Результат интеллектуальной деятельности: АНТИТЕЛА-АНТАГОНИСТЫ ПРОТИВ Notch3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С Notch3 ЗАБОЛЕВАНИЙ
Вид РИД
Изобретение
РОДСТВЕННЫЕ ЗАЯВКИ
По данной заявке испрашивается приоритет предварительной патентной заявки США No. 60/875597, поданной 18 декабря 2006, и предварительной патентной заявки США No. 60/879218, поданной 6 января 2007, описания которых приведены в настоящем документе в качестве ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам-антагонистам против Notch3 и их применению для облегчения, лечения или профилактики связанного с Notch3 заболевания или нарушения.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Ген Notch впервые описан в 1917, когда было обнаружено, что штамм плодовой мушки Drosophila melanogaster обладает выемчатыми крыловыми пластинками (Morgan, Am Nat 51:513 (1917)). Ген клонировали почти семьдесят лет спустя, и, как определили, он является рецептором клеточной поверхности, играющим ключевую роль в развитии многих различных типов клеток и тканей у Drosophila (Wharton et al., Cell 43:567 (1985)). Как вскоре обнаружили, путь передачи сигнала Notch представляет собой механизм передачи сигнала, опосредованный контактом клетка-клетка, и является эволюционно консервативным от Drosophila до человека. Как обнаружено, рецепторы Notch принимают участие во многих клеточных процессах, таких как дифференцировка, определение судьбы клеток, поддержание стволовых клеток, клеточная подвижность, пролиферация и апоптоз в различных типах клеток в ходе развития и в тканевом гомеостазе (для обзора см. Artavanis-Tsakonas, et al., Science 268:225 (1995)).
Млекопитающие обладают четырьмя белками рецептора Notch (обозначенными Notch1-Notch4) и пятью соответствующими лигандами (обозначенными Delta-1 (DLL-1), Delta-3 (DLL-3), Delta-4 (DLL-4), Jagged-1 и Jagged-2). Гены рецепторов Notch млекопитающих кодируют белки ~300 кД, которые подвергаются расщеплению при транспортировке к клеточной поверхности и существуют в виде гетеродимеров. Внеклеточная часть рецептора Notch обладает тридцатью четырьмя повторами, подобными фактору роста эпидермиса (EGF), и тремя цистеин-богатыми Notch/LIN12-повторами. Ассоциация двух расщепленных субъединиц опосредована последовательностями, расположенными непосредственно к N-концу и C-концу от участка расщепления, и эти две субъединицы образуют гетеродимеризационные (HD) домены Notch (Wharton, et al., Cell 43:567 (1985); Kidd, et al., Mol Cell Biol 6:3431 (1986); Kopczynski, et al., Genes Dev 2:1723 (1988); Yochem, et al., Nature 335:547 (1988)).
В настоящее время все еще остается неясным, как происходит регуляция передачи сигнала Notch различными рецепторами или как различаются пять лигандов в отношении передачи сигнала или регуляции. Различия в передаче сигнала и/или регуляции можно контролировать с помощью их паттернов экспрессии в различных тканях или с помощью разных сигналов окружающей среды. Установлено, что белки лигандов Notch, включая Jagged/Serrate- и Delta/Delta-подобные, специфически связываются с областью EGF-повтора и индуцируют рецептор-опосредованную передачу сигнала Notch (рассмотрено в Bray, Nature Rev Mol Cell Biol. 7:678 (2006), и в Kadesch, Exp Cell Res. 260:1 (2000)). Среди EGF-повторов повторы с 10-го по 12-й необходимы для связывания лиганда с рецептором Notch, и другие EGF-повторы могут усиливать взаимодействие рецептор-лиганд (Xu, et al., J Biol Chem. 280:30158 (2005); Shimizu, et al., Biochem Biophys Res Comm. 276:385 (2000)). Хотя LIN12-повторы и димеризационный домен не вовлечены непосредственно в связывание лиганда, они играют важные роли в сохранении гетеродимерного белкового комплекса, предотвращая лиганд-независимое расщепление протеазой и активацию рецептора (Sanche-Irizarry, et al., Mol Cell Biol. 24:9265 (2004); Vardar et al., Biochem. 42:7061 (2003)).
Экспрессия мутантных форм рецепторов Notch в ходе развития эмбриона Xenopus серьезно нарушает нормальное развитие (Coffman, et al., Cell 73:659 (1993)). Обнаружено, что нокаут Notch1 является эмбрионально летальным у мышей (Swiatek, et al., Genes & Dev 8:707 (1994)). У человека существует несколько генетических заболеваний, включая рак, связанных с различными мутациями рецептора Notch (Artavanis-Tsakonas, et al., Science 284:770 (1999)). Например, патологическая активация рецептора Notch1, вызываемая транслокацией, может приводить к T-клеточному лимфобластному лейкозу (Ellisen, et al., Cell 66:649 (1991)). Определенные мутации в HD-доменах рецептора Notch1 усиливают передачу сигнала без связывания лиганда (Malecki, et al., Mol Cell Biol 26:4642 (2006)), что дополнительно указывает на их роли в активации рецептора Notch. Сигнал, индуцируемый связыванием лиганда, передается в ядро в ходе процесса, включающего два протеолитических расщепления рецептора с последующей ядерной транслокацией внутриклеточного домена (Notch-IC). Хотя LIN12-повторы и HD-домены, как предполагали, предотвращают передачу сигнала в отсутствие лигандов, все еще остается неясным, как связывание лиганда способствует событиям протеолитического расщепления.
Рецепторы Notch связаны с широким рядом заболеваний, включая рак, неврологические нарушения и иммунные заболевания, как показано в сообщениях о сверхэкспрессии рецепторов Notch в различных тканях и линиях клеток при заболеваниях человека по сравнению с нормальными или незлокачественными клетками (Joutel, et al. Cell & Dev Biol 9:619 (1998); Nam, et al., Curr Opin Chem Biol 6:501 (2002)). Рецептор Notch3 сверхэкспрессируется в различных солидных опухолях, включая немелкоклеточный рак легкого (NSCLC) и рак яичника (Haruki, et al., Cancer Res 65:3555 (2005); Park, et al., Cancer Res 66:6312 (2006); Lu, et al., Clin Cancer Res 10:3291 (2004)), что позволяет предположить важность экспрессии рецептора Notch3 в солидных опухолях. Более того, экспрессия рецептора Notch3 активируется в новообразованиях из плазматических клеток, включая множественную миелому, плазмоклеточный лейкоз и экстрамедуллярную плазмацитому (Hedvat, et al., Br J Haematol 122:728 (2003)); рак поджелудочной железы (Buchler, et al., Ann Surg 242:791 (2005)); и T-клеточные острые лимфобластные лейкозы (T-ALL) (Bellavia, et al., Proc Natl Acad Sci USA 99:3788 (2002); Screpanti, et al., Trends Mol Med 9:30 (2003)). Рецептор Notch3 также экспрессируется в субпопуляции линий клеток нейробластомы и служит в качестве маркера этого типа опухоли, который обладает конституциональными или опухоль-специфичными мутациями в гене гомеобокса Phox2B (van Limpt, et al., Cancer Lett 228:59 (2005)). Другие симптомы и заболевания, которые связаны с экспрессией рецептора Notch3, включают неврологические нарушения (Joutel, et al., Nature 383:707 (1996)), диабет (Anastasi, et al., J Immunol 171:4504 (2003), ревматоидный артрит (Yabe, et al., J Orthop Sci 10:589 (2005)), связанные с сосудами заболевания (Sweeney, et al., FASEB J 18:1421 (2004)) и синдром Алажиля (Flynn, et al., J Pathol 204:55 (2004)).
Хотя сверхэкспрессию рецептора Notch3 (включая амплификацию гена) наблюдали в различных видах рака, до сих пор отсутствуют сообщения об активирующих мутациях. Вероятно, повышение уровня рецепторов Notch3 в опухолях может быть активировано разными лигандами в стромальных клетках или опухолевых клетках, и оно приводит к усилению передачи сигнала Notch3. В частности, лиганды Notch локализованы в сосудистом эндотелии в ходе как развития, так и онкогенеза (Mailhos, et al., Differentiation 69:135 (2001); Taichman, et al., Dev Dyn 225:166 (2002)), позволяя предположить, что эндотелиальные клетки могут предоставлять лиганды для активации рецептора Notch3 в опухолях. Сходное взаимное влияние опухоли и стромы, опосредованное лигандом и рецептором Notch, показано для различных типов рака (Houde, et al., Blood 104:3697 (2004); Jundt, et al., Blood 103:3511 (2004); Zeng, et al., Cancer Cell 8:13 (2005)). Усиление передачи сигнала Notch3, вызываемое сверхэкспрессией внутриклеточного Notch3 (Notch3-IC), может приводить к онкогенезу у животных моделей T-ALL и рака молочной железы (Vacca, et al., The EMBO J 25:1000 (2006); Hu, et al., Am J Pathol 168:973 (2006)).
Передача сигнала Notch3 и его роль в клеточном самообновлении имеют место в раковых стволовых клетках, которые представляют собой минорную популяцию в опухолях и могут инициировать образование опухоли (Reya, et al., Nature 414:105 (2001)). Нормальные стволовые клетки из многих тканей, включая кишечные и нейрональные стволовые клетки, зависят от передачи сигнала Notch3 в отношении самообновления и определения судьбы (Fre, et al., Nature, 435:964 (2005); van Es, et al., Nature, 435:959 (2005); Androutsellis-Theotokis, et al., Nature, 442:823 (2006)). Сходные механизмы могут существовать в раковых стволовых клетках, и, как показано, ингибирование передачи сигнала Notch3 ингибиторами γ-секретазы истощает количество раковых стволовых клеток и блокирует приживление для эмбриональных опухолей мозга (Fan, et al., Cancer Res 66:7445 (2006)).
Ингибирование передачи сигнала Notch3 ингибитором γ-секретазы обладает поразительными антинеопластическими эффектами в экспрессирующих Notch трансформированных клетках in vitro и в моделях ксенотрансплантата (Weijzen, et al., Nat Medicine 8:879 (2002); Bocchetta, et al., Oncogene 22:81 (2003); Weng, et al., Science, 306:269 (2004)). Позднее показано, что ингибитор γ-секретазы эффективно уничтожает аденомы толстой кишки у мышей Apc (min+) (van Es, et al., Nature, 435:959 (2005)), хотя терапевтическое окно, благодаря его эффекту на нормальные стволовые клетки и ингибированию нескольких путей Notch, является очень узким. В отличие от Notch1, нокаут гена Notch3 у мышей не являлся эмбрионально летальным и вызывал незначительное число дефектов (Domenga, et al., Genes & Dev 18:2730 (2004)), позволяя предположить, что Notch3 является потенциально лучшей терапевтической мишенью, чем Notch1.
У Tournier-Lasserve et al. (заявка США 2003/0186290) сообщается об ассоциации рецептора Notch3 и CADASIL. В заявке описаны различные мутации в гене Notch3 и их возможная ассоциация с заболеванием CADASIL. В заявке предложено применение диагностических антител для детектирования таких мутаций. В заявке также предложены терапевтические антитела для лечения CADASIL, т.е. агонистические антитела, но не описаны конкретные антитела, и не описано, как получать такие антитела.
Ввиду большого количества заболеваний человека, связанных с путем передачи сигнала Notch3, важно выявлять новые способы профилактики и лечения этих заболеваний. Настоящее изобретение относится к новым анти-Notch3 антителам, пригодным для этой нерешенной медицинской проблемы.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым антителам и их фрагментам, которые специфически связываются с конформационным эпитопом рецептора Notch3 человека, где эпитоп содержит домен LIN12 и гетеродимеризационный домен. Другой аспект изобретения относится к участку связывания эпитопа и антителам, которые связываются с тем же эпитопом, что и антитела по настоящему изобретению. Антитела по настоящему изобретению ингибируют индуцированную лигандом передачу сигнала посредством рецептора Notch3.
Изобретение относится к аминокислотным последовательностям вариабельной тяжелой и легкой цепи антител и соответствующим последовательностям их нуклеиновых кислот. Другой вариант осуществления изобретения относится к последовательностям CDR этих антител. Другой вариант осуществления относится к гуманизированным формам этих антител.
Другой вариант осуществления настоящего изобретения относится к линиям клеток и векторам, несущим последовательности антител по настоящему изобретению.
Настоящее изобретение также относится к конформационному эпитопу, распознаваемому антителами-антагонистами по изобретению. Настоящее изобретение также относится к антителам, которые связываются с этим конформационным эпитопом. Варианты осуществления относятся к конформационному эпитопу Notch3, содержащему домен LIN12, обладающий, по меньшей мере, 80%, 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 9, и димеризационный домен 2, обладающий, по меньшей мере, 80%, 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 18. Более конкретно, конформационный эпитоп Notch3 содержит аминокислотные остатки 1395-1396, 1402-1404 и 1420-1422 домена L1 LIN12 и аминокислотные остатки 1576-1578 и 1626-1628 димеризационного домена D2. Настоящее изобретение относится к антителам, которые связываются с этим конформационным эпитопом.
Другой вариант осуществления настоящего изобретения состоит в использовании любого из этих антител для получения лекарственного средства или композиции для лечения заболеваний и нарушений, связанных с активацией рецептора Notch3.
Другой вариант осуществления настоящего изобретения состоит в использовании любого из этих антител для лечения нарушений, связанных с активацией Notch3, включая ингибирование указанной активации, например, путем ингибирования передачи сигнала Notch3 или нейтрализации рецептора блокированием связывания с лигандом. Связанные с Notch3 нарушения могут включать в качестве неограничивающих примеров T-клеточный острый лимфобластный лейкоз, лимфому, заболевание печени, при котором имеет место патологическая васкуляризация, диабет, рак яичника, заболевания, при которых имеет место определение судьбы клеток сосудов, ревматоидный артрит, рак поджелудочной железы, немелкоклеточный рак легкого, новообразования из плазматических клеток (такие как множественная миелома, плазмоклеточный лейкоз и экстрамедуллярная плазмацитома) и нейробластому.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг.1 представлена аминокислотная последовательность Notch3. Область EGF-повтора простирается от аминокислотного остатка 43 до 1383; домен LIN12 простирается от аминокислотного остатка 1384 до 1503; и димеризационный домен простирается от аминокислотного остатка 1504 до 1640.
На фиг.2 (A-H) представлено сравнение аминокислотных последовательностей Notch1, Notch2, Notch3 и Notch4 человека.
На фиг.3 представлена процентная идентичность Notch1, Notch2, Notch3 и Notch4.
На фиг.4A и 4B представлены последовательности вариабельной области тяжелой и легкой цепи моноклонального антитела анти-Notch3 MAb 256A-4 (SEQ ID NO: 2) с подчеркнутыми областями CDR.
На фиг.5A и 5B представлены последовательности вариабельной области тяжелой и легкой цепи моноклонального антитела анти-Notch3 MAb 256A-8 (SEQ ID NO: 4) с подчеркнутыми областями CDR.
На фиг.6 представлен анализ с использованием люциферазного репортера из примера 5, показывающий ингибиторные эффекты анти-Notch3 MAb на лиганд Notch3 Jagged1.
На фиг.7 представлен анализ с использованием люциферазного репортера, показывающий ингибиторные эффекты анти-Notch3 MAb на лиганд Notch3 Jagged2.
На фиг.8 представлен анализ с использованием люциферазного репортера, показывающий ингибиторные эффекты анти-Notch3 MAb на лиганд Notch3 DLL4.
На фиг.9 представлен анализ с использованием люциферазного репортера, показывающий ингибиторные эффекты анти-Notch3 MAb на нативный Notch3 в клетках рака яичника. (9A) - линия клеток рака яичника человека OV/CAR3 и (9B) - линия клеток рака яичника человека A2780.
На фиг.10 представлен анализ апоптоза из примера 6, показывающий, что эффект выживания клеток, индуцированный Jagged1, ингибировался анти-Notch3 MAb.
На фиг.11 представлен ингибиторный эффект анти-Notch3 MAb на миграцию (11A) и инвазию (11B) клеток из примера 7.
На фиг.12 представлена схематическая диаграмма белка с перестановкой доменов Notch1-Notch3, экспрессируемого в виде слитого белка с IgG/Fc человека, связанного с C-концом.
На фиг.13A представлен ELISA с использованием контрольного антитела к Fc человека в качестве антитела для детекции, показывающий, что белки фиг.12 экспрессировались в кондиционированной среде. На фиг.13B представлен ELISA с использованием 256A-4 в качестве антитела для детекции. На фиг.13C представлен ELISA с использованием 256A-8 в качестве антитела для детекции. На фиг.13D представлен ELISA с использованием антитела 256A-13 для положительного контроля в качестве антитела для детекции.
На фиг.14 представлено сравнение сконструированной последовательности, кодирующей лидерный пептид Notch3, с нативной последовательностью, кодирующей лидерный пептид Notch3 (депозитарный No. NCBI GenBank NM_000435), показывающее замены нуклеотидов (14A), и транслируемая аминокислотная последовательность сконструированной последовательности лидерного пептида Notch (14B).
На фиг.15 представлено получение конструкции с перестановкой доменов с помощью способа PCR-SOE. Стрелки представляют собой праймеры для PCR. Незакрашенный участок - последовательность Notch3. Закрашенный участок - последовательность Notch1.
На фиг.16 представлены аминокислотные последовательности, использованные для картирования эпитопа домена LIN12 Notch3 MAb 256A-4 и 256A-8.
На фиг.17 представлены аминокислотные последовательности, использованные для картирования эпитопа димеризационного домена Notch3 MAb 256A-4 и 256A-8.
На фиг.18 представлена схема участка связывания эпитопа для MAb 256A-4 и 256A-8.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение не ограничивается описанными в настоящем документе конкретными способами, протоколами, линиями клеток, векторами или реагентами, поскольку они могут варьировать. Более того, используемая в настоящем документе терминология предназначена только для целей описания конкретных вариантов осуществления и не предназначена для ограничения объема настоящего изобретения. Как используют в настоящем описании и в прилагаемой формуле изобретения, формы единственного числа включают ссылки на формы множественного числа, если только из контекста ясно не следует иначе, например, ссылка на «клетку-хозяина» включает множество таких клеток-хозяев. Если только не указано иначе, все используемые в настоящем документе технические и научные термины и любые акронимы обладают одинаковыми значениями, общепринятыми для специалиста в данной области изобретения. Хотя любые способы и материалы, сходные с описанными в настоящем документе или эквивалентные им, можно использовать при практическом осуществлении настоящего изобретения, в настоящем документе описаны иллюстративные способы, устройства и материалы.
Все упомянутые в настоящем описании патенты и публикации приведены в качестве ссылки в пределах, разрешаемых законом, с целью описания и сообщения о рассмотренных там белках, ферментах, векторах, клетках-хозяевах и способах, которые можно применять с настоящим изобретением. Однако ничего в настоящем описании не следует истолковывать как допущение того, что изобретение не является основанием, чтобы предвосхищать такое описание на основании предшествующего изобретения.
Определения
Термины, используемые на протяжении данной заявки, следует толковать в общепринятом и типичном значении для специалистов в данной области. Однако заявители выражают желание, чтобы следующим терминам давали конкретные определения, как определено ниже.
Фразу «по существу идентичный» в отношении последовательности полипептидной цепи антитела можно толковать как цепь антитела, обладающую, по меньшей мере, 70%, или 80%, или 90%, или 95% идентичностью последовательности с референсной полипептидной последовательностью. Термин в отношении последовательности нуклеиновой кислоты можно толковать как последовательность нуклеотидов, обладающую, по меньшей мере, приблизительно 85% или 90%, или 95%, или 97% идентичностью последовательности с референсной последовательностью нуклеиновой кислоты.
Термин «идентичность» или «гомология» следует понимать как обозначение процентной доли аминокислотных остатков в рассматриваемой последовательности, которые идентичны остаткам соответствующей последовательности, с которой ее сравнивают, после выравнивания последовательностей и внесения пропусков, при необходимости достижения максимальной процентной идентичности с целой последовательностью, и, не рассматривая любые консервативные замещения, как имеющие отношение к идентичности последовательности. Ни N-, ни C-концевые добавочные последовательности, ни вставки не следует толковать как снижающие идентичность или гомологию. Способы и компьютерные программы для выравнивания хорошо известны в данной области. Идентичность последовательности можно измерять с использованием программного обеспечения для анализа последовательностей.
Термин «антитело» используют взаимозаменяемо в самом широком смысле, и он конкретно относится к моноклональным антителам (например, полноразмерным моноклональным антителам), поликлональным антителам и полиспецифическим антителам (например, биспецифическим антителам), и фрагментам антител до тех пор, пока они обладают требуемой биологической активностью. Антитела (Ab) и иммуноглобулины (Ig) представляют собой гликопротеины, обладающие одинаковыми структурными характеристиками. В то время как антитела обладают специфичностью связывания в отношении конкретной мишени, иммуноглобулины включают как антитела, так и другие антителоподобные молекулы, которые лишены специфичности в отношении мишени. Антитела по изобретению могут представлять собой любой тип (например, IgG, IgE, IgM, IgD, IgA и IgY), класс (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласс. Нативные антитела и иммуноглобулины обычно являются гетеротетрамерными гликопротеинами приблизительно 150000 Дальтон, образованными двумя идентичными легкими (L) цепями и двумя идентичными тяжелыми (H) цепями. Каждая тяжелая цепь содержит на одном конце вариабельный домен (VH), за которым следует ряд константных доменов. Каждая легкая цепь содержит вариабельный домен на одном конце (VL) и константный домен на другом конце.
Как используют в настоящем описании, «антитело анти-Notch3» означает антитело, которое специфически связывается с Notch3 человека таким образом, чтобы ингибировать или по существу снижать связывание Notch3 с его лигандами, или ингибировать передачу сигнала Notch3.
Термин «вариабельный» в контексте вариабельного домена антител, относится к тому факту, что последовательности определенных частей вариабельных доменов значительно различаются среди антител, и используется для связывания и специфичности каждого конкретного антитела в отношении его конкретной мишени. Однако вариабельность не распределена равномерно на протяжении вариабельных доменов антител. Она сконцентрирована в трех сегментах, называемых областями, определяющими комплементарность (CDR; т.е., CDR1, CDR2 и CDR3), также известными как гипервариабельные области в вариабельных доменах как легкой цепи, так и тяжелой цепи. Более высококонсервативные части вариабельных доменов называют каркасными областями (FR). Каждый из вариабельных доменов природных тяжелой и легкой цепей содержит четыре FR-области, в значительной степени принимающие конфигурацию β-слоя, соединенную тремя CDR, которые формируют петли, соединяющие структуру β-слоев и, в некоторых случаях, формирующие ее часть. CDR в каждой цепи расположены вместе в непосредственной близости с помощью областей FR и, совместно с CDR другой цепи, участвуют в формировании связывающего мишень участка антител (см. Kabat et al., Sequences of Protein of Immunological Interest, National Institute of Health, Bethesda, MD (1987)). Как используют в настоящем описании, нумерацию аминокислотных остатков иммуноглобулинов осуществляют согласно нумерации аминокислотных остатков иммуноглобулинов системы Kabat, et al., если не указано иначе.
Термин «фрагмент антитела» относится к части полноразмерного антитела, как правило, связывающей мишень или вариабельной области. Примеры фрагментов антител включают F(ab)-, F(ab')-, F(ab')2- и Fv-фрагменты. Фраза «функциональный фрагмент или аналог» антитела относится к соединению, обладающему количественной биологической активностью, подобной полноразмерному антителу. Например, функциональный фрагмент или аналог анти-Notch3 антитела представляет собой фрагмент или аналог, который может связываться с рецептором Notch3 таким образом, чтобы предотвращать или по существу снижать способность рецептора связываться со своими лигандами или инициировать передачу сигнала. Как используют в настоящем описании, «функциональный фрагмент» в отношении антител относится к Fv-, F(ab)- и F(ab')2-фрагментам. «Fv-фрагмент» состоит из димера вариабельного домена одной тяжелой и одной легкой цепи с прочной нековалентной связью (димер VH-VL). Именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют с образованием связывающего мишень участка на поверхности димера VH-VL. В совокупности шесть CDR придают антителу специфичность в отношении связывания мишени. Однако даже отдельный вариабельный домен (или половина Fv, содержащая только три CDR, специфичных в отношении мишени) обладает способностью распознавать и связывать мишень, хотя и с меньшей аффинностью, чем полноразмерный связывающий участок.
«Одноцепочечные Fv-» или «sFv-» фрагменты антитела содержат VH- и VL-домены антитела, где эти домены присутствуют в одной полипептидной цепи. Как правило, полипептид Fv дополнительно содержит полипептидный линкер между VH- и VL-доменами, который позволяет sFv формировать требуемую структуру для связывания мишени.
Термин «димерные антитела» относится к малым фрагментам антитела с двумя антигенсвязывающими участками, где фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) на той же самой полипептидной цепи. При использовании линкера, который является слишком коротким, чтобы обеспечить спаривание между двумя доменами на одной и той же цепи, домены вынуждены спариваться с комплементарными доменами другой цепи и формировать два антигенсвязывающих участка.
F(ab)-фрагмент содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. F(ab')-фрагменты отличаются от F(ab)-фрагментов добавлением нескольких остатков на карбоксильный конец тяжелой цепи домена CH1, включая один или несколько цистеинов из шарнирной области антител. F(ab')-фрагменты получают расщеплением дисульфидной связи между цистеинами шарнирной области продукта расщепления F(ab')2 пепсином. Дополнительные химические сшивки фрагментов антител известны специалистам в данной области.
Термин «моноклональное антитело», как используют в настоящем описании, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными за исключением возможных естественно возникающих мутаций, которые могут присутствовать в минорных количествах. Моноклональные антитела в данной работе, в частности, включают «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных из конкретных видов или принадлежащих конкретному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из других видов или принадлежащих другому классу или подклассу антител, а также фрагментов таких антител, при условии, что они обладают требуемой биологической активностью (патент США No. 4816567; и Morrison et al., Proc. Natl. Acad. Sci USA 81:6851-6855 (1984)). Моноклональные антитела являются высоко специфичными, будучи нацеленными на единственный участок мишени. Более того, в отличие от обычных (поликлональных) препаратов антител, которые, как правило, содержат различные антитела, нацеленные на различные детерминанты (эпитопы), каждое моноклональное антитело нацелено на единственный детерминант на мишени. В дополнение к специфичности моноклональные антитела обладают преимуществом в том, что их можно синтезировать в культуре гибридом, не контаминированной другими иммуноглобулинами. Модификатор «моноклональные» указывает на характер антитела, как полученного из по существу гомогенной популяции антител, и его не следует толковать как требующий получение антитела каким-либо конкретным способом. Например, моноклональные антитела для использования с настоящим изобретением можно выделять из фаговых библиотек антител с использованием хорошо известных способов. Родительские моноклональные антитела для использования согласно настоящему изобретению можно получать гибридомным способом, впервые описанным в Kohler, et al., Nature 256:495 (1975), или можно получать рекомбинантными способами.
«Гуманизированные» формы не являющихся человеческими (например, мышиных) антител представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие связывающиеся с мишенью подпоследовательности антител), которые содержат минимальную последовательность, полученную из иммуноглобулина, не являющегося человеческим. Как правило, гуманизированные антитела будут содержать по существу все, по меньшей мере, из одного и, как правило, двух вариабельных доменов, в которых все или по существу все области CDR соответствуют областям не являющегося человеческим иммуноглобулина, и все или по существу все области FR представляют собой области последовательности матрицы иммуноглобулина человека. Гуманизированное антитело может также содержать, по меньшей мере, часть константной области (Fc) иммуноглобулина, как правило, часть из выбранной матрицы иммуноглобулина человека.
Термины «клетка», «линия клеток» и «культура клеток» включают потомство. Также понятно, что все потомство может не являться в точности идентичным по содержанию ДНК благодаря преднамеренным или случайным мутациям. Включают вариантное потомство, которое обладает той же функцией или биологическим свойством, на которое осуществляли скрининг исходно трансформированной клетки. «Клетки-хозяева», используемые в настоящем изобретении, как правило, являются прокариотическими или эукариотическими хозяевами.
«Трансформация» клеточного организма, клетки или линии клеток ДНК означает введение ДНК в клетку-мишень таким образом, что ДНК может реплицироваться или в качестве внехромосомного элемента или путем интеграции в хромосому. «Трансфекция» клетки или организма ДНК означает поглощение ДНК, например, экспрессирующего вектора, клеткой или организмом, где фактически любые кодирующие последовательности или экспрессируются, или не экспрессируются. Термины «трансфицированная клетка-хозяин» и «трансформированная» означают клетку, в которую введена ДНК. Клетку обозначают «клетка-хозяин», и она может являться или прокариотической или эукариотической. Типичные прокариотические клетки-хозяева включают различные штаммы E. coli. Типичные эукариотические клетки-хозяева являются клетками млекопитающих, такими как клетки яичника китайского хомяка или клетки человеческого происхождения. Введенная последовательность ДНК может быть из того же вида, что и клетка-хозяин, или из другого вида, чем клетка-хозяин, или она может представлять собой гибридную последовательность ДНК, содержащую до некоторой степени чужеродную и до некоторой степени гомологичную ДНК.
Термин «вектор» означает конструкцию ДНК, содержащую последовательность ДНК, которая функционально связана с подходящей регуляторной последовательностью, способной осуществлять экспрессию последовательности ДНК в подходящем хозяине. Такие регуляторные последовательности включают промотор для осуществления транскрипции, необязательную операторную последовательность для регуляции такой транскрипции, последовательность, кодирующую подходящие участки связывания мРНК с рибосомой, и последовательности, которые управляют терминацией транскрипции и трансляции. Вектор может представлять собой плазмиду, фаговую частицу или просто потенциальную геномную вставку. При трансформации в подходящего хозяина, вектор может реплицироваться и функционировать независимо от генома хозяина, или, в некоторых случаях, может интегрироваться в сам геном. В настоящем описании термины «плазмида» и «вектор» иногда используют взаимозаменяемо, так как плазмида является наиболее обычно используемой формой вектора. Однако предполагается, что изобретение включает такие другие формы векторов, которые выполняют эквивалентные функции и которые известны или станут известными в данной области.
«Млекопитающее» для целей лечения относится к любому животному, классифицируемому в качестве млекопитающего, включая человека, домашних и сельскохозяйственных животных, не относящихся к человеку приматов и животных из зоопарка, спортивных или комнатных животных, таких как собаки, лошади, кошки, коровы и т.д.
Слово «метка» при использовании в настоящем документе означает детектируемое соединение или композицию, которую можно конъюгировать непосредственно или опосредованно с молекулой или белком, например, антителом. Метку можно детектировать саму по себе (например, радиоизотопные метки или флуоресцентные метки), или, в случае ферментативной метки, она может катализировать химическое превращение субстратного соединения или композиции, которые являются детектируемыми.
Как используют в настоящем описании, «твердая фаза» означает неводную матрицу, к которой могут прикрепляться антитела по настоящему изобретению. Примеры относящихся к данной работе твердых фаз включают фазы, образованные частично или полностью из стекла (например, стекла с контролируемым размером пор), полисахаридов (например, агарозы), полиакриламидов, полистирола, поливинилового спирта и силиконов. В определенных вариантах осуществления в зависимости от контекста, твердая фаза может содержать лунку планшета для анализа; в других она представляет собой колонку для очистки (например, колонку для аффинной хроматографии).
Как используют в настоящем описании, термин «опосредованное Notch3 нарушение» означает состояние или заболевание, которое характеризуется сверхэкспрессией и/или гиперчувствительностью рецептора Notch3. В частности, его можно толковать, как включающий состояния, связанные с видами рака, такими как немелкоклеточный рак легкого, рак яичника и T-клеточный острый лимфобластный лейкоз. Другие виды рака, включая рак поджелудочной железы, рак предстательной железы, новообразования из плазматических клеток (например, множественную миелому, плазмоклеточный лейкоз и экстрамедуллярную плазмацитому), нейробластому и экстрамедуллярную плазмацитому также входят в объем этого термина. Другие типы заболеваний включают лимфому, синдром Алажиля, заболевание печени, при котором имеет место патологическая васкуляризация, неврологические заболевания, диабет, заболевания, при которых имеет место определение судьбы клеток сосудов, и ревматоидный артрит.
ИММУНОГЕН НА ОСНОВЕ РЕЦЕПТОРА NOTCH3 ДЛЯ ПОЛУЧЕНИЯ АНТИТЕЛ
Растворимые мишени или их фрагменты можно использовать в качестве иммуногенов для получения антител. Антитело направлено против интересующей мишени. Предпочтительно, мишень представляет собой биологически важный полипептид, и введение антитела млекопитающему, страдающему заболеванием или нарушением, может оказывать благоприятное терапевтическое действие у этого млекопитающего. Целые клетки можно использовать в качестве иммуногена для получения антител. Иммуноген можно получать рекомбинантно или получать с использованием синтетических способов. Иммуноген можно также выделять из природного источника.
Для трансмембранных молекул, таких как рецепторы, их фрагменты (например, внеклеточный домен рецептора) можно использовать в качестве иммуногена. Альтернативно, клетки, экспрессирующие трансмембранную молекулу, можно использовать в качестве иммуногена. Такие клетки можно получать из природного источника (например, раковых линий клеток), или они могут являться клетками, которые трансформировали рекомбинантными способами для сверхэкспрессии трансмембранной молекулы. Другие формы иммуногена, пригодные для получения антител, будут очевидны для специалистов в данной области.
Альтернативно, ген или кДНК, кодирующие рецептор Notch3 человека, можно клонировать в плазмиду или другой экспрессирующий вектор и экспрессировать в любой из ряда экспрессионных систем согласно способам, хорошо известным специалистам в данной области. Известны способы клонирования и экспрессии рецептора Notch3 и последовательности нуклеиновой кислоты рецептора Notch3 человека (см., например, патенты США No. 5821332 и 5759546). Вследствие вырожденности генетического кода можно использовать несколько нуклеотидных последовательностей, кодирующих белок или полипептиды рецептора Notch3. Можно варьировать нуклеотидную последовательность путем отбора сочетаний на основе возможных выборов кодонов. Эти сочетания получают согласно стандартному триплетному генетическому коду в приложении к нуклеотидной последовательности, которая кодирует рецептор Notch3 природного происхождения, и можно рассматривать все такие варианты. Любой из этих полипептидов можно использовать для иммунизации животного для получения антител, которые связываются с рецептором Notch3 человека.
Рекомбинантные белки Notch3 из другого вида можно также использовать в качестве иммуногена для получения антител вследствие высокой степени консервативности аминокислотной последовательности Notch3. Сравнение Notch3 человека и мыши показало идентичность аминокислотных последовательностей свыше 90% между двумя видами.
Иммуноген на основе рецептора Notch3 можно, в благоприятной ситуации, экспрессировать в виде слитого белка, который содержит рецептор Notch3, присоединенный к слитому сегменту. Слитый сегмент часто помогает в очистке белка, например, позволяя выделять слитый белок и очищать его аффинной хроматографией, но его также можно использовать для усиления иммуногенности. Слитые белки можно получать культивированием рекомбинантной клетки, трансформированной слитой последовательностью нуклеиновой кислоты, которая кодирует белок, включая слитый сегмент, присоединенный или к C-концу и/или N-концу белка. Слитые сегменты могут включать в качестве неограничивающих примеров Fc-области иммуноглобулина, глутатион-S-трансферазу, β-галактозидазу, полигистидиновый сегмент, способный связываться с двухвалентным ионом металла, и связывающийся с мальтозой белок.
Рекомбинантный белок рецептора Notch3, как описано в примере 1, использовали для иммунизации мышей для получения гибридом, которые продуцируют моноклональные антитела по настоящему изобретению. Иллюстративные полипептиды содержат всю SEQ ID NO: 1 или ее часть, или ее варианты.
ПОЛУЧЕНИЕ АНТИТЕЛ
Антитела по настоящему изобретению можно получать любым подходящим способом, известным в данной области. Антитела по настоящему изобретению могут включать поликлональные антитела. Способы получения поликлональных антител известны специалисту (Harlow, et al., Antibodies: a Laboratory Manual, Cold spring Harbor Laboratory Press, 2nd ed. (1988), которое таким образом приведено в настоящем документе в качестве ссылки в полном объеме).
Например, иммуноген, как описано в примере 1, можно вводить различным животным-хозяевам, включая в качестве неограничивающих примеров кроликов, мышей, крыс и т.д., для индукции выработки сывороток, содержащих поликлональные антитела, специфичные в отношении антигена. Введение иммуногена может включать одну или несколько инъекций иммунизирующего средства и, при необходимости, адъюванта. Разнообразные адъюванты можно использовать для усиления иммунного ответа, в зависимости от вида хозяина, и они включают в качестве неограничивающих примеров адъювант Фрейнда (полный и неполный), минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, плюрониковые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианины морского блюдечка, динитрофенол и потенциально пригодные адъюванты человека, такие как BCG (бацилла Кальметта-Герена) и Corynebacterium parvum. Дополнительные примеры адъювантов, которые можно использовать, включают адъювант MPL-TDM (монофосфориллипид A, синтетический дикриномиколат трегалозы). Протоколы иммунизации хорошо известны в данной области, и их можно осуществлять любым способом, который вызывает иммунный ответ у выбранного животного-хозяина. Адъюванты также хорошо известны в данной области.
Как правило, иммуноген (с адъювантом или без него) инъецируют млекопитающему множественными подкожными или внутрибрюшинными инъекциями, или внутримышечно или посредством IV. Иммуноген может включать полипептид Notch3, слитый белок или их варианты. В зависимости от природы полипептидов (т.е. процента гидрофобности, процента гидрофильности, стабильности, суммарного заряда, изоэлектрической точки и т.д.) может оказаться пригодным конъюгировать иммуноген с белком, о котором известно, что он является иммуногенным у млекопитающих при иммунизации. Такая конъюгация включает или химическую конъюгацию посредством получения производных активных химических функциональных групп как иммуногена, так и иммуногенного белка, предназначенных для конъюгации, таким образом, чтобы сформировать ковалентную связь, или способом на основе слитого белка, или другими способами, известными специалисту. Примеры таких иммуногенных белков включают в качестве неограничивающих примеров гемоцианин морского блюдечка, овальбумин, сывороточный альбумин, бычий тироглобулин, соевый ингибитор трипсина и разнообразные T-хелперные пептиды. Разнообразные адъюванты можно использовать для усиления иммунного ответа, как описано выше.
Антитела по настоящему изобретению включают моноклональные антитела. Моноклональные антитела представляют собой антитела, которые распознают единственный антигенный участок. Их однородная специфичность делает моноклональные антитела гораздо более пригодными, чем поликлональные антитела, которые обычно содержат антитела, которые распознают множество различных антигенных участков. Моноклональные антитела можно получать с использованием гибридомной технологии, как например, описанной в Kohler, et al., Nature 256:495 (1975); патенте США No. 4376110; Harlow, et al., Antibodies: a Laboratory Manual, Cold spring Harbor Laboratory Press, 2nd ed. (1988) и Hammerling, et al., Monoclonal Antibodies and T-Cell Hybridomas, Elsevier (1981), способами рекомбинантных ДНК или другими способами, известными специалисту. Другие примеры способов, которые можно использовать для получения моноклональных антител, включают в качестве неограничивающих примеров способ на основе B-клеточных гибридом человека (Kosbor, et al., Immunology Today 4:72 (1983); Cole, et al., Proc Natl Acad Sci USA 80:2026 (1983)) и способ на основе EBV-гибридомы (Cole, et al., Monoclonal Antibodies and Cancer Therapy, pp. 77-96, Alan R. Liss (1985)). Такие антитела могут относиться к любому классу иммуноглобулинов, включая IgG, IgM, IgE, IgA, IgD и любой их подкласс. Гибридому, продуцирующую mAb по данному изобретению, можно культивировать in vitro или in vivo.
В модели гибридом, хозяина, такого как мышь, гуманизированная мышь, мышь с иммунной системой человека, хомяк, кролик, верблюд или любое другое подходящее животное-хозяин, иммунизируют для получения лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфически связываться с белком, используемым для иммунизации. Альтернативно, лимфоциты можно иммунизировать in vitro. Затем производят слияние лимфоцитов с миеломными клетками с использованием подходящего вещества, вызывающего слияние, такого как полиэтиленгликоль, для получения гибридомных клеток (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)).
Как правило, при получении продуцирующих антитела гибридом используют или лимфоциты периферической крови («PBL»), если требуются клетки человеческого происхождения, или клетки селезенки, или клетки лимфатического узла, если требуются источники из млекопитающих не относящихся к человеку. Затем производят слияние лимфоцитов с иммортализованной линией клеток с использованием подходящего вещества, вызывающего слияние, такого как полиэтиленгликоль, для получения гибридомной клетки (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)). Иммортализованные линии клеток, как правило, представляют собой трансформированные клетки млекопитающих, в частности, миеломные клетки, происходящие из грызунов, быка или человека. Как правило, используют миеломную линию клеток крысы или мыши. Гибридомные клетки можно культивировать в подходящей культуральной среде, которая предпочтительно содержит одно или несколько веществ, которые ингибируют рост или выживаемость неподвергшихся слиянию иммортализованных клеток. Например, если у родительских клеток отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), то культуральная среда для гибридом, как правило, будет включать гипоксантин, аминоптерин и тимидин («среда HAT»), вещества, которые предотвращают рост клеток с дефектом HGPRT.
Предпочтительные иммортализованные линии клеток представляют собой такие линии, которые эффективно подвергаются слиянию, поддерживают стабильную выработку антител на высоком уровне отобранными продуцирующими антитела клетками и являются чувствительными к среде, такой как среда HAT. Среди них миеломные линии клеток представляют собой мышиные миеломные линии, такие как линии, происходящие из опухолей мышей MOPC-21 и MPC-11, доступные в Salk Institute Cell Distribution Center, San Diego, Calif., и клетки SP-2/0 или X63-Ag8-653, доступные в American Type Culture Collection (ATCC), Manassas, VA, USA. Линии клеток миеломы человека и гетеромиеломы мыши-человека также описаны относительно получения человеческих моноклональных антител (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al, Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., pp. 51-63 (1987)). Также можно использовать мышиную миеломную линию клеток NSO (European Collection of Cell Cultures, Salisbury, Wilshire, UK).
Культуральную среду, в которой выращивают гибридомные клетки, анализируют относительно выработки моноклональных антител, направленных против Notch3. Специфичность связывания моноклональных антител, вырабатываемых гибридомными клетками, определяют иммунопреципитацией или анализом связывания in vitro, таким как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Такие способы известны в данной области и доступны для специалиста. Аффинность связывания моноклонального антитела к Notch3, например, можно определить с помощью анализа Скэтчарда (Munson et al., Anal. Biochem., 107:220 (1980)).
После идентификации тех гибридомных клеток, которые вырабатывают антитела требуемой специфичности, аффинности и/или активности, клоны можно субклонировать посредством способов серийных разведений и выращивать с помощью общепринятых способов (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, pp.59-103 (1986)). Подходящие культуральные среды для этой цели включают, например, среду Игла в модификации Дульбекко (D-MEM) или среду RPMI-1640. Кроме того, гибридомные клетки можно выращивать in vivo в виде асцитных опухолей в животном.
Моноклональные антитела, секретируемые субклонами, соответствующим образом выделяют из культуральной среды, асцитической жидкости или сыворотки с помощью общепринятых способов очистки иммуноглобулинов, таких как, например, протеин-A-сефароза, хроматография с гидроксилапатитом, гель-эксклюзионная хроматография, гель-электрофорез, диализ или аффинная хроматография.
В данной области существует множество способов продукции моноклональных антител, и, таким образом, изобретение не ограничено их продукцией исключительно в гибридомах. Например, моноклональные антитела можно получать способами рекомбинантных ДНК, такими как способы, описанные в патенте США No. 4816567. В этом контексте термин «моноклональное антитело» относится к антителу, полученному из одиночного эукариотического, фагового или прокариотического клона. ДНК, кодирующую моноклональные антитела по изобретению, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител, или такие цепи из человека, гуманизированные или из других источников) (Innis, et al. In PCR Protocols. A Guide to Methods and Applications, Academic (1990), Sanger, et al., Proc Natl Acad Sci 74:5463 (1977)). Гибридомные клетки служат в качестве источника такой ДНК. После выделения ДНК можно поместить в экспрессирующие векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки NS0, клетки COS обезьяны, клетки яичника китайского хомяка (CHO) или миеломные клетки, которые в противном случае не продуцируют белок иммуноглобулина, для получения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. ДНК также можно модифицировать, например, помещением кодирующей последовательности тяжелой и легкой цепи константных доменов человека вместо гомологичных последовательностей мыши (патент США No. 4816567; Morrison, et al., Proc Natl Acad Sci USA 81:6851 (1984)) или ковалентным присоединением к кодирующей последовательности иммуноглобулина всей кодирующей последовательности не являющегося иммуноглобулином полипептида или ее части. На такой не относящийся к иммуноглобулину полипептид можно заменить константные домены антитела по изобретению, или на него можно заменить вариабельные домены одного из антигенсвязывающих участков антитела по изобретению для получения химерного бивалентного антитела.
Антитела могут представлять собой моновалентные антитела. Способы получения моновалентных антител хорошо известны в данной области. Например, в одном из способов используют рекомбинантную экспрессию легкой цепи и модифицированной тяжелой цепи иммуноглобулина. Тяжелую цепь обрезают, как правило, в любой точке Fc-области таким образом, чтобы предотвратить перекрестное связывание тяжелой цепи. Альтернативно, соответствующие остатки цистеина замещают на другой аминокислотный остаток или удаляют таким образом, чтобы предотвратить перекрестное связывание.
Фрагменты антител, которые распознают специфические эпитопы, можно получать известными способами. Традиционно, эти фрагменты получали посредством протеолитического расщепления интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24:107-117 (1992); Brennan et al., Science, 229:81 (1985)). Например, фрагменты Fab и F(ab')2 по изобретению можно получать протеолитическим расщеплением молекул иммуноглобулина с использованием ферментов, таких как папаин (для получения Fab-фрагментов) или пепсин (для получения F(ab')2-фрагментов). F(ab')2-фрагменты содержат вариабельную область, константную область легкой цепи и CH1-домен тяжелой цепи. Однако эти фрагменты в настоящее время можно получать непосредственно с помощью рекомбинантных клеток-хозяев. Например, фрагменты антител можно выделять из фаговой библиотеки антител. Альтернативно, фрагменты F(ab')2-SH можно выделять непосредственно из E. coli и химически соединять с образованием фрагментов F(ab')2 (Carter et al., Bio/Technology 10:163-167 (1992)). В соответствии с другим подходом фрагменты F(ab')2 можно выделять непосредственно из рекомбинантной культуры клеток-хозяев. Другие способы получения фрагментов антител будут очевидны квалифицированному специалисту. В других вариантах осуществления антителом выбора является одноцепочечный Fv-фрагмент (Fv) (патентная заявка PCT WO 93/16185).
Для некоторых целей, включая применение антител in vivo у человека и анализы для детекции in vitro, может являться предпочтительным использование химерных, гуманизированных или человеческих антител. Химерное антитело представляет собой молекулу, в которой различные части антитела получены из различных видов животных, такое как антитела, содержащие вариабельную область, происходящую из мышиных моноклональных антител, и константную область иммуноглобулина человека. Способы получения химерных антител известны в данной области. См., например, Morrison, Science 229:1202 (1985); Oi, et al., BioTechniques 4:214 (1986); Gillies, et al., J Immunol Methods 125:191 (1989); патенты США No. 5807715, 4816567 и 4816397, которые приведены в настоящем документе в качестве ссылки в полном объеме.
Гуманизированное антитело разработано для получения большей степени гомологии с иммуноглобулином человека по сравнению с моноклональными антителами животного происхождения. Гуманизация представляет собой способ получения химерного антитела, где, по существу, менее интактный вариабельный домен человека замещен соответствующей последовательностью вида, не относящегося к человеку. Гуманизированные антитела представляют собой молекулы антител, полученные из не относящихся к человеку видов, которые связываются с требуемым антигеном и которые обладают одной или несколькими определяющими комплементарность областями (CDR) из не относящихся к человеку видов и каркасными (FR) областями из молекулы иммуноглобулина человека. Часто каркасные остатки в каркасных областях человека замещают на соответствующий остаток из CDR донорного антитела для изменения, предпочтительно, улучшения связывания с антигеном. Эти каркасные замещения идентифицируют способами, хорошо известными в данной области, например, моделированием взаимодействий CDR и каркасных остатков для идентификации каркасных остатков, важных для связывания с антигеном, и сравнением последовательностей для идентификации нестандартных каркасных остатков в конкретных положениях. См., например, патент США No. 5585089; Riechmann, et al., Nature 332:323 (1988), которые приведены в настоящем документе в качестве ссылки в полном объеме. Антитела можно гуманизировать с использованием множества способов, известных в данной области, включая, например, привитие CDR (EP 239400; публикация PCT WO 91/09967; патенты США No. 5225539, 5530101 и 5585089), рекомбинацию поверхностных остатков (EP 592106, EP 519596, Padlan, Molecular Immunology 28:489 (1991); Studnicka, et al., Protein Engineering 7:805 (1994); Roguska, et al., Proc Natl Acad Sci USA 91:969 (1994)) и рекомбинацию цепей (патент США No. 5565332).
Как правило, гуманизированные антитела содержат один или несколько аминокислотных остатков, введенных в них из не относящегося к человеку источника. Эти не принадлежащие человеку аминокислотные остатки часто обозначают как «импортные» остатки, которые, как правило, взяты из «импортного» вариабельного домена. Гуманизацию, главным образом, можно проводить в соответствии со способами Winter и коллег (Jones et al. Nature 321:522-525 (1986); Riechmann et al. Nature 332:323-327 (1988); Verhoeyen et al. Science 239:1534-1536 (1988)) посредством замены соответствующих последовательностей антитела человека не относящимися к человеку CDR или последовательностями CDR. Таким образом, такие «гуманизированные» антитела представляют собой химерные антитела (патент США No. 4816567), где, по существу, менее интактный вариабельный домен человека замещен соответствующей последовательностью вида, не относящегося к человеку. На практике, гуманизированные антитела, как правило, представляют собой антитела человека, в которых некоторые остатки CDR и, возможно, некоторые остатки FR замещены остатками из аналогичных участков антител грызунов.
Кроме того, важно, чтобы гуманизированные антитела сохраняли высокую аффинность в отношении антигена и другие положительные биологические свойства. Для достижения этой цели согласно предпочтительному способу гуманизированные антитела получают с помощью процесса анализа родительских последовательностей и различных предполагаемых гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются широко доступными и известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и выводят на экран возможные трехмерные конформационные структуры выбранных предполагаемых последовательностей иммуноглобулинов. Исследование этих выведенных на экран данных позволяет проводить анализ возможной роли определенных остатков в функционировании предполагаемой последовательности иммуноглобулина, т.е. анализ остатков, которые влияют на способность предполагаемого иммуноглобулина связывать свой антиген. Таким образом, остатки FR можно отбирать и комбинировать из реципиентной и импортированной последовательностей таким образом, чтобы максимизировать требуемые свойства антител, такие как повышенная аффинность к антигену(ам)-мишени(ям), хотя именно остатки CDR непосредственно и наиболее существенно влияют на связывание с антигеном.
Выбор вариабельных доменов человека, как легких, так и тяжелых цепей, для использования при получении гуманизированных антител является важным для снижения антигенности. В соответствии с иллюстративным способом, так называемым способом «наилучшего соответствия», последовательность вариабельного домена не относящегося к человеку антитела анализируют относительно целой библиотеки известных последовательностей вариабельных доменов человека. Затем последовательность человека, которая является наиболее сходной с последовательностью не относящегося к человеку родительского антитела, считают приемлемой в качестве FR человека для гуманизированного антитела (Sims et al., J. Immunol., 151:2296 (1993); Chothia et al, J. Mol. Biol., 196:901 (1987)). В другом способе используют конкретную каркасную область, полученную из консенсусной последовательности всех антител человека конкретной подгруппы легких или тяжелых цепей. Одну и ту же каркасную область можно использовать для нескольких различных гуманизированных антител (Carter et al, Proc. Natl. Acad. Sci. USA, 89:4285 (1992); Presta et al, J. Immunol., 151:2623 (1993)).
Полностью человеческие антитела, в частности, требуются для терапевтического лечения человека. Человеческие антитела можно получать множеством способов, известных в данной области, включая способы фагового дисплея, описанные выше с использованием библиотек антител, полученных из последовательностей иммуноглобулинов человека. См. также патенты США No. 4444887 и 4716111 и публикации PCT WO 98/46645, WO 98/50433, WO 98/24893, WO 98/16654, WO 96/34096, WO 96/33735 и WO 91/10741, каждая из которых приведена в настоящем документе в качестве ссылки в полном объеме. Способы Cole, et al. и Boerder, et al. также доступны для получения моноклональных антител человека (Cole, et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Riss (1985); и Boerner, et al., J Immunol 147:86 (1991)).
Человеческие антитела можно также получать с использованием трансгенных мышей, которые не способны экспрессировать функциональные эндогенные иммуноглобулины, но которые могут экспрессировать гены иммуноглобулинов человека. Например, генные комплексы тяжелой и легкой цепи иммуноглобулина человека можно вводить случайным образом или путем гомологичной рекомбинации в мышиные эмбриональные стволовые клетки. Альтернативно, вариабельную область, константную область и область разнообразия человека можно вводить в эмбриональные стволовые клетки мыши в дополнение к генам тяжелой и легкой цепи человека. Гены тяжелой и легкой цепи иммуноглобулина мыши можно сделать нефункциональными отдельно от введения или одновременно с введением локусов иммуноглобулина человека путем гомологичной рекомбинации. В частности, гомозиготная делеция JH-области предотвращает продукцию эндогенных антител. Модифицированные эмбриональные стволовые клетки размножают и микроинъецируют в бластоцисты для получения химерных мышей. Химерных мышей затем разводят для получения гомозиготного потомства, которое экспрессирует человеческие антитела. См., например, Jakobovits, et al., Proc Natl Acad Sci USA 90:2551 (1993); Jakobovits, et al., Nature 362:255 (1993); Bruggermann, et al., Year in Immunol 7:33 (1993); Duchosal, et al., Nature 355:258 (1992)). Трансгенных мышей иммунизируют общепринятым образом выбранным антигеном, например, полноразмерным полипептидом по изобретению или его частью. Моноклональные антитела, направленные против антигена, можно получать из иммунизированных трансгенных мышей с использованием общепринятой гибридомной технологии. Трансгены иммуноглобулина человека, которые несут трансгенные мыши, рекомбинируют в ходе дифференциации B-клеток и затем подвергаются мутациям с переключением класса и соматическим мутациям. Таким образом, с использованием такого способа возможно получение терапевтически пригодных антител IgG, IgA, IgM и IgE. Для обзора этой технологии получения человеческих антител см. Lonberg, et al., Int Rev Immunol 13:65-93 (1995). Для подробного рассмотрения этого способа получения человеческих антител и человеческих моноклональных антител и протоколов получения таких антител см., например, публикации PCT WO 98/24893, WO 92/01047, WO 96/34096, WO 96/33735, европейский патент No. 0598877, патенты США No. 5413923, 5625126, 5633425, 5569825, 5661016, 5545806, 5814318, 5885793, 5916771 и 5939598, которые приведены в настоящем документе в качестве ссылки в полном объеме. Кроме того, компании, такие как Abgenix, Inc. (Freemont, Calif.), Genpharm (San Jose, Calif.) и Medarex, Inc. (Princeton, N.J.), могут быть заняты получением человеческих антител, направленных против выбранного антигена с использованием технологии, сходной с описанной выше.
Также mAb человека можно получать путем иммунизации мышей, которым трансплантировали лейкоциты периферической крови, спленоциты или костный мозг человека (например, способы триом XTL). Полностью человеческие антитела, которые распознают выбранный эпитоп, можно получать с использованием способа, обозначенного как «управляемая селекция». В этом способе отобранное не относящееся к человеку моноклональное антитело, например мышиное антитело, используют для регурилования селекции полностью человеческого антитела, распознающего тот же самый эпитоп (Jespers, et al., Bio/technology 12:899 (1988)).
Кроме того, антитела к полипептидам по изобретению, в свою очередь, можно использовать для получения антиидиотипических антител, которые «мимикрируют» полипептиды по изобретению с использованием способов, хорошо известных специалистам в данной области (см., например, Greenspan, et al., FASEB J 7:437 (1989); Nissinoff, J Immunol 147:2429 (1991)). Например, антитела, которые связываются с полипептидом и конкурентно ингибируют мультимеризацию полипептида и/или связывание полипептида по изобретению с лигандом, можно использовать для получения антиидиотипов, которые «мимикрируют» мультимеризационный и/или связывающий домен полипептида и, как следствие, связываются с полипептидом и нейтрализуют полипептид и/или его лиганд. Такие нейтрализующие антиидиотипы или Fab-фрагменты таких антиидиотипов можно использовать в терапевтических схемах лечения для нейтрализации полипептидного лиганда. Например, такие антиидиотипические антитела можно использовать для связывания с полипептидом по изобретению и/или для связывания с его лигандами/рецепторами и, таким образом, блокировать его биологическую активность.
Антитела по настоящему изобретению могут представлять собой биспецифические антитела. Биспецифические антитела представляют собой моноклональные, предпочтительно человеческие или гуманизированные, антитела, которые обладают специфичностями связывания в отношении, по меньшей мере, двух различных антигенов. В настоящем изобретении одну из специфичностей связывания можно направить на Notch3, другая может относиться к любому другому антигену и, предпочтительно, белку клеточной поверхности, рецептору, субъединице рецептора, тканеспецифичному антигену, белку вирусного происхождения, кодируемому вирусом белку оболочки, белку бактериального происхождения или бактериальному поверхностному белку и т.д.
Способы получения биспецифических антител хорошо известны. Традиционно, рекомбинантное получение биспецифических антител основано на совместной экспрессии двух иммуноглобулиновых пар тяжелая цепь/легкая цепь, где две тяжелые цепи обладают различными специфичностями (Milstein et al., Nature, 305:537 (1983)). Вследствие случайной сборки тяжелых и легких цепей иммуноглобулинов такие гибридомы (квадромы) продуцируют смесь потенциально из 10 различных молекул антител, из которых только одна обладает правильной биспецифической структурой. Очистку правильных молекул обычно осуществляют посредством стадий аффинной хроматографии. Аналогичные способы описаны в WO 93/08829 и в Traunecker et al, EMBO J., 10:3655 (1991).
Вариабельные домены антител с требуемыми специфичностями связывания (связывающими участками антитело-антиген) можно подвергнуть слиянию с последовательностями константного домена иммуноглобулина. Слияние предпочтительно осуществляют с константным доменом тяжелой цепи иммуноглобулина, содержащим, по меньшей мере, часть шарнирной области, CH2- и CH3-области. Оно может содержать первую константную область тяжелой цепи (CH1), содержащую участок, необходимый для связывания легкой цепи, присутствующий, по меньшей мере, в одной из слитых конструкций. ДНК, кодирующие слитые конструкции тяжелой цепи иммуноглобулина и, при желании, легкую цепь иммуноглобулина, вставляют в отдельные экспрессирующие векторы и совместно трансформируют в организм подходящего хозяина. Дополнительные подробности получения биспецифических антител см., например, в Suresh, et al., Meth In Enzym 121:210 (1986).
Также настоящее изобретение относится к антителам-гетероконъюгатам. Антитела-гетероконъюгаты образованы двумя ковалентно связанными антителами. Такие антитела, например, предложены для нацеливания клеток иммунной системы на нежелательные клетки (патент США No. 4676980). Предполагается, что антитела можно получать in vitro с использованием известных способов синтетической белковой химии, включая способы с использованием поперечно-сшивающих средств. Например, иммунотоксины можно конструировать с использованием реакции дисульфидного обмена или путем образования тиоэфирной связи. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат, и реагенты, описанные, например, в патенте США No. 4676980.
Кроме того, можно получать однодоменные антитела к Notch3. Примеры этой технологии описаны в WO 94/25591 для антител, происходящих из тяжелой цепи Ig Camelidae, а также в US 2003/0130496, где описано получение однодоменных полностью человеческих антител из фаговых библиотек.
Также можно получать связывающие молекулы из одной пептидной цепи, в которых связаны Fv-области тяжелой и легкой цепи. Одноцепочечные антитела («scFv») и способ их конструирования описан в патенте США No. 4946778. Альтернативно, Fab можно конструировать и экспрессировать сходными способами. Все полностью и частично человеческие антитела менее иммуногенны, чем полностью мышиные mAb, и фрагменты и одноцепочечные антитела также менее иммуногенны.
Антитела или фрагменты антител можно выделять из фаговых библиотек антител, получаемых с использованием способов, описанных в McCafferty, et al., Nature 348:552 (1990). В Clarkson, et al., Nature 352:624 (1991) и Marks, et al., J Mol Biol 222:581 (1991) описано выделение мышиных и человеческих антител соответственно с использованием фаговых библиотек. В последующих публикациях описано получение высокоаффинных (порядок нМ) человеческих антител путем рекомбинации цепей (Marks, et al., Bio/Technology 10:779 (1992)), а также комбинаторной инфекции и рекомбинации in vivo в качестве стратегии для конструирования очень больших фаговых библиотек (Waterhouse, et al., Nuc Acids Res 21:2265 (1993)). Таким образом, для выделения моноклональных антител эти способы являются приемлемыми альтернативами традиционным гибридомным способам получения моноклональных антител.
ДНК также можно модифицировать, например, замещением последовательности, кодирующей константные домены тяжелой и легкой цепи человека вместо гомологичных мышиных последовательностей (патент США No. 4816567; Morrison, et al., Proc Natl Acad Sci USA 81:6851 (1984)).
Другой альтернативой является использование электрического слияния вместо химического слияния для получения гибридом. Этот способ хорошо налажен. Вместо слияния можно также трансформировать B-клетку, чтобы сделать ее иммортализованной с использованием, например, вируса Эпштейна-Барра или трансформирующего гена. См., например, «Continuously Proliferating Human Cell Lines Synthesizing Antibody of Predetermined Specificity», Zurawaki, et al., в Monoclonal Antibodies, ed. by Kennett, et al., Plenum Press, pp.19-33. (1980)). MAb анти-Notch3 можно получать иммунизацией грызунов (например, мышей, крыс, хомяков и морских свинок) белком Notch3, слитым белком или его фрагментами, экспрессируемыми или эукариотическими или прокариотическими системами. Для иммунизации можно использовать других животных, например, не относящихся к человеку приматов, трансгенных мышей, экспрессирующих иммуноглобулины, и мышей с тяжелым комбинированным иммунодефицитом (SCID), которым трансплантированы B-лимфоциты человека. Гибридомы можно получать общепринятыми способами слияния B-лимфоцитов из иммунизированных животных с миеломными клетками (например, Sp2/0 и NSO), как описано ранее (Kӧhler, et al., Nature 256:495 (1975)). Кроме того, антитела против Notch3 можно получать скринингом библиотек с рекомбинантными одноцепочечными Fv или Fab из B-лимфоцитов человека в системах фагового дисплея. Специфичность mAb к Notch3 можно тестировать с помощью ELISA, вестерн-иммуноблоттинга или других иммунохимических способов. Ингибиторную активность антител в отношении активации комплемента можно оценивать гемолитическими анализами с использованием сенсибилизированных RBC из курицы или овцы для классического пути комплемента. Гибридомы в положительных лунках клонируют лимитирующим разведением. Антитела очищают, чтобы охарактеризовать в отношении специфичности к Notch3 человека описанными выше анализами.
ИДЕНТИФИКАЦИЯ АНТИ-NOTCH3 АНТИТЕЛ
Настоящее изобретение относится к моноклональным антителам-антагонистам, которые ингибируют и нейтрализуют действие Notch3. В частности, антитела по настоящему изобретению связываются с Notch3 и ингибируют его активацию. Антитела по настоящему изобретению включают антитела, обозначенные как 256A-4 и 256A-8, которые описаны в данной работе. Настоящее изобретение также относится к антителам, которые связываются с тем же эпитопом, как и одно из этих антител.
Предполагаемые антитела против Notch3 тестировали твердофазным иммуноферментным анализом (ELISA), вестерн-иммуноблоттингом или другими иммунохимическими способами. Проделанные анализы для получения характеристик индивидуальных антител описаны в примерах.
Антитела по изобретению включают в качестве неограничивающих примеров поликлональные, моноклональные, моновалентные, биспецифические, гетероконъюгатные, полиспецифические, человеческие, гуманизированные или химерные антитела, одноцепочечные антитела, однодоменные антитела, Fab-фрагменты, F(ab')-фрагменты, фрагменты, получаемые из экспрессирующей Fab библиотеки, антиидиотипические (анти-Id) антитела (включая, например, анти-Id-антитела к антителам по изобретению) и эпитопсвязывающие фрагменты любого из вышеупомянутого.
Антитела могут представлять собой антигенсвязывающие фрагменты антител человека по настоящему изобретению и включают в качестве неограничивающих примеров Fab, Fab' и F(ab')2, Fd, одноцепочечные Fv (scFv), одноцепочечные антитела, связанные дисульфидной связью Fv (sdFv), и однодоменные антитела, содержащие или VL- или VH-домен. Антигенсвязывающие фрагменты антител, включая одноцепочечные антитела, могут содержать вариабельную область(и) отдельно или в сочетании со всем или частью из следующего: шарнирная область, CH1-, CH2- и CH3-домены. Также относятся к изобретению антигенсвязывающие фрагменты, содержащие любое сочетание вариабельной области(ей) с шарнирной областью, CH1-, CH2- и CH3-доменами. Антитела по изобретению могут происходить из любого животного, включая птиц и млекопитающих. Предпочтительно, антитела происходят из человека, не относящихся к человеку приматов, грызунов (например, мыши и крысы), осла, овцы, кролика, козла, морской свинки, верблюда, лошади или курицы.
Как используют в настоящем описании, «человеческие антитела» включают антитела, содержащие аминокислотную последовательность иммуноглобулина человека, и включают антитела, выделенные из библиотек иммуноглобулинов человека или из животных, трансгенных по одному или нескольким иммуноглобулинам человека, и которые не экспрессируют эндогенные иммуноглобулины, как описано ниже и, например, в патенте США No. 5939598, Kucherlapati, et al.
Антитела по настоящему изобретению могут являться моноспецифическими, биспецифическими, триспецифическими или обладают более высокой полиспецифичностью. Полиспецифические антитела могут являться специфическими в отношении различных эпитопов Notch3 или могут являться специфическими в отношении как Notch3, так и в отношении гетерологичного эпитопа, такого как гетерологичный полипептид или материал твердого носителя. См., например, публикации PCT WO 93/17715, WO 92/08802, WO 91/00360, WO 92/05793; Tutt, et al., J Immunol 147:60 (1991); патенты США No. 4474893, 4714681, 4925648, 5573920, 5601819; Kostelny, et al., J Immunol 148:1547 (1992).
Антитела по настоящему изобретению можно описать или определить в терминах эпитопа(ов) или части(ей) Notch3, которые они распознают или с которыми они специфически связываются. Эпитоп(ы) или полипептидную часть(и) можно определить, как описано в настоящем документе, например, по положениям N-конца и C-конца, по размеру смежных аминокислотных остатков, или как представлено в таблицах и на фигурах.
Антитела по настоящему изобретению можно также описать или определить в терминах их перекрестной активности. Антитела, которые связываются с полипептидами Notch3, которые обладают, по меньшей мере, 95%, по меньшей мере, 90%, по меньшей мере, 85%, по меньшей мере, 80%, по меньшей мере, 75%, по меньшей мере, 70%, по меньшей мере, 65%, по меньшей мере, 60%, по меньшей мере, 55% и, по меньшей мере, 50% идентичностью (как вычислено с использованием способов, известных в данной области и описанных в настоящем документе), с Notch3, также относятся к настоящему изобретению. Антитела против Notch3 могут также связываться с KD менее чем приблизительно 10-7 М, менее чем приблизительно 10-6 М или менее чем приблизительно 10-5 М с другими белками, такими как антитела против Notch3 из видов, отличных от тех, против которых направлены антитела против Notch3.
В конкретных вариантах осуществления антитела по настоящему изобретению перекрестно реагируют с гомологами Notch3 человека из обезьян и их соответствующими эпитопами. В конкретном варианте осуществления выше описанная перекрестная активность проявляется в отношении любого одиночного специфического антигенного или иммуногенного полипептида, или сочетания(ий) специфических антигенных и/или иммуногенных полипептидов, описанных в настоящем документе.
Кроме того, настоящее изобретение относится к антителам, которые связываются с полипептидами, кодируемыми полинуклеотидами, которые гибридизуются с полинуклеотидом, кодирующим Notch3, в строгих условиях гибридизации. Антитела по настоящему изобретению можно описать или определить в терминах их аффинности связывания с полипептидом по изобретению. Предпочтительные аффинности связывания включают аффинности с равновесной константой диссоциации или KD от 10-8 до 10-15 М, от 10-8 до 10-12 М, от 10-8 до 10-10 М или от 10-10 до 10-12 М. Изобретение также относится к антителам, которые конкурентно ингибируют связывание антитела с эпитопом по изобретению, как определяют любым способом, известным в данной области для определения конкурентного связывания, например, описанными в настоящем документе иммуноанализами. В предпочтительных вариантах осуществления антитело конкурентно ингибирует связывание с эпитопом, по меньшей мере, на 95%, по меньшей мере, на 90%, по меньшей мере, на 85%, по меньшей мере, на 80%, по меньшей мере, на 75%, по меньшей мере, на 70%, по меньшей мере, на 60% или, по меньшей мере, на 50%.
ВЕКТОРЫ И КЛЕТКИ-ХОЗЯЕВА
В другом аспекте настоящее изобретение относится к выделенным последовательностям нуклеиновой кислоты, кодирующим антитело, как описано в настоящем документе, векторным конструкциям, содержащим нуклеотидную последовательность, кодирующую антитела по настоящему изобретению, клеткам-хозяевам, содержащим такой вектор, и рекомбинантным способам получения антитела.
Для рекомбинантного получения антитела, кодирующую его нуклеиновую кислоту выделяют и вставляют в реплицирующийся вектор для последующего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую антитело, легко выделять и секвенировать с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Общепринятые способы клонирования и трансформации можно использовать для получения линий клеток, экспрессирующих антитела по настоящему изобретению.
ВЕКТОРЫ
Доступно множество векторов. Компоненты вектора, как правило, включают в качестве неограничивающих примеров одно или несколько из следующего: сигнальная последовательность, ориджин репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции. Рекомбинантные экспрессирующие векторы, содержащие нуклеотидную последовательность, кодирующую антитела по настоящему изобретению, можно получать с использованием хорошо известных способов. Экспрессирующие векторы содержат нуклеотидную последовательность, функционально связанную с подходящими транскрипционными или трансляционными регуляторными нуклеотидными последовательностями, такими как последовательности, происходящие из генов млекопитающих, микробных, вирусных генов или генов насекомых. Примеры регуляторных последовательностей включают транскрипционные промоторы, операторы, энхансеры, участки мРНК для связывания с рибосомами и/или другие подходящие последовательности, которые управляют транскрипцией и инициацией и терминацией трансляции. Нуклеотидные последовательности «функционально связаны» тогда, когда регуляторная последовательность функционально соотносится с нуклеотидной последовательностью подходящего полипептида. Таким образом, промоторная нуклеотидная последовательность функционально связана, например, с последовательностью тяжелой цепи антитела, если промоторная нуклеотидная последовательность управляет транскрипцией подходящей нуклеотидной последовательности.
Кроме того, последовательности, кодирующие подходящие сигнальные пептиды, которые не связаны с последовательностями тяжелой и/или легкой цепи антитела в природе, можно вставлять в экспрессирующие векторы. Например, можно произвести слияние нуклеотидной последовательности сигнального пептида (секреторной лидерной последовательности) в рамке с последовательностью полипептида таким образом, что происходит секреция антитела в периплазматическое пространство или в среду. Сигнальный пептид, который является функциональным в предполагаемых клетках-хозяевах, усиливает внеклеточную секрецию подходящего антитела. Сигнальный пептид можно отщеплять от полипептида при секреции антитела из клетки. Примеры таких секреторных сигналов хорошо известны и включают, например, сигналы, описанные в патентах США No. 5698435, 5698417 и 6204023.
Вектор может представлять собой плазмидный вектор, одно- или двухцепочечный фаговый вектор или одно- или двухцепочечный вектор на основе РНК- или ДНК-вируса. Такие векторы можно вводить в клетки в виде полинуклеотидов хорошо известными способами введения ДНК и РНК в клетки. Векторы, в случае фаговых и вирусных векторов, также можно вводить в клетки в виде упакованного или инкапсулированного вируса хорошо известными способами инфекции и трансдукции. Вирусные векторы могут являться компетентными в отношении репликации или дефектными в отношении репликации. В последнем случае размножение вируса, как правило, будет происходить только в комплементирующих клетках-хозяевах. Бесклеточные системы трансляции можно также применять для получения белка с использованием РНК, происходящих из настоящих конструкций ДНК. Такие векторы могут включать нуклеотидную последовательность, кодирующую константную область молекулы антитела (см., например, публикации PCT WO 86/05807 и WO 89/01036; и патент США No. 5122464), и вариабельный домен антитела можно клонировать в такой вектор для экспрессии полноразмерной тяжелой или легкой цепи.
КЛЕТКИ-ХОЗЯЕВА
Антитела по настоящему изобретению можно экспрессировать в подходящей клетке-хозяине. Примеры клеток-хозяев, пригодных в настоящем изобретении, включают прокариотические, дрожжевые клетки или клетки высших эукариот и включают в качестве неограничивающих примеров микроорганизмы, такие как бактерии (например, E. coli, B. subtilis), трансформированные экспрессирующими векторами на основе рекомбинантных бактериофаговых ДНК, плазмидных ДНК или космидных ДНК, содержащими кодирующие последовательности антител; дрожжи (например, Saccharomyces, Pichia), трансформированные рекомбинантными дрожжевыми экспрессирующими векторами, содержащими кодирующие последовательности антител; системы на основе клеток насекомых, инфицированных рекомбинантными вирусными экспрессирующими векторами (например, Baculovirus), содержащими кодирующие последовательности антител; системы на основе растительных клеток, инфицированных рекомбинантными вирусными экспрессирующими векторами (например, вирусом капустной мозаики, CaMV; вирусом табачной мозаики, TMV) или трансформированных рекомбинантными плазмидными экспрессирующими векторами (например, плазмидой Ti), содержащими кодирующие последовательности антител; или системы на основе клеток млекопитающих (например, клеток COS, CHO, BHK, 293, 3T3), несущих рекомбинантные экспрессионные конструкции, содержащие промоторы, происходящие из генома клеток млекопитающих (например, промотор металлотионеина) или из вирусов млекопитающих (например, поздний промотор аденовируса; промотор вируса коровьей оспы 7.5K).
Прокариоты, пригодные в качестве клеток-хозяев в настоящем изобретении, включают грамотрицательные или грамположительные организмы, такие как E. coli, B. subtilis, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, Serratia и Shigella, а также Bacilli, Pseudomonas и Streptomyces. Одним из предпочтительных хозяев для клонирования в E. coli является E. coli 294 (ATCC 31446), хотя подходят и другие штаммы, такие как E. coli B, E. coli X1776 (ATCC 31537) и E. coli W3110 (ATCC 27325). Эти примеры являются иллюстративными, а не ограничивающими.
Экспрессирующие векторы для использования в прокариотических клетках-хозяевах, как правило, содержат один или несколько маркерных генов для селекции по фенотипу. Маркерный ген для селекции по фенотипу, например, представляет собой ген, кодирующий белок, который придает устойчивость к антибиотикам или который удовлетворяет потребность в питательных веществах. Примеры пригодных экспрессирующих векторов для прокариотических клеток-хозяев включают векторы, происходящие из коммерчески доступных плазмид, таких как pKK223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden), pGEM1 (Promega Biotec, Madison, Wisconsin., USA) и серии векторов pET (Novagen, Madison, Wisconsin, USA) и pRSET (Invitrogen, Carlsbad, CA) (Studier, J Mol Biol 219:37 (1991); Schoepfer, Gene 124:83 (1993)). Промоторные последовательности, обычно используемые в рекомбинантных экспрессирующих векторах прокариотических клеток-хозяев, включают T7, (Rosenberg, et al., Gene 56:125 (1987)), промоторную систему β-лактамазы (пенициллиназы), лактозы (Chang, et al., Nature 275:615 (1978); Goeddel, et al., Nature 281:544 (1979)), промоторную систему триптофана (trp) (Goeddel, et al., Nucl Acids Res 8:4057 (1980)) и tac-промотор (Sambrook, et al., Molecular Cloning, A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory (1990)).
Дрожжи или мицелиальные грибы, пригодные в настоящем изобретении, включают организмы из рода Saccharomyces, Pichia, Actinomycetes, Kluyveromyces, Schizosaccharomyces, Candida, Trichoderma, Neurospora и мицелиальные грибы, такие как Neurospora, Penicillium, Tolypocladium и Aspergillus. Дрожжевые векторы часто содержат последовательность ориджина репликации из дрожжевой плазмиды 2µ, автономно реплицирующуюся последовательность (ARS), промоторную область, последовательности полиаденилирования, последовательности терминации транскрипции и маркерный ген для селекции. Подходящие промоторные последовательности дрожжевых векторов включают, среди прочих, промоторы металлотионеина, 3-фосфоглицераткиназы (Hitzeman, et al., J Biol Chem 255:2073 (1980)) или других гликолитических ферментов (Holland, et al., Biochem 17:4900 (1978)), таких как енолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа. Другие подходящие векторы и промоторы для использования в дрожжевой экспрессии дополнительно описаны в Fleer, et al., Gene 107:285 (1991). Другие подходящие промоторы и векторы дрожжей и способы трансформации дрожжей хорошо известны в данной области. Способы трансформации дрожжей хорошо известны. Один из таких способов описан в Hinnen, et al., Proc Natl Acad Sci 75:1929 (1978). В способе Hinnen отбирают трансформанты Trp+ в селективной среде.
Системы для культивирования клеток-хозяев из млекопитающих или насекомых можно также использовать для экспрессии рекомбинантных антител. В принципе, пригодна любая культура клеток высших эукариот, из культур клеток или позвоночных или беспозвоночных. Примеры клеток беспозвоночных включают клетки растений и насекомых (Luckow, et al., Bio/Technology 6:47 (1988); Miller, et al., Genetics Engineering, Setlow, et al., eds. Vol. 8, pp. 277-9, Plenam Publishing (1986); Mseda, et al., Nature 315:592 (1985)). Например, системы на основе Baculovirus можно использовать для продукции гетерологичных белков. В системе на основе насекомых вирус ядерного полигидроза Autographa californica (AcNPV) можно использовать в качестве вектора для экспрессии чужеродных генов. Вирус выращивают в клетках Spodoptera frugiperda. Последовательность, кодирующую антитело, можно индивидуально клонировать в нежизненноважные области (например, ген полигедрина) вируса и помещать под контроль промотора AcNPV (например, промотор полигедрина). Другие идентифицированные хозяева включают Aedes, Drosophila melanogaster и Bombyx mori. Множество вирусных штаммов для трансфекции являются общедоступными, например, вариант L-1 AcNPV и штамм Bm-5 NPV Bombyx mori, и такие вирусы можно использовать в качестве вируса в данной работе согласно настоящему изобретению, в частности, для трансфекции клеток Spodoptera frugiperda. Более того, культуры растительных клеток хлопка, кукурузы, картофеля, сои, петунии, томатов и табака также можно использовать в качестве хозяев.
Клетки позвоночных и размножение клеток позвоночных в культуре (культуре тканей) стало рутинной процедурой. См. Tissue Culture, Kruse, et al., eds., Academic Press (1973). Примеры пригодных линий клеток-хозяев млекопитающих представляют собой клетки почки мартышки; линии эмбриональной почки человека; почки новорожденного хомяка; клетки яичника китайского хомяка/-DHFR (CHO, Urlaub, et al., Proc Natl Acad Sci USA 77:4216 (1980)); клетки Сертоли мыши; клетки карциномы шейки матки человека (HELA); клетки почки собаки; клетки легкого человека; клетки печени человека; опухоль молочной железы мыши; и клетки NSO.
Клетки-хозяева трансформируют вышеописанными векторами для продукции антител и культивируют в общепринятых питательных средах, модифицированных соответствующим образом для индуцирования промоторов, последовательностей для управления транскрипцией и трансляцией, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности. Обычно используемые промоторные последовательности и энхансерные последовательности происходят из вируса полиомы, аденовируса 2, вируса обезьяны 40 (SV40) и цитомегаловируса человека (CMV). Последовательности ДНК, происходящие из вирусного генома SV40, можно использовать для получения других генетических элементов для экспрессии последовательности структурных генов в клетке-хозяине млекопитающих, например, ориджин, ранний и поздний промотор, энхансер, участки сплайсинга и полиаденилирования SV40. Вирусные ранние и поздние промоторы, в частности, пригодны вследствие того, что и те и другие легко получают из вирусного генома в виде фрагмента, который может также содержать вирусный ориджин репликации. Иллюстративные экспрессирующие векторы для использования в клетках-хозяевах млекопитающих являются коммерчески доступными.
Клетки-хозяева, используемые для продукции антитела по данному изобретению, можно культивировать во множестве сред. Коммерчески доступные среды, такие как среда Хэма F10 (Sigma, St Louis, MO), минимальная питательная среда (MEM, Sigma, St Louis, MO), RPMI-1640 (Sigma, St Louis, MO) и среда Игла в модификации Дульбекко (DMEM, Sigma, St Louis, MO) подходят для культивирования клеток-хозяев. Кроме того, в качестве культуральных сред для клеток-хозяев можно использовать любую из сред, описанных в Ham, et al., Meth Enzymol 58:44 (1979), Barnes, et al., Anal Biochem 102:255 (1980) и патентах США No. 4767704, 4657866, 4560655, 5122469, 5712163 или 6048728. В любую из этих сред можно добавить в случае необходимости гормоны и/или другие факторы роста (такие как инсулин, трансферрин или фактор роста эпидермиса), соли (такие как X-хлориды, где X представляет собой натрий, кальций, магний; и фосфаты), буферы (такие как HEPES), нуклеотиды (такие как аденозин и тимидин), антибиотики (такие как лекарственное средство ГЕНТАМИЦИН™), микроэлементы (определяемые как неорганические соединения, обычно присутствующие в конечных концентрациях порядка микромолей), и глюкозу или эквивалентный источник энергии. Любые другие необходимые добавки можно также вносить в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия, использованные ранее для клетки-хозяина, выбранной для экспрессии, и будут очевидны для специалиста.
ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ АНТИТЕЛА
Изобретение дополнительно относится к полинуклеотидам или нуклеиновым кислотам, например, ДНК, содержащим нуклеотидную последовательность, кодирующую антитело по изобретению и его фрагменты. Иллюстративные полинуклеотиды включают полинуклеотиды, кодирующие цепи антител, содержащие одну или несколько описанных в настоящем документе аминокислотных последовательностей. Изобретение также относится к полинуклеотидам, которые гибридизуются в строгих или менее строгих условиях гибридизации с полинуклеотидами, которые кодируют антитело по настоящему изобретению.
Полинуклеотиды можно получать и нуклеотидную последовательность полинуклеотидов определять любым способом, известным в данной области. Например, если известна нуклеотидная последовательность антитела, то полинуклеотид, кодирующий антитело, можно сконструировать из химически синтезированных олигонуклеотидов (например, как описано в Kutmeier, et al., Bio/Techniques 17:242 (1994)), что, в кратком изложении, предусматривает синтез перекрывающихся олигонуклеотидов, содержащих части последовательности, кодирующей антитело, отжиг и лигирование этих олигонуклеотидов и затем амплификацию лигированных олигонуклеотидов с помощью PCR.
Альтернативно, полинуклеотид, кодирующий антитело, можно получать из нуклеиновой кислоты из подходящего источника. Если клон, содержащий нуклеиновую кислоту, кодирующую конкретное антитело, не доступен, но последовательность молекулы антитела известна, то нуклеиновую кислоту, кодирующую иммуноглобулин, можно химически синтезировать или получить из подходящего источника (например, библиотеки кДНК антител или библиотеки кДНК, полученной из, или нуклеиновой кислоты, предпочтительно, поли-A+-РНК, выделенной из любой ткани или клеток, экспрессирующих антитело, таких как гибридомные клетки, отобранные по экспрессии антител по изобретению) амплификацией с помощью PCR с использованием синтетических праймеров, гибридизуемых с 3'- и 5'-концами последовательности, или клонированием с использованием олигонуклеотидного зонда, специфичного в отношении последовательности конкретного гена для идентификации, например, клона кДНК из библиотеки кДНК, который кодирует антитело. Амплифицированные нуклеиновые кислоты, полученные с помощью PCR, можно затем клонировать в реплицирующиеся клонирующие векторы с использованием любого способа, хорошо известного в данной области.
После определения нуклеотидной последовательности и соответствующей аминокислотной последовательности антитела нуклеотидной последовательностью антитела можно манипулировать с использованием способов, хорошо известных в данной области манипулирования нуклеотидными последовательностями, например, способов рекомбинантных ДНК, сайт-специфического мутагенеза, PCR и т.д. (см., например, способы, описанные в Sambrook, et al., Molecular Cloning, A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory (1990); Ausubel, et al., eds., Current Protocols in Molecular Biology, John Wiley & Sons (1998), оба из которых приведены в настоящем документе в качестве ссылки в полном объеме), для получения антител, обладающих другой аминокислотной последовательностью, например, для получения аминокислотных замещений, делеций и/или вставок.
В конкретном варианте осуществления аминокислотную последовательность вариабельных доменов тяжелой и/или легкой цепи можно исследовать для идентификации последовательностей CDR хорошо известными способами, например, сравнением с известными аминокислотными последовательностями других вариабельных областей тяжелой и легкой цепей для определения областей гипервариабельности последовательности. С использованием общепринятых способов рекомбинантных ДНК, одну или несколько CDR можно вставить в каркасные области, например, в каркасные области человека для гуманизации не относящегося к человеку антитела, как описано выше. Каркасные области могут быть природного происхождения или представляют собой консенсусные каркасные области и, предпочтительно, каркасные области человека (см., например, Chothia, et al., J Mol Biol 278:457 (1998) в отношении списка каркасных областей человека). Предпочтительно, полинуклеотид, полученный сочетанием каркасных областей и CDR, кодирует антитело, которое специфически связывается с полипептидом по изобретению. Предпочтительно, как рассмотрено выше, одно или несколько аминокислотных замещений можно осуществить внутри каркасных областей, и, предпочтительно, аминокислотные замещения улучшают связывание антитела со своим антигеном. Кроме того, такие способы можно использовать для осуществления аминокислотных замещений или делеций одного или нескольких остатков цистеина вариабельной области, принимающих участие во внутрицепочечной дисульфидной связи, для получения молекул антител, у которых отсутствуют одна или несколько внутрицепочечных дисульфидных связей. Другие изменения полинуклеотида относятся к настоящему изобретению и находятся в пределах компетенции в данной области.
Кроме того, можно использовать способы, разработанные для получения «химерных антител» (Morrison, et al., Proc Natl Acad Sci 81:851 (1984); Neuberger, et al., Nature 312:604 (1984); Takeda, et al., Nature 314:452 (1985)) путем сплайсирования генов молекулы антитела мыши подходящей антигенной специфичности вместе с генами молекулы антитела человека подходящей биологической активности. Как описано выше, химерное антитело представляет собой молекулу, в которой различные части происходят из различных видов животных, такие как части, содержащие вариабельную область, происходящую из мышиного mAb, и константную область иммуноглобулина человека, например, гуманизированные антитела.
Альтернативно, способы, описанные для получения одноцепочечных антител (патент США No. 4946778; Bird, Science 242:423 (1988); Huston, et al., Proc Natl Acad Sci USA 85:5879 (1988); и Ward, et al., Nature 334:544 (1989)), можно адаптировать для получения одноцепочечных антител. Одноцепочечные антитела образованы связыванием фрагментов тяжелой и легкой цепи Fv-области посредством аминокислотного мостика, что приводит к одноцепочечному полипептиду. Можно также использовать способы сборки функциональных Fv-фрагментов в E. coli (Skerra, et al., Science 242:1038 (1988)).
СПОСОБЫ ПОЛУЧЕНИЯ АНТИ-NOTCH3 АНТИТЕЛ
Антитела по изобретению можно получать любым способом, известным в данной области синтеза антител, в частности, химическим синтезом или, предпочтительно, способами рекомбинантной экспрессии.
Рекомбинантная экспрессия антитела по изобретению или его фрагмента, производного или аналога (например, тяжелой или легкой цепи антитела по изобретению или одноцепочечного антитела по изобретению) требует конструкции экспрессирующего вектора, содержащего полинуклеотид, который кодирует антитело или фрагмент антитела. После получения полинуклеотида кодирующего молекулу антитела, вектор для продукции антитела можно получить технологией рекомбинантных ДНК. Экспрессирующий вектор конструируют содержащим кодирующие последовательности антител и подходящие сигналы для управления транскрипцией и трансляцией. Эти способы включают, например, способы рекомбинантных ДНК in vitro, синтетические способы и генетическую рекомбинацию in vivo.
Экспрессирующий вектор переносят в клетку-хозяина общепринятыми способами, и трансфицированные клетки затем культивируют общепринятыми способами для получения антитела по изобретению. В одном из аспектов изобретения векторы, кодирующие как тяжелую, так и легкую цепи, можно совместно экспрессировать в клетке-хозяине для экспрессии полноразмерной молекулы иммуноглобулина, как подробно рассмотрено ниже.
Множество систем хозяин-экспрессирующий вектор можно использовать для экспрессии молекул антител по изобретению, как описано выше. Такие системы для экспрессии в хозяине представляют собой носители, с помощью которых интересующие кодирующие последовательности можно продуцировать и затем очищать, но также представляют собой клетки, которые при трансформации или трансфекции подходящими нуклеотидными кодирующими последовательностями могут экспрессировать молекулу антитела по изобретению in situ. Бактериальные клетки, такие как E. coli, и эукариотические клетки обычно используют для экспрессии рекомбинантной молекулы антитела, особенно для экспрессии полноразмерной рекомбинантной молекулы антитела. Например, клетки млекопитающих, такие как CHO, в сочетании с вектором, таким как промоторный элемент главного промежуточного раннего гена из цитомегаловируса человека, представляют собой эффективную систему для экспрессии антител (Foecking, et al., Gene 45:101 (1986); Cockett, et al., Bio/Technology 8:2 (1990)).
Кроме того, можно выбрать линию клетки-хозяина, которая модулирует экспрессию вставленных последовательностей или модифицирует и процессирует продукт гена желаемым специфическим образом. Такие модификации (например, гликозилирование) и процессирование (например, расщепление) белковых продуктов могут являться важными для функции белка. Различные клетки-хозяева обладают характерными и специфическими механизмами посттрансляционного процессинга и модификации белков и продуктов генов. Подходящие линии клеток или системы-хозяева можно выбрать для обеспечения правильной модификации и процессинга экспрессируемого чужеродного белка. Для этой цели можно использовать эукариотические клетки-хозяева, которые обладают клеточными механизмами правильного процессинга первичного транскрипта, гликозилирования и фосфорилирования продукта гена. Такие клетки-хозяева млекопитающих включают в качестве неограничивающих примеров клетки CHO, COS, 293, 3T3 или миеломы.
Для долговременной продукции с высоким выходом рекомбинантных белков предпочтительна стабильная экспрессия. Например, можно сконструировать линии клеток, которые стабильно экспрессируют молекулу антитела. Вместо использования экспрессирующих векторов, которые содержат вирусные ориджины репликации, клетки-хозяева можно трансформировать ДНК под управлением подходящих элементов для управления экспрессией (например, промоторных, энхансерных последовательностей, терминаторов транскрипции, участков полиаденилирования и т.д.) и селективного маркера. После введения чужеродной ДНК сконструированным клеткам можно дать возможность расти в течение одних или двух суток в обогащенных средах и затем перенести в селективные среды. Селектируемый маркер на рекомбинантной плазмиде сообщает устойчивость в отношении селекции и дает возможность клеткам стабильно интегрировать плазмиду в хромосомы и вырастать с образованием фокусов, которые, в свою очередь, можно клонировать и развивать в линии клеток. Этот способ можно успешно использовать для конструирования линий клеток, которые экспрессируют молекулу антитела. Такие сконструированные линии клеток, в частности, могут быть пригодны для скрининга и оценки соединений, которые взаимодействуют непосредственно или опосредованно с молекулой антитела.
Можно использовать ряд систем селекции, в том числе в качестве неограничивающих примеров можно использовать гены тимидинкиназы вируса простого герпеса (Wigler et al., Cell 11:223 (1977)), гипоксантин-гуанинфосфорибозилтрансферазы (Szybalska et al., Proc Natl Acad Sci USA 48:202 (1992)) и аденинфосфорибозилтрансферазы (Lowy et al., Cell 22:817 (1980)) в клетках tk, hgprt или aprt соответственно. Также, устойчивость к антиметаболитам можно использовать в качестве основы для селекции на следующие гены: dhfr, который придает устойчивость к метотрексату (Wigler, et al., Proc Natl Acad Sci USA 77:357 (1980); O'Hare, et al., Proc Natl Acad Sci USA 78:1527 (1981)); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan et al., Proc Natl Acad Sci USA 78:2072 (1981)); neo, который придает устойчивость к аминогликозиду G-418 (Wu et al., Biotherapy 3:87 (1991)); и hygro, который придает устойчивость к гигромицину (Santerre et al., Gene 30:147 (1984)). Способы, общеизвестные в данной области технологии рекомбинантных ДНК, можно обычным образом использовать для селекции требуемого рекомбинантного клона, и такие способы описаны, например, в Ausubel, et al., eds., Current Protocols in Molecular Biology, John Wiley & Sons (1993); Kriegler, Gene Transfer and Expression, A Laboratory Manual, Stockton Press (1990); и в Chapters 12 and 13, Dracopoli, et al., eds, Current Protocols in Human Genetics, John Wiley & Sons (1994); Colberre-Garapin, et al., J Mol Biol 150:1 (1981), которые приведены в настоящем документе в качестве ссылки в полном объеме.
Уровни экспрессии молекулы антитела можно повышать путем амплификации вектора (для обзора см. Bebbington, et al., «The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells», DNA Cloning, Vol.3. Academic Press (1987)). Если маркер в векторной системе, экспрессирующей антитело, способен к амплификации, то повышение уровня ингибитора, присутствующего в культуре клетки-хозяина, будет повышать количество копий маркерного гена. Так как амплифицируемая область ассоциирована с геном антитела, продукция антитела также возрастет (Crouse ef al., Mol Cell Biol 3:257 (1983)).
Клетку-хозяина можно совместно трансфицировать двумя экспрессирующими векторами по изобретению, где первый вектор кодирует полипептид, происходящий из тяжелой цепи, и второй вектор кодирует полипептид, происходящий из легкой цепи. Два вектора могут содержать идентичные селективные маркеры, которые обеспечивают одинаковую экспрессию полипептидов тяжелой и легкой цепи. Альтернативно, можно использовать одиночный вектор, который кодирует полипептиды как тяжелой, так и легкой цепи и способен их экспрессировать. В таких ситуациях легкую цепь следует помещать перед тяжелой цепью, чтобы избежать избытка токсичной свободной тяжелой цепи (Proudfoot, Nature 322:52 (1986); Kohler, Proc Natl Acad Sci USA 77:2197 (1980)). Кодирующие последовательности тяжелой и легкой цепей могут содержать кДНК или геномную ДНК.
После того как молекула антитела по изобретению продуцирована в животном, химически синтезирована или рекомбинантным образом экспрессирована, ее можно очищать любым способом, известным в данной области очистки молекулы иммуноглобулина, например, хроматографией (например, ионообменной, аффинной, в частности, по аффинности в отношении специфического антигена за белок A, и эксклюзионной хроматографией), центрифугированием, по разнице в растворимости, или любым другим общепринятым способом очистки белков. Кроме того, для облегчения очистки можно производить слияние антител по настоящему изобретению или их фрагментов с гетерологичными полипептидными последовательностями, описанными в настоящем документе или иным способом известным в данной области.
Настоящее изобретение относится к антителам, рекомбинантно слитым или химически конъюгированным (включая как ковалентную, так и нековалентную конъюгации) с полипептидом. Слитые или конъюгированные антитела по настоящему изобретению можно применять для облегчения очистки. См., например, публикацию PCT WO 93/21232; EP 439095; Naramura, et al., Immunol Lett 39:91 (1994); патент США No. 5474981; Gillies, et al., Proc Natl Acad Sci USA 89:1428 (1992); Fell, et al., J Immunol 146:2446 (1991), которые приведены в качестве ссылки в полном объеме.
Более того, для облегчения очистки можно производить слияние антител или их фрагментов по настоящему изобретению с маркерными последовательностями, такими как пептид. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой гексагистидиновый пептид, такой как метка, представленная в векторе pQE (QIAGEN, Inc., Valencia, CA), среди прочих, многие из которых коммерчески доступны. Как описано, например, в Gentz, et al., Proc Natl Acad Sci USA 86:821 (1989), гексагистидин обеспечивает удобную очистку слитого белка. Другие пептидные метки, пригодные для очистки, включают в качестве неограничивающих примеров метку «HA», которая соответствует эпитопу, происходящему из белка гемагглютинина гриппа (Wilson, et al., Cell 37:767 (1984)), и метку «flag».
ОЧИСТКА АНТИТЕЛ
При использовании рекомбинантных способов антитело можно продуцировать внутриклеточно, в периплазматическое пространство или непосредственно секретировать в среду. Если антитело продуцируют внутриклеточно, то в качестве первой стадии дебрис в виде частиц, или клетки-хозяева, или лизированные фрагменты можно удалить, например, центрифугированием или ультрафильтрацией. В Carter, et al., Bio/Technology 10:163 (1992) описан способ выделения антител, которые секретируются в периплазматическое пространство E. coli. В кратком изложении, клеточную массу оттаивают в присутствии ацетата натрия (pH 3,5), EDTA и фенилметилсульфонилфторида (PMSF) в течение приблизительно 30 минут. Клеточный дебрис можно удалить центрифугированием. При секреции антитела в среду супернатанты из таких экспрессионных систем, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрации белка, например, комплекта для ультрафильтрации Amicon или Millipore Pellicon. Можно вносить ингибитор протеаз, такой как PMSF, на любой из вышеописанных стадий для ингибирования протеолиза, и можно вносить антибиотики для предотвращения роста случайных контаминантов.
Композицию антител, полученную из клеток, можно очищать с использованием, например, хроматографии с гидроксилапатитом, гель-электрофореза, диализа и аффинной хроматографии, где аффинная хроматография является предпочтительным способом очистки. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, который находится в антителе. Белок A можно использовать для очистки антител, в основе которых лежат тяжелые цепи IgG1, IgG2 или IgG4 человека (Lindmark et al., J. Immunol. Methods 62:1-13 (1983)). Белок G рекомендован для всех изотипов мыши и для IgG3 человека (Guss et al., EMBO J. 5:1567-1575 (1986)). Матрица, к которой присоединен аффинный лиганд, является в наиболее частых случаях агарозой, однако доступны и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или полистиролдивинилбензол, обеспечивают более высокие скорости потока и более короткую продолжительность процесса, чем те, которые можно достичь с помощью агарозы. В случае, когда антитело содержит домен CH3, для очистки пригодна смола Bakerbond ABX™ (J. T. Baker, Phillipsburg, NJ). Также в зависимости от антитела, подлежащего выделению, доступны другие способы очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, HPLC с обращенной фазой, хроматография на кремнеземе, хроматография на гепарин-сефарозе™, хроматография на анионной или катионной обменной смоле (такой как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония.
После любой предварительной стадии(й) очистки смесь, содержащую интересующее антитело и примеси, можно подвергать хроматографии гидрофобного взаимодействия с низким значением pH с использованием буфера для элюции со значением pH приблизительно между 2,5-4,5, которую предпочтительно проводят при низких концентрациях соли (например, приблизительно 0-0,25 М соль).
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
Терапевтические композиции полипептида или антитела можно получать для хранения в виде лиофилизированных композиций или водных растворов смешиванием полипептида, обладающего требуемой степенью чистоты с оптимальными «фармацевтически приемлемыми» носителями, вспомогательными веществами или стабилизаторами, обычно используемыми в данной области (которые все обозначены как «вспомогательные вещества»), т.е. буферными веществами, стабилизаторами, консервантами, средствами для обеспечения изотоничности, неионными детергентами, антиоксидантами и другими разнообразными добавками. См. Remington's Pharmaceutical Sciences, 16th edition, Osol, Ed. (1980). Такие добавки должны быть нетоксичными для реципиентов в используемых дозировках и концентрациях.
Буферные вещества помогают поддерживать pH в диапазоне, который приближается к физиологическим условиям. Они предпочтительно присутствуют в концентрации, варьирующей от приблизительно 2 мМ до приблизительно 50 мМ. Подходящие буферные средства для использования с настоящим изобретением включают как органические, так и неорганические кислоты и их соли, такие как цитратные буферы (например, смесь цитрата мононатрия-цитрата динатрия, смесь лимонной кислоты-цитрата тринатрия, смесь лимонной кислоты-цитрата мононатрия и т.д.), сукцинатные буферы (например, смесь янтарной кислоты-сукцината мононатрия, смесь янтарной кислоты-гидроксида натрия, смесь янтарной кислоты-сукцината динатрия и т.д.), тартратные буферы (например, смесь виннокаменной кислоты-тартрата натрия, смесь виннокаменной кислоты-тартрата калия, смесь виннокаменной кислоты-гидроксида натрия и т.д.), фумаратные буферы (например, смесь фумаровой кислоты-фумарата мононатрия, смесь фумаровой кислоты-фумарата динатрия, смесь фумарата мононатрия-фумарата динатрия и т.д.), глюконатные буферы (например, смесь глюконовой кислоты-глюконата натрия, смесь глюконовой кислоты-гидроксида натрия, смесь глюконовой кислоты-глюконата калия и т.д.), оксалатные буферы (например, смесь щавелевой кислоты-оксалата натрия, смесь щавелевой кислоты-гидроксида натрия, смесь щавелевой кислоты-оксалата калия и т.д.), лактатные буферы (например, смесь молочной кислоты-лактата натрия, смесь молочной кислоты-гидроксида натрия, смесь молочной кислоты-лактата калия и т.д.) и ацетатные буферы (например, смесь уксусной кислоты-ацетата натрия, смесь уксусной кислоты-гидроксида натрия и т.д.). Кроме того, можно упомянуть фосфатные буферы, гистидиновые буферы и триметиламиновые соли, такие как трис.
Для задержки микробного роста можно добавлять консерванты, и их можно добавлять в количествах, варьирующих в пределах 0,2%-1% (мас./об.). Подходящие консерванты для использования с настоящим изобретением включают фенол, бензиловый спирт, мета-крезол, метилпарабен, пропилпарабен, октадецилдиметилбензиламмоний-хлорид, галогениды бензалкония (например, хлорид, бромид, иодид), гексаметоний-хлорид и алкилпарабены, такие как метил- или пропилпарабен, катехин, резорцин, циклогексанол и 3-пентанол.
Средства для обеспечения изотоничности, иногда известные как «стабилизаторы», можно добавлять для обеспечения изотоничности жидких композиций по настоящему изобретению, и они включают многоатомные сахароспирты, предпочтительно трехатомные или высшие сахароспирты, такие как глицерин, эритрит, арабит, ксилит, сорбит и маннит.
Стабилизаторы относятся к обширной категории вспомогательных веществ, которые могут варьировать по функции от наполнителя до добавки, которые солюбилизируют терапевтическое средство или помогают предотвратить денатурацию или сцепление со стенкой контейнера. Типичные стабилизаторы могут представлять собой многоатомные сахароспирты (перечисленные выше); аминокислоты, такие как аргинин, лизин, глицин, глутамин, аспарагин, гистидин, аланин, орнитин, L-лейцин, 2-фенилаланин, глутаминовая кислота, треонин и т.д.; органические сахара или сахароспирты, такие как лактоза, трегалоза, стахиоза, маннит, сорбит, ксилит, рибит, миоинозит, галактит, глицерин и т.п., включая циклические спирты, такие как инозит; полиэтиленгликоль; аминокислотные полимеры; серусодержащие восстанавливающие вещества, такие как мочевина, глутатион, тиоктовая кислота, тиогликолят натрия, тиоглицерин, альфа-монотиоглицерин и тиосульфат натрия; низкомолекулярные полипептиды (т.е. <10 остатков); белки, такие как человеческий сывороточный альбумин, бычий сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; моносахариды, такие как ксилоза, манноза, фруктоза, глюкоза; дисахариды, такие как лактоза, мальтоза, сахароза, и трисахариды, такие как раффиноза; и полисахариды, такие как декстран. Стабилизаторы могут присутствовать в диапазоне от 0,1 до 10000 мас. на часть массы активного белка.
Неионные поверхностно-активные вещества или детергенты (также известные как «смачивающие средства») можно добавлять для облегчения солюбилизации терапевтического средства, а также для защиты терапевтического белка от вызываемой перемешиванием агрегации, которые также позволяют композиции подвергаться натяжению на поверхности раздела без вызывания денатурации белка. Подходящие неионные поверхностно-активные вещества включают полисорбаты (20, 80 и т.д.), полиоксамеры (184, 188 и т.д.), полиолы плюроники™, моноэфиры полиоксиэтиленсорбитана (TWEEN-20®, TWEEN-80® и т.д.). Неионные поверхностно-активные вещества могут присутствовать в диапазоне приблизительно от 0,05 мг/мл до приблизительно 1,0 мг/мл, предпочтительно приблизительно от 0,07 мг/мл до приблизительно 0,2 мг/мл.
Дополнительные разнообразные вспомогательные вещества включают наполнители, (например, крахмал), хелатирующие средства (например, EDTA), антиоксиданты (например, аскорбиновую кислоту, метионин, витамин E) и сорастворители. Композиция также может содержать в настоящем документе более одного активного соединения, если это необходимо для конкретного подлежащего лечению симптома, предпочтительно соединения с комплементарными активностями, которые не оказывают отрицательное действие друг на друга. Например, может оказаться желательным дополнительное предоставление иммуносупрессорного средства. Такие молекулы соответствующим образом присутствуют в сочетании в количествах, которые эффективны для намеченной цели. Активные ингредиенты можно также заключать в микрокапсулу, полученную, например, способами коацервации или посредством межфазной полимеризации, например, микрокапсулу на основе гидроксиметилцеллюлозы или желатина и микрокапсулу на основе полиметилметакрилата, соответственно, в коллоидных системах для доставки лекарственного средства (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие способы описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Композиции для использования при введении in vivo должны быть стерильными. Этого легко достигают, например, посредством фильтрации через стерильные фильтрационные мембраны. Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где матрицы находятся в форме обладающих определенной формой изделий, например, пленок или микрокапсул. Примеры матриц для замедленного высвобождения включают сложные полиэфиры, гидрогели (например, поли-2-гидроксиэтилметакрилат или поливиниловый спирт), полилактиды (патент США No. 3773919), сополимеры L-глутаминовой кислоты и этил-L-глутамата, недеградируемый этиленвинилацетат, деградируемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту. В то время как полимеры, такие как на основе этиленвинилацетата и молочной кислоты-гликолевой кислоты, способны высвобождать молекулы в течение 100 суток, определенные гидрогели высвобождают белки в течение более коротких периодов времени. Если инкапсулированные антитела остаются в организме в течение длительного времени, то они могут денатурировать или агрегировать под действием влажности при 37°C, что приводит к утрате биологической активности и возможным изменениям иммуногенности. Можно разработать рациональные стратегии для стабилизации в зависимости от вовлеченного механизма. Например, если обнаружено, что механизм агрегации состоит в образовании межмолекулярных S-S связей посредством превращения тиола в дисульфид, то стабилизации можно достичь модификацией сульфгидрильных остатков, лиофилизацией из кислотных растворов, контролированием содержания воды с использованием подходящих добавок и разрабатывая специфические композиции на основе полимерных матриц.
Количество терапевтического полипептида, антитела или его фрагмента, которое будет эффективным при лечении конкретного нарушения или состояния, будет зависеть от природы нарушения или состояния, и его можно определять общепринятыми клиническими способами. Где возможно, желательно определять кривую доза-ответ и фармацевтические композиции по изобретению сначала in vitro и затем на пригодных системах животных моделей перед тестированием на человеке.
В предпочтительном варианте осуществления водный раствор терапевтического полипептида, антитела или его фрагмента вводят подкожной инъекцией. Каждая доза может варьировать от приблизительно 0,5 мкг до приблизительно 50 мкг на килограмм массы тела или, более предпочтительно, от приблизительно 3 мкг до приблизительно 30 мкг на килограмм массы тела.
Режим дозирования для подкожного введения может варьировать от одного раза в месяц, до ежедневного в зависимости от ряда клинических факторов, включая тип заболевания, тяжесть заболевания и чувствительность индивидуума к терапевтическому средству.
ТЕРАПЕВТИЧЕСКИЕ ИСПОЛЬЗОВАНИЯ АНТИ-NOTCH3 АНТИТЕЛ
Предполагается, что антитела по настоящему изобретению можно применять для лечения млекопитающего. В одном из вариантов осуществления антитело вводят не относящемуся к человеку млекопитающему, например, в целях получения доклинических данных. В качестве иллюстрации, не относящиеся к человеку млекопитающие, подлежащие лечению, включают не относящихся к человеку приматов, собак, кошек, грызунов и других млекопитающих, на которых осуществляют доклинические исследования. Такие млекопитающие могут являться общепринятыми животными моделями для заболевания, подлежащего лечению антителом, или их можно использовать для исследования токсичности интересующего антитела. В каждом из этих вариантов осуществления исследования повышения дозировки можно осуществлять на млекопитающем.
Антитело, с конъюгированным с ним терапевтическим компонентом или без него, вводимое по отдельности или в сочетании с цитотоксическим фактором(ами), можно применять в качестве терапевтического средства. Настоящее изобретение направлено на способы лечения на основе антител, которые включают введение антител по изобретению животному, млекопитающему или человеку для лечения опосредованного Notch3 заболевания, нарушения или состояния. Животное или индивидуум может представлять собой млекопитающее, нуждающееся в конкретном лечении, такое как млекопитающее, у которого диагностировали конкретное нарушение, например, связанное с Notch3. Антитела, направленные против Notch3, пригодны против рака и других связанных с Notch3 заболеваний, включая неврологические нарушения, диабет, ревматоидный артрит, связанные с сосудами заболевания и синдром Алажиля у млекопитающих, включая в качестве неограничивающих примеров коров, свиней, лошадей, кур, кошек, собак, не относящихся к человеку приматов и т.д., а также человека. Например, посредством введения терапевтически допустимой дозы антитела против Notch3, или антител по настоящему изобретению, или коктейля из настоящих антител, или в сочетании с другими антителами из различных источников, симптомы заболевания можно облегчать или предотвращать при лечении млекопитающего, в частности, человека.
Терапевтические соединения по изобретению включают в качестве неограничивающих примеров антитела по изобретению (включая их фрагменты, аналоги и производные, как описано в настоящем документе) и нуклеиновые кислоты, кодирующие антитела по изобретению, как описано ниже (включая их фрагменты, аналоги и производные, и антиидиотипические антитела, как описано в настоящем документе). Антитела по изобретению можно применять для лечения, ингибирования или профилактики заболеваний, нарушений или состояний, связанных с патологической экспрессией и/или активностью Notch3, включая в качестве неограничивающих примеров любое одно или несколько заболеваний, нарушений или состояний, описанных в настоящем документе. Лечение и/или профилактика заболеваний, нарушений или состояний, связанных с патологической экспрессией и/или активностью Notch3, включает в качестве неограничивающих примеров облегчение, по меньшей мере, одного из симптомов, связанных с этими заболеваниями, нарушениями или состояниями. Антитела по изобретению можно предоставлять в фармацевтически приемлемых композициях, как известно в данной области или как описано в настоящем документе.
Анти-Notch3 антитела по настоящему изобретению можно применять терапевтически при многих заболеваниях. Настоящее изобретение относится к способу профилактики или лечения опосредованных Notch3 заболеваний у млекопитающего. Способ включает введение млекопитающему количества антитела против Notch3, которое предотвращает заболевание или излечивает его. Антитело против Notch3 связывается с Notch3 и препятствует его функции. Передача сигнала Notch3 связана с различными заболеваниями, такими как различные виды рака (Haruki, et al., Cancer Res 65:3555 (2005); Park, et al., Cancer Res 66:6312 (2006); Lu, et al., Clin Cancer Res 10:3291 (2004)); Hedvat, et al., Br J Haematol 122:728 (2003); Buchler, et al., Ann Surg 242:791 (2005)); Bellavia, et al., Proc Natl Acad Sci USA 99:3788 (2002); Screpanti, et al., Trends Mol Med 9:30 (2003)); van Limpt, et al., Cancer Lett 228:59 (2005)), неврологические нарушения (Joutel, et al., Nature 383:707 (1996)), диабет (Anastasi, et al., J Immunol 171:4504 (2003), ревматоидный артрит (Yabe, et al., J Orthop Sci 10:589 (2005)), связанные с сосудами заболевания (Sweeney, et al., FASEB J 18:1421 (2004)), и синдром Алажиля (Flynn, et al., J Pathol 204:55 (2004)). Антитела против Notch3 также будут эффективны для профилактики вышеупомянутых заболеваний.
Количество антитела, которое будет эффективным для лечения, ингибирования и профилактики заболевания или нарушения, связанного с патологической экспрессией и/или активностью Notch3, можно определить общепринятыми клиническими способами. Дозировка будет зависеть от типа подлежащего лечению заболевания, тяжести и течения заболевания, вводят ли антитело с профилактическими или терапевтическими целями, предыдущего лечения, клинической истории пациента и ответа на антитело и выбора лечащего врача. Антитело можно вводить в режимах лечения, соответствующих заболеванию, например, однократной дозой или несколькими дозами в течение от одних до нескольких суток для облегчения состояния заболевания, или периодическими дозами в течение протяженного периода времени для ингибирования прогрессирования заболевания и профилактики рецидивирования заболевания. Кроме того, не обязательно, можно использовать анализы in vitro, чтобы способствовать определению оптимальных диапазонов дозировок. Точная доза для использования в композиции может также зависеть от способа введения и тяжести заболевания или нарушения, и ее следует определять согласно решению практикующего специалиста и обстоятельств, характерных для каждого пациента. Эффективные дозы можно экстраполировать исходя из кривых доза-ответ, получаемых исходя из тестовых систем in vitro или в виде животных моделей.
Для антител дозировка, вводимая пациенту, составляет, как правило, от 0,1 мг/кг до 150 мг/кг на массу тела пациента. Предпочтительно дозировка, вводимая пациенту, находится между 0,1 мг/кг и 20 мг/кг массы тела пациента, более предпочтительно от 1 мг/кг до 10 мг/кг массы тела пациента. Как правило, человеческие антитела обладают более длительным временем полужизни в организме человека, чем антитела других видов, благодаря иммунному ответу на чужеродные полипептиды. Таким образом, часто возможны более низкие дозировки человеческих антител и более низкая частота введения. Более того, дозировку и частоту введения антител по изобретению можно снизить усилением поглощения и проникновения в ткани (например, в мозг) антител путем модификаций, таких как, например, липидизация. Для повторных введений в течение нескольких суток или более, в зависимости от состояния, лечение поддерживают до тех пор, пока не произойдет требуемое подавление симптомов заболевания. Однако могут быть пригодны другие режимы дозирования. Прогресс этого лечения легко отслеживать общепринятыми способами и анализами.
Композицию антител будут составлять, дозировать и вводить способом, согласующимся с надлежащей медицинской практикой. Факторы для рассмотрения в данном контексте включают конкретное подлежащее лечению нарушение, конкретное подлежащее лечению млекопитающее, клиническое состояние индивидуального пациента, причину нарушения, участок доставки средства, способ введения, режим введения и другие факторы, известные практикующим медицинским специалистам. «Терапевтически эффективное количество» подлежащего введению антитела будет зависеть от таких соображений, и оно представляет собой минимальное количество, необходимое для профилактики, облегчения или лечения заболевания или нарушения. Не обязательно, из антитела получают композицию с одним или несколькими средствами, используемыми в настоящее время для профилактики или лечения рассматриваемого нарушения. Эффективное количество таких других средств зависит от количества антитела, присутствующего в композиции, типа нарушения или лечения и других рассмотренных выше факторов. Их, как правило, применяют в тех же дозировках и теми же способами введения, как отмечено выше, или приблизительно от 1 до 99% от ранее используемых дозировок.
Антитела по изобретению можно вводить по отдельности или в сочетании с другими типами лечения рака, включая общепринятые химиотерапевтические средства (паклитаксел, карбоплатин, цисплатин и доксорубицин), средства анти-EGFR (гефитиниб, эрлотиниб и цетуксимаб), антиангиогенные средства (бевацизумаб и сунитиниб), а также иммуномодулирующие средства, такие как интерферон-α и талидомид.
В предпочтительном аспекте антитело является по существу очищенным (например, по существу не содержащим веществ, которые ограничивают его эффект или вызывают нежелательные побочные эффекты).
Известны различные системы доставки, и их можно использовать для введения антитела по настоящему изобретению, включая инъекцию, например, заключение в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать соединение, рецептор-опосредованный эндоцитоз (см., например, Wu, et al., J Biol Chem 262:4429 (1987)), конструкцию нуклеиновой кислоты как часть ретровирусного или другого вектора и т.д.
Антитело против Notch3 можно вводить млекопитающему любым приемлемым способом. Способы введения включают в качестве неограничивающих примеров парентеральный, подкожный, внутрибрюшинный, внутрилегочный, интраназальный, эпидуральный, ингаляционный и пероральный способы и, если требуется, иммуносупрессивное лечение, введение внутрь очага. Парентеральные инфузии включают внутримышечное, интрадермальное, внутривенное, внутриартериальное или внутрибрюшинное введение. Антитела или композиции можно вводить любым удобным способом, например, инфузией или болюсной инъекцией, поглощением через эпителиальные или кожно-слизистые выстилки (например, слизистую ротовой полости, ректальную и интестинальную слизистую и т.д.), и их можно вводить вместе с другими биологически активными средствами. Введение может являться системным или местным. Кроме того, может являться желательным вводить терапевтические антитела или композиции по изобретению в центральную нервную систему любым подходящим способом, включая внутрижелудочковую и интратекальную инъекцию; внутрижелудочковую инъекцию можно облегчать внутрижелудочковым катетером, например, прикрепленным к резервуару, такому как резервуар Оммайя. Кроме того, антитело подходящим образом вводят пульсовой инфузией, в частности, с понижающимися дозами антитела. Предпочтительно, дозирование осуществляют инъекциями, наиболее предпочтительно, внутривенными или подкожными инъекциями, отчасти в зависимости от того, является ли введение кратковременным или хроническим.
Можно также использовать легочное введение, например, с использованием ингалятора или распылителя и композиции с распыляющим средством. Антитело можно также вводить в легкие пациента в форме композиции из сухого порошка (см. например, патент США No. 6514496).
В конкретном варианте осуществления может являться желательным введение терапевтических антител или композиций по изобретению местно в область, нуждающуюся в лечении; этого можно достичь, например и без ограничения, местной инфузией, местной аппликацией, инъекцией, посредством катетера, посредством суппозитория или посредством имплантата, где указанный имплантат является пористым, непористым или гелеобразным материалом, включающим мембраны, такие как силастиковые мембраны или волокна. Предпочтительно, при введении антитела по изобретению необходимо соблюдать осторожность при использовании материалов, которые не абсорбируют белок.
В другом варианте осуществления антитело можно доставлять в везикуле, в частности, липосоме (см. Langer, Science 249:1527 (1990); Treat, et al., в Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein, et al., eds., pp. 353-365 (1989); Lopez-Berestein, ibid., pp. 317-27; см. в основном ibid.).
В другом варианте осуществления антитело можно доставлять в системе с контролируемым высвобождением. В одном из вариантов осуществления можно использовать насос (см. Langer, Science 249:1527 (1990); Sefton, CRC Crit Ref Biomed Eng 14:201 (1987); Buchwald, et al., Surgery 88:507 (1980); Saudek, et al., N Engl J Med 321:574 (1989)). В другом варианте осуществления можно использовать полимерные материалы (см. Medical Applications of Controlled Release, Langer, et al., eds., CRC Press (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen, et al., eds., Wiley (1984); Ranger, et al., J Macromol Sci Rev Macromol Chem 23:61 (1983); см. также Levy, et al., Science 228:190 (1985); During, et al., Ann Neurol 25:351 (1989); Howard, et al., J Neurosurg 71:105 (1989)). В другом варианте осуществления систему с контролируемым высвобождением можно помещать вблизи терапевтической мишени.
Настоящее изобретение также относится к фармацевтическим композициям. Такие композиции содержат терапевтически эффективное количество антитела и физиологически приемлемый носитель. В конкретном варианте осуществления термин «физиологически приемлемый» означает одобренный контрольным органом федерального правительства или правительства штата или перечисленный в фармакопее США или другой общепризнанной фармакопее для использования у животных и, более конкретно, у человека. Термин «носитель» относится к разбавителю, адъюванту, вспомогательному веществу или носителю, с которым вводят терапевтическое средство. Такие физиологические носители могут представлять собой стерильные жидкости, такие как вода и масла, включая масла нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Вода является предпочтительным носителем, если фармацевтическую композицию вводят внутривенно. Солевые растворы и водные растворы декстрозы и глицерина также можно использовать в качестве жидких носителей, в частности, в растворах для инъекций. Подходящие фармацевтические вспомогательные вещества включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, глицерин-моностеарат, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропилен, гликоль, воду, этанол и т.п. Композиция, при желании, может также содержать минорные количества увлажняющих или эмульгирующих средств, или средств для забуферивания pH. Эти композиции могут принимать форму растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, композиций с замедленным высвобождением и т.п. Композицию можно также составлять в виде суппозитория, с традиционными связующими веществами и носителями, такими как триглицериды. Пероральная композиция может включать общепринятые носители, такие как маннит, лактозу, крахмал, стеарат магния, сахаринат натрия, целлюлозу, карбонат магния фармацевтической степени чистоты и т.д. Примеры подходящих носителей описаны в «Remington's Pharmaceutical Sciences» by E. W. Martin. Такие композиции будут содержать эффективное количество антитела, предпочтительно, в очищенной форме, вместе с подходящим количеством носителя таким образом, чтобы предоставить форму для подходящего введения пациенту. Композиция должна соответствовать способу введения.
В одном из вариантов осуществления композицию составляют согласно общепринятым способам в виде фармацевтической композиции, приспособленной для внутривенного введения человеку. Как правило, композиции для внутривенного введения представляют собой растворы в стерильном изотоническом водном буфере. При необходимости, композиция может также содержать солюбилизирующее средство и местный анестетик, такой как лигнокаин для уменьшения боли в участке инъекции. Как правило, ингредиенты поставляются или по отдельности или в смеси друг с другом в стандартной лекарственной форме, например, в виде сухого лиофилизированного порошка или безводного концентрата в герметически запаянном контейнере, таком как ампула или пакет с указанием количества активного средства. Если композицию следует вводить инфузией, то ее можно дозировать с помощью флакона для инфузии, содержащего стерильную воду фармацевтической степени чистоты или солевой раствор. Если композицию вводят инъекцией, то ампулу со стерильной водой для инъекций или солевым раствором можно предоставить таким образом, что ингредиенты можно смешать перед введением.
Изобретение также относится к фармацевтической упаковке или набору, содержащим один или несколько контейнеров, заполненных одним или несколькими ингредиентами фармацевтических композиций по изобретению. Не обязательно, к такому контейнеру(ам) может прилагаться сообщение в форме, предписанной правительственным учреждением, регулирующим производство, использование или продажу фармацевтических или биологических продуктов, где сообщение отражает одобрение учреждением по производству, использованию или продаже для введения человеку.
Кроме того, антитела по настоящему изобретению можно конъюгировать с различнми эффекторными молекулами, такими как гетерологичные полипептиды, лекарственные средства, радионуклеотиды или токсины. См., например, публикации PCT WO 92/08495, WO 91/14438, WO 89/12624, патент США No. 5314995 и EP 396387. Антитело или его фрагменты можно конъюгировать с терапевтическим компонентом, таким как цитотоксин (например, цитостатическим или цитотоксическим средством), терапевтическим средством или ионом радиоактивного металла (например, альфа-излучателями, такими как, например, 213Bi). Цитотоксин или цитотоксическое средство включает любое средство, которое вызывает повреждение клеток. Примеры включают паклитаксел, цитохалазин B, грамицидин D, этидий-бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи. Терапевтические средства включают в качестве неограничивающих примеров антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацилдекарбазин), алкилирующие средства (например, меклоретамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозоцин, митомицин C и цис-дихлородиамминоплатина (II) (DDP, цисплатин), антрациклины (например, даунорубицин (бывший дауномицин) и доксорубицин), антибиотики (например, дактиномицин (бывший актиномицин), блеомицин, митрамицин и антрамицин (AMC)) и антимитотические средства (например, винкристин и винбластин).
Способы конъюгирования таких терапевтических компонентов с антителами хорошо известны, см., например, Arnon, et al., «Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy», в Monoclonal Antibodies and Cancer Therapy, Reisfeld, et al. (eds.), pp. 243-56, Alan R. Liss (1985); Hellstrom, et al., «Antibodies For Drug Delivery», в Controlled Drug Delivery, 2nd ed., Robinson, et al., eds., pp. 623-53, Marcel Dekker (1987); Thorpe, «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review», in Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera, et al., eds., pp. 475-506 (1985); «Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy», в Monoclonal Antibodies For Cancer Detection and Therapy, Baldwin, et al., eds., pp. 303-16, Academic Press (1985); и Thorpe, et al., Immunol Rev 62:119 (1982). Альтернативно, антитело можно конъюгировать со вторыми антителом с образованием гетероконъюгата антител. См., например, патент США No. 4676980.
Конъюгаты по изобретению можно использовать для модификации данного биологического ответа, где компонент терапевтического средства или лекарственного средства не следует истолковывать как ограниченный классическими терапевтическими средствами. Например, компонент лекарственного средства может представлять собой белок или полипептид, обладающий требуемой биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин A, эндотоксин Pseudomonas или дифтерийный токсин; белок, такой как фактор некроза опухолей, α-интерферон, β-интерферон, фактор роста нервов, фактор роста тромбоцитов, тканевой активатор плазминогена, апоптотическое средство, например, TNF-α, TNF-β, AIM I (см. международную публикацию No. WO 97/33899), AIM II (см. международную публикацию No. WO 97/34911), Fas-лиганд (Takahashi, et al., Int Immunol, 6:1567 (1994)), VEGI (см. международную публикацию No. WO 99/23105); тромботическое средство; антиангиогенное средство, например, ангиостатин или эндостатин; или модификаторы биологического ответа, такие как, например, лимфокины, интерлейкин-1 («IL-1»), интерлейкин-2 («IL-2»), интерлейкин-6 («IL-6»), гранулоцитарно-макрофагальный колониестимулирущий фактор («GM-CSF»), гранулоцитарный колониестимулирующий фактор («G-CSF») или другие факторы роста.
ИЗДЕЛИЯ
Другой вариант осуществления изобретения относится к изделию, содержащему материалы, пригодные для лечения описанных выше нарушений. Изделие содержит контейнер и этикетку. Пригодные контейнеры включают, например, бутыли, флаконы, шприцы и пробирки. Контейнеры могут быть образованы из различных материалов, таких как стекло или пластмасса. Контейнер содержит композицию, которая является эффективной для профилактики или лечения состояния и может обладать отверстием для стерильного доступа (например, контейнер может представлять собой пакет для внутривенного раствора или флакон, обладающий пробкой, проницаемой для иглы для подкожных инъекций). Активное средство в композиции представляет собой антитело. На этикетке на контейнере или в связи с ним указано, что композицию используют для лечения выбранного состояния. Изделие может дополнительно содержать второй контейнер, содержащий фармацевтически приемлемый буфер, такой как фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Кроме того, оно может дополнительно содержать другие желательные с коммерческой и потребительской точки зрения материалы, включая другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши в упаковку с инструкциями для использования.
ГЕННАЯ ТЕРАПИЯ НА ОСНОВЕ АНТИТЕЛ
В другом аспекте изобретения нуклеиновые кислоты, содержащие последовательности, кодирующие антитела или их функциональные производные, вводят для лечения, ингибирования или профилактики заболевания или нарушения, связанного с патологической экспрессией и/или активностью Notch3, с помощью генной терапии. Генная терапия относится к терапии, осуществляемой введением индивидууму экспрессируемой или способной к экспрессии нуклеиновой кислоты. В этом варианте осуществления изобретения нуклеиновые кислоты продуцируют кодируемый ими белок, который опосредует терапевтический эффект. Любые доступные способы генной терапии можно использовать согласно настоящему изобретению. Иллюстративные способы описаны ниже.
Относительно общих обзоров способов генной терапии см. Goldspiel, et al., Clinical Pharmacy 12:488 (1993); Wu, et al., Biotherapy 3:87 (1991); Tolstoshev, Ann Rev Pharmacol Toxicol 32:573 (1993); Mulligan, Science 260:926 (1993); Morgan, et al., Ann Rev Biochem 62:191 (1993); May, TIBTECH 11:155 (1993).
В одном из аспектов соединение содержит последовательности нуклеиновой кислоты, кодирующие антитело, где указанные последовательности нуклеиновой кислоты являются частью экспрессирующих векторов, которые экспрессируют антитело или его фрагменты, или химерные белки, или тяжелые или легкие цепи в подходящем хозяине. В частности, такие последовательности нуклеиновой кислоты содержат промоторы, функционально связанные с кодирующей областью антитела, где указанный промотор является индуцибельным или конститутивным, и, не обязательно, тканеспецифичным.
В другом конкретном варианте осуществления используют молекулы нуклеиновой кислоты, в которых кодирующие последовательности антитела и любые другие требуемые последовательности фланкированы областями, которые способствуют гомологичной рекомбинации в требуемом участке в геноме, таким образом обеспечивая внутрихромосомную экспрессию кодирующих антитело нуклеиновых кислот (Koller, et al., Proc Natl Acad Sci USA 86:8932 (1989); Zijlstra, et al., Nature 342:435 (1989)). В конкретных вариантах осуществления экспрессируемая молекула антитела представляет собой одноцепочечное антитело; альтернативно, последовательности нуклеиновой кислоты включают последовательности, кодирующие как тяжелые, так и легкие цепи антитела или их фрагменты.
Доставка нуклеиновых кислот пациенту может быть или непосредственной, если пациенту непосредственно вводят нуклеиновую кислоту или несущие нуклеиновую кислоту векторы, или опосредованной, если сначала трансформируют клетки нуклеиновыми кислотами in vitro, затем трансплантируют пациенту. Эти два способа известны, соответственно, как генная терапия in vivo или ex vivo.
В конкретном варианте осуществления последовательности нуклеиновой кислоты непосредственно вводят in vivo, где они экспрессируются, продуцируя кодируемый продукт. Это можно осуществить любым из множества способов, известных в данной области, например, конструированием их в виде части подходящего вектора для экспрессии нуклеиновой кислоты и введением их таким образом, что они становятся внутриклеточными, например, путем инфицирования с использованием дефектных или аттенуированных ретровирусных или других вирусных векторов (см. патент США No. 4980286), или непосредственной инъекцией депротеинизированной ДНК, или с использованием обстрела микрочастицами (например, из генной пушки; Biolistic, Dupont), или покрыванием липидами или рецепторами клеточной поверхности, или трансфицирующими средствами, заключением в липосомы, микрочастицы или микрокапсулы, или введением их в связи с пептидом, о котором известно, что он проникает в ядро, введением их в связи с лигандом, подвергаемым рецептор-опосредованному эндоцитозу (см., например, Wu, et al., J Biol Chem 262:4429 (1987)) (который можно использовать для нацеливания на типы клеток, специфически экспрессирующих рецепторы) и т.д. В другом варианте осуществления можно образовать комплексы нуклеиновая кислота-лиганд, в которых лиганд содержит фузогенный вирусный пептид для разрушения эндосом, что позволяет нуклеиновой кислоте избежать лизосомальной деградации. В другом варианте осуществления нуклеиновую кислоту можно нацеливать in vivo для клеточно-специфичного поглощения и экспрессии путем конъюгирования со специфическим рецептором (см., например, публикации PCT WO 92/06180, WO 92/22635, WO 92/20316, WO 93/14188, WO 93/20221). Альтернативно, нуклеиновую кислоту можно вводить внутриклеточно и встраивать в ДНК клетки-хозяина для экспрессии путем гомологичной рекомбинации (Koller, et al., Proc Natl Acad Sci USA 86:8932 (1989); Zijlstra, et al., Nature 342:435 (1989)).
В конкретных вариантах осуществления используют вирусные векторы, которые содержат последовательности нуклеиновой кислоты, кодирующие антитело по изобретению. Например, можно использовать ретровирусный вектор (см. Miller, et al., Meth Enzymol 217:581 (1993)). Эти ретровирусные векторы содержат компоненты, необходимые для правильного упаковывания вирусного генома и интеграции его в ДНК клетки-хозяина. Последовательности нуклеиновой кислоты, кодирующие антитело для использования в генной терапии, клонируют в один или несколько векторов, которые облегчают доставку гена у пациента. Более подробно о ретровирусных векторах можно найти в Boesen, et al., Biotherapy 6:291 (1994), где описано использование ретровирусного вектора для доставки гена mdrl в гематопоэтические стволовые клетки для того, чтобы сделать стволовые клетки более устойчивыми к химиотерапии. Другие ссылки, иллюстрирующие использование ретровирусных векторов в генной терапии, представляют собой: Clowes, et al., J Clin Invest 93:644 (1994); Kiem, et al., Blood 83:1467 (1994); Salmons, et al., Human Gene Therapy 4:129 (1993); и Grossman, et al., Curr Opin Gen and Dev 3:110 (1993).
Аденовирусы можно использовать в настоящем изобретении. Аденовирусы являются особенно привлекательными носителями в настоящем изобретении для доставки антител в дыхательные эпителии. Аденовирусы естественным образом инфицируют дыхательные эпителии. Другими мишенями для систем доставки на основе аденовирусов являются печень, центральная нервная система, эндотелиальные клетки и мышцы. Аденовирусы обладают тем преимуществом, что они способны инфицировать неделящиеся клетки. В Kozarsky, et al., Curr Opin Gen Dev 3:499 (1993), представлен обзор генной терапии на основе аденовирусов. В Bout, et al., Human Gene Therapy 5:3 (1994), показано использование аденовирусных векторов для переноса генов в респираторные эпителии макаки-резуса. Другие случаи использования аденовирусов в генной терапии можно найти в Rosenfeld, et al., Science 252:431 (1991); Rosenfeld, et al., Cell 68:143 (1992); Mastrangeli, et al., J Clin Invest 91:225 (1993); публикация PCT WO94/12649; Wang, et al., Gene Therapy 2:775 (1995). Аденоассоциированный вирус (AAV) также предложен для использования в генной терапии (Walsh, et al., Proc Soc Exp Biol Med 204:289 (1993); патенты США No. 5436146, 6632670 и 6642051).
Другой способ генной терапии включает перенос гена в клетки в культуре тканей такими способами, как электропорация, липофекция, опосредованная фосфатом кальция трансфекция или вирусная инфекция. Как правило, способ переноса включает перенос селективного маркера в клетки. Клетки затем помещают в условия селекции для выделения таких клеток, которые захватили и экспрессируют перенесенный ген. Эти клетки затем вводят пациенту.
В этом варианте осуществления нуклеиновую кислоту вводят в клетку перед введением in vivo получаемой в результате рекомбинантной клетки. Такое введение можно осуществлять любым способом, известным в данной области, включая в качестве неограничивающих примеров трансфекцию, электропорацию, микроинъекцию, инфекцию вирусным или бактериофаговым вектором, содержащим последовательности нуклеиновой кислоты, слиянием клеток, хромосомно-опосредованным переносом гена, опосредованным микроклетками переносом гена, слиянием сферопластов и т.д. Множество способов известно в данной области для введения чужеродных генов в клетки (см., например, Loeffler, et al., Meth Enzymol 217:599 (1993); Cohen, et al., Meth Enzymol 217:618 (1993); Cline, Pharmac Ther 29:69 (1985)), и их можно использовать согласно настоящему изобретению при условии, что не нарушены необходимые, относящиеся к развитию, и физиологические функции клеток-реципиентов. Способ должен обеспечить стабильный перенос нуклеиновой кислоты в клетку таким образом, что нуклеиновая кислота способна экспрессироваться в клетке и, предпочтительно, наследоваться и экспрессироваться в потомстве этой клетки.
Получаемые в результате рекомбинантные клетки можно вводить пациенту различными способами, известными в данной области. Рекомбинантные клетки крови (например, гематопоэтические стволовые клетки или клетки-предшественники) предпочтительно вводят внутривенно. Количество клеток, предусмотренное для использования, зависит от требуемого эффекта, состояния пациента и т.д., и его может определить специалист в данной области.
Клетки, в которые можно вводить нуклеиновую кислоту с целью генной терапии, включают любой требуемый, доступный тип клеток и включают в качестве неограничивающих примеров эпителиальные клетки, эндотелиальные клетки, кератиноциты, фибробласты, мышечные клетки, гепатоциты; клетки крови, такие как T-лимфоциты, B-лимфоциты, моноциты, макрофаги, нейтрофилы, эозинофилы, мегакариоциты, гранулоциты; различные стволовые клетки или клетки-предшественники, в частности, гематопоэтические стволовые клетки или клетки-предшественники, например, которые получают из костного мозга, крови пуповины, периферической крови, плодной печени и т.д.
В одном из вариантов осуществления клетка, используемая для генной терапии, является аутологичной для пациента. Последовательности нуклеиновой кислоты, кодирующие антитело по настоящему изобретению, вводят в клетки таким образом, что они могут экспрессироваться клетками или их потомством, и рекомбинантные клетки затем вводят in vivo для терапевтического эффекта. В конкретном варианте осуществления используют стволовые клетки или клетки-предшественники. Любые стволовые клетки и/или клетки-предшественники, которые можно выделять и поддерживать in vitro, можно потенциально использовать согласно этому варианту осуществления настоящего изобретения (см., например, публикацию PCT WO 94/08598; Stemple, et al., Cell 71:973 (1992); Rheinwald, Meth Cell Bio 21A:229 (1980); Pittelkow, et al., Mayo Clinic Proc 61:771 (1986)).
ПРИМЕРЫ
ПРИМЕР 1: ПОЛУЧЕНИЕ ИММУНОГЕНА: СЛИТЫЙ БЕЛОК ВНЕКЛЕТОЧНОГО ДОМЕНА NOTCH3 И FC
Моноклональные антитела против Notch3, которые специфически связываются с LIN12/димеризационным доменом (в дальнейшем как «LD») Notch3 человека получали с использованием рекомбинантного слитого белка Notch3-Fc в качестве иммуногена, содержащего Notch3-LD, слитый с областью гамма-1-Fc на карбоксиконце. Конкретно, иммуноген содержал аминокислотные остатки слитого белка Notch3-LD с 1378 по 1640 (см. фиг.1) и γ1-Fc человека (Notch3-LD/Fc). Контрольное антитело получали содержащим область EGF-повторов Notch3 с аминокислотными остатками с 43 по 1377 (обозначено как 255A-79).
Последовательность белка Notch3 анализировали с использованием исследовательского программного обеспечения и сервиса на основе Интернет (Motif Search, http//motif.genome.jp/). РНК печени и поджелудочной железы человека (Ambion, Inc. Austin, TX) использовали в качестве матриц для синтезе первой цепи кДНК с использованием общепринятого коммерчески доступного набора для синтеза кДНК. кДНК, кодирующие Notch3-LD и область EGF-повторов, амплифицировали с помощью PCR в присутствии бетаина (1-2 M) и DMSO (5%). Синтезированный с помощью PCR фрагмент ДНК Notch3-LD (~0,8 kb) и фрагмент ДНК EGF-повторов Notch3 (~4 kb) клонировали в экспрессирующиеся векторы, содержащие His-γ1Fc в коммерчески доступном векторе pSec или в коммерчески доступном векторе pCD3.1, где каждый из векторов несет различнй антибиотиковый маркер. В результате клонирования получились две экспрессионные плазмиды, одна экспрессирует слитый белок Notch3-LD/Fc, и другая экспрессирует слитый белок Notch3-EGF/Fc.
Для облегчения конструирования плазмид и усиления экспрессии различных рекомбинантных белков Notch3 получали олигонуклеотиды, соответствующие последовательности лидерного пептида, содержащей первые 135 пар оснований последовательности нуклеиновой кислоты, кодирующей Notch3. Эти олигонуклеотиды, содержали некоторые замены в неоднозначных кодирующих положениях для снижения содержания GC. Все замены в нуклеотидной последовательности оказались молчащими, т.е. отсутствовали замены в аминокислотной последовательности (фиг.14A). После отжига олигонуклеотидов друг с другом сконструированную кодирующую последовательность лидерного пептида соединяли с остальной частью кодирующей последовательности с помощью PCR-SOE (Ho, et al., Gene 77:51 (1989); Horton, et al., BioMethods 8:528 (1990)) (см. фиг.15). Эту кодирующую последовательность лидерного пептида использовали в экспрессионных конструкциях Notch3-LD/Fc и Notch3. Таким образом, оба слитых с Fc белка содержали сигнальный пептид, связанный с N-концом и последовательность γ1Fc человека, слитую с C-концом. Аминокислотная последовательность Notch3-LD, включая лидерный пептид, представлена на фиг.14 и SEQ ID NO:6.
Экспрессию слитых белков Notch3-EGF/Fc и Notch3-LD/Fc подтверждали транзиентной трансфекцией плазмид для экспрессии Notch3 в клетки 293T (номер ATCC CRL-11268, Manassas, VA) и CHO (Invitrogen, Carlsbad, CA), соответственно. Перед трансфекцией клетки культивировали в среде для выращивания DMEM (Invitrogen, Carlsbad, CA), содержащей 10% эмбриональной бычьей сыворотки (FCS), 2 мМ глутамина и 1x раствор незаменимых аминокислот, с последующим высеванием в 6-луночный планшет из расчета приблизительно 3-5×105 клеток на лунку и выращиванием в течение приблизительно 24 часов. Клетки трансфицировали тремя микрограммами каждой из плазмид для экспрессии слитого белка Notch3 в каждой лунке с использованием системы для трансфекции LipoFectamine 2000 (Invitrogen, Carlsbad, CA), следуя протоколу производителя. После трансфекции клетки культивировали в свежей среде для выращивания и культивировали в CO2-инкубаторе в течение приблизительно 40-48 часов перед осуществлением анализа экспрессии слитого белка Notch3. Альтернативно, после трансфекции клетки культивировали в среде для выращивания в течение 3-4 часов, затем переносили в среду DMEM, содержащую 2% FCS, и культивировали в течение приблизительно 60-66 часов перед отбором кондиционированной среды для анализа на секретируемый белок.
Стабильные линии клеток получали как для Notch3-LD/Fc (вектор His-Fcγ/pSec), так и для Notch3-EGF/Fc (вектор His-Fcγ/pSec). Каждую плазмиду трансфицировали в клетки CHO. После трансфекции клетки культивировали в среде для выращивания DMEM в течение ночи, затем переносили в среду для выращивания с 800 мкг/мл гигромицина и культивировали, по меньшей мере, две недели до тех пор, пока клетки, не несущие плазмиду для экспрессии Notch3, не элиминировались с помощью антибиотиков. Кондиционированные среды из стабильных линий клеток подвергали анализу вестерн-блоттингом.
Стабильные или транзиентные трансфицированные клетки анализировали на экспрессию и секрецию слитого белка Notch3-LD/Fc или Notch3-EGF/Fc. Трансфицированные клетки, собранные из культуральных чашек, промывали один раз фосфатно-буферным солевым раствором (PBS) и ресуспендировали в деионизованной воде, смешанной с равным объемом 2x буфера для нанесения образца белка (BioRad, Hercules, CA), и затем нагревали при приблизительно 100°C в течение 10 минут. Секретируемый белок анализировали с использованием кондиционированной среды, смешанной с равным объемом 2x буфера для нанесения образца белка, и нагревали при 100°C в течение 10 минут. Образцы разделяли с использованием SDS-PAGE с градиентом 4-15%. Белки переносили из геля на мембрану PVDF (BioRad, Hercules, CA), которую блокировали в 5% обезжиренном сухом молоке в PBST (PBS с 0,05% твином-20®) в течение, по меньшей мере, одного часа перед переносом белка.
Слитые белки Notch3-EGF/Fc и Notch3-LD/Fc детектировали, инкубируя с γFc-специфическим HRP-конъюгированным антителом (Sigma, St Louis, MO) в блокирующем буфере в течение одного часа при комнатной температуре. Мембрану промывали три раза в PBST и проявляли с помощью хемилюминесцентного субстрата.
Для очистки слитого белка «домен Notch3/Fc», стабильные линии клеток CHO, как описано выше, культивировали в DMEM с 2% FCS вплоть до 5 суток. Один литр кондиционированной среды собирали и подвергали хроматографии на колонке, упакованной гранулами с белком A, для аффинного связывания. Колонку промывали PBS, и связавшиеся белки элюировали в 50 мМ цитратном буфере (pH 2,8), и pH доводили до нейтрального добавлением 1 М буфера Tris-HCl (pH 8). Чистоту белка оценивали анализом белка в геле с использованием SDS-PAGE с градиентом 4-15%. Концентрацию белка анализировали с использованием реагента Кумасси синий согласно протоколу производителя (Pierce, Rockford, IL). Посредством этой процедуры осуществляли очистку белка Notch3-LD/Fc и Notch3-EGF/Fc в количестве миллиграммов для иммунизации и анализов связывания ELISA.
ПРИМЕР 2: ПОЛУЧЕНИЕ АНТИ-NOTCH3 MAB
Самцов мышей A/J (Harlan, Houston, TX) в возрасте 8-12 недель инъецировали подкожно 25 мкг Notch3-EGF/Fc или Notch3-LD/Fc в полном адъюванте Фрейнда (Difco Laboratories, Detroit, MI) в 200 мкл PBS. Через две недели после инъекций и за трое суток до умерщвления мышей мышей снова инъецировали внутрибрюшинно 25 мкг того же антигена в PBS. Для каждой слитой конструкции получали суспензии одиночных клеток из селезенки иммунизированной мыши и использовали их для слияния с миеломными клетками Sp2/0; 5×108 клеток Sp2/0 и 5×108 клеток селезенки подвергали слиянию в среде, содержащей 50% полиэтиленгликоля (M.W. 1450) (Kodak, Rochester, NY) и 5% диметилсульфоксида (Sigma, St. Louis, MO). Затем клетки доводили до концентрации 1,5×105 клеток селезенки на 200 мкл суспензии в среде Iscove (Invitrogen, Carlsbad, CA), с добавлением 10% эмбриональной бычьей сыворотки, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина, 0,1 мкМ гипоксантина, 0,4 мкМ аминоптерина и 16 мкМ тимидина. Двести микролитров клеточной суспензии добавляли к каждой лунке в приблизительно шестьдесят 96-луночных планшетов. После приблизительно десяти суток отбирали культуральные супернатанты для скрининга на активность связывания с антителами с использованием ELISA.
96-луночные плоскодонные планшеты для микротестирования Immulon II (Dynatech Laboratories, Chantilly, VA) покрывали с использованием 100 мкл Notch3-EGF/Fc или Notch3-LD/Fc (0,1 мкг/мл в PBS), содержащих 1x феноловый красный и 3-4 капли pHix/литр (Pierce, Rockford, IL), и инкубировали в течение ночи при комнатной температуре. После удаления раствора для покрытия стряхиванием с планшета 200 мкл блокирующего буфера, содержащего 2% BSA в PBST, содержащего 0,1% мертиолята, добавляли к каждой лунке на один час для блокирования неспецифического связывания. Лунки затем промывали PBST. Пятьдесят микролитров культуральных супернатантов из каждой лунки для слияния собирали и смешивали с 50 мкл блокирующего буфера и затем добавляли к отдельным лункам планшетов для микротитрования. После инкубации в течение одного часа лунки промывали PBST. Связавшиеся мышиные антитела затем детектировали реакцией с Fc-специфическими козьими антимышиными IgG, конъюгированными с пероксидазой хрена (HRP) (Jackson ImmunoResearch Laboratories, West Grove, PA). Раствор субстрата для HRP, содержащий 0,1% 3,3,5,5-тетраметилбензидина и 0,0003% пероксида водорода, добавляли к лункам для развития цветной реакции в течение 30 минут. Реакцию останавливали добавлением 50 мл 2М H2SO4/лунка. Измеряли OD при 450 нм с помощью планшет-ридера для ELISA (Molecular Devices, Sunnyvale, CA).
Среди 185 гибридом, выделенных и проанализированных, два клона гибридом из мышей, иммунизированных Notch3-LD/Fc, продуцировали антитела, направленные против Notch3, которые в дальнейшем охарактеризовали. В ELISA с использованием супернатанта из двух клонов гибридом, продуцирующих MAb 256A-4 и 256A-8, показана сильная активность связывания с очищенным слитым белком Notch3-LD/FC, к которому они были получены, и они не связывались с Notch1-LD/Fc человека (LIN/димеризационный домен, слитый с Fc-областью на карбоксиконце) или с контрольным Fc-белком человека (данные не представлены) (таблица 1). В более поздних исследованиях с использованием функциональных анализов также показали, что MAb 256A-4 и 256A-8 специфически действуют против Notch3 по сравнению с Notch1 и Notch2 (данные не представлены).
|
Положительные клоны гибридом из этого первичного скрининга ELISA в последующем выделяли отбором одиночной колонии и осуществляли второй анализ ELISA, как описано выше, для подтверждения специфического связывания с выбранным иммуногеном. Подтвержденные клоны гибридом размножали в культурах в более крупном масштабе. Моноклональные антитела (MAb) очищали от среды этих крупномасштабных культур с использованием аффинной колонки с белком A. MAb анти-Notch3 затем охарактеризовывали с использованием клеточных анализов связывания, микроскопии, вестерн-блоттинга и анализа FACS.
ПРИМЕР 3: КЛЕТОЧНЫЕ АНАЛИЗЫ СВЯЗЫВАНИЯ АНТИ-NOTCH3 MAB
Для клеточных анализов связывания, используемых для характеристики анти-Notch3 MAb, потребовалось клонирование открытой рамки считывания полноразмерного Notch3 человека в вектор, в этом случае pcDNA3.1/Hygro (Invitrogen, Carlsbad, CA). Кодирующую Notch3 область синтезировали с помощью RT-PCR с использованием РНК из опухоли печени человека (Ambion, Inc., Austin, TX) в качестве матрицы. Окончательная конструкция плазмиды Notch3/Hygro экспрессировала полноразмерный белок Notch3, как представлено на фиг.1. Стабильную линию клеток, экспрессирующую Notch3, получали трансфекцией конструкцией плазмиды Notch3/Hygro в клетки 293T (ATCC No. CRL-11268) с использованием набора LipoFectamine 2000 после той же процедуры, как описано в примере 1. После трансфекции клетки культивировали в среде для выращивания DMEM в течение ночи, затем заново высевали в среду для выращивания с 200 мкг/мл гигромицина и культивировали в течение 12-14 суток. Хорошо изолированные отдельные колонии отбирали и выращивали в отдельных лунках до тех пор, пока количество клеток клонов не увеличится до достаточной степени. Стабильные клоны 293T, которые оказались устойчивы к селекции на гигромицин и экспрессировали белок Notch3 на высоком уровне, идентифицировали с помощью анализа вестерн-блоттингом и флуоресцентной электромикроскопией с использованием поликлональных антител против Notch3 (R&D Systems, Minneapolis, MN).
Плазмиду для экспрессии неполноразмерного Notch3, содержащую только LIN12/димеризационный домен (LD) и трансмембранный домен (TM) Notch, также конструировали с помощью PCR и субклонирования в pcDNA3.1 (Invitrogen, Carlsbad, CA). Эта плазмидная конструкция также содержит метку V5 на своем C-конце, и ее обозначили как Notch3-LDTM/V5. Стабильную линию клеток, экспрессирующую эту плазмиду, Notch3-LDTM/V5, получали согласно процедуре, описанной в примере 1.
Линию клеток Sup-T1 человека (ATCC No. CRL-1942), естественным образом экспрессирующую Notch3, также подтверждали вестерн-блоттингом. Клетки Sup-T1 выращивали в средах RPMI1640, содержащих 10% эмбриональную бычью сыворотку, 2 мМ глутамина и 1X раствор незаменимых аминокислот.
Связывание антител в клетках оценивали с использованием системы FMAT™ (флуоресцентный макроконфокальный высокопроизводительный скрининг) 8100 HTS (Applied Biosystems, Foster City, CA) согласно протоколу, предложенному производителем. Линии клеток, естественным образом экспрессирующие Notch3 или стабильно трансфицированные конструкциями для экспрессии Notch3, высевали в 96-луночные планшеты. Альтернативно, транзиентно трансфицированные клетки 293T или CHO высевали в 96-луночный планшет. Клетки высевали исходя из плотности 30000-50000 клеток на лунку. После 20-24 часов к лункам добавляли MAb анти-Notch3 и 1x реакционный буфер на основе PBS и инкубировали в течение одного часа при 37°C. Антимышиное антитело IgG, конъюгированное с Cy-5, добавляли в лунки после удаления первичных антител.
Связывание антител в клетках также оценивали с помощью флуоресцентно-активированного клеточного сортера (FACS) с использованием стабильной линии клеток 293T/Notch3 собственного получения и двух раковых линий, линий клеток Sup-T1 и A2780 человека (UK ECACC No. Cat. No. 93112519), которые обе естественным образом экспрессируют Notch3 (данные не представлены). Клетки сначала инкубировали с MAb анти-Notch3 в 1x PBS. После трех отмывок клетки инкубировали с вторичными антителами, конъюгированными с флуоресцентной молекулой. Клетки ресуспендировали, фиксировали в 1x PBS с 0,1% параформальдегидом и анализировали с помощью FACS (BD Sciences, Palo Alto, CA). Результаты указывали на то, что оба MAb связываются с рецептором Notch3, экспрессируемым или с рекомбинантных плазмидных конструкций, или в качестве нативного белка в культивируемых клетках (таблица 2). Однако с помощью вестерн-блоттинга показали, что при денатурации рецептора Notch3 или слитого белка Notch3-LD/Fc в SDS-PAGE и переносе на нейлоновую мембрану для блоттинга MAb анти-Notch3 перестают связываться, позволяя предположить конформационный эпитоп. Транзиентно трансфицированные клетки 293T, содержащие плазмиду Notch3/Hygro, также иммунофлуоресцентно окрашивали, как описано выше, и исследовали с помощью флуоресцентной микроскопии.
|
Клеточными анализами FMAT и FACS подтвердили, что оба MAb 256A-4 и 256A-8 действительно связываются с рецептором Notch3, экспрессируемым или с рекомбинантных плазмидных конструкций, или в виде нативного белка в культивируемых клетках (таблица 2 и таблица 3).
|
G3 представляет собой отрицательный контроль в виде Mab IgG1 человека. Положительный сигнал связывания определяли на основе значения сигнала FMAT, которое было значительно выше, чем у G3 и у других отрицательных клонов гибридом (p > 0,01). Значение отрицательного сигнала связывания G3 FMAT рассматривали в качестве фона. Транзиентно трансфицированные клетки 293T с плазмидой Notch3/Hygro также иммунофлуоресцентно окрашивали, как описано выше, и исследовали с помощью флуоресцентной микроскопии.
ПРИМЕР 4: АНАЛИЗ ВЕСТЕРН-БЛОТТИНГОМ АКТИВНОСТИ СВЯЗЫВАНИЯ АНТИ-NOTCH3 Mab
Вестерн-блоттинг осуществляли для оценки активности связывания анти-Notch3 MAb с Notch3 в денатурирующих условиях, а также уровней экспрессии Notch3 и других родственных Notch белков в линиях клеток человека. Очищенный слитый белок Notch3-LD/Fc смешивали с буфером для нанесения белка. Образцы белка также получали из транзиентно или стабильно трансфицированных клеток, описанных в примере 1, которые собирали из культуральных чашек, промывали один раз PBS, ресуспендировали в буфере для экстракции общего клеточного белка (Pierce, Rockford, IL) и прогревали при 100°C в течение 10 минут после добавления равного объема 2x буфера для нанесения образца белка. Все образцы разделяли электрофорезом SDS-PAGE в градиенте 4-15%. Белки переносили из геля на мембрану PVDF, и MAb анти-Notch3 наносили на мембрану для вестерн-блоттинга в качестве первичного антитела для детекции. Конъюгированное с HRP вторичное антитело использовали для детекции, и сигнал получали с использованием хемилюминесцентного субстрата, как описано выше. Антитела для положительного контроля против Fc человека, метки V5, Notch3 и Notch1 приобретали от Invitrogen, R&D Systems, Santa Cruz Biotechnologies и Orbigen.
Анализ вестерн-блоттингом показал, что MAb 256A-4 и 256A-8 не связываются с Notch3-LD/Fc в денатурирующих условиях, что значительно отличается от результатов, наблюдаемых в анализах ELISA и FACS, где LIN12/гетеродимеризационные домены Notch3 сохраняли нативную молекулярную конформацию. Таким образом, следует вывод о том, что MAb 256A-4 и 256A-8 связываются с несколькими эпитопами Notch3-LD, которые должны сохранять свою нативную конформацию. Это заключение подтверждено результатами по картированию эпитопов, рассмотренному в примере 8 ниже.
ПРИМЕР 5: ОЦЕНКА ФУНКЦИОНАЛЬНОСТИ АНТИ-NOTCH3 MAb АНАЛИЗОМ С ИСПОЛЬЗОВАНИЕМ ЛЮЦИФЕРАЗНОГО РЕПОРТЕРА
A. Плазмидные конструкции
Конструкцию для экспрессии полноразмерного Notch3, описанную в примере 3 выше, подтвердили секвенированием, и она оказалась идентичной опубликованной последовательности, представленной на фиг.1. Плазмиду с Jagged1 человека получали от OriGene (Rockville, MD), и при подтверждении секвенированием она оказалась идентичной NM_000214 (депозитарный номер NCBI/GenBank). Вследствие того, что плазмида OriGene Jagged1 не содержала маркера на селекцию с антибиотиком, фрагмент NotI, содержащий кодирующую последовательность Jagged1, переносили в pcDNA3.1/гигромицин. Субклон 3,7 т.п.о. кДНК Jagged2 человека получили синтезом первой цепи кДНК из линии клеток T-клеточного лейкоза, HH (ATCC No. CRL-2105), и амплифицировали с помощью PCR. Затем субклонировали кДНК Jagged2. Экспрессию Notch3, Jagged1 и Jagged2 подтверждали транзиентной трансфекцией и вестерн-блоттингом, как описано в примере 4.
Для получения репортерной люциферазной плазмиды для передачи сигнала Notch, два синтезированных комплементарных олигонуклеотидных праймера, содержащих тандемные повторы связывающего мотива CBF1, обладали следующими последовательностями:
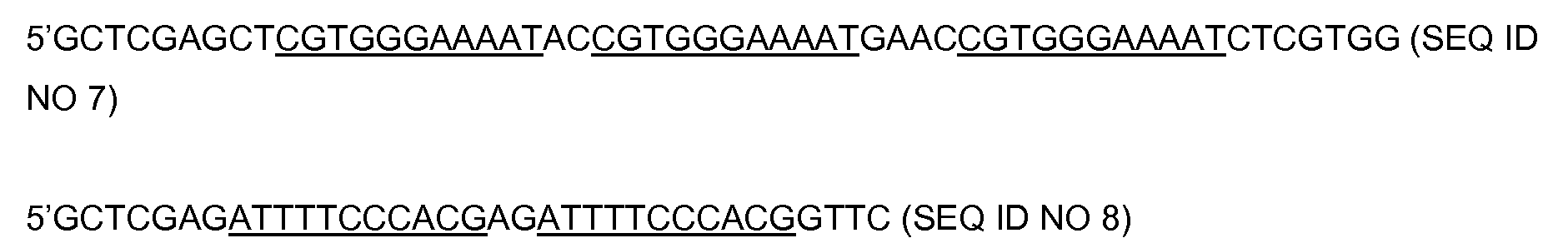
Эти два олигонуклеотидных праймера отжигали при 65°C в 100 мМ NaCl, где концентрация каждого олигонуклеотида составляла 4 мМ. После отжига друг с другом праймеры достраивали с помощью PCR. Продукт PCR клонировали в коммерчески доступный вектор. Вставку подтверждали секвенированием, где вставка содержала четыре тандемных повтора связывающего мотива CBF1 и два фланкирующих XhoI-участка. Вставку выщепляли с использованием XhoI и лигировали ниже кодирующей последовательности люциферазного репортера светляка. После анализа с использованием люциферазного репортера и анализа секвенированием отбирали плазмидные клоны с восемью повторами связывающих мотивов CBF1 и обозначали как CBF1-Luc.
B. Получение стабильной линии клеток
Две стабильные линии клеток получали для функциональных анализов с использованием линий клеток эмбриональной почки человека (HEK293). Одна из линий клеток содержала плазмиду для экспрессии Notch3 и репортерную плазмиду CBF1-Luc, интегрированную в ядерный геном. Эту линию клеток получали совместной трансфекцией плазмид Notch3/гигромицин и CBF1-Luc в клетки 293T с использованием LipoFectamine 2000 согласно протоколу производителя. Стабильные трансфицированные клоны клеток отбирали против 200 мкг/мл гигромицина в среде для выращивания DMEM и скринировали анализом с использованием люциферазного репортера и вестерн-блоттингом. Линию клеток с относительно высоким уровнем экспрессии Notch3 (на основе вестерн-блоттинга) и люциферазной активности отбирали для использования в функциональном анализе и обозначали как NC85.
Вторая стабильная линия клеток содержала конструкцию для экспрессии лиганда Notch, такого как Jagged1, или Jagged2, или pcDNA3.1, в качестве отрицательного контроля. Стабильные линии клеток, экспрессирующие Jagged1 человека или несущие pcDNA3.1, получали трансфекцией клеток 293T и селекцией против гигромицина, как описано выше. Jagged2 субклонировали, трансфицировали в линию клеток 293T, и предполагалось, что он интегрируется в специфический локус в геноме. Устойчивые к гигромицину клетки отбирали, как указано выше.
C. Анализ с использованием люциферазного репортера в условиях совместного культивирования
Клетки NC85 смешивали и совместно культивировали с другой линией клеток 293T, стабильно экспрессирующей Jagged1 человека (Jagged1/293T), Jagged2/293F или pcDNA3.1/293T соответственно в течение от 24 до 48 часов. В конце совместного культивирования среды удаляли отсасыванием, клетки лизировали в 1x буфере для пассивного лизиса (E1501, Promega, Madison, WI), и активности люциферазы анализировали с использованием системы для анализа люциферазы согласно протоколу производителя (E1501, Promega, Madison, WI) в люминометре TD-20/20 (Turner Designs Instrument, Sunnyvale, CA). Как представлено на фиг.6 и фиг.7, при совместном культивировании клеток NC85 с Jagged1/293T или с Jagged2/293F, люциферазная активность повышалась в 2-4 раза по сравнению с активностью при совместном культивировании с клетками pcDNA3.1/293T. Для оценки ингибиторного эффекта MAb анти-Notch3 антитела добавляли к культуре клеток в начале высевания и смешивания совместно культивируемых клеток (256-A, 256A-8 и контроль с доменом с EGF-повторами 255A-79).
D. Анализ с использованием люциферазного репортера при культивировании клеток в планшетах, покрытых лигандом Notch
Общепринятые 96-луночные планшеты для культуры тканей от Becton Dickinson Labware (#18779, Palo Alto, CA) покрывали Jagged1/Fc крысы, DLL-4 человека (R&D Systems, Minneapolis, MN) или Fc человека (Jackson ImmunoResearch, West Grove, PA), бычьим сывороточным альбумином (Sigma, St Louis, MO). По сто микролитров каждого белка (3 мкг/мл в PBS) распределяли в лунки и инкубировали при комнатной температуре или 4°C в течение, по меньшей мере, 8 часов до удаления покрывающего раствора перед использованием. Клетки NC85 или раковые клетки высевали из расчета 3-5×104 клеток на лунку и оставляли расти в течение 28-48 часов. Анализ с использованием люциферазного репортера и анализ ингибирования антителами осуществляли, как описано выше в разделе C. Анализ с использованием люциферазного репортера показал, что связывание MAb 256A-4 и 256A-8 с LIN12/димеризационным доменом почти полностью блокировало индуцированную Jagged1 и Jagged2 активность люциферазного репортера (фиг.6 и 7). В то же время специфическое связывание MAb с EGF-доменом Notch3 (255A-79), в качестве контроля, только ингибировало индуцированную Jagged1 активность люциферазного репортера (приблизительно, ингибирование на 60%, фиг.6), но
неиндуцированную Jagged2 активность люциферазного репортера (фиг.7). Способность MAb 256A-4 и 256A-8 блокировать индуцированную DLL-4 активность люциферазного репортера представлена на фиг.8.
С помощью дополнительных функциональных анализов показали, что MAb 256A-4 и 256A-8 ингибировали индуцированную лигандом положительную регуляцию генов-мишеней Notch. Клетки 293T, экспрессирующие рекомбинантный Notch3, культивировали в покрытых Jagged-1 планшетах. В присутствии MAb 256A-4 и 256A-8 происходило ингибирование положительной регуляции HES5 и HEY2, двух генов-мишеней Notch, при измерении с помощью количественной RT-PCR (данные не представлены).
Для подтверждения того, могут ли анти-Notch3 MAb связываться с нативным Notch3, экспрессируемым в раковых клетках человека, и блокировать обусловленную рецептором передачу сигнала, осуществляли репортерный анализ с использованием двух линий клеток рака яичника, OV/CAR3 и A2780. Оба 256A-4 и 256A-8 в значительной степени блокировали индуцированный Jagged1 Notch-сигналинг, опосредованный нативным Notch3 в клетках OV/CAR3 (фиг.9A). Сходным образом оба MAb ингибировали приблизительно 50% люциферазной активности, индуцируемой DLL-4, которым покрыт планшет (фиг.9B). Последний результат согласуется с тем фактом, что как Notch1, так и Notch3 экспрессируются в клетках A2780. Эти результаты позволяют предположить, что MAb анти-Notch3 может ингибировать опосредованную нативным Notch3 передачу сигнала в раковых клетках.
ПРИМЕР 6: АНАЛИЗ АПОПТОЗА
Аннексин V представляет собой ранний маркер апоптоза на клеточной поверхности, и популяцию апоптотических клеток можно маркировать меченным флуорофором антителом анти-аннексин V и количественно определять анализом FACS. Клетки NC85 высевали из расчета 5-6×104 клеток на лунку в покрытый Fc или Jagged1/Fc 96-луночный планшет, как описано выше, и инкубировали в бессывороточной среде DMEM в течение 24 часов. Апоптотические клетки окрашивали меченными FITC антителами анти-аннексин V (BD Biosciences, Palo Alto, CA) и анализировали с помощью FACS. Клетки, культивируемые на поверхности, покрытой Jagged1/Fc, содержали значительно меньшую популяцию апоптотических клеток по сравнению с клетками, культивируемыми на покрытом Fc планшете (фиг.10). Для исследования функционального эффекта антитела, MAb анти-Notch3 добавляли в культуру клеток в начале исследования. Как представлено на фиг.10, анти-Notch3 MAb 256A-4 и 256A-8 блокировали приблизительно на 50-65% эффект выживания клеток, индуцированный Jagged1.
ПРИМЕР 7: АНАЛИЗЫ КЛЕТОЧНОЙ МИГРАЦИИ, АНАЛИЗЫ ИНВАЗИИ И АНАЛИЗЫ МОРФОЛОГИИ
Анализы in vitro клеточной миграции и инвазии часто используют для оценки метастатического потенциала раковых клеток. Эти анализы осуществляли для анализа ингибиторного эффекта, оказываемого MAb анти-Notch3 на онкогенную стабильную линию клеток 293T/Notch3 (NC85). Анализ инвазии осуществляли с использованием 48-луночного планшета со вставкой Costar (Sigma-Aldrich, St. Louis, MO). Вставка разделяет лунку на верхнюю и нижнюю камеры, которые разделены пористой мембраной (диаметр пор = 8 мкм) на нижней части вставки. Осуществляли иммобилизацию лигандов Notch, Jagged1/Fc, DLL-4 или Fc человека на поверхности мембраны, как описано в вышеописанных разделах. Клетки NC85 высевали из расчета 100000 клеток на лунку и инкубировали в бессывороточной DMEM в верхней камере и 10% FCS/DMEM в нижней камере. После 10-24 часов удаляли клетки, которые оставались на верхней поверхности мембраны вставки, и клетки, которые проходили через мембрану, прикрепляясь на нижней части мембраны вставки, окрашивали 0,05% кристаллическим фиолетовым в PBS. Краситель экстрагировали из клеток 30% уксусной кислотой и регистрировали показатели поглощения при 590 нм. MAb анти-Notch3 добавляли к культуре клеток за 24 часа перед высевом клеток NC85 в планшет для анализа Costar и все MAb добавляли к культуре клеток за 24 часа перед высевом клеток NC85 в планшет для анализа Costar. Свежие MAb добавляли для поддержания той же самой концентрации в планшете для анализа миграции. Экспериментальные результаты представлены на фиг.11A.
Анализ инвазии осуществляли с использованием 48-луночного планшета с матригелем Becton Dickinson (BD Labware, Palo Alto, CA). Лунку с культурой клеток разделяли вставкой для лунки на верхнюю и нижнюю камеры, которые разделены пористой мембраной (диаметр пор = 8 мкм) в нижней части вставки для лунки. Матригелем исходя из оптимизированной плотности покрывали верхнюю поверхность мембраны, и покрытие фибронектином нижней поверхности мембраны осуществлено производителем. Клетки NC85, Jagged1/293T и pcDNA3.1/293T смешивали попарно так, как указано на фиг.11 B. Суммарно 6-10×104 клеток высевали в каждую лунку в 48-луночный планшет с матригелем и культивировали в среде для выращивания в течение 24 часов. Клетки, которые оставались на верхней части мембраны вставки в верхней камере, удаляли и клетки, которые прошли через мембрану, прикрепившись к нижней стороне мембраны вставки, окрашивали 0,05% кристаллическим фиолетовым в PBS. Краситель экстрагировали и измерения поглощения осуществляли, как описано в предыдущем разделе. MAb добавляли в начале смешивания культур клеток. результаты представлены на фиг.11B.
Результаты анализа клеточной миграции показали, что если клетки NC85 культивировали на покрытой Jagged1 мембране, то активация передачи сигнала Notch3 значительно усиливала клеточную миграцию, и MAb 256A-4 и 256A-8 отчетливо ингибировали миграцию (фиг.11A). В эксперименте по инвазии выявлена сходная тенденция (фиг.11B).
Кроме того, исследовали эффект MAb 256A-4 и 256A-8 на индуцированное Jagged-1 образование клеточных «сфер». Если клетки 293T, сверхэкспрессирующие Notch3, культивировали на покрытых Jagged-1 планшетах, то клетки образовывали слабо прикрепленные «клеточные шары» или «сферы». Однако в присутствии MAb 256A-4 и 256A-8 образование этих клеточных сфер ингибировалось (данные не представлены).
ПРИМЕР 8: КАРТИРОВАНИЕ ЭПИТОПА, СВЯЗЫВАЮЩЕГОСЯ С MAB ПРОТИВ NOTCH3
A. Стратегия перестановки доменов и обоснование
Во-первых, антагонистические анти-Notch3 MAb связываются с LIN12/димеризационным доменом (LD) Notch3, но не с гомологичным LIN12/димеризационным доменом Notch1 человека (см. фиг.12 и 13). Во-вторых, анти-Notch3 MAb не связываются с денатурированным белком Notch3 в вестерн-блоттинге, как рассмотрено в примере 4, указывая на то, что MAb связываются с конформационными эпитопами. В-третьих, Notch3 и Notch1 обладают приблизительно 55% гомологией аминокислотной последовательности в LIN12/димеризационном домене, и, таким образом, сделано заключение о том, что перестановка доменов между Notch3 и Notch1 внутри этой области может не нарушить конформацию белка.
B. Получение конструкций слитого белка с переставленными доменами
Анализ последовательности указывал на то, что Notch3 обладает тремя повторами LIN12 и его димеризационный домен разделен на два сегмента. Таким образом, получено пять конструкций белка с перестановкой доменов, где каждый из трех повторов LIN12 и два димеризационных сегмента заменили соответствующими доменами Notch1. Конструкции с перестановкой доменов получали с использованием PCR-SOE (Ho, et al., Gene 77:51 (1989); Horton, et al., BioMethods 8:528 (1990)), как проиллюстрировано на фиг.12. Реакции PCR и PCR-SOE осуществляли с использованием PCR с 1 М бетаином и 5% DMSO, добавленными в реакцию. Термоциклирование PCR являлось почти одинаковым для PCR и PCR-SOE, за исключением того, что стадию отжига для каждого цикла PCR увеличивали на одну минуту в PCR-SOE. Окончательный продукт PCR-SOE субклонировали и подтверждали секвенированием. Плазмидный клон с правильной вставленной последовательностью расщепляли NheI и XhoI для выщепления вставки, которую очищали в геле и субклонировали. Пять конструкций с перестановкой доменов Notch3/Notch1 проиллюстрированы на фиг.12. Для облегчения картирования эпитопа сигнальный пептид каппа-цепи IgG человека использовали в качестве лидерного пептида в конструкциях с переставленными доменами. Аминокислотные последовательности представлены на фиг.16 и 17.
кДНК Notch1-LD амплифицировали с помощью PCR с использованием PCR и способов, описанных в разделе выше. Первую цепь матрицы кДНК синтезировали исходя из суммарной РНК клеток PA-1 (ATCC No. CRL-1572). Кодирующую последовательность лидерного пептида каппа-цепи IgG человека амплифицировали с помощью PCR, используя ее в качестве лидерного пептида для связывания с 5'-Notch1-LD с помощью PCR-SOE, и субклонировали в His-γ1Fc/pSec.
На основе результатов анализа ELISA целевые домены L1, D1 и D2 далее разделяли на субдомены. Анализ связывания ELISA с использованием конструкций для экспрессии субдоменов показал, что только L1 и D2 необходимы для связывания MAb анти-Notch3. Домен D1 не являлся необходимым. Таким образом, домены L1 и D2 разделили на кластеры аминокислотных мутаций для последующего анализа специфического участка связывания. Получали конструкции, содержащие перестановку субдоменов L1 и D2 или кластеры аминокислотных мутаций, как представлено на фиг.16 и фиг.17.
C. Экспрессия слитого белка с перестановкой доменов Notch3/Notch1
Плазмиды с переставленными доменами Notch3/Notch1-LD транзиентно трансфицировали в клетки CHO с использованием LipoFectamine 2000. Клетки CHO высевали в среду для выращивания DMEM с 10% FCS из расчета 0,8-1×106 клеток на лунку в 6-луночный планшет, инкубировали в CO2-инкубаторе в течение ночи перед трансфекцией. Клетки после трансфекции переносили в среду для выращивания приблизительно на 3 часа, затем переносили в DMEM с 2% FCS и культивировали в течение трех суток. Кондиционированные среды собирали и центрифугировали при 3500 об/мин в течение 10 минут. Супернатант, содержащий белок с перестановкой домена Notch3-LD, секретируемый CHO, собирали и подготавливали для вестерн-блоттинга и анализов связывания ELISA. С помощью ELISA показали, что все слитые белки с перестановкой доменов экспрессировались и секретировались в кондиционированную среду (таблица 4), что далее подтверждено анализом с помощью вестерн-блоттинга (данные не представлены).
Данные ELISA с использованием антитела к Fc человека в качестве антитела для детекции показали, что все белки экспрессировались в кондиционированной среде. IgG/Fc человека использовали в качестве контроля. Начальная точка для IgG/Fc человека, которым была покрыта каждая лунка, составляет 100 нг.
|
Сокращения для белков, используемых в анализах связывания ELISA на таблице 4, включают: N1-LD - Notch1-LD/Fc, N3-LD - Notch3-LD/Fc, перестановка L1 - перестановка 1-го домена LIN12, перестановка L2 - перестановка 2-го домена LIN12, перестановка L3 - перестановка 3-го домена LIN12, перестановка D1 - перестановка 1-го димеризационного домена, перестановка D2 - перестановка 2-го димеризационного домена, hIgG-Fc, IgG-Fc человека.
D. Анализ связывания эпитопа с использованием ELISA
96-луночные плоскодонные планшеты для микротестирования Immulon II (Dynatech Laboratories, Chantilly, VA) покрывали антителами к Fc человека (Jackson ImmunoResearch) путем добавления 100 мкл антител (0,1 мкг/мл) в фосфатно-буферном солевом растворе (PBS), содержащем 1x феноловый красный и 3-4 капли pHix/литр (Pierce, Rockford, IL), и инкубировали в течение ночи при комнатной температуре. После покрывания раствор удаляли стряхиванием с планшета, добавляли к каждой лунке 200 мкл блокирующего буфера, содержащего 2% BSA в PBST и 0,1% мертиолята, на один час для блокирования неспецифического связывания. Лунки затем промывали PBST. Пятьдесят микролитров вышеуказанной кондиционированной среды из каждой трансфекции конструкцией с перестановкой доменов Notch3/Notch1 собирали, смешивали с 50 мкл блокирующего буфера и добавляли к отдельным лункам планшетов для микротитрования. После инкубации в течение одного часа белок с перестановкой доменов Notch3/Notch1-LD улавливали нанесенными антителами анти-Fc, и лунки промывали PBST. Анти-Notch3 MAb и подходящие по изотипу контрольные MAb последовательно разводили в блокирующем буфере, как описано выше, и добавляли к каждой лунке 50 мкл разведенных MAb для оценки связывания со связанным белком с перестановкой доменов Notch3/Notch1. Для детекции использовали конъюгированный с пероксидазой хрена (HRP) Fc-специфический козий антимышиный IgG. Раствор субстрата HRP, содержащий 0,1% 3,3,5,5-тетраметилбензидина и 0,0003% пероксида водорода, добавляли к лункам для развития цветного окрашивания в течение 30 минут. Реакцию останавливали добавлением 50 мл 2 М H2SO4/лунка. OD при 450 нм измеряли с помощью ELISA-ридера. Конструкции с перестановкой субдоменов и кластеры мутаций сходным образом исследовали с помощью вышеописанного анализа ELISA.
Эксперименты со связыванием в ELISA с использованием MAb 256A-4 и 256A-8 против белков с перестановкой доменов, показали, что перестановка 1-го домена LIN12 (L1) и 2-го димеризационного домена (D2) полностью устраняла связывание со всеми тремя MAb, в то время как перестановка 1-го димеризационного домена (D1) устраняла связывание с MAb 256A-4 и 256A-8 (фиг.13B и C). Перестановка 3-го домена LIN12 (L3) значительно ослабляла связывание. Тем не менее, оба MAb все еще сохраняли способность связываться со слитым белком. Перестановка 2-го домена LIN12 не нарушала связывание MAb (фиг.13B и C). Антитело в качестве положительного контроля, которое, как картировали ранее, связывается с 1-м доменом LIN12, связывается со всеми слитыми белками с перестановкой доменов за исключением L1 (фиг.13D). В то же время, изотипические антитела в качестве отрицательного контроля, G3, не связываются с любым из слитых белков с перестановкой доменов в анализе ELISA (данные не представлены). Сделан вывод из вышеописанных экспериментов о том, что 1-й домен LIN12 и 2-й димеризационный домен необходимы для связывания с MAb 256A-4 и 256A-8.
Для последующего картирования эпитопов в 1-м домене LIN12 (L1), с которым связываются MAb анти-Notch3, домен L1 далее разделили на три субдомена, L1-sub1, L1-sub2 и L1-sub3, и их заменили на соответствующие последовательности Notch1 (фиг.16). В анализе связывания ELISA показано, что перестановка L1-sub1 не обладает ингибиторными эффектами на активность связывания и перестановка L1-sub2 и L1-sub3 устраняла связывание (фиг.16). В областях L1-sub2 и L1-sub3 существует пять кластеров аминокислотных остатков, которые различаются у Notch3 и Notch1. Таким образом, конструкции слитого белка с перестановками получали в пределах этих пяти кластеров аминокислот (фиг.16). В анализе ELISA выявлено, что перестановка L1-кластер4 не обладала ингибированием на связывание со всеми тремя MAb. Оставшиеся четыре кластера перестановки частично или полностью устраняли связывание с MAb анти-Notch. Таким образом, эти четыре кластера аминокислотных остатков представляли четыре различных эпитопа, с которыми происходит связывание MAb. Необходимы L1-кластер3 (аминокислоты: DRE) и L1-кластер5 (аминокислоты: SVG). L1-кластер1 (аминокислоты: AKR) и кластер2 (аминокислоты: DQR) также играли роль в связывании с MAb анти-Notch3, мутации в которых значительно ослабляли связывание с MAb.
Для картирования эпитопов во 2-м димеризационном (D2) домене Notch3, с которым связываются MAb анти-Notch3, домен D2 далее разделили на пять субдоменов, D2-sub1, D2-sub2, D2-sub3, D2-sub4 и D2-sub5. Последовательности в этих субдоменах заменяли на соответствующие последовательности Notch1 (фиг.17). В анализе связывания ELISA показано, что MAb 256A-4 и 256A-8 обладают сильным связыванием с перестановкой D1-sub2 и D2-sub3, но не с перестановкой D2-sub1 и D2-sub4. Оба MAb обладали слабым связыванием с D2-sub5 (фиг.17). Таким образом, данные позволили предположить, что D2-sub1 и D2-sub4 необходимы для связывания с MAb анти-Notch3 и D2-sub5 может способствовать активности связывания.
Оба MAb 256A-4 и 256A-8 являются антагонистическими антителами, связывающимися с конформационным эпитопом, содержащим L1 и D2, в то время как другое антитело 256A-13, которое связывается только с L1, является агонистическим (см. рассматриваемую совместно заявку США No. 11/874682, поданную 18 октября, 2007). Более того, агонистическое антитело 256A-13 конкурирует с антагонистическим антителом 256A-4 за эпитоп внутри L1, и исследования по картированию эпитопа позволяют предположить, что они связываются с перекрывающимся эпитопом на L1. Основное отличие состоит в том, что антагонистические антитела также связываются с D2, в то время как агонистическое антитело не связывается. Для проверки гипотезы о том, что одновременное связывание с L1 и D2 отвечает за антагонистическую активность антитела, анализировали связывание 256A-2 с эпитопом в D2, сходным с эпитопом для 256A-4. MAb 256A-2 не является ни антагонистическим, ни агонистическим (данные не представлены). Исследования показали, что 256A-2 не конкурирует с 256A-13, и они могут связываться одновременно с Notch3. Более того, 256A-2 и 256A-13 по отдельности могут частично конкурировать с 256A-4, однако в сочетании эти два антитела полностью блокируют связывание 256A-4 с Notch3 (данные не представлены). Исследования также показали, что раздельное связывание двух антител с эпитопами в L1 и D2 не ведет к ингибированию лиганд-зависимой активации Notch3, позволяя предположить, что антагонистические антитела формируют мостик, возможно фиксируя и стабилизируя взаимодействие L1 и D2 и предотвращая индуцированные лигандом конформационные изменения (см. фиг.18).
ПРИМЕР 9: СЕКВЕНИРОВАНИЕ АНТИ-NOTCH3 MAB
Вследствие того, что характеристики связывания антител зависят от вариабельных областей как тяжелой цепи, так и легкой цепи, вариабельные последовательности 256A-4 и 256A-8 субтипировали и секвенировали. Подтип антитела IgG определяли с использованием набора Isostrip Mouse Monoclonal Antibody (Roche Diagnostics, Indianapolis, IN). Результаты показали, что оба MAb, 256A-4 и 256A-8, содержат тяжелую цепь IgG1 и легкую цепь каппа.
Последовательности вариабельной области тяжелой цепи и легкой цепи расшифровали с помощью RT-PCR и клонирования кДНК. Суммарную РНК из клонов гибридом 256A-4 и 256A-8 выделяли с использованием набора RNeasy Mini согласно протоколу производителя (QIAGEN, Valencia, CA). Первую цепь кДНК синтезировали с использованием матрицы РНК и набора Superscriptase III. кДНК вариабельной области легкой цепи и тяжелой цепи амплифицировали с помощью PCR с первой цепи кДНК с использованием вырожденных прямых праймеров, покрывающих 5'-конец кодирующей области каппа-цепи мыши, и обратного праймера, соответствующего константной области в месте соединения с 3'-концом вариабельной области, или с использованием вырожденных прямых праймеров, покрывающих 5'-конец кодирующей области тяжелой цепи мыши, и обратного праймера к константной области тяжелой цепи мыши. Продукт PCR клонировали в коммерчески доступный вектор и секвенировали с помощью Lone Star Lab (Houston, TX). Нуклеотидные последовательности анализировали с использованием компьютерного программного обеспечения DNAStar (DNASTAR, Inc., Madison, WI). Каждую последовательность MAb анти-Notch3 определяли с помощью последовательностей из нескольких клонов PCR, полученных из того же клона гибридомы.
MAb 256A-4 содержит 123 и 116 аминокислотных остатков в своей вариабельной области тяжелой цепи и легкой цепи, соответственно (фиг.4A и 4B). MAb 256A-8 состоит из 122 и 123 аминокислотных остатков в вариабельных областях тяжелой цепи и легкой цепи, соответственно (фиг.5A и 5B).
ПРИМЕР 10: ВЛИЯНИЕ АНТАГОНИСТИЧЕСКИХ АНТИТЕЛ К NOTCH3 НА РАСЩЕПЛЕНИЕ NOTCH3 МЕТАЛЛОПРОТЕАЗОЙ
Активация рецептора Notch включает индуцированное лигандом расщепление металлопротеазой в околомембранном участке (S2), приводя к образованию внеклеточной субъединицы. Это расщепление является существенной предпосылкой для отщепления S3 с высвобождением внутриклеточной области активированного Notch. Обнаружено, что для связывания как 256A-4, так и 256A-8 необходимы наличие, по меньшей мере, части доменов L1 и D2 Notch3. Эти два домена не расположены в непосредственной близости в линейной последовательности, но скорее расположены на двух отдельных полипептидах, позволяя предположить, что эти антитела могут стабилизировать неактивную, аутоингибированную конфигурацию Notch. Для тестирования того, могут ли антагонистические антитела ингибировать последовательные события активации Notch, включая два протеолитических расщепления, клетки 293T, стабильно экспрессирующие рекомбинантный рецептор Notch3 (клетки NC85), или подвергали воздействию иммобилизованного рекомбинантного Jagged-1 или совместно культивировали с клетками 293T, экспрессирующими Jagged-1. Растворимые внеклеточные субъединицы, получаемые при протеолитическом расщеплении в культуральной среде, детектируют анализом ELISA с использованием антитела, связанного с твердой поверхностью, которое распознает продукт расщепления Notch3. Как ожидают, антагонистические MAb к Notch3 уменьшают образование растворимых внеклеточных субъединиц Notch3 в кондиционированной среде, в то время как нефункциональные связывающиеся с Notch3 антитела не уменьшают.
Для непосредственной детекции фрагмента расщепления S2 осуществляли электрофорез в 7,5% SDS PAGE и вестерн-блоттинг с антителом к C-концу Notch3. Фрагмент S2 на 57 аминокислотных остатков меньше и мигрирует немного быстрее, чем нерасщепленная малая субъединица Notch3 (трансмембранная субъединица).
Для исследования того, ингибируют ли антагонистические MAb к Notch3 индуцированное лигандом расщепление металлопротеазой Notch3 в S2, клетки 293T, экспрессирующие рекомбинантный Notch3, обрабатывали соединением E ингибитора γ-секретазы (1 мкМ) в течение 4 часов, которое стабилизирует продукт расщепления в участке S2, обеспечивая его накопление. В присутствии MAb 256A-4 и 256A-8 индуцируемое Jagged-1 расщепление металлопротеазой Notch3 в S2 ингибировалось (данные не представлены).
ПРИМЕР 11: ИССЛЕДОВАНИЕ ЭФФЕКТИВНОСТИ С ИСПОЛЬЗОВАНИЕМ МОДЕЛЕЙ РАКА ЧЕЛОВЕКА У МЫШЕЙ С КСЕНОТРАНСПЛАНТАТОМ
A. Раковые клетки и онкогенные клетки человека
Раковые линии клеток человека с экспрессией Notch3, такие как HCC2429, HCC95, можно получить из академических институтов или из ATCC. 293T/pcDNA3.1 и 293T/Notch3 (NC85) получали трансфицированием 293T соответствующими генами и отбором с гигромицином, как описано в предыдущих разделах. Все клетки культивировали в среде DMEM или RPMI 1640 с 10% эмбриональной бычьей сывороткой, пируватом натрия, заменимыми аминокислотами, L-глутамином, раствором витаминов и пенициллином-стрептомицином (Flow Laboratories, Rockville, MD). Линии клеток инкубировали в смеси 5% CO2 и 95% воздуха при 37°C в инкубаторе. Культуры поддерживали в течение не более чем 3 недель после извлечения из замороженных запасов. Логарифмически растущие одноклеточные суспензии клеток с ≥90% жизнеспособностью использовали для инъекции опухолевых клеток после промывания PBS.
B. Животные
Мышей получали, например, от Animal Production Area из National Cancer Institute в Frederick Cancer Research and Development Center, Frederick, MD. Было осуществлено целевое разведение животных, и они не подвергались экспериментам до начала исследования. Мышей, отобранных для использования в исследованиях, выбирали таким образом, чтобы они по возможности были однородными по возрасту и массе. Их возраст составлял 6-8 недель, и их массы тела в начале измерения массы варьировали от приблизительно 18 до 25 граммов. Записи данных о рождении для животных, используемых в этом исследовании, сохранены в исходных данных исследования, и диапазон массы во время распределения по группам отмечен в сообщении. Каждого животного идентифицировали по пронумерованной метке на ухе. Осуществляли групповое содержание животных по группам исследования (4 мыши/клетка) в полистирольных одноразовых клетках типа ящика для обуви, содержащих целлюлозную подстилку, отвечающих нормативам NIH или превосходящих их. В ходе курса исследования окружающие условия в комнате с животными отслеживались и поддерживались в температурном диапазоне 18-26°C, и относительную влажность регистрировали ежедневно. 12-часовой цикл дневного/ночного освещения поддерживался в ходе исследования. Животных кормили высушенной пищей. Известно, что в пище отсутствовали контаминанты на уровнях, которые могут влиять на результаты исследования. Автоклавированная вода доступна для каждого животного посредством бутылок с водой. Известно, что в воде отсутствовали контаминанты на уровнях, которые могут влиять на результаты исследования. Перед назначением для исследования всех животных акклиматизировали к предназначенному для них содержанию, по меньшей мере, в течение 7 суток перед первым днем дозирования.
C. Модели опухоли и исследования эффективности
Мышей анестезировали с использованием фенобарбитала натрия (50 мг/кг массы тела) и располагали в положении лежа на правом боку. Раковые клетки, такие как линии клеток немелкоклеточного рака легкого (NSCLC), HCC2429 (Haruki, et al. Cancer Res. 65:3555 (2005)), HCC95 (от Dr. John Mina) и H2122 (ATCC No. CRL5985) в 50 мкл раствора Хэнка, содержащего 10% матригеля, инъецировали в левую долю легких. После инъекции опухолевых клеток мышей поворачивали в положение лежа на левом боку и наблюдали за ними в течение 45-60 мин до тех пор, пока они полностью не восстановились. Записи об инъекциях опухолевых клеток сохранены в исходных данных исследования.
За всеми животными производили наблюдение в их клетках, по меньшей мере, один раз в сутки в ходе исследования и клинические результаты записывали в исходных данных исследования. Животные, у которых наблюдали выраженные неблагоприятные эффекты, могли быть удалены из исследования, если бы это посчитали необходимым. Массу тела измеряли один раз в неделю в ходе лечения. Раковые ткани из каждой мыши, при доступности, собирали и хранили для возможного будущего получения биологических характеристик.
ПРИМЕР 12: АНАЛИЗ СВЯЗАННЫХ С NOTCH3 ЗАБОЛЕВАНИЙ
Для идентификации других связанных с Notch3 заболеваний можно секвенировать ген Notch3 из образцов пациентов, осуществить анализ FISH (флуоресцентной гибридизации in situ) и CGH (сравнительной геномной гибридизации) для поиска транслокации и амплификации гена с использованием клеток пациента или осуществить иммуногистохимию для проверки сверхэкспрессии рецептора Notch3 с использованием срезов ткани или опухоли пациента. Кроме того, можно выделить и культивировать клетки от пациента, у которого предполагают наличие связанного с Notch3 заболевания, и исследовать действие антагонистического антитела по настоящему изобретению на клеточную миграцию, инвазию, выживаемость и пролиферацию. Протоколы для анализа клеточной миграции и инвазии описаны в примере 7, и протокол для анализа апоптоза описан в примере 6. Для анализа клеточной пролиферации клетки, культивируемые из образцов пациента, высевают в 96-луночный планшет, покрытый и не покрытый лигандами Notch. Антагонистические антитела добавляют в начале культивирования. Количества клеток подсчитывают в конкретные моменты времени с использованием окрашивания трипановым синим. Анализ Notch3 с помощью FISH и CGH можно осуществить с использованием опубликованных протоколов Park, et al. (Cancer Res, 66:12 (2006)).
Специалисты в данной области примут во внимание или будут способны установить с использованием не более чем общепринятого набора экспериментов, многие эквиваленты описанных в настоящем документе конкретных вариантов осуществления изобретения. Предполагается, что такие эквиваленты охватываются следующей формулой изобретения.
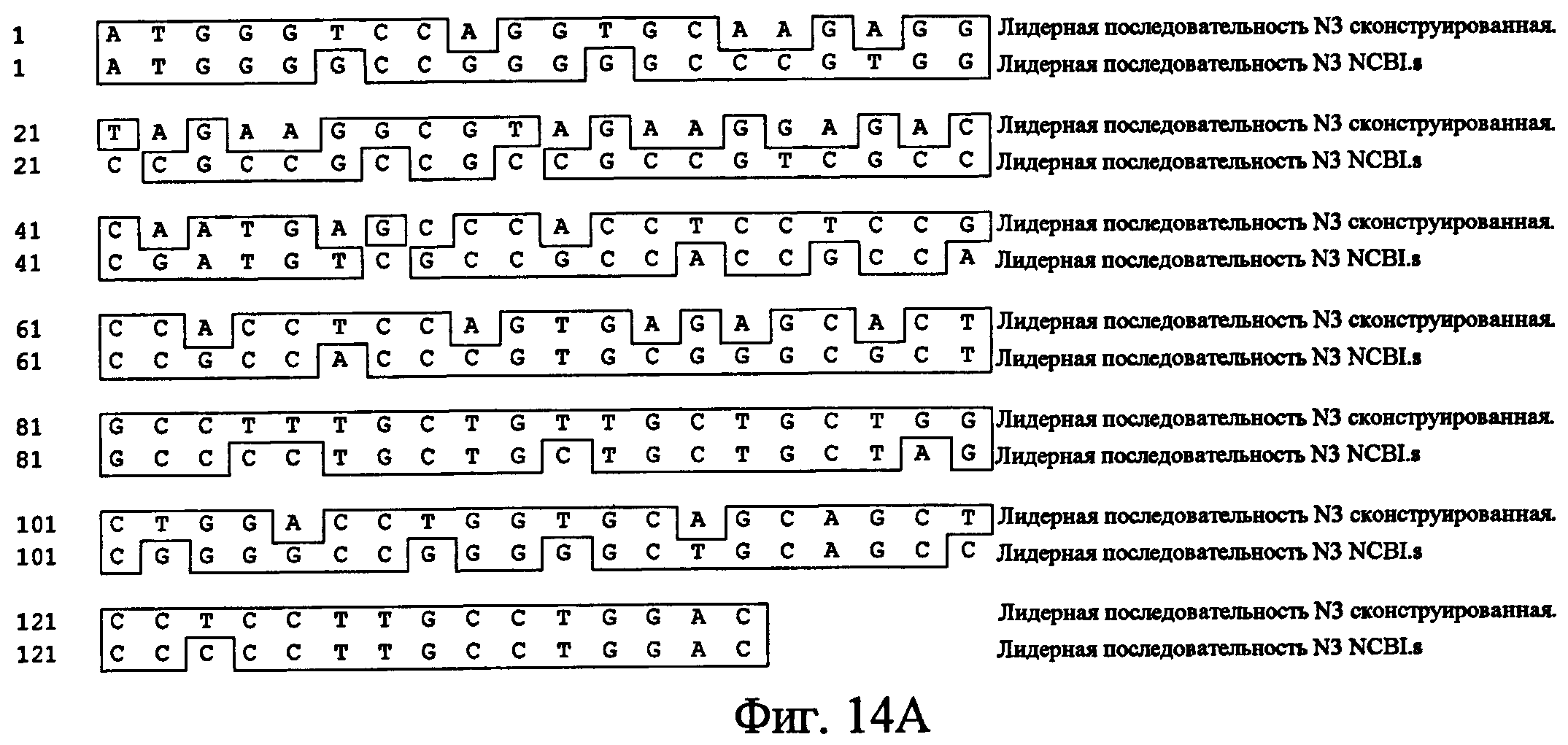


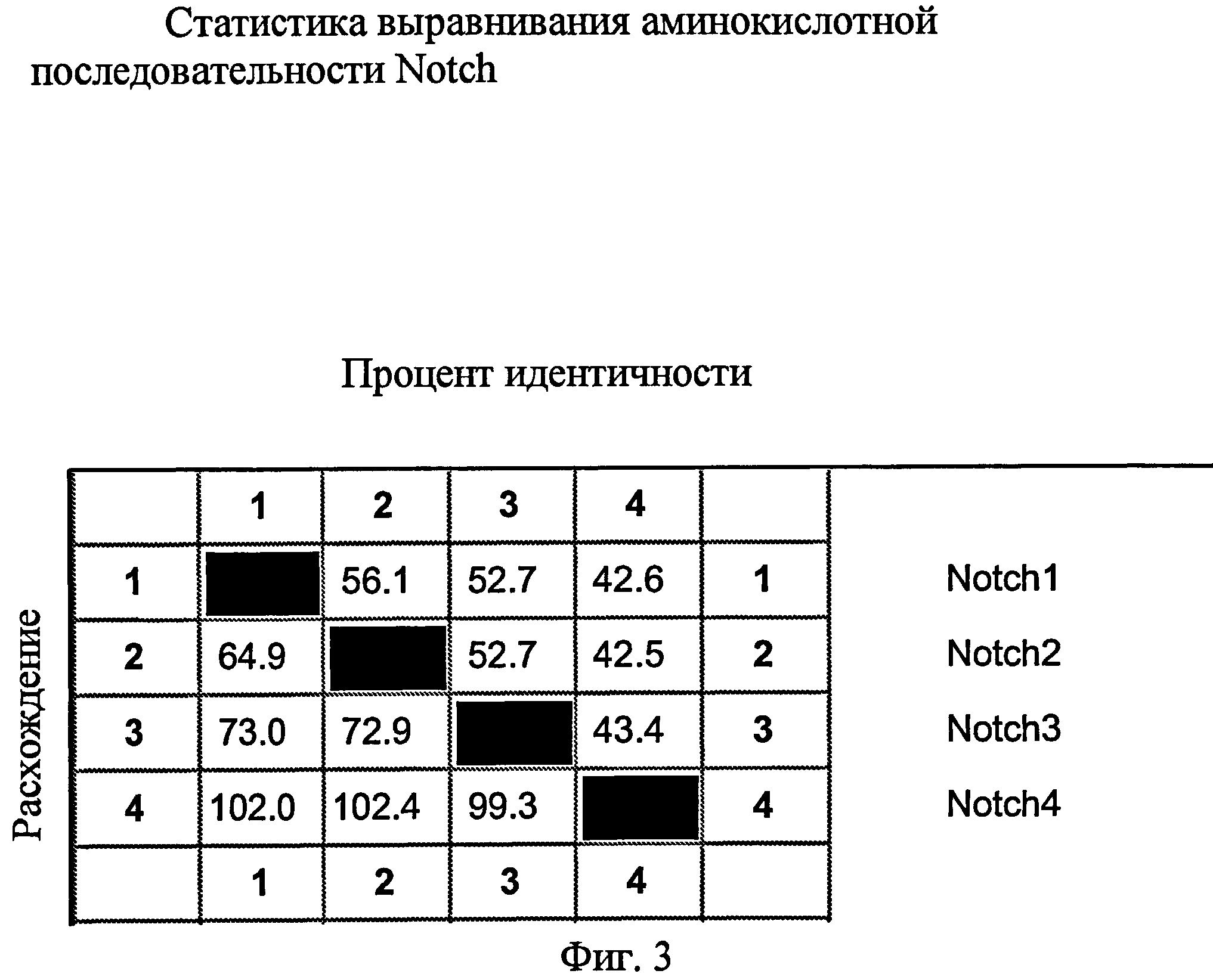
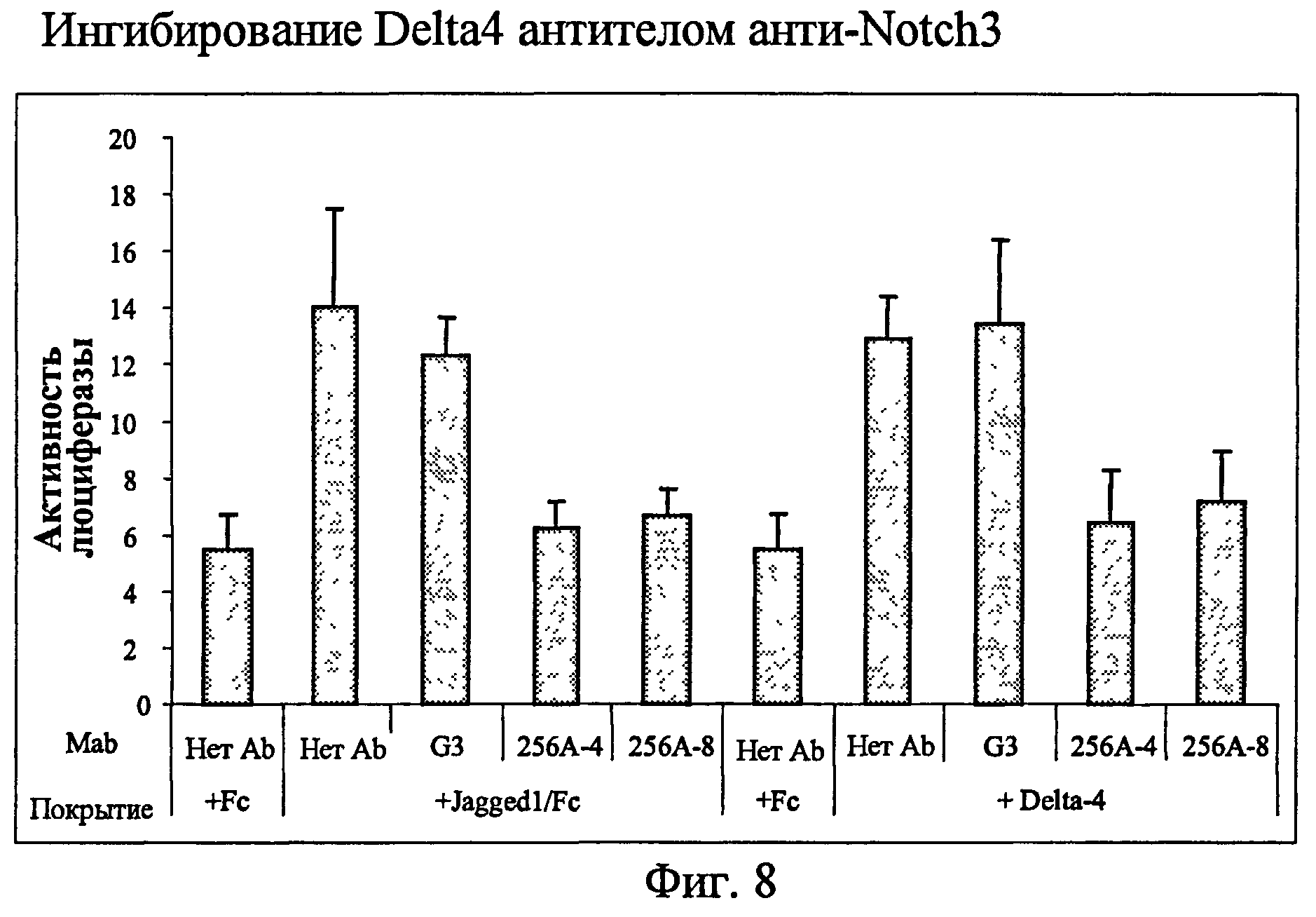





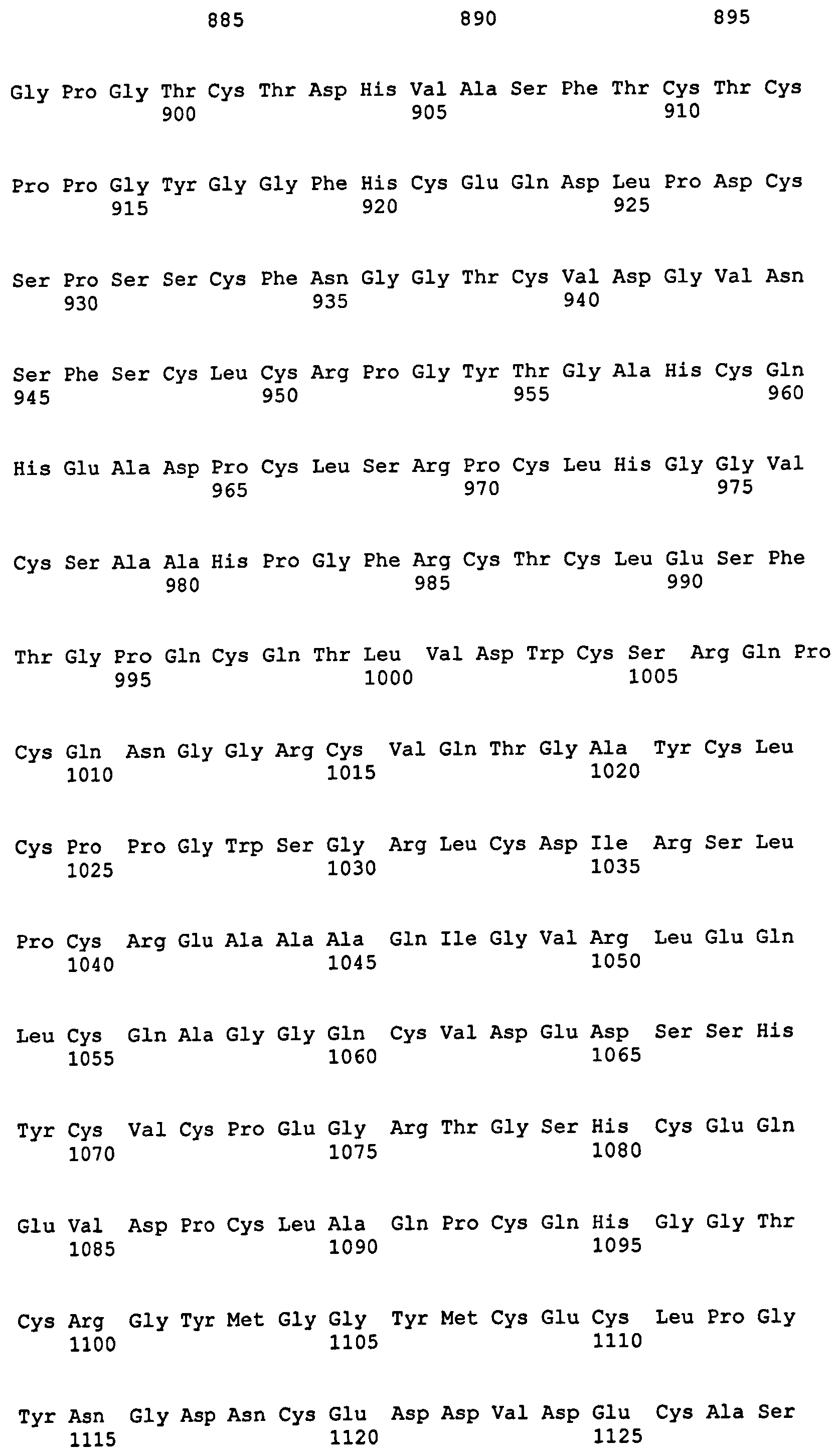

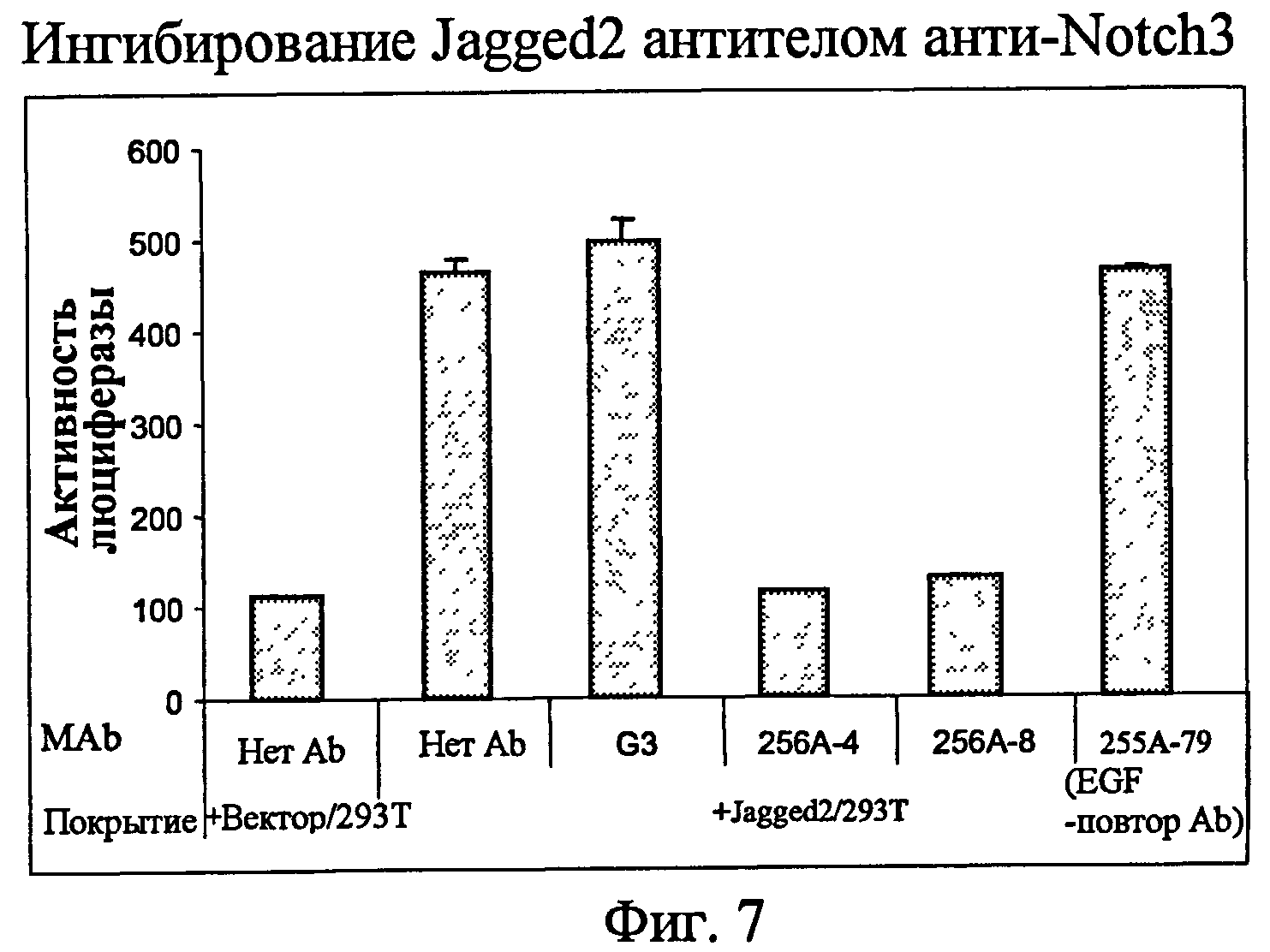

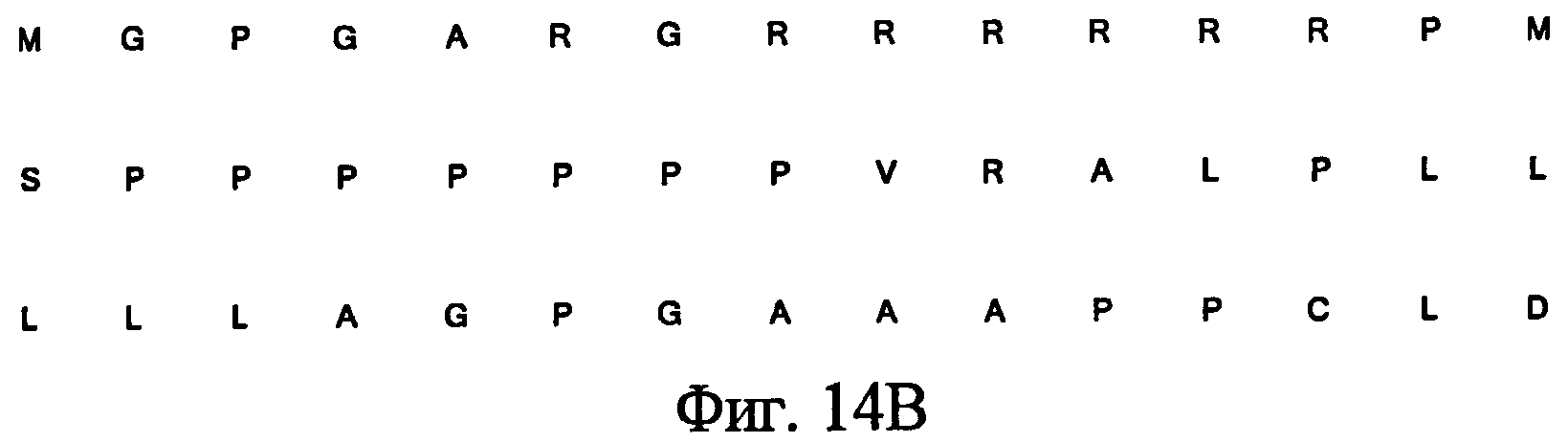


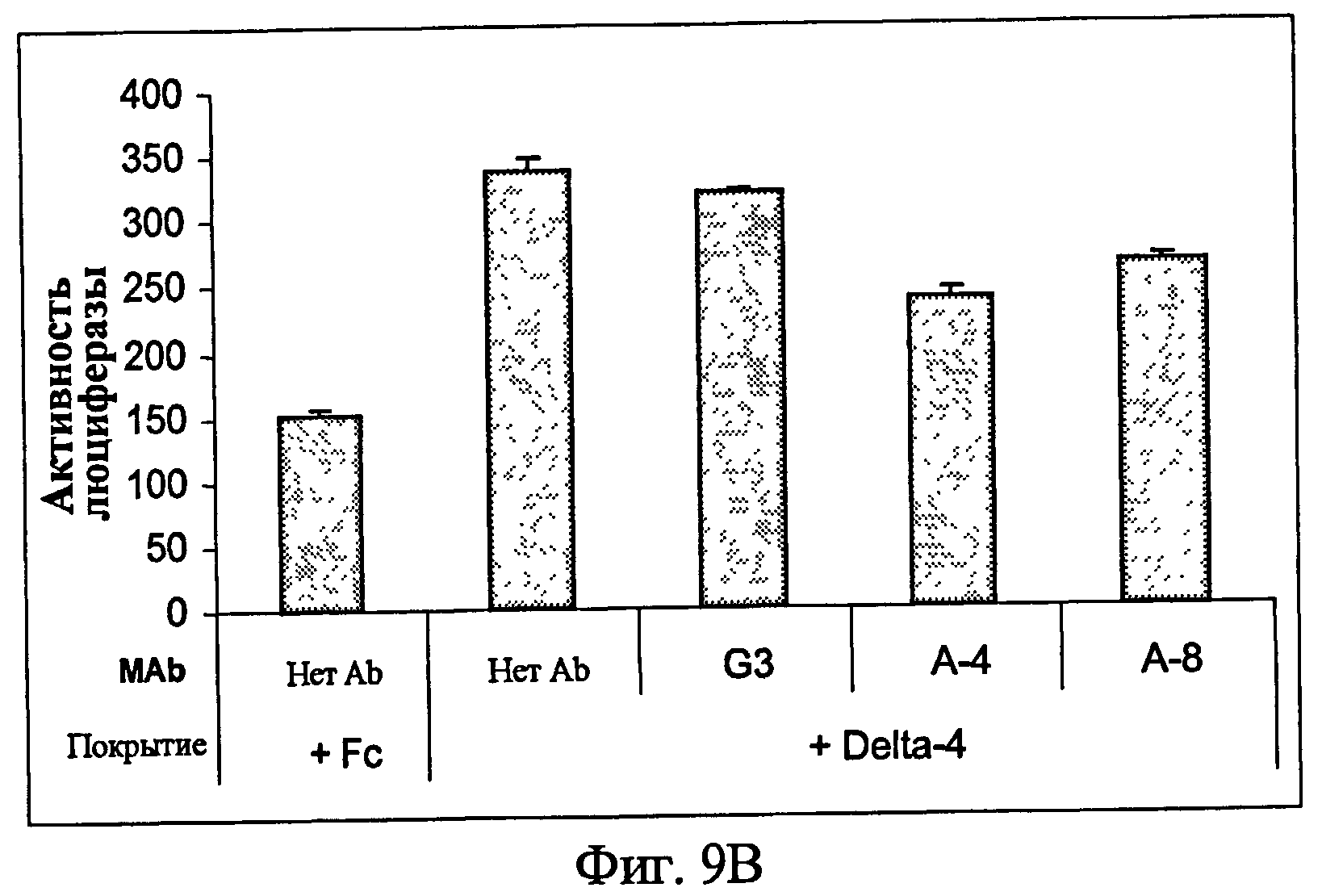

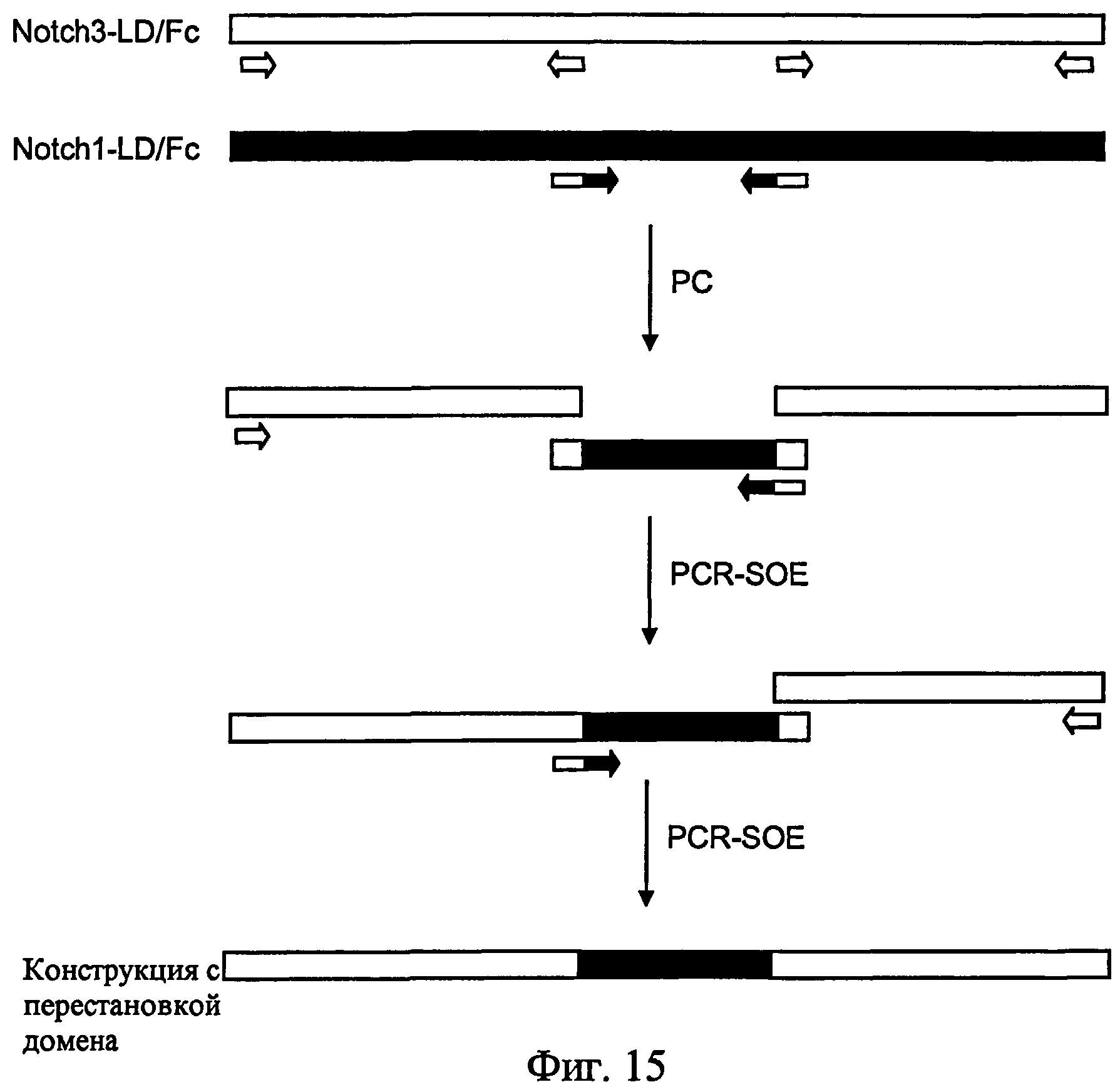
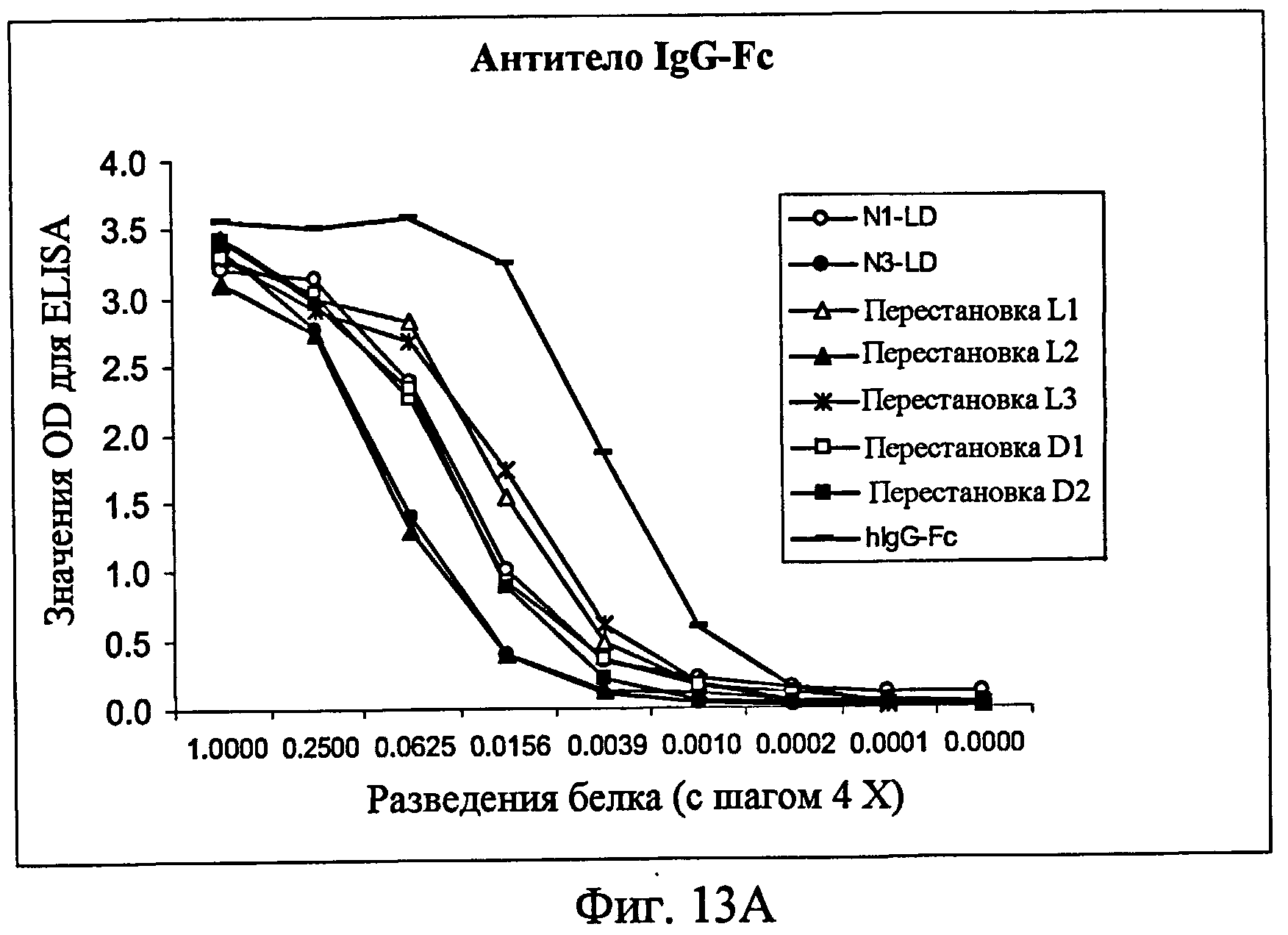






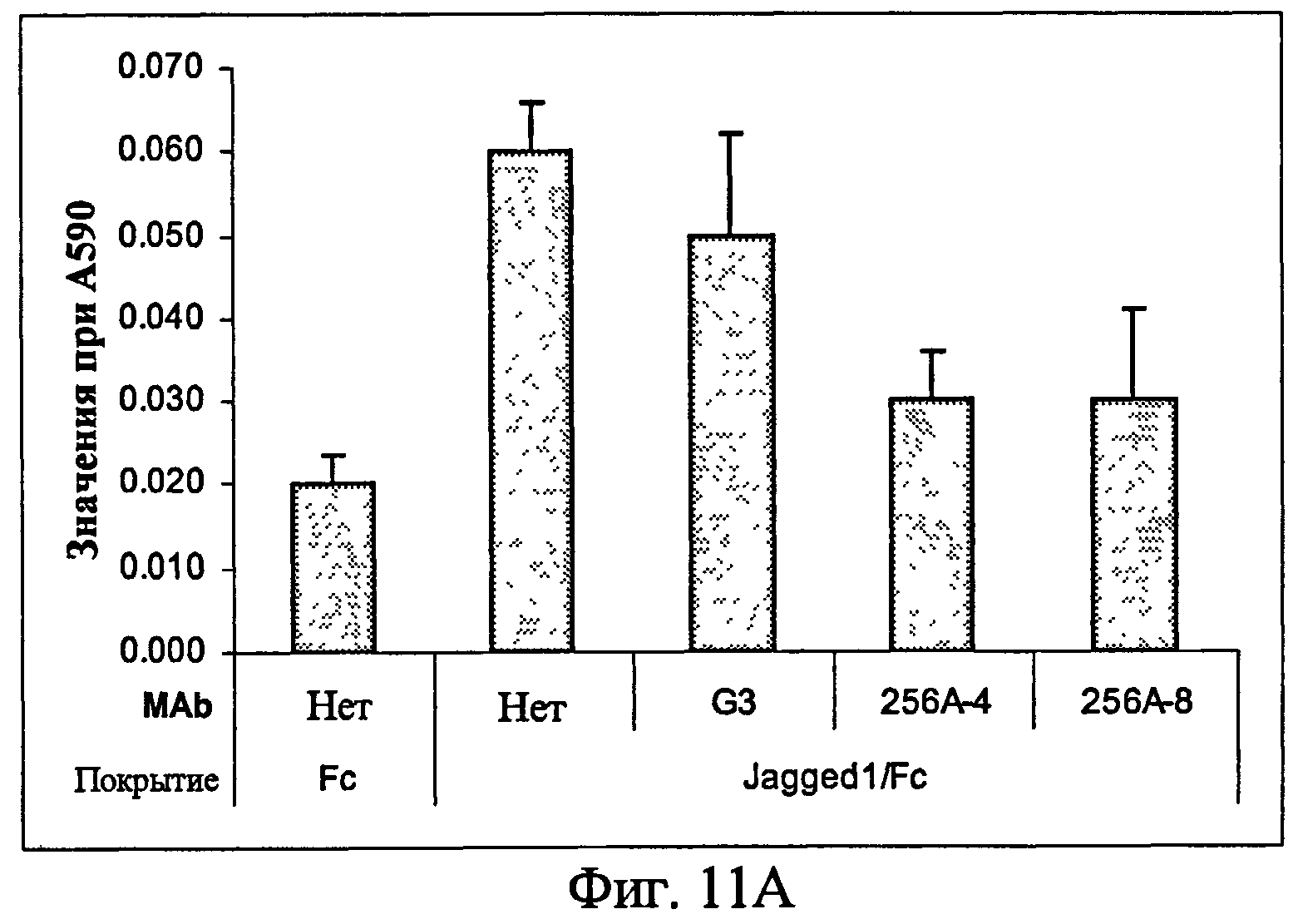
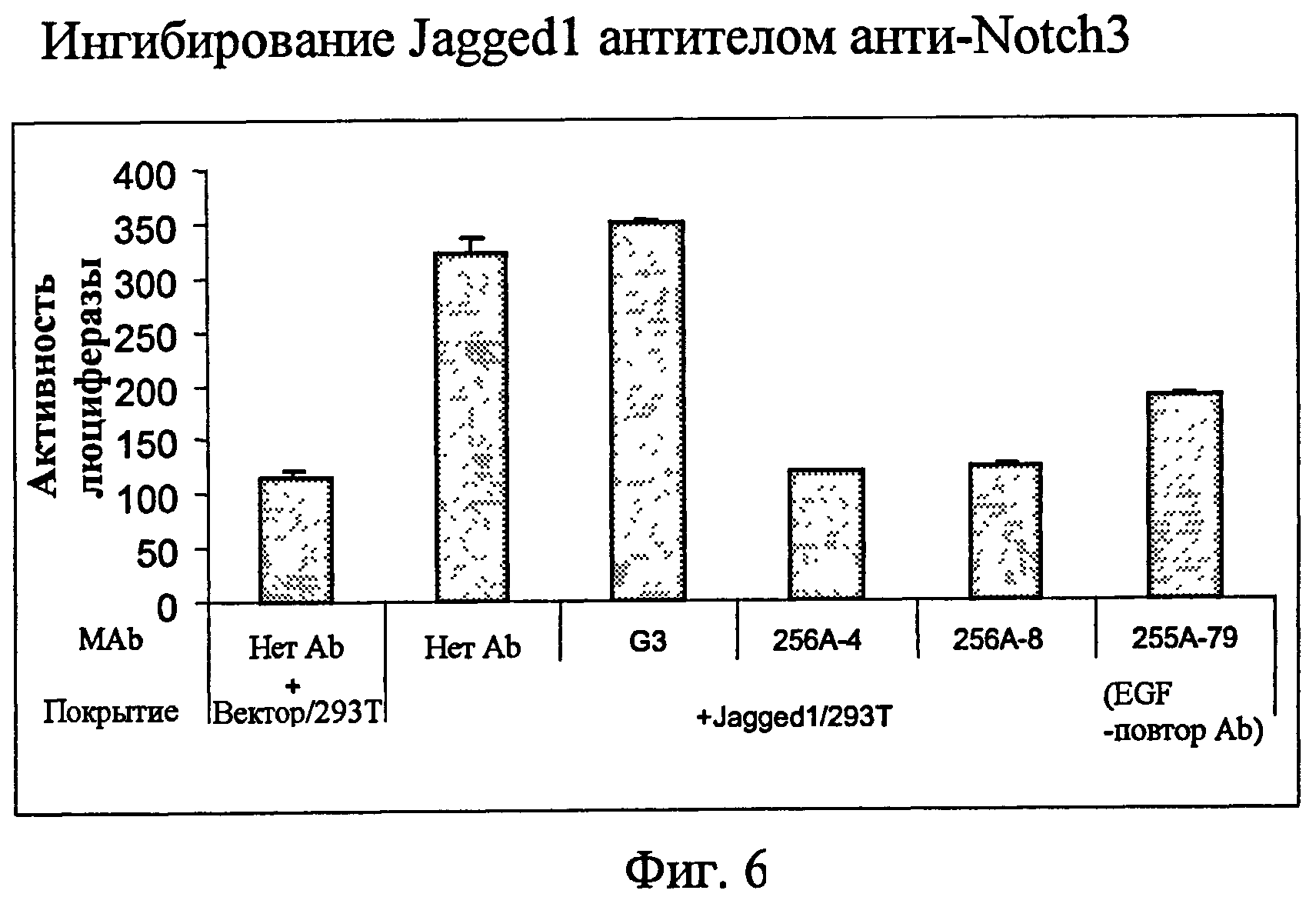
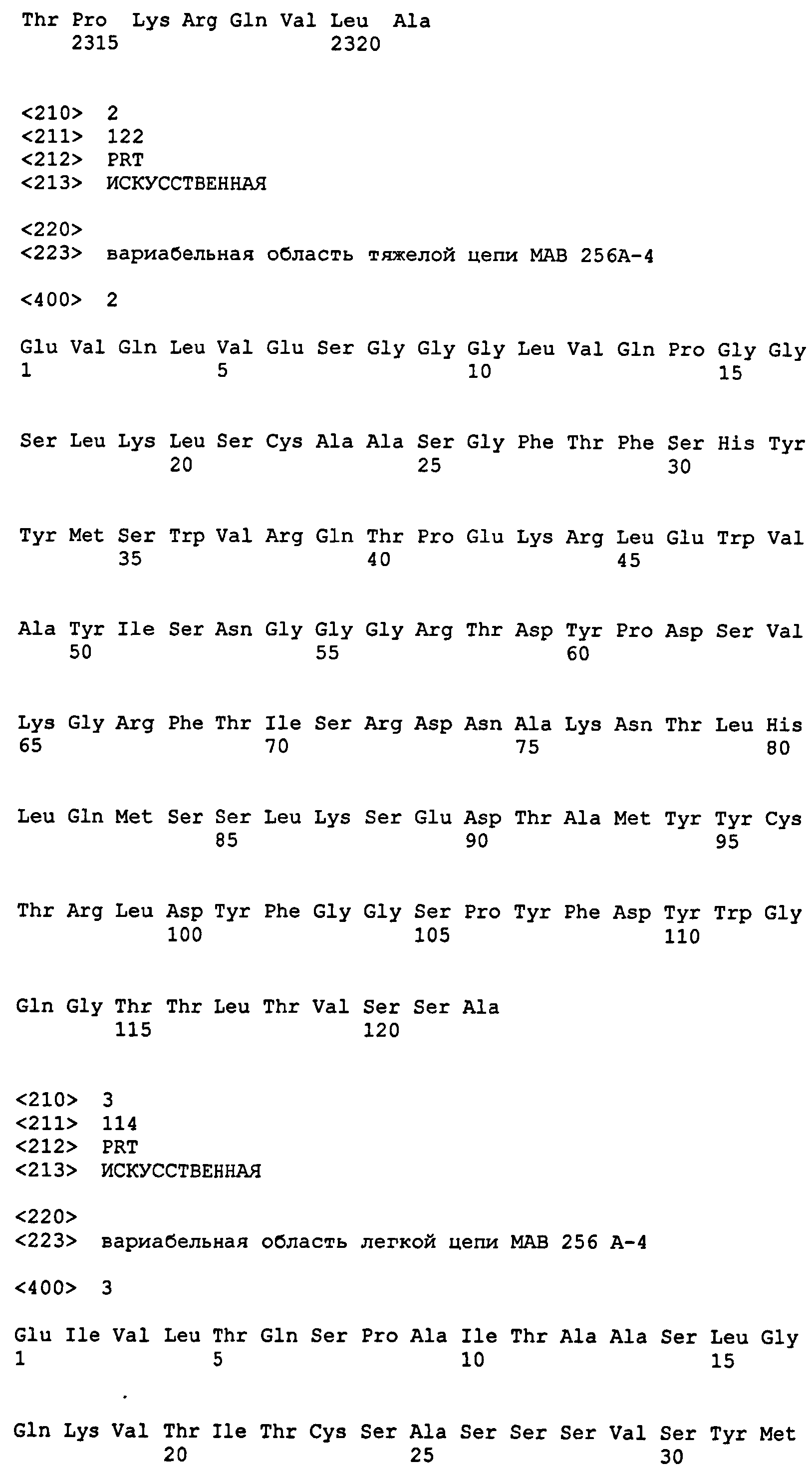
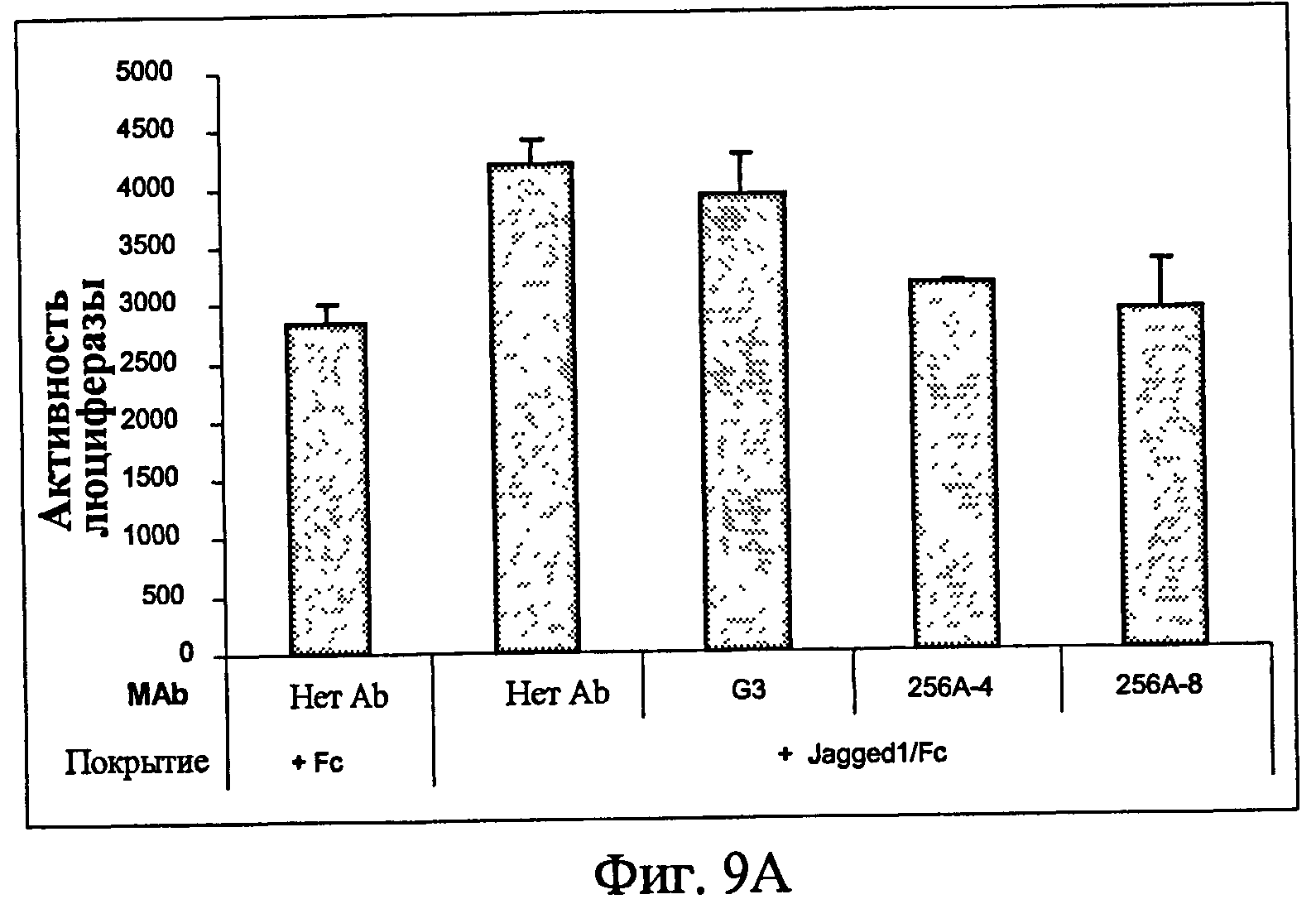







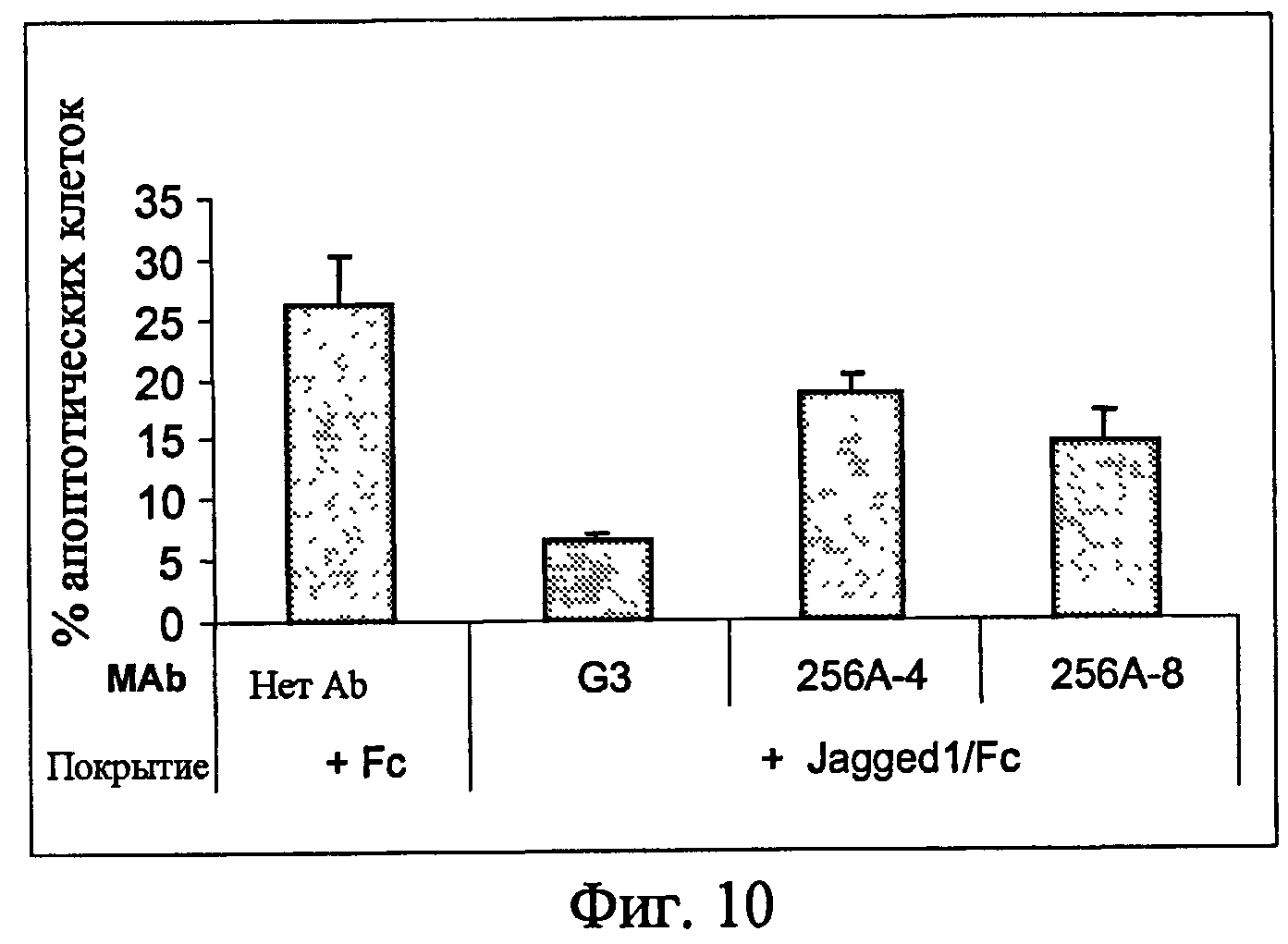
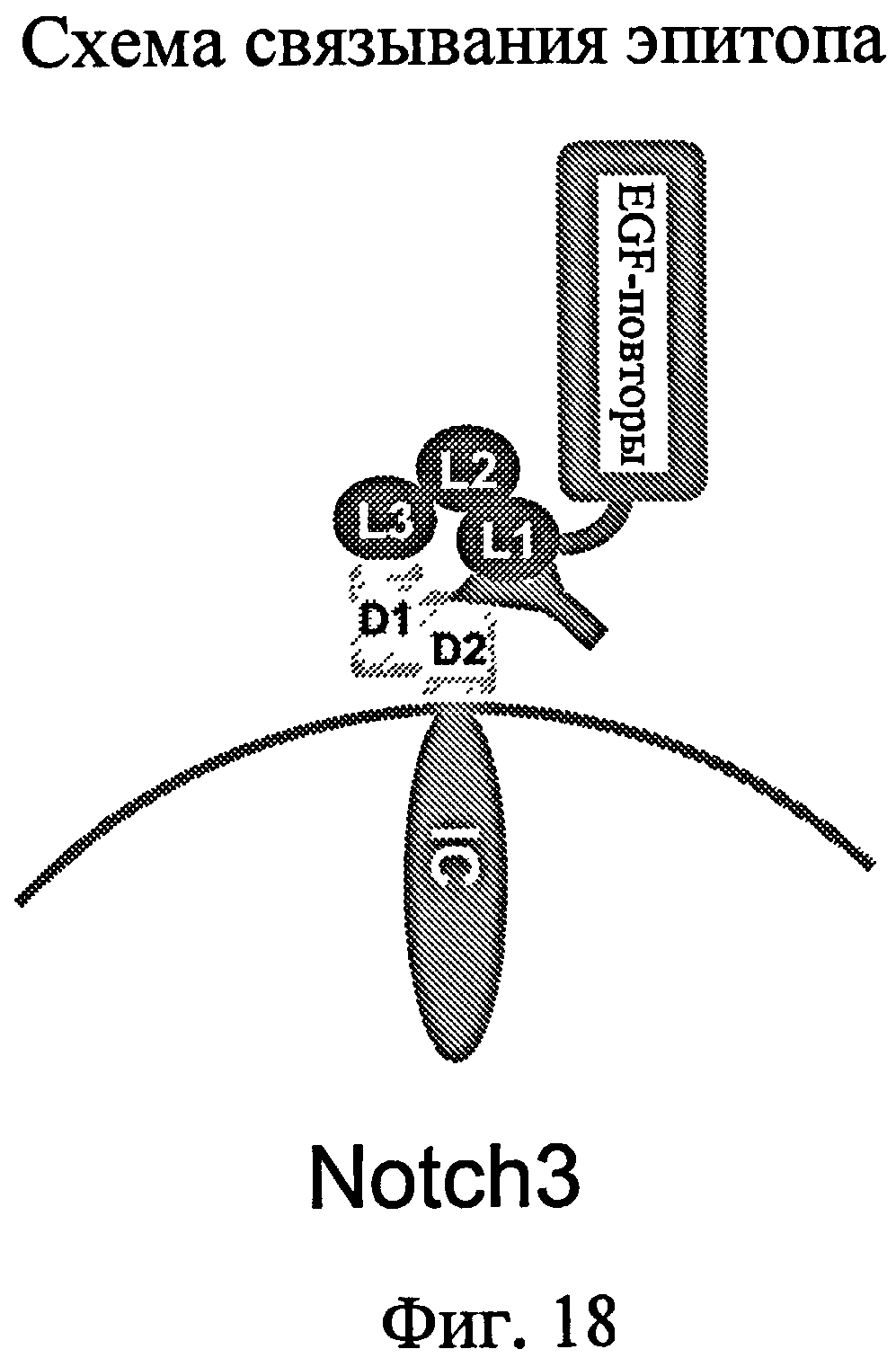
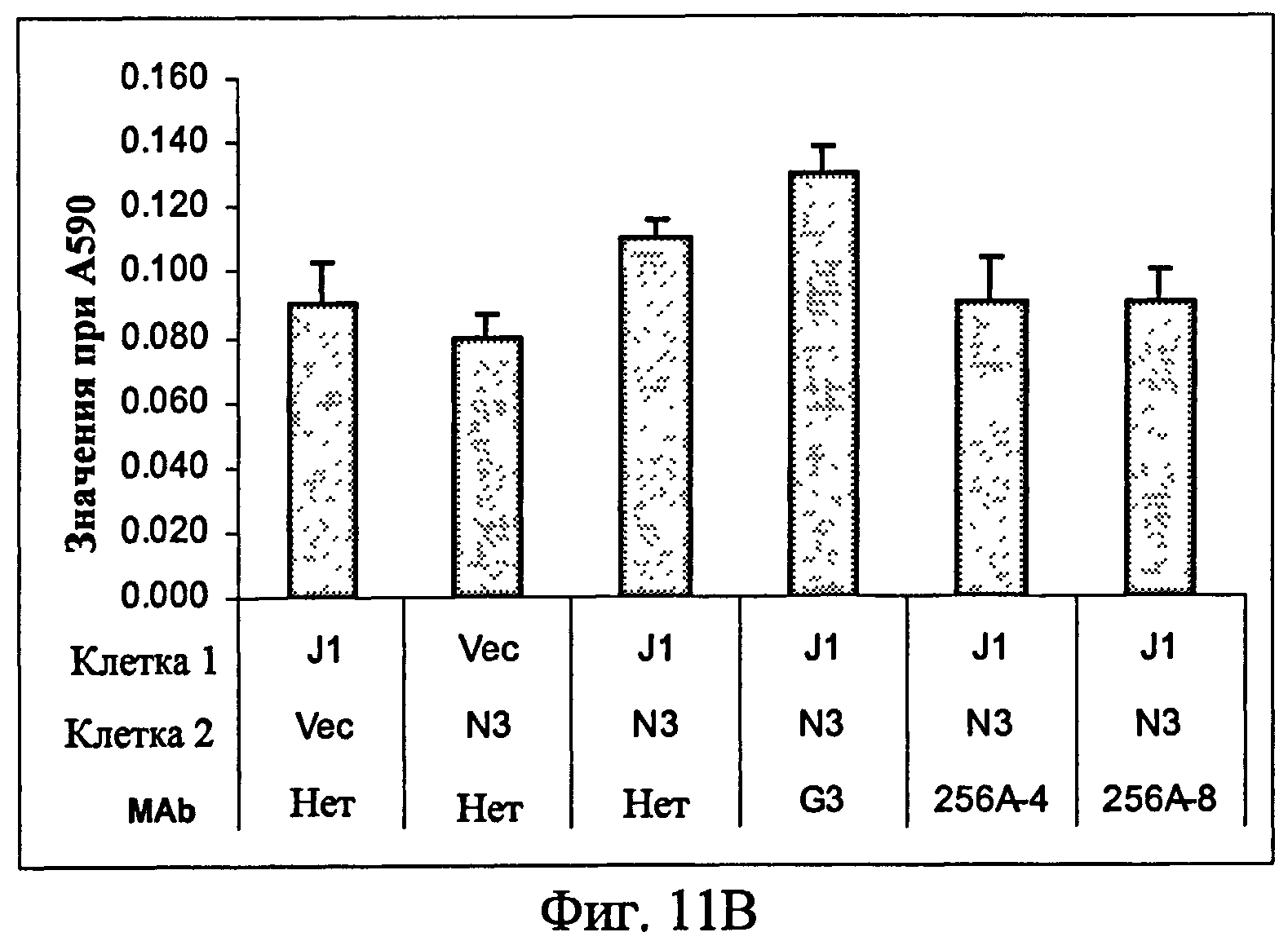



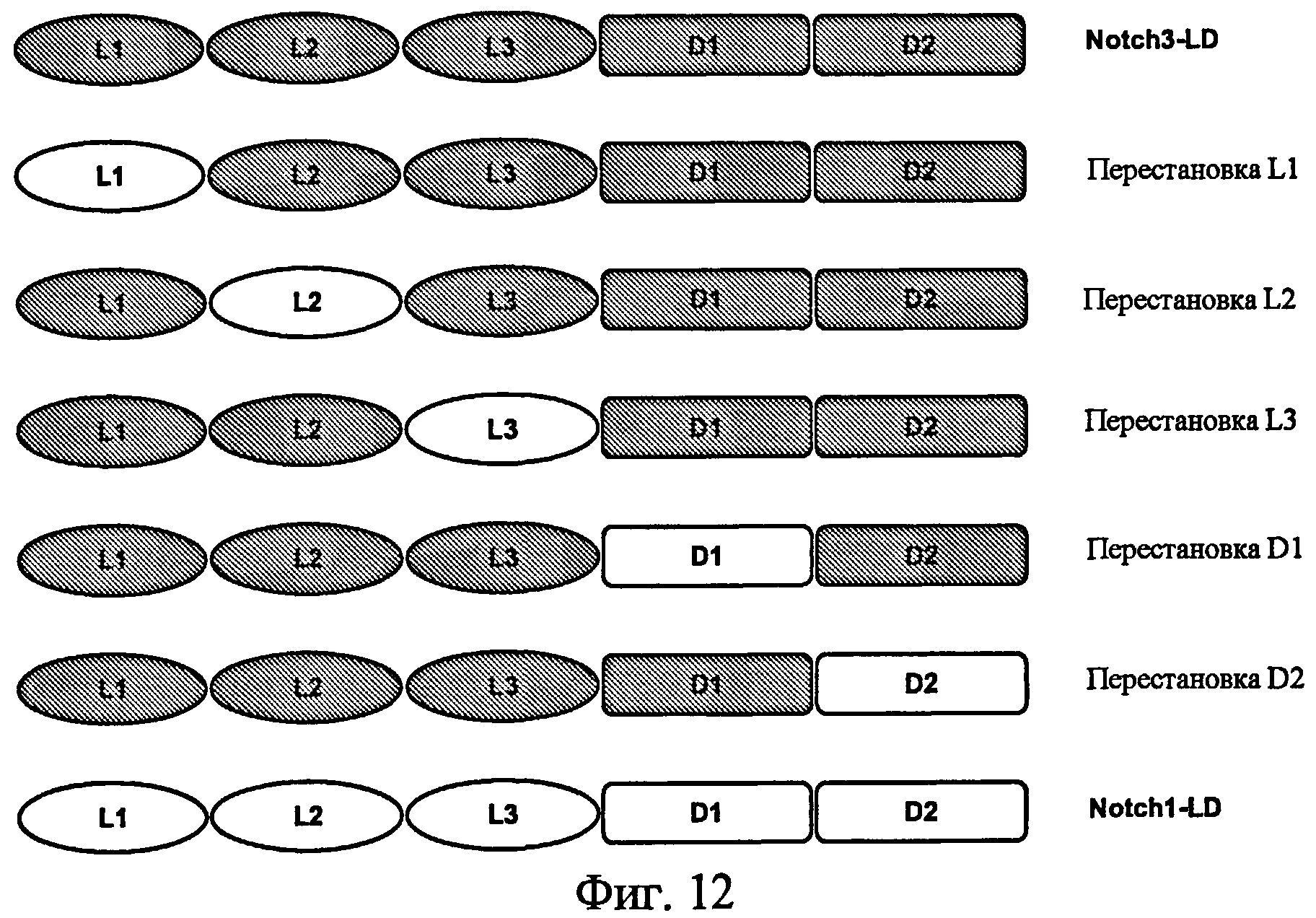


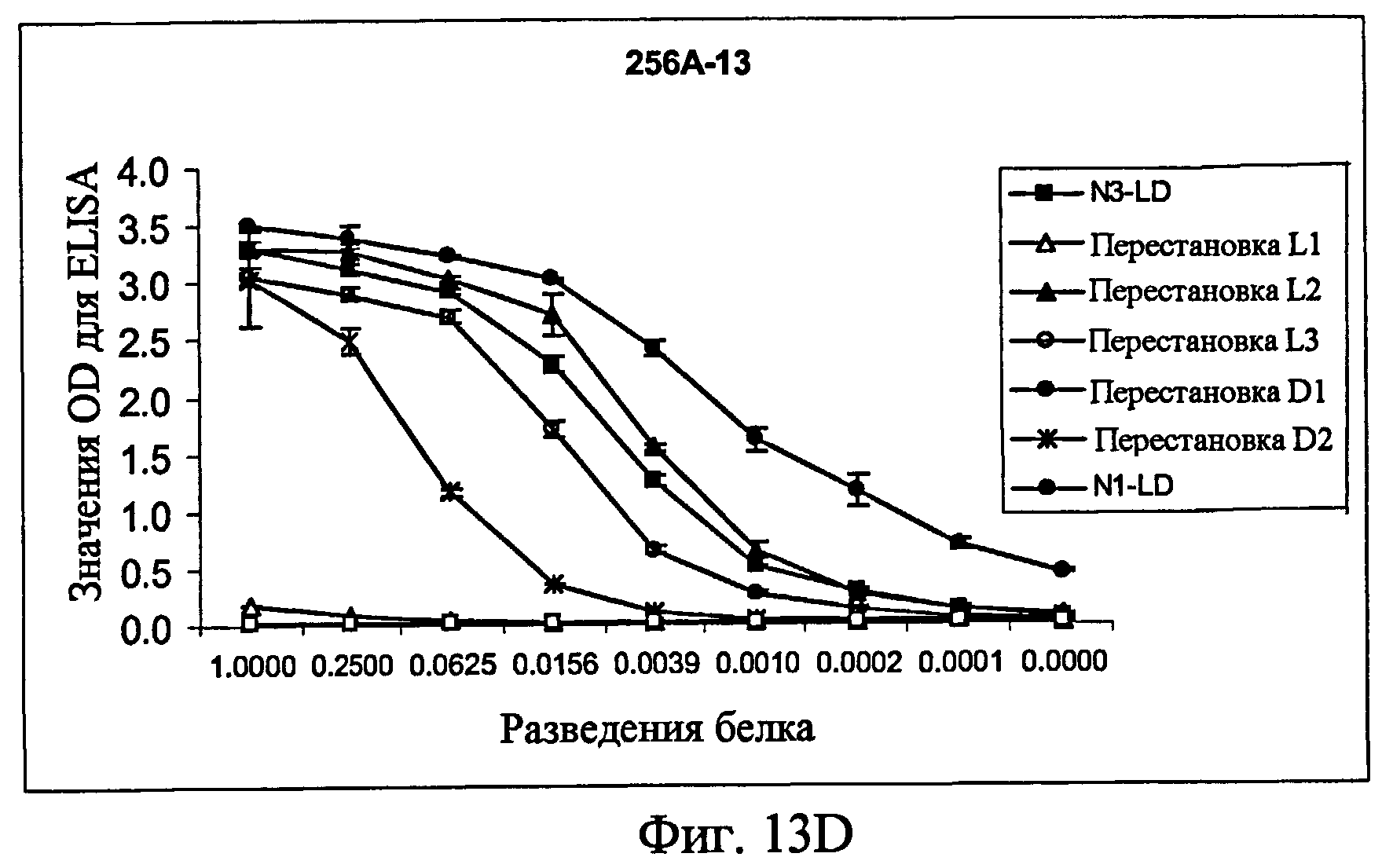
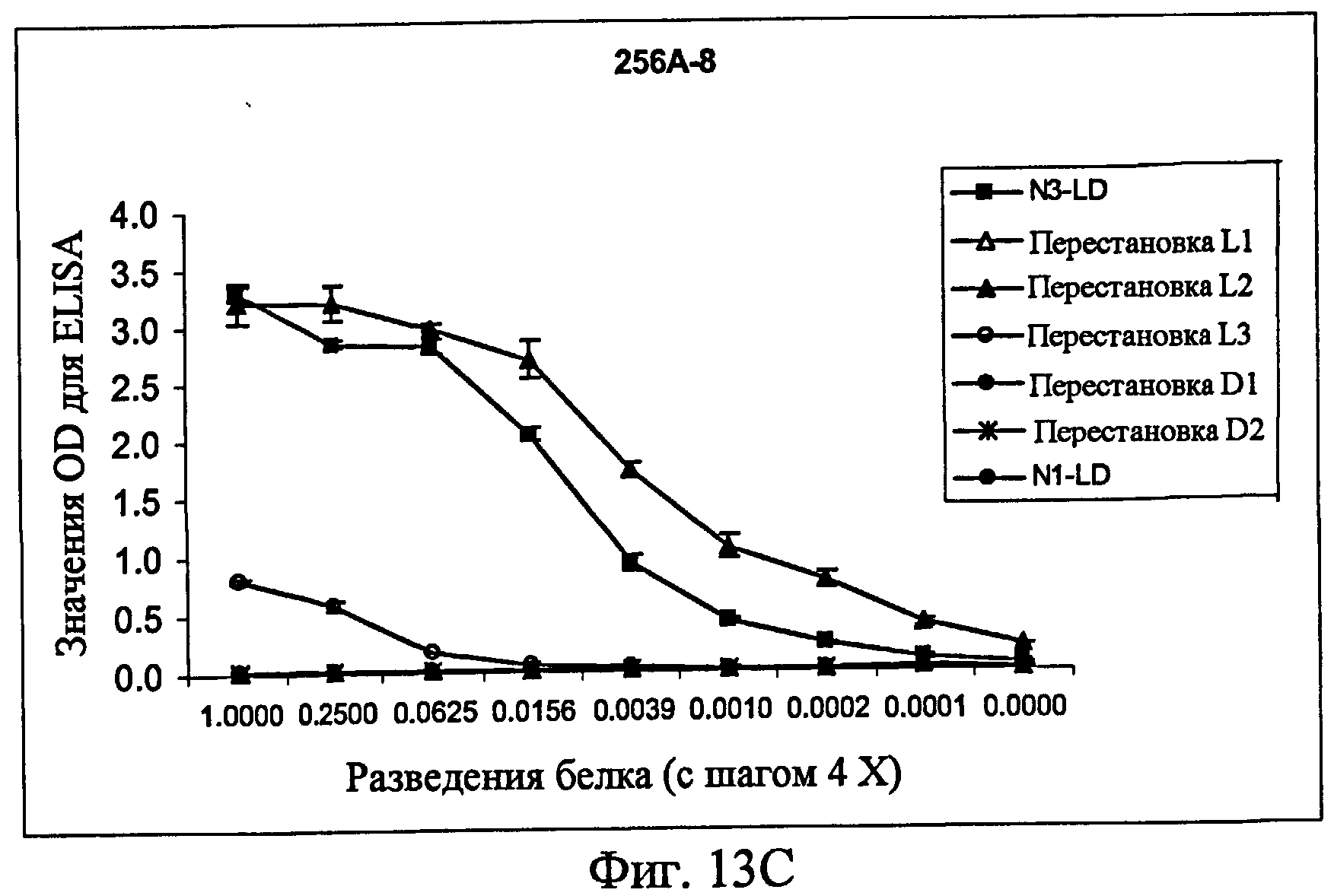
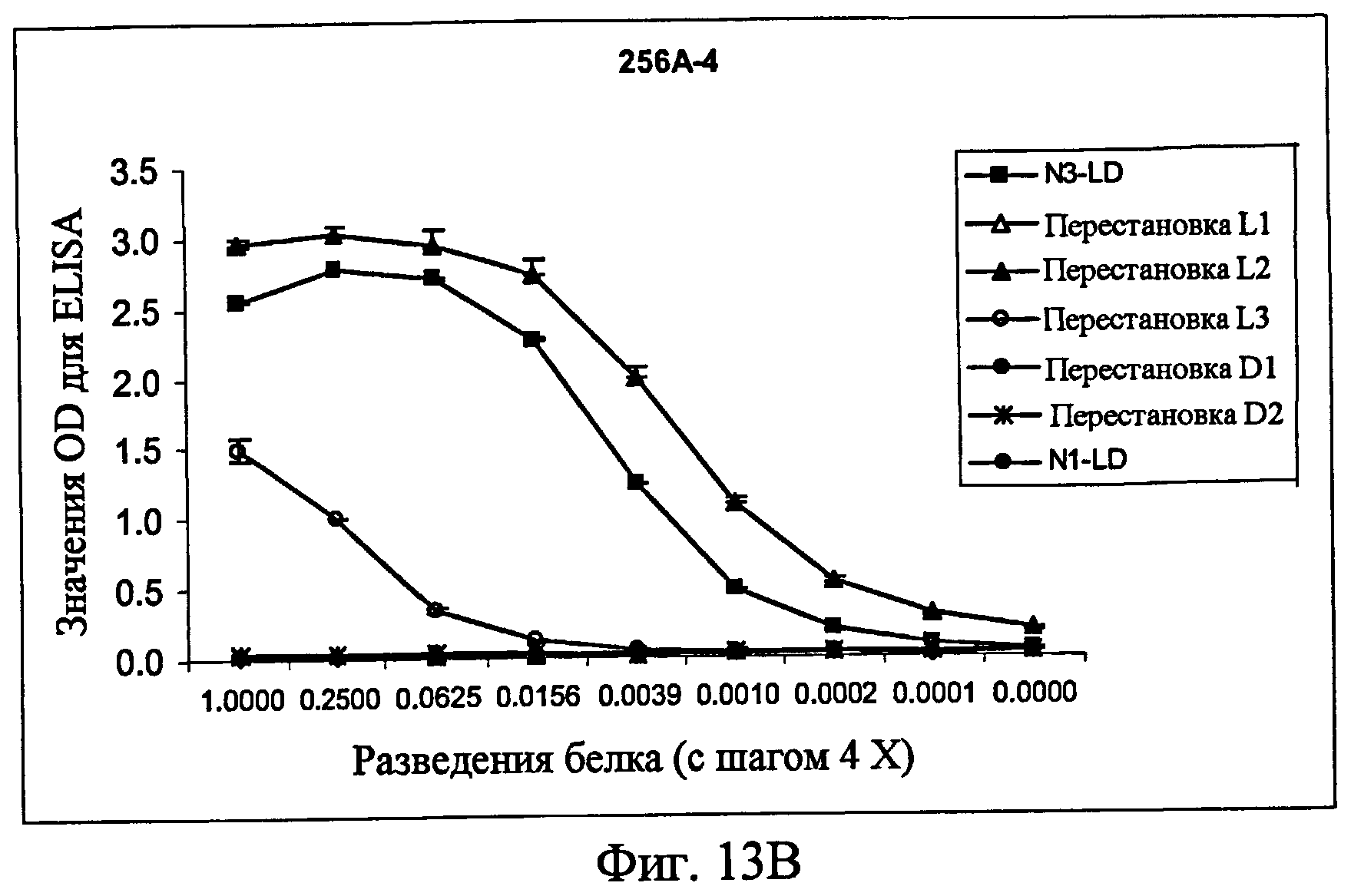

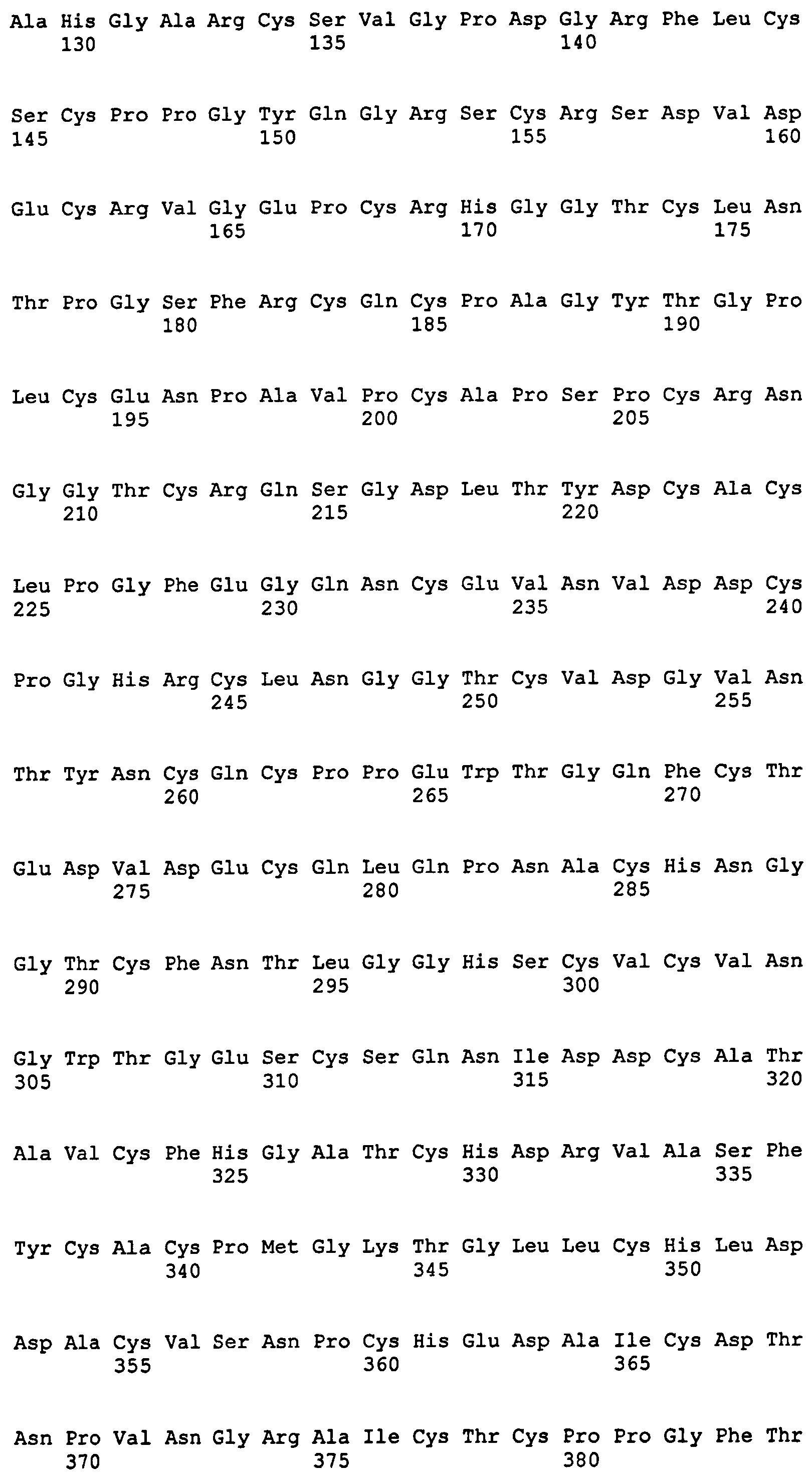




Ингибирование ангиогенеза
Пиримидиновые соединения, композиции и способы применения
Антитела против c3b и способы профилактики и лечения связанных с комплементом нарушений
Осаждение и очистка белков полиэлектролитами
Гуманизированные антитела к фактору d и их применения
Антитела против nrr notch1 и способы их применения
Гидроксилированные и метоксилированные циклопента[d]пиримидины в качестве ингибиторов акт протеинкиназ
Циклопента(d)пиримидины в качестве ингибиторов протеинкиназ акт
Vegf полиморфизм и анти-ангиогенезная терапия
Применение антитела против амилоида-бета при глазных заболеваниях
Ингибирование ангиогенеза
Пиримидиновые соединения, композиции и способы применения
Антитела против c3b и способы профилактики и лечения связанных с комплементом нарушений
Осаждение и очистка белков полиэлектролитами
Гуманизированные антитела к фактору d и их применения
Антитела против nrr notch1 и способы их применения
Гидроксилированные и метоксилированные циклопента[d]пиримидины в качестве ингибиторов акт протеинкиназ
Циклопента(d)пиримидины в качестве ингибиторов протеинкиназ акт
Vegf полиморфизм и анти-ангиогенезная терапия
Применение антитела против амилоида-бета при глазных заболеваниях

