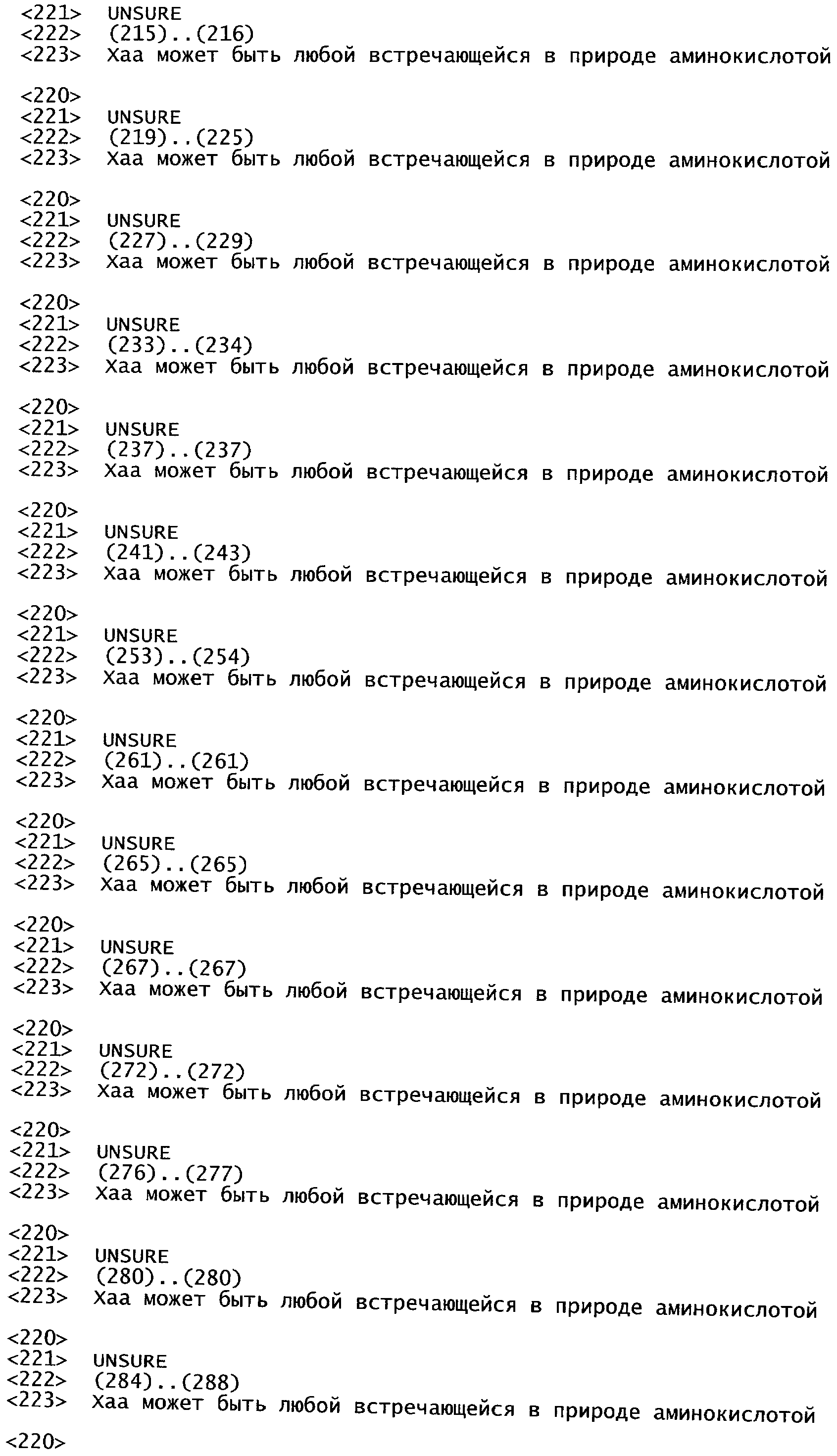
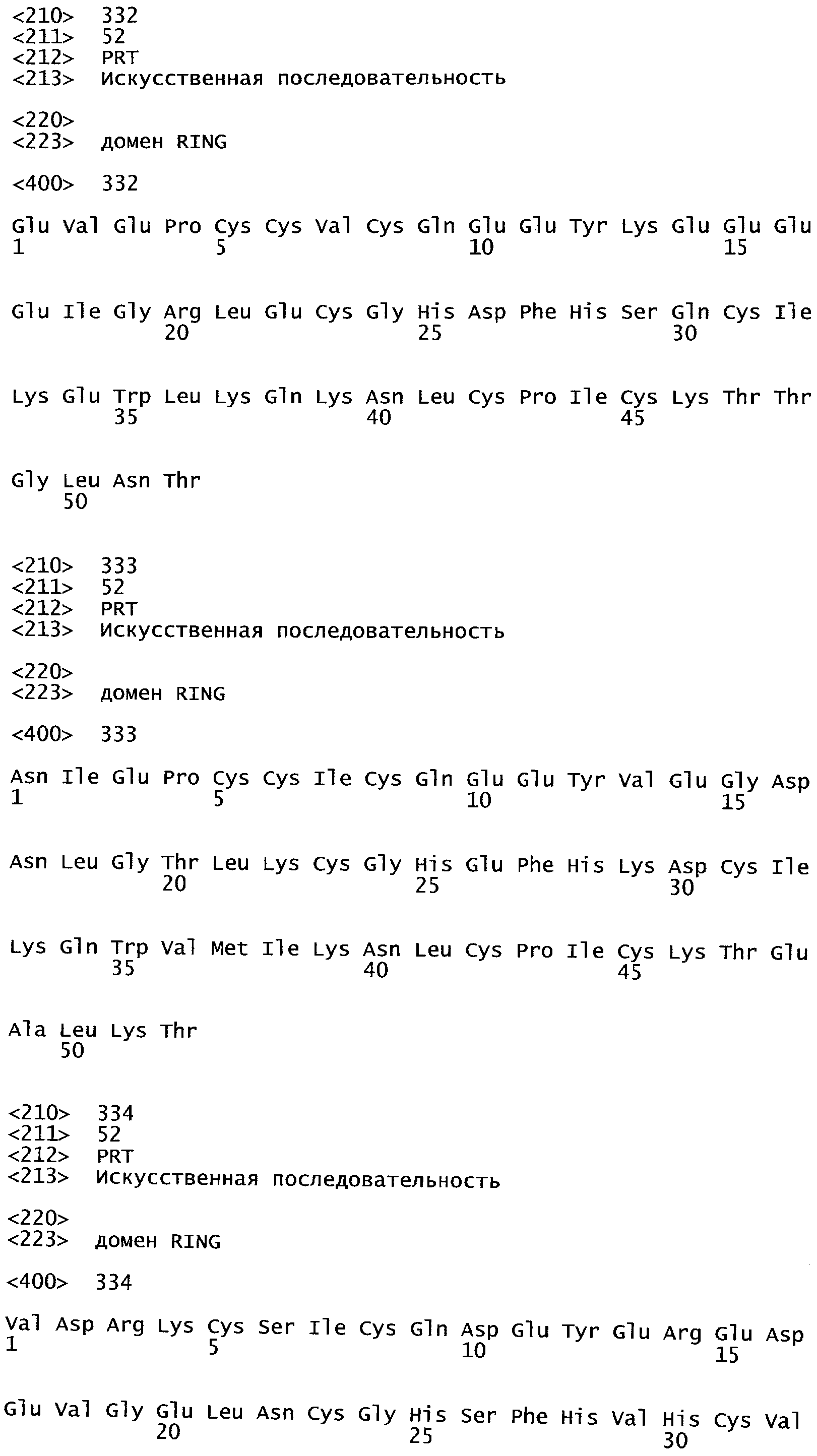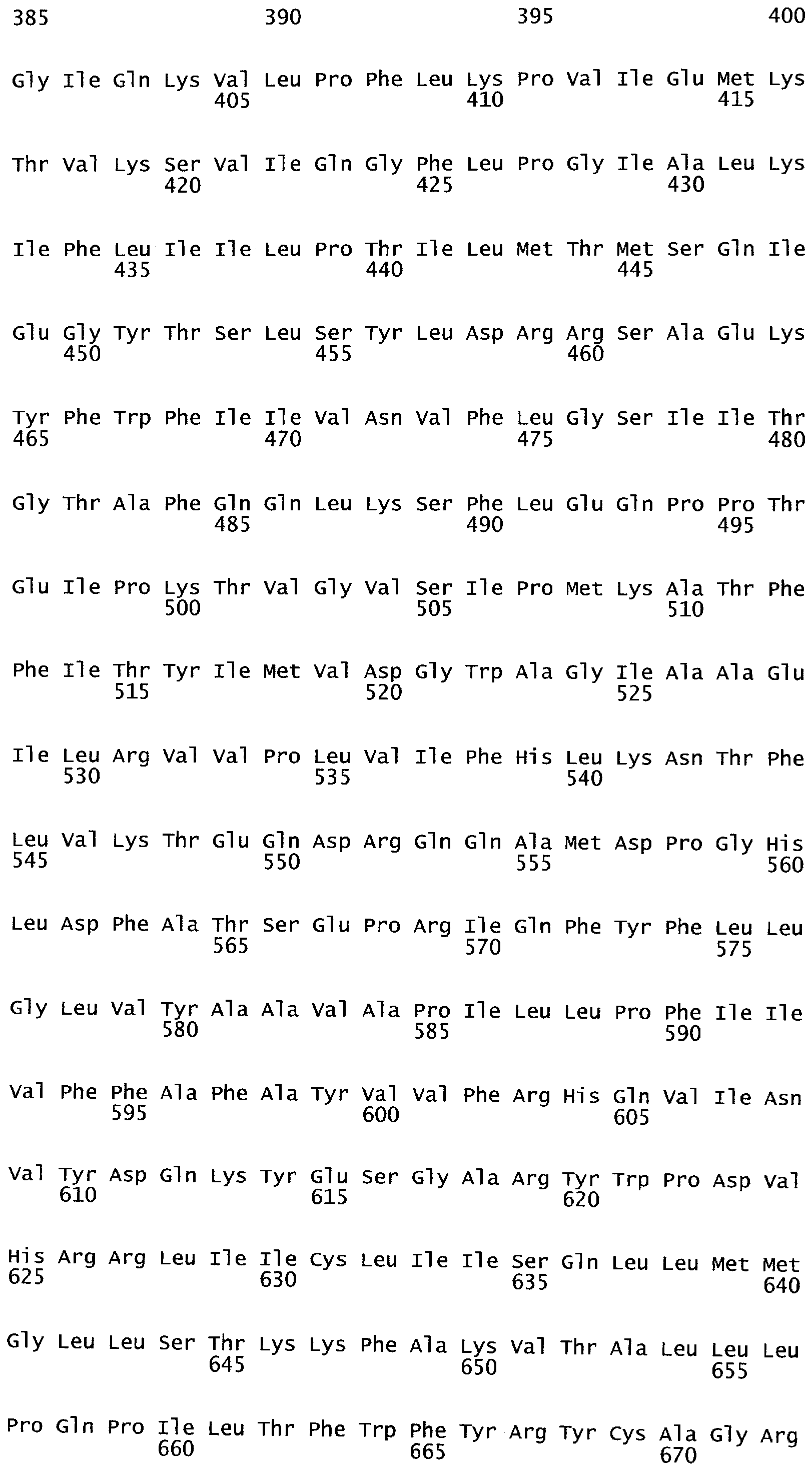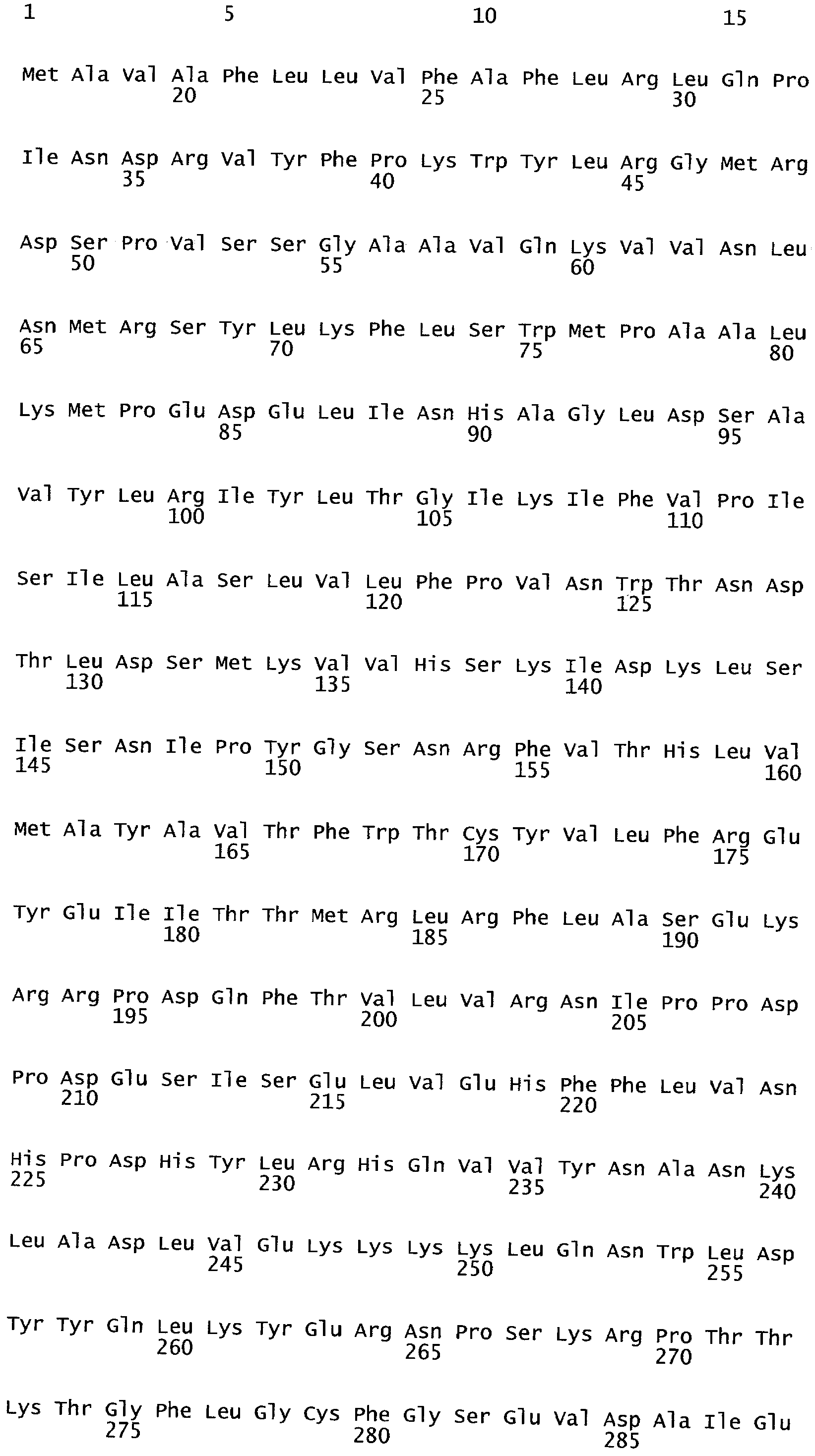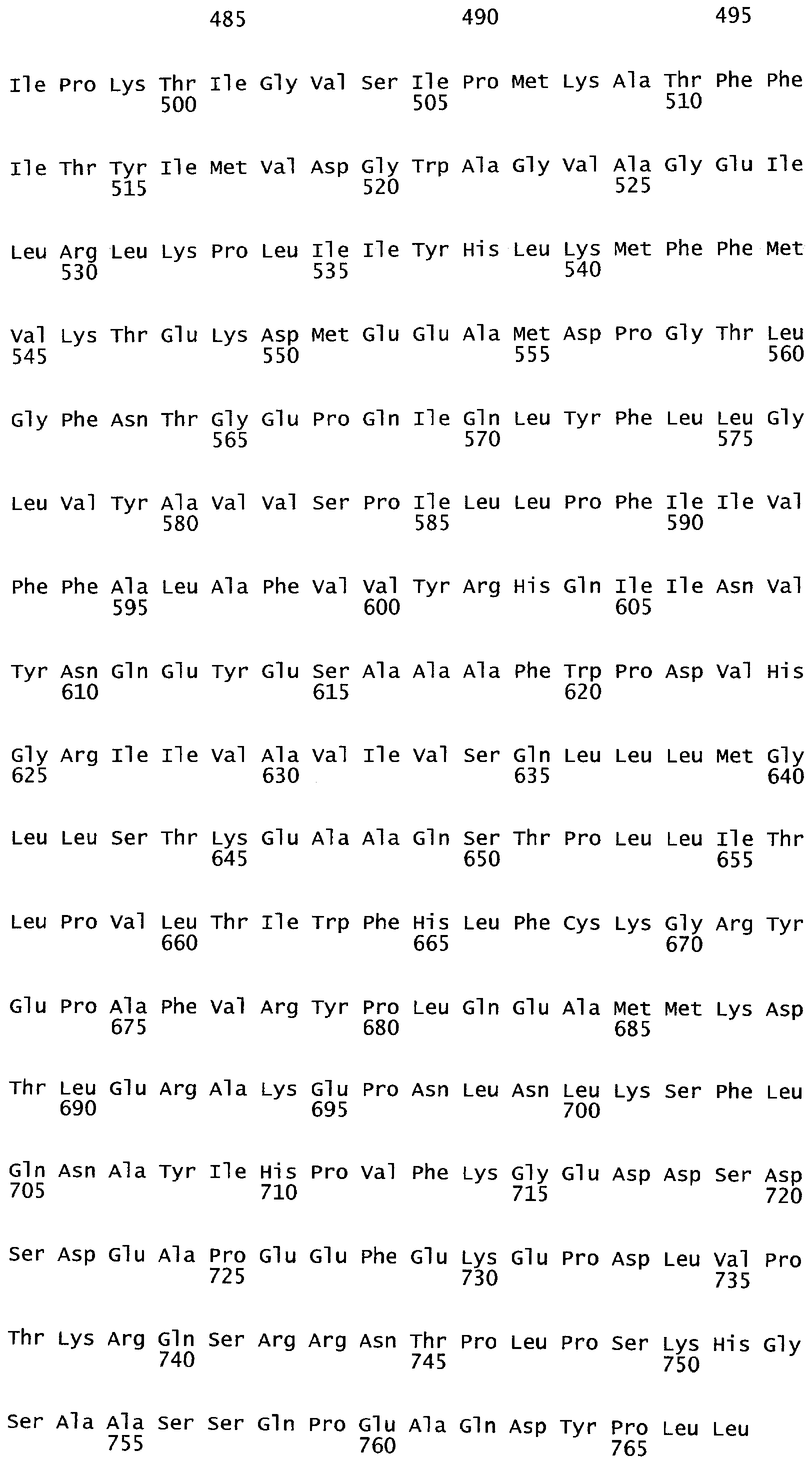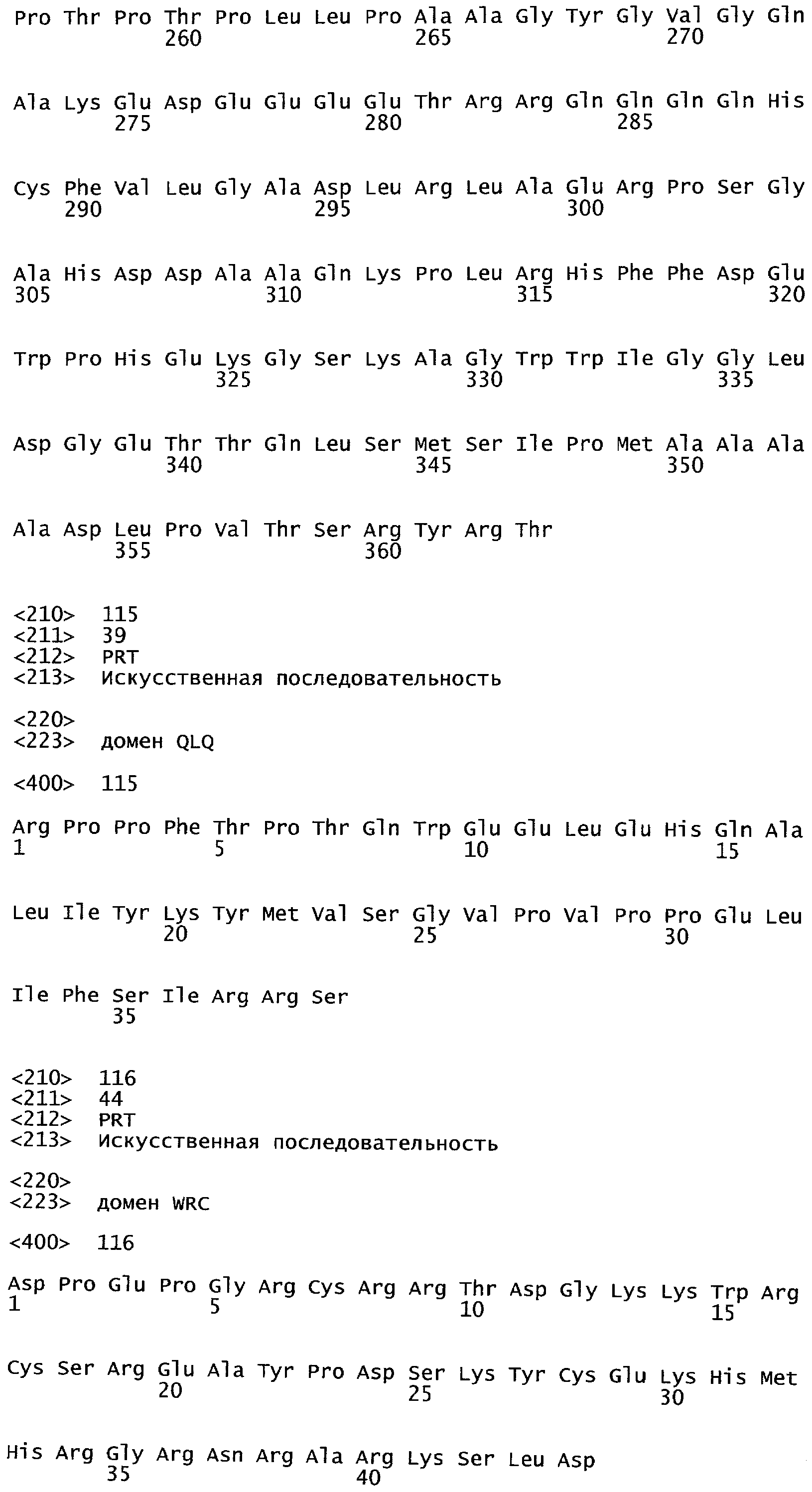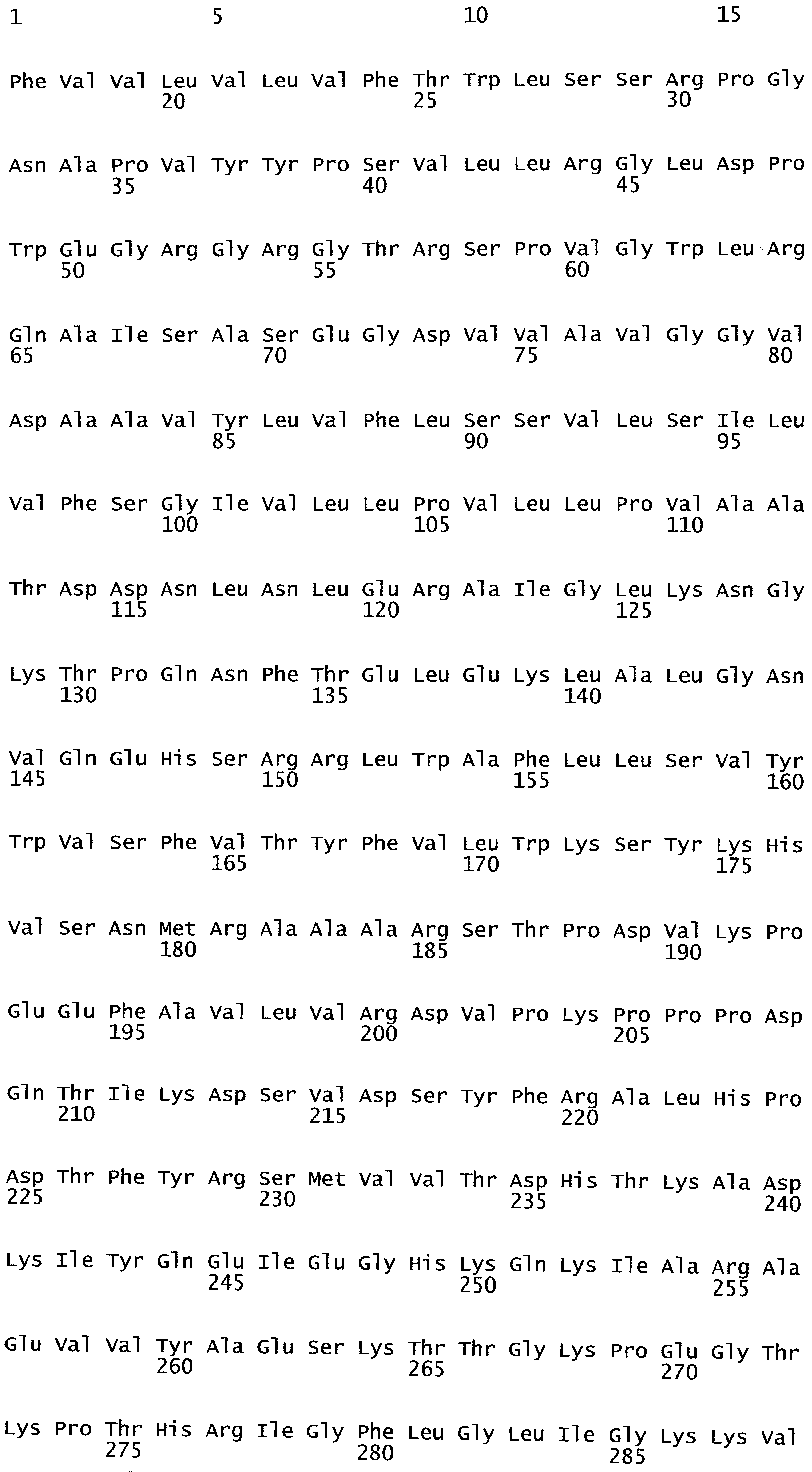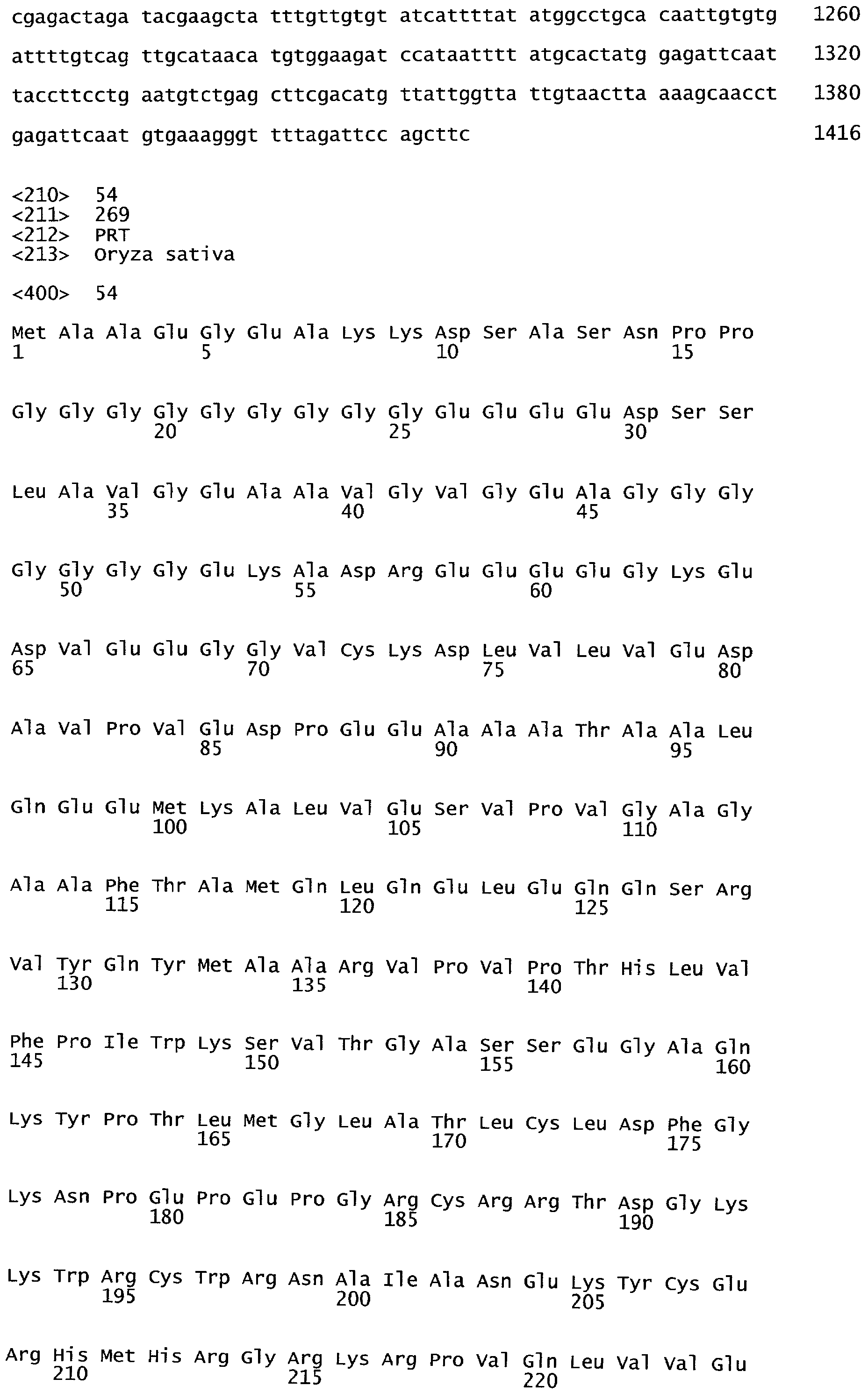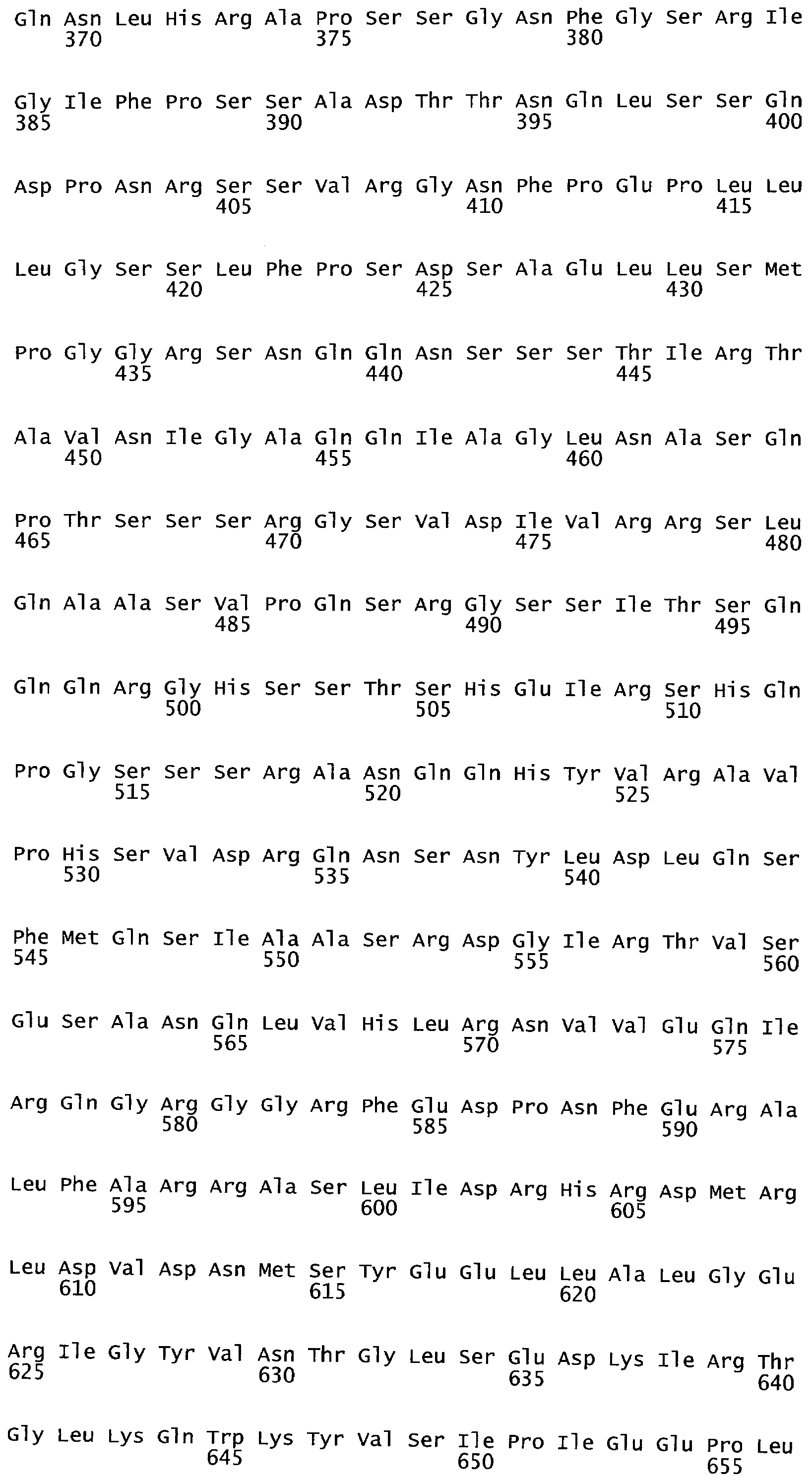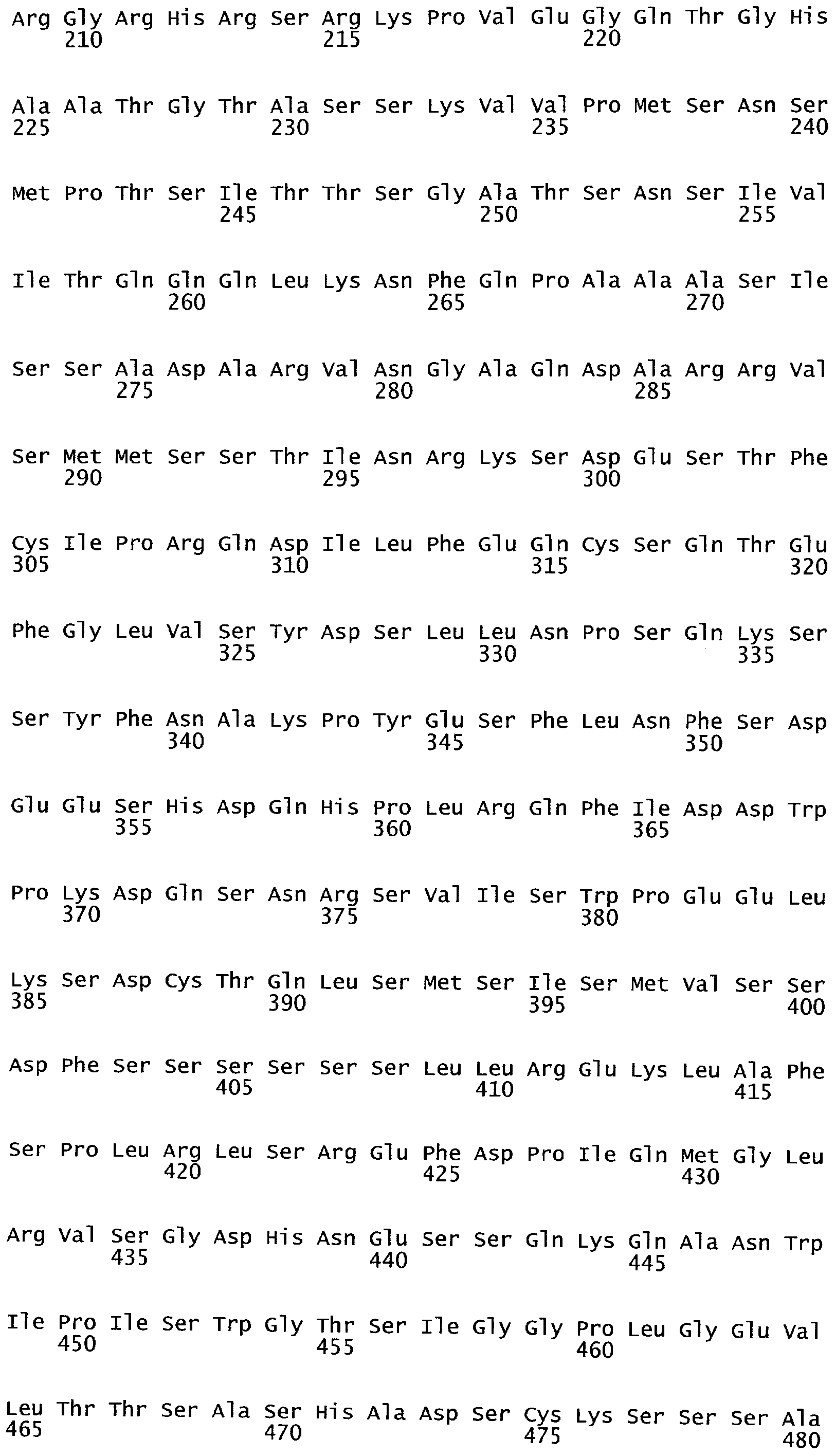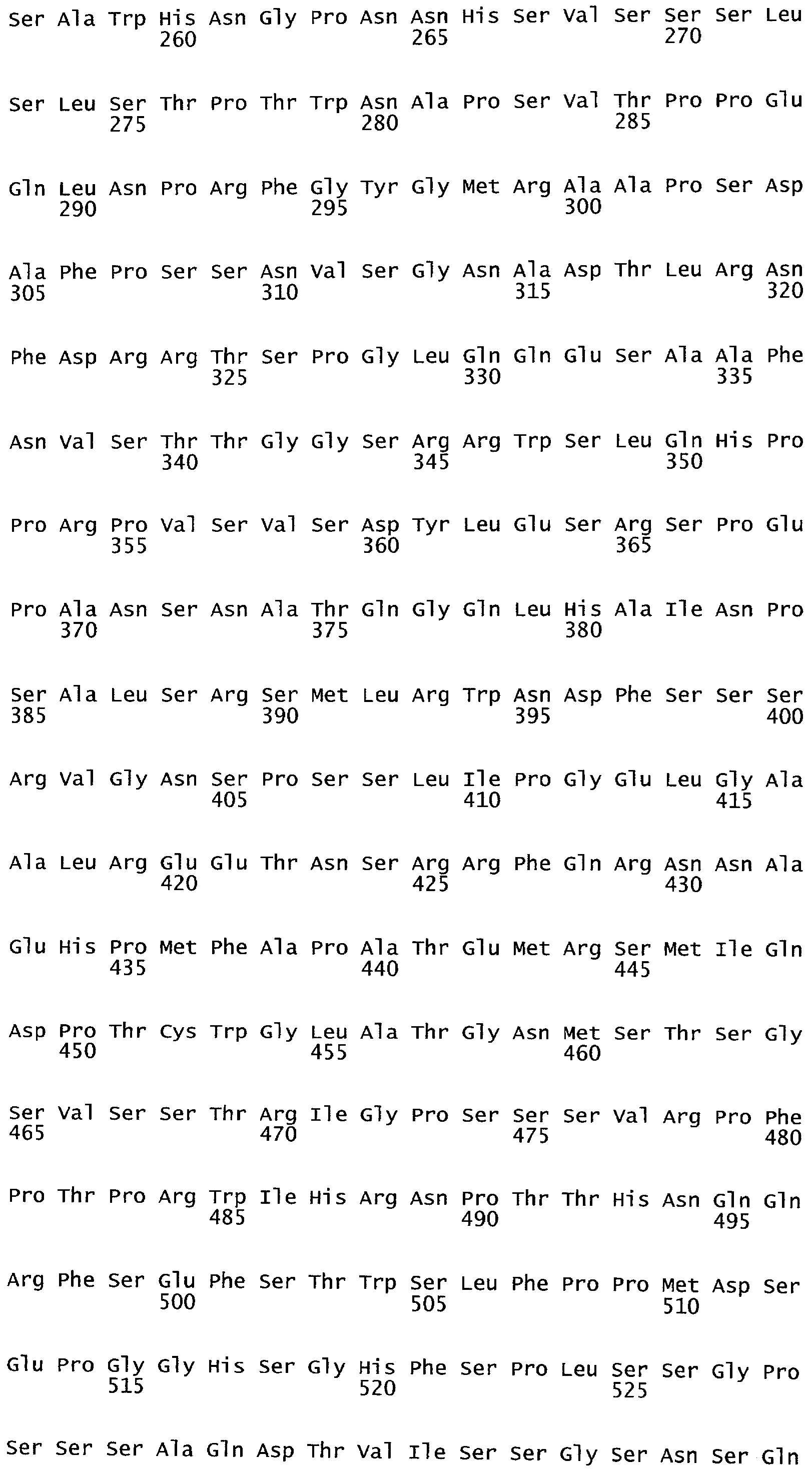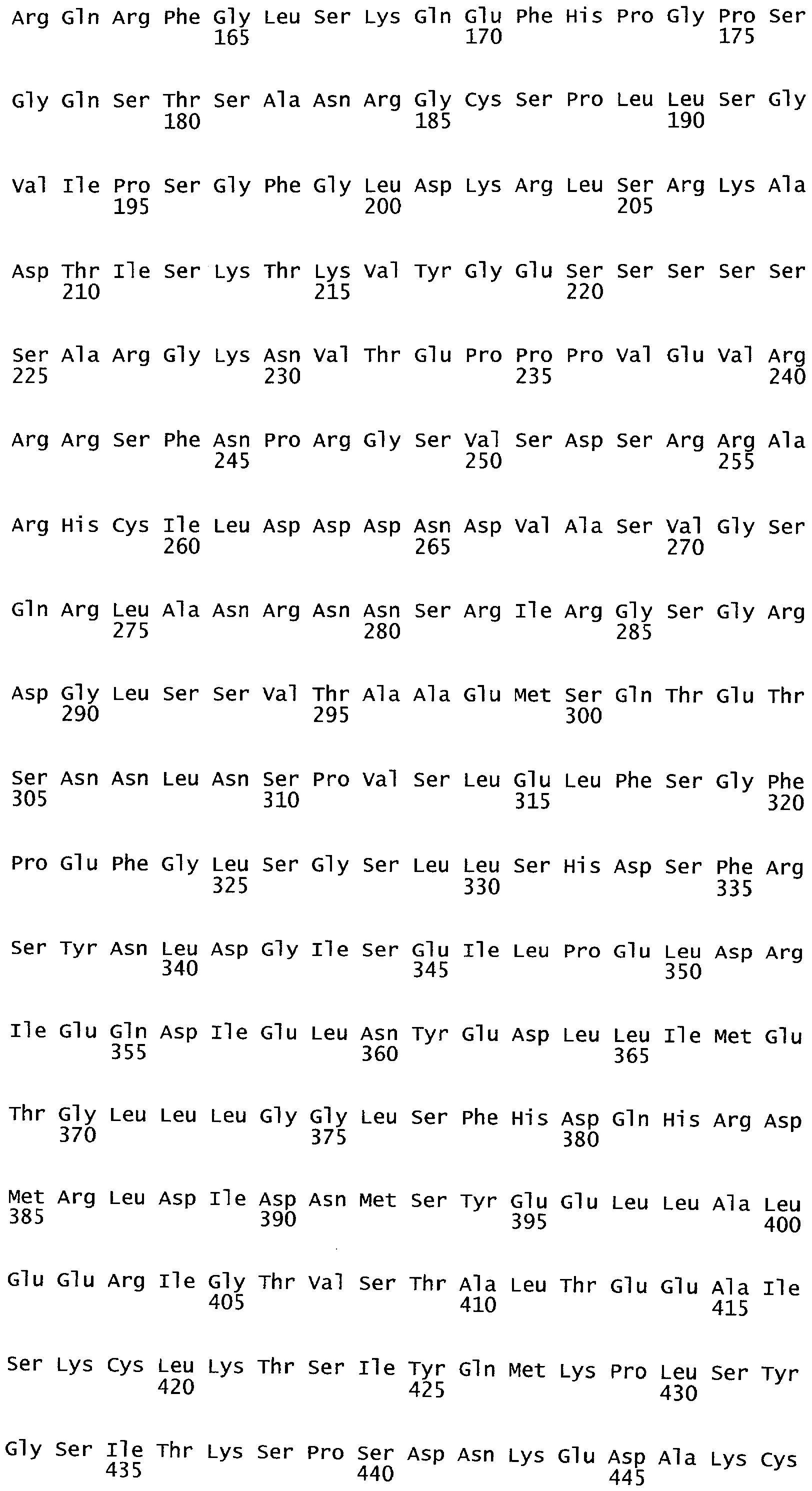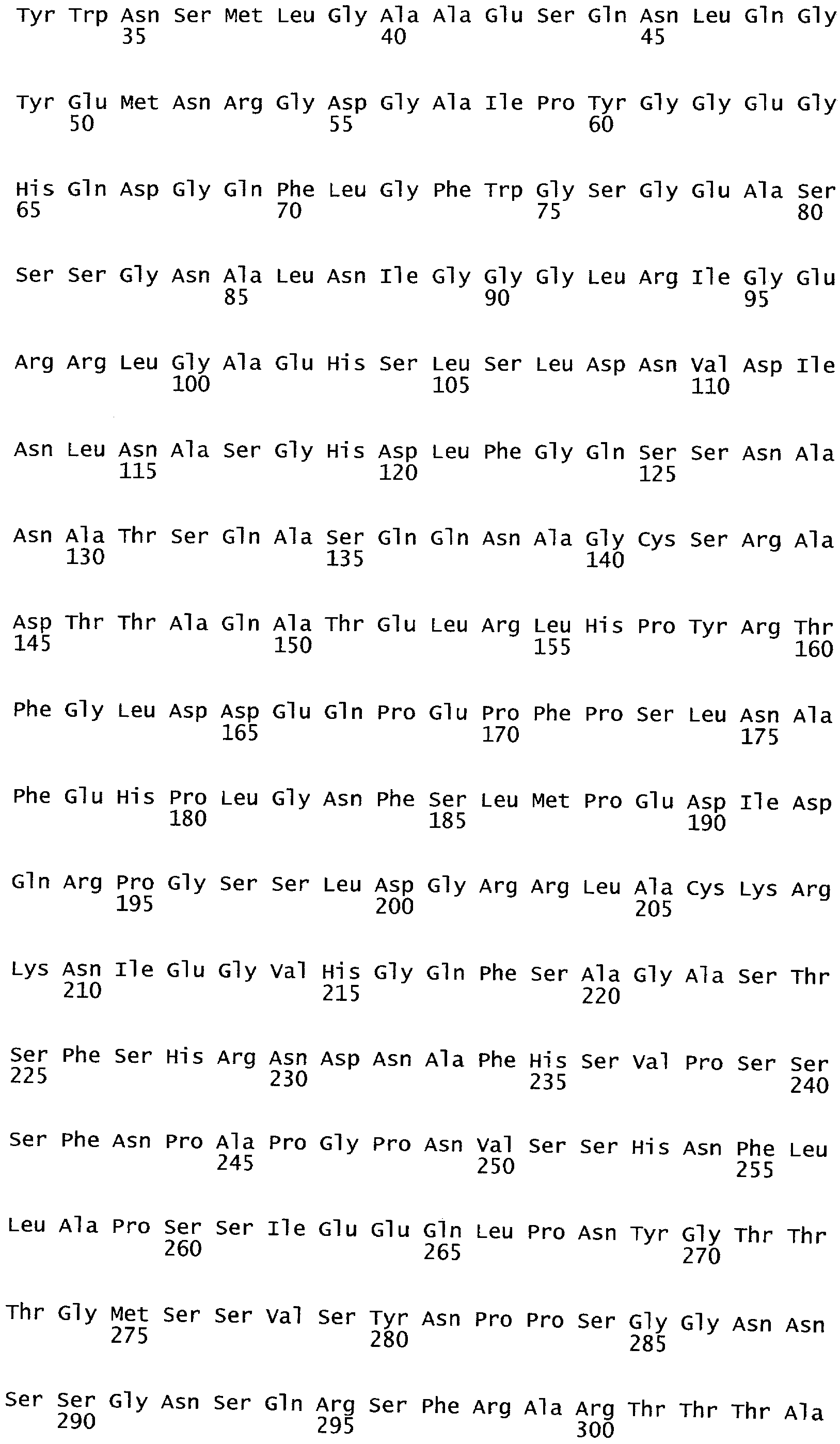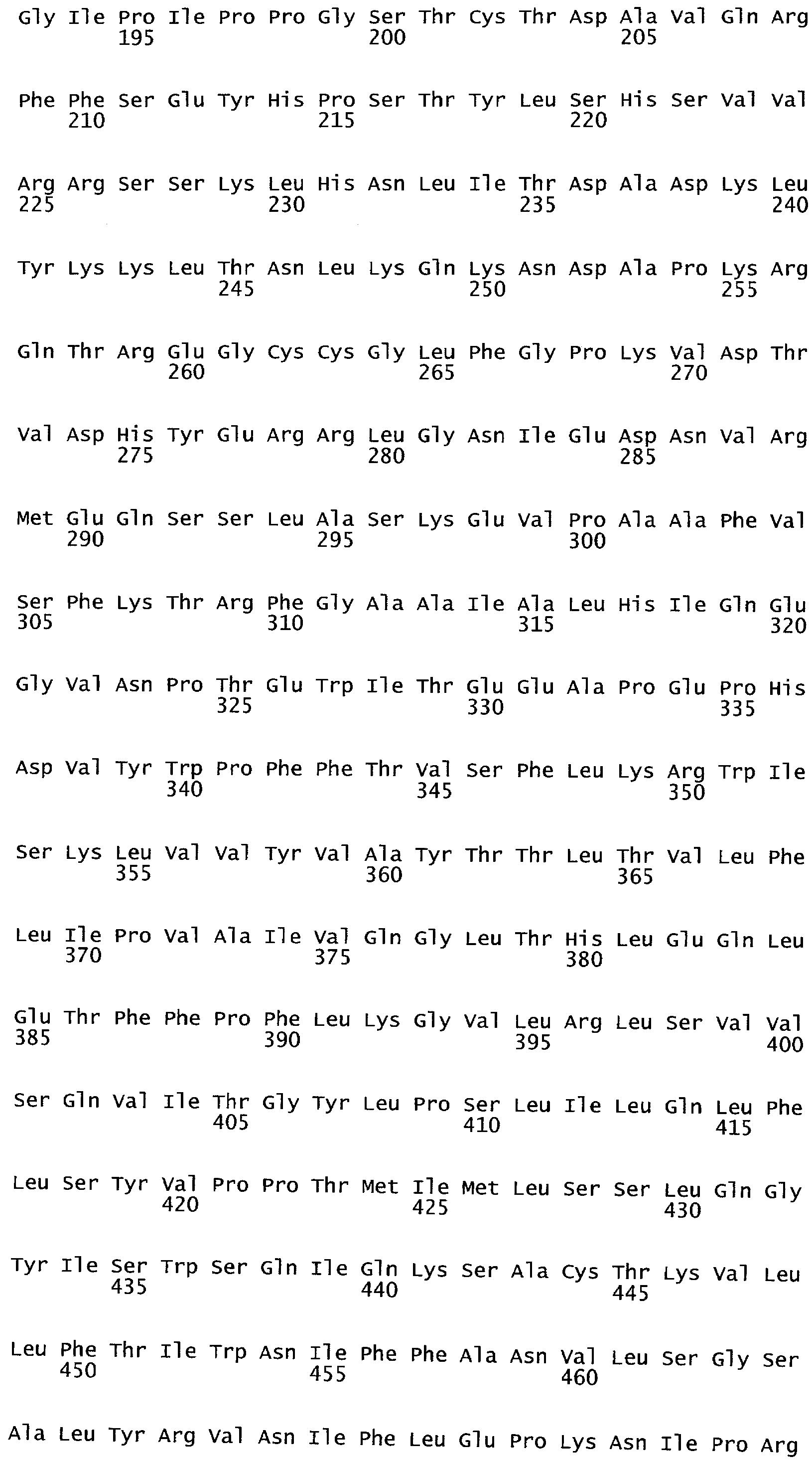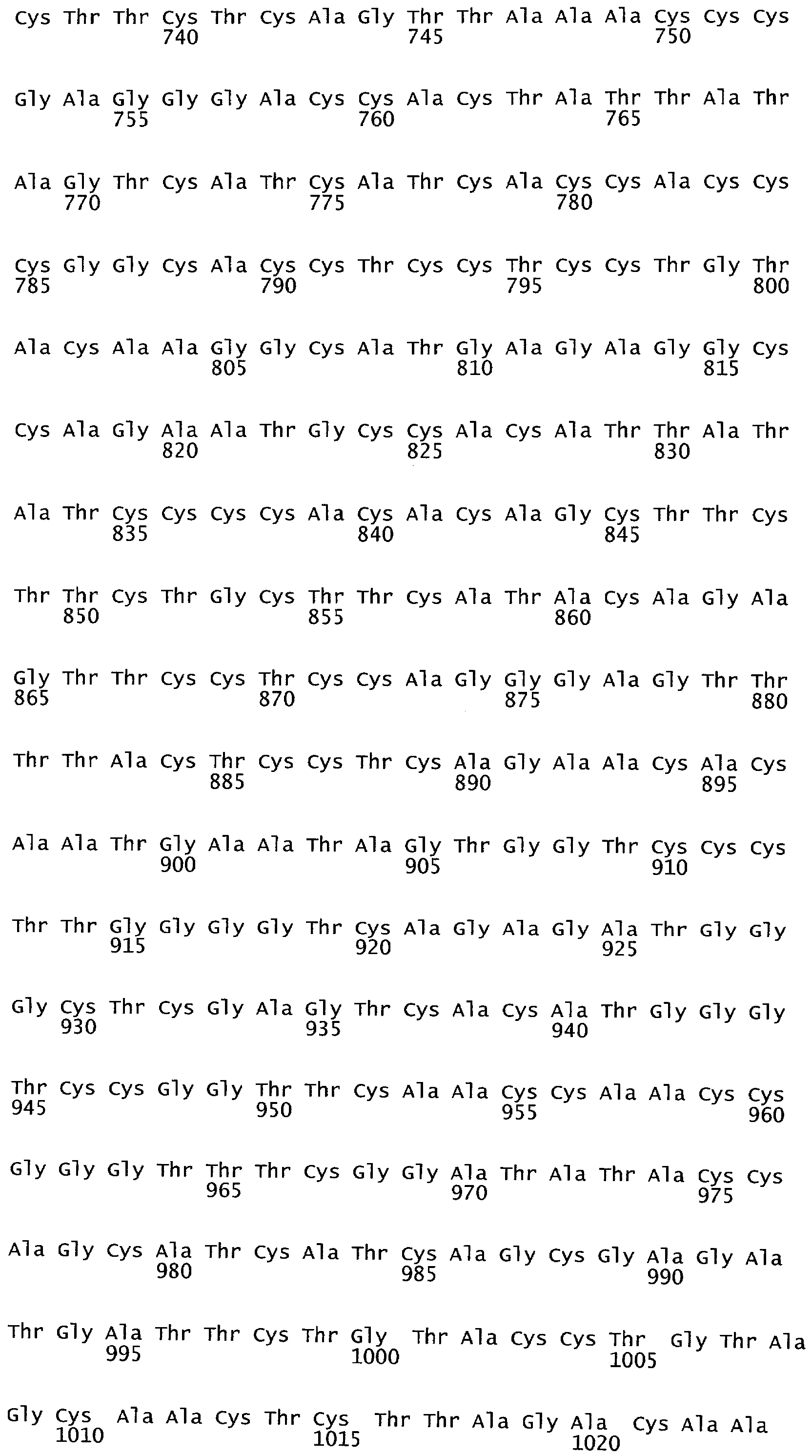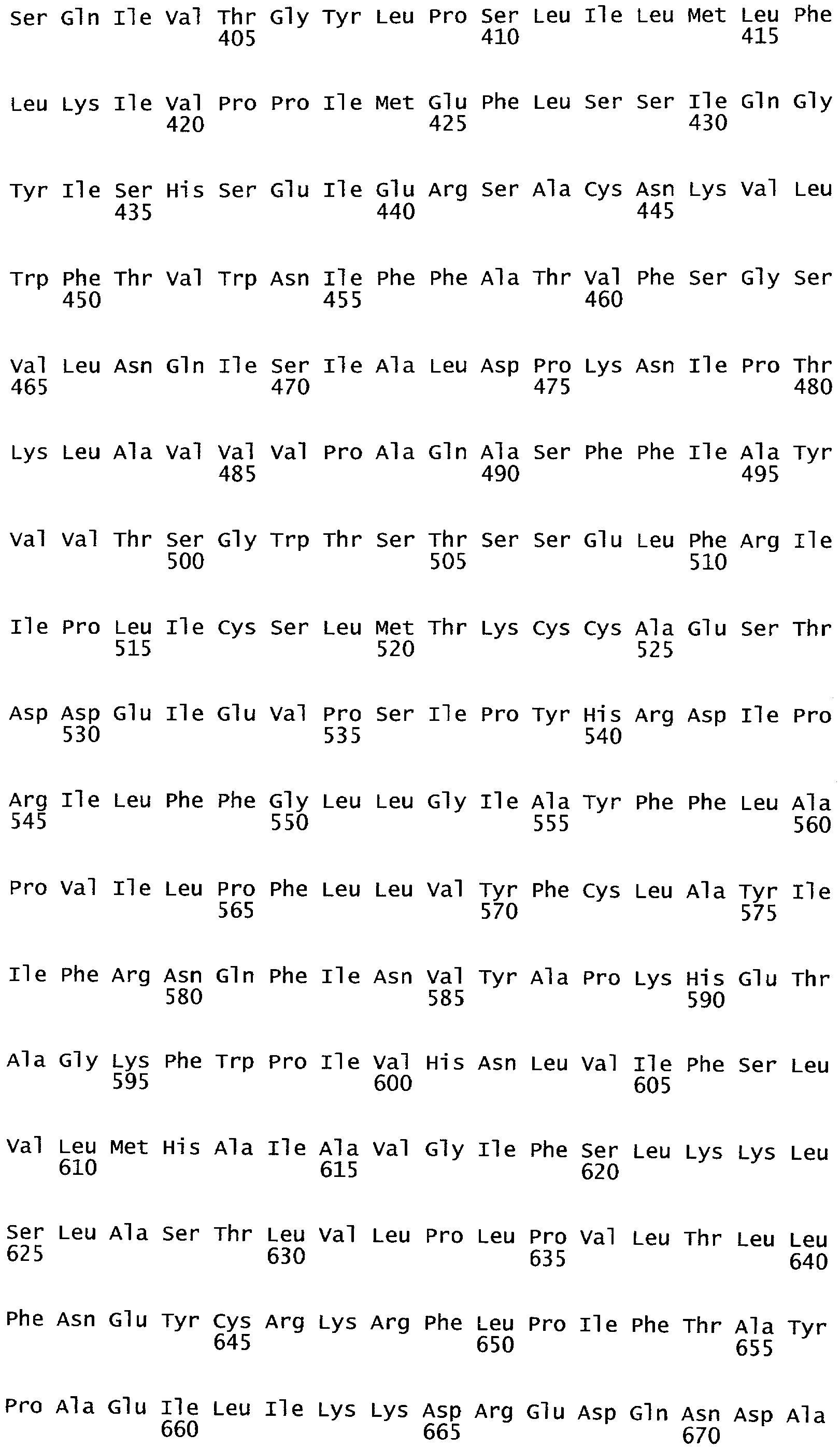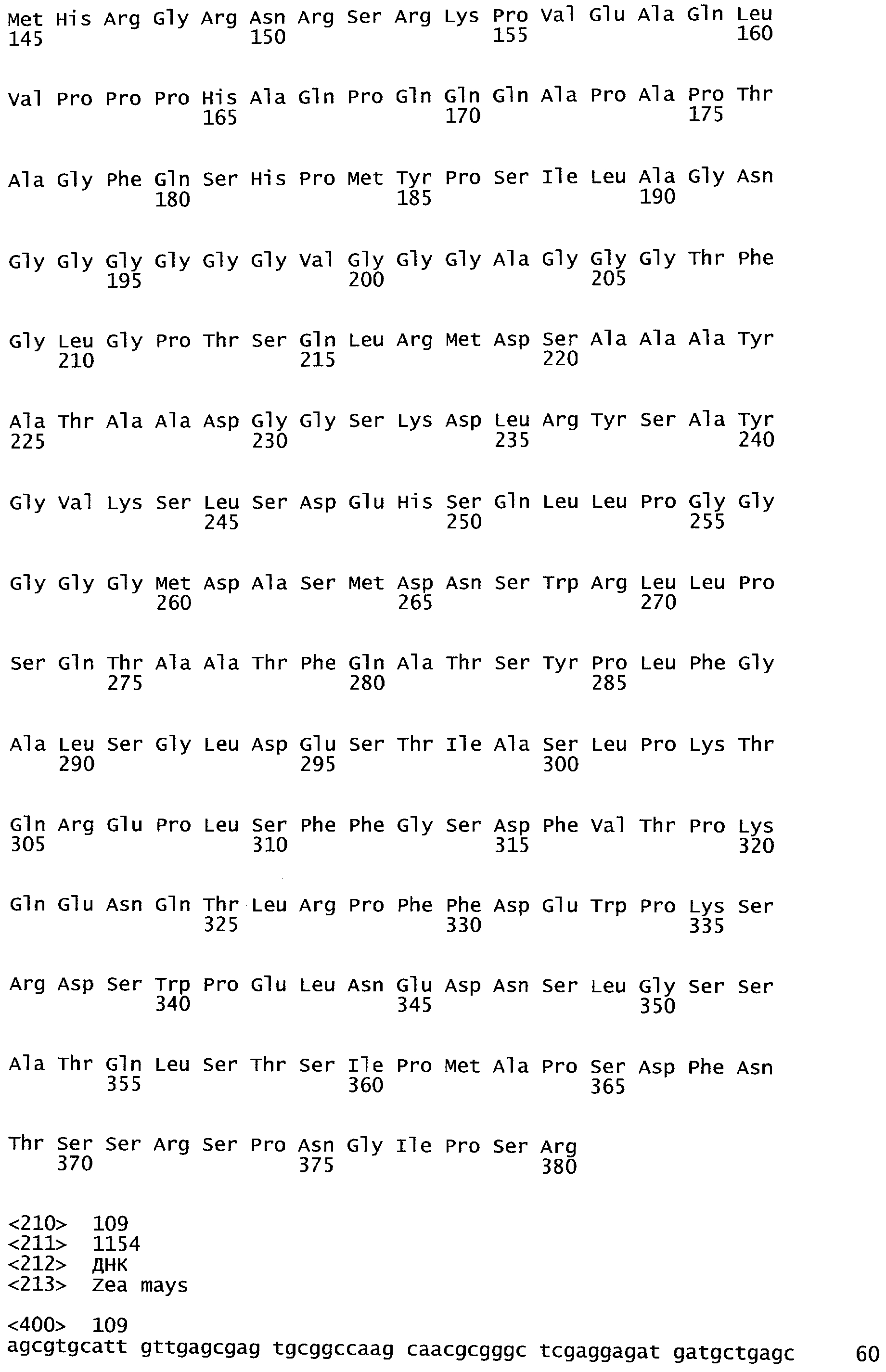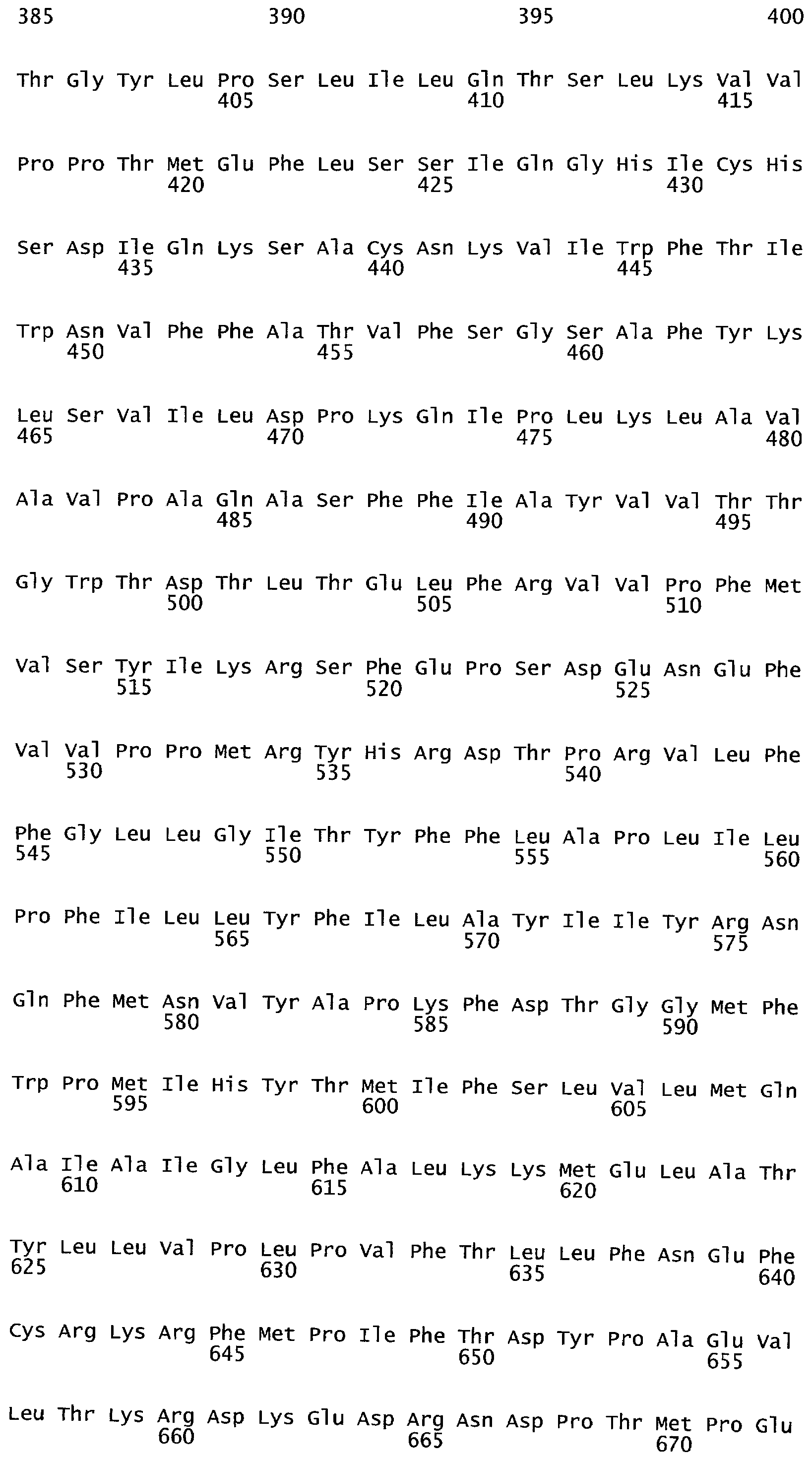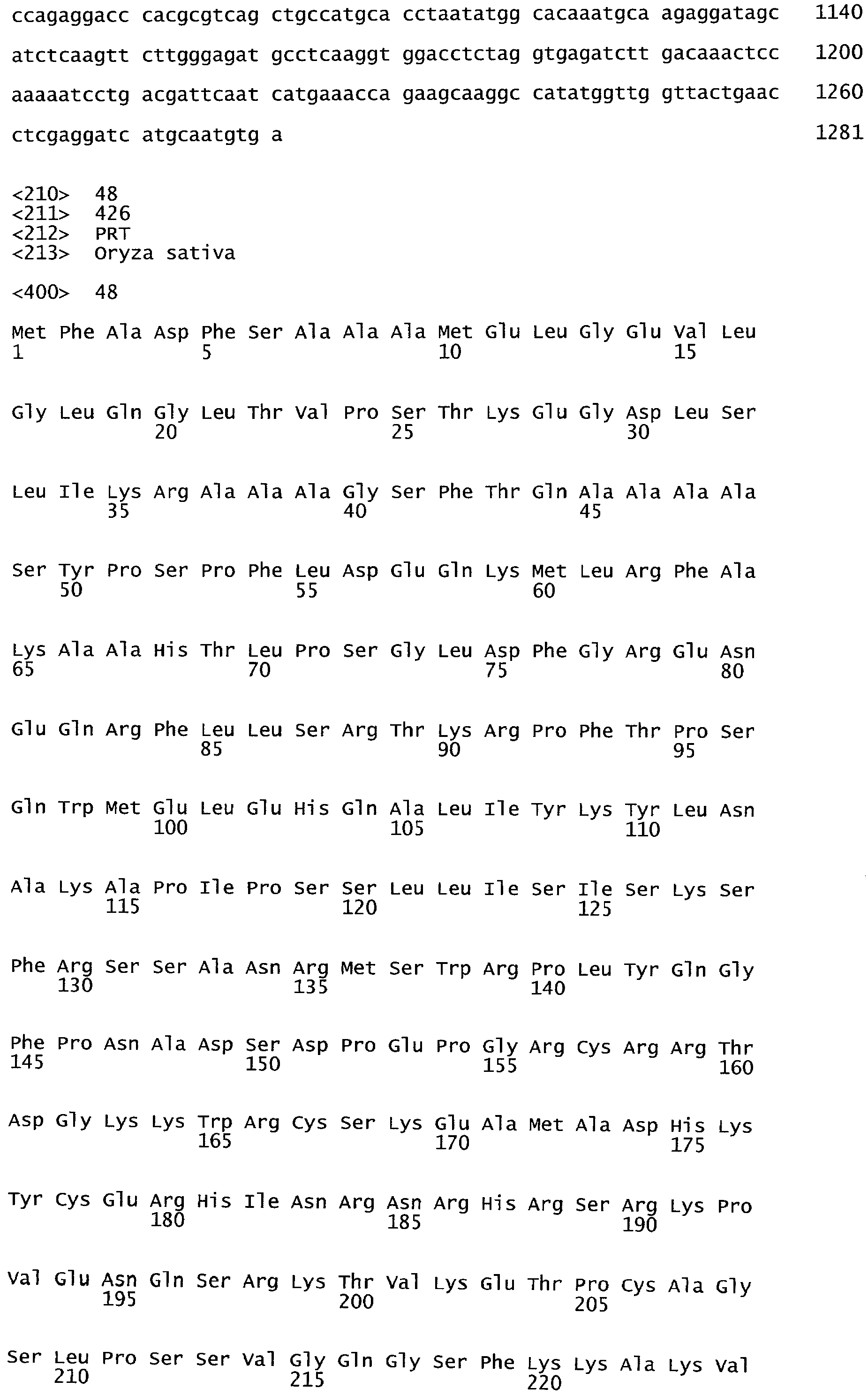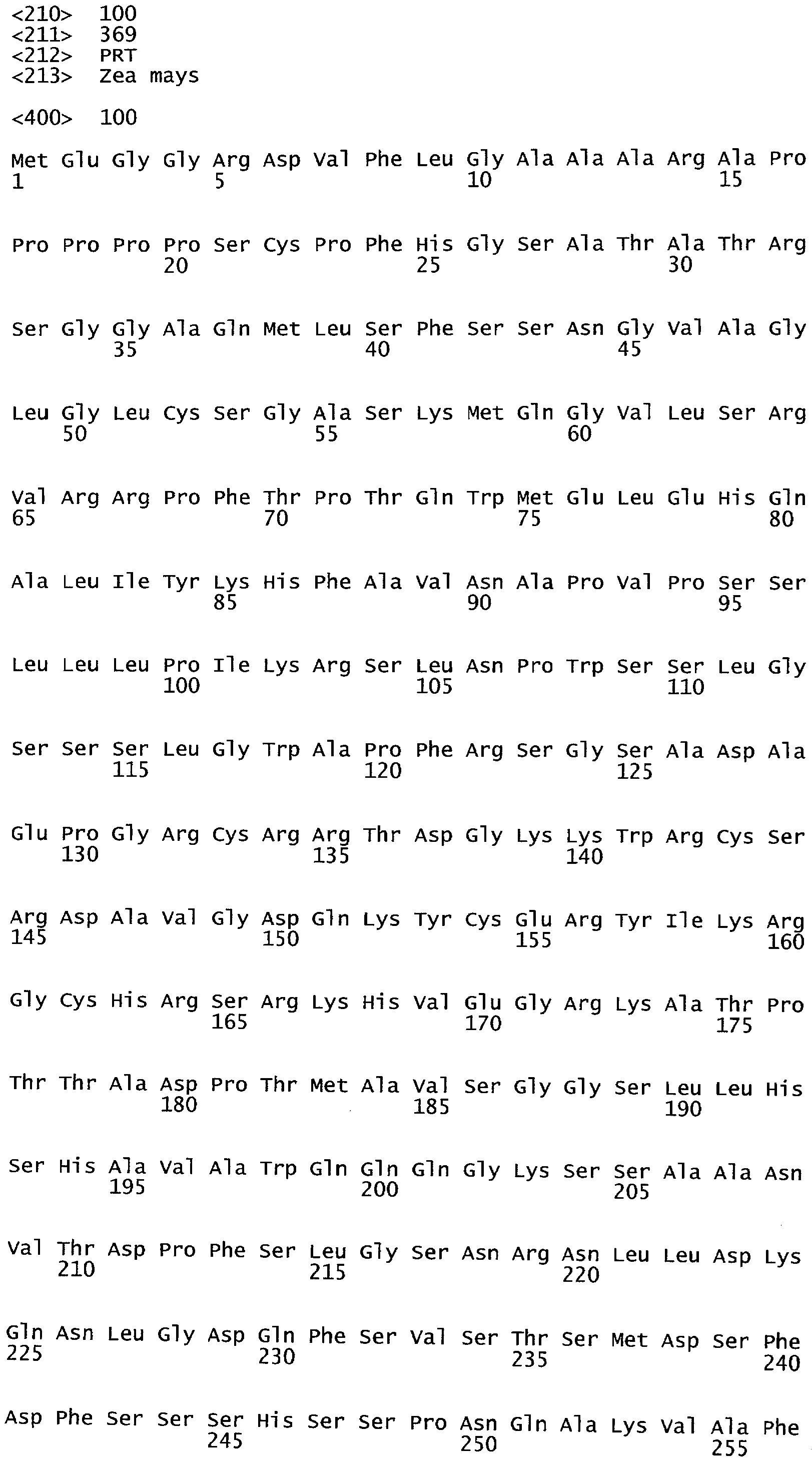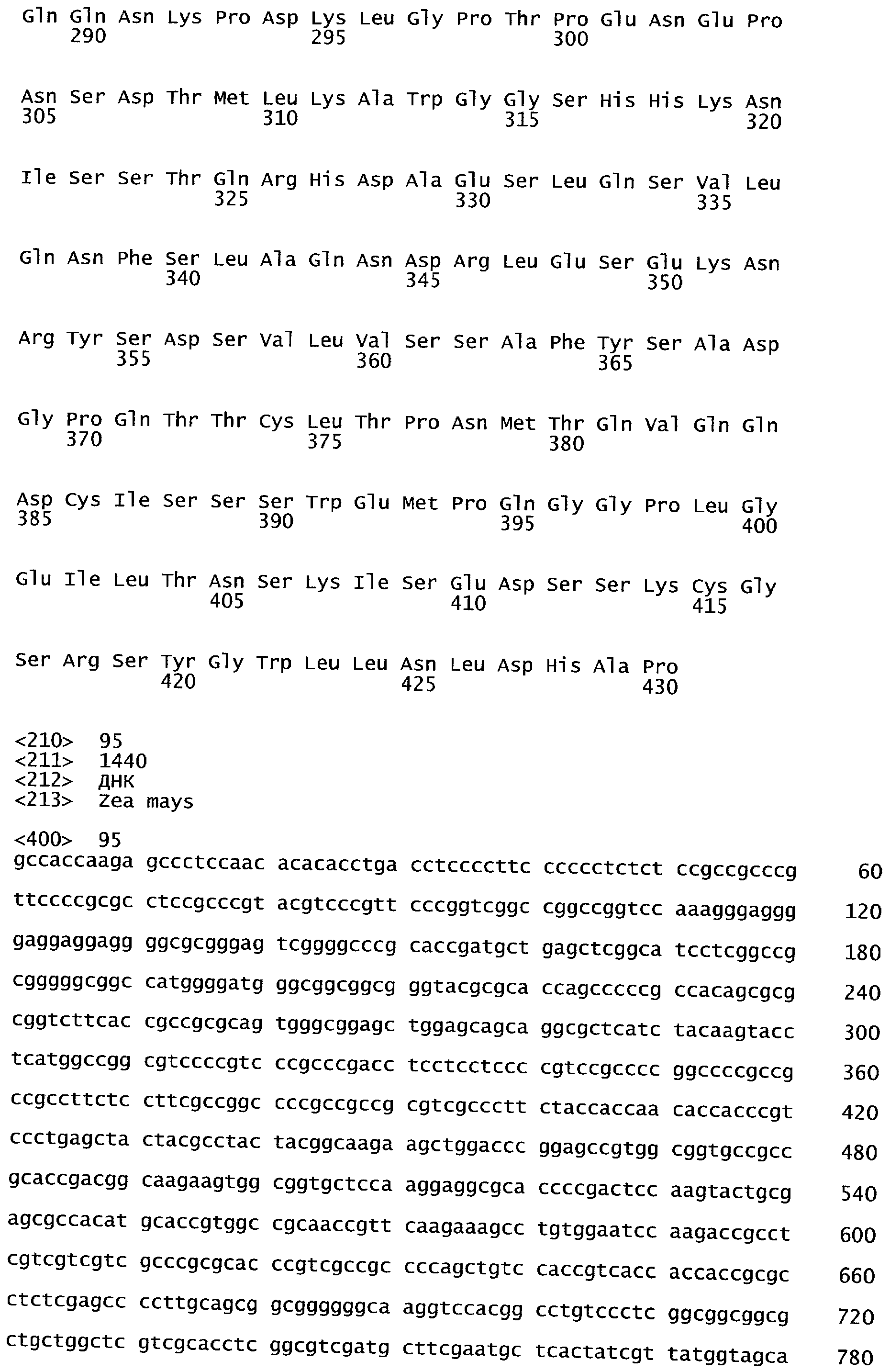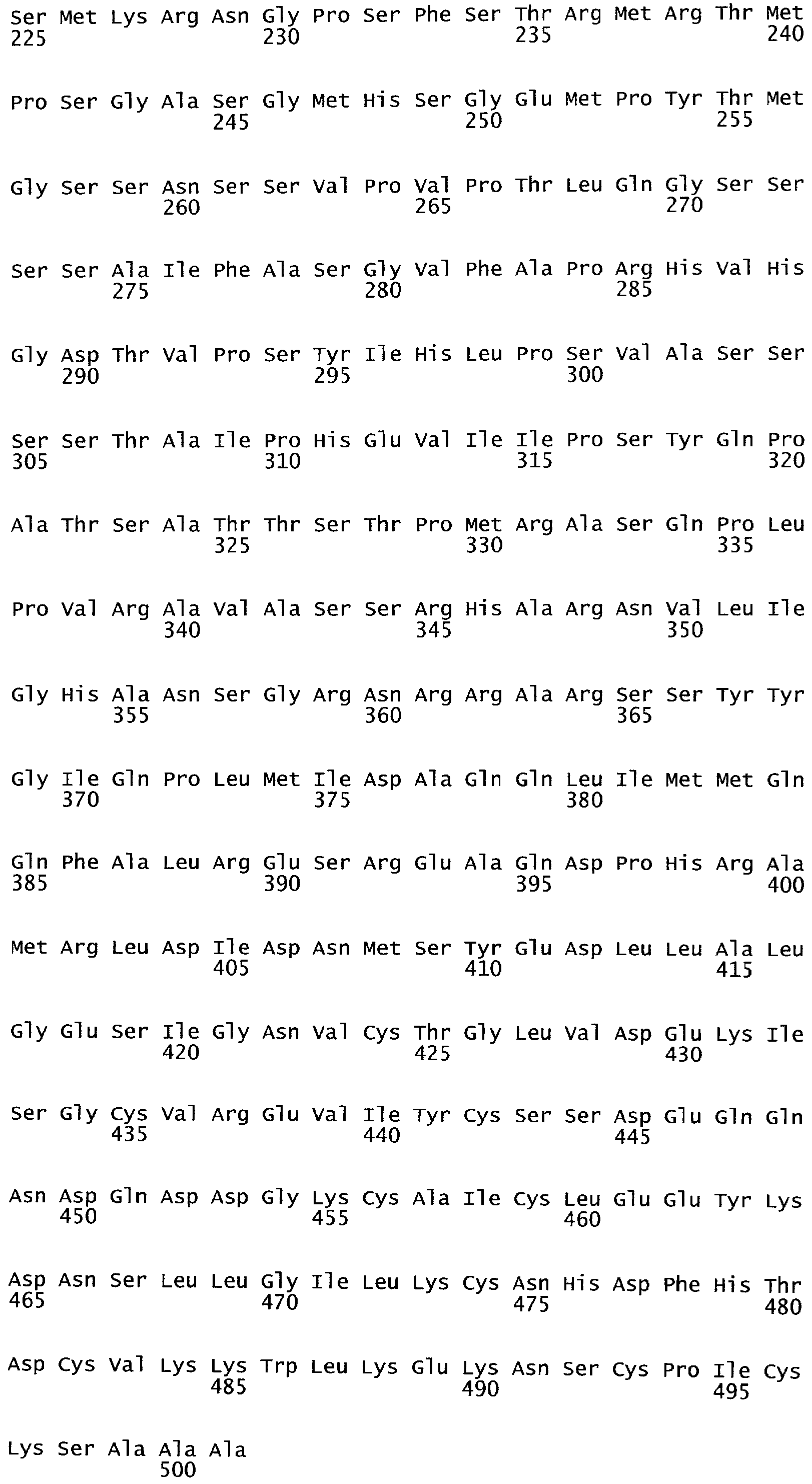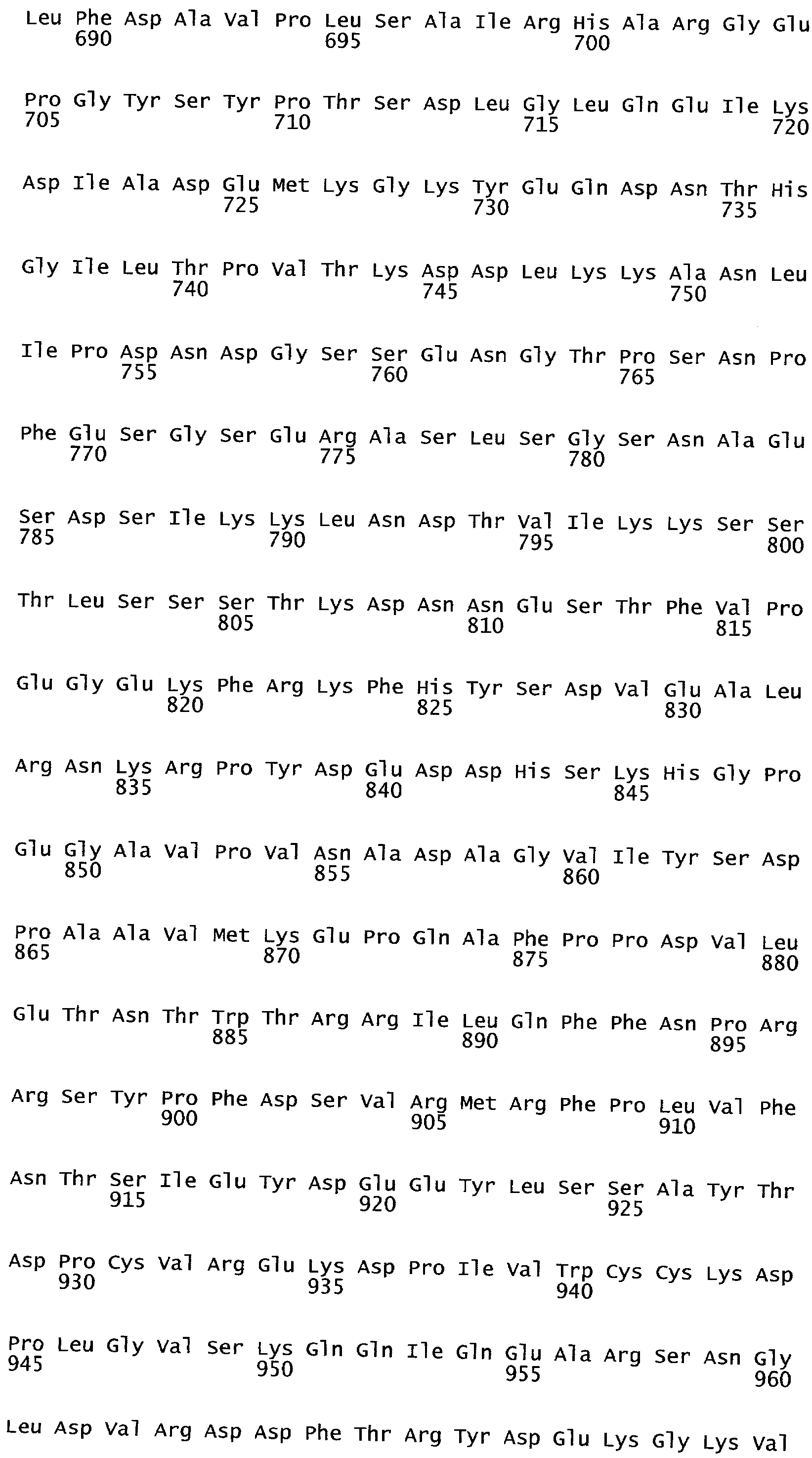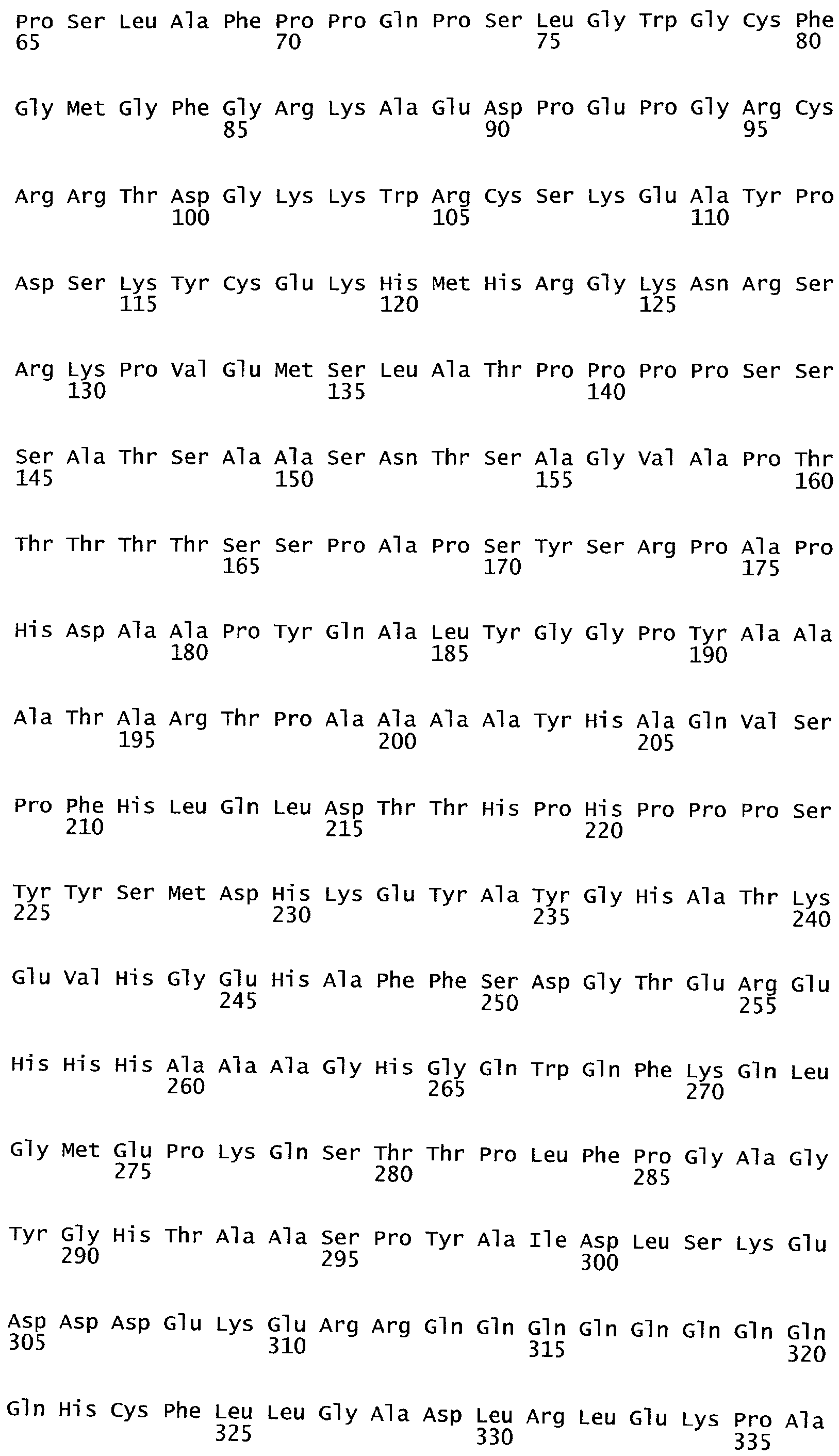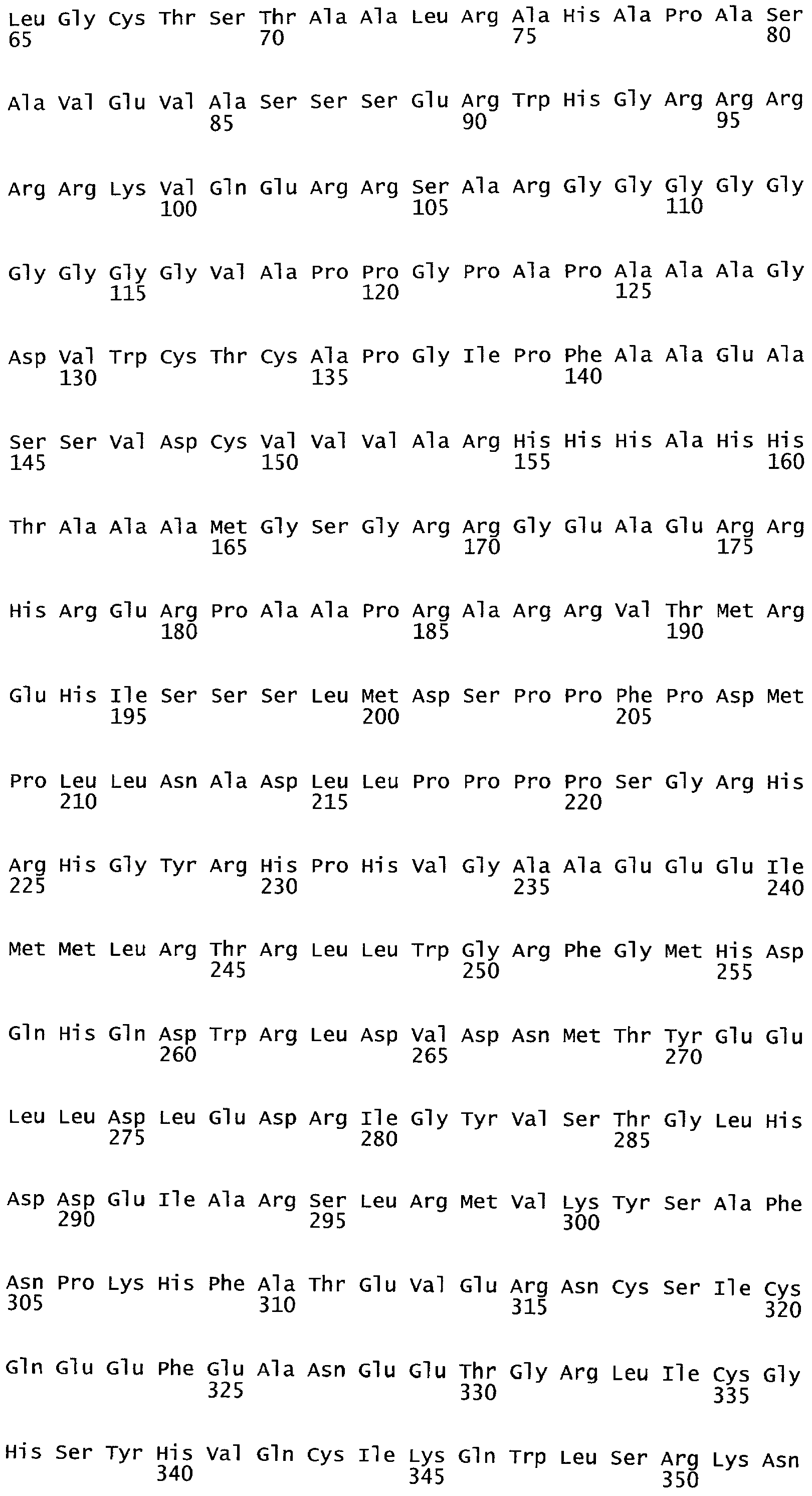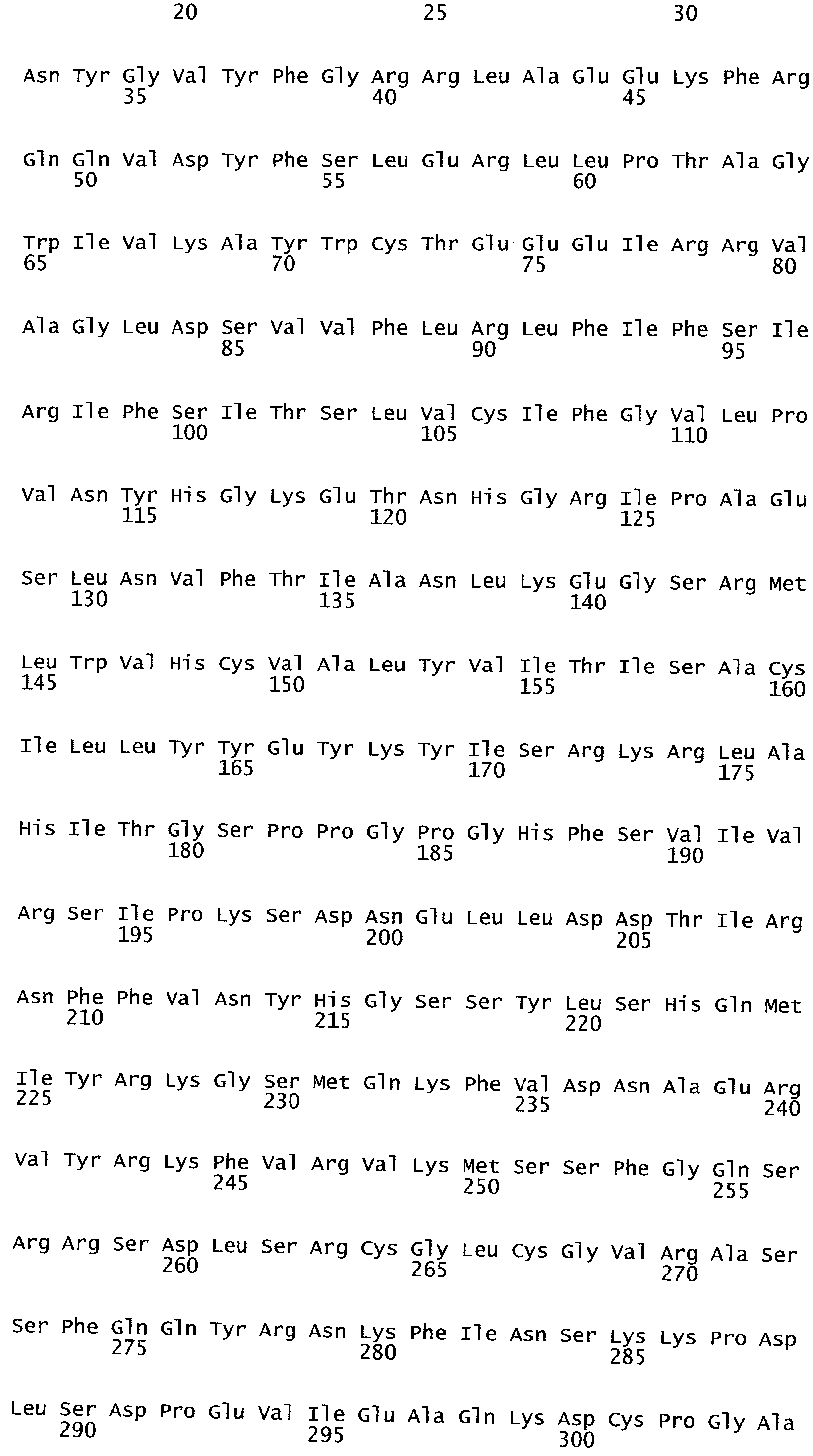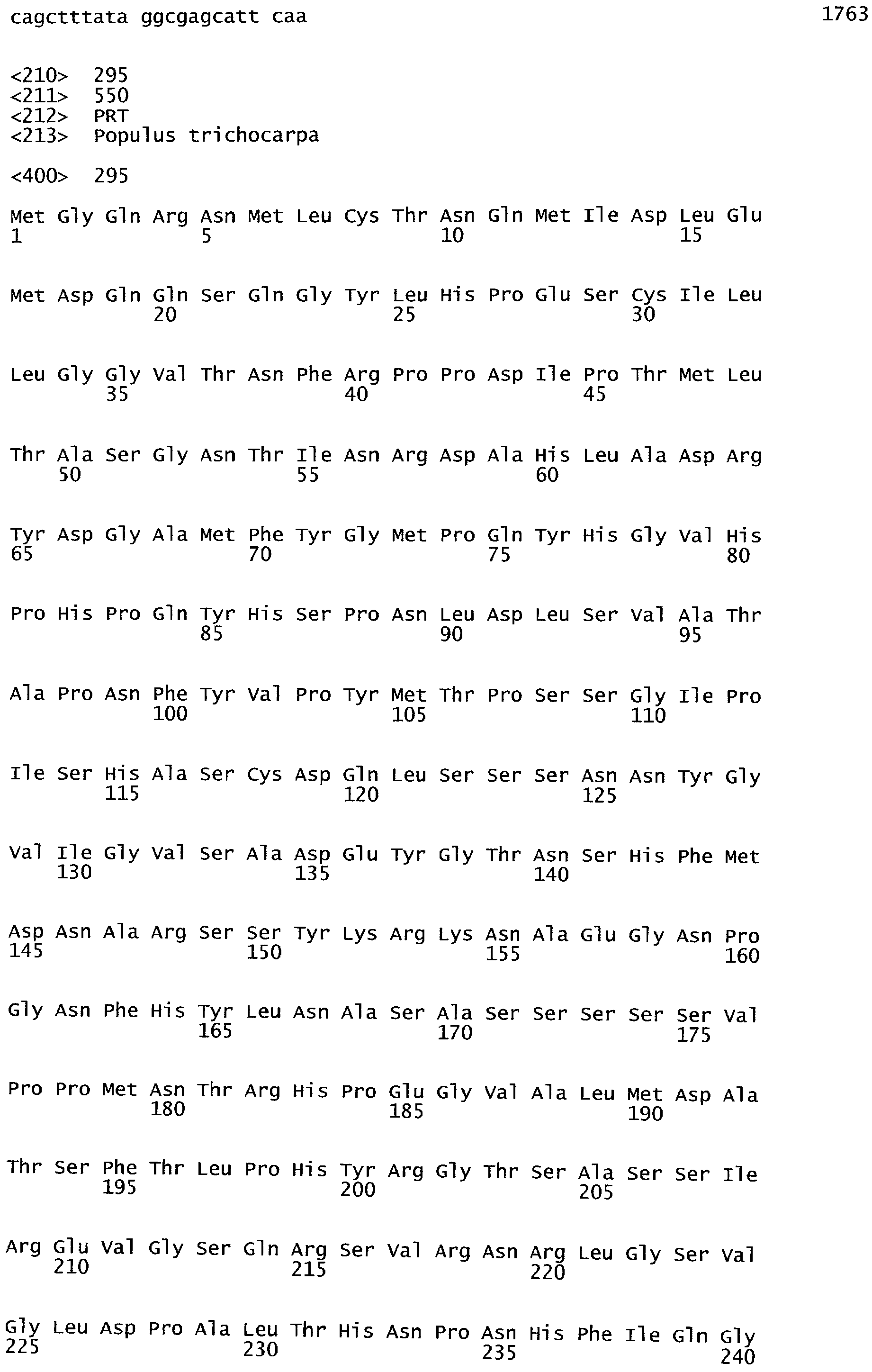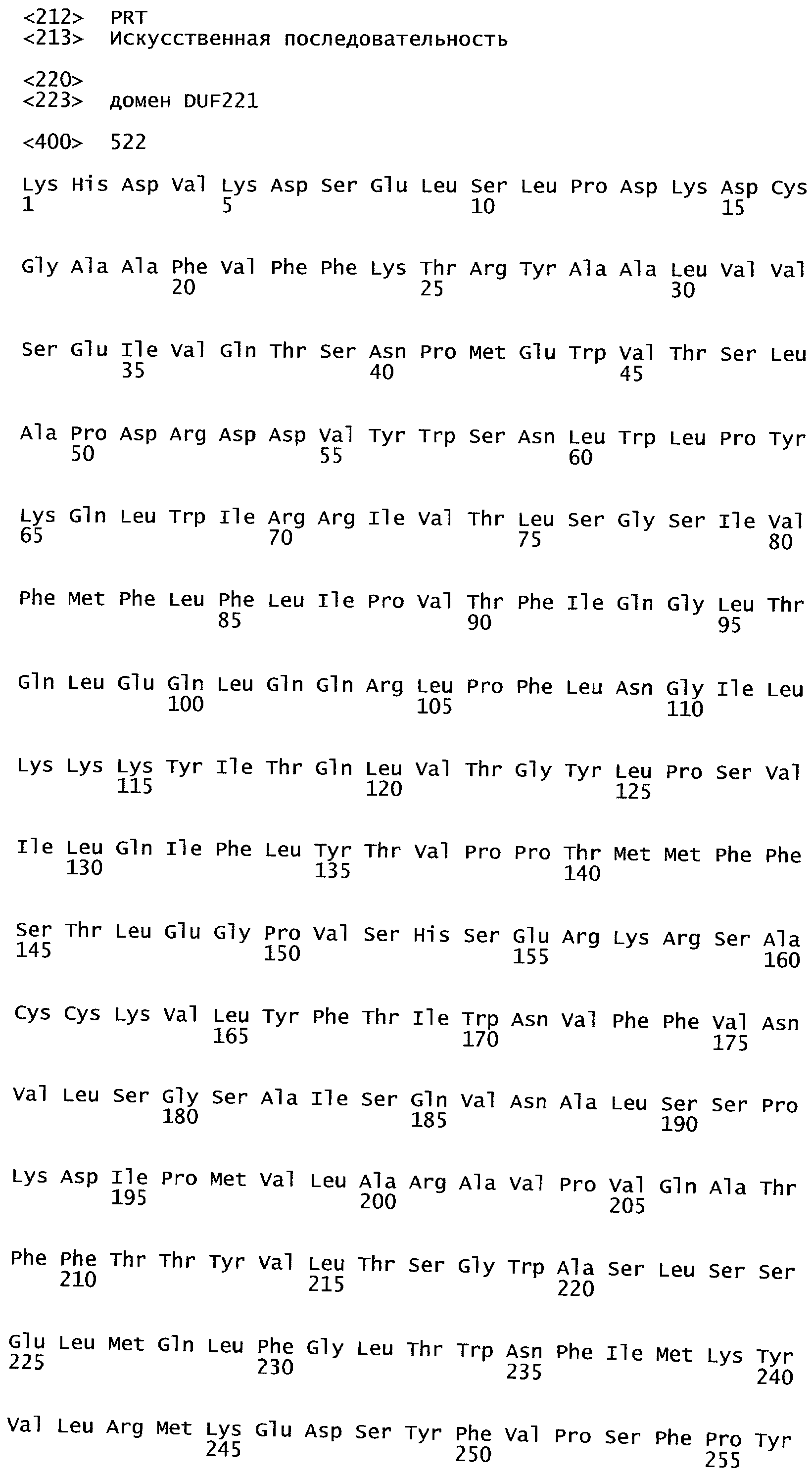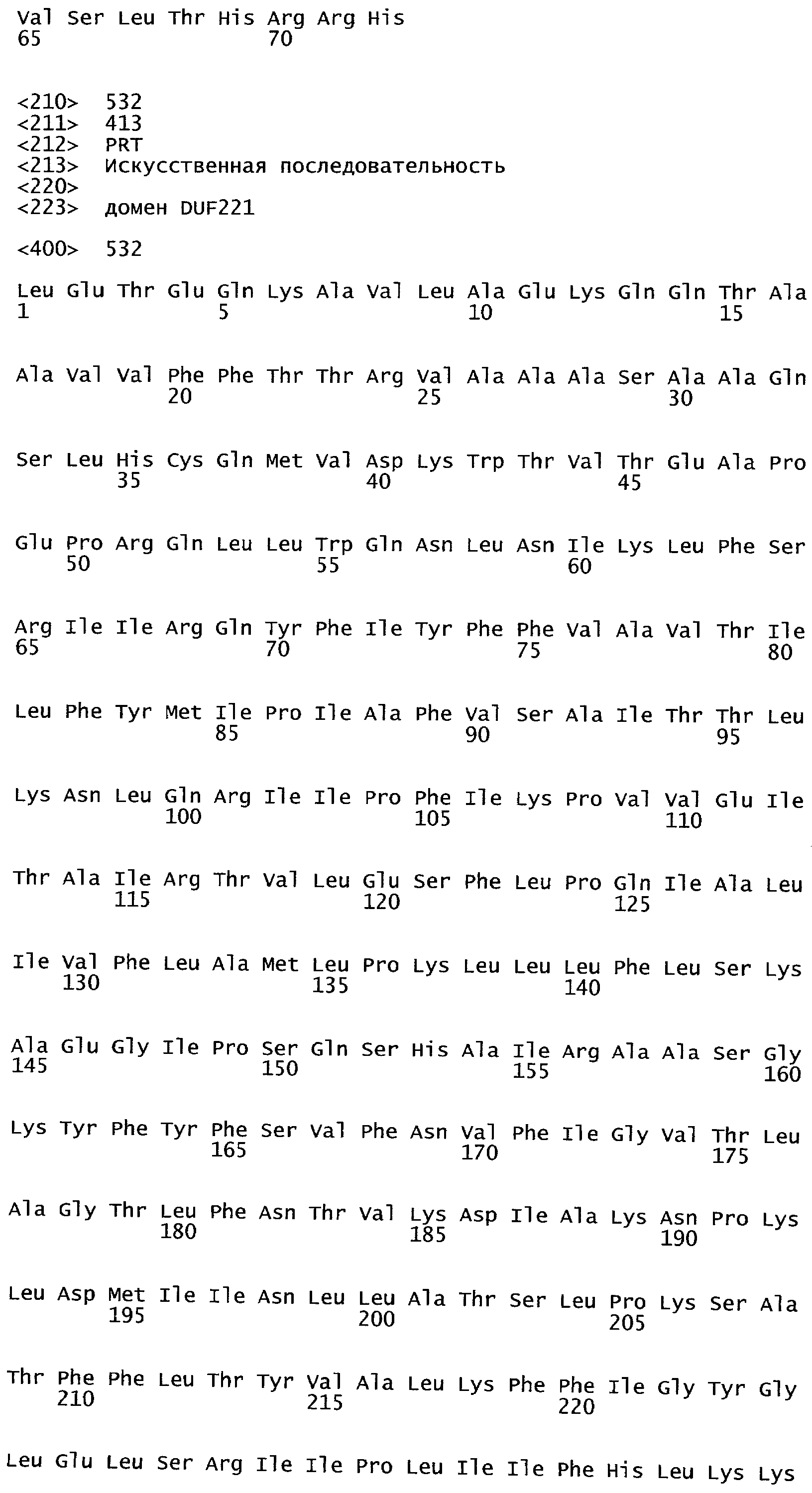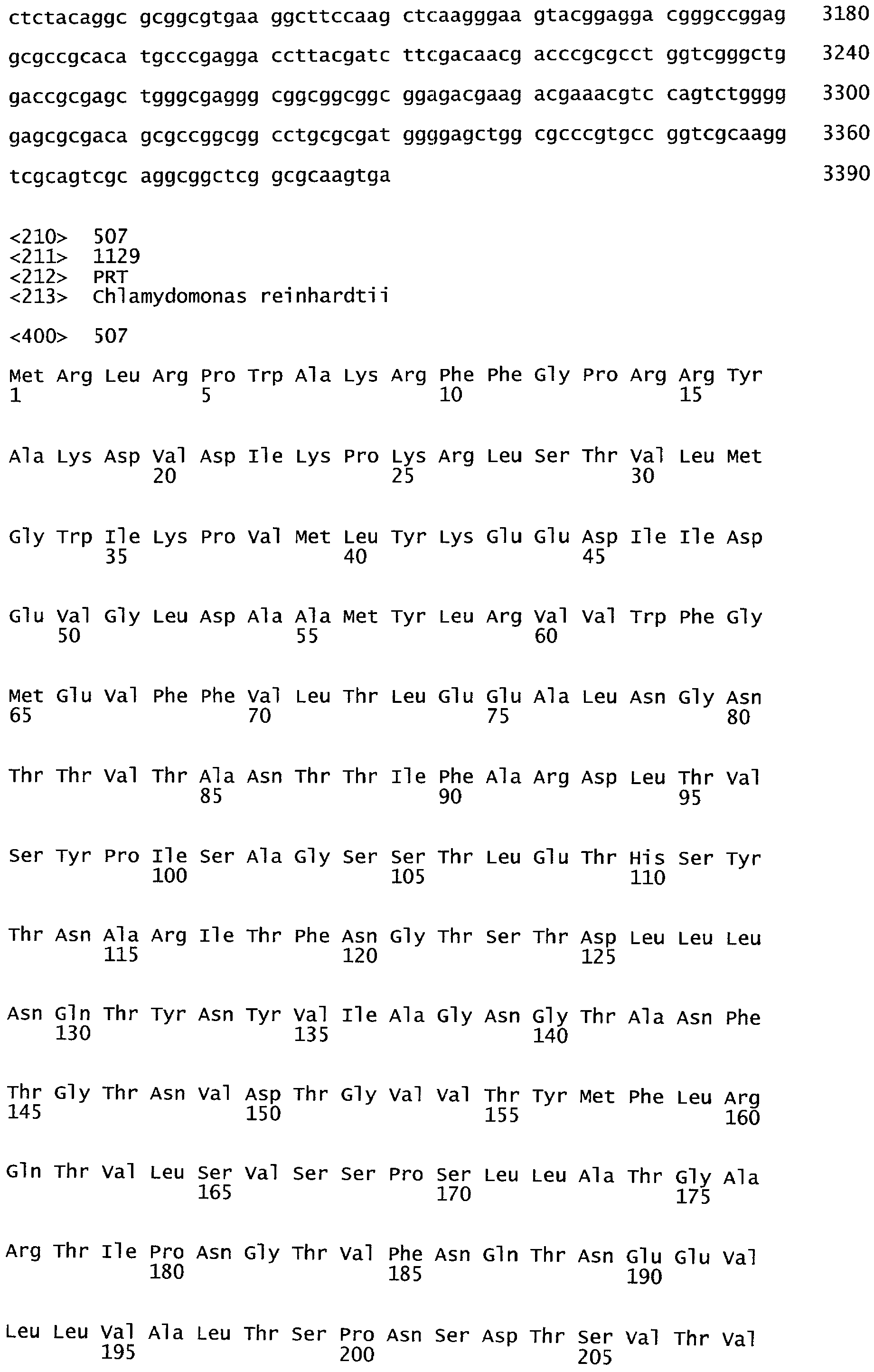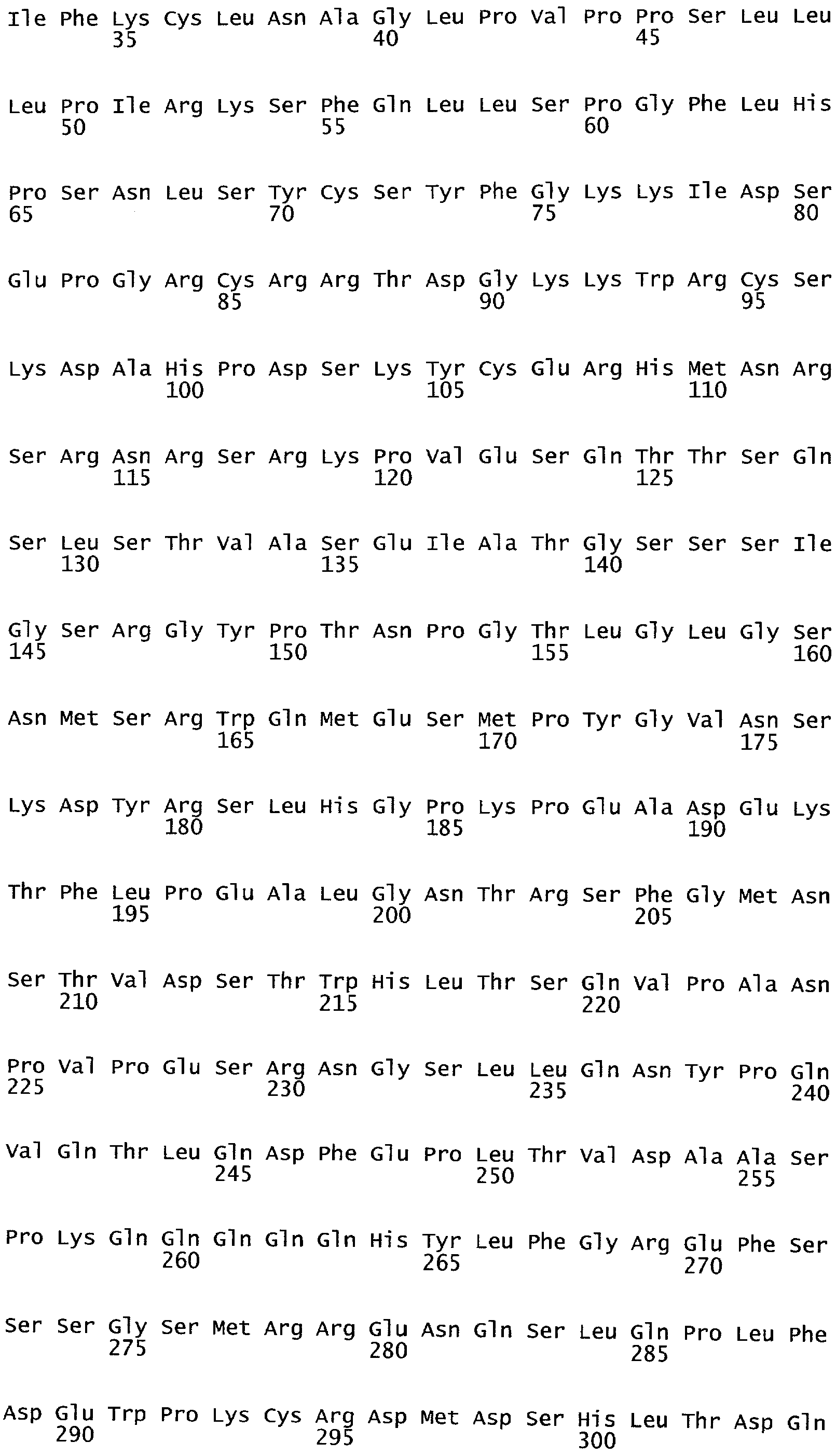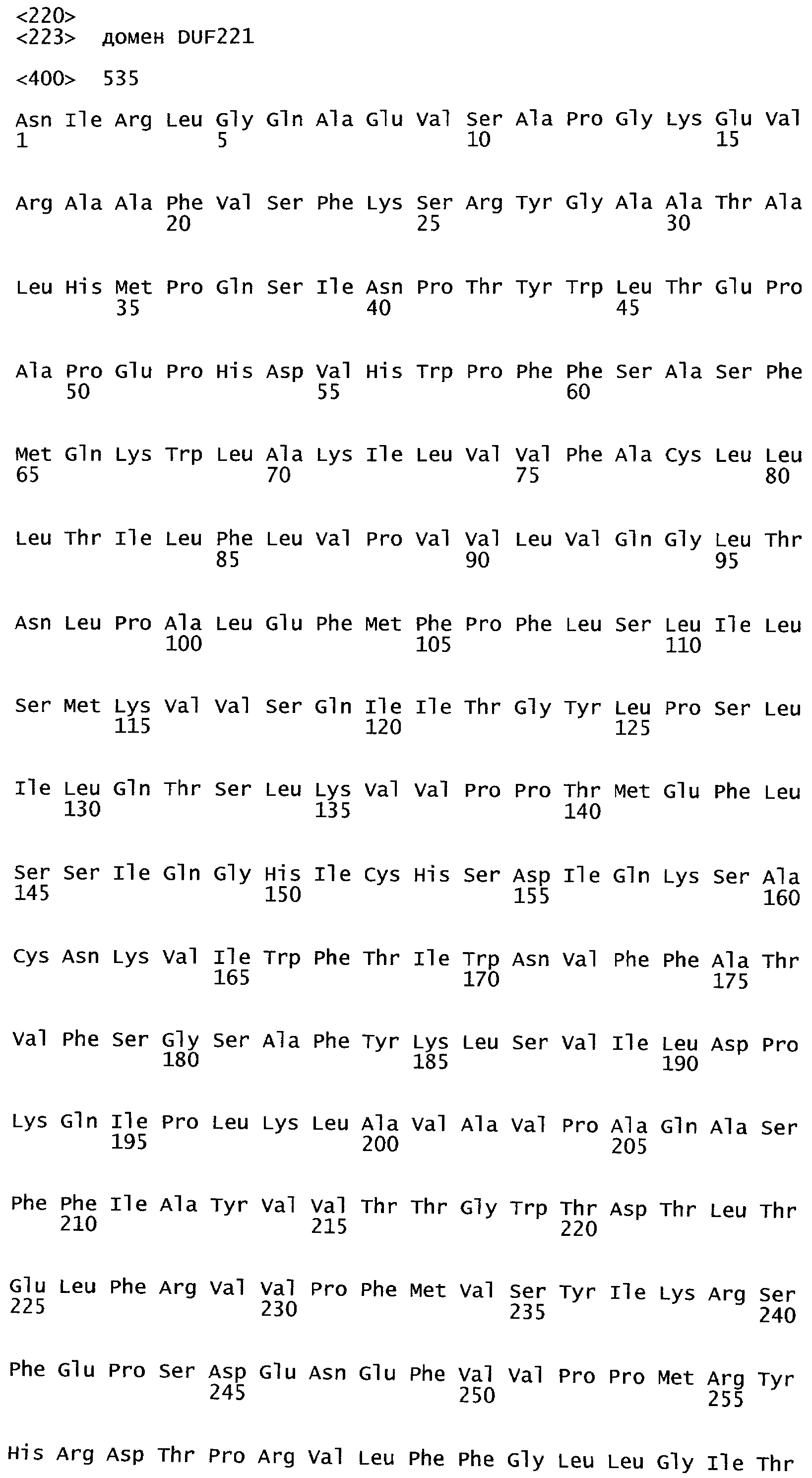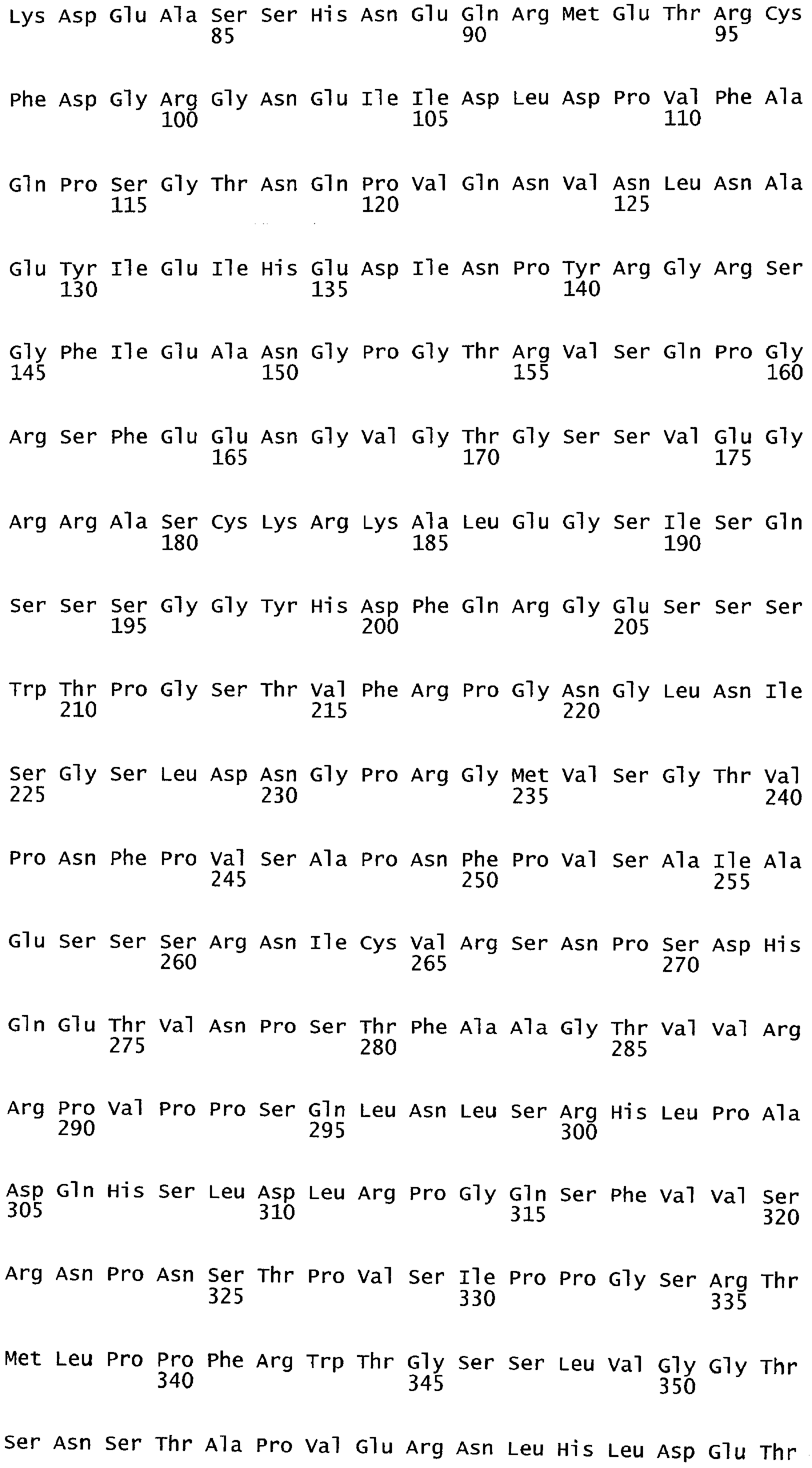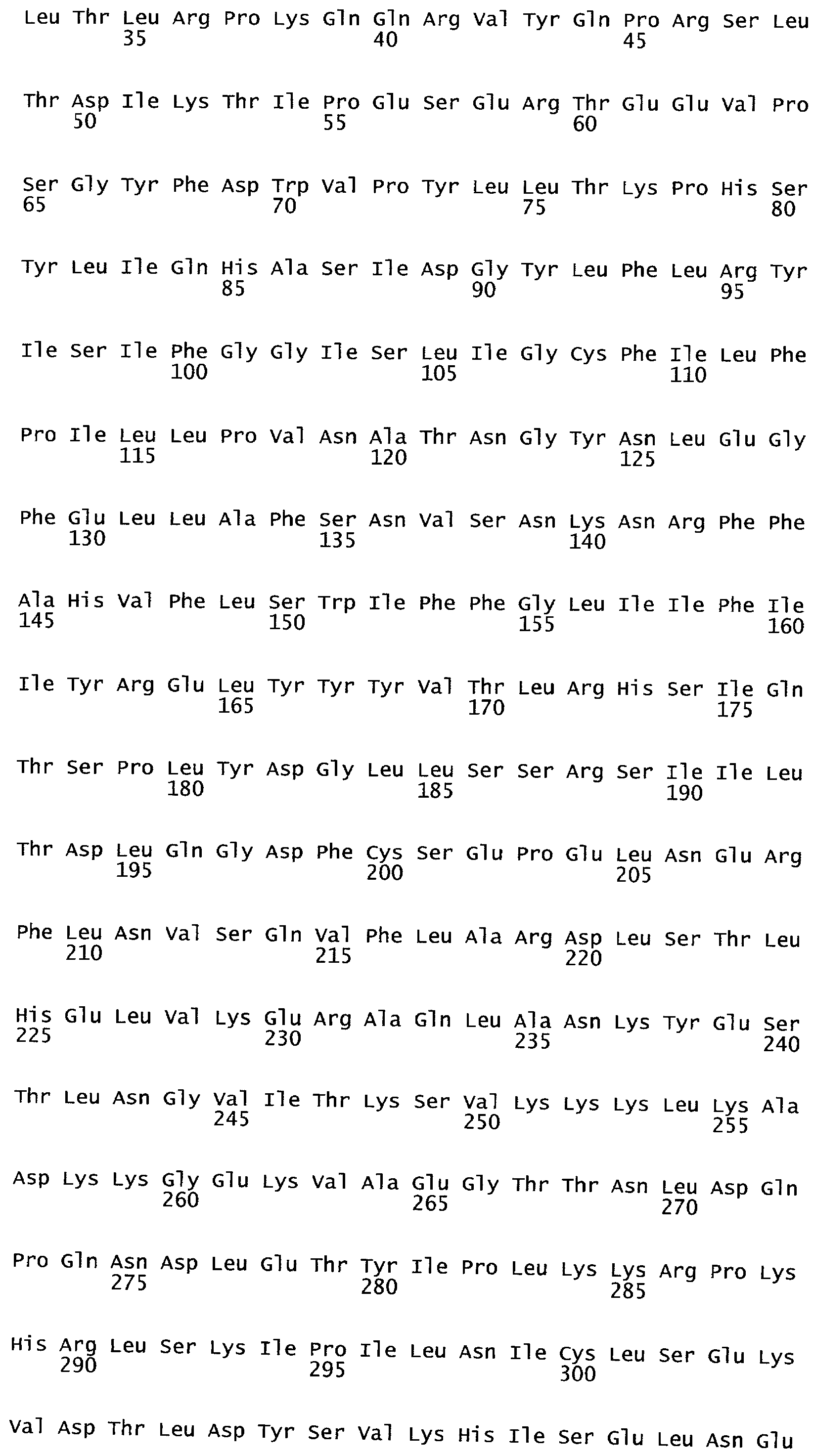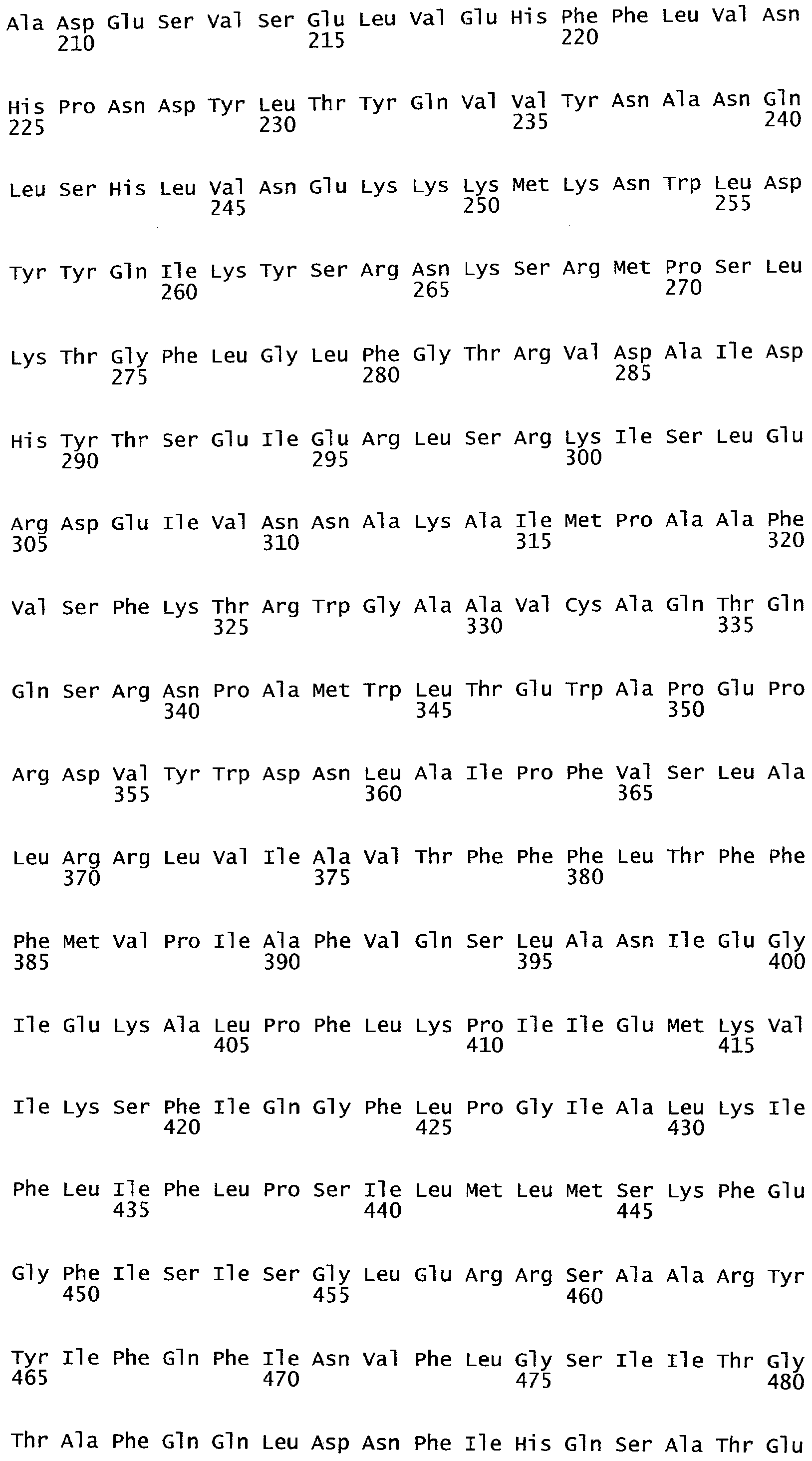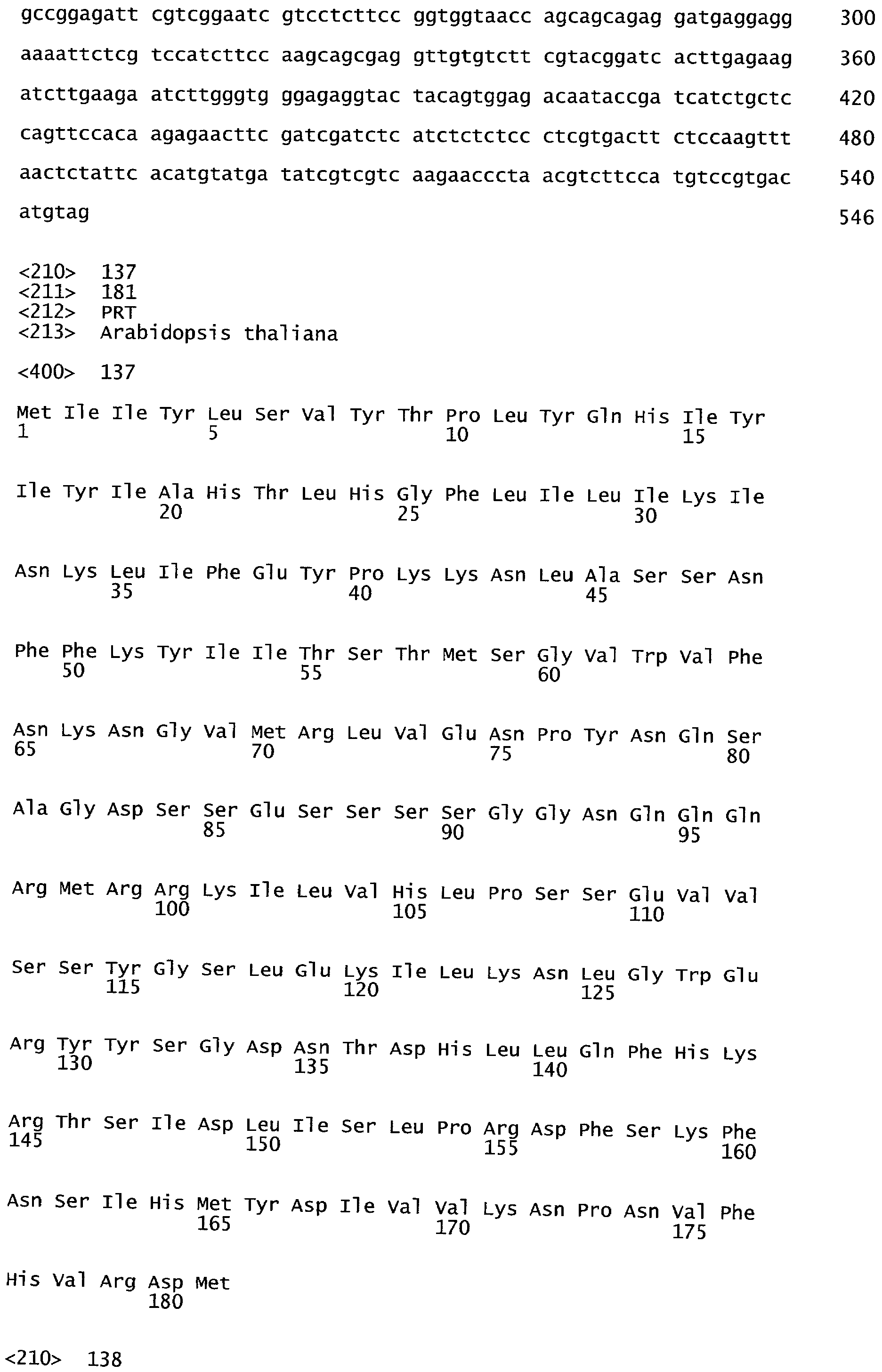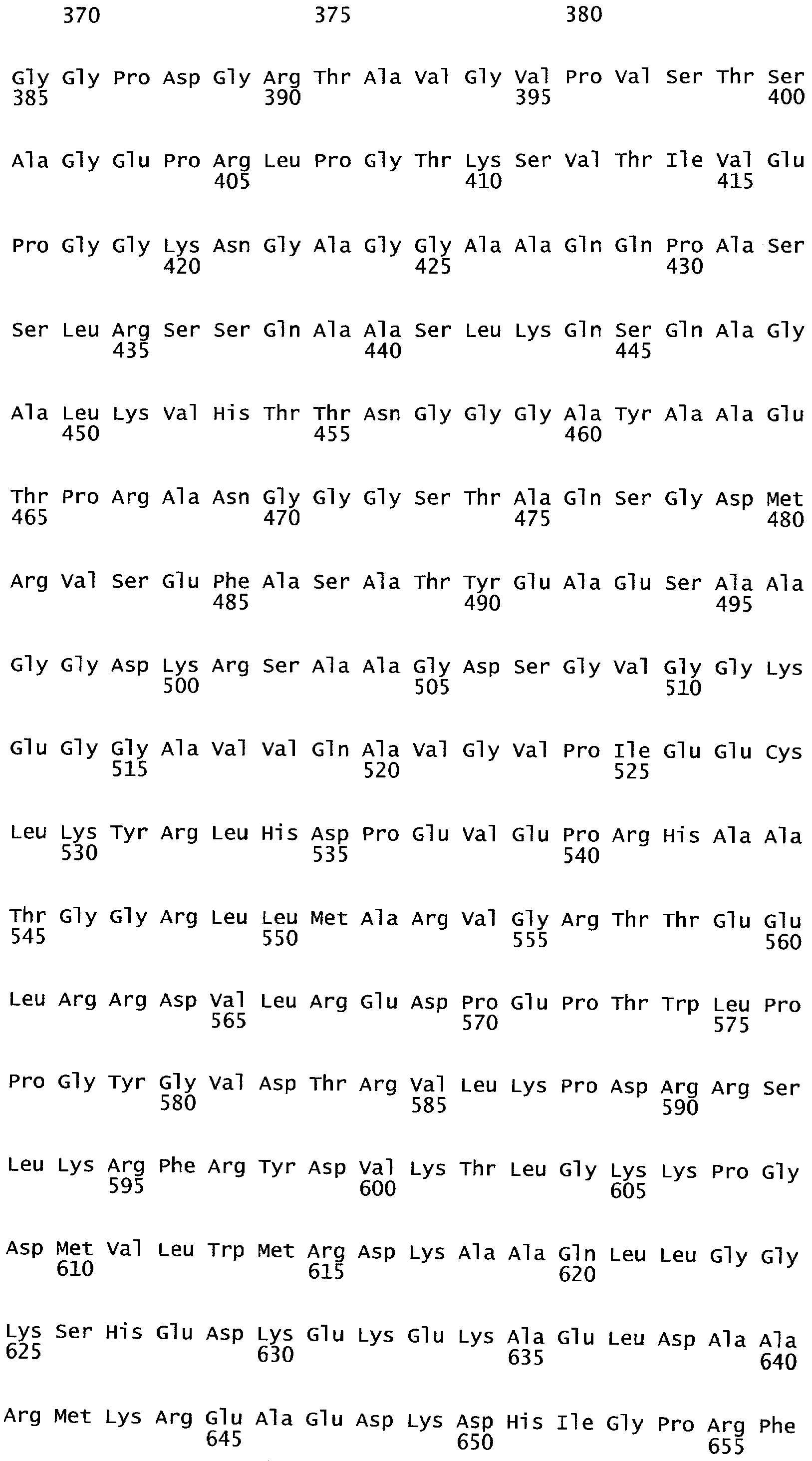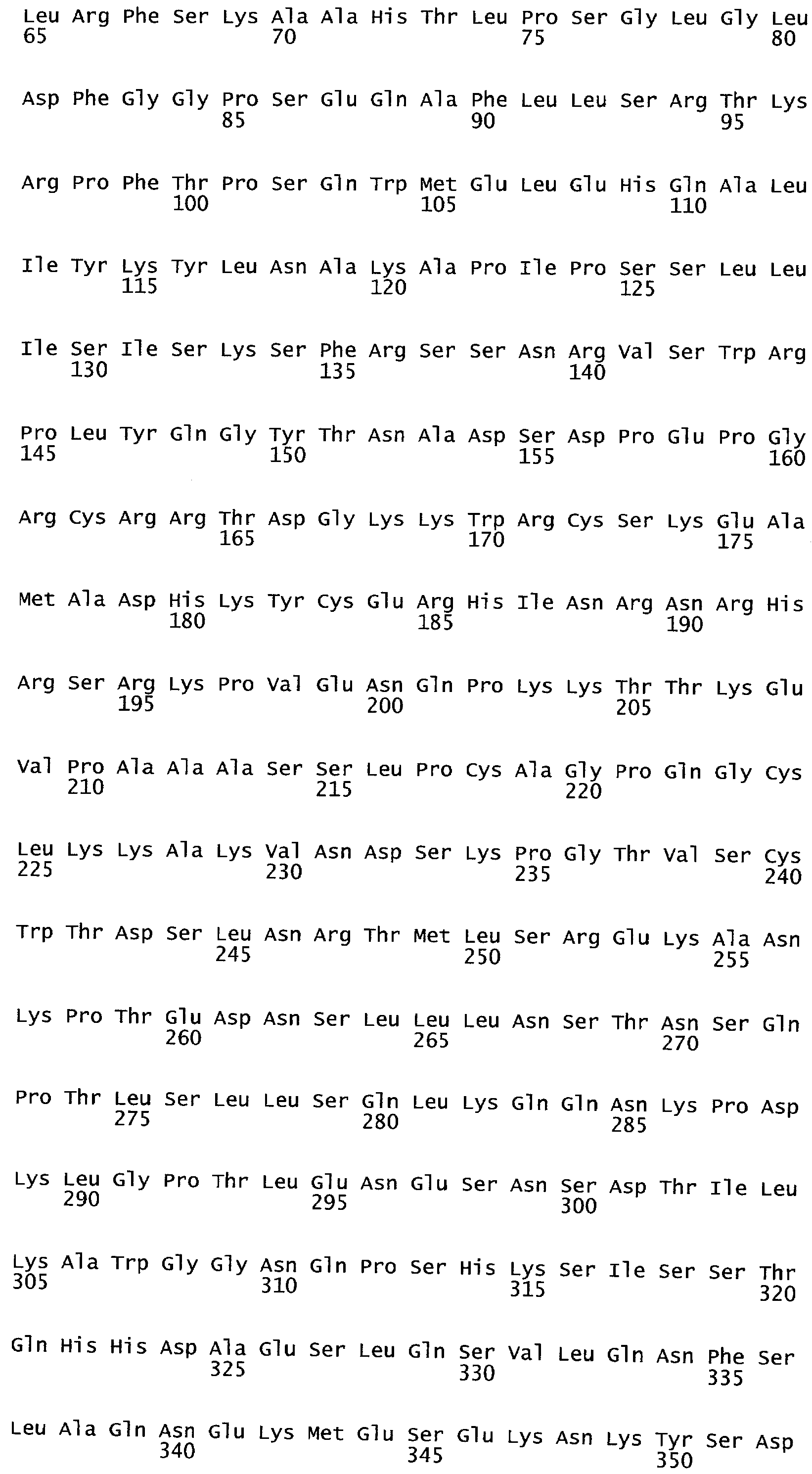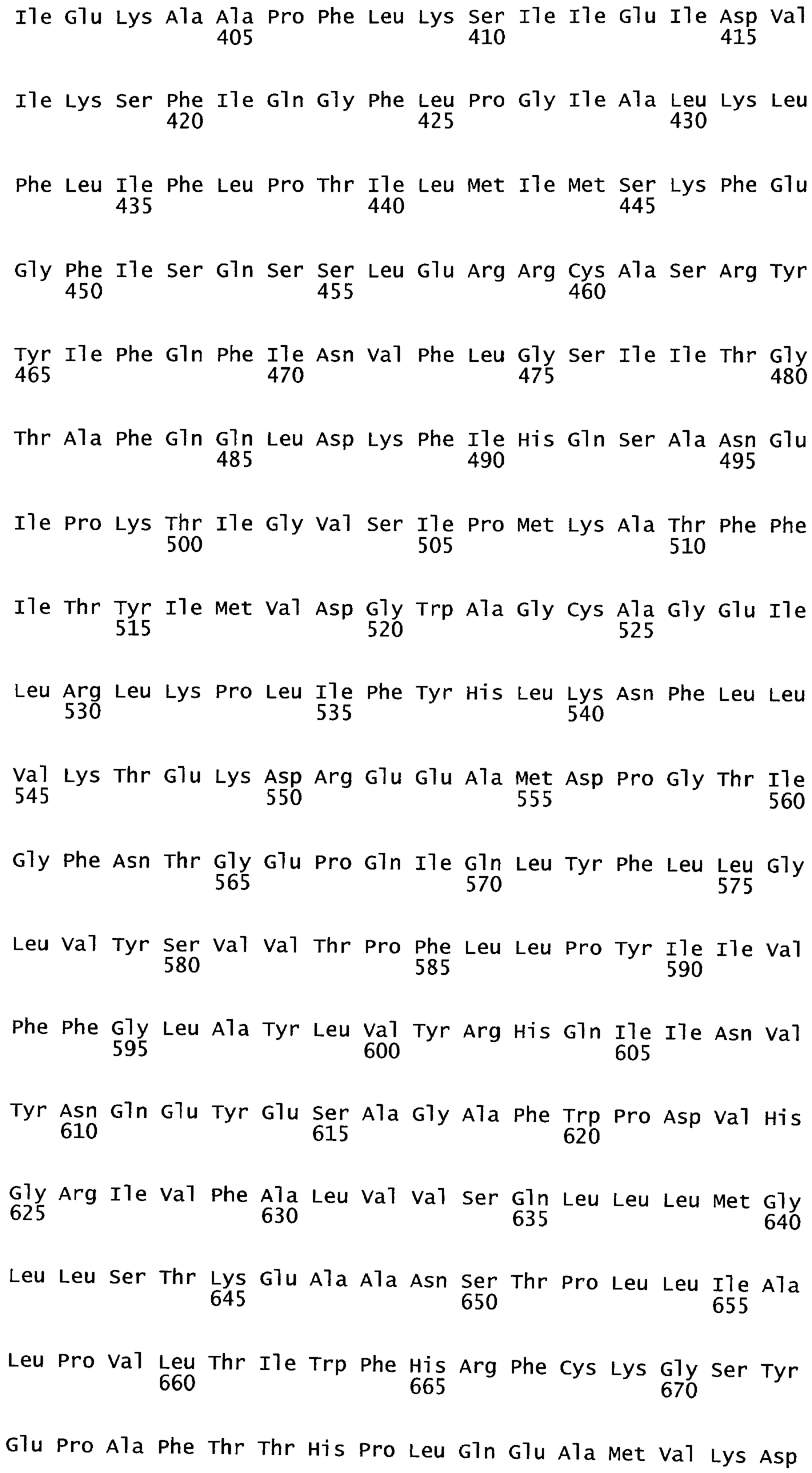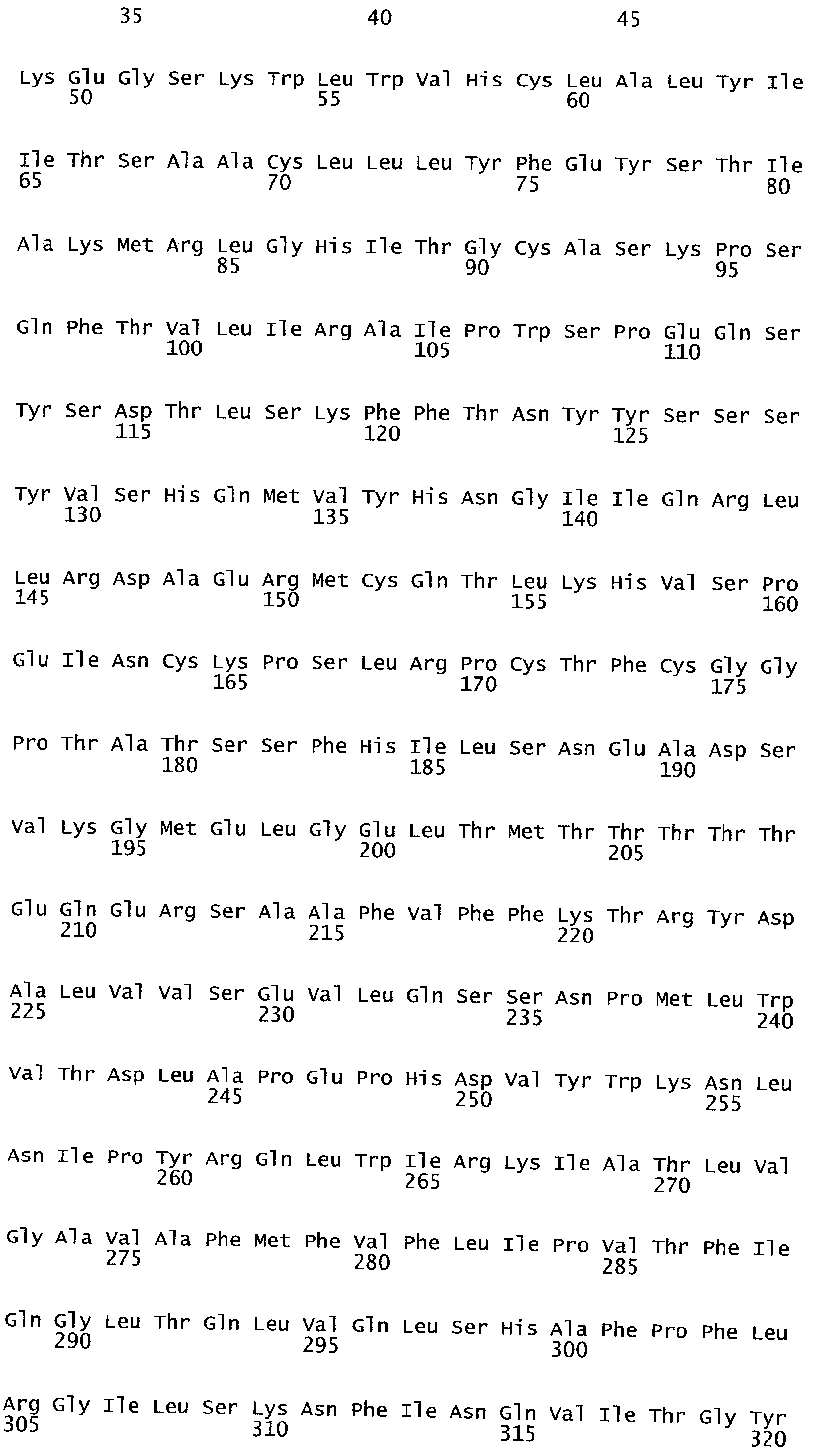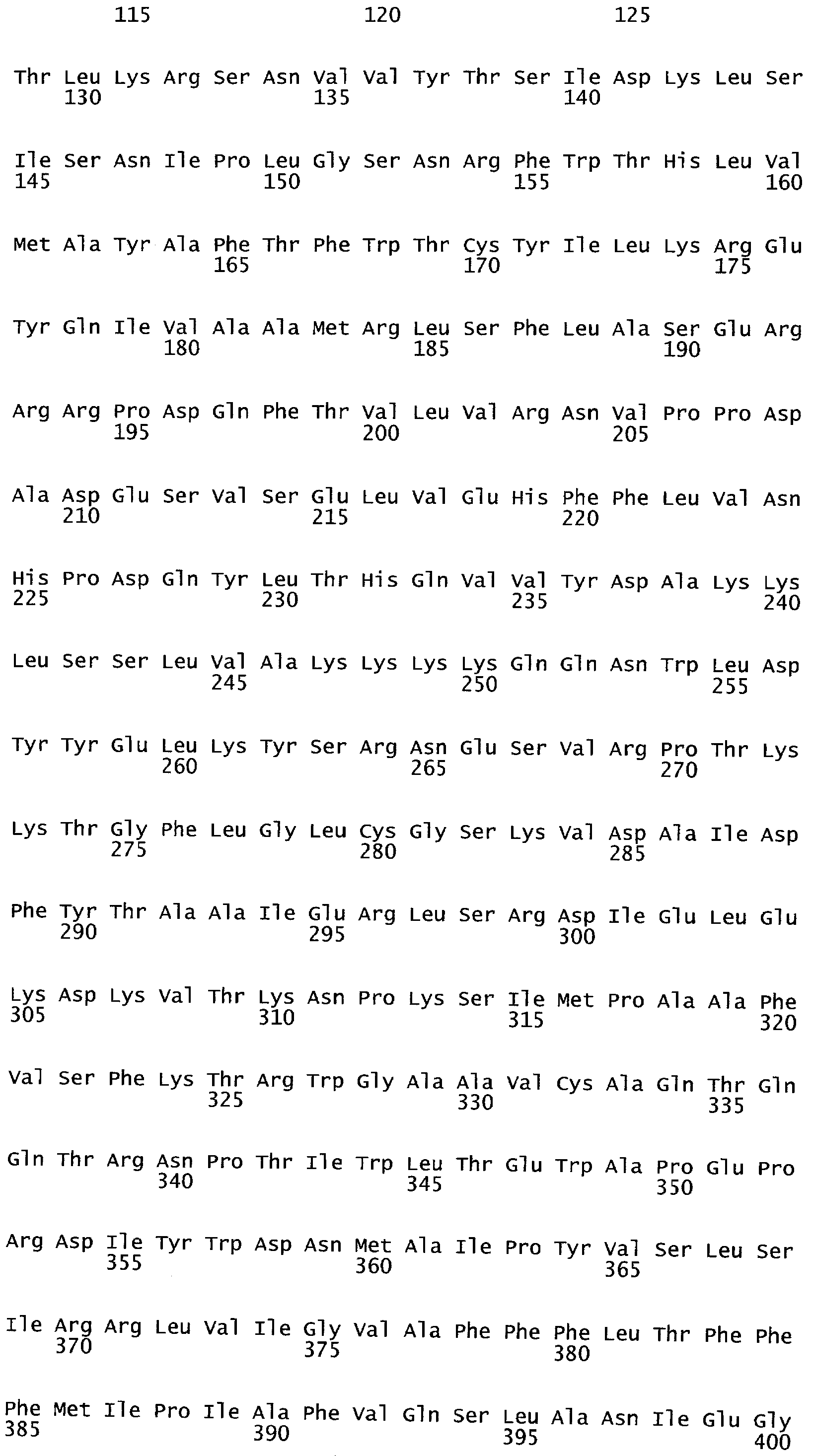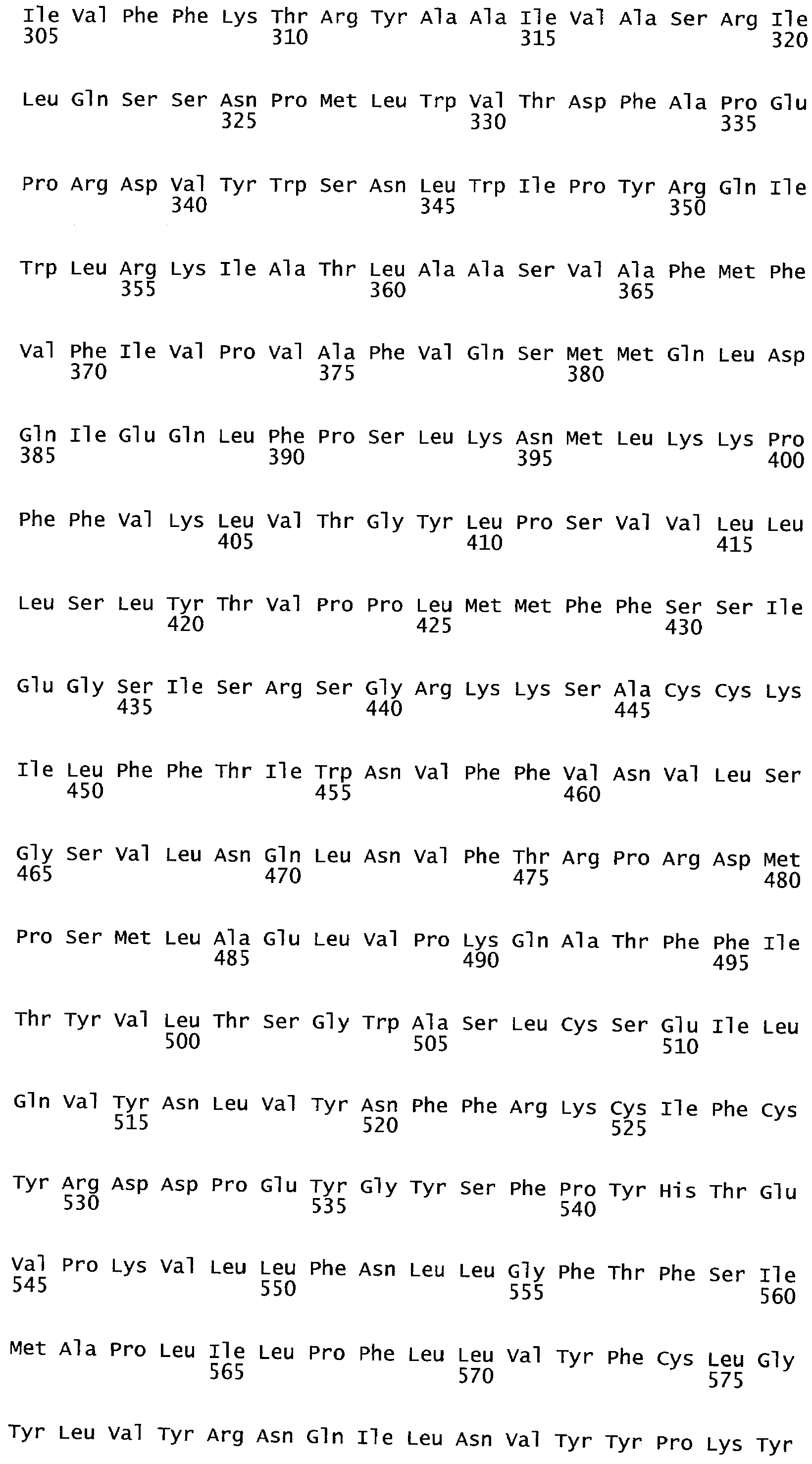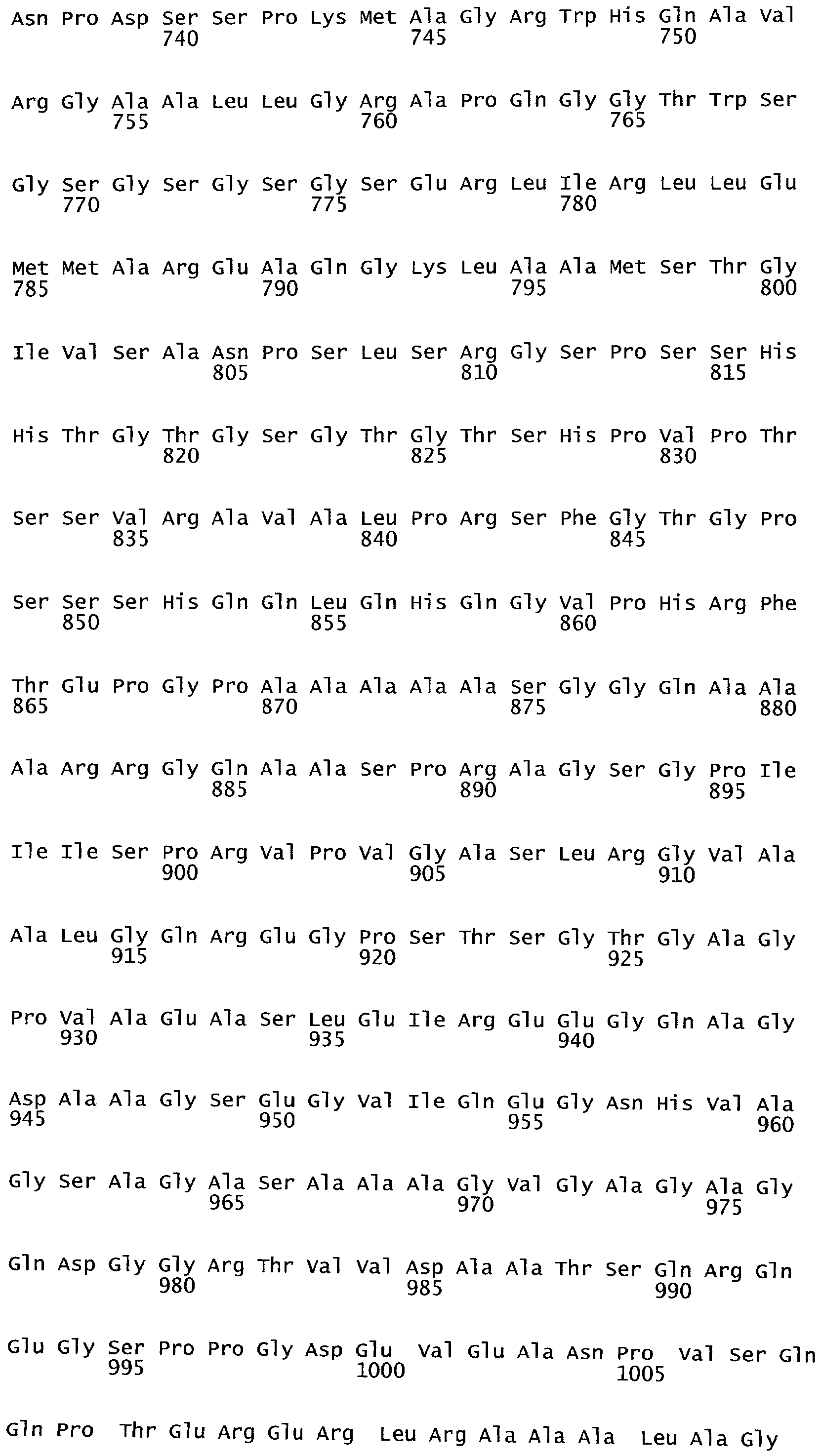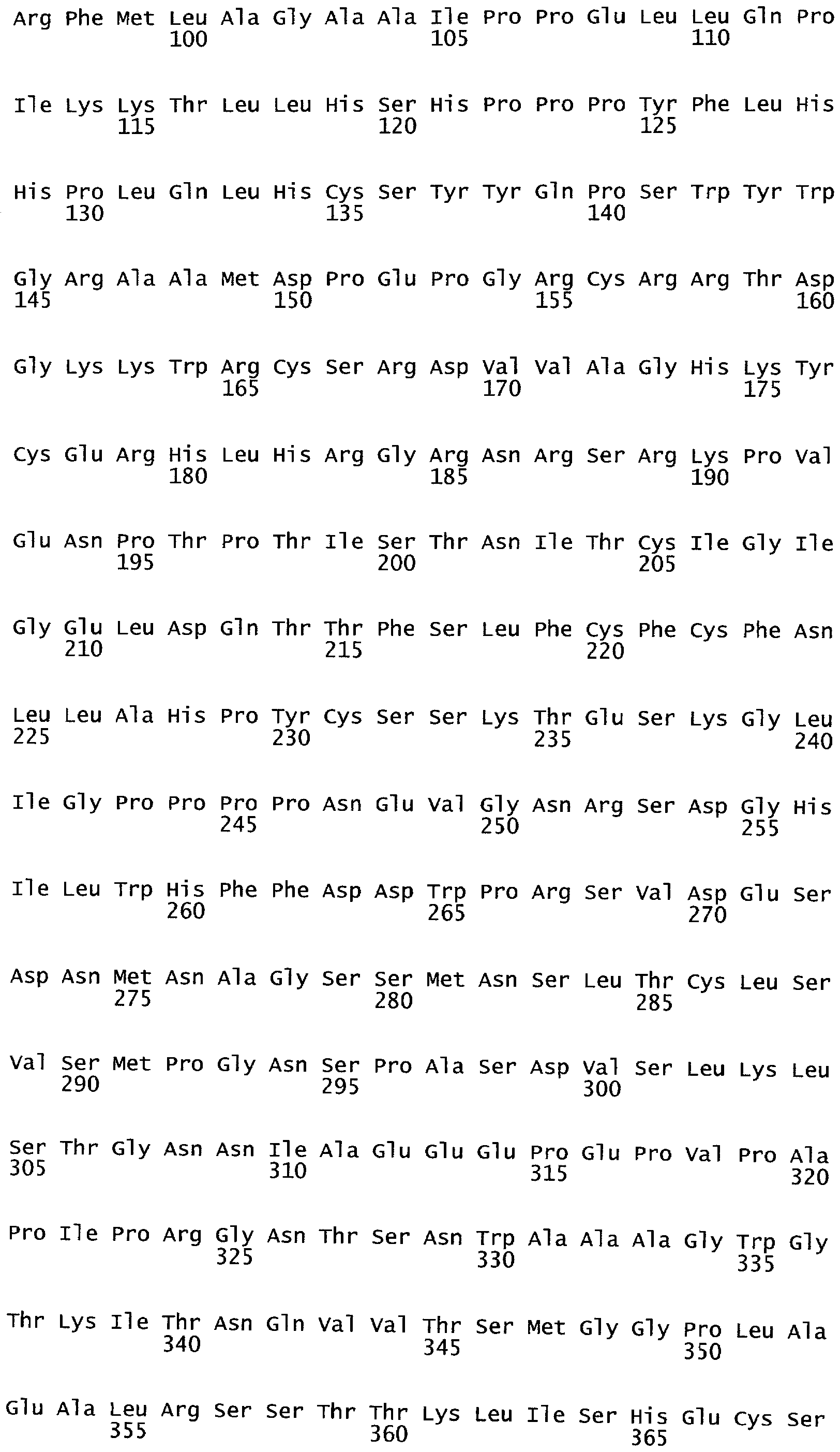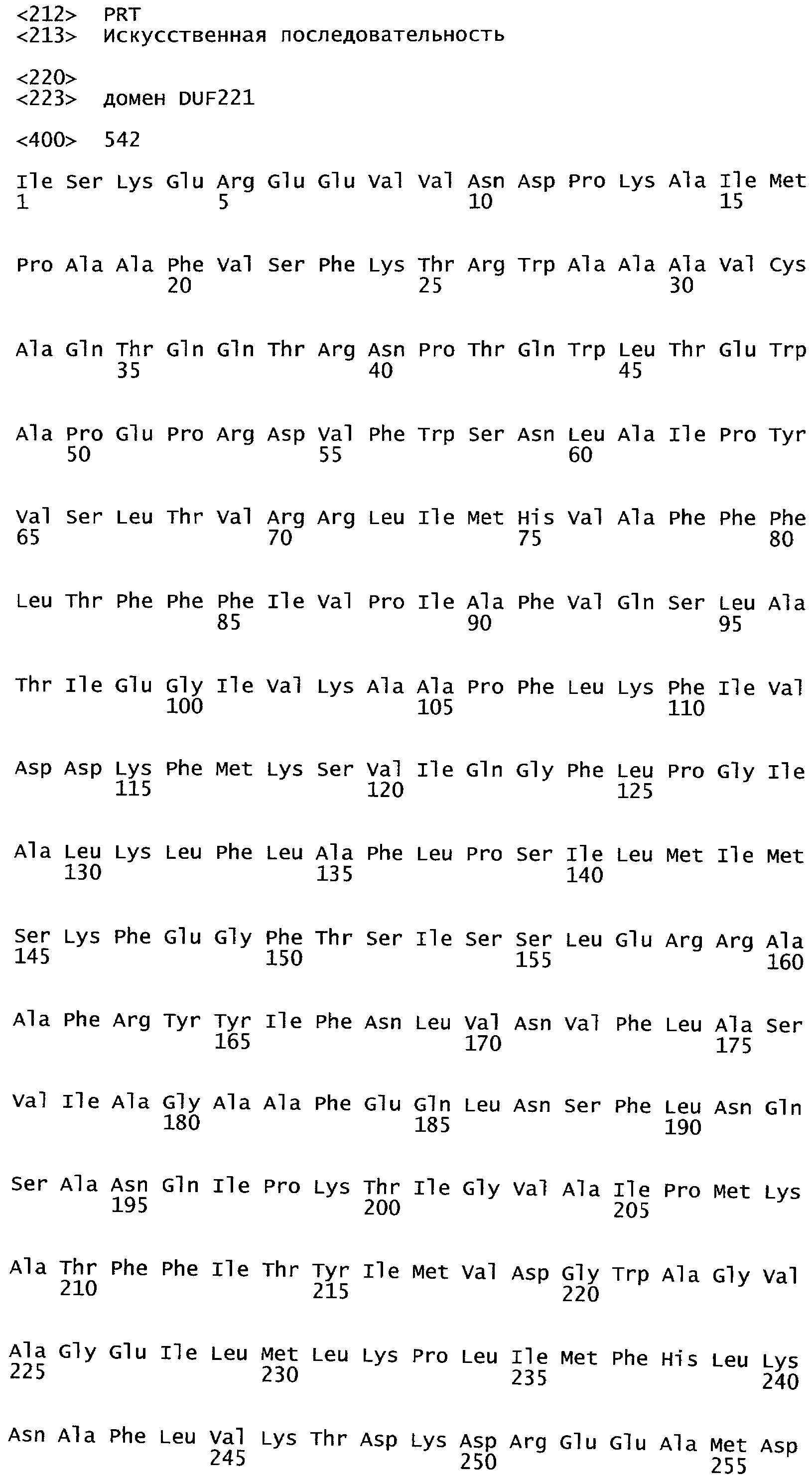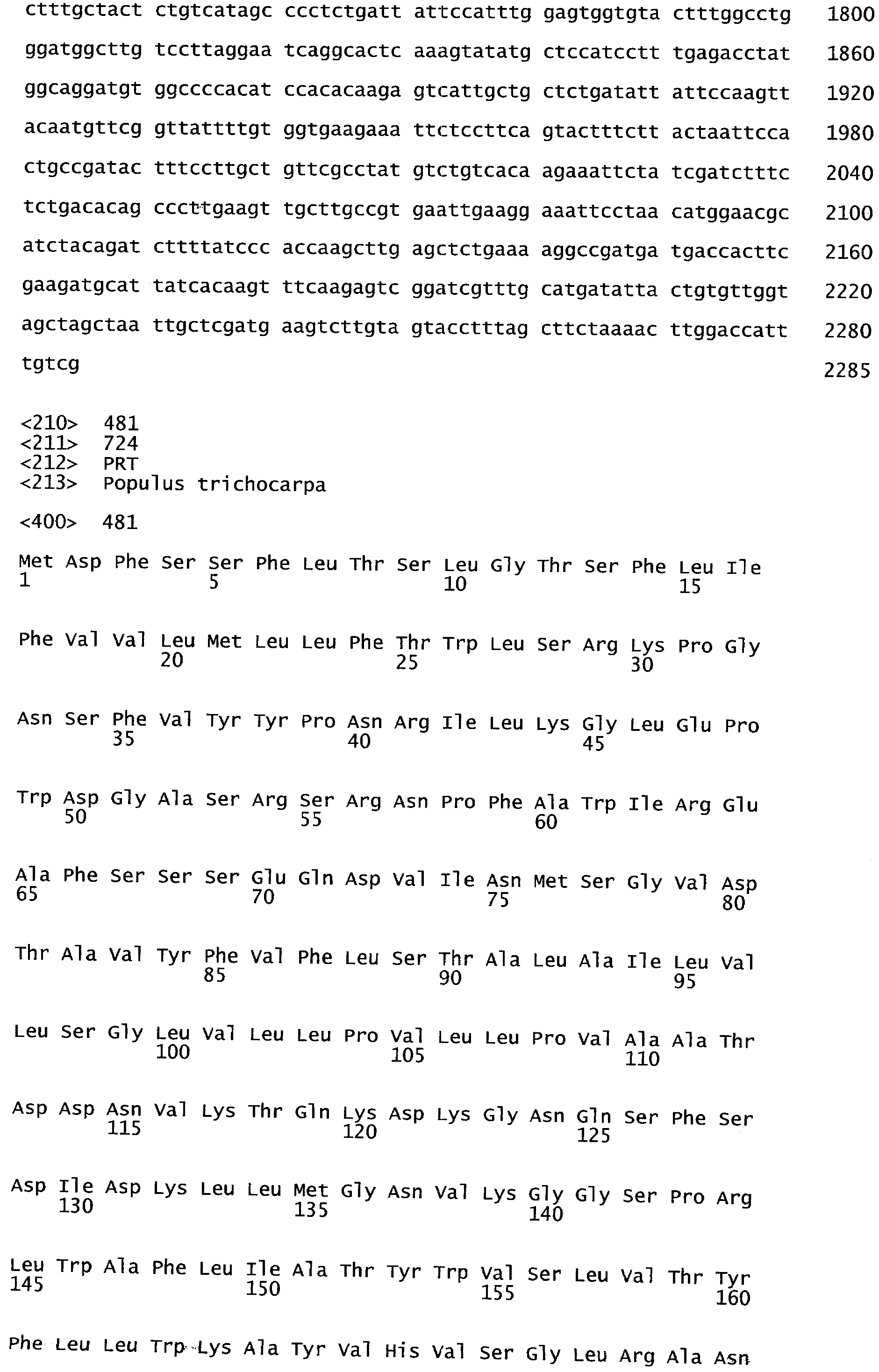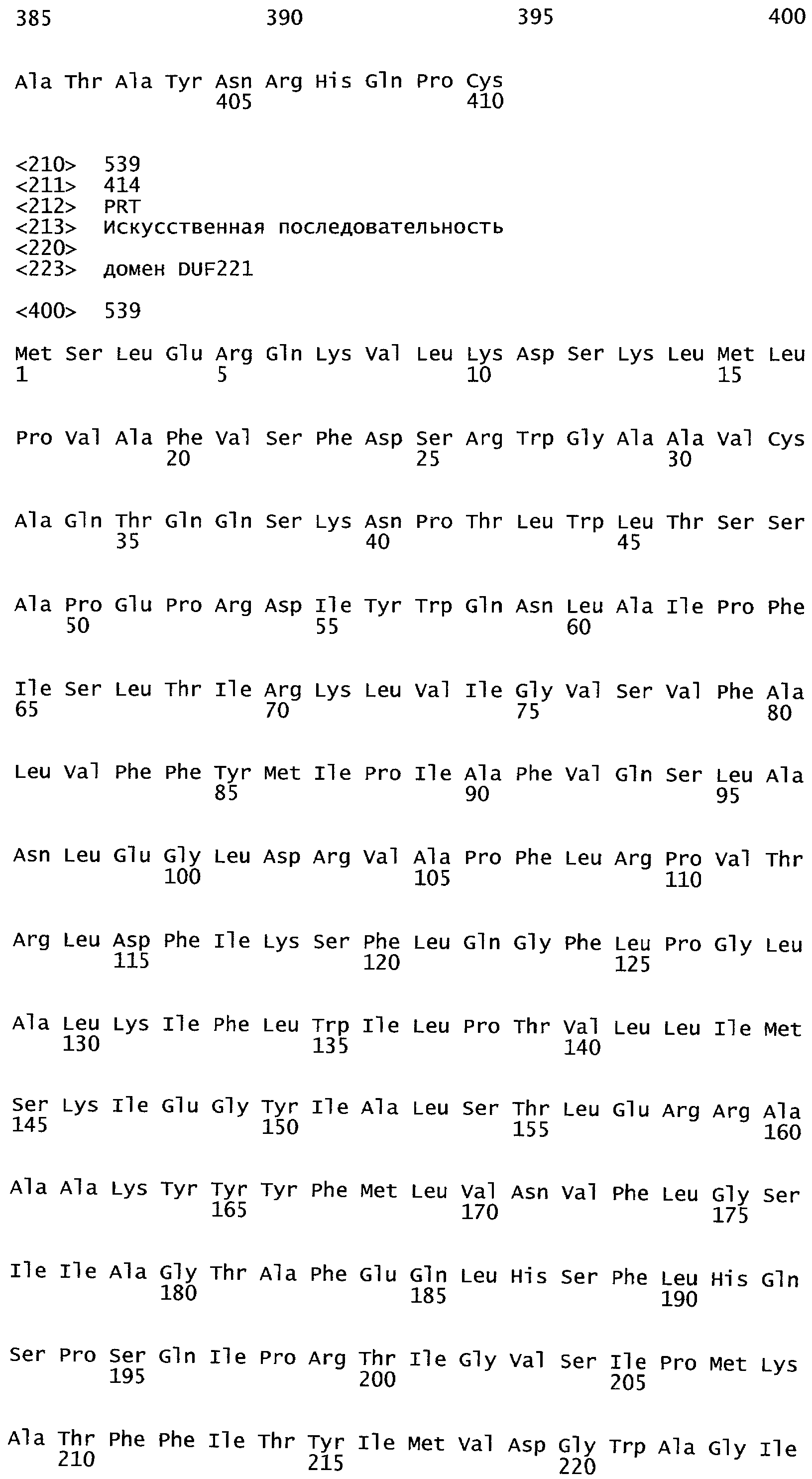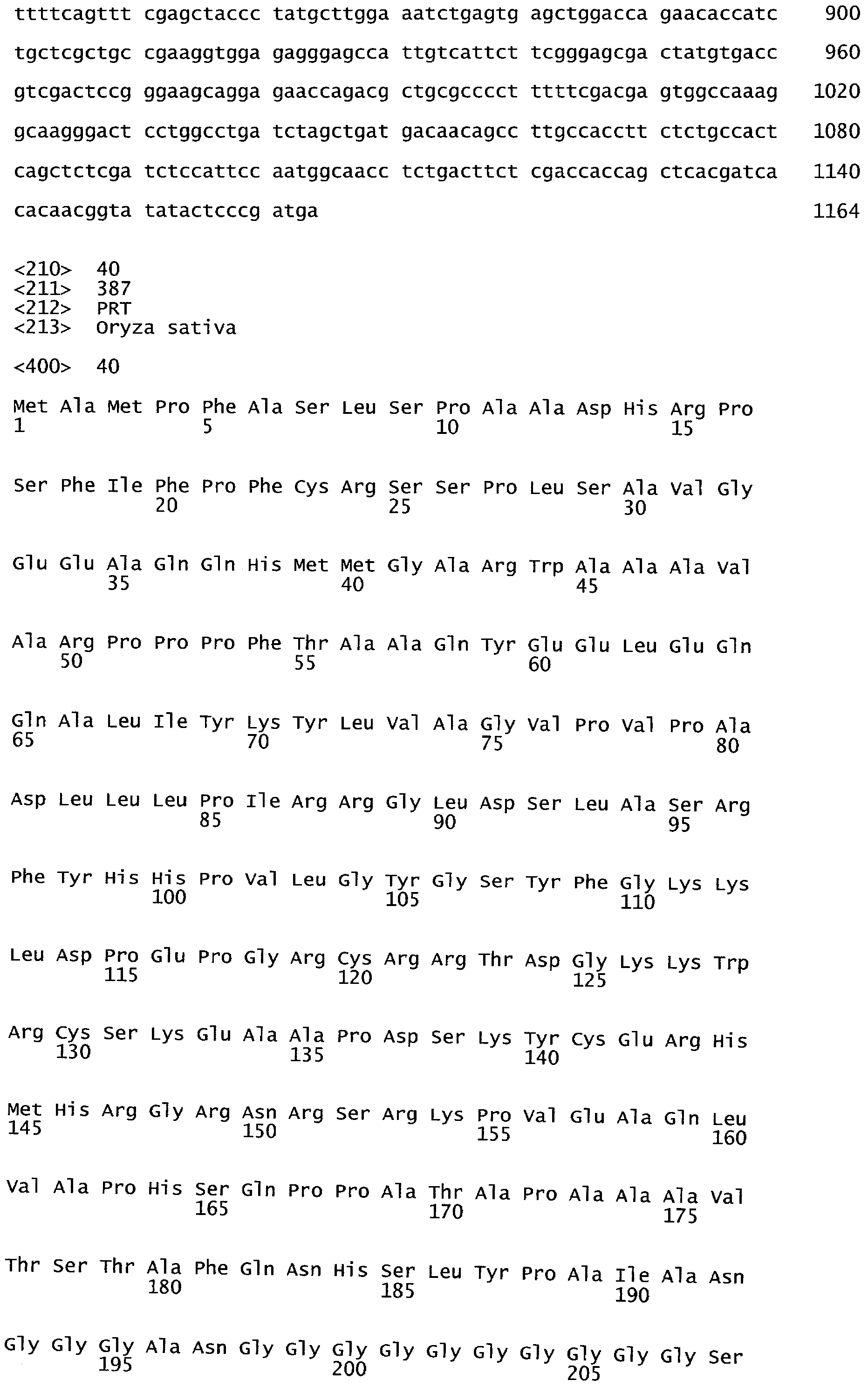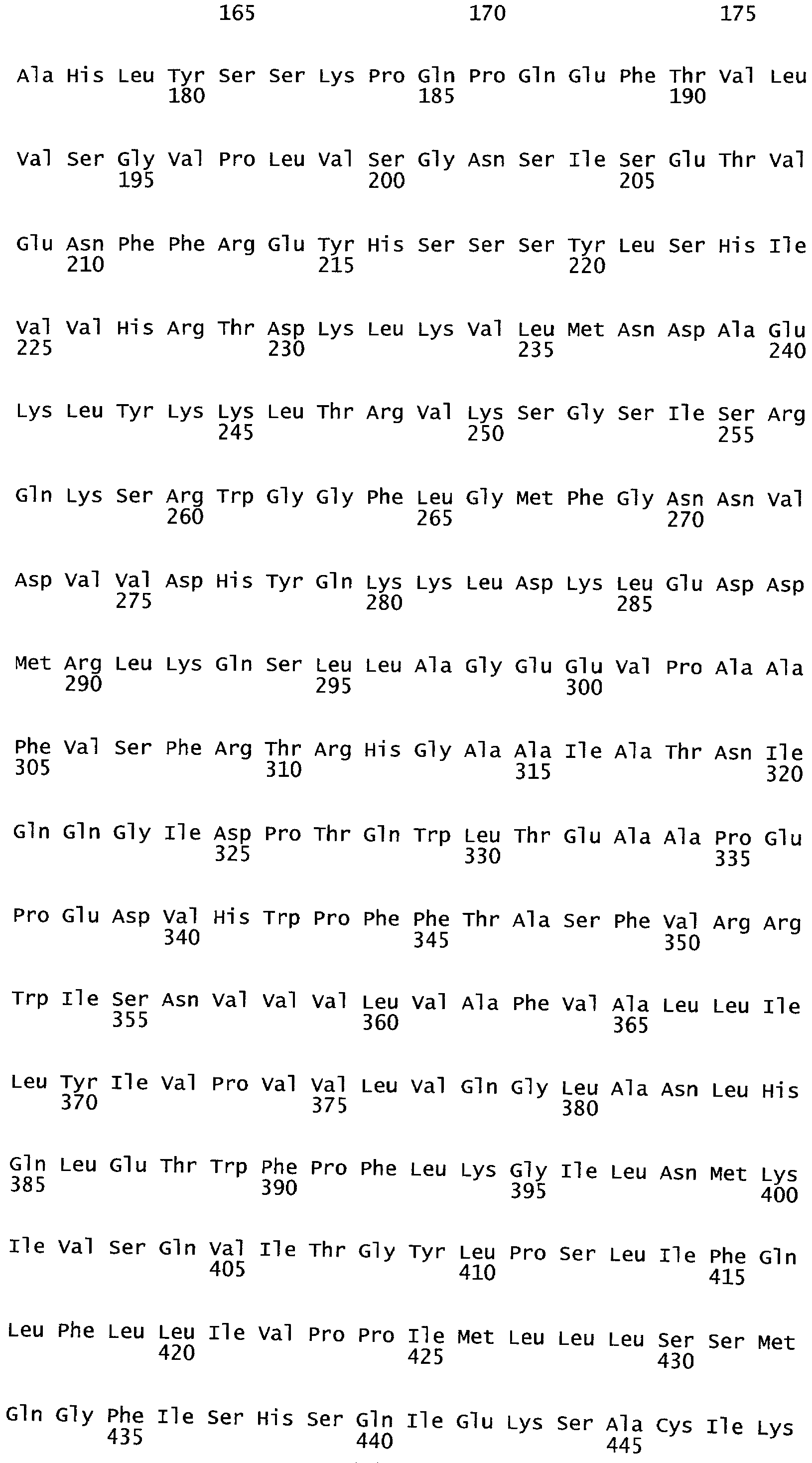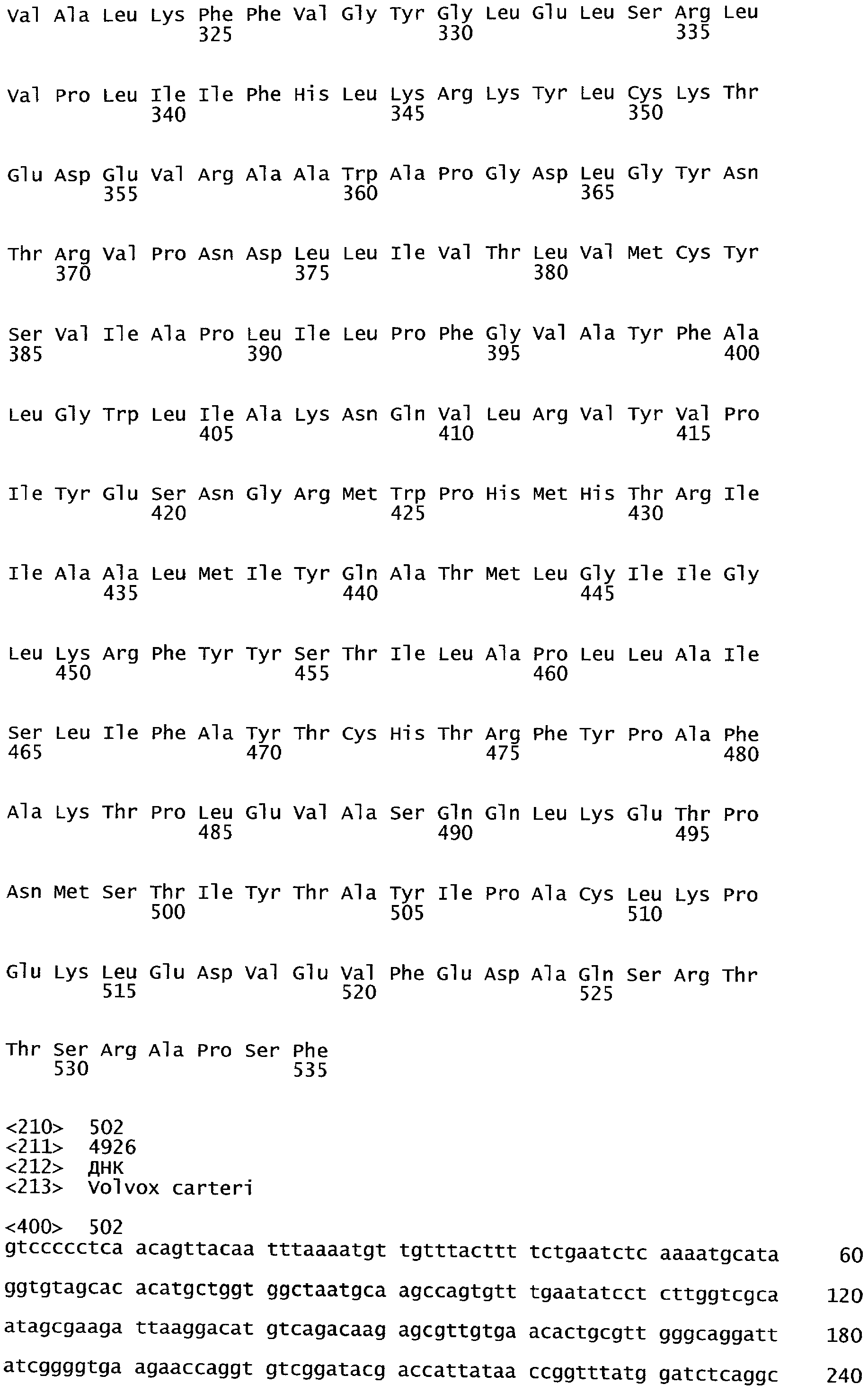РАСТЕНИЯ, ИМЕЮЩИЕ УСИЛЕННЫЕ ПРИЗНАКИ, СВЯЗАННЫЕ С УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ
Вид РИД
Изобретение
Настоящее изобретение относится в целом к области молекулярной биологии и касается способа усиления различных признаков растений, связанных с урожайностью, путем увеличения экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP. Настоящее изобретение также касается растений, имеющих увеличенную экспрессию последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP, при этом растения имеют усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями. Изобретение также предоставляет конструкции, применимые в способах по изобретению.
Постоянно увеличивающееся население мира и истощающийся запас пахотных земель, пригодных для сельского хозяйства, способствуют исследованиям по направлению к увеличению эффективности сельского хозяйства. Обычные средства для улучшения сельскохозяйственных и садовых культур используют методы селекционного разведения для определения растений, имеющих желательные характеристики. Однако такие методы селекционного разведения имеют некоторые недостатки, а именно эти методы обычно являются трудоемкими и приводят к растениям, которые часто содержат гетерогенные генетические компоненты, которые не всегда могут приводить к желательному признаку, переданному от родительских растений. Достижения в молекулярной биологии позволяют человечеству модифицировать зародышевую плазму животных и растений. Генетическая инженерия растений включает в себя выделение и манипуляции с генетическим материалом (обычно в форме ДНК или РНК) и последующее введение этого генетического материала в растение. Такая технология обладает способностью производить сельскохозяйственные культуры или растения, имеющие различные улучшенные экономические, агрономические или садоводческие признаки.
Признаком особого экономического интереса является увеличенная урожайность. Урожайность обычно определяют как измеряемую продукцию экономической ценности из сельскохозяйственной культуры. Она может определяться в терминах количества и/или качества. Урожайность непосредственно зависит от нескольких факторов, например числа и размера органов, структуры растений (например, число побегов), продукции семян, старения листьев и не только. Развитие корней, поглощение питательных веществ, устойчивость к стрессу и ранняя мощность могут также быть важными факторами в определении урожайности. Оптимизация вышеупомянутых факторов, следовательно, может способствовать увеличению урожайности сельскохозяйственных культур.
Урожайность семян является чрезвычайно важным признаком, так как семена многих растений являются важными для питания людей и животных. Сельскохозяйственные культуры, такие как кукуруза, рис, пшеница, канола и соя составляют более половины общего потребления калорий человеком или путем непосредственного употребления самих семян, или путем употребления мясных продуктов, выращенных на переработанных семенах. Они также являются источником сахаров, масел и многих видов метаболитов, используемых в промышленных процессах. Семена содержат зародыш (источник новых побегов и корней) и эндосперм (источник питательных веществ для роста зародыша во время прорастания и начального роста сеянцев). Развитие семени включает много генов и требует переноса метаболитов от корней, листьев и стеблей в растущее семя. Эндосперм, в частности, осуществляет ассимиляцию метаболических предшественников углеводов, жиров и белков и синтезирует их в запасные макромолекулы для наполнения зерна.
Растительная биомасса представляет собой урожай кормовых культур, таких как люцерна, силосная кукуруза и сено. Многие показатели урожая используются для зерновых культур. Важнейшими из них являются оценки размера растения. Размер растения может измеряться многими путями в зависимости от видов и стадии развития, но включает общую сухую массу растений, сухую надземную массу, сырую надземную массу, листовую поверхность, объем стебля, высоту растения, диаметр розетки, длину листа, длину корней, массу корней, число побегов и число листьев. Многие виды сохраняют консервативное соотношение между размером различных частей растения на определенной стадии развития. Эти аллометрические соотношения используются для экстраполяции от одного из этих показателей размера к другому (например, Tittonell et al (2005) Agric Ecosys & Environ 105:213). Размер растения на ранней стадии развития обычно может коррелировать с размером растения, более позднего по развитию. Более крупное растение с большей листовой поверхностью обычно может поглощать больше света и диоксида углерода, чем более маленькое растение, и, следовательно, может, вероятно, набирать более высокую массу в течение того же самого периода (Fasoula & Tollenaar 2005 Maydica 50:39). Это является дополнением к потенциальному продолжению преимущества микросреды или генетического преимущества, которые растение имеет изначально для достижения большего размера. Существует серьезный генетический компонент для размера растения и скорости роста (например, ter Steege et al. (2005) Plant Physiology 139:1078), и таким образом в пределах различных генотипов размер растения в одних условиях окружающей среды, вероятно, коррелирует с размером в других (Hittalmani et al. (2003) Theoretical Applied Genetics 107:679). Таким образом, стандартная окружающая среда используется как показатель для разнообразных и динамических условий окружающей среды, обнаруженных в различных местах и временах у сельскохозяйственных культур на поле.
Другим важным признаком для многих сельскохозяйственных культур является ранняя мощность. Улучшение ранней мощности является важным объектом современных программ селекции риса и для сортов умеренного климата, и для тропических сортов риса. Длинные корни важны для надлежащего закрепления в почве посеянного в воду риса. Там, где рис сеют непосредственно в затопленные поля, и там, где растения должны быстро подниматься через воду, более длинные побеги ассоциированы с мощностью. Когда практикуется рядовой посев, более длинные мезокотили и колеоптили важны для хорошего появления всходов. Способность создать раннюю мощность в растениях имела бы огромное значение в сельском хозяйстве. Например, плохая ранняя мощность является ограничением для введения гибридов кукурузы (Zea mays L.) на основе зародышевой плазмы кукурузного пояса в Атлантической Европе.
Индекс урожайности, отношение урожайности семян к сухой надземной массе, является относительно стабильным при многих условиях окружающей среды, и, следовательно, во многих случаях может быть получена прочная корреляция между размером растения и урожаем зерна (например, Rebetzke et al. (2002) Crop Science 42:739). Эти процессы по существу связаны, поскольку большая часть биомассы зерновых зависит от текущей или накопленной фотосинтетической продуктивности листьев и стебля растения (Gardener et al. (1985) Physiology of Crop Plants. Iowa State University Press, p.68-73). Следовательно, отбор по размеру растения, даже на ранних стадиях развития, используется как индикатор будущей потенциальной урожайности (например, Tittonell et al. (2005) Agric Ecosys & Environ 105:213). При исследовании влияния генетических различий на устойчивость к стрессу, способность стандартизировать свойства почвы, температуру, наличие воды и питательных веществ интенсивность света является важным преимуществом условий окружающей среды в теплицах или камерах для выращивания растений по сравнению с полевыми условиями. Однако искусственные ограничения по урожайности вследствие недостаточного опыления из-за отсутствия ветра или насекомых или недостаточного пространства для роста развитых корней или растительного полога могут ограничивать применение этих контролируемых условий окружающей среды для исследования различий по урожайности. Следовательно, измерения размера растений на ранней стадии развития в стандартизованных условиях в камере для выращивания или теплице являются стандартными приемами для обеспечения показателя потенциальных генетических преимуществ урожайности.
Другой важный признак соответствует улучшенной устойчивости к абиотическому стрессу. Абиотический стресс является основной причиной всемирных потерь сельскохозяйственных культур, уменьшая среднюю урожайность для большинства основных сельскохозяйственных культур более чем на 50% (Wang et al. (2003) Planta 218:1-14). Абиотические стрессы могут вызываться засухой, засолением, экстремальными значениями температур, химической токсичностью, избытком или недостаточностью питательных веществ (макроэлементов и/или микроэлементов), радиационным и окислительным стрессом. Возможность увеличить устойчивость растений к абиотическому стрессу явилась бы значительным экономическим преимуществом для фермеров по всему миру и позволила бы культивирование сельскохозяйственных культур во время неблагоприятных условий и на территориях, где культивирование сельскохозяйственных культур иначе невозможно.
Следовательно, урожайность сельскохозяйственных культур можно увеличить оптимизацией одного из вышеупомянутых факторов.
В зависимости от конечного применения модификация определенных признаков урожайности может являться предпочтительной по сравнению с другими. Например, для таких применений, как выращивание кормовых культур, или производство древесины, или источник биологического топлива, увеличение вегетативных частей растений может быть желательным, и для таких применений, как производство муки, крахмала или масла, увеличение параметров семян может быть особенно желательным. Даже среди параметров семян некоторые могут быть предпочтительнее, чем другие в зависимости от применения. Различные механизмы могут делать вклад в увеличение урожайности семян, а именно в форме увеличенного размера семян или увеличенного числа семян.
Один из подходов к усилению признаков, связанных с урожайностью (урожайность семян и/или биомассы), у растений может осуществляться через модификацию внутренних механизмов роста растения, таких как клеточный цикл или различные пути передачи сигналов, вовлеченные в рост растений или в защитные механизмы.
В настоящее время было обнаружено, что различные признаки, связанные с урожайностью семян, могут усиливаться в растениях по сравнению с контрольными растениями путем увеличения экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид фактора, регулирующего рост (GRF). Усиленные признаки, связанные с урожайностью, содержат один или более из: увеличенная ранняя мощность, увеличенная надземная биомасса, увеличенная общая урожайность семян на растение, увеличенный уровень наполнения семян, увеличенный индекс урожайности и увеличенная масса тысячи зерен.
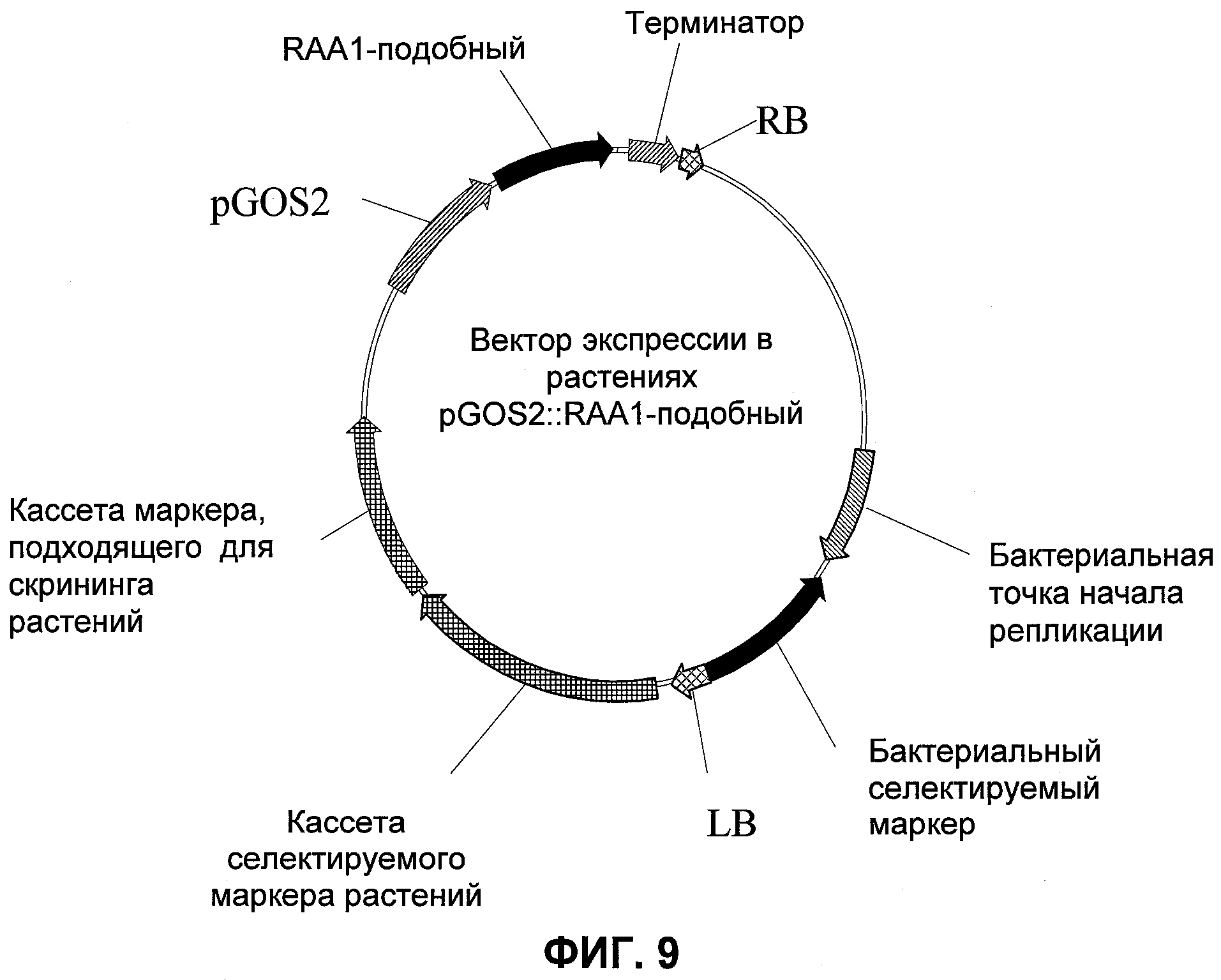
В настоящее время было обнаружено, что различные параметры роста могут быть улучшены у растений путем модуляции экспрессии в растении нуклеиновой кислоты, кодирующей RAA1-подобный (ассоциированный со строением корня, 1) полипептид в растении.
В настоящее время было обнаружено, что различные параметры роста, в частности увеличенная устойчивость к абиотическому стрессу, могут быть улучшены у растений модуляцией экспрессии в растении нуклеиновой кислоты, кодирующей белок регулятор урожайности семян (SYR).
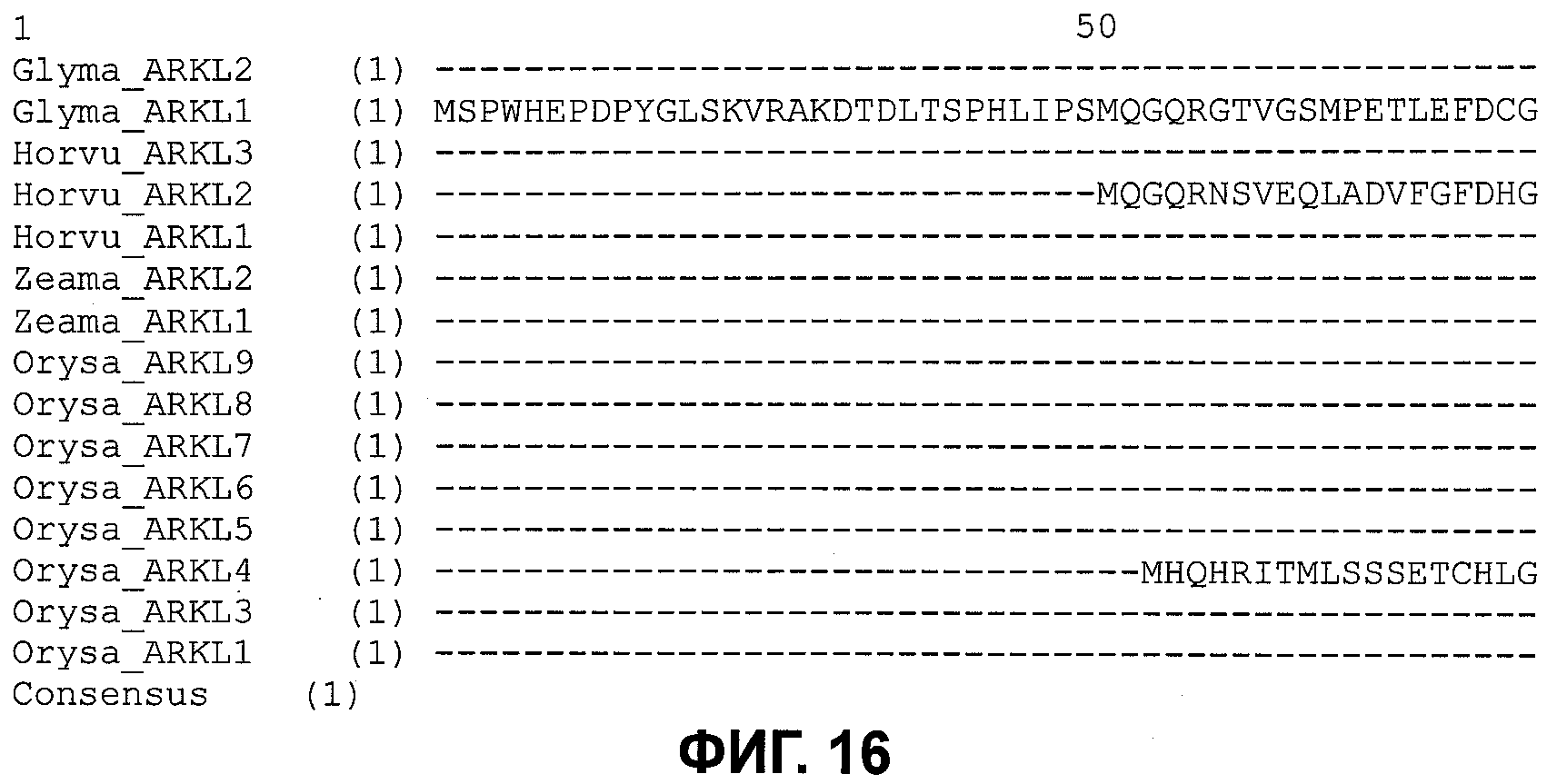
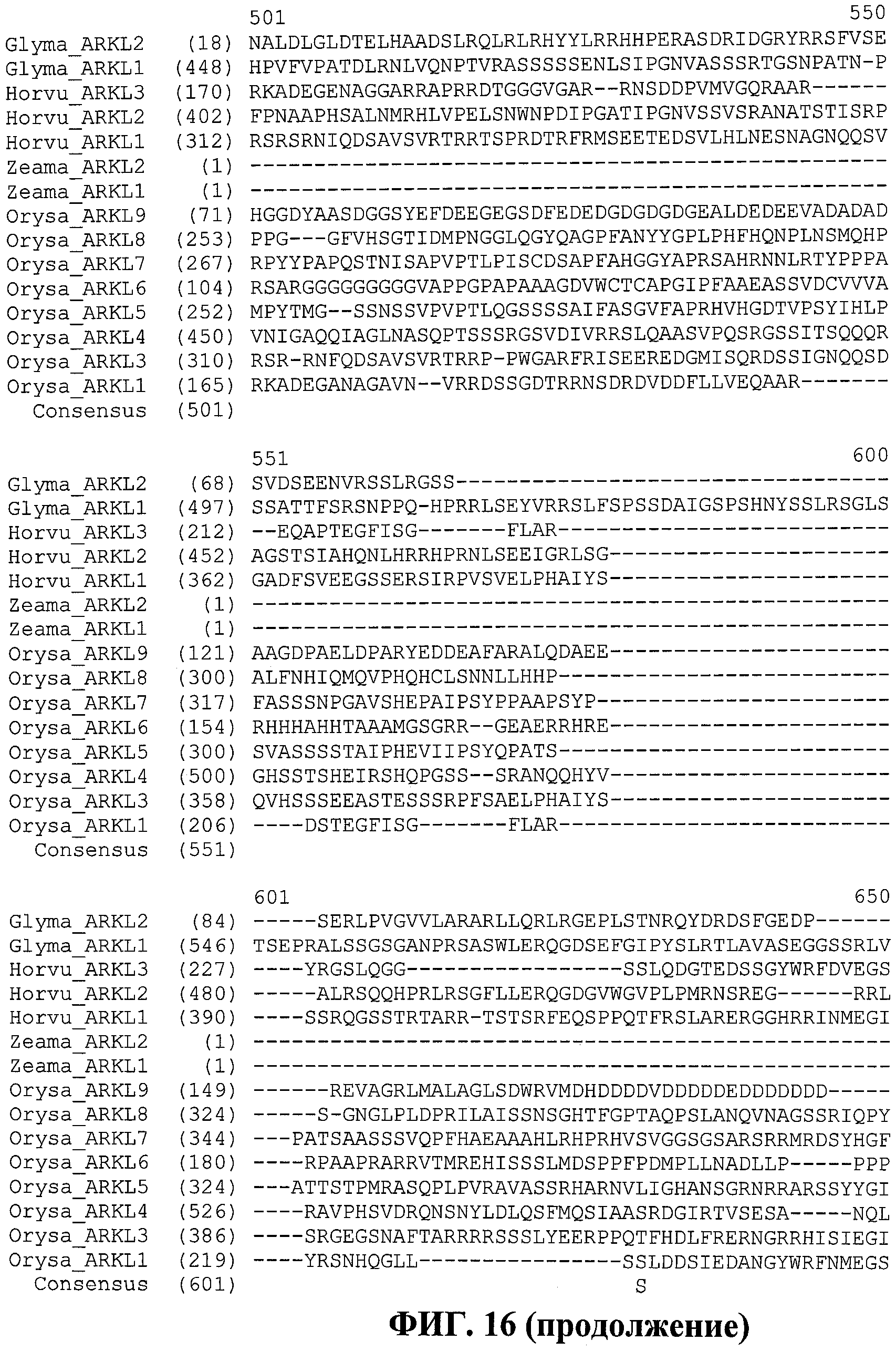
В настоящее время было обнаружено, что различные признаки, связанные с урожайностью семян, могут быть улучшены у растений модуляцией экспрессии в растении нуклеиновой кислоты, кодирующей ARKL (ARADIA подобный) полипептид в растении.
В настоящее время было обнаружено, что различные признаки, связанные с урожайностью семян, могут быть улучшены у растений модуляцией экспрессии в растении нуклеиновой кислоты, кодирующей YTP (трансмембранный белок урожайности) в растении.
Предшествующий уровень техники
ДНК-связывающие белки представляют собой белки, которые содержат любой из многочисленных ДНК-связывающих доменов и таким образом имеют специфическое или общее сродство к ДНК. ДНК-связывающие белки включают, например, факторы транскрипции, которые модулируют процесс транскрипции, нуклеазы, которые расщепляют молекулы ДНК, и гистоны, которые вовлечены в упаковку ДНК в ядре клетки.
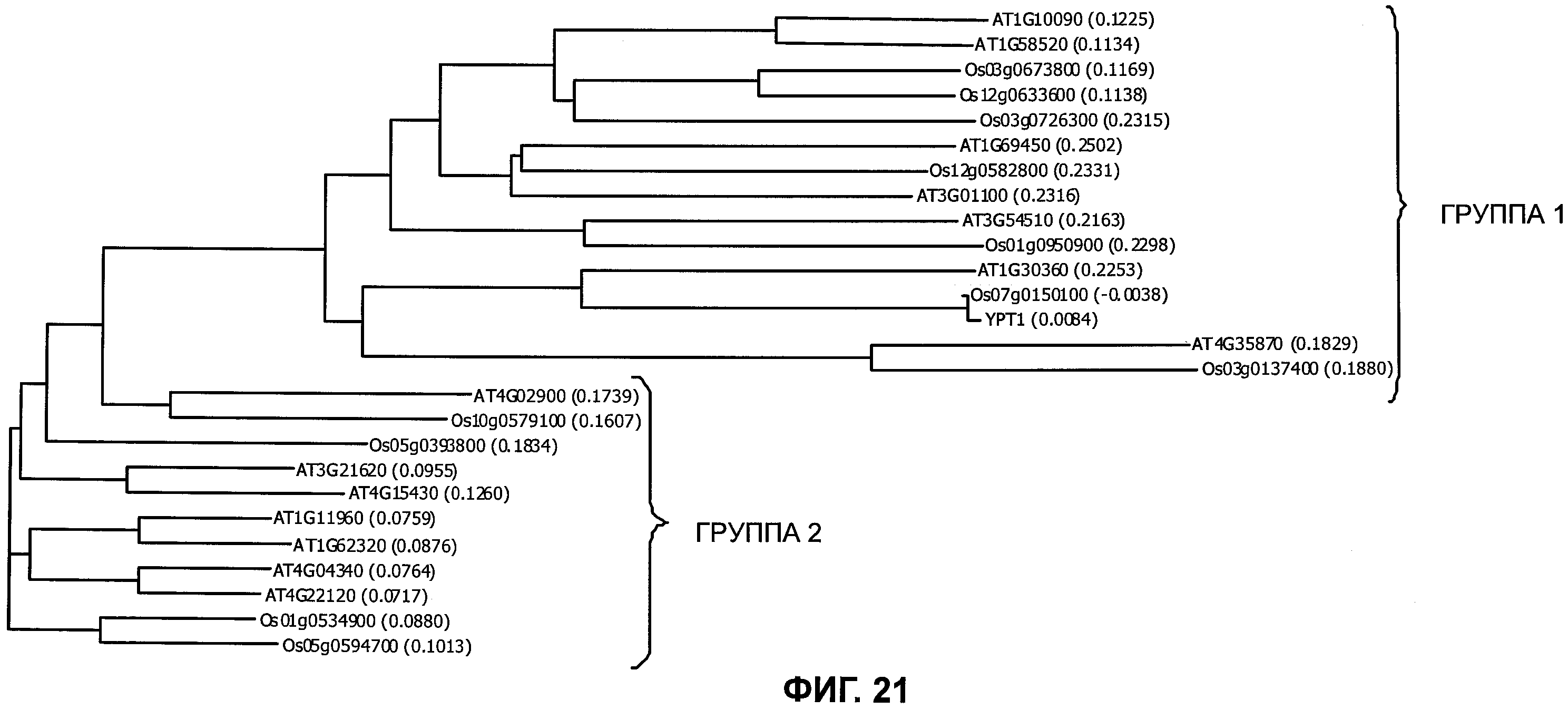
Факторы транскрипции обычно определяют как белки, которые демонстрируют специфическое для последовательности ДНК-связывающее сродство, и которые способны активировать и/или подавлять транскрипцию. Геном Arabidopsis thaliana кодирует по меньшей мере 1533 транскрипционных регуляторов, составляющих ~5.9% его предполагаемого общего числа генов (Riechmann et al. (2000) Science 290:2105-2109). База данных факторов транскрипции риса (DRTF) представляет собой коллекцию известных и предсказанных факторов транскрипции Oryza sativa L. ssp. indica и Oryza sativa L ssp. japonica и в настоящее время содержит 2025 предполагаемых моделей генов факторов транскрипции (TF) в indica и 2384 в japonica, распределенных в 63 семейства (Gao et al. (2006) Bioinformatics 2006, 22(10):1286-7).
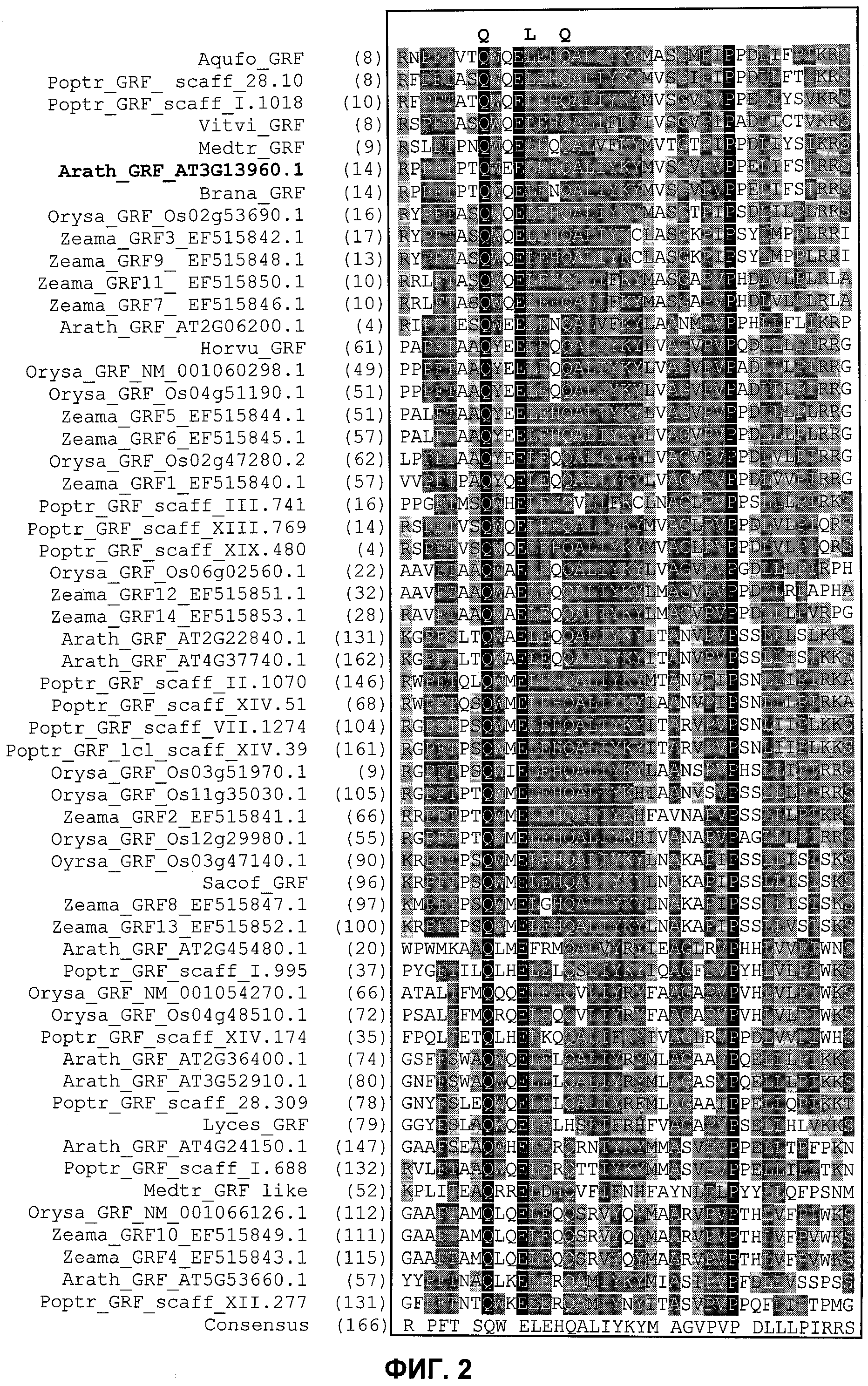
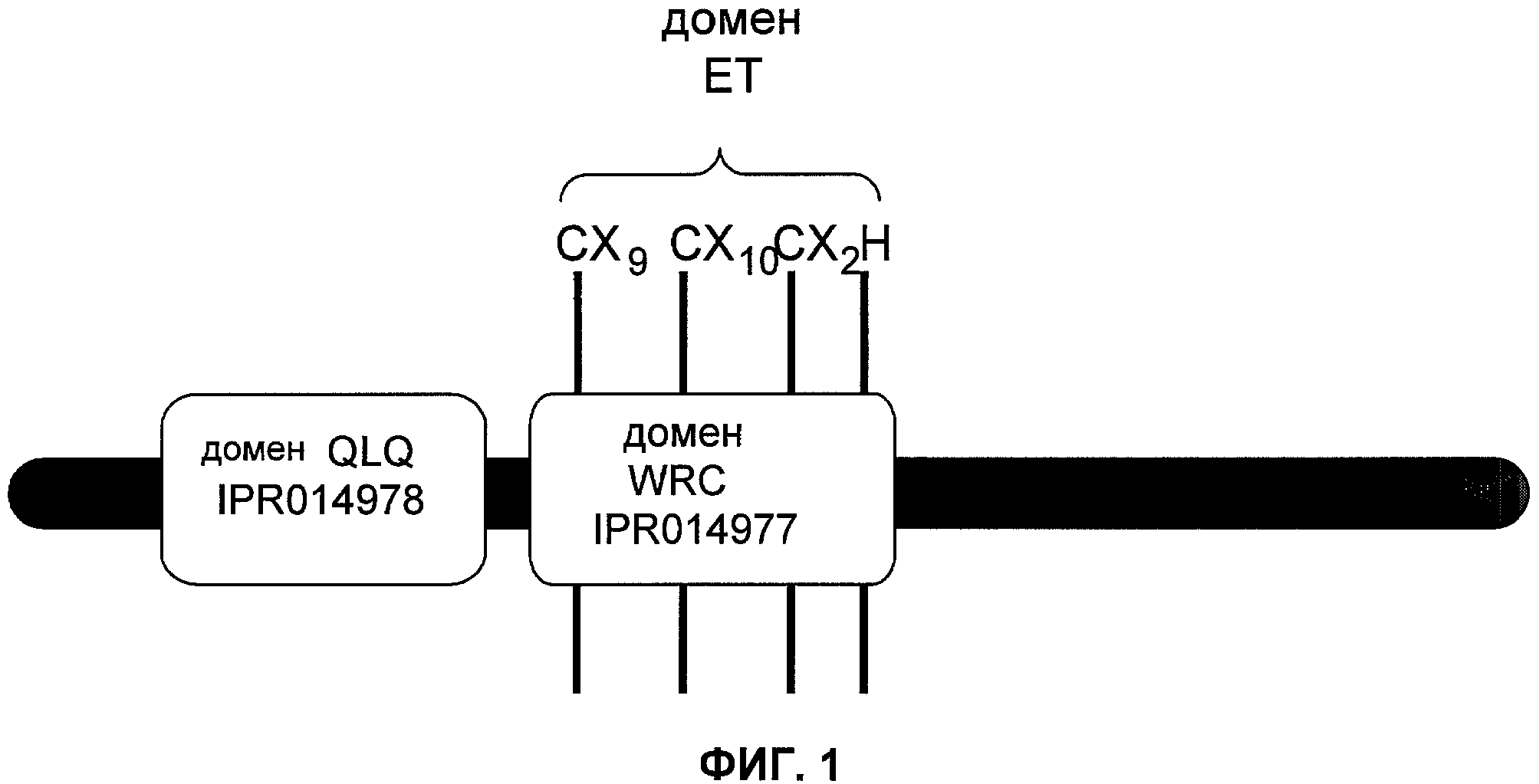
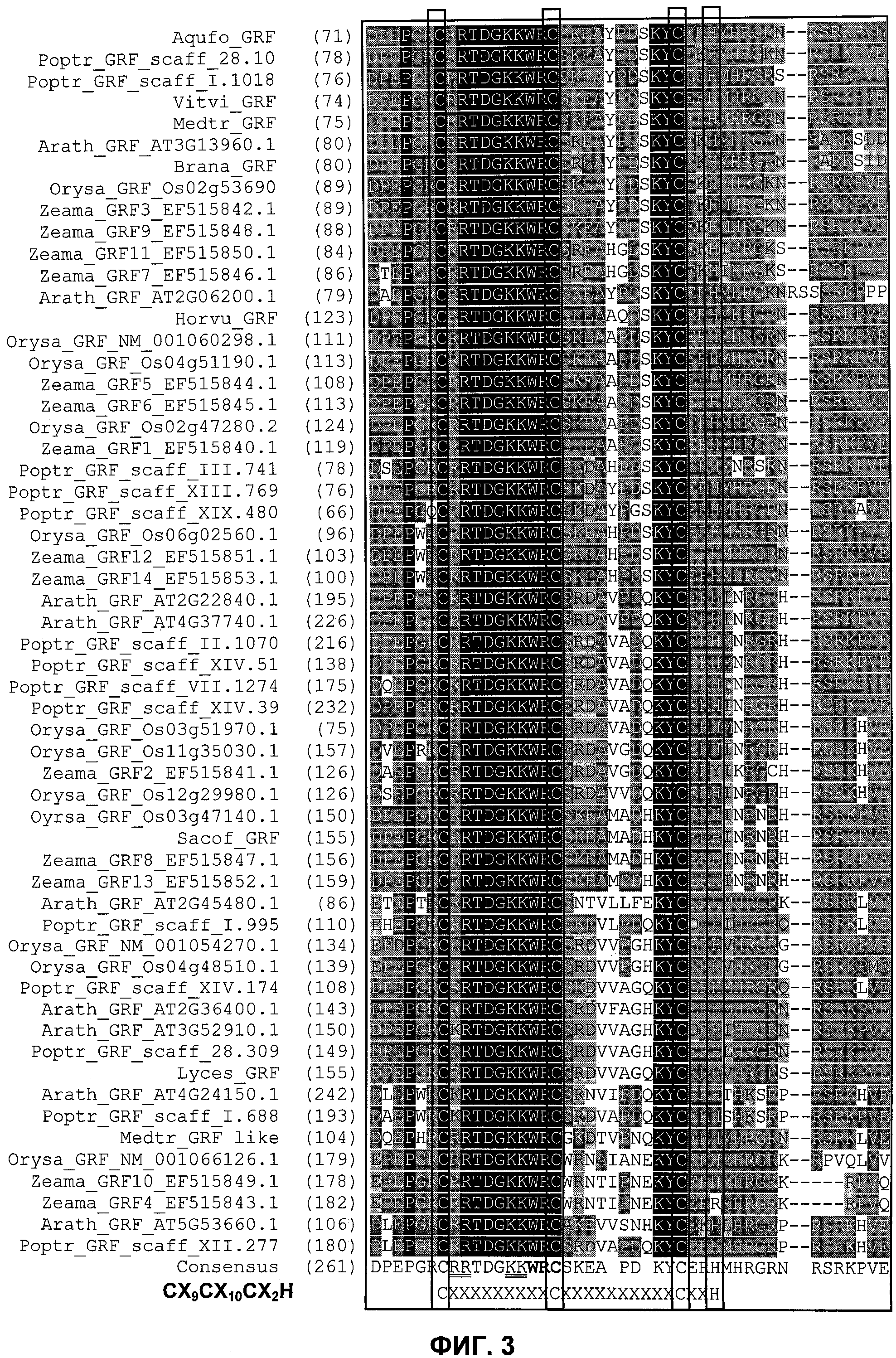
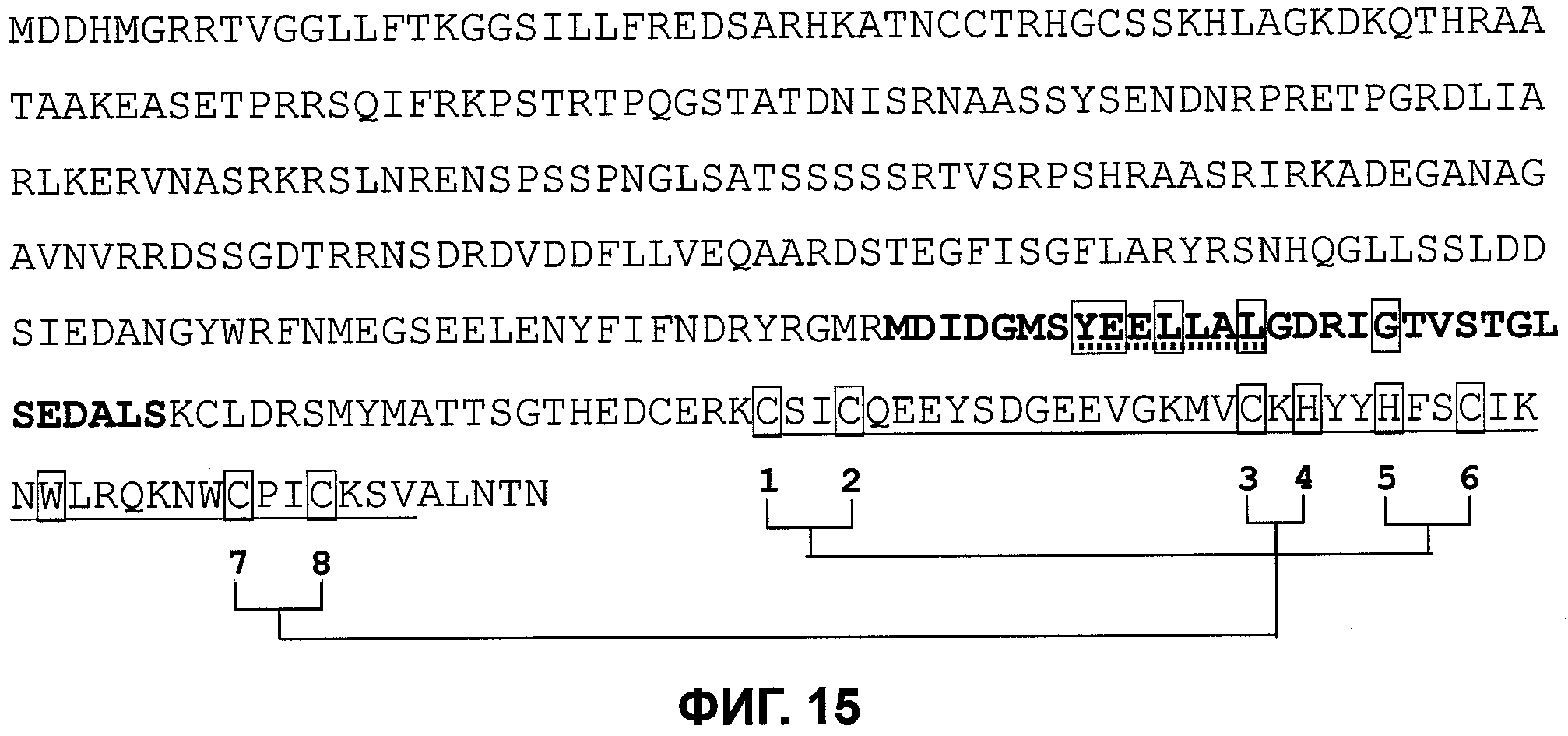
Одним из этих семейств является семейство факторов транскрипции, представляющее собой семейство фактора, регулирующего рост (GRF), которое является специфическим для растений. По меньшей мере девять полипептидов GRF идентифицировано в Arabidopsis thaliana (Kim et al. (2003) Plant J 36:94-104) и по меньшей мере двенадцать - в Oryza sativa (Choi et al. (2004) Plant Cell Physiol 45(7):897-904). Полипептиды GRF характеризуются присутствием на их N-концевой части по меньшей мере двух высококонсервативных доменов, названных в честь наиболее консервативных аминокислот внутри каждого домена: (i) домен QLQ (per. номер InterPro IPR014978, per. номер PFAM PF08880), где наиболее консервативные аминокислоты домена представляют собой Gln-Leu-Gln; и (ii) домен WRC (рег. номер InterPro IPR014977, рег. номер PFAM PF08879), где наиболее консервативные аминокислоты домена представляют собой Trp-Arg-Cys. Домен WRC также содержит два отличительных структурных признака, а именно домен WRC обогащен основными аминокислотами Lys и Arg и также содержит три Cys и один His остатки в консервативном участке (СХ9СХ10СХ2Н), обозначенном как домен эффектора транскрипции (ЕТ) (Ellerstrom et al. (2005) Plant Molec Biol 59:663-681). Консервативный участок цистеиновых и гистидиновых остатков в домене ЕТ напоминает участок белков «цинковые пальцы» (цинксвязывающие белки). Кроме того, сигнал ядерной локализации (NLS) обычно содержится в полипептидных последовательностях GRF.
Взаимодействие некоторых полипептидов GRF с небольшим семейством транскрипционных коактиваторов, GRF-взаимодействующих факторов (GIF1 - GIF3; также называемых полипептидами транслокации синовиальной саркомы (SYT) (synovial sarcoma translocation (SYT) polypeptide), SYT1 - SYT3), продемонстрировано с использованием анализа взаимодействия с помощью дрожжевых двойных гибридов (Kirn & Kende (2004) Proc Natl Acad Sci 101:13374-13379).
Название GRF также дано другому типу полипептидов, принадлежащих семейству 14-3-3 полипептидов (de Vetten & Ferl (1994) Plant Physiol 106:1593-1604), которые абсолютно не имеют отношения к полипептидам GRF, используемым при выполнении способов по изобретению.
Трансгенные растения Arabidopsis thaliana, трансформированные полипептидом GRF (OsGRF1) риса под контролем вирусного конститутивного промотора 35S CaMV, демонстрировали скрученные листья, сильно сниженное продолжение первичного цветения и задержку выметывания (van der Knapp et al. (2000) Plant Physiol 122:695-704). Трансгенные растения Arabidopsis thaliana, трансформированные одним из двух полипептидов GRF Arabidopsis (AtGRF1 и AtGRF2), развивавшие более крупные листья и семядоли, имели задержку в выметывании и были частично стерильны (вследствие отсутствия жизнеспособной пыльцы) по сравнению с растениями дикого типа (Kim et al. (2003) Plant J 36:94-104).
В патентной заявке США US2006/0048240 полипептид GRF Arabidopsis thaliana идентифицирован как SEQ ID NO:33421. В патентной заявке США US 2007/0022495 полипептид GRF Arabidopsis thaliana идентифицирован как SEQ ID NO:1803 (также он там упоминается как G1438). Трансгенные растения Arabidopsis, сверхэкспрессирующие G1438 с использованием промотора 35S CaMV, демонстрируют темно-зеленые листья.
Неожиданно в настоящее время было обнаружено, что увеличение экспрессии последовательности нуклеиновой кислоты, кодирующей полипептид GRF, дает растения, имеющие усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями.
Согласно одному варианту осуществления изобретения предоставляется способ усиления признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, предусматривающий увеличение экспрессии последовательности нуклеиновой кислоты, кодирующей полипептид GRF в растении. Усиленные признаки, связанные с урожайностью, содержат один или более из: увеличенная ранняя мощность, увеличенная надземная биомасса, увеличенная общая урожайность семян на растение, увеличенный уровень наполнения семян, увеличенный индекс урожайности и увеличенная масса тысячи зерен.
О молекулярной биологии формирования корней у однодольных растений известно немного. До настоящего времени было идентифицировано только несколько генов, которые влияют на развитие корней: примерами являются мутант rt1, который формирует малочисленные или не формирует корневые шейки и опорные корни (Jenkins, J. Hered. 21:79-80, 1930), мутант asr1, который демонстрирует дефектные зародышевые корешки (De Miranda et al., Maize Genet. Coop. News Lett. 54:18-19, 1980), мутант rtcs, не имеющий узловых (придаточных) корней (Hetz et al., Plant J. 10:845-857, 1996), мутант slr1 и мутант slr2 с укороченными боковыми корнями (Hochholdinger et al., Plant Physiol 125:1529-1539, 2001) или rum1, у которого нарушенная боковая инициация у первичного корня, а также инициация формирования зародышевого корешка (Woll et al., Plant Physiol., 139, 1255-1267, 2005). Liu et al. (Proteomics 6, 4300-4308, 2006) сделал протеомическое сравнение между первичными корнями сеянцев дикого типа и rum1 и обнаружил другие 12 генов, которые по-другому регулируются и вовлечены в биосинтез лигнина, защитные функции и цитратный цикл.
Другим геном, вовлеченным в формирование корней у однодольных растений, является гаа1, впервые изолированный из риса (Ge et al., Plant Physiol. 135, 1502-1513, 2004): этот ген кодирует 12.0 кДа белок, имеющий 58% гомологии с FPF1 (стимулирующий цветение фактор 1) Arabidopsis. В рисе RAA1 экспрессировался специфически в апикальной меристеме, зоне элонгации корневого кончика, стелах зоны ветвления и молодом боковом корне. Конститутивная сверхэкспрессия увеличивала число придаточных корней, но рост первичного корня снижался. Кроме того, увеличивалось содержание эндогенного ауксина. OsRAA1 также индуцировался ауксином; предполагается, что существует регуляция с положительной обратной связью между RAA1 и ауксином при развитии корней риса (Ge et al., 2004). Более того, растения, сверхэкспрессирующие OsRAA1, имели более длинные листья и стерильные цветки (Ge et al., 2004). WO 2006/067219 раскрывает применение FPF1 и родственных белков для увеличения получения углеводов в растениях, но трансгенные растения, сверхэкспрессирующие FPF1, не демонстрировали увеличенной урожайности семян и не сообщалось о влиянии на рост корней.
Неожиданно было обнаружено, что модуляция экспрессии нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, дает растения, имеющие улучшенные признаки, связанные с урожайностью, по сравнению с контрольными растениями.
Согласно одному варианту осуществления изобретения представляется способ улучшения признаков, связанных с урожайностью, у растения по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии нуклеиновой кислоты, кодирующей RAA1-подобный полипептид в растении. Улучшенные признаки, связанные с урожайностью, содержали увеличенные высоту, показатель побег/корень, толщину корня, индекс зелени, число цветков на метелку и увеличенную массу тысячи зерен. Улучшенные признаки, связанные с урожайностью, наблюдали при нормальных условиях роста, а также при стрессовых условиях.
Регулятор урожайности семян (SYR) представляет собой новый белок, который ранее не был охарактеризован. SYR демонстрирует некоторую гомологию (около 48% идентичности последовательности на уровне ДНК, около 45% на уровне белка) с белком Arabidopsis, названным ARGOS (Hu et al., Plant Cell 15, 1951-1961, 2003; US 2005/0108793). Hu et al. постулировал, что ARGOS представляет собой белок с уникальной функцией и кодирует один ген. Основные фенотипы сверхэкспрессии ARGOS в Arabidopsis представляются собой увеличенную биомассу листьев и отсроченное цветение. В отличие от этого сверхэкспрессия SYR в рисе в основном увеличивает урожайность семян, тогда как на биомассу листьев и период цветения не оказывается явного воздействия.
Неожиданно в настоящее время было обнаружено, что модуляция экспрессии в растении нуклеиновой кислоты, кодирующей белок регулятор урожайности семян (в дальнейшем именуемый SYR), дает растения, которые при выращивании в условиях абиотического стресса имеют улучшенную устойчивость к абиотическому стрессу по сравнению с контрольными растениями.
Следовательно, настоящее изобретение обеспечивает способ улучшения признаков, связанных с урожайностью, у растений, выращенных в условиях абиотического стресса, по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYR.
Полипептиды ARKL содержат домен RING-палец, который имеет сходство с обнаруженным в мышином белке ARKADIA, Е3 убиквитин-лигазой, вовлеченной в передачу сигнала Nodal во время эмбриогенеза (Mavrakis et al. 2007; PLoS Biol. 2007 Mar; 5(3):e67).
Убиквитилирование, способ, посредством которого белок модифицируется путем ковалентного присоединения убиквитина, является центральной и основной частью различных клеточных процессов у эукариот. У растений нарушения на этом пути вызывают многочисленные аберрации в развитии, измененную реакцию на внешние раздражители и модифицируют клеточный цикл и паттерны роста. Убиквитинированные белки предназначены для деградации через зависимый или не зависимый от протеасомы 268 путь. Модификация убиквитином играет роль в активации сигнальных белков, эндоцитозе, сортировке и модификации гистонов.
Судьба убиквитинированного белка определяется природой убиквитиновой связи. Один или множество убиквитинов могут присоединяться к мишени (моно- и полиубиквитинирование; специфический Lys остаток, использованный для формирования убиквитиновой цепочки, может влиять на конечную судьбу модифицированного белка, например, будет ли это деградация или активирование).
Присоединение убиквитина к белкам происходит в ходе многоэтапного процесса, включающего три фермента, именуемых Е1, Е2, Е3 (Glikcman and Ciechanover (2000) Physiol Rev 82:377-482). Вначале убиквитин связывается с белком АТФ-зависимым образом и затем переносится на цистеиновый акцептор в белке Е2 с образованием Е2-убиквитинового интермедиата, который действует как убиквитиновый донор в отношении белка-мишени в реакции, опосредуемой убиквитин-лигазой, также называемой лигазой Е3 или ферментом Е3. Существует множество типов лигаз Е3. Е3 лигазы типа RING характеризуются присутствием консервативного домена белка, названного RING-палец или RING-ZnF (Действительно Интересный Новый Ген (Really Interesting New Gene) - цинковый палец).
Цинк-связывающие мотивы представляют собой стабильные структуры, и они редко подвергаются конформационным изменениям при связывании со своей мишенью. Большая часть белков ZnF содержит множество пальцеподобных выступов, которые осуществляют тандемные контакты с их молекулой-мишенью, обычно распознавая вытянутые (extended) субстраты. RING-палец является специализированным цинк-связывающим доменом, который вероятно функционирует во взаимодействиях белок-белок. RING-палец составляет 40-60 остатков в длину и координирует два атома цинка. Он отличается от других цинковых пальцев, так как восемь аминокислотных остатков - лигандов металлов, которые координируют ион цинка, входят в специфическую структуру, называемую структурой «кросс скобки» (cross-brace) (Borden (2000). J MolBiol 295:1103-1112). Участок цистеинов/гистидинов, координирующих ионы цинка в таком домене, представляет собой С-х(2)-С-х(9-39)-С-х(1-3)-Н-х(2-3)-С-х(2)-С-х(4-48)-С-х(2)-С. Пары лигандов ионов металлов первая и третья координируются для связывания одного иона цинка, в то время как пары вторая и четвертая связывают второй.
Существуют два различных варианта, С3НС4-типа и С3Н2С3-типа, которые несомненно являются родственными, несмотря на различный цистеин/гистидин паттерн. Последний тип иногда называют 'RING-H2 палец'. В последнем координация иона цинка опосредуется 6 цистеинами и 2 гистидинами, в то время как в С3НС4 опосредуется 7 цистеинами и одним гистидином.
В Arabidopsis thaliana существуют по меньшей мере 477 предполагаемых белков, содержащих домен RING. Некоторые содержат множество доменов RING-пальцев. Домены RING классифицированы на восемь типов, основываясь на присутствующем остатке лиганда иона металла и/или числе аминокислот между ними (Stone at al. 2005) Plant Phys. 137, 13-30. Класс RING-H2 является самым большим классом в Arabidopsis. Основываясь на природе доменов и их организации, белки RING-пальцы в Arabidopsis также классифицируют на 30 групп, Группа 1 - Группа 30. Также распознаются подгруппы внутри некоторых групп, например подгруппы 2.1 и 2.2 группы 2 (Stone et al. 2005). Группу 1 относят к группе белка RING-палец, не имеющего ранее описанных доменов. Анализ последовательностей этих белков выявил области сходства между некоторыми белками за пределами домена RING, которые назвали DAR1 - DAR3 (домен, ассоциированный с RING). DAR1 и DAR3 составляют в длину приблизительно 40 аминокислот, a DAR2 - 120. Сообщали, что DAR1 встречается только в белках растительного происхождения (Stone et al. 2005). Присутствие общих консервативных доменов наводило на мысль о родственной функции белков, содержащих эти домены.
Неожиданно в настоящее время было обнаружено, что модуляция экспрессии нуклеиновой кислоты, кодирующей полипептид ARKL, дает растения, имеющие улучшенные признаки, связанные с урожайностью, в частности увеличенную урожайность по сравнению с контрольными растениями.
Согласно одному варианту осуществления изобретения предоставляется способ улучшения или увеличения признаков, связанных с урожайностью, у растения по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии нуклеиновой кислоты, кодирующей полипептид ARKL в растении.
Все эукариотические клетки содержат сложные системы внутренних мембран, которые устанавливают различные окруженные мембранами компартменты внутри клетки. Эндомембранная система представляет собой совокупность мембранных структур, вовлеченных в транспорт внутри клетки. Основными компонентами эндомембранной структуры являются эндоплазматический ретикулум, тельца Гольджи, пузырьки, клеточная мембрана и ядерная оболочка. Члены эндомембранной системы пропускают вещества через друг друга или посредством применения пузырьков. Общий признак всех клеток представляет собой наружную ограничивающую мембрану, называемую плазматической мембраной.
Клеточные мембраны построены из липидов и белков. Связь белков с мембраной может осуществляться через ковалентную связь, посредством которой белок присоединяется к липидам мембраны. В случае так называемых трансмембранных белков полипептидные цепочки белка в действительности проходят липидный бислой. Связь с мембраной может также происходить через соединение с белком, так называемым периферическим белком, посредством нековалентных связей с выступающими частями интегральных мембранных белков.
Трансмембранные белки (белки ТМ) имеют амфифильную природу с гидрофобными сегментами ТМ (TMS) и гидрофильными петлями. В трансмембранных белках часть внутри липидного бислоя состоит в основном из гидрофобных аминокислот. Они обычно организованы в альфа-спираль, так что полярные карбокси (-С=O) и амино (-NH) группы в пептидных связях могут взаимодействовать друг с другом, а не с их гидрофобным окружением. Те части полипептида, которые выступают за пределы бислоя, стремятся иметь высокий процент гидрофильных аминокислот. Следовательно, те, которые выступают во внеклеточное пространство, обычно являются гликозилированными.
Трансмембранная топология белка определена на основе экспериментальной рентгеновской кристаллографии, ЯМР, метода слияния генов, метода доступности замещенного цистеина, эксперимента по Asp(N)-связанному гликозилированию и других биохимических методов. Кроме того, разработан ряд методов предсказания трансмембранной топологии для определения структуры и функции ТМ белков из их аминокислотных последовательностей (Moller et al., 2001; Ikeda et al., 2002; Chen et al., 2002).
Анализ сходства белковых последовательностей между белками пользуется разработками в области геномики. Ряд доменов, консервативных между двумя или более белками, для которых никакой функции еще не установлено, могут исследоваться с использованием специфических алгоритмов. Один такой консервативный домен представляет собой так называемый домен DUF221 (домен с неизвестной функцией 221), как описано у Pfam (Finn et al. Nucleic Acids Research (2006) Database Issue 34:D247-D251). Этот домен обнаружен в семействе гипотетических трансмембранных белков, ни один из которых не имеет какой-либо известной функции, и выровненный участок составляет 538 остатков при максимальной длине. Этот домен встречается в ряде белков эукариотического происхождения. Сообщают, что экспрессия гена Arabidopsis, EDR4, кодирующего белок, содержащий DUF221, происходит вскоре после дегидратации (Kiyosue et al; Plant Mol Biol. 1994 25(5):791-8). Нокаутный мутант Arabidopsis, gfs10, no гену, кодирующему другой домен DUF221-содержащий белок, как сообщают, имеет фенотип, сходный с фенотипом мутантов вакуолярной сортировки (Fuji et al; 2007. Plant Cell. 2007. 19(2).-597-609).
Неожиданно в настоящее время было обнаружено, что модуляция экспрессии нуклеиновой кислоты, кодирующей полипептид YTP, дает растения, имеющие улучшенные признаки, связанные с урожайностью, в частности увеличенную урожайность по сравнению с контрольными растениями.
Согласно одному варианту осуществления изобретения предоставляется способ усиления (улучшения) признаков, связанных с урожайностью, у растения по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии нуклеиновой кислоты, кодирующей полипептид YTP в растении.
Определения
Полипептид(ы)/Белок(белки)
Термины "полипептид" и "белок" используются здесь взаимозаменяемо и относятся к аминокислотам в полимерной форме любой длины, соединенным вместе пептидными связями.
Полинуклеотид(ы)/Нуклеиновая(ые) кислота(ы)/ Последовательность(и) нуклеиновой(ых) кислот(ы)/нуклеотидная(ые) последовательность(и)
Термины "полинуклеотид(ы)", "последовательность(и) нуклеиновой(ых) кислот(ы)", "нуклеотидная(ые) последовательность(и)", "нуклеиновая(ые) кислота(ы)", "молекула нуклеиновой кислоты" используются здесь взаимозаменяемо и относятся к нуклеотидам, или рибонуклеотидам, или дезоксирибонуклеотидам, или комбинации их обоих в полимерной неразветвленной форме любой длины.
Контрольное(ые) растение(я)
Выбор подходящих контрольных растений является рутинной частью экспериментальной подготовки и может включать соответствующие растения дикого типа или соответствующие растения, не содержащие рассматриваемый ген. Контрольное растение обычно представляет собой растение того же вида или даже того же сорта, что и оцениваемое растение. Контрольное растение может также быть нуль-зиготой (nullizygote) оцениваемого растения. "Контрольное растение" при использовании здесь относится не только к целым растениям, но также к частям растений, включая семена и части семян.
Гомолог(и)
"Гомологи" белка охватывают пептиды, олигопептиды, полипептиды, белки и ферменты, имеющие аминокислотные замены, делеции и/или вставки по сравнению с немодифицированным рассматриваемым белком и имеющие схожую биологическую и функциональную активность как у немодифицированного белка, из которого они были получены.
Делеция относится к удалению одной или более аминокислот из белка.
Вставка относится к одному или более аминокислотным остаткам, введенным в заранее определенный участок белка. Вставки могут содержать N-концевые и/или C-концевые слияния, а также вставки внутри последовательности одной или множества аминокислот. Как правило, вставки внутри аминокислотной последовательности могут быть меньше, чем N- или С-концевые слияния, порядка около 1-10 остатков. Примеры N- или С-концевых слитых белков или пептидов включают связывающий домен или активирующий домен транскрипционного активатора, как использовано в системе дрожжевых двойных гибридов, белки оболочки фагов, (гистидин)-6-тэг, глутатион S-трансфераза-тэг, белок А, мальтоза-связывающий белок, дигидрофолатредуктазу, Tag·100 эпитоп, с-myc эпитоп, FLAG®-эпитоп, lacZ, CMP (кальмодулин-связывающий пептид), эпитоп гемагглютинина (НА), эпитоп белка С и эпитоп вируса везикулярного стоматита (VSV).
Замена относится к замещению аминокислот белка другими аминокислотами, имеющими схожие свойства (такие, как схожая гидрофобность, гидрофильность, антигенность, предрасположенность образовывать или разрушать α-спиральные структуры или β-складчатые структуры). Аминокислотные замены представляют собой обычно единичные остатки, но могут быть сгруппированы в зависимости от функциональных ограничений, установленных для полипептида; вставки могут быть обычно порядка около 1-10 аминокислотных остатков. Аминокислотные замены являются предпочтительно консервативными аминокислотными заменами. Таблицы консервативных замен хорошо известны в данной области техники (см., например, Creighton (1984) Proteins. W.H. Freeman and Company (Eds) и Таблицу 1 ниже).
|
Аминокислотные замены, делеции и/или вставки могут быть легко сделаны при использовании методов пептидного синтеза, хорошо известных в данной области техники, таких как твердофазный пептидный синтез и т.п., или путем манипуляции с рекомбинантными ДНК. Способы манипуляции с последовательностями ДНК для получения вариантов белка с заменами, вставками или делециями хорошо известны в данной области техники. Например, методы получения мутаций в виде замен в заранее определенных участках ДНК хорошо известны специалистам в данной области техники и включают М13-мутагенез, T7-GEN in vitro мутагенез (USB, Cleveland, ОН), QuickChange сайт-направленный мутагенез (Stratagene, San Diego, СА), ПЦР-опосредованный сайт-направленный мутагенез или другие протоколы сайт-направленного мутагенеза.
Производные
"Производные" включают пептиды, олигопептиды, полипептиды, которые могут по сравнению с аминокислотной последовательностью встречающейся в природе формы белка, такого как рассматриваемый белок, содержать замены аминокислот не встречающимися в природе аминокислотными остатками или добавления не встречающихся в природе аминокислотных остатков. "Производные" белка также охватывают пептиды, олигопептиды, полипептиды, которые содержат встречающиеся в природе измененные (гликозилированные, ацилированные, пренилированные, фосфорилированные, миристоилированные, сульфатированные и т.д.) или не встречающиеся в природе измененные аминокислотные остатки по сравнению с аминокислотной последовательностью встречающейся в природе формы полипептида. Производное может также содержать один(одно) или более аминокислотных заместителей или добавлений по сравнению с аминокислотной последовательностью, из которой оно получено, например репортерную молекулу или другой лиганд, ковалентно или нековалентно связанные с аминокислотной последовательностью, такие как репортерная молекула, которая присоединяется для облегчения ее обнаружения, и не встречающиеся в природе аминокислотные остатки по сравнению с аминокислотной последовательностью встречающегося в природе белка.
Кроме того, «производные» также включают слияния встречающейся в природе формы белка с пептидами для мечения, такими как FLAG, HIS6 или тиоредоксин (что касается обзора по пептидам для мечения, см. Terpe, Appl. Microbiol. Biotechnol. 60, 523-533, 2003).
Ортолог(и)/Паралог(и)
Ортологи и паралоги охватывают эволюционные понятия, используемые для описания наследственных взаимоотношений генов. Паралоги представляют собой гены внутри одного и того же вида, которые произошли путем удвоения предкового гена; ортологи представляют собой гены из различных организмов, которые произошли путем видообразования, и также образованы из общего предкового гена.
Домен
Термин "домен" относится к набору аминокислот, сохраняемых в определенных положениях на всем протяжении выравнивания последовательностей эволюционно родственных белков. Несмотря на то, что аминокислоты в других положениях могут различаться между гомологами, аминокислоты, которые весьма консервативны в определенных положениях, означают аминокислоты, которые вероятно являются важными для структуры, стабильности или функции белка. Будучи идентифицированы по их высокой степени консервативности в выровненных последовательностях семейства белковых гомологов, они могут использоваться как идентификаторы для определения, принадлежит ли любой рассматриваемый полипептид к ранее обнаруженному семейству полипептидов.
Мотив/ Консенсусная последовательность / Сигнатура (Signature)
Термин "мотив", или "консенсусная последовательность", или "сигнатура" относится к короткому консервативному участку последовательности эволюционно родственных белков. Мотивы часто являются высококонсервативными частями доменов, но могут также включать только часть домена или быть расположены за пределами консервативного домена (если все аминокислоты мотива выходят за пределы определенного домена).
Гибридизация
Термин "гибридизация", как здесь определено, означает процесс, в котором по существу гомологичные комплементарные нуклеотидные последовательности подвергаются отжигу друг с другом. Процесс гибридизации может происходить полностью в растворе, т.е. обе молекулы комплементарных нуклеиновых кислот находятся в растворе. Процесс гибридизации может также происходить с одной из молекул комплементарных нуклеиновых кислот, иммобилизованной на матрице, такой как магнитные шарики, гранулы сефарозы или любая другая смола. Процесс гибридизации может, кроме того, происходить с одной из молекул комплементарных нуклеиновых кислот, иммобилизованной на твердой подложке, такой как нитроцеллюлозная или нейлоновая мембрана, или иммобилизованной путем, например, фотолитографии на, например, подложку из кремниевого стекла (последнее известно как матрицы или микроматрицы нуклеиновых кислот или как чипы нуклеиновых кислот). Для того чтобы допустить возникновение гибридизации, молекулы нуклеиновой кислоты являются, как правило, термически или химически денатурированными для перехода двухцепочечной ДНК в две отдельные нити и/или для удаления шпилек или других вторичных структур из молекул одноцепочечных нуклеиновых кислот.
Термин "жесткость" относится к условиям, в которых происходит гибридизация. На жесткость гибридизации влияют такие условия, как температура, концентрация соли, ионная сила и состав буфера для гибридизации. Как правило, выбирают, чтобы условия низкой жесткости составляли примерно на 30°С ниже, чем температура плавления (Tm) для конкретной последовательности при определенных ионной силе и рН. Условия средней жесткости - когда температура составляет на 20°С ниже Tm, и условия высокой жесткости - когда температура на 10°С ниже Tm. Условия гибридизации с высокой жесткостью обычно используются для выделения гибридизующихся последовательностей, которые имеют высокое сходство с последовательностью-мишенью нуклеиновой кислоты. Однако последовательности нуклеиновых кислот могут отклоняться по составу последовательности и, тем не менее, кодировать по существу идентичный полипептид благодаря вырожденности генетического кода. Следовательно, условия гибридизации со средней жесткостью иногда могут быть необходимы для идентификации таких молекул нуклеиновых кислот.
Tm означает температуру при определенной ионной силе и рН, при которой 50% последовательности-мишени гибридизуется с абсолютно совпадающим зондом. Tm зависит от условий раствора, и состава оснований, и длины зонда. Например, более длинные последовательности гибридизуются, в частности, при более высоких температурах. Максимальную скорость гибридизации получают при от около 16°С до 32°С ниже Tm. Присутствие моновалентных катионов в гибридизационном растворе уменьшает электростатическое отталкивание между двумя нитями нуклеиновых кислот, таким образом способствуя образованию гибрида; этот эффект является заметным для концентраций натрия вплоть до 0.4М (для более высоких концентраций этот эффект может игнорироваться). Формамид уменьшает температуру плавления дуплексов ДНК-ДНК и ДНК-РНК от 0.6 до 0.7°С на каждый процент формамида, и добавление 50% формамида обеспечивает проведение гибридизации при от 30 до 45°С, хотя скорость гибридизации может снижаться. Ошибочно спаренные основания уменьшают скорость гибридизации и термическую стабильность дуплексов. В среднем и для больших зондов Tm снижается примерно на 1°С на % ошибочно спаренных оснований. Tm может быть рассчитана при использовании следующих уравнений в зависимости от типов гибридов:
1) ДНК-ДНК гибриды (Meinkoth and Wahl, Anal. Biochem., 138: 267-284, 1984):
Tm=81.5°C+16.6×log10[Na+]a+0.41×%[G/Cb]-500×[Lc]-1-0.61×% формамида
2) ДНК-РНК или РНК-РНК гибриды:
Tm=79.8+18.5 (log10[Na+]a)+0.58(%G/Cb)+11.8(%G/Cb)2-820/Lc
3) олиго-ДНК или олиго-РНКd гибриды:
Для <20 нуклеотидов: Tm=2 (In)
Для 20-35 нуклеотидов: Tm=22+1.46 (In)
а или для другого моновалентного катиона, но только точно в диапазоне 0.01-0.4 М.
b только точно для %GC в диапазоне 30% - 75%.
c L = длина дуплекса в парах оснований.
d олиго, олигонуклеотид; In, = эффективная длина праймера = 2×(число С/С) + (число А/Т).
Неспецифическое связывание можно контролировать, используя любую из ряда известных методик, таких как, например, блокирование мембраны растворами, содержащими белки, добавление гетерологичных РНК, ДНК и SDS (додецилсульфат натрия) в буфер гибридизации и обработка РНКазой. Для негомологичных зондов серии гибридизаций могут проводиться путем изменения одного из: (i) прогрессивное понижение температуры отжига (например, с 68°С до 42°С) или (ii) прогрессивное понижение концентрации формамида (например, с 50% до 0%). Специалист в данной области техники осведомлен о различных параметрах, которые могут изменяться во время гибридизации и которые будут или сохранять, или изменять условия жесткости.
Помимо условий гибридизации, специфичность гибридизации обычно также зависит от функции промывок после гибридизации. Для удаления фона, получающегося в результате неспецифической гибридизации, образцы промывают разбавленными солевыми растворами. Критические факторы таких промывок включают ионную силу и температуру финального промывочного раствора; чем ниже концентрация соли и выше температура промывки, тем выше жесткость промывки. Условия промывки обычно осуществляют при или ниже жесткости гибридизации. Положительная гибридизация дает сигнал, который представляет собой по меньшей мере удвоенный сигнал фона. Как правило, подходящие жесткие условия для анализов гибридизации последовательностей нуклеиновых кислот или процедур детектирования амплификации генов являются такими, как изложено выше. Могут также быть выбраны более или менее жесткие условия. Специалист в данной области техники осведомлен о различных параметрах, которые могут быть изменены во время промывки и которые будут или сохранять, или изменять условия жесткости.
Например, типичные условия гибридизации с высокой жесткостью для гибридов ДНК длиннее, чем 50 нуклеотидов, включают гибридизацию при 65°С в 1×SSC (раствор цитрата и хлорида натрия) или при 42°С в 1×SSC и 50% формамиде с последующей промывкой при 65°С в 0.3×SSC. Примеры условий гибридизации со средней жесткостью для гибридов ДНК длиннее 50 нуклеотидов включают гибридизацию при 50°С в 4×SSC или при 40°С в 6×SSC и 50% формамиде с последующей промывкой при 50°С в 2×SSC. Длина гибрида представляет собой предполагаемую длину молекулы гибридизующейся нуклеиновой кислоты. Когда гибридизуются молекулы нуклеиновых кислот с известной последовательностью, длина гибрида может определяться выравниванием последовательностей и определением консервативных участков, описанных здесь. 1×SSC означает 0.15М NaCl и 15 мМ цитрат натрия; гибридизационный раствор и промывочные растворы могут дополнительно включать 5× реагент Денхардта, 0.5-1.0% SDS, 100 мкг/мл денатурированной, фрагментированной ДНК из молок лососевых рыб, 0.5% пирофосфата натрия.
Применительно к определению уровня жесткости может быть сделана ссылка на Sambrook et al. (2001) Molecular Cloning: a laboratory manual, 3rd Edition, Cold Spring Harbor Laboratory Press, CSH, New York, или на Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989 and yearly updates).
Сплайс-вариант
Термин "сплайс-вариант" при использовании здесь охватывает варианты последовательности нуклеиновой кислоты, в которой выбранные интроны и/или экзоны вырезаются, замещаются, перемещаются или добавляются или в которой интроны укорачиваются или удлиняются. Такие варианты могут быть вариантами, в которых биологическая активность белка по существу сохраняется, это может быть достигнуто селективным сохранением функциональных частей белка. Такие сплайс-варианты могут обнаруживаться в природе или быть получены человеком. Способы прогнозирования и выделения таких сплайс-вариантов хорошо известны в данной области техники (см., например, Foissac and Schiex (2005) BMC Bioinformatics 6: 25).
Аллельный вариант
Аллели или аллельные варианты представляют собой альтернативные формы данного гена, локализованные в одинаковом хромосомном положении. Аллельные варианты включают однонуклеотидные полиморфизмы (SNP), а также небольшие инсерционно-делеционные полиморфизмы (INDEL). Размер INDEL обычно менее чем 100 пар оснований. SNP и INDEL формируют наибольшую совокупность вариантов последовательностей у встречающихся в природе полиморфных линий большинства организмов.
Перетасовка генов/направленная эволюция
Перетасовка генов или направленная эволюция состоит из повторений перетасовки ДНК с последующим соответствующим скринингом и/или селекцией для образования вариантов последовательностей нуклеиновых кислот или их частей, кодирующих белки, имеющие модифицированную биологическую активность (Castle et al., (2004) Science 304(5674): 1151-4; патенты США 5,811,238 и 6,395,547).
Регуляторный элемент/контрольная последовательность/промотор
Термины "регуляторный элемент", "контрольная последовательность" и "промотор" используются все здесь взаимозаменяемо и должны употребляться в широком контексте, чтобы ссылаться на последовательности регуляторных нуклеиновых кислот, способные к эффективной экспрессии последовательностей, с которыми они лигированы. Термин «промотор» обычно относится к контрольной последовательности последовательности нуклеиновой кислоты, расположенной выше точки начала транскрипции гена, которая вовлечена в узнавание и присоединение РНК полимеразы и других белков, таким образом направляя транскрипцию функционально связанной нуклеиновой кислоты. Вышеуказанными терминами охватываются транскрипционные регуляторные последовательности, полученные из классического эукариотического геномного гена (включая ТАТА-бокс, который необходим для правильной инициации транскрипции, с или без последовательности ССААТ-бокса), и дополнительные регуляторные элементы (т.е. вышерасположенные активирующие последовательности, усилители и сайленсеры), которые изменяют экспрессию гена в ответ на дифференцировочные (developmental) и/или внешние стимулы или тканеспецифичным образом. Также этот термин включает транскрипционную регуляторную последовательность классического прокариотического гена, и в этом случае он может включать транскрипционные регуляторные последовательности -35 бокса и/или -10 бокса.
Термин "регуляторный элемент" также охватывает синтетическую слитую молекулу или производное, которые предоставляют, активируют или увеличивают экспрессию молекулы последовательности нуклеиновой кислоты в клетке, ткани или органе.
"Растительный промотор" содержит регуляторные элементы, которые способствуют экспрессии участка кодирующей последовательности в растительных клетках. Соответственно, нет необходимости, чтобы растительный промотор был растительного происхождения, и он может происходить из вирусов или микроорганизмов, например из вирусов, которые атакуют растительные клетки. "Растительный промотор" предпочтительно происходит из растительной клетки, например из растения, которое трансформировано последовательностью нуклеиновой кислоты, экспрессируемой в способе согласно изобретению и описанной здесь. Это также применимо к другим "растительным" регуляторным сигналам, таким как "растительные" терминаторы. Промоторы, расположенные выше нуклеотидных последовательностей, пригодных в способах настоящего изобретения, могут быть модифицированы одним или более нуклеотидных замещений, вставок и/или делеций, не затрагивая функциональность или активность или промоторов, открытой рамки считывания (ORF), или 3'-регуляторного участка, такого как терминаторы или другие 3'-регуляторные участки, расположенные не в ORF. Кроме того, возможно, что активность промоторов увеличивается модификацией их последовательности или что они замещаются полностью более активными промоторами, даже промоторами из гетерологичных организмов. Во время экспрессии в растениях молекула последовательности нуклеиновой кислоты должна быть, как описано выше, функционально связана с подходящим промотором или содержать подходящий промотор, который экспрессирует ген в правильный момент времени и с требуемым пространственным паттерном экспрессии.
Для идентификации функционально эквивалентных промоторов сила промотора и/или паттерн экспрессии возможного промотора могут быть проанализированы, например, функциональным связыванием промотора с репортерным геном и анализом уровня экспрессии и паттерна репортерного гена в различных тканях растения. Подходящие хорошо известные репортерные гены включают, например, бета-глюкуронидазу или бета-галактозидазу. Активность промотора анализируют измерением ферментной активности бета-глюкуронидазы или бета-галактозидазы. Силу промотора и/или паттерн экспрессии можно затем сравнить с этими параметрами референсного промотора (такого, как используемый в способах настоящего изобретения). Альтернативно, силу промотора можно проанализировать количественным определением уровней мРНК или путем сравнения уровней мРНК последовательности нуклеиновой кислоты, использованной в способах настоящего изобретения, с уровнями мРНК генов домашнего хозяйства, такими как 18S рРНК, используя методы, хорошо известные в данной области техники, такие как Нозерн-блоттинг с денситометрическим анализом радиоавтографов, количественная ПЦР в реальном времени или ОТ-ПЦР (ПЦР с обратной транскрипцией) (Heid et al., 1996 Genome Methods 6: 986-994). Как правило, под "слабым промотором" подразумевается промотор, который проводит экспрессию кодирующей последовательности на низком уровне. Под "низким уровнем" подразумеваются уровни от около 1/10000 транскриптов до около 1/100000 транскриптов, до около 1/5000000 транскриптов на клетку. И наоборот, "сильный промотор" проводит экспрессию кодирующей последовательности на высоком уровне или от около 1/10 транскриптов до около 1/100 транскриптов, до около 1/1000 транскриптов на клетку. Как правило, под "промотором средней силы" подразумевается промотор, который проводит экспрессию кодирующей последовательности на уровне, который представлен во всех примерах ниже, который получен под контролем промотора 35S CaMV.
Функционально связанный
Термин "функционально связанный" при использовании здесь относится к такой функциональной связи между последовательностью промотора и рассматриваемым геном, что эта последовательность промотора способна инициировать транскрипцию рассматриваемого гена.
Конститутивный промотор
Термин "конститутивный промотор" относится к промотору, который является транскрипционно активным во время большей части, но не обязательно всех фаз роста и развития, и в большинстве условий окружающей среды, по меньшей мере в одной клетке, ткани или органе. Таблица 2а ниже дает примеры конститутивных промоторов.
|
Универсальный промотор
Универсальный промотор является активным по существу во всех тканях или клетках организма.
Промотор, регулируемый в процессе развития (Developmentally-regulated promoter)
Промотор, регулируемый в процессе развития, активен во время определенных стадий развития или в частях растения, которые подвергаются изменениям в процессе развития.
Индуцибельный промотор
Индуцибельный промотор вызывает или усиливает инициацию транскрипции в ответ на химические стимулы (для ознакомления см. Gatz 1997, Annu. Rev. Plant Physiol. Plant Mol. Biol., 48:89-108), стимулы окружающей среды или физические стимулы или может быть "стресс-индуцибельным", т.е. активируемым, когда растение подвергается воздействию различных стрессовых условий, или "патоген- индуцибельным", т.е. активируемым, когда растение подвергается воздействию различных патогенов.
Органоспецифичный/тканеспецифичный промотор
Органоспецифичный или тканеспецифичный промотор представляет собой промотор, который способен к предпочтительному инициированию транскрипции в определенных органах или тканях, таких как ткани листьев, корней, семян и т.д. Например, "промотор, специфичный для корней" означает промотор, который транскрипционно активен преимущественно в корнях растений, по существу исключая любые другие части растения, хотя и допуская еще какую-либо ликовую экспрессию в этих других частях растений. Промоторы, способные инициировать транскрипцию только в определенных клетках, указываются здесь как "клеточно-специфичные".
Примеры промоторов, специфичных для корней, перечислены в Таблице 2b ниже:
|
Специфичный для семян промотор транскрипционно активен преимущественно в ткани семян, но необязательно исключительно в ткани семян (в случаях ликовой экспрессии). Специфичный для семян промотор может быть активен во время развития семян и/или во время прорастания. Примеры специфичных для семян промоторов показаны в Таблице 2с ниже. Дополнительные примеры специфичных для семян промоторов даны в статье Qing Qu и Takaiwa (Plant Biotechnol. J. 2, 113-125, 2004), раскрытие которой включено сюда путем ссылки, как если бы полностью излагалось.
|
Промотор, специфичный для зеленых тканей растений, при определении здесь означает промотор, который транскрипционно активен преимущественно в зеленой ткани растений, по существу исключая любые другие части растения, хотя и допуская еще какую-либо ликовую экспрессию в этих других частях растений.
Примеры промоторов, специфичных для зеленых тканей растений, которые могут использоваться для осуществления способов изобретения, представлены в Таблице 2d ниже.
|
Другой пример тканеспецифичного промотора - это меристемспецифичный промотор, который транскрипционно активен преимущественно в меристематической ткани, по существу, исключая любые другие части растения, хотя и допуская еще какую-либо ликовую экспрессию в этих других частях растений. Примеры меристемспецифичных промоторов, которые могут использоваться для выполнения способов изобретения, представлены в Таблице 2е ниже.

Терминатор
Термин "терминатор" включает контрольную последовательность, которая является последовательностью ДНК на конце единицы транскрипции, которая дает сигнал к 3' процессингу и полиаденилированию первичного транскрипта и окончанию транскрипции. Терминатор может быть получен из природного гена, из множества других растительных генов или из Т-ДНК. Добавляемый терминатор может быть получен из, например, генов нопалинсинтазы или октопинсинтазы, или альтернативно из другого растительного гена, или, менее предпочтительно, из любого другого эукариотического гена.
Модуляция
Термин "модуляция" означает по отношению к экспрессии или экспрессии гена процесс, в котором уровень экспрессии меняется посредством указанной экспрессии гена по сравнению с контрольным растением, причем уровень экспрессии может увеличиваться или уменьшаться. Исходная, немодулированная, экспрессия может быть любым видом экспрессии структурной РНК (рРНК, тРНК) или мРНК с последующей трансляцией. Термин "модулирование активности" означает любое изменение экспрессии последовательностей нуклеиновых кислот или кодируемых белков по изобретению, которое приводит к увеличенной урожайности и/или увеличенному росту растений.
Экспрессия
Термин "экспрессия" или "экспрессия гена" означает транскрипцию определенного гена, или определенных генов, или определенной генетической конструкции. Термин "экспрессия" или "экспрессия гена", в частности, означает транскрипцию гена, или генов, или генетической конструкции в структурную РНК (рРНК, тРНК) или мРНК с или без последующей трансляции последней в белок. Процесс включает транскрипцию ДНК и процессинг получающегося продукта мРНК.
Увеличенная экспрессия /сверхэкспрессия
Термин "увеличенная экспрессия" или "сверхэкспрессия" при использовании здесь означает любую форму экспрессии, которая является добавочной к уровню исходной экспрессии дикого типа.
Способы увеличения экспрессии генов или генных продуктов хорошо задокументированы в данной области техники и включают, например, сверхэкспрессию, управляемую соответствующими промоторами, применение энхансеров транскрипции или энхансеров трансляции. Выделенные последовательности нуклеиновых кислот, которые служат как промоторные элементы или элементы энхансеров, могут вводиться в соответствующее положение (обычно выше) негетерологичной формы полинуклеотида для того, чтобы повышать экспрессию последовательности нуклеиновой кислоты, кодирующей рассматриваемый полипептид. Например, эндогенные промоторы могут изменяться in vivo путем мутации, делеции и/или замены (см., Kmiec, US 5,565,350; Zarling et al., WO9322443), или выделенные промоторы могут вводиться в растительную клетку в соответствующей ориентации и на соответствующем расстоянии от гена по настоящему изобретению с тем, чтобы контролировать экспрессию гена.
Если требуется экспрессия полипептида, как правило, желательно включить участок полиаденилирования на 3'-конце кодирующей области полинуклеотида. Участок полиаденилирования можно получать из природного гена, из множества других растительных генов или из Т-ДНК. Добавляемая последовательность на 3'-конце может быть получена из, например, генов нопалинсинтазы или октопинсинтазы, или альтернативно из другого растительного гена, или, менее предпочтительно, из любого другого эукариотического гена.
Интронная последовательность также может быть добавлена к 5' нетранслируемому участку (UTR) или кодирующей последовательности частичной кодирующей последовательности для увеличения количества зрелой мРНК, которая накапливается в цитозоле. Включение пригодного для сплайсинга интрона в единицу транскрипции как в растительных, так и животных экспрессионных конструкциях, как было показано, увеличивает экспрессию гена на уровне и мРНК и белка до 1000-кратной (Buchman and Berg (1988) Mol. Cell biol. 8:4395-4405; Callis et al. (1987) Genes Dev 1:1183-1200). Такое интронное усиление экспрессии гена является, как правило, наибольшим при размещении рядом с 5'-концом единицы транскрипции. Применение кукурузных интронов 1, 2 и 6 Adh1-S, интрона Bronze-1 известно в данной области техники. Для общей информации см.: The Maize Handbook, Chapter 116, Freeling and Walbot, Eds., Springer, N.Y. (1994).
Эндогенный ген
Ссылка здесь на "эндогенный" ген относится не только к изучаемому гену, как он обнаруживается в растении в его природной форме (т.е. без какого-либо вмешательства человека), но также относится к этому же гену (или по существу гомологичным нуклеиновой кислоте/гену) в выделенной форме, впоследствии введенному (вновь введенному) в растение (трансген). Например, трансгенное растение, содержащее такой трансген, может сталкиваться со значительным уменьшением экспрессии трансгена и/или значительным уменьшением экспрессии эндогенного гена.
Выделенный ген может быть выделен из организма или может быть получен человеком, например, химическим синтезом.
Сниженная экспрессия
Ссылка здесь на "сниженную экспрессию" или "уменьшение или значительное устранение" экспрессии употребляется для обозначения уменьшения экспрессии эндогенного гена, и/или уровней полипептидов, и/или активности полипептида по сравнению с контрольными растениями. Уменьшение или значительное устранение означают в возрастающем порядке предпочтения по меньшей мере 10%, 20%, 30%, 40% или 50%, 60%, 70%, 80%, 85%, 90% или 95%, 96%, 97%, 98%, 99% или большее уменьшение по сравнению с контрольными растениями.
Для уменьшения или значительного устранения экспрессии эндогенного гена в растении требуется достаточная длина по существу смежных нуклеотидов последовательности нуклеиновой кислоты. Для выполнения сайленсинга гена это может быть всего 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10 или менее нуклеотидов, альтернативно это может быть вплоть до целого гена (включая 5' и/или 3' UTR, или частично, или целые). Участок по существу смежных нуклеотидов может быть получен из последовательности нуклеиновой кислоты, кодирующей рассматриваемый белок (ген-мишень), или любой последовательности нуклеиновой кислоты, способной кодировать ортолог, паролог или гомолог рассматриваемого белка. Предпочтительно, участок по существу смежных нуклеотидов способен формировать водородные связи с геном-мишенью (или смысловой, или антисмысловой цепью), более предпочтительно, участок по существу смежных нуклеотидов имеет, в возрастающем порядке предпочтения, 50%, 60%, 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 100% идентичности последовательности с геном-мишенью (или смысловой, или антисмысловой цепью). Последовательность нуклеиновой кислоты, кодирующая (функциональный) полипептид, не является необходимым условием для различных способов, обсуждаемых здесь, для уменьшения или значительного устранения экспрессии эндогенного гена.
Такое уменьшение или значительное устранение экспрессии может достигаться при использовании рутинных средств и методов. Способ уменьшения или значительного устранения экспрессии эндогенного гена осуществляется путем РНК-опосредованного сайленсинга с использованием инвертированного повтора последовательности нуклеиновой кислоты или ее части (в этом случае участок по существу смежных нуклеотидов, полученных из рассматриваемого гена или из любой последовательности нуклеиновой кислоты, способной кодировать ортолог, паралог или гомолог рассматриваемого белка), предпочтительно способного к формированию шпилечной структуры. Другой пример метода РНК-сайленсинга включает введение последовательностей нуклеиновых кислот или их частей (в этом случае участок по существу смежных нуклеотидов, полученных из рассматриваемого гена или из любой последовательности нуклеиновой кислоты, способной кодировать ортолог, паралог или гомолог рассматриваемого белка) в смысловой ориентации в растение. Другой пример метода РНК-сайленсинга включает применение антисмысловых последовательностей нуклеиновых кислот. Сайленсинг генов может также достигаться инсерционным мутагенезом (например, вставкой Т-ДНК или вставкой транспозона) или путем стратегий, как описано, среди прочих, Angell и Baulcombe ((1999) Plant J 20(3): 357-62), (Amplicon VIGS WO 98/36083) или Baulcombe (WO 99/15682). Другие способы, такие как применение антител, направленных против эндогенного полипептида, для ингибирования его функции в растении или вмешательство в сигнальный путь, в который вовлечен полипептид, могут быть хорошо известны специалисту в данной области техники. Могут использоваться искусственные и/или природные микроРНК (miPHK) для выключения экспрессии гена и/или трансляции мРНК. Эндогенные miPHK являются одноцепочечными небольшими РНК обычно длиной 19-24 нуклеотидов.
Такое уменьшение или значительное устранение экспрессии может достигаться при использовании рутинных средств и методов. Предпочтительный способ уменьшения или значительного устранения экспрессии эндогенного гена заключается во введении и экспрессии в растении генетической конструкции, в которой нуклеиновая кислота (в этом случае участок по существу смежных нуклеотидов, полученных из рассматриваемого гена или из любой нуклеиновой кислоты, способной кодировать ортолог, паралог или гомолог любого рассматриваемого белка) клонирована в виде инвертированного повтора (частично или полностью), разделенного спейсером (некодирующая ДНК).
В таком предпочтительном способе экспрессия эндогенного гена уменьшена или по существу устранена через РНК-опосредованный сайленсинг с использованием инвертированного повтора последовательности нуклеиновой кислоты или ее части (в этом случае участок по существу смежных нуклеотидов, полученных из рассматриваемого гена или из любой последовательности нуклеиновой кислоты, способной кодировать ортолог, паралог или гомолог рассматриваемого белка), предпочтительно способного к формированию шпилечной структуры. Инвертированный повтор клонируют в вектор экспрессии, содержащий контрольные последовательности. Последовательность нуклеиновой кислоты некодирующей ДНК (спейсер, например фрагмент участка связывания с матриксом (MAR), интрон, полилинкер и т.д.) расположена между двумя инвертированными нуклеиновыми кислотами, формируя инвертированный повтор. После транскрипции инвертированного повтора формируется химерная РНК с самокомплементарной структурой (частичная или полная). Эту двухцепочечную структуру РНК упоминают как шпилечную РНК (hpРНК). ПрРНК процессируется растением в siPHK, которые включены в РНК-индуцированный комплекс сайленсинга (RISC). RISC затем расщепляет транскрипты мРНК, таким образом существенно уменьшая число транскриптов мРНК, транслируемых в полипептиды. Для дополнительных общих подробностей см., например, Grierson et al. (1998) WO 98/53083; Waterhouse et al. (1999) WO 99/53050).
Выполнение способов по изобретению не основывается на введении и экспрессии в растении генетической конструкции, в которую нуклеиновая кислота клонирована в виде инвертированного повтора, но любой один или более из нескольких хорошо известных способов "сайленсинга генов" может использоваться для достижения таких же результатов.
Один такой метод уменьшения экспрессии эндогенного гена представляет собой РНК-опосредованный сайленсинг экспрессии гена (негативная регуляция). Сайленсинг в этом случае запускается в растении последовательностью двухцепочечной РНК (dsRNA), которая по существу схожа с эндогенным геном-мишенью. Эта dsPHK затем процессируется растением в последовательности от около 20 до около 26 нуклеотидов, названных малыми интерферирующими РНК (siPHK). siPHK включены в РНК-индуцированный комплекс сайленсинга (RISC), который расщепляет транскрипт мРНК эндогенного гена-мишени, таким образом по существу уменьшая число транскриптов мРК, транслируемых в полипептид. Предпочтительно, последовательность двухцепочечной РНК соответствует гену-мишени.
Другой пример метода РНК-сайленсинга включает введение последовательностей нуклеиновых кислот или их частей (в этом случае участок по существу смежных нуклеотидов, полученных из рассматриваемого гена или из любой последовательности нуклеиновой кислоты, способной кодировать ортолог, паралог или гомолог рассматриваемого белка) в смысловой ориентации в растение. "Смысловая ориентация" относится к ДНК-последовательности, которая гомологична ее транскрипту мРНК. Следовательно, в растение была бы введена по меньшей мере одна копия последовательности нуклеиновой кислоты. Последовательность дополнительной нуклеиновой кислоты будет уменьшать экспрессию эндогенного гена, вызывая явление, известное как косупрессия. Уменьшение экспрессии гена будет более выраженное, если несколько дополнительных копий последовательности нуклеиновой кислоты будет введено в растение, поскольку существует положительная корреляция между высокими уровнями транскриптов и запуском косупрессии.
Другой пример метода РНК-сайленсинга включает применение антисмысловых последовательностей нуклеиновых кислот."Антисмысловая" последовательность нуклеиновой кислоты содержит нуклеотидную последовательность, которая комплементарна "смысловой" последовательности нуклеиновой кислоты, кодирующей белок, т.е. комплементарна кодирующей цепи молекулы двухцепочечной кДНК или комплементарна последовательности транскрипта мРНК. Антисмысловая последовательность нуклеиновой кислоты предпочтительно комплементарна «выключенному» эндогенному гену. Комплементарность может располагаться в "кодирующей области" и/или в "некодирующей области" гена. Термин "кодирующая область" относится к области нуклеотидной последовательности, содержащей кодоны, которые транслируются в аминокислотные остатки. Термин "некодирующая область" относится к 5'- и 3'- последовательностям, которые фланкируют кодирующую область и которые транскрибируются, но не транслируются в аминокислоты (их также называют 5'- и 3'-нетранслируемыми областями).
Антисмысловые последовательности нуклеиновых кислот могут быть сконструированы согласно правилам спаривания оснований Уотсона и Крика. Антисмысловая последовательность нуклеиновой кислоты может быть комплементарна полной последовательности нуклеиновой кислоты (в этом случае участок по существу смежных нуклеотидов, полученных из рассматриваемого гена или из любой последовательности нуклеиновой кислоты, способной кодировать ортолог, паралог или гомолог рассматриваемого белка), но может быть также олигонуклеотидом, который является антисмысловым относительно только части последовательности нуклеиновой кислоты (включая мРНК 5' и 3' UTR). Например, антисмысловая олигонуклеотидная последовательность может быть комплементарной области, окружающей сайт инициации трансляции транскрипта мРНК, кодирующего полипептид. Длина подходящей антисмысловой олигонуклеотидной последовательности известна в данной области техники и может начинаться от около 50, 45, 40, 35, 30, 25, 20, 15 или 10 нуклеотидов в длину или менее. Антисмысловая последовательность нуклеиновой кислоты согласно изобретению может быть сконструирована с использованием реакций химического синтеза и ферментативного лигирования, применяя способы, известные в данной области техники. Например, антисмысловая последовательность нуклеиновой кислоты (например, антисмысловая олигонуклеотидная последовательность) может быть химически синтезирована с использованием встречающихся в природе нуклеотидов или различным образом модифицированных нуклеотидов, сконструированных для увеличения биологической стабильности молекул или для увеличения физической стабильности дуплекса, сформированного между антисмысловой и смысловой последовательностями нуклеиновых кислот, например, могут использоваться фосфоротиоатные производные и замещенные акридином нуклеотиды. Примеры модифицированных нуклеотидов, которые могут быть использованы для получения антисмысловых последовательностей нуклеиновых кислот, хорошо известны в данной области техники. Известные нуклеотидные модификации включают метилирование, циклизацию и «кэпы» и замещение одного или более встречающихся в природе нуклеотидов аналогом, таким как инозин. Другие модификации нуклеотидов хорошо известны в данной области техники.
Антисмысловая последовательность нуклеиновой кислоты может быть получена биологически с использованием вектора экспрессии, в который последовательность нуклеиновой кислоты субклонирована в антисмысловой ориентации (т.е. РНК, транскрибированная с инсертированной нуклеиновой кислотой, будет иметь антисмысловую ориентацию относительно рассматриваемой нуклеиновой кислоты-мишени). Предпочтительно, получение антисмысловых последовательностей нуклеиновых кислот у растений происходит посредством стабильно интегрированной конструкции нуклеиновой кислоты, содержащей промотор, функционально связанный антисмысловой олигонуклеотид и терминатор.
Молекулы нуклеиновых кислот, использованные для сайленсинга в способах по изобретению (или введенные в растение, или генерированные in situ), гибридизуются или связываются с транскриптами мРНК и/или геномной ДНК, кодирующей полипептид, чтобы посредством этого ингибировать экспрессию белка, например, путем ингибирования транскрипции и/или трансляции. Гибридизация может происходить вследствие стандартной нуклеотидной комплементарности с формированием стабильного дуплекса или, например, в случае антисмысловой последовательности нуклеиновой кислоты, которая связывается с дуплексами ДНК, через специфические взаимодействия в большой бороздке двойной спирали. Антисмысловые последовательности нуклеиновых кислот могут быть введены в растение трансформацией или прямой инъекцией в определенный участок ткани. Альтернативно, антисмысловые последовательности нуклеиновых кислот могут быть модифицированы для выбранных клеток-мишеней и затем введены системно. Например, для системного введения антисмысловые последовательности нуклеиновых кислот могут быть модифицированы таким образом, что они специфически связываются с рецепторами или антигенами, экспрессированными на выбранной клеточной поверхности, например, путем связывания антисмысловой последовательности нуклеиновой кислоты с пептидами или антителами, которые присоединены к рецепторам или антигенам клеточной поверхности. Антисмысловые последовательности нуклеиновых кислот могут также доставляться в клетки, используя векторы, описанные здесь.
Согласно дополнительному аспекту антисмысловая последовательность нуклеиновой кислоты является последовательностью α-аномерной нуклеиновой кислоты. Последовательность α-аномерной нуклеиновой кислоты образует специфические двухцепочечные гибриды с комплементарной РНК, в которых, в противоположность обычным β-единицам, цепи идут параллельно друг другу (Gaultier et al. (1987) Nucl Ac Res 15: 6625-6641). Антисмысловая последовательность нуклеиновой кислоты может также содержать 2'-о-метилрибонуклеотид (Inoue et al. (1987) Nucl Ac Res 15, 6131-6148) или химерный аналог РНК-ДНК (Inoue et al. (1987) FEBS Lett. 215, 327-330).
Уменьшение или значительное устранение экспрессии эндогенного гена может быть также осуществлено с использованием рибозимов. Рибозимы представляют собой каталитические молекулы РНК с рибонуклеазной активностью, которые способны расщеплять одноцепочечную последовательность нуклеиновой кислоты, такую как мРНК, к которой они имеют комплементарную область. Таким образом, рибозимы (например, рибозимы типа головки молотка (описанные у Haselhoff and Gerlach (1988) Nature 334, 585-591) могут использоваться для каталитического расщепления транскриптов мРНК, кодирующих полипептид, таким образом по существу уменьшая количество транскриптов мРНК, транслируемых в полипептид. Рибозим, имеющий специфичность к последовательности нуклеиновой кислоты, может быть сконструирован (см., например, Cech et al. Патент США 4,987,071; и Cech et al. патент США 5,116,742). Альтернативно, транскрипты мРНК, соответствующие последовательности нуклеиновой кислоты, могут использоваться для отбора каталитической РНК, имеющей специфическую рибонуклеазную активность, из пула молекул РНК (Bartel and Szostak (1993) Science 261, 1411-1418). Применение рибозимов для сайленсинга генов в растениях известно в данной области техники (например, Atkins et al. (1994) WO 94/00012; Lenne et al. (1995) WO 95/03404; Lutziger et al. (2000) WO 00/00619; Prinsen et al. (1997) WO 97/13865 и Scott et al. (1997) WO 97/38116).
Сайленсинг генов может также достигаться инсерционным мутагенезом (например, вставкой Т-ДНК или вставкой транспозона) или путем стратегий, как описано, среди прочих, Angell и Baulcombe ((1999) Plant J 20(3):357-62), (Amplicon VIGS WO 98/36083) или Baulcombe (WO 99/15682).
Сайленсинг гена может также происходить, если существует мутация на эндогенном гене и/или мутация на выделенном гене/нуклеиновой кислоте, впоследствии введенных в растение. Уменьшение или значительное устранение могут вызываться нефункциональным полипептидом. Например, полипептид может связываться с различными взаимодействующими белками; одна(о) или более мутаций и/или усечений могут, следовательно, обеспечить полипептид, который еще способен связываться с взаимодействующими белками (такими, как рецепторные белки), но который не может проявлять свою нормальную функцию (например, сигнальный лиганд).
Дополнительный подход к сайленсингу генов заключается в нацеливании последовательностей нуклеиновых кислот, комплементарных к регуляторной области гена (например, промотору и/или энхансерам), с образованием тройных спиральных структур, которые препятствуют транскрипции гена в клетках-мишенях. См. Helene, С., Anticancer Drug Res. 6, 569-84, 1991; Helene et al., Ann. N.Y. Acad. Sci. 660, 27-36 1992; и Maher, L.J. Bioassays 14, 807-15, 1992.
Другие способы, такие как применение антител, направленных против эндогенного полипептида для ингибирования его функции в растении, или вмешательство в сигнальный путь, в который вовлечен полипептид, могут быть хорошо известны специалисту в данной области техники. В частности, можно представить, что искусственные молекулы могут использоваться для ингибирования биологической функции полипептида-мишени или для вмешательства в сигнальный путь, в который вовлечен полипептид-мишень.
Альтернативно, можно установить программу скрининга для идентификации в популяции растений природных вариантов гена, причем эти варианты кодируют полипептиды с уменьшенной активностью. Такие природные варианты могут также использоваться, например, для выполнения гомологичной рекомбинации.
Могут использоваться искусственные и/или природные микроРНК (miPHK) для выключения экспрессии гена и/или трансляции мРНК. Эндогенные miPHK являются одноцепочечными небольшими РНК обычно длиной 19-24 нуклеотидов. В основном они функционируют для регуляции экспрессии генов и/или трансляции мРНК. Большинство растительных микроРНК (miRNA) имеют полную или почти полную комплементарность с их последовательностями-мишенями. Однако существуют природные мишени, имеющие до 5 несовпадений. Они процессируются с более длинных некодирующих РНК с характерными fold-back структурами при помощи двухцепочечных специфических РНКаз семейства Dicer. После процессинга они включаются в РНК-индуцированный комплекс сайленсинга (RISC), связываясь с его основным компонентом белком Argonaute. MiPHK служат компонентами специфичности RISC, так как они спариваются с нуклеиновыми кислотами-мишенями, главным образом мРНК, в цитоплазме. Последующие регуляторные события включают расщепление мРНК-мишени и деструкцию и/или ингибирование трансляции. Результаты сверхэкспрессии miPHK таким образом часто отражаются в уменьшенных уровнях мРНК генов-мишеней.
Искусственные микроРНК (amiPHK), которые обычно составляют 21 нуклеотид в длину, могут быть созданы методами генетической инженерии для негативной регуляции экспрессии генов одного или множества рассматриваемых генов. Определяющие факторы целевой селекции микроРНК растений хорошо известны в данной области техники. Эмпирические параметры целевой идентификации определены и могут использоваться, помогая при конструировании специфических amiPHK (Schwab et al., (2005) Dev. Cell 8(4):517-27). Подходящие средства для конструирования и образования amiPHK и их предшественников также доступны общественности (Schwab et al., (2006) Plant Cell 18(5):1121-33).
Для оптимального выполнения методики сайленсинга генов, используемые для уменьшения экспрессии у растений эндогенного гена, требуют применения последовательностей нуклеиновых кислот из однодольных растений для трансформации однодольных растений и из двудольных растений для трансформации двудольных растений. Предпочтительно, последовательность нуклеиновой кислоты из любого данного вида растений вводится тому же самому виду. Например, последовательность нуклеиновой кислоты из риса трансформируется в рисовое растение. Однако это не является абсолютным требованием, что вводимая последовательность нуклеиновой кислоты происходит из того же вида растений, как и растение, в которое она будет вводиться. Является достаточным, если существует значительная гомология между эндогенным геном-мишенью и вводимой последовательностью нуклеиновой кислоты.
Вышеописанное представляет собой примеры различных способов уменьшения или значительного устранения экспрессии у растений эндогенного гена. Специалист в данной области техники был бы способен легко адаптировать вышеуказанные способы сайленсинга с тем, чтобы достигнуть уменьшения экспрессии эндогенного гена у целого растения или его частей путем применения, например, соответствующего промотора.
Селектируемый маркер (ген)/Репортерный ген
"Селектируемый маркер", "Селектируемый маркерный ген" или "репортерный ген" включает любой ген, который обеспечивает фенотип клетке, в которой он экспрессируется, облегчающий идентификацию и/или отбор клеток, которые трансфицируются или трансформируются конструкцией последовательности нуклеиновой кислоты по изобретению. Эти маркерные гены дают возможность идентификации удачного переноса молекул последовательностей нуклеиновой кислоты через серию различных начал (principles). Подходящие маркеры могут быть выбраны из маркеров, которые придают устойчивость к антибиотикам или гербицидную устойчивость, которые вводят новый метаболический признак или позволяют визуальный отбор. Примеры генов селектируемых маркеров включают гены, обеспечивающие устойчивость к антибиотикам (например, nptll, который фосфорилирует неомицин и канамицин, или hpt, фосфорилирующий гидромицин, или гены, обеспечивающие устойчивость к, например, блеомицину, стрептомицину, тетрациклину, хлорамфениколу, ампициллину, гентамицину, генетицину (G418), спектиномицину или бластицидину), к гербицидам (например, bar, который обеспечивает устойчивость к Basta®; aroA или дох, обеспечивающий устойчивость к глифозату, или гены, придающие устойчивость, например, к имидазолинону, фосфинотрицину или сульфонилмочевине), или гены, которые обеспечивают метаболический признак (например, manA, который позволяет растениям использовать маннозу как единственный источник углерода, или ксилоза изомераза для утилизации ксилозы, или антипитательные маркеры, такие как устойчивость к 2-дезоксиглюкозе). Экспрессия визуальных маркерных генов приводит к образованию окраски (например, β-глюкуронидаза, GUS или β-галактозидаза с их окрашенными субстратами, например, X-Gal), люминесценции (например, система люциферин/люцефераза) или флуоресценции (зеленый флуоресцентный белок, GFP и его производные). Этот перечень представляет только небольшое число возможных маркеров. Специалист в данной области техники знаком с такими маркерами. Разные маркеры являются предпочтительными в зависимости от организма и метода отбора.
Известно, что в результате стабильной или временной интеграции последовательностей нуклеиновых кислот в растительные клетки только меньшая часть клеток принимает чужеродную ДНК и, при желании, интегрирует ее в свой геном в зависимости от использованного вектора экспрессии и использованной методики трансфекции. Для определения и отбора таких интегрантов ген, кодирующий селектируемый маркер (такой, как описанные выше), обычно вводится в клетки-хозяева вместе с рассматриваемым геном. Эти маркеры могут, например, использоваться в мутантах, в которых эти гены не являются функциональными в результате, например, делеции стандартными способами. Кроме того, молекулы нуклеиновых кислот, кодирующие селектируемый маркер, могут вводиться в клетку-хозяина в том же векторе, который содержит последовательность, кодирующую полипептиды по изобретению или использованную в способах по изобретению, или же в отдельном векторе. Клетки, которые стабильно трансфицированы введенной последовательностью нуклеиновых кислот, могут быть идентифицированы, например, отбором (например, клетки, которые интегрировали селектируемый маркер, выживают, тогда как другие клетки умирают).
Поскольку маркерные гены, в частности гены устойчивости к антибиотикам и гербицидам, не требуются больше или являются нежелательными в трансгенной клетке-хозяине, когда последовательности нуклеиновых кислот успешно введены, способ согласно изобретению введения последовательностей нуклеиновых кислот преимущественно применяет методики, которые делают возможным удаление или вырезание этих маркерных генов. Один такой метод известен как совместная трансформация. Метод совместной трансформации применяет одновременно два вектора для трансформации, при этом один вектор несет последовательность нуклеиновой кислоты согласно изобретению, а второй вектор несет маркерный(е) ген(ы). Большая часть трансформантов получает или, в случае растений, содержит (до 40% или более трансформантов) оба вектора. В случае трансформации агробактериями трансформанты обычно получают только часть вектора, то есть последовательность, фланкированную Т-ДНК, которая обычно представляет экспрессионную кассету. Маркерные гены могут быть затем удалены из трансформированного растения проведением скрещиваний. В другом способе маркерные гены, интегрированные в транспозон, используются для трансформации вместе с желательной последовательностью нуклеиновой кислоты (известен как технология Ac/Ds). Трансформанты могут скрещивать с источником транспозазы, или трансформанты трансформируют конструкцией последовательности нуклеиновой кислоты, предоставляющей экспрессию транспозазы, временно или стабильно. В нескольких случаях (прибл. 10%) транспозон «выпрыгивает» из генома клетки-хозяина после того, как трансформация прошла удачно, и теряется. В дополнительном ряде случаев транспозон «перепрыгивает» в другое место. В этих случаях маркерный ген должен устраняться проведением скрещиваний. В микробиологии разработаны методы, которые делают возможным или облегчают обнаружение таких случаев. Дополнительный полезный способ основывается на том, что известно как рекомбинационные системы; их преимущество заключается в том, что можно обойтись без устранения путем скрещивания. Самая известная система такого типа известна как система Cre/lox. Cre1 означает рекомбиназу, которая удаляет последовательности, расположенные между последовательностями loxP. Если маркерный ген интегрирован между последовательностями loxP, он удаляется после того, как трансформация произошла успешно, путем экспрессии рекомбиназы. Дополнительными рекомбинационными системами являются системы HIN/HIX, FLP/FRT и REP/STB (Tribble et al., J. Biol. Chem, 275, 2000: 22255-22267; Velmurugan et al, J. Cell Biol., 149, 2000:553-566). Возможна сайт-специфическая интеграция в геном растения последовательностей нуклеиновых кислот согласно изобретению. Естественно, эти способы могут также быть применимы к микроорганизмам, таким как дрожжи, грибы или бактерии.
Трансгенный/Трансген/Рекомбинантный
Для целей настоящего изобретения "трансгенный", "трансген" или "рекомбинантный" означают в отношении, например, последовательности нуклеиновой кислоты экспрессионную кассету, генную конструкцию или вектор, содержащие последовательность нуклеиновой кислоты, или организм, трансформированный последовательностями нуклеиновых кислот, экспрессионными кассетами или векторами согласно изобретению, при этом все эти конструкции получены рекомбинантными методами, в которых или
(a) последовательности нуклеиновых кислот, кодирующие белки, пригодные в способах изобретения, или
(b) генетическая(ие) контрольная(ые) последовательность(и), которые функционально связаны с последовательностью нуклеиновой кислоты согласно изобретению, например промотор, или
(c) а) и b)
не расположены в их природном генетическом окружении или модифицированы рекомбинантными методами, и модификация может принимать форму, например, замены, добавления, делеции, инверсии или вставки одного или более нуклеотидных остатков. Природное генетическое окружение понимается как означающее природный геномный или хромосомный локус в первоначальном растении или присутствие в геномной библиотеке. В случае геномной библиотеки природное генетическое окружение последовательности нуклеиновой кислоты предпочтительно сохраняется по меньшей мере частично. Окружение фланкирует последовательность нуклеиновой кислоты по меньшей мере с одной стороны и имеет длину последовательности по меньшей мере 50 п.н., предпочтительно 500 п.н., особенно предпочтительно по меньшей мере 1000 п.н., особенно предпочтительно по меньшей мере 5000 п.н. Встречающая в природе экспрессионная кассета, например встречающаяся в природе комбинация природного промотора последовательностей нуклеиновых кислот с соответствующей последовательностью нуклеиновой кислоты, кодирующей полипептид, используемой в способах настоящего изобретения, как определено выше, - становится трансгенной экспрессионной кассетой, когда эта кассета модифицирована неприродными, синтетическими ("искусственными") способами, такими как, например, мутагенная обработка. Подходящие способы описаны, например, в US 5,565,350 или WO 00/15815.
Таким образом, трансгенное растение для целей данного изобретения понимается как означающее, по вышесказанному, что последовательности нуклеиновых кислот, использованные в способе изобретения, не находятся на их природном локусе генома указанного растения, при этом последовательности нуклеиновых кислот могут экспрессироваться гомологично или гетерологично. Однако, как упомянуто, "трансгенный" также означает, что в то время как последовательность нуклеиновой кислоты согласно изобретению или использованная в способе по изобретению находятся в своем природном положении в геноме растения, последовательность модифицирована относительно природной последовательности, и/или что регуляторные последовательности природных последовательностей модифицированы. "Трансгенный" предпочтительно понимается как означающий экспрессию последовательностей нуклеиновых кислот согласно изобретению в неприродном локусе генома, т.е. происходит гомологичная или предпочтительно гетерологичная экспрессия последовательностей нуклеиновых кислот. Предпочтительные трансгенные растения здесь упомянуты.
Трансформация
Термин "введение" или "трансформация" при упоминании здесь охватывает перенос экзогенного полинуклеотида в клетку-хозяина, независимо от способа, использованного для переноса. Ткань растения, способная к последующему клональному размножению, путем органогенеза или эмбриогенеза, может трансформироваться генетической конструкцией по настоящему изобретению, и из нее регенерируется целое растение. Выбираемая специфическая ткань может изменяться в зависимости от систем клонального размножения, доступных для или наиболее подходящих к трансформируемому специфическому виду. Типичные ткани-мишени включают листовые диски, пыльцу, зародыши, семядоли, гипокотили, макрогаметофиты, каллусную ткань, существующую меристематическую ткань (например, апикальная меристема, пазушные почки и корневые меристемы) и индуцированную меристематическую ткань (например, меристема семядолей и меристема гипокотилей). Полинуклеотид может быть временно или стабильно введен в клетку-хозяина и может поддерживаться неинтегрированным, например, как плазмида. Альтернативно, он может интегрироваться в геном хозяина. Получившаяся трансформированная растительная клетка может затем использоваться для регенерации трансформированного растения известным специалисту в данной области техники образом.
Перенос чужеродных генов в геном растения называется трансформацией. Трансформация видов растений в настоящее время является довольно рутинной методикой. Преимущественно любой из нескольких способов трансформации может использоваться для введения рассматриваемого гена в подходящую исходную клетку. Способы, описанные для трансформации и регенерации растений из растительных тканей или растительных клеток, могут использоваться для временной или стабильной трансформации. Способы трансформации включают применение липосом, электропорацию, химикаты, увеличивающие поглощение свободной ДНК, инъекцию ДНК непосредственно в растение, бомбардировку частицами с помощью «генной пушки», трансформацию с использованием вирусов или пыльцы и микропроекцию (microprojection). Способы могут выбираться из кальциевого/полиэтиленгликолевого способа для протопластов (Krens, F.A. et al., (1982) Nature 296, 72-74; Negrutiu I et al. (1987) Plant Mol Biol 8:363-373); электропорации протопластов (Shillito R.D. et al. (1985) Bio/Technol 3, 1099-1102); микроинъекции в растительный материал (Crossway A et al., (1986) Mol. Gen Genet 202:179-185); бомбардировки частицами, покрытыми ДНК или РНК (Klein TM et al., (1987) Nature 327:70); заражения (неинтегративными) вирусами и т.п. Трансгенные растения, включая трансгенные сельскохозяйственные культуры, предпочтительно получают путем Agrobacterium-опосредованной трансформации. Предпочтительным способом трансформации является трансформация in planta. С этой целью можно, например, позволить агробактериям воздействовать на семена растений или инокулировать меристему растений агробактериями. Оказалось особенно целесообразно в соответствии с изобретением позволить суспензии трансформированных агробактерий действовать на интактное растение или по меньшей мере на примордий цветка. Растение затем выращивают до тех пор, пока не получают семена обработанного растения (Clough and Bent, Plant J. (1998) 16, 735-743). Способы Agrobacterium-опосредованной трансформации риса включают хорошо известные способы трансформации риса, такие как описанные в любом из следующих источников: Европейская патентная заявка ЕР 1198985 А1, Aldemita and Hodges (Planta 199:612-617, 1996); Chan et al. (Plant Mol Biol 22 (3):491-506, 1993), Hiei et al. (Plant J 6 (2):271-282, 1994), раскрытия которых включены здесь путем ссылки, как если бы были полностью изложены. В случае трансформации кукурузы предпочтительным способом является или описанный Ishida et al. (Nat. Biotechnol 14(6):745-50, 1996), или Frame et al. (Plant Physiol 129(1):13-22, 2002), раскрытия которых включены здесь путем ссылки, как если бы были полностью изложены. Указанные способы также описаны, к примеру, у В. Jenes et al., Techniques for Gene Transfer, in: Transgenic Plants, Vol.1, Engineering and Utilization, eds. S.D. Kung and R. Wu, Academic Press (1993) 128-143 и в Potrykus Annu. Rev. Plant Physiol. Plant Molec. Biol. 42 (1991) 205-225. Экспрессируемые последовательности нуклеиновых кислот или конструкция предпочтительно клонируются в вектор, который подходит для трансформации Agrobacterium tumefaciens, например pBin19 (Bevan et al., Nucl. Acids Res. 12 (1984) 8711). Агробактерии, трансформированные таким вектором, могут затем использоваться известным образом для трансформации растений, например растений, используемых как модель, подобно Arabidopsis (Arabidopsis thaliana в объеме настоящего изобретения не рассматривается как сельскохозяйственная культура), или сельскохозяйственных культур, таких как, к примеру, табак, например, путем погружения мятых или порезанных листьев в агробактериальный раствор и затем культивирования их в подходящей среде. Трансформация растений посредством Agrobacterium tumefaciens описана, например, у Hofgen и Willmitzer в Nucl. Acid Res. (1988) 16, 9877, или известна среди прочего из F.F. White, Vectors for Gene Transfer in Higher Plants; in Transgenic Plants, Vol.1, Engineering and Utilization, eds. S.D. Kung and R. Wu, Academic Press, 1993, pp.15-38.
Дополнительно к трансформации соматических клеток, которые затем должны регенерироваться в интактные растения, также можно трансформировать клетки меристем растений и в частности те клетки, которые развиваются в гаметы. В этом случае трансформированные гаметы следуют естественному развитию растения, приводя к трансгенным растениям. Таким образом, например, семена Arabidopsis обрабатываются агробактериями, и из развивающихся растений получают семена, из которых определенная часть трансформирована и таким образом является трансгенной [Feldman, KA and Marks MD (1987). Mol Gen Genet 208:274-289; Feldmann K (1992). In: С Koncz, N-H Chua and J Shell, eds, Methods in Arabidopsis Research. Word Scientific, Singapore, pp.274-289]. Альтернативные способы основаны на повторяющемся удалении соцветий и инкубировании вырезанного участка в центре розетки с трансформированными агробактериями, посредством чего трансформированные семена могут быть также получены в более поздний момент времени (Chang (1994). Plant J. 5:551-558; Katavic (1994). Mol Gen Genet, 245:363-370). Однако особенно эффективным способом является метод вакуумной инфильтрации с его модификациями, такими как метод "погружения цветков". В случае вакуумной инфильтрации Arabidopsis интактные растения при пониженном давлении обрабатывают агробактериальной суспензией [Bechthold, N (1993). С R Acad Sci Paris Life Sci, 316:1194-1199], в то время как в случае метода "погружения цветков" ткань развивающихся цветков в течение короткого периода инкубируют с агробактериальной суспензией, обработанной поверхностно-активным веществом [Clough, SJ and Bent AF (1998) The Plant J. 16, 735-743]. Определенную часть трансгенных семян получали в обоих случаях, и эти семена можно отличить от нетрансгенных семян при выращивании в вышеуказанных селективных условиях. Кроме того, стабильная трансформация пластид является удобной, так как пластиды наследуются по материнской линии в большинстве сельскохозяйственных культур, снижая или устраняя риск «утечки» трансгена через пыльцу. Трансформация генома хлоропластов, как правило, выполняется способом, который схематически показан у Klaus et al., 2004 [Nature Biotechnology 22 (2), 225-229]. Вкратце, подлежащие трансформации последовательности клонируют вместе с селектируемым маркером между фланкирующими последовательностями, гомологичными геному хлоропластов. Эти гомологичные фланкирующие последовательности направляют сайт-специфичную интеграцию в пластом. Пластидная трансформация описана для многих различных видов растений, и обзор представлен у Bock (2001) Transgenic plastids in basic research and plant biotechnology. J Mol Biol. 2001 Sep 21; 312 (3):425-38 или Maliga, P (2003) Progress towards commercialization of plastid transformation technology. Trends Biotechnol. 21 20-28. Недавно сообщали о дальнейшем биотехнологическом прогрессе в виде безмаркерных пластидных трансформантов, которые могут быть получены при помощи временного совместно интегрированного маркерного гена (Klaus et al., 2004, Nature Biotechnology 22(2), 225-229).
Активационное мечение Т-ДНК
Активационное мечение Т-ДНК (Hayashi et al. Science (1992) 1350-1353) включает вставку Т-ДНК, обычно содержащей промотор (может быть также усилитель трансляции или интрон) в геномную область рассматриваемого гена или выше, или ниже на 10 т.п.н. кодирующей области гена в такой конфигурации, что промотор направляет экспрессию целевого гена. Как правило, регуляция экспрессии целевого гена его природным промотором нарушается, и ген попадает под контроль нововведенного промотора. Промотор обычно вставлен в Т-ДНК. Эта Т-ДНК случайным образом вставляется в геном растения, например, путем заражения агробактериями, и приводит к модифицированной экспрессии генов около вставленной Т-ДНК. Полученные трансгенные растения демонстрируют доминантные фенотипы благодаря модифицированной экспрессии генов рядом с введенным промотором.
TILLING
Термин "TILLING" означает аббревиатуру "направленные индуцированные локальные повреждения в геномах" и относится к технологии мутагенеза, пригодной для получения и/или определения последовательностей нуклеиновых кислот, кодирующих белки с модифицированной экспрессией и/или активностью. TILLING также позволяет отбор растений, несущих такие мутантные варианты. Эти мутантные варианты могут проявлять модифицированную экспрессию с точки зрения или силы, или расположения, или тайминга (если мутации влияют на промотор, например). Эти мутантные варианты могут проявлять более высокую активность, чем проявляемая геном в его природной форме. TILLING комбинирует мутагенез высокой плотности (high-density mutagenesis) с методами высокопроизводительного скрининга. Как правило, в TILLING следующие стадии: (а) EMS-мутагенез (мутагенез с помощью этан метилсульфоната) (Redei GP and Koncz С (1992) In Methods in Arabidopsis Research, Koncz C, Chua NH, Schell J, eds. Singapore, World Scientific Publishing Co, pp.16-82; Feldmann et al., (1994) In Meyerowitz EM, Somerville CR, eds, Arabidopsis. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp 137-172; Lightner J and Caspar Т (1998) In J Martinez-Zapater, J Salinas, eds, Methods on Molecular Biology, Vol.82. Humana Press, Totowa NJ, pp 91-104); (b) получение ДНК и объединение в пул представителей; (с) ПЦР-амплификация рассматриваемого участка; (d) денатурация и отжиг с образованием гетеродуплексов; (е) денатурирующая высокоэффективная жидкостная хроматография (DHPLC), где присутствие гетеродуплекса в пуле обнаруживается как дополнительный пик на хроматограмме; (f) идентификация мутантного представителя; и (g) секвенирование мутантного ПЦР-продукта. Способы TILLING хорошо известны в данной области техники (McCallum et al., (2000) Nat Biotechnol 18:455-457; reviewed by Stemple (2004) Nat Rev Genet 5(2):145-50).
Гомологичная рекомбинация
Гомологичная рекомбинация обеспечивает введение в геном выбранной последовательности нуклеиновой кислоты в определенное выбранное положение. Гомологичная рекомбинация является стандартной технологией, обычно используемой в биологических науках для низших организмов, таких как дрожжи или мох Physcomitrella. Способы проведения гомологичной рекомбинации у растений описаны не только для растений-моделей (Offringa et al. (1990) EMBO J 9(10):3077-84), но также для сельскохозяйственных культур, например риса (Terada et al. (2002) Nat Biotech 20(10):1030-4; lida and Terada (2004) Curr Opin Biotech 15(2):132-8), и существуют подходы, которые, как правило, применимы независимо от организма-мишени (Miller et al, Nature Biotechnol. 25, 778-785, 2007).
Урожайность
Термин "урожайность" в общем означает измеримый продукт с экономической ценностью, обычно зависимый от определенной сельскохозяйственной культуры, площади и периода времени. Отдельные части растения непосредственно делают вклад в урожайность на основании их количества, размера и/или массы, или фактическую урожайность - это урожайность на квадратный метр для сельскохозяйственной культуры за год, которая определяется делением общей продукции (включает и собранную, и оцененную продукцию) на засаженные квадратные метры. Термин "урожайность" растения может относиться к вегетативной биомассе (биомассе корней и/или побегов), репродуктивным органам растений и/или растительным материалам, используемым для размножения (таким, как семена).
Ранняя мощность
"Ранняя мощность" относится к активному, здоровому, хорошо сбалансированному росту, особенно во время ранних стадий роста растений, и может являться результатом увеличенной приспособленности растения благодаря тому, например, что растения лучше адаптированы к их окружающей среде (т.е. оптимизируют использование источников энергии и распределение между побегом и корнем). Растения, имеющие раннюю мощность, также демонстрируют повышенное выживание сеянцев и лучшую приживаемость сельскохозяйственной культуры, что часто приводит к высокооднородным полям (с сельскохозяйственной культурой, растущей однородным образом, т.е. с большинством растений, достигающих различных стадий развития по существу за одно и то же время), и часто лучшую и более высокую урожайность. Следовательно, ранняя мощность может определяться измерением различных факторов, таких как масса тысячи зерен, всхожесть в процентах, прорастание в процентах, рост сеянцев, высота сеянцев, длина корней, биомасса корней и побегов и многое другое.
Увеличение/Улучшение/Усиление
Термины "увеличение", "улучшение" или "усиление" являются взаимозаменяемыми и могут означать в ключе изобретения по меньшей мере на 5%, 6%, 7%, 8%, 9% или 10%, предпочтительно по меньшей мере 15% или 20%, более предпочтительно 25%, 30%, 35% или 40% больше урожайности и/или роста по сравнению с контрольными растениями, как здесь определено.
Урожайность семян
Увеличенная урожайность семян может проявлять себя как одно из следующего: а) увеличение биомассы семян (общая масса семян), которая может быть отдельно для семян, и/или на растение, и/или на гектар или акр; b) увеличенное число цветков на метелку и/или на растение; с) увеличенное число (наполненных) семян; d) увеличенный уровень наполнения семян (который выражается как отношение числа наполненных семян к общему числу семян); е) увеличенный индекс урожайности, который выражается как отношение урожайности заготавливаемых частей, таких как семена, к общей биомассе; f) увеличенное число первичных метелок; g) увеличенная масса тысячи зерен (TKW), которая экстраполируется из числа подсчитанных наполненных семян и их общей массы. Увеличенная TKW может являться результатом увеличенного размера семян и/или массы семян и может также являться результатом увеличения размера зародыша и/или эндосперма.
Увеличение урожайности семян может также проявлять себя как увеличение размера семян и/или объема семян. Кроме того, увеличение урожайности может также проявлять себя как увеличение площади семян, и/или длины семян, и/или ширины семян, и/или периметра семян. Увеличенная урожайность семян может также приводить к модифицированной структуре или может возникать вследствие модифицированной структуры.
Индекс зелени
"Индекс зелени" при использовании здесь рассчитывается из цифровых изображений растений. Для каждого пикселя, принадлежащего растительному объекту на изображении, рассчитывается соотношение значения зеленого цвета и значения красного цвета (для модели RGB для кодирования цвета). Индекс зелени выражается как процент пикселей, для которых соотношение зеленого и красного превышает заданный порог. В нормальных условиях роста, в условиях роста при солевом стрессе и в условиях роста при пониженной доступности питательных веществ индекс зелени растений измеряется на последнем изображении перед цветением. В отличие от этого в условиях роста при стрессе, вызванном засухой, индекс зелени растений измеряется на первом изображении после засухи.
Растение
Термин "растение" при использовании здесь охватывает целые растения, предков и потомство этих растений и части растений, включая семена, побеги, стебли, листья, корни (включая клубни), цветки, и ткани, и органы, причем каждый из вышеупомянутых содержит рассматриваемые ген/последовательность нуклеиновой кислоты. Термин "растение" также охватывает клетки растений, суспензионные культуры, каллусную ткань, зародыши, меристематические участки, гаметофиты, спорофиты, пыльцу и микроспоры, где в свою очередь каждый из вышеупомянутых содержит рассматриваемые ген/последовательность нуклеиновой кислоты.
Растения, которые особенно полезны в способах по изобретению, включают все растения, которые принадлежат надсемейству Viridiplantae, в частности однодольные и двудольные растения, включая кормовые или фуражные бобовые растения, декоративные растения, продовольственные культуры, деревья или кустарники, выбранные из перечня, содержащего, среди прочих, Acer spp., Actinidia spp., Abelmoschus spp., Agave sisalana, Agropyron spp., Agrostis stolonifera, Allium spp., Amaranthus spp., Ammophila arenaria, Ananas comosus, Annona spp., Apium graveolens, Arachis spp, Artocarpus spp., Asparagus officinalis, Avena spp. (например, Avena sativa, Avena fatua, Avena byzantina, Avena fatua var. sativa, Avena hybrida), Averrhoa carambola, Bambusa sp., Benincasa hispida, Bertholletia excelsea, Beta vulgaris, Brassica spp. (например, Brassica napus, Brassica rapa ssp. канола, масличный рапс, репа масличная), Cadaba farinosa, Camellia sinensis, Canna indica, Cannabis sativa, Capsicum spp., Carex elata, Carica papaya, Carissa macrocarpa, Carya spp., Carthamus tinctorius, Castanea spp., Ceiba pentandra, Cichorium endivia, Cinnamomum spp., Citrullus lanatus, Citrus spp., Cocos spp., Coffea spp., Colocasia esculenta, Cola spp., Corchorus sp., Coriandrum sativum, Corylus spp., Crataegus spp., Crocus sativus, Cucurbita spp., Cucumis spp., Cynara spp., Daucus carota, Desmodium spp., Dimocarpus longan, Dioscorea spp., Diospyros spp., Echinochloa spp., Elaeis (например, Elaeis guineensis, Elaeis oleifera), Eleusine coracana, Erianthus sp., Eriobotrya japonica, Eucalyptus sp., Eugenia uniflora, Fagopyrum spp., Fagus spp., Festuca arundinacea, Ficus carica, Fortunella spp., Fragaria spp., Ginkgo biloba, Glycine spp. (например, Glycine max, Soja hispida или Soya max), Gossypium hirsutum, Helianthus spp. (например, Helianthus annuus), Hemerocallis fulva, Hibiscus spp., Hordeum spp. (например, Hordeum vulgare), Ipomoea batatas, Juglans spp., Lactuca sativa, Lathyrus spp., Lens culinaris, Linum usitatissimum, Litchi chinensis, Lotus spp., Luffa acutangula. Lupinus spp., Luzula sylvatica, Lycopersicon spp. (например, Lycopersicon esculentum, Lycopersicon lycopersicum, Lycopersicon pyriforme), Macrotyloma spp., Malus spp., Malpighia emarginata, Mammea americana, Mangifera indica, Manihot spp., Manilkara zapota, Medicago sativa, Melilotus spp., Mentha spp., Miscanthus sinensis, Momordica spp., Morus nigra, Musa spp., Nicotiana spp., Olea spp., Opuntia spp., Omithopus spp., Oryza spp. (например, Oryza sativa, Oryza latifolia), Panicum miliaceum, Panicum virgatum, Passiflora edulis, Pastinaca sativa, Pennisetum sp., Persea spp., Petroselinum crispum, Phalaris arundinacea, Phaseolus spp., Phleum pratense, Phoenix spp., Phragmites australis, Physalis spp., Pinus spp., Pistacia vera, Pisum spp., Poa spp., Populus spp., Prosopis spp., Prunus spp., Psidium spp., Punica granatum, Pyrus communis, Quercus spp., Raphanus sativus, Rheum rhabarbarum, Ribes spp., Ricinus communis, Rubus spp., Saccharum spp., Salix sp., Sambucus spp., Secale cereale, Sesamum spp., Sinapis sp., Solarium spp. (например, Solanum tuberosum, Solanum integrifolium или Solanum lycopersicum), Sorghum bicolor, Spinacia spp., Syzygium spp., Tagetes spp., Tamarindus indica, Theobroma cacao, Trifolium spp., Triticale sp., Triticosecale rimpaui, Triticum spp. (например, Triticum aestivum, Triticum durum, Triticum turgidum, Triticum hybernum, Triticum macha, Triticum sativum или Triticum vulgare), Tropaeolum minus, Tropaeolum majus, Vaccinium spp., Vicia spp., Vigna spp., Viola odorata, Vitis spp., Zea mays, Zizania palustris, Ziziphus spp.
Подробное описание изобретения
Неожиданно в настоящее время было обнаружено, что увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF, дает растения, имеющие усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями. Согласно первому варианту осуществления настоящее изобретение предоставляет способ усиления признаков, связанных с урожайностью семян, у растений по сравнению с контрольными растениями, предусматривающий увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF.
Предпочтительный способ увеличения экспрессии последовательности нуклеиновой кислоты, кодирующей полипептид GRF, осуществляется путем введения и экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF.
В одном варианте осуществления любая ссылка в дальнейшем на "белок, используемый в способах по изобретению" делается для обозначения полипептида GRF, как здесь определено. Любая ссылка ниже на "последовательность нуклеиновой кислоты, используемую в способах изобретения" делается для обозначения последовательности нуклеиновой кислоты, способной кодировать такой полипептид GRF. Последовательность нуклеиновой кислоты, которую вводят в растение (и, следовательно, используемая при выполнении способов по изобретению), представляет собой любую последовательность нуклеиновой кислоты, кодирующую тип полипептида, который будет сейчас описан, в дальнейшем также именуемую "последовательность нуклеиновой кислоты GRF" или "ген GRF".
"Полипептид GRF", как здесь определено, относится к любому полипептиду, содержащему: (i) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом QLQ, как представлено SEQ ID NO:115; и (ii) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом WRC, как представлено SEQ ID NO:116.
Альтернативно или дополнительно "полипептид GRF", как здесь определено, относится к любому полипептиду, содержащему: (i) домен QLQ с рег. номером IPR014978 в InterPro (рег. номер PF08880 в PFAM); (ii) домен WRC с рег. номером IPR014977 в InterPro (рег. номер PF08879 в PFAM); и (iii) домен эффектора транскрипции (ЕТ), содержащий три Cys и один His остатки в консервативном участке (СХ9СХ10СХ2Н).
Альтернативно или дополнительно "полипептид GRF", как здесь определено, относится к любому полипептиду, имеющему в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с полипептидом GRF, как представлено SEQ ID NO:2, или с любой из полноразмерных полипептидных последовательностей, приведенных здесь в Таблице А.
Альтернативно или дополнительно "полипептид GRF" взаимодействует с полипептидами GRF-взаимодействующих факторов (GIF) (называемыми также полипептидами транслокации синовиальной саркомы (SYT)) в анализе взаимодействия с помощью дрожжевых двойных гибридов.
Неожиданно в настоящее время было обнаружено, что модуляция экспрессии в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, дает растения, имеющие улучшенные признаки, связанные с урожайностью, по сравнению с контрольными растениями. Согласно первому варианту осуществления настоящее изобретение обеспечивает способ улучшения признаков, связанных с урожайностью, в растениях по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид.
Предпочтительный способ модуляции (предпочтительно, увеличения) экспрессии нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, представляет собой введение и экспрессию в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид.
В одном варианте осуществления изобретения любая ссылка в дальнейшем на "белок, используемый в способах по изобретению" делается для обозначения RAA1-подобного полипептида, как здесь определено. Любая ссылка ниже на "последовательность нуклеиновой кислоты, используемую в способах изобретения" делается для обозначения последовательности нуклеиновой кислоты, способной кодировать такой RAA1-подобный полипептид. Последовательность нуклеиновой кислоты, которую вводят в растение (и, следовательно, используемая при выполнении способов по изобретению), представляет собой любую последовательность нуклеиновой кислоты, кодирующую тип белка, который будет сейчас описан, в дальнейшем также именуемую "RAA1-подобной нуклеиновой кислотой" или "RAA1-подобным геном".
"RAA1-подобный полипептид", как определено здесь, относится к любому полипептиду, представленному SEQ ID NO:121, и к его ортологам и паралогам. RAA1-подобные белки являются небольшими (молекулярная масса между 10 и 21 кДа) и основными полипептидами (pl выше 8.5) и обычно не имеют или содержат один Cys остаток в последовательности, которая выравнивается с SEQ ID NO:121, когда используется стандартная программа выравнивания Нидлмана и Вунша с установками по умолчанию.
Предпочтительно, RAA1-подобный полипептид содержит два или более из следующих консервативных мотивов последовательности:
SEQ ID NO:162, мотив 1: GVW(V/L)F
SEQ ID NO:163, мотив 2: LGW(E/S)RY(Y/F)
SEQ ID NO:164, мотив 3: (D/H)L(L/I)S(I/V/L)P(R/K/A)(S/D)F
SEQ ID NO:165, мотив 4: (H/Y)(F/M)YD(V/I)WK(N/T)(R/P)
Альтернативно, гомолог белка RAA1 имеет в возрастающем порядке предпочтения по меньшей мере 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% общей идентичности последовательности с аминокислотной последовательностью, представленной SEQ ID NO:121, при условии, что гомологичный белок содержит консервативные мотивы 1 (a, b, с или d), 2 и 3, и обогащенный лейцином домен, как описано выше. Общую идентичность последовательности определяют, используя алгоритм глобального выравнивания, такой как алгоритм Нидлмана-Вунша в программе GAP (GCG Wisconsin Package, Accelrys), предпочтительно с параметрами по умолчанию.
Предпочтительно, полипептидная последовательность при использовании в построении филогенетического дерева, такого как показано на Фиг.8 (Ge et al., 2004), образует кластеры с группой RAA1-подобных полипептидов, содержащей аминокислотную последовательность, представленную SEQ ID NO:121, а не с любой другой группой.
Неожиданно в настоящее время было обнаружено, что модуляция экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYR, дает растения, которые при выращивании в условиях абиотического стресса имеют улучшенные признаки, связанные с урожайностью, по сравнению с контрольными растениями. Согласно первому варианту осуществления настоящее изобретение предоставляет способ улучшения признаков, связанных с урожайностью, у растений, выращенных в условиях абиотического стресса, по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYR.
Предпочтительный способ модуляции (предпочтительно увеличения) экспрессии нуклеиновой кислоты, кодирующей полипептид SYR, осуществляется путем введения и экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYR.
В одном варианте осуществления любая ссылка в дальнейшем на "белок, используемый в способах по изобретению" делается для обозначения полипептида SYR, как здесь определено. Любая ссылка ниже на "нуклеиновую кислоту, используемую в способах изобретения" делается для обозначения нуклеиновой кислоты, способной кодировать такой полипептид SYR. Нуклеиновая кислота, которую вводят в растение (и, следовательно, используемая при выполнении способов по изобретению), представляет собой любую нуклеиновую кислоту, кодирующую тип белка, который будет сейчас описан, в дальнейшем также именуемую "нуклеиновой кислотой SYR" или "геном SYR".
Термин "белок SYR или его гомолог", как здесь определено, относится к полипептиду от около 65 до около 200 аминокислот, содержащему (i) обогащенный лейцином домен, который похож на лейциновую молнию на C-концевой части белка, при этом перед обогащенным лейцином доменом (ii) находится трипептид с последовательностью YFS (консервативный мотив 5а, SEQ ID NO:173), или YFT (консервативный мотив 5b, SEQ ID NO:174), или YFG (консервативный мотив 5с, SEQ ID NO:175), или YLG (консервативный мотив 5d, SEQ ID NO:176), а после (iii) следует консервативный мотив 6 ((V/A/I)LAFMP(T/S), SEQ ID NO:177). Предпочтительно, консервативный домен 6 представляет собой (A/V)LAFMP(T/S), более предпочтительно консервативный домен представляет собой VLAFMPT. "Белок SYR или его гомолог" предпочтительно также имеет консервативный C-концевой пептид, заканчивающийся консервативным мотивом 7 (SYL или PYL, SEQ ID NO:178). Обогащенный лейцином домен белка SYR или его гомолога составляет от около 38 до 48 аминокислот в длину, начинаясь непосредственно после консервативного мотива 5 и заканчиваясь непосредственно перед консервативным мотивом 6, и содержит по меньшей мере 30% лейцина. Обогащенный Leu домен предпочтительно имеет мотив, который похож на мотив «лейциновая молния» (L-X6-L-X6-L-X6-L, где Х6 представляет собой последовательность из 6 последовательных аминокислот). Предпочтительный пример белка SYR представлен SEQ ID NO:169, общие сведения о его доменах даны на Фиг.11.
Также предпочтительно, белки SYR имеют два трансмембранных домена, при этом N-концевая часть и C-концевая часть белка расположены с внутренней стороны, и часть между трансмембранными доменами расположена с наружной стороны.
Альтернативно, гомолог белка SYR имеет в увеличивающемся порядке предпочтения по меньшей мере 27%, 28%, 29%, 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% общей идентичности последовательности с аминокислотной последовательностью, представленной SEQ ID NO:169, при условии, что гомологичный белок содержит консервативные мотивы 5 (а, b, с or d), 6 и 7, и обогащенный лейцином домен, как описано выше. Общую идентичность последовательности определяют, используя алгоритм глобального выравнивания, такой как алгоритм Нидлмана-Вунша в программе GAP (GCG Wisconsin Package, Accelrys), предпочтительно с параметрами по умолчанию. По сравнению с общей идентичностью последовательности идентичность последовательности будет, как правило, выше, когда рассматриваются только консервативные домены или мотивы.
Неожиданно в настоящее время было обнаружено, что модуляция экспрессии у растения нуклеиновой кислоты, кодирующей полипептид ARKL, дает растения, имеющие улучшенные признаки, связанные с урожаем, по сравнению с контрольными растениями. Согласно первому варианту осуществления настоящее изобретение обеспечивает способ улучшения признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид ARKL.
Предпочтительный способ модуляции (предпочтительно, увеличения) экспрессии нуклеиновой кислоты, кодирующей полипептид ARKL, осуществляется путем введения и экспрессии в растении нуклеиновой кислоты, кодирующей полипептид ARKL.
Любая ссылка в дальнейшем на "белок, используемый в способах по изобретению" делается для обозначения полипептида ARKL, как здесь определено. Любая ссылка ниже на "нуклеиновую кислоту, используемую в способах изобретения" делается для обозначения нуклеиновой кислоты, способной кодировать такой полипептид ARKL. Нуклеиновая кислота, которую вводят в растение (и, следовательно, используемая при выполнении способов по изобретению), представляет собой любую нуклеиновую кислоту, кодирующую тип белка, который будет сейчас описан, в дальнейшем также именуемую "нуклеиновой кислотой ARKL" или "геном ARKL".
"Полипептид ARKL", как здесь определено, относится к любому полипептиду, содержащему консервативный домен цинкового пальца типа RING и при необходимости домен DAR1. Цинковый палец типа RING, обнаруженный в полипептиде ARKL, содержит традиционный тип домена цинкового пальца С3Н2С3.
Он также может быть классифицирован в тип RING-H2 внутри группы I, как определено Stone et al. 2005.
Консенсусная последовательность, представляющая домен RING-H2, описывается как представленная СХ(2)СХ(9-39)СХ(1-3)НХ(2-3)НХ(2)СХ(4-48)СХ(2)С (SEQ ID NO:400). Длина вариабельных петель в полипептидах ARKL составляет обычно 14-15 аминокислот между металлическими лигандами 2 и 3, и 10 аминокислот между металлическими лигандами 6 и 7 (Фиг.1). Специфические аминокислотные остатки, исключая те, которые вовлечены в непосредственное координирование ионов Zn2+, являются высококонсервативными в домене RING-H2 полипептидов ARKL (Фиг.1). SEQ ID NO:401 представляет консенсусную последовательность, консервативную между большинством полипептидов ARKL.
Предпочтительный полипептид ARKL, используемый в способах по изобретению, относится к полипептиду, содержащему RING-домен цинкового пальца ZfC3H2C3, при этом такой домен представлен SEQ ID NO:400 или полипептидом, имеющим в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более идентичности последовательности с одним или более доменов ZfC3H2C3, как представлено SEQ ID NO:306 - SEQ ID NO:351. Кроме того, полипептид ARKL по изобретению предпочтительно содержит домен ZfC3H2C3, как представлено SEQ ID NO:401.
Полипептиды ARKL обычно содержат дополнительный домен, именуемый DAR1 (домен, ассоциированный с RING), который, как было ранее описано, находится за пределами RING-домена в небольшом числе RING-белков растительного происхождения (Stone et al. 2005). Домен DAR1 обычно обнаруживается на N-конце RING-домена. Обычно домены DAR1 содержат консервативную аминокислотную сигнатуру, как представлено SEQ ID NO:399 (мотив 8).
Дополнительный предпочтительный полипептид ARKL, используемый в способах по изобретению, относится к полипептиду, содержащему домен DAR1, имеющий в увеличивающемся порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более идентичности последовательности с одним или более доменов DAR1, как представлено SEQ ID NO:352 - SEQ ID NO:398. Еще более предпочтительно полипептид ARKL по изобретению содержит мотив 8, как представлено SEQ ID NO:399.
Домены RING-типа цинковых пальцев и DAR1 могут быть найдены в базах данных белков, специализированных на белковых семействах, доменах и функциональных сайтах, таких как Pfam (Finn et al. Nucleic Acids Research (2006) Database Issue 34:D247-D251) или InterPro, которые включают базы данных белковых сигнатур: PROSITE, PRINTS, ProDom, Pfam, SMART, TIGRFAMs, PIRSF, SUPERFAMILY, GeneSD и PANTHER (Mulder et al. 2007 Nucleic Acids Research, 2007, Vol.35, Database issue D224-D228). В Pfam собрана большая коллекция множественных выравниваний последовательностей и скрытых марковских моделей (НММ), включающих много распространенных белковых доменов и семейств, и она доступна в Институте Сенгера в Великобритании. Достоверные совпадения, как предполагается в базе данных Pfam, представляют собой те, чьи веса выравнивания последовательностей выше, чем пороговое значение (gathering cutoff threshold). Пороговое значение (gathering cut-off threshold) отсечения домена RING-H2 (рег. номер Pfam: PF00097) в методе Pfam HMM_fs составляет 16.0, в методе Pfam HMM_ls составляет 15.2. Однако потенциальные совпадения, содержащие истинные домены RING-H2, могут еще подпадать под пороговое значение (gathering cut-off). Предпочтительно, полипептид ARKL, используемый в способах по изобретению, представляет собой белок, имеющий один или более доменов в своей последовательности, которые превышают пороговое значение (gathering cut-off) семейства белковых доменов Pfam PF000097, также известных как цинковый палец, домен семейства С3НС4 типа (RING-палец).
Альтернативно, домены цинкового пальца RING-типа и DAR1 в полипептиде могут определяться путем проведения сравнения последовательностей с известными полипептидами, содержащими домены цинкового пальца RING-типа и/или DAR1, и установления сходства в области указанных доменов. Последовательности могут быть выровнены с использованием любого из способов, хорошо известных в данной области техники, таких как алгоритмы Blast. Вероятность того, что выравнивание произойдет для данной последовательности, принимается как основа для идентификации схожих полипептидов. Параметром, который обычно используется для представления такой вероятности, называется значение Е. Значение Е - это мера достоверности веса выравнивания (score) S. Вес выравнивания S - это мера сходства запрошенной последовательности с показанной последовательностью. Значение Е описывает, как часто данный вес S, как предполагается, встречается случайно. Отсечение значения Е может быть до 1.0. Типичное пороговое значение для достоверного значения Е из результата поиска BLAST при использовании полипептида ARKL в качестве запрошенной последовательности составляет меньше чем е-5(=10-5), 1.е-10, 1.е-15, 1.е-20, 1.е-25, 1.е-50, 1.е-75, 1.е-100, 1.е-200, 1.е-300, 1.е-400, 1.е-500, 1.е-600, 1.е-700 и 1.е-800. Предпочтительно, полипептиды ARKL, используемые в способах по изобретению, содержат последовательность, имеющую в увеличивающемся порядке предпочтения значение Е ниже чем е-5(=10-5), 1.е-10, 1.е-15, 1.е-20, 1.е-25, 1.е-50, 1.е-75, 1.е-100, 1.е-200, 1.е-300, 1.е-400, 1.е-500, 1.е-600, 1.е-700 и 1.е-800, в выравнивании с доменами цинкового пальца RING-типа и/или DAR1, как обнаружено в известных полипептидах ARKL, таких как, например, SEQ ID NO:213.
Примеры полипептидов ARKL, используемых в способах по изобретению, представлены в Таблице А. Последовательность, содержащая домены RING-H2 и DAR1, присутствующие в типичных полипептидах ARKL Таблицы А, представлена в SEQ ID NO:306 - SEQ ID NO:351 и SEQ ID NO:352 - SEQ ID NO:398, соответственно. Аминокислотные координаты положения доменов RING-H2 и DAR1, присутствующих в выборке полипептидов ARKL Таблицы А, представлены в Примере 4.
Кроме того, предпочтительные полипептиды ARKL, используемые в способах по изобретению, представляют собой полипептиды, имеющие в увеличивающемся порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%; 98% или более идентичности последовательностей с любыми из полипептидов, представленных в Таблице А.
Неожиданно в настоящее время было обнаружено, что модуляция экспрессии в растении нуклеиновой кислоты, кодирующей полипептид YTP, дает растения, имеющие улучшенные признаки, связанные с урожайностью, по сравнению с контрольными растениями. Согласно первому варианту осуществления настоящее изобретение обеспечивает способ улучшения признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид YTP.
Предпочтительный способ модуляции (предпочтительно, увеличения) экспрессии нуклеиновой кислоты, кодирующей полипептид YTP, представляет собой введение и экспрессию в растении нуклеиновой кислоты, кодирующей YTP.
В одном варианте осуществления любая ссылка в дальнейшем на "белок, используемый в способах по изобретению" делается для обозначения полипептида YTP, как здесь определено. Любая ссылка ниже на "нуклеиновую кислоту, используемую в способах изобретения" делается для обозначения нуклеиновой кислоты, способной кодировать такой полипептид YTP. Нуклеиновая кислота, которую вводят в растение (и, следовательно, используемая при выполнении способов по изобретению), представляет собой любую нуклеиновую кислоту, кодирующую тип белка, который будет сейчас описан, в дальнейшем также именуемую "нуклеиновой кислотой YTP" или "геном YTP".
"Полипептид YTP", как здесь определено, относится к полипептиду, содержащему по меньшей мере один трансмембранный домен и часть по меньшей мере из 50 смежных аминокислот домена DUF221. Дополнительно, полипептид YTP может содержать Мотив 9, как представлено SEQ ID NO:546.
Трансмембранные белки имеют амфифильную структуру с гидрофобными сегментами, пересекающими мембраны, и гидрофильными петлями, которые могут располагаться на любой стороне мембраны (см. Фиг.20). Петли представляют собой сегменты (участки) белка, расположенные между двумя доменами ТМ. Петли среднего размера, расположенные на внутренней стороне мембраны, обычно более отрицательно заряжены, чем расположенные с внешней стороны мембраны.
Трансмембранный домен образует вторичную структуру (обычно, альфа- или бета-спираль), как правило, из 12-35 аминокислотных остатков. Петли между трансмембранными доменами в основном короче, чем 60 аминокислотных остатков, хотя могут также встречаться длинные глобулярные участки. Число трансмембранных доменов в полипептиде YTP меняется, но в основном составляет между 2 и 20.
Трансмембранный домен, обнаруженный в полипептидах YTP, составляет предпочтительно между 8 и 50 аминокислотами, более предпочтительно 8, 12, 14 16, 18, 2, 22, 24, 26, 28, 30, 32, 34, 35 или 36 аминокислот. Петля, обнаруженная в полипептидах YTP, имеет предпочтительно более 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 100 аминокислотных остатков.
Предпочтительный полипептид YTP, используемый в способах по изобретению, содержит в возрастающем порядке предпочтения более чем 1, 2, 4, 5, 6, 8, 10, 12 трансмембранных доменов.
Трансмембранные домены являются высокогидрофобными белками, обогащенными неполярными аминокислотами. Таблица 3 демонстрирует классификацию аминокислот в соответствии со свойствами боковых цепей. Указаны гидрофобные аминокислоты. Гидрофобный характер пептида может определяться способами, хорошо известными в данной области техники, как, например, сообщается Kyte и Doolittle (1982) J. Mol. Biol., 157:105-132.
Полипептиды YTP, используемые в способах по изобретению, предпочтительно содержат трансмембранные домены, имеющие по меньшей мере 20%, 30%, 40%, 50%, 60% или более неполярных аминокислот. В Таблице 3 приведена полярность 20 основных аминокислот.
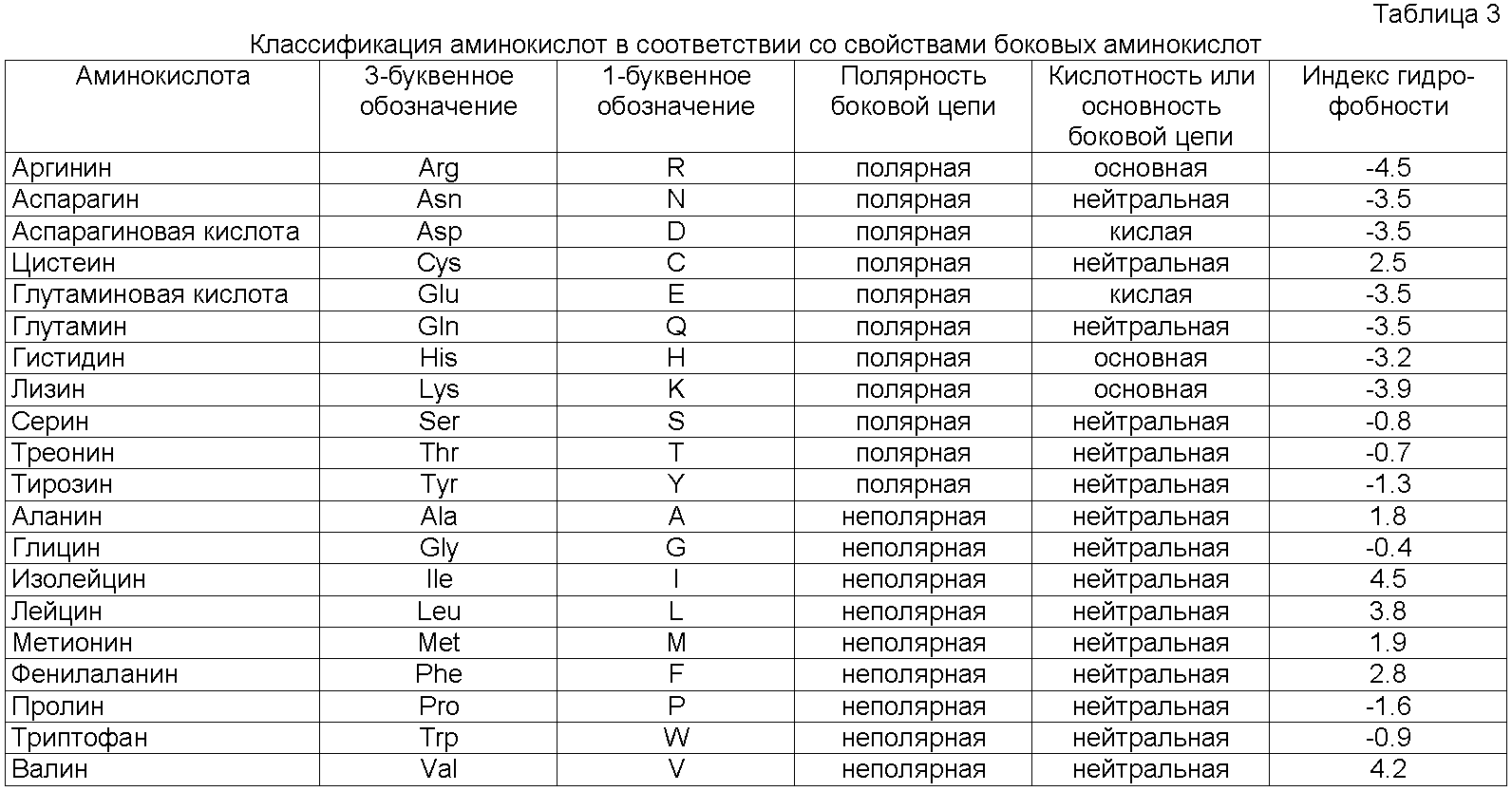
Трансмембранный домен в белке может идентифицироваться с использованием ряда методик, хорошо известных в данной области техники, таких как рентгеновская кристаллография, ЯМР, методы слияния генов, методы доступности замещенного цистеина, эксперименты по Asp(N)-связанному гликозилированию. Дополнительно или альтернативно компьютерные алгоритмы могут использоваться для предсказания трансмембранных доменов. Примеры таких доменов описаны и доступны в институтах, обеспечивающих биоинформатические услуги (Moller et al. 2001. Bioinformatics 17, 646). Применение одного из таких алгоритмов для предсказания трансмембранных доменов в полипептиде YTP показано здесь в разделе примеров.
Домен DUF221 относится к консервативной аминокислотной последовательности, обнаруживаемой в некоторых белках эукариотического происхождения. Домены DUF221 обычно составляют от 350 до 550 остатков в длину. Примеры доменов DUF221, включенных в полипептиды YTP, происходящие из Arabidopsis thaliana и Oryza sativa, представлены SEQ ID NO:518 - SEQ ID NO:543. Консенсусная последовательность, представляющая последовательность SEQ ID NO:518 - SEQ ID NO:543, приведена в SEQ ID NO:544.
Предпочтительный полипептид YTP по изобретению содержит по меньшей мере 50 смежных аминокислот домена DUF221, имеющего в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности с любым из доменов, представленных SEQ ID NO:518 - SEQ ID NO:544. Сходство последовательностей предпочтительно устанавливается в локальном выравнивании с использованием алгоритмов, хорошо известных в данной области техники, таких как Blast.
Полипептид YTP можно легко идентифицировать проведением поиска в специализированных базах данных, содержащих консервативные белковые домены, таких как Pfam (Finn et al. Nucleic Acids Research (2006) Database Issue 34:D247-D251). Инструменты, используемые для проведения поиска в таких базах данных, хорошо известны в этой области техники, например, INTERPRO (Европейский институт биоинформатики, UK), который позволяет проводить поиск в нескольких базах данных белковых доменов одновременно.
Домен DUF221 может идентифицироваться сравнением последовательностей с известными полипептидами, содержащими домен DUF221, и установлением сходства в процентах на участке домена DUF221. Последовательности могут быть выровнены, используя любые способы, хорошо известные в данной области техники, такие как алгоритмы Blast (для локального выравнивания) или BestFit (для глобального выравнивания). Вероятность выравнивания с данной последовательностью принимается как основа для идентификации схожих полипептидов. Параметр, который обычно используют для представления такой вероятности, называют значением Е. Значение Е представляет собой меру достоверности веса выравнивания "S". "S" означает меру сходства между двумя выровненными последовательностями. Значение Е описывает, как часто данный вес "S", как предполагается, встречается случайно. Отсечение значения Е может быть до 1.0. Типичное пороговое значение для достоверного (истинного) хита, показывающего значительную гомологию последовательностей для запрошенной последовательности и получившейся при поиске BLAST, составляет ниже чем 1.е-5, в некоторых примерах берется даже более низкое пороговое значение, например, 1.е-10 или даже ниже.
Предпочтительно, полипептиды YTP, используемые в способах по изобретению, содержат по меньшей мере 50 смежных аминокислот домена DUF221, имеющего в увеличивающемся порядке предпочтения значение Е ниже чем 1.е-5, 1.е-10, 1.е-15, 1.е-20, 1.е-25, 1.е-50, 1.е-75, 1.е-100, 1.е-200, 1.е-300, 1.е-400, 1.е-500, 1.е-600, 1.е-700 и 1.е-800 в локальном выравнивании с доменом DUF221, обнаруженным в известном полипептиде YTP, таком как любой из полипептидов Таблицы А.
Следует понимать, что нуклеиновые кислоты, кодирующие полипептид YTP согласно изобретению, не ограничиваются последовательностями природного происхождения. Нуклеиновая кислота может кодировать "de novo" сконструированный полипептид YTP.
Альтернативно или дополнительно белок YTP имеет в увеличивающемся порядке предпочтения по меньшей мере 25%, 26%, 27%, 28%, 29%, 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% общей идентичности последовательности с аминокислотной последовательностью, представленной SEQ ID NO:409.
Общая идентичность последовательности определяется, используя алгоритм глобального выравнивания, такой как алгоритм Нидлмана и Вунша в программе GAP (GCG Wisconsin Package, Accelrys), предпочтительно с параметрами по умолчанию. По сравнению с общей идентичностью последовательности идентичность последовательности будет, как правило, выше, когда рассматриваются только консервативные домены или мотивы.
Предпочтительно, последовательность полипептида YTP при использовании в построении филогенетического дерева, такого как показано на Фиг.21, образует кластеры с Группой 1, содержащей аминокислотную последовательность, представленную SEQ ID NO:409, а не с полипептидами YTP в Группе 2.
Термины "домен" и "мотив" определены здесь в разделе "определения". Существуют специализированные базы данных для идентификации доменов, например, SMART (Schultz et al. (1998) Proc. Natl. Acad. Sci. USA 95, 5857-5864; Letunic et al. (2002) Nucleic Acids Res 30, 242-244, InterPro (Mulder et al., (2003) Nucl. Acids. Res. 31, 315-318), Prosite (Bucher and Bairoch (1994), A generalized profile syntax for biomolecular sequences motifs and its function in automatic sequence interpretation. (In) ISMB-94; Proceedings 2nd International Conference on Intelligent Systems for Molecular Biology. Altman R., Brutlag D., Karp P., Lathrop R., Searls D., Eds., pp53-61, AAAI Press, Menio Park; Hulo et al., Nucl. Acids. Res. 32:D134-D137, (2004)) или Pfam (Bateman et al., Nucleic Acids Research 30(1): 276-280 (2002)). Набор средств для анализа in silico белковых последовательностей доступен на протеомическом сервере ExPASy (Swiss Institute of Bioinformatics (Gasteiger et al., ExPASy: the proteomics server for in-depth protein knowledge and analysis, Nucleic Acids Res. 31:3784-3788(2003)).
Анализ полипептидной последовательности SEQ ID NO:2 представлен здесь ниже в Примерах 2 и 4. Например, полипептид GRF, как представлено SEQ ID NO:2, содержит домен QLQ с InterPro регистрационным номером IPR014978 (регистрационный номер PF08880 в PFAM) и домен WRC с InterPro регистрационным номером IPR014977 (регистрационный номер PF08879 в PFAM) в базе данных доменов InterPro. Домены могут также идентифицироваться с использованием рутинных методик, таких как выравнивание последовательностей. Выравнивание домена QLQ полипептидов из Таблицы А здесь представлено на Фиг.2, и выравнивание домена WRC полипептидов из Таблицы А здесь представлено на Фиг.3. Такие выравнивания полезны для определения наиболее консервативных аминокислот между полипептидами GRF, таких как аминокислотные остатки QLQ и WRC.
Способы для выравнивания последовательностей для сравнения хорошо известны в данной области техники, при этом такие способы включают GAP, BESTFIT, BLAST, FASTA и TFASTA. GAP использует алгоритм Нидлмана и Вунша ((1970) J Mol Biol 48:443-453) для обнаружения глобального (т.е. охватывающего полные последовательности) выравнивания двух последовательностей, которое увеличивает число совпадений и доводит до минимума число гэпов. Алгоритм BLAST (Altschul et al. (1990) J Mol Biol 215:403-10) рассчитывает идентичность последовательности и проводит статистический анализ сходства между двумя последовательностями. Программное обеспечение для выполнения анализа BLAST открыто доступно через Национальный центр биотехнологической информации (NCBI). Гомологи могут легко идентифицироваться, используя, например, алгоритм множественного выравнивания последовательностей ClustalW (версия 1.83) с параметрами попарного выравнивания по умолчанию и методом количественной оценки в процентах. Общие проценты сходства и идентичности могут также быть определены при использовании одного из способов, доступного в пакете программ MatGAT (Campanella et al., BMC Bioinformatics. 2003 Jul 10;4:29. MatGAT: an application that generates similarity/identity matrices using protein or DNA sequences). Небольшое ручное редактирование может быть выполнено для оптимизации выравнивания между консервативными мотивами, которое, вероятно, является очевидным специалисту в данной области техники. Кроме того, вместо применения полноразмерных последовательностей для идентификации гомологов можно также использовать специфические домены. Значения идентичности последовательности могут определяться для полной последовательности нуклеиновой кислоты или полипептидной последовательности, или для выбранных доменов или консервативного(ых) мотива(ов), применяя программы, упомянутые выше, с использованием параметров по умолчанию.
Для локальных выравниваний в частности применим алгоритм Смита-Ватермана (Smith TF, Waterman MS (1981) J. Mol. Biol 147(1); 195-7).
По общему мнению за пределами домена QLQ и домена WRC полипептиды GRF имеют низкую идентичность аминокислотной последовательности. Пример 3 описывает здесь в Таблице В идентичность в процентах между полипептидом GRF, как представлено SEQ ID NO:2, и полипептидами GRF, перечисленными в Таблице А, которая может быть такой низкой, как 15% идентичности аминокислотной последовательности. Идентичность в процентах может существенно увеличиваться, если вычисление идентичности проводится между доменом QLQ из SEQ ID NO:2 (как представлено SEQ ID NO:115, включенной в SEQ ID NO:2; домен QLQ полипептидов GRF Таблицы А, представленный на Фиг.2) и доменами QLQ полипептидов, используемых при выполнении изобретения. Подобным образом идентичность в процентах может существенно увеличиваться, если вычисление идентичности проводится между доменом WRC из SEQ ID NO:2 (как представлено SEQ ID NO:116, включенной в SEQ ID NO:2; домен WRC полипептидов GRF из Таблицы А, представленный на Фиг.3) и доменами WRC полипептидов, используемых при выполнении изобретения. Идентичность в процентах в пределах домена QLQ среди полипептидных последовательностей, используемых при проведении способов по изобретению, находится в диапазоне между 25% и 99% аминокислотной идентичности, и идентичность в процентах в пределах домена WRC среди полипептидных последовательностей, используемых при проведении способов по изобретению, находится в диапазоне между 60% и 99% аминокислотной идентичности. Как можно также наблюдать на Фиг.3, домен WRC в большей степени консервативен среди различных полипептидов GRF, чем домен QLQ, как показано на Фиг.2.
Задача предсказания субклеточной локализации белка является важной и хорошо изученной. Знание локализации белка помогает объяснить его функцию. Экспериментальные способы для определения локализации белка простираются от иммунолокализации до мечения белков с применением зеленого флуоресцентного белка (GFP) или бета-глюкуронидазы (GUS). Такие способы являются точными, хотя и трудоемкими по сравнению с вычислительными методами. В последнее время был сделан большой прогресс в вычислительном предсказании локализации белка из данных о последовательности. Среди алгоритмов, хорошо известных специалисту в данной области техники, являются доступными среди протеомических инструментальных средств ExPASy, размещенных Swiss Institute for Bioinformatics, например, PSort, TargetP, ChloroP, Predotar, LipoP, MITOPROT, PATS, PTS1, SignalP и другие.
Кроме того, полипептиды GRF, используемые в способах настоящего изобретения (по меньшей мере, в их нативной форме) обычно, но необязательно, имеют транскрипционную регуляторную активность и способность взаимодействовать с другими белками. Следовательно, полипептиды GRF с уменьшенной транскрипционной регуляторной активностью, без транскрипционной регуляторной активности, со сниженной способностью взаимодействия белок-белок или без способности взаимодействия белок-белок могут в равной степени использоваться в способах по настоящему изобретению. DNA-связывающая активность и взаимодействия белок-белок могут легко определяться in vitro или in vivo с использованием методик, хорошо известных в данной области техники (например, Current Protocols in Molecular Biology, Volumes 1 and 2, Ausubel et al. (1994), Current Protocols). Полипептиды GRF способны к транскрипционной активации репортерных генов в дрожжевых клетках (Kim & Kende (2004) Proc Natl Acad Sci 101(36): 13374-13379). Полипептиды GRF также способны взаимодействовать с полипептидами GRF-взаимодействующих факторов (GIF1 - GIF3; также называемые полипептидами транслокации синовиальной саркомы (SYT), SYT1 - SYT3) in vivo в дрожжевых клетках, при использовании анализа взаимодействия белок-белок с помощью дрожжевых двойных гибридов (Kim & Kende, supra). Анализы связывания in vitro также используются для демонстрации того, что полипептиды GRF и полипептиды GIF (также называемые SYT) являются взаимодействующими партнерами (Kim & Kende, supra).
Настоящее изобретение демонстрируется в одном из вариантов осуществления путем трансформации растений последовательностью нуклеиновой кислоты, представленной SEQ ID NO:1, кодирующей полипептидную последовательность GRF согласно SEQ ID NO:2. Однако выполнение изобретения не ограничивается этими последовательностями; способы по изобретению могут успешно осуществляться с использованием любой последовательности нуклеиновой кислоты, кодирующей полипептид GRF, как здесь определено.
Настоящее изобретение демонстрируется в одном из вариантов осуществления путем трансформации растений последовательностью нуклеиновой кислоты, представленной SEQ ID NO:120, кодирующей полипептидную последовательность SEQ ID NO:121. Однако выполнение изобретения не ограничивается этими последовательностями; способы по изобретению могут успешно осуществляться с использованием любой нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, или RAA1-подобного полипептида, как здесь определено.
Дополнительно, RAA1-подобные полипептиды, будучи экспрессированы в рисе согласно способам настоящего изобретения, как описано в примерах, дают растения, имеющие усиленные признаки, связанные с урожайностью, в частности увеличенный показатель корни/побеги, увеличенное число цветков на метелку и увеличенную массу тысячи зерен.
Трансмембранные домены составляют от около 15 до 30 аминокислот в длину и обычно состоят из гидрофобных остатков, которые формируют альфа-спираль. Они обычно предсказываются на основе гидрофобности (например, Klein et al., Biochim. Biophys. Acta 815, 468, 1985; или Sonnhammer et al., In J. Glasgow, T. Littlejohn, F. Major, R. Lathrop, D. Sankoff, and C. Sensen, editors, Proceedings of the Sixth International Conference on Intelligent Systems for Molecular Biology, pages 175-182, Menlo Park, CA, 1998. AAAI Press.).
Примеры белков, подпадающих под определение "полипептид SYR или его гомолог", представлены в Таблице А в разделе примеров и включают последовательности из различных однодольных растений, таких как рис (SEQ ID NO:169, SEQ ID NO:179 и SEQ ID NO:180), кукуруза (SEQ ID NO:181), пшеница (SEQ ID NO:182), ячмень (SEQ ID NO:183), сахарный тростник (SEQ ID NO:184 и SEQ ID NO:185), сорго (SEQ ID NO:186); и из двудольных растений, таких как Arabidopsis (SEQ ID NO:187 и SEQ ID NO:188), виноград (SEQ ID NO:189), цитрусовые (SEQ ID NO:190) или томат (SEQ ID NO:191 и SEQ ID NO:192). Представляется, что обогащенный Leu домен является важным для функционирования белка, следовательно, белки с обогащенным Leu доменом, но без консервативных мотивов 5 или 6, могут также использоваться в способах по настоящему изобретению; примеры таких белков представлены в SEQ ID NO:201 и 202.
Следует понимать, что термин "полипептид SYR или его гомолог" не ограничивается последовательностью, представленной SEQ ID NO:169 или гомологами, перечисленными как SEQ ID NO:179 - SEQ ID NO:192, но что любой полипептид от около 65 до около 200 аминокислот, отвечающий критериям содержания обогащенного лейцином домена, как определено выше, перед которым находится консервативный трипептидный мотив 5 (а, b, с или d), и за которым следует консервативный мотив 6 и предпочтительно также консервативный мотив 7; или имеющий по меньшей мере 38% идентичности последовательности по отношению к последовательности SEQ ID NO:169, может подходить для использования в способах по изобретению.
Активность белка SYR или его гомолога может анализироваться экспрессией белка SYR или его гомолога под контролем промотора GOS2 в Oryza sativa, которая приводит к растениям с увеличенными биомассой и/или урожайностью семян без отсрочки периода цветения при выращивании в условиях недостатка азота или условиях стресса, вызванного засухой, и по сравнению с соответствующими растениями дикого типа. Это увеличение урожайности семян может быть измерено несколькими способами, например, как увеличение общей массы семян, числа наполненных семян, фильтрата, индекса урожайности или массы тысячи зерен.
Настоящее изобретение иллюстрируется в одном из вариантов осуществления изобретения трансформацией растений последовательностью нуклеиновой кислоты, представленной SEQ ID NO:168, кодирующей полипептидную последовательность SEQ ID NO:169. Однако выполнение изобретения не ограничивается этими последовательностями; способы по изобретению могут успешно осуществляться с использованием SYR-кодирующей нуклеиновой кислоты или полипептида SYR, как здесь определено.
Кроме того, полипептиды ARKL (по меньшей мере, в их нативной форме), как правило, имеют Е3 убиквитин-протеин лигазную активность. Инструменты и методики для измерения Е3 убиквитин-протеин лигазной активности хорошо известны в данной области техники (патент США 6737244; WO/2001/075145; Miura et al. (2005) Proc Natl Acad Sci USA. 102(21): 7760-7765; Kawasaki et al. (2005) The Plant Journal 44, 258-270). Вкратце, Е3 убиквитин-протеин лигазная активность может быть проанализирована инкубированием белка ARKL с Е1 и Е2 белками и меченым убиквитином. Убиквитинированные белки могут обнаруживаться после SDS-PAGE электрофореза и блоттинга с использованием антитела для мечения убиквитина. Примеры белков Е1 и Е2, которые могут быть использованы в анализе, представляют собой Е1 белок пшеницы и AtUBC1 Е2 белок Arabidopsis thaliana. Меченый убиквитин с гистидином и антителами для его обнаружения доступен в продаже (Calbiochem, San Diego, CA, USA).
Дополнительно, полипептиды ARKL, при экспрессии в рисе согласно способам по настоящему изобретению, как описано в примерах, дают растения, имеющие усиленные признаки, связанные с урожайностью, в частности массу тысячи зерен, общую урожайность семян, раннюю мощность и/или индекс урожайности.
Настоящее изобретение иллюстрируется в одном варианте осуществления трансформацией растений последовательностью нуклеиновой кислоты, представленной SEQ ID NO:212, кодирующей полипептидную последовательность SEQ ID NO:213. Однако выполнение изобретения не ограничивается этими последовательностями; способы по изобретению могут успешно выполняться, используя любую нуклеиновую кислоту, кодирующую ARKL, или полипептид ARKL, как здесь определено.
Кроме того, полипептиды YTP, как правило, имеют активность, увеличивающую урожайность семян. Инструменты и методики для измерения активности, увеличивающей (или улучшающей) урожайность, хорошо известны в данной области техники. Дополнительные подробности представлены здесь в разделе Примеры.
Дополнительно, полипептиды YTP при экспрессии и фенотипической оценке в рисе согласно способам настоящего изобретения, как описано в Примерах 10-15, дают растения, имеющие усиленные признаки, связанные с урожайностью, в частности один или более из общей массы семян, массы тысячи зерен, числа цветков на метелку, уровня наполнения семян и индекса урожайности.
Настоящее изобретение иллюстрируется в одном варианте осуществления трансформацией растений последовательностью нуклеиновой кислоты, представленной SEQ ID NO:408, кодирующей полипептидную последовательность SEQ ID NO:409. Однако выполнение изобретения не ограничивается этими последовательностями; способы по изобретению могут успешно выполняться, используя любую нуклеиновую кислоту, кодирующую YTP, или полипептид YTP, как здесь определено.
Примеры последовательностей нуклеиновых кислот, кодирующих полипептиды по изобретению, приведены здесь в Таблице А Примера 1, особенно последовательностей нуклеиновых кислот, кодирующих полипептиды, выбранные из группы, состоящей из:
полипептид GRF, представленный в Таблице А1,
RAA1-подобный полипептид, представленный в Таблице А2,
полипептид SYR, представленный в Таблице A3,
полипептид ARKL, представленный в Таблице А4, и
полипептид YTP, представленный в Таблице А5, соответственно.
Такие последовательности нуклеиновых кислот используются при проведении способов по изобретению. Полипептидные последовательности, приведенные в Таблице А Примера 1, являются последовательностями-примерами ортологов и паралогов полипептида, выбранного из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и YTP, представленные SEQ ID NO:2, 121, 169, 213 или 409 соответственно, при этом термины "ортологи" и "паралоги" являются такими, как здесь определено. Дополнительные ортологи и паралоги могут быть легко идентифицированны путем проведения так называемого реципрокного blast поиска. Обычно это содержит первый BLAST, включая проведение BLAST для запрошенной последовательности (например, используя любую из последовательностей, перечисленных в Таблице А Примера 1) относительно любой базы данных с последовательностями, например открыто доступной базы данных NCBI. BLASTN или TBLASTX (при использовании стандартных значений по умолчанию), как правило, используются, когда исходят из нуклеотидной последовательности, и BLASTP или TBLASTN (при использовании стандартных значений по умолчанию), когда исходят из белковой последовательности. Результаты BLAST при необходимости можно фильтровать. Для полноразмерных последовательностей в случае или фильтрованных результатов, или нефильтрованных результатов затем проводят обратный BLAST (второй BLAST) относительно последовательностей из организма, из которого получена запрошенная последовательность (если запрошенная последовательность означает SEQ ID NO:1, 120, 168 или 408 соответственно или SEQ ID NO:2, 121, 169, 213 или 409 соответственно, второй BLAST, следовательно, проводили бы относительно последовательностей Arabidopsis thaliana). Результаты первого и второго BLAST затем сравниваются. Паралог идентифицируется, если высший хит из первого blast происходит из того же вида, из которого получена запрошенная последовательность, а обратный BLAST затем в идеале приводит к запрошенной последовательности относительно наивысших хитов; ортолог идентифицируется, если высший хит в первом BLAST происходит не из того вида, из которого получена запрошенная последовательность, и предпочтительно приводит при обратном BLAST к запрошенной последовательности среди наивысших хитов.
Термин "таблица А", употребляемый в данном описании, используется, чтобы определить содержание таблицы А1, А2, A3, А4 и/или А5. Термин "таблица А1", употребляемый в данном описании, используется, чтобы определить содержание таблицы А1. Термин "таблица А2", употребляемый в данном описании, используется, чтобы определить содержание таблицы А2. Термин "таблица A3", употребляемый в данном описании, используется, чтобы определить содержание таблицы A3. Термин "таблица А4", употребляемый в данном описании, используется, чтобы определить содержание таблицы А4. Термин "таблица А5", употребляемый в данном описании, используется, чтобы определить содержание таблицы А5.
В одном предпочтительном варианте осуществления изобретения термин "таблица А" означает таблицу А1. В одном предпочтительном варианте осуществления изобретения термин "таблица А" означает таблицу А2. В одном предпочтительном варианте осуществления изобретения термин "таблица А" означает таблицу A3. В одном предпочтительном варианте осуществления изобретения термин "таблица А" означает таблицу А4. В одном предпочтительном варианте осуществления изобретения термин "таблица А" означает таблицу А5.
Высшие хиты - это те, которые имеют низкое значение Е. Чем ниже значение Е, тем более значительный вес выравнивания (score) (или, другими словами, тем ниже возможность, что хит обнаружили случайно). Вычисление значения Е хорошо известно в данной области техники. Дополнительно к значениям Е сравнения также оцениваются идентичностью в процентах. Идентичность в процентах относится к числу идентичных нуклеотидов (или аминокислот) между двумя сравниваемыми последовательностями нуклеиновых кислот (или полипептидов) на определенной длине. В случае больших семейств может использоваться ClustalW с последующим деревом, построенным методом присоединения соседа, чтобы помочь визуализировать выделение кластера родственных генов и идентифицировать ортологи и паралоги.
Варианты нуклеиновых кислот могут также использоваться в осуществлении на практике способов по изобретению. Примеры таких вариантов включают последовательности нуклеиновых кислот, кодирующие гомологи и производные любой из полипептидных последовательностей, приведенных в Таблице А Примера 1, при этом термины "гомолог" и "производное" являются такими, как здесь определено. Также применимыми в способах по изобретению являются последовательности нуклеиновых кислот, кодирующие гомологи и производные ортологов или паралогов любой из полипептидных последовательностей, представленных в Таблице А Примера 1. Гомологи и производные, применимые в способах настоящего изобретения, имеют по существу такую же биологическую и функциональную активность, как немодифицированный белок, из которого они получены.
В одном варианте осуществления изобретения предпочтительное производное, используемое в способах по изобретению, представляет собой полипептид ARKL, имеющий остаток цистеина в положении лиганда 5 (см. Фиг.15) в домене RING-пальца, координирующем один из ионов цинка. Другое предпочтительное производное, используемое в способах изобретения, представляет собой полипептид ARKL, имеющий семь цистеинов и один гистидин (ZfC3HC4) в качестве остатков в положениях лигандов ионов цинка.
Дополнительные варианты нуклеиновых кислот, используемые в осуществлении на практике способов по изобретению, включают части последовательностей нуклеиновых кислот, кодирующих полипептиды, выбранные из группы, состоящей из: полипептиды GRF, RAAI-подобные полипептиды, полипептиды SYR, полипептиды ARKL и полипептиды YTP соответственно, последовательности нуклеиновых кислот, гибридизующиеся с последовательностями нуклеиновых кислот, кодирующими полипептиды, выбранные из группы, состоящей из:
полипептиды GRF, RAA1-подобные полипептиды, полипептиды SYR, полипептиды ARKL и полипептиды YTP соответственно, сплайс-варианты последовательностей нуклеиновых кислот, кодирующих полипептиды, выбранные из группы, состоящей из: полипептиды GRF, RAA1-подобные полипептиды, полипептиды SYR, полипептиды ARKL и полипептиды YTP соответственно, аллельные варианты нуклеиновых кислот, кодирующих полипептиды, выбранные из группы, состоящей из: полипептиды GRF, RAA1-подобные полипептиды, полипептиды SYR, полипептиды ARKL и полипептиды YTP соответственно, и варианты нуклеиновых кислот, кодирующих полипептиды, выбранные из группы, состоящей из: полипептиды GRF, RAA1-подобные полипептиды, полипептиды SYR, полипептиды ARKL и полипептиды YTP соответственно, полученные перетасовкой генов. Термины «гибридизующаяся последовательность», «сплайс-вариант», «аллельный вариант» и «перетасовка генов» означают такие, как здесь описано.
Последовательности нуклеиновых кислот, кодирующие полипептиды, выбранные из группы, состоящей из: полипептиды GRF, RAA1-подобные полипептиды, полипептиды SYR, полипептиды ARKL и полипептиды YTP соответственно, необязательно являются полноразмерными последовательностями нуклеиновых кислот, так как выполнение способов изобретения не основывается на применении полноразмерных последовательностей нуклеиновых кислот. Согласно настоящему изобретению предоставляется способ усиления признаков, связанных с урожайностью семян, у растений, предусматривающий введение и экспрессию в растении части любой из последовательностей нуклеиновых кислот, приведенных в Таблице А Примера 1, или части последовательности нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из полипептидных последовательностей, приведенных в Таблице А Примера 1.
Часть последовательности нуклеиновой кислоты может быть получена, например, осуществлением одной или более делеций в последовательности нуклеиновой кислоты. Части могут использоваться в выделенной форме или могут быть слиты с другими кодирующими (или некодирующими) последовательностями для того, чтобы, например, получить белок, который комбинирует несколько активностей. При слиянии с другими кодирующими последовательностями получающийся полипептид, полученный при трансляции, может быть больше, чем предсказанный для белковой части.
Части, используемые в способах изобретения, кодируют в одном варианте осуществления изобретения полипептид GRF, как здесь определено, и имеют по существу ту же самую биологическую активность, как полипептидные последовательности, представленные в Таблице А1 Примера 1. Предпочтительно, часть представляет собой часть любой из последовательностей нуклеиновых кислот, представленных в Таблице А1 Примера 1, или представляет собой часть последовательности нуклеиновой кислоты, кодирующей ортолог или паралог любой из полипептидных последовательностей, представленных в Таблице А1 Примера 1. Предпочтительно, часть означает в увеличивающемся порядке предпочтения по меньшей мере 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1190 последовательных нуклеотидов в длину, при этом последовательные нуклеотиды принадлежат любой из последовательностей нуклеиновых кислот, представленных в Таблице А1 Примера 1, или последовательности нуклеиновой кислоты, кодирующей ортолог или паралог любой из полипептидных последовательностей, представленных в Таблице А1 Примера 1. Предпочтительно, часть представляет собой часть нуклеиновой последовательности, кодирующей полипептид полипептидной последовательности, содержащий: (i) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом QLQ, как представлено SEQ ID NO:115; и (ii) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом WRC, как представлено SEQ ID NO:116. Наиболее предпочтительно, часть представляет собой часть последовательности нуклеиновой кислоты SEQ ID NO:1.
Части, используемые в способах по изобретению, кодируют в одном варианте осуществления RAAI-подобный полипептид, как здесь определено, и имеют по существу такую же биологическую активность, как аминокислотные последовательности, представленные в Таблице А2 Примера 1. Предпочтительно, часть представляет собой часть любой из нуклеиновых кислот, представленных в Таблице А2 Примера 1, или часть нуклеиновой кислоты, кодирующей ортолог или паралог любой из аминокислотных последовательностей, представленных в Таблице А2 Примера 1. Предпочтительно, часть означает по меньшей мере 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900, 2000, 2100, 2200 последовательных нуклеотидов в длину, при этом последовательные нуклеотиды принадлежат любой из последовательностей нуклеиновых кислот, представленных в Таблице А2 Примера 1, или нуклеиновой кислоте, кодирующей ортолог или паралог любой из полипептидных последовательностей, представленных в Таблице А2 Примера 1. Наиболее предпочтительной является часть нуклеиновой кислоты согласно SEQ ID NO:120. Предпочтительно часть кодирует фрагмент аминокислотной последовательности, которая при использовании в построении филогенетического дерева, такого как изображенное на Фиг.8, образует кластеры с группой RAA1-подобных полипептидов, содержащей аминокислотную последовательность, представленную SEQ ID NO:121, а не с любой другой группой.
Части, используемые в способах по изобретению, кодируют в одном варианте осуществления полипептид SYR, как здесь определено, и имеют по существу такую же биологическую активность, как аминокислотные последовательности, представленные в Таблице A3 Примера 1. Предпочтительно, часть представляет собой часть любой из нуклеиновых кислот, представленных в Таблице A3 Примера 1, или часть нуклеиновой кислоты, кодирующей ортолог или паралог любой из аминокислотных последовательностей, представленных в Таблице A3 Примера 1. Предпочтительно, часть означает по меньшей мере 150, 200, 250, 300, 350, 400, 450, 500, 550, 600 последовательных нуклеотидов в длину, при этом последовательные нукпеотиды принадлежат любой из последовательностей нуклеиновых кислот, представленных в Таблице A3 Примера 1, или нуклеиновой кислоте, кодирующей ортолог или паралог любой из полипептидных последовательностей, представленных в Таблице A3 Примера 1. Наиболее предпочтительно часть представляет собой часть нуклеиновой кислоты SEQ ID NO:168. Предпочтительно, часть кодирует полипептид от около 65 до около 200 аминокислот, содержащий обогащенный лейцином домен, как определено выше, перед которым находится консервативный трипептидный мотив 5 (а, b, с или d), и за которым следует консервативный мотив 6 и предпочтительно также консервативный мотив 7; или имеющий по меньшей мере 38% идентичности последовательности по отношению к последовательности SEQ ID NO:169.
Части, используемые в способах по изобретению, кодируют в одном варианте осуществления полипептид ARKL, как здесь определено, и имеют по существу такую же биологическую активность, как аминокислотные последовательности, представленные в Таблице А4 Примера 1. Предпочтительно, часть представляет собой часть любой из нуклеиновых кислот, представленных в Таблице А4 Примера 1, или часть нуклеиновой кислоты, кодирующей ортолог или паралог любой из аминокислотных последовательностей, представленных в Таблице А4 Примера 1. Предпочтительно, часть означает по меньшей мере 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000 последовательных нуклеотидов в длину, при этом последовательные нуклеотиды принадлежат любой из последовательностей нуклеиновых кислот, представленных в Таблице А4 Примера 1, или нуклеиновой кислоте, кодирующей ортолог или паралог любой из полипептидных последовательностей, представленных в Таблице А4 Примера 1. Наиболее предпочтительно часть представляет собой часть нуклеиновой кислоты SEQ ID NO:212. Предпочтительно часть кодирует фрагмент аминокислотной последовательности, которая при использовании в построении филогенетического дерева, такого как изображенное на Фиг.17, образует кластеры с группой полипептидов ARKL, содержащей аминокислотную последовательность, представленную SEQ ID NO:213, а не с любой другой группой, такой как представленная последовательностью Musmu_Goliath.
Части, используемые в способах по изобретению, кодируют в одном варианте осуществления полипептид YTP, как здесь определено, и имеют по существу такую же биологическую активность, как аминокислотные последовательности, представленные в Таблице А5 Примера 1. Предпочтительно, часть представляет собой часть любой из нуклеиновых кислот, представленных в Таблице А5 Примера 1, или часть нуклеиновой кислоты, кодирующей ортолог или паралог любой из аминокислотных последовательностей, представленных в Таблице А5 Примера 1. Предпочтительно, часть означает по меньшей мере 300, 400, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1500, 2000, 2500, 3000 последовательных нуклеотидов в длину, при этом последовательные нуклеотиды принадлежат любой из последовательностей нуклеиновых кислот, представленных в Таблице А5 Примера 1, или нуклеиновой кислоте, кодирующей ортолог или паралог любой из полипептидных последовательностей, представленных в Таблице А5 Примера 1. Наиболее предпочтительно часть представляет собой часть нуклеиновой кислоты согласно SEQ ID NO:408. Предпочтительно часть кодирует фрагмент аминокислотной последовательности, которая при использовании в построении филогенетического дерева, такого как изображенное на Фиг.21, образует кластеры с группой 1, содержащей аминокислотную последовательность, представленную SEQ ID NO:409, а не с полипептидами YTP в группе 2.
Другой вариант последовательности нуклеиновой кислоты, используемой в способах по изобретению, представляет собой последовательность нуклеиновой кислоты, способную к гибридизации в условиях пониженной жесткости, предпочтительно в жестких условиях, с последовательностью нуклеиновой кислоты, кодирующей полипептид, увеличивающий урожайность, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как здесь определено, или с частью, как здесь определено.
Согласно настоящему изобретению предоставляется способ усиления признаков, связанных с урожайностью семян, у растений, предусматривающий введение и экспрессию в растении последовательности нуклеиновой кислоты, способной к гибридизации с любой из последовательностей нуклеиновых кислот, представленных в Таблице А Примера 1, или предусматривающий введение и экспрессию в растении последовательности нуклеиновой кислоты, способной к гибридизации с последовательностью нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из последовательностей нуклеиновых кислот, представленных в Таблице А Примера 1.
Гибридизующиеся последовательности, применимые в способах изобретения, кодируют полипептид, выбранный из группы: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как здесь определено, имеющий по существу такую же биологическую активность, как полипептидные последовательности, данные в Таблице А Примера 1. Предпочтительно, гибридизующаяся последовательность способна к гибридизации с любой из последовательностей нуклеиновых кислот, представленных в Таблице А Примера 1, или с частью любой из этих последовательностей, при этом часть является такой, как определено выше, или гибридизующаяся последовательность способна к гибридизации с последовательностью нуклеиновой кислоты, кодирующей ортолог или паралог любой из полипептидных последовательностей, приведенных в Таблице А Примера 1.
Предпочтительно, в одном варианте осуществления гибридизующаяся последовательность способна к гибридизации с последовательностью нуклеиновой кислоты, кодирующей полипептидную последовательность, содержащую: (i) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом QLQ, как представлено SEQ ID NO:115; и (ii) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом WRC, как представлено SEQ ID NO:116. Наиболее предпочтительно, гибридизующаяся последовательность способна гибридизоваться с последовательностью нуклеиновой кислоты, как представлено SEQ ID NO:1, или с ее частью.
Предпочтительно, в одном варианте осуществления гибридизующаяся последовательность способна гибридизоваться с нуклеиновой кислотой, как представлено SEQ ID NO:120, или с ее частью. Предпочтительно, гибридизующаяся последовательность кодирует полипептид с аминокислотной последовательностью, которая, когда является полноразмерной и используется в построении филогенетического дерева, такого как показано на Фиг.8, образует кластеры с группой RAA1-подобных полипептидов, содержащей аминокислотную последовательность, представленную SEQ ID NO:121, а не с любой другой группой.
Предпочтительно, в одном варианте осуществления гибридизующаяся последовательность способна гибридизоваться с нуклеиновой кислотой, как представлено SEQ ID NO:168, или с ее частью. Предпочтительно, гибридизующаяся последовательность кодирует полипептид от около 65 до около 200 аминокислот, содержащий обогащенный лейцином домен, как определено выше, перед которым находится консервативный трипептидный мотив 5 (а, b, с или а), и за которым следует консервативный мотив 6 и предпочтительно также консервативный мотив 7; или имеющий по меньшей мере 38% идентичности последовательности по отношению к последовательности SEQ ID NO:169.
Предпочтительно, в одном варианте осуществления гибридизующаяся последовательность способна гибридизоваться с нуклеиновой кислотой, как представлено SEQ ID NO:212, или с ее частью. Предпочтительно, гибридизующаяся последовательность кодирует полипептид с аминокислотной последовательностью, которая, когда является полноразмерной и используется в построении филогенетического дерева, такого как показано на Фиг.17, образует кластеры с группой полипептидов ARKL, содержащей аминокислотную последовательность, представленную SEQ ID NO:213, а не с любой другой группой, такой как представлено последовательностью Musmu_Goliath.
Предпочтительно, в одном варианте осуществления гибридизующаяся последовательность способна гибридизоваться с нуклеиновой кислотой, как представлено SEQ ID NO:408, или с ее частью.
Предпочтительно, гибридизующаяся последовательность кодирует полипептид с аминокислотной последовательностью, которая, когда является полноразмерной и используется в построении филогенетического дерева, такого как показано на Фиг.21, образует кластеры с группой 1, содержащей аминокислотную последовательность, представленную SEQ ID NO:409, а не с полипептидами YTP в Группе 2.
Другим вариантом последовательности нуклеиновой кислоты, используемым в способах по изобретению, является сплайс-вариант, кодирующий полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как определено выше, при этом сплайс-вариант является таким, как здесь определено.
Согласно настоящему изобретения предоставляется способ усиления признаков, связанных с урожайностью, предусматривающий введение и экспрессию в растении сплайс-варианта любой из последовательностей нуклеиновых кислот, представленных в Таблице А Примера 1, или сплайс-варианта последовательности нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из полипептидных последовательностей, представленных в Таблице А Примера 1.
Предпочтительные сплайс-варианты в одном варианте осуществления изобретения представляют собой сплайс-варианты последовательности нуклеиновой кислоты, представленной SEQ ID NO:1, или сплайс-вариант последовательности нуклеиновой кислоты, кдирующей ортолог или паралог SEQ ID NO:2. Предпочтительно, сплайс-вариант представляет собой сплайс-вариант последовательности нуклеиновой кислоты, кодирующей полипептидную последовательность, содержащую: (i) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом QLQ, как представлено SEQ ID NO:115; и (ii) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом WRC, как представлено SEQ ID NO:116.
Предпочтительными сплайс-вариантами в одном варианте осуществления изобретения являются сплайс-варианты нуклеиновой кислоты, представленной SEQ ID NO:120, или сплайс-вариант нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:121. Предпочтительно, аминокислотная последовательность, кодируемая сплайс-вариантом, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.8, объединяется в кластер с группой RAA1-подобных полипептидов, содержащей аминокислотную последовательность, представленную SEQ ID NO:121, а не с любой другой группой.
Предпочтительными сплайс-вариантами в одном варианте осуществления изобретения являются сплайс-варианты нуклеиновой кислоты, представленной SEQ ID NO:168, или сплайс-вариант нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:169. Предпочтительно, аминокислотная последовательность, кодируемая сплайс-вариантом, представляет собой полипептид от около 65 до около 200 аминокислот, содержащий обогащенный лейцином домен, как определено выше, перед которым находится консервативный трипептидный мотив 5 (а, b, с или d), и за которым следует консервативный мотив 6 и предпочтительно также консервативный мотив 7; или имеющий по меньшей мере 38% идентичности последовательности по отношению к последовательности SEQ ID NO:169.
Предпочтительными сплайс-вариантами в одном варианте осуществления изобретения являются сплайс-варианты нуклеиновой кислоты, представленной SEQ ID NO:212, или сплайс-вариант нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:213. Предпочтительно, аминокислотная последовательность, кодируемая сплайс-вариантом, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.17, объединяется в кластер с группой полипептидов ARKL, содержащей аминокислотную последовательность, представленную SEQ ID NO:213, а не с любой другой группой, такой как представленная последовательностью Musmu_Goliath.
Предпочтительными сплайс-вариантами в одном варианте осуществления изобретения являются сплайс-варианты нуклеиновой кислоты, представленной SEQ ID NO:408, или сплайс-вариант нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:409. Предпочтительно, аминокислотная последовательность, кодируемая сплайс-вариантом, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.21, объединяется в кластер с Группой 1, содержащей аминокислотную последовательность, представленную SEQ ID NO:409, а не с полипептидами YTP в Группе 2.
Другим вариантом последовательности нуклеиновой кислоты, используемым при выполнении способов по изобретению, является аллельный вариант последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как определено выше, при этом аллельный вариант является таким, как здесь определено.
Согласно настоящему изобретению предоставляется способ усиления признаков, связанных с урожайностью, предусматривающий введение и экспрессию в растении аллельного варианта любой из последовательностей нуклеиновых кислот, представленных в Таблице А Примера 1, или предусматривающий введение и экспрессию в растении аллельного варианта последовательности нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из полипептидных последовательностей, представленных в Таблице А Примера 1.
Аллельные варианты, используемые в одном варианте осуществления настоящего изобретения, имеют по существу такую же биологическую активность, как полипептид GRF согласно SEQ ID NO:2 и любая из полипептидных последовательностей, указанных в Таблице А Примера 1. Аллельные варианты существуют в природе, и способами по настоящему изобретению охватывается применение этих природных аллелей. Предпочтительно, аллельный вариант представляет собой аллельный вариант SEQ ID NO:1 или аллельный вариант последовательности нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:2. Предпочтительно, аллельный вариант представляет собой аллельный вариант полипептидной последовательности, содержащей: (i) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом QLQ, как представлено SEQ ID NO:115; и (ii) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом WRC, как представлено SEQ ID NO:116.
Аллельные варианты, используемые в одном варианте осуществления настоящего изобретения, имеют по существу такую же биологическую активность, как RAA1-подобный полипептид согласно SEQ ID NO:121 и любая из аминокислотных последовательностей, указанных в Таблице А Примера 1. Аллельные варианты существуют в природе, и способами по настоящему изобретению охватывается применение этих природных аллелей. Предпочтительно, аллельный вариант представляет собой аллельный вариант SEQ ID NO:120 или аллельный вариант последовательности нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:121. Предпочтительно, аминокислотная последовательность, кодируемая аллельным вариантом, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.8, объединяется в кластер с группой RAA1-подобных полипептидов, содержащей аминокислотную последовательность, представленную SEQ ID NO:121, а не с любой другой группой.
Аллельные варианты, используемые в одном варианте осуществления настоящего изобретения, имеют по существу такую же биологическую активность, как полипептид SYR согласно SEQ ID NO:169 и любая из аминокислотных последовательностей, указанных в Таблице А Примера 1. Аллельные варианты существуют в природе, и способами по настоящему изобретению охватывается применение этих природных аллелей. Предпочтительно, аллельный вариант представляет собой аллельный вариант SEQ ID NO:168 или аллельный вариант последовательности нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:169. Предпочтительно, аминокислотная последовательность, кодируемая аллельным вариантом, представляет собой полипептид от около 65 до около 200 аминокислот, содержащий обогащенный лейцином домен, как определено выше, перед которым находится консервативный трипептидный мотив 5 (а, b, с или а), и за которым следует консервативный мотив 6 и предпочтительно также консервативный мотив 7; или имеющий по меньшей мере 38% идентичности последовательности по отношению к последовательности SEQ ID NO:169.
Аллельные варианты, используемые в одном варианте осуществления настоящего изобретения, имеют по существу такую же биологическую активность, как полипептид ARKL согласно SEQ ID NO:213 и любая из аминокислотных последовательностей, указанных в Таблице А Примера 1. Аллельные варианты существуют в природе, и способами по настоящему изобретению охватывается применение этих природных аллелей. Предпочтительно, аллельный вариант представляет собой аллельный вариант SEQ ID NO:212 или аллельный вариант последовательности нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:213. Предпочтительно, аминокислотная последовательность, кодируемая аллельным вариантом, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.17, объединяется в кластер с группой полипептидов ARKL, содержащей аминокислотную последовательность, представленную SEQ ID NO:213, а не с любой другой группой, такой как представленная последовательностью Musmu_Goliath.
Аллельные варианты, используемые в одном варианте осуществления настоящего изобретения, имеют по существу такую же биологическую активность, как полипептид YTP согласно SEQ ID NO:409 и любая из аминокислотных последовательностей, указанных в Таблице А Примера 1. Аллельные варианты существуют в природе, и способами по настоящему изобретению охватывается применение этих природных аллелей. Предпочтительно, аллельный вариант представляет собой аллельный вариант SEQ ID NO:408 или аллельный вариант последовательности нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO:409. Предпочтительно, аминокислотная последовательность, кодируемая аллельным вариантом, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.21, объединяется в кластер с Группой 1, содержащей аминокислотную последовательность, представленную SEQ ID NO:409, а не с полипептидами YTP в Группе 2.
Перетасовка генов или направленная эволюция может также использоваться для получения вариантов последовательностей нуклеиновых кислот, кодирующих полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как определено выше; термин "перетасовка генов" является таким, как определено здесь.
Согласно настоящему изобретению предоставляется способ усиления признаков, связанных с урожайностью, предусматривающий введение и экспрессию в растении варианта любой из последовательностей нуклеиновых кислот, представленных в Таблице А Примера 1, или предусматривающий введение и экспрессию в растении варианта последовательности нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из полипептидных последовательностей, представленных в Таблице А Примера 1, при этом последовательность нуклеиновой кислоты варианта получают перетасовкой генов.
Предпочтительно, в одном варианте осуществления изобретения последовательность нуклеиновой кислоты варианта, полученная перетасовкой генов, кодирует полипептидную последовательность, содержащую: (i) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом QLQ, как представлено SEQ ID NO:115; и (ii) домен имеющий, по меньшей мере, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом WRC, как представлено SEQ ID NO:116.
Предпочтительно, в одном варианте осуществления изобретения аминокислотная последовательность, кодируемая нуклеиновой кислотой варианта, полученной перетасовкой гена, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.8, объединяется в кластер с группой RAA1-подобных полипептидов, содержащей аминокислотную последовательность, представленную SEQ ID NO:121, а не с любой другой группой.
Предпочтительно, в одном варианте осуществления изобретения аминокислотная последовательность, кодируемая нуклеиновой кислотой варианта, полученной перетасовкой гена, представляет собой полипептид от около 65 до около 200 аминокислот, содержащий обогащенный лейцином домен, как определено выше, перед которым находится консервативный трипептидный мотив 5 (а, b, с или а) и за которым следует консервативный мотив 6 и предпочтительно также консервативный мотив 7; или имеющий по меньшей мере 38% идентичности последовательности по отношению к последовательности SEQ ID NO:169.
Предпочтительно, в одном варианте осуществления изобретения аминокислотная последовательность, кодируемая нуклеиновой кислотой варианта, полученной перетасовкой гена, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.17, объединяется в кластер с группой полипептидов ARKL, содержащей аминокислотную последовательность, представленную SEQ ID NO:213, а не с любой другой группой, такой как представленная последовательностью Musmu_Goliath.
Предпочтительно, в одном варианте осуществления изобретения аминокислотная последовательность, кодируемая нуклеиновой кислотой варианта, полученной перетасовкой гена, при использовании в построении филогенетического дерева, такого как изображенное на Фиг.21, объединяется в кластер с Группой 1, содержащей аминокислотную последовательность, представленную SEQ ID NO:409, а не с полипептидами YTP в Группе 2.
Кроме того, варианты последовательностей нуклеиновых кислот могут также быть получены сайт-направленным мутагенезом. Доступно несколько способов для выполнения сайт-направленного мутагенеза, наиболее распространенными являются способы на основе ПЦР (Current Protocols in Molecular Biology, Wiley Eds.)
Последовательности нуклеиновых кислот, кодирующие полипептиды GRF, могут происходить из любого природного или искусственного источника. Последовательность нуклеиновой кислоты может быть модифицирована из ее нативной формы в составе и/или геномном окружении путем преднамеренной манипуляции человека. Предпочтительно, последовательность нуклеиновой кислоты, кодирующая полипептид GRF, происходит из растения, более того, предпочтительно из двудольного растения, более предпочтительно из семейства Brassicaceae, наиболее предпочтительно, последовательность нуклеиновой кислоты происходит из Arabidopsis thaliana.
Нуклеиновые кислоты, кодирующие RAA1-подобные полипептиды, могут происходить из любого природного или искусственного источника. Нуклеиновая кислота может быть модифицирована из ее нативной формы в составе и/или геномном окружении путем преднамеренной манипуляции человека. Предпочтительно, нуклеиновая кислота, кодирующая RAA1-подобный полипептид, происходит из растения, более того, предпочтительно из однодольного растения, более предпочтительно из семейства Роасеае, наиболее предпочтительно, нуклеиновая кислота происходит из Oryza sativa.
Нуклеиновые кислоты, кодирующие полипептиды SYR, могут происходить из любого природного или искусственного источника. Нуклеиновая кислота может быть модифицирована из ее нативной формы в составе и/или геномном окружении путем преднамеренной манипуляции человека. Предпочтительно, нуклеиновая кислота, кодирующая полипептид SYR, происходит из растения, более того, предпочтительно из однодольного растения, более предпочтительно из семейства Роасеае, наиболее предпочтительно, нуклеиновая кислота происходит из Oryza sativa.
Нуклеиновые кислоты, кодирующие полипептиды ARKL, могут происходить из любого природного или искусственного источника. Нуклеиновая кислота может быть модифицирована из ее нативной формы в составе и/или геномном окружении путем преднамеренной манипуляции человека. Предпочтительно, нуклеиновая кислота, кодирующая полипептид ARKL, происходит из растения, более того, предпочтительно из однодольного растения, более предпочтительно из семейства Роасеае, наиболее предпочтительно, нуклеиновая кислота происходит из Oryza sativa.
Нуклеиновые кислоты, кодирующие полипептиды YTP, могут также кодироваться de novo сконструированным полипептидом YTP, т.е. не происходить из природного источника. Нуклеиновая кислота может быть модифицирована из ее нативной формы в составе и/или геномном окружении путем преднамеренной манипуляции человека. Предпочтительно, нуклеиновая кислота, кодирующая полипептид YTP, происходит из растения, более того, предпочтительно из однодольного растения, более предпочтительно из семейства Роасеае, наиболее предпочтительно, нуклеиновая кислота происходит из Oryza sativa.
Выполнение способов по изобретению дает в одном варианте осуществления изобретения растения, имеющие улучшенные признаки, связанные с урожайностью. В частности, выполнение способов по изобретению дает растения, имеющие увеличенную урожайность, главным образом увеличенную урожайность семян по сравнению с контрольными растениями. Термины "урожайность" и "урожайность семян" описаны здесь более детально в разделе "определения".
Выполнение способов по изобретению дает в одном варианте осуществления изобретения растения, имеющие увеличенную устойчивость к абиотическому стрессу (или толерантность к абиотическому стрессу, эти термины используются взаимозаменяемо), осуществленную как улучшенные признаки, связанные с урожайностью, по сравнению с контрольными растениями при выращивании при абиотическом стрессе. В частности, выполнение способов по изобретению дает растения, имеющие увеличенную урожайность, главным образом увеличенную урожайность семян и увеличенную биомассу по сравнению с контрольными растениями. Термины "урожайность" и "урожайность семян" описаны здесь более детально в разделе "определения".
Предполагается, что ссылка здесь на улучшенные признаки, связанные с урожайностью, означает увеличение биомассы (массы) одной или более частей растения, которые могут включать надземные (заготавливаемые) части и/или подземные (заготавливаемые) части.
В одном варианте осуществления изобретения такими заготавливаемыми частями являются семена, и выполнение способов по изобретению приводит к растениям, имеющим увеличенную урожайность семян по сравнению с урожайностью семян контрольных растений.
В одном варианте осуществления изобретения такими заготавливаемыми частями являются корни, цветки и/или семена, и выполнение способов по изобретению приводит к растениям, имеющим увеличенные биомассу и/или урожайность семян по сравнению с урожайностью семян контрольных растений.
В одном варианте осуществления изобретения такими заготавливаемыми частями являются семена, и выполнение способов по изобретению приводит к растениям, имеющим увеличенные урожайность, общую массу семян, уровень наполнения семян, число цветков (или отдельных цветков), индекс урожайности и массу тысячи зерен по сравнению с контрольными растениями.
Если взять кукурузу в качестве примера, увеличение урожайности может проявляться как одно или более из следующего, среди прочих: увеличение числа растений, установленное на гектар или акр, увеличение числа початков на растение, увеличение числа рядов, числа зерен на ряд, массы зерен, массы тысячи зерен, длины початка/диаметра початка, увеличение уровня наполнения семян (который означает число наполненных семян, деленное на общее число семян и умноженное на 100). Если взять рис в качестве примера, увеличение урожайности может проявляться как увеличение одного или более из следующего, среди прочих: число растений на гектар или акр, число метелок на растение, число колосков на метелку, число цветков (отдельных цветков) на метелку (которое выражается как отношение числа наполненных семян к числу первичных метелок), увеличение уровня наполнения семян (который означает число наполненных семян, деленное на общее число семян и умноженное на 100), увеличение массы тысячи зерен.
Настоящее изобретение предоставляет способ усиления признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, при этом способ предусматривает увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептид GRF и RAA1-подобный полипептид соответственно, как здесь определено.
Настоящее изобретение предоставляет способ увеличения устойчивости к абиотическому стрессу у растений, приводящий к увеличенной урожайности, главным образом, урожайности семян и/или увеличенной биомассе растений, по сравнению с контрольными растениями, при выращивании в условиях абиотического стресса, при этом способ предусматривает модуляцию экспрессии, предпочтительно увеличение экспрессии, в растении нуклеиновой кислоты, кодирующей полипептид SYR, как здесь определено.
Настоящее изобретение предоставляет способ увеличения урожайности, главным образом урожайности семян растений, по сравнению с контрольными растениями, при этом способ предусматривает модуляцию экспрессии у растения нуклеиновой кислоты, кодирующей полипептид ARKL, как здесь определено.
Настоящее изобретение предоставляет способ увеличения урожайности, главным образом урожайности семян растений, по сравнению с контрольными растениями, при этом способ предусматривает модуляцию экспрессии у растения нуклеиновой кислоты, кодирующей полипептид YTP, как здесь определено.
Поскольку трансгенные растения согласно настоящему изобретению имеют усиленные признаки, связанные с урожайностью семян, вероятно, что эти растения показывают повышенную скорость роста (в течение по меньшей мере части их жизненного цикла) по сравнению со скоростью роста контрольных растений на соответствующей стадии их жизненного цикла.
Кроме того, увеличенная производительность, увеличенная эффективность поглощения питательных веществ могут также способствовать увеличению урожайности. Обнаружено, что растения согласно настоящему изобретению демонстрируют более высокую эффективность поглощения питательных веществ. Увеличенная эффективность поглощения питательных веществ обеспечивает лучший рост растения, когда растение находится в стрессовых условиях. Также обнаружено, что трансгенные растения согласно настоящему изобретению имеют увеличенную толерантность к стрессу, вызванному засухой, позволяя растениям продолжать расти в условиях, при которых замедляется или ингибируется рост контрольных растений.
Повышенная скорость роста может быть специфична для одной или более частей растения (включая семена) или может быть по существу по всему растению целиком. Растения, имеющие повышенную скорость роста, могут иметь более короткий жизненный цикл. Предполагается, что жизненный цикл растения означает время, необходимое для роста из сухого зрелого семени вплоть до стадии, где растение производит сухие зрелые семена, схожие с исходным материалом. На этот жизненный цикл можно влиять такими факторами, как ранняя мощность, скорость роста, индекс зелени, период цветения и скорость созревания семян. Повышение скорости роста может иметь место на одной или более стадий жизненного цикла растения или в течение по существу всего жизненного цикла растения. Увеличенная скорость роста во время ранних стадий жизненного цикла растения может отражать улучшенную (раннюю) мощность. Увеличение скорости роста может изменять цикл урожая растения, давая возможность высевать растения позже и/или собирать быстрее, чем было бы возможно иным образом (схожий эффект может быть получен для более раннего периода цветения; отсроченное цветение обычно не является желательным признаком у сельскохозяйственных культур). Если скорость роста достаточно увеличена, она может позволить дополнительный сев семян того же вида растений (например, сев и сбор рисовых растений с последующими севом и сбором дополнительных рисовых растений, и все во время одного стандартного периода роста). Также, если скорость роста достаточно увеличена, это может позволить дополнительный сев семян различных видов растений (например, сев и сбор кукурузных растений с последующим, например, севом и дополнительным сбором сои, картофеля или любого другого подходящего растения). Также возможны дополнительные разы сбора с одного подвоя в случае некоторых сельскохозяйственных культур. Изменение цикла урожая растения может привести к увеличению годовой продукции биомассы на акр (благодаря увеличению числа раз (например, за год), которое любое определенное растение может расти и собираться). Увеличение скорости роста может также позволить культивирование трансгенных растений на более широкой географической площади, чем их аналогов дикого типа, так как территориальные ограничения роста сельскохозяйственной культуры часто определяются неблагоприятными условиями окружающей среды или во время посадки (раннеспелые сорта) или во время сбора (позднеспелые). Таких неблагоприятных условий можно избежать, если укоротить цикл урожая. Скорость роста может быть определена получением различных параметров из кривых роста, такими параметрами могут быть, среди прочих: T-Mid (время, требуемое растениям для достижения 50% их максимального размера) и Т-90 (время, требуемое растениям для достижения 90% их максимального размера).
Согласно предпочтительному признаку настоящего изобретения выполнение способов по изобретению дает растения, имеющие увеличенную скорость роста по сравнению с контрольными растениями. Следовательно, согласно настоящему изобретению предоставляется способ увеличения скорости роста растений, при этом способ предусматривает увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF, или RAA1-подобный полипептид, или полипептид YTP, или полипептид ARKL, как здесь определено.
Согласно предпочтительному признаку настоящего изобретения выполнение способов по изобретению дает растения, имеющие увеличенную скорость роста по сравнению с контрольными растениями при выращивании в условиях абиотического стресса. Следовательно, согласно настоящему изобретению предоставляется способ увеличения скорости роста растений в условиях абиотического стресса, при этом способ предусматривает модуляцию экспрессии, предпочтительно увеличение экспрессии, в растении последовательности нуклеиновой кислоты, кодирующей полипептид SYR, как здесь определено.
Усиленные признаки, связанные с урожайностью семян, возникают, находится ли растение в условиях, не приводящих к стрессу, или растение подвергается воздействию различных стрессов по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Растения обычно отвечают на воздействие стресса более медленным ростом. В условиях сильного стресса растение может даже остановить рост полностью. С другой стороны, легкий стресс определен здесь как любой стресс, которому подвергается растение, который не приводит к растению, полностью прекратившему расти без возможности возобновить рост. Легкий стресс в рамках изобретения приводит к снижению роста растений, подвергшихся стрессу, меньше чем на 40%, 35% или 30%, предпочтительно меньше чем на 25%, 20% или 15%, более предпочтительно меньше чем на 14%, 13%, 12%, 11% или 10% или меньше по сравнению с контрольным растением в условиях, не приводящих к стрессу. Благодаря достижениям в агрономических приемах (ирригация, удобрение, обработки пестицидами) сильные стрессы не часто встречаются у культивируемых сельскохозяйственных культур. Как следствие, нарушенный рост, вызванный легким стрессом, часто является нежелательным признаком для сельского хозяйства. Легкие стрессы представляют собой ежедневные биотические и/или абиотические (относящиеся к окружающей среде) стрессы, которым подвергается растение. Абиотические стрессы могут быть вследствие засухи или избытка воды, анаэробного стресса, солевого стресса, химической токсичности, окислительного стресса и высоких, низких или отрицательных температур. Абиотический стресс может являться осмотическим стрессом, вызванным водным стрессом (особенно вследствие засухи), солевым стрессом, окислительным стрессом или ионным стрессом. Биотические стрессы обычно представляют собой стрессы, вызванные патогенами, такими как бактерии, вирусы, грибы, нематоды и насекомые. Термин "не приводящие к стрессу" условия при использовании здесь означает такие условия окружающей среды, которые обеспечивают оптимальный рост растений. Специалисты в данной области техники осведомлены о нормальных грунтовых условиях и климатических условиях для данного местоположения.
В частности, способы настоящего изобретения могут выполняться в условиях, не приводящих к стрессу, или в условиях умеренной засухи, давая растения, имеющие увеличенную урожайность по сравнению с контрольными растениями. Как сообщалось у Wang et al. (Planta (2003) 218:1-14), абиотический стресс приводит к сериям морфологических, физиологических, биохимических и молекулярных изменений, которые неблагоприятно влияют на рост растения и продуктивность. Известно, что засуха, засоленность, экстремальные температуры и окислительный стресс взаимосвязаны и могут вызывать нарушения роста и клеточные повреждения через похожие механизмы. Rabbani et al. (Plant Physiol (2003) 133:1755-1767) описывает особенно высокую степень "перекрестных влияний" между стрессом, вызванным засухой, и стрессом, вызванным высокой засоленностью. Например, засуха и/или засоление проявляются первоначально как осмотический стресс, приводящий к нарушению гомеостаза и распределения ионов в клетке. Окислительный стресс, который часто сопровождает высокую или низкую температуру, стресс, вызванный засухой, или стресс, вызванный засоленностью, могут вызывать денатурацию функциональных и структурных белков. Как следствие, эти разнообразные стрессы под влиянием условий среды часто активируют схожие клеточные пути передачи сигналов и клеточные ответы, например, продуцирование белков стрессов, активацию антиоксидантов, накопление совместимых осмолитов и задержку роста. Термин "не приводящие к стрессу" условия при использовании здесь означает такие условия окружающей среды, которые обеспечивают оптимальный рост растений. Специалисты в данной области техники осведомлены о нормальных грунтовых условиях и климатических условиях для данного местоположения.
Увеличение урожайности и/или скорости роста возникают, находится ли растение в условиях, не приводящих к стрессу, или растение подвергается воздействию различных стрессов по сравнению с контрольными растениями. Растения обычно отвечают на воздействие стресса более медленным ростом. В условиях сильного стресса растение может даже остановить рост полностью. С другой стороны, легкий стресс определен здесь как любой стресс, которому подвергается растение, который не приводит к растению, полностью прекратившему расти без возможности возобновить рост. Благодаря достижениям в агрономических приемах (ирригация, удобрение, обработки пестицидами) сильные стрессы не часто встречаются у культивируемых сельскохозяйственных культур. Как следствие, нарушенный рост, вызванный легким стрессом, часто является нежелательным признаком для сельского хозяйства. Легкие стрессы представляют собой ежедневные биотические и/или абиотические (относящиеся к окружающей среде) стрессы, которым подвергается растение. Абиотические стрессы могут быть вследствие засухи или избытка воды, анаэробного стресса, солевого стресса, химической токсичности, окислительного стресса и высоких, низких или отрицательных температур. Абиотический стресс может являться осмотическим стрессом, вызванным водным стрессом (особенно вследствие засухи), солевым стрессом, окислительным стрессом или ионным стрессом. Биотические стрессы обычно представляют собой стрессы, вызванные патогенами, такими как бактерии, вирусы, грибы, нематоды и насекомые.
Выполнение способов по изобретению дает в одном варианте осуществления изобретения растения, выращенные в условиях, не приводящих к стрессу, или в условиях легкого стресса, имеющие усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ усиления признаков, связанных с урожайностью, у растений, выращенных в условиях, не приводящих к стрессу, или в условиях легкого стресса, при этом способ предусматривает увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF.
Выполнение способов согласно настоящему изобретению приводит к растениям, выращенным в условиях абиотического стресса, имеющим усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями, выращенными в сопоставимых стрессовых условиях. Как сообщалось у Wang et al. (Planta (2003) 218:1-14), абиотический стресс приводит к сериям морфологических, физиологических, биохимических и молекулярных изменений, которые неблагоприятно влияют на рост растения и продуктивность. Известно, что засуха, засоленность, экстремальные температуры и окислительный стресс взаимосвязаны и могут вызывать нарушения роста и клеточные повреждения через похожие механизмы. Rabbani et al. (Plant Physiol (2003) 133:1755-1767) описывает особенно высокую степень "перекрестных влияний" между стрессом, вызванным засухой, и стрессом, вызванным высокой засоленностью. Например, засуха и/или засоление проявляются первоначально как осмотический стресс, приводящий к нарушению гомеостаза и распределения ионов в клетке. Окислительный стресс, который часто сопровождает высокую или низкую температуру, стресс, вызванный засухой, или стресс, вызванный засоленностью, могут вызывать денатурацию функциональных и структурных белков. Как следствие, эти разнообразные стрессы под влиянием условий среды часто активируют схожие клеточные пути передачи сигналов и клеточные ответы, например, продуцирование белков стрессов, активацию антиоксидантов, накопление совместимых осмолитов и задержку роста. Так как разнообразные стрессы под влиянием условий среды активируют схожие пути, иллюстративные примеры настоящего изобретения со стрессом, вызванным засухой, не следует рассматривать как ограничение стрессом, вызванным засухой, а в большей степени как тест, чтобы показать участие полипептидов GRF, как определено выше, в усилении признаков, связанных с урожайностью, по сравнению с контрольными растениями, выращенными в сопоставимых стрессовых условиях, в абиотических стрессах вообще.
Предполагается, что термин "абиотический стресс", как определено здесь, означает любой один или более из: водный стресс (вследствие засухи или избытка воды), анаэробный стресс, солевой стресс, температурный стресс (вследствие высоких, низких или отрицательных температур), стресс, вызванный химической токсичностью, и окислительный стресс. Согласно одному из аспектов изобретения абиотический стресс означает осмотический стресс, выбранный из водного стресса, солевого стресса, окислительного стресса и ионного стресса. Предпочтительно, водный стресс означает стресс, вызванный засухой. Термин «солевой стресс» не ограничивается поваренной солью (NaCl), но может являться любым стрессом, вызванным одним или более из, среди прочих: NaCl, KCl, LiCl, MgCl2, CaCl2.
Выполнение способов по изобретению дает растения, имеющие усиленные признаки, связанные с урожайностью семян, в условиях абиотического стресса по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ усиления признаков, связанных с урожайностью семян, у растений, выращенных в условиях абиотического стресса, при этом способ предусматривает увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF. Согласно одному аспекту изобретения абиотический стресс представляет собой осмотический стресс, выбранный из одного или более из следующих: водный стресс, солевой стресс, окислительный стресс и ионный стресс.
Другой пример абиотического стресса под влиянием условий среды представляет собой сниженную доступность одного или более питательных веществ, которые необходимы для усвоения растениями для роста и развития. Вследствие сильного влияния эффективности усвоения питательных веществ на урожайность растений и качество продуктов, огромное количество удобрений выливается на поля для оптимизирования роста и качества растений. Продуктивность растений обычно ограничивается тремя основными питательными веществами, фосфором, калием и азотом, который обычно является элементом, лимитирующим скорость, для роста растений из этих трех. Следовательно, главным питательным элементом, требуемым для роста растений, является азот (N). Он является составной частью многочисленных важных соединений, обнаруживаемых в живых клетках, включая аминокислоты, белки (ферменты), нуклеиновые кислоты и хлорофилл. От 1.5% до 2% сухого растительного вещества составляет азот и приблизительно 16% от общего растительного белка. Таким образом, доступность азота является главным лимитирующим фактором для роста и продукции сельскохозяйственных растений (Frink et al. (1999) Proc Natl Acad Sci USA 96(4):1175-1180), а также имеет основное влияние на накопление белка и аминокислотный состав. Следовательно, большой интерес представляют сельскохозяйственные растения с усиленными признаками, связанными с урожайностью, которые выращены в условиях лимитирования по азоту.
Выполнение способов по изобретению дает растения, выращенные в условиях сниженной доступности питательных веществ, в частности в условиях сниженной доступности азота, имеющие усиленные признаки, связанные с урожайностью семян, по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ усиления признаков, связанных с урожайностью семян, у растений, выращенных в условиях сниженной доступности питательных веществ, предпочтительно сниженной доступности азота, при этом способ предусматривает увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF. Сниженная доступность питательных веществ может являться результатом недостатка или избытка питательных веществ, таких как азот, фосфаты и другие содержащие фосфор соединения, калий, кальций, кадмий, магний, марганец, железо и бор, среди прочих. Предпочтительно, сниженная доступность питательных веществ представляет собой сниженную доступность азота.
Выполнение способов по изобретению дает в одном варианте осуществления изобретения растения, выращенные в условиях, не приводящих к стрессу, или в условиях умеренной засухи, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности у растений, выращенных в условиях, не приводящих к стрессу, или в условиях умеренной засухи, при этом способ предусматривает модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид.
Выполнение способов по изобретению дает в одном варианте осуществления растения, выращенные в условиях недостаточности питательных веществ, в частности в условиях азотной недостаточности, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности у растений, выращенных в условиях недостаточности питательных веществ, при этом способ предусматривает модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид. Недостаточность питательных веществ может являться результатом дефицита питательных веществ, таких как азот, фосфаты и другие содержащие фосфор соединения, калий, кальций, кадмий, магний, марганец, железо и бор, среди прочих.
Выполнение способов по изобретению дает растения, выращенные в условиях абиотического стресса, таких как условия, вызванные засухами, от умеренной засухи до сильной, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности в растениях, выращенных в условиях абиотического стресса, таких как условия, вызванные засухами, от умеренной засухи до сильной, при этом способ предусматривает увеличение экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYR. Термины "условия, вызванные сильной засухой" или "стресс, вызванный сильной засухой", как использовано здесь, представляют собой те условия, вызванные засухой, которые вызывают снижение урожайности на 50% или более в контрольных растениях по сравнению с урожайностью контрольных растениях, выращенных в условиях, не приводящих к стрессу.
Выполнение способов по изобретению дает в одном варианте осуществления растения, выращенные в условиях недостаточности питательных веществ, в частности в условиях азотной недостаточности, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности у растений, выращенных в условиях недостаточности питательных веществ, при этом способ предусматривает увеличение экспрессии у растения нуклеиновой кислоты, кодирующей полипептид SYR. Недостаточность питательных веществ может являться результатом дефицита питательных веществ, таких как азот, фосфаты и другие содержащие фосфор соединения, калий, кальций, кадмий, магний, марганец, железо и бор, среди прочих.
Выполнение способов по изобретению дает растения, выращенные в условиях, не приводящих к стрессу, или в условиях умеренной засухи, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности у растений, выращенных в условиях, не приводящих к стрессу, или в условиях умеренной засухи, при этом способ предусматривает модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид ARKL.
Выполнение способов по изобретению дает в одном варианте осуществления растения, выращенные в условиях недостаточности питательных веществ, в частности в условиях азотной недостаточности, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности у растений, выращенных в условиях недостаточности питательных веществ, при этом способ предусматривает увеличение экспрессии у растения нуклеиновой кислоты, кодирующей полипептид ARKL. Недостаточность питательных веществ может являться результатом дефицита питательных веществ, таких как азот, фосфаты и другие содержащие фосфор соединения, калий, кальций, кадмий, магний, марганец, железо и бор, среди прочих.
Выполнение способов по изобретению дает растения, выращенные в условиях, не приводящих к стрессу, или в условиях умеренной засухи, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности у растений, выращенных в условиях, не приводящих к стрессу, или в условиях умеренной засухи, при этом способ предусматривает модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид YTP.
Выполнение способов по изобретению дает растения, выращенные в условиях недостаточности питательных веществ, в частности в условиях азотной недостаточности, имеющие увеличенную урожайность по сравнению с контрольными растениями, выращенными в сопоставимых условиях. Следовательно, согласно настоящему изобретению предоставляется способ увеличения урожайности у растений, выращенных в условиях недостаточности питательных веществ, при этом способ предусматривает увеличение экспрессии у растения нуклеиновой кислоты, кодирующей полипептид YTP. Недостаточность питательных веществ может являться результатом дефицита питательных веществ, таких как азот, фосфаты и другие содержащие фосфор соединения, калий, кальций, кадмий, магний, марганец, железо и бор, среди прочих.
Настоящее изобретение охватывает растения, или их части (включая семена), или их клетки, получаемые способами согласно настоящему изобретению. Растения, или их части, или их клетки содержат трансген нуклеиновой кислоты, кодирующий полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как определено выше.
Изобретение также предоставляет генетические конструкции и векторы для облегчения введения и/или увеличенной экспрессии в растениях последовательностей нуклеиновых кислот, кодирующих полипептиды, выбранные из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно. Генные конструкции могут вставляться в векторы, которые могут иметься в продаже, пригодные для трансформации в растения и для экспрессии рассматриваемого гена в трансформированных клетках. Изобретение также обеспечивает применение генной конструкции, как здесь определено, в способах по изобретению.
Более конкретно, настоящее изобретение обеспечивает в одном варианте осуществления изобретения конструкцию, содержащую:
(a) последовательность нуклеиновой кислоты, кодирующую полипептид GRF, как определено выше;
(b) одну или более контрольных последовательностей, способных увеличивать экспрессию последовательности нуклеиновой кислоты (а); и при необходимости
(c) последовательность терминации транскрипции.
Предпочтительно, последовательность нуклеиновой кислоты, кодирующая полипептид GRF, является такой, как определено выше. Термины "контрольная последовательность" и "последовательность терминации" являются такими, как здесь определено.
Предпочтительно, одна из контрольных последовательностей конструкции представляет собой конститутивный промотор, выделенный из генома растения. Примером растительного конститутивного промотора является промотор GOS2, предпочтительно промотор GOS2 риса, более предпочтительно промотор GOS2, как представлено SEQ ID NO:117.
Более конкретно, настоящее изобретение обеспечивает в одном варианте осуществления изобретения конструкцию, содержащую:
(a) нуклеиновую кислоту, кодирующую RAA1-подобный полипептид, как определено выше;
(b) одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а); и при необходимости
(c) последовательность терминации транскрипции.
Предпочтительно, нуклеиновая кислота, кодирующая RAA1-подобный полипептид, является такой, как определено выше. Термины "контрольная последовательность" и "последовательность терминации" являются такими, как здесь определено.
Более конкретно, настоящее изобретение обеспечивает в одном варианте осуществления изобретения конструкцию, содержащую:
1. нуклеиновую кислоту, кодирующую полипептид SYR, как определено выше;
2. одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а); и при необходимости
3. последовательность терминации транскрипции.
Предпочтительно, нуклеиновая кислота, кодирующая полипептид SYR, является такой, как определено выше. Термины "контрольная последовательность" и "последовательность терминации" являются такими, как здесь определено.
Более конкретно, настоящее изобретение обеспечивает в одном варианте осуществления изобретения конструкцию, содержащую:
1) нуклеиновую кислоту, кодирующую полипептид ARKL, как определено выше;
2) одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а) и при необходимости
3) последовательность терминации транскрипции.
Предпочтительно, нуклеиновая кислота, кодирующая полипептид ARKL, является такой, как определено выше. Термины "контрольная последовательность" и "последовательность терминации" являются такими, как здесь определено.
Более конкретно, настоящее изобретение обеспечивает в одном варианте осуществления изобретения конструкцию, содержащую:
1) нуклеиновую кислоту, кодирующую полипептид YTP, как определено выше;
2) одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а) и при необходимости
3) последовательность терминации транскрипции.
Предпочтительно, нуклеиновая кислота, кодирующая полипептид YTP, является такой, как определено выше. Термины "контрольная последовательность" и "последовательность терминации" являются такими, как здесь определено.
Растения трансформируют вектором, содержащим любую из последовательностей нуклеиновых кислот, описанных выше. Специалист в данной области техники хорошо осведомлен о генетических элементах, которые должны присутствовать в векторе для того, чтобы удачно трансформировать, отбирать и размножать клетки-хозяева, содержащие рассматриваемую последовательность. Рассматриваемая последовательность функционально связана с одной или более контрольных последовательностей (по меньшей мере, с промотором).
Преимущественно любой тип промотора, природный или синтетический, может использоваться для увеличения экспрессии последовательности нуклеиновой кислоты. Конститутивный промотор в частности применим в способах, предпочтительно конститутивный промотор выделен из генома растения. Растительный конститутивный промотор управляет экспрессией кодирующей последовательности на уровне, который во всех примерах ниже, чем полученный под контролем вирусного промотора 35S CaMV.
Другие органоспецифичные промоторы, например, для предпочтительной экспрессии в листьях, стеблях, клубнях, меристемах, семенах (зародыше и/или эндосперме) применимы для выполнения способов по изобретению. Смотри здесь раздел «Определения» относительно определений различных типов промоторов.
Предпочтительно в одном варианте осуществления конститутивный промотор является также убиквитарным промотором. Смотри здесь раздел «Определения» относительно определений различных типов промоторов.
Очевидно, что применимость настоящего изобретения не ограничена последовательностью нуклеиновой кислоты, кодирующей полипептид GRF, как представлено SEQ ID NO:1, также применимость изобретения не ограничена экспрессией последовательности нуклеиновой кислоты, кодирующей полипептид GRF, которая управляется конститутивным промотором.
Очевидно, что применимость настоящего изобретения не ограничена нуклеиновой кислотой, кодирующей RAA1-подобный полипептид, представленной SEQ ID NO:120, также применимость изобретения не ограничена экспрессией нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, которая управляется конститутивным промотором.
Конститутивный промотор является предпочтительно промотором GOS2, предпочтительно промотором GOS2 из риса. Также предпочтительно конститутивный промотор представлен последовательностью нуклеиновой кислоты, по существу сходной с SEQ ID NO:124, наиболее предпочтительно промотор GOS2 представлен SEQ ID NO:124 или SEQ ID:211. Согласно другому предпочтительному признаку изобретения, конститутивный промотор представляет собой промотор белка группы с высокой подвижностью (HMGP), предпочтительно промотор HMGP из риса, более предпочтительно, по существу сходный с SEQ ID NO:125, наиболее предпочтительно, идентичный SEQ ID NO:125. См. Таблицу 2 здесь в разделе "Определения" на предмет дополнительных примеров конститутивных промоторов.
Очевидно, что применимость настоящего изобретения не ограничена нуклеиновой кислотой, кодирующей полипептид SYR, представленной SEQ ID NO:168, также применимость изобретения не ограничена экспрессией нуклеиновой кислоты, кодирующей полипептид SYR, которая управляется конститутивным промотором.
Конститутивный промотор является предпочтительно промотором GOS2, предпочтительно промотором GOS2 из риса. Также предпочтительно конститутивный промотор представлен последовательностью нуклеиновой кислоты, по существу сходной с SEQ ID NO:172 или SEQ ID NO:211, наиболее предпочтительно конститутивный промотор представлен SEQ ID NO:172 или SEQ ID NO:211. См. Таблицу 2 здесь в разделе "Определения" на предмет дополнительных примеров применимых конститутивных промоторов.
Очевидно, что применимость настоящего изобретения не ограничена нуклеиновой кислотой, кодирующей полипептид ARKL, представленной SEQ ID NO:212, также применимость изобретения не ограничена экспрессией нуклеиновой кислоты, кодирующей полипептид ARKL, которая управляется конститутивным промотором или которая управляется промотором, специфичным для корней.
Конститутивный промотор является предпочтительно промотором GOS2, предпочтительно промотором GOS2 из риса. Также предпочтительно конститутивный промотор представлен последовательностью нуклеиновой кислоты, по существу сходной с SEQ ID NO:406, наиболее предпочтительно конститутивный промотор представлен SEQ ID NO:406 или SEQ ID NO:211. См. Таблицу 2 здесь в разделе "Определения" на предмет дополнительных примеров конститутивных промоторов.
Очевидно, что применимость настоящего изобретения не ограничена нуклеиновой кислотой, кодирующей полипептид YTP, представленной SEQ ID NO:1 также применимость изобретения не ограничена экспрессией нуклеиновой кислоты, кодирующей полипептид YTP, которая управляется конститутивным промотором.
Конститутивный промотор является предпочтительно промотором GOS2, предпочтительно промотором GOS2 из риса. Также предпочтительно конститутивный промотор представлен последовательностью нуклеиновой кислоты, по существу сходной с SEQ ID NO:548, наиболее предпочтительно конститутивный промотор представлен SEQ ID NO:548 или SEQ ID NO:211. См. Таблицу 2 здесь в разделе "Определения" на предмет дополнительных примеров конститутивных промоторов.
При необходимости, одна или более терминаторных последовательностей могут использоваться в конструкции, введенной в растение. Дополнительные регуляторные элементы могут включать транскрипционные, а также трансляционные усилители. Специалисты в данной области техники осведомлены о последовательностях терминаторов и усилителей, которые могут подходить для применения при выполнении изобретения. Также может быть добавлена интронная последовательность на 5' нетранслируемый участок (UTR) или в кодирующую последовательность, чтобы увеличить количество зрелой мРНК, которая накапливается в цитозоле, как описано в разделе определений. Другие контрольные последовательности (кроме промотора, усилителя, сайленсера, интронных последовательностей, 3'UTR и/или 5'UTR участков) могут представлять собой белковые и/или РНК-стабилизирующие элементы. Такие последовательности могут быть известны или легко получены специалистом в данной области техники.
При необходимости, одна или более терминаторных последовательностей могут использоваться в конструкции, введенной в растение. Предпочтительно, конструкция содержит экспрессионную кассету, по существу схожую или идентичную с SEQ ID NO:166, содержащую промотор GOS2, нуклеиновую кислоту, кодирующую RAA1-подобный полипептид. В альтернативном варианте осуществления конструкция содержит экспрессионную кассету, по существу схожую или идентичную с SEQ ID NO:167, содержащую промотор HMGP, нуклеиновую кислоту, кодирующую RAA1-подобный полипептид.
При необходимости, одна или более терминаторных последовательностей могут использоваться в конструкции, введенной в растение. Предпочтительно, конструкция содержит экспрессионную кассету, по существу схожую или идентичную с SEQ ID NO:407, содержащую промотор GOS2, нуклеиновую кислоту, кодирующую полипептид Orysa_ARKL1 и последовательность терминатора транскрипции Т-зеин + T-rubisco.
При необходимости, одна или более терминаторных последовательностей могут использоваться в конструкции, введенной в растение. Предпочтительно, конструкция содержит экспрессионную кассету, по существу схожую или идентичную с SEQ ID NO:549, содержащую промотор GOS2, нуклеиновую кислоту, кодирующую полипептид Orysa_ARKL1 и последовательность терминатора транскрипции T-zein + T-rubisco.
Дополнительные регуляторные элементы могут включать транскрипционные, а также трансляционные энхансеры. Специалисты в данной области техники осведомлены о последовательностях терминаторов и энхансеров, которые могут подходить для применения при выполнении изобретения. Также может быть добавлена интронная последовательность на 5' нетранслируемый участок (UTR) или в кодирующую последовательность, чтобы увеличить количество зрелой мРНК, которая накапливается в цитозоле, как описано в разделе определений. Другие контрольные последовательности (кроме промотора, энхансера, сайленсера, интронных последовательностей, 3'UTR и/или 5'UTR участков) могут представлять собой белковые и/или РНК-стабилизирующие элементы. Такие последовательности могут быть известны или легко получены специалистом в данной области техники.
Генетические конструкции по изобретению могут дополнительно включать последовательность точки начала репликации, которая требуется для поддержания и/или репликации в определенном типе клеток. Одним из таких примеров является случай, когда генетическую конструкцию необходимо поддерживать в бактериальной клетке как эписомальный генетический элемент (например, молекулы плазмиды или космиды). Предпочтительные точки начала репликации включают, но не ограничиваются этим, f1-ori и colE1.
Для обнаружения удачного переноса последовательностей нуклеиновых кислот, как, например, использованных в способах изобретения, и/или отбора трансгенных растений, содержащих эти последовательности нуклеиновых кислот, предпочтительно использовать маркерные гены (или репортерные гены). Следовательно, генетическая конструкция может при необходимости содержать селектируемый маркерный ген. Селектируемые маркеры описываются более подробно здесь в разделе "определения".
Маркерные гены могут удаляться или вырезаться из трансгенной клетки после того, как они больше не нужны. Методики по удалению маркерных генов известны в данной области техники, и применимые методики описаны выше в разделе определений.
Известно, что при стабильной или временной интеграции последовательностей нуклеиновых кислот в растительные клетки только меньшая часть клеток принимает чужеродную ДНК и, если желательно, интегрирует ее в свой геном в зависимости от использованных вектора экспрессии и метода трансфекции. Для определения и отбора таких интегрантов ген, кодирующий селектируемый маркер (такой как описанные выше), обычно вводят в клетки-хозяева вместе с рассматриваемым геном. Эти маркеры могут, например, использоваться в мутантах, в которых эти гены являются нефункциональными в результате, например, делеции стандартными способами. Кроме того, молекулы последовательностей нуклеиновых кислот, кодирующих селектируемый маркер, могут вводиться в клетку-хозяина в том же векторе, который содержит последовательность, кодирующую полипептиды по изобретению, или используемую в способах по изобретению, или еще в отдельном векторе. Клетки, которые стабильно трансфицированы введенной последовательностью нуклеиновой кислоты, могут обнаруживаться, например, отбором (например, клетки, которые интегрировали селектируемый маркер, выживают, тогда как другие клетки погибают). Маркерные гены могут удаляться или вырезаться из трансгенной клетки после того, как они больше не нужны. Методики по удалению маркерных генов известны в данной области техники, и применимые методики описаны выше в разделе определений.
Изобретение также обеспечивает способ получения трансгенных растений, имеющих усиленные признаки, связанные с урожайностью семян, по сравнению с контрольными растениями, предусматривающий введение и экспрессию в растении любой последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как здесь определено выше.
Более конкретно, настоящее изобретение предоставляет в одном варианте осуществления изобретения способ получения трансгенных растений, имеющих усиленные признаки, связанные с урожайностью семян, по сравнению с контрольными растениями, причем этот способ предусматривает:
(i) введение и экспрессию в растении, части растения или растительной клетке последовательности нуклеиновой кислоты, кодирующей полипептид GRF, под контролем растительного конститутивного промотора;
(ii) культивирование растительной клетки, части растения или растения в условиях, способствующих росту и развитию растений.
Последовательность нуклеиновой кислоты (i) может быть любой из последовательностей нуклеиновых кислот, способных кодировать полипептид GRF, как здесь определено.
Более конкретно, настоящее изобретение предоставляет в одном варианте осуществления изобретения способ получения трансгенных растений, имеющих усиленные, улучшенные признаки, связанные с урожайностью семян, в частности увеличенные биомассу и/или урожайность семян, причем этот способ предусматривает:
i) введение и экспрессию в растении или растительной клетке нуклеиновой кислоты, кодирующей RAA1-подобный полипептид;
ii) культивирование растительной клетки в условиях, способствующих росту и развитию растений.
Нуклеиновая кислота (i) может быть любой из нуклеиновых кислот, способных кодировать RAA1-подобный полипептид, как здесь определено.
Более конкретно, настоящее изобретение предоставляет в одном варианте осуществления изобретения способ получения трансгенных растений, имеющих усиленные, улучшенные признаки, связанные с урожайностью семян, в частности увеличенную урожайность (семян) и/или увеличенную биомассу, причем этот способ предусматривает:
(i) введение и экспрессию в растении или растительной клетке нуклеиновой кислоты, кодирующей полипептид SYR;
(ii) культивирование растительной клетки в условиях, способствующих росту и развитию растений.
Последовательность нуклеиновой кислоты (i) может быть любой из нуклеиновых кислот, способных кодировать полипептид SYR, как здесь определено.
Более конкретно, настоящее изобретение предоставляет в одном варианте осуществления изобретения способ получения трансгенных растений, имеющих усиленные, улучшенные признаки, связанные с урожайностью семян, в частности увеличенную урожайность (семян), причем этот способ предусматривает:
(i) введение и экспрессию в растении или растительной клетке нуклеиновой кислоты, кодирующей полипептид ARKL;
(ii) культивирование растительной клетки в условиях, способствующих росту и развитию растений.
Нуклеиновая кислота (i) может быть любой из нуклеиновых кислот, способных кодировать полипептид ARKL, как здесь определено.
Более конкретно, настоящее изобретение предоставляет в одном варианте осуществления изобретения способ получения трансгенных растений, имеющих усиленные, улучшенные признаки, связанные с урожайностью семян, в частности увеличенную урожайность (семян), причем этот способ предусматривает:
i) введение и экспрессию в растении или растительной клетке нуклеиновой кислоты, кодирующей полипептид YTP;
ii) культивирование растительной клетки в условиях, способствующих росту и развитию растений.
Нуклеиновая кислота (i) может быть любой из нуклеиновых кислот, способных кодировать полипептид YTP, как здесь определено.
Последовательность нуклеиновой кислоты может вводиться непосредственно в растительную клетку или в само растение (включая введение в ткань, орган или любую другую часть растения). Согласно предпочтительному признаку настоящего изобретения последовательность нуклеиновой кислоты предпочтительно вводится в растение путем трансформации. Термин "трансформация" описывается более подробно здесь в разделе "определения".
Генетически модифицированные растительные клетки могут регенерироваться посредством всех способов, с которыми знаком специалист в данной области техники. Подходящие способы могут быть обнаружены в вышеупомянутых публикациях S.D. Kung и R. Wu, Potrykus или Hofgen и Willmitzer.
Как правило, после трансформации растительные клетки или группы клеток отбираются по присутствию одного или более маркеров, которые кодируются экспрессируемыми в растении генами, перенесенными совместно с рассматриваемым геном, после чего трансформированный материал регенерируется в целое растение. Чтобы отобрать трансформированные растения, растительный материал, полученный при трансформации, подвергают, как правило, влиянию селективных условий для того, чтобы трансформированные растения можно было отличить от нетрансформированных растений. Например, семена, полученные вышеуказанным способом, могут быть посажены и после начального периода вегетации подвергнуты подходящему отбору путем опрыскивания. Дополнительная возможность заключается в выращивании семян, если уместно, после стерилизации, на чашках с агаровой средой с использованием подходящего агента отбора для того, чтобы только трансформированные семена могли вырастать в растения. Альтернативно, трансформированные растения подвергают скринингу на присутствие селектируемого маркера, как, например, маркеров, описанных выше.
После переноса ДНК и регенерации предполагаемые трансформированные растения могут также оцениваться, например, используя Саузерн-анализ, на присутствие рассматриваемого гена, число копий и/или геномную организацию. Альтернативно или дополнительно, за уровнями экспрессии нововведенной ДНК можно наблюдать, используя Нозерн- и/или Вестерн-анализ, обе эти методики хорошо известны специалистам в данной области техники.
Полученные трансформированные растения могут размножаться различными способами, например, клональным размножением или классическими методами селекции. Например, трансформированное растение первого поколения (или Т1) может самоопыляться, и отбираются гомозиготные трансформанты второго поколения (или Т2), и растения Т2 могут затем дополнительно размножаться посредством классических методов селекции. Полученные трансформированные организмы могут принимать различные формы. Например, они могут быть химерами трансформированных клеток и нетрансформированных клеток; клональными трансформантами (например, все трансформированные клетки, содержащие экспрессионную кассету); трансплантатами трансформированных и нетрансформированных тканей (например, в растениях трансформированный подвой с привитым нетрансформированным привоем).
Настоящее изобретение, несомненно, распространяется на любую растительную клетку или растение, полученные любым из способов, описанных здесь, и на все их части растений и растительные материалы, используемые для размножения. Кроме того, настоящее изобретение также охватывает потомство первичных трансформированных или трансфицированных клетки, ткани, органа или целого растения, которое получено любым из вышеуказанных способов, причем единственное требование заключается в том, что потомство проявляет такой(ие) же генотипический(е) и/или фенотипический(е) признак(и), как производимые родителем в способах согласно изобретению.
Изобретение также включает клетки-хозяева, содержащие выделенную последовательность нуклеиновой кислоты, кодирующую полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, как здесь определено выше, функционально связанную с растительным конститутивным промотором. Предпочтительными клетками-хозяевами согласно настоящему изобретению являются растительные клетки. Растения-хозяева для нуклеиновых кислот или вектора, использованных в способе согласно изобретению, экспрессионной кассеты, или конструкции, или вектора представляют собой в принципе преимущественно все растения, которые способны синтезировать полипептиды, используемые в способе по изобретению.
Способы по изобретению являются преимущественно применимыми к любому растению. Растения, которые в частности применимы в способах по изобретению, включают все растения, которые принадлежат к суперсемейству Viridiplantae, в частности однодольные и двудольные растения, включающие кормовые или фуражные бобовые растения, декоративные растения, продовольственные культуры, деревья или кустарники. Согласно предпочтительному варианту осуществления изобретения растение представляет собой сельскохозяйственную культуру. Примеры сельскохозяйственных культур включают сою, подсолнечник, канолу, люцерну, семена рапса, хлопок, томат, картофель и табак. Кроме того, растение предпочтительно является однодольным растением. Примеры однодольных растений включают сахарный тростник. Более предпочтительно, растение является зерновой культурой. Примеры зерновых культур включают рис, кукурузу, пшеницу, ячмень, просо, рожь, тритикале, сорго и овес.
Изобретение также распространяется на заготавливаемые части растения, содержащего выделенную последовательность нуклеиновой кислоты, кодирующую полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно (как определено здесь выше), функционально связанную с растительным конститутивным промотором, такие как, но не ограничиваясь этим, семена, листья, плоды, цветки, стебли, корневища, клубни и луковицы. Изобретение также относится к продуктам, полученным, предпочтительно непосредственно полученным, из заготавливаемой части такого растения, например, к сухим гранулам или порошкам, маслу, жиру и жирным кислотам, крахмалу или белкам.
Согласно предпочтительному признаку изобретения модулированная экспрессия представляет собой увеличенную экспрессию. Способы увеличения экспрессии последовательностей нуклеиновых кислот, или генов, или генных продуктов хорошо задокументированы в данной области, и примеры предоставлены в разделе определений.
Как упомянуто выше, предпочтительный способ увеличения экспрессии последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, заключается во введении и экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно; однако результаты выполнения способа, т.е. усиление признаков, связанных с урожайностью семян, могут также достигаться при использовании других хорошо известных методик, включая, но не ограничиваясь этим, активационное мечение Т-ДНК, TILLING, гомологичную рекомбинацию. Описание этих методов представлено в разделе определений.
Настоящее изобретение также охватывает в одном варианте осуществления применение последовательностей нуклеиновых кислот, кодирующих полипептиды GRF, как здесь описано, и применение этих полипептидов GRF в усилении любых из вышеупомянутых признаков, связанных с урожайностью, в растениях в нормальных условиях роста, в условиях роста при абиотическом стрессе (предпочтительно, в условиях роста при осмотическом стрессе) и в условиях роста при сниженной доступности питательных веществ, предпочтительно при условиях сниженной доступности азота.
Настоящее изобретение также охватывает в одном варианте осуществления применение нуклеиновых кислот, кодирующих RAA1-подобные полипептиды, как здесь описаны, и применение этих RAA1-подобных полипептидов в улучшении любого из вышеупомянутых признаков, связанных с урожайностью, у растений.
Настоящее изобретение также охватывает в одном варианте осуществления применение нуклеиновых кислот, кодирующих полипептиды SYR, как здесь описано, и применение этих полипептидов SYR в улучшении любого из вышеупомянутых признаков, связанных с урожайностью, у растений при выращивании в условиях абиотического стресса.
Настоящее изобретение также охватывает в одном варианте осуществления применение нуклеиновых кислот, кодирующих полипептиды ARKL, как здесь описано, и применение этих полипептидов ARKL в улучшении любого из вышеупомянутых признаков, связанных с урожайностью, у растений.
Настоящее изобретение также охватывает в одном варианте осуществления применение нуклеиновых кислот, кодирующих полипептиды YTP, как здесь описано, и применение этих полипептидов YTP в улучшении любого из вышеупомянутых признаков, связанных с урожайностью, у растений.
Нуклеиновые кислоты, кодирующие полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, описанные здесь, или сами полипептиды по изобретению могут найти применение в селекционных программах, в которых идентифицируется ДНК-маркер, который может быть генетически связан с геном, кодирующим полипептид GRF. Гены/ последовательности нуклеиновых кислот или сами полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно могут использоваться для определения молекулярного маркера. Этот ДНК- или белковый маркер может затем использоваться в селекционных программах для отбора растений, имеющих усиленные признаки, связанные с урожайностью, как определено выше, в способах по изобретению.
Аллельные варианты гена/ последовательности нуклеиновой кислоты, кодирующих полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, могут также находить применение в селекционных программах, выполняемых с помощью маркера. Такие селекционные программы иногда требуют введения аллельного варианта мутагенной обработкой растений с применением, например, EMS-мутагенеза; альтернативно, программа может начинать с коллекции аллельных вариантов так называемого "природного" происхождения, вызванного умышленно. Затем имеет место идентификация аллельных вариантов, например, путем ПЦР. За этим следует стадия отбора лучших аллельных вариантов рассматриваемой последовательности, которые дают увеличенную урожайность. Отбор обычно проводят путем контроля за характеристиками роста растений, содержащих различные аллельные варианты рассматриваемой последовательности. Характеристики роста могут контролироваться в теплице или на поле. Дополнительные необязательные стадии включают скрещивающиеся растения, в которых лучший аллельный вариант присоединяется к другому растению. Это может использоваться, например, для получения комбинации интересных фенотипических признаков.
Последовательности нуклеиновых кислот, кодирующие полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, могут также использоваться как зонды для генетического и физического картирования генов, частью которых они являются, и как маркеры для признаков, связанных с этими генами. Такая информация может использоваться в селекции растений для того, чтобы создавать линии с желательными фенотипами. Такое применение последовательностей нуклеиновых кислот, кодирующих полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, требует только последовательность нуклеиновой кислоты по меньшей мере 15 нуклеотидов в длину. Последовательности нуклеиновых кислот, кодирующие полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно, могут использоваться как маркеры полиморфизма длины фрагментов рестрикции (RFLP). Саузерн-блоты (Sambrook J, Fritsch EF and Maniatis Т (1989) Molecular Cloning, A Laboratory Manual) расщепленной рестрикцией геномной ДНК растений могут зондироваться последовательностями нуклеиновых кислот, кодирующими полипептид, выбранный из группы, состоящей из: полипептид GRF, RAA1-подобный полипептид, полипептид SYR, полипептид ARKL и полипептид YTP соответственно. Получающиеся паттерны распределения полос могут затем подвергаться генетическому анализу с использованием компьютерных программ, таких как MapMaker (Lander et al. (1987) Genomics 1:174-181), для того, чтобы сконструировать генетическую карту. Кроме того, последовательности нуклеиновых кислот могут использоваться для зондирования Саузерн-блотов, содержащих геномные ДНК, обработанные рестриктазами, группы объектов, представляющих родителя и потомство определенного генетического скрещивания. Сегрегацию ДНК-полиморфизмов отмечают и используют для расчета положения последовательности нуклеиновой кислоты, кодирующей полипептид GRF, на генетической карте, предварительно полученной с использованием этой популяции (Botstein et al. (1980) Am. J. Hum. Genet. 32:314-331).
Получение и применение зондов из генов растений для использования в генетическом картировании описывается у Bernatzky и Tanksley (1986) Plant Mol. Biol. Reporter 4:37-41. Многочисленные публикации описывают генетическое картирование специфических кДНК-клонов с применением методологии, описанной выше, или ее вариаций. Например, F2 популяции от интеркросса, популяции от обратного скрещивания, популяции от случайного скрещивания, близко изогенные линии и другие группы объектов могут использоваться для картирования. Такие методологии хорошо известны специалисту в данной области техники.
Зонды последовательностей нуклеиновых кислот можно также использовать для физического картирования (т.е. размещения последовательностей на физических картах; см. Hoheisel et al. In: Non-mammalian Genomic Analysis: A Practical Guide, Academic press 1996, pp.319-346, и упомянутые там ссылки).
В другом варианте осуществления изобретения зонды последовательностей нуклеиновых кислот могут использоваться в картировании путем прямой флуоресцентной гибридизации in situ (FISH) (Trask (1991) Trends Genet. 7:149-154). Хотя текущие способы FISH-картирования предпочитают использование больших клонов (от нескольких т.п.н. до нескольких сотен т.п.н.; см. Laan et al. (1995) Genome Res. 5:13-20), усовершенствования в чувствительности могут позволить проведение FISH-картирования при использовании более коротких зондов.
Ряд способов генетического и физического картирования на основе амплификации последовательностей нуклеиновых кислот может проводиться с использованием последовательностей нуклеиновых кислот. Примеры включают аллель-специфическую амплификацию (Kazazian (1989) J. Lab. Clin. Med 11:95-96), полиморфизм ПЦР-амплифицированных фрагментов (CAPS; Sheffield et al. (1993) Genomics 16:325-332), аллель-специфическое лигирование (Landegren et al. (1988) Science 241:1077-1080), реакции удлинения нуклеотидных цепей (Sokolov (1990) Nucleic Acid Res. 18:3671), картирование с помощью облученных гибридов (Walter et al. (1997) Nat. Genet. 7:22-28) и HAPPY картирование (Dear and Cook (1989) Nucleic Acid Res. 17:6795-6807). В случае этих способов последовательность последовательности нуклеиновой кислоты используется для конструирования и получения пары праймеров для применения в реакции амплификации или реакциях удлинения праймеров. Конструирование таких праймеров хорошо известно специалистам в данной области техники. В способах, использующих генетическое картирование на основе ПЦР, может быть необходимо обнаружить различия последовательностей ДНК между родителями картирующего скрещивания в участке, соответствующем текущей последовательности нуклеиновой кислоты. Это, однако, не является необходимым для способов картирования.
Способы согласно настоящему изобретению приводят в одном варианте осуществления к растениям, имеющим усиленные признаки, связанные с урожайностью семян, как описано выше. Эти признаки могут также комбинироваться с другими экономически выгодными признаками, такими как дополнительные увеличивающие урожайность признаки, устойчивость к другим абиотическим и биотическим стрессам, устойчивость к гербицидам, инсектицидам, признаки, модифицирующие различные структурные признаки, и/или биохимические, и/или физиологические признаки.
Способы согласно настоящему изобретению приводят в одном варианте осуществления к растениям, имеющим улучшенные признаки, связанные с урожайностью, как здесь описано выше. Эти признаки могут также комбинироваться с другими экономически выгодными признаками, такими как дополнительные улучшающие урожайность признаки, устойчивость к другим абиотическим и биотическим стрессам, признаки, модифицирующие различные структурные признаки, и/или биохимические, и/или физиологические признаки.
Способы стэкинга генов в трансгенных растениях хорошо известны в данной области техники (смотри, например, обзор Halpin (2005) Plant Biotech J (3):141-155). Стэкинг генов может выполняться посредством итеративных стадий, где два или более трансгенов могут последовательно вводиться в растение скрещиванием растения, содержащего один трансген, с особями, содержащими другие трансгены, или, альтернативно, повторной трансформацией (или супертрансформацией (super-transforming)) растения, содержащего один трансген, новыми генами. Единственное ограничение этой итеративной процедуры заключается в том, что эти трансгены не связаны и будут располагаться в различных произвольных локусах растительного генома. Следствием является то, что два локуса могут выделяться отдельно в последующих поколениях, что имеет значение для селекционных программ.
Альтернативно, стэкинг генов может происходить путем котрансформации, который является более быстрым и может использоваться во всем диапазоне методик трансформации. Например, при использовании трансформации Agrobacterium трансгены (по меньшей мере два) могут присутствовать в ряде структур:
(i) кодирующие последовательности слиты с образованием единого полипептида при трансляции, и помещены под контроль одного промотора;
(ii) кодирующие последовательности последовательно помещены ниже одного промотора и разделены сигналами нуклеиновых кислот (nucleic acid signals), которые влияют на синтез мРНК (внутренние сайты связывания рибосомы (IRES), 2A прерывающие (stuttering) сигналы и т.д.), или на полипептидный синтез (полипротеины, разделенные сайтами субстрата протеаз и т.д.);
(iii) кодирующие последовательности независимо управляются отдельными промоторами, и комбинации промотор-кодирующая последовательность расположены в одной и той же Т-ДНК;
(iv) кодирующие последовательности независимо управляются отдельными промоторами, и комбинации промотор-кодирующая последовательность расположены в различных Т-ДНК в одной и той же плазмиде;
(v) кодирующие последовательности независимо управляются отдельными промоторами, и комбинации промотор-кодирующая последовательность расположены в различных Т-ДНК в различных плазмидах в одном и том же или отдельных штаммах Agrobacterium.
В другом варианте осуществления настоящее изобретение обеспечивает способ усиления признаков, связанных с урожайностью, по сравнению с контрольными растениями, предусматривающий увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF, и модуляцию экспрессии в том же растении последовательности нуклеиновой кислоты, кодирующей второй полипептид.
Неожиданно было обнаружено, что увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF, дает растения, имеющие усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями. Согласно первому варианту осуществления настоящее изобретение обеспечивает способ усиления признаков, связанных с урожайностью, в растениях по сравнению с контрольными растениями, предусматривающий увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF.
В одном варианте осуществления изобретение относится к объектам, суммированным следующим образом:
1. Способ усиления признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, предусматривающий увеличение экспрессии в растении последовательности нуклеиновой кислоты, кодирующей полипептид фактора, регулирующего рост (GRF), при этом полипептид GRF содержит: (i) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом QLQ, как представлено SEQ ID NO:115; и (ii) домен, имеющий по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с доменом WRC, как представлено SEQ ID NO:116, и при необходимости отбор растений, имеющих усиленные признаки, связанные с урожайностью.
2. Способ по п.1, где указанный полипептид GRF содержит: (i) домен QLQ с регистрационным номером IPR014978 в InterPro (рег. номер PF08880 в PFAM); (ii) домен WRC с рег. номером IPR014977 в InterPro (рег. номер PF08879 в PFAM); и (iii) домен эфсректора транскрипции (ЕТ), содержащий три Cys и один His остатки в консервативном участке (СХ9СХ10СХ2Н).
3. Способ по п.1 или п.2, где указанный полипептид GRF имеет в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более идентичности аминокислотной последовательности с полипептидом GRF, как представлено SEQ ID NO:2, или с любой из полипептидных последовательностей, приведенных здесь в Таблице А.
4. Способ по любому из предшествующих пунктов, где указанная последовательность нуклеиновой кислоты, кодирующая полипептид GRF, представлена любой из последовательностей нуклеиновых кислот SEQ ID NO, приведенных в Таблице А, или ее частью, или последовательностью, способной гибридизоваться с любой из последовательностей нуклеиновых кислот SEQ ID NO, представленных в Таблице А.
5. Способ по любому из предшествующих пунктов, где указанная последовательность нуклеиновой кислоты кодирует ортолог или паралог любой из полипептидных последовательностей SEQ ID NO, приведенных в Таблице А.
6. Способ по любому из предшествующих пунктов, где на указанную увеличенную экспрессию влияют одним или более из: активационное мечение Т-ДНК, TILLING, гомологичная рекомбинация.
7. Способ по любому из предшествующих пунктов, где на указанную увеличенную экспрессию влияют введением и экспрессией в растении последовательности нуклеиновой кислоты, кодирующей полипептид GRF.
8. Способ по любому из предшествующих пунктов, где указанный усиленный признак, связанный с урожайностью, представляет собой один или более из: (i) увеличенная ранняя мощность, (ii) увеличенная надземная биомасса, (iii) увеличенная общая урожайность семян на растение, (iv) увеличенный уровень наполнения семян, (v) увеличенный индекс урожайности или (vi) увеличенная масса тысячи зерен (TKW).
9. Способ по любому из предшествующих пунктов, где указанная последовательность нуклеиновой кислоты функционально связана с конститутивным промотором, предпочтительно с растительным конститутивным промотором, более предпочтительно с промотором GOS2, наиболее предпочтительно с промотором GOS2 из риса, как представлено SEQ ID NO:117.
10. Способ по любому из предшествующих пунктов, где указанная последовательность нуклеиновой кислоты, кодирующая полипептид GRF, имеет растительное происхождение, предпочтительно из двудольного растения, также предпочтительно из семейства Brassicaceae, наиболее предпочтительно из Arabidopsis thaliana.
11. Растения, их части (включая семена) или растительные клетки, получаемые способом по любому предшествующему пункту, где указанное растение, его часть или клетка содержат выделенный трансген нуклеиновой кислоты, кодирующий полипептид GRF, функционально связанный с растительным конститутивным промотором.
12. Конструкция, содержащая:
1. последовательность нуклеиновой кислоты, кодирующую полипептид GRF, как определено в любом из пп.1-5;
2. одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а); при необходимости
3. последовательность терминации транскрипции.
13. Конструкция по п.12, где указанная контрольная последовательность представляет собой растительный конститутивный промотор, предпочтительно промотор GOS2, более предпочтительно промотор GOS2, как представлено SEQ ID NO:117.
14. Применение конструкции по п.12 или п.13 в способе получения растений, имеющих усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями, при этом усиленные признаки, связанные с урожайностью, представляют собой один или более из: (i) увеличенная ранняя мощность, (ii) увеличенная надземная биомасса, (iii) увеличенная общая урожайность семян на растение, (iv) увеличенный уровень наполнения семян, (v) увеличенный индекс урожайности или (vi) увеличенная масса тысячи зерен (TKW).
15. Растение, часть растения или растительная клетка, трансформированные конструкцией по п.12 или п.13.
16. Способ получения трансгенных растений, имеющих усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями, предусматривающий:
(i) введение и экспрессию в растении, части растения или растительной клетке последовательности нуклеиновой кислоты, кодирующей полипептид GRF, как определено в любом из пп.1-5, под контролем растительного конститутивного промотора;
(ii) культивирование растительной клетки, части растения или растения в условиях, способствующих росту и развитию растений.
17. Трансгенное растение, имеющее усиленные признаки, связанные с урожайностью, по сравнению с контрольными растениями, являющиеся результатом увеличенной экспрессии последовательности нуклеиновой кислоты, кодирующей полипептид GRF, как определено в любом из пп.1-5, функционально связанной с растительным конститутивным промотором, или клетка трансгенного растения, или часть трансгенного растения, полученные из указанного трансгенного растения.
18. Трансгенное растение по п.11, 15 или 17, где указанное растение представляет собой сельскохозяйственную культуру, или однодольное растение, или зерновую культуру, такую как рис, кукуруза, пшеница, ячмень, просо, рожь, тритикале, сорго и овес, или клетка трансгенного растения, полученная из указанного трансгенного растения.
19. Заготавливаемые части, содержащие выделенную последовательность нуклеиновой кислоты, кодирующую полипептид GRF, растения по п.18, где указанные заготавливаемые части являются предпочтительно семенами.
20. Продукты, полученные из растения по п.18 и/или из заготавливаемых частей растения по п.19.
21. Применение последовательности нуклеиновой кислоты, кодирующей полипептид GRF, как определено в любом из пп.1-5, в усилении признаков, связанных с урожайностью, содержащих один или более из: (i) увеличенная ранняя мощность, (ii) увеличенная надземная биомасса, (iii) увеличенная общая урожайность семян на растение, (iv) увеличенный уровень наполнения семян, (v) увеличенный индекс урожайности или (vi) увеличенная масса тысячи зерен (TKW).
В одном из вариантов осуществления изобретение относится к объектам, суммированным следующим образом:
22. Способ улучшения признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, где указанный RAA1-подобный полипептид содержит два или более из следующих мотивов:
(i) мотив 1: GVW(V/L)F (SEQ ID NO:162),
(ii) мотив 2: LGW(E/S)RY(Y/F) (SEQ ID NO:163),
(iii) мотив 3: (D/H)L(L/I)S(I/V/L)P(R/K/A)(S/D)F (SEQ ID NO:164),
(iv) мотив 4: (H/Y)(F/M)YD(V/I)WK(N/T)(R/P) (SEQ ID NO:165).
23. Способ по п.22, где указанный RAA1-подобный полипептид также имеет молекулярную массу между 10 и 21 кДа и pl выше 8.5.
24. Способ по п.22 или 23, где на указанную модулированную экспрессию влияют введением и экспрессией в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид.
25. Способ по любому из предшествующих пунктов, где указанная нуклеиновая кислота, кодирующая RAA1-подобный полипептид, кодирует любой из белков, перечисленных в Таблице А, или представляет собой часть такой нуклеиновой кислоты, или нуклеиновую кислоту, способную гибридизоваться с такой нуклеиновой кислотой.
26. Способ по любому из предшествующих пунктов, где указанная последовательность нуклеиновой кислоты кодирует ортолог или паралог любого из белков, представленных в Таблице А.
27. Способ по любому из предшествующих пунктов, где указанные улучшенные признаки, связанные с урожайностью, содержат увеличенную урожайность, предпочтительно увеличенную биомассу и/или увеличенную урожайность семян по сравнению с контрольными растениями.
28. Способ по любому из пп.22-27, где указанные улучшенные признаки, связанные с урожайностью, получают в условиях, не приводящих к стрессу.
29. Способ по любому из пп.22-27, где указанные улучшенные признаки, связанные с урожайностью, получают в условиях недостаточности азота.
30. Способ по любому из пп.24-29, где указанная нуклеиновая кислота функционально связана с конститутивным промотором.
31. Способ по п.30, где указанный конститутивный промотор представляет собой промотор GOS2 или промотор HMGP, предпочтительно промотор GOS2 или промотор HMGP из риса.
32. Способ по любому из предшествующих пунктов, где указанная нуклеиновая кислота, кодирующая полипептид RAA1, имеет растительное происхождение, предпочтительно из двудольного растения, также предпочтительно из семейства Роасеае, боле предпочтительно из рода Oryza, наиболее предпочтительно из Oryza sativa.
33. Растение или его часть, включая семена, получаемые способом по любому из предшествующих пунктов, где указанное растение или его часть содержат рекомбинантную нуклеиновую кислоту, кодирующую RAA1-подобный полипептид.
34. Конструкция, содержащая:
(i) нуклеиновую кислоту, кодирующую RAA1-подобный полипептид;
(ii) одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а); при необходимости
(iii) последовательность терминации транскрипции.
35. Конструкция по п.34, где одна из контрольных последовательностей представляет собой конститутивный промотор.
36. Конструкция по п.35, где указанный конститутивный промотор представляет собой промотор GOS2 или промотор HMGP, предпочтительно промотор GOS2 или промотор HMGP из риса.
37. Применение конструкции по любому из пп.34-36 в способе получения растений, имеющих увеличенную урожайность, в частности увеличенную биомассу и/или увеличенную урожайность семян по сравнению с контрольными растениями.
38. Растение, часть растения или растительная клетка, трансформированные конструкцией по любому из пп.34-36.
39. Способ получения трансгенного растения, имеющего увеличенную урожайность, в частности увеличенную биомассу и/или увеличенную урожайность семян по сравнению с контрольными растениями, предусматривающий:
(i) введение и экспрессию в растении нуклеиновой кислоты, кодирующей RAA1-подобный полипептид;
(ii) культивирование растительной клетки в условиях, способствующих росту и развитию растений.
40. Трансгенное растение, имеющее увеличенную урожайность, в частности увеличенную биомассу и/или увеличенную урожайность семян, по сравнению с контрольными растениями, являющуюся результатом модулированной экспрессии нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, или клетка трансгенного растения, полученная из указанного трансгенного растения.
41. Трансгенное растение по п.33, 38 или 39, или клетка трансгенного растения, полученная из него, где указанное растение представляет собой сельскохозяйственную культуру, или однодольное растение, или зерновую культуру, такую как рис, кукуруза, пшеница, ячмень, просо, рожь, тритикале, сорго и овес.
42. Заготавливаемые части растения по п.41, где указанные заготавливаемые части представляют собой предпочтительно биомассу корней и/или семена.
43. Продукты, полученные из растения по п.41 и/или из заготавливаемых частей растения по п.42.
44. Применение нуклеиновой кислоты, кодирующей RAA1-подобный полипептид, для увеличения урожайности, в частности для увеличения урожайности семян и/или биомассы корней в растениях, по сравнению с контрольными растениями.
В одном из вариантов осуществления изобретение относится к объектам, суммированным следующим образом:
45. Способ увеличения устойчивости к абиотическому стрессу у растений по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYR, при этом полипептид SYR содержит обогащенный лейцином домен, перед которым находится консервативный трипептидный мотив 5 (один из SEQ ID NO:173, 174, 175 или 176), и за которым следует консервативный мотив 6 (SEQ ID NO:177), где указанная увеличенная устойчивость к абиотическому стрессу представляет собой увеличенную эффективность поглощения питательных веществ и/или увеличенную толерантность к стрессу, вызванному засухой, по сравнению с контрольными растениями.
46. Способ по п.45, где указанный полипептид SYR имеет в увеличивающемся порядке предпочтения по меньшей мере 27%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более идентичности последовательности с полипептидом SYR, представленным SEQ ID NO:169.
47. Способ по п.45 или п.46, где указанная нуклеиновая кислота, кодирующая полипептид SYR, представлена любой из нуклеиновых кислот SEQ ID NO, приведенных в Таблице А, или ее частью, или последовательностью, способной гибридизоваться с любой из нуклеиновых кислот SEQ ID NO, приведенных в Таблице А.
48. Способ по любому из пп.45-47, где указанная последовательность нуклеиновой кислоты кодирует ортолог или паралог любой из SEQ ID NO, приведенных в Таблице А.
49. Способ по любому из предшествующих пунктов, где указанный белок SYR содержит, кроме того, консервативный мотив 7 (SEQ ID NO:178).
50. Способ по любому из предшествующих пунктов, где указанная эффективность поглощения питательных веществ приводит к увеличенной урожайности семян и/или увеличенной биомассе.
51. Способ по п.50, где указанная увеличенная урожайность семян содержит по меньшей мере увеличенные общую массу семян, массу тысячи зерен и/или увеличенное число наполненных семян.
52. Способ по п.50, где указанная увеличенная биомасса представляет собой увеличенную биомассу побегов и/или увеличенную биомассу корней.
53. Способ по любому из предшествующих пунктов, где указанная увеличенная эффективность поглощения питательных веществ имеет место в условиях умеренной засухи.
54. Способ по любому из предшествующих пунктов, где указанная увеличенная толерантность к стрессу, вызванному засухой, приводит к увеличенной урожайности семян.
55. Способ по п.54, где указанная увеличенная урожайность семян содержит по меньшей мере увеличенные общую массу семян, фильтрат и/или индекс урожайности.
56. Способ по любому из предшествующих пунктов, где на указанную модулированную экспрессию влияют введением и экспрессией в растении нуклеиновой кислоты, кодирующей полипептид SYR.
57. Способ по п.56, где указанная нуклеиновая кислота функционально связана с конститутивным промотором, предпочтительно с промотором GOS2.
58. Способ по любому предшествующему пункту, где указанная нуклеиновая кислота, кодирующая полипептид SYR, имеет растительное происхождение, предпочтительно из однодольного растения, также предпочтительно из семейства Роасеае, более предпочтительно из рода Oryza, наиболее предпочтительно из Oryza sativa.
59. Применение конструкции в способе получения растений, имеющих увеличенную устойчивость к абиотическому стрессу, при этом указанная конструкция содержит
(a) нуклеиновую кислоту, кодирующую полипептид SYR, как определено в любом из пп.45-49;
(b) одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а); при необходимости
(c) последовательность терминации транскрипции,
и где одна из контрольных последовательностей представляет собой конститутивный промотор, предпочтительно промотор GOS2, и где указанная увеличенная устойчивость к абиотическому стрессу представляет собой увеличенную эффективность поглощения питательных веществ и/или увеличенную толерантность к стрессу, вызванному засухой, по сравнению с контрольными растениями.
60. Применение нуклеиновой кислоты, кодирующей полипептид SYR, в способе увеличения устойчивости к абиотическому стрессу у растений по сравнению с контрольными растениями, где указанная увеличенная устойчивость к абиотическому стрессу представляет собой увеличенную эффективность поглощения питательных веществ и/или увеличенную толерантность к стрессу, вызванному засухой, по сравнению с контрольными растениями.
61. Применение по п.60, где указанная эффективность поглощения питательных веществ приводит к увеличенной урожайности семян и/или увеличенной биомассе.
62. Применение по п.60, где указанная увеличенная толерантность к стрессу, вызванному засухой, приводит к увеличенной урожайности семян.
В одном из вариантов осуществления изобретение относится к объектам, суммированным следующим образом:
63. Способ улучшения признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид ARKL.
64. Способ по п.63, где указанный полипептид ARKL содержит один или более из следующих доменов:
(i) домен ZfC3H2C3, как представлено SEQ ID NO:400, или домен, имеющий в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более идентичности последовательности с одним или более из доменов ZfC3H2C3, как представлено SEQ ID NO:95 - SEQ ID NO. 351;
(II) домен DAR1, имеющий в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более идентичности последовательности с одним или более из доменов PfamB2828, как представлено SEQ ID NO:352 - SEQ ID NO:398.
65. Способ по п.63 и п.64, где указанный полипептид ARKL содержит один или более из следующих:
(i) домен ZfC3H2C3, как представлено SEQ ID NO:401;
(ii) мотив 8, как представлено SEQ ID NO:399.
66. Способ по пп.63-65, где на указанную модулированную экспрессию влияют введением и экспрессией в растении нуклеиновой кислоты, кодирующей полипептид ARKL.
67. Способ по любому из предшествующих пунктов, где указанная нуклеиновая кислота, кодирующая полипептид ARKL, кодирует любой из белков, перечисленных в Таблице А, или представляет собой часть такой нуклеиновой кислоты, или нуклеиновую кислоту, способную гибридизоваться с такой нуклеиновой кислотой.
68. Способ по любому из предшествующих пунктов, где указанная последовательность нуклеиновой кислоты кодирует ортолог или паралог любого из белков, приведенных в Таблице А.
69. Способ по любому из предшествующих пунктов, где указанные улучшенные признаки, связанные с урожайностью, содержат увеличенную урожайность, предпочтительно увеличенную урожайность семян по сравнению с контрольными растениями.
70. Способ по любому из предшествующих пунктов, где указанные улучшенные признаки, связанные с урожайностью, получают в условиях, не приводящих к стрессу.
71. Способ по любому из предшествующих пунктов, где указанные улучшенные признаки, связанные с урожайностью, получают в условиях стресса, вызванного засухой.
72. Способ по любому из предшествующих пунктов, где указанная нуклеиновая кислота функционально связана с конститутивным промотором, предпочтительно с промотором GOS2, наиболее предпочтительно с промотором GOS2 из риса.
73. Способ по любому из предшествующих пунктов, где указанная нуклеиновая кислота, кодирующая полипептид ARKL, имеет растительное происхождение, предпочтительно из однодольного растения, также предпочтительно из семейства Роасеае, более предпочтительно из рода Oryza, наиболее предпочтительно из Oryza sativa.
74. Растение или его часть, включая семена, получаемые способом по любому из предшествующих пунктов, где указанное растение или его часть содержит рекомбинантную нуклеиновую кислоту, кодирующую полипептид ARKL.
75. Конструкция, содержащая:
(i) нуклеиновую кислоту, кодирующую полипептид ARKL, как определено в пп.63-65;
(ii) одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а); и при необходимости
(iii) последовательность терминации транскрипции.
76. Конструкция по п.75, где одна из контрольных последовательностей представляет собой конститутивный промотор, предпочтительно промотор GOS2, наиболее предпочтительно промотор GOS2 из риса.
77. Применение конструкции по п.75 или п.76 в способе получения растений, имеющих улучшенные признаки, связанные с урожайностью, предпочтительно увеличенную урожайность, более предпочтительно увеличенную урожайность семян по сравнению с контрольными растениями.
78. Растение, часть растения или растительная клетка, трансформированные конструкцией по п.75 или п.76.
79. Способ получения трансгенного растения, имеющего увеличенную урожайность, в частности увеличенную урожайность семян по сравнению с контрольными растениями, предусматривающий:
(i) введение и экспрессию в растении нуклеиновой кислоты, кодирующей полипептид ARKL, как определено в пп.63-65; и
(ii) культивирование растительной клетки в условиях, способствующих росту и развитию растений.
80. Трансгенное растение, имеющее увеличенную урожайность, в частности увеличенную урожайность семян, по сравнению с контрольными растениями, являющуюся результатом увеличенной экспрессии нуклеиновой кислоты, кодирующей полипептид ARKL, как определено в пп.63-65, или клетка трансгенного растения, полученная из указанного трансгенного растения.
81. Трансгенное растение по пп.74, 78 или 80 или клетка трансгенного растения, полученная из него, где указанное растение представляет собой сельскохозяйственную культуру, или однодольное растение, или зерновую культуру, такую как рис, кукуруза, пшеница, ячмень, просо, рожь, тритикале, сорго и овес.
82. Заготавливаемые части растения по п.81, где указанные заготавливаемые части представляют собой предпочтительно биомассу побегов и/или семена.
83. Продукты, полученные из растения по п.81 и/или из заготавливаемых частей растения по п.82.
84. Применение нуклеиновой кислоты, кодирующей полипептид ARKL, для улучшения признаков, связанных с урожайностью, предпочтительно для увеличенной урожайности, более предпочтительно для увеличенной урожайности семян по сравнению с контрольными растениями.
В одном из вариантов осуществления изобретение относится к объектам, суммированным следующим образом:
85. Способ улучшения признаков, связанных с урожайностью, у растений по сравнению с контрольными растениями, предусматривающий модуляцию экспрессии в растении нуклеиновой кислоты, кодирующей полипептид YTP, содержащий
(i) по меньшей мере один трансмембранный домен и
(ii) по меньшей мере часть домена DUF221.
86. Способ по п.85, где указанная часть имеет в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности с любым из доменов, представленным SEQ ID NO:518 - SEQ ID NO:544.
87. Способ по п.85 или п.86, где указанная нуклеиновая кислота кодирует полипептид, имеющий в возрастающем порядке предпочтения по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с любым из полипептидов Таблицы А.
88. Способ по любому из предшествующих пунктов, где указанный полипептид YTP также содержит мотив 9 (SEQ ID NO:545).
89. Способ по любому из предшествующих пунктов, где на указанную модулированную экспрессию влияют введением и экспрессией в растении нуклеиновой кислоты, кодирующей полипептид YTP.
90. Способ по любому из предшествующих пунктов, где указанная нуклеиновая кислота, кодирующая полипептид YTP, кодирует любой из белков перечисленных в Таблице А или представляет собой часть такой нуклеиновой кислоты или нуклеиновую кислоту, способную гибридизоваться с такой нуклеиновой кислотой.
91. Способ по любому из предшествующих пунктов, где указанная последовательность нуклеиновой кислоты кодирует ортолог или паралог любого из белков, приведенных в Таблице А, или представляет собой часть такой нуклеиновой кислоты или нуклеиновую кислоту, способную гибридизоваться с такой нуклеиновой кислотой.
92. Способ по любому из предшествующих пунктов, где указанные улучшенные признаки, связанные с урожайностью, содержат увеличенную урожайность, предпочтительно увеличенную урожайность семян по сравнению с контрольными растениями.
93. Способ по любому из пп.85-92, где указанные улучшенные признаки, связанные с урожайностью, получают в условиях, не приводящих к стрессу.
94. Способ по любому из пп.89-93, где указанная нуклеиновая кислота функционально связана с конститутивным промотором, предпочтительно с промотором GOS2, наиболее предпочтительно с промотором GOS2 из риса.
95. Способ по любому из предшествующих пунктов, где указанная нуклеиновая кислота, кодирующая полипептид YTP, имеет растительное происхождение, предпочтительно из двудольного растения, также предпочтительно из семейства Роасеае, более предпочтительно из рода Oryza, наиболее предпочтительно из Oryza sativa.
96. Растение или его часть, включая семена, получаемое способом по любому предшествующему пункту, где указанное растение или его часть содержат рекомбинантную нуклеиновую кислоту, кодирующую YTP полипептид.
97. Конструкция, содержащая:
(i) нуклеиновую кислоту, кодирующую полипептид YTP, как определено в пп.85-88,
(ii) одну или более контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а); и при необходимости
(iii) последовательность терминации транскрипции.
98. Конструкция по п.97, где одна из контрольных последовательностей представляет собой конститутивный промотор, предпочтительно промотор GOS2, более предпочтительно промотор GOS2 из риса.
99. Применение конструкции по п.97 или п.98 в способе получения растений, имеющих увеличенную урожайность, в частности увеличенную урожайность семян, по сравнению с контрольными растениями.
100. Растение, часть растения или растительная клетка, трансформированные конструкцией по п.97 или п.98.
101. Способ получения трансгенного растения, имеющего увеличенную урожайность, в частности увеличенную урожайность семян, по сравнению с контрольными растениями, предусматривающий:
(i) введение и экспрессию в растении нуклеиновой кислоты, кодирующей полипептид YTP, как определено в пп.85-88; и
(ii) культивирование растительной клетки в условиях, способствующих росту и развитию растений.
102. Трансгенное растение, имеющее увеличенную урожайность, в частности увеличенную урожайность семян, по сравнению с контрольными растениями, являющуюся результатом модулированной экспрессии нуклеиновой кислоты, кодирующей полипептид YTP, как определено в пп.85-88, или клетка трансгенного растения, полученная из указанного трансгенного растения.
103. Трансгенное растение по п.96, 100 или 102 или клетка трансгенного растения, полученная из него, где указанное растение представляет собой сельскохозяйственную культуру, или однодольное растение, или зерновую культуру, такую как рис, кукуруза, пшеница, ячмень, просо, рожь, тритикале, сорго и овес.
104. Заготавливаемые части растения по п.103, где указанные заготавливаемые части представляют собой предпочтительно биомассу побегов и/или семена.
105. Продукты, полученные из растения по п.103 и/или из заготавливаемых частей растения по п.104.
106. Применение нуклеиновой кислоты, кодирующей полипептид YTP, для увеличения урожайности, в частности для увеличения урожайности семян и/или биомассы побегов у растений по сравнению с контрольными растениями.
Описание чертежей
Настоящее изобретение будет сейчас описано со ссылкой на следующие чертежи, на которых:
Фиг.1 представляет собой рисунок полипептида GRF, как представлено SEQ ID NO:2, который содержит следующие признаки: (i) домен QLQ с рег. номером IPR014978 в InterPro (рег. номер PF08880 в PFAM); (ii) домен WRC с рег. номером IPR014977 в InterPro (рег. номер PF08879 в PFAM); и (iii) домен эффектора транскрипции (ЕТ), содержащий три Cys и один His остатки в консервативном участке (СХ9СХ10СХ2Н), расположенный в домене WRC.
Фиг.2 демонстрирует множественное выравнивание последовательностей AlignX (из Vector NTI 10.3, Invitrogen Corporation) домена QLQ полипептидов GRF из Таблицы А (как представлено SEQ ID NO:115 для SEQ ID NO:2). Консервативные аминокислотные остатки QLQ расположены в верхней части множественного выравнивания. Два других высококонсервативных остатка (обозначены черным) представляют собой Е (Glu) и Р (Pro).
Фиг.3 демонстрирует множественное выравнивание последовательностей AlignX (из Vector NTI 10.3, Invitrogen Corporation) домена WRC полипептидов GRF из Таблицы А (как представлено SEQ ID NO:116 для SEQ ID NO:2). Консервативные аминокислотные остатки выделены жирным шрифтом в консенсусной последовательности. Три Cys и один His остатки в консервативном участке (СХ9СХ10СХ2Н), обозначенном как домен эффектора транскрипции (ЕТ), заключены в рамку вертикально поперек выравнивания и также обозначены в нижней части выравнивания. Предполагаемый сигнал ядерной локализации (NLS), заключенный в домен WRC, обозначен двойным подчеркиванием.
Фиг.4 демонстрирует бинарный вектор для увеличенной экспрессии в Oryza sativa последовательности нуклеиновой кислоты, кодирующей полипептид GRF, под контролем промотора GOS2 (pGOS2) из риса.
Фиг.5 подробно описывает примеры последовательностей, используемых при выполнении способов согласно настоящему изобретению.
Фиг.6 представляет последовательность рисового RAA1-подобного белка (SEQ ID NO:121) с консервативными последовательностями сигнатур, обозначенными жирным шрифтом с подчеркиванием.
Фиг.7 демонстрирует множественное выравнивание различных RAA1-подобных белков. NP_001046787 соответствует QOE1D7, NP_001052368 соответствует QOJEF5, AAR97604 соответствует Q6RIBO, NP_001042631 соответствует Q9LGE3, NP_001045304 соответствует Q8LR63, NP_974763 соответствует Q9LXB5, NP_197868 соответствует O23624, NP_194866 соответствует Q5QOB3, NP_001060595 соответствует Q8H475.
Фиг.8 демонстрирует филогенетическое дерево RAA1-подобных полипептидов (Ge et al., 2004). OsRAA1 соответствует SEQ ID NO:121.
Фиг.9 представляет бинарный вектор для увеличенной экспрессии в Oryza sativa нуклеиновой кислоты, кодирующей RAA1-подобный пептид, под контролем рисового промотора GOS2 (pGOS2).
Фиг.10 подробно описывает примеры последовательностей, используемых при выполнении способов согласно настоящему изобретению.
Фиг.11 дает обзор консервативных мотивов, присутствующих в SEQ ID NO:169. Обогащенный лейцином домен подчеркнут, консервативные мотивы 5, 6 и 7 указаны жирным шрифтом, а последовательность курсивом представляет предполагаемый сайт N-гликозилирования с предполагаемым сайтом фосфорилирования протеинкиназы С.
Фиг.12 демонстрирует множественное выравнивание различных белков SYR. Звездочки показывают одинаковые аминокислотные остатки, двоеточия представляют высококонсервативные замены, и точки представляют менее консервативные замены. Со сведениями из Фиг.11 различные домены и консервативные мотивы в SEQ ID NO:171 могут быть легко идентифицированы в других белках SYR.
Фиг.13 демонстрирует бинарный вектор pGOS2::SYR для трансформации и экспрессии в Oryza sativa нуклеиновой кислоты Oryza sativa SYR под контролем рисового промотора GOS2.
Фиг.14 подробно описывает примеры последовательностей, используемых при выполнении способов согласно настоящему изобретению, или используемых для выделения таких последовательностей. Последовательности могут получаться из общедоступных наборов EST с менее качественным секвенированием. Как следствие, может ожидаться небольшое количество нуклеотидных замен. И 5', и 3' UTR могут также использоваться для выполнения способов по изобретению. SEQ ID NO:193 представляет белковую последовательность ARGOS (рег. номер AY305869 в GenBank).
Фиг.15 представляет аминокислотную последовательность SEQ ID NO:213. Консервативные домены pfamB2828 и ZfC3H2C3 (pfam00097) выделены жирным шрифтом и подчеркнутыми буквами соответственно. Высококонсервативный мотив 8 обозначен подчеркиванием пунктирной линией. Наиболее высококонсервативные остатки в полипептидах ARKL заключены в рамку. Продемонстрированы консервативные положения аминокислотных остатков - лигандов металлов (цифры), и цинк (Zn2+)-координирующие аминокислотные пары.
Фиг.16 представляет множественное выравнивание выбранных полипептидов ARKL. Высококонсервативные аминокислотные остатки указаны в консенсусной последовательности. Как показано на Фиг.16, последовательность С-конца полипептидов ARKL более сильно консервативна, чем N-конец.
Фиг.17 демонстрирует филогенетическое дерево полипептидов ARKL, основанное на выравнивании домена RING-пальца (pfam00097), включенного в полипептиды ARKL, как представлено SEQ ID NO:306-314, 316-318, 322, 323, 402 (SEQ ID NO:402 содержит домен pfam00097 (цинковый RING-палец), присутствующий в полипептиде Akadia у Mus musculus, и SEQ ID NO:403 представляет домен pfam00097 как присутствующий в полипептиде Goliath у Mus musculus). Использованные аббревиатуры: Os: Oryza sativa (Orysa); Hv: Hordeum vulgare (Horvu); Gm: Glycine max (Glyma); Zm: Zea mays (Zeama); Musmu: Mus musculus.
Фиг.18 представляет бинарный вектор для увеличенной экспрессии нуклеиновой кислоты OS_ARKL1, как представлено SEQ ID NO:212 под контролем рисового промотора GOS2 (pGOS2).
Фиг.19 подробно описывает примеры последовательностей, используемых при выполнении способов согласно настоящему изобретению.
Фиг.20 представляет последовательность YTP1 (SEQ ID NO:409). Трансмембранные домены обведены в рамки. Часть домена DUF221 из 124 аминокислотных остатков выделена жирным шрифтом. Мотив 8 подчеркнут.
Инвариантные остатки мотива 8 обозначены буквами большего размера. Первая и третья петли, как предполагается, располагаются на внешней стороне мембраны; вторая петля - на внутренней.
Фиг.21 демонстрирует филогенетическое дерево отбора полипептидов YTP.
Фиг.22 представляет множественное выравнивание отобранных полипептидов YTP. Приведена консенсусная последовательность, как представлено SEQ ID NO:544. Консервативные аминокислотные остатки указаны в консенсусной последовательности; пропуски представляют собой места низкой консервативности.
Фиг.23 представляет бинарный вектор увеличенной экспрессии в Oryza sativa нуклеиновой кислоты, кодирующей YTP1, под контролем рисового промотора GOS2 (PGOS2).
Фиг.24 подробно описывает примеры последовательностей, используемых при выполнении способов согласно настоящему изобретению.
Примеры
Настоящее изобретение будет сейчас описано со ссылкой на следующие примеры, которые представлены исключительно с целью иллюстрации. Не подразумевается, что следующие примеры полностью определяют или ограничивают иным способом объем изобретения.
Манипуляции с ДНК: если не указано иное, методы рекомбинантных ДНК осуществляются согласно стандартным протоколам, описанным в (Sambrook (2001) Molecular Cloning: a laboratory manual, 3rd Edition Cold Spring Harbor Laboratory Press, CSH, New York) или в томах 1 и 2 Ausubel et al. (1994), Current Protocols in Molecular Biology, Current Protocols. Стандартные материалы и методы для молекулярных исследований на растениях описаны в Plant Molecular Biology Labfax (1993) by R.D.D. Croy, опубликованной BIOS Scientific Publications Ltd (UK) и Blackwell Scientific Publications (UK).
Пример 1: Идентификация последовательностей, родственных последовательности нуклеиновой кислоты, используемой в способах по изобретению
Последовательности (полноразмерная кДНК, последовательности EST или геномные), родственные последовательности нуклеиновой кислоты, использованной в способах по настоящему изобретению, идентифицировали среди последовательностей, содержащихся в базе данных нуклеотидных последовательностей Entrez в National Center for Biotechnology Information (NCBI), с использованием инструментов для поиска последовательностей в базе данных, таких как Basic Local Alignment Tool (BLAST) (Altschul et al. (1990) J. Mol. Biol. 215:403-410; и Altschul et al. (1997) Nucleic Acids Res. 25:3389-3402). Эта программа используется для обнаружения участков локального сходства между последовательностями сравнением последовательностей нуклеиновых кислот или полипептидных последовательностей с базами данных последовательностей и расчетом статистической значимости совпадений. Например, полипептид, кодируемый последовательностью нуклеиновой кислоты по настоящему изобретению, использовали для алгоритма TBLASTN с настройками по умолчанию и отключенным фильтром для игнорирования последовательностей с малой сложностью. Результат анализа оценивали попарным сравнением и классифицировали согласно оценке вероятности (значение Е), где оценка отражает вероятность, что определенное выравнивание встречается случайно (чем ниже значение Е, тем больше значимость хита). В дополнение к значениям Е сравнения также оценивали на идентичность в процентах. Идентичность в процентах относится к числу идентичных нуклеотидов (или аминокислот) между двумя сравниваемыми последовательностями нуклеиновых кислот (или полипептидными последовательностями) на определенной длине. В некоторых случаях параметры по умолчанию могут регулироваться, чтобы модифицировать строгость поиска. Например, значение Е может быть увеличено, чтобы показать менее строгие совпадения. Таким образом, могут быть идентифицированы короткие, почти точные совпадения.
Таблица А предоставляет перечень последовательностей нуклеиновых кислот, родственных с последовательностью нуклеиновой кислоты, используемой в способах по настоящему изобретению.
|
В дополнение к общедоступным последовательностям нуклеиновых кислот, имеющимся в NCBI, поиск проводится также по частным базам данных последовательностей, следуя той же процедуре, как описанная здесь выше.
Таблица A3 обеспечивает перечень последовательностей нуклеиновых кислот и белков, родственных последовательности нуклеиновой кислоты, как представлено SEQ ID NO:168, и белковой последовательности, представленной SEQ ID NO:169.
|
Таблица А5 предоставляет перечень последовательностей нуклеиновых кислот, родственных с последовательностью нуклеиновых кислот, используемой в способах настоящего изобретения.
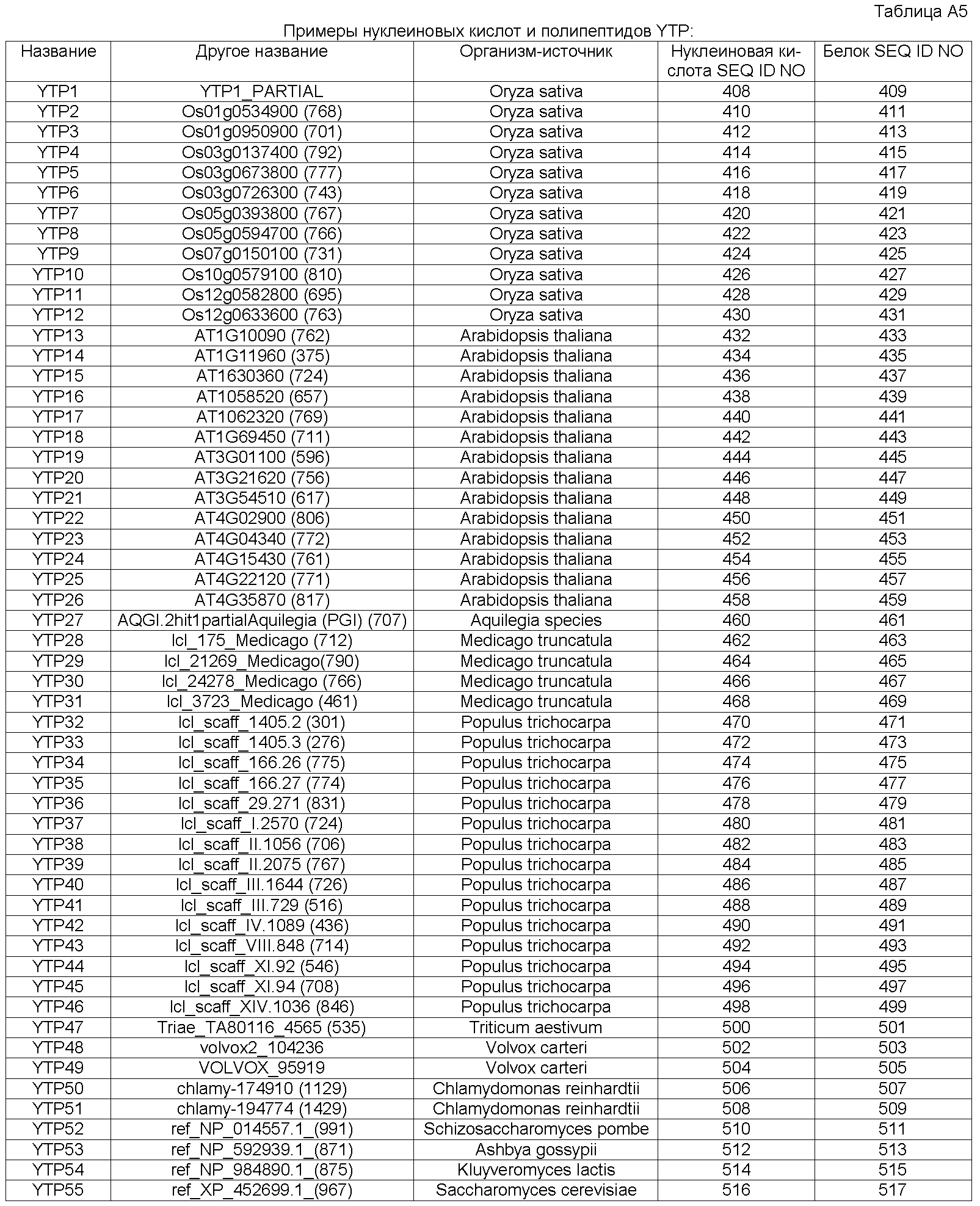
В некоторых случаях родственные последовательности предварительно собраны и открыто предоставлены исследовательскими институтами, такими как Институт геномных исследований (The Institute for Genomic Research (TIGR)). База данных ортологов эукариотических генов (EGO) может использоваться для обнаружения таких родственных последовательностей или поиском по ключевому слову, или путем использования алгоритма BLAST для рассматриваемых последовательности нуклеиновой кислоты или полипептидной последовательности. В других случаях специальные базы данных последовательностей нуклеиновых кислот создаются для определенных организмов, например, у Joint Genome Institute, например, для тополя и Ostreococcus tauri.
Пример 2:
a) Выравнивание полипептидных последовательностей GRF
Множественное выравнивание последовательностей всех полипептидных последовательностей GRF в Таблице А выполняли, используя алгоритм AlignX (из Vector NTI 10.3, Invitrogen Corporation). Результаты выравнивания для домена QLQ полипептидов GRF из Таблицы А (как представлено SEQ ID NO:115 для SEQ ID NO:2) показаны на Фиг.2 настоящей заявки. Консервативные аминокислотные остатки QLQ расположены на верху множественного выравнивания. Два других высококонсервативных остатка (обозначены черным) представляют собой Е (Glu) и Р (Pro). Результаты выравнивания для домена WRC полипептидов GRF из Таблицы А (как представлено SEQ ID NO:116 для SEQ ID NO:2) показаны на Фиг.3 настоящей заявки. Консервативные аминокислотные остатки WRC отмечены жирным шрифтом в консенсусной последовательности. Три остатка Cys и один His в консервативном участке (СХ9СХ10СХ2Н), обозначенном как домен эффектора транскрипции (ЕТ), заключены в рамку вертикально поперек выравнивания и также обозначены в нижней части выравнивания.
b) Выравнивание RAA 1-подобных полипептидных последовательностей
Выравнивание полипептидных последовательностей проводили, используя программу AlignX из Vector NTI (Invitrogen), которая основана на популярном алгоритме Clustal W прогрессивного выравнивания (Thompson et al. (1997) Nucleic Acids Res 25:4876-4882; Chenna et al. (2003). Nucleic Acids Res 31:3497-3500). Значения по умолчанию представляют собой штраф за открытие гэпа 10, штраф за удлинение гэпа 0.1, и выбранной матрицей весов является Blosum 62 (если полипептиды выравниваются). Небольшое ручное редактирование может быть сделано для дополнительной оптимизации выравнивания. Консервативность последовательностей между RAA1-подобными полипептидами существует главным образом на всем протяжении последовательности за исключением обогащенного Gly и/или Ser участка на N-концевой части белка. RAA1-подобные полипептиды выровнены на Фиг.7.
c) Выравнивание полипептидных последовательностей SYR
AlignX (Vector NTI, Invitrogen) основан на популярном алгоритме Clustal прогрессивного выравнивания (Thompson et al. (1997) Nucleic Acids Res 25:4876-4882; Chenna et al. (2003). Nucleic Acids Res 31:3497-3500). Филогенетическое дерево может быть построено при использовании алгоритма кластеризации присоединением соседа. Значения по умолчанию представляют собой штраф за открытие гэпа 10, штраф за удлинение гэпа 0.1, и выбранной матрицей весов является Blosum 62 (если полипептиды выравниваются).
Результат множественного выравнивания последовательностей с использованием полипептидов, релевантных в идентифицировании полипептидов, применимых при выполнении способов по изобретению, продемонстрирован на Фиг.12. Повтор, обогащенный лейцином, и консервативные мотивы могут легко распознаваться в различных последовательностях.
d) Выравнивание полипептидных последовательностей ARKL
Выравнивание полипептидных последовательностей проводили, используя программу AlignX из Vector NTI (Invitrogen), которая основана на популярном алгоритме Clustal W прогрессивного выравнивания (Thompson et al. (1997) Nucleic Acids Res 25:4876-4882; Chenna et al. (2003). Nucleic Acids Res 31:3497-3500). Значения по умолчанию представляют собой штраф за открытие гэпа 10, штраф за удлинение гэпа 0.1, и выбранной матрицей весов является Blosum 62 (если полипептиды выравниваются). Небольшое ручное редактирование может быть сделано для дополнительной оптимизации выравнивания. Консервативность последовательностей между полипептидами ARKL имеется по существу на С-конце вдоль DAR1 и RING-H2 домена полипептидов, при этом N-концевой домен обычно является более вариабельным по длине последовательности и составу. Полипептиды ARKL выровнены на Фиг.16.
Филогенетическое дерево полипептидов ARKL (Фиг.17) строили, используя алгоритм кластеризации присоединением соседа, как предусмотрено в программе AlignX из Vector NTI (Invitrogen).
е) Выравнивание полипептидных последовательностей УТР
Выравнивание полипептидных последовательностей проводили, используя программу AlignX из Vector NTI (Invitrogen), которая основана на популярном алгоритме Clustal W прогрессивного выравнивания (Thompson et al. (1997) Nucleic Acids Res 25:4876-4882; Chenna et al. (2003). Nucleic Acids Res 31:3497-3500). Значения по умолчанию представляют собой штраф за открытие гэпа 10, штраф за удлинение гэпа 0.1, и выбранной матрицей весов является Blosum 62 (если полипептиды выравниваются). Небольшое ручное редактирование может быть сделано для дополнительной оптимизации выравнивания. Консервативность последовательностей среди полипептидов YTP выше на С-конце белка полипептидов вдоль домена DUF221. N-концевой домен обычно является более вариабельным по длине последовательности и составу. Полипептиды YTP выровнены на Фиг.22. Высококонсервативные аминокислотные остатки указаны в консенсусной последовательности (см. Фиг.22).
Филогенетическое дерево полипептидов YTP (Фиг.21) строили, используя алгоритм кластеризации присоединением соседа, как предусмотрено в программе AlignX из Vector NTI (Invitrogen). Как показано на Фиг.21, Группа 1 содержит полипептиды YTP, кластеризующиеся с SEQ ID NO:409 (YTP1 на Фиг.21).
Пример 3: Вычисление общей идентичности в процентах между полипептидными последовательностями, используемыми в выполнении способов по изобретению
Общие сходство и идентичность в процентах между полноразмерными полипептидными последовательностями, используемыми в выполнении способов по изобретению, определяли, используя один из способов, доступных в данной области техники, программу MatGAT (Matrix Global Alignment Tool) (BMC Bioinformatics. 2003 4:29. MatGAT: an application that generates similarity/identity matrices using protein or DNA sequences. Campanella JJ, Bitincka L, Smalley J; software hosted by Ledion Bitincka). Программа MatGAT производит матрицы сходства/идентичности для последовательностей ДНК или белков без необходимости в предварительном выравнивании данных. Программа выполняет серии попарных выравниваний, используя алгоритм глобального выравнивания Майерса и Миллера (штраф за открытие гэпа 12 и штраф за удлинение гэпа 2), рассчитывает сходство и идентичность, используя, например, Blosum 62 (для полипептидов) и затем помещает результаты в матрицу расстояний. Сходство последовательностей демонстрируется в нижней половине от разделительной линии, а идентичность последовательностей демонстрируется в верхней половине от диагональной разделительной линии.
Параметры, использованные в сравнении, представляли собой:
|
Результаты анализа с помощью программного обеспечения представлены в Таблице В для общих сходства и идентичности по всей длине полипептидных последовательностей (исключая частичные полипептидные последовательности).



Идентичность в процентах между полноразмерными полипептидными последовательностями, используемыми при проведении способов по изобретению, может составлять всего 15% аминокислотной идентичности по сравнению с SEQ ID NO:2.
Идентичность в процентах может быть существенно увеличена, если вычисление идентичности проводить между доменом QLQ SEQ из ID NO:2 (как представлено SEQ ID NO:115, включенной в SEQ ID NO:2; домен QLQ полипептидов GRF из Таблицы А, представленный на Фиг.2) и доменами QLQ полипептидов, используемых при выполнении изобретения. Аналогично, идентичность в процентах может быть существенно увеличена, если вычисление идентичности проводить между доменом WRC SEQ из ID NO:2 (как представлено SEQ ID NO:116, включенной в SEQ ID NO:2; домен WRC полипептидов GRF из Таблицы А, представленный на Фиг.3) и доменами WRC полипептидов, используемых при выполнении изобретения. Идентичность в процентах на протяжении домена QLQ между полипептидными последовательностями, используемыми при выполнении способов по изобретению, находится в диапазоне между 25% и 99% аминокислотной идентичности, и идентичность в процентах на протяжении домена WRC между полипептидными последовательностями, используемыми при выполнении способов по изобретению, находится в диапазоне между 60% и 99% аминокислотной идентичности. Как можно увидеть на Фиг.3, домен WRC более консервативен, чем домен QLQ между различными полипептидами GRF, как показано на Фиг.2.
Проценты аминокислотной идентичности между доменами QLQ и идентичность в процентах между доменами WRC значительно выше, чем аминокислотная идентичность в процентах, вычисленная между полноразмерными полипептидными последовательностями GRF.
Результаты анализа с помощью программного обеспечения, касающегося RAA1-подобного полипептида, представлены в Таблице В2 для общих сходства и идентичности по всей длине полипептидных последовательностей. Сходство в процентах приведено над диагональю, а идентичность в процентах дана ниже диагонали (жирный шрифт).
Идентичность в процентах между RAA1-подобными полипептидными последовательностями, используемыми при выполнении способов по изобретению, может составлять всего 31% аминокислотной идентичности по сравнению с SEQ ID NO:121, не учитывая Q9LXB6 (SEQ ID NO:155).
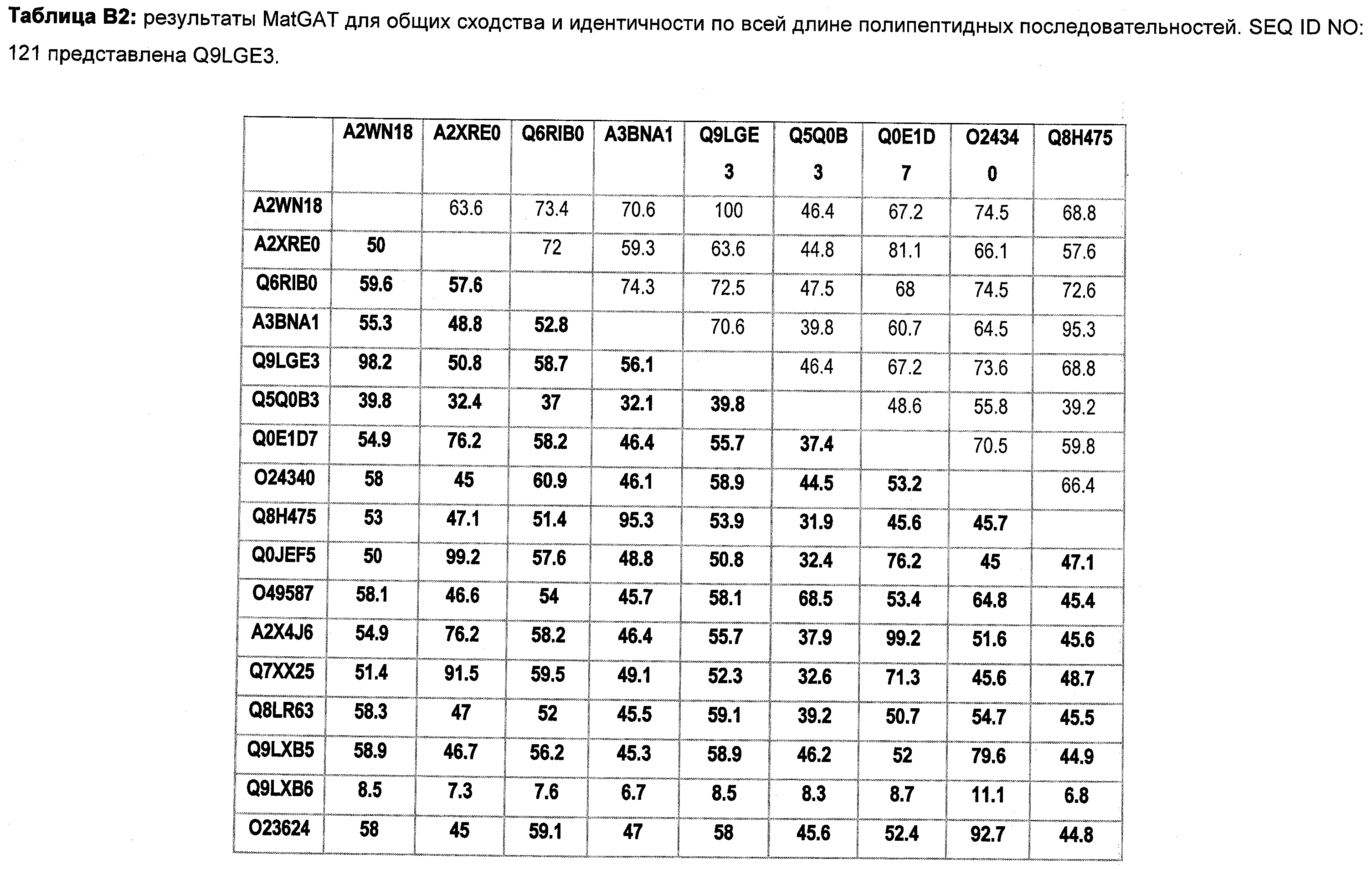
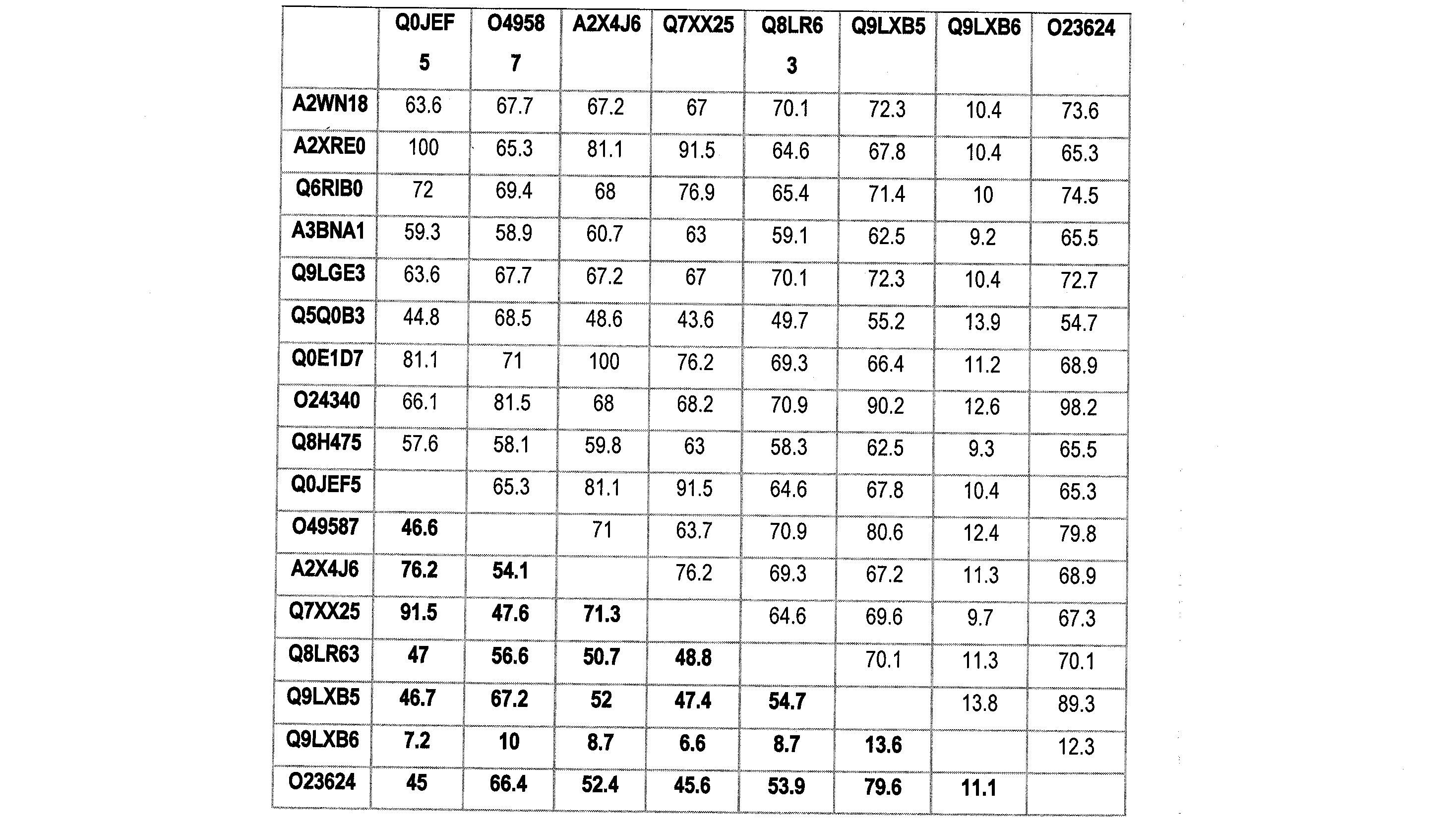
Идентичность в процентах между полипептидными последовательностями, относящимся к полипептидам SYR, используемым при выполнении способов по изобретению, может быть всего 27% аминокислотной идентичности по сравнению с SEQ ID NO:169.
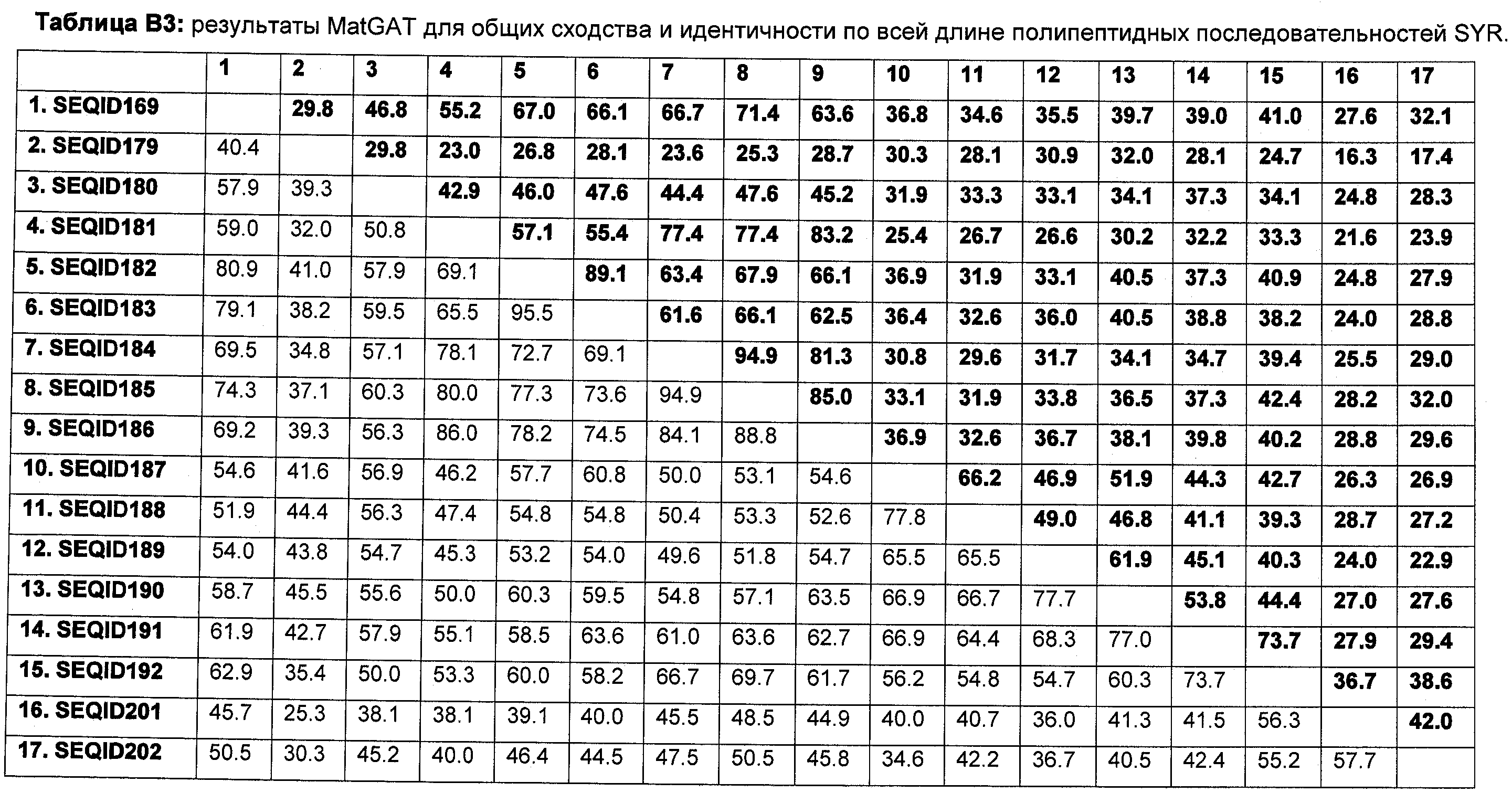
Идентичность в процентах между полипептидными последовательностями ARKL, используемыми при выполнении способов по изобретению, может быть всего 10% аминокислотной идентичности по сравнению с SEQ ID NO:213 (Orysa_ARKL1).
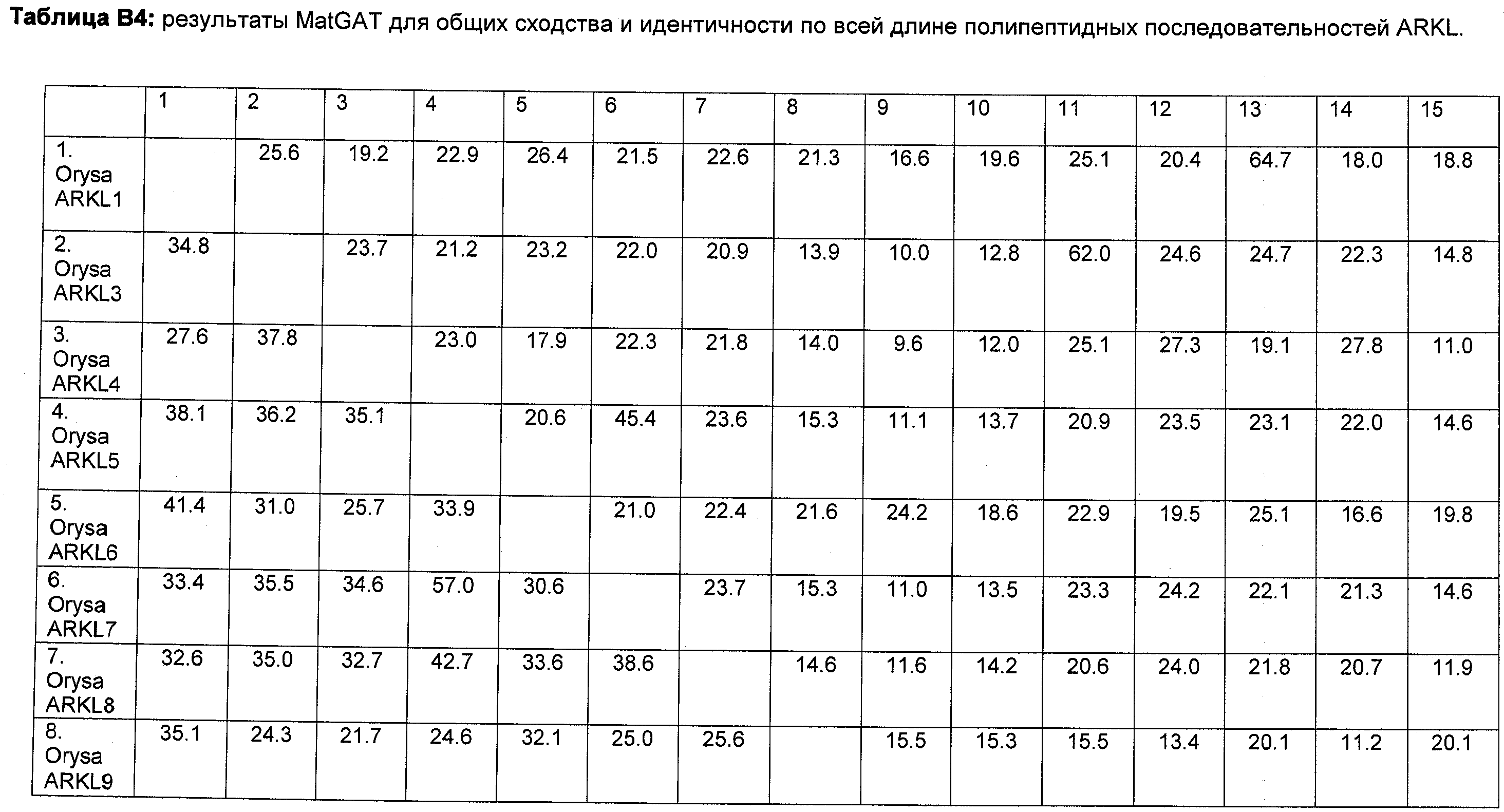
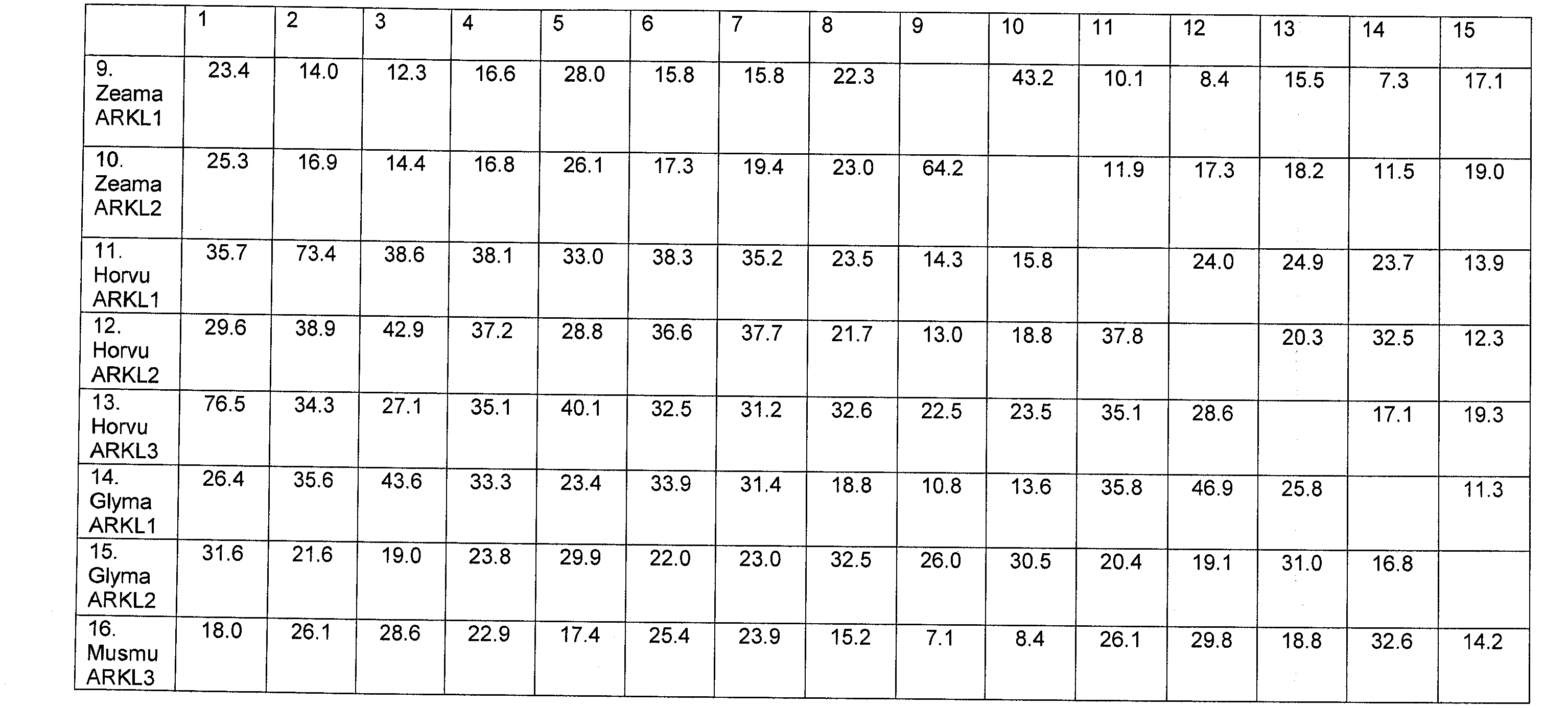
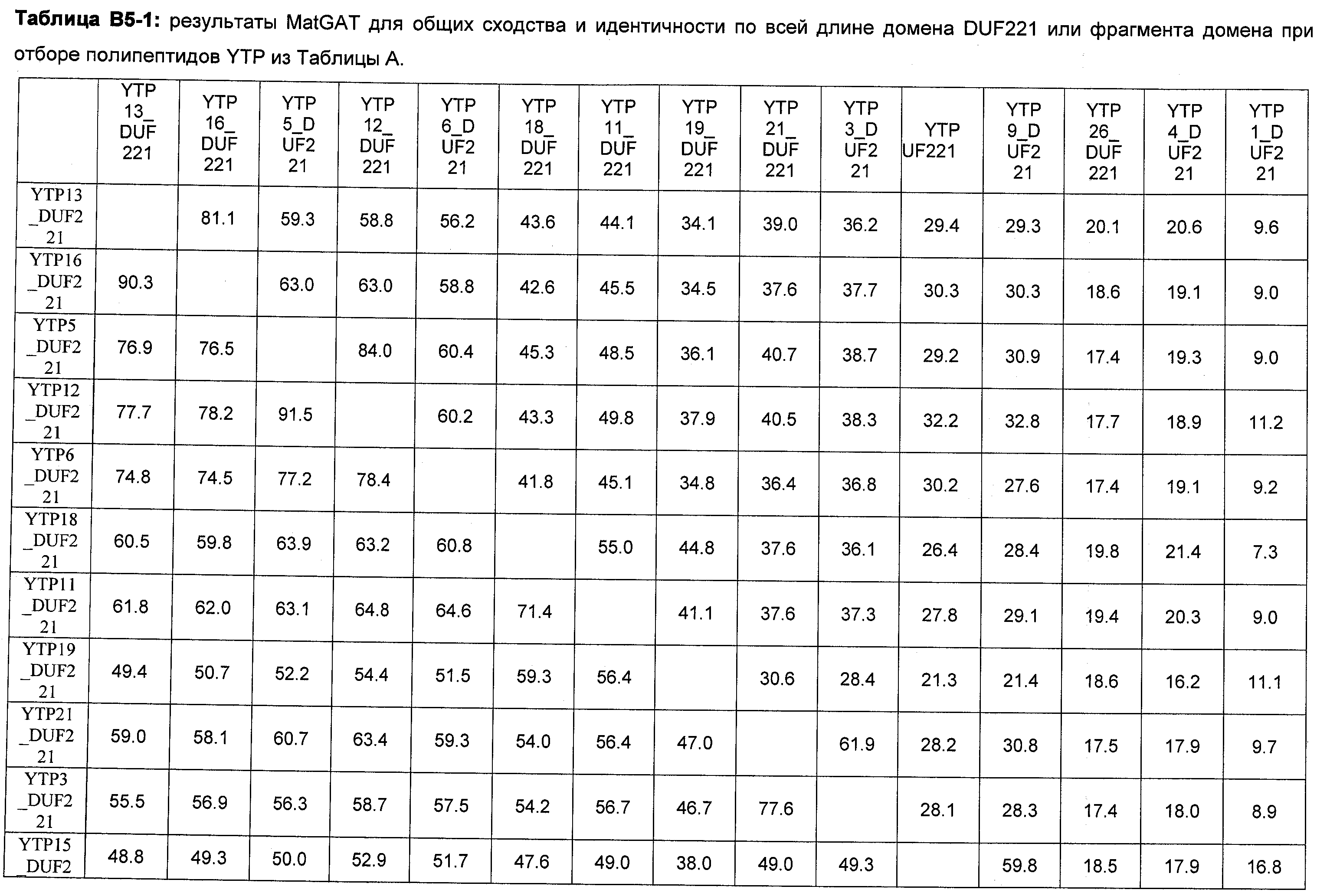
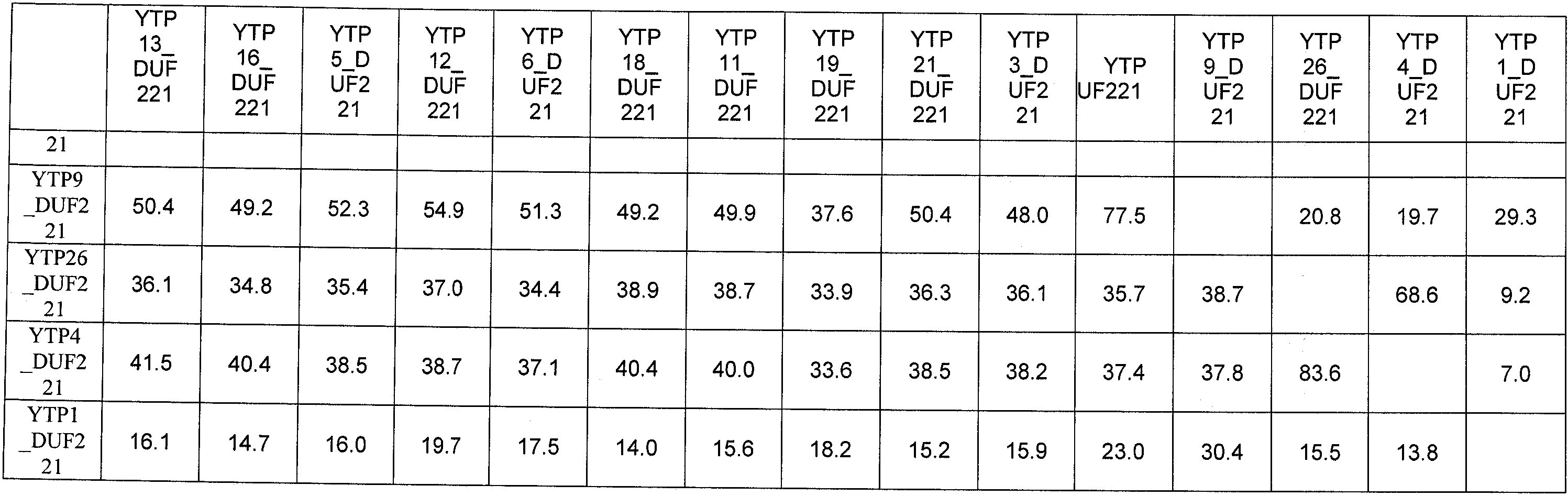
Таблица В5-2 демонстрирует SEQ ID NO:, соответствующие последовательностям, использованным в Таблице В5-1.
|
Пример 4: Идентификация доменов, содержащихся в полипептидных последовательностях, применимых для выполнения способов по изобретению
База данных Integrated Resource of Protein Families, Domains and Sites (Объединенный ресурс белковых семейств, доменов и сайтов) (InterPro) представляет собой объединенный интерфейс для обычно используемых баз данных сигнатур для поисков на основе текста и последовательностей. База данных InterPro объединяет эти базы данных, которые используют различные методологии и различающиеся уровни биологической информации по хорошо изученным белкам для получения белковых сигнатур. Сотрудничающие базы данных включают SWISS-PROT, PROSITE, TrEMBL, PRINTS, ProDom и Pfam, Smart и TIGRFAMs. InterPro размещена в European Bioinformatics Institute (Европейский институт биоинформатики) в Великобритании.
Результаты сканирования InterPro полипептидной последовательности, как представлено SEQ ID NO:2, демонстрируются в Таблице С.
|
Результаты сканирования InterPro полипептидной последовательности, как представлено SEQ ID NO:213, демонстрируются в Таблице С1.

Таблица С2 приводит SEQ ID NO:, содержащие консервативный RING-домен (ZfC3HC4) и DAR1 (PfamB2828) в полипептидах ARKL Таблицы А.
|
Результаты поиска Pfam для полипептидной последовательности, как представлено SEQ ID NO:409, приведены в Таблице С3-1 (достоверные совпадения) и в Таблице С2 (совпадения с Pfam-B).
|
|
Пример 5: Предсказание субклеточной локализации полипептидных последовательностей, применимых для выполнения способов по изобретению
Экспериментальные способы для определения локализации белка простираются от иммунолокализации до мечения белков с применением зеленого флуоресцентного белка (GFP) или бета-глюкуронидазы (GUS). Например, полипептид GRF, слитый с репортерным геном GUS, использовали, чтобы транзиентно трансформировать эпидермальные клетки лука (van der Knapp et al. (2000) Plant Phys 122:695-704). Ядро определили как субклеточный компартмент полипептида GRF. Такие способы для определения субклеточной компартментализации полипептидов GRF хорошо известны в данной области техники.
Предсказанный сигнал ядерной локализации (NLS) обнаруживали посредством множественного выравнивания последовательностей, за которым следует проверка «на глаз» (eye inspection), в домене WRC (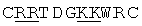 ) полипептида GRF из Таблицы А. NLS представляет собой одну или более коротких последовательностей положительно заряженных лизинов или аргининов.
) полипептида GRF из Таблицы А. NLS представляет собой одну или более коротких последовательностей положительно заряженных лизинов или аргининов.
Выполняли компьютерное предсказание локализации белка на основе данных о последовательности. Из числа алгоритмов, хорошо известных специалистам в данной области техники, доступными являются на ExPASy Proteomics tools, размещенных Swiss Institute for Bioinformatics, например, PSort, TargetP, ChloroP, LocTree, Predotar, LipoP, MITOPROT, PATS, PTS1, SignalP и другие.
LOCtree представляет собой алгоритм, который может предсказывать субклеточную локализацию и предрасположенность к ДНК-связыванию немембранных белков у нерастительных и растительных эукариот, а также прокариот. LOCtree распределяет эукариотические животные белки в один из пяти субклеточных классов, тогда как растительные белки распределяются в один из шести классов, а прокариотические белки распределяются в один из трех классов. Таблица D ниже демонстрирует результат LOCtree при использовании информации о полипептидной последовательности согласно SEQ ID NO:2. Предсказания с высокой достоверностью имеют значения показателя надежности более чем 5.

Предсказанный субклеточный компартмент полипептида GRF, как представлено SEQ ID NO:2, при использовании алгоритма LOCTree представляет собой ядро.
Пример 6: Анализ, касающийся полипептидных последовательностей, используемых для выполнения способов по изобретению
Полипептиды GRF, используемые в способах настоящего изобретения (по меньшей мере, в их нативной форме), обычно, но необязательно, имеют транскрипционную регуляторную активность и способность взаимодействовать с другими белками. ДНК-связывающая активность и взаимодействия белок-белок могут быть легко определены in vitro или in vivo при использовании методик, хорошо известных в данной области техники (например, в Current Protocols in Molecular Biology, Volumes 1 and 2, Ausubel et al. (1994), Current Protocols). Полипептиды GRF способны к транскрипционной активации репортерных генов в дрожжевых клетках (Kim & Kende (2004) Proc Natl Acad Sci 101(36): 13374-13379). Полипептиды GRF также способны взаимодействовать с полипептидами GRF-взаимодействующих факторов (GIF1 - GIF3; также называемые SYT1 - SYT3) in vivo в дрожжевых клетках при использовании анализа взаимодействия белок-белок с помощью дрожжевых двойных гибридов (Kim & Kende, supra). Анализы связывания in vitro также используют для демонстрации, что полипептиды GRF и полипептиды GIF (также называемые SYT) являются взаимодействующими партнерами (Kim & Kende, supra). Эксперименты, описанные в этой публикации, применимы для характеризования полипептидов GRF и хорошо известны в данной области техники.
Пример 7: Предсказание топологии полипептидных последовательностей, используемых при выполнении способов по изобретению
TargetP 1.1 предсказывает субклеточную локализацию эукариотических белков. Установление локализации основывается на предсказанном присутствии любой из N-концевых препоследовательностей: транзитного пептида хлоропласта (сТР), митохонд-риального направляющего пептида (mitochondrial targeting peptide (mTP)) или сигнального пептида секреторного пути (SP). Веса выравнивания (scores), на которых основывается конечное предсказание, не являются реальными вероятностями, и они не добавляются обязательно. Однако локализация с наивысшим весом выравнивания является наиболее вероятной согласно TargetP, и взаимоотношение между весами выравнивания (класс достоверности) может являться признаком, насколько определенным является предсказание. Класс достоверности (RC) ранжируется от 1 до 5, где 1 указывает на наиболее твердое предсказание. TargetP поддерживается на сервере Технического Университета Дании (Technical University of Denmark).
Для последовательностей, для которых предсказано содержание N-концевой препоследовательности, может быть также предсказан потенциальный сайт расщепления.
Отбирали ряд параметров, таких как группа организмов (не растение или растение), установки точки отсечения (cutoff) (нет, заранее определенная установка точек отсечения или определенная пользователем установка точек отсечения) и вычисление предсказания сайтов расщепления (да или нет).
Результаты анализа TargetP 1.1 полипептидной последовательности, как представлено SEQ ID NO:121, приведены в Таблице Е1. Выбрана группа организмов "растение", не заданы точки отсечения, и запрошена предсказанная длина транзитного пептида. Субклеточная локализация полипептидной последовательности, как представлено SEQ ID NO:121, вероятно представляет собой цитоплазму, и транзитный пептид (SignalP) или сигнал ядерной локализации (PredictNLS) не предсказываются.
|
Многие другие алгоритмы могут использоваться для выполнения таких анализов, включая:
- ChloroP 1.1, размещенный на сервере Технического Университета Дании;
- Protein Prowler Subcellular Localisation Predictor version 1.2, размещенный на сервере Института Молекулярной Бионауки (Institute for Molecular Bioscience), Университет Квинслэнда (University of Queensland), Brisbane, Австралия;
- PENCE Proteome Analyst PA-GOSUB 2.5, размещенный на сервере Университета Альберты (University of Alberta), Edmonton, Alberta, Канада;
- TMHMM, размещенный на сервере Технического Университета Дании.
Результаты анализа TargetP 1.1 полипептидной последовательности, как представлено SEQ ID NO:169, приводятся в Таблице Е2. Выбрана группа организмов "растение", не заданы точки отсечения, и запрошена предсказанная длина транзитного пептида. Субклеточная локализация полипептидной последовательности, как представлено SEQ ID NO:169, может представлять собой митохондрию; однако следует отметить, что класс достоверности составляет 5 (т.е. низший класс достоверности).
|
Два трансмембранных домена идентифицированы программой TMHMM, размещенной на сервере Центра анализа биологических последовательностей (Center for Biological Sequence Analysis), Технический Университет Дании. Вероятность того, что N-конец расположен внутри, составляет 0.997. Дополнительные детали по ориентации представлены в Таблице F:
|
Многие другие алгоритмы могут использоваться для выполнения таких анализов, включая:
- ChloroP 1.1, размещенный на сервере Технического Университета Дании;
- Protein Prowler Subcellular Localisation Predictor version 1.2, размещенный на сервере Института Молекулярной Бионауки, Университет Квинслэнда, Brisbane, Австралия;
- PENCE Proteome Analyst PA-GOSUB 2.5, размещенный на сервере Университета Альберты, Edmonton, Alberta, Канада.
Пример 8: Функциональный анализ полипептидов ARKL
Анализ убиквитинирования проводится по существу как описано Stone et al, 2005. Белок ARKL, меченный GST, инкубируют при 30°С и рН 7.5 с дрожжевым Е1, очищенным Е2 At UBCC8 и убиквитином (Sigma). Реакцию останавливают и анализируют электрофорезом SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) с последующим Вестерн-блоттингом с использованием антител к убиквитину.
Эксперименты по хелатообразованию с цинком проводятся путем инкубирования белка GST-ARKL, связанного с TPEN-обработанными бусинами, с ZnCl2.
Пример 9: Предсказание трансмембранной топологии полипептидных последовательностей, используемых при выполнении способов по изобретению
Алгоритм ТМНММ V 2.O (Krogh et al. 2001 J Mol Biol, 305, 567-580) использовали для предсказания трансмембранных спиралей в SEQ ID NO:409.
Как показано ниже, существуют 4 предсказанных трансмембранных спирали. Положение аминокислотных остатков спиралей также показывается. Как предполагается, петли между трансмембранными спиралями располагаются с внутренней стороны мембраны в отношении петель между остатками 28-85 и 172-373 и с внешней стороны в отношении петли между остатками 109-151.
# Sequence Number of predicted TMHs: 4
# Sequence Exp number of AAs in TMHs: 89.26923
# Sequence Exp number, first 60 AAs: 22.14249
# Sequence Total prob of N-in: 0.04519
# Sequence POSSIBLE N-term signal sequence
|
Пример 10:
a) Клонирование последовательности нуклеиновой кислоты, как представлено SEQ ID NO:1
Если не указано иное, методы рекомбинантных ДНК осуществляют согласно стандартным протоколам, описанным в Sambrook (2001) Molecular Cloning: a laboratory manual, 3rd Edition Cold Spring Harbor Laboratory Press, CSH, New York или в томах 1 и 2 у Ausubel et al. (1994), Current Protocols in Molecular Biology, Current Protocols. Стандартные материалы и методы для работ на молекулярном уровне с растениями описываются в Plant Molecular Biology Labfax (1993) by R.D.D. Croy, опубликованной by BIOS Scientific Publications Ltd (UK) and Blackwell Scientific Publications (UK).
кДНК Arabidopsis thaliana, кодирующую последовательность полипептида GRF, как представлено SEQ ID NO:2, амплифицировали ПЦР, используя в качестве матрицы банк кДНК Arabidopsis, синтезированный из мРНК, извлеченной из смешанных тканей растения. Для ПЦР амплификации использовали следующие праймеры, которые включают AttB-сайты для рекомбинации Gateway;
1) Prm 10010 (SEQ ID NO:118, смысловой):
5'-GGGGACAAGTTTGTACAAAAAAGCAGGCTTAAACAATGATGAGTCTAAGTGGAAGTAG-3'
2) Prm 10011 (SEQ ID NO:119, обратный, комплементарный): 5'-GGGGACCACTTTGTACAAGAAAGCTGGGTAGCTCTACTTAATTAGCTACCAG-3'
ПЦР выполняли, используя Hifi Taq ДНК-полимеразу в стандартных условиях. ПЦР-фрагмент ожидаемой длины (включая attB-сайты) амплифицировали и очищали, также используя стандартные способы. Затем проводилась первая стадия процедуры Gateway, реакция ВР, во время которой ПЦР-фрагмент рекомбинировал in vivo с плазмидой pDONR201 с получением, согласно терминологии Gateway, "клона ввода (entry clone)". Плазмиду pDONR201 приобретали в Invitrogen, как часть технологии Gateway®.
b) Клонирование последовательности нуклеиновой кислоты, кодирующей RAAI-подобные полипептиды, использованные в способах по изобретению
Последовательность нуклеиновой кислоты, использованную в способах по изобретению, амплифицировали посредством ПЦР, используя в качестве матрицы сделанную по заказу библиотеку кДНК сеянцев Oryza sativa (в pCMV Sport 6.0; Invitrogen, Paisley, UK). ПЦР выполняли, применяя Hifi Taq ДНК-полимеразу в стандартных условиях, используя 200 нг матрицы в 50 мкл ПЦР-смеси. Использованные праймеры представляли собой prm09129 (SEQ ID NO:122; смысловой, инициирующий кодон выделен жирным шрифтом):
5' ggggacaagtttgtacaaaaaagcaggcttaaacaatgtcaggggtttgggtg 3' и prm09988 (SEQ ID NO:123; обратный, комплементарный):
5' ggggaccactttgtacaagaaagctgggttgtcgcataggtcaatttagg 3',
которые включают AttB-сайты для рекомбинации Gateway. Амплифицированный ПЦР-фрагмент очищали, также используя стандартные способы. Затем проводилась первая стадия процедуры Gateway, реакция ВР, во время которой ПЦР-фрагмент рекомбинирует in vivo с плазмидой pDONR201 с получением, согласно терминологии Gateway, "клона ввода". Плазмиду pDONR201 приобретали в Invitrogen, как часть технологии Gateway®.
с) Генетическое планирование последовательности нуклеиновой кислоты, кодирующей полипептиды SYR
Манипуляции с ДНК: если не указано иное, методы рекомбинантных ДНК осуществляются согласно стандартным протоколам, описанным в (Sambrook (2001) Molecular Cloning: a laboratory manual, 3rd Edition Cold Spring Harbor Laboratory Press, CSH, New York) или в томах 1 и 2 Ausubel et al. (1994), Current Protocols in Molecular Biology, Current Protocols. Стандартные материалы и методы для молекулярных исследований на растениях описаны в Plant Molecular Biology Labfax (1993) by R.D.D. Croy, опубликованной BIOS Scientific Publications Ltd (UK) и Blackwell Scientific Publications (UK).
Ген SYR Oryza sativa амплифицировали посредством ПЦР, используя в качестве матрицы библиотеку кДНК сеянцев Oryza sativa (Invitrogen, Paisley, UK). После обратной транскрипции РНК, извлеченной из сеянцев, кДНК клонировали в pCMV Sport 6.0. Средний размер вставки из банка составлял 1.5 т.п.н., и первоначальное число клонов составляло порядка 1.59×107 КОЕ. Первоначальный титр, как было определено, составлял 9.6×105 КОЕ/мл после первой амплификации 6×1011 КОЕ/мл. После выделения плазмиды 200 нг матрицы использовали в 50 мкл ПЦР-смеси. Для ПЦР амплификации использовали праймеры prm08170 (SEQ ID NO:170; смысловой, инициирующий кодон выделен жирным шрифтом, AttB1-сайт выделен курсивом):
5'-ggggacaagtttgtacaaaaaagcaggcttaaacaatggaaggtgtaggtgctagg-3') и prm08171 (SEQ ID NO:171; обратный, комплементарный, AttB2-сайт выделен курсивом):
5'-ggggaccactttgtacaagaaagctgggtcaaaaacaaaaataaattcccc-3'), которые включают AttB-сайты для рекомбинации Gateway. ПЦР выполняли, применяя Hifi Taq ДНК-полимеразу в стандартных условиях. Амплифицированный ПЦР-фрагмент соответствующего размера амплифицировали и очищали, также используя стандартные способы. Затем проводилась первая стадия процедуры Gateway, реакция ВР, во время которой ПЦР-фрагмент рекомбинирует in vivo с плазмидой pDONR201 с получением, согласно терминологии Gateway, "клона ввода". Плазмиду pDONR201 приобретали в Invitrogen, как часть технологии Gateway®.
d) Клонирование последовательности нуклеиновой кислоты, кодирующей полипептиды ARKL, использованные в способах по изобретению
Последовательность нуклеиновой кислоты, использованную в способах по изобретению, амплифицировали ПЦР, используя в качестве матрицы сделанную по заказу библиотеку кДНК сеянцев и метелок Oryza sativa (в pCMV Sport 6.0; Invitrogen, Paisley, UK). ПЦР выполняли, применяя Hifi Taq ДНК-полимеразу в стандартных условиях, используя 200 нг матрицы в 50 мкл ПЦР-смеси. Использованные праймеры представляли собой prm04873(SEQ ID NO:404; смысловой, инициирующий кодон выделен жирным шрифтом):
5'-ggggacaagtttgtacaaaaaagcaggcttaaacaatggatgatcacatgggaaga-3' и prm04874 (SEQ ID NO:405; обратный, комплементарный):
5'-ggggaccactttgtacaagaaagctgggttttggtttctgaagaagcacc-3', которые включают AttB-сайты для рекомбинации Gateway. Амплифицированный ПЦР-фрагмент очищали, также используя стандартные способы. Затем проводилась первая стадия процедуры Gateway, реакция ВР, во время которой ПЦР-фрагмент рекомбинирует in vivo с плазмидой pDONR201 с получением, согласно терминологии Gateway, "клона ввода". Плазмиду pDONR201 приобретали в Invitrogen, как часть технологии Gateway®.
e) Клонирование нуклеиновой кислоты, кодирующей полипептиды УТР, использованные в способах по изобретению
Последовательность нуклеиновой кислоты, использованную в способах по изобретению, амплифицировали ПЦР, используя в качестве матрицы сделанную по заказу библиотеку кДНК сеянцев Oryza sativa (в pCMV Sport 6.0; Invitrogen, Paisley, UK). ПЦР выполняли, применяя Hifi Taq ДНК-полимеразу в стандартных условиях, используя 200 нг матрицы в 50 мкл ПЦР-смеси. Использованные праймеры представляли собой (SEQ ID NO:546; смысловой, инициирующий кодон выделен жирным шрифтом):
5'-ggggacaagtttgtacaaaaaagcaggcttaaacaatggacaccgcgtcgt-3' and (SEQ ID NO:547; обратный, комплементарный):
5'-ggggaccactttgtacaagaaagctgggtcagcacttgcattagatggat-3', которые включают AttB-сайты для рекомбинации Gateway. Амплифицированный ПЦР-фрагмент очищали, также используя стандартные способы. Затем проводилась первая стадия процедуры Gateway, реакция ВР, во время которой ПЦР-фрагмент рекомбинирует in vivo с плазмидой pDONR201 с получением, согласно терминологии Gateway, "клона ввода". Плазмиду pDONR201 приобретали в Invitrogen, как часть технологии Gateway®.
Пример 11:
a) Конструирование вектора экспрессии с использованием последовательности нуклеиновой кислоты, как представлено SEQ ID NO:1
Клон ввода, содержащий SEQ ID NO:1, затем использовали в реакции LR с вектором назначения (destination vector), применяемым для трансформации Oryza sativa. Этот вектор содержал в качестве функциональных элементов внутри границ Т-ДНК: селектируемый маркер растений; экспрессионную кассету маркера, подходящего для скрининга; и кассету Gateway, предназначенную для рекомбинации LR in vivo с рассматриваемой последовательностью нуклеиновой кислоты, уже клонированной в клон ввода. Рисовый промотор GOS2 (SEQ ID NO:117) для конститутивной экспрессии располагался выше данной кассеты Gateway.
После стадии рекомбинации LR получающийся вектор экспрессии pGOS2::GRF (Фиг.4) трансформировали в штамм Agrobacterium LBA4044 согласно способам, хорошо известным в данной области техники.
b) Конструирование вектора экспрессии с использованием последовательности нуклеиновой кислоты, как представлено SEQ ID NO:120
Клон ввода, содержащий SEQ ID NO:120, затем использовали в реакции LR с вектором назначения, применяемым для трансформации Oryza sativa. Этот вектор содержал в качестве функциональных элементов внутри границ Т-ДНК: селектируемый маркер растений; экспрессионную кассету маркера, подходящего для скрининга; и кассету Gateway, предназначенную для рекомбинации LR in vivo с рассматриваемой последовательностью нуклеиновой кислоты, уже клонированной в клон ввода. Рисовый промотор GOS2 (SEQ ID NO:124) для конститутивной экспрессии располагался выше данной кассеты Gateway. В альтернативном варианте осуществления рисовый промотор HMPG (SEQ ID NO:125) для конститутивной экспрессии располагался выше кассеты Gateway.
После стадии рекомбинации LR получающийся вектор экспрессии pGOS2::RAA1-подобный (Фиг.9) трансформировали в штамм Agrobacterium LBA4044 согласно способам, хорошо известным в данной области техники.
c) Конструирование вектора экспрессии с использованием последовательности нуклеиновой кислоты, кодирующей полипептиды SYR
Клон ввода pSYR затем использовали в реакции LR с вектором назначения, применяемым для трансформации Oryza sativa. Этот вектор содержал в качестве функциональных элементов внутри границ Т-ДНК: селектируемый маркер растений; экспрессионную кассету маркера, подходящего для скрининга; и кассету Gateway, предназначенную для рекомбинации LR in vivo с рассматриваемой последовательностью нуклеиновой кислоты, уже клонированной в клон ввода. Рисовый промотор GOS2 (SEQ ID NO:211) для конститутивной экспрессии располагался выше данной кассеты Gateway. Получали схожую конструкцию вектора, но с промотором белка группы с высокой подвижностью (HMGP, SEQ ID NO:200 или SEQ ID NO:210) вместо промотора GOS.
После стадии рекомбинации LR получающиеся векторы экспрессии pGOS2::SYR (с промотором GOS2) и pHMGP::SYR (с промотором HMGP), оба для конститутивной экспрессии SYR, трансформировали в штамм Agrobacterium LBA4044 и затем в растения Oryza sativa.
d) Конструирование вектора экспрессии с использованием последовательности нуклеиновой кислоты, как представлено SEQ ID NO:212
Клон ввода, содержащий SEQ ID NO:212, затем использовали в реакции LR с вектором назначения, применяемым для трансформации Oryza sativa. Этот вектор содержал в качестве функциональных элементов внутри границ Т-ДНК: селектируемый маркер растений; экспрессионную кассету маркера, подходящего для скрининга; и кассету Gateway, предназначенную для рекомбинации LR in vivo с рассматриваемой последовательностью нуклеиновой кислоты, уже клонированной в клон ввода. Рисовый промотор GOS2 (SEQ ID NO:406) для экспрессии, специфичной для корней, располагался выше данной кассеты Gateway.
После стадии рекомбинации LR получающийся вектор экспрессии pGOS2::ARKL (Фиг.18) трансформировали в штамм Agrobacterium LBA4044 согласно способам, хорошо известным в данной области техники.
e) Конструирование вектора экспрессии с использованием последовательности нуклеиновой кислоты, как представлено SEQ ID NO:408
Клон ввода, содержащий SEQ ID NO:408, затем использовали в реакции LR с вектором назначения, применяемым для трансформации Oryza sativa. Этот вектор содержал в качестве функциональных элементов внутри границ Т-ДНК: селектируемый маркер растений; экспрессионную кассету маркера, подходящего для скрининга; и кассету Gateway, предназначенную для рекомбинации LR in vivo с рассматриваемой последовательностью нуклеиновой кислоты, уже клонированной в клон ввода. Рисовый промотор GOS2 (SEQ ID NO:548) для экспрессии, специфичной для корней, располагался выше данной кассеты Gateway.
После стадии рекомбинации LR получающийся вектор экспрессии pGOS2::YTP1 (Фиг.23) трансформировали в штамм Agrobacterium LBA4044 согласно способам, хорошо известным в данной области техники.
Пример 12: Трансформация растений
Трансформация риса
Агробактерии, содержащие вектор экспрессии, использовали для трансформации растений Oryza sativa. Вылущивали зрелые сухие семена риса японского сорта Nipponbare. Стерилизацию проводили инкубированием в течение одной минуты в 70% этаноле с последующими 30 минутами в 0.2% HgCl2, после чего следовала 6-кратная 15-минутная промывка стерильной дистиллированной водой. Стерильные семена затем проращивали в среде, содержащей 2,4-D (среда для индукции каллуса). После инкубации в темноте в течение 4 недель эмбриогенные, образованные из щитка каллусы срезали и размножали в той же среде. Через две недели каллусы увеличивали или размножали субкультивированием на той же среде в течение еще 2 недель. Кусочки эмбриогенного каллуса субкультивировали на свежей среде 3 дня перед совместным культивированием (для увеличения активности деления клеток).
Штамм агробактерий LBA4404, содержащий каждый индивидуальный вектор экспрессии, использовали независимо для совместного культивирования. Агробактерии инокулировали в среду АВ с подходящими антибиотиками и культивировали в течение 3 дней при 28°С. Затем бактерии собирали и суспендировали в жидкой среде для совместного культивирования до плотности (OD600) примерно 1. Суспензию затем переносили на чашку Петри, и каллусы погружали в суспензию на 15 минут. Каллусные ткани затем подвергали сухому блоттингу на фильтровальную бумагу и переносили на затвердевшую среду для совместного культивирования, и инкубировали в течение 3 дней в темноте при 25°С. Совместно культивированные каллусы выращивали на 2,4-D-содержащей среде в течение 4 недель в темноте при 28°С в присутствии агента селекции. Во время этого периода развивались островки быстро растущего устойчивого каллуса. После переноса этого материала на регенерационную среду и инкубации на свету высвобождался эмбриогенный потенциал, и побеги развивались во время следующих 4-5 недель. Побеги срезали с каллусов и инкубировали в течение 2-3 недель на среде, содержащей ауксин, с которой они переносились в почву. Затвердевшие побеги выращивали при высокой влажности и коротких днях в теплице.
Приблизительно 35 независимых рисовых трансформантов ТО получали для каждой конструкции. Первичные трансформанты переносили из камеры для культивирования тканей в теплицу. После количественного ПЦР-анализа для проверки числа копий вставки Т-ДНК только трансгенные растения с одной копией, которые демонстрировали устойчивость к агенту селекции, сохраняли для сбора семян Т1. Семена затем собирали через 3-5 месяцев после пересадки. Этот способ давал трансформанты с одним локусом на уровне свыше 50% (Aldemita and Hodges1996, Chan et al. 1993, Hiei et al. 1994).
Пример 13: Процедура фенотипической оценки
13.1-1 Общая информация по параметрам оценки
Получали приблизительно 35 независимых рисовых трансформантов ТО. Первичные трансформанты переносили из камеры для культивирования тканей в теплицу для выращивания и сбора семян Т1. Поддерживали шесть случаев, у которых потомство Т1 имело расщепление 3:1 по присутствию/отсутствию трансгена. Для каждого из этих случаев приблизительно 10 сеянцев Т1, содержащих трансген (гетеро- и гомозиготы), и приблизительно 10 сеянцев Т1, потерявших трансген (нуль-зиготы), отбирали путем мониторинга экспрессии визуального маркера. Трансгенные растения и соответствующие нуль-зиготы выращивали вместе при произвольно выбранном расположении. Тепличные условия представляют собой короткие дни (12 часов света), 28°С на свету и 22°С в темноте, и относительную влажность 70%.
13.1-2 Параметры оценки для растений, трансформированных SYR под контролем рисового промотора GOS2 или промотора HMGP
Получали приблизительно 15-20 независимых рисовых трансформантов Т0. Первичные трансформанты переносили из камеры для культивирования тканей в теплицу для выращивания и сбора семян Т1. Поддерживали восемь случаев, у которых потомство Т1 имело расщепление 3:1 по присутствию/отсутствию трансгена. Для каждого из этих случаев приблизительно 10 сеянцев Т1, содержащих трансген (гетеро- и гомозиготы), и приблизительно 10 сеянцев Т1, потерявших трансген (нуль-зиготы), отбирали путем мониторинга экспрессии визуального маркера. Отобранные растения Т1 переносили в теплицу. Каждое растение получало индивидуальную штрихкодовую метку, чтобы однозначно связать фенотипические данные с соответствующим растением. Выбранные растения Т1 выращивали в земле в горшках диаметром 10 см при следующих установках: длина светового дня = 11.5 ч, интенсивность дневного света = 30000 люкс или более, температура в дневное время = 28°С или выше, температура в ночное время = 22°С, относительная влажность = 60-70%.
Общие параметры
От стадии посева до стадии зрелости растения пропускали несколько раз через камеру для получения цифровых изображений. В каждый момент времени цифровые изображения (2048×1536 пикселей, 16 миллионов цветов) получали для каждого растения по меньшей мере с 6 различных углов.
Четыре случая Т1 дополнительно оценивали в поколении Т2, следуя такой же процедуре оценки, как для поколения Т1, но с большим количеством представителей на случай. От стадии посева до стадии зрелости растения пропускали несколько раз через камеру для получения цифровых изображений. В каждый момент времени цифровые изображения (2048×1536 пикселей, 16 миллионов цветов) получали для каждого растения по меньшей мере с 6 различных углов.
13.2 Статистический анализ: F-критерий
Двухфакторный анализ ANOVA (анализ вариантов) использовали как статистическую модель для общей оценки фенотипических признаков растения. F-критерий определялся для всех параметров, измеренных для всех растений всех случаев, трансформированных геном по настоящему изобретению. F-критерий определяли, чтобы проверить воздействие гена на все случаи трансформации и подтвердить общее воздействие гена, также известное как общий эффект гена. Порог значимости для достоверного общего эффекта гена устанавливали при 5% уровне вероятности для F-критерия. Значимое значение F-критерия указывает на эффект гена, подразумевая, что это не только всего лишь присутствие или расположение гена, которое вызывает различия в фенотипе.
Поскольку производились два эксперимента с перекрывающимися событиями, проводился факторный анализ. Это применимо для проверки согласованности эффектов в двух экспериментах, и если дело обстоит так, для суммирования доказательств из двух экспериментов, чтобы увеличить достоверность заключения. Использованный способ представляет собой принцип смешанной модели, который учитывает многоуровневую структуру данных (т.е. эксперимент - событие - сегреганты).
Р-значения получали сравнением критерия отношения правдоподобия с распределениями хи-квадрат.
13.3 Измеренные параметры
Измерение параметров, связанных с биомассой (общий способ)
От стадии посева до стадии зрелости растения пропускали несколько раз через камеру для получения цифровых изображений. В каждый момент времени цифровые изображения (2048×1536 пикселей, 16 миллионов цветов) получали для каждого растения по меньшей мере с 6 различных углов.
Площадь надземной части растения (или лиственную биомассу) определяли подсчетом общего числа пикселей на цифровых изображениях надземных частей растений, отличных от фона. Это значение усреднялось для снимков, сделанных в одно и то же время под различными углами, и преобразовывалось в величину физической поверхности, выраженную в кв. мм, путем калибровки. Эксперименты демонстрируют, что измеренная таким образом площадь надземных частей растений коррелирует с биомассой надземных частей растений. Площадь надземных частей представляет собой площадь, измеренную в момент времени, в который растение достигало своей максимальной лиственной биомассы. Ранняя мощность представляет собой площадь надземной части растения (сеянца) через три недели после прорастания. Увеличение корневой биомассы выражается как увеличение общей биомассы корней (измеренной как максимальная биомасса корней, обнаруженных во время продолжительности жизни растений); или как увеличение показателя корни/побеги (измеренного как соотношение между массой корней и массой побегов в период активного роста корней и побегов).
Раннюю мощность определяли подсчетом общего числа пикселей надземных частей растений, отличных от фона. Это значение усреднялось для снимков, сделанных в одно и то же время под различными углами, и преобразовывалось в величину физической поверхности, выраженную в кв. мм, путем калибровки. Результаты, описанные ниже, даны для растений через три недели после прорастания.
Измерение параметров, связанных с семенами (общий способ)
Собирали зрелые первичные метелки, подсчитывали, упаковывали в пакеты, метили штрихкодом и затем высушивали в течение трех дней в печи при 37°С. Затем метелки обмолачивали, и все семена собирали и подсчитывали. Наполненные пленки отделяли от пустых, используя воздуходувное устройство. Пустые пленки отбрасывали и оставшуюся фракцию пересчитывали вновь. Наполненные пленки взвешивали на аналитических весах. Число наполненных семян определяли подсчетом числа наполненных пленок, которые оставались после стадии разделения. Общая масса семян на растение измерялась взвешиванием всех наполненных пленок, собранных с растения. Общее число семян на растение измеряли подсчетом числа пленок, собранных с растения. Масса тысячи зерен (TKW) экстраполировалась из числа подсчитанных наполненных семян и их общей массы. Индекс урожайности (HI) в настоящем изобретении определяли как соотношение между общей массой семян на растение и площадью надземных частей (мм2), умноженное на 106. Общее число цветков на метелку, как определено в настоящем изобретении, представляет собой соотношение между общим числом семян и числом зрелых первичных метелок. Уровень наполнения семян, как определено в настоящем изобретении, представляет собой содержание (выраженное в %) числа наполненных семян относительно общего числа семян (или цветков).
Скрининг эффективности применения азота (для растений, трансформированных SYR)
Рисовые растения из семян Т2 выращивали в почве для горшечных культур при нормальных условиях за исключением питательного раствора. Горшки поливали от пересадки до созревания особым питательным раствором, имеющим сниженное содержание азота (N), обычно в 7-8 раз меньше. Остальное культивирование (созревание растения, сбор семян) являлось таким же, как для растений, не выращивавшихся при абиотическом стрессе. Параметры роста и урожайности регистрируются, как подробно описано для роста при нормальных условиях.
Скрининг при стрессе, вызванном засухой (для растений, трансформированных SYR)
Растения из Т1, Т2 или дальнейших поколений выращивали в почве для горшечных культур при нормальных условиях до тех пор, пока они не достигали фазы выметывания. Затем их переносили в "сухую" зону, где приостанавливали полив. Зонды измерения влажности вставляли в случайным образом выбранные горшки для наблюдения за содержанием воды в почве (SWC). Когда SWC опускалось ниже определенного порога, растения автоматически непрерывно повторно поливают до тех пор, пока снова не достигают нормального уровня. Растения затем повторно переносили снова в нормальные условия. Остальное культивирование (созревание растения, сбор семян) являлось таким же, как для растений, не выращивавшихся в условиях абиотического стресса. Параметры роста и урожайности регистрировали, как подробно описано для роста при нормальных условиях. Применяемые условия засухи представляли собой "условия сильной засухи", как определено выше.
Пример 14: Результаты фенотипической оценки трансгенных рисовых растений, экспрессирующих последовательность нуклеиновой кислоты, кодирующую полипептид GRF, как представлено SEQ ID NO:2
Ниже представлены результаты оценки трансгенных рисовых растений, экспрессирующих последовательность нуклеиновой кислоты, кодирующую полипептид GRF, как представлено SEQ ID NO:2, под контролем промотора GOS2 для конститутивной экспрессии, выращенных при нормальных условиях роста.
Существовало значительное увеличение ранней мощности, надземной биомассы, общей урожайности семян на растение, уровня наполнения семян, индекса урожайности и массы тысячи зерен (TKW) у трансгенных растений по сравнению с соответствующими нуль-зиготами (контроли), как показано в Таблице G.
|
Пример 15:
а) Результаты фенотипической оценки трансгенных рисовых растений, экспрессирующих последовательности нуклеиновых кислот, кодирующие другие полипептиды GRF
Получали трансгенные рисовые растения, независимо экспрессирующие последовательности нуклеиновых кислот, кодирующие другие полипептиды GRF, как показано в Таблице Н ниже, под контролем промотора GOS2 для конститутивной экспрессии.
Существовало значительное увеличение массы тысячи зерен (TKW) для семян трансгенных растений по сравнению с соответствующими нуль-зиготами (контроли) для трех конструкций. Это увеличение было менее выражено, чем у семян трансгенных растений, экспрессирующих последовательность нуклеиновой кислоты, кодирующую полипептид GRF, как представлено SEQ ID NO:2.
|
b) Результаты фенотипической оценки трансгенных растений, экспрессирующих RAA1-подобную нуклеиновую кислоту
Результаты оценки трансгенных рисовых растений, экспрессирующих RAA1-подобную нуклеиновую кислоту под контролем конститутивного промотора (GOS2 или HMGP) в условиях, не приводящих к стрессу, были следующими: увеличение по меньшей мере на 2% было обнаружено для массы тысячи зерен и увеличение более чем на 5% было обнаружено для по меньшей мере одного из следующих параметров: показатель корни/побеги, общая биомасса корней, цветки на метелку. Также в условиях сниженной доступности азота было обнаружено увеличение одного или более из: биомасса корней, высота и индекс зелени.
с1) измерение связанных с урожайностью параметров для трансформантов pGOS2::SYR, выращенных в условиях недостаточности питательных веществ:
В результате анализа семян, как описано выше, изобретатели обнаружили, что растения, трансформированные генной конструкцией pGOS2::SW и выращенные при стрессе, вызванном недостаточностью питательных веществ, имели более высокую урожайность семян, выраженную как число наполненных семян (увеличение более чем 5%), общая масса семян (увеличение более чем 5%) и TKW (увеличение более чем 2.5%), по сравнению с растениями, не имеющими трансгена SYR. Также было обнаружено увеличение биомассы побегов (более чем 5%) и биомассы корней (несколько линий - более чем 5%).
с2) измерение связанных с урожайностью параметров для трансформантов pGOS2::SYR, выращенных в условиях стресса, вызванного сильной засухой:
В результате анализа семян, как описано выше, изобретатели обнаружили, что растения, трансформированные генной конструкцией pGOS2::SYR и выращенные при стрессе, вызванном сильной засухой, имели более высокую урожайность семян, выраженную как общая масса семян (увеличение более чем на 5%), уровень наполнения (увеличение более чем 5%) и индекс урожайности (увеличение более чем 5%), по сравнению с растениями, не имеющими трансгена SYR.
d) Результаты фенотипической оценки трансгенных растений, экспрессирующих нуклеиновую кислоту Orysa_ARKL1
Результаты оценки трансгенных рисовых растений, экспрессирующих нуклеиновую кислоту Orysa_ARKL1, в условиях, не приводящих к стрессу, представлены ниже. Увеличение по меньшей мере на 5% было обнаружено для мощности прорастания (emergence vigour) (ранняя мощность), общей урожайности семян, числа наполненных семян, массы тысячи зерен и индекса урожайности, и на 3% для массы тысячи зерен.
|
Трансгенные рисовые растения, экспрессирующие нуклеиновую кислоту Orysa_ARKL1, также оценивали в условиях стресса, вызванного засухой, как описано выше. Те же параметры (увеличение урожайности семян, числа наполненных семян, ранней мощности и индекса урожайности) также увеличивались у трансгенных растений по сравнению с соответствующим контрольным растением, хотя в более низкой степени.
е) Результаты фенотипической оценки трансгенных растений, экспрессирующих нуклеиновую кислоту YTP1
Результаты оценки трансгенных рисовых растений, экспрессирующих нуклеиновую кислоту YTP1, в условиях, не приводящих к стрессу, представлены ниже. Увеличение по меньшей мере на 5% было обнаружено для общей урожайности семян, уровня наполнения семян, числа цветков на метелку, индекса урожайности, и на 2% для массы тысячи зерен.
Результаты оценки трансгенных рисовых растений, экспрессирующих нуклеиновую кислоту YTP1, в условиях, не приводящих к стрессу, представлены ниже. Увеличение было обнаружено для общей массы семян, числа наполненных семян, уровня наполнения, индекса урожайности и массы тысячи зерен (Таблица J).
|
Пример 16: Примеры трансформации других сельскохозяйственных культур Трансформация кукурузы
Трансформация кукурузы (Zea mays) проводится с модификацией способа, описанного Ishida et al. (1996) Nature Biotech 14(6): 745-50. Трансформация является зависимой от генотипа у кукурузы, и только определенные генотипы поддаются трансформации и регенерации. Инбредная линия А188 (Университет Миннесоты (University of Minnesota)) или гибриды с А188 в качестве исходного растения являются хорошими источниками донорного материала для трансформации, но другие генотипы могут также удачно использоваться. Початки собирают с кукурузного растения приблизительно через 11 дней после опыления (DAP), когда длина незрелого зародыша составляет от около 1 до 1.2 мм. Незрелые зародыши культивируют совместно с Agrobacterium tumefaciens, содержащими вектор экспрессии, и трансгенные растения восстанавливают органогенезом. Срезанные зародыши выращивают на среде индукции каллуса, затем на среде регенерации кукурузы, содержащей агент селекции (например, имидазолинон, но могут использоваться различные маркеры селекции). Чашки Петри инкубируются на свету при 25°С в течение 2-3 недель, или до развития побегов. Зеленые побеги переносятся из каждого зародыша в среду для укоренения кукурузы и инкубируются при 25°С в течение 2-3 недель до тех пор, пока разовьются корни. Укорененные побеги пересаживаются в почву в теплицу. Семена Т1 получают из растений, которые проявляют устойчивость к агенту селекции и которые содержат одну копию вставки Т-ДНК.
Трансформация пшеницы
Трансформацию пшеницы проводят способом, описанным Ishida et al. (1996) Nature Biotech 14(6): 745-50. Как правило, культурный сорт Bobwhite (доступен от CIMMYT, Mexico) используется для трансформации. Незрелые зародыши совместно культивируют с Agrobacterium tumefaciens, содержащими вектор экспрессии, и трансгенные растения восстанавливают органогенезом. После инкубации с агробактериями зародыши выращивают in vitro на среде индукции каллуса, затем на среде для регенерации, содержащей агент селекции (например, имидазолинон, но могут использоваться различные маркеры селекции). Чашки Петри инкубируются на свету при 25°С в течение 2-3 недель, или до развития побегов. Зеленые побеги переносятся из каждого зародыша в среду для укоренения и инкубируются при 25°С в течение 2-3 недель до тех пор, пока разовьются корни. Укорененные побеги пересаживаются в почву в теплицу. Семена Т1 получают из растений, которые проявляют устойчивость к агенту селекции и которые содержат одну копию вставки Т-ДНК.
Трансформация сои
Сою трансформируют согласно модификации способа, описанного в патенте US 5,164,310, Texas A&M. Несколько коммерческих сортов сои поддаются трансформации этим способом. Как правило, для трансформации используется культурный сорт Jack (доступный от Illinois Seed foundation). Семена сои стерилизуют для посева in vitro. Гипокотиль, зародышевый корень и одну семядолю вырезают из молодых семидневных сеянцев. Эпикотиль и оставшуюся семядолю дополнительно выращивают для развития добавочных узлов. Эти добавочные узлы вырезают и инкубируют с Agrobacterium tumefaciens, содержащими вектор экспрессии. После процедуры совместного культивирования эксплантаты промывают и переносят в селекционную среду. Регенерированные побеги срезают и помещают в среду для удлинения побегов. Побеги не длиннее чем 1 см помещают на среду для укоренения до тех пор, пока разовьются корни. Укорененные побеги пересаживают в почву в теплицу. Семена Т1 получают из растений, которые проявляют устойчивость к агенту селекции и которые содержат одну копию вставки Т-ДНК.
Трансформация рапса/канолы
Черешки семядолей и гипокотили 5-6-дневного сеянца используют как эксплантаты для тканевой культуры и трансформируют согласно Babic et al. (1998, Plant Cell Rep 17:183-188). Коммерческий культурный сорт Westar (Agriculture Canada) является стандартным сортом, используемым для трансформации, но другие сорта также могут быть использованы. Семена канолы подвергают поверхностной стерилизации для посева in vitro. Эксплантаты в виде черешков семядолей с присоединенной семядолей вырезают из in vitro сеянцев и инокулируют агробактериями (содержащими вектор экспрессии) погружением срезанного конца черешкового эксплантата в бактериальную суспензию. Эксплантаты затем культивируют в течение 2 дней на среде MSBAP-3, содержащей 3 мг/л ВАР (бензиламинопурин), 3% сахарозы, 0.7% Phytagar при 23°С, 16 ч света. Через два дня совместного культивирования с агробактериями черешковые эксплантаты пересаживают на среду MSBAP-3, содержащую 3 мг/л ВАР, цефотаксим, карбенициллин или тиментин (300 мг/л), на 7 дней и затем культивируют на среде MSBAP-3 с цефотаксимом, карбенициллином или тиментином и агентом селекции до регенерации побегов. Когда побеги достигают 5-10 мм в длину, их срезают и пересаживают на среду элонгации побегов (MSBAP-0.5, содержащая 0.5 мг/л ВАР). Побеги примерно 2 см в длину пересаживают на среду для укоренения (MSO) для индукции корней. Укорененные побеги пересаживают в почву в теплицу. Семена Т1 получают из растений, которые проявляют устойчивость к агенту селекции и которые содержат одну копию вставки Т-ДНК.
Трансформация люцерны
Регенерирующий клон люцерны (Medicago sativa) трансформируют, используя способ McKersie et al., 1999 Plant Physiol 119:839-847. Регенерация и трансформация люцерны является зависимой от генотипа, и следовательно, требуется регенерирующее растение. Способы получения регенерирующих растений описаны. Например, они могут быть выбраны из культурного сорта Rangelander (Agriculture Canada) или других коммерческих сортов люцерны, как описано у Brown DCW и A Atanassov (1985. Plant Cell Tissue Organ Culture 4:111-112). Альтернативно, сорт RA3 (Университет Висконсина (University of Wisconsin)) выбирают для применения в тканевой культуре (Walker et al., 1978 Am J Bot 65:654-659). Черешковые эксплантаты совместно культивируют с ночной культурой Agrobacterium tumefaciens C58C1 рМР90 (McKersie et al., 1999 Plant Physiol 119:839-847) или LBA4404, содержащими вектор экспрессии. Эксплантаты совместно культивируют в течение 3 дней в темноте на среде индукции SH (среда Шенка и Хильдебрандта), содержащей 288 мг/л Pro, 53 мг/л тиопролина, 4.35 г/л K2SO4 и 100 мкМ ацетосирингона. Эксплантаты промывают в среде Мурасиге и Скуга с половинной концентрацией (Murashige and Skoog, 1962) и помещают на чашки на такую же среду индукции SH без ацетосирингона, но с подходящим агентом селекции и подходящим антибиотиком для ингибирования роста агробактерий. Через несколько недель соматические зародыши пересаживают в среду для развития BOi2Y, не содержащую никаких регуляторов роста, никаких антибиотиков, и содержащую 50 г/л сахарозы. Соматические зародыши впоследствии развиваются на среде Мурасиге и Скуга с половинной концентрацией. Укоренившиеся сеянцы пересаживают в горшки и выращивают в теплице. Семена Т1 получают из растений, которые проявляют устойчивость к агенту селекции и которые содержат одну копию вставки Т-ДНК.
Трансформация хлопка
Трансформация хлопка (Gossypium hirsutum L.) проводится с использованием Agrobacterium tumefaciens на эксплантатах из гипокотиля. Коммерческие культурные сорта, такие как Coker 130 или Coker 312 (SeedCo, Lubbock, TX) являются стандартными сортами, используемыми для трансформации, но другие сорта могут также использоваться. Семена подвергают поверхностной стерилизации и проращивают в темноте. Эксплантаты из гипокотиля вырезают из проращенных сеянцев длиной около 1-1.5 см. Эксплантат из гипокотиля погружают в инокулят Agrobacterium tumefaciens, содержащий вектор экспрессии, в течение 5 минут, затем совместно культивируют в течение около 48 часов на MS+1.8 мг/л KNO3+2% глюкозы при 24°С в темноте. Эксплантаты переносят на такую же среду, содержащую подходящие бактериальные и растительные селектируемые маркеры (обновляемую несколько раз) до обнаружения эмбриогенных каллусов. Каллусы отделяют и субкультивируют до появления соматических зародышей. Ростки, полученные из соматических зародышей, созревают на среде для укоренения до появления корней. Укорененные побеги пересаживают в почву для горшечных культур в теплицу. Семена Т1 получают из растений, которые проявляют устойчивость к агенту селекции и которые содержат одну копию вставки Т-ДНК.
Пример 17: Примеры скринингов при абиотических стрессах
Скрининг при засухе
Растения из выбранного числа случаев выращивают в почве для горшечных культур при нормальных условиях до тех пор, пока они не достигали фазы выметывания. Затем их переносят в "сухую" зону, где приостанавливают полив. Зонды измерения влажности вставляют в случайным образом выбранные горшки для наблюдения за содержанием воды в почве (SWC). Когда SWC опускается ниже определенного порога, растения автоматически непрерывно повторно поливают до тех пор, пока снова не достигают нормального уровня. Растения затем повторно переносят снова в нормальные условия. Остальное культивирование (созревание растения, сбор семян) является таким же, как для растений, не выращивавшихся в условиях абиотического стресса. Параметры роста и урожайности регистрируются, как подробно описано для роста при нормальных условиях.
Скрининг при солевом стрессе
Растения выращивают на субстрате, полученном из кокосовых волокон и Argex (соотношение 3 к 1). Используется стандартный питательный раствор в течение первых двух недель после пересадки ростков в теплицу. После первых двух недель добавляется 25 мМ соли (NaCl) в питательный раствор до тех пор, пока растения не соберут. Параметры роста и урожайности регистрируются, как подробно описано для роста при нормальных условиях.
Скрининг при сниженной доступности питательных веществ (азота)
Растения из шести случаев (семена Т2) выращивают в почве для горшечных культур при нормальных условиях за исключением питательного раствора. Горшки поливают от пересадки до созревания особым питательным раствором, имеющим сниженное содержание азота (N), обычно в 7-8 раз меньше. Остальное культивирование (созревание растения, сбор семян) является таким же, как для растений, не выращивавшихся при абиотическом стрессе. Параметры роста и урожайности регистрируются, как подробно описано для роста при нормальных условиях.