613
Gaubitz M, Schorat A, Schotte H, Kern P, Domschke W. (1999) Mycophenolate mofetil for the treatment of systemic lupus erythematosus: an open pilot trial. Lupus 8:731-736.
Gerard C, Rollins, BJ(2001). Chemokines and disease. Nat. Immunol. 2:108
Gong X, Gong W, Kuhns DB, Ben-Baruch A, Howard OM, Wang JM (1997). Monocyte chemotactic protein-2 (MCP-2) uses CCR1 and CCR2B as its functional receptors. J. Biol. Chem. 272:11682
Gonzalo JA, Lloyd CM, Wen D, Albar JP, Wells TNC, Proudfoot A, Martinez-A C, Dorf M, Bjerke T, Coyle AJ, Gutierrez-Ramos JC (1998). The coordinated action of CC chemokines in the lung orchestrates allergic inflammation and airway hyperresponsiveness. J. Exp. Med. 188:157
Gordillo GM, Onat D, Stockinger M, Roy S, Atalay M, Beck FM, Sen CK (2004). A key angiogenic role of moncyte chemoattractant protein-1 in hemangioendothelioma proliferation. Am. J. Physiol. Cell Physiol. 287:C866
Gourley MF, Austin HA III, Scott D, et al. (1996) Methylprednisolone and cyclophosphamide, alone or in combination, in patients with lupus nephritis. Ann Intern Med 125:549-557.
Green LS et al. (1995). Chem Biol 2:683
Handel TM, Domaille PJ (1996). Heteronuclear (1H, 13C, 15N) NMR assignments and solution structure of the monocyte chemoattractant protein-1 (MCP-1) dimer. Biochemistry 35:6569
Harigai M, Hara M, Yoshimura T, Leonard EJ, Inoue K, Kashiwazaki S (1993). Monocyte chemoattractant protein-1 (MCP-1) in inflammatory joint diseases and its involvement in the cytokine network of rheumatoid synovium. Clin. Immunol. Immunopathol. 69:83
Hasegawa H, Kohno M, Sasaki M, Inoue A, Ito MR, Terada M, Hieshima K, Maruyama H, Miyazaki J, Yoshie O, Nose M, Fujita S (2003). Antagonist of monocyte chemoattractant protein 1 ameliorates the initiation and progression of lupus nephritis and renal vasculitis in MRL/lpr mice. Arthritis Rheum. 48:2555
Hatano S, Strasser R (1975). Primary pulmonary hypertension. Geneva: World Heath Organization.
Heath H, Qin S, Rao P, Wu L, LaRosa G, Kassam N, Ponath PD, Mackay CR. (1997). Chemokine receptor usage by human eosinophils. The importance of CCR3 demonstrated using an antagonistic monoclonal antibody. J Clin Invest 99:178
Holdsworth SR, Kitching AR, Tipping PG (2000). Chemokines as therapeutic targets in renal disease. Curr. Opin. Nephrol. Hypertens. 9:505
Holgate ST, Bodey KS, Janezic A, Frew AJ, Kaplan AP, Teran LM (1997). Release of RANTES, MIP-1α, and MCP-1 into asthmatic airways following endobronchial allergen challenge. Am. J. Respir. Crit. Care Med. 156:1377
Hopkins N, McLoughlin P (2002) The structural basis of pulmonary hypertension in chronic lung disease: remodelling, rarefaction or angiogenesis? J Anat. 201(4):335-48.
Hosaka S, Akahoshi T, Wada C, Kondo H. (1994). Expression of the chemokine superfamily in rheumatoid arthritis. Clin Exp Immunol 97:451
Huang DR, Wang J, Kivisakk P, Rollins BJ, Ransohoff RM (2001). Absence of monocyte chemoattractant protein 1 in mice leads to decreased local macrophage recruitment and antigen-specific T helper cell type 1 immune response in experimental autoimmune encephalomyelitis. J. Exp. Med. 193:713
Hulkower K, Brosnan CF, Aquino DA, Cammer W, Kulshrestha S, Guida MP, Rapoport DA, Berman JW (1993). Expression of CSF-1, c-fms, and MCP-1 in the central nervous system of rats with experimental allergic encephalomyelitis. J. Immunol. 150:2525
Humbert M, Ying S, Corrigan C, Menz G, Barkans J, Pfister R, Meng Q, Van Damme J, Opdenakker G, Durham SR, Kay AB (1997). Bronchial mucosal expression of the genes encoding chemokines RANTES and MCP-3 in symptomatic atopic and nonatopic asthmatics: relationship to the eosinophil-active cytokines interleukin (IL)-5, granulocyte macrophage-colony-stimulating factor, and IL-3. Am J Respir Cell Mol Biol 16:1
Ihm CG, Park JK, Hong SP, Lee TW, Cho BS, Kim MJ, Cha DR, Ha H (1998). A high glucose concentration stimulates the expression of monocyte chemotactic peptide 1 in human mesangial cells. Nephron 79:33
Ioannidis JPA, Boki KA, Katsorida EM, et al. (2000) Remission, relapse, and re-remission of proliferative lupus nephritis treated with cyclophosphamide. Kidney Int 57:258-264.
Itoh T, Nagaya N, Ishibashi-Ueda H, Kyotani S, Oya H, Sakamaki F, Kimura H, Nakanishi N (2006), Increased plasma monocyte chemoattractant protein-1 level in idiopathic pulmonary arterial hypertension, Respirology. 11(2):158-63.
Iyonaga K, Takeya M, Saita N, Sakamoto O, Yoshimura T, Ando M, Takahashi K (1994). Monocyte chemoattractant protein-1 in idiopathic pulmonary fibrosis and other interstitial lung diseases. Hum. Pathol. 25:455
Johrer K, Zelle-Rieser C, Perathoner A, Moser P, Hager M, Ramoner R, Gander H, Holtl L, Bartsch G, Greil R, Thurnher M (2005). Up-regulation of functional chemokine receptor CCR3 in human renal cell carcinoma. Clin Cancer Res 11:2459
Jolicoeur C, Lemay A, Akoum A (2001). Comparative effect of danazol and a GnRH agonist on monocyte chemotactic protein-1 expression by endometriotic cells. Am. J. Reprod. Immunol. 45:86
Jose PJ, Griffiths-Johnson DA, Collins PD, Walsh DT, Moqbel R, Totty NF, Truong O, Hsuan JJ, Williams TJ (1994). Eotaxin: a potent eosinophil chemoattractant cytokine detected in a guinea pig model of allergic airways inflammation. J. Exp. Med. 179:881
Kaburagi Y, Shimada Y, Nagaoka T, Hasegawa M, Takehara K, Sato S (2001). Enhanced production of CC-chemokines (RANTES, MCP-1, MIP-1α, MIP-1β, and eotaxin) in patients with atopic dermatitis. Arch. Dermatol. Res. 293:350
Karim MY, Alba P, Cuadrado MJ, et al. (2002) Mycophenolate mofetil for systemic lupus erythematosus refractory to other immunosuppressive agents. Rheumatology (Oxford) 41:876-882.
Kawasaki AM et al. (1993). J Med Chem 36:831
Kennedy KJ, Strieter RM, Kunkel SL, Lukacs NW, Karpus WJ (1998). Acute and relapsing experimental autoimmune encephalomyelitis are regulated by differential expression of the CC chemokines macrophage inflammatory protein-1α and monocyte chemotactic protein-1. J. Neuroimmunol. 91:98
Kim JS, Gautam SC, Chopp M, Zaloga C, Jones ML, Ward PA, Welch KM (1995). Expression of monocyte chemoattractant protein-1 and macrophage inflammatory protein-1 after focal cerebral ischemia in the rat. J. Neuroimmunol. 56:127
Kingdon EJ, McLean AG, Psimenou E, et al. (2001) The safety and efficacy of MMF in lupus nephritis: a pilot study. Lupus 10:606-611.
Kitamoto S, Egashira K (2003). Anti-monocyte chemoattractant protein-1 gene therapy for cardiovascular diseases. Expert Rev. Cardiovasc. Ther. 1:393
Kleinhans M, Tun-Kyi A, Gilliet M, Kadin ME, Dummer R, Burg G, and Nestle FO (2003). Functional expression of the eotaxin receptor CCR3 in CD30+ cutaneous T-cell lymphoma. Blood 101:1487
Ko FW, Lau CY, Leung TF, Wong GW, Lam CW, Hui DS (2006) Exhaled breath condensate levels of 8-isoprostane, growth related oncogene alpha and monocyte chemoattractant protein-1 in patients with chronic obstructive pulmonary disease. Respir Med. 100(4):630-8.
Koch AE, Kunkel SL, Harlow LA, Johnson B, Evanoff HL, Haines GK, Burdick MD, Pope RM, Strieter RM (1992). Enhanced production of monocyte chemoattractant protein-1 in rheumatoid arthritis. J. Clin. Invest. 90:772
Korbet SM, Lewis EJ, Schwartz MM, Reichlin M, Evans J, Rohde RD. (2000) Factors predictive of outcome in severe lupus nephritis. Am J Kidney Dis 35:904-914.
Kouno J, Nagai H, Nagahata T, Onda M, Yamaguchi H, Adachi K, Takahashi H, Teramoto A, and Emi M (2004). Up-regulation of CC chemokine, CCL3L1, and receptors, CCR3, CCR5 in human glioblastoma that promotes cell growth. J Neurooncol 70:301
Kurihara T, Warr G, Loy J, Bravo R (1997). Defects in macrophage recruitment and host defense in mice lacking the CCR2 chemokine receptor. J. Exp. Med. 186:1757
Kusser W (2000). Chemically modified nucleic acid aptamers for in vitro selections: evolving evolution. J Biotechnol 74:27-38
Kuziel WA, Morgan SJ, Dawson TC, Griffin S, Smithies O, Ley K, Maeda N (1997). Severe reduction in leukocyte adhesion and monocyte extravasation in mice deficient in CC chemokine receptor 2. Proc. Natl Acad. Sci. U S A 94:12053
Lehman TJ, Sherry DD, Wagner-Weiner L, et al. (1989) Intermittent intravenous cyclophosphamide therapy for lupus nephritis. J Pediatr 114:1055-1060.
Lesnik EA, Guinosso CJ, Kawasaki AM, Sasmor H, Zounes M, Cummins LL, Ecker DJ, Cook PD, Freier SM. (1993). Oligodeoxynucleotides containing 2'-O-modified adenosine: synthesis and effects on stability of DNA:RNA duplexes. Biochemistry 32:7832
Lloyd CM, Minto AW, Dorf ME, Proudfoot A, Wells TNC, Salant DJ, Gutierrez-Ramos JC (1997). RANTES and monocyte chemoattractant protein-1 (MCP-1) play an important role in the inflammatory phase of crescentic nephritis, but only MCP-1 is involved in crescent formation and interstitial fibrosis. J. Exp. Med. 185:1371
Lu BB, Rutledge BJ, Gu L, Fiorillo J, Lukacs NW, Kunkel SL, North R, Gerard C, Rollins BJ (1998). Abnormalities in monocyte recruitment and cytokine expression in monocyte chemoattractant protein-1 deficient mice. J. Exp. Med. 187:601
Lubkowski J, Bujacz G, Boque L, Domaille PJ, Handel TM, Wlodawer A (1997). The structure of MCP-1 in two crystal forms provides a rare example of variable quaternary interactions. Nat Struct Biol 4:64
Mack M, Cihak J, Simonis C, Luckow B, Proudfoot AE, Plachy J, Bruhl H, Frink M, Anders HJ, Vielhauer V, Pfirstinger J, Stangassinger M, Schlöndorff D (2001). Expression and characterization of the chemokine receptors CCR2 and CCR5 in mice. J. Immunol. 166:4697
Martinelli R, Sabroe I, LaRosa G, Williams TJ, Pease JE. The CC chemokine eotaxin (CCL11) is a partial agonist of CC chemokine receptor 2b. J Biol Chem 276:42957
Matsushima K, Morishita K, Yoshimura T, Lavu S, Kobayashi Y, Lew W, Appella E, Kung HF, Leonard EJ, Oppenheim JJ (1989). Molecular cloning of a human monocyte-derived neutrophil chemotactic factor (MDNCF) and the induction of MDNCF mRNA by interleukin 1 and tumor necrosis factor. J. Exp. Med. 167:1883
McGinnis S, Madden TL (2004). BLAST: at the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 32(Web Server issue):W20-5.
Meyer TW (2003). Immunosuppression for diabetic glomerular disease? Kidney Int. 63:377
Miller LE et al. (1993). J Physiol 469:213
Miller MD, Krangel MS (1992). Biology and biochemistry of the chemokines: a family of chemotactic and inflammatory cytokines. Crit. Rev. Immunol. 12:17
Mok CC, Ying KY, Tang S, et al. Predictors and outcome of renal flares after successful cyclophosphamide treatment for diffuse proliferative lupus glomerulonephritis. (2004) Arthritis Rheum 50:2559-2568.
Mora C, Navarro JF (2005). The role of inflammation as a pathogenic factor in the development of renal disease in diabetes. Curr. Diab. Rep. 5:399
Morii T, Fujita H, Narita T, Shimotomai T, Fujishima H, Yoshioka N, Imai H, Kakei M, Ito S (2003). Association of monocyte chemoattractant protein-1 with renal tubular damage in diabetic nephropathy. J. Diabetes Complications 17:11
Murphy PM, Baggiolini M, Charo IF, Hebert CA, Horuk R, Matsushima K, Miller LH, Oppenheim JJ, Power CA (2000). International union of pharmacology. XXII. Nomenclature for chemokine receptors. Pharmacol. Rev. 52:145
Myers SJ, Wong LM, Charo IF (1995). Signal transduction and ligand specificity of the human monocyte chemoattractant protein-1 receptor in transfected embryonic kidney cells. J. Biol. Chem. 270:5786
Nakamura H, Weiss ST, Israel E, Luster AD, Drazen JM, Lilly CM (1999). Eotaxin and impaired lung function in asthma. Am J Respir Crit Care Med 160:1952
Nakazawa T, Hisatomi T, Nakazawa C, Noda K, Maruyama K, She H, Matsubara A, Miyahara S, Nakao S, Yin Y, Benowitz L, Hafezi-Moghadam A, Miller JW (2007). Monocyte chemoattractant protein 1 mediated retinal detachment-induced photoreceptor apoptosis. Proc Natl. Acad. Sci. U S A 104:2425
Navarro JF, Mora C, Maca M, Garca J (2003). Inflammatory parameters are independently associated with urinary albumin in type 2 diabetes mellitus. Am. J. Kidney Dis. 42:53
Needleman & Wunsch (1970), A general method applicable to the search for similarities in the amino acid sequence of two proteins. J Mol Biol. 48(3):443-53.
Nelken NA, Coughlin SR, Gordon D, Wilcox JN (1991). Monocyte chemoattractant protein-1 in human atheromatous plaques. J. Clin. Invest. 88:1121
Neote K, DiGregorio D, Mak JY, Horuk R, Schall TJ (1993). Molecular cloning, functional expression, and signaling characteristics of a C-C chemokine receptor. Cell 72:415
Ninichuk V, Gross O, Reichel C, Khandoga A, Pawar RD, Ciubar R, Segerer S, Belemezova E, Radomska E, Luckow B, de Lema GP, Murphy PM, Gao JL, Henger A, Kretzler M, Horuk R, Weber M, Krombach F, Schlondorff D, Anders HJ (2005). Delayed chemokine receptor 1 blockade prolongs survival in collagen 4A3-deficient mice with Alport disease. J. Am. Soc. Nephrol. 16:977
Ogata H, Takeya M, Yoshimura T, Takagi K, Takahashi K (1997). The role of monocyte chemoattractant protein-1 (MCP-1) in the pathogenesis of collagen-induced arthritis in rats. J. Pathol. 182:106
Okuno T, Andoh A, Bamba S, Araki Y, Fujiyama Y, Fujiyama M, Bamba T (2002). Interleukin-1β and tumor necrosis factor-α induce chemokine and matrix metalloproteinase gene expression in human colonic subepithelial myofibroblasts. Scand. J. Gastroenterol. 37:317
Oppenheim JJ, Zachariae CO, Mukaida N, Matsushima K (1991). Properties of the novel proinflammatory supergene "intercrine" cytokine family. Annu. Rev. Immunol. 9:617
Pawar RD, Patole PS, Zecher D, Segerer S, Kretzler M, Schlöndorff D, Anders HJ (2006). Toll-like receptor-7 modulates immune complex glomerulonephritis. J. Am. Soc. Nephrol. 17:141
Pearson & Lipman (1988), Improved tools for biological sequence comparison. Proc. Nat'l. Acad. Sci. USA 85: 2444
Peinado VI, Pizarro S, Barberà JA. Pulmonary Vascular Involvement in COPD (2008) Chest. 134:808-814
Perez de Lema G, Maier H, Franz TJ, Escribese M, Chilla mS, Segerer S, Camarasa N, Schmid H, Banas B, Kalaydjiev S, Busch DH, Pfeffer K, Mampaso F, Schlöndorff D, Luckow B (2005). Chemokine receptor CCR2 deficiency reduces renal disease and prolongs survival in MRL/lpr lupus-prone mice. J. Am. Soc. Nephrol. 16:3592
Perez de Lema G, Maier H, Nieto E, Vielhauer V, Luckow B, Mampaso F, Schlöndorff D (2001). Chemokine expression precedes inflammatory cell infiltration and chemokine receptor and cytokine expression during the initiation of murine lupus nephritis. J. Am. Soc. Nephrol. 12:1369
Ponath PD, Qin S, Post TW, Wang J, Wu L, Gerard NP, Newman W, Gerard C, Mackay CR (1996b). Molecular cloning and characterization of a human eotaxin receptor expressed selectively on eosinophils. J. Exp. Med. 183:2437
Ponath PD, Qin S, Ringler DJ, Clark-Lewis I, Wang J, Kassam N, Smith H, Shi X, Gonzalo JA, Newman W, Gutierrez-Ramos JC, Mackay CR (1996a). Cloning of the human eosinophil chemoattractant, eotaxin. Expression, receptor binding, and functional properties suggest a mechanism for the selective recruitment of eosinophils. J. Clin. Invest. 97:604
Power CA, Meyer A, Nemeth K, Bacon KB, Hoogewerf AJ, Proudfoot AE, Wells TN (1995). Molecular cloning and functional expression of a novel CC chemokine receptor cDNA from a human basophilic cell line. J. Biol. Chem. 270:19495
Qi Z, Whitt I, Mehta A, Jin J, Zhao M, Harris RC, Fogo AB, Breyer MD (2004). Serial determination of glomerular filtration rate in conscious mice using FITC-inulin clearance. Am. J. Physiol. Renal Physiol. 286:F590
Qin S, LaRosa G, Campbell JJ, Smith-Heath H, Kassam N, Shi X, Zeng L, Buthcher EC, Mackay CR (1996). Expression of monocyte chemoattractant protein-1 and interleukin-8 receptors on subsets of T cells: correlation with transendothelial chemotactic potential. Eur. J. Immunol. 26:640
Ransohoff RM, Hamilton TA, Tani M, Stoler MH, Shick HE, Major JA, Estes ML, Thomas DM, Tuohy VK. (1993). Astrocyte expression of mRNA encoding cytokines IP-10 and JE/MCP-1 in experimental autoimmune encephalomyelitis FASEB J 7:592
Raport CJ, Gosling J, Schweickart VL, Gray PW, Charo IF (1996). Molecular cloning and functional characterization of a novel human CC chemokine receptor (CCR5) for RANTES, MIP-1β, and MIP-1α. J. Biol. Chem. 271:17161
Ritz E, Rychlik I, Locatelli F, Halimi S (1999). End-stage renal failure in type 2 diabetes: A medical catastrophe of worldwide dimensions. Am. J. Kidney Dis. 34:795-808
Rollins BJ (1996). Monocyte chemoattractant protein 1: a potential regulator of monocyte recruitment in inflammatory disease. Mol. Med. Today 2:198
Rollins BJ, Stier P, Ernst T, Wong GG (1989). The human homolog of the JE gene encodes a monocyte secretory protein. Mol. Cell Biol. 9:4687
Rovin BH, Rumancik M, Tan L, Dickerson J (1994). Glomerular expression of monocyte chemoattractant protein-1 in experimental and human glomerulonephritis. Lab. Invest. 71:536
Rubin LJ (1997) Primary pulmonary hypertension. N Engl J Med. 336(2):111-7
Ruffing N, Sullivan N, et al. (1998). CCR5 has an expanded ligand-binding repertoire and is the primary receptor used by MCP-2 on activated T cells. Cell Immunol 189:160
Salcedo R, Ponce ML, Young HA, Wasserman K, Ward JM, Keinman HK, Oppenheim JJ, Murphy WJ (2000). Human endothelial cells express CCR2 and respond to MCP-1: direct role of MCP-1 in angiogenesis and tumor progression. Blood 96:34
Samson M, Labbe O, Mollereau C, Vassart G, Parmentier M (1996). Molecular cloning and functional expression of a new human CC-chemokine receptor gene. Biochemistry 35:3362
Schall TJ, Bacon KB (1994). Chemokines, leukocyte trafficking, and inflammation. Curr. Opin. Immunol. 6:865
Schneider A, Panzer U, Zahner G, Wenzel U, Wolf G, Thaiss F, Helmchen U, Stahl RA (1999). Monocyte chemoattractant protein-1 mediates collagen deposition in experimental glomerulonephritis by transforming growth factor-beta. Kidney Int. 56:135
Schwarting A, Paul K, Tschirner S, Menke J, Hansen T, Brenner W, Kelly VR, Relle M, Galle PR (2005). Interferon-beta: a therapeutic for autoimmune lupus in MRL-Faslpr mice. J. Am. Soc. Nephrol. 16:3264
Schwartz CJ, Valente AJ, Sprague EA (1993). A modern view of atherogenesis. Am. J. Cardiol. 71:9B
Segerer S, Nelson PJ, Schlöndorff D (2000). Chemokines, chemokine receptors, and renal disease: from basic science to pathophysiologic and therapeutic studies. J. Am. Soc. Nephrol. 11:152
Shimizu S, Nakashima H, Masutani K, Inoue Y, Miyake K, Akahoshi M, Tanaka Y, Egashira K, Hirakata H, Otsuka T, Harada M (2004). Anti-monocyte chemoattractant protein-1 gene therapy attenuates nephritis in MRL/lpr mice. Rheumatology (Oxford) 43:1121
Simonneau G, Galiè N, Rubin LJ, et al (2004). Clinical classification of pulmonary hypertension". J. Am. Coll. Cardiol. 43 (12 Suppl S): 5S-12S.
Smith & Waterman (1981), Adv. Appl. Math. 2: 482
Springer TA (1995). Traffic signals on endothelium for lymphocyte recirculation and leukocyte emigration. Annu. Rev.Physiol. 57:827
Steinberg AD, Steinberg SC. Long-term preservation of renal function in patients with lupus nephritis receiving treatment that includes cyclophosphamide versus those treated with prednisone only. (1991) Arthritis Rheum 34:945-950.
Steinman L (2004). Immune therapy for autoimmune diseases. Science 305:212
Svensson M, Sundkvist G, Arnqvist HJ, Bjork E, Blohme G, Bolinder J, Henricsson M, Nystrom L, Torffvit O, Waernbaum I, Ostman J, Eriksson JW (2003). Signs of nephropathy may occur early in young adults with diabetes despite modern diabetes management: Results from the nationwide population-based Diabetes Incidence Study in Sweden (DISS). Diabetes Care 26:2903
Takebayashi K, Matsumoto S, Aso Y, Inukai T (2006). Association between circulating monocyte chemoattractant protein-1 and urinary albumin excretion in nonobese Type 2 diabetic patients. J. Diabetes Complications 20:98
Takeya M, Yoshimura T, Leonard EJ, Takahashi K (1993). Detection of monocyte chemoattractant protein-1 in human atherosclerotic lesions by an anti-monocyte chemoattractant protein-1 monoclonal antibody. Hum. Pathol. 24:534
Tang WW, Qi M, Warren JS (1996). Monocyte chemoattractant protein 1 mediates glomerular macrophage infiltration in anti-GBM Ab GN. Kidney Int. 50:665
Tashiro K, Koyanagi I, Saitoh A, Shimizu A, Shike T, Ishiguro C, Koizumi M, Funabiki K, Horikoshi S, Shirato I, Tomino Y (2002). Urinary levels of monocyte chemoattractant protein-1 (MCP-1) and interleukin-8 (IL-8), and renal injuries in patients with type 2 diabetic nephropathy. J. Clin. Lab. Anal. 16:1
Tesch GH, Maifert S, Schwarting A, Rollins BJ, Kelley VR (1999). Monocyte chemoattractant protein 1-dependent leukocytic infiltrates are responsible for autoimmune disease in MRL-Fas(lpr) mice. J. Exp. Med. 190:1813
Torres F (2007) Systematic review of randomised, double-blind clinical trials of oral agents conducted in patients with pulmonary arterial hypertension. Int J Clin Pract. 61(10):1756-65.
Traves SL, Culpitt SV, Russell RE, Barnes PJ, Donnelly LE (2002) Increased levels of the chemokines GROalpha and MCP-1 in sputum samples from patients with COPD. Thorax. 57(7):590-5.
Tuaillon N, Shen de F, Berger RB, Lu B, Rollins BJ, Chan CC (2002). MCP-1 expression in endotoxin-induced uveitis. Invest. Ophthalmol. Vis. Sci. 43:1493
Tuttle KR (2005). Linking metabolism and immunology: diabetic nephropathy is an inflammatory disease. J. Am. Soc. Nephrol. 16:1537
Uguccioni M, Mackay CR et al. (1997). High expression of the chemokine receptor CCR3 in human blood basophils. Role in activation by eotaxin, MCP-4, and other chemokines. J Clin Invest 100:1137
United States Renal Data System (2004). Annual data report: Incidence and prevalence 2004. Am. J. Kidney Dis. 45:S77
Utimura R, Fujihara CK, Mattar AL, Malheiros DM, Noronha IL, Zatz R (2003). Mycophenolate mofetil prevents the development of glomerular injury in experimental diabetes. Kidney Int. 63:209
Valeri A, Radhakrishnan J, Estes D, et al. (1994) Intravenous pulse cyclophosphamide treatment of severe lupus nephritis: a prospective five-year study. Clin Nephrol 42:71-78.
Van Riper G, Siciliano S, Fischer PA, Meurer R, Springer MS, Rosen H (1993). Characterization and species distribution of high affinity GTP-coupled receptors for human rantes and monocyte chemoattractant protein 1. J. Exp. Med. 177:851
Venkatesan N et al. (2003). Curr Med Chem 10:1973
Vestergaard C, Just H, Baumgartner Nielsen J, Thestrup-Pedersen K, Deleuran M (2004). Expression of CCR2 on monocytes and macrophages in chronically inflamed skin in atopic dermatitis and psoriasis. Acta Derm. Venereol. 84:353
Viedt C, Orth SR (2002). Monocyte chemoattractant protein-1 (MCP-1) in the kidney: does it more than simply attract monocytes? Nephrol. Dial. Transplant. 17:2043
Vielhauer V, Anders HJ (2006). Blockade of chemokine-mediated tissue injury in lupus nephritis. Endocr Metab Immune Disord Drug Targets. 6(4):313-21.
Voelkel NF, Tuder RM. (1995) Cellular and molecular mechanisms in the pathogenesis of severe pulmonary hypertension (1995) Eur Respir J. 8(12):2129-38.
Wada T, Furuichi K, Segada-Takaeda C, Ahimizu M, Sakai N, Takeda SI, Takasawa K, Kida H, Kobayashi KI, Mukaida N, Ohmoto Y, Matsushima K, Yokoyama H (1999). MIP-1α and MCP-1 contribute to crescents and interstitial lesions in human crescentic glomerulonephritis. Kidney Int. 56:995
Wada T, Yokoyama H, Furuichi K, Kobayashi KI, Harada K, Naruto M, Su SB, Akiyama M, Mukaida N, Matsushima K (1996). Intervention of crescentic glomerulonephritis by antibodies to monocyte chemotactic and activating factor (MCAF/MCP-1). FASEB J. 10:1418
Wada T, Yokoyama H, Matsushima K, Kobayashi KI (2001). Chemokines in renal diseases. Int. Immunopharmacol. 1:637
Wang X, Yue TL, Barone FC, Feuerstein GZ (1995). Monocyte chemoattractant protein-1 messenger RNA expression in rat ischemic cortex. Stroke 26:661
Wenzel U, Schneider A, Valente AJ, Abboud HE, Thaiss F, Helmchen UM, Stahl RA (1997). Monocyte chemoattractant protein-1 mediates monocyte/macrophage influx in anti-thymocyte antibody-induced glomerulonephritis. Kidney Int. 51:770
Yamagishi S, Inagaki Y, Okamoto T, Amano S, Koga K, Takeuchi M, Makita Z (2002). Advanced glycation end product-induced apoptosis and overexpression of vascular endothelial growth factor and monocyte chemoattractant protein-1 in human-cultured mesangial cells. J. Biol. Chem. 277:20309
Ying S, Meng Q, Zeibecoglou K, Robinson DS, Macfarlane A, Humbert M, Kay AB (1999). Eosinophil chemotactic chemokines (eotaxin, eotaxin-2, RANTES, monocyte chemoattractant protein-3 (MCP-3), and MCP-4), and C-C chemokine receptor 3 expression in bronchial biopsies from atopic and nonatopic (Intrinsic) asthmatics. J Immunol 163:6321
Ying S, Robinson DS, Meng Q, Rottman J, Kennedy R, Ringler DJ, Mackay CR, Daugherty BL, Springer MS, Durham SR, Williams TJ, Kay AB (1997). Enhanced expression of eotaxin and CCR3 mRNA and protein in atopic asthma. Association with airway hyperresponsiveness and predominant co-localization of eotaxin mRNA to bronchial epithelial and endothelial cells. Eur J Immunol 27:3507
Yla-Herttuala S, Lipton BA, Rosenfeld ME, Sarkioja T, Yoshimura T, Leonard EJ, Witztum JL, Steinberg D (1991). Expression of monocyte chemoattractant protein 1 in macrophage-rich areas of human and rabbit atherosclerotic lesions. Proc. Natl Acad. Sci. U S A 88:5252
Yoshimura T, Robinson EA, Tanaka S, Appella E, Leonard EJ (1989). Purification and amino acid analysis of two human monocyte chemoattractants produced by phytohemagglutinin-stimulated human blood mononuclear leukocytes. J. Immunol. 142:1956
Yozai K, Shikata K, Sasaki M, Tone A, Ohga S, Usui H, Okada S, Wada J, Nagase R, Ogawa D, Shikata Y, Makino H (2005). Methotrexate prevents renal injury in experimental diabetic rats via anti-inflammatory actions. J. Am. Soc. Nephrol. 16:3326
Zimmet P, Alberti KG, Shaw J (2001). Global and societal implications of the diabetes epidemic. Nature 414:782
Признаки настоящего изобретения, описанного в описании, формуле изобретении и/или на рисунках, как по отдельности, так и в любой их комбинации, могут быть материалом для осуществления изобретения в различных его формах.

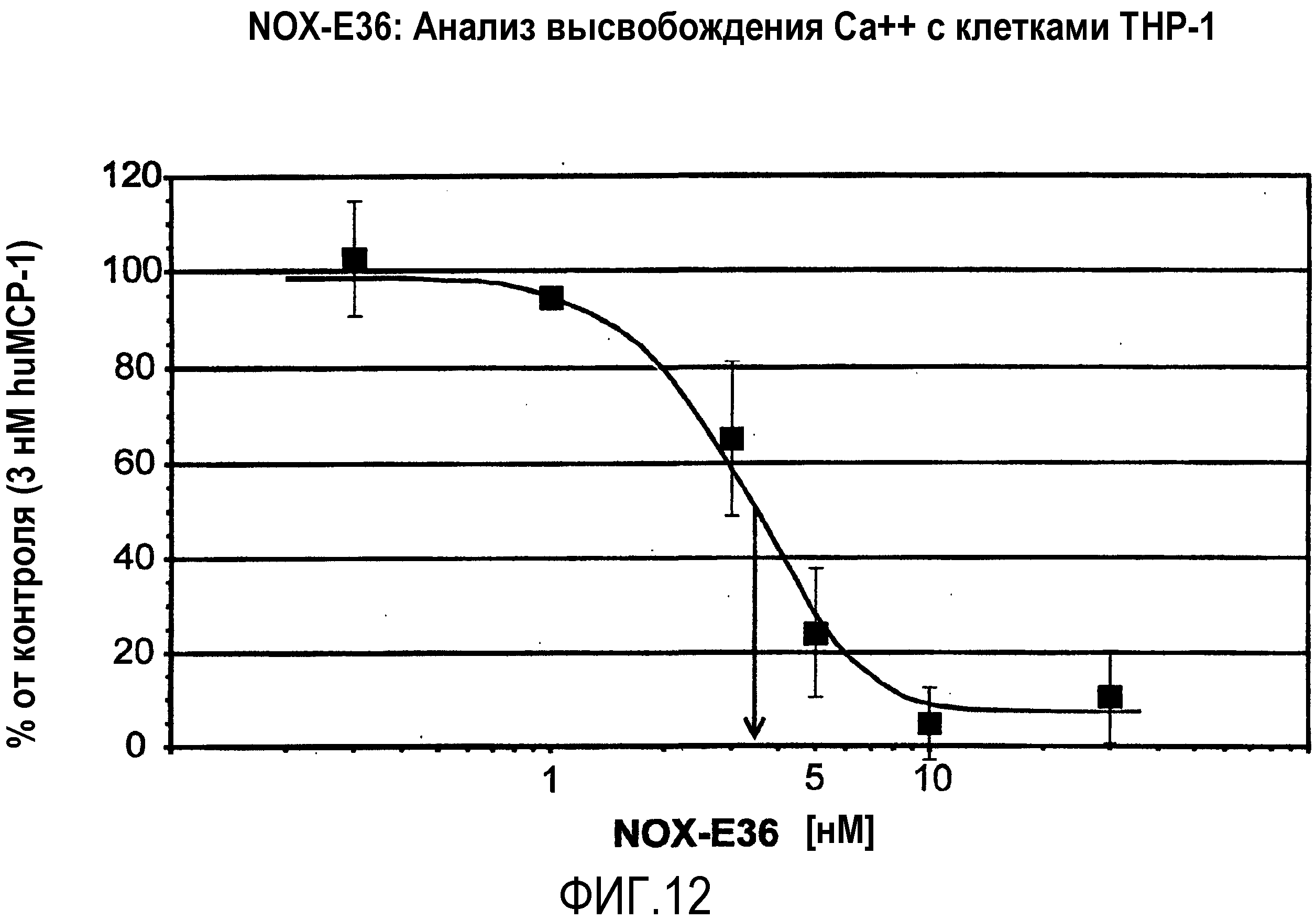

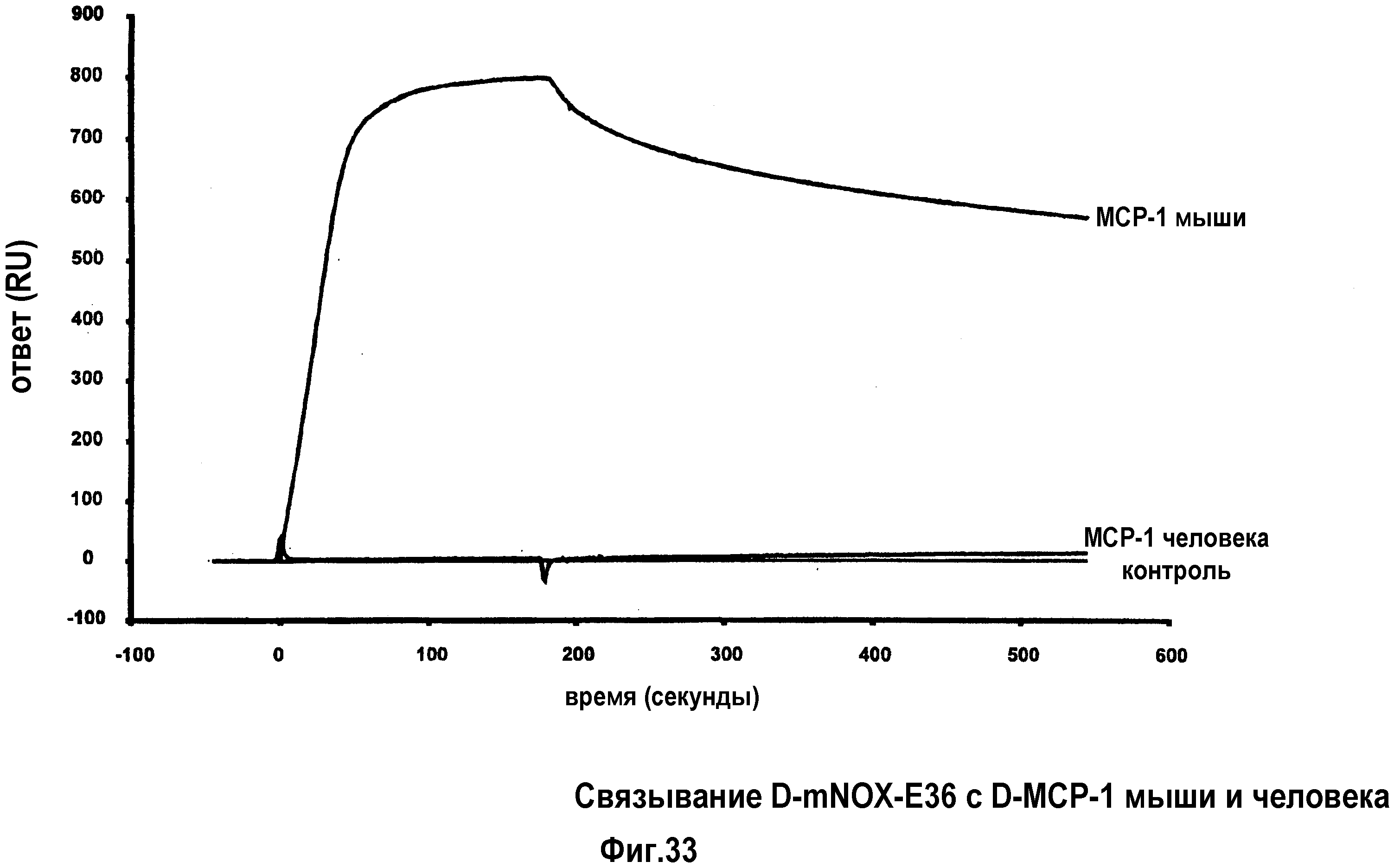
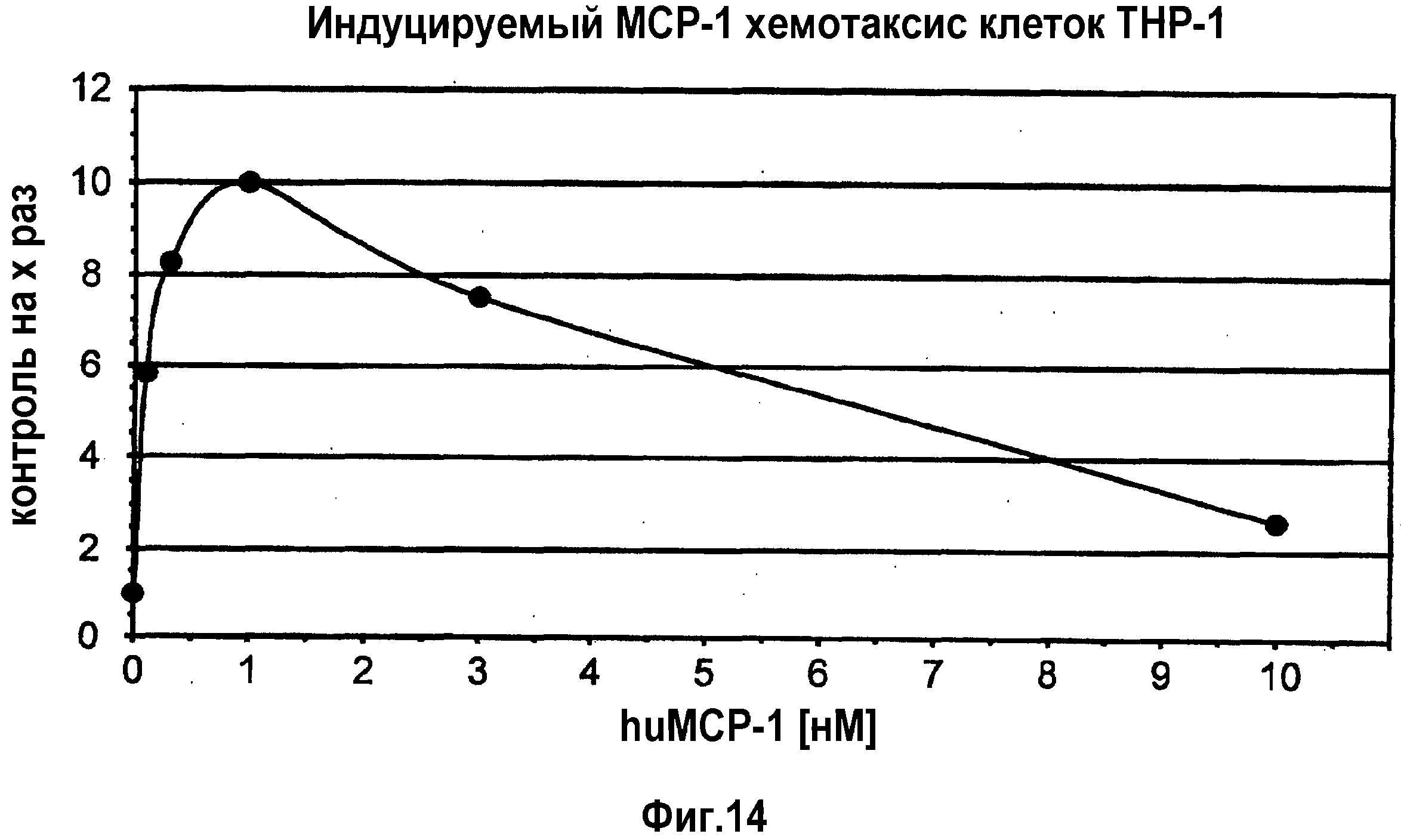
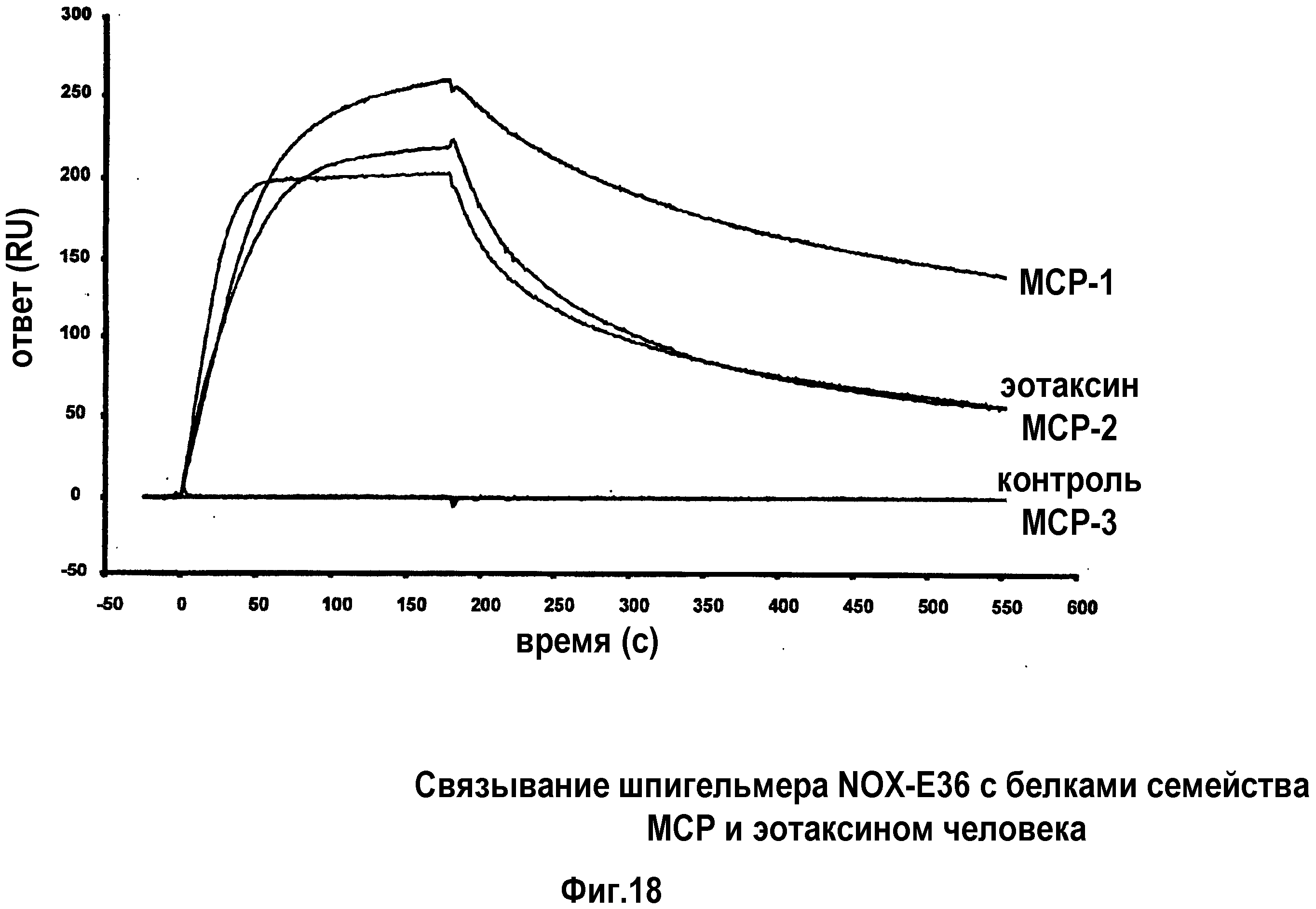
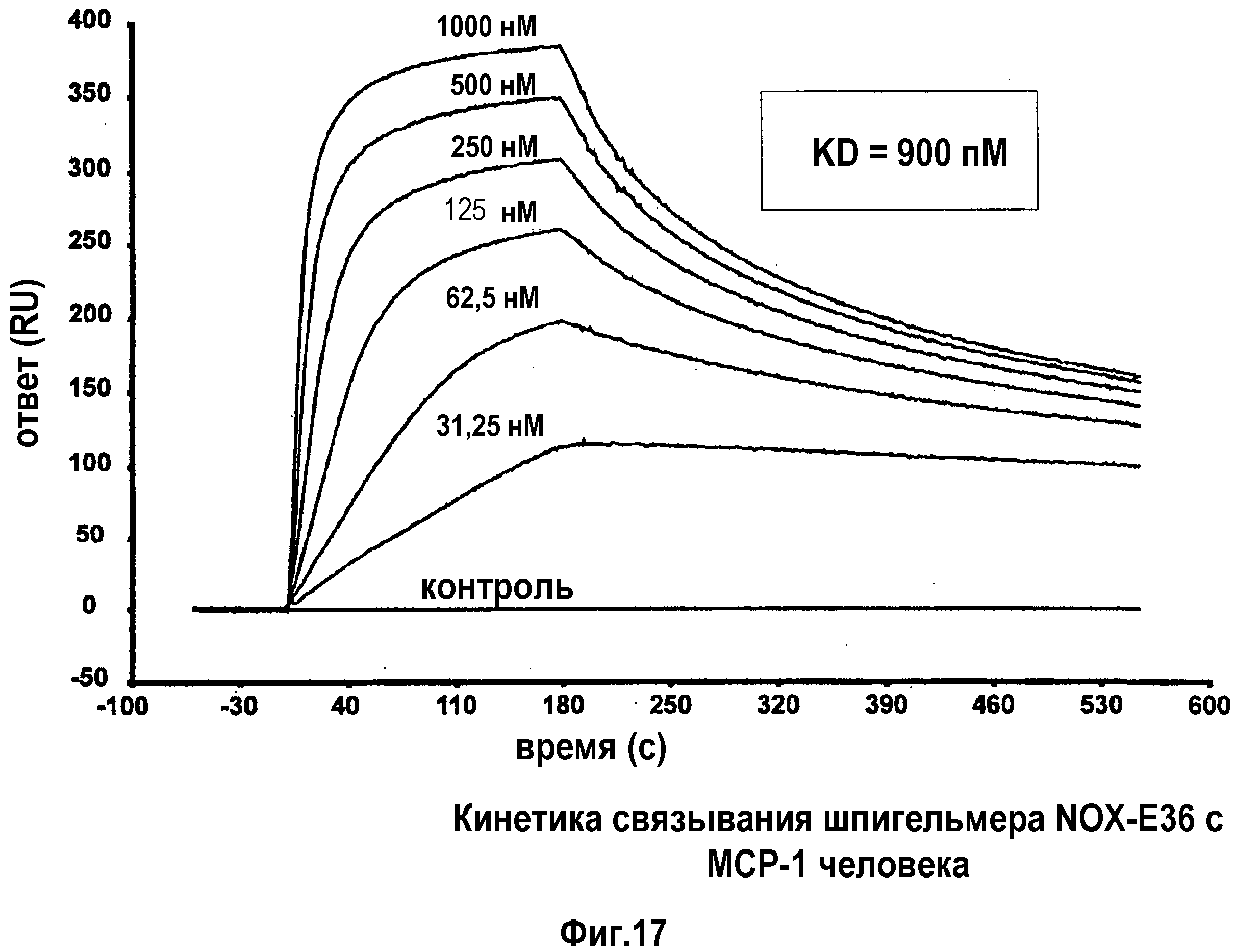

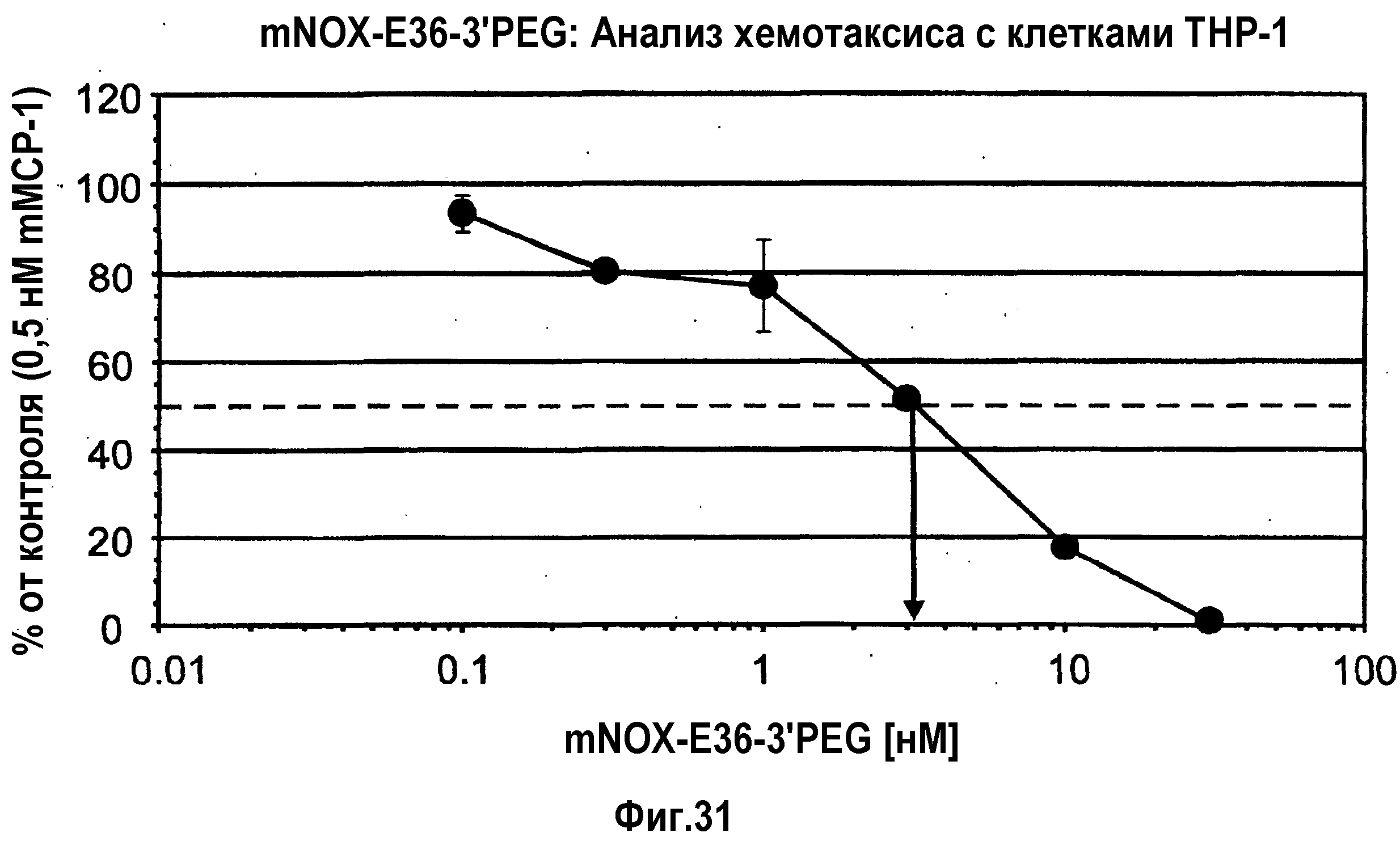

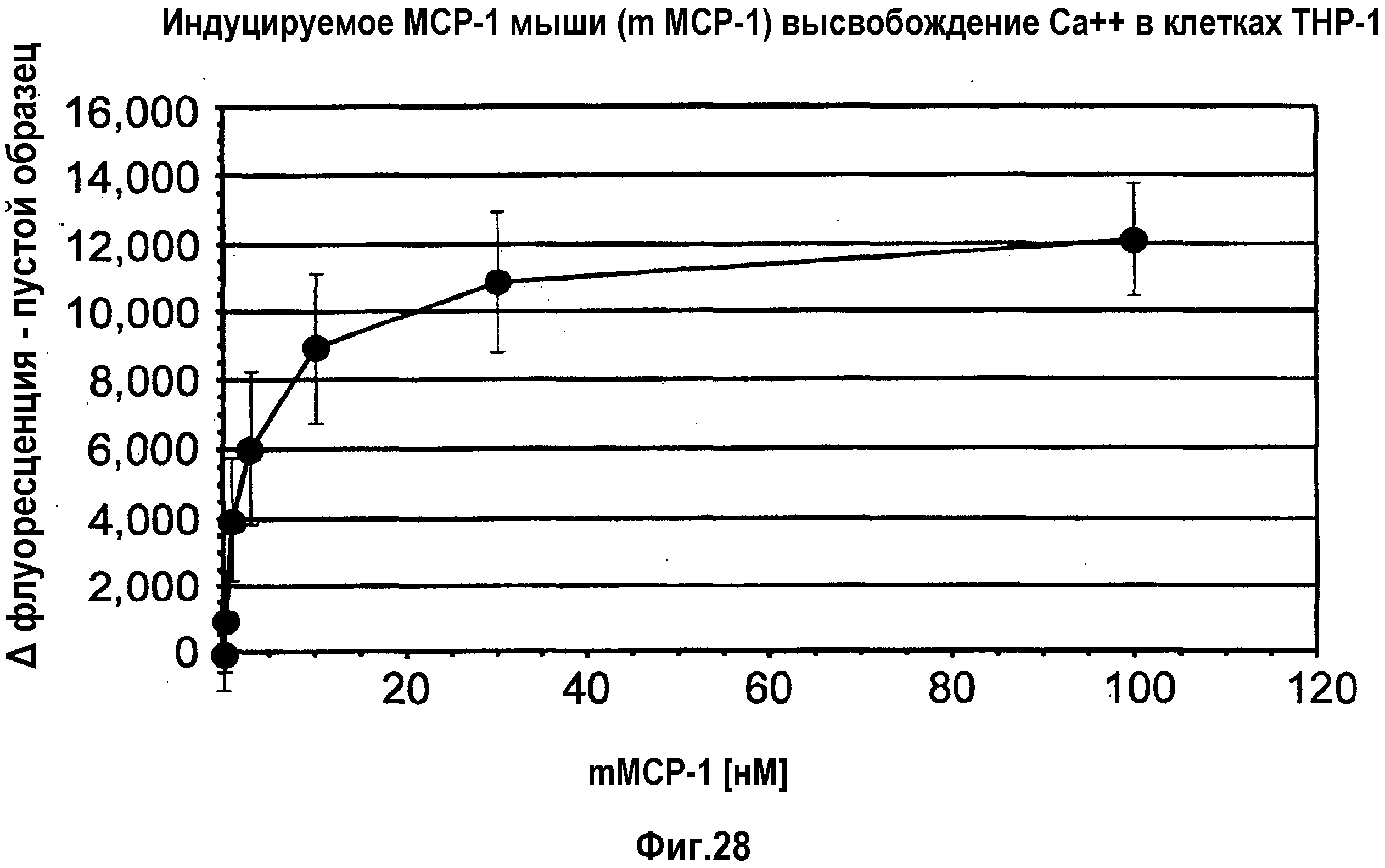





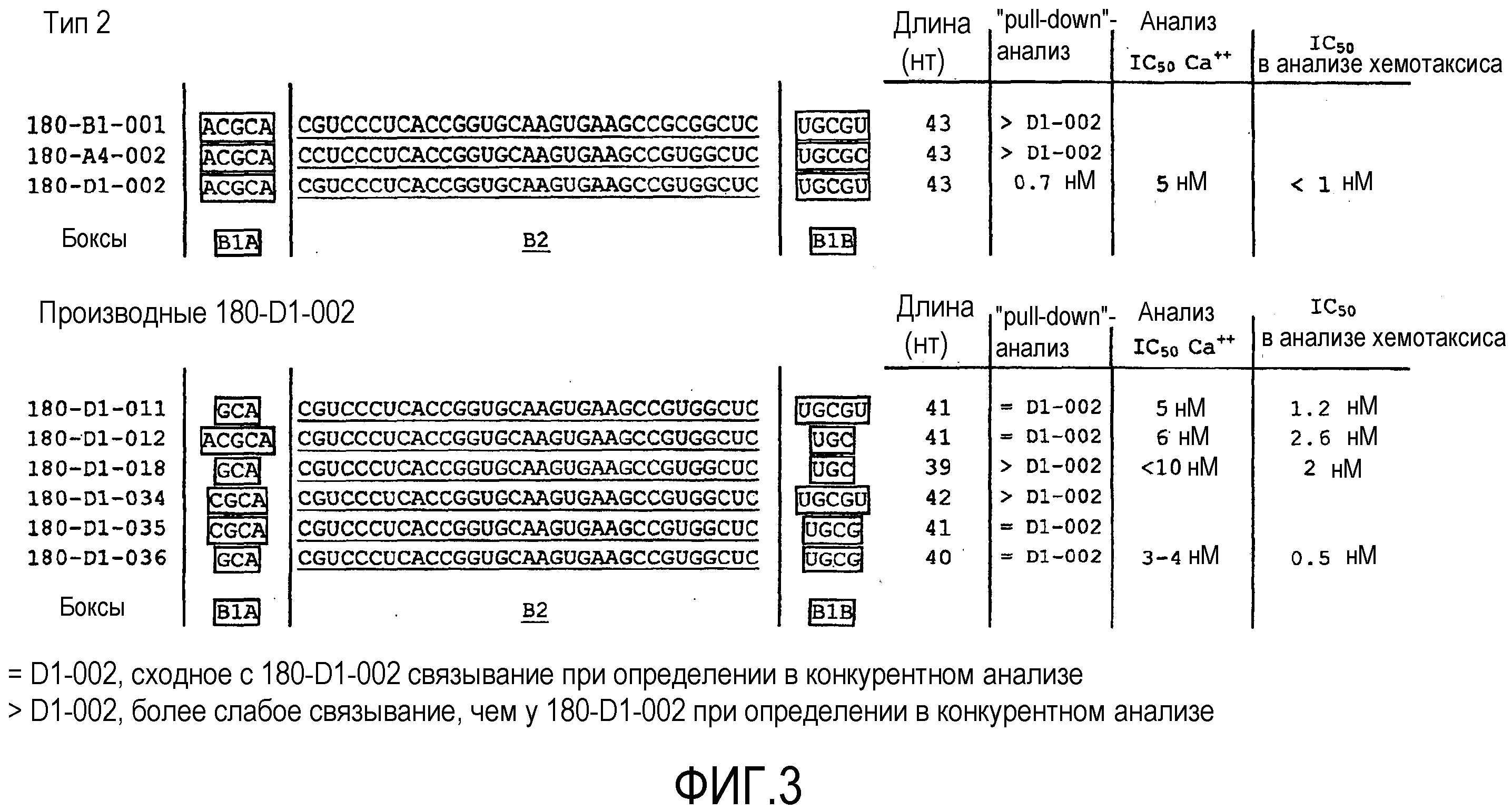
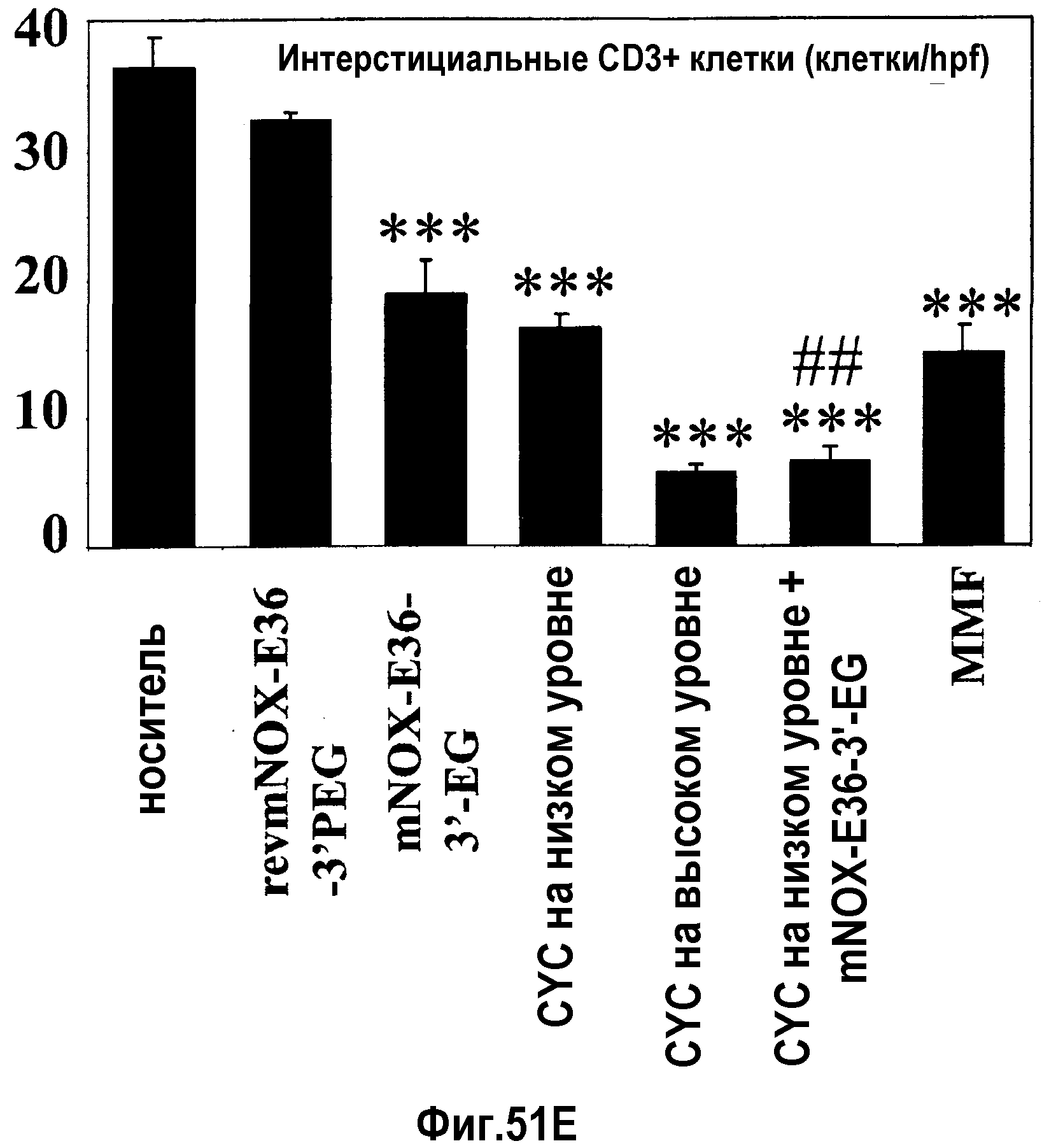
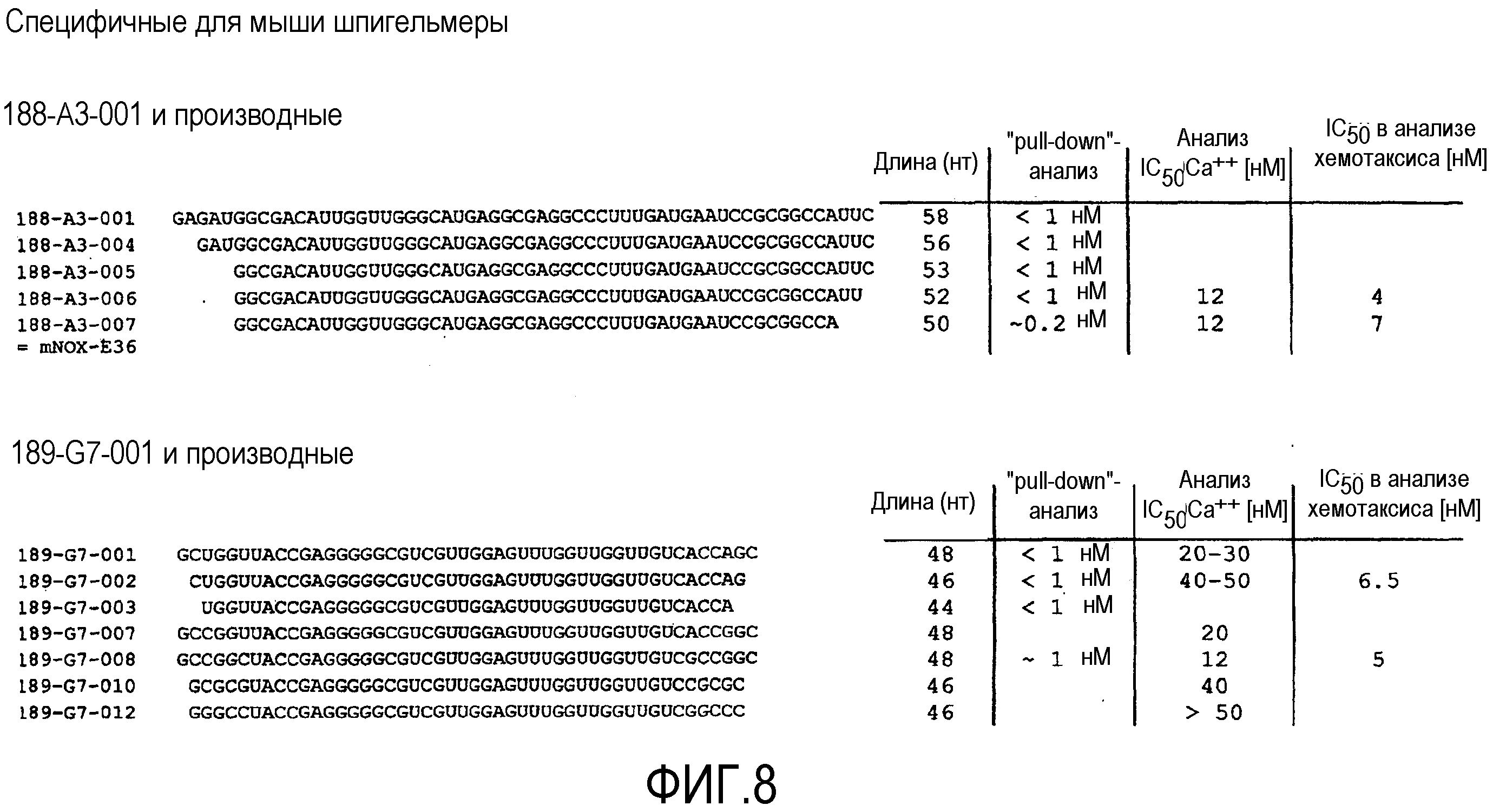
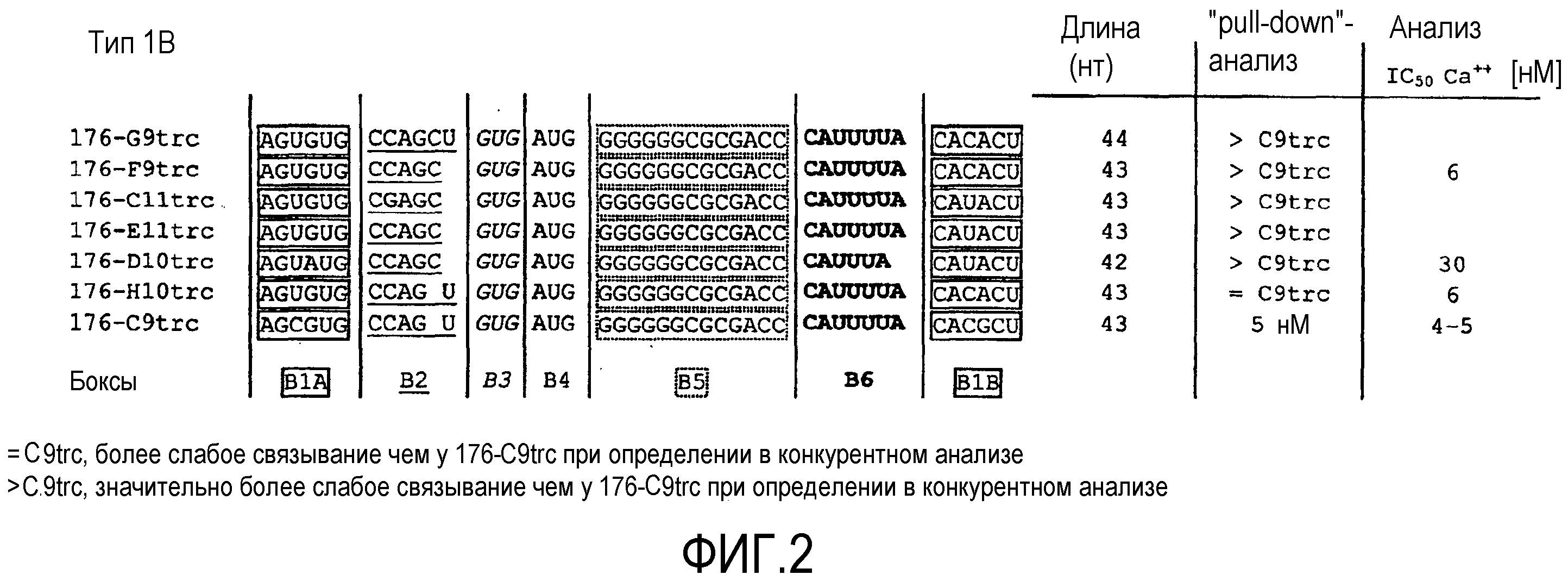
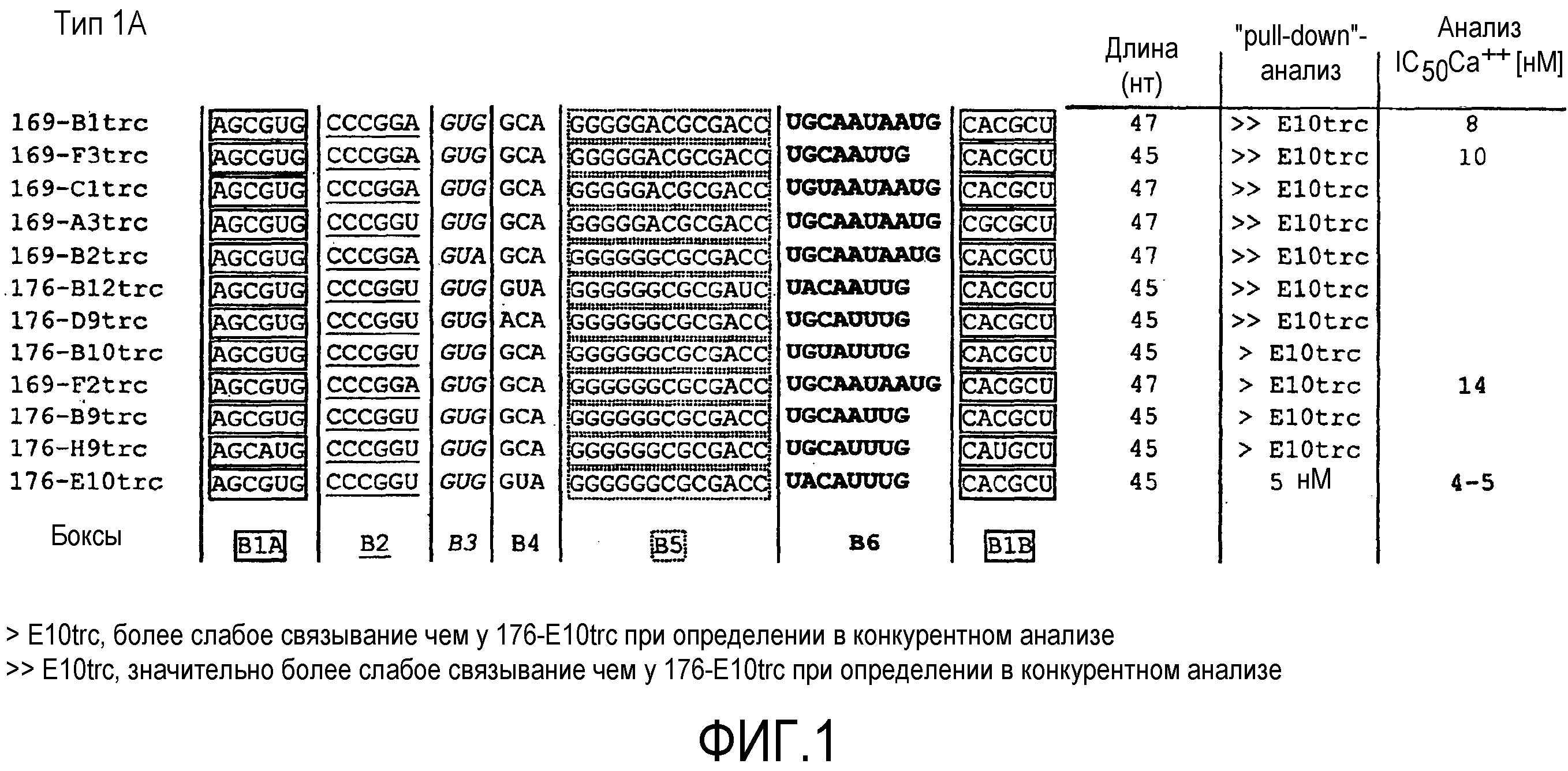

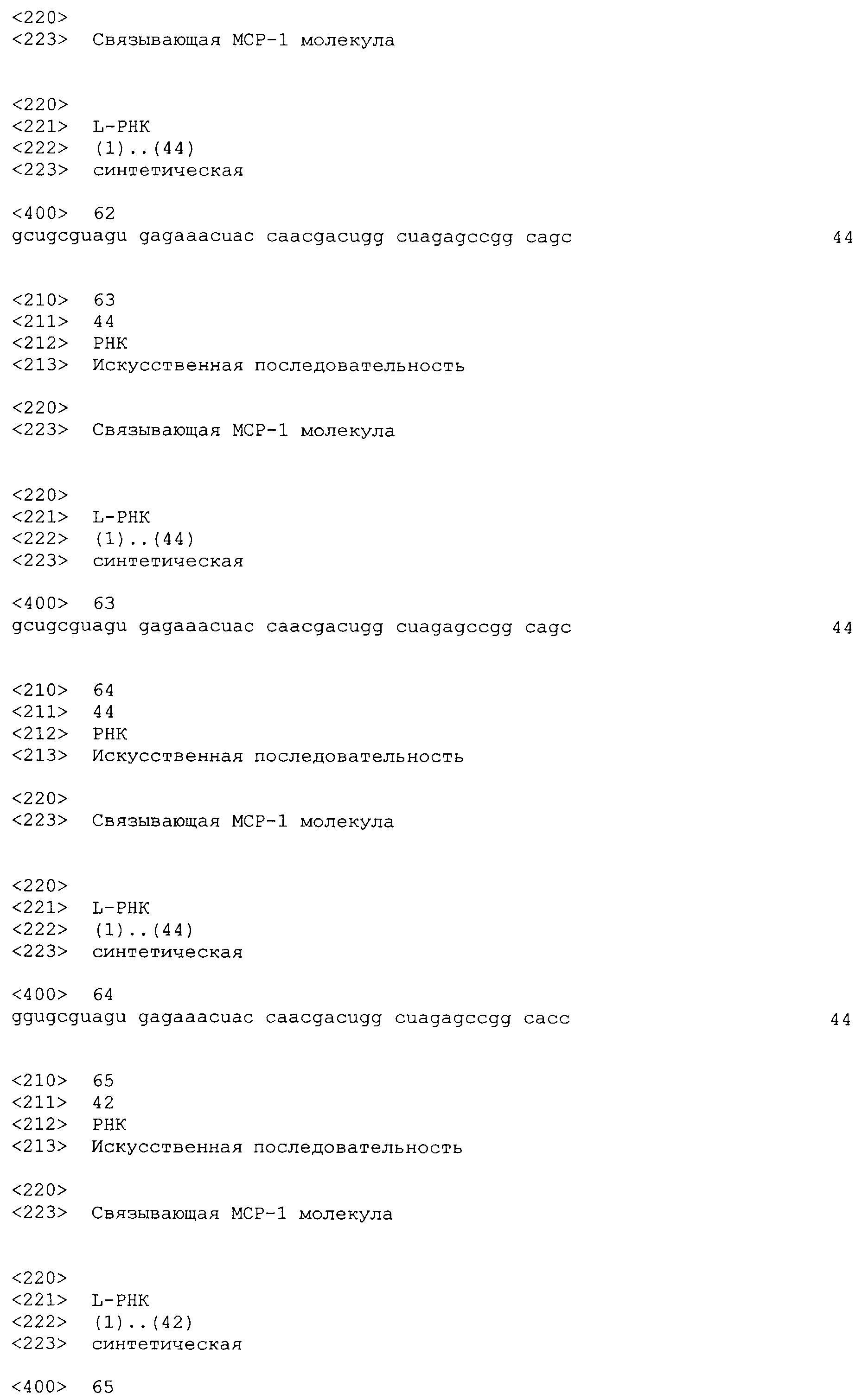
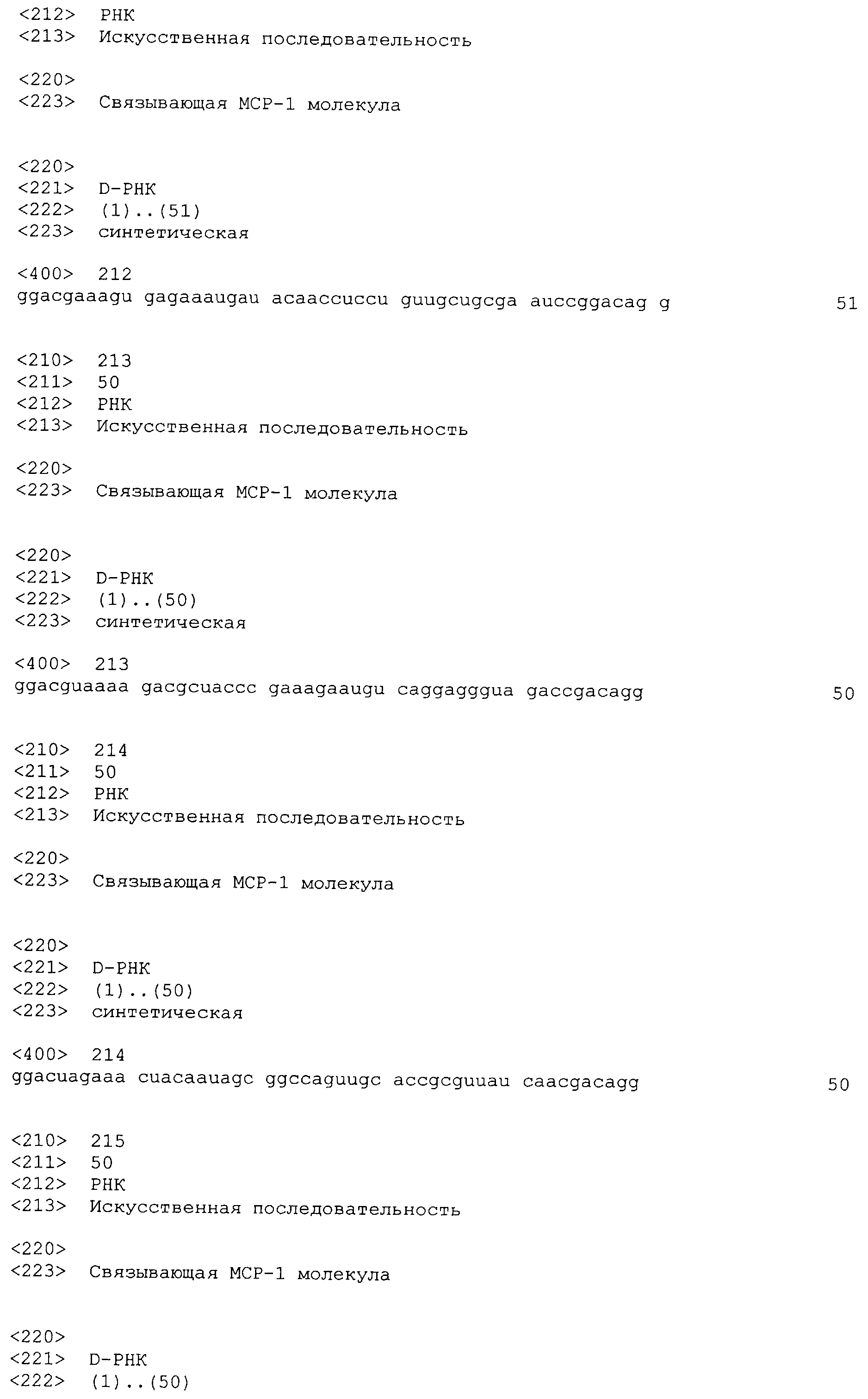
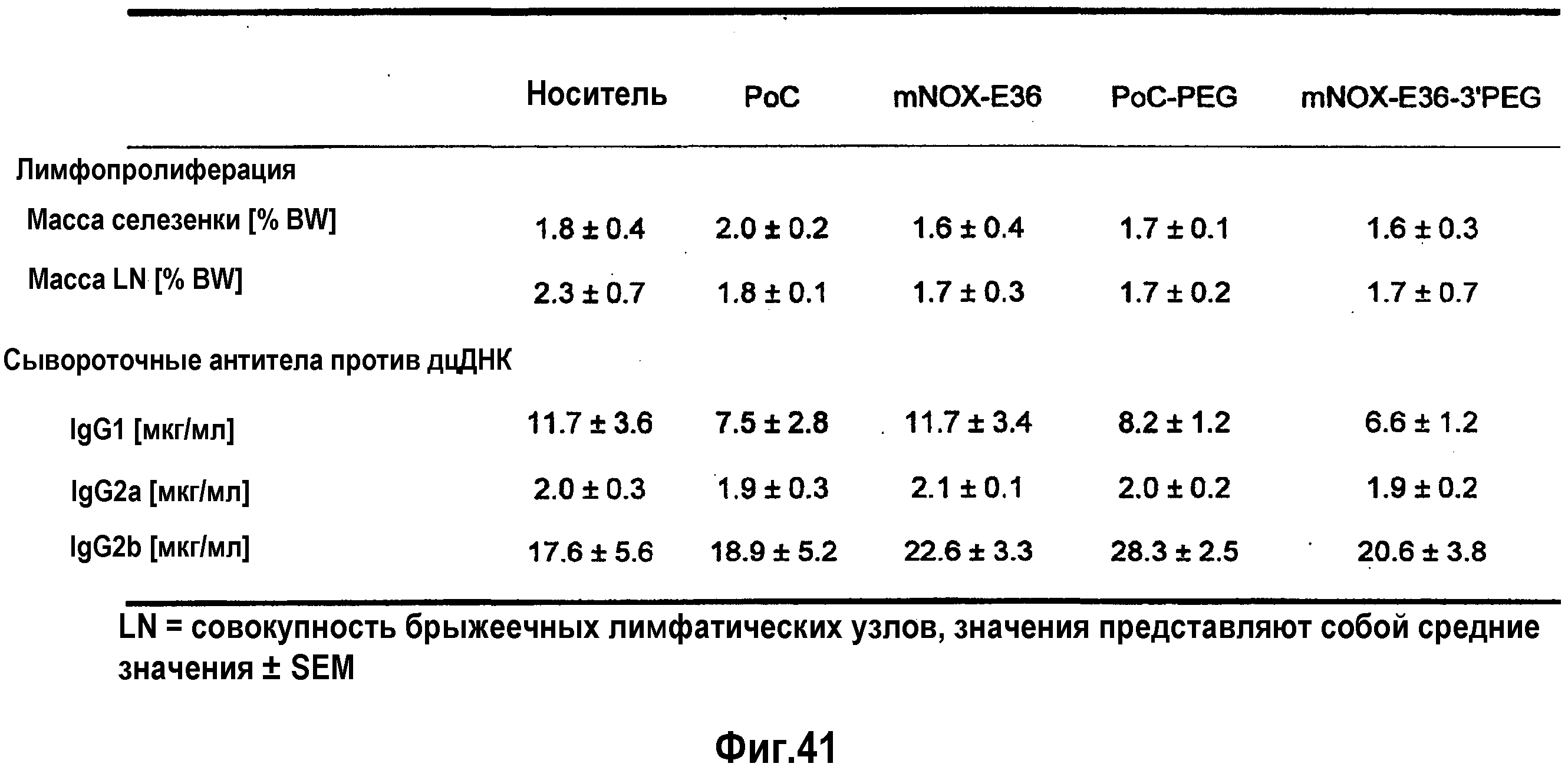



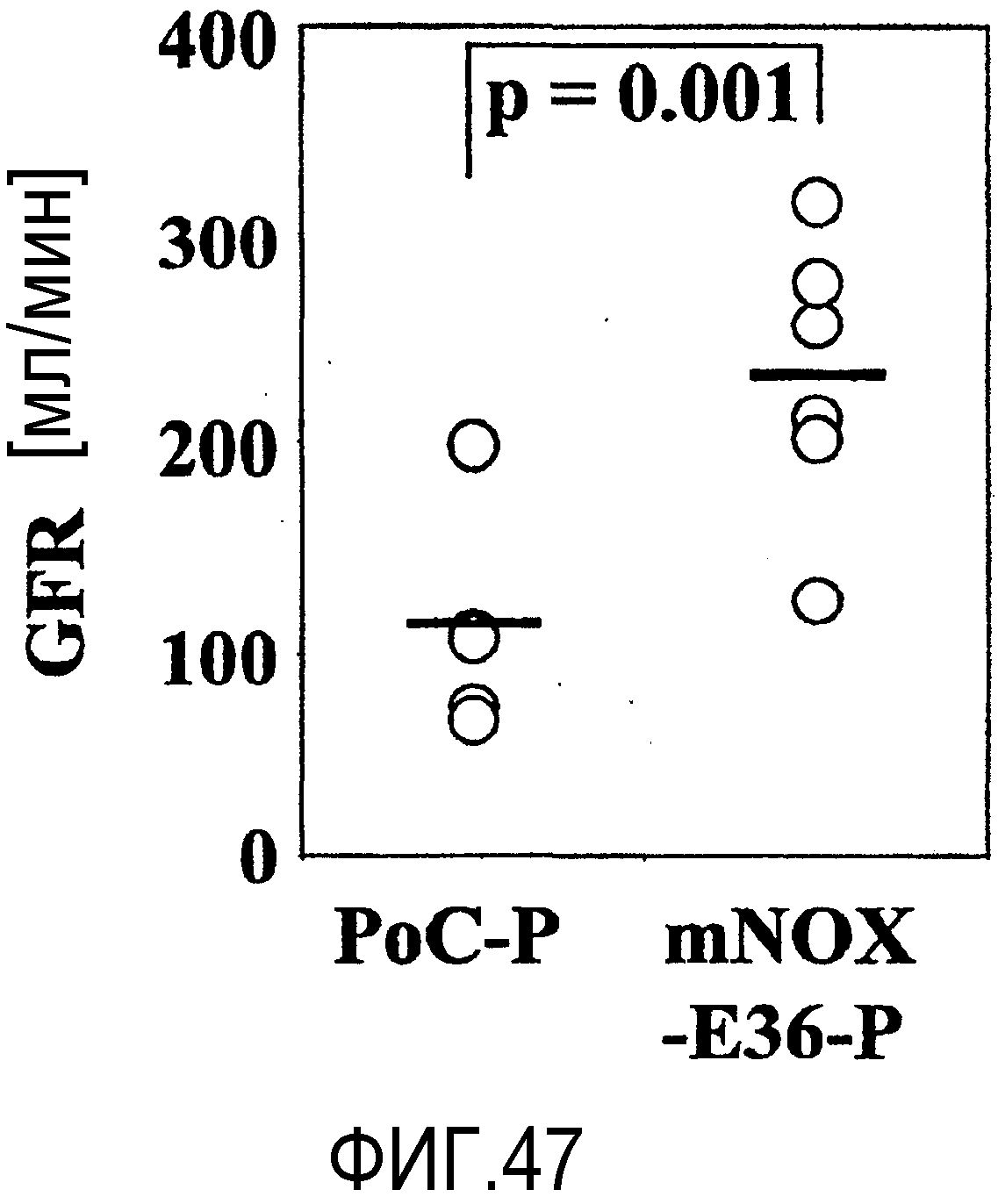




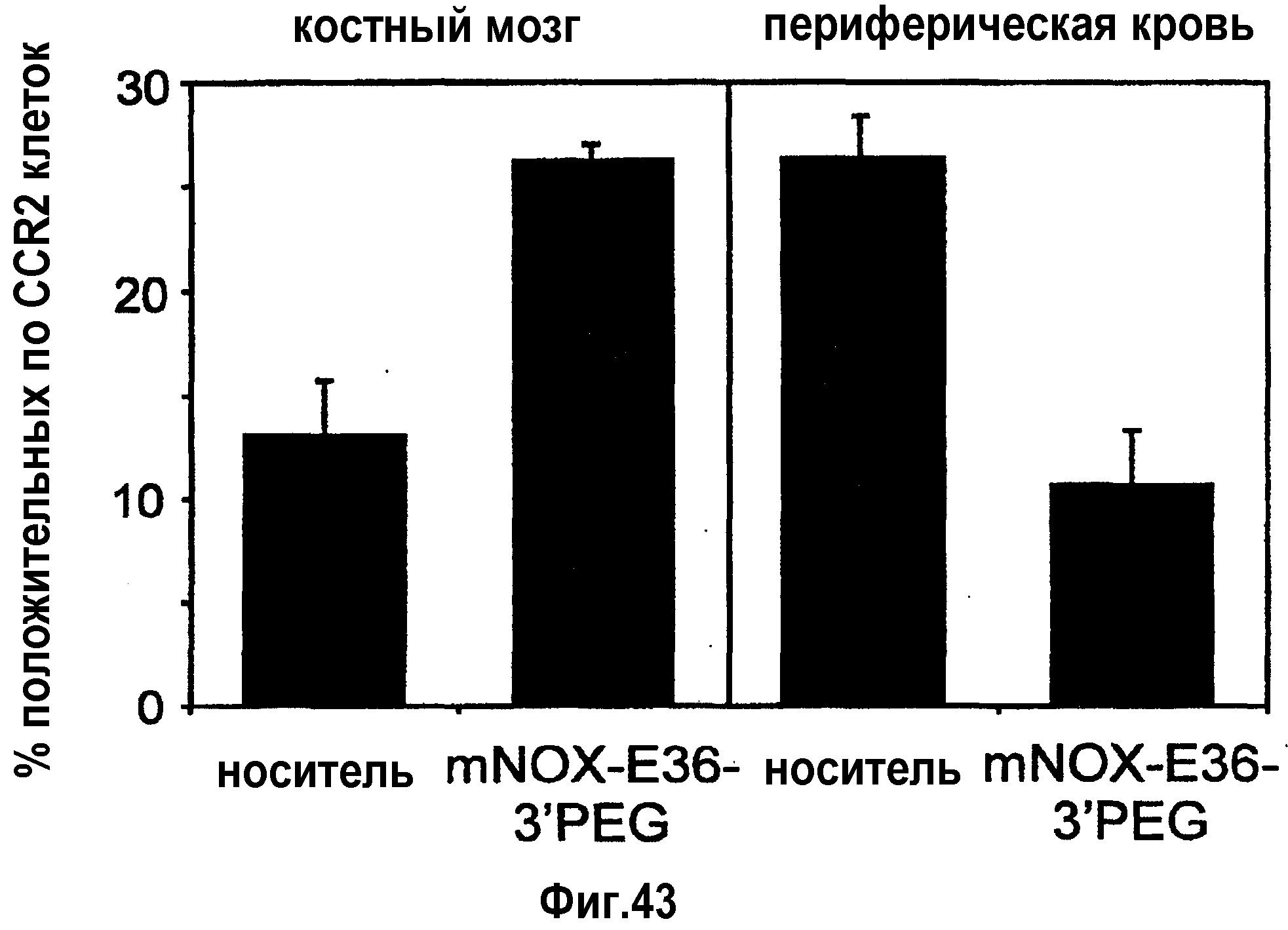



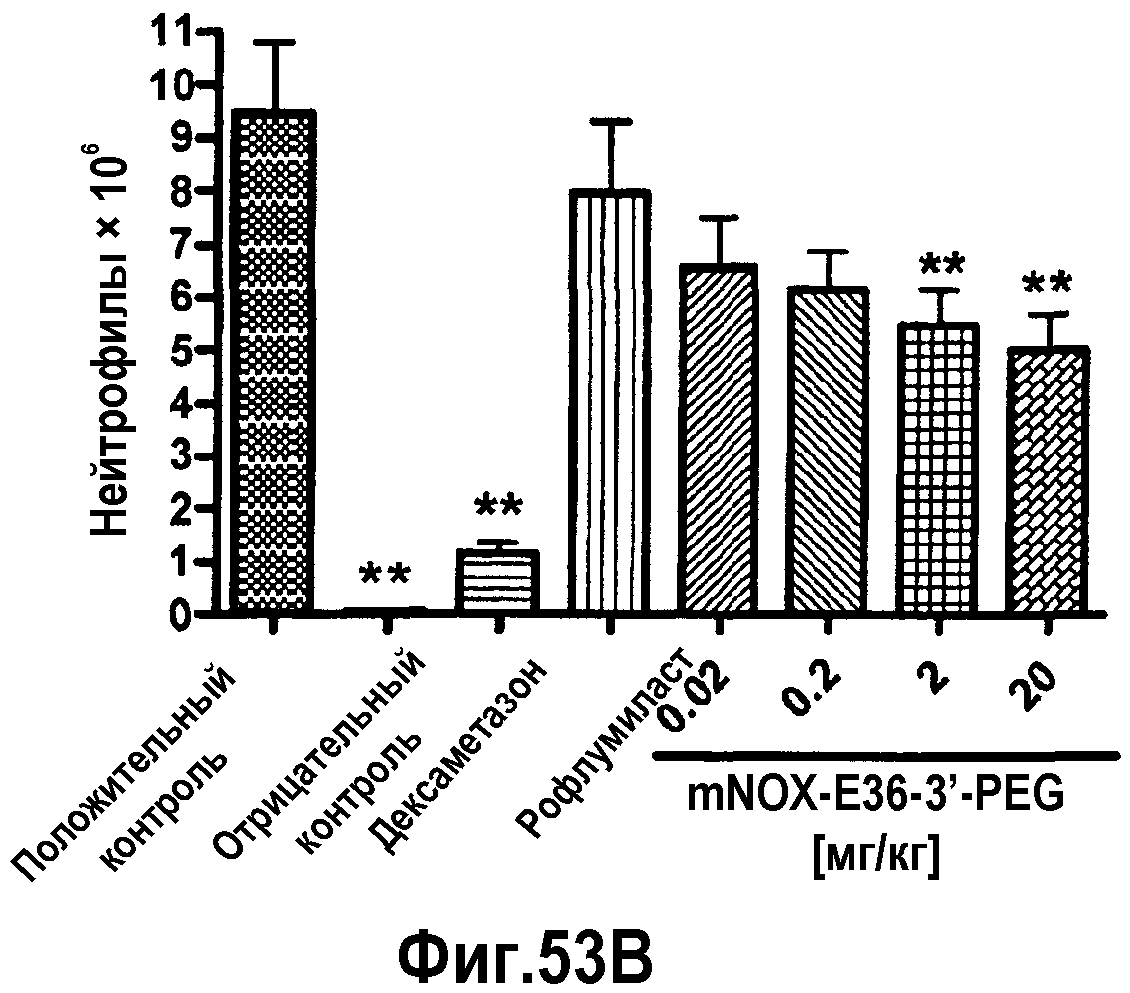



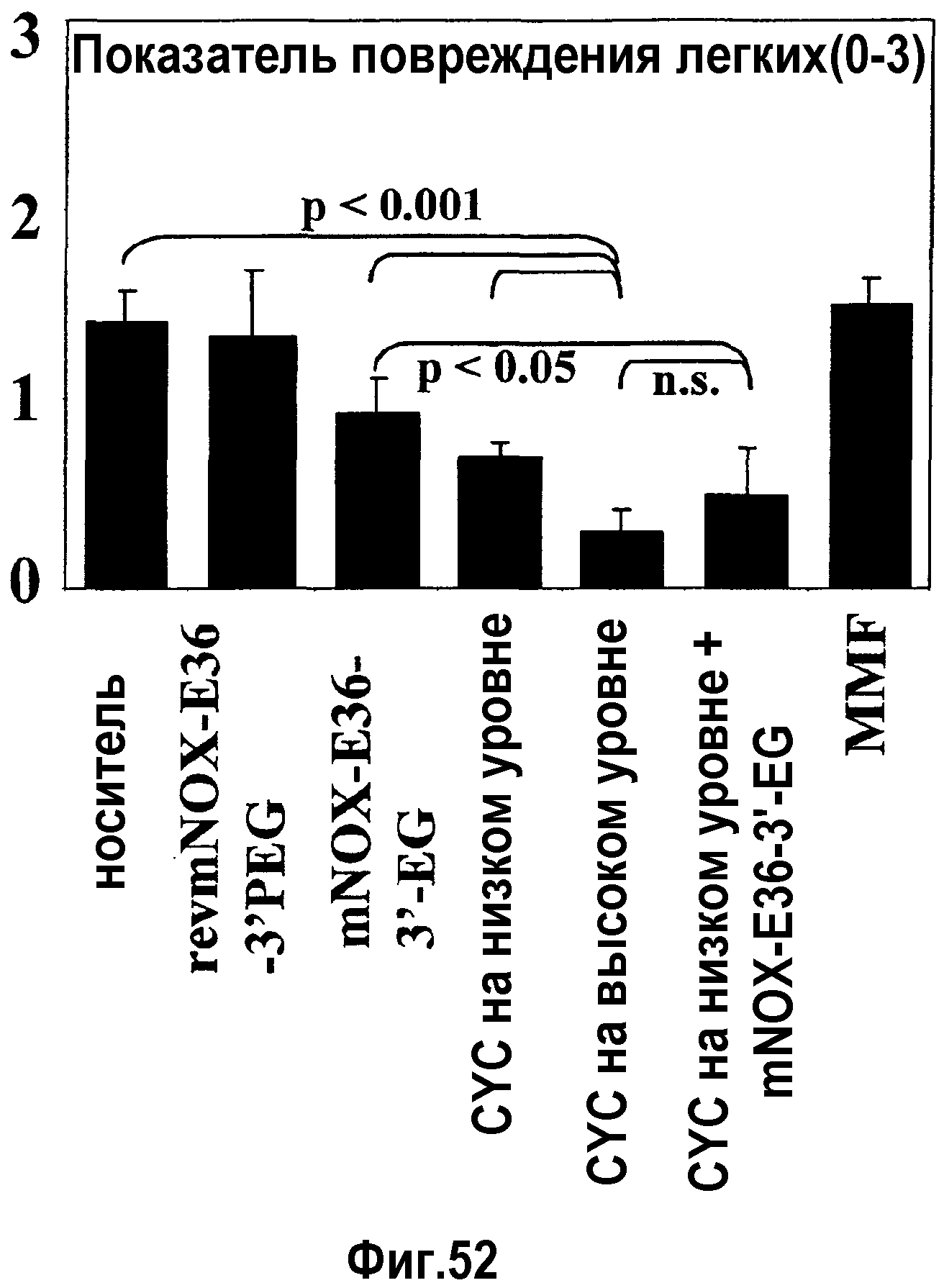


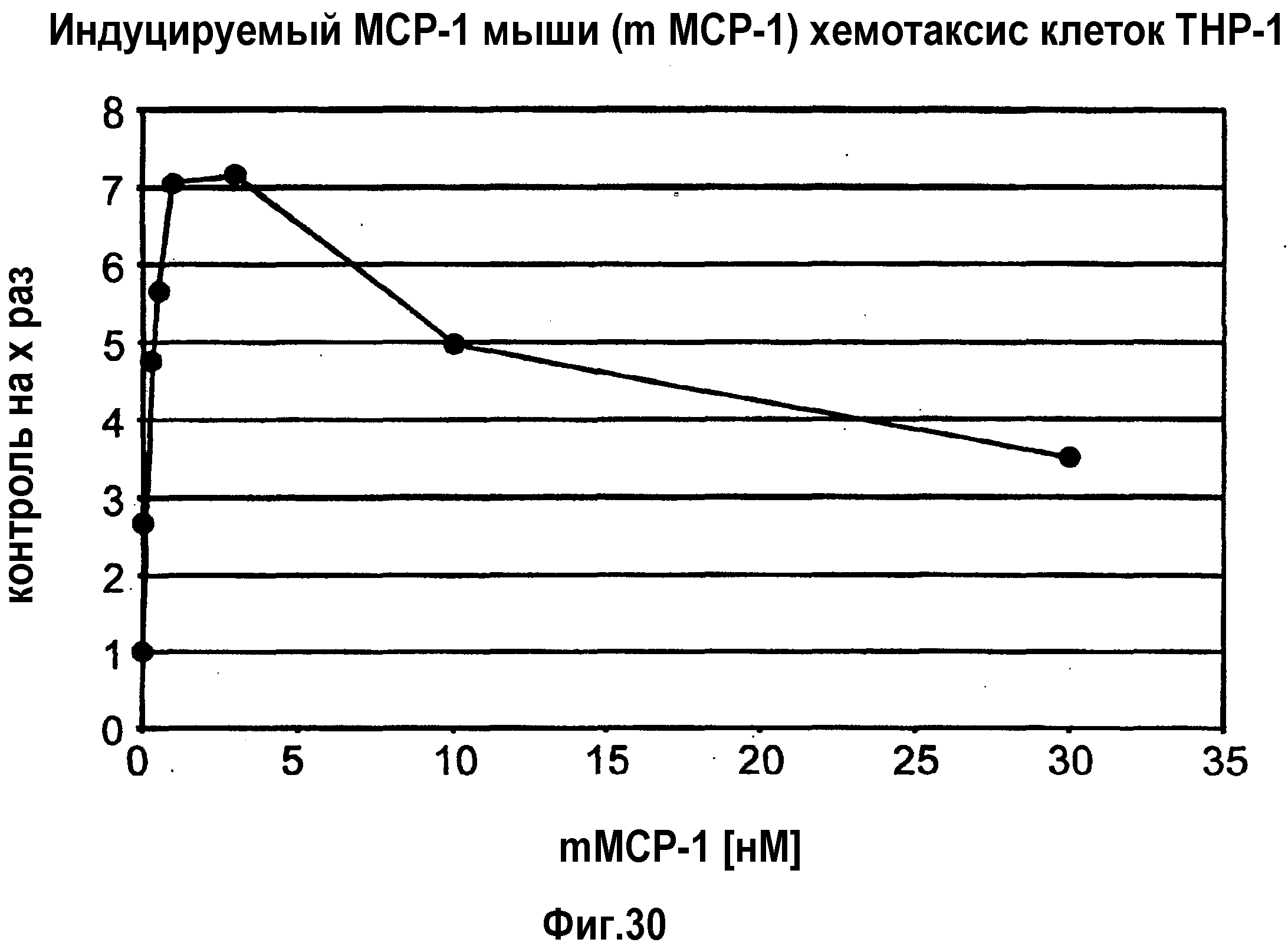



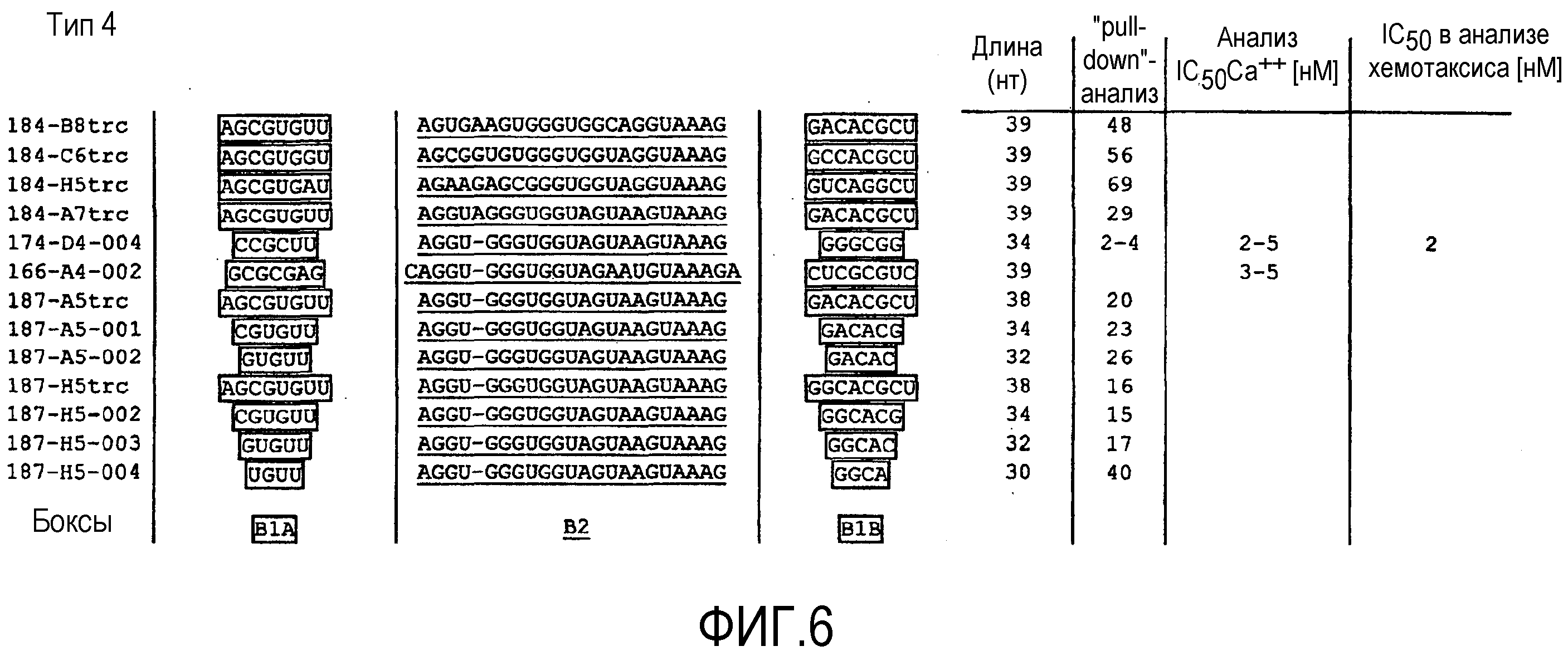
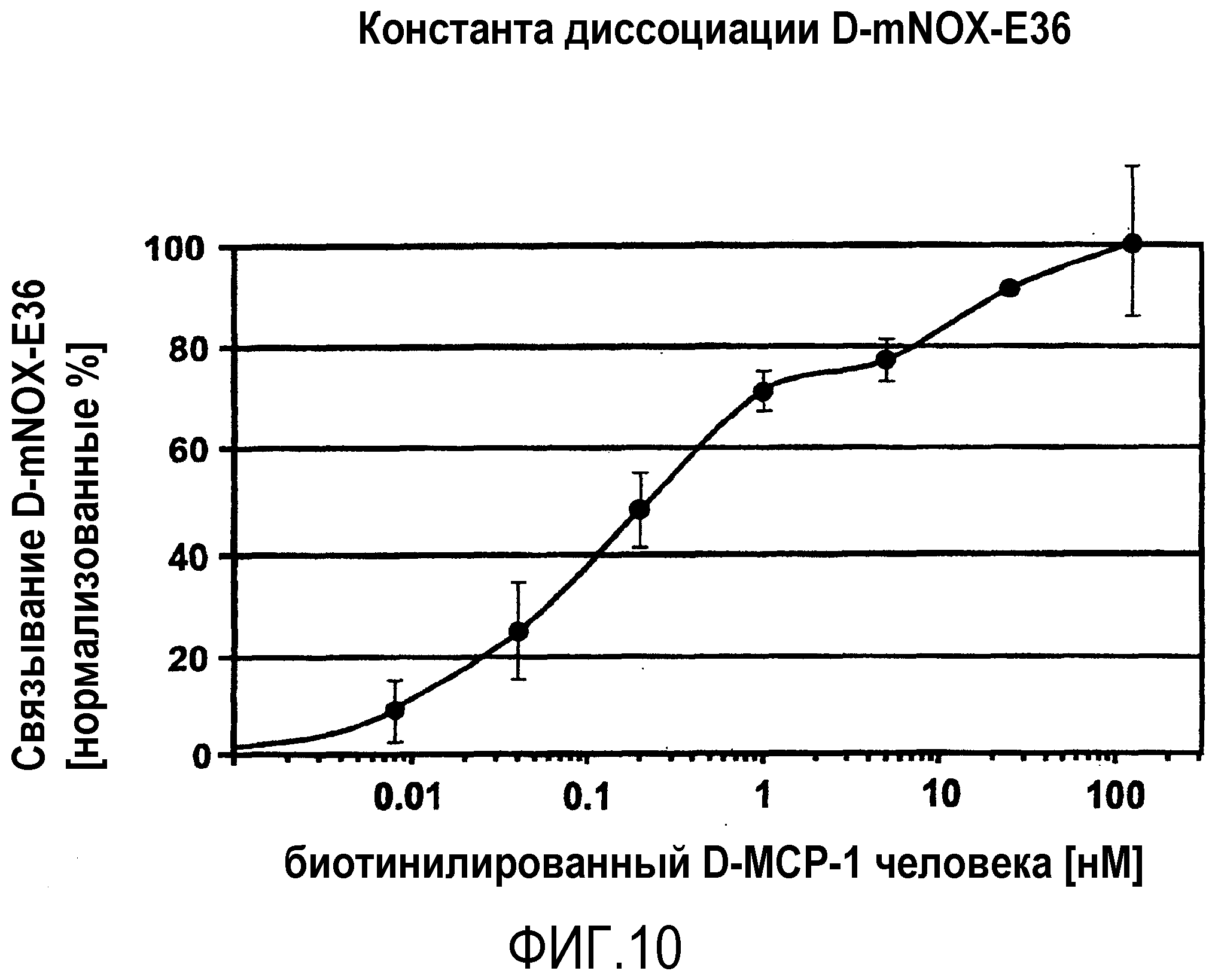

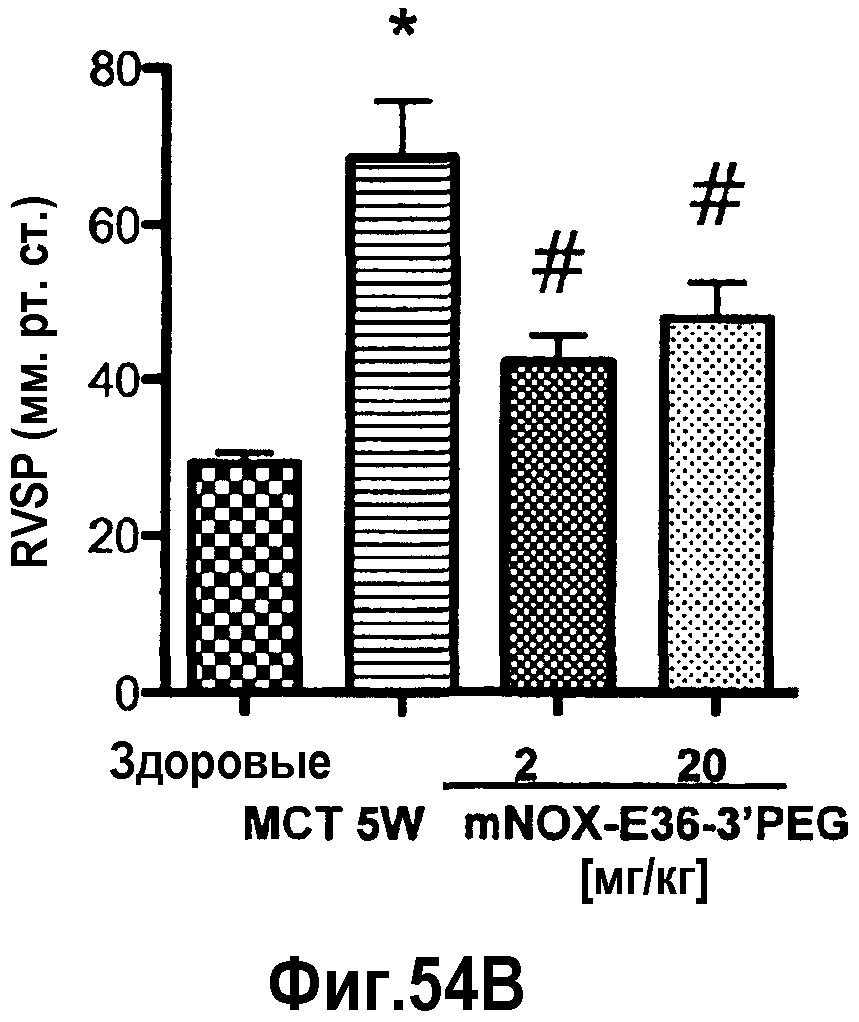
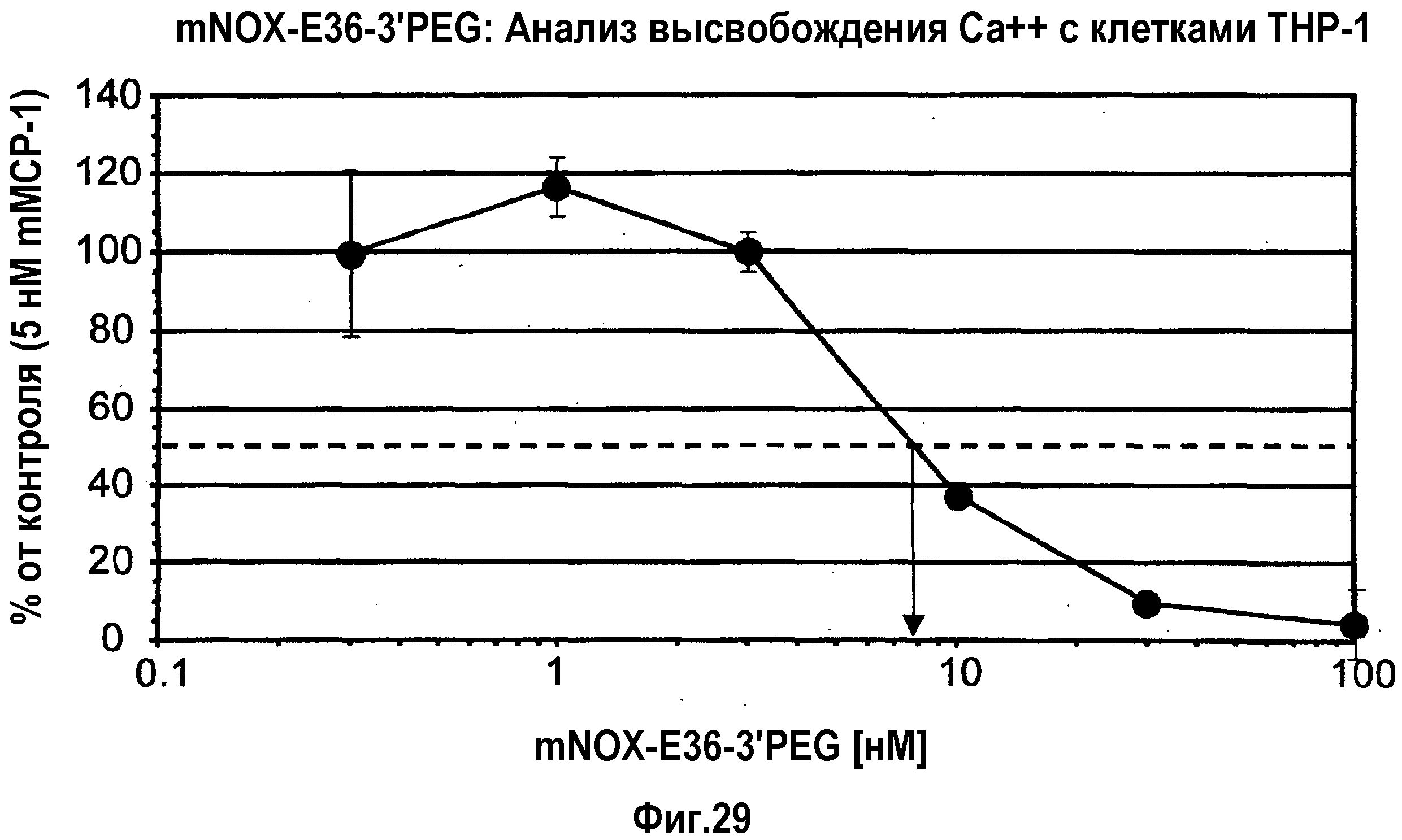
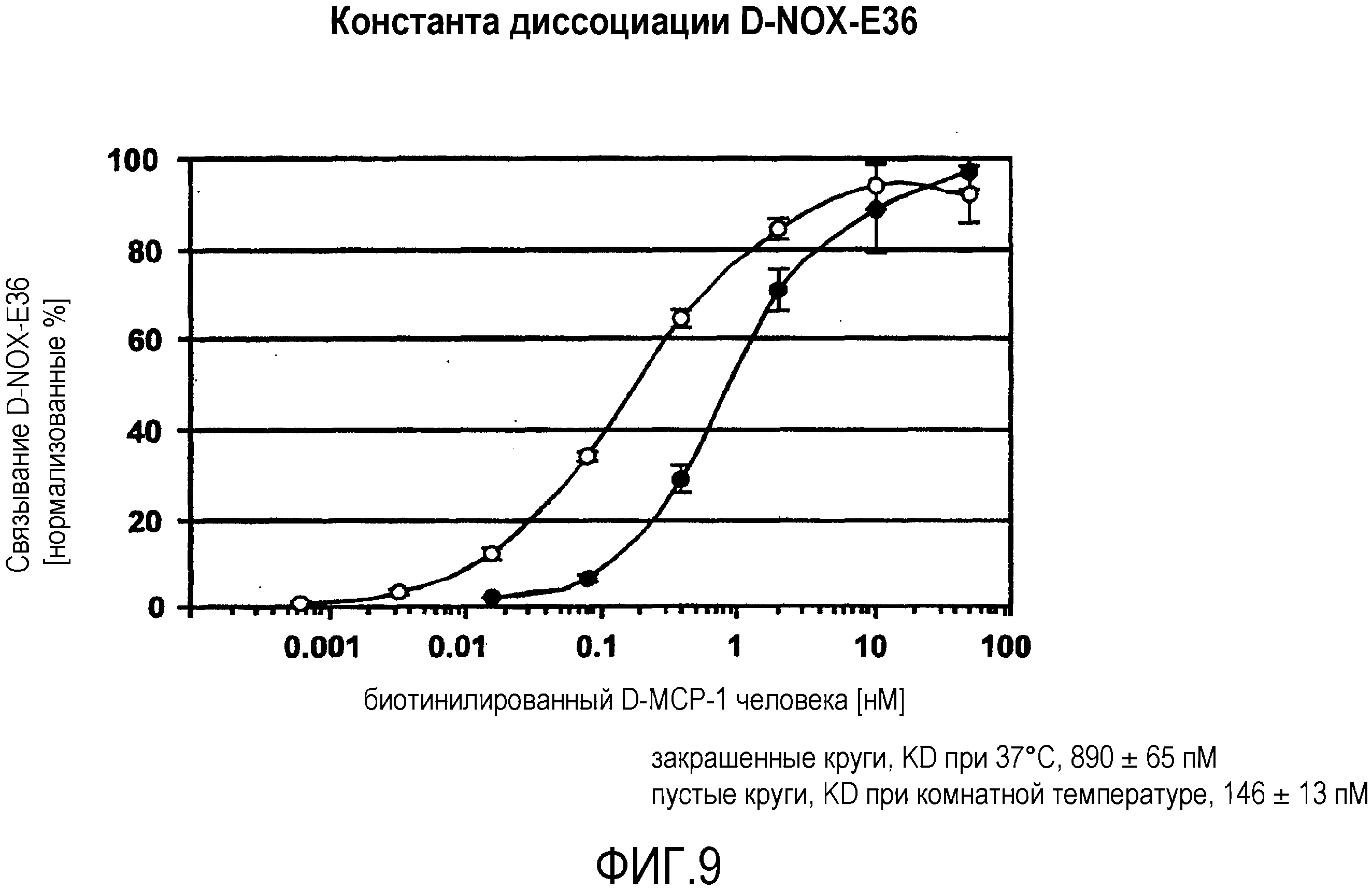





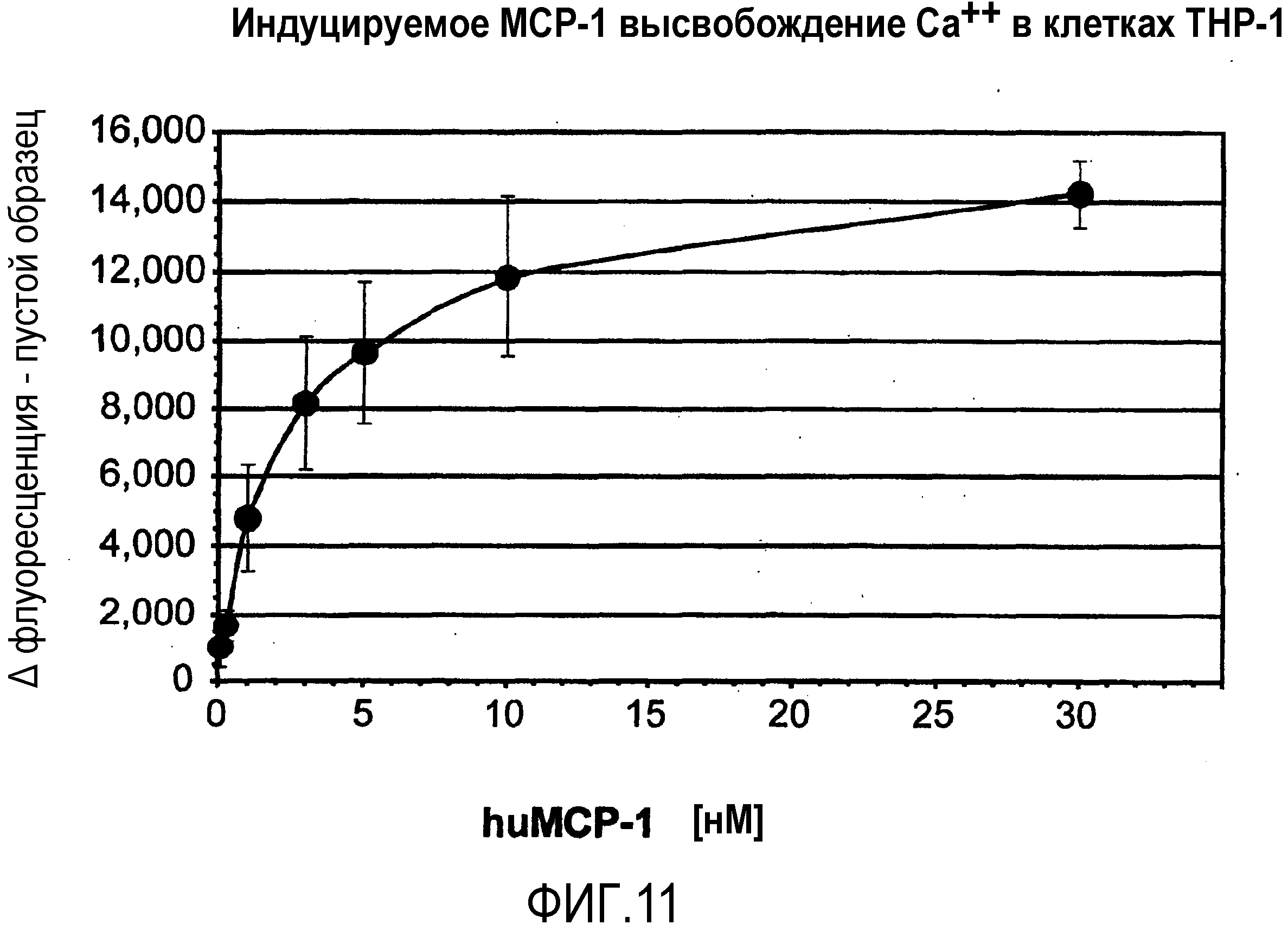
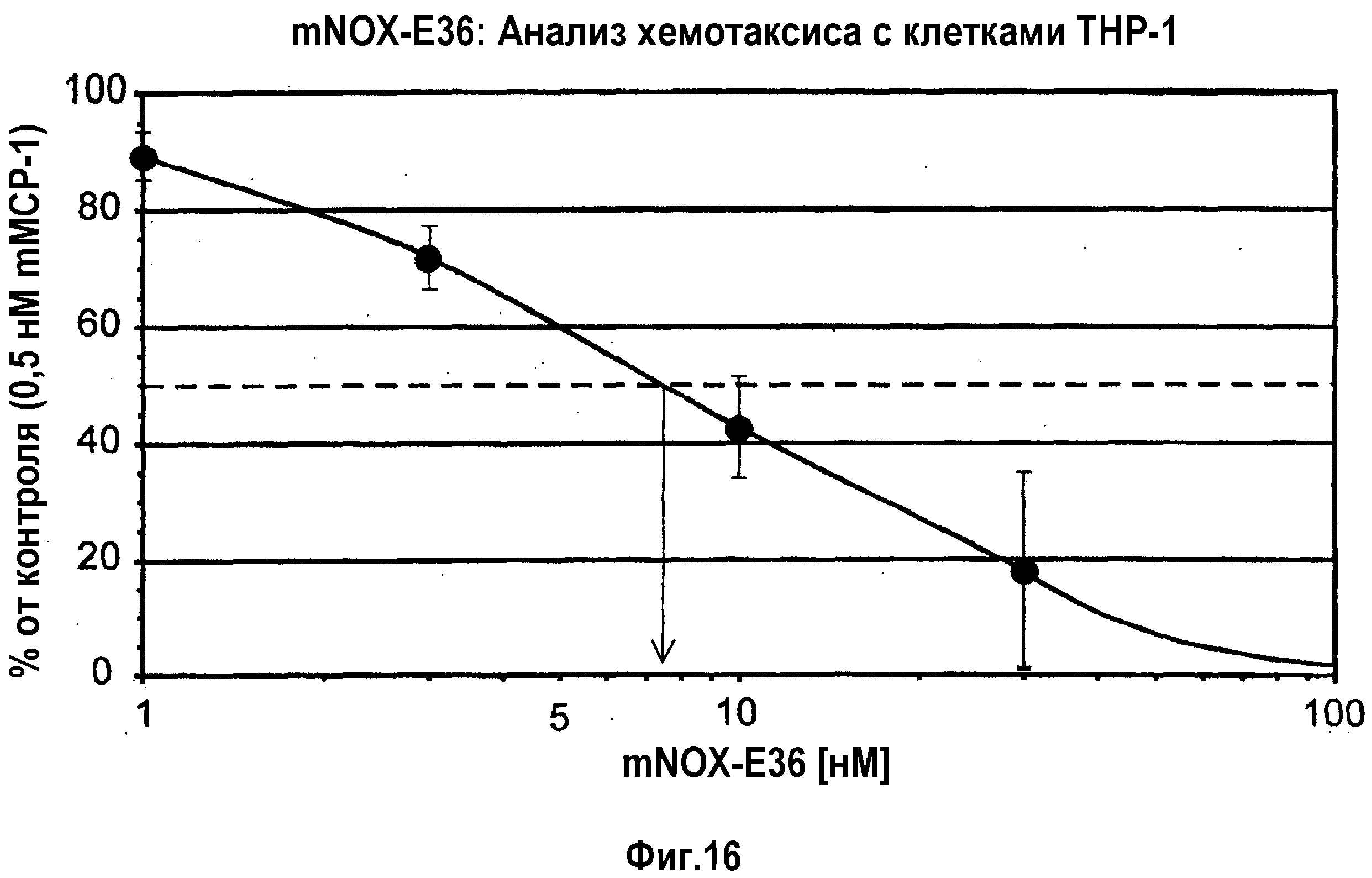
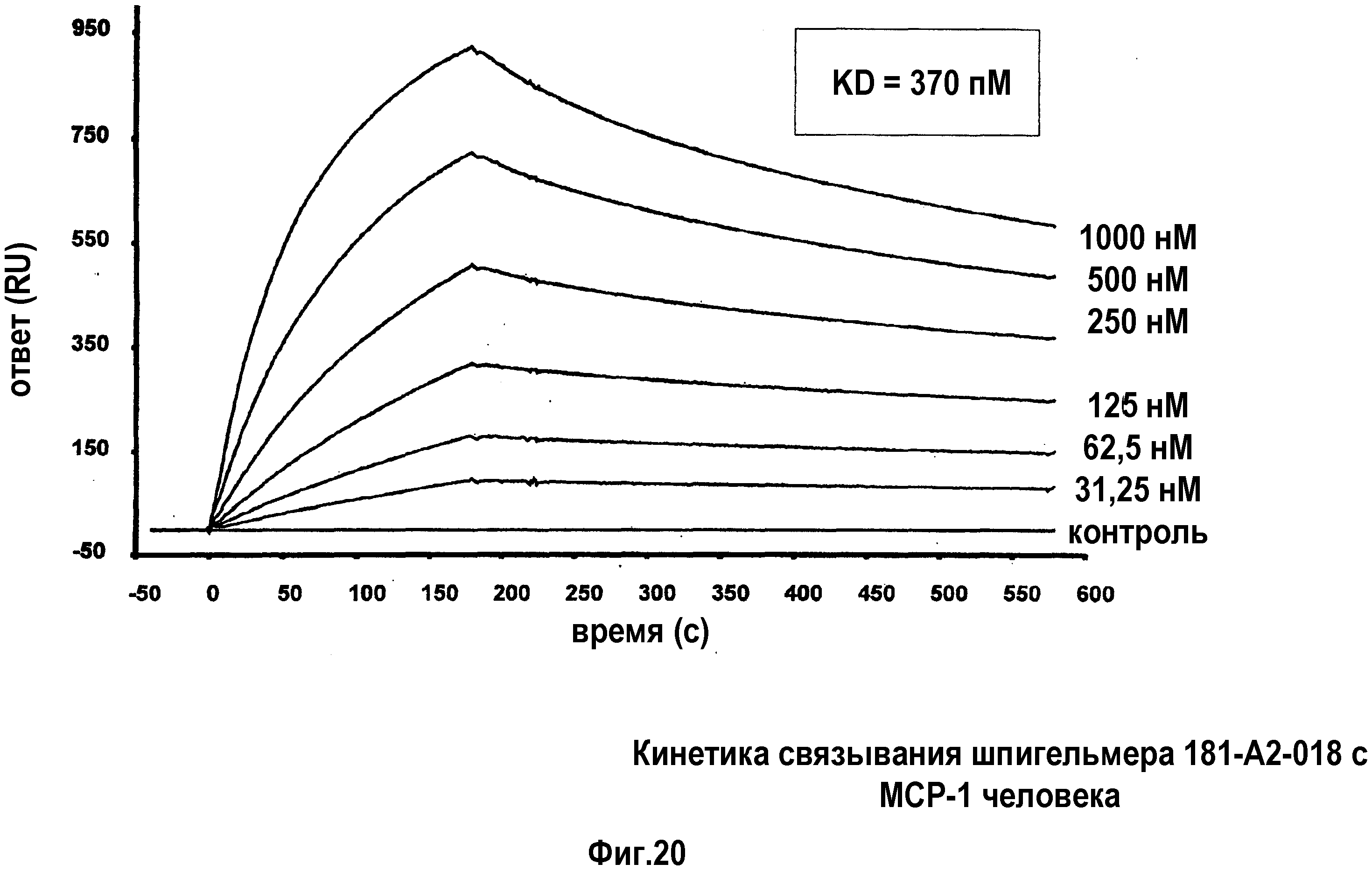
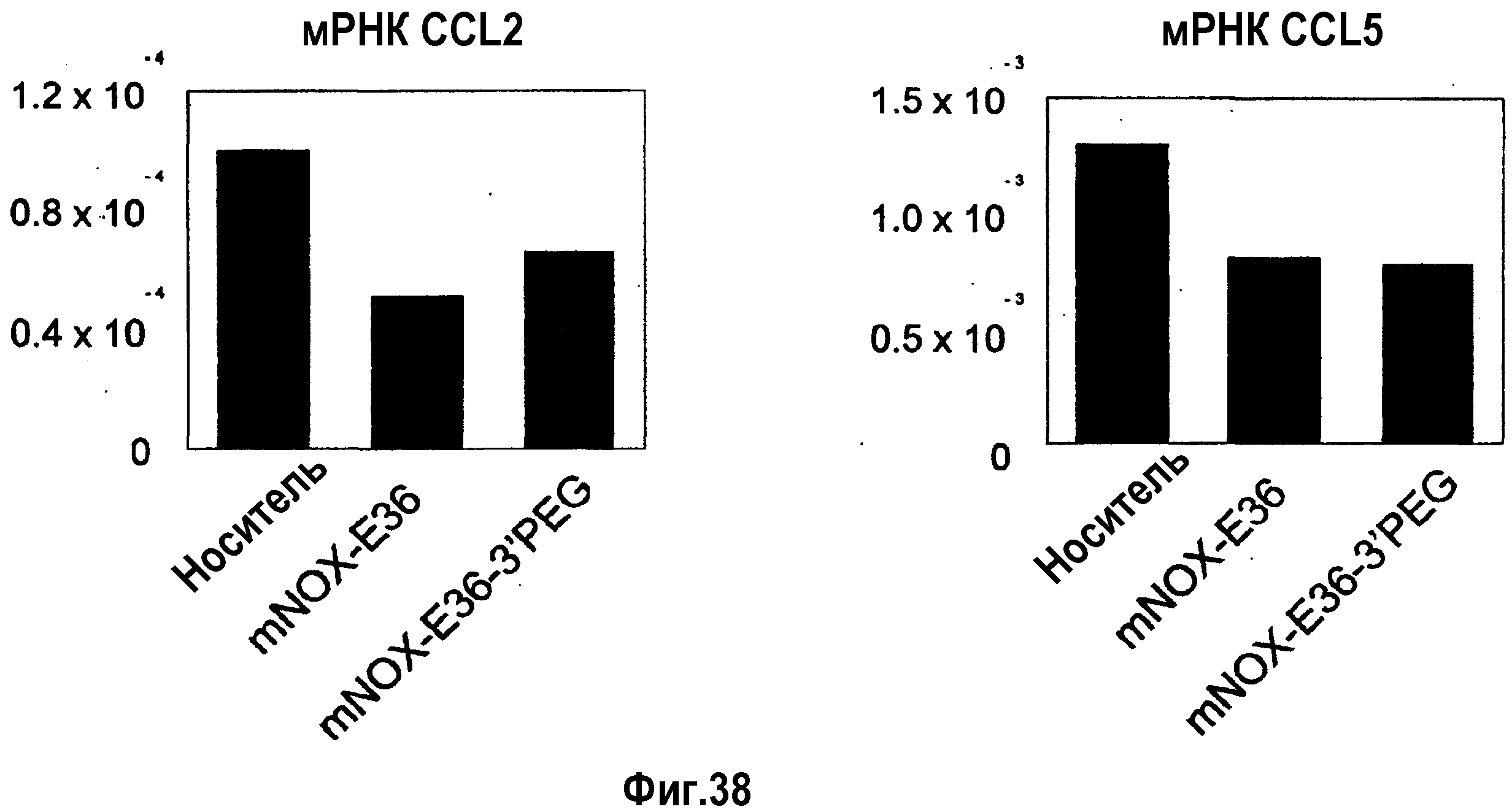
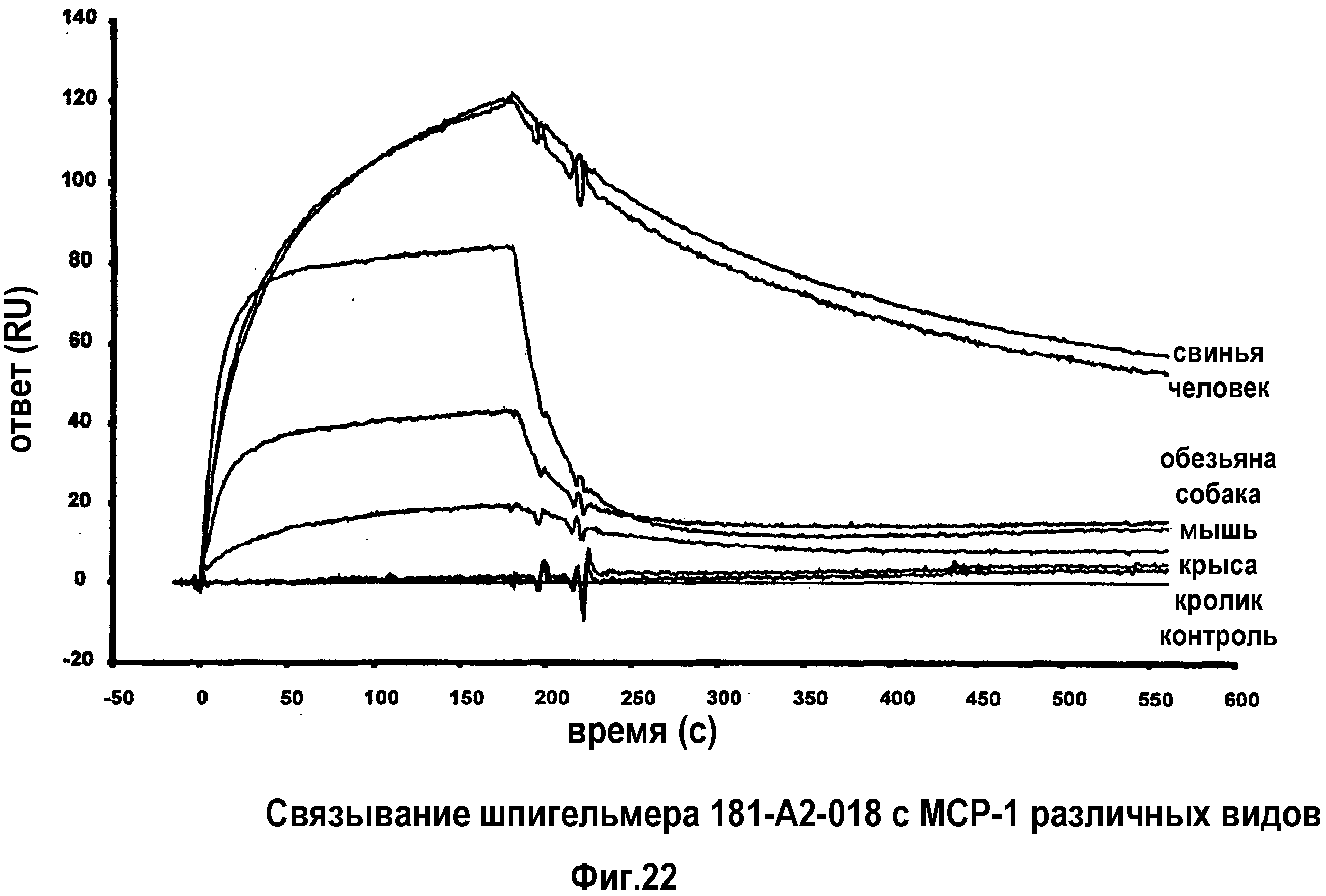

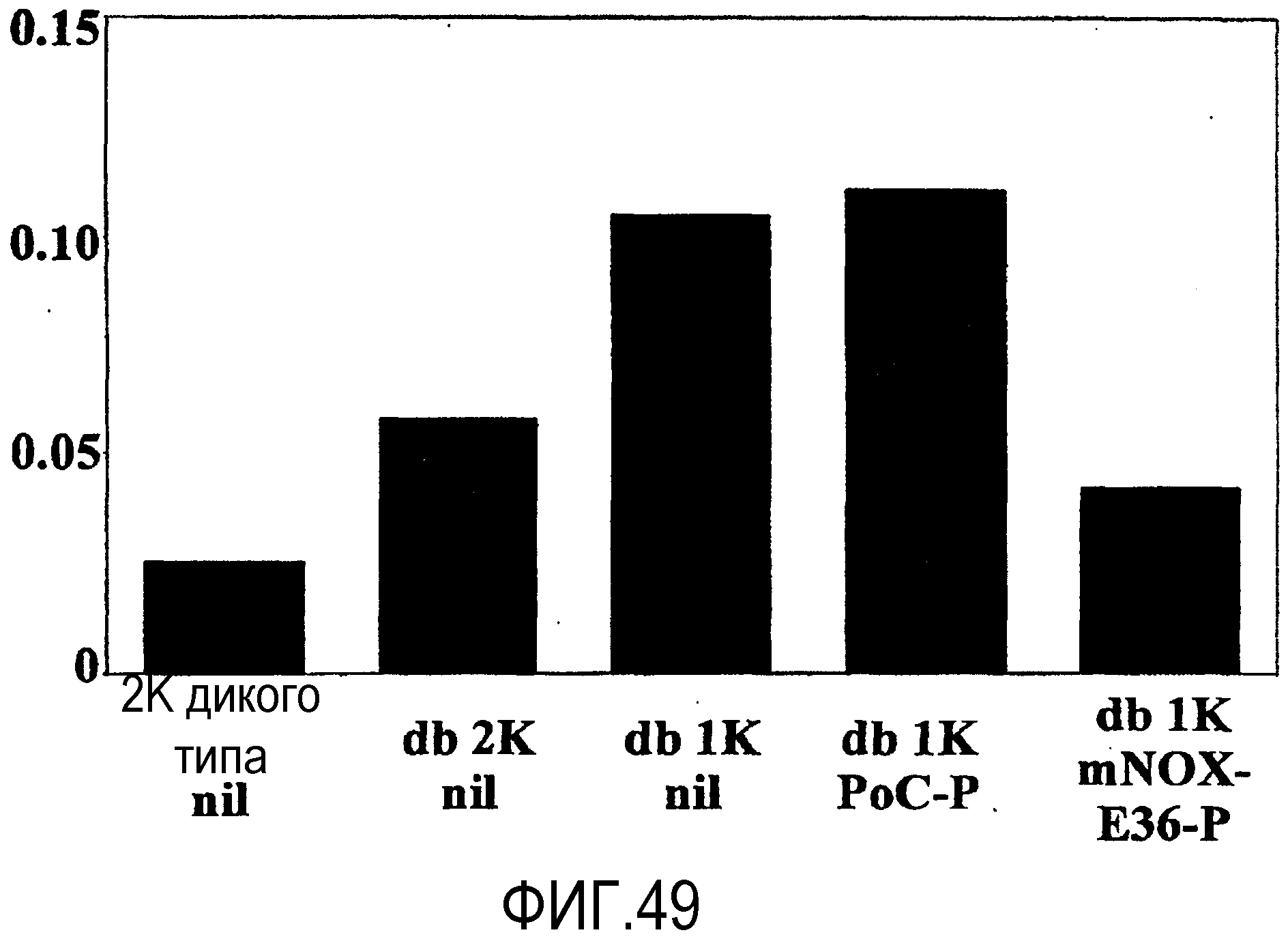
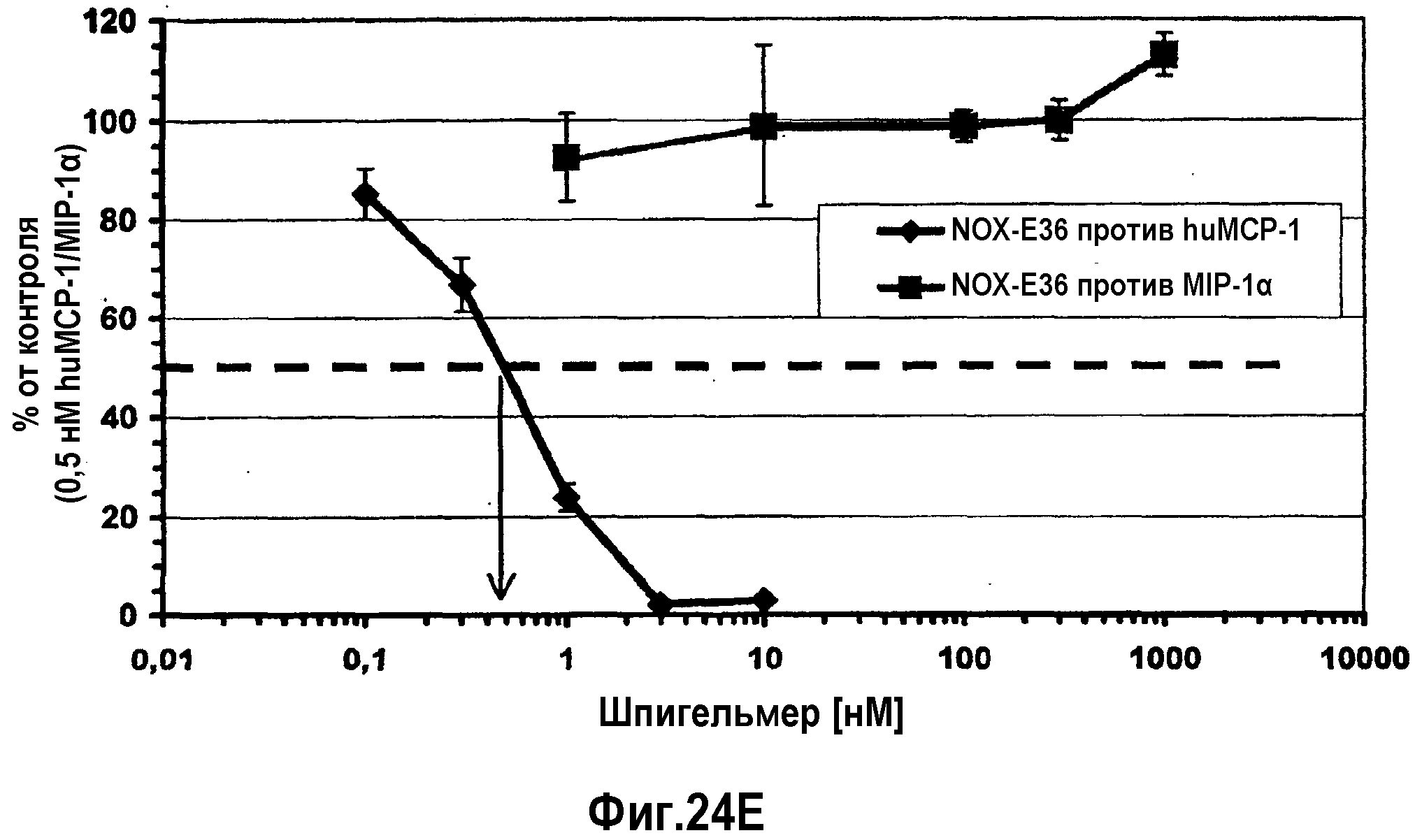




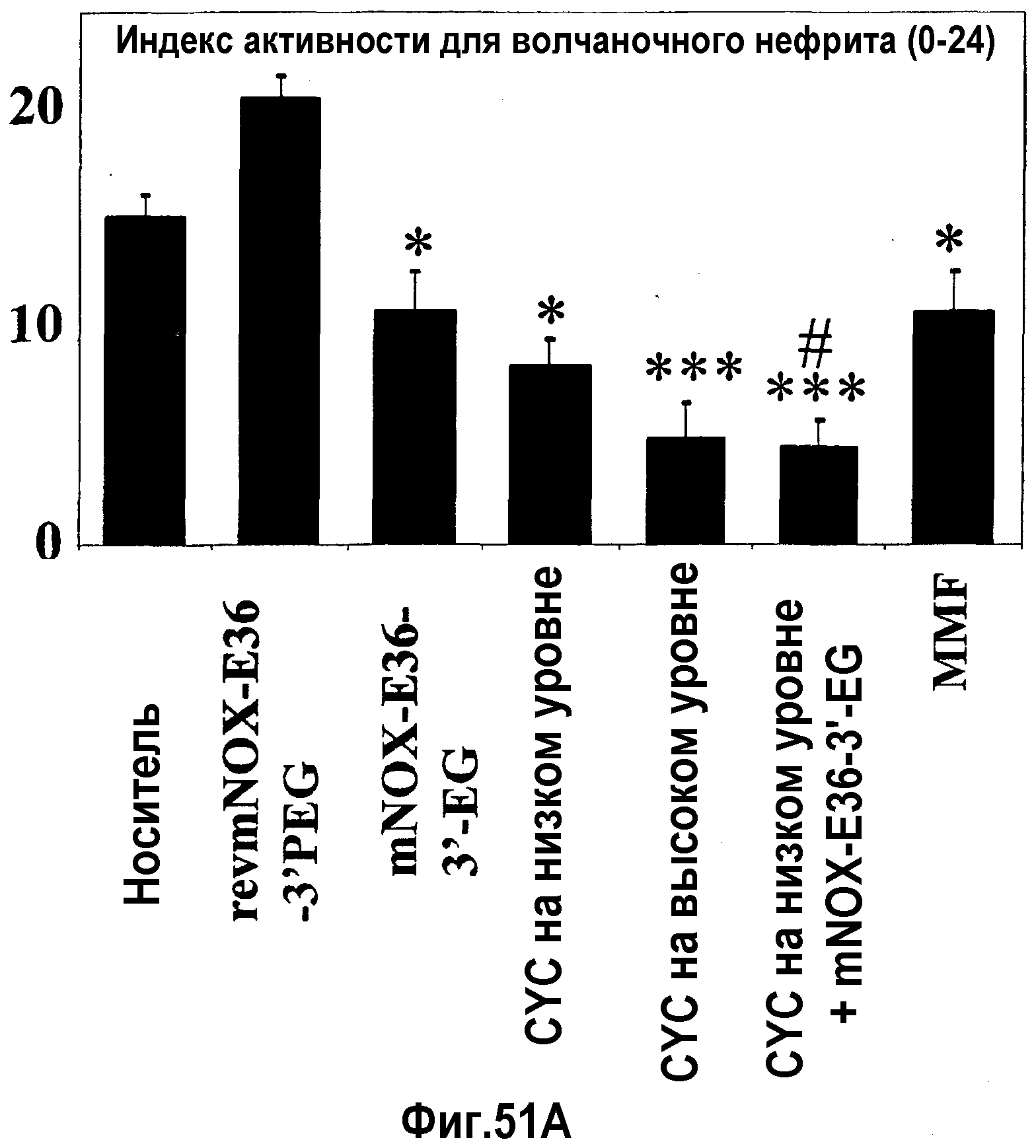

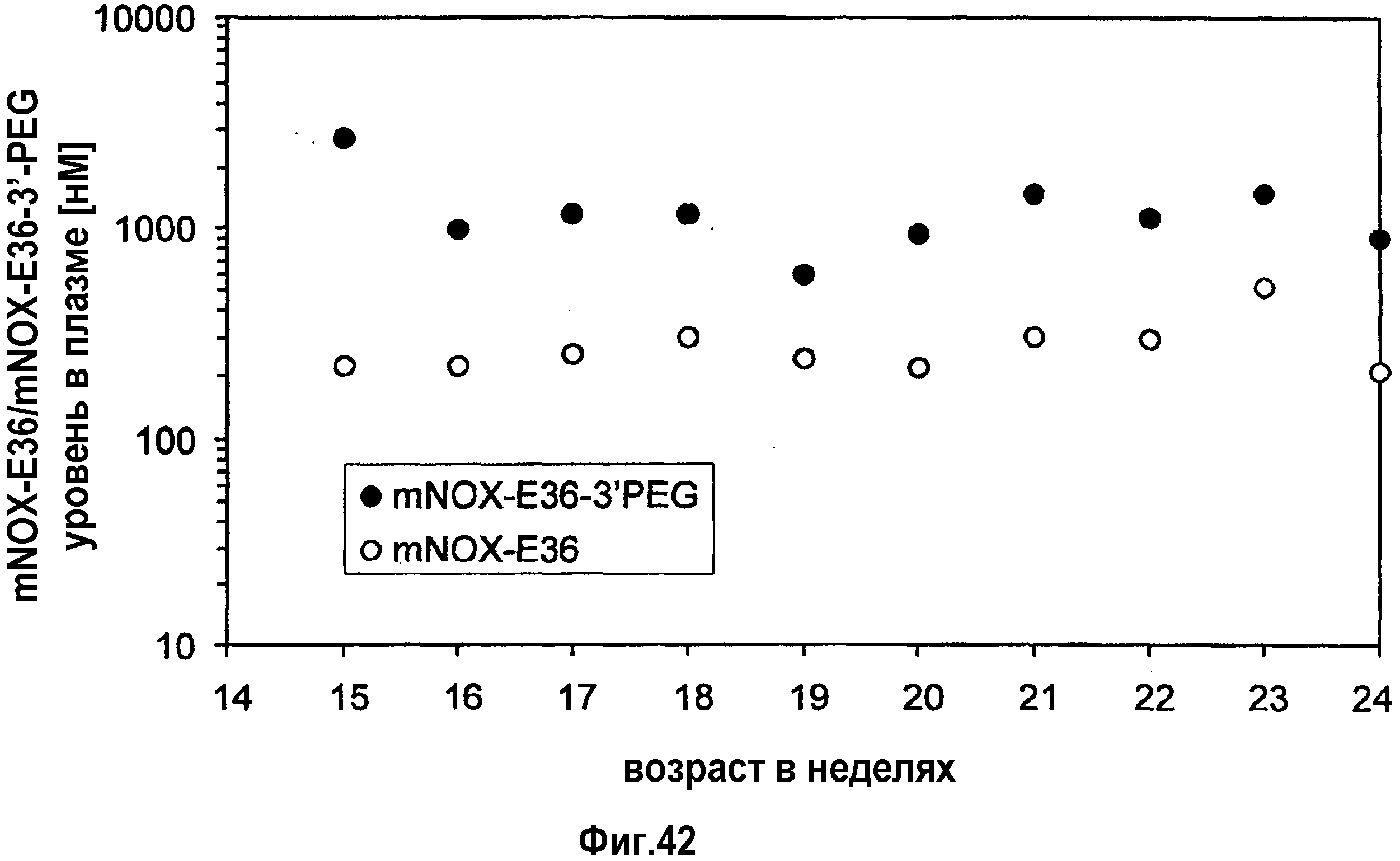
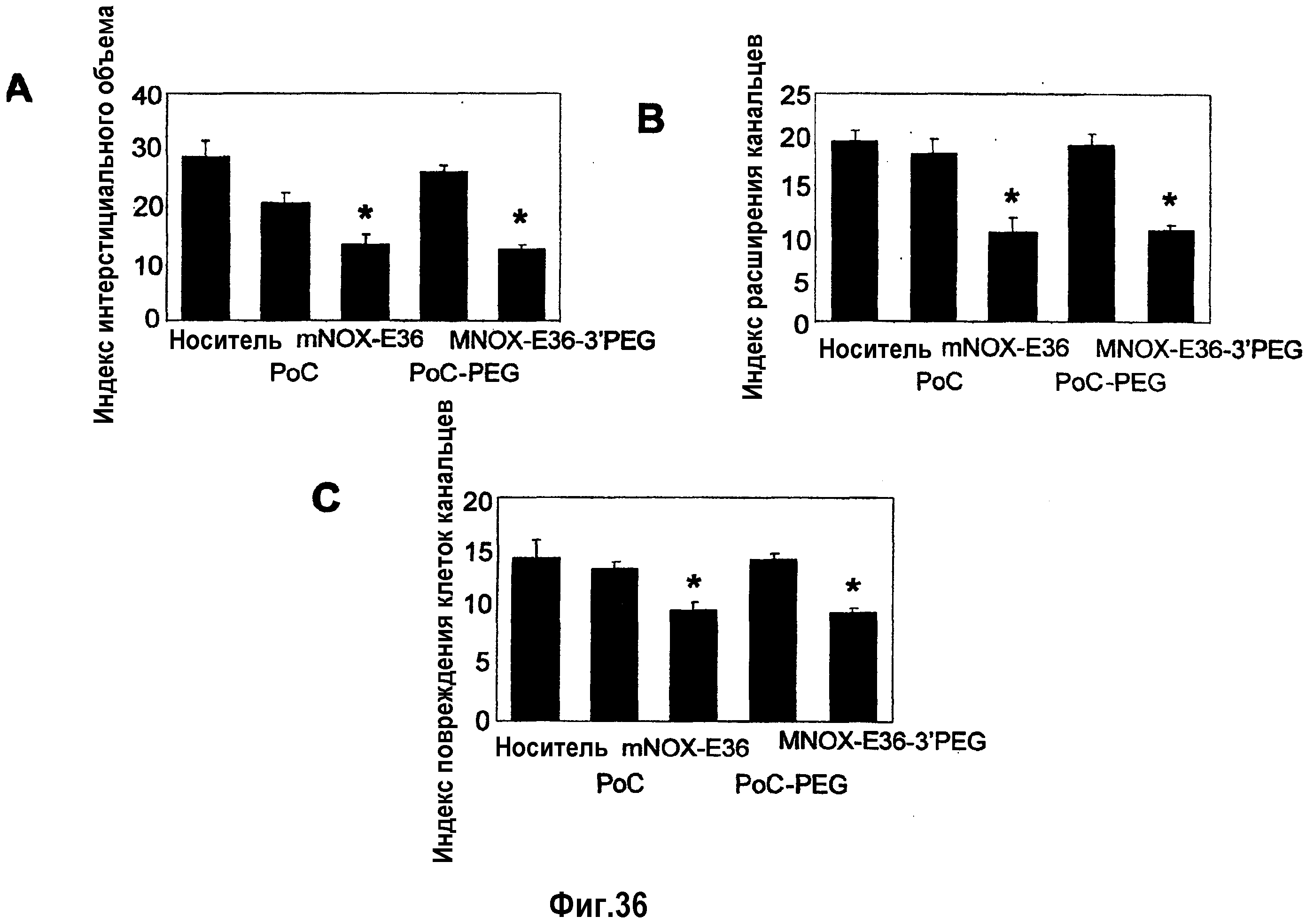

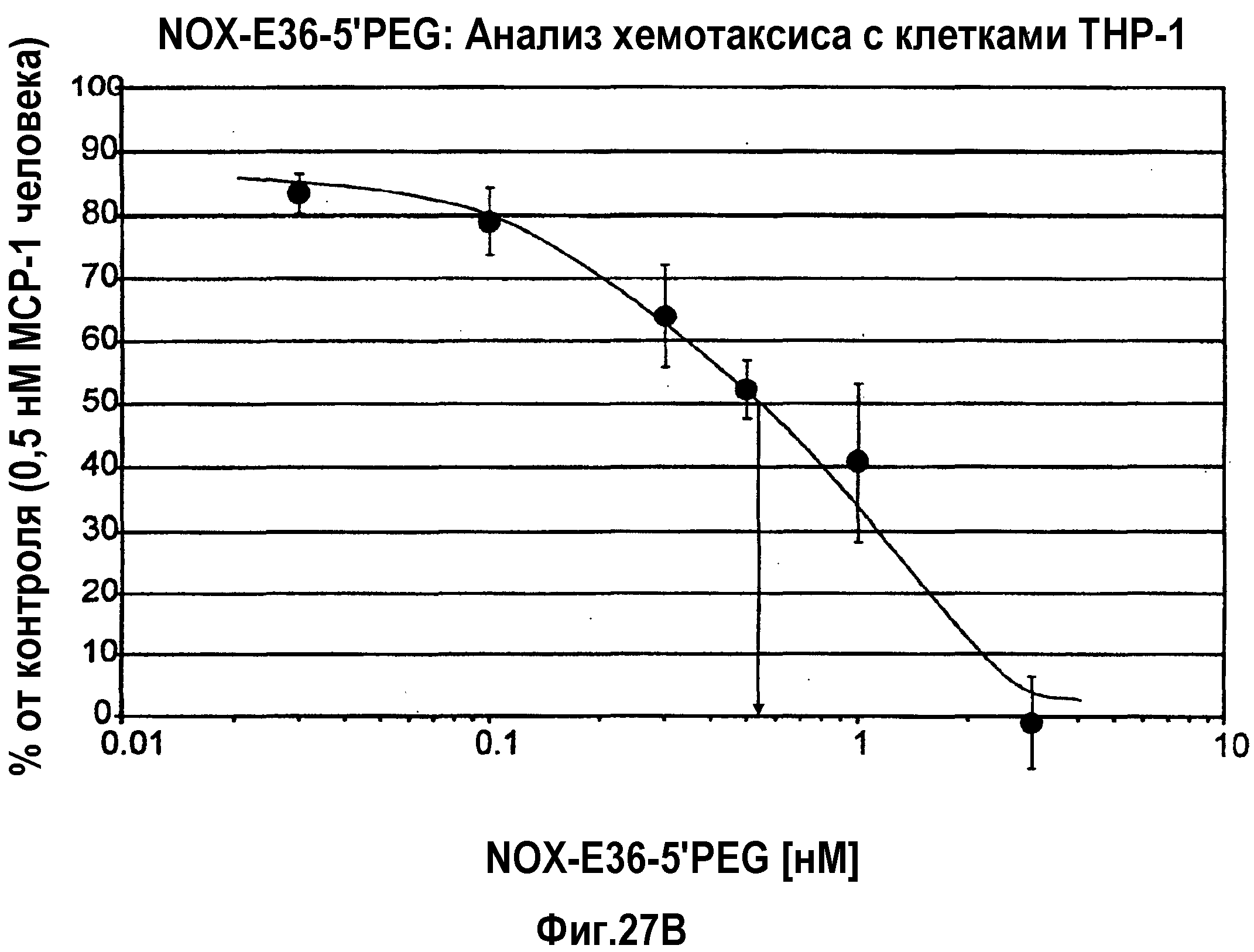


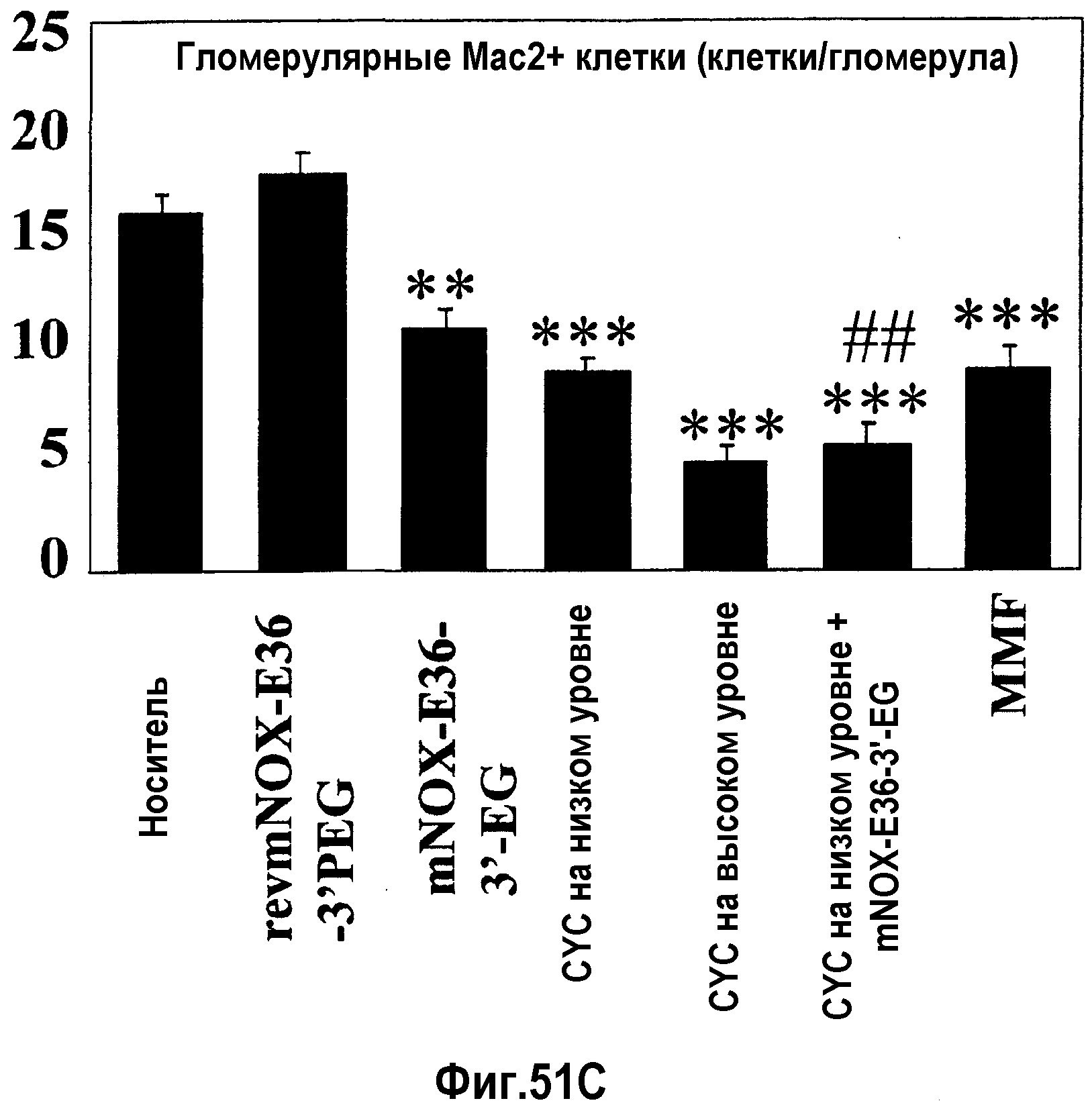
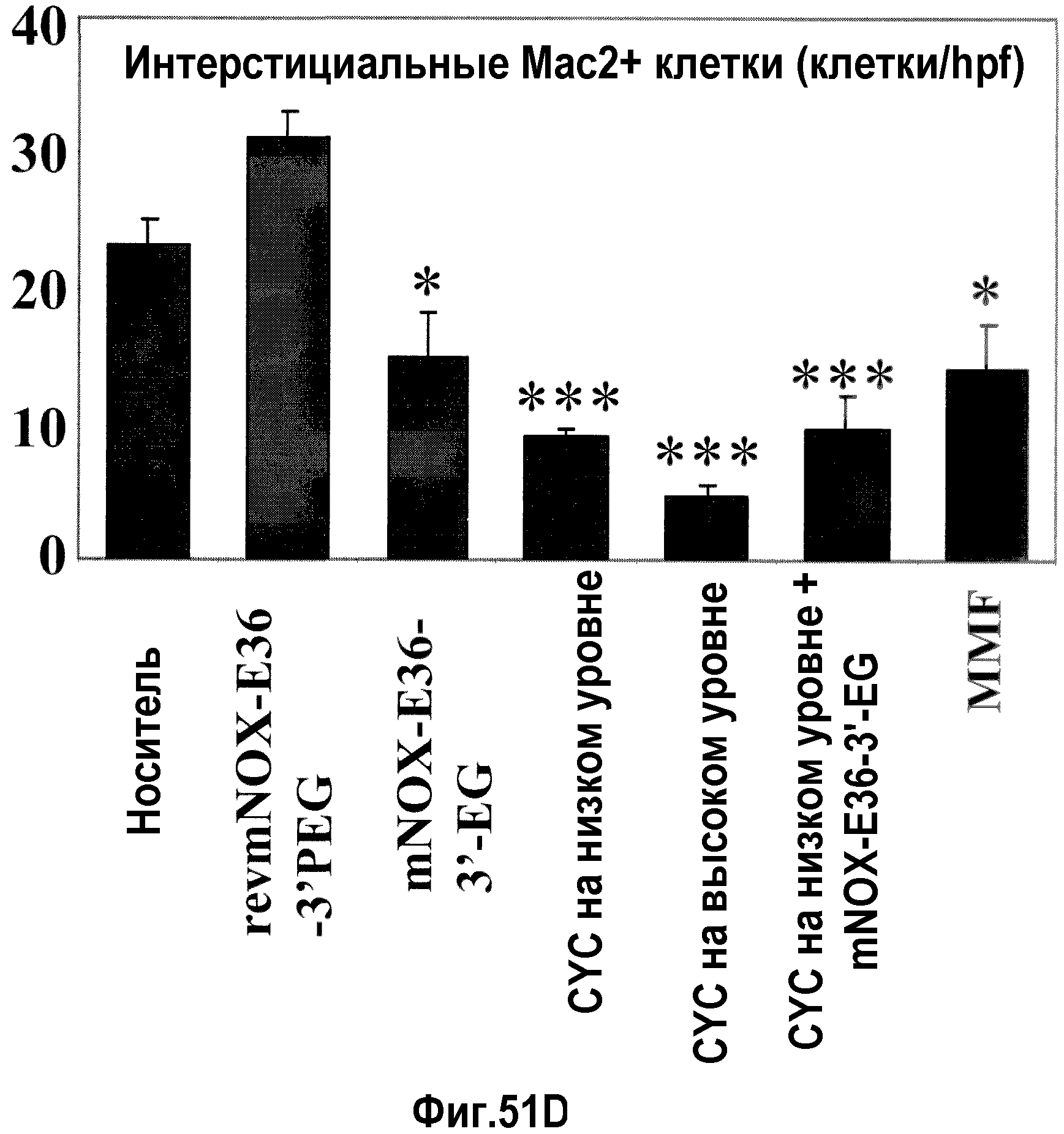





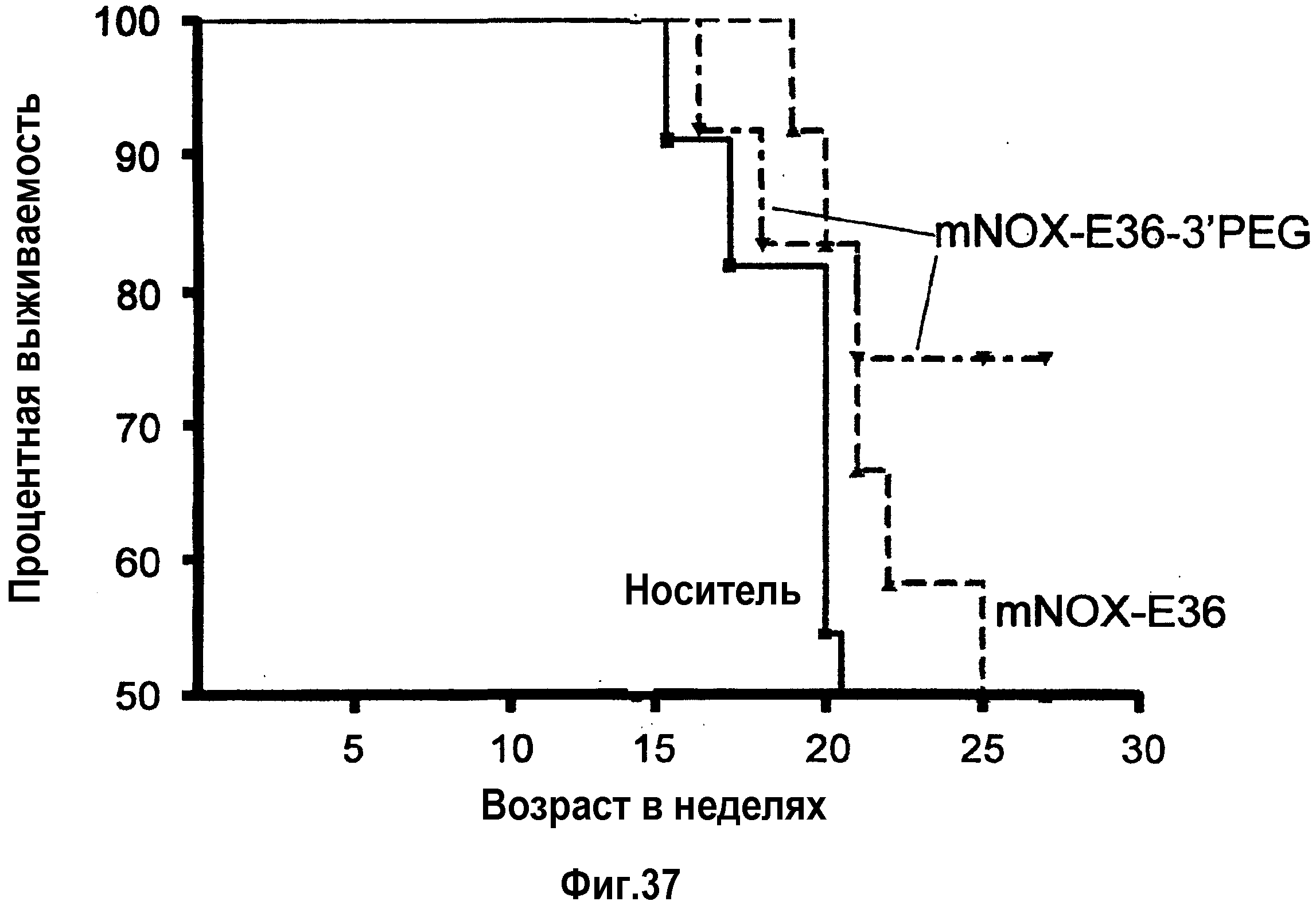







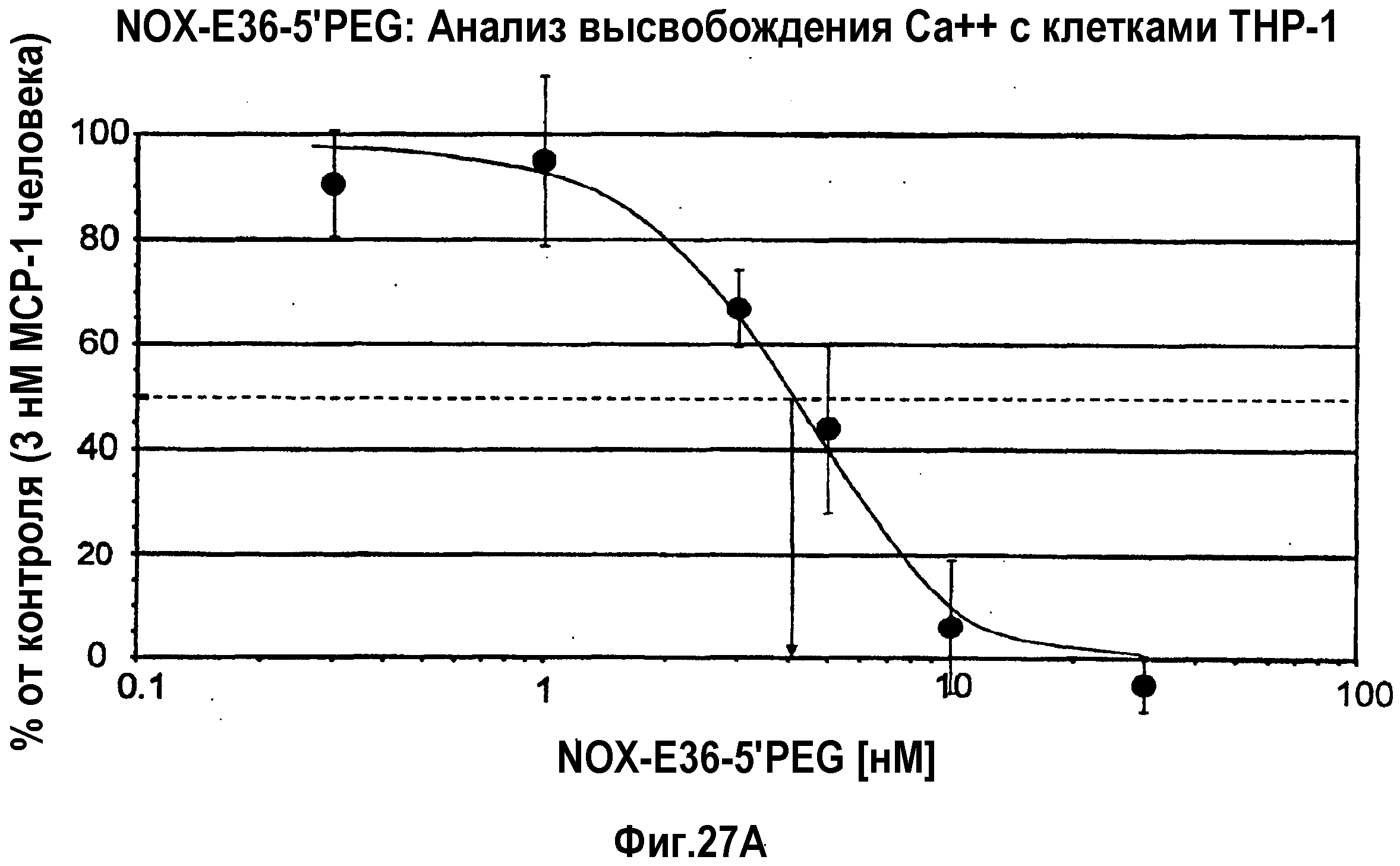
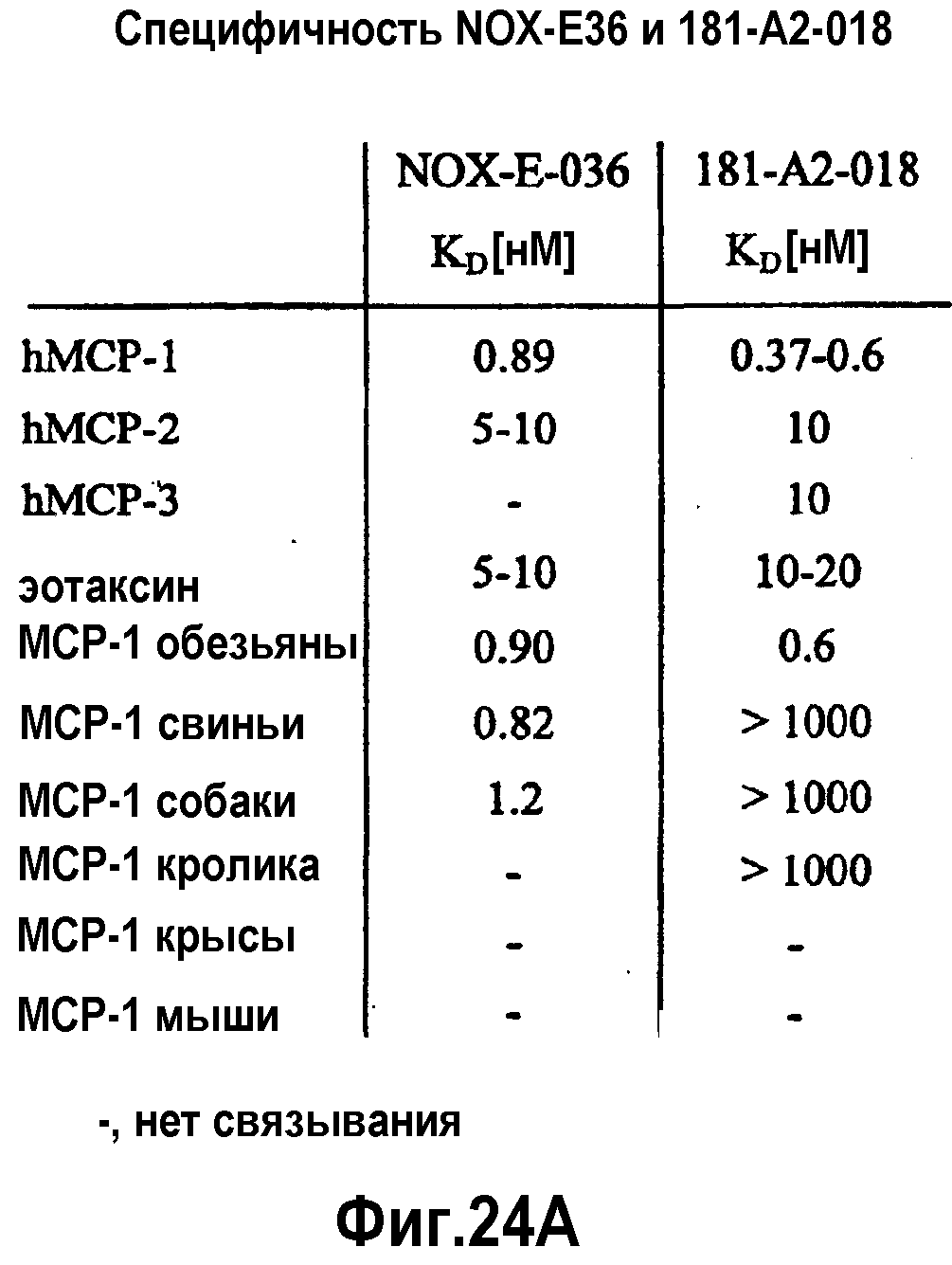

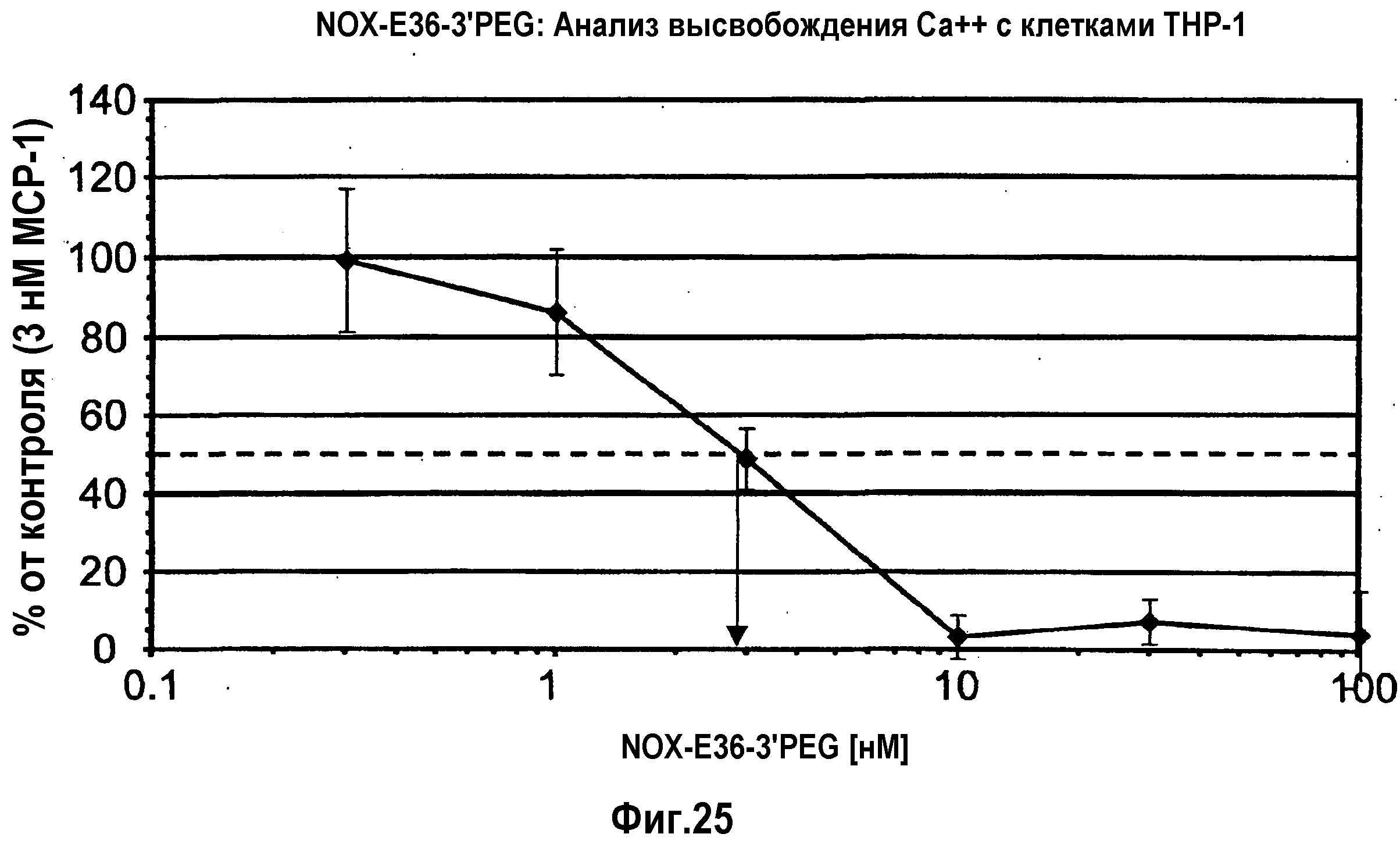

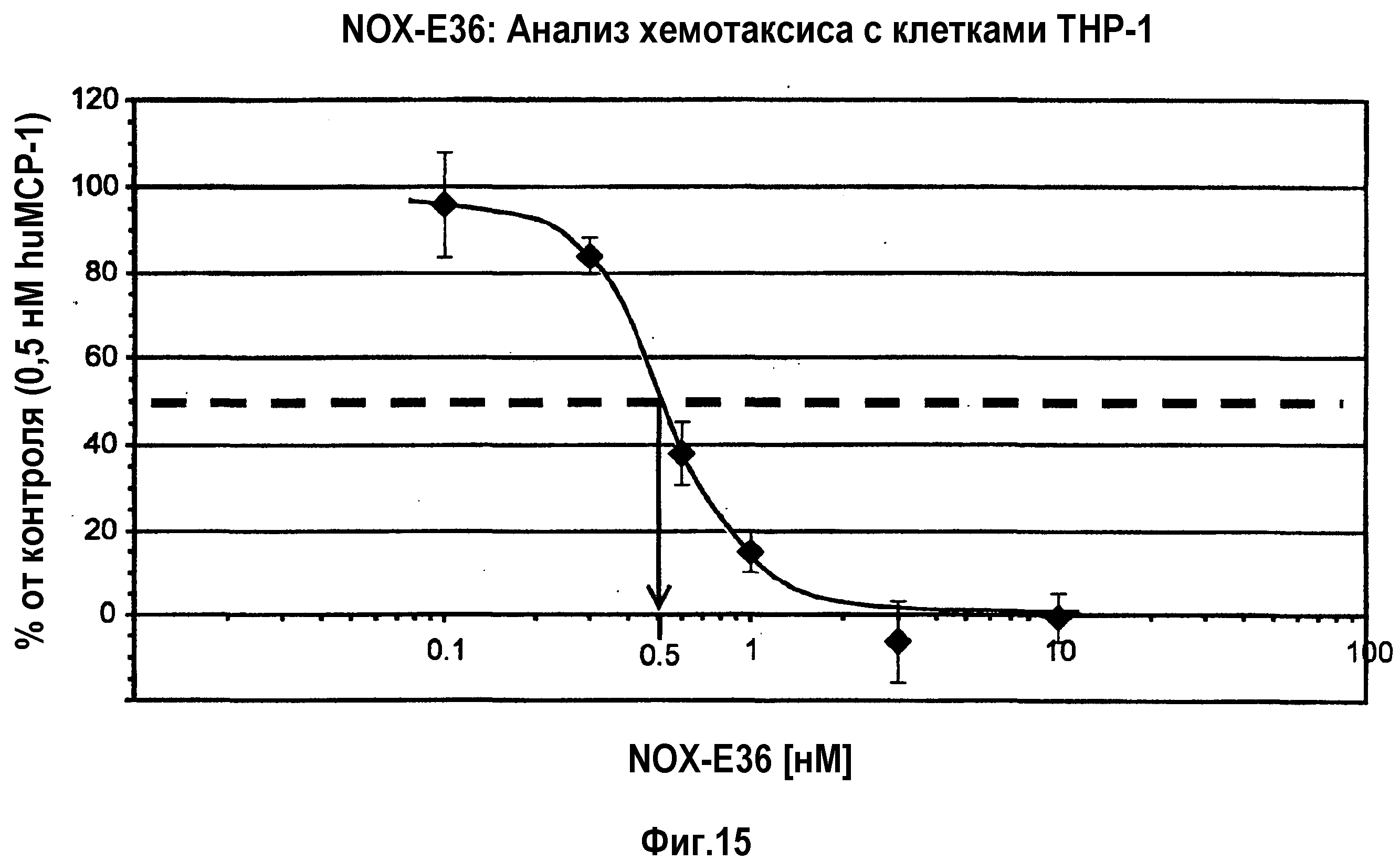










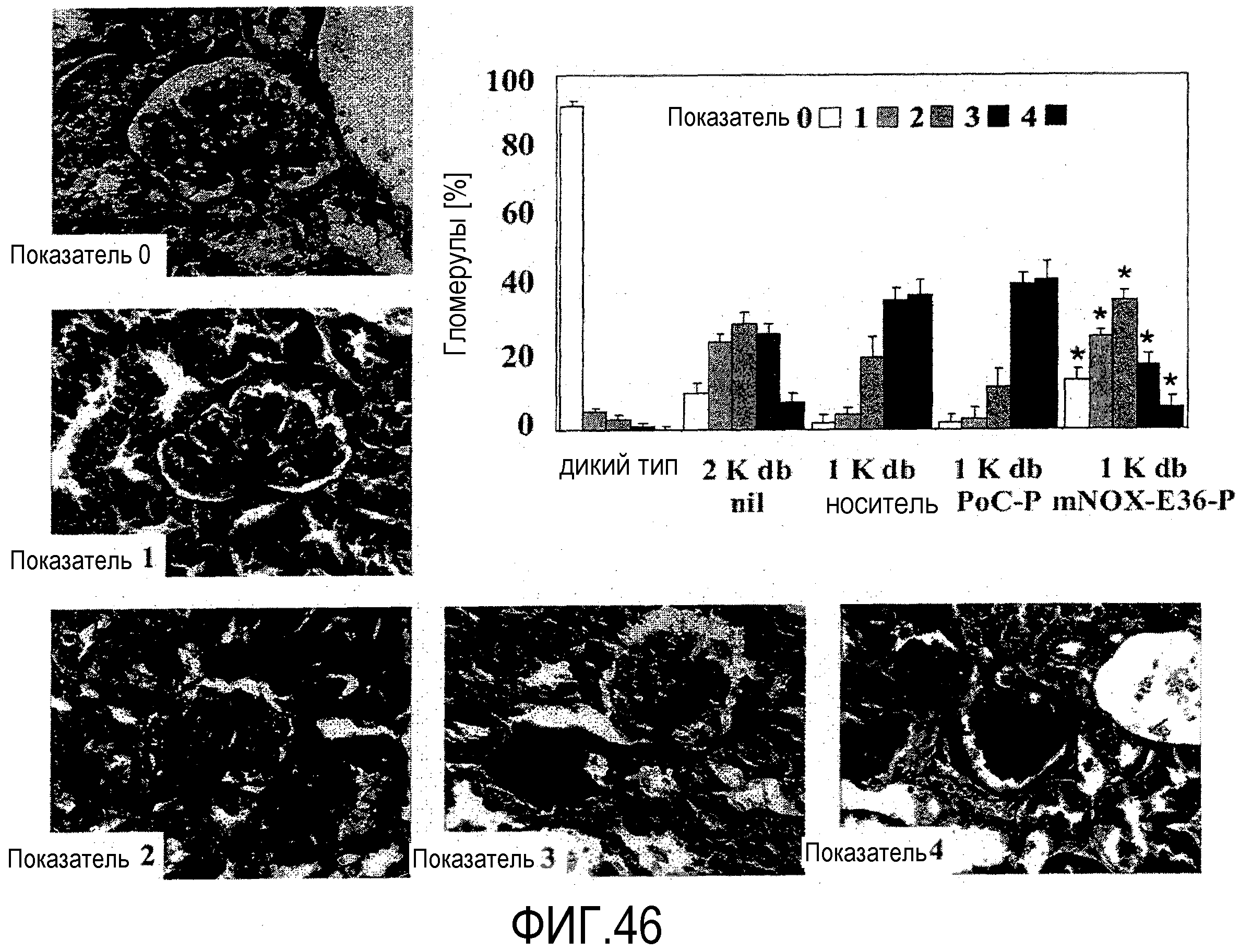

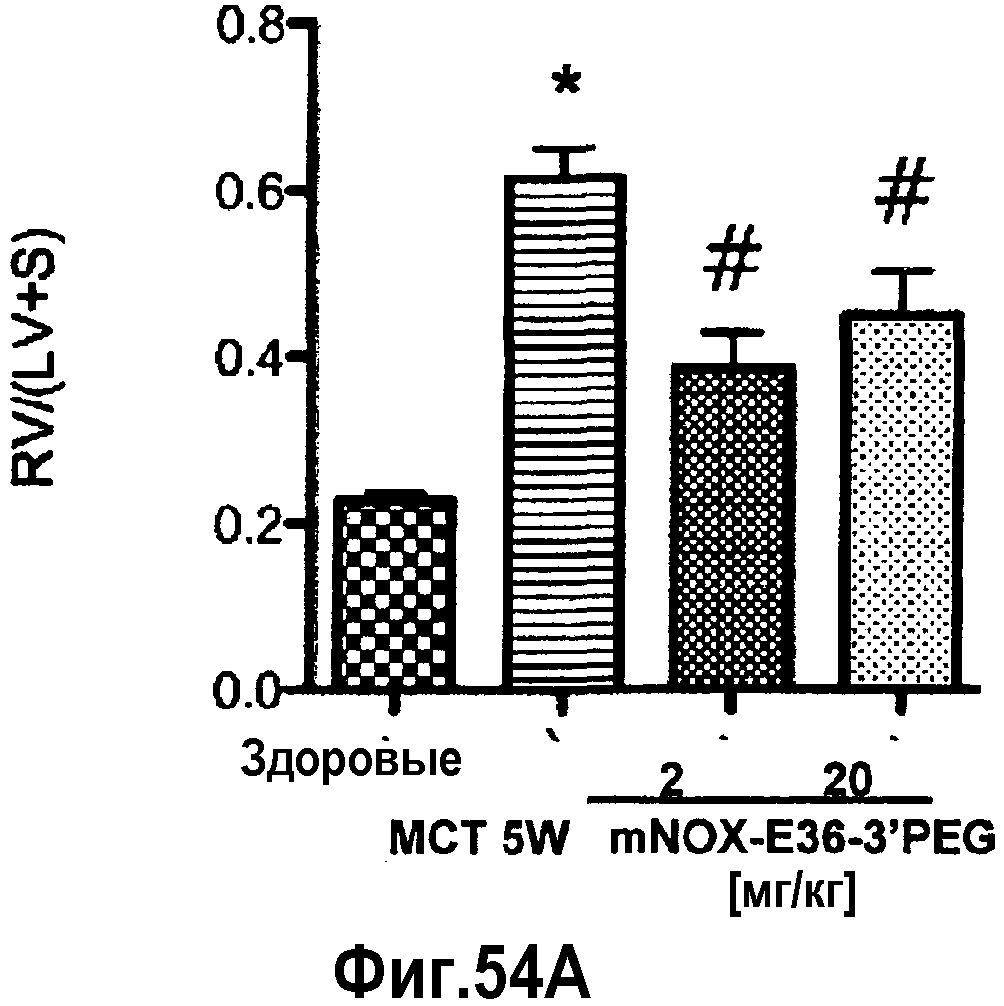
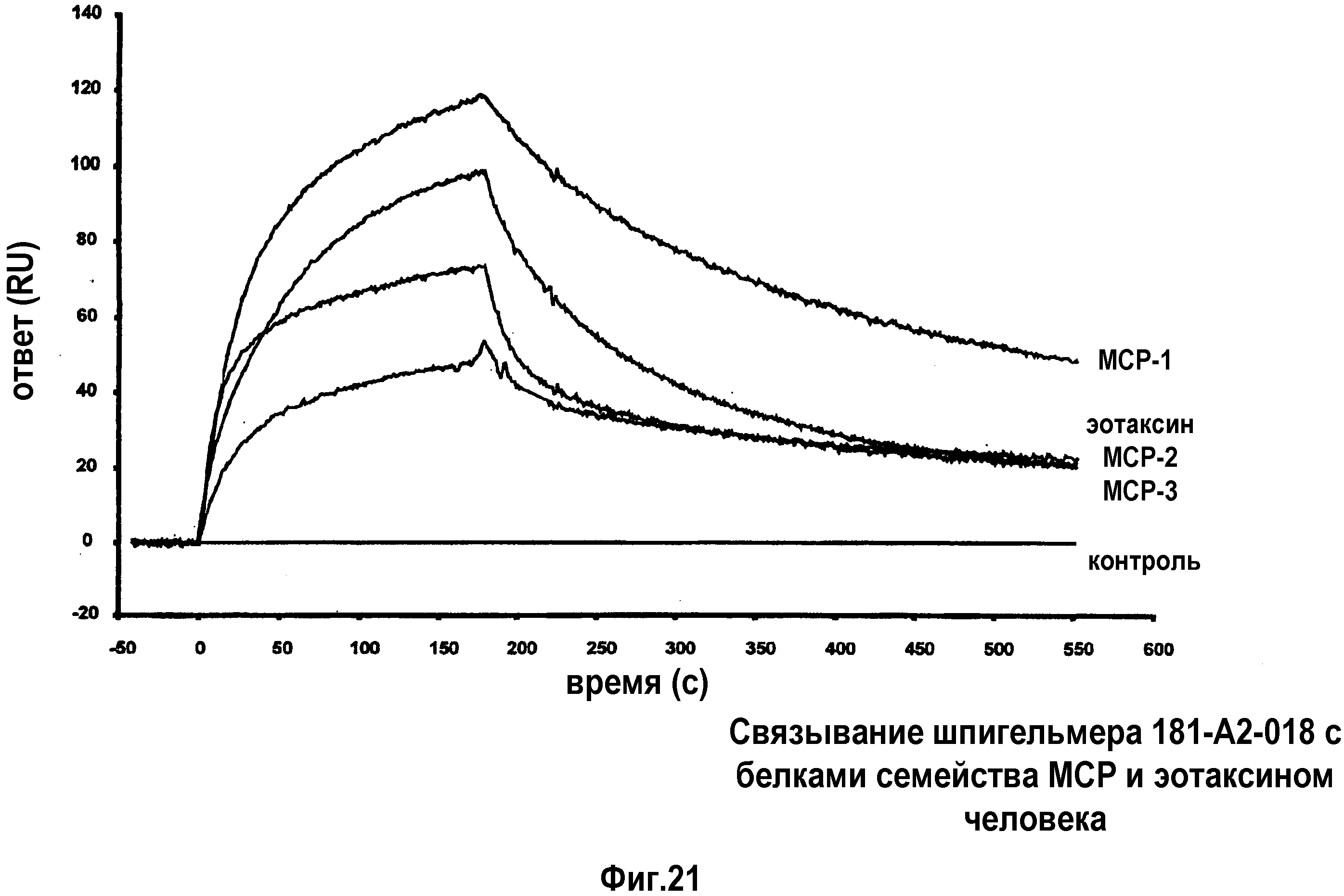

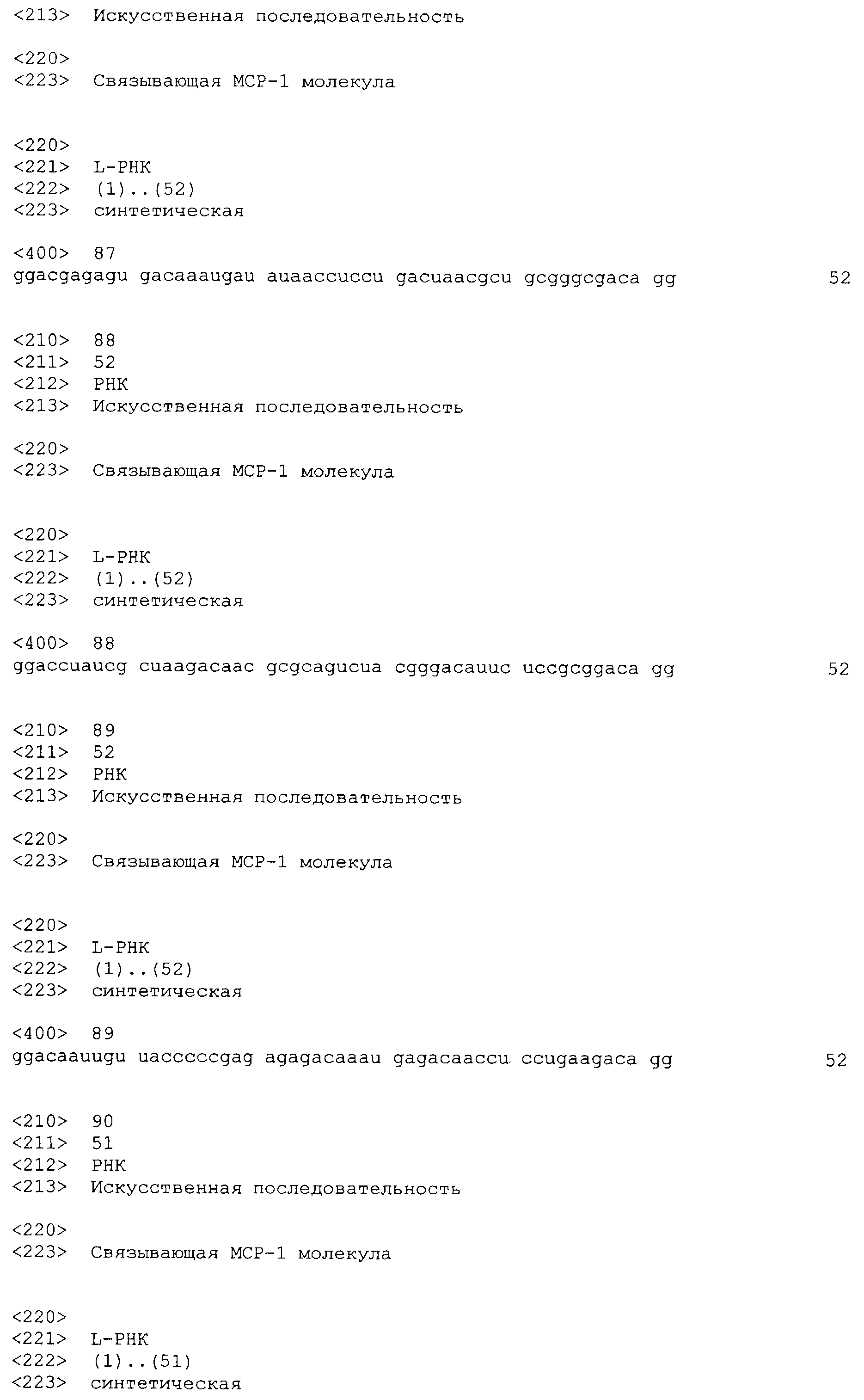





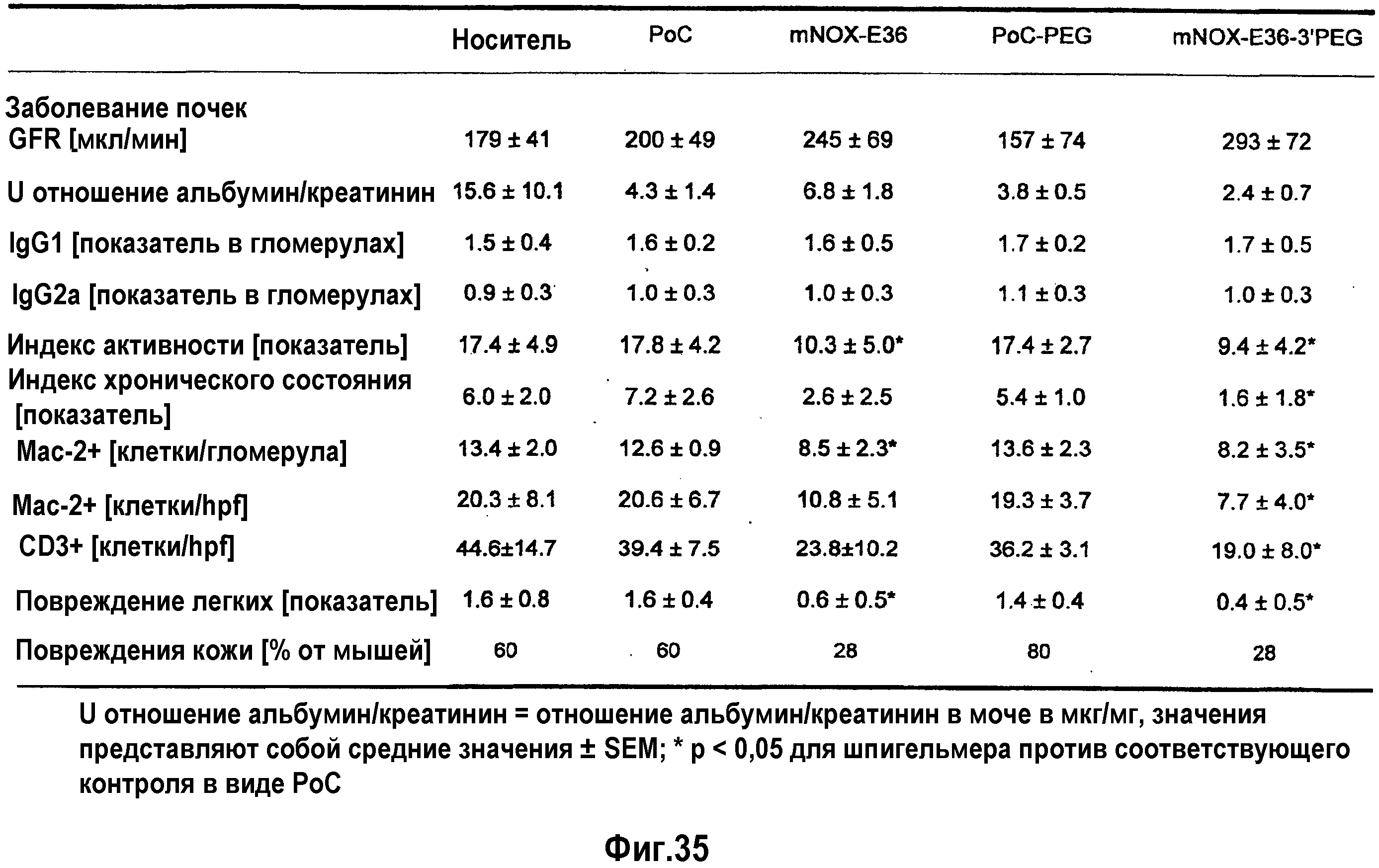


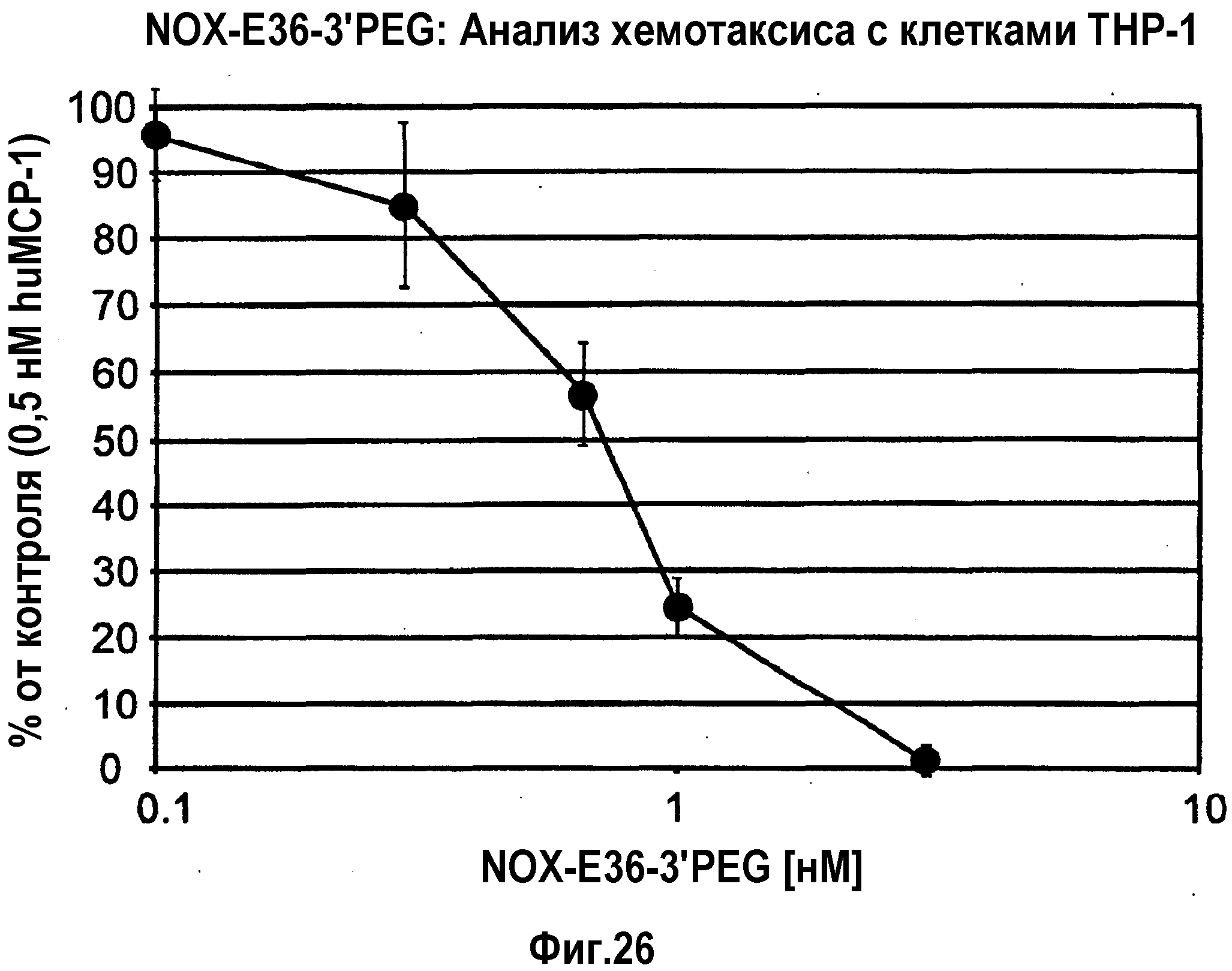
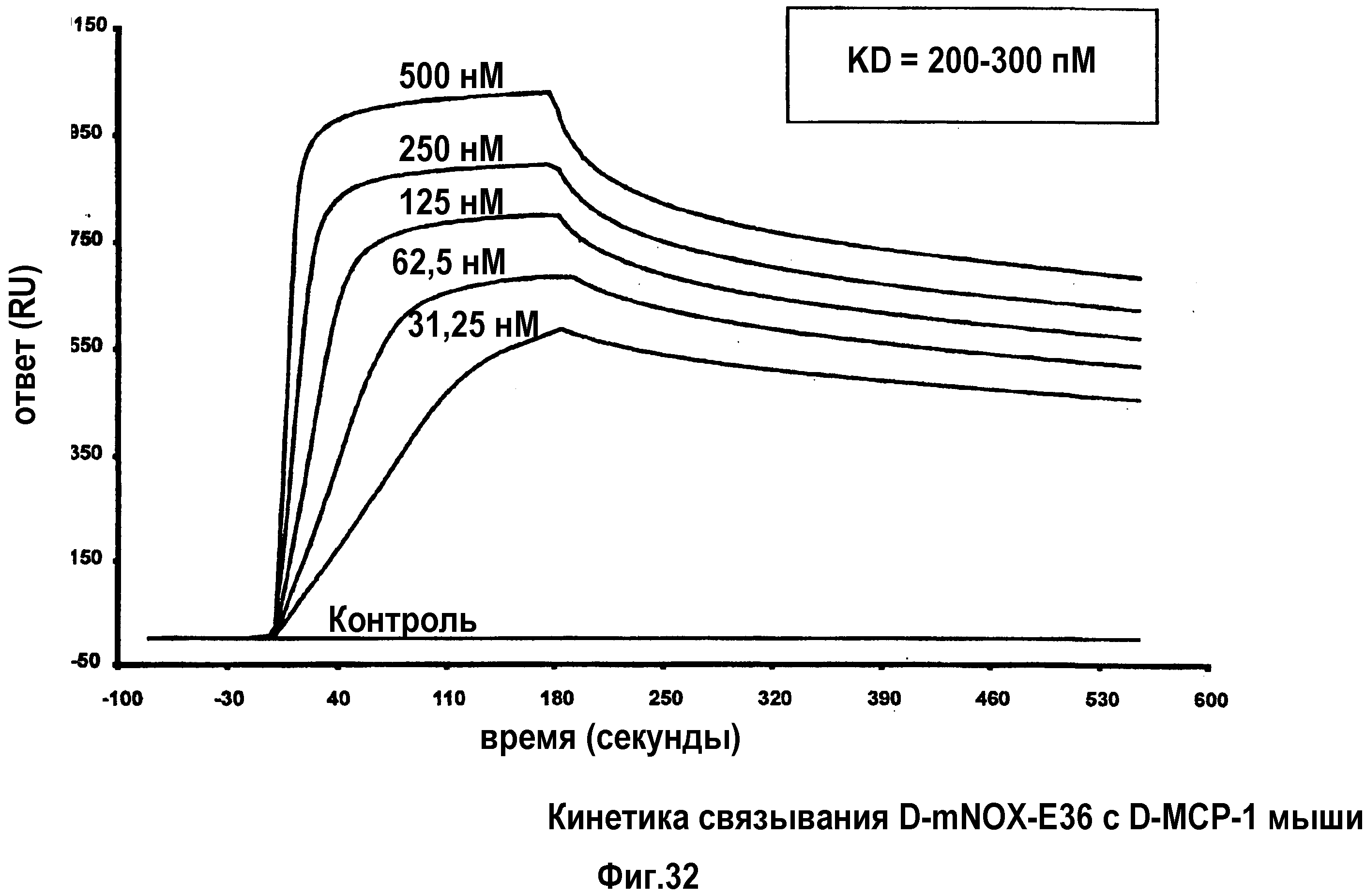




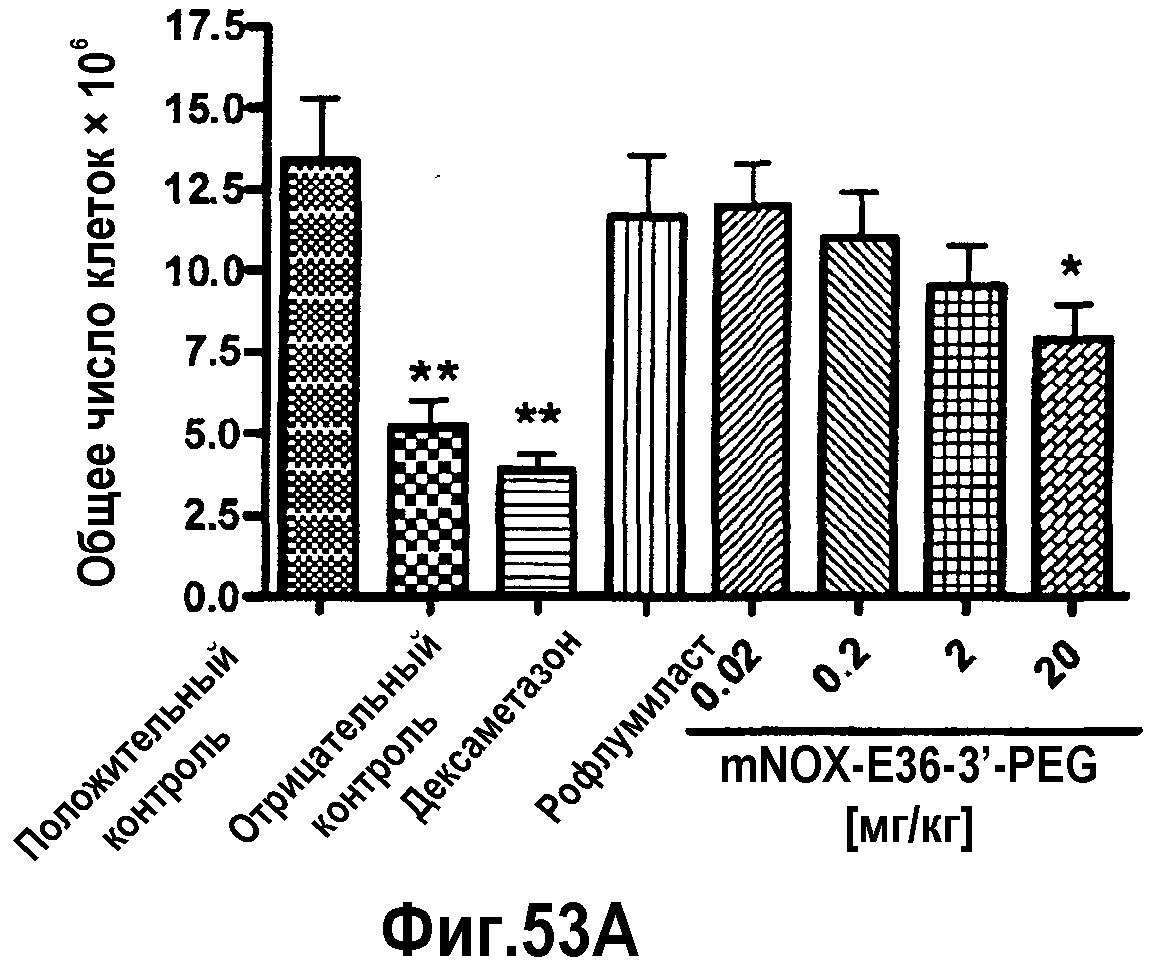
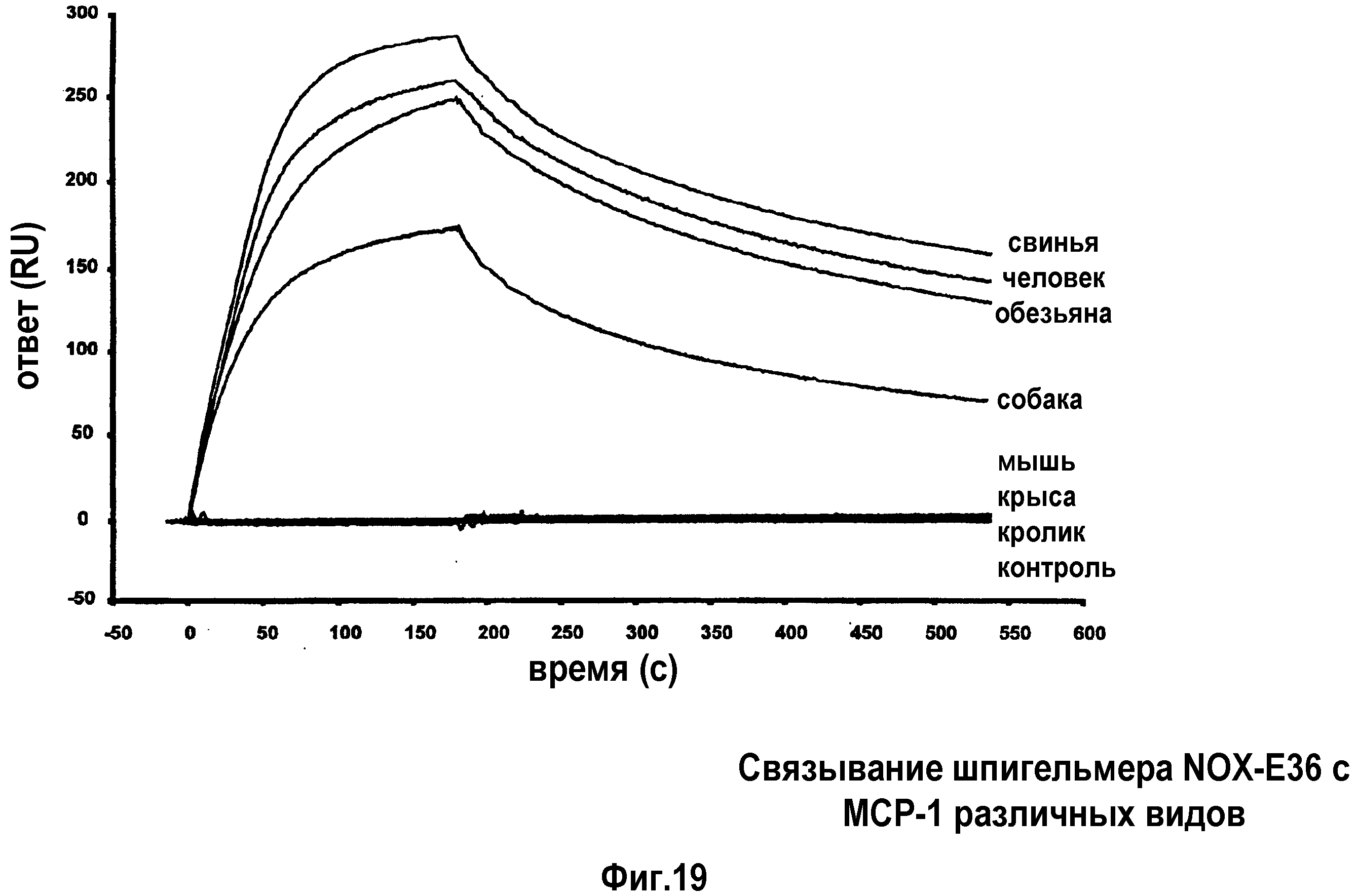
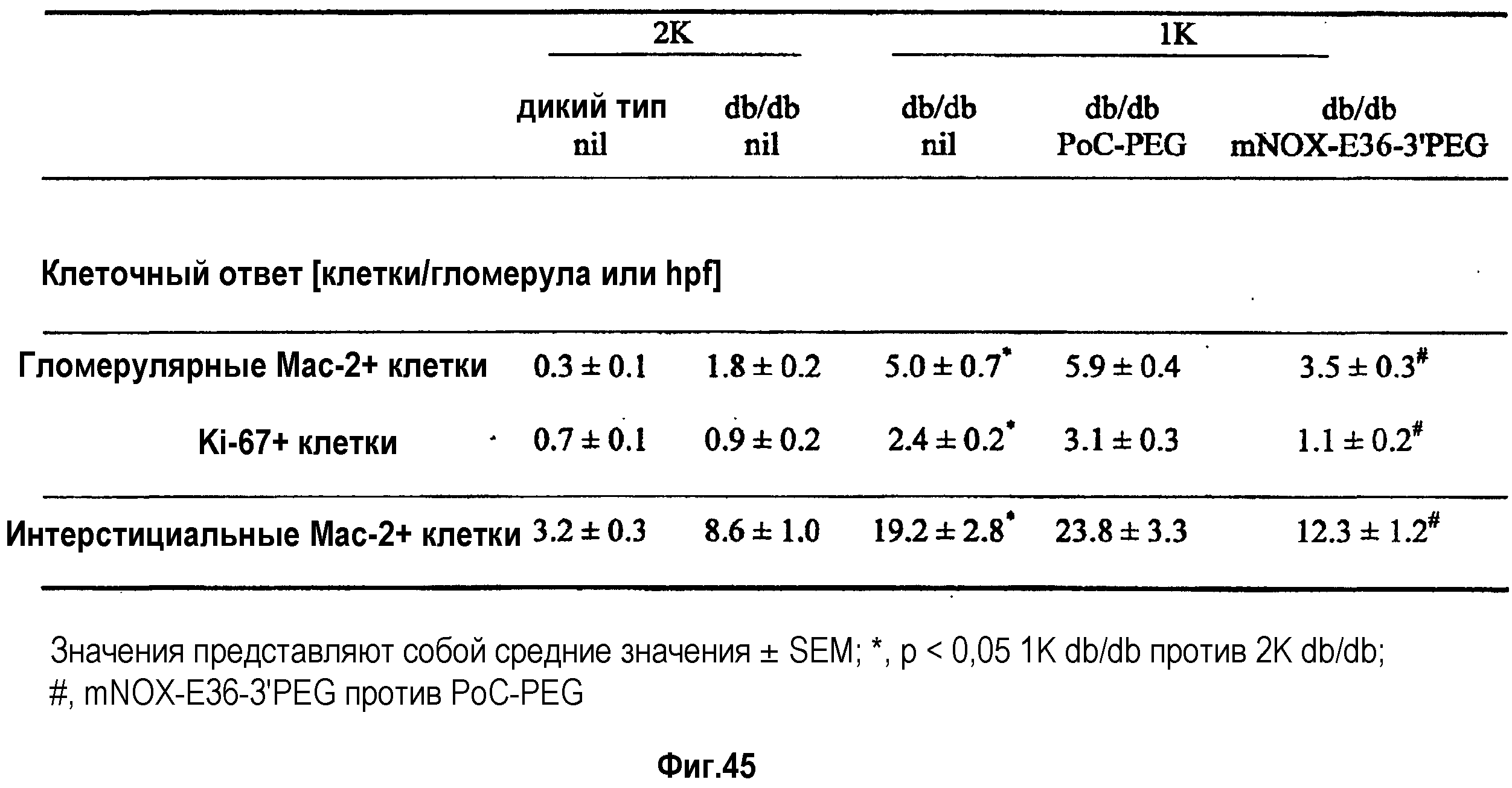
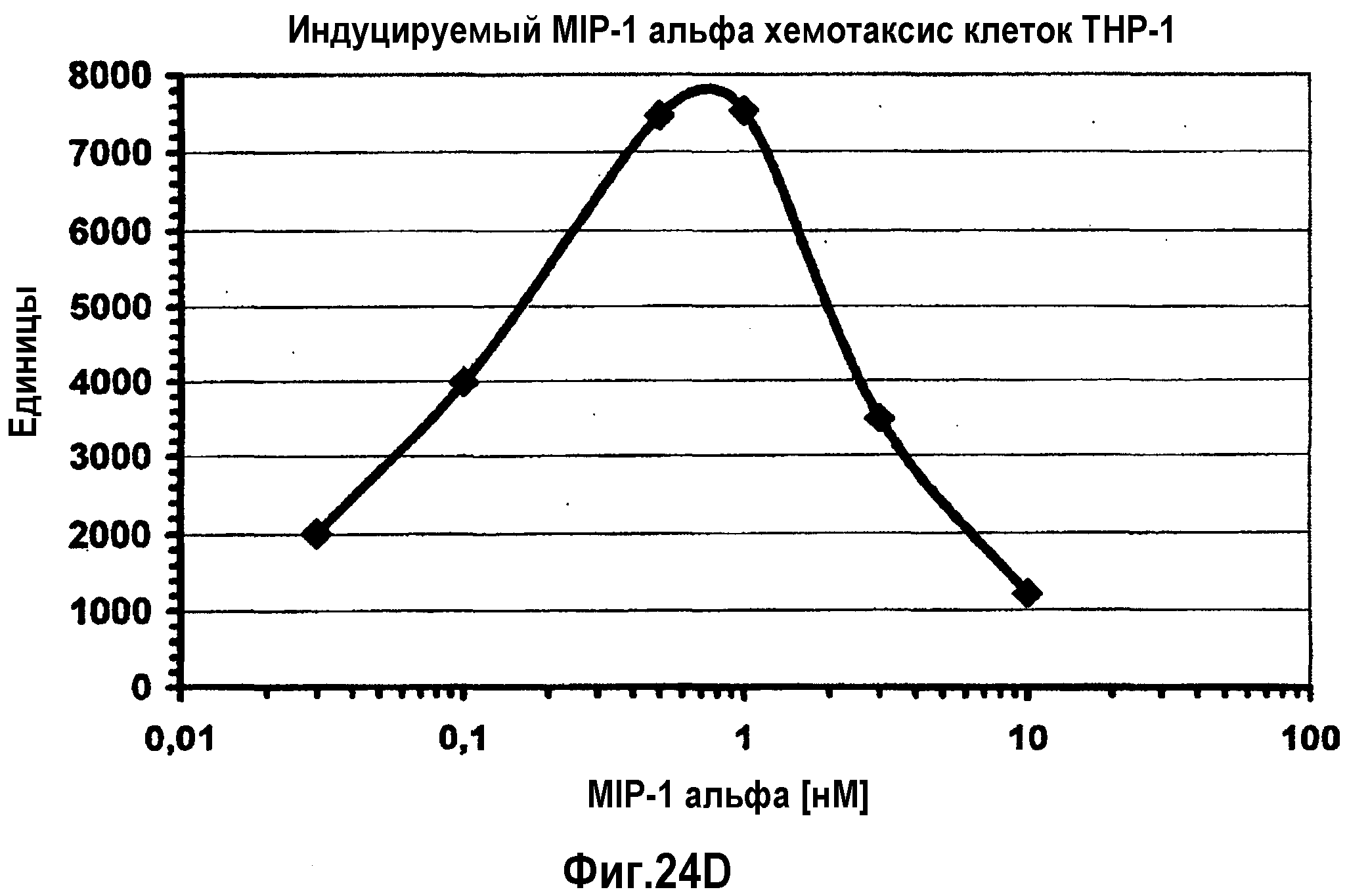

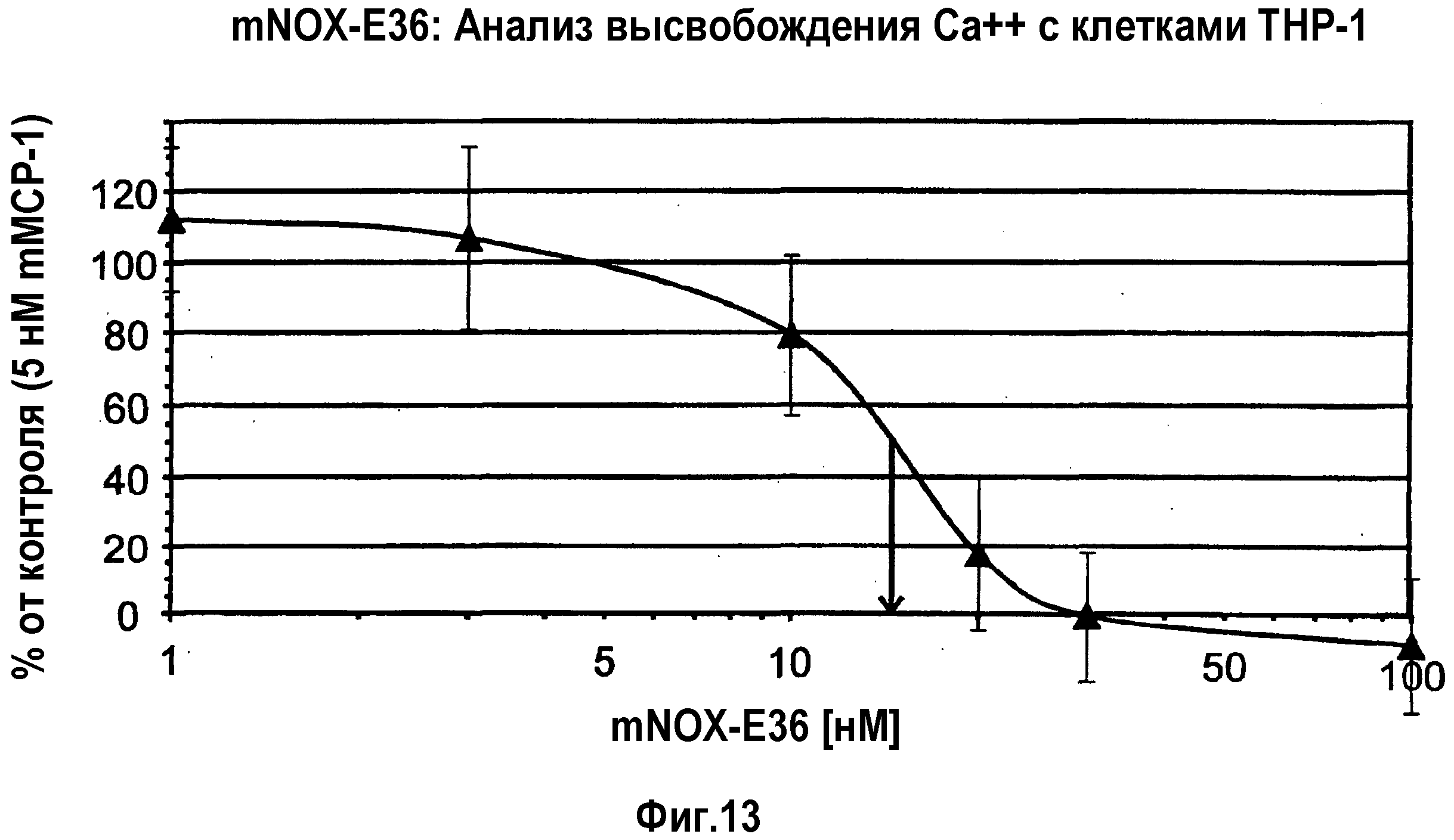





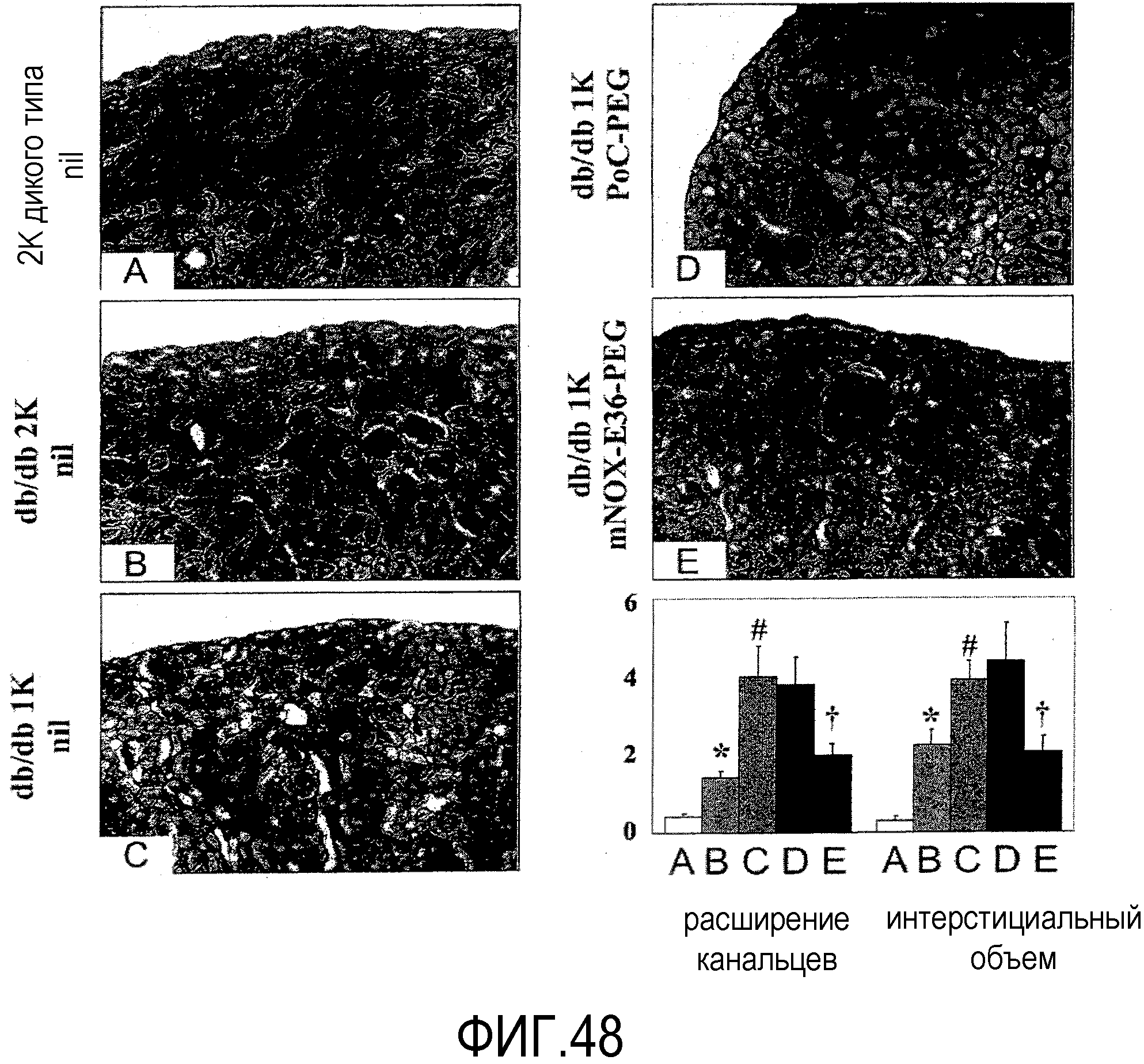
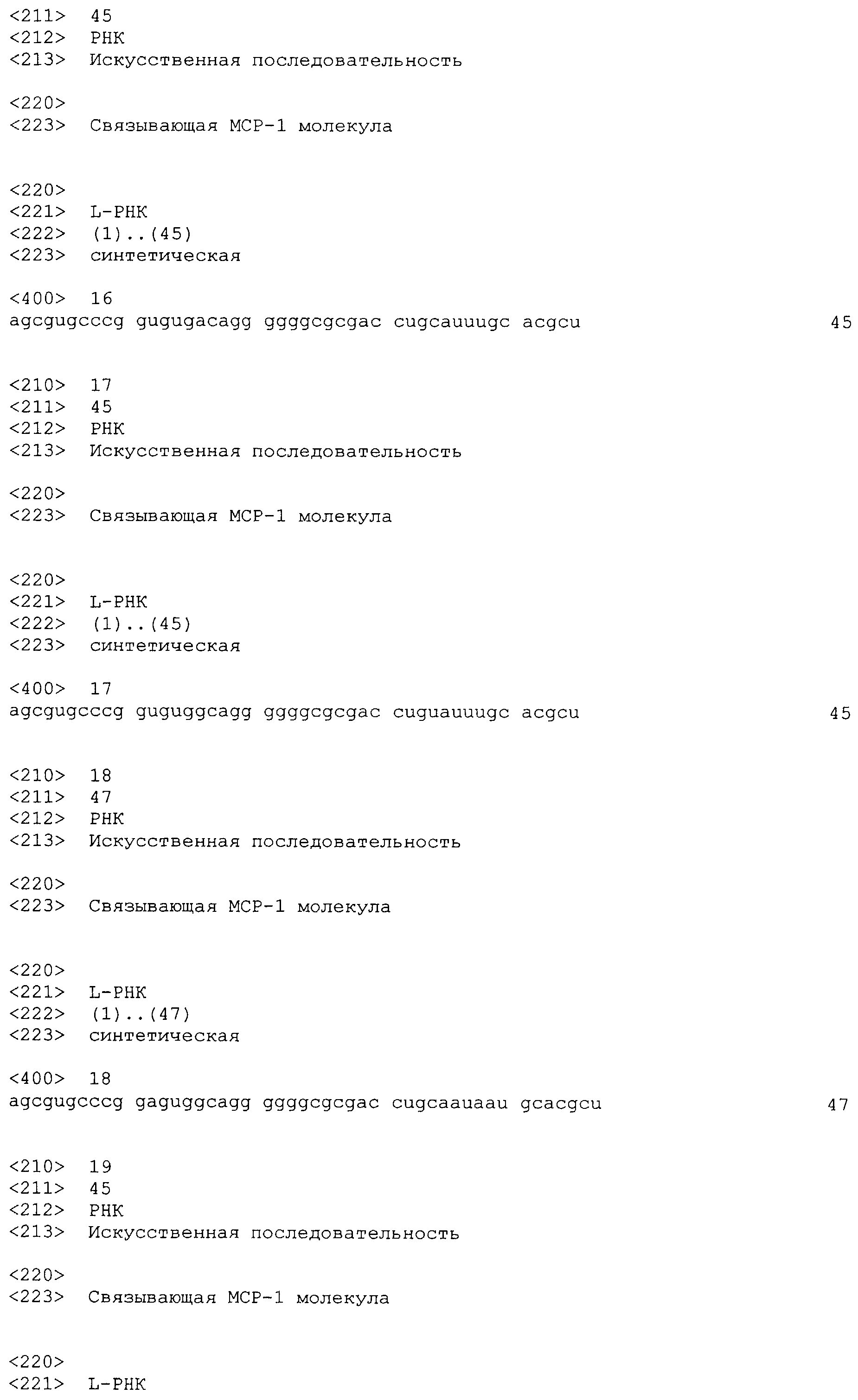


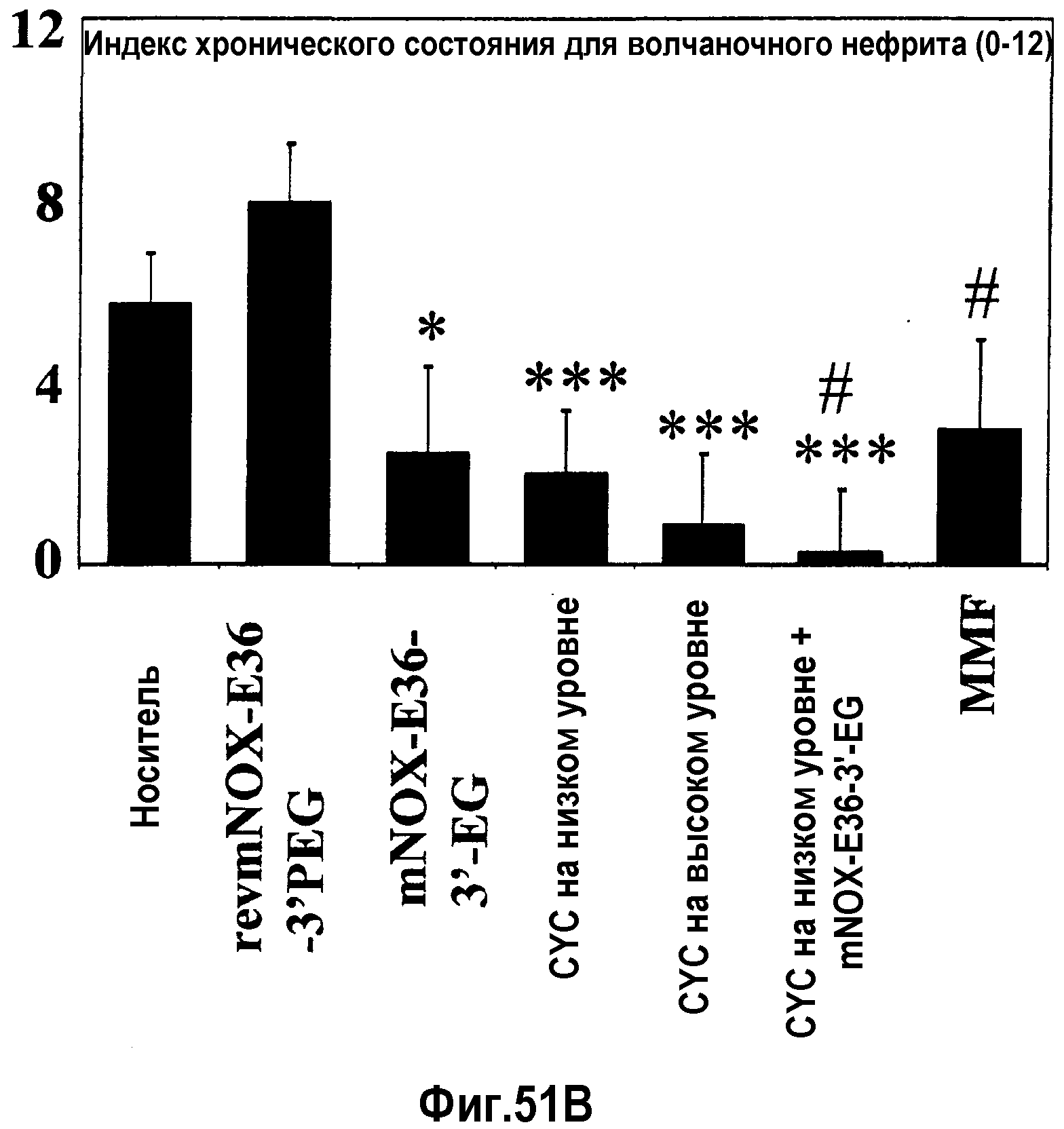

Нуклеиновые кислоты, связывающиеся с мср-1
Sdf-1-связывающие нуклеиновые кислоты
Связывающие sdf-1 нуклеиновые кислоты и их применение
Новые связывающие c5a нуклеиновые кислоты
Связывающиеся с sdf1 нуклеиновые кислоты и их применение при лечении рака
Нуклеиновые кислоты, связывающиеся с мср-1
Sdf-1-связывающие нуклеиновые кислоты
Связывающие sdf-1 нуклеиновые кислоты и их применение
Новые связывающие c5a нуклеиновые кислоты

