Результат интеллектуальной деятельности: СИНТЕТИЧЕСКИЕ 5 UTR (НЕТРАНСЛИРУЕМЫЕ ОБЛАСТИ), ЭКСПРЕССИОННЫЕ ВЕКТОРЫ И СПОСОБ ПОВЫШЕНИЯ ТРАНСГЕННОЙ ЭКСПРЕССИИ
Вид РИД
Изобретение
Область изобретения
Это изобретение относится к области биотехнологии. В частности, относится к повышению посттранскрипционного контроля экспрессии гена в эукариотических клетках.
Уровень техники
После транскрипции первичных мРНК с ДНК экспрессия эукариотических генов проходит несколько этапов контроля. Первичный транскрипт мРНК включает в себя кодирующие (экзоны) и некодирующие (интроны) зоны. Во время сплайсинга мРНК интроны транскрипта вырезаются и удаляются, а экзоны соединяются вместе, образуя зрелую матричную РНК (мРНК). Сплайсинг служит контрольным этапом в продуцировании множественных изоформ белка отдельного гена посредством дополнения и удаления экзонов в разных комбинациях. Этот процесс, называемый альтернативным сплайсингом, происходит в четко регулируемых, многокомпонентных структурах, называемых сплайсосомами, которые находятся под управлением внутриклеточных и внеклеточных сигнальных путей.
Альтернативный сплайсинг внутри кодирующей белок зоны приводит к образованию сложных изоформ с разнообразными функциями. Кроме того, было показано, что сплайсинг значительно увеличивает синтез белков в клетках млекопитающих (Huang и Gorman, 1990. Nucleic Acids Research 18(4):937-947). Механизм этого процесса неизвестен. Альтернативный сплайсинг также может происходить в нетранслируемых зонах транскрипта, что может содействовать энхансеру или стабилизации доменов в конечном транскрипте, в результате чего увеличивается трансляция белка.
Было показано, что включение элементов сплайсинга в 5'-регуляторную область синтетической конструкции гена увеличивает экспрессию генов, гипотетически в связи с улучшением транспорта мРНК из ядра в цитоплазму (см. выше Huang и Gorman; Choi et al., 1991. Molecular and Cellular Biology 11(6):3070-3074). В результате этого процесса интроны часто включают между промотором и сайтом множественного клонирования коммерчески доступных экспрессивных векторов млекопитающих. Однако комбинации интронов с другими регуляторными зонами в целях увеличения экспрессии генов не были исследованы.
Краткое описание изобретения
В настоящем изобретении предложены синтетические полинуклеотидные последовательности 5'UTR, которые предназначены для увеличения экспрессии трансгенного компонента синтетической конструкции гена в клетке-хозяине. Будучи не связаны с теорией, синтетические 5'UTR области сконструированы таким образом, чтобы экспрессия трансгена могла быть повышена за счет увеличения транспорта и стабильности РНК.
Синтетические последовательности 5'UTR содержат полинуклеотидный фрагмент, который включает сайт сплайсинга, одного эукариотического гена, соединенный с полинуклеотидным фрагментом, кодирующим последовательность 5'UTR, другого эукариотического гена, который стабилен на уровне РНК и белка. В одном из вариантов осуществления изобретения синтетическая последовательность 5'UTR является химерной последовательностью, содержащей полинуклеотидный фрагмент, включающий сайт сплайсинга гена кальциевой АТФазы сакроплазматического/эндоплазматического ретикулума, и полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5'UTR гена казеина.
Синтетические полинуклеотидные последовательности 5'UTR согласно изобретению применяли в целях увеличения экспрессии интересующей последовательности или интересующей кодирующей зоны внутри синтетической конструкции гена. С помощью технологии рекомбинантной ДНК синтетическую последовательность 5'UTR можно включить в вирусные или невирусные векторы между промотором и интересующей нуклеотидной последовательностью. Синтетические последовательности 5'UTR можно расположить по концам нуклеотидных последовательностей, имеющих сайты эндонуклеаз рестрикции и другие нуклеотиды, необходимые для активации эндонуклеаз рестрикции. Фланкирующие последовательности могут содержать сайты клонирования внутри вектора.
В настоящем изобретении также предложены векторы, содержащие синтетические 5'UTR области. В одном из вариантов осуществления изобретения вектор является эукариотическим экспрессионным вектором.
В настоящем изобретении предложены также способы увеличения трансгенной экспрессии в эукариотической клетке. Указанные способы включают этапы создания синтетической последовательности 5'UTR путем слияния полинуклеотидного фрагмента одного гена эукариотической клетки, содержащего сайт сплайсинга, и полинуклеотидного фрагмента другого гена эукариотической клетки, содержащего, по меньшей мере, фрагмент 5'UTR, с целью получить химерную полинуклеотидную последовательность, и этапы включения химерной полинуклеотидной последовательности внутрь экспрессионного вектора между промотором и интересующей последовательностью.
Заявителем было неожиданно обнаружено, что синтетическая последовательность 5'UTR, созданная путем соединения полинуклеотидного фрагмента, содержащего интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, с полинуклеотидным фрагментом, содержащим, по меньшей мере, фрагмент гена казеина, в результате ведет к увеличению экспрессии гена. Как подробно описано в настоящей заявке, два разных варианта синтетической 5'UTR увеличивают экспрессию репортерного гена по сравнению с контролем в двух разных типах клеток, трансфицированных экспрессионным вектором, содержащим синтетическую 5'UTR.
Таким образом, один объект изобретения состоит в применении синтетической последовательности 5'UTR, которая содержит полинуклеотидный фрагмент, включающий сайт сплайсинга, соединенный с полинуклеотидным фрагментом, содержащим, по меньшей мере, часть гетерологической 5' нетранслируемой области для того, чтобы увеличить трансгенную экспрессию в эукариотической клетке.
Другой объект изобретения состоит в применении синтетической последовательности 5'UTR, которая содержит полинуклеотидный фрагмент, включающий интрон, соединенный с полинуклеотидным фрагментом, содержащим, по меньшей мере, часть гетерологической 5' нетранслируемой области для того, чтобы увеличить трансгенную экспрессию в эукариотической клетке.
Еще один объект изобретения состоит в применении синтетической последовательности 5'UTR, которая содержит полинуклеотидный фрагмент, включающий интрон, который имеет прилегающие с 3'- и 5'-концов фрагменты близлежащих экзонов, соединенный с полинуклеотидным фрагментом, содержащим, по меньшей мере, часть гетерологической 5' нетранслируемой области для того, чтобы увеличить трансгенную экспрессию в эукариотической клетке.
Другой объект изобретения предлагает синтетическую последовательность 5'UTR, совместимую с включением в вектор.
Другой объект изобретения предлагает векторы, содержащие синтетические 5'UTRs.
Другой объект изобретения предлагает клетки-хозяева, содержащие синтетические 5'UTR области.
Описание последовательностей
SEQ ID NO:1 представлена вариантом синтетической последовательности 5'UTR, содержащей: сайт рестрикции MluI, SEQ ID NO:2, сайт рестрикции KpnI, SEQ ID NO:3, сайт рестрикции MfeI. SEQ ID NO:1 также упоминается в настоящей заявке как 5U2.
SEQ ID NO:2 представлена вариантом последовательности интрона 2 гена собаки SERCA2, мутированным предполагаемым консенсусным поли-А сайтом с частью экзона 2, расположенной в 5'-конце, и частью экзона 3, расположенной в 3'-конце. SEQ ID NO:2 является мутированной неполной последовательностью 26-й хромосомы Canis familiaris, полученной путем механического фрагментирования целого генома (общий номер доступа NC-06608.2).
SEQ ID NO:3 представлена вариантом последовательности 5'UTR бычьего казеина. SEQ ID NO:3 является неполной последовательностью полноразмерной мРНК бета-казеина Bos taurus (общий номер доступа NM_181008).
SEQ ID NO:4 представлена нормальным вариантом последовательности интрона 2 SERCA2 собаки с частью экзона 2, расположенной в 5'-конце, и частью экзона 3, расположенной в 3'-конце. SEQ ID NO:4 является неполной последовательностью 26-й хромосомы Canis familiaris и получена путем механического фрагментирования целого генома (общий номер доступа NC_006608.2).
SEQ ID NO:5 представлена нормальным вариантом последовательности интрона 2 SERCA2 человека с частью экзона 2, расположенной в 5'-конце, и частью экзона 3, расположенной в 3'-конце. SEQ ID NO:5 является неполной последовательностью 12-й хромосомы Homo sapiens, совокупность реперов, полная последовательность (общий номер доступа NC_000012).
SEQ ID NO:6 представлена нормальным вариантом последовательности интрона 2 SERCA2 мыши с частью экзона 2, расположенной в 5'-конце, и частью экзона 3, расположенной в 3'-конце. SEQ ID NO:6 является последовательностью 5-й хромосомы Mus musculus, совокупность реперов (общий номер доступа NC_000071).
SEQ ID NO:7 представлена синтетической последовательностью 5'UTR, содержащей сайт рестрикции AscI, сайт рестрикции MluI, SEQ ID NO:4, сайт рестрикции KpnI, SEQ ID NO:3, сайт рестрикции MfeI. SEQ ID NO:7 также упоминается в настоящей заявке как INXN-1.
SEQ ID NO:8 представлена вариантом последовательности 5'UTR казеина мыши. SEQ ID NO:8 является неполной последовательностью мРНК бета-казеина Mus musculus (кДНК клон MGC:91065) (общий номер доступа ВС080709).
SEQ ID NO:9 представлена вариантом последовательности 5'UTR казеина крысы. SEQ ID NO:9 является неполной последовательностью мРНК казеина Rattus norvegicus (Csn2) (общий номер доступа NM_017120).
SEQ ID NO:10 представлена вариантом 5'UTR последовательности казеина овцы. SEQ ID NO:10 является неполной последовательностью мРНК бета-казеина Ovis aries (CSN2) (общий номер доступа NM_001009373).
SEQ ID NO:11 представлена экзоном 3 SECRA2 собаки. SEQ ID NO:11 является неполной последовательностью 26-й хромосомы Canis familiaris и получена путем механического фрагментирования целого генома (общий номер доступа NC_006608).
SEQ ID NO:12 представлена вариантом векторной последовательности, содержащей синтетический 5'UTR. Вектор, представленный SEQ ID NO:12, включает SEQ ID NO:1 и схематично изображен на Фигуре 10.
SEQ ID NO:13 представлена другим вариантом векторной последовательности, содержащей синтетическую 5'UTR. Вектор, представленный SEQ ID NO:13, включает SEQ ID NO:7 и схематично изображен на Фигуре 11.
SEQ ID NO:14 представлена вектором, содержащим контрольную (поли-G) синтетическую 5'UTR, и схематично изображена на Фигуре 9.
В любой из этих последовательностей Т (тимидин) может быть замещен на U (урацил).
Описание фигур
На ФИГУРЕ 1А показано схематическое изображение полинуклеотида, имеющего последовательность SEQ ID NO:4.
На ФИГУРЕ 1В показано схематическое изображение полинуклеотида, имеющего последовательность SEQ ID NO:5, и полинуклеотида, имеющего последовательность SEQ ID NO:6.
На ФИГУРЕ 1C показаны полинуклеотиды, имеющие последовательности SEQ ID NO:2 и SEQ ID NOS:4-6. Второй интрон SECRA2 выделен черным цветом. Прилежащие экзоны или их части не окрашены.
На ФИГУРЕ 2А показано схематическое изображение полинуклеотида, имеющего последовательность SEQ ID NO:1.
На ФИГУРЕ 2В показано схематическое изображение полинуклеотида, имеющего последовательность SEQ ID NO:7.
На ФИГУРЕ 3А показано схематическое изображение синтетической 5'UTR, включенной в экспрессионный вектор между промотором и интересующей последовательностью.
На ФИГУРЕ 3В показано схематическое изображение синтетических 5'UTR, включенных в экспрессионный вектор между промотором и клонирующим сайтом.
На ФИГУРЕ 4 изображены результаты исследования синтетических 5'UTR вариантов осуществления изобретения в клетках НЕК-293.
На ФИГУРЕ 5 изображены результаты исследования синтетических 5'UTR вариантов осуществления изобретения в клетках 1080.
На ФИГУРЕ 6 изображены результаты исследований синтетических 5'UTR вариантов осуществления изобретения в виде кратного прироста относительно контроля в клетках НЕК-293 и клетках 1080, при этом контрольные значения приведены к 1.
На ФИГУРЕ 7 изображен контрольный вектор, используемый в Примере 1 (VVN-2712), при этом кодирующая последовательность бета-галактозидазы (LacZ) не имеет 5'UTR и функционально связана с CMV промотором.
На ФИГУРЕ 8 изображен контрольный вектор, используемый в Примере 1 (VVN-2713), при этом указанный вектор не имеет 5'UTR и LacZ.
На ФИГУРЕ 9 изображен вектор, используемый в Примере 1 (VVN-8318), при этом кодирующая последовательность бета-галактозидазы (LacZ) функционально связана с поли-G 5'UTR и CMV промотором.
На ФИГУРЕ 10 изображен вектор, используемый в Примере 1 (VVN-8277), при этом кодирующая последовательность бета-галактозидазы (LacZ) функционально связана с 5'UTR согласно изобретению (5U2) и CMV промотором.
На ФИГУРЕ 11 изображен вектор, используемый в Примере 1 (VVN-8276), при этом кодирующая последовательность бета-галактозидазы (LacZ) функционально связана с 5'UTR согласно изобретению (INXN-1) и CMV промотором.
На ФИГУРЕ 12 представлена таблица, содержащая данные Примера 1, изображенного на ФИГУРАХ 4-6.
На ФИГУРЕ 13 изображен участок линейной структуры геномной и мРНК последовательностей SECRA2 лошади домашней, содержащие второй интрон и экзоны 2 и 3. 5'- и 3'-концы второго интрона обозначены стрелками.
На ФИГУРАХ 7-11 были использованы следующие обозначения: про CMV=промотор цитомегаловируса, LacZ=LacZ кодирующая последовательность, SV40pA=SV40 поли-А, Amp=ген устойчивости к ампициллину, Neo=ген устойчивости к неомицину, MCS=сайт множественного клонирования, SPL-1=часть экзона 2 SECRA2+интрон 2 SECRA2+часть экзона 3 SECRA2, UTR-1=часть 5' UTR казеина.
Подробное описание изобретения
Следующие определения применимы ко всему описанию, фигурам и формуле изобретения. Однако термины, используемые в описании и формуле изобретения, которым не даны определения в настоящей заявке, имеют общепринятые значения, известные в данной области техники.
При использовании в этом описании терминов «один» имеется в виду значение «по меньшей мере, один» или «один или более», если не указано другое.
Термины «нуклеиновая кислота», «молекула нуклеиновой кислоты», «последовательность нуклеиновых кислот», «олигонуклеотид», « последовательность олигонуклеотида», «нуклеотидная последовательность», «полинуклеотид» и «полинуклеотидная последовательность» в настоящей заявке взаимозаменяемы и относятся к фосфатным эфирам полимерных форм рибонуклеозидов (аденозин, гуанозин, уридин или цитидин; «РНК молекулы») или дезоксирибонуклеозидов (дезоксиаденозин, дезоксигуанин, дезокситимидин или дезоксицитизин; «ДКН молекулы») или к другим их фосфоэфирным аналогам, таким как фосфотиоаты и тиоэфиры, в одноцепочечной форме либо в форме двухцепочечной спирали. Возможно существование двухцепочечных ДНК-ДНК, ДНК-РНК и РНК-РНК спиралей. Термин молекула нуклеиновой кислоты, в частности ДНК или РНК молекула, относится только к первичной или вторичной структуре молекулы и не касается других отдельных третичных форм. Таким образом, этот термин включает двойную спираль ДНК, в том числе, линейные или кольцевые ДНК молекулы (например, фрагменты рестрикции), плазмиды, сверхспирали ДНК и хромосомы. При описании структуры отдельных двухцепочечных молекул ДНК в настоящей заявке согласно принятому обозначению последовательность можно описать только как последовательность вдоль нетранскрибируемой цепи ДНК в направлении от 5'-к 3'-концу (то есть цепи, имеющей последовательность, гомологичную мРНК). «Рекомбинантная молекула ДНК» - это молекула ДНК, которая претерпевает молекулярно-биологическое преобразование. ДНК включает, но не ограничивается указанными, кДНК, геномную ДНК, ДНК плазмиды, синтетическую ДНК и полусинтетическую ДНК.
Термин «фрагмент», используемый применительно к полинуклеотидной последовательности (например, «полинуклеотидный фрагмент»), относится к нуклеотидной последовательности, длина которой уменьшена по отношению к длине рассматриваемой нуклеиновой кислоты и которая содержит помимо общей части нуклеотидную последовательность, идентичную рассматриваемой нуклеиновой кислоте. Такой фрагмент нуклеиновой кислоты согласно изобретению при необходимости можно включить в более длинный полинуклеотид, составной частью которого он будет являться. Согласно изобретению такие фрагменты содержат или, в качестве альтернативы, состоят из полинуклеотидов, длина которых находится в пределах, по меньшей мере, от 6, 8, 9, 10, 12, 15, 18, 20, 21, 22, 23, 24, 25, 30, 39, 40, 42, 45, 48, 50, 51, 54, 57, 60, 63, 66, 70, 75, 78, 80, 90, 100, 105, 120, 135, 150, 200, 300, 500, 720, 900, 1000 или 1500 следующих друг за другом нуклеотидов нуклеиновой кислоты.
Термин «химерный» означает содержание фрагментов, не встречающихся вместе в их естественном состоянии. Например, химерный полинуклеотид является полинуклеотидом, содержащим фрагменты, не граничащие друг с другом в их естественном состоянии.
Термин «синтетический», используемый применительно к полинуклеотидной последовательности, относится к ненатуральному полинунуклеотиду (или части полинуклеотида), который отличается от нормальной полинуклеотидной последовательности. Например, синтетический ген (или часть гена) может содержать одну или более последовательностей нуклеиновых кислот, в природе не расположенных совместно друг с другом (химерные последовательности), и/или могут включать замещения, инсерции, делеции и их комбинации.
Термин «ген» относится к полинуклеотиду, содержащему нуклеотиды, которые кодируют функциональную молекулу (например, полипептид или РНК), и включает кДНК или нуклеиновые кислоты геномной ДНК. В общем смысле следует понимать, что геномная ДНК, кодирующая полипептид или РНК, содержит некодирующие зоны (то есть интроны), которые вырезаются из зрелой мРНК, и, следовательно, не представлены в кДНК, кодирующей тот же полипептид или РНК. «Ген» может содержать фрагмент из нуклеиновых кислот, который экспрессирует специфическую РНК, белок или полипептид. Дополнительно, «ген» может содержать регуляторные последовательности, предшествующие (5' некодирующие последовательности) и следующие за (3' некодирующие последовательности) кодирующей последовательностью. «Ген» может также содержать триплекс-образующие олигонуклеотиды (TFO). Термин «нативный ген» относится к гену, существующему в природе с его собственными регуляторными последовательностями. «Химерный ген» или «рекомбинантный ген» относится к любому ненативному гену, содержащему регуляторные и/или кодирующие последовательности, которые не встречаются в природе совместно. Следовательно, химерный ген может содержать регуляторные последовательности и кодирующие последовательности, полученные из разных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные в порядке, отличающемся от естественного. «Эндогенный ген» относится к нативному гену в его естественном местоположении в геноме организма.
«Чужеродный» ген, «экзогенный» ген, или «гетерологичный» ген, или «трансген» относится к гену, который обычно не присутствует в клетке-хозяине или организме, но который вводят в клетку-хозяин или организм путем переноса гена. Трансгены могут содержать как нативные гены, включенные в ненативный организм, так и химерные или синтетические гены. Также трансген может быть версией кДНК эндогенного гена. Также трансген может быть немутированным вариантом мутированного эндогенного гена или мутированной версией немутированного эндогенного гена. Также трансген может быть терапевтическим геном или опытным геном, таким как репортер. Трансген может быть введен напрямую в клетки-мишени организма-хозяина или опосредованно через перенос трансформированных клеток, например аутологичных клеток, в организм хозяина.
Под термином «5'-концевая нетранслируемая область» или «5'UTR» гена следует понимать часть гена, которая транскрибируется в первичный транскрипт РНК (пре-мРНК) и расположена перед кодирующей последовательностью. Полученный путем транскрипции ДНК первичный транскрипт является начальным продуктом РНК, содержащим интроны и экзоны. Большое количество транскриптов должны подвергнуться РНК-процессингу для того, чтобы сформировать физиологически активные типы РНК. Процессинг зрелой мРНК может включать тримминг концов, удаление интронов, кэпирование и/или вырезание отдельных молекул рРНК из их РНК-предшественников. 5'UTR область мРНК, следовательно, представляет собой ту часть мРНК, которая не транслируется в белок и расположена перед кодирующей последовательностью. В геномной последовательности 5'UTR обычно определяется как область между сайтом инициации транскрипции и инициирующим кодоном. 5' нетранслируемые области (5'UTRs) мРНК позвоночных могут содержать от нескольких десятков до нескольких сотен оснований в длину (Crowe et al., 2006. BMC Genomics 7:16).
«Синтетическая 5'UTR» - это искусственная 5'UTR, отличающаяся от нормальной полинуклеотидной последовательности 5'UTR. Синтетическая 5'UTR может содержать одну или более последовательностей нуклеиновых кислот, в природе не расположенных совместно друг с другом (химерные последовательности), и/или может включать замещения, инсерции, делеции и их комбинации.
«Граница сплайсинга», «интрон-экзонная граница сплайсинга» или «сайт сплайсинга» - это области на границах интрона в эукариотических пре-мРНК, распознаваемые клеточными аппаратами сплайсинга, где два близлежащих экзона соединяются, а интрон удаляется. Сайты сплайсинга представлены консервативными последовательностями на 5'- и 3'-границах интрон/экзон. Для подавляющего большинства интронов наиболее консервативными последовательностями являются GU, ограничивающая 5'-конец, и AG с 3'-конца. Однако также известны исключения среди этих типичных последовательностей, такие как интроны с AU-AC сайтами сплайсинга. 5'-концевой сайт сплайсинга на границе интрон/экзон известен как донорный сайт сплайсинга. 3'-концевой сайт сплайсинга на границе интрон/экзон известен как акцепторный сайт сплайсинга.
«Сплайсосома» - это большой рибонуклеопротеиновый комплекс, который служит в качестве клеточного аппарата сплайсинга. Сплайсосомы состоят из субъединиц малых ядерных рибонуклеопротеинов (мяРНП), которые собираются на пре-мРНК. Сами мяРНП состоят из малых ядерных РНК (мяРНК) и нескольких протеиновых субъединиц. Во время процесса сплайсинга распознавание сайтов сплайсинга внутри пре-мРНК осуществляется путем спаривания оснований с мяРНК.
«Гетерологичную» ДНК относят к ДНК, которая обычно не присутствует в клетках или хромосомных сайтах клетки. Следовательно, гетерологичная ДНК включает чужеродный клетке ген. «Гетерологичная» ДНК также может включать ген, обычно присутствующий в клетке, но находящийся в неестественном местоположении. Более того, «гетерологичной» молекулой ДНК может быть молекула ДНК, содержащая чужеродный сегмент ДНК, функционально связанный с сегментом ДНК хозяина, например с промотором транскрипции. С другой стороны, гетерологичная молекула ДНК может содержать эндогенный ген, функционально связанный с экзогенным промотором. Далее, «гетерологичную» ДНК можно отнести к ДНК-молекуле или фрагменту, которые получены из гена, не имеющего общего эволюционного происхождения с рассматриваемой ДНК-молекулой или фрагментом.
Термин «геном» включает хромосомную, а также митохондриальную, хлоропластную и вирусную ДНК или РНК.
Термин «зонд» относится к одноцепочечной молекуле нуклеиновой кислоты, которая может гибридизоваться с целевой комплементарной одноцепочечной молекулой нуклеиновой кислоты, образуя двухцепочечную молекулу.
«Кодирующая последовательность» ДНК относится к двухцепочечной последовательности ДНК, которая кодирует полипептид и которая может быть транскрибирована и транслирована в полипептид в клетке in vitro или in vivo или внеклеточно, например, в пробирке, когда эта последовательность контролируется соответствующими регуляторными последовательностями. «Соответствующие регуляторные последовательности» относятся к нуклеотидным последовательностям, расположенным до (5' некодирующие последовательности), внутри или после (3' кодирующие последовательности) кодирующей последовательности, и которые влияют на транскрипцию, процессинг или стабильность РНК или трансляцию ассоциированной кодирующей последовательности. Регуляторные последовательности могут включать промоторы, лидерные последовательности трансляции, интроны, последовательности распознавания полиаденилирования, сайт процессинга РНК, центр связывания эффектора и структуру типа «стебель-петля». Границы кодирующей последовательности определены инициирующим кодоном в 5'(амино)-конце и стоп-кодоном трансляции в 3'(карбоксильном)-конце. Кодирующая последовательность может включать, но не ограничивается указанными, эукариотические, прокариотические и химерные последовательности, кДНК с мРНК, последовательности геномной ДНК и даже синтетические последовательности ДНК.
«Открытая рамка считывания», обозначенная аббревиатурой ORF, относится к длине последовательности нуклеиновой кислоты, любой (ДНК, кДНК или РНК), которая содержит сайт инициации трансляции, или инициирующий кодон, такой как ATG или AUG, и терминирующий кодон, и потенциально может быть транслирована в полипептидную последовательность.
Термин «по ходу транскрипции» относится к нуклеотидной последовательности, которая расположена с 3'-конца относительно рассматриваемой нуклеотидной последовательности. В частности, расположенные по ходу транскрипции нуклеотидные последовательности обычно относятся к последовательностям, которые следуют за сайтом инициации транскрипции. Например, инициирующий кодон трансляции гена находится по ходу транскрипции относительно сайта инициации транскрипции.
Термин «против хода транскрипции» относится к нуклеотидной последовательности, которая расположена с 5'-конца относительно рассматриваемой нуклеотидной последовательности. В частности, расположенные против хода транскрипции нуклеотидные последовательности обычно относятся к последовательностям, которые находятся с 5'-стороны кодирующей последовательности или сайта инициации транскрипции. Например, большинство промоторов расположены против хода транскрипции относительно сайта инициации транскрипции.
Термин «химически синтезированный», в отношении последовательности ДНК, значит, что сборка цепи ДНК из составляющих ее нуклеотидов была проведена in vitro. Неавтоматический химический синтез ДНК может быть выполнен с помощью хорошо отлаженных технологий, или можно осуществить автоматический химический синтез с помощью одного из нескольких коммерчески доступных приборов. Следовательно, гены можно изготовить для получения их оптимальной экспрессии, основанной на оптимизации нуклеотидной последовательности, чтобы отразить особенности состава кодонов клетки-хозяина. Специалист в данной области техники примет во внимание вероятность успешной экспрессии гена, если частота кодона смещена относительно тех кодонов, предпочтительных для хозяина. Определение предпочтительных кодонов может быть основано на исследовании генов, полученных из клетки-хозяина, если информация о последовательностях доступна.
Термины «эндонуклеаза рестрикции» и «фермент рестрикции» в настоящей зваявке взаимозаменяемы и относятся к ферменту, который связывается и разрезает специфичную нуклеотидную последовательности внутри двухцепочечной ДНК.
Термины «полипептид», «пептид» и «белок» в настоящей заявке взаимозаменяемы и относятся к полимерному соединению, содержащему ковалентно связанные аминокислотные остатки. Аминокислоты имеют следующую основную структуру.
«Полимеразная цепная реакция», сокращено ПЦР, относится к методу ферментативной амплификации in vitro нуклеотидных последовательностей. ПЦР включает повторяющиеся серии температурных циклов, каждый из которых состоит из трех стадий: денатурация матричной нуклеотидной последовательности, имеющей целью разделить цепи целевой молекулы, отжиг одноцепочечного олигонуклеотидного праймера с матричной нуклеотидной последовательностью и удлинение подвергнутого отжигу праймера (праймеров) ДНК-полимеразой.
Термин «гомология» относится к проценту идентичности между двумя полинуклеотидами или двумя полипептидными цепями. Соответствие между двумя последовательностями можно определить с помощью методик, известных специалистам в данной области. Например, гомологию можно выявить путем прямого сравнения известных последовательностей двух полипептидных молекул с помощью «выравнивания» последовательностей и использования доступных компьютерных программ. В качестве альтернативы, гомологию можно установить путем гибридизации полинуклеотидов в условиях, когда образуются стабильные дуплексы между гомологичными областями, с последующим расщеплением специфичной к одноцепочечным участкам нуклеазой и определением размеров расщепленных фрагментов.
Используемый в настоящей заявке термин «гомологичный», во всех его грамматических формах и вариациях в написании, относится к сходству между белками, имеющими «общее эволюционное происхождение», включая белки из суперсемейств (например, суперсемейство иммуноглобулинов) и гомологичные протеины разных видов (например, легкая цепь миозина и прочее) (Reeck et al., Cell 50:667 (1987)). Последовательности таких белков (и кодирующих их генов) обладают гомологией, что отражено в их высокой степени подобия. Однако термин «гомологичный» в общем употреблении и в настоящем применении, в особенности используемый с наречием «высоко», можно отнести к подобию последовательностей, но не к общему эволюционному происхождению. Таким образом, термин «подобие последовательностей», во всех его грамматических формах, относится к степени идентичности или соответствия между нуклеотидными последовательностями или аминокислотными последовательностями белков, которые могут иметь, а могут и не иметь общего эволюционного происхождения (см. Reeсk et al., Cell 50:667 (1987)). В одном из вариантов две последовательности ДНК «гомологичны в значительной степени» или «подобны в значительной степени» в случае, когда, по меньшей мере, около 21% (желательно, по меньшей мере, около 50% и еще более предпочтительно, по меньшей мере, около 75%, 90%, 95%, 96%, 97%, 98% или 99%) нуклеотидов совпадают на определенной длине последовательности ДНК. Последовательности, которые в значительной степени гомологичны, можно идентифицировать путем сравнения последовательностей с использованием имеющегося в базах данных последовательностей стандартного программного обеспечения, или в эксперименте по Саузерн-гибридизации, например, при жестких условиях, которые определяются для такой системы отдельно. Задание соответствующих условий для гибридизации может определить специалист в этой области техники (см. например, Sambrook et al., 1989).
Используемый в настоящей заявке термин «подобный в значительной степени» относится к фрагментам нуклеотидных последовательностей, где изменения в одном или более нуклеотидных основаниях приводят к замещению одной или более аминокислот, но не оказывают влияния на функциональные свойства белков, кодируемых ДНК-последовательностью. «Подобный в значительной степени» также относится к фрагментам нуклеиновых кислот, при этом изменения в одном или более нуклеотидных основаниях не влияют на способность фрагмента нуклеотидной последовательности опосредовать изменения экспрессии генов путем антисмысловой или косупрессионной технологии. «Подобный в значительной степени» также относится к модификациям фрагментов нуклеотидных последовательностей данного изобретения, таким как делеция или инсерция одного или более нуклеотидных оснований, не оказывающим значительного влияния на функциональные свойства получаемого транскрипта. Следовательно, подразумевается, что изобретением охвачены больше, чем специфические последовательности, приведенные в примерах. Каждая из представленных модификаций хорошо известна специалисту в данной области, так же как определение сохранения биологической активности кодируемых продуктов.
Более того, специалисту в данной области известно, что подобные в значительной степени последовательности, включенные в это изобретение, также характеризуются их способностью к гибридизации в жестких условиях. Молекула нуклеиновой кислоты способна гибридизоваться с другой молекулой нуклеиновой кислоты, такой как кДНК, геномная ДНК или РНК, когда одноцепочечная форма нуклеиновой кислоты может комплементарно связаться с другой молекулой нуклеиновой кислоты при соответствующих температурных условиях и ионной силе раствора (см. Sambrook et al., 1989). Условия гибридизации и промывки хорошо известны и приведены в пособии Sambrook, J., Fritsch, E.F. And Maniatis, T. Molecular Cloning. A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor (1989), в особенности в Главе 11 и Таблице 11.1. Температурные условия и ионная сила определяют «жесткость» гибридизации.
Жесткость условий можно подобрать таким образом, чтобы проводить поиск умеренно подобных фрагментов, таких как гомологичные последовательности эволюционно далеких организмов, либо высокоподобных фрагментов, таких как гены, которые являются копиями функциональных ферментов от близкородственных организмов. Для предварительного скрининга гомологичных нуклеиновых кислот могут быть использованы нежесткие условия гибридизации, соответствующие температуре 55°С и, например, 5-кратному SSC, 0.1% SDS, 0.25% молока, в отсутствие формамида; или 30% формамид, 5-кратный SSC, 0.5% SDS. Умеренная жесткость условий гибридизации соответствует более высокой температуре и, например, 40% формамида с 5- или 6-кратным SSC (буфер для денатурации). Высокая жесткость условий гибридизации соответствует самой высокой температуре и, например, 50% формамида, 5- или 6-кратному SSC.
Для гибридизации необходимо, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя в зависимости от жесткости условий гибридизации между основаниями возможны несоответствия. Термин «комплементарный» применяют для описания связи между нуклеотидными основаниями, которые способны гибридизоваться друг с другом. Например, касательно ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, настоящее изобретение также включает отдельные фрагменты нуклеиновых кислот, которые комплементарны полным последовательностям, как здесь было описано или применено, а также тем в значительной степени подобным последовательностям нуклеиновых кислот.
В одном из вариантов осуществления изобретения полинуклеотиды выявляют путем применения условий гибридизации, содержащих этап гибридизации при 55°С, и с использованием условий, какие были описаны выше. В другом варианте осуществления изобретения температура составляет 60°С; в отдельных случаях температура достигает 63°С или 65°С.
Промывки после гибридизации также определяют жесткость условий. В одном варианте предпочтительных условий применяют серию промывок, начиная с 6-кратного SSC, 0.5% SDS при комнатной температуре в течение 15 мин, затем повторяют в 2-кратном SSC, 0.5% SDS при 45°С в течение 30 мин, а потом дважды в 0.2XSSC, 0.5% SDS при 50°С в течение 30 мин. В другом примере жестких условий применяют более высокие температуры, и где промывки идентичны упомянутым выше, за исключением температуры последних двух 30-мин промывок в 0.2XSSC, 0.5% SDS, которую увеличили до 60°С. Еще в одном примере очень жестких условий две последние промывки проводят в 0.1XSSC, 0.1% SDS при 65°С. Для гибридизации необходимо, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя в зависимости от жесткости гибридизации между основаниями возможны несоответствия.
Соответствующая жесткость для подвергающихся гибридизации нуклеиновых кислот зависит от длины нуклеиновых кислот и степени комплементарности (комплементации), переменных, хорошо известных специалистам в данной области. Чем больше степень подобия или гомологии между двумя нуклеотидными последовательностями, тем выше значение температуры для получения гибридов нуклеиновых кислот, имеющих такие последовательности. Относительная стабильность (соответствующая более высокой температуре) гибридизации нуклеиновых кислот уменьшается в следующем порядке: РНК:РНК, ДНК:РНК, ДНК:ДНК. Формулы для вычисления температуры для гибридов длиной более 100 нуклеотидов были получены (см. Sambrook et al., 9.50-0.51). Для гибридизации более коротких нуклеиновых кислот, например олигонуклеотидов, расположение несоответствий становится более важным и длина олигонуклеотидов определяет ее (гибридизации) специфичность (см. Sambrook et al., 11.7-11.8).
В одном из вариантов полинуклеотиды выявляют путем использования условий гибридизации, состоящих из этапа гибридизации при концентрации солевого раствора менее чем 500 мМ и, по меньшей мере, 37°С и этапа промывки в 2-кратном SSPE не менее чем при 63°С. В другом варианте условия гибридизации включают концентрацию солевого раствора менее 200 мМ и, по меньшей мере, 37°С для этапа гибридизации. В отдельных случаях условия гибридизации включают 2-кратный SSPE и 63°С и для этапа гибридизации, и для промывки.
Длина поддающихся гибридизации нуклеиновых кислот составляет, к примеру, не менее 10 нуклеотидов. Минимальная длина способных к гибридизации нуклеиновых кислот может быть, по меньшей мере, около 15 нуклеотидов; не менее 20 нуклеотидов или не менее 30 нуклеотидов. Более того, специалисту в данной области известно, что температуру и концентрацию промывочного солевого раствора при необходимости можно подобрать в соответствии с факторами, такими как длина зонда.
Подобные в значительной степени фрагменты нуклеиновых кислот согласно настоящему изобретению - это те фрагменты нуклеиновых кислот, ДНК-последовательности которых, по меньшей мере, на 70% идентичны ДНК-последовательностям фрагментов нуклеиновых кислот, описанных в настоящей заявке. Фрагменты нуклеиновых кислот согласно настоящему изобретению включают фрагменты нуклеиновых кислот, ДНК-последовательности которых, по меньшей мере, на 80%, 90%, 95%, 96%, 97%, 98% и 99% идентичны ДНК-последовательностям фрагментов нуклеиновых кислот, описанных в настоящей заявке.
Термин «соответствующий чему-либо» используют здесь, чтобы сослаться на подобные или гомологичные последовательности, вне зависимости от того, будет ли точная позиция идентична или отличаться от позиции в молекуле, по отношению к которой подобие или гомология измеряется. Линейное соответствие последовательностей нуклеиновых кислот или аминокислот может включать пустоты. Таким образом, термин «соответствующий» относится к подобию последовательности, а не к количеству аминокислотных остатков или нуклеотидный оснований.
«Значительная часть» аминокислотной или нуклеотидной последовательности содержит в себе достаточную часть аминокислотной последовательности полипептида или нуклеотидной последовательности гена, чтобы предположительно идентифицировать этот полипептид или ген либо путем анализа последовательности специалистом в данной области вручную, либо путем автоматизированного сравнения и определения последовательностей с помощью компьютера с использованием алгоритмов, таких как BLAST (Basic Local Alignment Search Tool; Altschul et al., J. Mol. Biol. 215:403 410 (1993)); BLAST общедоступен во Всемирной сети Интернет. Как правило, необходима последовательность из десяти или более смежных аминокислот или тридцати или более нуклеотидов для того, чтобы предположительно идентифицировать полипептид или последовательность нуклеиновых кислот как гомологичные по отношению к известному протеину или гену. Более того, по отношению к нуклеотидным последовательностям, специфичным к гену олигонуклеотидным зондам, содержащим 20 или 30 смежных полинуклеотидов, могут быть применены в зависимых от последовательности методах генной идентификации (например, при Саузерн-гибридизации) и изоляции (например, в гибридизации in situ бактериальных колоний и бляшек бактериофагов). Кроме того, короткие олигонуклеотиды из 12-15 оснований можно использовать как амплификационные праймеры в ПЦР, чтобы обнаружить отдельные фрагменты нуклеиновых кислот, содержащие последовательности праймеров. Таким образом, «значительная часть» нуклеотидной последовательности содержит достаточную часть последовательности, чтобы точно идентифицировать и/или изолировать фрагмент нуклеиновой кислоты, содержащий последовательность.
Термин «процент подобия», как известно в данной области, это соответствие между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, которое установлено путем сравнения последовательностей. В данной области «идентичность» также означает степень родства последовательностей между полипептидными и нуклеотидными последовательностями, что в определенном случае может быть установлено путем совпадения между нитями таких последовательностей. «Идентичность» и «подобие» можно без труда подсчитать известными методиками, включающими, в частности, те, которые описаны в: Compulation Molecular Biology (Lesk, A.M., ed.) Oxford University Press, New York (1988); Biocomputing: Infomatics and Genome Projects (Smith, D.W., ed.) Academic Press, New York (1993); Computer Analysis of Sequence Data, Part I (Griffin, A.M., and Griffin, H. G., eds.) Humana Press, New Jersey (1994); Sequence Analysis in Molecular Biology (von Heinje, G., ed.) Academic Press (1987); и Sequence Analysis Primer (Gribskov, M. and Devereux, J., eds.) Stockton Press, New York (1991). Способы определения идентичности созданы таким образом, чтобы найти наилучшее соответствие между тестируемыми последовательностями. Способы определения идентичности и подобия систематизированы в общедоступных компьютерных программах. Анализ линейного соответствия последовательностей и расчет процента идентичности можно выполнить с применением программы Megalign из пакета LASERGENE для вычислительной биоинформатики (DNASTAR Inc., Madison, WI). Множественный анализ линейного соответствия можно осуществить с помощью метода Clustal (Higgins et al., CABIOS. 5:151 153 (1989)) с параметрами, используемыми по умолчанию (GAP PENALTY=10, GAP LENGH=10). Параметры, используемые по умолчанию, для парного анализа линейного соответствия при применении метода Clustal можно выбрать следующие: KTUPLE 1, GAP PENALTY=3, WINDOW=5 и DIAGONALS SAVED=5.
Термин «программное обеспечение для анализа последовательностей» относится к любому вычислительному алгоритму или программному обеспечению, которое успешно применяется для анализа нуклеотидных или аминокислотных последовательностей. «Программное обеспечение для анализа последовательностей» может быть коммерчески доступно или независимо разработано. Типичное программное обеспечение для анализа последовательностей включает, но не ограничивается этим, следующее: пакет программ GCG (Wisconsin Package Version 9.0, Genetics Computer Group (GCG), Madison, WI), BLASTP, BLASTN, BLASTX (Altschul et al., J. Mol. Biol. 215:403 410 (1990)), и DNASTAR (DNASTAR, Inc. 1228 S. Park St. Madison, WI 53715 USA). Употребление в контексте данного документа подразумевает, что при применении программного обеспечения для анализа последовательностей, результаты анализа будут основаны на «значениях, установленных по умолчанию» упомянутой программы, если не оговорено иное. Используемый в настоящей заявке термин «значения, установленные по умолчанию» означает любой набор значений или параметров, которые загружаются при первоначальном запуске программного обеспечения.
Применяемый в настоящей заявке термин «экспрессия» или «экспрессия гена» относится к процессу преобразования генетической информации, закодированной в гене, в РНК (например, мРНК, рРНК, тРНК или мяРНК) посредством «транскрипции» гена (то есть через механизм действия фермента РНК-полимеразы), а для белок-кодирующих генов - в белок посредством «трансляции» мРНК. На многих этапах процесса экспрессия гена может регулироваться. «Позитивная регуляция» или «активация» относится к регуляции, которая увеличивает выработку продуктов экспрессии гена (то есть РНК или белка), в то время как «негативная регуляция» или «репрессия» относится к регуляции, которая подавляет эту выработку. Факторы (например, факторы транскрипции), которые участвуют в позитивной или негативной регуляции, часто называют «активаторами» и «репрессорами» соответственно. Для целей изобретения экспрессия гена-мишени может быть подавлена на посттранскрипционном уровне (то есть на уровне РНК транскрипта) посредством специфического взаимодействия с негативно регулирующей молекулой РНК.
Термин «контролирующие последовательности транскрипции и трансляции» относится к регуляторным последовательностям ДНК, таким как промоторы, энхансеры, терминаторы и т.п., которые обеспечивают экспрессию кодирующей последовательности в клетке-хозяине.
Термин «функционально связанный» относится к объединению последовательностей нуклеиновых кислот в один фрагмент нуклеиновой кислоты таким образом, чтобы функция одной последовательности находилась под влиянием другой. Например, промотор функционально связан с кодирующей последовательностью, когда он (промотор) способен влиять на экспрессию кодирующей последовательности (то есть кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующая последовательность может быть функционально связана с регуляторными последовательностями в смысловой или антисмысловой ориентации.
Термин «вектор» относится к любой конструкции для клонирования и/или перенесения в клетку-хозяин нуклеиновой кислоты. Вектор может быть репликоном, к которому может быть присоединен другой сегмент ДНК с тем, чтобы вызвать репликацию присоединенного сегмента. «Репликон» имеет отношение к любому генетическому элементу (например, плазмида, фаг, космида, хромосома, вирус), действующему как автономная единица репликации ДНК in vivo, то есть способна к самоконтролируемой репликации. Термин «вектор» может включать вирусные и невирусные конструкции для введения нуклеиновой кислоты в клетку-хозяина in vitro, ex vivo или in vivo. Термин «вектор» может также включать мини-кольца ДНК. Например, вектор может быть плазмидой без бактериальных последовательностей ДНК. Как было показано, удаление бактериальных последовательностей ДНК, богатых ГЦ-областями, снижает угнетение трансгенной экспрессии и ведет к более устойчивой экспрессии плазмидных ДНК-векторов (см., например, Ehrhardt, A. et al. (2003) Hum Gene Ther 10:215-25; Yet, N.S. (2002) Mol Ther 5:731-38; Chen, Z.Y. et al. (2004) Gene Ther 11:856-64). Термин «вектор» также может включать транспозоны, такие как Спящая Красавица (Izsvak et al., J. Mol. Biol. 302:93-102 (2000)), или искусственные хромосомы.
Большое количество векторов, известных в данной области, можно применять с целью воздействия на нуклеиновые кислоты, включения элементов отклика и промоторов в гены и т.д. или перенесения нуклеиновой кислоты в клетку-хозяина. Векторы могут включать, например, плазмиды или модифицированные вирусы, включая, например, такие бактериофаги, как производные фага лямбда, или такие плазмиды, как pBR322 или производные плазмиды pUC, или вектор Bluescript. Более крупные векторы, такие как искусственные хромосомы (бактериальные (ВАС), дрожжевые (YAC) или хромосомы человека (НАС)), можно применить для осуществления более крупных включений. Например, инсерция фрагментов ДНК, соответствующих элементам отклика или промоторам в подходящий вектор, может быть проведена за счет лигирования соответствующих фрагментов ДНК в выбранный вектор, который имеет комплементарные «липкие» концы. В качестве альтернативы концы ДНК молекул могут быть модифицированы ферментами, либо можно получить любой сайт путем лигирования нуклеотидных последовательностей (линкеров) к концам ДНК. Такие векторы можно создать искусственно таким образом, чтобы они содержали селектируемые маркерные гены, которые обеспечат отбор клеток, трансфицированных или трансформированных вектором. Рекомбинантный вектор, содержащий полинуклеотид согласно изобретению, может включать один или более сайтов инициации для репликации в клетках-хозяевах, в которых необходимо достичь его амплификации или экспрессии, а также маркеры или селектируемые маркеры.
Термин «селектируемый маркер» относится к идентифицирующему фактору, обычно это ген устойчивости к антибиотикам или химическим агентам, по отношению к которому может производиться селекция на основании эффекта маркерного гена, например устойчивости к антибиотику, устойчивости к гербициду, колориметрическим маркерам, ферментам, флуоресцентным маркерам и т.п., где эффект используют для того, чтобы проследить наследование интересующей нуклеиновой кислоты и/или идентифицировать или отобрать клетку или организм, который наследовал интересующую нуклеиновую кислоту. Примеры селектируемых маркерные генов, известных и применяемых в данной области, включают: гены, обеспечивающие устойчивость к ампициллину, стрептомицину, гентамицину, канамицину, гигромицину, гербициду биалафос, сульфонамиду и т.п.; и гены, применяемые как фенотипические маркеры, например антоцианин-регуляторные гены, ген изопентанил-трансферазы и т.п.
Термин «репортерный ген» относится к нуклеиновой кислоте, кодирующей идентифицирующий фактор, который можно распознать на основе эффекта репортерного гена, где эффект используют для того, чтобы проследить наследование интересующей нуклеиновой кислоты, идентифицировать клетку или организм, который унаследовал интересующую нуклеиновую кислоту, и/или оценить индукцию экспрессии гена или транскрипцию гена. Примеры репортерных генов, известных и используемых в данной области техники, включают: люцеферазу (Luc), флуоресцентные белки, такие как зеленый флуоресцентный белок (GFP), хлорамфеникол-ацетилтрансфераза (CAT), бета-галактозидаза (LacZ), бета-глукуронидаза (Gus) и т.п. Селектируемые маркерные гены также могут считаться репортерными генами.
Термин «плазмида» относится к экстра-хромосомному элементу, часто несущему ген, который не является частью центрального метаболизма клетки, и обычно находится в форме двухцепочечных кольцевых молекул ДНК. Такими элементами могут быть автономно реплицирующиеся последовательности, интегрирующиеся в геном последовательности, фаги или нуклеотидные последовательности, линейные, кольцевые или сверхспиральные одноцепочечные или двухцепочечные ДНК или РНК, полученные из любого источника, при этом ряд нуклеотидных последовательностей связываются или рекомбинируются в уникальную конструкцию, которая способна внедрять в клетку промоторный фрагмент и последовательность ДНК-продукта выбранного гена вместе с соответствующей 3'-концевой нетранслируемой последовательностью.
«Вектор для клонирования» относится к «репликону», который является единицей длины нуклеиновой кислоты, например ДНК, которая реплицируется частями и содержит сайт инициации репликации, такому как плазмида, фаг или космида, к которому может быть присоединен сегмент другой нуклеиновой кислоты таким образом, чтобы это привело к репликации присоединенного сегмента. Векторы для клонирования могут обладать способностью к репликации в одном типе клеток и к экспрессии - в другом («шаттл-вектор»).
Термин «экспрессионный вектор» относится к вектору, плазмиде или конструкции, предназначенной для экспрессии включенной последовательности нуклеиновой кислоты, следующей за трансформацией в клетку-хозяин. Клонируемый ген, то есть включенная последовательность нуклеиновой кислоты, обычно помещается под управление регулирующих элементов, таких как промотор, минимальный промотор, энхансер и т.п. Регулирующие инициацию области или промоторы, которые способны контролировать экспрессию нуклеиновой кислоты в клетке-хозяине, многочисленны и хорошо знакомы специалистам в данной области.
Векторы можно ввести в желаемые клетки-хозяева методами, известными специалистам в данной области, например, путем трансфекции, электропорации, микроинъекции, трансдукции, клеточного слияния, с использованием метода с диэтиламинэтилдекстраном (DEAE-декстран), преципитации фосфатом кальция, липофекцией (лизосомное слияние), с применением генной пушки или переносчика ДНК-вектора (см., например, Wu et al., J. Biol. Chem. 267:963-967 (1992); Wu et al., J. Biol. Chem. 263:14621-14624 (1988); и Hartmut et al., Canadian Patent Application No.2,012,311, filed March 15, 1990).
Примеры эукариотических векторов включают, но не ограничиваются указанными, pW-LNEO, pSV2CAT, pOG44, pXT1 и pSG, имеющиеся в наличии у Stratagene; pSVK3, pBPV, pMSG и pSVL, имеющиеся в наличии у Amersham Pharmacia Biotech; pCMVDsRed2-express, pIRES2-DsRed2, pDsRed2-Mito, pCMV-EGFP, имеющиеся в наличии у Clontech. Многие другие векторы хорошо известны и коммерчески доступны.
Например, полезные векторы, содержащие молекулярные центры инсерции для быстрой инсерции или удаления элементов генетических программ, описаны в опубликованной патентной заявке США No.2004/0185556, в патентной заявке США No.11/233,246 и международных опубликованных заявках WO 2005/040336 и WO 2005/116231.
Термины «промотор» и «промоторная последовательность» в настоящей заявке взаимозаменяемы и относятся к ДНК-последовательности, способной контролировать экспрессию кодирующей последовательности или функциональную РНК. Как правило, кодирующая последовательность расположена в 3'-области относительно промоторной последовательности. Промоторы можно получить полностью из нативного гена или создать их из разных элементов, взятых из разных промоторов, имеющихся в природе, и даже включить синтетические ДНК-сегменты. Специалистам в данной области известно, что разные промоторы могут направлять экспрессию гена в разных тканях или типах клеток, или на разных этапах развития, или в ответ на разные физиологические условия или условия окружающей среды.
Промоторы, которые вызывают экспрессию гена в большинстве типов клеток в большинстве случаев, обычно относят к «конститутивным промоторам». Промоторы, которые вызывают экспрессию гена в специфических типах клеток, обычно относят к «условным промоторам». Примерами условных промоторов являются «клеточно-специфичные промоторы» или «тканеспецифичные промоторы». Промоторы, которые вызывают экспрессию гена на специфической стадии развития или клеточной дифференциации, обычно относят к «промоторам, специфичным к стадии развития» или к «промоторам, специфичным к стадии клеточной дифференцировки». Промоторы, которые индуцируются и вызывают экспрессию гена, вслед за экспозицией или обработкой клетки агентом, биологической молекулой, химическим веществом, лигандом, светом и т.п., что вызывает индукцию промотора, обычно относят к «индуцируемым промоторам» или «регулируемым промоторам». Примером индуцируемого промотора является TetO-индуцируемый промотор. Далее, так как известно, что в большинстве случаев точные границы регуляторных последовательностей полностью не определены, ДНК-фрагменты разной длины могут иметь идентичную промоторную активность.
Последовательность промотора обычно ограничена с 3'-конца сайтом инициации транскрипции и тянется против хода транскрипции (5'-направление) настолько, чтобы включить минимальное количество оснований или элементов, необходимых для инициации транскрипции на детектируемых (относительно фона) уровнях. В пределах последовательности промотора находят сайт инициации транскрипции (традиционно определяемый при помощи, например, картирования с применением нуклеазы S1), а также белок-связывающие домены (консенсунсные последовательности), ответственные за связывание РНК-полимеразы.
Кодирующая последовательность «контролируется» в клетке транскрипционными и трансляционными контролирующими последовательностями, когда РНК-полимераза транскрибирует кодирующую последовательность в мРНК, которая затем подвергается сплайсингу с участием транс-РНК (если кодирующая последовательность содержит интроны) и транслируются в белок, закодированный кодирующей последовательнрстью.
Терминирующие регуляторные области, то есть терминатор или поли-А последовательности, можно получить из разных генов, присущих предпочтительным хозяевам. Сайт терминации возможно может отсутствовать, однако может быть включен. В одном из вариантов осуществления изобретения терминирующая контролирующая область может быть составной или может быть получена из синтетической последовательности, синтетического сигнала полиаденилирования, позднего сигнала полиаденилирования SV40, сигнала полиаденилирования SV40, сигнала полиаденилирования гена бычьего гормона роста (BGH), последовательностей терминаторов вирусов и т.п.
Термин «трансфекция» относится к поглощению клеткой экзогенных или гетерогенных РНК или ДНК. Клетка является «трансфицированной» экзогенными или гетерогенными РНК или ДНК, когда такие РНК или ДНК введены внутрь клетки. Трансфицированные РНК или ДНК могут быть интегрированы (ковалентно связаны) в хромосомную ДНК, таким образом пополняя геном клетки-хозяина.
«Трансформация» относится к переносу фрагмента нуклеиновой кислоты в геном организма-хозяина, в результате чего происходит генетически стабильное наследование.
Термины «модулировать» и «модулирует» означают стимулировать, снижать или ингибировать экспрессию нуклеиновой кислоты или гена, результатом чего являются соответствующие индукция, редукция или ингибирование выработки белка или полипептида.
Термин «РНК-транскрипт» относится к продукту, полученному в результате катализируемой РНК-полимеразой транскрипции ДНК-последовательности. Когда РНК-транскрипт является абсолютно комплементарной копией ДНК-последовательности, его называют первичным транскриптом, также он может быть РНК-последовательностью, полученной в результате посттранскрипционного процессинга первичного транскрипта, и тогда его называют зрелой РНК. «Матричная РНК (мРНК)» относится к РНК, у которой отсутствуют интроны и которая может быть транслирована клеткой в белок. «кДНК» относится к двухцепочечной ДНК, которая комплементарна мРНК и из нее же получена. «Смысловая» РНК относится к РНК-транскрипту, в том числе к мРНК, и таким образом может быть транслирована клеткой в белок.
Одним из вариантов осуществления настоящего изобретения является синтетический 5'UTR полинуклеотид, содержащий первый полинуклеотидный фрагмент и второй полинуклеотидный фрагмент, где:
а. первый нуклеотидный фрагмент содержит, по меньшей мере, один сайт сплайсинга первого эукариотического гена;
b. второй нуклеотидный фрагмент содержит, по меньшей мере, часть 5'-концевой нетранслируемой области второго эукариотического гена; и
с. первый полинуклеотидный фрагмент локализован в 5'-конце второго полинуклеотидного фрагмента.
В другом варианте осуществления настоящего изобретения синтетический 5'UTR является химерным полинуклеотидом, который содержит первый полинуклеотидный фрагмент и второй полинуклеотидный фрагмент, при этом:
а. первый полинуклеотидный фрагмент содержит второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума;
b. второй полинуклеотидный фрагмент содержит, по меньшей мере, часть 5'-концевой нетранслируемой области (5'UTR) гена казеина; и
с. первый полинуклеотидный фрагмент локализован в 5'-конце второго полинуклеотидного фрагмента.
Полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, можно получить из любого эукариотического гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума. Одним из вариантов осуществления настоящего изобретения является полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, полученный из гена SERCA2. В других вариантах осуществления изобретения его получали из генов SERCA1 и SERCA3. Ген кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, который является источником полинуклеотидного фрагмента, содержащего второй интрон, может быть получен от любого вида эукариот. В одном из вариантов осуществления настоящего изобретения ген кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума был получен от млекопитающих. В другом варианте осуществления изобретения был взят птичий ген кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума. В еще одном варианте осуществления настоящего изобретения использовался ген кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума рыб. В конкретных вариантах настоящего изобретения полинуклеотидный фрагмент, содержащий второй интрон, получали из гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума человека, собаки или мыши. В других конкретных вариантах осуществления настоящего изобретения полинуклеотидный фрагмент, содержащий второй интрон, получали из гена саркоплазматического/эндоплазматического ретикулума крысы, шимпанзе, цыпленка, лошади, коровы, лося, свиньи, кота, макаки-резус или полосатого данио.
В другом варианте изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, включает часть экзона 2, расположенную в 5'-конце, и часть экзона 3, расположенную в 3'-конце. В еще одном варианте настоящего изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, включает полный экзон 2, расположенный в 5'-конце, и полный экзон 3, расположенный в 3'-конце.
Полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, может составлять, по меньшей мере, примерно 50 нуклеотидов в длину. В других вариантах осуществления изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, может составлять, по меньшей мере, около 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240 или 250 нуклеотидов в длину.
В другом варианте осуществления изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, мутирован в предполагаемом консенсусном поли-А сайте. В еще одном варианте осуществления изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, расположенную в 5'-конце часть экзона 2 и расположенную в 3'-конце часть экзона 3, мутирован в предполагаемом консенсусном поли-А сайте и получен из гена собаки SERCA2. В конкретном варианте осуществления изобретения последовательность представлена SEQ ID NO:2.
В других вариантах осуществления настоящего изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, является нормальной или мутированной неполной SERCA2-последовательностью. Полинуклеотидный фрагмент можно получить из любого полноразмерного гена SERCA2 любого вида. Например, в одном из вариантов осуществления изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, является нормальной неполной геномной последовательностью SERCA2 собаки, которая содержит часть экзона 2, расположенную в 5'-конце, и часть экзона 3, расположенную в 3'-конце. В конкретном варианте осуществления она представлена последовательностью SEQ ID NO:4. SEQ ID NO:4 схематично изображена на ФИГУРЕ 1А и ФИГУРЕ 1C. В другом варианте осуществления изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, является нормальной неполной геномной последовательностью SERCA2 человека, которая содержит часть экзона 2, расположенную в 5'-конце, и часть экзона 3, расположенную в 3'-конце. В конкретном варианте осуществления она представлена последовательностью SEQ ID NO:5. В другом варианте осуществления полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, является нормальной неполной геномной последовательностью SERCA2 мыши, которая содержит часть экзона 2, расположенную в 5'-конце, и часть экзона 3, расположенную в 3'-конце. В конкретном варианте осуществления она представлена последовательностью SEQ ID NO:6. SEQ ID NO:5 и SEQ ID NO:6 схематично изображены на ФИГУРЕ 1В и ФИГУРЕ 1C. В других вариантах осуществления настоящего изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, является мутантной или нормальной неполной последовательностью SERCA2 Rattus norvegicus, последовательностью SERCA2 Equus caballus, последовательностью SERCA2 Bos Taurus, последовательностью SERCA2 Pan troglodytes, последовательностью SERCA2 Felis catus, последовательностью SERCA2 Ortolagus cuniculus, последовательностью SERCA2 Sus scrofa, последовательностью SERCA2 Macaca mulatta, последовательностью SERCA2 Cervus elaphus, последовательностью SERCA2 Gallus gallus или последовательностью SERCA2 Danio rerio.
Полинуклеотидная последовательность, содержащая, по меньшей мере, часть 5' нетранслируемой области гена казеина, может быть получена из любого вида млекопитающих. В одном из вариантов изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области, получен из гена бета-казеина быка. В конкретном варианте осуществления он представлен последовательностью SEQ ID NO:3. В другом варианте осуществления изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области, получен из гена бета-казеина мыши. В конкретном варианте осуществления он представлен последовательностью SEQ ID NO:8. В другом варианте осуществления изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области, получен из гена бета-казеина крысы. В конкретном варианте осуществления он представлен последовательностью SEQ ID NO:9. В другом варианте осуществления изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области, получен из гена бета-казеина овцы. В конкретном варианте осуществления он представлен последовательностью SEQ ID NO:10. В других вариантах осуществления изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области, получен из гена бета-казеина Bubalus bubalus, гена бета-казеина Carpa hircus, гена бета-казеина Equus cabalus, гена бета-казеина Sus scrofa, гена бета-казеина Camelus dromedaries, гена бета-Oryctolagus cuniculus или гена бета-казеина Canis lupus.
Полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области гена казеина, может быть, по меньшей мере, около 25 нуклеотидов в длину. В других вариантах осуществления полинуклеотидный фрагмент гена казеина, содержащий, по меньшей мере, часть 5'UTR, может быть, по меньшей мере, около 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 100 или более нуклеотидов в длину. В другом варианте изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5'UTR гена казеина, может представлять, по меньшей мере, около 50% натуральной последовательности 5'UTR. В других вариантах осуществления полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5'UTR гена казеина, может представлять, по меньшей мере, около 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более натуральной последовательности 5'UTR. В другом варианте изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5'UTR гена казеина, может представлять собой полную натуральную последовательность 5'UTR.
В иных вариантах осуществления изобретения функциональные варианты отдельных компонентов (полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, и полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5'UTR гена казеина) используют для создания синтетической 5'UTR. Функциональные варианты включают варианты с заменами, инсерциями и делениями и их комбинациями. Вариантами с заменами являются те, в которых, по меньшей мере, одно основание в нуклеотидной последовательности было удалено, а другое основание введено на его место. Вариантами нуклеиновой кислоты с инсерциями являются те, в которых один или более нуклеотидов введены в определенный сайт последовательности. Варианты нуклеиновой кислоты с делениями характеризуются удалением одного или более нуклеотидов из нуклеиновой кислоты. Могут иметь место любые комбинации замены (замен), инсерции (инсерций) или делеции (делеций) при условии, что функциональность компонента остается по существу такой же, а именно функциональный вариант, который используют в синтетической 5'UTR области настоящего изобретения, вызывает увеличение экспрессии интересующей последовательности, синтетического гена или трансгена.
Далее, последовательности, гомологичные конкретным вариантам полинуклеотидного фрагмента, содержащего второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, описанным здесь (SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:5 и SEQ ID NO:6), и последовательности, гомологичные конкретным вариантам полинуклеотидного фрагмента, содержащего, по меньшей мере, часть 5'UTR казеина (SEQ ID NO:3, SEQ ID NO:8, SEQ ID NO:9 и SEQ ID NO:10), могут быть использованы для создания синтетической 5'UTR. Как упоминалось ранее, подходящие источники фрагмента для создания синтетической 5'UTR содержатся в гене кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума любых видов эукариот и в гене казеина любых видов млекопитающих. В одном из вариантов изобретения полинуклеотидный фрагмент, содержащий второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, получают из ортолога SERCA2 собаки, SERCA2 мыши и SERCA2 человека. В другом варианте осуществления полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5'UTR гена казеина, получен из ортолога бета-казеина быка, бета-казеина мыши, бета-казеина крысы и бета-казеина овцы.
Способы изучения и идентификации гомологов кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума или гомологов 5'UTR казеина должны быть хорошо известны специалистам в данной области. Такие методы включают сравнение последовательностей, представленных SEQ ID NO:2-6 и 8-10 в читаемом формате компьютера, с последовательностями, которые доступны в публичнцх базах данных, имеющихся во Всемирной сети Интернет, таких как MIPS, GenBank или раза Данных Последовательностей Нуклеотидов EMBL, при помощи хорошо известных в данной области алгоритмов для выравнивания или сравнения последовательностей, таких как GAP (Needleman and Wunsch, J. Mol. Biol. 48; 443453 (1970)), BESTFIT (Miller, W., Myers, E.W. & Lipman, D.J., J. Mol. Biol. 215:403-410 (1990)), FASTA и TFASTA (W.R.Pearson and D.J.Lipman Proc. Natl. Acad. See. USA 85:2444-2448 (1988)). Программное обеспечение для BLAST-анализа находится в открытом доступе в Национальном Центре Биотехнологической Информации. Соответствующие гомологи могут быть идентифицированы с помощью параметров BLAST по умолчанию (BLOSUM62 matrix, gap opening penalty 11 и gap extension penalty 1).
Далее, гомологи SERCA2 собаки, мыши и человека могут быть также идентифицированы путем изучения консервативных последовательностей в гене SERCA2. Например, полная последовательность экзона 3 SERCA2 собаки, такая как SEQ ID NO:11, может быть использована как искомая последовательность в BLAST-анализе. Можно ожидать, что использование последовательности экзона в качестве искомой последовательности в BLAST-анализе будет выявлять большее количества гомологов SERCA2, чем при использовании последовательности, содержащей последовательность интрона. Подобным образом могут быть идентифицированы гомологи к бета-казеину быка, мыши, крысы и овцы с использованием кодирующей части в качестве искомой последовательности.
Также возможен анализ геномных последовательностей для идентификации гомологов кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума и гомологов 5'UTR казеина. Существуют алгоритмы и программные средства для идентификации генов в необработанной последовательности ДНК. Обычно такие способы сочетают в себе анализ статистических параметров в последовательности ДНК с основанными на гомологии способами для идентификации гомологичных последовательностей в базе данных. Несмотря на то, что по отдельности ни один из этих методов не является достаточно надежным для убедительного прогноза, комбинация различных программ обычно дает хорошие результаты. Хорошо известными примерами таких программных средств, которые находятся в открытом доступе во Всемирной сети Интернет, являются GeneMark (Borodovsky, M. аnd McIninch J.DeneMark: Параллельное распознавание генов для обеих цепей ДНК. Computers & Chemistry, 17, 123-133 (1993)), Gene Locator and Interpolated Markov Modeler (GLIMMER) (A.L.Delcher et fit. Improved microbial gene identification with GLIMMER. Nucleic Acids Research, 27, 4636-4641 (1991)), Gene Recognition and Assembly Internet Link (GRAIL), GenScan (Burge, C. and Karlin, S. Prediction of complete gene structures in human genomic DNA. J. Mol. Biol. 268, 78-94 (1997)) и GeneBuilder (Milanesi L. et al. GeneBuilder: interactive in silico prediction genes structure. Bioinformatics. 15 (7):612-621 (1999)). Комбинированный анализ можно осуществлять с помощью TIGR Combiner Program (J.E.Allen et al. Computational gene prediction using multiple sources of evidence. Genome Research, 14(1), 142-148 (2004)), который прогнозирует модели генов с применением выходных данных других аннотационных программ, таких как GeneMark, GlimmerM, GRAIL, GenScan и Fgenes. Эта программа использует статистический алгоритм для идентификации структур доказательств, соответствующих генетическим моделям.
Второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума может быть идентифицирован с помощью стандартных методов, таких как сравнение геномной последовательности ДНК гена с последовательностью его мРНК или кДНК в программе для проведения парного линейного сравнения. Области гомологии представлены экзонами, в то время как промежуточные последовательности, которые отсутствуют в последовательности кДНК, но имеются в геномной ДНК, представлены интронами. Начало и конец последовательности интрона могут быть идентифицированы с помощью расположенных в 5'-конце ГТ и 3'-конце АГ. Применяя такой подход, последовательность мРНК SERCA2 собаки, представленная общим номером доступа NM_001003214, будет полезна для идентификации интронов в геномной последовательности SERCA2 собаки, в то время как последовательности мРНК SERCA2 человека и мыши (NM_170665 и NM_009722 соответственно) могут быть использованы для того, чтобы идентифицировать интроны в их соответствующих геномных последовательностях.
Гомологи кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума и гомологи 5'UTR казеина могут быть также идентифицированы путем скринирования библиотек геномных или кДНК-фрагментов других видов. Например, геномная ДНК интересующего вида может быть разбита на фрагменты примерно одного размера для инсерции в выбранный вектор, такой как плазмида или лямбда-вектор. Затем вектор разрезается соответствующими ферментами рестрикции, после чего дотируется с полной смесью геномных фрагментов. Бактериальные клетки трансформируются вектором и затем размещаются на агарозных чашках. После этого ДНК колонии или фаговой бляшки прикрепляют к мембране. В одном варианте осуществления фрагменты, представленные последовательностями SEQ ID NO:2-6 и 8-10 или их частями, используют в качестве меченых зондов для гибридизации с ДНК клона из библиотеки, который содержит гомологичную последовательность. Подобные процедуры можно использовать для скринирования библиотеки кДНК. Далее, гомологи можно идентифицировать с применением фрагментов, представленных последовательностями SEQ ID NO:2-6 и 8-10 или их частями, в качестве меченых зондов для Саузерн-гибридизации с геномной или кДНК. В других вариантах осуществления более консервативные последовательности, в том числе включающие кодирующую область гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума (такие, как SEQ ID NO:11) или гена казеина или их части, используют как зонды в эксперименте Саузерн-гибридизации или для скрининга библиотек.
Соответствующие условия гибридизации могут быть выбраны таким образом, чтобы позволять гибридизацию фрагмента с зондом другого вида (частично несоответствующие гибриды зонд-мишень), путем снижения жесткости гибридизации эксперимента через подходящую комбинацию температуры, концентрации солевого раствора и % формамида. Например, жесткость эксперимента гибридизации может быть уменьшена путем снижения температуры с увеличением концентрации солевого раствора. Методики определения соответствующих условий гибридизации хорошо известны специалистам в данной области техники и описаны в Sambrook (2001) Molecular Cloning: a laboratory manual, 3rd Edition Cold Spring Harbor Laboratory Press, CSH, New York.
Другим вариантом осуществления изобретения является полинуклеотид, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный полинуклеотиду, представленному одной из последовательностей SEQ ID NOS:1-10.
Другим вариантом осуществления изобретения является синтетическая 5'UTR, содержащая полинуклеотид, представленный одной из последовательностей SEQ ID NOS:2 и 4-6, и полинуклеотид, представленный одной из последовательностей SEQ ID NOS:3 и 8-10.
Другим вариантом осуществления изобретения является синтетическая 5'UTR, содержащая полинуклеотид, который, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичен полинуклеотиду, представленному одной из последователньостей SEQ ID NOS:2 и 4-6, и полинуклеотид, который, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичен полинуклеотиду, представленному одной из последовательностей SEQ ID NOS:3 и 8-10.
Другим вариантом осуществления изобретения является синтетическая конструкция гена, содержащая полинуклеотид, который, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичен полинуклеотиду, представленному последовательностью SEQ ID NO:1.
Другим вариантом осуществления изобретения является синтетическая конструкция гена, содержащая полинуклеотид, который, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичен полинуклеотиду, представленному последовательностью SEQ ID NO:7.
В другом варианте изобретения синтетическая последовательность 5'UTR не имеет сайтов рестрикции, что будет препятствовать инсерции в UltraVector Production System (Intrexon Corp., Blacksburg, VA), как описано в публикации W02007/038276, включенной в настоящую заявку в качестве ссылки. В конкретном варианте осуществления синтетическая последовательность 5'UTR не имеет внутренних последовательностей распознавания для следующих эндонуклеаз рестрикции: AsiS I, Рас I, Sbf I, Fse I, Asc I, Mlu I, SnaB I, Not I, Sal I, Swa I, Rsr II, BsiW I, Mfe I, Nhe I, Nsi I, Cla I, Nde I, Nsi I, Kpn I, Nco I and Pst I.
Синтетическая последовательность 5'UTR факультативно включает сайты рестрикции в 5'- и 3'-концах, что способствует клонированию в вектор. В конкретном варианте осуществления синтетическая последовательность 5'UTR включает последовательности распознавания для Mlu I в 5'-конце и последовательности распознавания для Mfe I в 3'-конце.
В конкретном варианте осуществления синтетическая 5'UTR представлена последовательностью SEQ ID NO:1. SEQ ID NO:1 содержит в себе следующие элементы:
сайт рестрикции MluI, SEQ ID NO:2, сайт рестрикции KpnI, SEQ ID NO:3, сайт рестрикции MfeI. SEQ ID NO:1 схематично представлена на ФИГУРЕ 2.
В другом конкретном варианте синтетическая 5'UTR представлена последовательностью SEQ ID NO:7. SEQ ID NO:7 содержит следующие элементы: сайт рестрикции AscI, сайт рестрикции MluI, SEQ ID NO:4, сайт рестрикции KpnI, SEQ ID NO:3, сайт рестрикции MfeI. SEQ ID NO:7 схематично представлена на ФИГУРЕ 2В.
В одном из вариантов осуществления изобретения синтетическая последовательность 5'UTR составляет меньше чем около 500 нуклеотидов в длину. В другом варианте синтетическая последовательность 5'UTR составляет меньше чем около 400 нуклеотидов в длину. В другом варианте осуществления синтетическая последовательность 5'UTR составляет меньше чем около 350 нуклеотидов в длину. В другом варианте осуществления синтетическая последовательность 5'UTR составляет меньше чем около 300 нуклеотидов в длину. В другом варианте осуществления синтетическая последовательность 5'UTR составляет меньше чем около 240 нуклеотидов в длину. В другом варианте осуществления синтетическая последовательность 5'UTR составляет меньше чем около 200 нуклеотидов в длину.
В другом варианте осуществления изобретения синтетический полинуклеотид 5'UTR является компонентом экспрессионного вектора эукариот, содержащего первый нуклеотидный фрагмент и второй полинуклеотидный фрагмент, где:
а. первый полинуклеотидный фрагмент содержит, по меньшей мере, один сайт сплайсинга от первого эукариотического гена;
b. второй полинуклеотидный фрагмент содержит, по меньшей мере, часть 5' нетранслируемой области (5'UTR) второго эукариотического гена; и
с. первый полинуклеотидный фрагмент располагается в 5'-конце второго полинуклеотидного фрагмента.
В одном из вариантов осуществления изобретения полинуклеотидный фрагмент, содержащий, по меньшей мере, один сайт сплайсинга, - это фрагмент эукариотического гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, описанный в настоящей заявке. В другом варианте осуществления полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области, - это фрагмент гена казеина, описанный в настоящей заявке.
Настоящее изобретение также предлагает векторы, содержащие синтетическую 5'UTR, описанную в настоящей заявке. В векторы возможно включить любой из вариантов синтетической полинуклеотидной последовательности 5'UTR, описанных в настоящей заявке. Например, вариантом осуществления изобретения является вектор, содержащий синтетическую полинуклеотидную последовательность 5'UTR, включающую полинуклеотидный фрагмент, содержащий второй интрон эукариотического гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, и полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5'UTR гена казеина.
Другим вариантом изобретения является вектор, содержащий полинуклеотид, который, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичен полинуклеотиду, представленному одной из последовательностей SEQ ID NOS:1-10.
В другом варианте осуществления изобретения вектор является экспрессионным вектором, содержащим синтетическую конструкцию гена, включающую в себя синтетическую 5'UTR. Синтетическая конструкция гена может содержать промотор, расположенный на одном конце синтетической 5'UTR, и интересующую экспрессируемую последовательность, расположенную на другом конце синтетической 5'UTR. Синтетическая конструкция гена может также включать в себя сайт полиаденилирования.
Например, другим вариантом осуществления изобретения является экспрессионный вектор, содержащий синтетическую конструкцию гена, заключающую в себе, в направлении от 5'- к 3'-концу, промотор, химерный полинуклеотид и интересующую экспрессируемую последовательность, при этом:
а. химерный полинуклеотид состоит из полинуклеотидного фрагмента первого эукариотического гена, содержащего, по меньшей мере, один сайт сплайсинга, и полинуклеотидного фрагмента второго эукариотического гена, содержащего, по меньшей мере, часть 5' нетранслируемой области; и
b. химерный полинуклеотид расположен между промотором и интересующей экспрессируемой последовательностью, где полинуклеотидный фрагмент первого эукариотического гена находится ближе к промотору, а полинуклеотидный фрагмент второго эукариотического гена расположен ближе к интересующей экспрессируемой последовательности.
На ФИГУРЕ 3А схематично представлен вариант синтетической конструкции гена, помещенной в каркас вектора. В этом варианте SPL на ФИГУРЕ 3А относится к полинуклеотидному фрагменту, содержащему, по меньшей мере, один сайт сплайринга, а UTR относится к полинуклеотидному фрагменту, содержащему, по меньшей мере, часть 5' нетранслируемой области. Вместе SPL и UTR составляют синтетическую 5'UTR. Промотор синтетической конструкции гена помещен для того, чтобы направлять экспрессию РНК синтетической 5'UTR и интересующей последовательности. Дополнительно, интересующая экспрессируемая последовательность содержит старт-кодон для запуска трансляции.
Образцы экспрессионных векторов, содержащие такое построение, схематично изображены на ФИГУРЕ 10 и ФИГУРЕ 11. Последовательность вектора на ФИГУРЕ 10 представлена в SEQ ID NO:12; последовательность вектора на ФИГУРЕ 11 представлена в SEQ ID NO:13.
В другом варианте осуществления вектор является экспрессионным вектором, содержащим, в направлении от 5'- к 3'-концу, промотор, химерный полинуклеотид и сайт клонирования, где:
а. химерный полинуклеотид состоит из полинуклеотидного фрагмента первого эукариотического гена, содержащего, по меньшей мере, один сайт сплайсинга, и полинуклеотидного фрагмента второго эукариотического гена, содержащего, по меньшей мере, часть 5' нетранслируемой области; и
b. химерный полинуклеотид расположен между промотором и сайтом клонирования, при этом полинуклеотидный фрагмент первого эукариотического гена находится ближе к промотору, а полинуклеотидный фрагмент второго эукариотического гена находится ближе к сайту клонирования.
Сайт клонирования экспрессионного вектора может содержать один или более уникальный сайтов рестрикции для того, чтобы можно было включить интересующую последовательность. В другом варианте осуществления сайт клонирования включает сайты связывания рекомбиназы для того, чтобы интересующие последовательности могли быть включены путем сайт-специфической рекомбинации.
Вариант экспрессионного вектора схематично изображен на ФИГУРЕ 3В. На ФИГУРЕ 3В SPL относится к полинуклеотидному фрагменту, содержащему сайт сплайсинга, a UTR относится к полинуклеотидному фрагменту, заключающему в себе, по меньшей мере, часть 5' нетранслируемой области. SPL и UTR вместе образуют синтетическую 5'UTR.
Полинуклеотидный фрагмент, содержащий сайт сплайсинга в экспрессионном векторе, может быть фрагментом любого эукариотического гена. Примеры полинуклеотидных фрагментов, которые могут быть использованы в экспрессионном векторе, включают те из описанных в настоящей заявке фрагментов, которые содержат в себе сайт сплайсинга эукариотического гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, описанного в настоящей заявке. Далее, полинуклеотидный фрагмент, содержащий, по меньшей мере, часть 5' нетранслируемой области, может быть фрагментом любого эукариотического гена. Примеры полинуклеотидных фрагментов, которые могут быть использованы в экспрессионном векторе, включают те из описанных в настоящей заявке фрагментов, которые содержат в себе, по меньшей мере, часть 5'UTR гена казеина, описанного в настоящей заявке.
Экспрессионные векторы изобретения в дальнейшем могут содержать одну или более дополнительных полинуклеотидных последовательностей, расположенных после интересующей последовательности или сайга клонирования, для того, чтобы создать слитную конструкцию с полипептидом, закодированным интересующей последовательностью. Например, дополнительные полинуклеотиды, расположенные после интересующей последовательности, могут кодировать эпитопную метку, репортер, метку для специфичной очистки. Эпитопные метки известны специалистам в данной области и включают myc, гемаглютинин (НА) и FLAG. Примеры репортеров включают зеленый флуоресцирующий белок и его варианты, бета-галактозидазу (LacZ), бета-глукоронидазу (Gus), хлорамфеникол ацетилтрансферазу (CAT) и люциферазу. Примеры меток для специфичной очистки включают His6 и GST. Экспрессионный вектор может также содержать полиА-сайт, расположенный после интересующей последовательности или сайта клонирования.
В зависимости от желаемого результата промоторная часть экспрессионного вектора, содержащая синтетическую 5'UTR, может быть конститутивным промотором, неконститутивным промотором, тканеспецифическим промотором (конститутивным или неконститутивным), промотором, связанным с патогенезом или заболеванием, промотором, специфичным к стадии развития, или избирательно контролируемым промотором, таким как индуцируемый промотор. Разные избирательно контролируемые промоторы находятся под управлением разных механизмов. Например, тетрациклин-индуцируемый промотор активируется для экспрессии расположенной за ним кодирующей последовательности, когда клетки, содержащие промотор и другие необходимые клеточные факторы, обработаны тетрациклином. Другие индуцируемые промоторы активируются другими лекарственными веществами или факторами. RHEOSWITCH - это система индуцируемого промотора, производимая New England Biolabs (Ipswich, MA). Чувствительные к температуре промоторы также могут быть использованы для увеличения или уменьшения экспрессии гена. Вариант осуществления изобретения включает генную конструкцию, заключающую в себе синтетическую 5'UTR, экспрессия которой контролируется индуцируемым промотором.
Изобретение включает варианты, где каркас вектора, содержащий синтетическую 5'UTR, включает последовательности, подходящие для экспрессии интересующей последовательности в эукариотической клетке. В одном из вариантов изобретения каркас вектора, содержащего синтетическую 5'UTR, включает последовательности, подходящие для экспрессии интересующей последовательности в клетке млекопитающих. Экспрессионные векторы млекопитающих могут содержать нетранскрибируемые элементы, такие как сайт инициации репликации, пригодный промотор и энхансер, связанные с интересующей экспрессируемой последовательностью, другие расположенные в 5'- или 3'-конце нетранскрибируемые последовательности и 5' или 3' нетранслируемые последовательности, такие как необходимые сайты связывания рибосом, сайт полиаденилирования и транскрипционные терминирующие последовательности. Примеры экспрессионных векторов млекопитающих хорошо известны специалистам в данной области и включают pcDNA3 (Инвитроген) и pRSVneo (АТТС).
Например, промоторная часть экспрессионного вектора, содержащая синтетическую 5'UTR, может быть промотором животных или млекопитающих. Примеры промоторов животных или млекопитающих включают раннюю (SV40e) промоторную область SV40, промотор, содержащийся в 3' длинном концевом повторе (LTR) вируса саркомы Рауса (RSV), промоторы Е1А или главный поздний промотор (MLP) генов аденовирусов (Ad), ранний промотор цитомегаловируса (CMV), промотор гена тимидинкиназы вируса простого герпеса (HSV), промотор фактора элонгации 1 альфа (EF1), промотор гена фосфоглицераткиназы (PGK), промотор убиквитина (Ubc), промотор альбумина, регуляторные последовательности промотора металлотионеина-L мыши и транскрипционные контрольные области, убиквитарные промоторы (гипоксантингуанин-фосфорибозилтрансферазы (HPRT), виментина, бета-актина, тубулина и т.п.), промоторы промежуточных филаментов (десмина, нейрофиламентов, кератина, GFAP и т.п.), промоторы терапевтических генов (MDR, CFTR или фактора VIII типа и т.п.), связанные с патогенезом или заболеванием промоторы и промоторы, которые проявляют тканеспецифичность и были использованы в трансгенных животных, такие как контрольная область гена эластазы I, которая активна в панкреатических ацинарных клетках; активная в панкреатических бета-клетках контрольная область гена инсулина, активная в лимфоидных клетках контрольная область гена иммуноглобулина, активная в тестикулярных, лимфоидных и тучных клетках и клетках молочных желез контрольная область гена вируса рака молочных желез мышей; активные в печени контрольные области гена альбумина, Аро AI и Аро AII, активная в печени контрольная область гена альфа-фетопротеина, активная в печени контрольная область гена альфа 1-антитрипсина, активная в миелоидных клетках контрольная область гена бета-глобина, активная в олигодендроцитарных клетках мозга контрольная область гена основного миелинового белка, активная в скелетной мышце контрольная область гена легкой миозиновой цепи-2 и активная в гипоталамусе контрольная область гена гонадотропного рилизинг-гормона, промотор гена пируват-киназы, промотор гена виллина, промотор гена интестинального белка, связывающего жирные кислоты, промотор гена бета-актина в гладко-мышечных клетках и т.п. Промоторы в экспрессионном векторе могут быть модифицированы путем включения энхансера или регуляторных последовательностей и т.п.
Интересующая последовательность, как часть экспрессионного вектора, содержащего синтетическую 5'UTR, может быть последовательностью любого эукариотического гена, или его частью, или естественной или искусственной кодирующей или некодирующей последовательностью. Примеры кодирующих последовательностей, которые могут быть использованы в настоящем изобретении, включают последовательности, кодирующие репортеры (например, люцифераза, бета-галактозидаза, флуоресцентные белки), эпитопы, опытные полипептиды или терапевтические полипептиды. Дополнительные примеры интересующих последовательностей нуклеиновых кислот включают молекулы РНК, такие как малые РНК, микро-РНК, рибосомальные РНК, терапевтические РНК и рибозимы.
В другом аспекте изобретения множественные неизбыточные синтетические 5'UTR применяются в контексте мультигенных конструкций внутри одного вектора.
В другом варианте осуществления изобретения вектор является вектором для генной терапии, содержащим синтетическую конструкцию гена, в которой находится синтетическая 5'UTR. Вектор для генной терапии может быть любым вектором для генной терапии, известным в данной области и включающим вирусные или невирусные векторы, такие как вектор на основе аденовируса, вектор на основе аденоассоциированного вируса (AAV) или вектор на основе ретровируса. Синтетическая конструкция гена может содержать промотор, расположенный на 5'-конце синтетической 5'UTR, и интересующий терапевтический ген, расположенный на 3'-конце синтетической 5'UTR, как показано на ФИГУРЕ 3А. Промотор может быть конститутивным промотором, тканеспецифическим промотором, индуцируемым промотором или иным промотором, описанным здесь. Примеры классов терапевтических генов интереса, которые могут быть включены в вектор для генной терапии, включают, но не ограничиваются указанными, гены, кодирующие цитокины, хемокины, гормоны, антитела, изобретенные иммуноглобулиноподобные молекулы, одноцепочечные антитела, слитные белки, ферменты, иммунные костимулирующие молекулы, иммуномодулирующие молекулы, трансдоминантные негативные мутанты целевых белков, токсины, условные токсины, хемотерапевтические или радиотерапевтические сенсибилизирующие вещества, антигены, опухолевые белки-супрессоры, факторы роста, мембранные белки, вазоактивные белки и пептиды, антивирусные белки или их варианты.
Конкретных примеров интересующих терапевтических генов множество, они включают, но не ограничиваются указанными, эритропоэтин, инсулин, VEGF, FGF, Фактор XVIII, Фактор ГХ, эндостатин, ангиостатин, GDNF, BDNF, NGF, EFG, CFTR, PEGF, IFN-alfa, IFN-gamma, IL-1, IL-2, IL-4, IL-6, IL-7, IL-10. IL-12, IL-21, GM-CSF, M-CSF, TNF-α, TNF-β, TGF-α, TGF-β, CD40, гирудин и т.п.
Настоящее изобретение также предлагает наборы, содержащие полинуклеотидные последовательности изобретения. Например, в одном из вариантов осуществления настоящее изобретение предлагает набор, содержащий, по меньшей мере, один полинуклеотид, который, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичен полинуклеотиду, представленному одной из последовательностей SEQ ID NOS:1-10. Наборы могут содержать вышеупомянутые векторы или компоненты, которые позволяют их сборку. Например, набор может включать вектор, который может быть переведен в линейную форму путем расщепления ферментом рестрикции в сайте, который позволяет пользователю включать синтетическую 5'UTR или отдельные компоненты синтетической 5'UTR. В дальнейшем набор может содержать дополнительные компоненты для сборки вектора, такие как фермент рестрикции, лигазу, буфер и т.п.
Дополнительно настоящее изобретение предлагает способы экспрессии продукта гена или интересующей последовательности в клетке-хозяине.
Например, другим вариантом осуществления изобретения является способ экспрессии продукта гена, включающий трансфекцию клетки-хозяина синтетической конструкцией гена, содержащей синтетическую 5'UTR, описанную в настоящей заявке.
Другим вариантом осуществления изобретения является способ экспрессии продукта гена, включающий трансфекцию клетки-хохяина синтетической конструкцией гена, содержащей полинуклеотид, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный полинуклеотиду, представленному последовательностью SEQ ID NO:7.
Другим вариантом осуществления изобретения является способ экспрессии интересующей последовательности в клетке-хозяине, включающий этапы:
а. трансфекции клетки-хозяина экспрессионным вектором, содержащим синтетическую конструкцию гена, описанную в настоящей заявке; и
b. культивирования вышеупомянутой клетки-хозяина в условиях, подходящих для экспрессии вышеуказанной интересующей последовательности.
В другом способе экспрессии интересующей последовательности в клетке-хозяине интересующая последовательность может быть включена в описанный в настоящей заявке экспрессионный вектор, содержащий промотор, синтетическую 5'UTR и сайт клонирования. Например, другим вариантом осуществления изобретения является способ экспрессии интересующей последовательности в клетке-хозяине, включающий этапы:
а. включения интересующей экспрессируемой последовательности в описанный в настоящей заявке экспрессионный вектор, содержащий в клонирующем сайте промотор, синтетическую 5'UTR и клонирующий сайт, где интересующая экспрессируемая последовательность содержит РНК или полипептидную кодирующую последовательность; включения интересующей экспрессируемой последовательности в описанный здесь экспрессионный вектор, содержащий промотор, синтетическую 5'UTR и сайт клонирования, в сайт клонирования, где интересующая экспрессируемая последовательность содержит кодирующую последовательность РНК или полипептида;
b. трансфекции клетки-хозяина экспрессионным вектором; и
с. культивирования вышеуказанной клетки-хозяина в условиях, подходящих для получения экспрессии вышеупомянутой интересующей последовательности.
В другом способе экспрессии интересующей последовательности в клетке-хозяине синтетическая 5'UTR может быть включена в экспрессионный вектор между промотором и интересующей последовательностью таким образом, что часть, содержащая элемент сплайсинга эукариотического гена внутри синтетической 5'UTR, располагается ближе к промотору, а часть, содержащая, по меньшей мере, часть 5'UTR другого эукариотического гена, находится ближе к интересующей последовательности.
Например, другим вариантом осуществления изобретения является споосб экспрессии интересующей последовательности в клетке-хозяине, содержащий этапы:
а. включения синтетической 5'UTR, описанной в настоящей заявке, в экспрессионный вектор между промотором и интересующей экспрессируемой последовательностью, где интересующая экспрессируемая последовательность включает кодирующую последовательность РНК или полипептида;
b. трансфекции клетки-хозяина вышеупомянутым экспрессионным вектором; и
с. культивирования вышеуказанной клетки-хозяина в условиях, подходящих для экспрессии вышеупомянутой интересующей последовательности.
Другим вариантом осуществления изобретения является метод экспрессии интересующей последовательности в клетке-хозяине, содержащий этапы:
а. включения полинуклеотида, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичного полинуклеотиду, представленному последовательностью SEQ ID NO:1 или SEQ ID NO:7, в экспрессионный вектор между промотором и интересующей экспрессируемой последовательностью, где интересующая экспрессируемая последовательность включает кодирующую последовательность РНК или полипептида;
b. трансфекции клетки-хозяина вышеуказанным экспрессионным вектором; и
с. культивирования вышеупомянутой клетки-хозяина в условиях, подходящих для экспрессии вышеуказанной интересующей последовательности.
Способы согласно изобретению осуществляют с применением стандартных технологий. Если не указано иное, технологии рекомбинантной ДНК осуществляются согласно стандартным методикам, описанным в (Sambrook, supra) или в Томе 1 и 2 Ausubel et al. (1994), Current Protocols in Molecular Biology, Current Protocols.
Настоящее изобретение также предлагает клетки-хозяева, содержащие синтетическую 5'UTR. Клетки-хозяева подготовлены для включения любого из вариантов полинуклеотидных последовательностей синтетических 5'UTR, описанных в настоящей заявке. Например, другим вариантом осуществления изобретения является клетка-хозяин, содержащая синтетическую полинуклеотидную последовательность 5'UTR, включающую полинуклеотидный фрагмент, заключающий в себе второй интрон гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, слитый с полинуклеотидным фрагментом, содержащим, по меньшей мере, часть области 5'UTR гена казеина.
В одном из вариантов изобретения клетка-хозяин является клеткой-хозяином млекопитающих. В конкретных вариантах осуществления клетка-хозяин является клеткой хомяка, клеткой мыши, клеткой крысы, клеткой кролика, клеткой кошки, клеткой собаки, клеткой быка, клеткой козы, клеткой коровы, клеткой свиньи, клеткой лошади, клеткой овцы, клеткой макаки, клеткой обезьяны, клеткой шимпанзе или клеткой человека. Конкретные примеры клеток-хозяев изобретения, содержащие синтетическую 5'UTR, включают, но не ограничиваются указанными, А549, ARPE-19, CH3/10T1/2, С2С12, асо2, COS7, FL-83B, НЕК-293, HEPG2, HeLa, HT-1080, MDCK, P19, SH-SY5Y, Sol 8 и U87.
Клетки-хозяева согласно изобретению трансфицируют полинуклеотидом, содержащим синтетическую 5'UTR. Трансфекция клетки-хозяина хорошо известна специалистам в данной области и может быть достигнута с помощью разнообразных методик, включающих в качестве неограниченных примеров электропорацию, вирусное инфицирование, трансфекцию системой плазмида/вектор, опосредованную невирусным вектором трансфекцию, бомбардировку частицами и т.п. Экспрессия желаемых продуктов гена включает культивирование трансфицированных клеток-хозяев в подходящих условиях и оценку экспрессии трансфицированного гена. Протоколы условий культивирования и экспрессии гена хорошо известны специалистам в данной области техники.
Настоящее изобретение также предлагает организмы-хозяева, содержащие синтетические 5'UTR, описанные в настоящей заявке. Синтетические 5'UTR могут быть включены in vivo непосредственно в геном хозяйского организма. Например, в одном из вариантов осуществления изобретения синтетическую 5'UTR вводят в организм-хозяин с целью заменить нормальную 5'UTR на непосредственно введенный в организм-хозяин вектор, который содержит синтетическую 5'UTR изобретения, фланкированную последовательностями, гомологичными последовательностям, фланкирующим нормальную 5'UTR, которую необходимо заменить. В другом варианте синтетическую конструкцию гена, содержащую синтетическую 5'UTR, включают в геном организма-хозяина путем введения в организм-хозяин интегрирующегося вектора, заключающего в себе конструкцию гена. Тканеспецифичностью встраивания можно управлять, например, с помощью способа введения вектора. В другом варианте синтетическая конструкция гена, содержащая синтетическую 5'UTR, вводится в организм-хозяин путем введения в организм-хозяин неинтегрирующегося вектора, включающего в себя конструкцию гена.
Синтетическую 5'UTR или синтетическую конструкцию гена, содержащую синтетическую 5'UTR, можно также ввести в организм-хозяин способами ех vivo. Например, аутологичные или неаутологичные клетки можно трансформировать вектором, содержащим синтетическую 5'UTR, а затем ввести в организм-хозяин.
В другом варианте осуществления синтетическую 5'UTR вводят в геном клетки-хозяина или организм, используя систему сайт-специфичной рекомбиназы, такую как система Cre/loxP. В этом варианте стабильная интреграция в геном фрагмента ДНК, такого как фрагмент, содержащий синтетическую 5'UTR или синтетическую конструкцию гена, заключающего в себе синтетическую 5'UTR, достигается путем введения модифицированных lox-сайтов в геном и донорный вектор, что препятствует реэксцизии интрегрированной ДНК (см. Metzger and Feil, Current Opinion in Biotechnology, 10:470-476 (1999), включенный в настоящее описание посредством ссылки). Далее, гетероспецифичные loxP-сайты могут быть введены ('floxed') в геном с целью фланкировать область, которую необходимо заместить (Metzger and Feil, выше), такую как эндогенная 5'UTR и синтетическая 5'UTR в донорной плазмиде. Трансгенных животных с хромосомными сайтами lox P, введенными в геном, можно скрестить с трансгенными мышами, экспрессирующими Сrе-рекомбиназу под управлением тканевого или клеточного промотора, известными специалистам в данной области техники. Введение донорной плазмиды, содержащей синтетическую 5'UTR, в потомство, получаемое в результате такого скрещивания, приведет к интеграции ее внутри специфических типов тканей и клеток.
В других вариантах иные сайт-специфичные рекомбинационные системы, такие как те, что описаны в публикации заявки на патент США No.20060172377, включенной в настоящее описание посредством ссылки, применяются для введения синтетической 5'UTR в геном организма-хозяина.
Другим вариантом изобретения является организм, не относящийся к человеку, содержащий полинуклеотид, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный полинуклеотиду, представленному последователньостью SEQ ID NOS:1-10.
Другим вариантом осуществления изобретения является организм, не относящийся к человеку, содержащий синтетическую конструкцию гена, заключающую в себе полинуклеотид, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный полинуклеотиду, представленному последовательностью SEQ ID NOS:1-10.
Дополнительно, можно получить трансгенный организм, такой как организм мыши, содержащий синтетическую 5'UTR, или конструкцию гена, содержащую синтетическую 5'UTR. Например, трансгенные мыши могут быть получены путем инъекции соответствующего вектора, содержащего синтетическую 5'UTR, в пронуклеусы оплодотворенных ооцитов мыши. В качестве альтернативы вектор может быть введен в эмбриональные стволовые клетки мыши, которые затем при помощи микроинъекций вводят в бластоцисты мыши. Трансформированные зиготы или бластоциты далее трансплантируют в женскую особь с ложной беременностью. Полученных детенышей подвергают скринингу на наличие полинуклеотидов при помощи ПЦР или Саузерн-блоттинга. Гетерозиготные трансгенные животные затем скрещиваются друг с другом с целью получения гомозигот. В одном из вариантов в трансгенном животном синтетическая 5'UTR замещает эндогенную 5'UTR интересующего гена путем гомологичной рекомбинации.
Так, другим вариантом осуществления изобретения является трансгенное животное, содержащее синтетическую полинуклеотидную последовательность 5'UTR, заключающую в себе фрагмент эукариотического гена кальциевой АТФазы саркоплазматического/эндоплазматического ретикулума, который содержит второй интрон, слитый с фрагментом, содержащим, по меньшей мере, часть 5'UTR гена казеина.
Другим вариантом изобретения является трансгенный организм, содержащий полинуклеотид, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный полинуклеотиду, представленному последовательностью SEQ ID NOS:1-10.
Другим вариантом осуществления изобретения является трансгенный организм, содержащий синтетическую конструкцию гена, заключающую в себе полинуклеотид, по меньшей мере, на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный полинуклеотиду, представленному последовательностью SEQ ID NOS:1-10.
Следующие примеры иллюстрируют, но не ограничивают, варианты осуществления настоящего изобретения. Другие применимые модификации и адаптации, которые очевидны специалистам в данной области, не выходят за рамки настоящего изобретения и соответствуют его сущности.
Примеры
Пример 1
Были получены и включены в вектор три разных варианта синтетической 5'UTR, при этом промотор цитомегаловируса (CMV) человека управляет экспрессией репортера бета-галактозидазы (кодирующая последовательность LacZ). В варианте один полиG (12) в качестве синтетической 5'UTR встраивали между промотором и репортером бета-галактозидазы (ФИГУРА 9). В варианте два, называемом 5U2, SEQ ID NO:1 включали между промотором и репортером бета-галактозидазы в качестве синтетической 5'UTR (ФИГУРА 10). В варианте 3, называемом INXN-I, SEQ ID NO:7 встраивали между промотором и репортером бета-галактозидазы в качестве синтетической 5'UTR (ФИГУРА 11). Каждый трансген имел последовательность полиаденилирования SV40 в 3' регуляторной области. Общий вариант каждого вектора, содержащего синтетическую 5'UTR 5U2 или INXN-I, схематично показан на ФИГУРЕ 3А. Клетки НЕК-293 и клетки 1080 были временно трансфицированы каждым вектором.
Бета-галактозидазу из клеток оценивали с использованием Системы Галакто-Стар™ от Applied Biosystems (Cat Nos. BM100S, BM300S, BM2500S, BY100S, BY300S, BY2500S) согласно следующей методике. Клетки лизировали в 50 мкМ лизисного раствора (Система Галакто-Стар™) в течение 10 мин при комнатной температуре. 100 мкМ субстрата Галакто-Стар® распределяли аликвотами по лункам белой непрозрачной 96-луночной микропланшеты. 10 мкМ клеточного лизиса добавляли к 100 мкМ субстрата Галакто-Стар® и инкубировали в течение 30 мин. Световой сигнал оценивали с помощью микропланшетного люминометра.
Результаты для клеток НЕК-293 показаны на ФИГУРЕ 4. Экспрессия репортера бета-галактозидазы была явно увеличена в НЕК-293 клетках, трансфицированных векторами, содержащими синтетическую 5'UTR INXN-1 или 5U2, по сравнению с векторами, содержащими полиG в качестве 5'UTR. Увеличенная экспрессия репортера в клетках 1080, трансфицированных векторами, содержащими синтетическую 5'UTR INXN-1 или 5U2, по сравнению с векторами, содержащими полиG в качестве 5'UTR, показаны на ФИГУРЕ 5. Уровень экспрессии трансгенов в векторах, содержащих полиG в качестве 5'UTR, был подобен контролю, где не было 5'UTR (VVN-2712, ФИГУРА 7, данные на ФИГУРЕ 12). Отрицательным контролем при оценке фонового уровня был вектор без бета-галактозидазы (VVN-2713, ФИГУРА 8, данные на ФИГУРЕ 12). На ФИГУРЕ 6 показано, что синтетический ген-репортер бета-галактозидазы, содержащий синтетическую 5'UTR INXN-1, был экспрессирован примерно в 7.5 раз выше в клетках НЕК-293 и примерно в 2.5 раз выше в клетках 1080 по сравнению с синтетическим геном-репортером, содержащим полиG в качестве 5'UTR, в то время как синтетический ген-репортер, содержащий синтетическую 5'UTR INXN-1 или 5U2, был экспрессирован примерно в 8 раз выше в клетках НЕК-293 и примерно в 2.5 раза выше в клетках 1080 по сравнению с синтетическим геном-репортером, содержащим полиG в качестве 5'UTR.
Пример 2.
В результате поиска в публичной неизбыточной нуклеотидной базе данных GenBank с применением blastn-алгоритма с параметрами по умолчанию с использованием SEQ ID NO:4 в качестве запрашиваемой последовательности, были найдены следующие типичные гомологи SERCA2, перечисленные в Таблице 1. SEQ ID NO:4 представляет собой компонент синтетической 5'UTR, представленной SEQ ID NO:7.
|
Результаты идентифицированного с помощью BLASTN-анализа фрагмента генома Equus caballus, приведенные в Таблице 1 (общий номер доступа AM 13 7440), сравнивали с последовательностью мРНК SERCA2 NM_001081765 Equus caballus, при помощи парного анализа линейного соответствия в общедоступной программе DNA Stricter 1.4f17 (см. описания DNA Strider в Mark, С., Исследования нуклеиновых кислот 16(5):1829-183 7 (1988) и Douglas, D, Молекулярные биотехнологии 3(1):37-45 (1995)). Последовательности выравнивали с помощью метода блоков с использованием параметров по умолчанию для mismatch penalty and gap penalty. Часть линейного сравнения геномной и мРНК последовательности Equus caballus показаны на ФИГУРЕ 13, где стрелками отмечены начало и конец второго интрона гена SERCA2 Equus caballus. Область гомологии, которая находится 5' относительно интрона, представлена экзоном 2, а область гомологии, которая находится 3' относительно интрона, представлена экзоном 3.
Олигонуклеотид, представленный фрагментом гена SERCA2 Equus caballus, содержащий второй интрон, затем был синтезирован и соединен с SEQ ID NO:3 с помощью технологий рекомбинантной ДНК, чтобы получить синтетическую 5'UTR. После синтетическую 5'UTR встраивали в вектор между промотором цитомегаловируса человека и репортерным геном люциферазы. Вектор и контрольный вектор с полиG 5'UTR затем использовали для трансфекции 3Т3-клеток. Активность люциферазы оценивали с помощью люминометра, а относительные световые единицы сравнивали между двумя группами клеток. Экспрессия репортера в клетках, трансфицированных содержащим синтетическую 5'UTR вектором, увеличивалась по сравнению с экспрессией в клетках, трансфицированных содержащим полиG 5'UTR вектором.
Пример 3
В результате поиска в публичной неизбыточной нуклеотидной базе данных GenBank с применение blastn-алгоритма с параметрами по умолчанию, с использованием SEQ ID NO:3 в качестве запрашиваемой последовательности были найдены следующие типичные гомологи, перечисленные в Таблице 2. SEQ ID NO:3 представляет собой компонент 5'UTR казеина быка, входящий в синтетическую 5'UTR, представленную SEQ ID NO:1 и SEQ ID NO:7.
|
Второй интрон каждого типичного представителя гена SERCA2, перечисленного в Таблице 1, идентифицировали с помощью программы для парного линейного сравнения путем сравнения геномной последовательности с ее соответствующей последовательностью мРНК. Была синтезирована первая группа олигонуклеотидов, содержащих второй интрон SERCA2 для каждого гомолога SERCA2. Затем синтезировали вторую группу олигонуклеотидов, содержащих, по меньшей мере, часть 5'UTR для каждого гомолога бета-казеина, идентифицированного в Таблице 2. Часть 5'UTR содержит часть покрытия запроса, идентифицированного в BLAST-результатах Таблицы 2. После группу синтетических 5'UTR получали с помощью технологии рекомбинантной ДНК путем слияния уникального представителя из первой группы олигонуклеотидов, содержащих второй интрон SERCA2, или олигонуклеотида, представленного последовательностями SEQ ID NOS:2 и 4-6, с уникальным представителем из второй группы олигонуклеотидов, содержащих, по меньшей мере, часть 5'UTR, или олигонуклеотидом, представленным последовательностями SEQ ID NOS:3 и 8-10, где олигонуклеотид, содержащий второй интрон SERCA2, соединен 5'-концом с олигонуклеотидом, содержащим часть 5'UTR. Затем с помощью ПЦР к каждому концу синтетической 5'UTR добавляли сайты рестрикции. Далее каждую уникальную синтетическую 5'UTR встраивали в вектор между промотором цитомегаловируса человека и промотором LacZ. После чего в 96-луночных планшетах клетки 1080 трансфицировали каждым вектором, а также контрольным вектором с полиG 5'UTR. Далее активность бета-галактозидазы оценивали с помощью анализа, описанного в Примере 1, уровень экспрессии оценивали относительно полиG 5'UTR и затем сравнивали.
После ознакомления с полным описанием изобретения специалистам в данной области техники будет ясно, что то же самое можно проделать в широком и эквивалентном круге условий и других параметров без влияния на объем изобретения или любые его варианты.
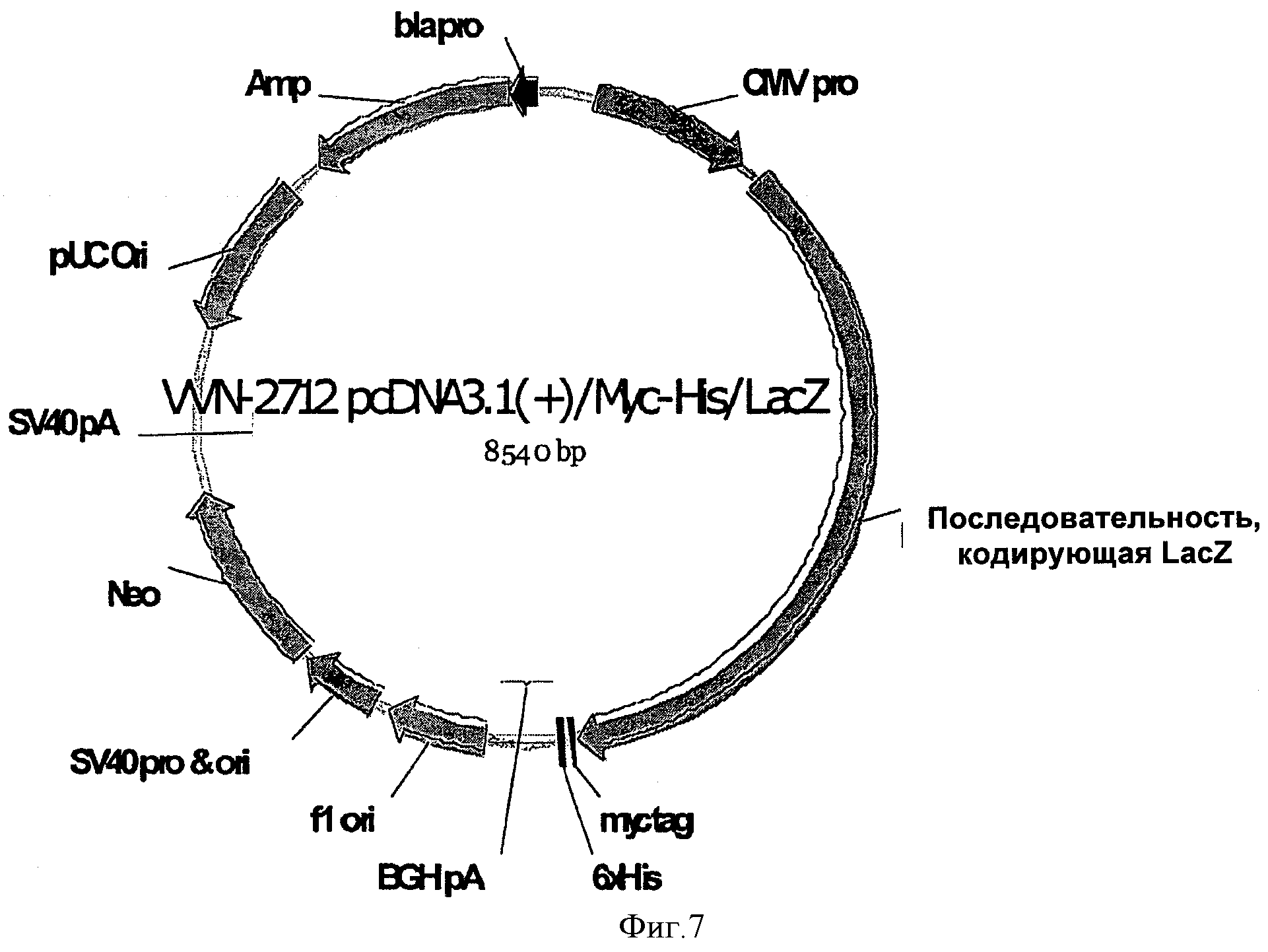

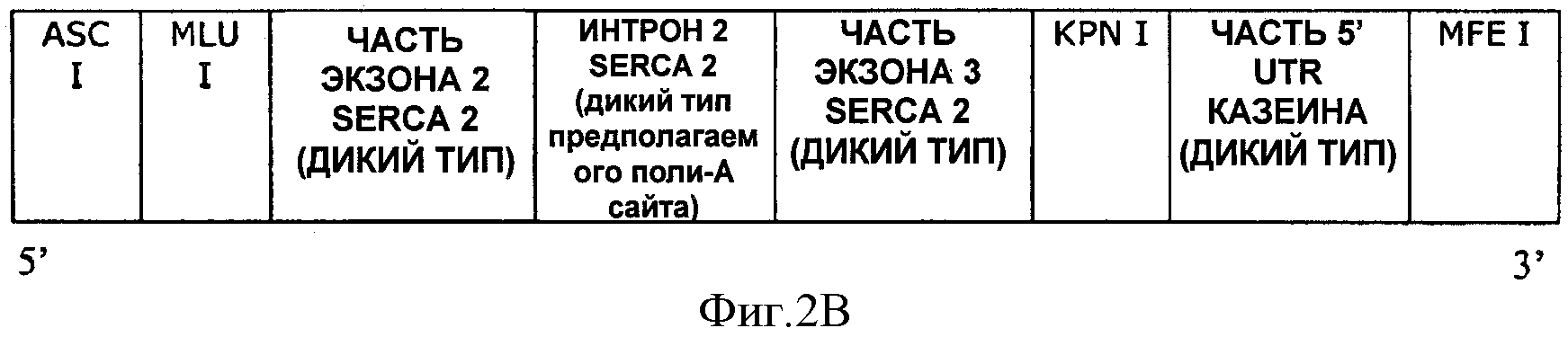

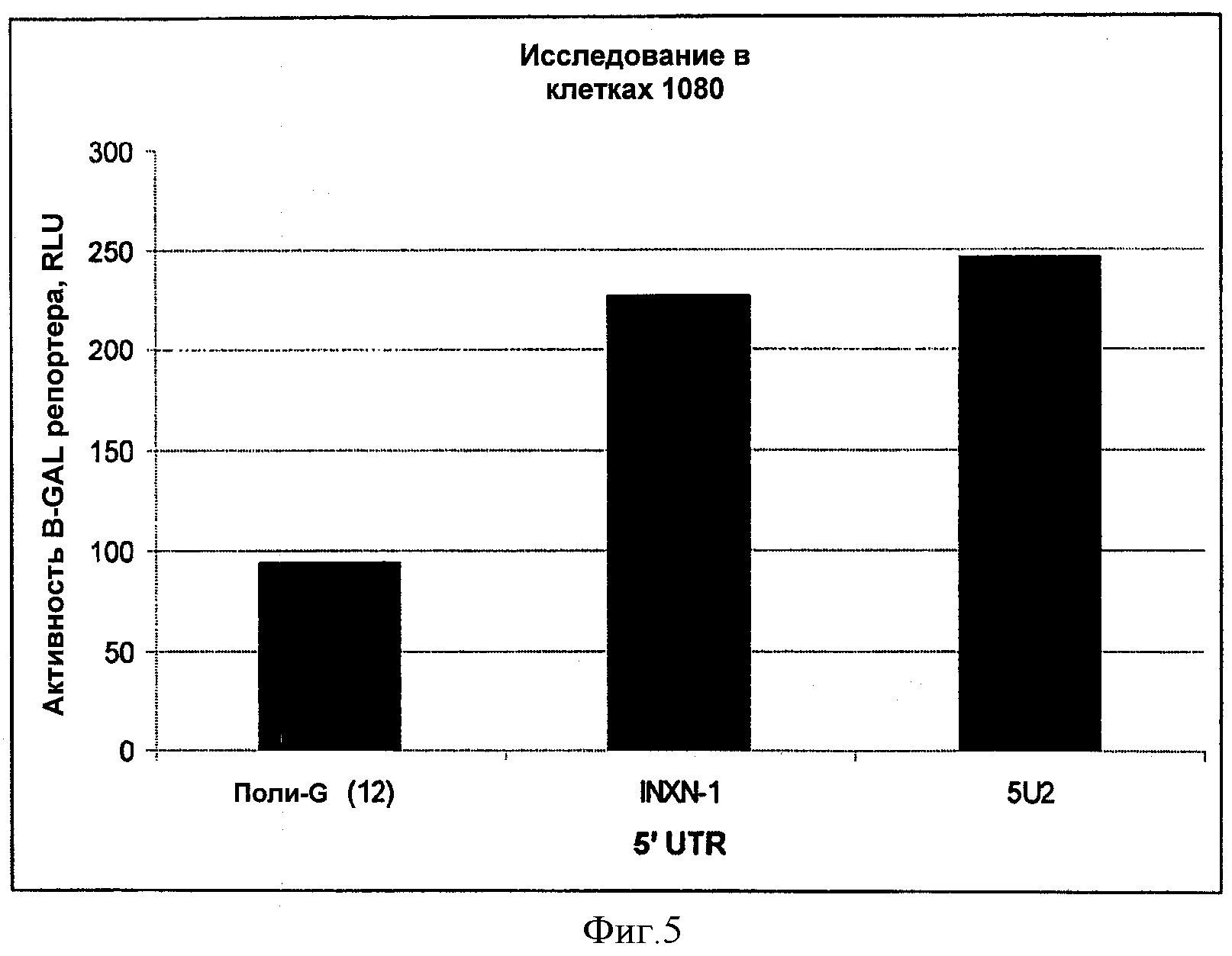

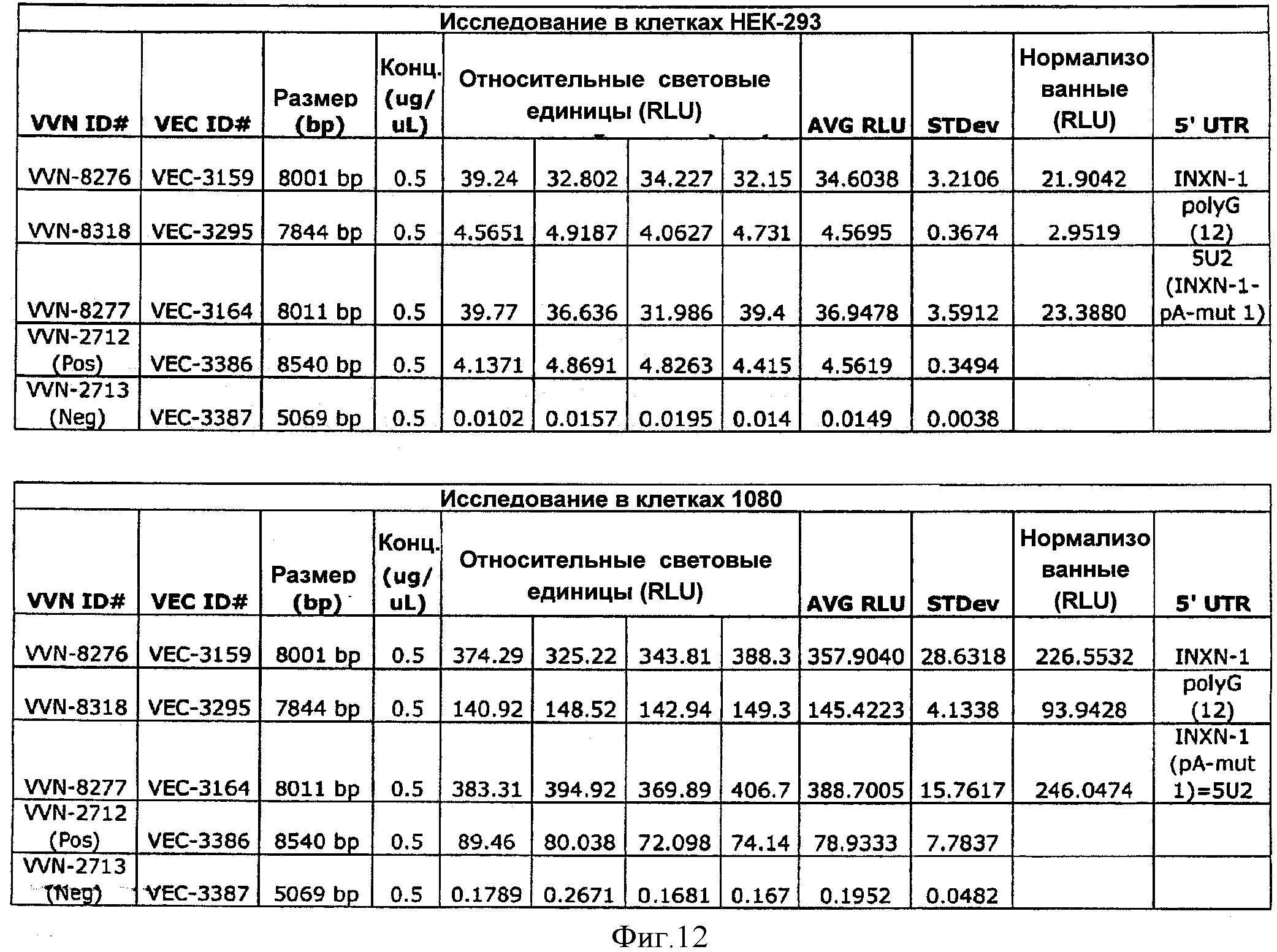


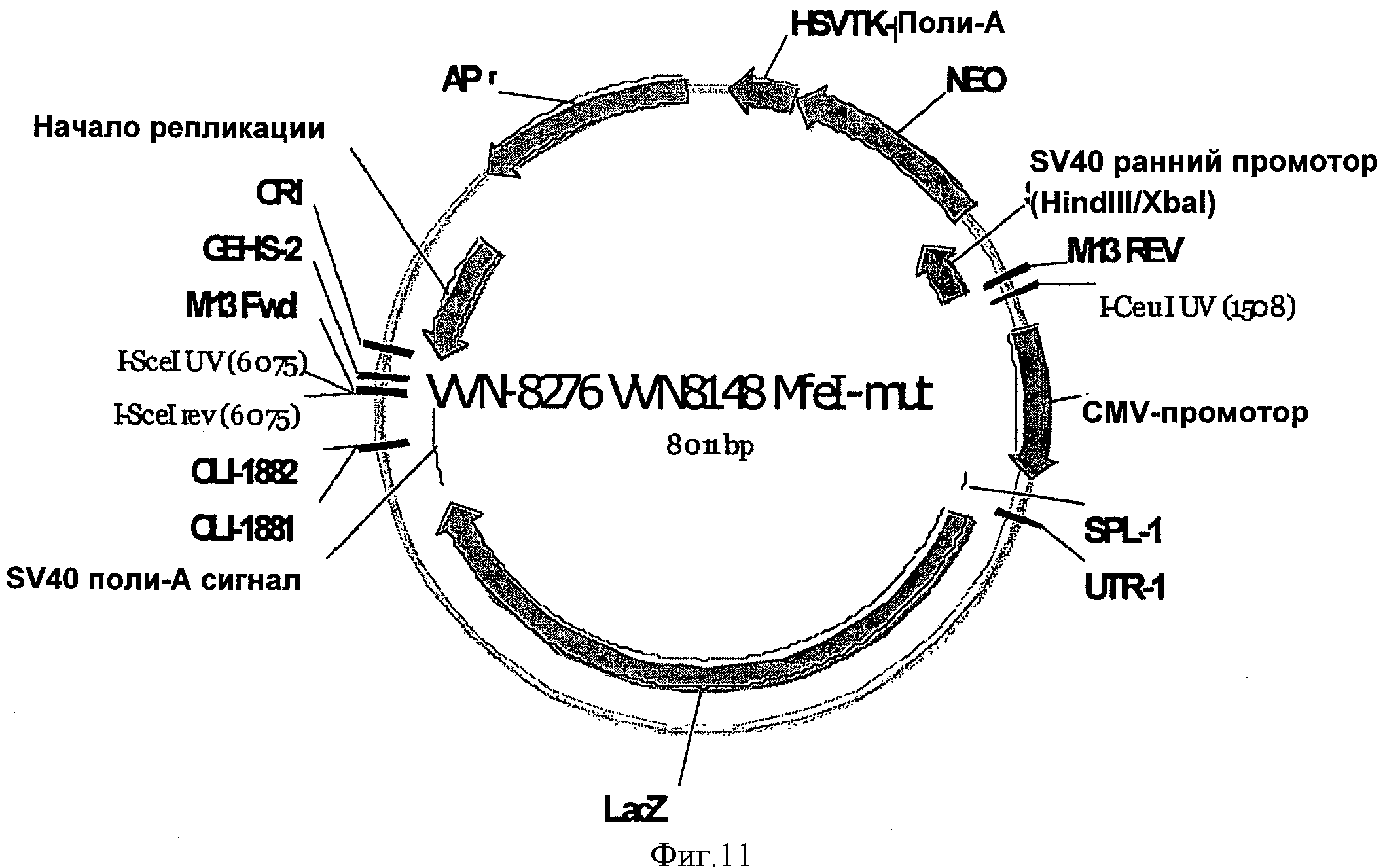
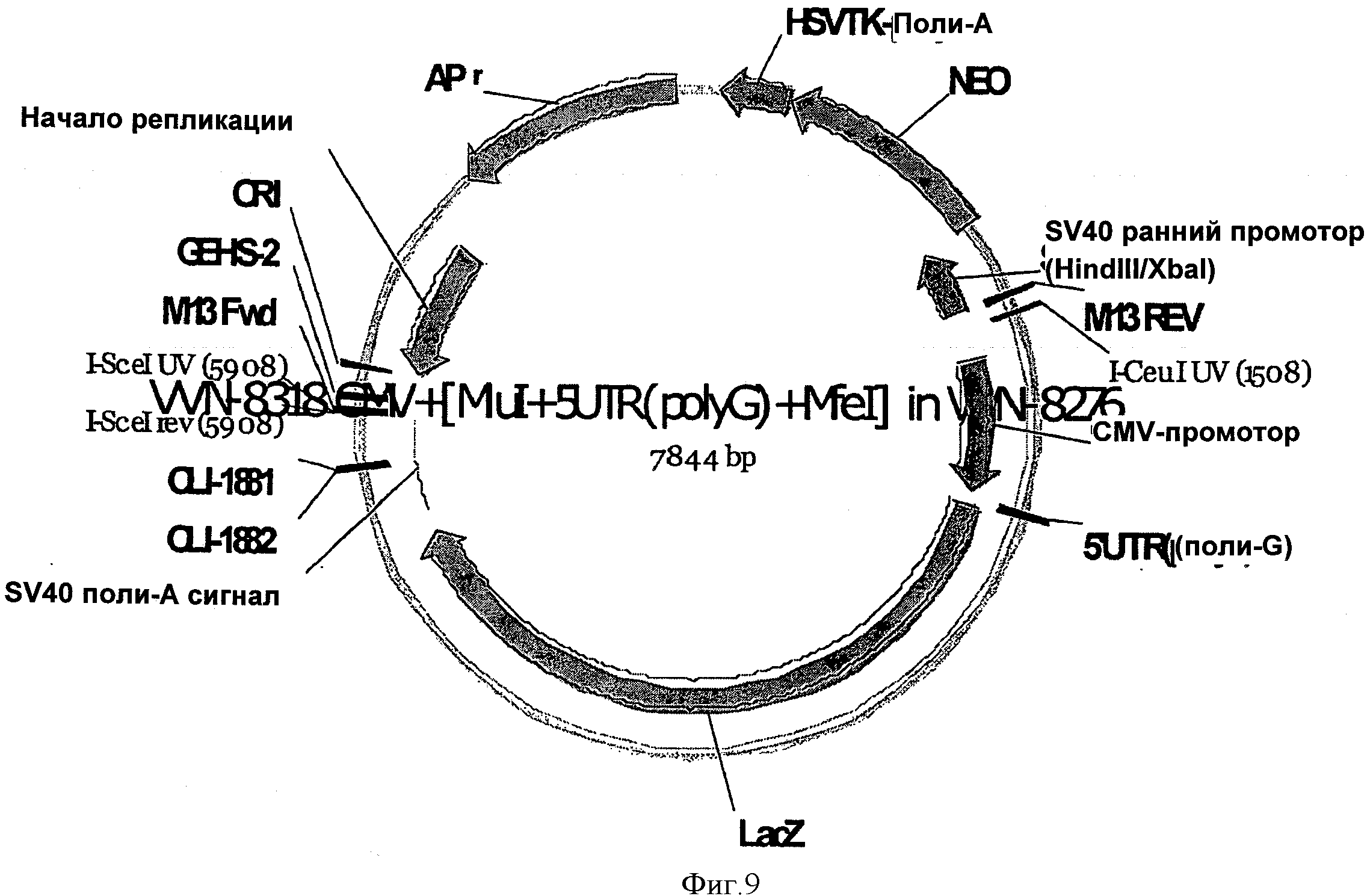





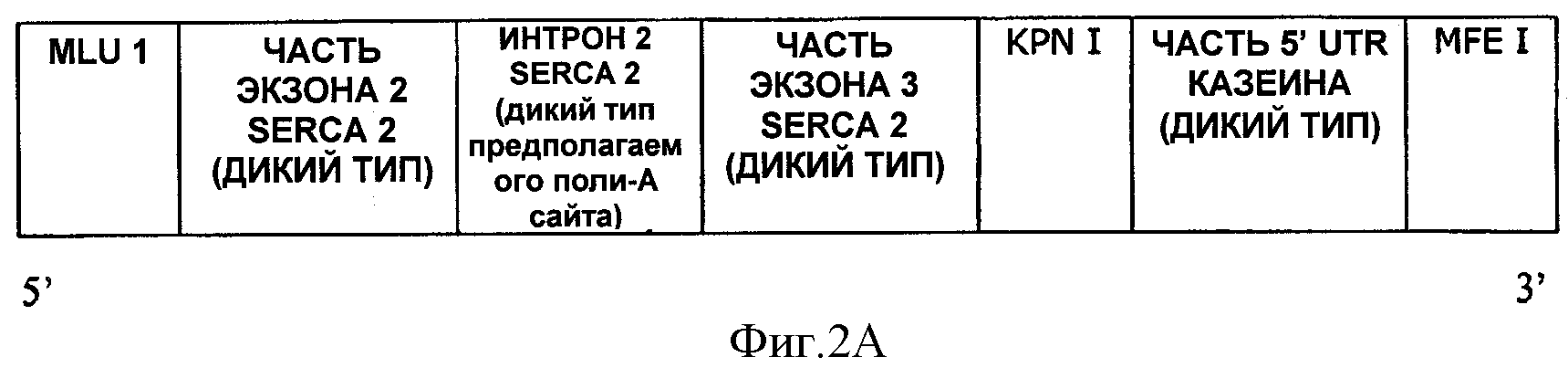
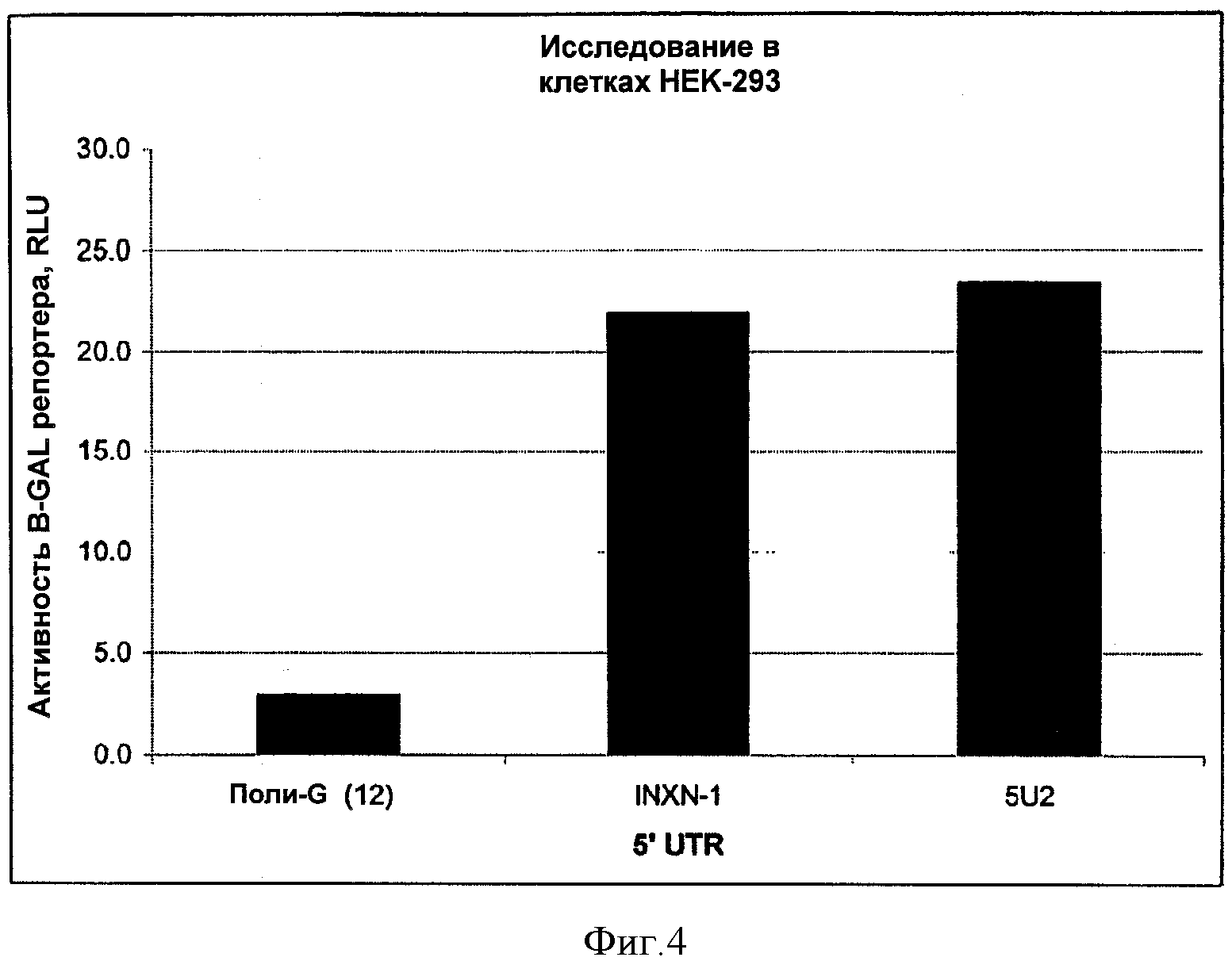
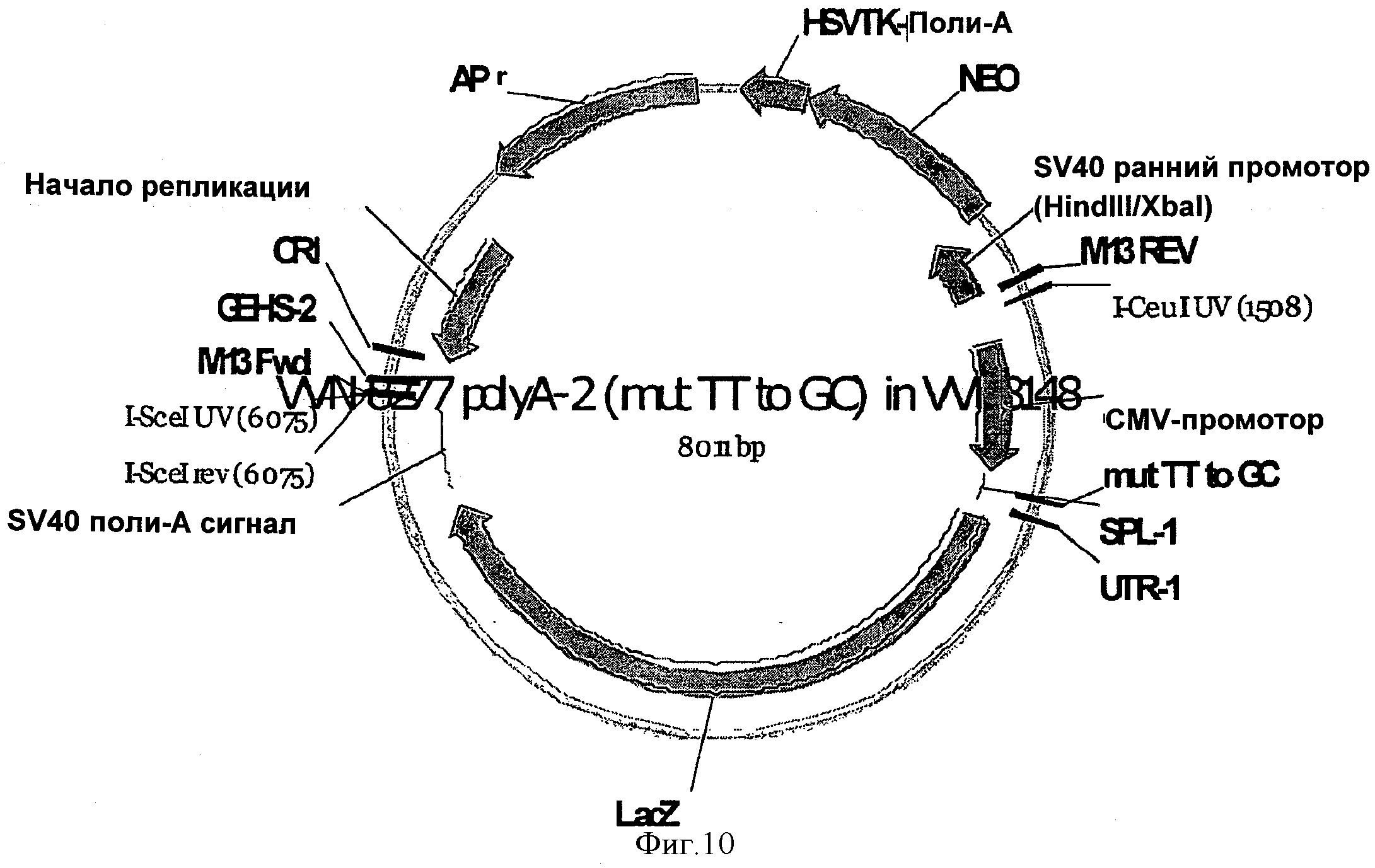
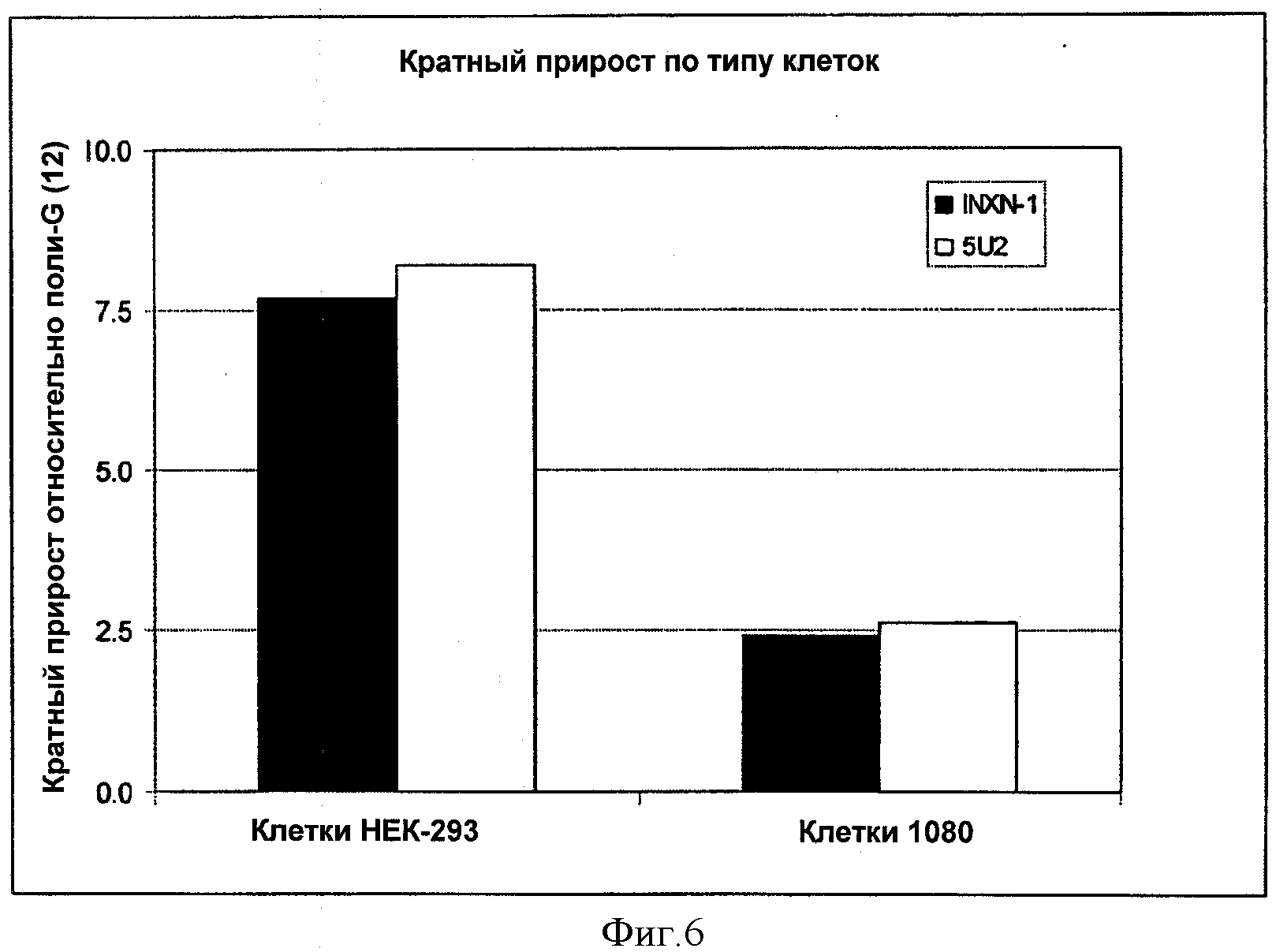


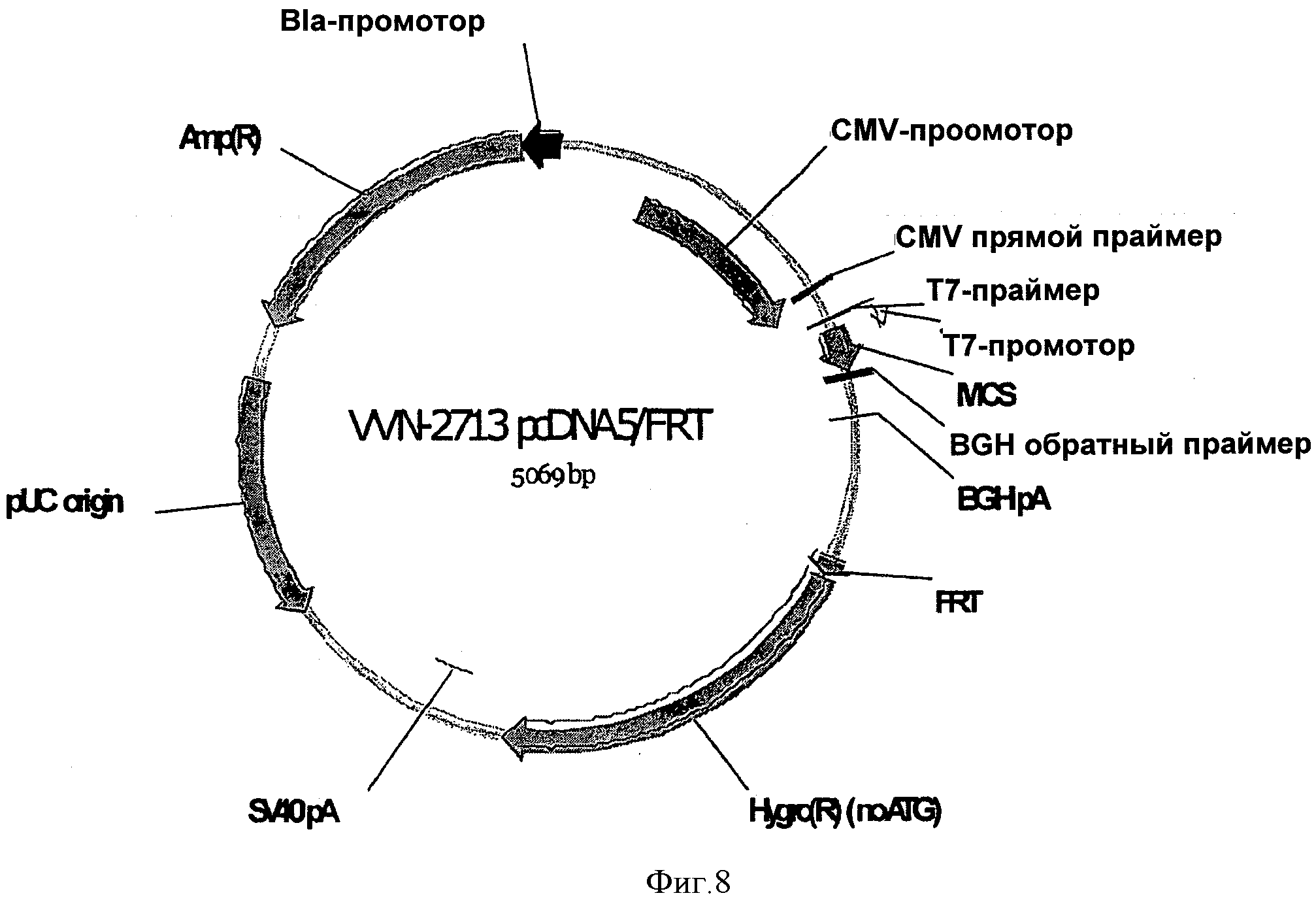


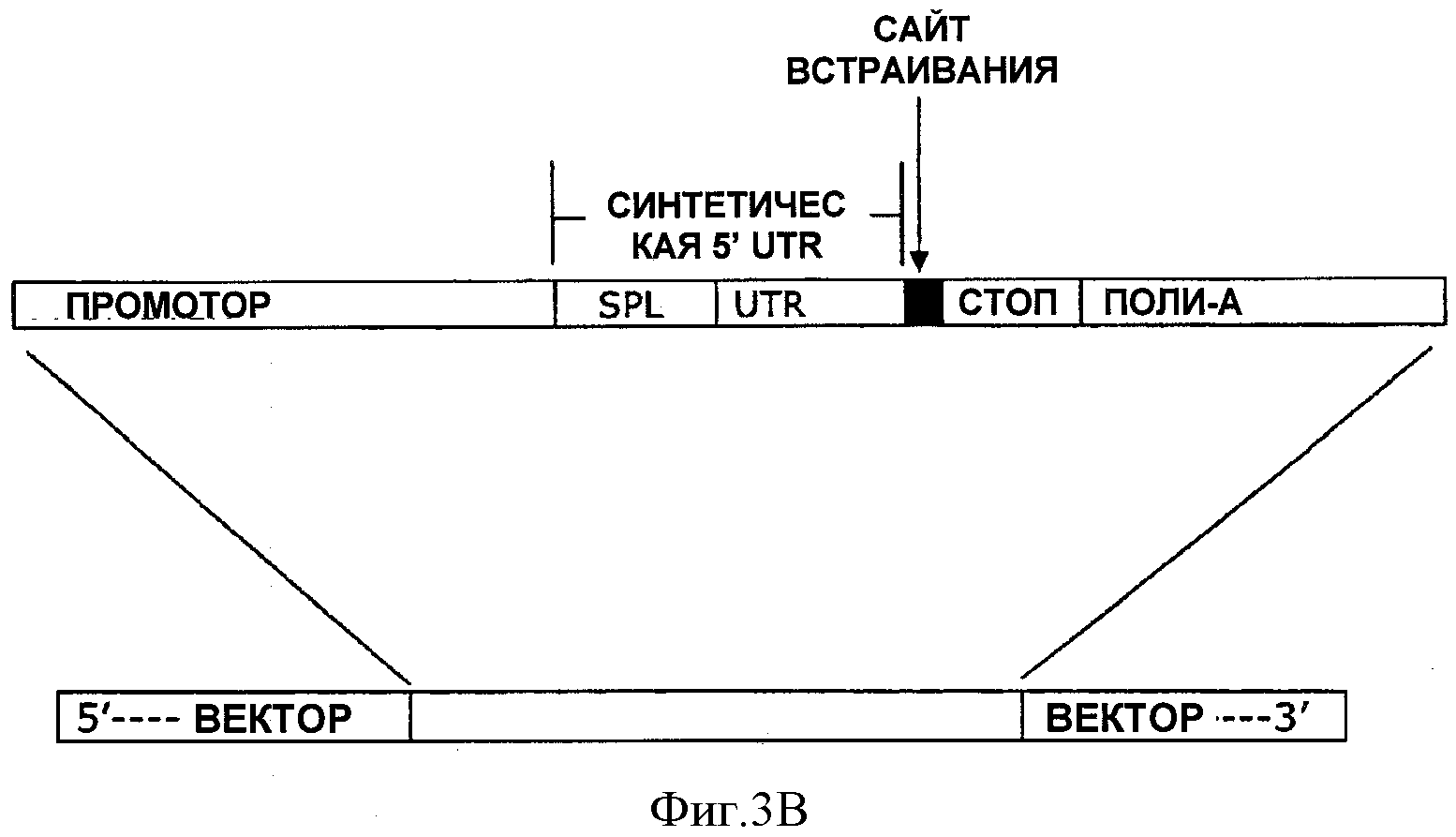

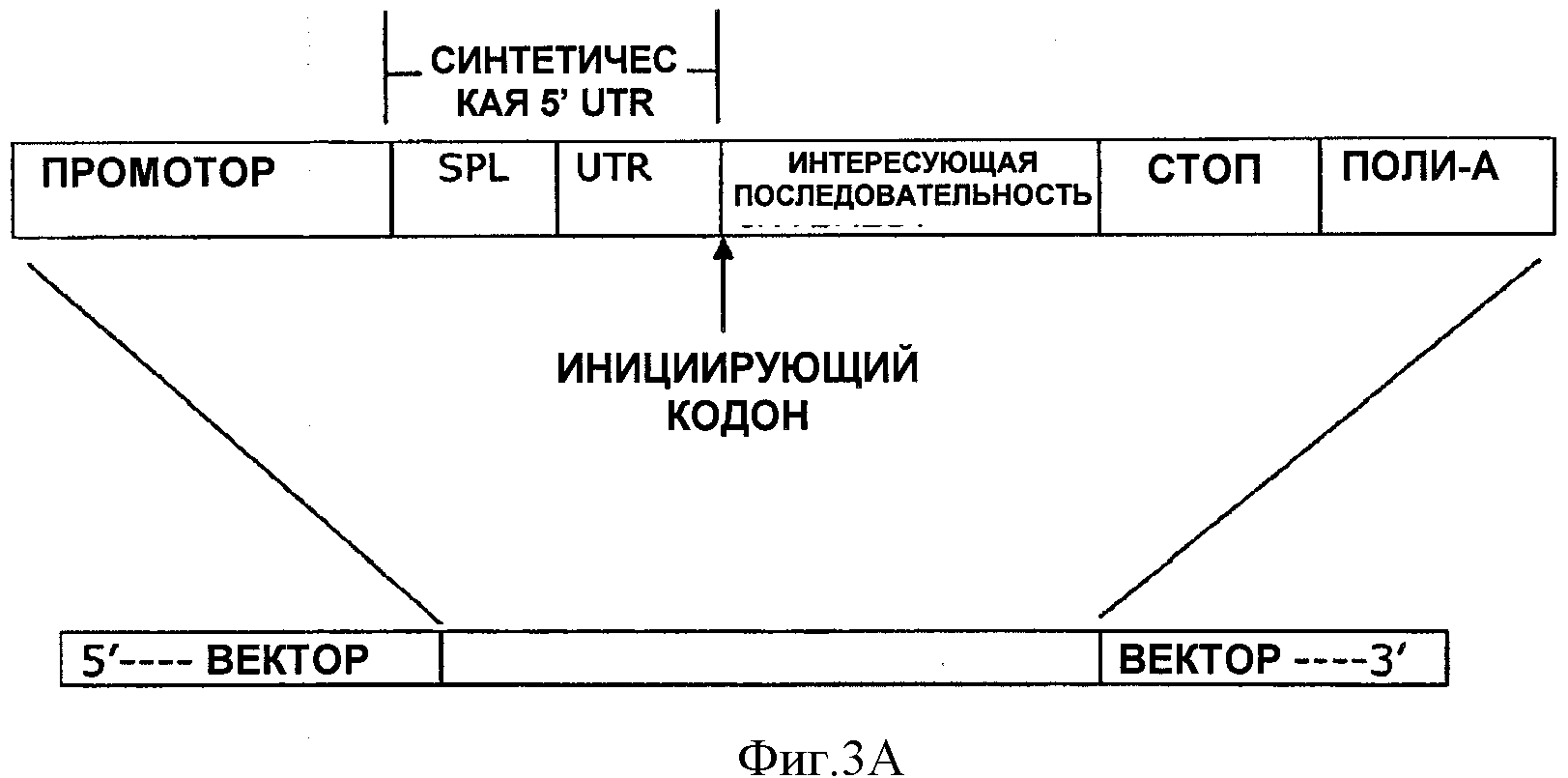



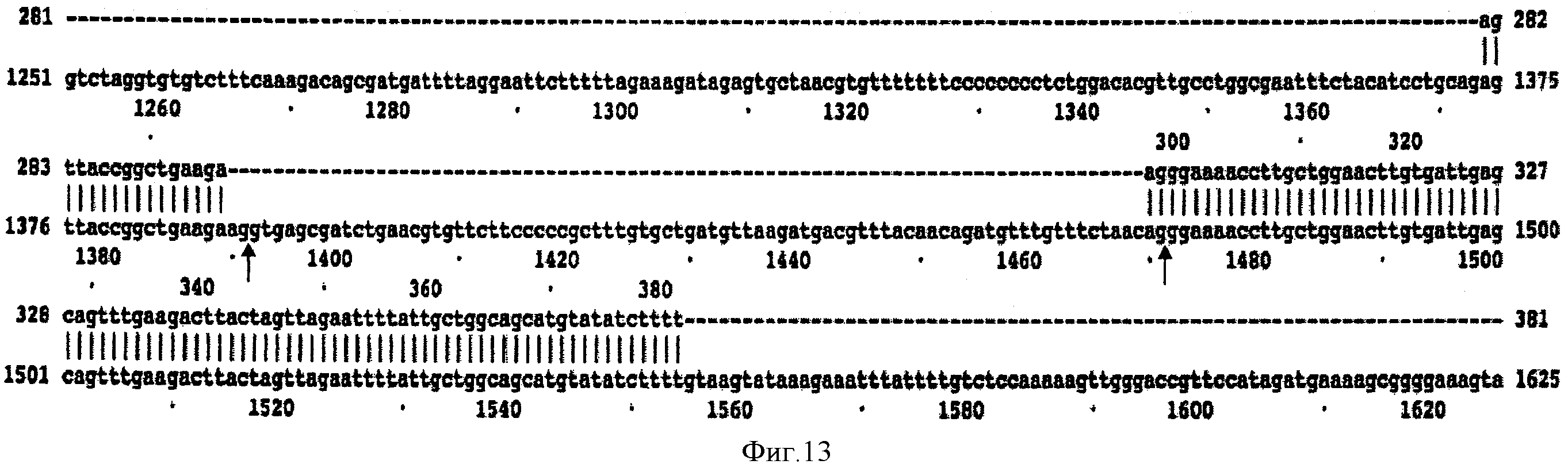
Модифицированные дендритные клетки и их применение в лечении злокачественных опухолей
Стероидные лиганды и их применение для модуляции переключения генов
Векторные плазмиды для клонирования модулей днк и способы их применения
Конструкции терапевтического переключения генов и биореакторы для экспрессии биотерапевтических молекул и их применение
Мутантные рецепторы и их использование в системе экспрессии индуцибельного гена на основе ядерного рецептора
Сконструированные клетки, экспрессирующие множественные иммуномодуляторы, и их применения
Векторы, условно экспрессирующие терапевтические белки, клетки-хозяева, содержащие указанные векторы, и их применение
Кристаллический диацилгидразин и его применение
Борсодержащие диацилгидразины
Модифицированные дендритные клетки и их применение в лечении злокачественных опухолей
Модифицированные дендритные клетки и их применение в лечении злокачественных опухолей
Стероидные лиганды и их применение для модуляции переключения генов
Векторные плазмиды для клонирования модулей днк и способы их применения
Конструкции терапевтического переключения генов и биореакторы для экспрессии биотерапевтических молекул и их применение
Мутантные рецепторы и их использование в системе экспрессии индуцибельного гена на основе ядерного рецептора
Сконструированные клетки, экспрессирующие множественные иммуномодуляторы, и их применения
Векторы, условно экспрессирующие терапевтические белки, клетки-хозяева, содержащие указанные векторы, и их применение
Кристаллический диацилгидразин и его применение
Борсодержащие диацилгидразины
Модифицированные дендритные клетки и их применение в лечении злокачественных опухолей

