Результат интеллектуальной деятельности: ПРИМЕНЕНИЕ РЕКОМБИНАНТНОЙ БЕТА-ГАЛАКТОЗИДАЗЫ AsBgl 1390 ИЗ АРХЕИ SACCHAROVORANS В КАЧЕСТВЕ БЕТА-ГЛЮКОЗИДАЗЫ, БЕТА-КСИЛОЗИДАЗЫ И БЕТА-МАННОЗИДАЗЫ
Вид РИД
Изобретение
Область применения
Изобретение относится к области биотехнологии и касается термостабильных ферментов с активностью гликозил-гидролаз. Предлагается использовать бета-галактозидазу AsBgl_1390 из термофильной археи Acidilobus saccharovorans по новому назначению, а именно в качестве бета-глюкозидазы, бета-ксилозидазы или бета-маннозидазы.
Актуальность
Ферменты - биокатализаторы широко используются в различных отраслях промышленности, сельского хозяйства и медицины. В настоящее время гликозил-гидролазы имеют широкое применение в пищевой промышленности (производство сыров, получение этанола, получение глюкозы, галактозы, мальтозы и других моносахаридов, в технологиях дозревания и хранения фруктов, и др.), биоремедиации, в биосенсорных технологиях для медицины и пищевой промышленности (диагностика лактозы в молоке, глюкозы в крови и т.п.), в диагностике и лечении ряда заболеваний.
Гликозил-гидролазы, гидролизующие различные полисахариды (крахмал, целлюлоза, гемицеллюлозы, хитин и т.д.) до моносахаридов, представляют большой интерес для промышленного гидролиза целлюлозы, являющейся одним из наиболее распространенных в природе сахаров. Деградация целлюлозы до глюкозы достигается совместным действием гидролаз трех классов: эндоглюканаз, экзоглюканаз и бета-глюкозидаз. В качестве продукта гидролиза целлюлозы можно получить глюкозу, а затем этиловый спирт и бутанол, используемые как биотоплива.
Бета-глюкозидазы (EC 3.2.1.21) - ферменты класса гидролаз (подкласс гликозил-гидролаз), катализирующие гидролиз бета-глюкозидной связи в природных и синтетических β-глюкозидах и олигосахаридах. В природе эти ферменты участвуют в реакции гидролиза полисахаридов, обеспечивая клетку моносахаридами для биосинтетических целей и получения энергии. Бета-глюкозидазы осуществляют заключительную стадию деградации целлюлозы, расщепляя целлобиозу с образованием глюкозы. Помимо целлюлозы, одним из основных компонентов древесины, а именно ее гемицеллюлозной фракции, являются полисахариды ксилан и маннан. В силу сложной структурной организации ксилана его гидролиз требует совместного действия эндоксиланаз, бета-ксилозидаз и альфа-арабинозидаз. Гидролиз маннанов также требует активности различных гидролитических ферментов, в том числе эндоманнаназы, бета-маннозидазы, бета-глюкозидазы и альфа-галактозидазы. Термостабильные гликозил-гидролазы этих классов имеют большой потенциал применения для переработки растительного сырья в сахара при высоких температурах.
В настоящее время в этих областях биотехнологии используют гликозил-гидролазы, выделенные из мезофильных микроорганизмов. Основные проблемы используемых в настоящее время гликозил-гидролаз - это низкая активность и нестабильность при высоких температурах реакции. Один из способов решения этой проблемы - это использование ферментов из термофильных микроорганизмов.
Поэтому актуальной задачей, стоящей перед крупными исследовательскими центрами и ведущими биотехнологическими компаниями, является поиск гликозил-гидролаз, обладающих высокой термостабильностью в сочетании с высокой удельной активностью и широкой субстратной специфичностью.
Уровень техники
В течение последних 10 лет активно велись работы по выделению и характеристике гликозил-гидролаз из различных микроорганизмов, в первую очередь грибов Aspergillus и Penicillium (Nagy et al. 2001; Haider and Husain 2007), дрожжей Kluyveromyces (Kim et al. 2004b), бактерий Bacillus и Bifidobacterium (Phan Tran et al. 1998; Moller et al. 2001), архей Sulfolobus and Pyrococcus (Petzelbauer et al. 1999). Термостабильные бета-глюкозидазы из Sulfolobus solfataricus (Pisani et al. 1990), Pyrococcus woesei (Xiong et al. 2007), Thermotoga maritima (Kim et al. 2004a) и Thermus sp. T2 (Ladero et al. 2003) перспективны для практического использования благодаря высоким скоростям реакции, низкому риску загрязнения посторонней микрофлорой, высокой термостабильностью, ограниченным ингибированием продуктами реакции и высокому выходу при их получении (Petzelbauer et al. 1999; Haki and Rakshit 2003; Ji et al. 2005).
Например, в работе (Park et al., 2010) описана рекомбинантная гликозил-гидролаза из гипертермофильной археи Sulfolobus acidocaldarius. Максимальная активность наблюдалась при pH 5.5 и температуре 90°C. Времена полуинактивации фермента при температурах 70, 80 и 90°C составляли соответственно 494, 60 и 0.2 часа. Фермент проявлял гидролитическую активность в отношении следующих производных п-нитрофенила, в порядке уменьшения активности: p-nitrophenyl-β-D-fucopyranoside, pNP-β-D-glucopyranoside, pNP-β-D-galactopyranoside, pNP-β-D-mannopyranoside, pNP-β-D-xylopyranoside, но не в отношении арил-β-глюкозидов и pNP-β-L-arabinofuranoside. Фермент осуществлял гидролиз целлобиозы и, с меньшей эффективностью, лактозы. Авторы делают заключение о том, что новый фермент может быть использован как для гидролиза целлобиозы, так и для синтеза галактоолигосахаридов.
В работе (Kim et al. 2009) представлена новая гликозил-гидролаза из термоацидофильной археи Thermoplasma acidophilum. Максимум активности фермента наблюдался при pH 6.0 и температуре 90°C. Особенностью фермента является его нестабильность при низких pH: при pH 6.0 время его полуинактивации при 75°C составляло 28 дней, а при pH 4.5 - только 13 часов. Фермент проявлял гидролитическую активность в отношении следующих производных п-нитрофенила, в порядке уменьшения активности: pNP-β-D-fucopyranoside, pNP-β-Dglucopyranoside, pNP-β-D-galactopyranoside и pNP-β-D-mannopyranoside. Фермент может быть использован для гидролиза целло-олигосахаридов до глюкозы в процессах осахаривания целлюлозосодержащего сырья.
Источником новых ферментов, перспективных для решения биотехнологических задач, могут являться гипертермофильные микроорганизмы, в частности архея Acidilobus saccharovorans (Prokofeva et al., 2009), выделенная из термального источника кальдеры вулкана Узон на Камчатке. Анализ определенной нами полной нуклеотидной последовательности генома археи Acidilobus saccharovorans 345-15T (Mardanov et al., 2010) выявил наличие гена ASAC_1390, предположительно кодирующего бета-галактозидазу, состоящую из 490 аминокислот, с расчетной молекулярной массой 55.5 кДа. Аминокислотная последовательность этого белка помещена в GenBank под номером ADL19795.1.
Настоящее изобретение касается получения и функциональной характеристики этого фермента, в результате которой было обнаружено, что он является многофункциональной гликозил-гидролазой и может быть использован по новому назначению, а именно в качестве бета-глюкозидазы, бета-ксилозидазы и бета-маннозидазы.
Раскрытие изобретения
Методом культивирования рекомбинантного штамма Е.coli DLT1270(pRARE2), трансформированного вектором pQE60, содержащим нуклеотидную последовательность гена ASAC_1390 (стартовый кодон TTG природного гена ASAC_1390 заменен на стандартный стартовый кодон ATG для экспрессии в Е.coli), был выделен и очищен рекомбинантный фермент AsBgl_1390.
Функциональная характеристика этого фермента показала, что он обладает активностями сразу нескольких видов гликозил-гидролаз и проявляет гидролитическую активность в отношении широкого спектра хроменгенных субстратов: 2-Nitrophenyl-β-D-galactopyranoside (ONPG), 4-Nitrophenyl β-D-galactopyranoside (4-NPGal), 4-Nitrophenyl β-D-glucopyranoside (4NPGluc), 4-Nitrophenyl β-D-xylopyranoside (4-NPXyl), 4-Nitrophenyl β-D-mannopyranoside (4-NPMan), т.е. обладает активностями бета-галактозидазы (EC 3.2.1.23), бета-глюкозидазы (EC 3.2.1.21), бета-ксилозидазы (EC 3.2.1.37) и бета-маннозидазы (EC 3.2.1.25).
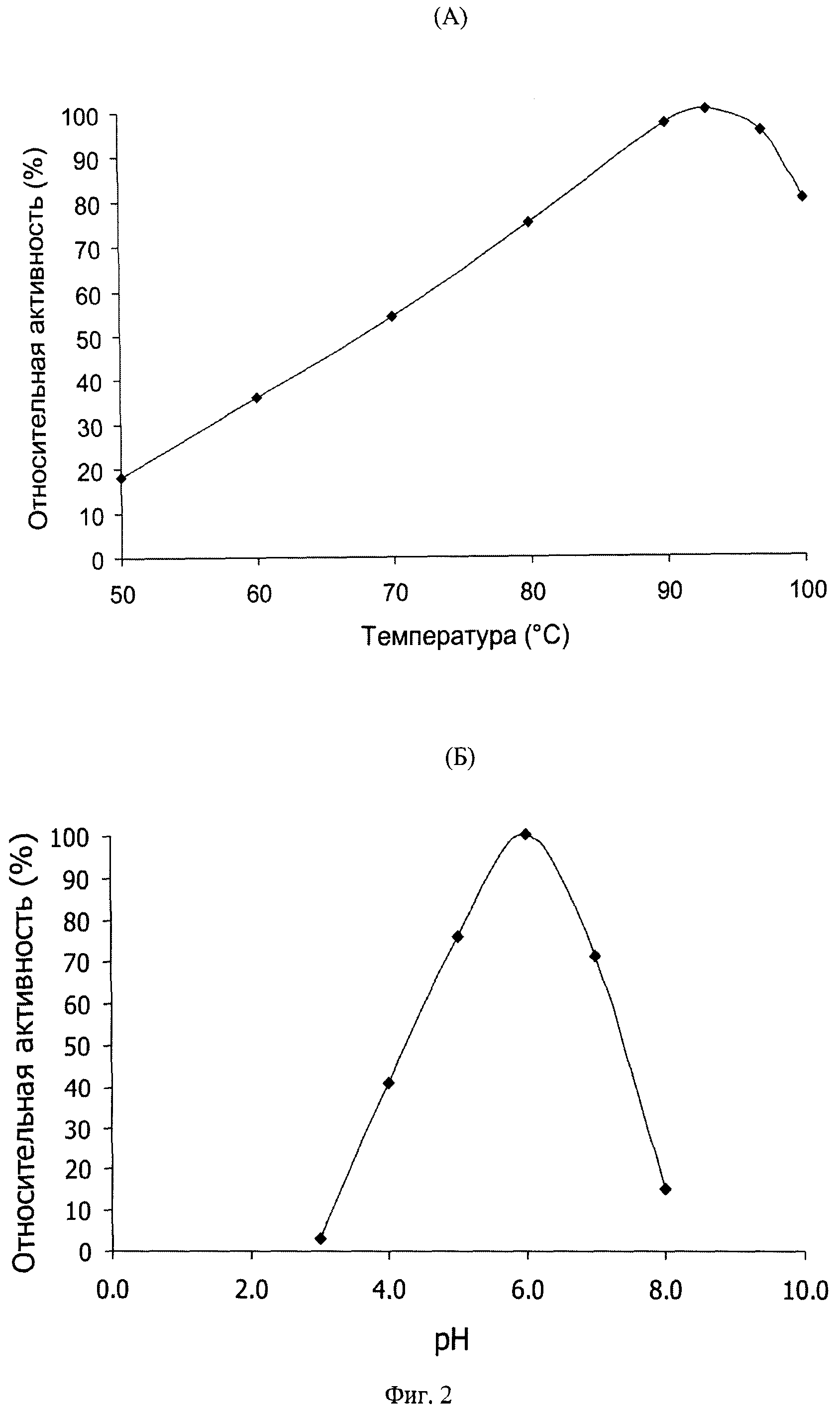
AsBgl_1390 проявляет максимальную активность при pH 6.0 и температуре 93°C. Удельная активность фермента в отношении ONPG в оптимальных условиях составляет 550 ед./мг (1 ед. активности определяется как количество фермента, освобождающее 1 микромоль нитрофенола в минуту). Более 70% максимальной активности сохраняется в диапазоне pH от 5.0 до 7.0, при pH 4.0 фермент проявляет более 40% активности. Не менее 80% максимальной активности сохраняется при температуре 100°C и около 75% - при 80°C. AsBgl_1390 обладает исключительно высокой термостабильностью, сохраняя более 70% активности после инкубации при температуре 90°C в течение 4 часов.
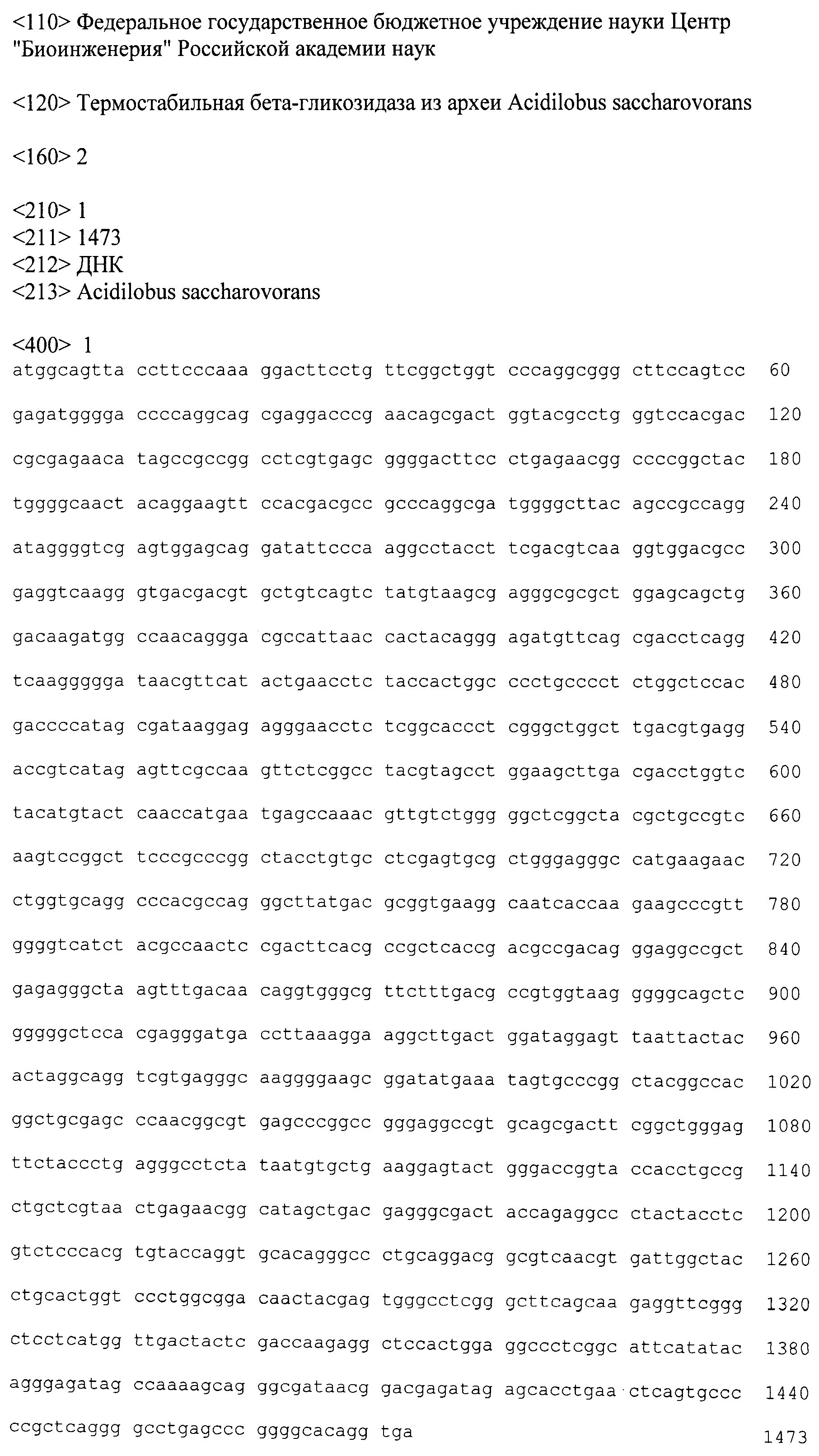
Представленная в настоящем изобретении гликозил-гидролаза содержит аминокислотную последовательность, представленную в GenBank под номером ADL19795.1, или ее фрагмент. Этот фермент может кодироваться последовательностью ДНК, представленной в GenBank под номером CP001742 (участок с 1372868-го по 1374340-й нуклеотиды), или фрагментом этой последовательности, или вырожденной последовательностью, полученной из нее. Термин "вырожденная" подразумевает последовательность ДНК, обладающую по меньшей мере 50% гомологией с представленной в GenBank под номером CP001742 (участок с 1372868-го по 1374340-й нуклеотиды), предпочтительно гомологией на уровне от 70 до 100% и наиболее предпочтительно гомологией на уровне от 90 до 100%. Такая последовательность ДНК может содержать нуклеотидные замены, вставки и удаления, результатом которых является изменение менее 35%, предпочтительно менее 25% и наиболее предпочтительно менее 10% аминокислотной последовательности, приведенной в GenBank под номером ADL19795.1. Следствием нуклеотидных замен могут быть консервативные аминокислотные замены.
Таким образом, AsBgl_1390 является ферментом, обладающим высокой удельной активностью при высоких температурах, высокой термостабильностью и при этом, помимо использования в качестве бета-галактозидазы, может быть использован по новому назначению, а именно в качестве бета-глюкозидазы, бета-ксилозидазы и бета-маннозидазы.
Краткое описание чертежей
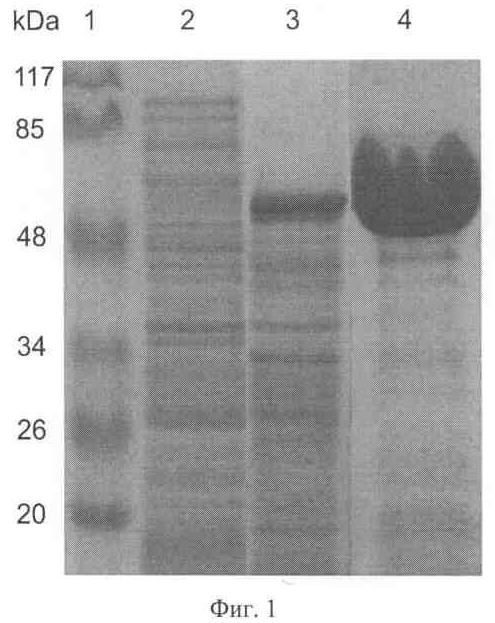
Фиг.1 - экспрессия AsBgl_1390 в Е.coli и очистка белка.
Приведены результаты электрофоретического анализа белковых препаратов:
1 - маркер молекулярного веса;
2 - суммарный белковый препарат, выделенный из клеток штамма DLT1270(pRARE2), содержащего плазмиду pQE60_ASAC_1390 до индукции синтеза белка;
3 - суммарный белковый препарат, выделенный из клеток штамма DLT1270(pRARE2), содержащего плазмиду pQE60_ASAC_1390, через 19 часов после индукции синтеза AsBgl_1390 внесением в среду 1 мМ изопропил-бета-D-тиогалактозида (ИПТГ);
4 - очищенный препарат AsBgl_1390.
Фиг.2 - зависимость активности AsBgl_1390 от температуры (A) и pH (Б).
В качестве субстрата использовали 2-Nitrophenyl-β-D-galactopyranoside.
Реакцию проводили в течение 5 минут в буфере с pH 7.0 при указанной температуре (A) или в течение 5 минут в буфере с указанным рН при температуре 80°C (Б).
Фиг.3 - термостабильность AsBgl_1390. Фермент инкубировали в реакционном буфере с pH 7.0 при 90°C в течение указанного времени, затем проводили реакцию гидролиза с 2-Nitrophenyl-β-D-galactopyranoside при 50°C в течение 20 мин.
Осуществление изобретения
Пример 1. Анализ аминокислотной последовательности гена ASAC_1390 термофильной археи Acidilobus saccharovorans
Термофильная архея Acidilobus saccharovorans (Prokofeva et al., 2009) была выделена из кислого горячего источника кальдеры вулкана Узон (Камчатка, Россия). Нами определена полная нуклеотидная последовательность генома этого микроорганизма, проведен анализ генома и идентифицированы белок-кодирующие гены (Mardanov et al., 2010). Сравнение аминокислотных последовательностей предсказанных белков A.saccharovorans с представленными в GenBank аминокислотными последовательностями выявило наличие в геноме A.saccharovorans гена ASAC_1390, предположительно кодирующего бета-галактозидазу. Аминокислотная последовательность продукта этого гена, представленная в GenBank под номером ADL19795.1, имеет 70% идентичности с последовательностью функционально охарактеризованной бета-галактозидазы из археи Sulfolobus solfataricus (Pisani et al., 1990), 54% - с последовательностью функционально-охарактеризованной бета-глюкозидазы из археи Pyrococcus furiosus (Bruins et al., 2009), 45% - с последовательностью бета-маннозидазы из Pyrococcus furiosus (Park et al., 2011), также функционально охарактеризованной.
Анализ аминокислотной последовательности ASAC_1390 показал, что фермент относится к семейству GH1 гликозил-гидролаз согласно классификации, принятой в базе данных ферментов, активных в отношении углеводов (carbohydrate-active enzymes, CAZy) (Cantarel et al., 2009). Отсутствие в белке N-концевой сигнальной последовательности, которая могла бы обеспечивать его секрецию, указывает на внутриклеточную локализацию в A.saccharovorans.
Пример 2. Получение штамма Е.coli - продуцента рекомбинантной гликозил-гидролазы, выделение и очистка фермента
Праймеры ASAC_1390-NcoI (5′-TACCATGGCAGTTACCTTCCCAAAGGAC-3′) и ASAC_1390-BglII (5′-ATGAGATCTACCAGGCGCTGGGCTGCT-3′) были использованы для ПЦР-амплификации полноразмерного гена ASAC_1390, в качестве матрицы использовали геномную ДНК A.saccharovorans. Полученный фрагмент обрабатывали рестриктазами NcoI и BglII и клонировали по сайтам NcoI и BamHI в экспрессионном векторе pQE60. В результате был получен экспрессионный вектор pQE60_ASAC1390, обеспечивающий продукцию рекомбинантного фермента AsBgl_1390.
Для продукции рекомбинантной гликозил-гидролазы AsBgl_1390 экспрессионным вектором pQE60_ASAC1390 трансформировали штамм Е.coli DLT1270(pRARE2). Указанный штамм, содержащий pQE60_ASAC1390, выращивали в среде LB с добавлением ампициллина (100 мг/л) и хлорамфеникола (20 мг/л) на шейкере при 37°C до середины логарифмической фазы роста (OD600 ~0.5), затем индуцировали синтез рекомбинантного белка, внося изопропил-β-D-тиогалактопиранозид (ИПТГ) до 1 мМ, и продолжали выращивать культуру в течение 19 ч при 37°C.
Клетки штамма-продуцента собирали центрифугированием, выделение и очистку рекомбинантной гликозил-гидролазы AsBgl_1390 осуществляли в соответствии со следующим протоколом.
1. Сбор биомассы штамма - продуцента
По окончании индукции клетки штамма-продуцента (~1000 мл) осаждали центрифугированием в течение 15 мин при 4000 g с охлаждением до +4°C.
2. Разрушение клеток штамма-продуцента
Собранный осадок биомассы клеток ресуспендировать в 100 мМ натрий-фосфатного буфера, pH 7.0, 1 мг/мл лизоцима. Буфер добавляли из расчета 10 мл на 1 г сырой биомассы (стандартно - 20 мл). Клетки разрушали при +4°C на ультразвуковом дезинтеграторе Bandelin Sonopuls HD2200 (4 цикла по 30 сек), клеточный дебрис удаляли центрифугированием (12000 g, 20 мин, +4°C).
3. Удаление белков Е.coli
Для удаления термолабильных белков Е.coli полученный на предыдущей стадии бесклеточный экстракт прогревали на водяной бане в течение 30 мин при температуре 75°C. Осадок (денатурированные белки Е.coli) удаляли центрифугированием (12000 g, 20 мин, +4°C). Надосадок повторно прогревали на водяной бане в течение 30 мин при температуре 85°C. Осадок (денатурированные белки Е.coli) удаляли центрифугированием (12000 g, 20 мин, +4°C). Отбирали супернатант, содержащий очищенный белок. В случае необходимости проводили диализ препарата против необходимого буфера.
Гомогенность препарата AsBgl_1390 и соответствие молекулярного веса рекомбинантного белка расчетному значению 55,5 кДа подтверждены при помощи SDS-PAGE (фиг.1).
Пример 3. Ферментативная активность и субстратная специфичность AsBgl_1390
Для определения активности AsBgl_1390 использовали модифицированный метод Кравена (Craven et al., 1965), применяющийся практически во всех работах, описывающих выделение и функциональную характеристику бета-галактозидаз и родственных им ферментов. В качестве субстрата для проведения реакции используется o-nitrophenyl-beta-D-galactopyranoside (ONPG) или другие производные нитрофенила, гидролитическая активность гликозил-гидролазы приводит к отщеплению нитрофенола, вещества желтого цвета, концентрация которого количественно определяется спектрофотометрически путем измерения поглощения при длине волны 420 нм. Использование вместо ONPG других производных нитрофенила (4-Nitrophenyl-beta-D-galactopyranoside, 4-Nitrophenyl-beta-D-glucopyranoside, 4-Nitrophenyl-beta-D-xylopyranoside, 4-Nitrophenyl-beta-D-mannopyranoside и др.) позволяет проводить характеристику субстратной специфичности фермента.
Определение активности фермента проводили в реакции:
к раствору 50-мкл раствора фермента (количество белка - 0.08 мкг, в 0.1М натрий-фосфатном буфере pH 7.0), прединкубированного при 80°C в течение 5 мин, добавляли 1076 мкл раствора субстрата (2.32 mM ONPG) в 0.1М натрий-фосфатном буфере pH 7.0, прединкубированного при 80°C в течение 5 мин.
Реакцию (в 3-кратной повторности) проводили при температуре 80°C до появления видимой желтой окраски. Реакцию останавливали, добавляя 375 мкл холодного раствора 1М карбоната натрия. Результат реакции детектировали спектрофотометрически, определяя поглощение на длине волны 420 нм относительно таким же образом проведенной контрольной реакции (не содержащей фермент).
Активность фермента определяется в ед./мг (одна единица активности фермента определяется как количество фермента, освобождающее 1 микромоль нитрофенола в минуту) по формуле:
A (ед./мг)=[(OD/Е)×V]/(T×M),
где A - активность фермента в ед./мг;
OD - поглощение на длине волны 420 нм для o-нитрофенола или 405 нм - для 4-нитрофенола (за вычетом контроля);
E - коэффициент экстинкции (4.5 мл·мкмоль-1·см-1 для o-нитрофенола, 18.3 мл·мкмоль-1·см-1 для 4-нитрофенола);
V - объем реакционной смеси в мл (1,5 мл);
T - время реакции в мин;
M - количество фермента в реакции в мг (0.08·10-3).
Эксперименты по тестированию бета-галактозидазной активности AsBgl_1390 на ONPG показали, что фермент осуществляет гидролиз этого субстрата, причем активность в оптимальных условиях реакции (93°C, 0.1М натрий-фосфатный буфер с pH 6.0) составляла 550 ед./мг.
Для сравнения отметим, что активность бета-галактозидазы из археи P.woesei составляла для нативного фермента 51,1 ед./мг. Активость бета-галактозидазы из бактерии Caldicellulosiruptor saccharolyticus составляла около 211 ед./мг (на 4-NPGal, см. Park and Oh, 2010), из Alicyclobacillus acidocaldarius - около 230 ед./мг (на ONPG, см. Yuan et al., 2008), из Sulfolobus acidocaldarius - около 60 ед./мг (на ONPG, см. Park et al., 2010). Таким образом, по своей активности AsBgl_1390 превышает характеристики известных функциональных аналогов.
Для характеристики субстратной специфичности фермента его активность измеряли в следующих условиях: 93°C, 0.1 М натрий-фосфатный буфер (pH 7.0), концентрация взятого на реакцию субстрата - 2.32 mM, количество фермента - 0.08 мкг. В качестве субстратов использовали 4-Nitrophenyl-beta-D-galactopyranoside (4NPGal), 4-Nitrophenyl-beta-D-glucopyranoside (4NPGlu), 4-Nitrophenyl-beta-D-xylopyranoside (4-NPXyl), 4-Nitrophenyl-beta-D-mannopyranoside (4-NPMan) и ONPG. Спектрофотометрическое измерение проводили на длине волны 405 нм для всех субстратов, кроме ONPG (для ONPG измерение проводили на длине волны 420 нм). Было установлено (Табл.1), что AsBgl_1390 с разной эффективностью гидролизует все вышеуказанные субстраты. Высокая активность проявлялась в отношении ONPG, 4NPGal и 4NPGlu, в меньшей степени фермент гидролизовал NPXyl и 4-NPMan. Таким образом, AsBgl_1390 является многофункциональной гликозил-гидролазой и обладает активностями бета-глюкозидазы, бета-галактозидазы, бета-кислозидазы и бета-маннозидазы.
|
Пример 4. Влияние температуры и рН на активность рекомбинантной гликозил-гидролазы AsBgl_1390
Для определения зависимости активности AsBgl_1390 от температуры проводили реакцию гидролиза ONPG в 0.1М натрий-фосфатном буфере (pH 7.0) с концентрацией субстрата 2.32 mM при различных температурах в диапазоне от 50 до 100°C. Компоненты реакции прединкубировали при выбранной температуре в течение 5 мин, затем проводили реакцию в фосфатно-натриевом буфере (pH 7.0) в течение 5 мин, используя 0.08 мкг фермента на реакцию. Контрольные пробы инкубировали в тех же условиях без фермента. Полученные результаты (фиг.2(A)) показывают, что фермент проявляет максимальную активность при температуре 93°C, не менее 80% максимальной активности сохраняется при температуре 100°C и около 75% - при температуре 80°C. Таким образом, оптимальная для проявления активности фермента температура является одной из самых высоких среди охарактеризованных гликозил-гидролаз (более высокий температурный оптимум лишь у эуриархеи Pyrococcus furiosus DSM3638 - 100°C; Park et al., 2011).
Для характеристики зависимости активности ASAC_1390 от pH среды реакцию проводили в стандартных условиях при температуре 80°C, меняя pH буфера в диапазоне от 3 до 8. В диапазоне рН 3-5 использовали 0.1М ацетатный буфер, в диапазоне pH 6-8 - 0.1М фосфатный. Полученные результаты (фиг.2(Б)) показывают, что фермент активен в диапазоне значений pH от 4.0 до 7.0, дальнейшее понижения pH ниже 4.0 или повышение выше 7.0 вызывает быстрое снижение активности фермента. Такая зависимость активности от pH позволит использовать AsBgl_1390 в технологических процессах гидролиза при нейтральных и кислых значениях pH.
Для характеристики термостабильности AsBgl_1390 препарат фермента инкубировали в реакционном буфере при 90°C различное время, затем охлаждали до 50°C, вносили ONPG и проводили реакцию при 50°C в стандартном реакционном буфере в течение 20 мин. Как видно из полученных результатов (фиг.3), период полуинактивации фермента более 4 ч. После 4 ч инкубации при 90°C сохраняется около 70% максимальной активности фермента.
Для сравнения отметим, что период полуинактивации бета-галактозидазы из термофильной бактерии Thermotoga maritima составляет 16 мин при 90°C (Kim et al., 2004a), из термофильной бактерии Caldicellulosiruptor saccharolyticus - 2 ч при 80°C (Park and Oh, 2010), из кренархеии Sulfolobus solfataricus - 30 мин при 90°C (Pisani et al., 1990), из кренархеи Sulfolobus acidocaldarius - 12 мин при 90°C (Park et al., 2010), из эуриархеи Pyrococcus furiosus DSM3638 - 6.63 ч при 90° (Park et al., 2011). Активность бета-галактозидазы из термофильной бактерии Alicyclobacillus acidocaldarius при 90°C падает до нуля меньше чем за 5 мин (Yuan et al., 2008). Период полуинактивации бета-галактозидазы из термофильной бактерии Thermus sp. A4 при 90°C составляет 1 ч (Ohtsu, et al., 1998).
Пример 5. Влияние глюкозы и галактозы на активность рекомбинантной гликозил-гидролазы AsBgl_1390
Одним из перспективных направлений использования ферментов, обладающих бета-галактозидазной активностью, является гидролиз лактозы до глюкозы и галактозы в молочных продуктах. Известно, что эти моносахариды могут оказывать ингибирующее влияние на активность бета-галактозидаз (Kim et al. 2004a, b; Cowan et al., 1984), поэтому мы протестировали влияние глюкозы и галактозы на активность AsBgl_1390. Для этого реакцию гидролиза ONPG проводили в присутствии глюкозы и галактозы. Полученные результаты (Табл. 2) показывают, что глюкоза и галактоза не оказывают существенного ингибирующего влияния на активность AsBgl_1390.
|
СПИСОК ЛИТЕРАТУРЫ
1. Bruins М.Е., Meersman F., Janssen A.E., Heremans K., Boom R.M. (2009) Increased susceptibility of beta-glucosidase from the hyperthermophile Pyrococcus furiosus to thermal inactivation at higher pressures. FEBS J. 276: 109-117.
2. Cantarel B.L., Coutinho P.M., Rancurel C., Bernard T., Lombard V., Henrissat. B. (2009) The Carbohydrate-Active EnZymes database (CAZy): an expert resource for glycogenomics. Nucleic Acids Res 37: D233-238.
3. Cowan D.A., Daniel R.M., Martin A.M., and Morgan H.W. (1984) Some Properties of a beta-Galactosidase from an Extremely Thermophilic Bacterium. Biotechnology and Bioengineering. Biotechnol Bioeng. 26: 1141-1 145.
4. Craven G.R., Steers E., Anfinsen C.B. (1965) Purification, composition and molecular weight of the beta-galactosidase of Escherichia coli K12. J Biol Chem 240: 2468-2477.
5. Ji E.S., Park N.H., Oh D.K. (2005) Galacto-oligosaccharide production by a thermostable recombinant β-galactosidase from Thermotoga maritima. World J Microbiol Biotechnol 21: 759-764.
6. Haider Т., Husain Q. (2007) Calcium alginate entrapped preparations of Aspergillus oryzae beta galactosidase: its stability and applications in the hydrolysis of lactose. Int J Biol Macromol 41: 72-80.
7. Haki G.D., Rakshit S.K. (2003) Developments in industrially important thermostable enzymes: a review. Bioresour Technol 89: 17-34.
8. Nagy Z., Kiss Т., Szentirmai A., Biro S. (2001) Beta-galactosidase of Penicillium chrysogenum: production, purification, and characterization of the enzyme. Protein Expr Purif 21: 24-29.
9. Kim C.S., Ji E.S., Oh D.K. (2004a) Characterization of a thermostable recombinant β-galactosidase from Thermotoga maritima. J Appl Microbiol 97: 1006-1014.
10. Kim C.S., Ji E.S., Oh D.K. (2004b) A new kinetic model of recombinant β-galactosidase from Kluyveromyces lactis for both hydrolysis and transgalactosylation reactions. Biochem Biophys Res Commun 316: 738-743.
11. Kim H.J., Park A.R., Lee J.K., Oh D.K. (2009) Characterization of an acid-labile, thermostable beta-glycosidase from Thermoplasma acidophilum. Biotechnol Lett 31: 1457-1462.
12. Ladero M., Perez M.T., Santos A., Garcia-Ochoa F. (2003) Hydrolysis of lactose by free and immobilized beta-galactosidase from Thermus sp. strain T2. Biotechnol Bioeng 81: 241-252.
13. Mardanov A.V., Svetlitchnyi V.A., Beletsky A.V., Prokofeva М.I., Bonch-Osmolovskaya E.A., Ravin N.V., Skryabin K.G. (2010) The genome sequence of the crenarchaeon Acidilobus saccharovorans supports a new order, Acidilobales, and suggests an important ecological role in terrestrial acidic hot springs. Appl. Environ. Microbiol., 76, 5652-5657.
14. Moller P.L., Jorgensen F., Hansen O.C., Madsen S.M., Stougaard P. (2001) Intra- and extracellular beta-galactosidases from Bifidobacterium bifidum and B. infantis: molecular cloning, heterologous expression, and comparative characterization. Appl Environ Microbiol 67: 2276-2283.
15. Ohtsu N., Motoshima H., Goto K., Tsukasaki F., Matsuzawa H. (1998) Thermostable beta-galactosidase from an extreme thermophile, Thermus sp. A4: enzyme purification and characterization, and gene cloning and sequencing. Biosci Biotechnol Biochem. 62: 1539-1545.
16. Park A.R., Oh D.K. (2010) Effects of galactose and glucose on the hydrolysis reaction of a thermostable β-galactosidase from Caldicellulosiruptor saccharolyticus. Appl Microbiol Biotechnol 85: 1427-1435.
17. Park A.R., Kim H.J., Lee J.K., Oh D.K. (2010) Hydrolysis and transglycosylation activity of a thermostable recombinant beta-glycosidase from Sulfolobus acidocaldarius. Appl Biochem Biotechnol 160: 2236-2247.
18. Park S.H., Park K.H., Oh B.C., Alli I., Lee B.H. (2011) Expression and characterization of an extremely thermostable β-glycosidase (mannosidase) from the hyperthermophilic archaeon Pyrococcus furiosus DSM3638. N Biotechnol. 28: 639-648.
19. Petzelbauer I., Nidetzky В., Haltrich D., Kulbe K.D. (1999) Development of an ultra-high-temperature process for the enzymatic hydrolysis of lactose. I. The properties of two thermostable β-glycosidases. Biotechnol Bioeng 64: 322-332.
20. Phan Tran L.S., Szabo L., Fulop L., Orosz L., Sik Т., Holczinger A. (1998) Isolation of a beta-galactosidase-encoding gene from Bacillus licheniformis: purification and characterization of the recombinant enzyme expressed in Escherichia coli. Curr Microbiol 37: 39-43.
21. Pisani F.M., Rella R., Raia C.A., Rozzo C., Nucci R., Gambacorta A., De Rosa M., Rossi M. (1990) Thermostable beta-galactosidase from the archaebacterium Sulfolobus solfataricus. Purification and properties. Eur J Biochem 187: 321-328.
22. Prokofeva M.I., Kostrikina N.A., Kolganova T.V., Tourova T.P., Lysenko A.M., Lebedinsky A.V., and Bonch-Osmolovskaya E.A. (2009) Isolation of the anaerobic thermoacidophilic crenarchaeote Acidilobus saccharovorans sp. nov. and proposal of Acidilobales ord. nov., including Acidilobaceae fam. nov. and Caldisphaeraceae fam. nov. Int. J. Syst. Evol. Microbiol. 59: 3116-3122.
23. Xiong A.S., Peng R.H., Zhuang J., Li X., Xue Y., Liu J.G., Gao F., Cai В., Chen J.M., Yao Q.H. (2007) Directed evolution of a beta-galactosidase from Pyrococcus woesei resulting in increased thermostable beta-glucuronidase activity. Appl Microbiol Biotechnol 77: 569-578.
24. Yuan Т., Yang P., Wang Y., Meng K., Luo H., Zhang W., Wu N., Fan Y., Yao B. (2008) Heterologous expression of a gene encoding a thermostable beta-galactosidase from Alicyclobacillus acidocaldarius. Biotechnol Lett 30: 343-348.
Применение рекомбинантной бета-галактозидазы AsBgl_1390 из археи Acidilobus saccharovorans в качестве бета-глюкозидазы, бета-ксилозидазы и бета-маннозидазы при гидролизе соответствующих углеводов этим ферментом при температурах от 50 до 100 градусов цельсия и значениях pH среды от 3 до 8.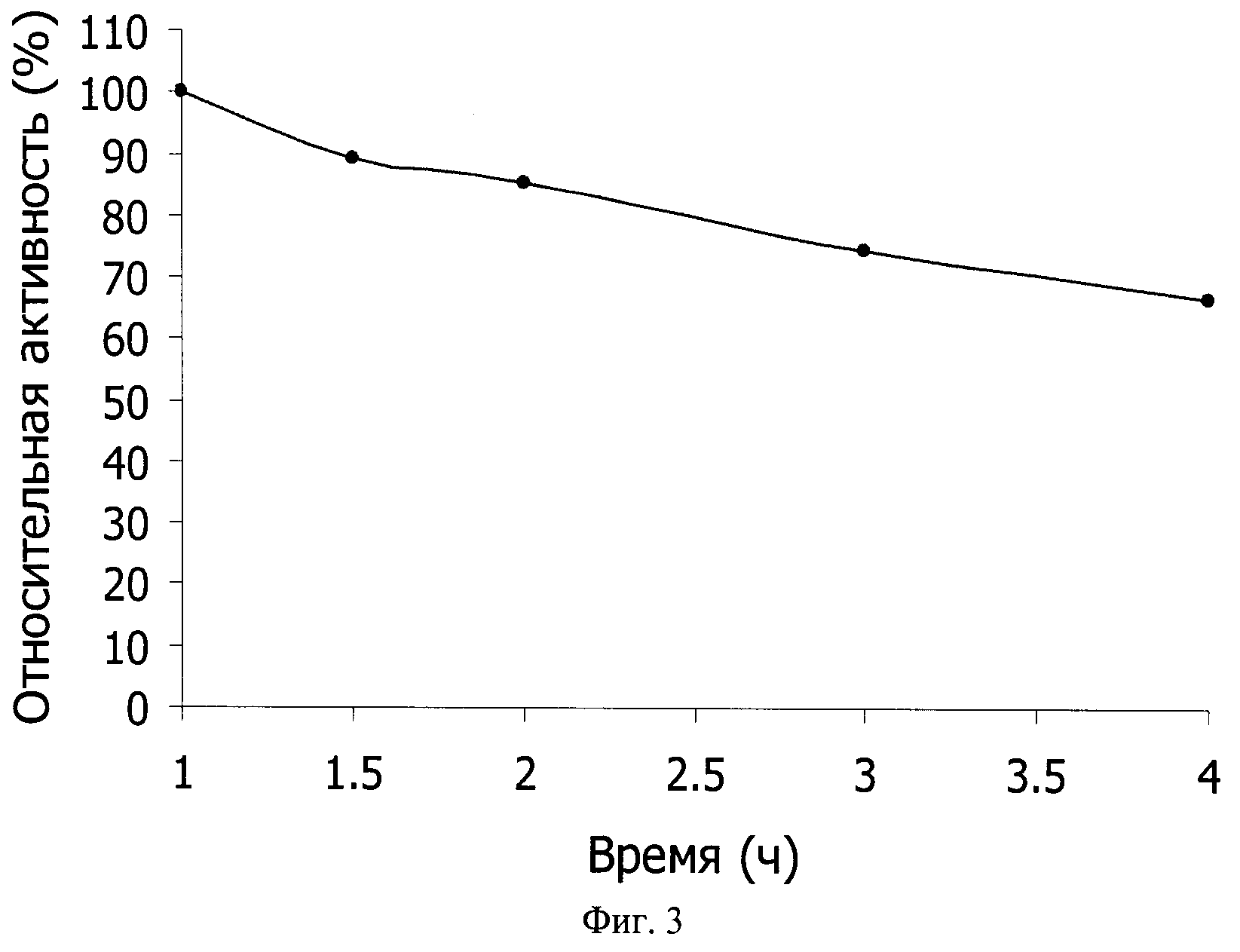
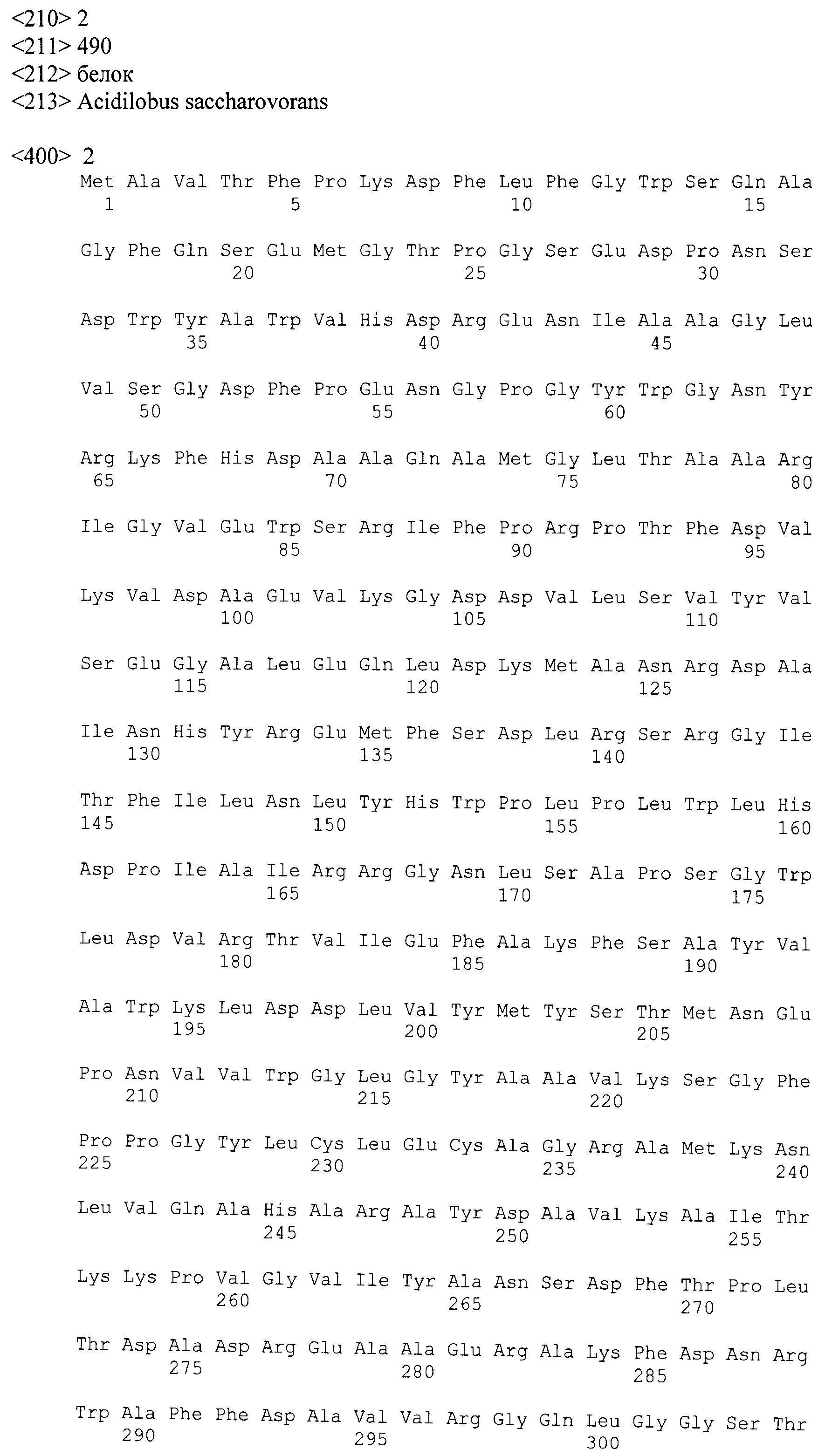
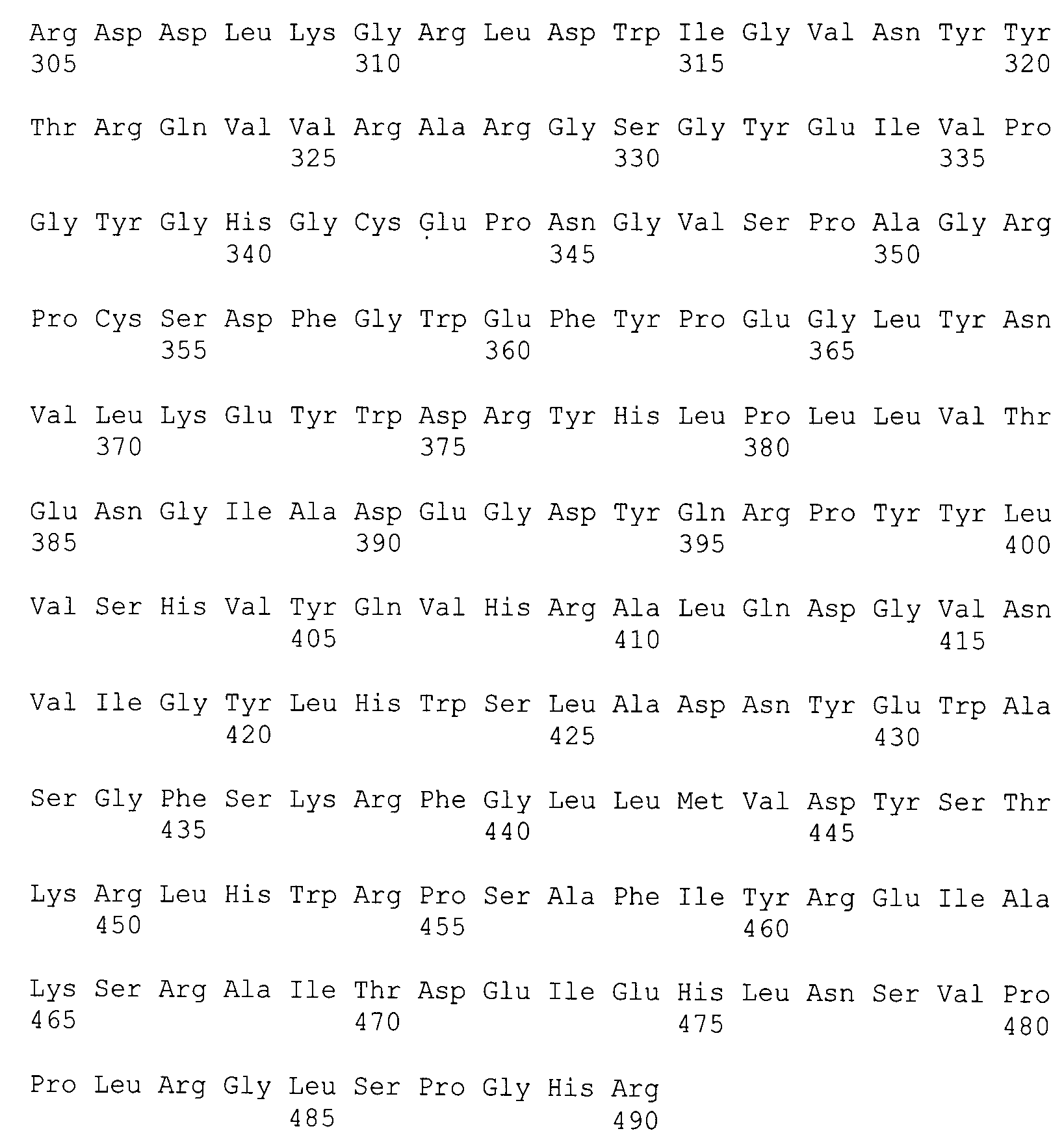
Термостабильная липаза из бактерии thermosyntropha lipolytica, активная в щелочной среде
Усовершенствованный способ очистки правастатина
Биологический днк маркер для идентификации гена устойчивости к х вирусу картофеля
Способ получения полифункциональных магнитных наночастиц на основе магнетосом бактериального происхождения
Высокоспецифичный днк-маркер, используемый в качестве эндогенного референсного контроля для обнаружения геномной днк картофеля в растительном материале и пищевых продуктах, в том числе при идентификации гмо
Ацидофильный штамм desulfosporosinus sp. для очистки загрязненных экосистем с экстремально кислыми значениями от ионов тяжелых металлов
Вектор на основе репликона бактериофага n15 и рекомбинантный вектор для регулируемой экспрессии целевого гена в клетках escherichia coli, штамм escherichia coli, обеспечивающий возможность регуляции числа копий вектора, и система экспрессии
Термостабильная липаза из бактерии thermosyntropha lipolytica, активная в щелочной среде
Усовершенствованный способ очистки правастатина
Биологический днк маркер для идентификации гена устойчивости к х вирусу картофеля
Способ получения полифункциональных магнитных наночастиц на основе магнетосом бактериального происхождения
Высокоспецифичный днк-маркер, используемый в качестве эндогенного референсного контроля для обнаружения геномной днк картофеля в растительном материале и пищевых продуктах, в том числе при идентификации гмо
Ацидофильный штамм desulfosporosinus sp. для очистки загрязненных экосистем с экстремально кислыми значениями от ионов тяжелых металлов
Способ подготовки шахтных вод для выделения днк
Термостабильная алкогольдегидрогеназа из археи thermococcus sibiricus
Вирусный вектор для продукции рекомбинантных белков в растениях
Термостабильная днк-лигаза из археи рода thermococcus, способ ее получения и нуклеотидная последовательность днк, кодирующая эту днк-лигазу

