Результат интеллектуальной деятельности: СПОСОБ СОЗДАНИЯ МОДЕЛИ ПЕРЕКИСНОГО ОКИСЛЕНИЯ ЛИМФОЦИТОВ
Вид РИД
Изобретение
Известны способы оценки эффективности модели перекисного окисления путем определения роста общего содержания SH-групп моделированных белков [Microtubule dynamics and glutathione metabolism in phagocytizing human polymorphonuclear leukocytes / B.R. Burchill, J.M. Oliver, С.B. Pearson et al. // J. of Cell Biology. - 1978. - Vol.76, №2. - P. 439-447.], но данная модель не позволяет оценить уровень белково-связанного глутатиона, а следовательно, оценить эффективность моделирования перекисного окисления лимфоцитов. Известен также способ косвенного определения содержания восстановленного глутатиона по активности глутатионпероксидазы [Медицинские лабораторные технологии: В 2-х томах./Под ред. А.И. Карпищенко - Т. 2 / А.И. Карпищенко. - СПб.: Интермедика. - 1999. - 656 с], однако, активность этого фермента зависит от конформации активного центра фермента. При изменении конформации активного центра фермента его активность меняется, что не позволяет с высокой точностью оценить уровень восстановленного и белково-связанного глутатиона, а следовательно, оценить эффективность моделирования перекисного окисления. Известен также способ косвенного определения концентрации восстановленного глутатиона по активности глутатионредуктазы [Медицинские лабораторные технологии: В 2-х томах./Под ред. А.И. Карпищенко - Т. 2 / А.И. Карпищенко. - СПб.: Интермедика. - 1999. - 656 с], однако активность этого фермента зависит от конформации активного центра фермента. При изменении конформации активного центра фермента его активность меняется, что не позволяет достоверно в любой ситуации оценить с высокой точностью уровень восстановленного и белково-связанного глутатиона, а следовательно, оценить эффективность модели переокисления липидов.
Известен также способ оценки эффективности модели переокисления по содержанию восстановленного глутатиона, предложенный М.Е. Anderson (1985) в модификации S. Kojima et al. (2004) [Low dose gamma-rays activate immune functions via induction of glutathione and delay tumor growth / S. Kojima, K. Nakayama, H. Ishida // J. Radiat. Res. - 2004. - Vol.45, №1. - P. 33-39.], основаный на взаимодействии восстановленного глутатиона (GSH) с 5,5′-дитио-бис(2-нитробензойной) кислотой (ДТНБ). При этом образуется окисленный глутатион (GSSG), который затем восстанавливается и вновь взаимодействует с ДТНБ. Данный способ является наиболее близким к предлагаемому по технической сущности и достигаемому результату и выбран в качестве прототипа.
Недостатком данного способа является невозможность оценки эффективности модели перекисного окисления липидов в мембране лимфоцитов.
Целью предлагаемого изобретения является повышение оценки эффективности и точности способа.
Указанная цель достигается дополнительной предварительной обработкой лимфоцитов перекисью водорода в конечной концентрации 0,5 мМ для последующей оценки уровня белково-связанного глутатиона лимфоцитов.
Новым в данном способе является внесение в инкубационную среду лимфоцитов перекиси водорода в конечной концентрации 0,5 мМ, без которой невозможна оценка эффективности модели перекисного окисления липидов в мембране лимфоцитов.
Следовательно, только комплексная модернизация способа-прототипа позволяет получить желаемый результат. Только внесение в инкубационную среду лимфоцитов перекиси водорода в концентрации 0,5 мМ позволяет эффективно оценить модель перекисного окисления липидов в мембране лимфоцитов, а именно при одноразовом увеличении уровня белково-связанного глутатиона в 6 раз и более считают модель эффективной, а при увеличении уровня белково-связанного глутатиона в 2 раза и менее считают неэффективной.
В настоящее время известно, что ряд патологических состояний, в том числе гипоксия, нейродегенеративные процессы, онкологические заболевания сопровождаются наработкой активных форм кислорода (АФК), нарушением окислительно-восстановительного баланса клетки с последующим формированием окислительного стресса (ОС) и дизрегуляцией апоптоза. Для изучения процесса дизрегуляции апоптоза в условиях ОС и поиска молекулярных мишеней воздействия на клеточную гибель используются модели, созданные in vitro, получившие в экспериментальной медицине и биологии широкое распространение.
Для интеграции теории в практику необходимы экспериментальные модели патологических процессов, которые создаются на нормально функционирующих клеточных или тканевых системах.
Модель экспериментального перекисного окисления лимфоцитов формировали на лимфоцитах крови. Для индукции переокисления в эксперименте использовали различные концентрации пероксида водорода: 0,3; 0,5; 1 и 2 мМ. Оптимальным был выбран уровень 0,5 мМ (см. таблицы 1 и 2).
|
|
Для оценки клеточного ответа на воздействие пероксида водорода в изучаемых концентрациях проводили определение: содержания внутриклеточных АФК методом проточной цитофлуориметрии с помощью красителя с заблокированной флуоресценцией - дихлорфлуоресцеина диацетата. Дихлорфлуоренцеин диацетат, изначально не флуоресцирующий, пассивно проникает внутрь клетки и под действием эстераз переходит в полярное соединение, не способное диффундировать обратно из клетки. После реакции с H2O2 он превращается во флуоресцирующий метаболит. Количество аннексин-положительных клеток оценивается методом проточной цитофлуориметрии по способности аннексина V специфически связываться с фосфатидилсерином, экспрессированным на внешней стороне цитоплазматической мембраны апоптозных клеток и количества пропидий иодид-положительных клеток, оцениваемых по способности пропидия йодида (PI) проникать в поврежденные клетки и интерколировать с дефрагментированной ДНК при некрозе клетки.
Поддержание внутри клетки содержания на определенном уровне АФК важно для регуляции жизнедеятельности. Активные формы кислорода участвуют в реализации танатогенной программы посредством изменения редокс-регуляции экспрессии генов и проапоптотических белков.
При активации реакций переокисления происходит окисление белков, что способствует развитию повреждений на уровне молекул, клеток и целого организма.
Первые сведения об окислении белковых молекул АФК получены A.J. Swallow (1960), W.M. Garrison, М.Е. Jayko (1962), которые описали модификацию 17-ти белков под действием ·ОН и 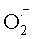 , генерируемых при радиолизе воды. В настоящее время интерес к изучению механизмов взаимодействия АФК с белками значительно возрос. Механизмы повреждений белков различными АФК включают нарушение третичной структуры, агрегацию и денатурацию с изменением функциональной активности и антигенных свойств молекул.
, генерируемых при радиолизе воды. В настоящее время интерес к изучению механизмов взаимодействия АФК с белками значительно возрос. Механизмы повреждений белков различными АФК включают нарушение третичной структуры, агрегацию и денатурацию с изменением функциональной активности и антигенных свойств молекул.
В любых белках наиболее чувствительны к окислению остатки триптофана, цистеина, тирозина и гистидина. Атака функциональных групп аминокислот приводит к образованию первичных радикалов, которые вступают в реакции с соседними остатками аминокислот, создавая сложную картину повреждающего действия АФК на макромолекулы белка. Окислительные изменения структуры полипептидной цепи могут касаться не только функциональных групп в боковых радикалах аминокислотных остатков, но и непосредственно пептидных связей, приводя к фрагментации молекул.
Поэтому в настоящее время окислительное повреждение белков рассматривается как «ранний индикатор повреждения тканей при различных патологических процессах», в том числе при гипоксии и должен использоваться для создания модели перекисного окисления лимфоцитов.
Карбонилированные производные белков (КПБ) образуются при окислении остатков аргинина, лизина, гистидина, пролина, треонина. Вместе с тем, карбонильные группы могут появиться вследствие реакций белка с альдегидными продуктами перекисного окисления липидов, а именно 4-гидрокси-2,3-ноненалем, малоновым диальдегидом с участием гликозилирования и гликооксидации.
Определение уровня окислительно-модифицированных белков (глутатионилирования) в клетках, чувствительных к гипоксии, основывалось на определении: а) содержания SH-групп белков, основанного на способности тиоловых соединений при взаимодействии с ДТНБ образовывать окрашенное соединение - тио-2-нитробензойную кислоту, водный раствор которой имеет максимум поглощения при длине волны 412 нм; б) содержания белково-связанного глутатиона, основанного на способности боргидрата натрия высвобождать глутатион из связи с белками; в) уровня карбонилированных аминокислот модифицированных белков методом иммуноферментного анализа.
Оптимальным для оценки эффективности создания модели перекисного окисления лимфоцитов было определение уровня белково-связанного глутатиона. При этом оптимальная конечная концентрация перекиси водорода была установлена экспериментальным путем.
Таким образом, при гипоксии и переокислении существенная доля образованных в клетках активных форм кислорода не утилизируется и оказывает цитотоксические эффекты. Накопление окислительных повреждений белковых и липидных молекул в результате окислительной модификации белков приводит к возникновению окислительного стресса - составного элемента гипоксии. Накопление окислительно-модифицированных производных белковых молекул не только снижает функциональную активность ион-транспортных систем, но и может являться стимулом для запуска программированной гибели в изучаемых клетках.
При этом антиоксидантная система направлена на эффективную нейтрализацию прооксидантов и снижение токсичной для организма гидроперекиси. Основным компонентом антиокидантной системы является восстановленная форма глутатиона.
Глутатион - трипептид (L-γ-глутамил-L-цистеилглицин) с молекулярной массой 307 Da занимает особое место среди SH-содержащих соединений. Наличие γ-глутамильной связи защищает трипептид от ферментативной деградации. В организме глутатион присутствует в двух формах: окисленной - GSSG и восстановленной - GSH, причем содержание GSH в клетках на несколько порядков выше, чем GSSG [Колесниченко Л.С., 1989; Wu G. et al., 2004; Смирнова Г.В., Октябрьский О.Н., 2005; Марри Р. и соавт., 2009]. По данным P. Pietarinen-Runtti et al. (2000), концентрация GSH в нейтрофилах составляет около 5 нмоль/мг белка. Содержание глутатиона в сыворотке крови здоровых людей незначительно, поэтому клетки основную потребность в GSH обеспечивают путем нематричного синтеза [Wu G. et al., 2004] в ходе двух последовательных реакций, катализируемых γ-глутамилцистеин-синтетазой (КФ 6.3.2.2) и глутатион-синтетазой (КФ 6.3.2.3) [Кулинский В.И., 1990; Смирнова Г.В., Октябрьский О.Н., 2005; Марри Р. и соавт., 2009]. Лимитирующим звеном синтеза является образование γ-глутамилцистеина, зависящее от наличия L-цистеина и его способности окисляться в L-цистин [Зенков Н.К. и соавт., 2001]. В то же время недостаточность глутатион-синтетазы способствует развитию окислительных повреждений в нейтрофилах [Spielberg S.P. et al., 1979].
Глутатион при физиологических значениях pH имеет две анионные карбоксигруппы, положительно заряженную аминогруппу и SH-группу цистеинового остатка, которая придает GSH свойства восстановителя и способность быстро обезвреживать свободные радикалы и АФК [Day R.M., 2005; Zhu Y., 2007; Circu C.L. et al., 2009]. Глутатин является типичным тиолом и, участвуя в одноэлектронных восстановительных реакциях, становится GS·, который димеризуется до GSSG, легко реагирующего со свободными SH-группами. Второй тип окислительно-восстановительных превращений с участием GSH - это реакции тиолдисульфидного обмена, которые известны как основной путь образования смешанных дисульфидов глутатиона с белками (белок-SSG) и играют роль в регуляции биологических процессов [Chai Y.C. et al., 1994]. В реакциях третьего типа происходит двухэлектронное окисление глутатиона с образованием интермедиата, который реагирует со второй молекулой GSH (получение GSSG) или иной молекулой (синтез смешанного дисульфида) [Смирнова Г.В., Октябрьский О.Н., 2005].
GSH является ингибитором активных форм кислорода (АФК) и стабилизатором мембран [Биленко М.В, 1989; Udupi V., 1992; Trudel S. et al., 2009]. Он защищает клеточные структуры нейтрофилов от высокотоксичного OCI-, производимого МПО [Carr А.С, Winterbourn С.С, 1997], при этом GSH превращается в глутатион-сульфонамид и дегидроглутатион [Harwood D.T. et al., 2006]. Связывая NO, глутатион образует токсичные для клетки нитрозильные комплексы. Моно нитрозоглутатион может активировать апоптоз [Turpaev К.Т. et al., 1997].
Не всегда восстановительного потенциала GSH достаточно для полной нейтрализации прооксидантов. Существует мнение, что взаимодействие GSH с органическими радикалами эффективно только в условиях удаления
, поэтому глутатион образует с супероксиддисмутазой своеобразную антиоксидантную систему, ибо в противном случае развиваются реакции образования H2O2 и GS· [Панкин В.З. и соавт., 1997; Меньшикова Е.Б. и соавт., 2006]. В сочетании с витамином B12 глутатион, а также N-ацетилцистеин, могут потенцировать прооксидантное и цитотоксическое действие на клетку [Соловьева М.Е. и соавт., 2007].
Основной антиоксидантный эффект GSH реализует посредством участия в работе ферментов. Глутатион выступает донором водорода при восстановлении H2O2 и перекисей липидов глутатион-пероксидазами и глутатион-S-трансферазами (ГТ) [Hirayama К., 1989; Sies Н. et al., 1997; Кулинский В.И., 1990; Hayes J.D. et al., 2005; Зенков H.K.. и соавт., 2009; Liu G. et al., 2010]. Высокая активность глутатион-редуктазы и накопление GSH оказывает протекторный эффект в отношении альвеолярных макрофагов, инкубируемых с прооксидантами in vitro [Pietarinen Р.К. 1995].
С изменением окислительно-восстановительного баланса сопряжено большое количество реакций, поэтому поддержание оптимального редокс-состояния цитозоля выступает важным условием нормальной жизнедеятельности клеток. Высокая концентрация глутатиона в цитоплазме, его редокс-активность и возможность поддержания в восстановленном состоянии делают систему GSH/GSSG важнейшим внутриклеточным редокс-буфером [Reed М.С. et al., 2008]. Концентрация GSH в клетке в 500-1000 раз превышает уровень НАДФН и других внутриклеточных редокс-систем, поэтому изменения соотношения GSH/GSSG прямо отражают изменения редокс-статуса клетки [Кулинский В.И., 2007; Asian М., Canatan D., 2008; Reed М.С., 2008]. Считают, что буферная емкость системы глутатиона защищает репликативную систему клетки, а дефицит GSH в условиях высокой генерации АФК приводит к снижению синтеза ДНК и белков [Poot М., 1991; Ланкин В.З., 1997; Day R.M., Suzuki Y.J., 2005; Liu G. et al., 2010].
Исходя из вышесказанного, целесообразно создание модели перекисного окисления липидов путем предварительной обработки лимфоцитов перекисью водорода в конечной концентрации 0,5 мМ с последующим определением белково-связанного глутатиона (табл.1), а также окисленного и восстановленного глутатиона (табл.2). Конечная концентрация перекиси водорода подобрана экспериментально. Лимфоциты обрабатывались перекисью водорода в конечной концентрации 0,3 мМ, 0,5 мМ, 1 мМ, 2 мМ (таблица 1, табл.2) Следовательно, комплексная модернизация способа-прототипа позволяет повысить точность создания модели перекисного окисления лимфоцитов.
Каждый вновь введенный в формулу изобретения признак выполняет функцию повышения точности и эффективности способа: дополнительная обработка лимфоцитов перекисью водорода в конечной концентрации 0,5 мМ, подобранная экспериментально для последующего определения белково-связанного глутатиона.
Защита лимфоцитов от продуктов перекисного окисления осуществляется антиоксидантами. Общепринятой номенклатуры антиоксидантов в настоящее время не существует, хотя ряд авторов [Dimascio Р., 1990; Kalra V. et al., 2001; Зайцев В.Г. и соавт., 2003] выделяет два класса: превентивные, снижающие скорость инициации цепной реакции окисления, и гасящие (прерывающие цепь), препятствующие развитию цепной реакции. К превентивным относят каталазу и пероксидазы, разрушающие ROOH, а также агенты, образующие хелатные комплексы с металлами переменной валентности, к прерывающим цепь - фенолы, ароматические амины. В условиях in vivo главными гасящими антиоксидантами являются витамин Е, нейтрализующий ROO· в липидной фазе мембран [Jore D. et al., 1990; Hong J.H. et al., 2004], фермент СОД, улавливающий
в водной фазе клетки [Fridovich 1., 1989; Dimascio P., 1990; CiureaD., 1992], и церулоплазмин - белок острой фазы, выполняющий антирадикальную функцию в крови [Marklund S.L., 1987; AtanasiuRL. et al., 1998].
Более известно деление антиоксидантов на ферменты и соединения неферментативной природы. Последние в определенных концентрациях всегда присутствуют в липидной фазе мембран и водных средах организма и расходуются первыми при устранении проявлений окислительного стресса [Droge W., 2002; Blokhina О., 2003]. Ферменты наиболее активно присоединяется к АОЗ после включения механизмов индукции [Лущак В.П., 2001]. При возникновении ОС расход антиоксидантов возрастает и меняется экспрессия генов, кодирующих белковые компоненты АОЗ [Дубинина Е.Е., 2006]. Между ферментами и неферментативными элементами АОЗ существует равновесие, причем последние при ряде патологических состояний организма могут выступать в качестве прооксидантов [Зенков Н.К. и др., 2001].
Главную роль среди неферментативных антиоксидантных систем защиты отводят глутатиону.
В настоящее время в лабораторной практике наиболее распространен способ оценки эффективности антиоксидантной активности с помощью определения глутатиона.
Содержание глутатиона определяют методом, предложенным М.Е. Anderson (1985) в модификации S. Kojima et al. (2004) [Kojima, S. Low dose gamma-rays activate immune functions via induction of glutathione and delay tumor growth / S. Kojima, K. Nakayama, H. Ishida // J. Radiat. Res. - 2004. - Vol.45, №1. - P.33-39.]. Принцип метода основан на взаимодействии GSH с 5,5'-дитио-бис(2-нитробензойной) кислотой (ДТНБ) с образованием тио-2-нитробензойной кислоты, водный раствор которой имеет максимум поглощения при длине волны 412 нм. При этом образуется GSSG, который восстанавливается глутатионредуктазой до GSH и вновь взаимодействует с ДТНБ. Скорость образования окрашенного продукта пропорциональна содержанию общего глутатиона. Для определения содержания GSSG пробы прединкубируются с блокатором SH-групп 2-винилпиридином («Wako», Япония), который необратимо связывает GSH, и, следовательно, скорость образования окрашенного продукта пропорциональна содержанию GSSG.
Лизат лимфоцитов крови готовят на 5% сульфосалициловой кислоте, которая осаждает белки, но не ингибирует активность глутатионредуктазы. Количество общего глутатиона (GSH и GSSG) определяют в пробе, содержащей 0,1 М Na-фосфатный буфер (pH 7,5) с 1 мМ ЭДТА, 0,4 мМ НАДФН2, 0,3 мМ ДТНБ и 1 U/мл глутатион-редуктазы («Wako», Япония). При измерении уровня GSSG супернатант предварительно инкубируют 30 мин с 10 мМ 2-винилпиридином, после чего определяют скорость реакции в описанной выше инкубационной среде. Расчет содержания общего и окисленного глутатиона производят с помощью калибровочных графиков, для построения которых растворы GSH и GSSG («Sigma», США) в концентрации от 3 до 100 мкМ обрабатывают аналогично опытным пробам. Концентрацию GSH рассчитывают как разницу между концентрацией общего глутатиона и GSSG, выражая результат в нмоль/мг белка.
Концентрацию белка в лимфоцитах крови определяют методом [A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding / M.M. Bradford // Analyt. Biochem. - 1976. - Vol.7, №1, 2. - P. 248-254.], основанным на взаимодействии Кумасси голубого G-250 с остатками аргинина и лизина в белках. Свободный краситель красного цвета (максимум поглощения - 495 нм) при образовании комплекса с белком переходит в синюю форму (максимум поглощения - 595 нм).
К 0,1 мл лизата лимфоцитов крови добавляют 1,0 мл раствора Кумасси голубого (100 мг красителя, 50 мл 96° этанола, 100 мл 85% Н3РО4, Н2О до 1,0 л), перемешивают, инкубируют 3 мин при комнатной температуре и измеряют оптическую плотность проб (длина волны 595 нм) против контроля, содержащего 0,1 мл воды и 1,0 мл раствора Кумасси голубого. Содержание белка рассчитывают по калибровочной кривой, построенной по разведениям стандартного раствора альбумина (1,0 мг/мл) и выражают в мг/мл.
Функционирование лимфоцитов связано с уровнем белково-связанного глутатиона. Определение белково-связанного глутатиона основано на способности боргидрата натрия (NaBH4) высвобождать из связи с белками глутатион, который при взаимодействии с ДТНБ образует окрашенное соединение, а именно тио-2-нитробензойную кислоту, водный раствор которой имеет максимум поглощения при длине волны 412 нм [Burchill, B.R. Microtubule dynamics and glutathione metabolism in phagocytizing human polymorphonuclear leukocytes [Text] / B.R. Burchill, J.M. Oliver, C.B. Pearson et al. // J. of Cell Biology. - 1978. - Vol.76, №2. - P. 439-447.].
В настоящее время крайне важно оценить эффективность модели перекисного окисления липидов мембран лимфоцитов с последующим определением белково-связанного глутатиона. Для решения этой задачи предложен новый способ оценки эффективности модели перекисного окисления липидов мембран лимфоцитов после дополнительной предварительной обработки лимфоцитов перекисью водорода в конечной концентрации 0,5 мМ.
Все сказанное свидетельствует о крайней важности разработки способа оценки эффективности модели перекисного окисления липидов мембран лимфоцитов для оценки их функциональной активности.
Популярность указанного выше способа обоснована его высокой чувствительностью, простотой осуществления и достаточной адекватностью получаемых результатов, лежащих в основе определения белково-связанного глутатиона.
Существенные признаки, характеризующие изобретение, проявили в заявленной совокупности новые свойства, явным образом не вытекающие из уровня техники в данной области, и не являются очевидными для специалиста.
Идентичной совокупности признаков не обнаружено при изучении патентной и научной медицинской литературы.
Данное изобретение может быть использовано в медицинской практике для повышения точности оценки функции лимфоцитов при окислительном стрессе и при различных заболеваниях. Таким образом, следует считать предлагаемое изобретение соответствующим условиям патентоспособности: «новизна», «изобретательский уровень», «промышленная применимость».
Метод основан на определении восстановленного глутатиона.
Способ осуществляется следующим образом поэтапно.
1. Выделение лимфоцитов крови из венозной крови.
Пробирки с венозной гепаринизированной кровью (25 Ед/мл) выдерживали при температуре 37°C в течение 40 минут для отделения плазмы и эритроцитов. Затем пробирки переносят в стерильный ламинарный шкаф для выполнения процедуры выделения лимфоцитов крови. Полученную плазму наслаивают на градиент плотности Ficoll-Paque («Pharmacia», Швеция) (ρ=1,077 г/см3) в соотношении 1:2 и центрифугируют при 500 g в течение 20 минут [Bignold L.P., 1987]. После центрифугирования собирают образовавшееся интерфазное кольцо из смеси мононуклеарных клеток в стерильную центрифужную пробирку с 4,5 мл питательной среды ((90% RPMI-1640 («Вектор-Бест», Россия), 10% эмбриональной телячьей сыворотки («Биолот», Россия), инактивированной при температуре 56°C в течение 30 мин, 0,3 мг/мл L-глутамина («Борисовский ЗМП», Беларусь), 100 мкг/мл гентамицина, 2 ммоль/мл Hepes («Flow», Великобритания)), затем центрифугируют 10 минут при 500 g. Процедуру отмывки повторяют дважды: последовательно ресуспендируя клетки и затем центрифугируя в течение 10 минут при 500 g. Выделение лимфоцитов из мононуклеарной фракции клеток проводят на двойном градиенте Перколла [Ulmer A.J., 1979]. Стандартный изоосмотический раствор Перколла (SIP) получают смешиванием одного объема 10х PBS (фосфатно-солевого буфера (pH 7,4)) с девятью объемами Перколла («Sigma», США) (плотность полученного раствора - 1,130 г/мл). Затем готовят 47,5% стандартный изоосмотический раствор Перколла (47,5%о SIP) и 15,0%) стандартный изоосмотический раствор Перколла (15,0% SIP). К клеточной суспензии добавляют 1,5 мл SIP (4°C), перемешивают и переносят в новую стерильную пробирку. Сверху наслаивают 5 мл 47,5% SIP (4°C). Создают верхнюю фазу посредством 2 мл 15,0% SIP (4°C). Центрифугируют при 1500 g и 4°C 45 минут. Собирают интерфазное кольцо (лимфоцитарную фракцию клеток). Объем доводят до 5 мл питательной средой ((90% RPMI-1640 («Вектор-Бест», Россия), 10% эмбриональной телячьей сыворотки («Биолот», Россия), инактивированной при температуре 56°C в течение 30 мин, 0,3 мг/мл L-глутамина («Борисовский ЗМП», Беларусь), 100 мкг/мл гентамицина, 2 ммоль/мл Hepes («Flow», Великобритания)), температура раствора должна соответствовать 37°C. Далее проводят центрифугирование при 700 g и 20°C в течение 10 минут. Затем удаляют супернатант до конечного объема 1 мл.
2. Количественное определение численности жизнеспособных лимфоцитов крови с помощью окраски трипановым синим микроскопическим методом.
Лимфоциты крови ресуспендируют в 1 мл клеточной взвеси. Отбирают 100 мкл ресуспендированной клеточной суспензии и добавляют 100 мкл 0,1% раствора трипанового синего на физ. растворе, хорошо перемешивают и заполняют камеру Горяева. Предварительно к камере притирают покровное стекло так, чтобы появлялись радужные ньютоновые кольца (только при этих условиях соблюдался правильный объем камеры). Каплю клеточной взвеси с красителем вносят под притертое покровное стекло. Подсчет клеток производят в 5 больших квадратах по диагонали камеры Горяеева. Расчет клеточности лимфоцитов крови производят по формуле
А×106=(число клеток)/4
где А - клеточность лимфоцитов крови.
3. Методика создания модели перекисного окисления
В среду культивирования лимфоцитов добавляют перекись водорода в конечной крнцентрации 0,5 мМ.
4. Биохимическое исследование белково-связанного, восстановленного и окисленного глутатиона.
Лизат лимфоцитов готовят на 5% сульфосалициловой кислоте, которая осаждает белки, но не ингибирует активность глутатионредуктазы. Количество общего глутатиона (GSH и GSSG) определяют в пробеЮ содержащей 0,1 М Na-фосфатный буфер (pH 7,5) с 1 мМ ЭДТА, 0,4 мМ НАДФН2, 0,3 мМ ДТНБ и 1 U/мл глутатионредуктазы («Wako». Япония). Окисленный глутатион определяют аналогичным способом в клеточном лизате после предварительной инкубации пробы в течение 30 мин с 10 мМ 2-винилпиридином. Расчет содержания общего и окисленного глутатиона производят с помощью калибровочных графиков, для построения которых используют растворы GSH и GSSG («МР», США) в концентрации от 3 до 100 мкМ, обработанные аналогично опытным пробам. Концентрацию GSH рассчитывают как разницу между концентрацией общего глутатиона и GSSG, выражая результат в нмоль/мг белка. Определение белково-связанного глутатиона.
После инкубации клетки центрифугируют 5 минут при 4°C и 1500 об/мин для их осаждения. Удаляют супернатант. Добавляют 1 мл охлажденного PBS (рН 7,4). Ресуспендируют на вортексе. Центрифугируют 5 минут при 4°C и 1500 об/мин. Удаляют супернатант. Осадок клеток ресуспендируют в 1 мл 5% сульфосалициловой кислоты для получения клеточного лизата. Центрифугируют при 3000 об/мин 10 мин. 1,0 мл осадка белка инкубируют 1 ч при 50°C с 1,0 мл 1% NaBU4. Далее оставшийся белок осаждают добавлением 0,4 мл 30% ТХУ. Пробу инкубируют 15 мин при 50°C. Затем пробу охлаждают 5 мин (0°C). Центрифугируют 10 мин при 3000 об/мин. Супернатант смешивают с 2,5 мл PBS (рН 7,4) и добавляют 2,0 мл ацетона для полного окисления NaBH4. Перемешивают. Центрифугируют 10 мин при 3000 об/мин. Удаляют верхнюю фазу. К нижней фазе добавляют равный объем диэтилового эфира (для удаления ТХУ). Перемешивают. Центрифугируют 10 мин при 3000 об/мин. Затем снова удаляют верхнюю фазу. Далее процедуру отмывки пробы от ТХУ с помощью диэтилового эфира производят четырехкратно. Затем отбирают 0,1 мл жидкости (из нижней фазы) и смешивают с 0,4 мл 0,01 М фосфатного буфера (рН 7,0). В пробу добавляют 0,1 мл 0,4 мг/мл ДТНБ. Пробу спектрофотометрируют при 412 нм против контроля, содержащего воду вместо раствора осажденного белка.
Расчет производят с учетом коэффициента молярной экстинкции 13·103 М-1 см-1. Результаты определения концентрации белково-связанного глутатиона выражают в нмоль/мг белка.
Исследование антиоксидантной активности по способу-прототипу и предлагаемому способу выполнялось 40 раз. Результаты исследования обработаны статистически с использованием пакета программ Stat Soft Statistica 6.0.
При проведении исследования по способу-прототипу уровень белково-связанного глутатиона после инкубации лимфоцитов возрастал в 1,5 раза и менее, а при оценке эффективности модели перекисного окисления липидов мембран лимфоцитов по предлагаемому способу в случае эффективно созданной модели перекисного окисления липидов мембран лимфоцитов уровень белково-связанного глутатиона возрастал в 6,2±0,5 раза, а в случае неэффективно созданной модели перекисного окисления липидов мембран лимфоцитов уровень белково-связанного глутатиона возрастал в 2,0±0,2 раза (табл.3).
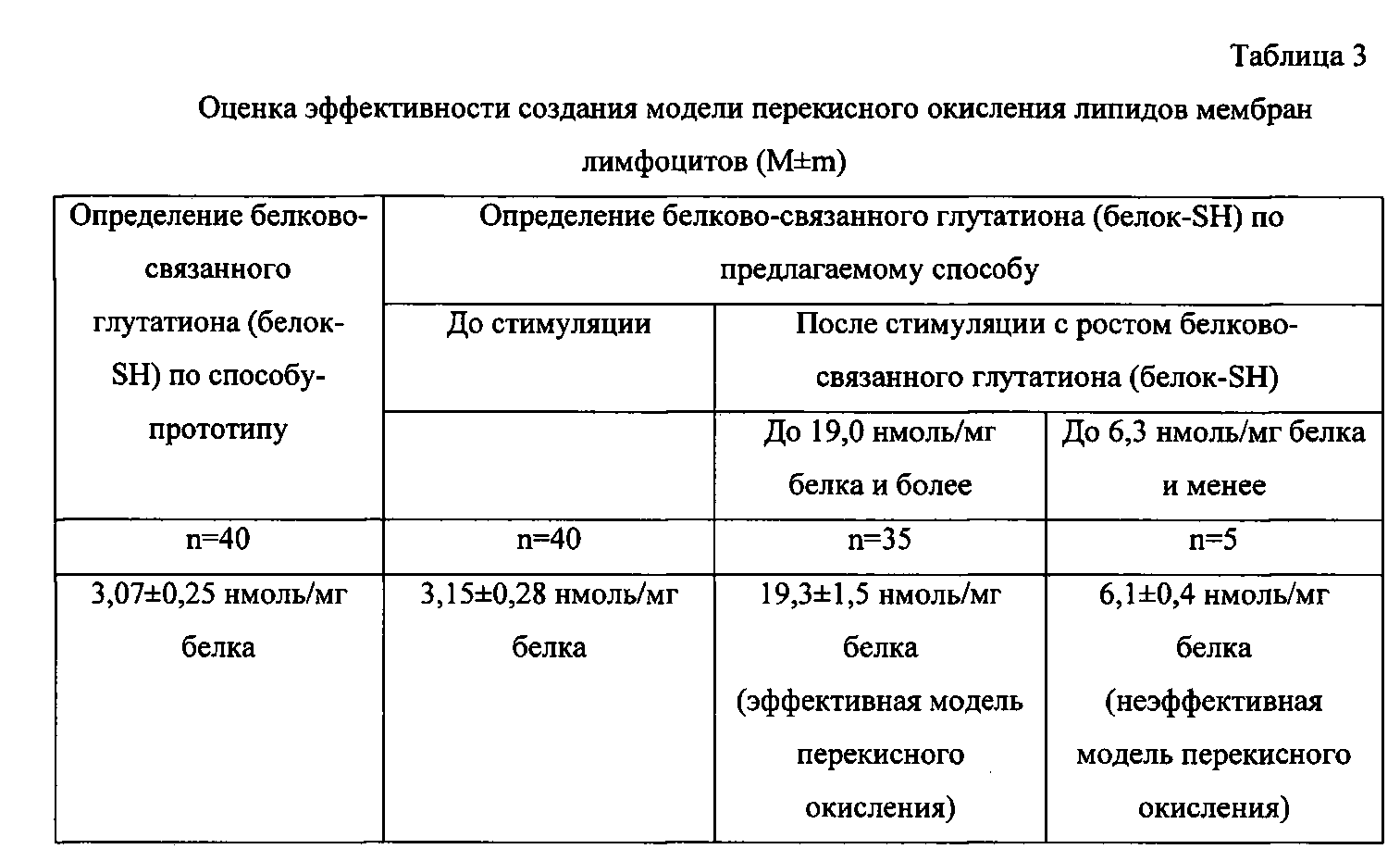
Полученные результаты уровня белково-связанного глутатиона соответствуют данным литературы [Смирнова Г.В., Октябрьский О.Н., 2005].
Итак, при применении способа-прототипа был получен недостаточно точный результат, не позволивший оценить эффективность модели перекисного окисления липидов мембран лимфоцитов, что связано с отсутствием биохимической стимуляции процесса перекисью водорода в конечной концентрации 0,5 мМ, а наиболее эффективным и точным был предлагаемый способ.
При этом предлагаемый способ прост в исполнении и интерпретации полученных результатов.
Способ оценки эффективности модели перекисного окисления липидов мембран лимфоцитов путем определения белково-связанного глутатиона, отличающийся тем, что предварительно обрабатывают лимфоциты перекисью водорода в конечной концентрации 0.5 мМ и при увеличении уровня белково-связанного глутатиона в 6 раз и более считают модель эффективной, а при увеличении уровня белково-связанного глутатиона в 2 раза и менее считают неэффективной.Способ иммобилизации биомолекул на поверхности магнитоуправляемых наночастиц железа покрытых углеродной оболочкой
Интерференционный переключатель резонансного свч компрессора
Устройство для измерения температуры
Вакуумно-дуговой генератор с жалюзийной системой фильтрации плазмы от микрочастиц
Способ оценки эффективности стимуляции антиоксидантной активности
Способ прогнозирования течения липидемии
Способ диагностики значимых коронарных стенозов у пациентов с нарушениями миокардиальной перфузии по данным однофотонной эмиссионной компьютерной томографии миокарда с 99mtc-технитрилом
Комплексный препарат для профилактики и лечения кишечных инфекций
Способ прогнозирования течения ишемической болезни сердца
Способ подземной газификации
Сверхпроводящий размыкатель
Способ оценки степени метаболической и кардиореспираторной адаптации кардиохирургических больных
Способ лечения резистентной артериальной гипертонии
Способ оценки вероятности пристеночного тромбоза правых камер сердца - правого предсердия и правого желудочка
Средство, увеличивающее устойчивость сердца к ишемическим и последующим реперфузионным повреждениям
Способ получения порошка нитрида титана
Переставная опалубка из объемных блоков на несущих металлических трубах (варианты)
Способ получения гранулированной алюминиевой пудры
Устройство для подрезания блоков горных пород высоковольтными разрядами
Способ определения осмия инверсионно-вольтамперометрическим методом в природном и техногенном сырье на графитовом электроде, модифицированном золотом

