Результат интеллектуальной деятельности: СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ
Вид РИД
Изобретение
ПРИОРИТЕТ
По данной заявке испрашивается приоритет согласно 35 U.S.C. §119 по заявке США с серийным № 61/051866, поданной 9 мая 2008 года, содержание которой включено в настоящий документ в качестве ссылки в полном объеме.
ИНФОРМАЦИЯ О ГРАНТЕ
Это изобретение было осуществлено при поддержке правительства грантом под номером NIH 1 PO1 AT004511-02, выданным National Institute of Health. Правительство США имеет определенные права на это изобретение.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к применению экстракта виноградных косточек для профилактики и лечения нейродегенеративного заболевания. В частности, настоящее изобретение относится к способу лечения пациента, у которого диагностировано нейродегенеративное заболевание или который имеет риск его развития, путем введения фармацевтической композиции, содержащей экстракт виноградных косточек или одно или несколько полученных из него соединений пациенту в эффективном количестве для уменьшения накопления, агрегации и/или отложения амилоида-бета или его олигомеров и/или для уменьшения неправильного сворачивания, накопления и/или агрегации тау-белков или других белков.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Нейродегенеративные заболевания ассоциированы с состояниями, при которых нейрональные клетки повреждаются, утрачивают функцию и часто погибают. Поскольку они, как правило, являются прогрессирующими, последствия нейродегенеративных заболеваний часто являются тяжелыми. Пациенты с нейродегенеративным заболеванием могут страдать тяжелым нарушением познавательных или двигательных навыков.
В результате качество их жизни и продолжительность жизни могут значительно снижаться. У человека эти заболевания включают, но не ограничиваются ими, болезнь Альцгеймера, болезнь Паркинсона, боковой амиотрофический склероз, болезнь Гентингтона, лобно-височную деменцию и кортико-базальную дегенерацию, среди прочих.
Болезнь Паркинсона представляет собой прогрессирующее нарушение, которое поражает нейроны головного мозга, контролирующие движение мышц. Эти нейроны продуцируют дофамин, который является важным химическим веществом для передачи сигналов между клетками, обеспечивающих движение тела. Таким образом, утрата этих нейронов ведет к двигательным нарушениям, таким как тремор и нарушение речи, которые, как правило, проявляются у пациентов с болезнью Паркинсона. Согласно National Institute of Neurological Disorders and Stroke, по меньшей мере 1 миллион человек страдает болезнью Паркинсона, и каждый год в США описывают приблизительно 50000 новых случаев.
В патологию болезни Паркинсона вовлечен высоко консервативный пресинаптический белок, α-синуклеин. Полагают, что конформационные изменения α-синуклеина приводят к скоплению белка и образованию фибрилл, характерным для этого заболевания. Смотрите патент США № 7045290 (выданный Lindquist et al). Современные способы лечения включают леводопу и агонисты дофамина. Однако эти лекарственные средства дают только временное смягчение симптомов и имеют тяжелые побочные эффекты, если их используют в больших количествах. Депренил, ингибитор моноаминоксидазы B, является первым лекарственным средством, предложенным для этиотропной терапии болезни Паркинсона путем смягчения симптомов и замедления прогрессирования заболевания, однако терапевтическая эффективность депренила является спорной. Смотрите патент США № 6417177 (выданный Nelson).
Болезнь Гентингтона (HD) представляет собой генетическое неврологическое нарушение с симптомами аномальных движений тела и сниженных умственных способностей. Болезнь Гентингтона вызывается экспансией тринуклеотидных повторов в гене хантингтина (htt), с которого, в свою очередь, продуцируется мутантная форма белка htt, имеющая патологическую экспансию полиглутаминовой (PolyQ) последовательности. Мутантный белок htt неправильно сворачивается и образует агрегаты в головном мозге и других пораженных тканях, что приводит к гибели нейронов (Wolfgang et al., Proc Nat Acad Sci 2005; 102: 11563-11568). Большинство лекарственных средств, используемых для лечения болезни Гентингтона, имеют побочные эффекты, такие как усталость, беспокойство или повышенная возбудимость.
Болезнь Альцгеймера (AD) представляет собой прогрессирующее заболевание головного мозга, общеизвестное как старческая деменция. Более 4,5 миллиона имеют диагноз AD, и ожидается, что это количество утроится в последующие 40-50 лет (Lyketsos et al., Am J Geriatr Psychiatry 2006; 14(7): 561-72).
Полагают, что патофизиологическая причина AD состоит, частично, в нарушении процессинга или в мутации амилоидного белка-предшественника. Белок с нарушением процессинга может образовывать увеличенное количество амилоидных бета-пептидов (Aβ) или их вариантных форм. Накопление Aβ ведет к отложению нерастворимых бляшек Aβ, и, в конечном итоге, к синаптической недостаточности, повреждению нейронов, образованию клубков гиперфосфорилированного тау-белка и апоптотической гибели нейронов. Повреждение или гибель нейронов приводят к потере множества нейротрансмиттеров, которая, в свою очередь, приводит к возникновению когнитивных и функциональных симптомов заболевания.
Доступные в настоящее время лекарственные средства обеспечивают относительно небольшую симптоматическую пользу для некоторых пациентов и не замедляют прогрессирование заболевания. Таким образом, существуют различные симптоматические стратегии лечения или профилактики AD. Например, было выявлено, что AD ассоциирована с воспалением головного мозга, и, таким образом, для снижения риска развития AD используют нестероидные противовоспалительные лекарственные средства, такие как ибупрофен и индометацин. Однако эти лекарственные средства имеют долгосрочный риск желудочно-кишечного кровотечения и заболевания почек, и они ассоциированы с редкой сердечно-сосудистой токсичностью. Связь свободных радикалов кислорода с AD также привела к возможности терапии антиоксидантами. Как American Psychiatric Association, так и American Academy of Neurology Treatment Guidelines for AD, рекомендовали высокие дозы витамина E в качестве дополнительного средства для лечения. Однако эта рекомендация ограничивается недавними открытиями, что терапия витамином E не замедляет прогрессирование мягкого когнитивного нарушения, ассоциированного с AD, и что витамин E в очень высоких дозах повышает смертность пожилых людей. Смотрите Lyketsos et al., 2006, выше.
Другим вариантом лечения AD является снижение естественной деградации ацетилхолина ферментом, известным как ацетилхолинэстераза (AChE). Ингибирование AChE приводит к повышению уровней ацетилхолина. Food and Drug Administration США одобрили для лечения AD четыре лекарственных средства на основе ингибитора холинэстеразы: такрин, донепезил, ривастигмин и галантамин. Кратковременные (вплоть до 6 месяцев) клинические испытания ингибиторов холинэстеразы показали, что эти лекарственные средства приводили к улучшению или замедляли снижение когнитивной функции, ассоциированное с AD, однако результаты клинических испытаний их длительной пользы не являются окончательными. При очень мягкой или более тяжелой AD, польза ингибиторов холинэстеразы является менее существенной. Смотрите Lyketsos et al., 2006, выше.
Разрабатывается лечение AD, которое нацелено на удаление Aβ из головного мозга. Например, в патенте США № 7262223 (выданном Kong et al.) описано применение соединений амидина для лечения связанных с амилоидом заболеваний; в патенте США № 7279501 (выданном Kim) описано применение природных соединений, выделенных из растений (например, куркумы, гинкго двухлопастного и имбиря) и их синтетических химических аналогов для лечения индуцируемого Aβ заболевания. Последние данные указывают на то, что умеренное употребление красного вина может снизить заболеваемость AD и может уменьшить когнитивное нарушение по типу AD и амилоидную нейропатологию (Dartigues et al, Therapie 1993; 48: 185-187; Dorozynski, BMJ 1997; 314: 997; Luchsinger et al, J Am Geriatr Soc 2004; 52: 540-546). Накопление растворимых внеклеточных высокомолекулярных (HMW) олигомерных молекул Aβ в головном мозге считается главным фактором риска возникновения и прогрессирования когнитивного нарушения (Klyubin et al, Nat Med 2005; 11: 556-561; Selkoe, J Alzheimer's Dis 2001; 3: 75-80). Было предположено, что происходящие из винограда полифенольные соединения могут ингибировать олигомеризацию Aβ in vitro (Porat et al, Chem Biol Drug Des 2006; 67: 27-37). Однако до сегодняшнего дня исследования были ограничены тестированием in vitro.
Многие типы нейродегенеративных заболеваний связаны с аномальным сворачиванием белка, накоплением, агрегацией и/или отложением белков. Например, существует два типа аномальных отложений белка в головном мозге пациентов с болезнью Альцгеймера. Существуют амилоидные бляшки, состоящие из амилоидных бета-пептидов, которые депонируются внутриклеточно в паренхиме головного мозга и вокруг стенок сосудов головного мозга, и нейрофибриллярные клубки, которые состоят из агрегатов гиперфосфорилированного тау-белка, расположенного в цитоплазме дегенерирующих нейронов. У пациентов с болезнью Паркинсона в цитоплазме нейронов черной субстанции наблюдают тельца Леви. Основными составляющими телец Леви являются фрагменты белка, называющегося α-синуклеином. Типичным признаком головного мозга пациентов с болезнью Гентингтона являются внутриядерные отложения богатого полиглутамином варианта мутантного белка хантингтина. Пациенты с наследственным боковым амиотрофическим склерозом имеют агрегаты, главным образом, состоящие из супероксиддисмутазы, в телах клеток и аксонах двигательных нейронов. Кроме того, различные формы трансмиссивной губчатой энцефалопатии характеризуются скоплениями устойчивых к протеазе агрегатов прионного белка.
Данные биохимических, генетических и нейропатологических исследований указывают на активное вовлечение неправильного сворачивания и/или агрегации белка в патологию нейродегенеративных заболеваний. Например, присутствие аномальных агрегатов, как правило, встречается в областях головного мозга, наиболее поврежденных заболеванием. Мутации в гене, кодирующем неправильно свернутый белок, вызывают врожденные формы заболевания, которые, как правило, имеют более раннее начало и более тяжелый фенотип, чем спорадические формы. У трансгенных животных, у которых экспрессируется мутантный ген человека для неправильно свернутого белка, развиваются некоторые из типичных нейропатологических и клинических признаков заболевания человека. Также неправильно свернутые агрегаты белка, продуцируемые in vitro, являются нейротоксичными и индуцируют гибель клеток.
Таупатии представляют собой семейство нейродегенеративных заболеваний, в которые вовлечено нарушение функции тау-белков (семейство близкородственных внутриклеточных ассоциированных с микротрубочками белков). Эти нейродегенеративные заболевания включают, например, болезнь Альцгеймера, прогрессирующий супрануклеарный паралич, кортикобазальную дегенерацию, болезнь аргирофильных зерен, болезнь Пика, а также другие заболевания. Общими признаками таупатий является аномальное гиперфосфорилирование тау и скопления тау в устойчивых к детергенту внутриклеточных включениях, известных как нейрофибриллярные клубки (NFT) среди нейронов или глиальных клеток головного мозга. Аномально гиперфосфорилированные тау-белки легко диссоциируют от микротрубочек и агрегируют в олигомерные парные спиралевидные тау-фибриллы, которые в конечном итоге откладываются в качестве внутриклеточных NFT. (Mi, K. et al, Curr Alzheimer Res 2006; 3: 449-463). Образование олигомеров служит в качестве участков ядрообразования, которые объединяют дополнительный гиперфосфорилированный тау, а также нормальный нефосфорилированный тау, в фибриллярные агрегаты. (Sorrentino et al, Neurol Sci 2007; 28: 63-71). Таким образом, теория тау-опосредуемой нейродегенерации основана на модели "токсического приобретения функции", в которой аномально фосфорилированные тау-белки стимулируют удаление как гиперфосфорилированных, так и нормальных белков тау из микротрубочек. Это приводит к нестабильности микротрубочек и изменениям опосредуемых микротрубочками процессов, таких как аксональный транспорт, которые, в свою очередь, приводят к нарушению функции и снижению жизнеспособности нейрональных и глиальных клеток в головном мозге (Sorrentino et al., 2007, выше).
В публикации патентной заявки США № 2007/0122504 (выданной Moon et al.) описан процесс производства экстракта виноградных косточек и способы применения такого экстракта виноградных косточек для лечения нейродегенеративных заболеваний, включая AD. Экстракт получают путем (1) экстракции виноградных косточек в щелочном растворе, имеющем pH от 8 до 11, при предпочтительно 20-50°C с получением растворимого в щелочи вещества; (2) нейтрализации кислотного раствора для доведения до pH в диапазоне от 2 до 4, и центрифугирования полученного раствора и получения осажденного слоя; (3) добавления низшего спирта и получения слоя супернатанта, затем концентрирования слоя супернатанта; и (4) добавления неполярного растворителя и удаления растворимого в неполярном растворителе слоя с получением очищенной фракции и проведения повторяющейся очистки и лиофилизации для получения высушенного экстракта виноградных косточек. (смотрите Moon, абзацы [0030]-[0035]).
Вследствие распространенности нейродегенеративных заболеваний и отсутствия подтвержденных эффективных фармацевтических композиций или способов лечения симптомов, ассоциированных с нейродегенеративными заболеваниями, все еще существует необходимость в улучшенных фармацевтических композициях и способах их лечения и профилактики.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу лечения нейродегенеративного заболевания у индивида путем введения индивиду, нуждающемуся в этом, фармацевтической композиции, где фармацевтическая композиция содержит экстракт виноградных косточек или одно или несколько соединений, полученных из него. В определенных вариантах осуществления, фармацевтическая композиция также может содержать активный ингредиент, выбранный из группы, состоящей из антиоксиданта, ингибитора ацетилхолинэстеразы и их комбинаций. В конкретном варианте осуществления, экстракт виноградных косточек содержит менее чем приблизительно 12% по массе галлоилированного проантоцианидина в расчете на общую массу проантоцианидинов в экстракте.
В одном варианте осуществления фармацевтическую композицию вводят перорально. Пероральные дозированные формы включают порошок, таблетку, капсулу, диспергирующуюся в полости рта таблетку, мягкую капсулу, водное лекарственное средство, сироп, эликсир или саше. В другом варианте осуществления, фармацевтическую композицию вводят трансдермально. В другом варианте осуществления, фармацевтическую композицию вводят трансназально.
В конкретных вариантах осуществления индивидом является человек. Частота введения представляет собой введение раз в месяц, раз в две недели, раз в неделю или раз в сутки, и введение можно проводить в однократной дозе или разделенными дозами. Эффективное количество соединений экстракта виноградных косточек представляет собой дозировку от приблизительно 100 до приблизительно 1000 мг в сутки, предпочтительно от приблизительно 200 до приблизительно 600 мг в сутки.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1A-1F. На фиг.1 проиллюстрирован результат анализа компонентов продукта экстракта виноградных косточек (GSE), MegaNatural®-AZ (или MNG-AZ). На фиг.1A проиллюстрирована молекулярная структура типичного гетерополимерного проантоцианидина. На фиг.1B представлен анализ MNG-AZ посредством ВЭЖХ на нормальной фазе. На фиг.1C схематично представлен гомотетрамерный проантоцианидин, состоящий из галлата эпикатехина (левая панель), и полученный дегаллоилированный проантоцианидин вместе отделенной структурой галловой кислоты (правая панель). На фиг.1D показан процент галлоилированных проантоцианидинов (из всех проантоцианидинов) в MNG-AZ по сравнению с четырьмя другими коммерчески доступными препаратами GSE: MegaNatural®-Gold, торговое название GSE A, торговое название GSE B и торговое название GSE C (фиг.1D) (торговое название A представляет собой "Activin", GSE, полученный от San Joaquin Valley Concentrates; торговое название B представляет собой "Masquelie® OPC", GSE из Франции; и торговое название C представляет собой GSE от Indena S.p.A., Италия). На фиг.1E и 1F представлены уровни галлоилированного проантоцианидина в MNG-AZ и некоторые другие коммерчески доступные GSE.
Фиг.2A-2D. На фиг.2 представлен эффект GSE MNG-AZ на превращение Aβ-пептидов в их растворимые олигомерные формы in vitro. На фиг.2A и 2B представлен полиакриламидный гель-электрофорез (SDS-PAGE) с додецилсульфатом натрия продукта инкубации Aβ1-42 (2A) и Aβ1-40 (2B) с различными количествами GSE MNG-AZ. На фиг.2C и 2D продемонстрирован результат SDS-PAGE Aβ1-42 (2C) и Aβ1-40 (2D) в присутствии или отсутствие GSE MNG-AZ после проведения химической реакции фотоиндуцированного поперечного сшивания немодифицированных белков (PICUP).
Фиг.3A-3D. На фиг.3 представлен эффект ингибирования MNG-AZ в отношении Aβ, исследованный спектроскопией кругового дихроизма (CD). На фиг.3A и 3C представлены спектры CD необработанных Aβ1-40 и Aβ1-42, соответственно; на фиг.3B и 3D представлены спектры CD Aβ1-40 и Aβ1-42, соответственно, обработанных MNG-AZ. Спектры получали сразу после начала периода инкубации 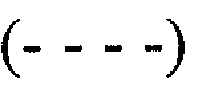 и через 2
и через 2 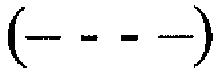 , 3
, 3 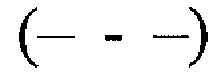 , 6
, 6 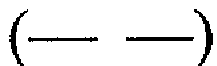 и 7
и 7 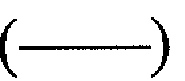 суток. Спектры, представленные для каждого момента времени, соответствуют спектрам, полученным в ходе каждого из 3 независимых экспериментов.
суток. Спектры, представленные для каждого момента времени, соответствуют спектрам, полученным в ходе каждого из 3 независимых экспериментов.
Фиг.4A-4D. На фиг.4 представлен эффект MNG-AZ в отношении ингибирования Aβ, при исследовании в анализе связывания ThT. На фиг.4A и 4C представлены спектры флуоресценции необработанных Aβ1-40 и Aβ1-42, при различных концентрациях контрольного соединения Med1, соответственно; на фиг.4B и 4D представлены спектры флуоресценции Aβ1-40 и Aβ1-42, соответственно, обработанных MNG-AZ. На каждой из фиг.4A-4D концентрации Med1 (или MNG-AZ) представляют собой 0 (♦), 5 (■) или 25 (▲) мкМ.
Фиг.5A-5F. На фиг.5 проиллюстрирован эффект MNG-AZ в отношении ингибирования Aβ при исследовании электронной микроскопией. На фиг.5A и 5B представлена иллюстративная морфология необработанных Aβ1-40 и Aβ1-42, соответственно. На фиг.5C и 5E представлена морфология Aβ1-40 в присутствии более низких (5 мкМ) и более высоких (25 мкМ) концентраций MNG-AZ, соответственно. На фиг.5D и 5F представлена морфология Aβ1-42, в присутствии более низкой (5 мкМ) и более высокой (25 мкМ) концентрации MNG-AZ, соответственно.
Фиг.6A-6D. На фиг.6 проиллюстрирован эффект MNG-AZ в отношении ингибирования Aβ при исследовании по метаболизму МТТ. На фиг.6A (и 6C) представлена токсичность низкомолекулярного Aβ1-40 (и Aβ1-42) и эффект MNG-AZ в отношении снижения токсичности. На фиг.6B (и 6D) представлена токсичность агрегации Aβ1-40 (и агрегации Aβ1-42) и эффект MNG-AZ в отношении снижения токсичности.
Фиг.7A-7G. На фиг.7 проиллюстрированы эффекты GSE MNG-AZ на нейропатологию у мышей Tg2576. На фиг.7A-7B представлены эффекты GSE MNG-AZ на массу тела (7A) или употребление жидкости (7B). На фиг.7C-7D представлена оценка содержания растворимого внеклеточного пептида HMW-Aβ в головном мозге мышей Tg2576. На фиг.7E представлена оценка концентраций пептида Aβ1-42 и Aβ1-40 в головном мозге у мышей, которым вводили GSE MNG-AZ, и контрольных мышей. На фиг.7F представлена стереологическая оценка нагрузки Aβ-амилоидными бляшками коры головного мозга и парагиппокампальной извилины у мышей, которым вводили GSE MNG-AZ, и у контрольных мышей. На фиг.7G проиллюстрирована оценка концентраций пептидов Aβ1-42 и Aβ1-40 в головном мозге мышей, которым вводили MegaNatural®-Gold, и у контрольных мышей.
Фиг.8A-8F. На фиг.8 представлены результаты различных экспериментов по установлению потенциальных механизмов благоприятных эффектов GSE MNG-AZ. На фиг.8A проиллюстрирован вестерн-анализ тотальной экспрессии APP у мышей Tg2576, которым вводили MNG-AZ в течение приблизительно 5 месяцев. На фиг.8B проиллюстрирована оценка активности α-, β- и γ-секретазы. На фиг.8C проиллюстрирован вестерн-анализ экспрессии растворимого APPα и APPβ у мышей Tg2576, которым вводили GSE MNG-AZ, относительно контрольной группы. На фиг.8D и 8E представлена экспрессия деградирующего Aβ фермента неприлизина и деградирующего инсулин фермента в головном мозге мышей Tg2576, которым вводили GSE MNG-AZ, относительно контрольных мышей. На фиг.8F проиллюстрирована оценка содержания в сыворотке Aβ1-40 (левая панель) и Aβ1-42 (правая панель) посредством твердофазного иммуноферментного анализа (ELISA).
Фиг.9A-9C. На фиг.9 проиллюстрировано ослабление когнитивного нарушения у мышей Tg2576, которым вводили GSE MNG-AZ. На фиг.9A и 9B представлено влияние GSE MNG-AZ на связанную с Aβ пространственную память у мышей Tg2576 при определении с помощью тестов в водном лабиринте Морриса. На фиг.9C представлена оценка содержания растворимого внеклеточного высокомолекулярного пептида Aβ в головном мозге мышей Tg2576.
Фиг.10A-10B. На фиг.10A и 10B проиллюстрирован эффект введения GSE MNG-AZ на когнитивную функцию у мышей дикого типа, совпадающих по линии, возрасту и полу, при определении в тестах с водным лабиринтом Морриса.
Фиг.11A-11B. На фиг.11 проиллюстрирована кинетика агрегации тау-пептида в отсутствие или присутствии GSE MNG-AZ. На фиг.11A представлены зависимые от времени спектры флуоресценции ThS для агрегированного тау при различных концентрациях MNG-AZ; на фиг.11B представлено максимальное накопление агрегатов тау при различных концентрациях MNG-AZ.
Фиг.12A-12B. На фиг.12 проиллюстрирован эффект GSE MNG-AZ на диссоциацию заранее сформированных агрегатов тау-пептида. На фиг.12A представлены зависимые от времени спектры флуоресценции ThS для агрегированного тау при различных концентрациях MNG-AZ; На фиг.12B представлены скорости диссоциации агрегатов тау как функция концентрации MNG-AZ.
Фиг.13A-13D. На фиг.13 проиллюстрированы преимущества GSE MNG-AZ в модели с Drosophila. На фиг.13A представлен результат развития глаз у Drosophila в отсутствие GSE; на фиг.13B представлен результат развития глаз у Drosophila в присутствии GSE; на фиг.13C представлена визуальная оценка глаз самцов Drosophila в иллюстративном эксперименте (в присутствии и в отсутствие GSE); на фиг.13D представлено количество отсутствующих глаз в том же испытании, что и на фиг.13C.
Фиг.14. На фиг.14 проиллюстрирован процент выживаемости на протяжении данного количества суток в модели болезни Гентингтона у Drosophila. Пустые круги соответствуют результатам для группы, в которой вводили экстракт виноградных косточек, и а закрашенные круги соответствуют результатам для контрольной группы.
Фиг.15A-15C. На фиг.15 проиллюстрирован эффект GSE MNG-AZ на олигомеризацию Aβ. На фиг.15A представлены результаты для Aβ1-40; на фиг.15B представлены результаты для Aβ1-42; на фиг.15C представлены результаты для глутатион-S-трансферазы. Дорожки 1: маркеры молекулярной массы; дорожки 2: белок отдельно, без поперечного сшивания; дорожки 3: белок отдельно; дорожки 4: белок вместе с Med1 (25 мкМ); дорожки 5: белок вместе с Med1 (250 мкМ); дорожки 6: белок вместе с MNG-AZ (25 мкМ); дорожки 7: белок вместе с MNG-AZ (250 мкМ). Гель соответствует каждому из трех независимых экспериментов.
Фиг.16. На фиг.16 проиллюстрирован эффект GSE MNG-AZ на агрегацию тау-пептида с использованием анализа PICUP. Гель демонстрирует иллюстративный анализ 25 мкМ тау-пептида, поперечно-сшитого в присутствии (дорожки 2, 4, 6, 8) или отсутствие (дорожки 1, 3, 5, 7) равного количества моль (25 мкМ) GSE. Вследствие ожидаемого неэффективного окрашивания небольших пептидов, мономерные тау-пептиды не поддаются детекции в этом эксперименте. Полосы размером ~2,1 и ~3,5 кДа соответствуют, соответственно, тримерным и пентамерным агрегатам тау-пептидов. CTR: не поперечно-сшитый тау-пептид; дорожки 1-8: тау-пептид с персульфатом аммония (APS) и 1× (дорожки 1, 2), 2× (дорожки 3, 4), 3× (дорожки 5, 6) и 4× (дорожки 7, 8) Ru(Bpy).
Фиг.17A и 17B. На фиг.17 проиллюстрирован эффект GSE MNG-AZ на агрегацию тау-пептида с использованием спектроскопии кругового дихроизма. Агрегация тау-пептида в отсутствие MNG-AZ представлена на фиг.17A, в то время как агрегация тау-пептида в присутствии MNG-AZ в молярном отношении 1:1 относительно тау-пептидов представлена на фиг.17B. Кривые, обозначенные с помощью легенд d0, d1, d2 и d3, на фиг.17A и 17B соответствуют спектрам, полученным на 0, 1, 2, 3 сутки в процессе инкубации синтетического тау-пептида (при 37°C), соответственно. Стрелки указывают на спектры, характерные для упорядоченных конформеров.
Фиг.18A и 18B. На фиг.18 проиллюстрирован эффект GSE MNG-AZ на морфологию тау-фибрилл с использованием электронной микроскопии. Морфология тау-фибрилл в отсутствие MNG-AZ представлена на фиг.18A; морфология тау-фибрилл в присутствии MNG-AZ представлена на фиг.18B. Планками масштаба показано 100 нм.
Фиг.19A-19D. На фиг.19 проиллюстрирован эффект GSE MNG-AZ на ультраструктурные характеристики нативных парных спиралевидных фибрилл (PHF), выделенных из образцов головного мозга при AD. На фиг.19A представлена электронная микрофотография очищенных PHF в отсутствие GSE MNG-AZ; на фиг.19B и 19C представлены электронные микрофотографии очищенных PHF в присутствии 100 мкМ GSE MNG-AZ в течение 5 с (фиг.19B) или 1 ч (фиг.19C). На фиг.19A и 19C электронноплотные частицы соответствуют мечению pSer214tau (стрелки на C). На фиг.19D представлен количественный анализ PHF при обработке GSE как функция времени обработки (от 5 до 60 мин), где столбец диаграммы соответствует средней максимальной ширине со стандартным отклонением; измеренные количества PHF представлены в скобках. Статистический анализ проводили посредством одностороннего ANOVA (P<0,0001), с последующим анализом с помощью критерия множественных сравнений Бонферрони, **p<0,001 по сравнению с необработанными PHF (время 0).
Фиг.20A-20D. На фиг.20 проиллюстрирован эффект GSE MNG-AZ на расщепление трипсином PHF. На фиг.20A представлена электронная микрофотография нативных PHF, выделенных из головного мозга при AD, не инкубированных с трипсином; на фиг.20B представлена электронная микрофотография PHF, предварительно обработанных MNG-AZ (100 мкМ, 1 ч), не инкубированных с трипсином; на фиг.20C представлена электронная микрофотография нативных PHF, инкубированных с трипсином (1 мкг/мл, 10 мин); на фиг.20D представлена электронная микрофотография PHF, предварительно инкубированных с MNG-AZ (100 мкМ, 1 ч) и инкубированных с трипсином (1 мкг/мл, 10 мин).
Фиг.21A-21G. На фиг.21 проиллюстрирован эффект GSE MNG-AZ на аномальные фенотипы глаз в модели Drosophila с мутантным тау. На фиг.21A и 21D представлены иллюстративные фенотипы глаз у мух дикого типа; на фиг.21B и 21E представлены глаза мух с мутантным тау R406W в отсутствие введения GSE; на фиг.21C и 21F представлены глаза мух с мутантным тау R406W, которым вводили MNG-AZ; на фиг.21G представлен количественный анализ морфологии взрослого глаза, с использованием четырехточечной системы оценки (где 0=нет глаза и 4=нормальный глаз) у самцов и самок мух в трех независимых испытаниях. Указано количество оцененных мух на испытание. Столбиковые диаграммы соответствуют среднему значению +SEM.
Фиг.22A-22D. На фиг.22 представлена схема оценки с использованием анализа вытягивания задних конечностей в модели таупатии на трансгенных мышах JNPL3. Природную тенденцию животных к вытягиванию их задних конечностей латерально, когда их подвешивают в перевернутом состоянии за их хвосты, оценивают согласно четырехточечной системе оценки: 4=нормальная функция (фиг.22A), 3=мягкое нарушение (фиг.22B), 2=умеренное нарушение (фиг.22C) и 1=тяжелое нарушение (фиг.22D).
Фиг.23A и 23B. На фиг.23 проиллюстрирован эффект введения GSE в модели таупатии на трансгенных мышах JNPL3. На фиг.23A представлено двигательное нарушение у мышей JNPL3 в возрасте 5 месяцев и 13 месяцев, соответственно, при введении GSE по сравнению с мышами, которым не вводили GSE. На фиг.23B представлено сравнение уровня смертности среди мышей JNPL3, которым вводили GSE и которым не вводили GSE, где линейные графики соответствуют % выживанию с течением времени.
Фиг.24A и 24B. На фиг.24 проиллюстрирован эффект GSE MNG-AZ в отношении уменьшения агрегата белка htt с использованием флуоресцентной микроскопии. На фиг.24A представлены изображения контрольных клеток (Ctrl), обработанных носителем, и клеток, обработанных 12,5 мкМ и 25 мкМ GSE после индукции муристероном A. На фиг.24B представлен вестерн-блот анализ агрегации агрегатов слитого белка GFP-Htt в высокомолекулярные агрегаты в отсутствие (Ctrl) или в присутствии обработки 12,5 мкМ и 25 мкМ GSE (левая панель: вестерн-блот, исследованный с помощью антитела против GFP, для идентификации агрегации слитого белка GFP-Htt в более высокомолекулярные частицы; правая панель: денситометрический анализ вестерн-блота, демонстрирующий распределение белка GFP-Htt и более высокомолекулярных агрегатов htt).
Фиг.25A и 25B. На фиг.25 проиллюстрирован эффект введения GSE на двигательные нарушения в модели HD на Drosophila при оценке анализом подъема. На фиг.25A и 25B проиллюстрирован результат анализа подъема на 9 сутки и на 16 сутки, соответственно. В каждый день тестирования проводили три независимых испытания подъема. Столбиковые диаграммы соответствуют среднему значению +SEM для % мух, которые успешно выполнили задачу подъема. Статистический анализ проводили с помощью t-критерия Стьюдента, **p<0,001, путем сравнения группы введения GSE с группой без введения GSE.
Фиг.26. На фиг.26 проиллюстрирован процент выживания с течением данного количества суток в модели HD у Drosophila. Закрашенные перевернутые треугольники соответствуют результатам для группы, в которой вводили GSE MNG-AZ, а закрашенные ромбы соответствуют результатам для контрольной группы. Данные соответствуют результатам 4 независимых испытаний.
Фиг.27. На фиг.27 проиллюстрирован эффект обработки GSE на двигательное нарушение в модели HD на мышах, оцененный с использованием анализа на вращающемся стержне в возрасте, составляющем различное количество недель. Данные соответствуют результатам, полученным для 3 независимых испытаний.
Фиг.28. На фиг.28 проиллюстрирован эффект введения GSE на смертность в модели HD на мышах. Линейные графики соответствуют % выживанию с течением времени.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение преимущественно относится к способу снижения неправильного сворачивания, накопления, агрегация или отложения белков Aβ, олигомерного Aβ, тау или других белков, ассоциированных с нейродегенеративным заболеванием. Способ вовлекает введение эффективного количества фармацевтической композиции, содержащей экстракт виноградных косточек или одно или несколько соединений, полученных из него. Эти и другие аспекты изобретения подробно описаны в описании и примерах, предоставленных ниже.
Настоящее изобретение основано на открытии, что соединения из экстракта виноградных косточек функционируют в качестве эффективных ингибиторов неправильного сворачивания, накопления, агрегации и/или отложения Aβ, тау-белка и других белков, которые ассоциированы с различными нейродегенеративными заболеваниями. Конкретно, изобретение основано, частично, на открытии, что конкретные типы экстракта виноградных косточек снижали или ингибировали (1) образование олигомеров синтетических Aβ1-40 (Aβ40) и Aβ1-42 (Aβ42) in vitro; (2) количество олигомерного Aβ в головном мозге мышей Tg2576 (трансгенных мышей, экспрессирующих мутантный белок амилоидного предшественника и проявляющих когнитивное нарушение по типу AD) и приводили к заметному улучшению при утрате когнитивной функции или ее замедлению у мышей Tg2576 (по сравнению с мышами без введения); (3) инициацию ядрообразования, ведущего к агрегации тау в структуры, характеризующиеся парными спиралевидными фибриллами, встречающимися при различных таупатиях, а также стабильность агрегатов тау, in vitro; (4) вредоносные эффекты белка в фенотипе трансгенных R406W Drosophila, а также вредоносные эффекты тау-белка в модели на трансгенных мышах JNPL3, in vivo, (5) агрегацию содержащих полиглутамин белковых частиц htt in vitro; (6) вредоносные эффекты мутантного белка htt в трансгенном фенотипе elav>Q93httexonl Drosophila, а также вредоносные эффекты мутантного белка htt в модели трансгенных мышей R6/2, in vivo. Эти наблюдения неожиданно демонстрируют, что экстракт виноградных косточек или соединения, полученные из них, можно использовать для снижения образования амилоида, htt, и связанной с тау нейропатологии.
Таким образом, настоящее изобретение относится к фармацевтическим композициям, содержащим экстракт виноградных косточек или одно или несколько соединений, полученных из них, и к способам применения таких фармацевтических композиций для лечения или профилактики нейропатологических признаков нейродегенеративного заболевания, таких как нейродегенерация, клеточная токсичность, когнитивное нарушение или повреждение и двигательное нарушение. Предпочтительно экстракт виноградных косточек характеризуется наличием менее чем приблизительно 12% по массе галлоилированных проантоцианидинов в расчете на общее количество проантоцианидинов.
Термины, использованные в данном описании, как правило, обладают значениями, общепринятыми для них в данной области, в контексте этого изобретения и в конкретном контексте, где каждый термин используется. Некоторые термины определены ниже для обеспечения дополнительного руководства для описания композиций и способов по изобретению.
Определения
Термин "деменция" относится к клиническому синдрому, ассоциированному с общим когнитивным ухудшением памяти и других областей познавания.
Термин "дегенеративное заболевание" относится к заболеванию, в котором функция или структура поврежденных тканей или органов постепенно ухудшается с течением времени, в противоположность инфекционным заболеваниям.
Термин "нейродегенеративное заболевание" относится к состоянию или нарушению, при котором нейрональные клетки утрачиваются вследствие гибели клеток.
Термин "болезнь Альцгеймера" (или "старческая деменция") относится к умственному нарушению, ассоциированному с конкретным дегенеративным заболеванием головного мозга, которое характеризуется старческими бляшками, клубками нейритов и прогрессирующей утратой нейронов.
Термин "болезнь Паркинсона" представляет собой хроническое и прогрессирующее дегенеративное нарушение центральной нервной системы, которое часто нарушает двигательные навыки и речь. Болезнь Паркинсона относится к группе состояний, называемых двигательными нарушениями, и характеризуется ригидностью мышц, тремором, замедлением движения тела и, в крайних случаях, утратой движения тела.
Термин "болезнь Гентингтона" относится к наследственному неврологическому нарушению, вызываемому экспансией тринуклеотидных повторов в гене, кодирующем белок хантингтин. Симптомы болезни Гентингтона включают аномальные движения тела и отсутствие координации.
Термин "таупатия" относится к семейству нейродегенеративных заболеваний, которые вовлекают нарушение функции тау-белков (семейство близкородственных внутриклеточных ассоциированных с микротрубочками белков). Эти нейродегенеративные заболевания (таупатии) включают, например, болезнь Альцгеймера, прогрессирующий супрануклеарный паралич, кортикобазальную Дегенерацию, болезнь аргирофильных гранул, болезнь Пика и семейную лобно-височную деменцию.
Термин "амилоид-бета" (Aβ) относится к пептиду, продуцируемому расщеплением белка-предшественника амилоида-бета (APP), накопление и отложение которого формирует бляшки в головном мозге индивида. Наиболее распространенными изоформами Aβ являются Aβ1-40 (Aβ40) и Aβ1-42 (Aβ42). Выражение "олигомер Aβ" относится к пептиду, имеющему более одного элемента Aβ, связанного химическими связями, или множество пептидов Aβ, связанных химическими связями и/или ассоциированных физическими силами. Термин "олигомеризация" относится к комбинированию или сборке множества меньших химических или биологических молекул, таких как Aβ, в более крупный агрегат путем химического связывания и/или физической ассоциации.
Термин "снижать" относится к уменьшению или сокращению количества или концентрации химического или биологического вещества, или к замедлению или реверсии протекающего химического или физического процесса.
Термин "накопление" относится к увеличению концентрации или количества химического или биологического вещества, такого как пептид, в определенной области или пространстве.
Термин "агрегация" относится к комбинированию или сборке множества меньших химических или биологических молекул, или их агрегата, в более крупный агрегат путем химического связывания и/или физической ассоциации.
Термин "отложение" относится к прикреплению химического или биологического вещества к биологической поверхности, такой как мембрана клетки или стенка кровеносного сосуда.
Термин "полифенол" или "полифенольное соединение" относится к соединению, характеризующемуся наличием более одной группы фенола на молекулу.
Термины "терапевтически эффективная доза" или "терапевтически эффективное количество", или "эффективное количество" относятся к количествам экстракта виноградных косточек или соединений, содержащихся в них, которые являются достаточными для того, чтобы вызывать терапевтический ответ. Терапевтический ответ может представлять собой любой ответ, который пользователь (например, врач) признает в качестве эффективного ответа на лечение, например, путем оценки симптомов и заменяющих их клинических маркеров. Таким образом, терапевтический ответ, как правило, представляет собой смягчение одного или нескольких симптомов заболевания или нарушения.
Выражение "фармацевтически приемлемый" относится к молекулам и композициям, которые являются физиологически переносимыми и, как правило, не вызывают неблагоприятных реакций при введении человеку. Предпочтительно, как используют в настоящем документе, термин "фармацевтически приемлемый" означает "одобренный органом регулирования федерального правительства или правительства штата" или "приведенный в фармакопее США или других общепризнанных фармакопеях для применения у животных и, более конкретно, у человека".
Выражение "фармацевтически приемлемые соли" относится к производным соединений, модифицированным путем получения их кислых или основных солей. Примеры фармацевтически приемлемых солей включают соли минеральных или органических кислот с основными остатками, такими как амины; и щелочами, или органические соли кислотных остатков, таких как карбоновые кислоты. Фармацевтически приемлемые соли включают общепринятые нетоксические соли или четвертичные соли аммония исходного соединения, образованные, например, из нетоксических неорганических или органических кислот. Такие общепринятые нетоксичные соли включают соли, образованные из неорганических кислот, таких как хлористоводородная, бромистоводородная, серная, сульфаминовая, фосфорная и азотная кислоты; и соли, полученные из органических кислот, таких как уксусная, пропионовая, янтарная, гликолевая, стеариновая, молочная, яблочная, виннокаменная, лимонная, аскорбиновая, пальмоевая, малеиновая, гидроксималеиновая, фенилуксусная, глутаминовая, бензойная, салициловая, сульфанильная, 2-ацетоксибензойная, фумаровая, толуолсульфоновая, метансульфоновая, этандисульфоновая, щавелевая и изетионовая кислота. Фармацевтически приемлемые соли можно синтезировать из исходного соединения, которое содержит основную или кислотную группу, общепринятыми химическими способами. Как правило, такие соли можно получать реакцией форм свободной кислоты или основания этих соединений со стехиометрическим количеством соответствующего основания или кислоты в водном или в органическом растворителе, или в их смеси.
Термины "носитель" или "фармацевтический носитель" относятся к разбавителю, адъюванту, эксципиенту или носителю, с которыми вводят соединение. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла. Предпочтительно в качестве носителей используют воду или водные солевые растворы и водные растворы декстрозы и глицерина. Пригодные фармацевтические носители описаны в "Remington's Pharmaceutical Sciences", E. W. Martin, 18 издание или другие издания.
Термин "антиоксидант" относится к серии химических веществ, способных ингибировать или нейтрализовывать вредоносные свободные радикалы в организме индивида.
Термин "индивид" включает живые организмы, в которых может происходить неправильное сворачивание, накопление, агрегация или отложение амилоида-бета, олигомера амилоида-бета, тау-белка или других белков. Термин "млекопитающее" относится к любому организму класса Млекопитающих высших позвоночных, который кормит свое потомство молоком, секретируемым молочными железами, например к мыши, крысе, кролику, собаке, кошке, и, в частности, к человеку. Термин "человек" относится к представителю вида Homo Sapiens. Термин "пациент" относится к человеку, которому проводят лечение композициями в соответствии с настоящим изобретением.
Термин "лечить" относится к введению композиции по настоящему изобретению индивиду для ослабления, замедления прогрессирования, отсрочивания или реверсии состояния и/или одного или нескольких симптомов, ассоциированных с нейродегенеративным заболеванием и/или неправильным сворачиванием, накоплением, агрегацией или отложением белков, включая, но не ограничиваясь ими, амилоид бета, олигомер амилоида бета, тау-белки, α-синуклеин и т.д.
Термин "предупреждать" относится к введению композиций по настоящему изобретению индивиду до возникновения состояния или симптома, ассоциированного с неправильным сворачиванием, накоплением, агрегацией или отложением амилоида-бета, олигомера амилоида-бета, тау-белков или других белков, чтобы предотвратить возникновение состояния или симптома.
Термин "снижение риска" возникновения состояния или симптома у индивида означает, что вероятность развития у индивида состояния или симптома является меньшей, чем вероятность их развития по сравнению с контрольным индивидом, например, когда индивиду вводят фармацевтическую композицию по изобретению, а контрольному индивиду не вводят ничего или вводят плацебо.
Термин "приблизительно" или "приближенно" означает нахождение в пределах приемлемого диапазона погрешностей для конкретного значения, который определяет специалист в данной области и который зависит, частично, от того, как значение измеряют или определяют, т.е. от ограничений системы измерения. Например, "приблизительно" может означать в пределах 3 или более 3 стандартных отклонений, согласно практике в данной области. Альтернативно "приблизительно" может означать диапазон вплоть до 20%, предпочтительно вплоть до 10%, более предпочтительно вплоть до 5% и более предпочтительно вплоть до 1% от данного значения. Альтернативно, в частности, в отношении биологических систем или процессов термин может означать "в пределах порядка величины", предпочтительно в пределах 5-кратного, и более предпочтительно в пределах 2-кратного значения величины.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ
Экстракт виноградных косточек
Один из аспектов настоящего изобретения относится к применению фармацевтических композиций, полученных из экстракта виноградных косточек, для лечения или профилактики нейродегенеративных заболеваний, ассоциированных с неправильным сворачиванием, накоплением, агрегацией и/или отложением белков. Как используют в настоящем документе, термин "экстракт виноградных косточек (GSE)" относится к материалу или одному или нескольким соединениям, которые экстрагируются из косточек, кожуры или жмыха винограда.
Экстракт виноградных косточек можно получать из различных источников. Например, Polyphenolics (отделение Constellation Wines U.S., Inc.) выпускает на рынок серию продуктов экстракта виноградных косточек под торговым названием MegaNatural®. Примеры коммерческих продуктов MegaNatural® включают MegaNatural® GSKE Grape Pomace Extract, MegaNatural®-BP и MegaNatural®-Gold. Экстракт виноградных косточек также можно получить с помощью некоторых конкретных способов экстракции и/или очистки. Например, экстракт виноградных косточек можно получить с использованием способа, описанного в патенте США № 6544581 (выданном Shrikhande et al, патент '581), или публикации патентной заявки США 2007/0071871 (выданной Shrikhande et al.), описания которых включены в настоящий документ в качестве ссылок в полном объеме.
MegaNatural®-AZ (или MNG-AZ), который является экспериментальным и не доступен коммерчески, имеет уникальные признаки, которые позволяют ему легко всасываться через слизистую оболочку кишечника вследствие удаления галлатной группы из полифенольных составляющих. В процессе изготовления MNG-AZ, неочищенный полифенольный экстракт подвергают ферментации в смешанной дрожжевой культуре в течение периода времени, требуемого для гидролиза галловой кислоты из мономеров и олигомеров проантоцианидинов. Кроме того, экстракт перерабатывают в порошковую форму, содержащую более 90% по массе полифенолов и более 3% по массе галловой кислоты (смотрите патент '581). Дрожжевую культуру выбирают вследствие танназной активности для высвобождения галловой кислоты из мономеров и полимеров виноградных косточек. Альтернативно неочищенные танназные ферменты можно получать способом ферментации с использованием дрожжей и плесеней и их можно добавлять в неочищенный экстракт виноградных косточек для высвобождения галловой кислоты. Полученный MNG-AZ характеризуется наличием менее чем приблизительно 12% по массе галлоилированных проантоцианидинов, исходя из общего количества проантоцианидинов. Без связи с какой-либо конкретной теорией, полагают, что удаление боковых групп галловой кислоты значительно повышает биодоступность MNG-AZ.
Полифенолы, важное семейство соединений в экстракте виноградных косточек, признаны в качестве эффективных антиоксидантов. Проантоцианидины, подкласс полифенолов, представляют собой полимерные соединения, происходящие из основных элементов катехина и эпикатехина и их соответствующих производных (например, галлата эпикатехина, в котором эпикатехин модифицирован присоединением галловой кислоты). Результат анализа компонентов экстракта виноградных косточек MNG-AZ проиллюстрирован на фиг.1. Молекулярная структура типичного гетерополимерного проантоцианидина содержит катехин, эпикатехин, эпигаллокатехин и их производные (эпигаллокатехин и галлат эпикатехина) (фиг.1A). Анализ MNG-AZ с помощью ВЭЖХ на нормальной фазе также указывает на присутствие мономерных и полимерных элементов проантоцианидинов (фиг.1B).
Проантоцианидины, содержащие галлат эпикатехина, можно дегаллоилировать путем микробиологического или ферментативного превращения (фиг.1C). Это удаление боковых цепей галловой кислоты обеспечивает главное отличие между MNG-AZ и другими коммерчески доступными экстрактами виноградных косточек. MNG-AZ содержит небольшое количество боковых цепей галловой кислоты или не содержит их, на что указывает процент галлоилированных проантоцианидинов (от всех проантоцианидинов) в MNG-AZ по сравнению с четырьмя другими коммерчески доступными препаратами GSE (фиг.1E и 1F). Показано, что в настоящем изобретении, MNG-AZ имеет неожиданную биологическую активность in vivo в отношении связанных с Aβ и тау нейродегенеративных заболеваний в моделях заболеваний на животных.
В конкретном варианте осуществления настоящего изобретения фармацевтическая композиция содержит конкретный экстракт виноградных косточек, а именно MegaNatural®-AZ. Фармацевтическая композиция по настоящему изобретению также может содержать одно или несколько соединений, полученных из экстракта виноградных косточек. Одно или несколько соединений могут включать, но не ограничиваться этим, один или несколько полифенолов, один или несколько проантоцианидинов или их смеси. Иллюстративные полифенолы включают, но не ограничиваются ими, мономерные основные элементы катехина и эпикатехина.
В других вариантах осуществления композиция по настоящему изобретению дополнительно содержит носитель. Предпочтительно, чтобы указанный носитель использовали в качестве пригодного вещества в соответствии со способом употребления и применения. Например, для перорального введения пригодные фармацевтические носители по настоящему изобретению включают, но не ограничиваются ими, лактозу, декстрозу, сахарозу, сорбит, маннит, крахмал, гуммиарабик, ксилит, эритрит, альгинат, желатин, фосфат кальция, силикат кальция, целлюлозу, метилцеллюлозу, поливинилпирролидон, воду, метилгидроксибензоат, пропилгидроксибензоат, стеарат магния и минеральное масло. Кроме того, композиция может включать наполнители, средства против агглютинации, смазывающие вещества, смачивающие вещества, вкусовые добавки, эмульгаторы, консерванты и т.п.
Фармацевтическую композицию по настоящему изобретению можно получать в качестве пероральной формы, включая, но не ограничиваясь ими, порошок, таблетку, капсулу, диспергирующуюся в полости рта таблетку, мягкую капсулу, водное лекарственное средство, сироп, эликсир и саше.
Альтернативно фармацевтическую композицию можно вводить трансдермально. Композиции по настоящему изобретению можно наносить прямо на кожу или непрямо через приспособление для чрескожной доставки. Композиции по настоящему изобретению можно изготавливать в качестве прямых чрескожных дозированных форм, таких как гель, крем, лосьон, эмульсия, масло, мазь, суспензия, аэрозоль, спрей или сходные с ними. Композиции по настоящему изобретению можно получать в виде непрямой чрескожной дозированной формы в качестве компонента приспособления для чрескожной доставки, включающего пластырь, повязку, липкую ленту или другую герметичную повязку. Кроме того, фармацевтическую композицию можно вводить трансназально, например, в качестве трансназального спрея. Также предусматриваются другие пассивные или активные приспособления для чрескожной доставки для всасывания через кожу или слизистую поверхность.
Пригодные фармацевтические носители для чрескожного введения композиций по настоящему изобретению могут представлять собой любой фармацевтически приемлемый материал носителя, пригодный для чрескожного введения лекарственного средства. Такие носители включают материалы, известные в данной области, такие как жидкость, гелеобразный растворитель, жидкий разбавитель, солюбилизатор, или сходные с ними. Пригодные носители являются нетоксичными и не взаимодействуют с другими компонентами композиции неблагоприятным образом. Примеры пригодных носителей для применения в этом случае включают воду, силикон, жидкие сахара, воски, вазелиновое масло. Также носители могут включать стабилизаторы, адъюванты, усилители проникновения или другие типы добавок, пригодные для способствования чрескожной доставке лекарственного средства.
В определенных вариантах осуществления, когда это применимо, соединения по настоящему изобретению можно использовать в форме их фармацевтически приемлемых солей, а также их можно использовать отдельно или в соответствующей комбинации, а также в комбинации с другими фармацевтически активными соединениями. В конкретных вариантах осуществления, композиция по настоящему изобретению, кроме того, может содержать антиоксидант и/или ингибитор холинэстеразы.
Способы лечения
Фармацевтическую композицию по настоящему изобретению можно вводить индивиду, имеющему фактор риска или состояние, ассоциированные с нейродегенеративными заболеваниями. Индивид может представлять собой человека или низшее млекопитающее, включая, но не ограничиваясь ими, кошку, собаку, крысу, мышь, овцу, козу, корову, обезьяну, шимпанзе и их трансгенные линии. Фармацевтическую композицию вводят индивиду в терапевтически эффективном количестве, в таких количествах и в течение такого времени, которые необходимы для достижения желаемых результатов. Нейродегенеративные заболевания, предусмотренные в настоящем документе, как правило, характеризуются повышенными уровнями одного или нескольких белков или пептидов в головном мозге индивида, включая их неправильное сворачивание, накопление, агрегацию и/или отложение. Эти заболевания включают, но не ограничиваются ими, болезнь Альцгеймера, болезнь Паркинсона, боковой амиотрофический склероз, болезнь Гентингтона, лобно-височную деменцию и кортико-базальную дегенерацию и/или таупатию. Таупатия может представлять собой болезнь Альцгеймера, прогрессирующий супрануклеарный паралич, кортикобазальную дегенерацию, болезнь аргирофильных гранул, болезнь Пика и семейную лобно-височную деменцию, среди прочих.
Как используют в настоящем документе, один или несколько белков (или, используемых взаимозаменяемо с "пептидами"), на которые нацелены способы лечения по настоящему изобретению, относятся к молекулам, состоящим из множества аминокислотных элементов, соединенных пептидными связями, где молекулы ассоциированы с одним или несколькими нейродегенеративными заболеваниями, как описано. Белки включают как белки дикого типа, так и мутантные, трансгенные и синтетические белки. Например, они могут включать, но не ограничиваться ими, конкретные белки, ассоциированные с конкретными нейродегенеративными заболеваниями. Например, белок амилоида-бета (например, Aβ1-40, Aβ1-42) и/или нейрофибриллярный клубок представляет собой белок-мишень(белки-мишени) у пациентов с болезнью Альцгеймера. Также мутантный белок htt является белком-мишенью у пациентов с болезнью Гентингтона, белок α-синуклеин является белком-мишенью у пациентов с болезнью Паркинсона и тау-белок является белком-мишенью у пациентов с таупатией.
Кроме того, предусматривается, что способы лечения согласно изобретению охватывают лечение индивидов, где заболевание, ассоциированное с повышенным уровнем белка или нарушенным сворачиванием, существует, но где индивиды не обладают внешним проявлением симптомов. Более того, способы лечения по настоящему изобретению предусматривают лечение симптомов существующих заболеваний, где индивиды обладают внешним проявлением симптомов.
Доза композиции по настоящему изобретению может варьировать в зависимости от массы и состояния индивида, формы композиции, способа и периода введения, и ее могут определять специалисты в данной области. Оптимальную дозу соединения(ий) можно определять в соответствии с количеством, требуемым для максимального эффекта снижения концентрации нежелательных или неправильно свернутых белков в конкретных областях головного мозга. Например, диапазон дозировок может составлять от приблизительно 100 до приблизительно 1000 мг в сутки. Предпочтительно диапазон дозировок составляет от приблизительно 200 до приблизительно 600 мг в сутки. Композицию можно вводить раз в месяц, раз в две недели, раз в неделю, раз в сутки или несколько раз в сутки в одной или разделенных дозах.
ПРИМЕРЫ
Следующие примеры являются только иллюстративными для настоящего изобретения, и их не следует истолковывать, как ограничивающие объем изобретения, никоим образом.
Пример 1. Доказательства эффектов экстракта виноградных косточек на образование олигомеров синтетического Aβ in vitro
Настоящий пример обеспечивает доказательства эффектов композиции согласно одному варианту осуществления настоящего изобретения в отношении уменьшения олигомеризации Aβ in vitro.
Материалы и способы
Анализ агрегации Aβ1-40 и Aβ1-42 in vitro. Продукт экстракта виноградных косточек, MegaNatural®-AZ, приобретали от Phenolics (Madera, CA). Пептиды Aβ1-40 и Aβ1-42 для анализов агрегации Aβ1-40 и Aβ1-42 in vitro приобретали от American Peptides (Sunnyvale, CA). Пептиды солюбилизировали в HFIP (Sigma), сушили в течение ночи при комнатной температуре и быстро вакуумировали в течение 10 минут. Пептиды растворяли в dH2O в концентрации 1 мг/мл, и исходный раствор GSE MNG-AZ растворяли в H2O в концентрации 400 мкМ. Aβ1-40 и Aβ1-42 (100 мкг/мл) смешивали с различными концентрациями GSE MNG-AZ в объеме 1:1 и инкубировали при 37°C в течение 3 суток. Эффект MNG-AZ на агрегацию Aβ анализировали вестерн-блот-анализом с использованием антитела 6E10.
Анализ фотоиндуцированного поперечного сшивания немодифицированных белков (PICUP). Свежевыделенный низкомолекулярный (LMW) пептид Aβ1-42 или Aβ1-40 смешивали с 1 мкл 1(×1), 2(×2), 5(×5) или 10(×10) мМ трис(2,2'-бипиридил)дихлоррутения(II) (Ru(bpy)) и 1 мкл 20(×1), 40(×2), 100(×5) или 200(×10) мМ персульфата аммония (APS) в присутствии или отсутствие 50 мкМ GSE MNG-AZ в 10 мМ фосфате при pH 7,4. Смесь облучали в течение 1 секунды и сразу гасили 10 мкл буфера для образца Tricine (Invitrogen, CA), содержащего 5% β-меркаптоэтанол. Реакционную смесь подвергали SDS-PAGE и визуализировали окрашиванием серебром (SilverXpress, Invitrogen, CA). Глутатион-S-трансферазу поперечно сшивали в сходных условиях и использовали в качестве контрольного пептида.
Результаты и обсуждение
Эффект MNG-AZ на ингибирование олигомеризации Aβ проиллюстрирован на фиг.2. Олигомеризация синтетических Aβ1-42 (2A) и Aβ1-40 (2B) ингибировалась MNG-AZ зависимым от концентрации образом, как проиллюстрировано на SDS-PAGE (дорожки 1-6 на фиг.2A и 2B: 0, 0,2, 1, 5, 25 и 100 мкМ Aβ; CTR представляют собой образцы без инкубации). Сходные результаты наблюдали в случае SDS-PAGE для Aβ1-42 (фиг.2C) и Aβ1-40 (фиг.2D) после химической реакции PICUP (дорожки 1 и 2 на фиг.2C и 2D: Aβ-пептид с 1×Ru(Bpy) и APS в присутствии и отсутствие MNG-AZ, соответственно; дорожки 3 и 4: Aβ пептид с 2×Ru(Bpy) и APS в присутствии или отсутствие GSE MNG-AZ, соответственно; CTR: не поперечно-сшитый Aβ1-42 (2C) или Aβ1-40 (2D), использованный в качестве мономерного контроля.
Указанные выше результаты согласованно указывают на то, что MNG-AZ может эффективно ингибировать олигомеризацию Aβ in vitro. Более того, ингибиторный эффект MNG-AZ, по-видимому, является зависимым от концентрации или дозы. Эти результаты демонстрируют, что GSE MNG-AZ может предупреждать или лечить заболевания, ассоциированные с накоплением, агрегацией или отложением Aβ.
Пример 2. Оценка нейропатологии типа AD у мышей TG2576
В данном примере иллюстрируются эффекты in vivo введения композиции согласно одному варианту осуществления настоящего изобретения на нейропатологию Aβ в модели на трансгенных мышах.
Материалы и способы
Мыши Tg2576 и введение GSE MNG-AZ. Взрослых самок мышей Tg2576 (Taconic, Germantown Inc.) распределяли на две различные группы: группу введения GSE MNG-AZ и группу водного контроля. GSE MNG-AZ доставляли в их питьевой воде в концентрации 1,2 г/л, что приводило к конечному употреблению 200 мг/кг/сутки. Это было эквивалентно дозе у человека 1 г/сутки при использовании критериев FDA для преобразования эквивалентных дозировок лекарственных средств между видами, исходя из площади поверхности тела (эквивалентная доза для человека в мг/кг=доза для животного в мг/кг × (масса животного в кг/масса человека в кг)0,33) (http://www.fda.gov/cber/gdlns/dose.htm). Животные имели свободный доступ к жидкости и стандартный корм. Питьевые растворы заменяли каждые трое суток. На протяжении исследования каждую неделю проводили мониторинг употребления жидкости и массы тела животного. После введения в течение 5 месяцев, мышей подвергали анестезии общим анестетиком кетамином HCL и ксилазином (Fort Dodge Animal Health, Fort Dodge, Iowa) и умерщвляли декапитацией. Головной мозг извлекали и разрезали вдоль полушарий. Одно полушарие фиксировали в 4% параформальдегиде в течение 24 часов для морфологических исследований. Гиппокамп и неокортекс отделяли от противоположного полушария, быстро замораживали, растирали в жидком азоте и хранили при 80°C для биохимических исследований.
Оценка амилоидной нейропатии по типу AD. Для количественной оценки Aβ-пептидов головного мозга замороженную растертую ткань гомогенизировали в буфере на основе гуанидина в концентрации 5 моль/л, разбавленного 1:10 в фосфатно-солевом буфере, содержащем 0,05% (об./об.) Tween®-20 и 1 ммоль/л ингибиторов протеаз Pefabloc (Roche Applied Science, Indianapolis, IN) и центрифугировали в течение 20 минут при 4°C. Общий Aβ1-40 или Aβ1-42 количественно определяли с помощью сэндвич-ELISA (BioSource, Camarillo, CA). Для стереологической оценки амилоидной нагрузки по типу AD у мышей Tg2576 свежевзятые полушария головного мозга фиксировали погружением в 4% параформальдегид в течение ночи и нарезали на срезы во фронтальной плоскости на вибратоме при номинальной толщине 50 мкм. Каждый 15 срез, начиная со случайного исходного положения, отбирали и обрабатывали для окрашивания тиофлавином-S. Все стереологические анализы проводили с использованием фотомикроскопа Zeiss Axiophot, оборудованного столиком Zeiss с электроприводом и контроллером столика MSP65, цифровой камерой Zeiss ZVS-47E высокого разрешения и компьютером Macintosh G3, выполняющим разработанное по заказу программное обеспечение NeuroZoom. Амилоидную нагрузку оценивали с использованием принципа Кавальери с сеткой малого размера (50×50 мкм) для точечного подсчета; эта процедура обеспечила неискаженную оценку долевого объема, занимаемого амилоидными бляшками - выраженного в качестве процента от объема неокортекса или гиппокампа. Оценку объема бляшек проводили с использованием процедуры систематического случайного взятия образцов при увеличении ×40.
Анализ растворимого олигомера Aβ головного мозга. Уровень растворимых олигомеров Aβ измеряли как с помощью дот-блот анализа, так и с помощью вестерн-блот-анализа. Конкретно, растворимый амилоидный пептид экстрагировали растворением растертой ткани коры в PBS, дополненном ингибитором протеаз (Roche Applied Science, Indianapolis, IN). После центрифугирования при 78500×g в течение 1 часа при 4°C, супернатант анализировали. 5 мкг общего белка наносили на нитроцеллюлозную мембрану, блокировали 10% обезжиренным молоком и инкубировали с антителом A11 (Invitrogen, CA), которое специфично распознает олигомерную форму Aβ. После инкубации при комнатной температуре в течение двух часов, блот инкубировали с конъюгированным с HRP антителом козы против антител кролика, и иммунореактивные сигналы визуализировали с использованием усиленной хемилюминесцентной детекции (SuperSignal Chemiluminescent Detection Kit, Pierce, Rockford, IL) и количественно определяли путем денситометрии (Quantity One, Bio-Rad). Тот же образец также использовали для вестерн-анализа. 75 мкг всех белков разделяли в геле 10-20% Tris-Tricine и переносили на нитроцеллюлозную мембрану, блокированную в течение 1 часа 10% обезжиренным молоком. Мембраны инкубировали либо с 6E10 (Signet), либо с A11. Иммунореактивные сигналы визуализировали с использованием усиленной хемилюминесцентной детекции и количественно определяли путем денситометрии.
Процессинг APP и активность α-, β-, γ-секретазы. Экспрессию голо-APP исследовали вестерн-блот-анализом с антителом C8 (индуцированным против AA676-695 цитоплазматического домена APP человека). Для детекции sAPP-α и sAPP-β проводили иммунопреципитацию, как описано ранее (Wang et al., FASEB J 2005; 19: 659-661). Активность α-, β-, γ-секретазы оценивали с использованием коммерчески доступных наборов (R & D Systems). Экспрессию неприлизина и деградирующего инсулин фермента анализировали вестерн-блоттингом с использованием коммерчески доступного антитела.
Статистический анализ. Все данные и величины в этих примерах выражали в качестве среднего значения и стандартной ошибки среднего значения (SEM). Различия между средними значениями анализировали с использованием либо 2-сторонних показателей ANOVA, либо 2-стороннего t-критерия Стьюдента. Во всех анализах нулевую гипотезу отвергали на уровне 0,05. Все статистические анализы проводили с использованием программы Prism Stat (GraphPad Software, Inc., San Diego CA).
Результаты и обсуждение
Эффекты GSE MNG-AZ на нейропатологию у мышей Tg2576 проиллюстрированы на фиг.7. На фиг.7А и 7В представлен эффект GSE MNG-AZ на массу тела (7А) или употребление жидкости (7В) у мышей Tg2576 после введения в течение 5 месяцев. На фиг.7С представлена оценка содержания растворимого внеклеточного пептида HMW-Aβ в головном мозге с использованием антитела, специфичного к олигомерным HMW-пептидам Аβ в дот-блот-анализе. На фиг.7D представлен вестерн-блот-анализ растворимого внеклеточного олигомерного HMW-пептида Аβ (антитело A11) и мономерного пептида Аβ (антитело 6Е10) в головном мозге мышей Tg2576. На фиг.7Е представлена оценка концентраций пептидов Aβ1-42 и Aβ1-40 в головном мозге мышей, которым вводили GSE MNG-AZ, и контрольных мышей. На фиг.7F представлена стереологическая оценка нагрузки Аβ-амилоидными бляшками коры головного мозга и парагиппокампальной извилины у мышей, которым вводили GSE MNG-AZ, и у контрольных мышей, выраженная в качестве положительного по тиофлавину-S объема как процента объема области. На вставке на фиг.7F представлены иллюстративные фотографии нейропатологии положительных по тиофлавину-S бляшек амилоида Аβ в неокортексе (СТХ) и парагиппокамиальной извилине (Hippo) у контролей без введения (верхняя панель) и у мышей Tg2576, которым вводили MNG-AZ (нижняя панель). Величины соответствуют групповому среднему значению ±SEM, n=5-6 мышей на группу. *р<0,05, **р<0,01, посредством анализа с помощью двухстороннего t-критерия Стьюдента. На фиг.7G проиллюстрирована оценка концентраций пептидов Aβ1-42 и Aβ1-40 в головном мозге при параллельных исследованиях эффективности MegaNatural®-Gold in vivo.
Эти результаты показали, что GSE MNG-AZ не вызывает поддающихся детекции побочных эффектов, включая изменение массы тела (фиг.7А) или употребления воды (фиг.7В). Нейропатология у мышей Tg2576 после введения в течение 5 месяцев показала снижение олигомеризации эндогенных пептидов Аβ в HMW-частицы Аβ приблизительно в два-три раза, согласно оценке вестерн-блот-анализом с использованием антитела, специфичного к олигомерам Аβ (р<0,05, фиг.7С) и вестерн-блоттингом с использованием антител A11 (p<0,01, фиг.7D). Было выявлено, что снижение иммунореактивных к A11 олигомерных HMW-частиц Аβ в головном мозге мышей Tg2576 совпадает с соответствующим увеличением количества мономерных пептидов Аβ (р<0,05, фиг.7D), что указывает на то, что GSE MNG-AZ благоприятно влияет на AD посредством предотвращения олигомеризации Аβ.
На фиг.7G показано, что в параллельных исследованиях введение мышам Tg2576 другого коммерчески доступного препарата GSE, а именно MegaNatural®-Gold, не модулировало накопление пептидов Aβ1-42 и Aβ1-40 в парагиппокамиальной извилине мышей Tg2576 по сравнению с контрольными мышами Тg2576. Результат, представленный на фиг.7G, указывает на то, что MNG-AZ является уникальным среди доступных в настоящее время препаратов GSE в отношении его эффективности модулирования нейродегенерации амилоидного типа в головном мозге вследствие его по существу более низкого содержания галлоилированных проантоцианидинов.
Недавние наблюдения указывают на то, что предотвращение олигомеризации Аβ в HMW-частицы в головном мозге может привести к компенсаторному снижению содержания всех пептидов Аβ и, в конечном итоге, содержания амилоидных нейритных бляшек в головном мозге, возможно, в результате предпочтительного выведения мономерных пептидов Aβ из головного мозга относительно олигомерных Aβ-частиц (Morelli et al., Biochem 2005: 38: 129-145). В соответствии с этой гипотезой, результаты данного примера показали, что, в дополнение к сниженным уровням олигомерных HMW-частиц Aβ (фиг.7C, 7D), длительное введение GSE MNG-AZ также значимо снижало количества пептидов Aβ1-42 (фиг.7E, левая панель) и Aβ1-40 (фиг.7E, правая панель) и нагрузку амилоидными нейритными бляшками (фиг.7F), относительно совпадающих по возрасту и полу контрольных мышей, которым вводили воду.
Кроме того, оценивали альтернативные потенциальные механизмы, которые могли привести к благоприятным эффектам GSE MNG-AZ на амилоидную нейропатологию. Не было выявлено поддающихся детекции изменений содержания голо-APP (фиг.8A), а также не было выявлено каких-либо изменений ферментативной активности α-, β- и γ-секретазы, или содержания растворимого APPα и растворимого APPβ (фиг.8B, 8C). Более того, не наблюдали поддающихся детекции изменений содержания неприлизина (фиг.8D) или деградирующего инсулин фермента (фиг.8E), которые являются основными протеолитическими ферментами, ответственными за деградацию Aβ. Наконец, не было выявлено поддающихся детекции изменений уровней пептидов Aβ1-42 и Aβ1-40 в периферической сыворотке (фиг.8F).
Эти данные указывают на то, что GSE MNG-AZ может оказывать его благоприятный эффект in vivo, главным образом, путем предотвращения олигомеризации Aβ в растворимые HMW-частицы, как выявлено in vitro.
Пример 3. Оценка когнитивной функции AD-типа у мышей TG2576
Этот пример демонстрирует эффекты введения композиций согласно некоторым вариантам осуществления настоящего изобретения на когнитивную функцию трансгенных мышей Tg2576.
Материалы и способы
Мышам Tg2576 вводили GSE MNG-AZ в течение 5 месяцев и оценивали их когнитивную функцию в возрасте 11 месяцев. Память при пространственном обучении оценивали с помощью поведенческого теста в водном лабиринте Морриса, как описано ранее (Morris, J Neurosci Methods 1984; 11: 47-60). Пространственную память оценивают путем регистрации латентного периода для выхода животного из воды на погруженную платформу для выхода как функции обучающих испытаний в ходе фазы обучения. Через 24 часа после фазы обучения мышей исследовали в пробном испытании путем удаления платформы для выхода без изменения визуальных ориентиров. Анализ поведения последовательно проводили в течение последних 4 часов дневной части светового цикла в условиях с минимальными стимулами (например, шум, движение или изменения света или температуры).
Результаты и обсуждение
На фиг.9 проиллюстрировано снижение когнитивного нарушения у мышей Tg2576, которым вводили GSE MNG-AZ, совпадающее со снижением внеклеточных олигомерных HMW-Aβ. На фиг.9A и 9B представлено влияние GSE MNG-AZ на связанную с Aβ пространственную память у мышей Tg2576 при определении в тестах водного лабиринта Морриса. На фиг.9A представлен показатель латентности (соответствующий времени, затраченному на выход на платформу из воды) как функция времени введения. На фиг.9B представлен процент времени пробного испытания, которое мыши Tg2576 провели в заданном секторе (вычисленное как отношение времени, проведенного в области заданного сектора относительно времени, проведенного в остальной части ванны). На фиг.9C представлена оценка содержания растворимого внеклеточного HMW-пептида Aβ в головном мозге мышей Tg2576 с использованием антитела, специфичного к олигомерным HMW-пептидам Aβ, в дот-блот анализе. На вставке на фиг.9C представлен иллюстративный дот-блот содержания растворимого HMW-Aβ. Величины соответствуют групповому среднему значению ±SEM, n=7-9 мышей на группу; на фиг.9B, ***p<0,0001; на фиг.9C, *p<0,01 при анализе с помощью 2-стороннего t-критерия Стьюдента.
Мыши Tg2576 в возрасте 11 месяцев проявляли значительные нарушения функции долговременной памяти, что отражалось их неспособностью учиться использовать основные ориентиры для нахождения скрытой платформы для выхода в процессе обучающих испытаний в тесте водного лабиринта Морриса (фиг.9A). Напротив, мыши Tg2576, которым вводили MNG-AZ, проходили тест поведенческой функции, связанной с пространственной памятью, значительно лучше и были способны учиться использовать пространственные основные ориентиры для нахождения платформы для выхода, что отражалось значимым снижением латентного времени для выхода при последовательных обучающих испытаниях (двухсторонний повторяющийся показатель ANOVA; GSPE против контрольной группы: F1,11=4,90; p=0,049 для введения GSPE, F7,77=4,25; p=0,0005 для времени и F7,77=1,63; p=0,140 для взаимодействия) (фиг.9A). Индуцированное MNG-AZ снижение когнитивного нарушения у мышей Tg2576 подтверждали путем анализа сохранения пространственной памяти в пробном испытании, показавшем, что мыши, которым вводили MNG-AZ, проводили значительно больше времени в области заданного сектора относительно контрольных мышей, которым вводили воду (фиг.9B). Это улучшение когнитивной функции совпало со значимым снижением количества олигомерных HMW-частиц Aβ в головном мозге мышей Tg2576, которым вводили MNG-AZ, относительно контрольных мышей (фиг.5C).
В контрольном исследовании было выявлено, что введение GSE MNG-AZ не влияет на когнитивную функцию у совпадающих по линии, возрасту и полу животных дикого типа (фиг.10A и 10B). Указанные выше результаты указывают на то, что GSE MNG-AZ может быть полезен селективно при дефицитах долговременной пространственной памяти у мышей Tg2576, путем ослабления опосредуемого Aβ AD-типа ответа в головном мозге.
Пример 4. Эффект MNG-AZ на агрегацию тау-белков in vitro
Данный пример иллюстрирует эффекты in vitro композиции согласно одному варианту осуществления настоящего изобретения на агрегацию тау-пептида, диссоциацию предварительно сформированных агрегатов тау и стабильность нативных тау-фибрилл, полученных из образца головного мозга при AD.
Материалы и способы
Оценка биологической активности MNG-AZ против агрегации тау in vitro. N-ацетилированный тау-пептид из 6-аминокислот (Ac-306VQIVYK311), соответствующий остаткам 306-311 тау, приобретают коммерчески. Этот синтетический тау-пептид представляет собой короткий пептидный сегмент, встречающийся в связывающей микротрубочки области тау-белка. Данные указывают на то, что этот короткий пептидный сегмент необходим для полимеризации тау. (Goux et al, J. Biol. Chem. 2004; 279: 26868-26875). Это подтверждается биофизическим наблюдением in vitro, что короткий пептид Ac-306VQIVYK311 спонтанно агрегирует в фибриллярные структуры в присутствии соли (Goux et al., 2004, выше). Олигомеризация пептида Ac-306VQIVYK311 была по существу такой, как описано в Goux et al., 2004, выше. В кратком изложении, синтетический тау-пептид растворяли в 20 мМ MOPS, pH 7,2. Полимеризацию тау-пептида проводили в 75 мкл конечных растворов 20 мМ MOPS (pH 7,2), содержащих 2,2 мкМ пептид и 10 мкМ тиофлавин-S (ThS). Реакцию запускали добавлением соли до конечной концентрации 0,15 M. Кинетику агрегации тау-пептида в отсутствие или в присутствии различных концентраций MNG-AZ оценивали в течение 1 часа по последующему повышению флуоресценции ThS при связывании ThS с агрегированными пептидными частицами; флуоресцентное возбуждение индуцировали при 436 нм и флуоресцентное испускание определяли при 470 нм.
Кроме того, также для исследования влияния MNG-AZ на исходные белок-белковые взаимодействия, необходимые для образования агрегатов тау-пептида, использовали PICUP, спектроскопию кругового дихроизма (CD) и способы электронной микроскопии.
Для анализа PICUP, 25 мкМ тау-пептид поперечно сшивали в присутствии или отсутствие равного количества моль (25 мкМ) MNG-AZ, и мультимерные тау-пептиды разделяли посредством SDS-PAGE и визуализировали окрашиванием серебром.
Для спектроскопии CD, тау-пептид инкубировали при 37°С в течение 1-3 суток в присутствии 10 мМ фосфата, pH 7,4, и спектры CD получали в начале инкубации (0 сутки), и каждые из следующих 3 суток.
Для электронной микроскопии, тау-пептид инкубировали при 37°C в течение 3 суток в 10 мМ фосфате натрия, pH 7,4, в отсутствие или в присутствии GSE в молярном отношении 1:1 относительно тау-пептидов. После инкубации раствор центрифугировали при 16000×g в течение 5 мин, а затем 200 мкл супернатантов фракционировали на колонке для гель-фильтрации. Тау-фибриллы выявляли и выделяли при времени элюирования ~12 мин по УФ-поглощению при 254 нм.
Оценка способности MNG-AZ диссоциировать предварительно сформированные агрегаты тау in vitro. Синтетический тау-пептид Ac-306VQIVYK311 агрегировал в отсутствие GSE MNG-AZ. После образования агрегатов тау в реакционные смеси добавляли различные концентрации MNG-AZ и проводили мониторинг изменений содержания агрегатов тау в ответ на добавление GSE по последующей флуоресценции ThS.
Оценка способности GSE MNG-AZ вызывать дестабилизацию нативных тау-фибрилл, полученных из образцов головного мозга при AD. PHF выделяли и очищали из полученных при аутопсии образцов головного мозга при случаях AD, и некоторые образцы PHF обрабатывали MNG-AZ (100 мкМ) в течение 5 с или 1 ч. Результаты наблюдали с помощью электронной микроскопии (Hitachi H700).
Оценка эффектов GSE MNG-AZ на расщепление трипсином тау-фибрилл. PHF выделяли из образцов головного мозга при AD. Некоторые образцы PHF обрабатывали 100 мкМ MNG-AZ в течение 10 мин. Некоторые образцы PHF (которые предварительно обрабатывали MNG-AZ) далее инкубировали с 1 мкг трипсина в течение 10 мин. Результаты наблюдали с помощью электронной микроскопии (Hitachi H700).
Результаты и обсуждение
MNG-AZ ингибирует агрегацию тау. Результаты ингибирования агрегации тау посредством GSE MNG-AZ проиллюстрированы на фиг.11 (следует отметить, что MNG-AZ обозначен как MegN на фиг.11). Синтетический тау-пептид Ac-306VQIVYK311 легко образует агрегаты с течением времени в присутствии соли, что отражается повышением флуоресценции ThS как функции времени реакции (фиг.11A, где по ThS-флуоресценции оценивали накопление агрегатов тау как функцию времени; концентрации GSE на уровне 0,22 мкМ, 2,2 мкМ и 22 мкМ соответствуют, соответственно, молярному отношению GSE относительно тау-пептидов 1:10, 1:1 и 10:1).
Добавление MegaNatural®-AZ GSE в количестве от 0,22 до 22 мкМ значимо препятствовало агрегации тау-пептида дозозависимым образом (односторонний ANOVA, p<0,0001); вычисленное среднее значение максимального флуоресцентного испускания в отсутствие GSE составляет 122,9±1,3 единиц, по сравнению с вычисленным средним значением флуоресцентного испускания, составляющим 114,2±1,1, 109,6±0,6 и 100,6±0,3 единиц, соответственно, в присутствии 0,22, 2,2 и 22 мкМ MegaNatural®-AZ GSE (как показано на фиг.11B, где максимальное накопление агрегатов тау вычислено как средняя единица флуоресценции в диапазоне 6-10 мин). Более того, каждое пошаговое повышение содержания GSE MNG-AZ приводило к значимому пошаговому снижению флуоресценции ThS (апостериорный парный анализ Tukey, p<0,001 для отсутствия MNG-AZ против 0,22 мкМ MNG-AZ, 0,22 против 2,2 мкМ MNG-AZ, и 2,2 против 22 мкМ MNG-AZ). Интересно, что поддающееся детекции сниженное накопление агрегатов тау-пептида наблюдали при низком содержании GSE MNG-AZ, составляющем 0,22 мкМ, которое соответствует молярному отношению GSE относительно тау-пептидов 1:10. Агрегация тау-пептида полностью ингибировалась при 22 мкМ GSE, где GSE присутствовал в молярном избытке 10:1 относительно тау-пептидов (фиг.11A и 11B).
Уровень агрегатов тау значимо снижался в присутствии MNG-AZ, как показано в результатах анализа PICUP (фиг.16, где концентрации реагентов являются следующими. Контроль: нет Ru(Bpy) и нет APS; дорожки 1, 2: 1 мкл 1 мМ Ru(Bpy) и 1 мкл 20 мМ APS; дорожки 3, 4: 2 мкл 1 мМ Ru(Bpy) и 2 мкл 20 мМ APS; дорожки 5, 6: 3 мкл 1 мМ Ru(Bpy) и 3 мкл 20 мМ APS; дорожки 7, 8: 4 мкл 1 мМ Ru(Bpy) и 4 мкл 20 мМ APS). Это указывает на то, что MNG-AZ может ингибировать агрегацию тау-пептида, частично, препятствуя начальным стадиям самоассоциации тау-пептида.
Конформации тау-пептидов в процессе инкубации тау-пептидов (1-3 сутки), как показано в результатах спектроскопии кругового дихроизма (фиг.17A и 17B), также указывают на то, что MNG-AZ влияет на сборку тау-пептида. В отсутствие GSE случайная ассоциация тау-пептида постепенно преобразовывалась в упорядоченные конформеры β-слоя после инкубации в течение 2-3 суток, как показано по растущей величине части спектров с центром на уровне ~198 нм (фиг.17A). Напротив, в присутствии молярного отношения GSE относительно тау-пептидов 1:1, коинкубация тау-пептида с GSE препятствовала преобразованию тау-пептидов в упорядоченные вторичные структуры (фиг.17B).
Как исследовали посредством электронной микроскопии, морфология конформеров тау-пептида в отсутствие и в присутствии MNG-AZ показала, что тау-пептид спонтанно агрегирует в спиралевидные протофибриллы (фиг.18A). Напротив, присутствие GSE полностью ингибировало образование протофибрилл тау-пептида (фиг.18B).
В совокупности, указанные выше данные указывают на то, что MNG-AZ препятствовали агрегации тау-белка в олигомерные PHF.
MNG-AZ способствует диссоциации предварительно сформированных агрегатов тау. Показано, что MNG-AZ вызывает диссоциацию предварительно сформированных агрегатов тау-пептида Ac-306VQIVYK311. В частности, добавление 1,1 мкМ MNG-AZ, соответствующего молярному отношению MNG-AZ относительно тау-пептидов 1:1, было способно снизить содержание предварительно сформированных агрегатов тау-пептидов, что отражалось все в большей мере сниженными количествами ThS-положительных агрегатов тау как функция времени (на фиг.12A представлен график флуоресценции ThS для содержимого агрегата тау при различных концентрациях MNG-AZ (0,55-4,4 мкМ)). Как и ожидалось, в параллельных исследованиях с использованием более высоких концентраций MNG-AZ (2,2 мкМ и 4,4 мкМ MNG-AZ) также происходила диссоциация предварительно сформированного агрегата тау-пептида. Следует отметить, что на фиг.12A и фиг.12B, ниже, MNG-AZ обозначен как MegN. Эффективность ответа на дозу MNG-AZ в отношении диссоциации предварительно сформированных агрегатов тау исследовали путем вычисления скорости диссоциации агрегата тау в присутствии различных концентраций GSE MNG-AZ посредством линейного регрессионного анализа флуоресцентного испускания ThS (фиг.12B). Оказалось, что добавление возрастающих концентраций GSE MNG-AZ от 1,1 до 4,4 мкМ стимулировало диссоциацию предварительно сформированных агрегатов тау-пептида дозозависимым образом. Иными словами, скорость диссоциации агрегированных тау-пептидов прямо коррелировала с концентрацией GSE MNG-AZ (критерий Пирсона R=0,96654, p<0,05).
MNG-AZ препятствует стабильности тау-фибрилл. Эффекты MNG-AZ на стабильность тау-фибрилл при исследовании электронной микроскопией проиллюстрированы на фиг.19. На фиг.19A представлены PHF при AD, которые проявляют типичные организованные фибриллярные структуры со средней шириной 18,9+3,4 нм и средней длиной витка спирали 81,3+10,8 нм, и иммунологическое мечение золотом, с использованием антитела PHF1, определило локализацию эпитопа фосфосерина 396/404 проксимально от плотного центра PHF. Интересно, что инкубация выделенных PHF с GSE индуцировала пошаговое разворачивание PHF при увеличении длительности воздействия GSE (фиг.19B-19D); инкубация в течение 1 ч с GSE привела к 67% увеличению ширины фибрилл (до 31,6+3,8 нм) без влияния на средний виток спирали (средняя ширина и виток спирали обработанных GSE PHF составили, соответственно, 31,6+3,8 и 77,4+10,8 нм) (фиг.19C, 19D). Более того, было выявлено, что обработка GSE маскировала иммунореактивность выделенных тау-фибрилл к антителу PHF1 (фиг.19B и 19C, по сравнению с фиг.19A).
Было выявлено, что обработка выделенных тау-филаментов GSE стимулирует расщепление трипсином тау-филаментов (фиг.20). PHF, выделенные из головного мозга при AD, инкубировали с 1 мкг/мл трипсина (фиг.20C и 20D) или без него (фиг.20A, 20B) в течение 10 мин. Образцы на фиг.20B и 20D предварительно обрабатывали GSE (100 мкМ GSE, 1 ч). После обработки трипсином PHF сохраняли их филаментный вид (смотрите вставку на фиг.20C). Предварительная обработка GSE перед расщеплением трипсином вызывала дезинтеграцию PHF на аморфные фрагменты тау, иммунореактивные к антителу против тау AH-1 (смотрите стрелки и вставку на фиг.20D). Эти результаты указывают на то, что опосредуемое GSE модулирование конформации филаментов тау может стимулировать деградацию тау клеточными протеазами.
В общем, было выявлено, что GSE MNG-AZ ингибирует агрегацию синтетического тау-пептида в филаменты и вызывает диссоциацию предварительно сформированных агрегатов тау. Это указывает на то, что взаимодействия MNG-AZ с тау могут ослабить накопление отложений агрегатов тау, которое, в свою очередь, является ключевым нейропатологическим признаком множества ассоциированных с тау нейродегенеративных нарушений. Более того, также было выявлено, что GSE MNG-AZ смягчает опосредуемые тау фенотипы путем препятствования образованию и/или стабильности нейротоксических протофибрилл тау. Таким образом, указанные выше наблюдения обеспечивает убедительные доказательства того, что MNG-AZ или составляющие его соединения можно использовать в качестве профилактической меры для снижения возникновения таупатий или в качестве лекарственных средств для лечения ассоциированных с тау нейродегенеративных заболеваний.
Пример 5. Эффект экстракта виноградных косточек in vivo на тау и формы пептида HTT, расширенные полиглутамином
Модели на Drosophila с использованием индуцибельной системы Gal4/UAS (Brand, et al, Development, 1993; 118: 401-415) для трансгенной сверхэкспрессии ассоциированных с заболеванием склонных к агрегации белков успешно моделируют аспекты таупатии путем сверхэкспрессии мутантного тау R406W и болезни Гентингтона путем сверхэкспрессии Q93httexon1 (смотрите, например, Sang et al, NeuroRx. 2005; 2: 438-446.; Berger et al, Hum Mol Genet 2006; 15: 433-442). В частности, сверхэкспрессия R406W в клетках, которые формируют глаз (ey>R406W) приводит к значительному снижению или полному отсутствию глаз; глаза, которые формируются, демонстрируют аномальную морфологию. Более того, экспрессия Gal4 во всей нейронной системе (elav-Gal4) в трансгенных линиях, содержащих UAS-Q93httexon1, приводит к возникновению нейродегенерации во взрослом состоянии и снижению продолжительности жизни.
В данном примере проиллюстрирована польза экстракта виноградных косточек in vivo в отношении фенотипа Drosophila, несущих мутантный тау (R406W) или расширенные полиглутамином формы htt (Q93httexon1), которые моделируют определенные формы таупатии и болезни Гентингтона, соответственно.
Материалы и способы
Исследование фенотипа глаза мух ey>R406W. Яйца ey>R406W откладывались и выводились в контрольном корме (рецепт среды для мух быстрого приготовления 4-24 blue) или в корме, дополненном 2,8 мкг/мл GSE MNG-AZ (или GSPE, на фиг.13 и 14). Глаза Drosophila наблюдали под микроскопом.
Мониторинг продолжительности жизни мух elav>httQ3. Самцов мышей, сверхэкспрессирующих Q93httexon1 во всей нейрональной системе с использованием elav-Gal4 (elav>Q93httexon1) собирали в течение одних суток после вылупления и помещали по 10 в колбу с контрольным кормом или кормом, дополненным 2,8 мкг/мл GSE MNG-AZ. Выживших мух подсчитывали каждые сутки и переносили в свежие колбы каждые несколько суток.
Результаты и обсуждение
GSE подавляет сверхэкспрессию тау R406W в глазах Drosophila. Сверхэкспрессия R406W рано в развитии глаза приводила к небольшим глазам или отсутствию глаз (фиг.13A), однако введение GSE подавляло уменьшение размера глаз (фиг.13B) (показаны глаза самцов). Диапазон фенотипов ey>R406W варьировал от испытания к испытанию, так что испытания оценивали по отдельности. Визуальную оценку глаз самцов из иллюстративного эксперимента получали в пределах 3 суток после вылупления (0=нет глаз, 4=практически нормальный глаз) (фиг.13C). Количество отсутствующих глаз снижалось при введении GSE (справа) в том же испытании (фиг.13D).
Введение GSE продлевает продолжительность жизни Drosophila elav>httQ3. Было выявлено, что мухи elav>Q93httexon1, которым вводили GSE, характеризуются значительным повышением общей продолжительности жизни по сравнению с контролями без введения (фиг.14). В среднем 50% самцов elav>Q93httexon1 на контрольном корме погибали к 20 суткам, по сравнению только с 20% самцов на GSE.
Указанные выше результаты показывают, что введение GSE подавляет две различных модели нейродегенерации у Drosophila, вовлекающие агрегацию белка in vivo. Данные указывают на то, что GSE может иметь терапевтическую ценность в отношении профилактики и/или лечения склонных к агрегации белка нейродегенеративных нарушений.
Пример 6. Эффекты экстракта виноградных косточек на самосборку и цитотоксичность Aβ
В данном примере предоставлены доказательства эффектов композиции согласно одному варианту осуществления настоящего изобретения на снижение самосборки и цитотоксичности Aβ.
Материалы и способы
Химические вещества и реагенты. Химические вещества приобретали от Sigma-Aldrich Co. (St. Louis, MO) и они имели наиболее высокую доступную чистоту. Medysin #1 ("Med1") приобретали от Aurora Fine Chemicals Ltd., Graz, Austria. MegaNatural-AZ (MNG-AZ) приобретали от Polyphenolics (Madera, CA). (Как MNG-AZ, так и Med1 иногда называют "соединением", когда упоминают их смеси с Aβ). Воду дважды дистиллировали и деионизировали с использованием системы Milli-Q (Millipore Corp., Bedford, MA).
Пептиды и белки. Пептиды Aβ синтезировали, очищали и охарактеризовывали, как описано ранее (Walsh, et al., J Biol Chem 1997; 272: 22364-22372). В кратком изложении, синтез проводили на автоматизированном устройстве для синтеза пептидов (модель 433A, Applied Biosystems, Foster City, CA) с использованием способов на основе 9-флуоренилметоксикарбонила на предварительно нагруженных смолах Ванга. Пептиды очищали с использованием обращенно-фазовой высокоэффективной жидкостной хроматографии (ОФ-ВЭЖХ). Количественный анализ аминокислот и масс-спектрометрия дали ожидаемые составы и молекулярные массы, соответственно, для каждого пептида. Очищенные пептиды хранили в качестве лиофилизатов при -20°С. Исходный раствор глутатион-S-трансферазы (GST; Sigma-Aldrich, St. Louis, MO) получали растворением лиофилизата до концентрации 250 мкМ в 60 мМ NaOH. Перед применением аликвоты разбавляли в 10 раз в 10 мМ фосфате натрия, pH 7,4.
Получение исходных растворов Aβ. Не содержащие агрегатов исходные растворы Aβ получали с использованием гель-фильтрации (SEC). Номинальную мономерную фракцию назвали низкомолекулярным (LMW) Aβ, поскольку при экспериментальных концентрациях пептидов эта фракция содержит смесь мономера и низкомолекулярных олигомеров в быстром равновесии. Для получения Aβ 200 мкл раствора пептида в концентрации 2 мг/мл (номинальная концентрация) в диметилсульфоксиде обрабатывали ультразвуком в течение 1 мин с использованием устройства для обработки ультразвуком на бане (Branson Ultrasonics, Danbury, CT), а затем центрифугировали в течение 10 мин при 16000×g. Полученный супернатант фракционировали на колонке Superdex 75 HR с использованием 10 мМ фосфатного буфера, pH 7,4, при низкой скорости потока 0,5 мл/мин. Среднюю часть пика Aβ собирали в течение 50 с и использовали сразу для всех экспериментов. Аликвоту объемом 10 мкл отбирали для анализа аминокислот для количественного определения концентрации пептидов в каждом препарате. Как правило, концентрации Aβ1-40 и Aβ1-42 составили 30-40 мкМ и 10-20 мкМ, соответственно.
Инкубация Aβ. Образцы Aβ приготавливали, как описано выше, а затем аликвоты объемом 0,5 мл помещали в 1-мл центрифужные микропробирки. Тестируемые соединения растворяли в этаноле до конечной концентрации 2,5 мМ, а затем разбавляли 10 мМ фосфатом, pH 7,4, с получением концентраций 10 и 50 мкМ. Затем 0,5 мл каждого соединения добавляли в отдельные пробирки с Aβ с получением конечных концентраций пептида ~20 мкМ (Aβ1-40) и ~10 мкМ (Aβ1-42) и конечных концентраций ингибитора 5 и 25 мкМ. Таким образом, соотношения соединение : пептид составляли ~1:4 (Aβ1-40) и ~1:2 (Aβ1-42) при более низкой концентрации соединения и 5:4 (Aβ1-40) и 5:2 (Aβ1-42) при более высокой концентрации соединения. В контрольные пробирки с пептидом отдельно добавляли 0,5 мл буфера. Пробирки инкубировали при 37°C в течение 0-7 суток без перемешивания.
Химическое поперечное сшивание и распределение частот олигомера. Сразу после приготовления образцы подвергали поперечному сшиванию с использованием способа PICUP. В кратком изложении, к 18 мкл раствора белка добавляли 1 мкл 1 мМ Ru(bpy) и 1 мкл 20 мМ персульфата аммония (APS). Конечные соотношения белок:Ru(bpy):APS Aβ1-40 и Aβ1-42 составили 0,29:1:20 и 0,16:1:20, соответственно. Смесь облучали в течение 1 с видимым светом, а затем реакционную смесь гасили 10 мкл буфера для образца Tricine (Invitrogen, Carlsbad, CA), содержащего 5% β-меркаптоэтанол. Определение распределения мономеров и олигомеров проводили с использованием SDS-PAGE и окрашивания серебром. В кратком изложении, 20 мкл каждого поперечно-сшитого образца подвергали электрофорезу на 10-20% градиентном геле с Tricine и визуализировали окрашиванием серебром (SilverXpress, Invitrogen). В качестве контролей в каждом эксперименте использовали не поперечно-сшитые образцы. Для получения профилей интенсивности и вычисления относительных количеств каждого типа олигомера проводили денситометрию, и для определения площадей пиков скорректированных по исходному уровню данных использовали программное обеспечение One-Dscan (v. 2.2.2; BD Biosciences Bioimaging, Rockville, MD). В некоторых экспериментах, молярные количества Ru(bpy) и APS были повышены относительно пептида на коэффициенты 2, 5, 10 и 20.
Спектроскопия CD. Спектры CD растворов Aβ получали сразу после приготовления образца и после инкубации в течение 2, 3, 6 или 7 суток. Измерения CD проводили путем извлечения аликвоты реакционной смеси объемом 200 мкл, добавления аликвоты в кювету для CD с длиной пути 1 мм (Hellma, Forest Hills, NY), и получением спектров в спектрополяриметре J-810 (JASCO, Tokyo, Japan). Кюветы CD хранили на льду перед помещением в спектрометр. После уравновешивания температур, спектры CD регистрировали при 22°C при ~190-260 нм с разрешением 0,2 нм и скоростью сканирования 100 нм/мин. Получали десять изображений и для каждого образца данные усредняли. Необработанные данные обрабатывали путем сглаживания и вычитания спектров буфера согласно инструкциям изготовителей.
Анализ связывания тиофлавина T (ThT). Десять мкл образца добавляли к 190 мкл ThT, растворенного в 10 мМ фосфатном буфере (pH 7,4), и смесь кратковременно встряхивали. Флуоресценцию определяли три раза с интервалами 10 секунд с использованием флуориметра Hitachi F-4500. Длины волн возбуждения и испускания составляли 450 и 482 нм соответственно. Флуоресценцию образца определяли усреднением трех полученных наборов данных и вычитанием флуоресценции пустого образца с ThT.
Электронная микроскопия (EM). Аликвоты каждого образца объемом 10 мкл наносили точечно на покрытые углеродом решетки Formvar с тлеющим разрядом (Electron Microscopy Sciences, Hatfield, PA) и инкубировали в течение 20 минут. Затем каплю вытесняли равным объемом 2,5% (об./об.) глутаральдегида в воде и инкубировали в течение дополнительных 5 минут. Наконец, пептид окрашивали 8 мкл 1% (об./об.) отфильтрованного (0,2 мкм) уранилацетата в воде (Electron Microscopy Sciences, Hatfield, PA). Этот раствор удаляли и решетку сушили воздухом. Образцы исследовали с использованием трансмиссионной электронной микроскопии JEOL CX100.
Метаболизм бромид 3-[4,5-диметилтиазол-2-ил]-2,5-дифенилтетразолия (MTT). Клетки феохромацитомы крысы PC12 культивировали в 75-см2 флаконах (#430641, Corning Inc., Corning, NY) в среде F-12K (ATCC, Manassas, VA), содержащей 15% (об./об.) сыворотку лошади, 2,5% (об./об.) эмбриональную телячью сыворотку, 100 единиц/мл пенициллина, 0,1 мг/мл стрептомицина и 25 мкг/мл амфотерицина B при 37°С в 5% (об./об.) CO2 на воздухе. Для приготовления клеток для анализа среду удаляли и клетки осторожно промывали один раз средой F-12K, содержащей 0,5% (об./об.) эмбриональную телячью сыворотку, 100 единиц/мл пенициллина, 0,1 мг/мл стрептомицина и 25 мкг/мл амфотерицина B. Затем получали суспензию клеток добавлением этой указанной среды, но содержащей 100 мкг/мл фактора роста нервов (Invitrogen, CA), с последующим встряхиванием флакона. После подсчета клеток с использованием трипанового синего клетки высевали с плотностью 30000 клеток/лунка (общий объем/лунка 90 мкл) в 96-луночных планшетах для анализа (Costar #3610, Corning Inc., Corning, NY). Индуцированной фактором роста нервов дифференцировке клеток позволяли протекать в течение 48 ч, и в этот момент проводили анализы токсичности.
Токсичность Aβ оценивали двумя способами. Пептиды предварительно инкубировали либо с 0, либо с 25 мкМ MNG-AZ в 10 мМ фосфате натрия, pH 7,4 при 37°C в течение 0, 2, 3 или 7 суток, и в это время в лунки добавляли 10 мкл раствора пептида. Альтернативно Aβ инкубировали как описано выше, но в отсутствие MNG-AZ. В этом случае, растворы пептидов смешивали с 0 или 25 мкМ MNG-AZ непосредственно перед добавлением к клеткам. Клетки обрабатывали в течение 24 ч при конечной концентрации 0 или ~2 мкМ только Aβ или обработанного MNG-AZ Aβ, содержавшего 2,5 мкМ MNG-AZ. Соотношения пептид/соединение для пептидов Aβ1-40 и Aβ1-42 составляли 0,72 и 0,39, соответственно. На практике, образцы "нулевого времени" для каждой альтернативной экспериментальной процедуры были эквивалентными, поскольку все компоненты смешивали с клетками одновременно.
Для определения токсичности в каждую лунку добавляли 15 мкл раствора MTT (Promega, Madison, WI) и планшет держали в инкубаторе с CO2 в течение дополнительных 3,5 часа. Затем клетки подвергали лизису посредством добавления 100 мкл раствора для солюбилизации (Promega, Madison, WI) с последующей инкубацией в течение ночи. Восстановление MTT оценивали путем измерения поглощения при 570 нм (с коррекцией по фоновому поглощению при 630 нм) с использованием устройства для считывания микропланшетов BioTek Synergy HT (Bio-Tek Instruments, Winooski, Vermont). Контроли включали среду с фосфатом натрия ("отрицательный"), фибриллы ("положительный") и 1 мкМ стауроспорин ("максимально положительный"). Фибриллярные Aβ1-40 и Aβ1-42 добавляли к клеткам в конечных концентрациях 10 мкМ и 5 мкМ, соответственно. Те же препараты фибрилл использовали для всех экспериментов, и они служили для контроля вариаций между анализами. Для осуществления сравнения между анализами, сначала определяли токсичность в каждом эксперименте. Для каждой группы введения проводили шесть повторений и данные 3 независимых экспериментов объединяли и описывали в качестве среднего значения ±S.E. Процентная токсичность T=((AAβ-Aсреда)/(Астауроспорин-Aсреда))×l00; где AAβ, Aсреда, Астауроспорин представляли собой величины поглощения содержащих Aβ образцов, среды отдельно или стауроспорина отдельно, соответственно.
Статистический анализ. Для статистического анализа использовали односторонний долевой ANOVA и критерии множественного сравнения с использованием статистических процедур GraphPad Prism (version 4.0a, GraphPad Software, Inc., San Diego, CA). Значимым считали значение p<0,05.
Результаты и обсуждение
Олигомеризация Aβ. В отсутствие поперечного сшивания PICUP были выявлены только мономеры Aβ1-40 (фиг.15A, дорожка 2) и мономеры и тримеры Aβ1-42 (фиг.15B, дорожка 2). Было показано, что полоса тримера Aβ1-42 является индуцированным SDS артефактом. После поперечного сшивания Aβ1-40 существовал в качестве смеси мономеров и олигомеров порядка 2-4 (фиг.15A, дорожка 3), в то время как Aβ1-42 содержал мономеры и олигомеры порядка 2-6 (фиг.15B, дорожка 3).
Когда MNG-AZ смешивали с Aβ1-40 в отношении соединение:пептид ~5:4, олигомеризация блокировалась практически полностью (фиг.15A, дорожка 6). Полоса тримера была слабо видимой, и интенсивность димера также была минимальной. Повышение в десять раз отношения соединение:пептид приводило к сходным уровням ингибирования (фиг.15A, дорожка 7). Эффект MNG-AZ на олигомеризацию Aβ1-42 был в равной степени значимым (фиг.15B). При отношении соединение:пептид ~5:2, MNG-AZ приводил к распределениям олигомеров, практически идентичным распределениям для необработанного Aβ1-42, что согласуется по существу с полным ингибированием олигомеризации (смотрите дорожки 6 (обработанные) и 2 (не обработанные) фиг.15B). Повышение отношения соединение:пептид в десять раз приводило к сходным уровням ингибирования (фиг.15B, дорожка 7). Эти данные указывают на то, что при отношениях ~5:2 или менее может быть достигнуто полное ингибирование олигомеризации Aβ.
В качестве контрольного соединения использовали Med1, неактивную полициклическую молекулу со структурой, отличающейся от MNG-AZ. Как показано на фиг.15A и 15B, дорожки 4, олигомеризация Aβ1-40 и Aβ1-42 в присутствии Med1 приводила к распределениям олигомеров, неотличимым от распределений для каждого пептида отдельно. Повышение отношения соединение:пептид в десять раз показало сходные распределения олигомеров (фиг.15A и 15B, дорожки 5).
Ранее считалось возможным, что сильное ингибирование олигомеризации Aβ могло быть следствием эффекта ингибитора на саму химическую реакцию PICUP. Для оценки этой возможности, реакции поперченного сшивания также проводили на глутатион-S-трансферазе (GST; ~26 кДа), положительном контроле для химической реакции поперечного сшивания. Не поперечно-сшитый GST проявлял интенсивную полосу мономера и относительно слабую полосу димера (фиг.15C, дорожка 2). Поперечное сшивание приводило к интенсивной полосе димера, которая была ожидаемой, поскольку GST обычно существует в качестве гомодимера, а также поперечно-сшитых молекул более высокого порядка. В присутствии Med1 не наблюдали изменений поперечного сшивания GST при любом из двух протестированных отношений соединение:белок, 1:1 (фиг.15C, дорожка 4) или 10:1 (фиг.15C, дорожка 5). Количественно сходное распределение также наблюдали для MNG-AZ при отношении 1:1 (фиг.15C, дорожка 6). Существенный эффект MNG-AZ на олигомеризацию GST наблюдали только при отношении 10:1, которое в 4-8 раз превышало наиболее высокое отношение концентраций, использованное в экспериментах с Aβ. Этот эффект может быть следствием прямых эффектов соединения:GST или эффектов на химическую реакцию. Однако, эффект на химическую реакцию не может объяснить сильное ингибирование олигомеризации Aβ1-40 и Aβ1-42 и отсутствие сильного ингибирования олигомеризации GST, наблюдаемого на дорожке 6 фиг.15 (а также отсутствие ингибиторной активности в других анализах). Это обеспечивает убедительные доказательства того, что MNG-AZ эффективно ингибирует олигомеризацию как Aβ1-40, так и Aβ1-42.
Спектры кругового дихроизма. Эффект MNG-AZ на вторичную структуру Aβ и его олигомеры исследовали по спектрам CD (фиг.3). Aβ1-40 и Aβ1-42, когда их инкубировали по отдельности, давали исходные спектры, характерные для в большой степени неупорядоченных конформеров (фиг.3A и 3C). Основным признаком этих спектров был большой минимум с центром на уровне ~198 нм. В процессе инкубации в течение последующих трех суток, происходил большой конформационный переход, который в конечном итоге приводил к совокупной конформеров смешанного характера α-спирали и β-слоя (смотрите перегибы при ~195, ~210 и ~220 нм). Напротив, в присутствии MNG-AZ переходов не наблюдали (фиг.3B и 3D). Все спектры обработанных MNG-AZ Aβ1-40 и Aβ1-42 выявили совокупности конформеров, которые были в большой степени дезорганизованными, указывая на то, что MNG-AZ успешно препятствовал образованию вторичных структур, особенно β-слоев, Aβ, которые вовлечены в аномальную агрегацию Aβ.
Связывание ThT. Связывание ThT использовали для определения уровня структуры β-слоя в препаратах Aβ1-40 и Aβ1-42. В отсутствие соединений, Aβ1-40 имел псевдо-сигмоидальную кривую связывания, характеризующуюся периодом задержки приблизительно 2 суток, и периодом последовательного повышения связывания ThT приблизительно 3 суток (что коррелировало с образованием фибрилл), и с плато связывания приблизительно через 5 суток (фиг.4). Когда Aβ1-40 инкубировали с Med1, в соотношении соединение:пептид либо 1:4, либо 5:4, кривые связывания были идентичны кривым связывания для необработанного пептида, в пределах экспериментальной ошибки (фиг.4A). Напротив, значимых эффектов достигали с помощью MNG-AZ (фиг.4B). Они включали зависимое от концентрации MNG-AZ увеличение времени задержки, снижение темпов роста β-слоя, и снижение конечных уровней β-слоя (таблица 1). При использовании более высокой (25 мкМ) концентрации MNG-AZ наблюдали практически полное ингибирование сборки Aβ1-40.
|
Необработанные и обработанные Med1 образцы Aβ1-42 имели сборку, аналогичную сборке Aβ1-40. (фиг.4C). В пределах временной разрешающей способности анализа, наблюдали небольшое время задержки или не наблюдали времени задержки появления флуоресценции, которая возрастала псевдо-линейным образом в течение 4 суток, а затем оставалась постоянной. Эффект MNG-AZ на сборку Aβ1-42 был еще большим, чем эффект на сборку Aβ1-40. Когда Aβ1-42 инкубировали с MNG-AZ при отношении соединение:пептид 1:2, наблюдали задержку 1 сутки, и максимальное связывание ThT, которое происходило только через 1 сутки, было в шесть раз ниже, чем максимальное связывание ThT необработанным пептидом (фиг.4D; таблица 1). Однако при отношении соединение:пептид 5:2, не наблюдали образования β-слоя. Таким образом, MNG-AZ ингибировал образование β-слоя как Aβ1-40, так и Aβ1-42, зависимым от концентрации образом и ингибирование было значительно более эффективным для Aβ1-42.
Результаты EM. Параметры вторичной структуры коррелировали с состоянием сборки Aβ, однако сами по себе они не определяли четвертичные структуры сборки, которые лучше наблюдались с помощью электронной спектроскопии (фиг.5). Классические амилоидные фибриллы наблюдали в образцах необработанных Aβ1-40 и Aβ1-42 (фиг.5A и 5B, соответственно). Фибриллы Aβ1-40 представляли собой неразветвленные, спиральные нити диаметрами ~7 нм, которые проявляли периодичность спирали ~220 нм. Aβ1-42 формировал неразветвленные нити шириной ~8 нм и с различными степенями спиральности. Кроме того, для сборки Aβ42 наблюдали более толстые прямые неразветвленные нити шириной ~12 нм. При более низкой концентрации MNG-AZ (5 мкМ), фибриллы были тоньше (4 против 8 нм), чем фибриллы, образованные необработанным Aβ (фиг.5C). Кроме того, наблюдали множество небольших относительно аморфных агрегатов. Обработка Aβ1-40 25 мкМ MNG-AZ значительно снижала количество фибрилл и повышала относительные количества коротких фибрилл и аморфных агрегатов (фиг.5E). Эффекты MNG-AZ на сборку Aβ1-42 были сходны в том, что количество и длина фибрилл снижались и возрастала частота аморфных агрегатов (фиг.5D и 5F).
Метаболизм MTT (анализ токсичности). Результаты анализа токсичности Aβ проиллюстрированы на фиг.6. На фиг.6A-6D, группы слева соответствуют результатам токсичности после инкубации в течение 0 суток (для обоих пептидов Aβ1-40 и Aβ1-42); группы в центре соответствуют результатам токсичности после инкубации в течение 3 суток для Aβ1-40 (фиг.6A и 6B) или инкубации в течение 2 суток для Aβ1-42 (фиг.6C и 6D); группы справа соответствуют результатам токсичности после инкубации в течение 7 суток (для обоих пептидов). В одной группе экспериментов (фиг.6A и 6C), пептиды коинкубировали с Med1 или MNG-AZ. Во второй группе экспериментов (фиг.6B и 6D), Med1 и MNG-AZ добавляли после инкубации и непосредственно перед добавлением смесей к дифференцированным клеткам PC12.
Было показано, что Aβ1-40 является токсичным для клеток, и токсичность составила ~20% от токсичности фибрилл (фиг.6A). Смесь Aβ1-40 и Med1 также была токсичной ~20%. Напротив, MNG-AZ приводил к тому, что Aβ1-40 был нетоксичным. Инкубация Aβ1-40 в течение 3 суток, в течение которых образуются олигомеры, протофибриллы и фибриллы, приводила к композиции, которая была значительно более токсичной (~60%, средняя группа фиг.6A). Обработка Aβ1-40 посредством MNG-AZ снижала эту токсичность до <10% (p<0,005). Ту же качественную взаимосвязь между тремя экспериментальными группами наблюдали после инкубации в течение 7 суток (правая группа фиг.6A): как необработанные, так и обработанные Med1 Aβ1-40 были токсичными на ~35-40%, в то время как обработанный MNG-AZ Aβ1-40 был токсичным на <10%. Сходные наблюдения были сделаны в экспериментах с Aβ1-42 (фиг.6C). Aβ1-42 был более токсичным, чем Aβ1-40, во все моменты времени.
Эффекты MNG-AZ на индуцированную Aβ1-40 токсичность по мере прогрессирования сборки пептида проиллюстрированы на фиг.6B. Во все три периода времени инкубации, необработанный и обработанный Med1 пептид давал сходные уровни токсичности. Напротив, обработанный MNG-AZ пептид был либо нетоксичным (0 сутки) или значительно менее токсичным (3 или 7 сутки). Качественно сходные результаты были получены в исследованиях Aβ1-42 (фиг.6D). Эффекты MNG-AZ на Aβ1-42 были более выраженными, чем в случае Aβ1-40.
Указанные выше результаты согласованно указывают на то, что MNG-AZ эффективно ингибирует олигомеризацию Aβ и его токсичность в головном мозге. Эти данные доказывают, что MNG-AZ может предупреждать или лечить заболевания, ассоциированные с неправильным сворачиванием, накоплением, агрегацией или отложением Aβ.
Пример 7. Эффект in vivo экстракта виноградных косточек в модели таупатий на трансгенных Drosophila и мышах
Настоящий пример иллюстрирует пользу in vivo композиции согласно одному варианту осуществления настоящего изобретения на фенотипах Drosophila, обладающих мутантным тау (R406W), которые моделируют определенные формы таупатии, и на модели таупатии на трансгенных мышах JNPL3.
Материалы и способы
Исследование фенотипа глаз мух ey>R406W. Это исследование представляло собой продолжение исследования, представленного в примере 5. В кратком изложении, яйца ey>R406W откладывали и выводили на среде для мух быстрого приготовления 4-24, дополненной 2,8 мкг/мл GSE MNG-AZ (или GSPE, фиг.21) или на контрольном корме, дополненном эквивалентным объемом воды (растворитель GSE, контроль в виде носителя). Мухам вводили GSE (или носитель) постоянно до взрослого состояния. Глаза Drosophila наблюдали под микроскопом.
Оценка нарушения двигательной функции и смертности у мышей JNPL3. Модель на мышах JNPL3 создавали для экспрессии семейного мутантного тау P301L человека, который приводит к связанной со старением нейродегенерации, которая отражается двигательной дисфункцией. Мышам JNPL3 вводили 150 мг/кг массы тела/сутки GSE MNG-AZ, начиная с возраста приблизительно семь месяцев, что раньше возникновения опосредуемого мутантным тау двигательного нарушения, которое, как правило, начинает развиваться к возрасту ~12 месяцев. Для оценки нарушения двигательной функции использовали тест вытягивания задних конечностей (проиллюстрированный на фиг.22), исходя из оценочной системы на основе 4 точек.
Результаты и обсуждение
Введение GSE подавляло аномальные фенотипы глаз у мух с мутантным тау R406W. Глаза мух с мутантным тау R406W характеризуются уменьшенным размером и аномальной морфологией (фиг.21B и 21E) по сравнению со взрослыми мухами дикого типа (фиг.21A и 21D). Напротив, мухи с мутантным тау, которым вводили GSE, обладали меньшей аномалией размера глаз (фиг.21C и 21F).
В количественном анализе морфологии глаз взрослых мух с использованием четырехточечной оценочной системы для изменений морфологии глаз, где 0 означает отсутствие глаза и 4 означает нормальный глаз, было показано, что введение GSE значительно улучшает фенотипы глаз у взрослых самцов и самок мух с мутантным тау R406W в трех независимых испытаниях (фиг.21G) (ANOVA: P<0,0005, F=57,29; DF=1,531; *P<0,05, сравнение мух, которым вводили GSE, и мух, которым его не вводили, в индивидуальных испытаниях).
Введение GSE ослабляло доклинические фенотипы таупатии у мышей JNPL3. Длительное введение GSE не приводило к выявляемым побочным эффектам у мышей JNPL3, включая изменение массы тела или употребления воды (не показано). Введение GSE мышам JNPL3 снижало тяжесть двигательного нарушения, которое обычно происходит при старении в этой модели на мышах (фиг.23A). Аналогично ослаблению двигательного нарушения, введение GSE также значимо снижало смертность мышей JNPL3 относительно контрольной группы JNPL3 без введения (на фиг.23B показано, что уровень смертности мышей JNPL3 был снижен на ~30% через 13 месяцев) (логарифмическая ранговая статистика, p=0,05; смертность: мыши без введения=27%, мыши с введением GSE=0%).
В совокупности, данные in vitro и in vivo, представленные в примерах 4, 5 и 7 указывают на то, что GSE MNG-AZ может благоприятным образом модулировать опосредуемые тау нейропатологические фенотипы, препятствуя неправильному сворачиванию тау в агрегаты тау, что подтверждает потенциальное применение GSE для профилактики и/или лечения связанных с тау нейродегенеративных нарушений, включая AD, прогрессирующий супрануклеарный паралич, кортикобазальную дегенерацию, болезнь аргирофильных гранул, болезнь Пика, FTDP-17 и т.д. Более того, доказательства того, что GSE препятствует как опосредуемым Aβ (как проиллюстрировано в примерах 1-3 и 6), так и опосредуемым тау, нейропатологическим механизмам, строго подтверждают возможность применения MNG-AZ в профилактике и/или лечении AD.
Пример 8. Эффект in vitro экстракта виноградных косточек на опосредуемые htt с полиглутамином нейропатологические фенотипы
В данном примере проиллюстрированы эффекты in vitro композиции согласно одному варианту осуществления настоящего изобретения на агрегацию содержащих полиглутамин молекул белка htt.
Материалы и способы
Молекулы белка htt получали с использованием клеточной линии PC-12, содержащей индуцируемый экдизоном содержащий полиглутамин слитый белок Htt, содержащий первые 17 аминокислот белка htt и 103 остатка глутамина, слитые с усиленным GFP (Htt103Q-EGFP), которая экспрессирует слитый белок htt при индукции аналогом экдизона, муристероном A, и образует флуоресцентные агрегаты htt (Apostol et al., Proc Nat Acad Sci 2003; 100:5950-5955). Слитый белок GFP-Htt индуцировали 0,2 мкМ муристероном A, и агрегацию слитого белка GFP-Htt в отсутствие и в присутствии обработки GSE MNG-AZ (12,5 мкМ и 25 мкМ) оценивали флуоресцентной микроскопией и вестерн-блот-анализом. Накопление агрегатов htt в отсутствие и в присутствии GSE отражалось флуоресцентным испусканием после привлечения слитого белка GFP-Htt в агрегаты.
Результаты и обсуждение
Обработка GSE значимо снижала накопление флуоресцентных агрегатов htt дозозависимым образом: более высокая дозировка обработки GSE приводила к более выраженному снижению высокомолекулярных агрегатов Htt (фиг.24A). Этот результат также подтверждается независимым вестерн-блот-анализом (фиг.24B).
Указанные выше результаты обеспечивают доказательства in vitro, что MNG-AZ может предотвращать опосредуемые содержащим полиглутамин htt нейропитологические фенотипы HD.
Пример 9. Эффект экстракта виноградных косточек in vivo на болезнь Гентингтона на модели трансгенных Drosophila и мышах
Настоящий пример иллюстрирует пользу in vivo композиции согласно одному варианту осуществления настоящего изобретения в модели HD на Drosophila elav>Q93httexon1, и в модели HD на трансгенных мышах JNPL3 R6/2.
Материалы и способы
Оценка двигательной функции и смертности мух с HD. В этих исследованиях использовали модель HD на Drosophila elav>Q93httexon1. Эта модель HD вовлекает регуляторную систему elav-Gal4/UAS для достижения селективной, общей нейрональной сверхэкспрессии укороченного мутантного белка htt человека, кодируемого экзоном 1 гена htt человека, который содержит 93 полиглутамильных остатка (Sang et al., NeuroRx, 2005; 2:438-446). Это приводит к возникающей во взрослом состоянии нейродегенерации, включая разрушение фоторецепторных клеток глаза, нарушение способности подниматься и снижение продолжительности жизни (Sang et al., 2005, выше).
Для оценки двигательного нарушения, взрослых мух elav>Q93httexon1 собирали в течение одних суток после вылупления (появления взрослой мухи из ее куколочной оболочки) и помещали по 10 на колбу на контрольном корме или на корме с добавлением GSE MNG-AZ (n=30 на группу). На 9 сутки и 16 сутки, соответственно, оценивали двигательную активность, осторожно постукивая по дну колбы с мухами и осуществляя мониторинг процента мух, успешно поднявшихся по стенке колбы выше заданной высоты (например, до отметки 7 см на 9 сутки или 2 см на 16 сутки) в течение 8 секунд. При оценке уровня смертности, самцов мух Elav>Q93httexon1 собирали в пределах одних суток после вылупления и помещали по 10 на флакон на стандартный (контрольный) корм или корм с добавлением GSE. Живых мух подсчитывали каждые сутки.
Оценка двигательных функций и смертности мышей с HD. Эти исследования проводили с использованием модели HD на мышах R6/2, первоначально полученных Dr. Bates и его коллегами (Mangiarini et al. Cell, 1996; 87:493-506), которая являются наиболее часто используемой моделью трансгенных мышей с HD. Мыши R6/2 экспрессируют фрагмент экзона 1 htt, который содержит 148-153 повтора полиглутамина. Регуляция мутантного htt запускается промотором htt человека. Мыши R6/2 проявляют очень агрессивный неврологический фенотип и обеспечивают явные экспериментальные конечные точки, которые идеальны для предварительного доклинического исследования (Ramaswamy et al. ILAR J, 2007; 48:356-73).
Самок мышей с трансплантированным яичником, полученных от Jackson's Laboratories, случайным образом разделяли на две группы (группа, в которой вводили 100 мг/кг/сутки GSE MNG-AZ, и контрольная группа, в которой вводили H2O) и скрещивали с самцами мышей дикого типа. Отлученных от молока детенышей постоянно кормили с одинаковыми режимами введения GSE или контроля.
Для оценки влияния введения GSE на изменение координации движений у мышей R6/2 в процессе связанного со старением развития и прогрессирования фенотипов HD использовали тесты с вращающимся стержнем. Трансгенных мышей с HD тренировали, чтобы они оставались на узком стержне в устройстве с ускоряющимся стержнем (4 об./мин. - 40 об./мин. в течение 10 минут) в возрасте 6 недель, и мониторинг параметров на вращающемся стержне проводили один раз в неделю, начиная с возраста 8 недель. Для каждого данного теста проводили три испытания и регистрировали среднее значение для трех испытаний. Утрата двигательной функции отражается сниженным периодом перед падением животного с устройства. При оценке смертности мышей R6/2, мышам вводили либо 100 мг/кг/сутки GSE MNG-AZ, либо H2O (контрольная группа), и уровень выживания регистрировали каждые сутки.
Результаты и обсуждение
Двигательная функция и смертность в модели HD на Drosophila. При оценке двигательной активности мух elav>Q93httexon1 на 9 сутки, когда мухи проявляли фенотипы мягкой HD, было выявлено, что GSE улучшает двигательную активность в анализе подъема на 9 сутки (~40% контрольных мух без введения и ~50% мух, которым вводили GSE, успешно выполнили задание подъема) (фиг.25A). Однако это наблюдение не достигло статистической значимости. При оценке на 16 сутки, когда у мух elav>Q93httexon1 развивалось более тяжелое двигательное нарушение, значительно больший процент мух, которым вводили GSE, успешно выполнили задание подъема по сравнению с контрольными мухами (~15% мух без введения и ~47% мух, которым вводили GSE, успешно выполнили задание подъема) (фиг.25B). Эти результаты указывают на то, что GSE обладает биоактивностью in vivo и смягчает двигательные нарушения в этой модели HD на Drosophila.
Помимо двигательного нарушения экспрессия мутантного белка htt в этой модели HD на Drosophila приводила к уменьшению продолжительности жизни. Введение GSE продолжали (относительно того, как описано в примере 5) для оценки его влияния на повышение продолжительности жизни в этой модели на Drosophila. В соответствии с предшествующим наблюдением, что введение GSE значимо снижало опосредуемое мутантным Htt двигательное нарушение (фиг.25), введение GSE также на значимом уровне обеспечивало увеличение продолжительности жизни мух elav>Q93httexon1 (анализ Каплана-Мейера выживания мух для всех четырех испытаний продемонстрировал, что введение GSE в значительной степени обеспечивало увеличение продолжительности жизни у мух elav>Q93httexon1: χ2=21,73, df=1, p=0,0001) (фиг.26).
Двигательные функции и смертность у мышей с HD. В исследовании двигательной функции мышей R6/2 не было выявлено поведенческих отличий между контрольной группой и группой введения GSE в возрасте 6 недель, когда животные были, главным образом, в предсимптомном состоянии. Двигательные функции мышей продолжали исследовать в возрасте 9 недель в ходе возникновения двигательных нарушений, и в возрасте 11 недель, когда фенотипы HD прогрессировали до умеренных двигательных нарушений. Введение GSE на значимом уровне улучшало двигательные функции у мышей R6/2 с HD как при возникновении клинического заболевания (в возрасте 9 недель), так и при прогрессировании (в возрасте 11 недель), как показано на фиг.27 (столбики соответствуют среднему значению +SEM времени (с), в течение которого животные были способны оставаться на вращающемся стержне. Статистический анализ проводили с помощью t-критерия Стьюдента, *p<0,05, сравнивая группу введения GSE с контрольной группой без введения).
В этом исследовании смертности мышей R6/2 было выявлено, что введение GSE значимо продлевает выживание мышей R6/2 HD. Средняя продолжительность жизни у мышей R6/2 без введения составила 90 суток, в то время как введение GSE значимо увеличивало среднюю продолжительность жизни мышей HD до 100 суток (анализ Каплана-Мейера продемонстрировал, что введение GSE в обеспечивало значимое увеличение продолжительности жизни у мышей с HD: X2=4,018, df=1, p=0,045) (фиг.28).
Таким образом, указанные выше исследования in vivo с использованием экспериментальных моделей HD, полученных для независимых филогенетически отдаленных видов Drosophila и мыши, продемонстрировали эффективность GSE MNG-AZ в отношении ослабления опосредуемых мутантным Htt патологических фенотипов. В комбинации с доказательствами из исследований in vitro (рассмотренными в примерах 5 и 8), эти данные подтверждают потенциальную ценность применения GSE для профилактики и/или лечения HD.
Все патенты, патентные заявки, публикации, описания продуктов и протоколы, цитированные этом описании, включены в настоящий документ в качестве ссылок в полном объеме. В случае противоречия терминов, следует руководствоваться настоящим описанием.
В то время как очевидно, что изобретение, описанное в настоящем документе, рассчитано на достижение пользы и преимуществ, указанных выше, настоящее изобретение не ограничивается объемом конкретных вариантов осуществления, описанных в настоящем документе. Понятно, что возможны модификации, варианты и изменения изобретения без отклонения от его сущности.
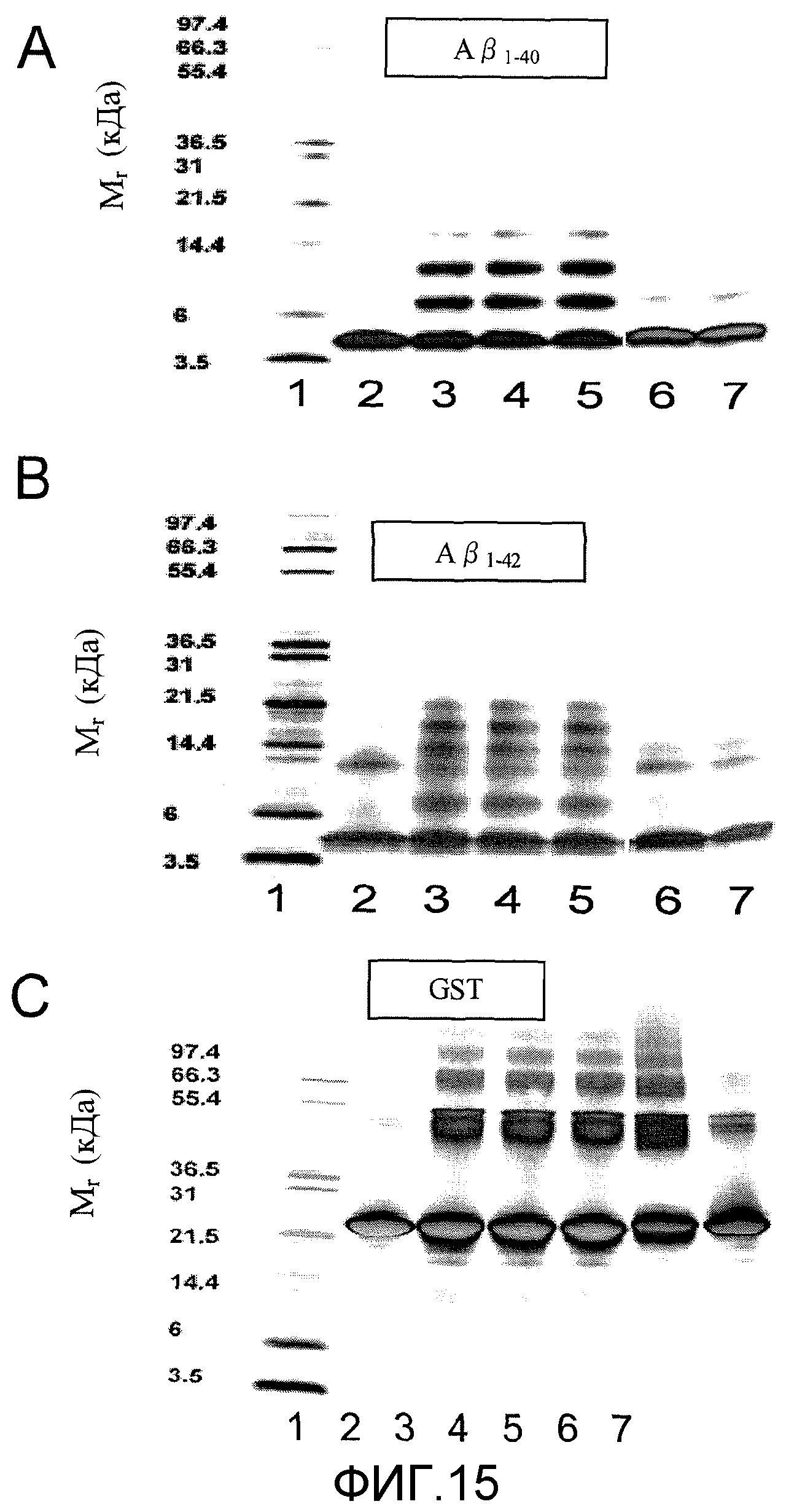
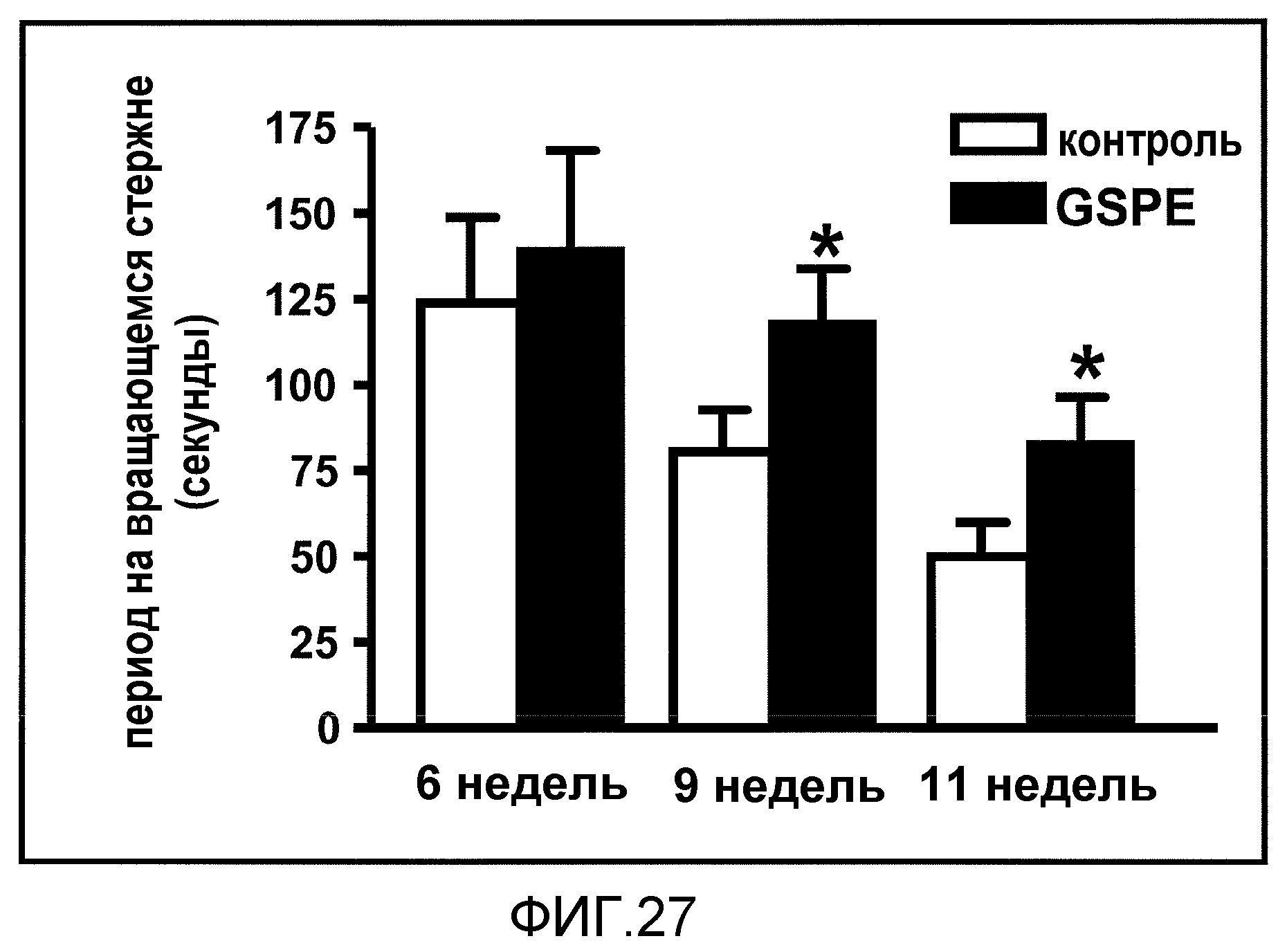
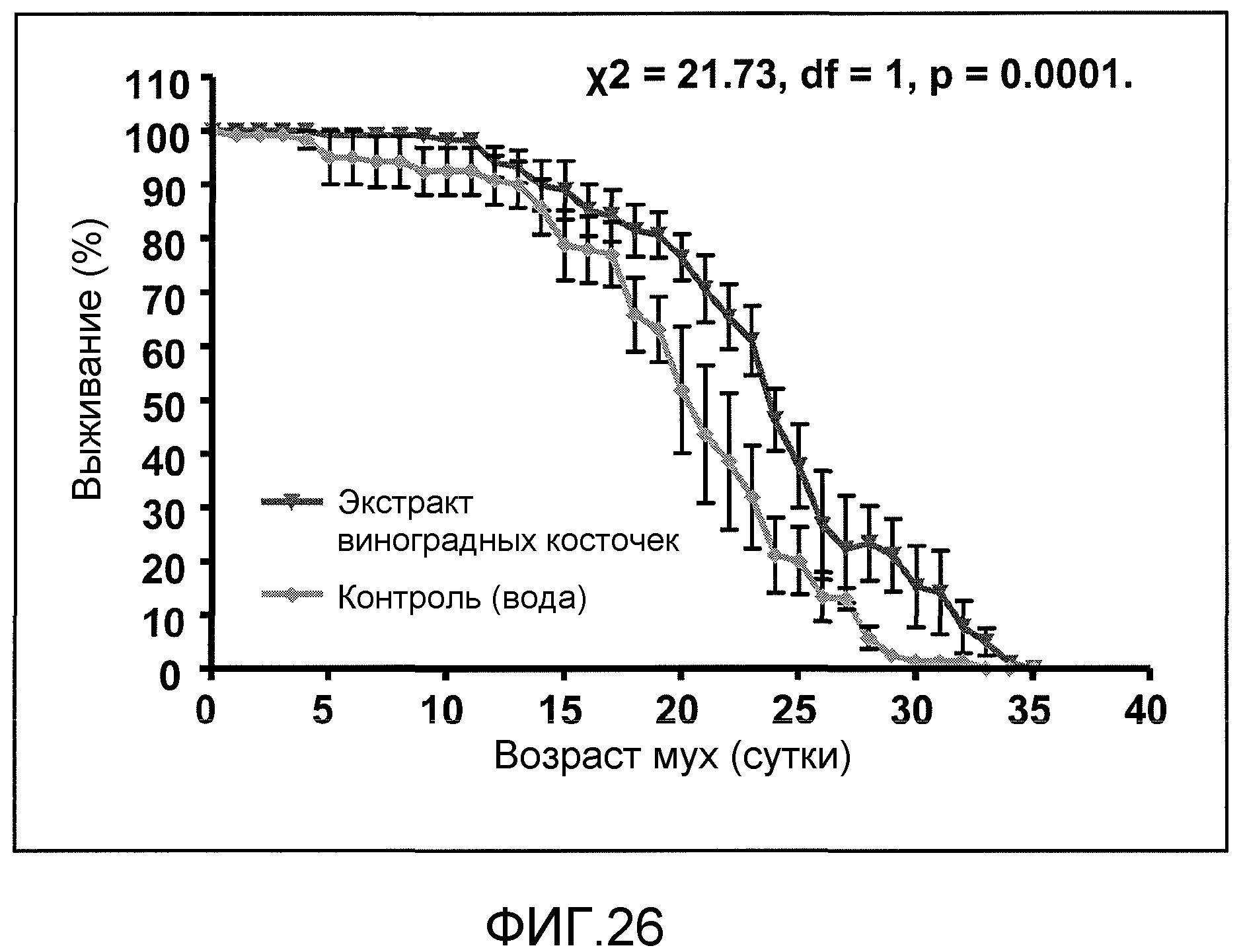
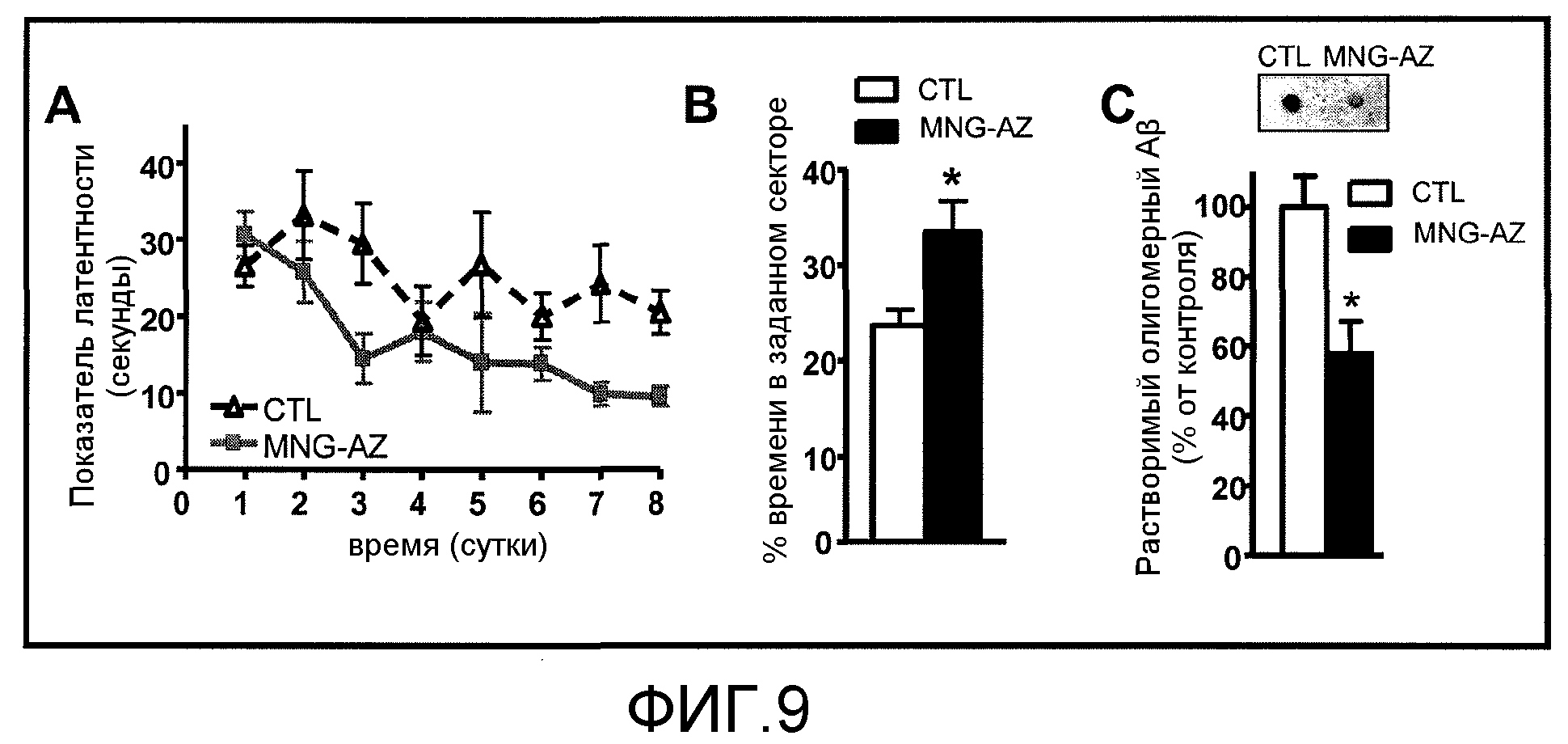
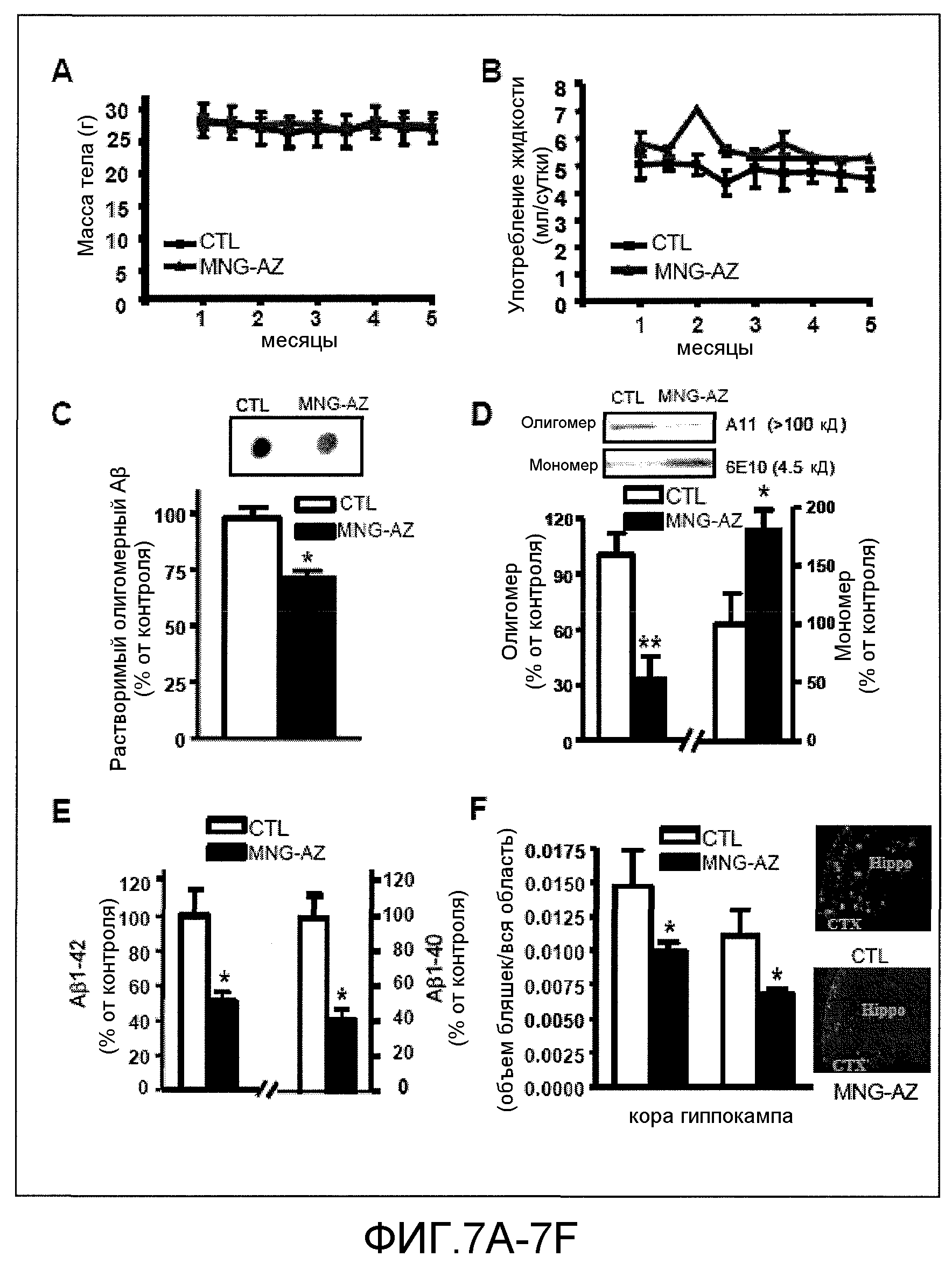
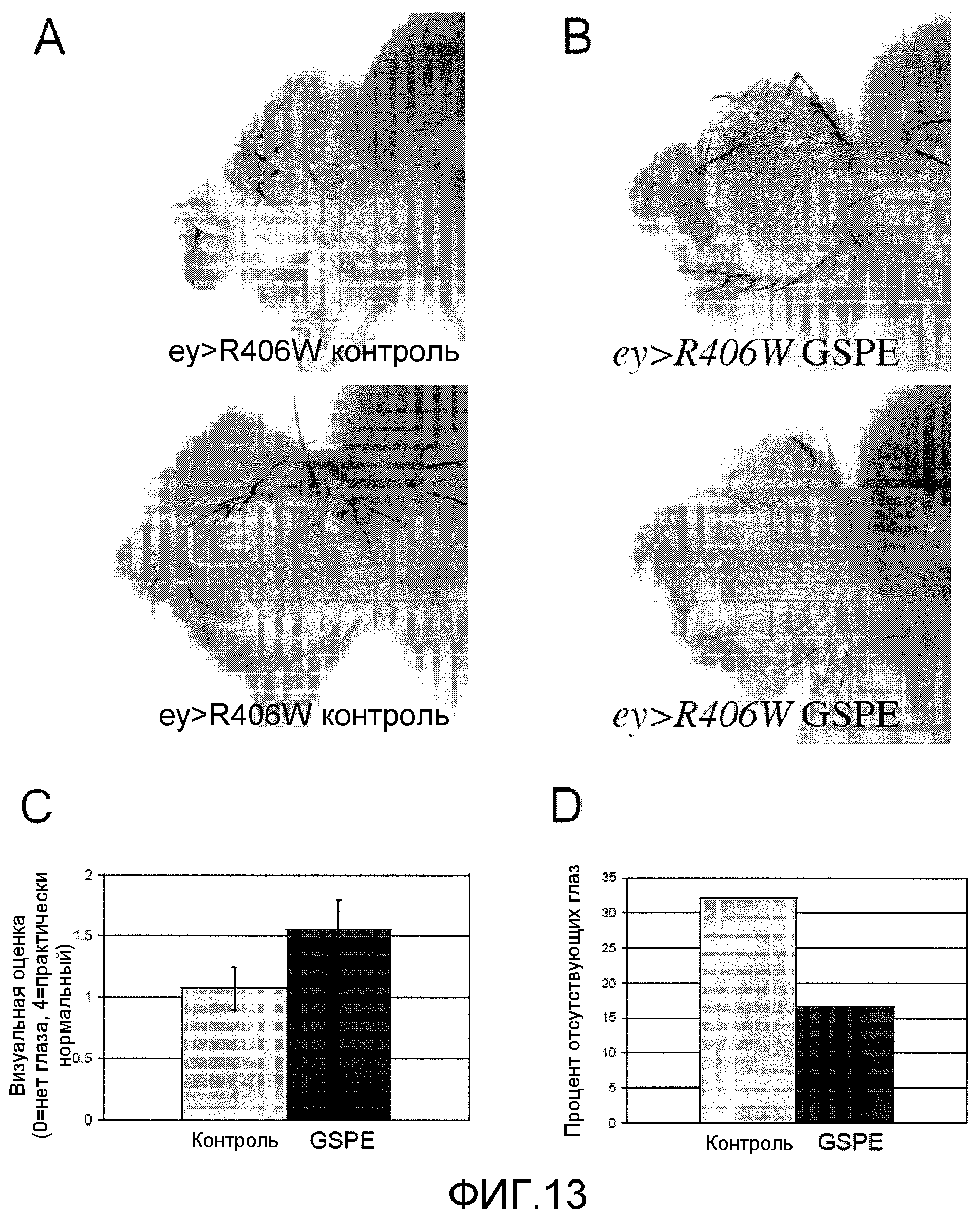
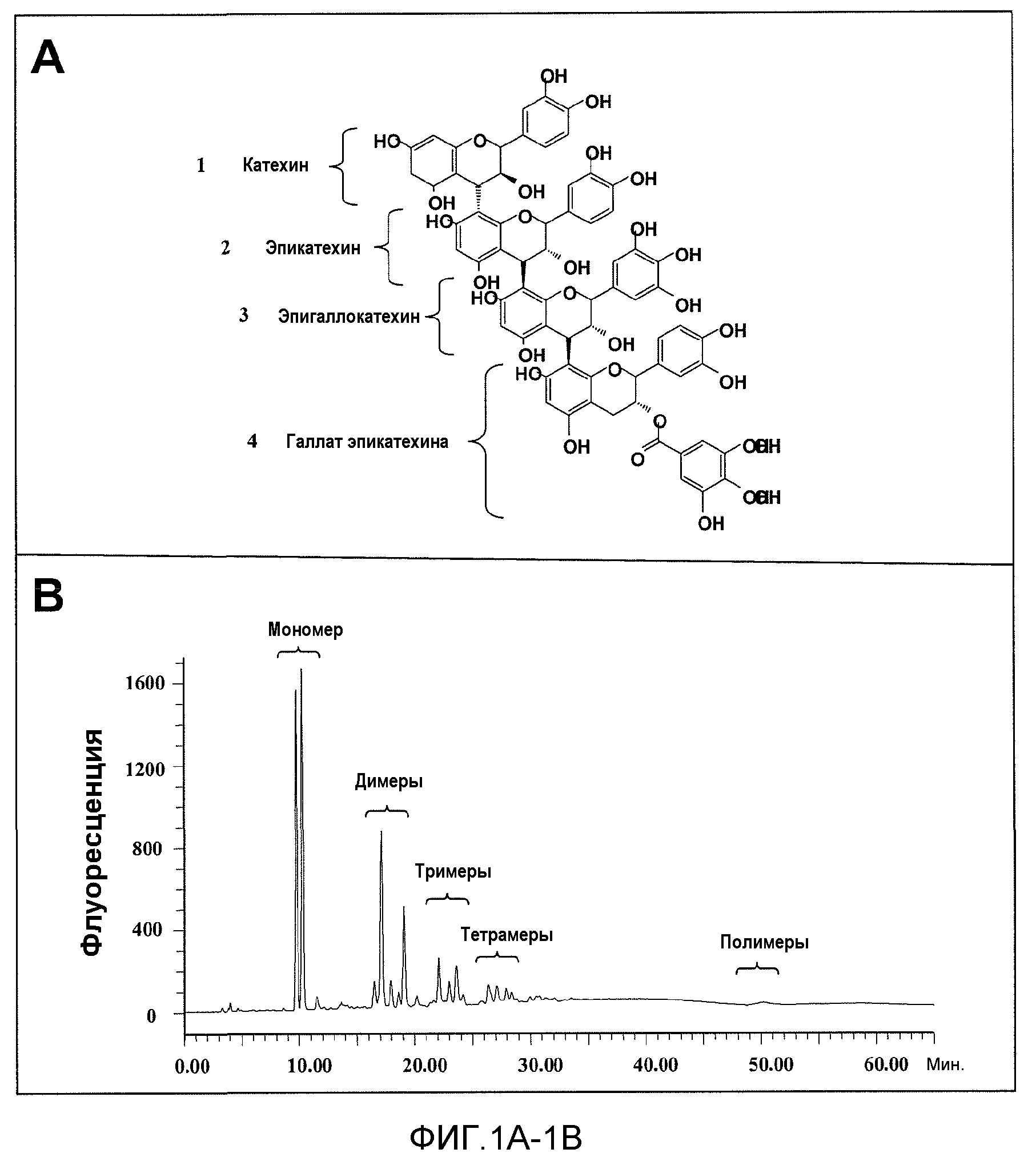
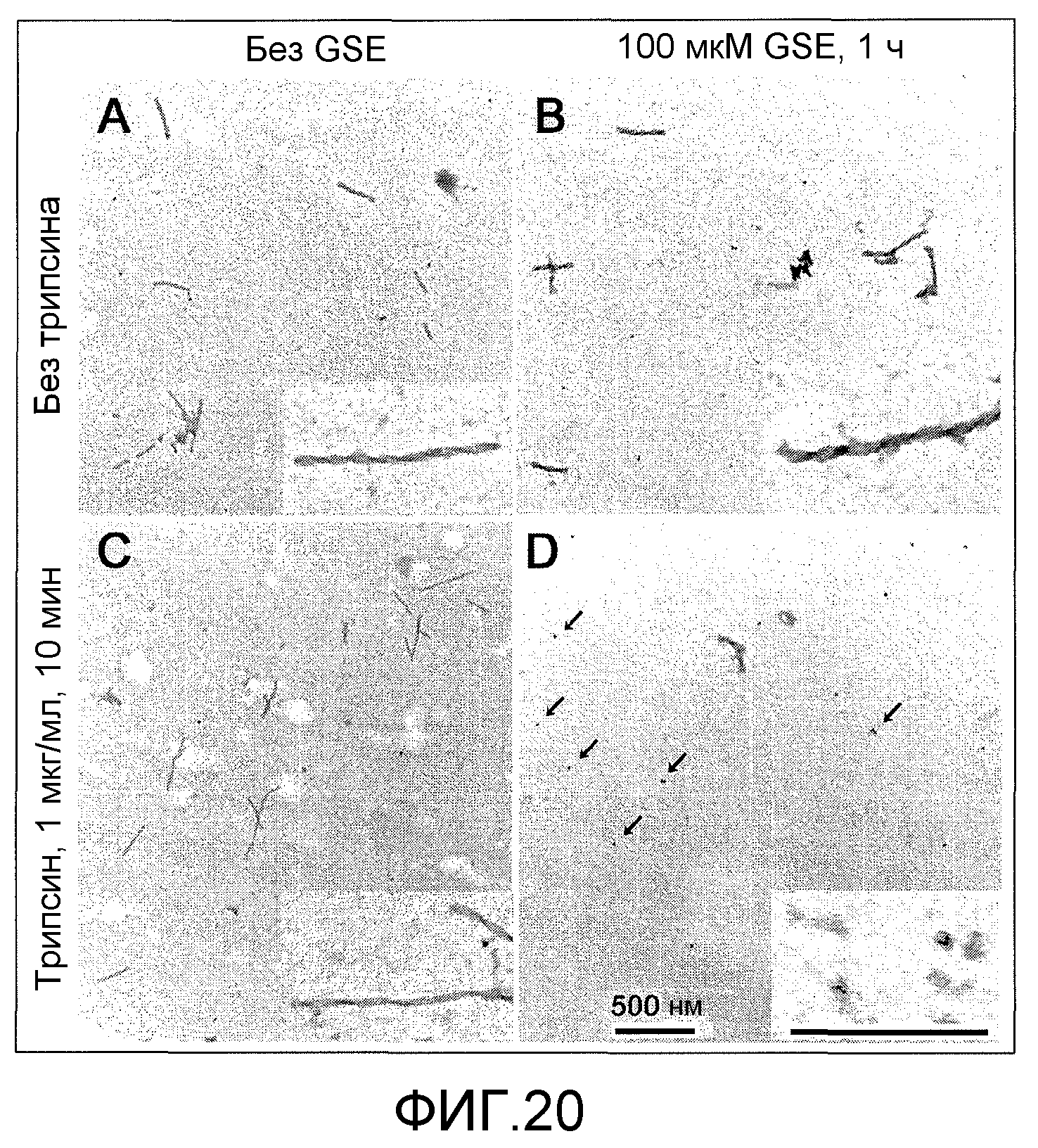
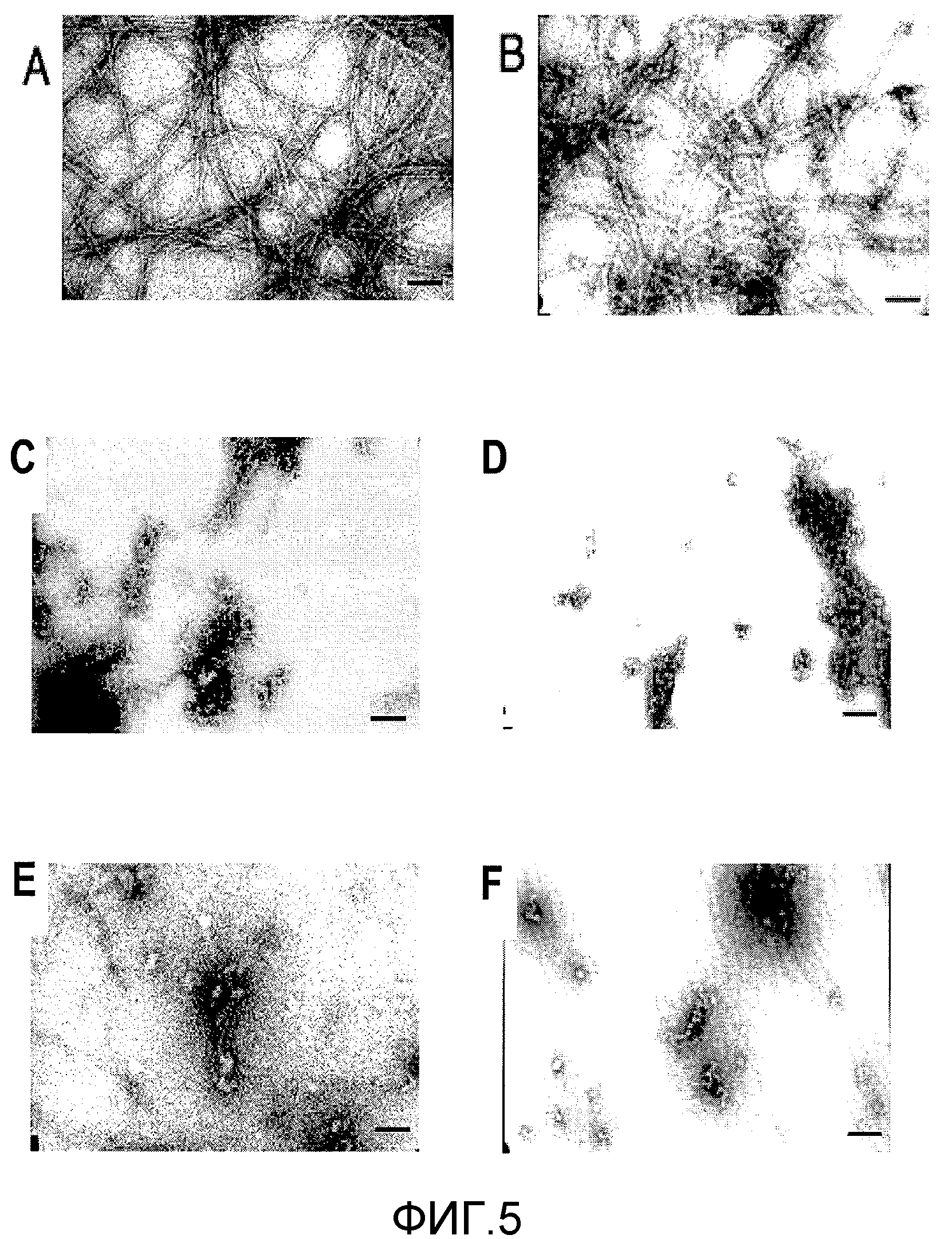
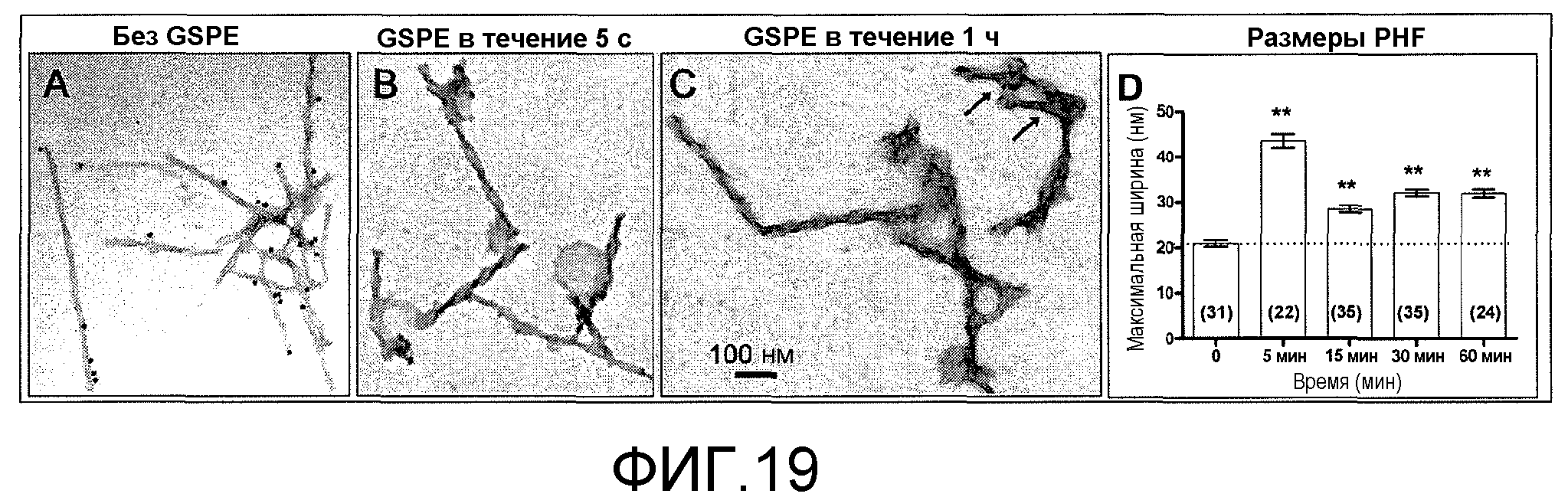
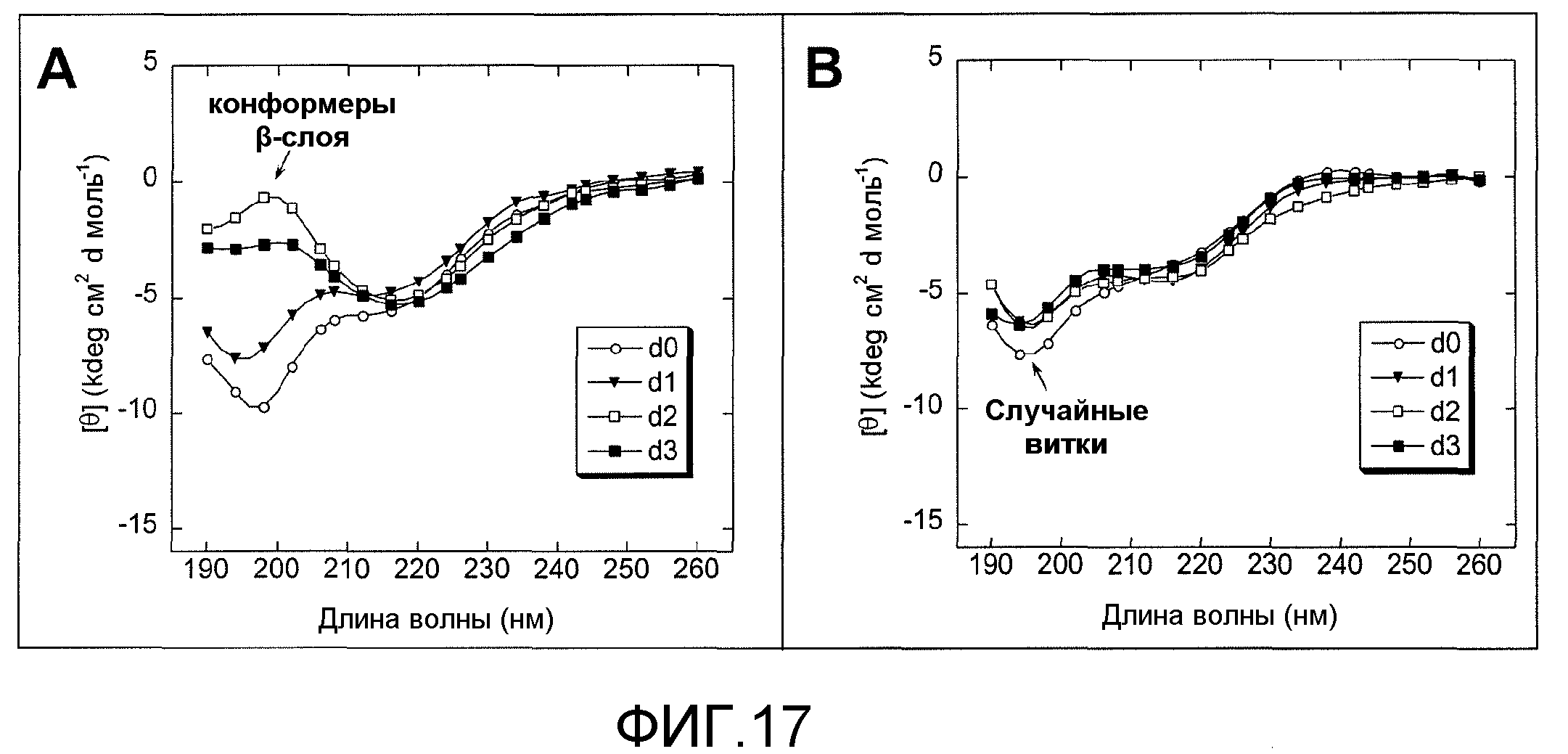

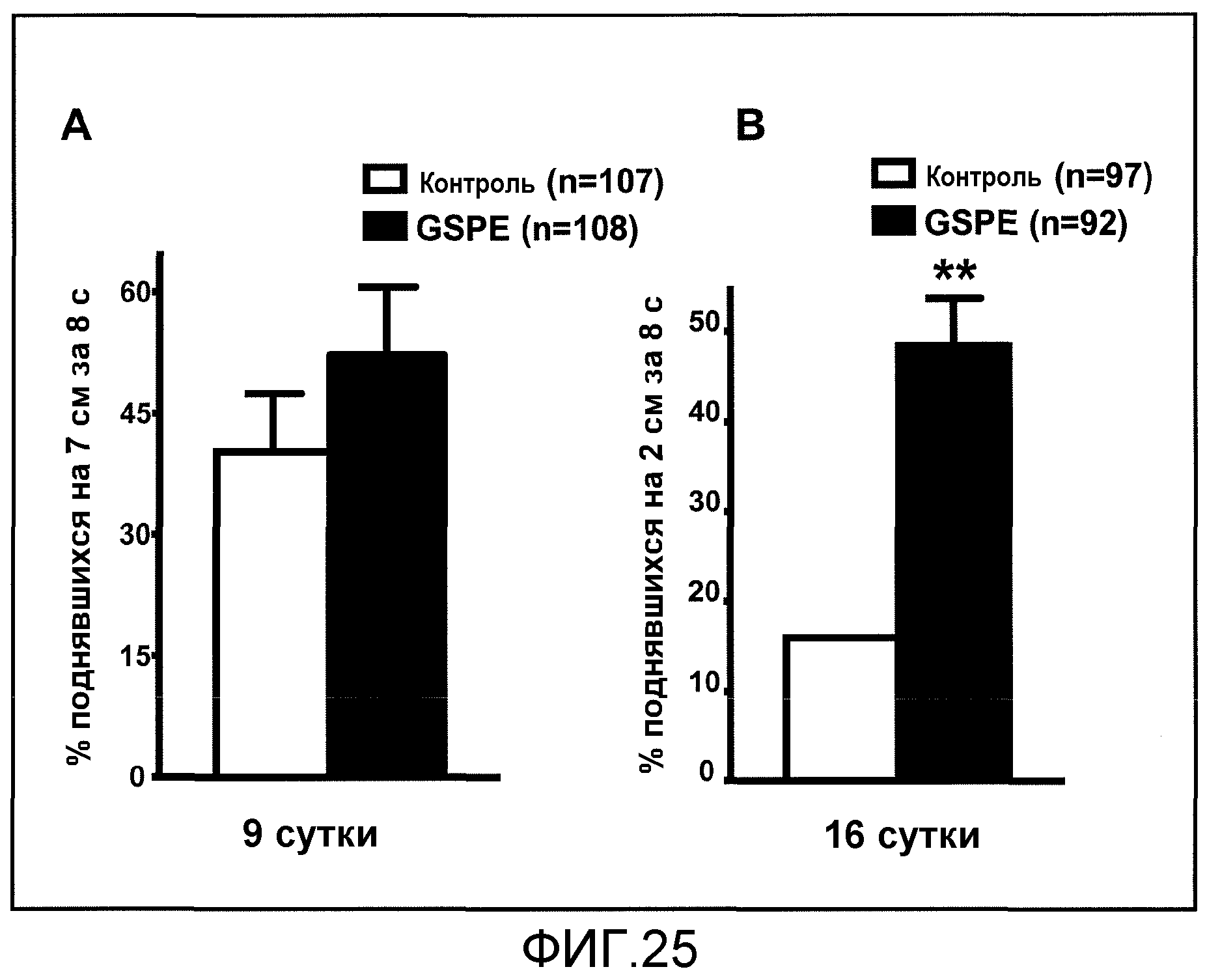
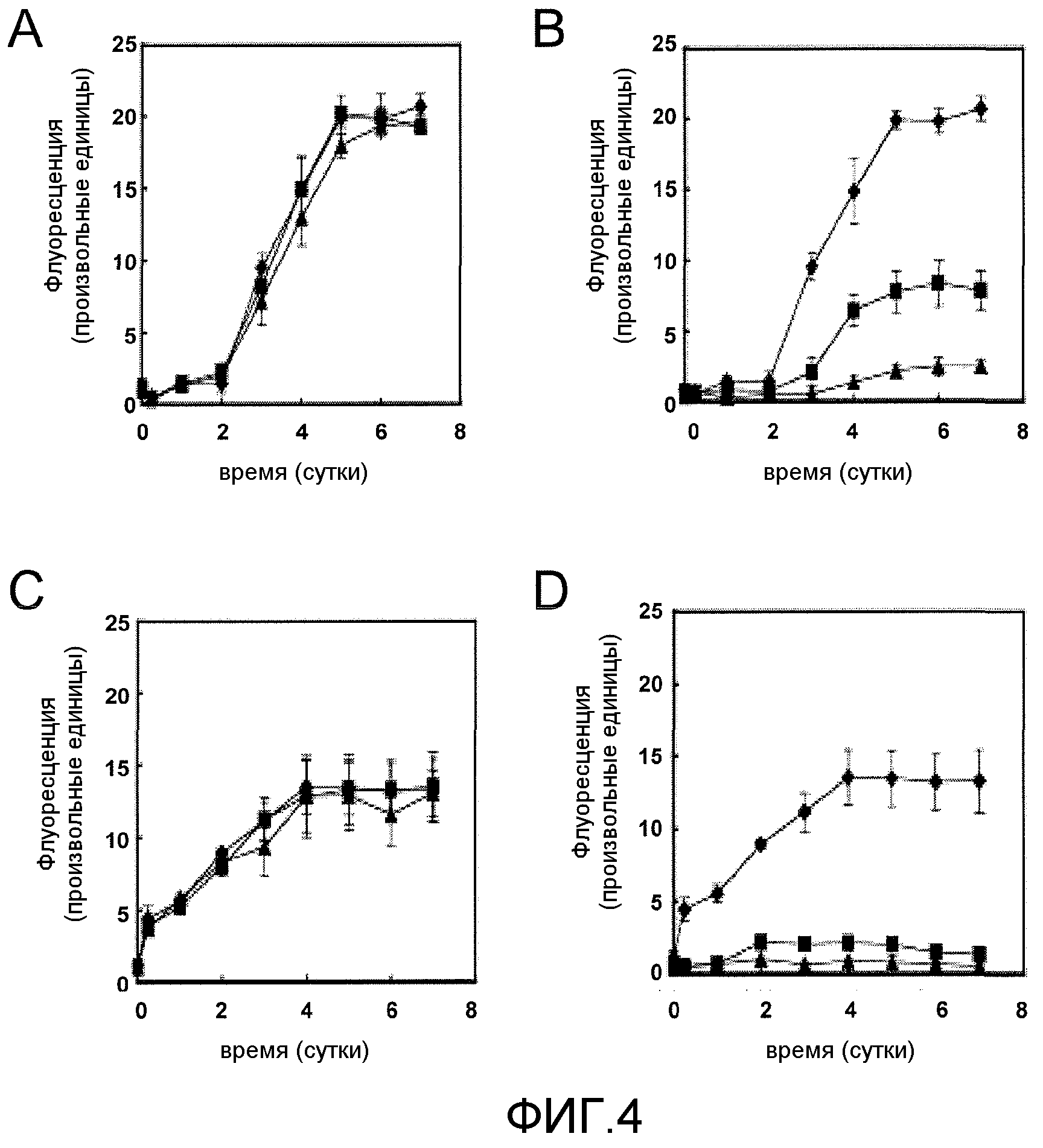
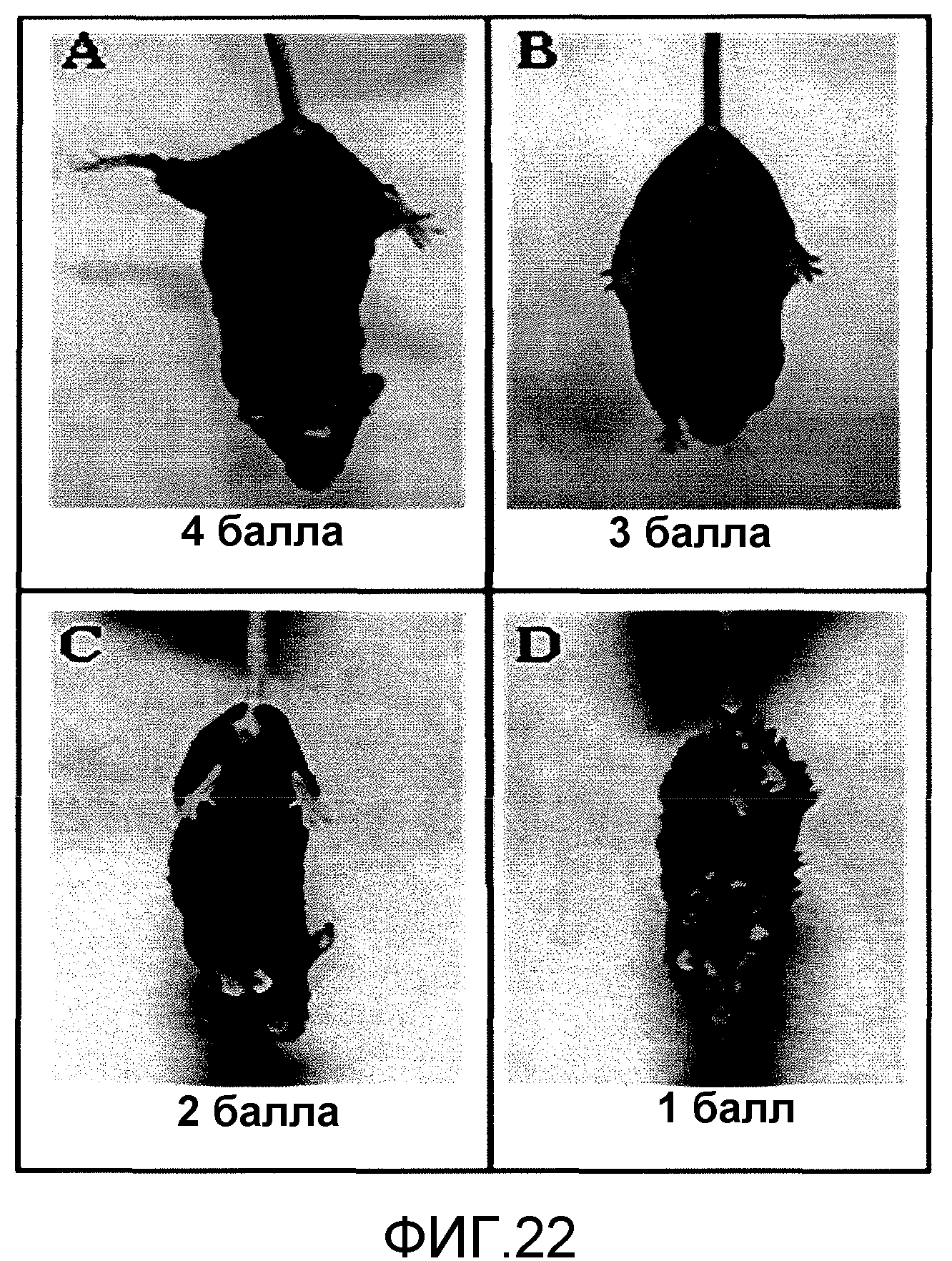

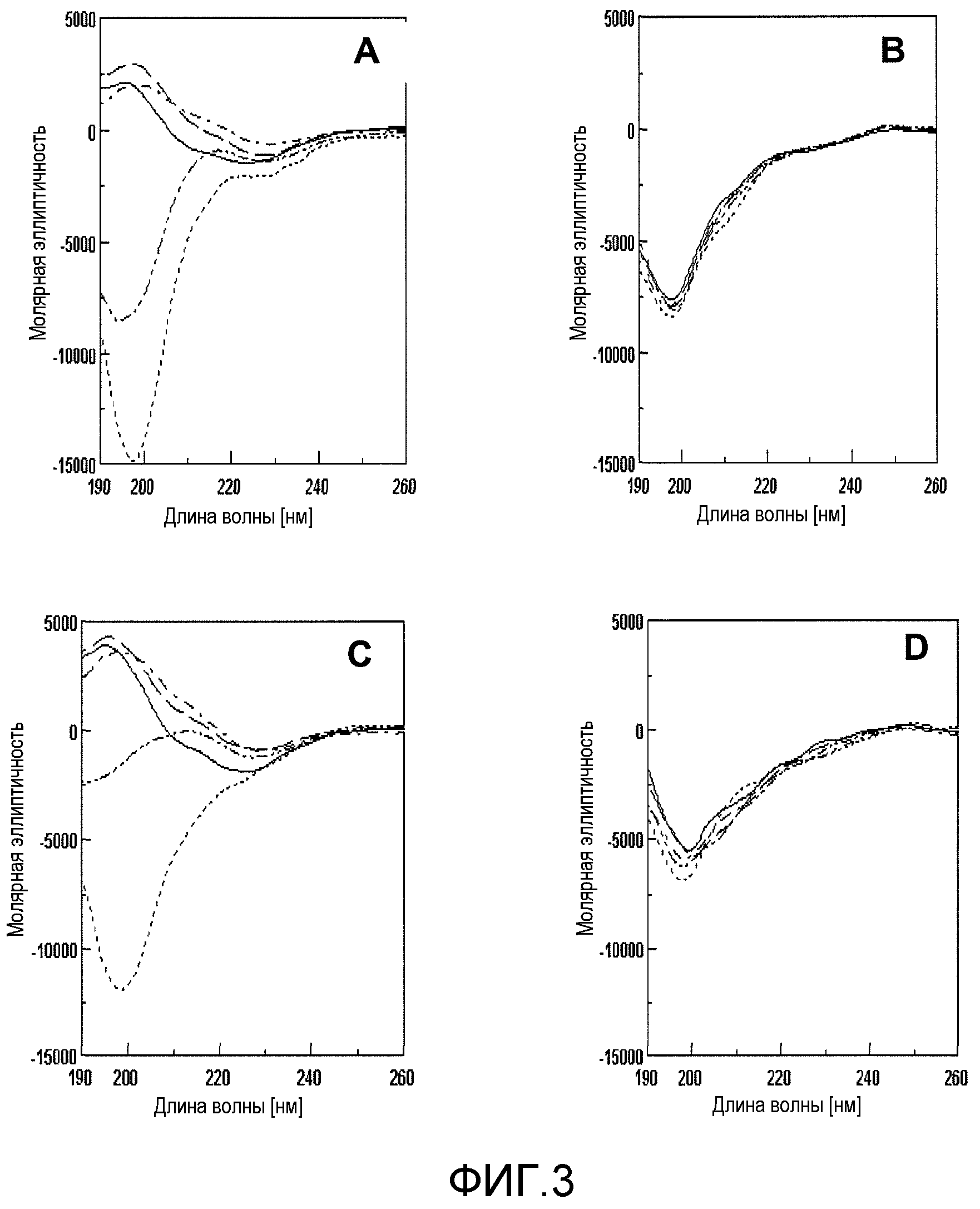
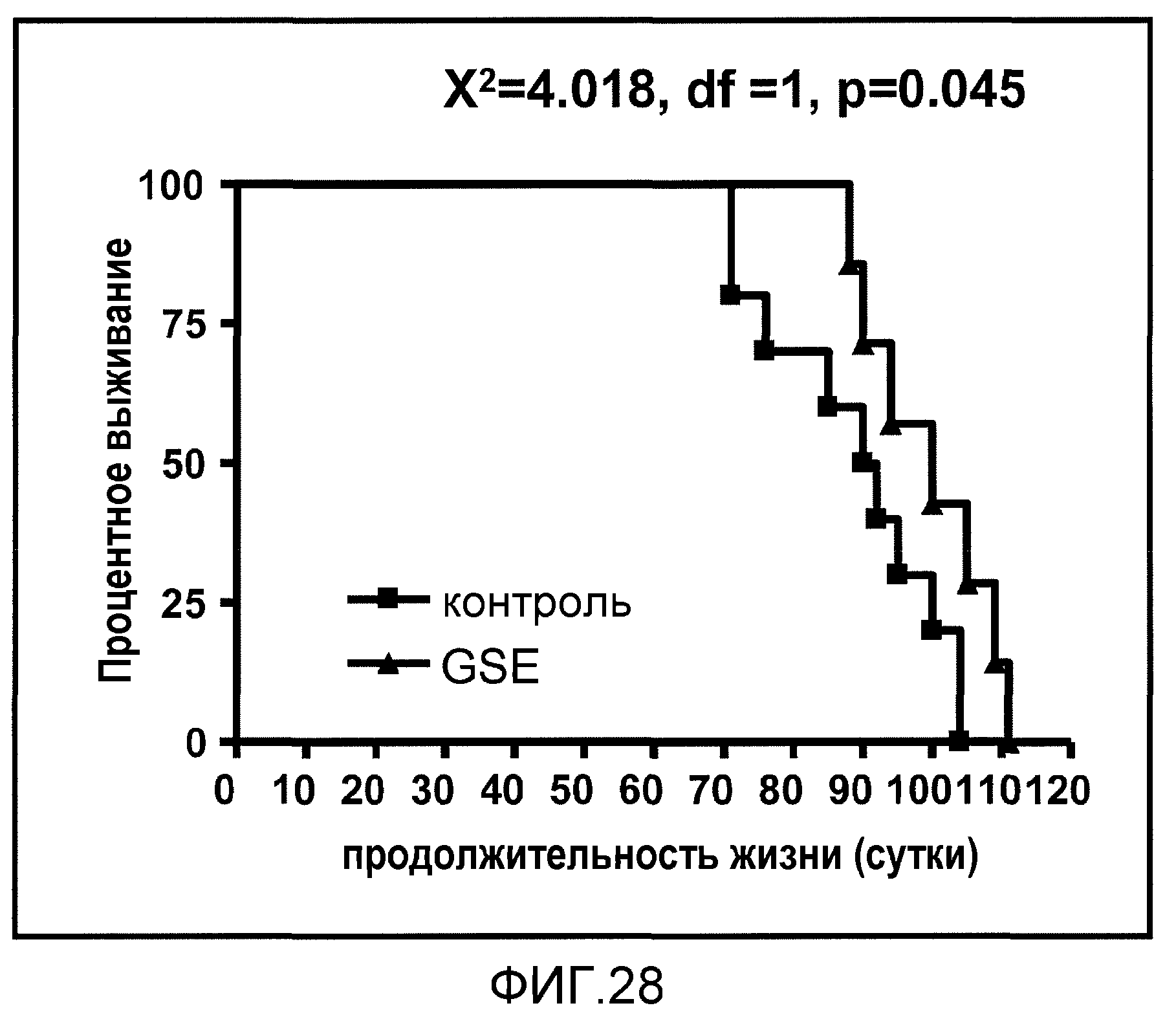
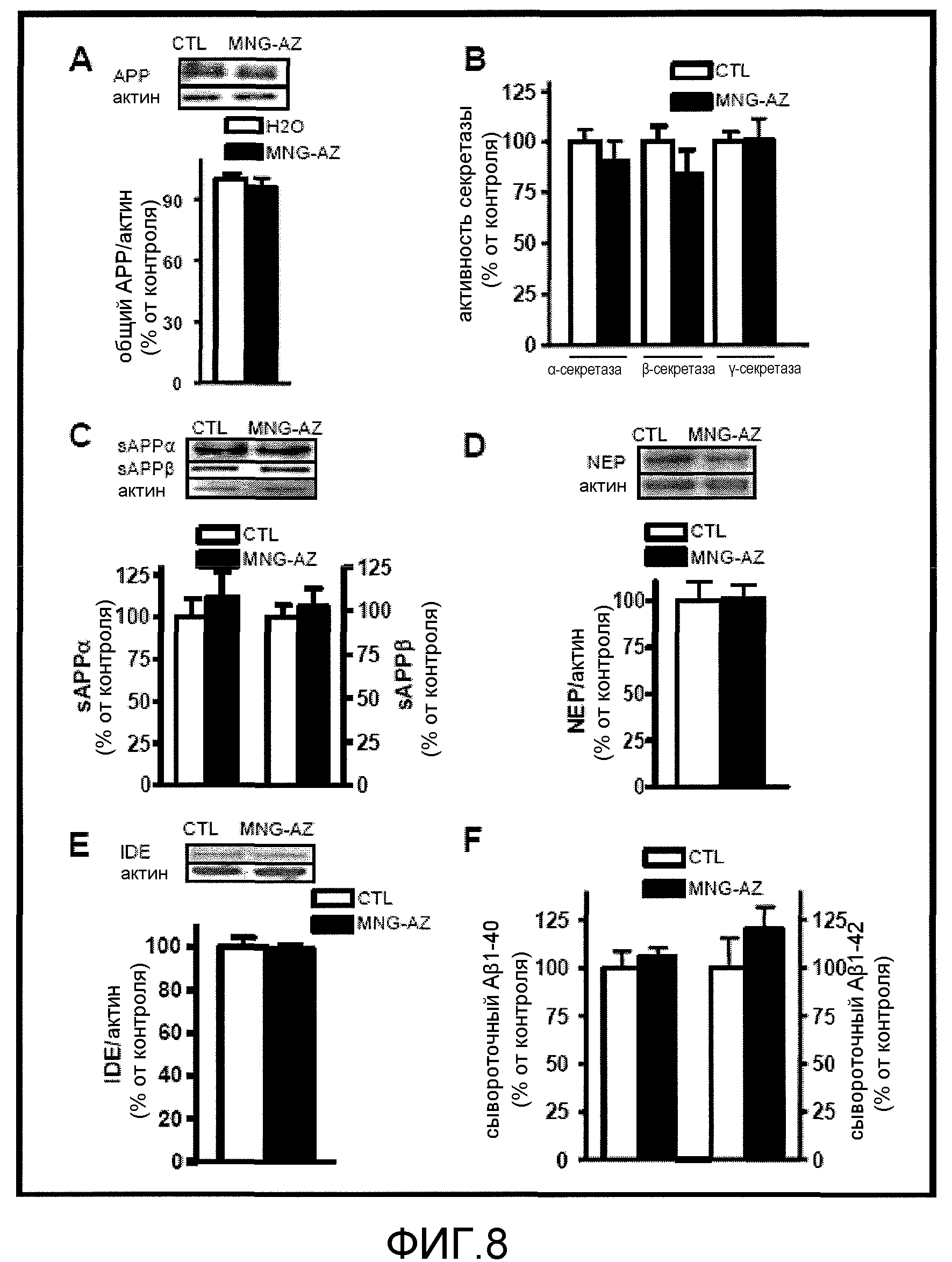
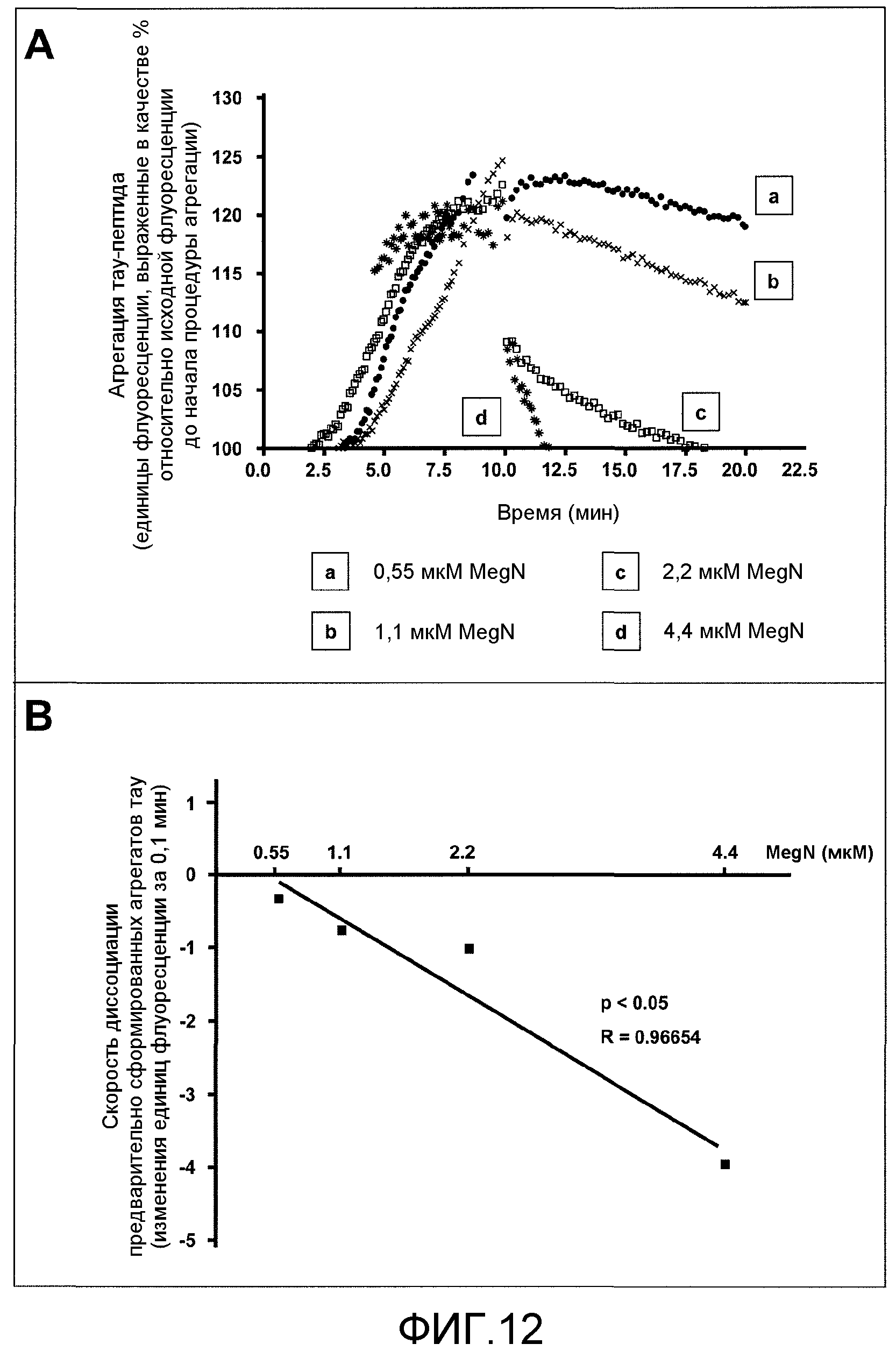
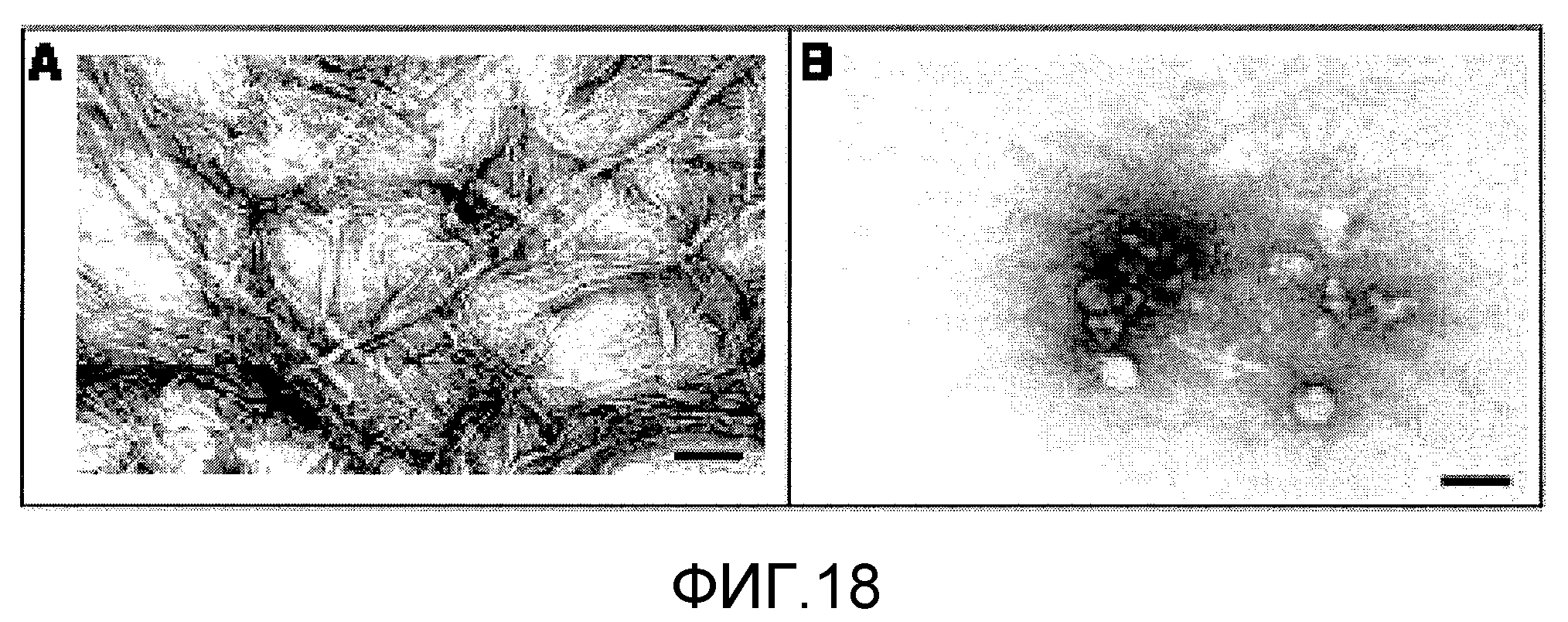
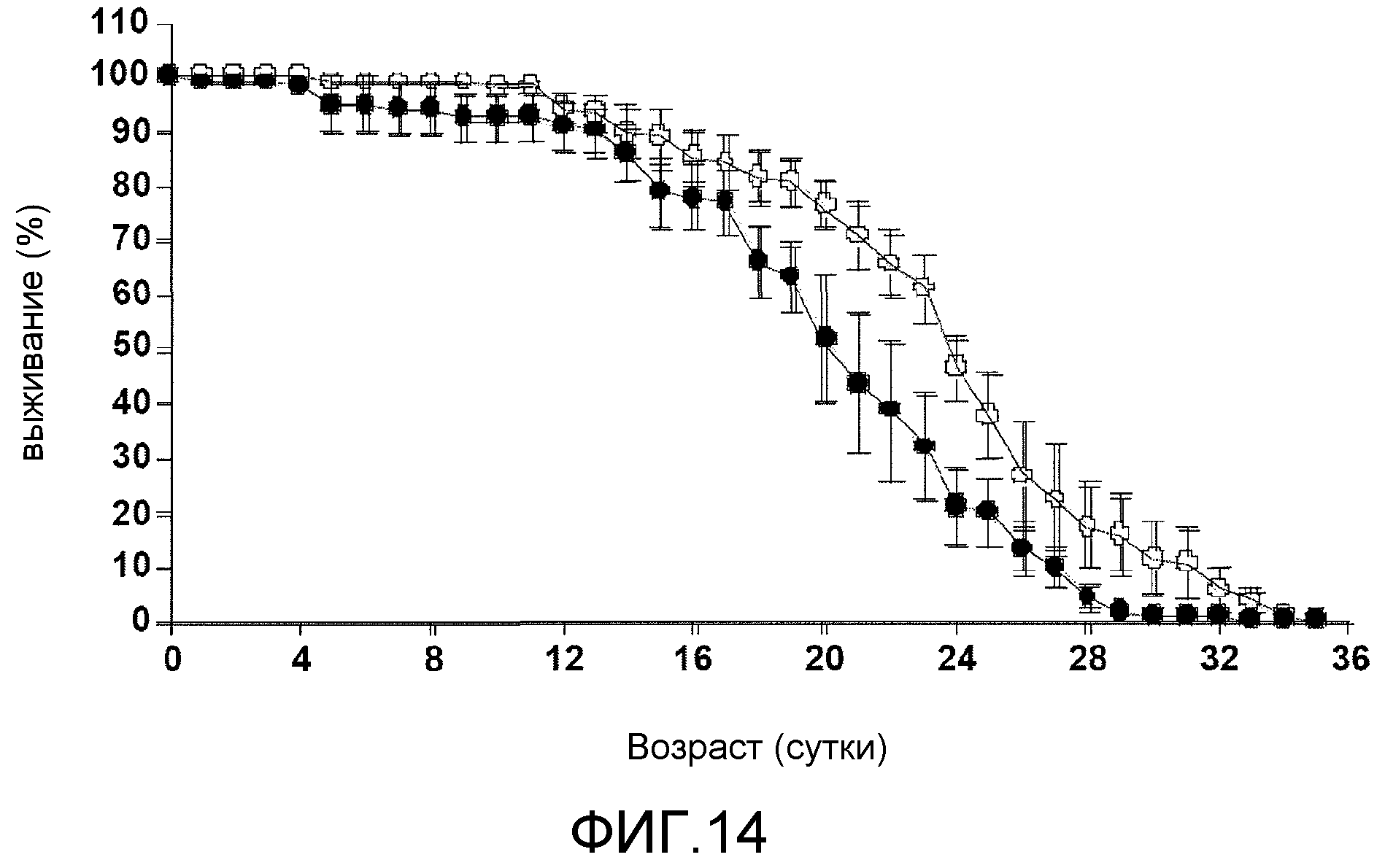
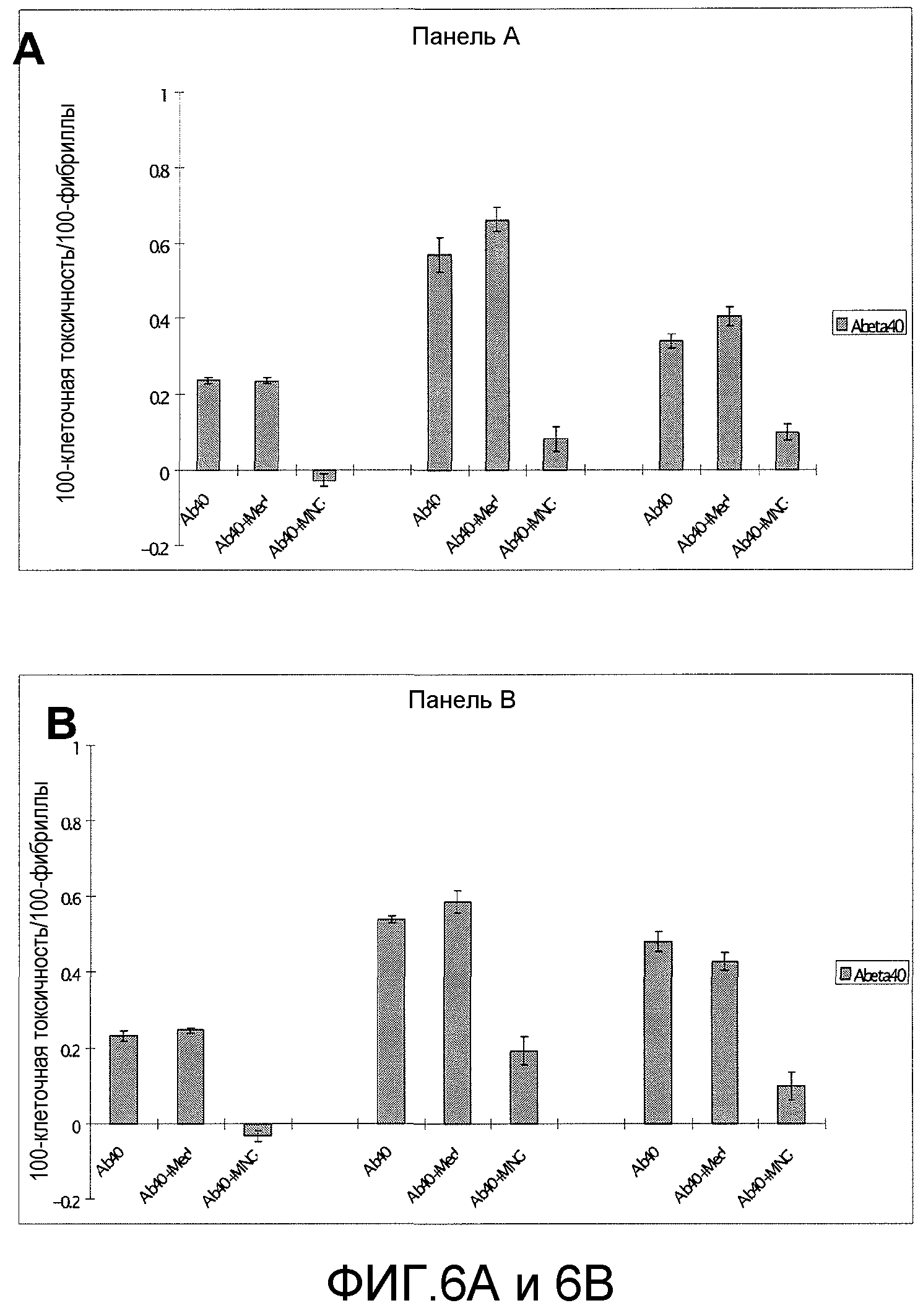
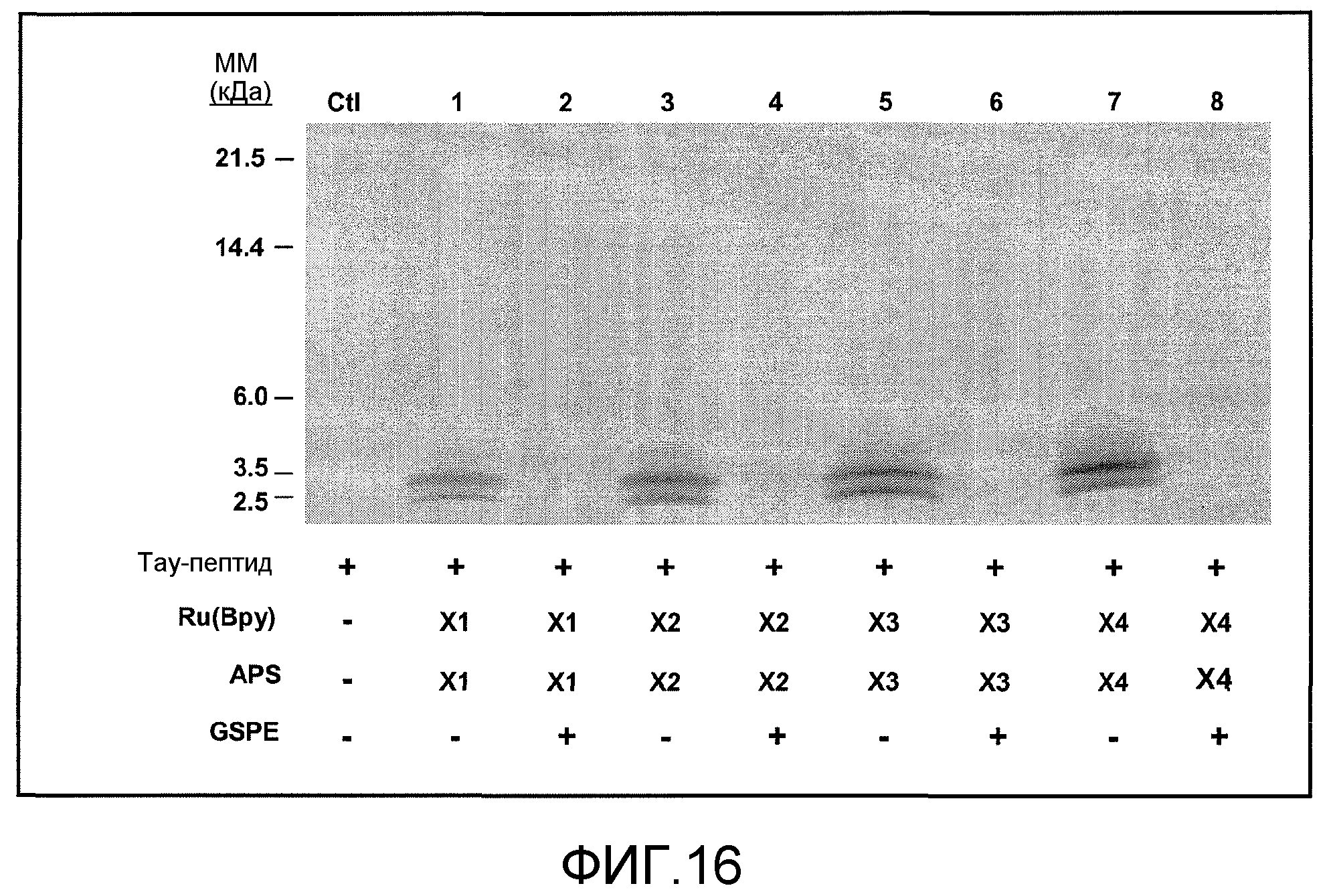
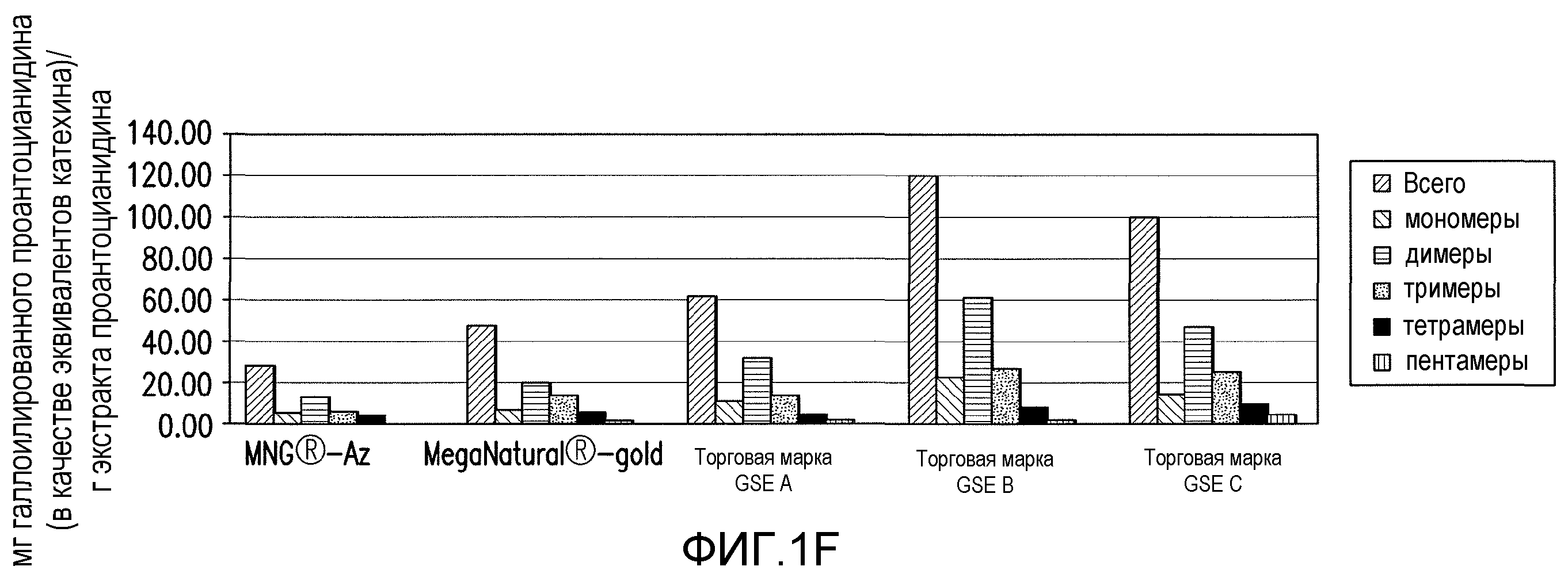
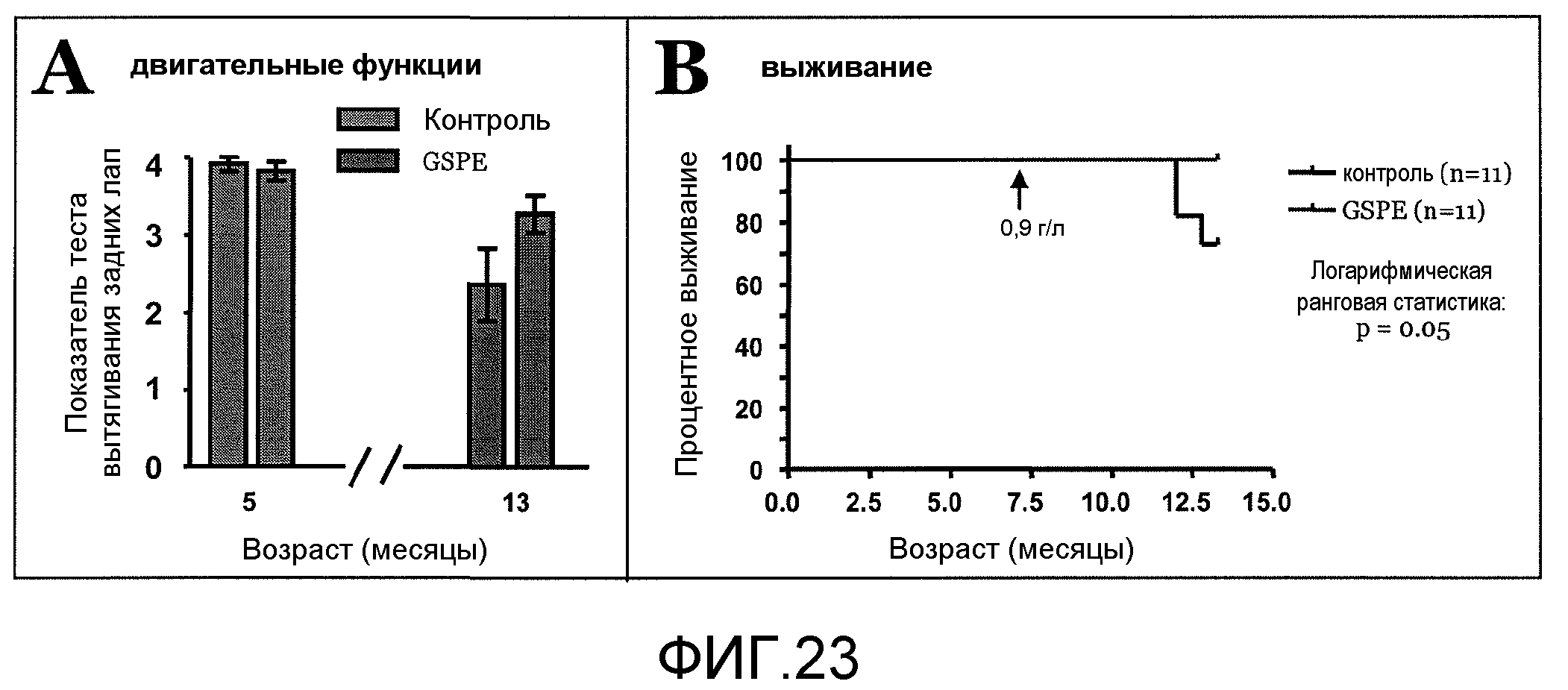
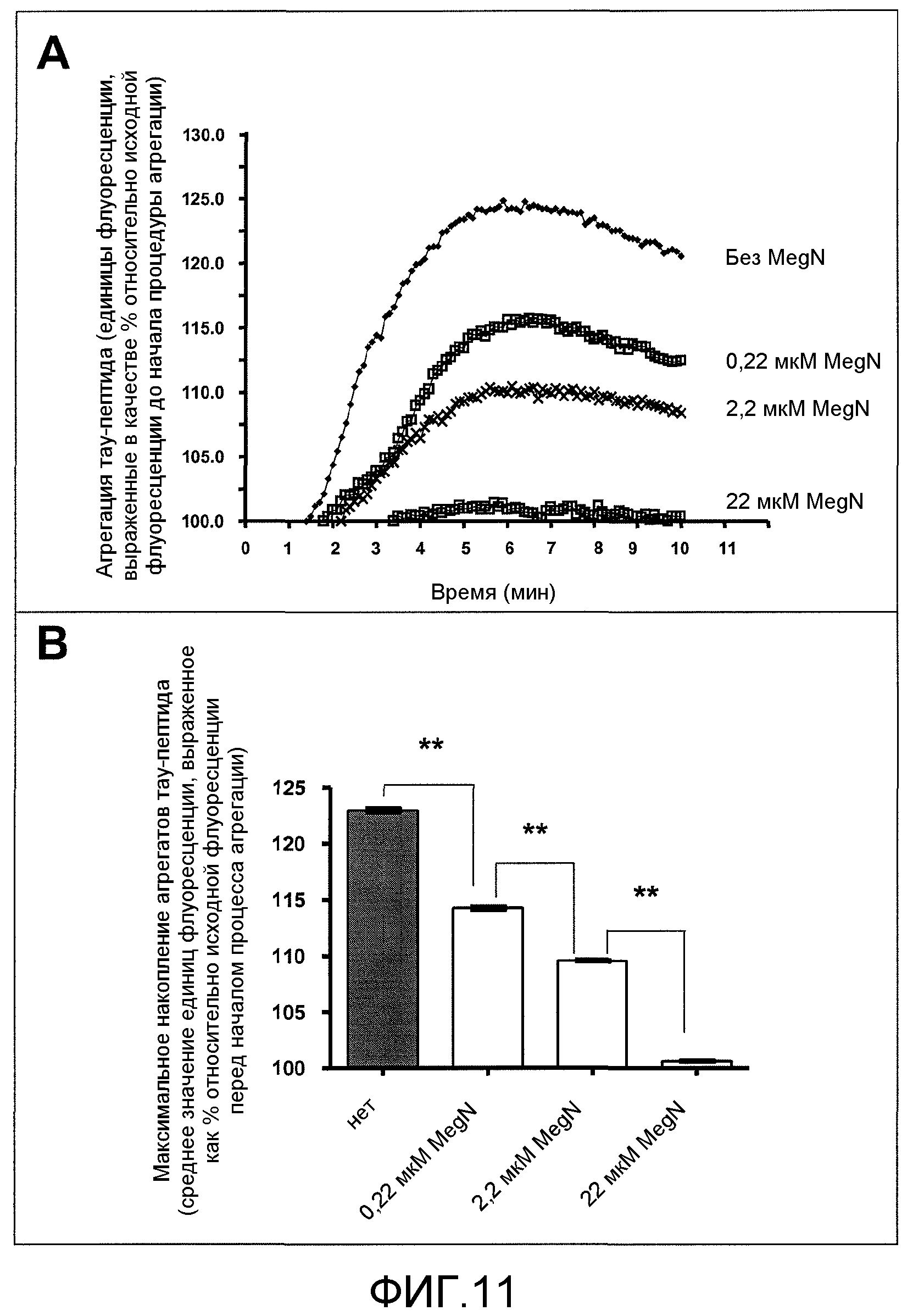
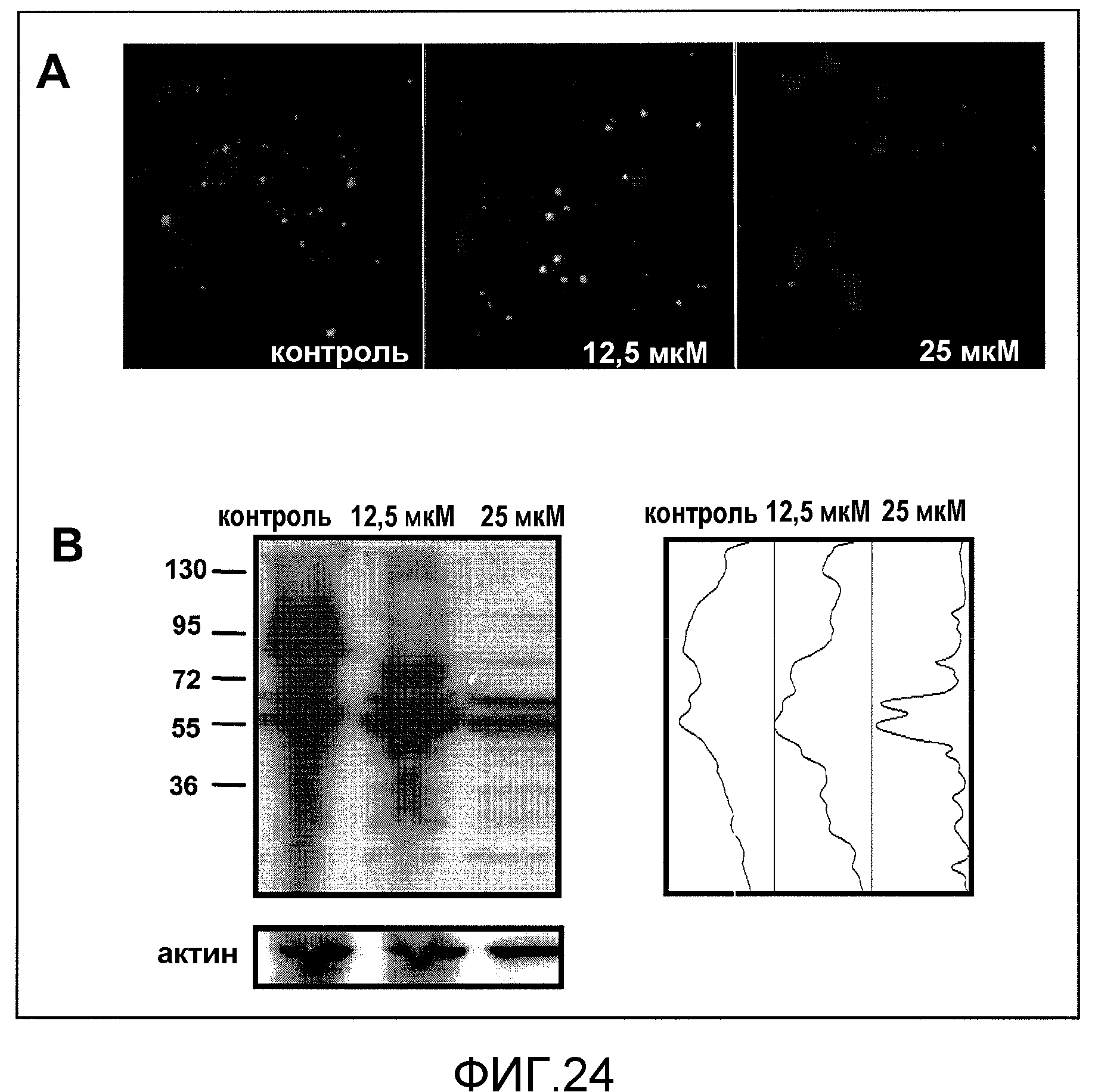
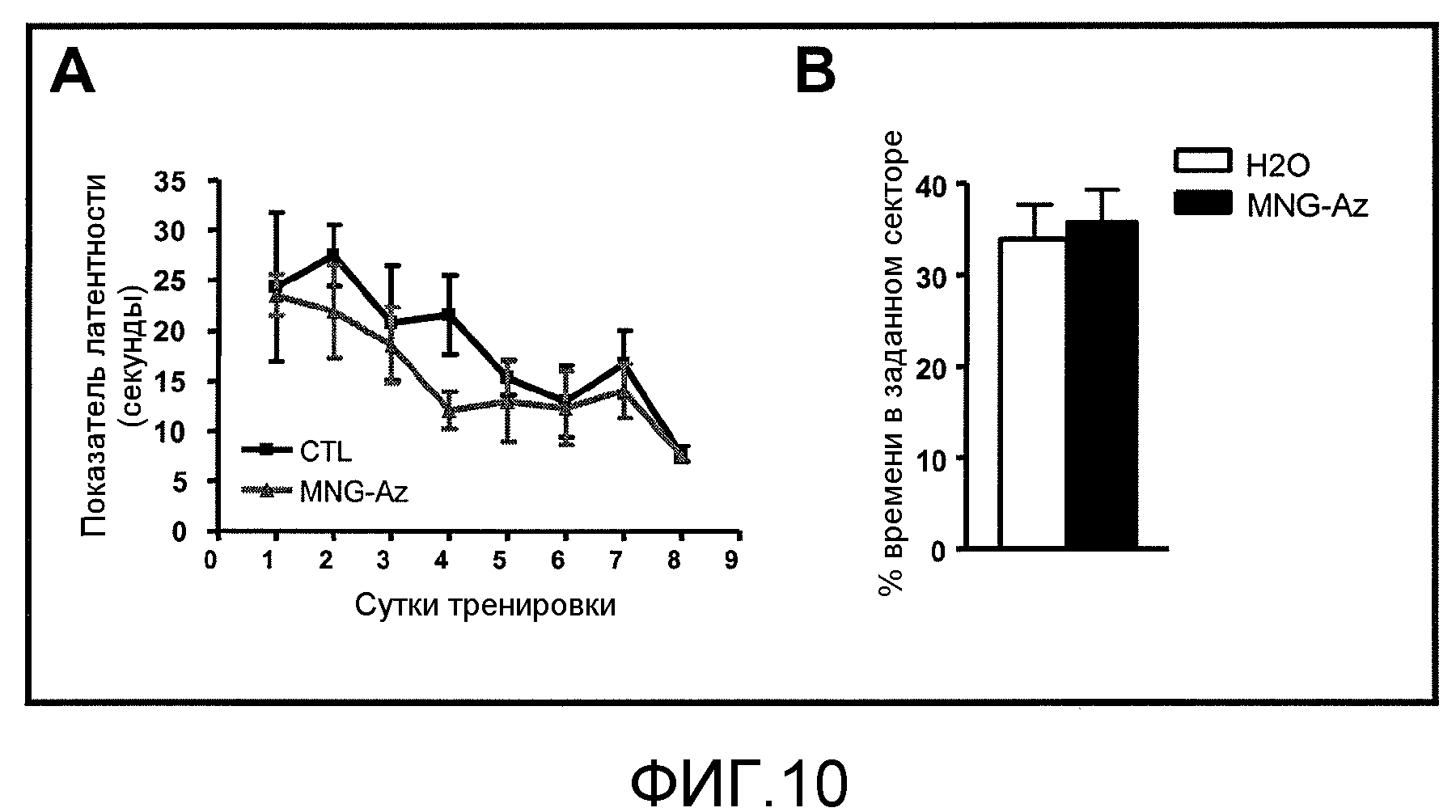
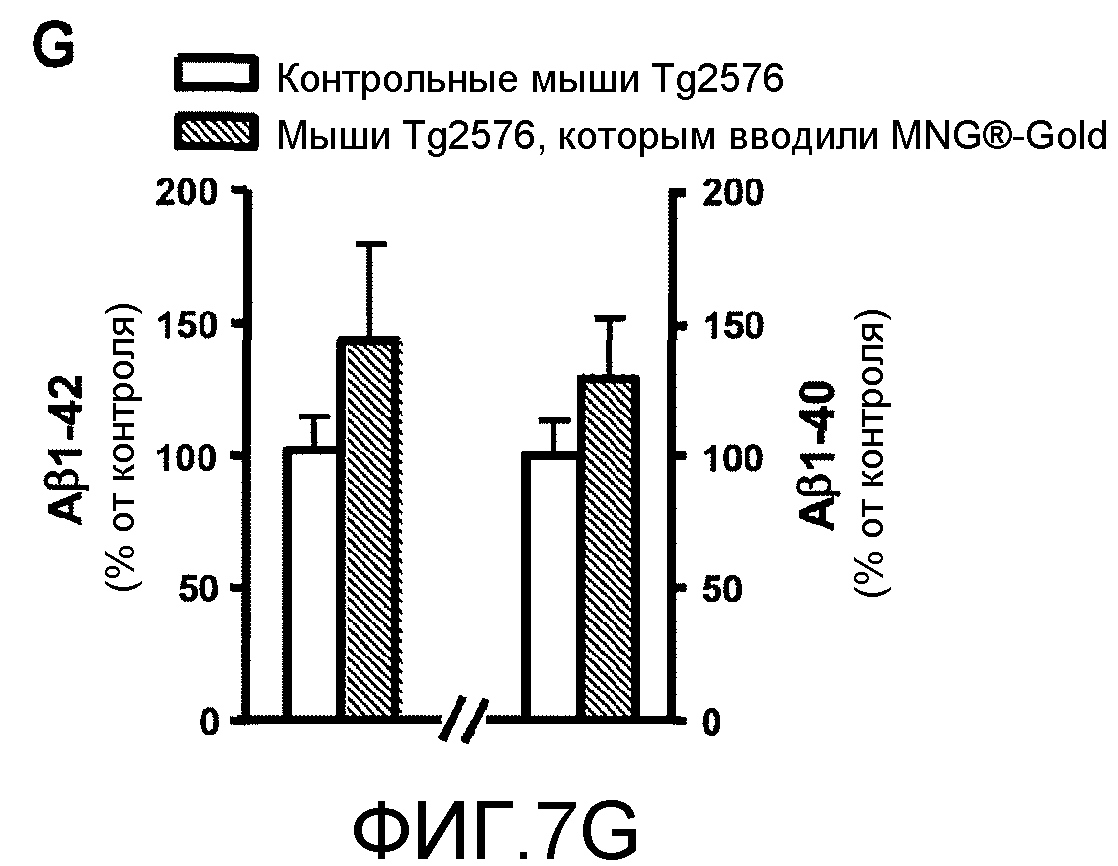
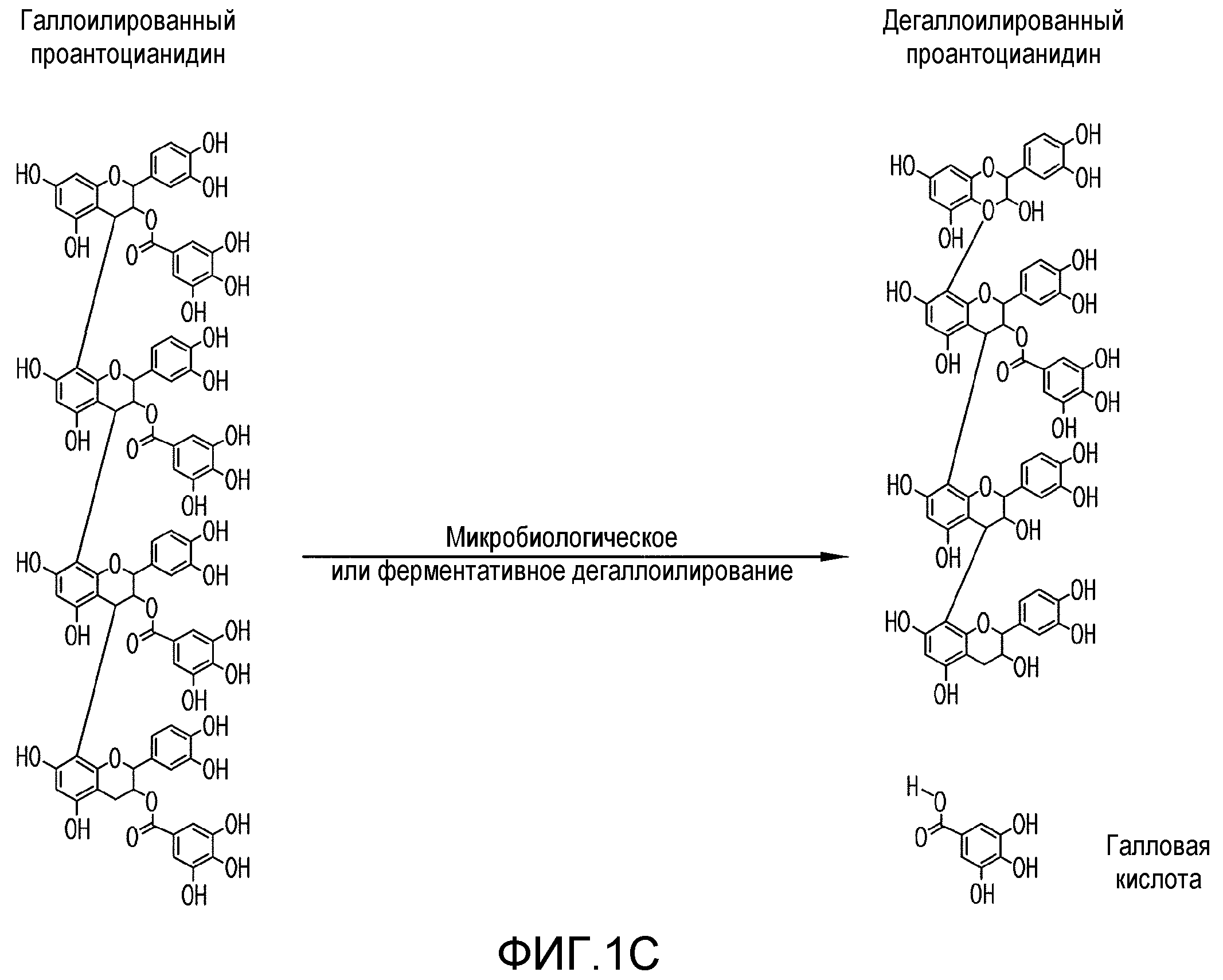
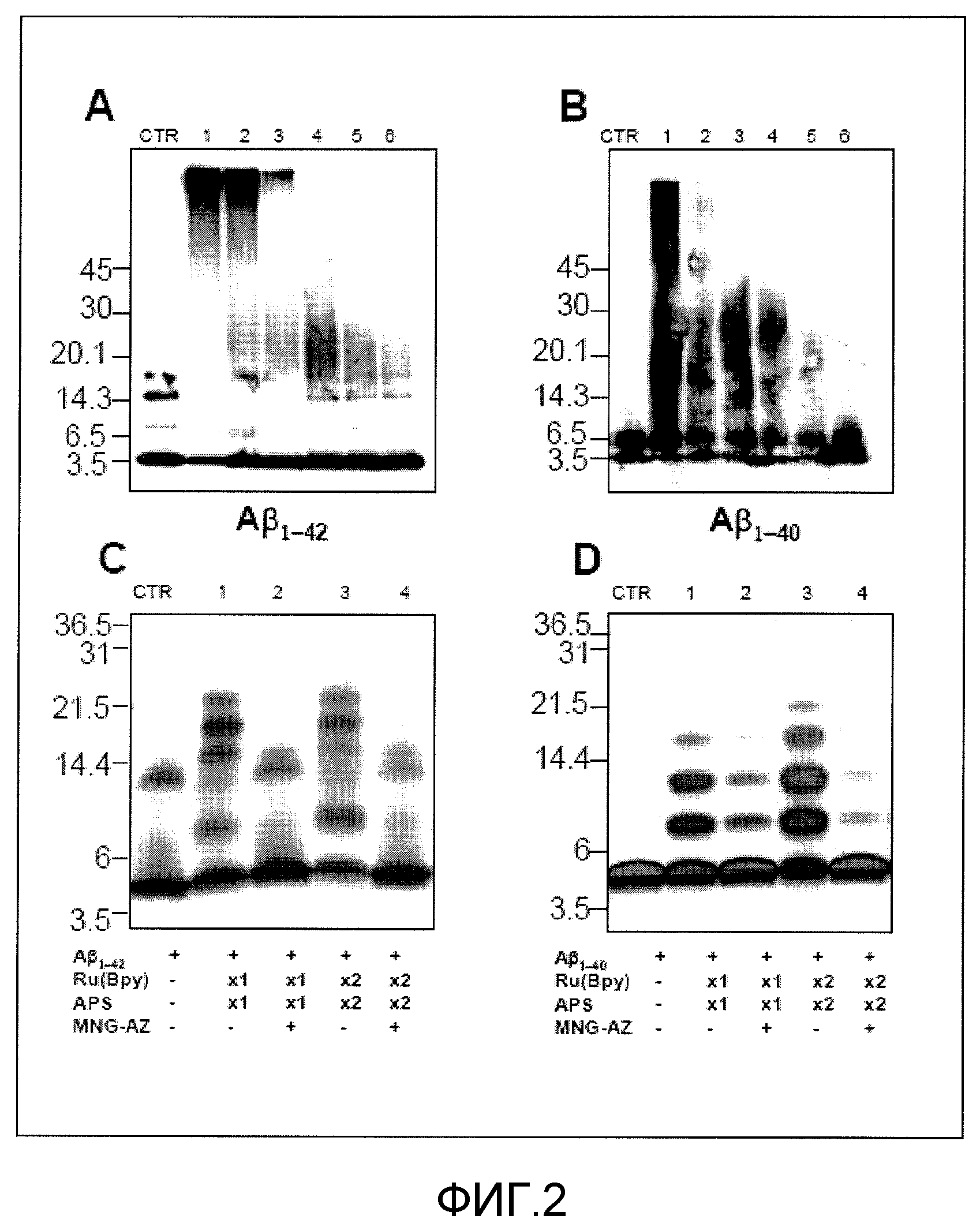
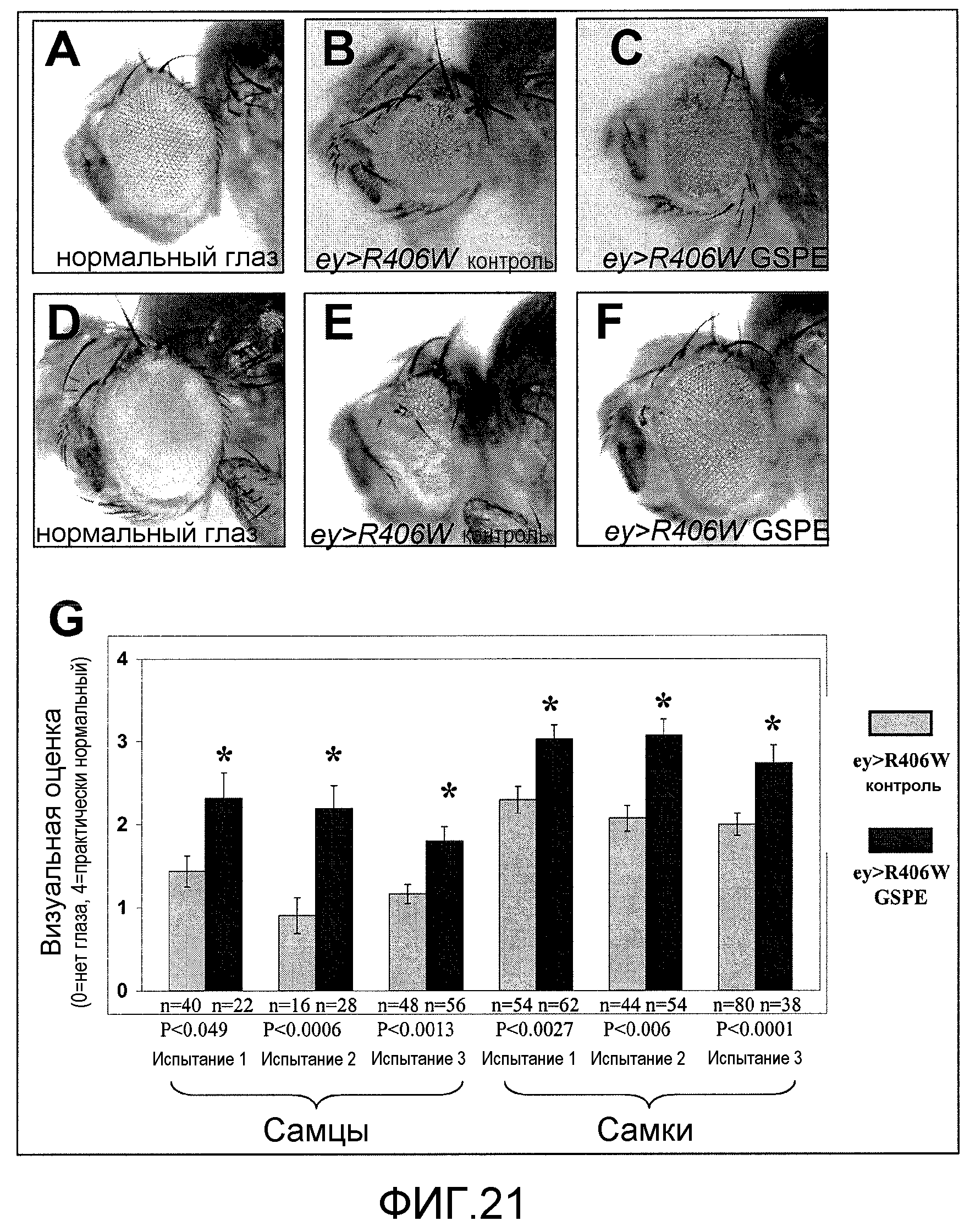
Реконструкция изображения, включающая в себя коррекцию изменяющегося со смещением размытия
Ферментная заместительная терапия с увеличением дозы для лечения недостаточности кислой сфингомиелиназы
Упаковка, имеющая единый корпус, включающий отламываемый колпачок
Упаковка, имеющая цельный корпус, включающий в себя отламывающийся колпачок
Аналоги азацитидина и их применение
Аналоги азацитидина и их применение
Реконструкция изображения, включающая в себя коррекцию изменяющегося со смещением размытия
Ферментная заместительная терапия с увеличением дозы для лечения недостаточности кислой сфингомиелиназы
Упаковка, имеющая единый корпус, включающий отламываемый колпачок
Упаковка, имеющая цельный корпус, включающий в себя отламывающийся колпачок
Профиль пользователя, политика и распределение ключей pmip в сети беспроводной связи
Конфигурирование информации о качестве обслуживания
Система, способ и устройство обслуживания местоположения (омп) на основании плоскости пользователя

