Результат интеллектуальной деятельности: ВЫДЕЛЕННЫЕ ПОПУЛЯЦИИ МИЕЛОПОДОБНЫХ КЛЕТОК И СПОСОБЫ ЛЕЧЕНИЯ С ИСПОЛЬЗОВАНИЕМ ТАКИХ ПОПУЛЯЦИЙ
Вид РИД
Изобретение
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка является частичным продолжением международной заявки на патент № PCT/US2006/06411, поданной 24 февраля 2006 г., по которой испрашивается приоритет по предварительной патентной заявке США №60/656037, поданной 24 февраля 2005 г. и включенной в данное описание в качестве ссылки.
СПРАВКА О ГОСУДАРСТВЕННЫХ ВЛОЖЕНИЯХ
Часть описанной в данном документе работы была проведена при поддержке грантов EY11254, EY12598, EY13916 и EY14174, выданных Национальным институтом глаза, относящимся к Национальным институтам здравоохранения. Правительство Соединенных Штатов имеет определенные права на данное изобретение.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к выделенным клеткам млекопитающих. Более конкретно, данное изобретение относится к популяциям выделенных клеток, которые имеют характерные особенности миелоидных клеток и могут внедряться в сосудистую сеть сетчатки после введения в стекловидное тело глаза. Данное изобретение также относится к способам лечения дегенеративных заболеваний глаза путем введения миелоподобных клеток в глаз млекопитающего.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Возрастная дегенерация желтого пятна (ARMD) и диабетическая ретинопатия (DR), приводящие к аномальному образованию новых сосудов в сетчатке, являются основными причинами потери зрения у населения стран с развитой промышленностью. Поскольку сетчатка состоит из четко определенных слоев нейронных, глиальных и сосудистых элементов, относительно небольшие нарушения, такие как наблюдающиеся при пролиферации сосудов или отеке, могут приводить к значительной потере зрительной функции. Наследственные дегенеративные заболевания сетчатки, такие как пигментная дегенерация сетчатки (RP), также связаны с сосудистыми аномалиями, такими как сужение артериол и атрофия сосудов. Большинство наследственных дегенеративных заболеваний сетчатки человека специфически поражают палочковидные фоторецепторы, но также сопровождаются утратой колбочек, основным клеточным компонентом пятнышка, участка сетчатки у людей, который отвечает за центральную, тонкую остроту зрения. Недавно описаны колбочко-специфичные факторы выживания (Mohand-Said et al., 1998, Proc. Natl. Acad. Sci USA, 95: 8357-8362), которые могут облегчить выживание колбочек у мышиных моделей дегенерации сетчатки.
Наследственные дегенеративные заболевания сетчатки поражают до 1 субъекта из 3500 и характеризуются прогрессирующей ночной слепотой, уменьшением поля зрения, атрофией оптического нерва, утоньшением артериол, изменением проницаемости сосудов и утратой центрального зрения, часто прогрессирующей до полной слепоты (Heckenlively, J. R., editor, 1988; Retinitis Pigmentosa, Philadelphia: JB Lippincott Co.). Молекулярный генетический анализ данных заболеваний, проведенный для относительно небольшого числа известных пораженных субъектов, позволил идентифицировать мутации более чем в 110 различных генах (Humphries et al., 1992, Science 256:804-808; Farrar et al., 2002, EMBO J. 21:857-864). Многие из этих мутаций связаны с ферментными и структурными компонентами механизма фотопреобразования, включающими родопсин, фосфодиэстеразу цГМФ, периферин rds и RPE65. Несмотря на указанные наблюдения еще не существуют эффективные способы лечения, позволяющие замедлить или прекратить развитие этих дегенеративных заболеваний сетчатки. Последние достижения в генной терапии привели к успешному устранению фенотипов rds (Ali et al 2000, Nat. Genet. 25:306-310) и rd (Takahashi et al., 1999, J. Virol. 73:7812-7816) у мышей и фенотипа RPE65 у собак (Acland et al., 2001, Nat. Genet. 28:92-95), после доставки трансгена дикого типа к фоторецепторам или пигментированному эпителию сетчатки (RPE) животных, несущих специфическую мутацию.
Давно известно, что популяция стволовых клеток существует в кровотоке и костном мозге нормальных взрослых субъектов. Разные субпопуляции этих клеток могут дифференцироваться по гематопоэтической положительной (Lin+) или отрицательной (Lin-) линиям дифференцировки. Кроме того, недавно было показано, что популяция гематопоэтических стволовых клеток отрицательной линии дифференцировки (HSC) содержит предшественники эндотелиальных клеток (EPC), способные формировать кровеносные сосуды in vitro и in vivo (See Asahara et al. 1997, Science 275: 964-7). Данные клетки могут участвовать в нормальном и патологическом постнатальном ангиогенезе (см. Lyden et al., 2001 Nat. Med. 7, 1194-201; Kalka et al., 2000, Proc. Natl. Acad. Sci U.S.A. 97:3422-7; и Kocher et al., 2001, Nat. Med. 7: 430-6), а также дифференцироваться в ряд неэндотелиальных клеточных типов, включающих гепатоциты (см. Lagasse et al., 2000, Nat. Med. 6:1229-34), микроглию (см. Priller et al. 2002 Nat. Med. 7:1356-61), кардиомиоциты (см. Orlic et al., 2001, Proc. Natl. Acad. ScL U.S.A. 98: 10344-9) и эпителиальные клетки (см. Lyden et al., 2001, Nat. Med. 7: 1194-1201). Хотя такие клетки используют в нескольких экспериментальных моделях ангиогенеза, механизм направления EPC к участку образования новых сосудов неизвестен, и не разработана стратегия, позволяющая эффективно увеличивать число клеток, которые вносят вклад в конкретную сосудистую сеть.
Гематопоэтические стволовые клетки костного мозга в настоящее время являются единственным типом стволовых клеток, широко используемым для терапевтического применения. HSC костного мозга используют для трансплантации в течение 40 лет. В настоящее время исследуется возможность применения усовершенствованных методов сбора очищенных стволовых клеток для разработки способов лечения лейкоза, лимфомы и наследственных заболеваний крови. На ограниченном числе пациентов-людей исследуется возможность клинического применения стволовых клеток для лечения диабета и запущенного рака почки у человека.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение предлагает популяцию миелоподобных клеток, полученную путем положительной селекции клеток, экспрессирующих CD44, CD11b и индуцируемый гипоксией фактор 1α (HIF-1α), выделенную из костного мозга млекопитающих. Такие клетки обладают полезной сосудисто-трофической и нейротрофической активностью при внутриглазном введении млекопитающему, в частности млекопитающему, страдающему дегенеративным заболеванием глаза. Популяцию миелоподобных клеток по данному изобретению можно выделить путем обработки клеток костного мозга (например, клеток костного мозга человека) антителом против CD44 (рецептор гиалуроновой кислоты), антителом против CD11b, антителом против CD33, CD14, или сочетанием антител против указанных антигенов, причем положительно отбираются клетки, которые способны к иммунному взаимодействию с антителом или антителами, в зависимости от обстоятельств (для разделения клеток используют, например, проточную цитометрию или гранулы, покрытые антителами, или связанные с антителами). Такие клетки, полученные из костного мозга, в данном описании называют миелоподобные клетки костного мозга (MLBM). Альтернативно, популяции миелоподобных клеток по данному изобретению можно выделить из периферической крови (например, из периферической крови человека) или из крови пуповины (например, из крови пуповины человека). Большая часть клеток из популяции миелоподобных клеток по данному изобретению экспрессирует антиген CD44, антиген CD11b и индуцируемый гипоксией фактор 1α (HIF-1α).
Настоящее изобретение также предлагает способ лечения сосудисто-трофических и нейротрофических заболеваний сетчатки у млекопитающего. Способ включает введение выделенных клеток из популяции миелоподобных клеток в больной глаз млекопитающего, предпочтительно, путем внутриглазной инъекции. Предпочтительно, популяция миелоподобных клеток является аутологичной по отношению к млекопитающему, подлежащему лечению (т.е., популяцию миелоподобных клеток выделяют из костного мозга, периферической крови или крови пуповины млекопитающего, подлежащего лечению). Способ лечения по настоящему изобретению уменьшает дегенерацию сосудов и фоторецепторных нейронов в сетчатке млекопитающего, страдающего заболеванием глаза. Клетки вводят в количестве, достаточном для замедления сосудистой и невральной дегенерации сетчатки. Благоприятный эффект оказывает внедрение клеток из популяции миелоподобных клеток в сосудистую сеть сетчатки, хотя в то же время они внедряются в нейронную сеть, уменьшая дегенерацию колбочек в сетчатке. Выделенная популяция миелоподобных клеток млекопитающих содержит клетки, которые селективно воздействуют на активированные астроциты сетчатки при введении в стекловидное тело глаза и остаются стабильно внедренными в сеть новообразованных сосудов и нейронную сеть глаза. Предпочтительно, млекопитающее представляет собой человека.
В предпочтительном воплощении, по меньшей мере, приблизительно 75 процентов клеток выделенной популяции миелоподобных клеток экспрессирует CD44, более предпочтительно, по меньшей мере, приблизительно 90 процентов.
В одном предпочтительном воплощении клетки из популяции миелоподобных клеток трансфицируют терапевтически полезным геном. Например, в способе клеточной генотерапии клетки можно трансфицировать полинуклеотидами, функционально кодирующими нейротрофические средства или антиангиогенные средства, которые селективно воздействуют на новообразованную сосудистую сеть и ингибируют образование новых сосудов, не влияя на уже существующие сосуды. В одном воплощении выделенная популяция миелоподобных клеток по данному изобретению включает ген, который кодирует пептид, ингибирующий ангиогенез. Ангиогенез-ингибирующие клетки из популяции миелоподобных клеток можно использовать для модуляции аномального роста кровеносных сосудов при таких заболеваниях, как ARMD, DR и некоторые дегенеративные заболевания сетчатки, связанные с аномальной сосудистой сетью. В другом предпочтительном воплощении выделенные клетки из популяции миелоподобных клеток по настоящему изобретению трансфицируют геном, кодирующим нейротрофический пептид. Нейротрофические трансфицированные миелоподобные клетки можно использовать для спасения нейронов при глазных заболеваниях, включающих нейронную дегенерацию сетчатки, таких как глаукома, пигментная дегенерация сетчатки и т.п.
Конкретным преимуществом способов лечения глазных заболеваний выделенной популяцией миелоподобных клеток по настоящему изобретению является эффект сосудисто-трофического и нейротрофического спасения, наблюдающийся в глазах, обработанных через стекловидное тело клетками из популяции миелоподобных клеток. Нейроны и фоторецепторы сетчатки, в особенности колбочки, сохраняются, и в глазах, обработанных клетками из популяции миелоподобных клеток по данному изобретению, в некоторой степени сохраняется зрительная функция.
Настоящее изобретение также предлагает способ выделения популяции миелоподобных клеток из костного мозга путем негативной селекции по клеточному маркеру. Данный способ включает контактирование совокупности клеток костного мозга с антителами, специфичными к Ter119, CD45RB220 и CD3e, удаление из совокупности клеток костного мозга клеток, способных к иммунному взаимодействию с антителами против Ter119, CD45RB220 и CD3e, и выделение миелоподобных клеток костного мозга, которые не содержат клеток, экспрессирующих Ter119, CD45RB220 и CD3e. С помощью данного способа можно выделить популяцию клеток, в которой более 90 процентов клеток экспрессируют CD44.
Предпочтительно, если больная сетчатка, подлежащая лечению с помощью популяции миелоподобных клеток и способов по данному изобретению, содержит активированные астроциты. Этого можно достичь путем ранней обработки глаза, когда присутствует ассоциированный глиоз, или путем стимуляции локальной пролиферации активированных астроцитов с помощью лазера.
Помимо терапевтического применения выделенные популяции миелоподобных клеток костного мозга по данному изобретению можно использовать в качестве инструментов исследования физиологии развития сосудов глаза, а также для доставки конкретных генов к конкретным участкам (например, к астроцитам) глаза. Такое применение является ценным инструментом для исследования функции гена и потенциальных терапевтических механизмов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 изображены схематические диаграммы развития сетчатки мыши. (a) Развитие первичного сплетения. (b) Вторая фаза образования сосудов сетчатки. GCL - слой ганглионарных клеток; IPL - внутренний плексиформный (сплетениевидный) слой; INL - внутренний нуклеарный слой; OPL - наружный плексиформный слой; ONL - наружный нуклеарный (зернистый) слой; RPE - пигментный эпителий сетчатки; ON - зрительный нерв; P - периферия. В разделе (c) приведены результаты анализа методом проточной цитометрии разделенных клеток костного мозга Lin+HSC и Lin-HSC. Верхний ряд: гистограмма распределения немеченных антителом клеток, где R1 обозначает область измеримого селектируемого положительного PE-окрашивания; R2 обозначает GFP-положительную область. Средний ряд: клетки Lin-HSC (C57B/6); и нижний ряд: клетки Lin+HSC (C57B/6), где все клеточные линии помечены PE-конъюгированными антителами против Sca-1, c-kit, Flk-1/KDR, CD31. Данные Tie-2 получают с использованием мышей Tie-2-GFP. Проценты указывают долю меченных клеток во всей популяции Lin-HSC или Lin+HSC.
Фиг.2 иллюстрирует приживление Lin-HSC на развивающейся сетчатке мыши. (a) Через четыре дня после введения (P6) в стекловидное тело клетки eGFP+ Lin-HSC присоединяются к сетчатке и дифференцируются. (b) Lin-HSC (мыши B6.129S7-Gtrosa26, окрашенные антителом против β-gal) размещаются впереди сосудистой сети, окрашенной антителом против коллагена IV (звездочка указывает на оконечность сосудистой сети). (c) Большая часть клеток Lin+HSC (eGFP+) через четыре дня после инъекции (P6) не способна дифференцироваться. (d) Мезентериальные мышиные eGFP+ EC через четыре дня после инъекции (P6). (e) Lin-HSC (eGFP+), введенные в глаза взрослых мышей. (f) Небольшое накопление eGFP+ Lin-HSC (стрелки), перемещающихся к существующей астроцитарной матрице трансгенной мыши GFAP-GFP и дифференцирующихся по данному пути. (g) Повышенное накопление ассоциации клеток Lin- (eGFP) и исходных астроцитов (стрелки). (h) Трансгенный контроль, не получающий GFAP-GFP. (I) Через четыре дня после инъекции (P6) eGFP+ Lin-HSC мигрируют к области будущего глубокого сплетения и подвергаются дифференциации. В левой части показана активность Lin-HSC в целой закрепленной сетчатке; в правой части показано расположение клеток Lin- (стрелки) в сетчатке (вверху сторона, обращенная к стекловидному телу, внизу сторона, обращенная к склере), (j) Двойное мечение антителами против α-CD31-PE и α-GFP-alexa 488. Через семь дней после введения клетки Lin-HSC (eGFP, красный) внедряют в сосудистую сеть (CD31). Стрелки указывают участки внедрения. (k) Клетки eGFP+ Lin-HSC образуют сосуды через четырнадцать дней после введения (P17). (l и m) Введение родамин-декстрана в сердце показывает, что сосуды являются интактными и функциональными как в первичном (l), так и в глубоком сплетении (m).
На фиг.3 показано, что клетки eGFP+ Lin-HSC перемещаются к участку глиоза (определяемому по GFAP-экспрессирующим астроцитам, крайнее левое изображение), индуцированного либо лазерным (a), либо механическим (b) повреждением взрослой сетчатки (звездочка указывает поврежденный участок). На крайних правых изображениях видно повышенное скопление, демонстрирующее тесную связь Lin-HSC с астроцитами. Калибровочная шкала=20 мкМ.
На фиг.4 показано, что клетки that Lin-HSC восстанавливают сосудистую сеть мыши с дегенерацией сетчатки. (a-d) Сетчатки на 27 день после инъекции (P33) с окрашиванием коллагеном IV; (a) и (b) сетчатки, обработанные клетками Lin-HSC (Balb/c): отсутствует отличие от сосудистых сетей нормальных мышей FVB; (c) и (d) сетчатки, обработанные клетками Lin-HSC (Balb/c): густая сосудистая сеть, аналогичная сосудистой сети мышей дикого типа; (a) и (c) замороженные срезы целой сетчатки (вверху сторона, обращенная к стекловидному телу, внизу сторона, обращенная к склере) с окрашиванием DAPI; (b) и (d) глубокое сплетение в тотальном препарате сетчатки; (e) столбчатая диаграмма, иллюстрирующая увеличение васкуляризации глубокого сосудистого сплетения, образованного в сетчатках, обработанных клетками Lin-HSC (n=6). Степень глубокой васкуляризации определяют путем вычисления общей длины сосудов в каждом изображении. Сравнивают общую длину сосудов/область высокой мощности (в микронах) для Lin-HSC, Lin+HSC или контрольных сетчаток. (f) Сравнение длины глубокого сосудистого сплетения после введения клеток Lin-HSC (R - правый глаз) или Lin+HSC (L - левый глаз) мыши rd/rd. Показаны результаты, полученные от шести независимых мышей (каждый цвет обозначает независимую мышь), (g) и (h) Клетки Lin-HSC (Balb/c) также восстанавливают сосудистую сеть rd/rd после введения в глаза P15. Показано промежуточное и глубокое сосудистое сплетение в сетчатках, обработанных клетками Lin-HSC (G) или Lin+HSC (H) (через месяц после введения).
На фиг.5 приведены микрофотографии ткани сетчатки мыши: (a) тотальный препарат глубокого слоя сетчатки (мышь rd/rd), пятый день после инъекции (P11) с визуализацией eGFP+ Lin-HSC (серый), (b) и (c) сосудистая сеть сетчатки P60 мышей Tie-2-GFP (rd/rd), получающих клетки Lin- Balb/c (b) или инъекцию клеток Lin+HSC (c) на P6. В левых секциях (b) и (c) видны только эндогенные эндотелиальные клетки (окрашенные GFP). В средних секциях (b) и (c) показаны клетки, окрашенные антителом против CD31; стрелки указывают сосуды, окрашенные CD31, но не GFP, в правых секциях (b) и (c) показаны клетки, окрашенные и GFP и CD31. (d) Окрашивание α-SMA сетчатки, обработанной клетками Lin-HSC (левая секция), и контрольной сетчатки (правая секция).
На фиг.6 показано, что T2-TrpRS-трансфицированные Lin-HSC ингибируют развитие сосудистой сети мышиной сетчатки. (a) Схематическое изображение человеческих TrpRS, T2-TrpRS и T2-TrpRS, содержащих на амино-концах сигнальную последовательность Igk. (b) Сетчатки, обработанные клетками Lin-HSC, трансфицированными T2-TrpRS, экспрессируют белок T2-TrpRS in vivo. (1) Рекомбинантный T2-TrpRS, полученный в E. coli; (2) Рекомбинантный T2-TrpRS, полученный в E. coli; (3) Рекомбинантный T2-TrpRS, полученный в E. coli; (4) контрольная сетчатка; (5) сетчатка, обработанная Lin-HSC+pSecTag2A (только вектор); (6) сетчатка, обработанная Lin-HSC+pKLe135 (Igk-T2-TrpRS в pSecTag). (a) Эндогенный TrpRS. (b) Рекомбинантный T2-TrpRS. (c) T2-TrpRS сетчатки, обработанные клетками Lin-HSC. (c-f) Типичные первичные (поверхностные) и вторичные (глубокие) сплетения сетчаток через семь дней после инъекции. (c) и (d) Глаза, обработанные клетками Lin-HSC, трансфицированными пустой плазмидой, развиваются нормально. (e) и (f) В большинстве глаз, обработанных клетками Lin-HSC, трансфицированными T2-TrpRS, наблюдается ингибирование образования глубокого сплетения. (c) и (e) Первичное (поверхностное) сплетение. (d) и (f) вторичное (глубокое) сплетение). Слабое очертание сосудов, наблюдающееся в секции (f), обусловлено "проступающими" изображениями первичной сети сосудов, показанными в секции (e).
На фиг.7 показана последовательность ДНК, кодирующая His6-меченный T2-TrpRS, SEQ ID NO: 1.
На фиг.8 показана аминокислотная последовательность His6-меченного T2-TrpRS, SEQ ID NO: 2.
На фиг.9 показаны микрофотографии и электроретинограммы (ERG) сетчаток мышей, глаза которых были обработаны Lin-HSC и Lin+HSC (контроли).
На фиг.10 изображены статистические графики, отражающие взаимозависимость восстановления нейронов (ось y) и восстановления сосудов (ось x) для промежуточных (Int.) и глубоких сосудистых слоев глаз мышей rd/rd, обработанных Lin-HSC.
На фиг.11 изображены статистические графики, отражающие отсутствие корреляции между восстановлением нейронов (ось y) и восстановлением сосудов (ось x) для глаз мышей rd/rd, обработанных Lin+HSC.
На фиг.12 изображена столбчатая диаграмма, отражающая длину сосудов (ось y), выраженную в произвольных относительных единицах, для глаз мышей rd/rd, обработанных Lin-HSC (темные столбики), и для необработанных глаз мышей rd/rd (светлые столбики) через 1 месяц (1M), 2 месяца (2M) и 6 месяцев (6M) после введения.
Фиг.13 включает три столбчатых диаграммы, отражающих число ядер во внешнем нейронном слое (ONR) мышей rd/rd через 1 месяц (1M), 2 месяца (2M) и 6 месяцев (6M) после введения, и демонстрирует значительное увеличение числа ядер в глазах, обработанных Lin-HSC (темные столбики), по сравнению с контрольными глазами, обработанными Lin+HSC (светлые столбики).
На фиг.14 изображены графики, отражающие число ядер во внешнем нейронном слое отдельных мышей rd/rd и сравнивающие правый глаз (R, обработанный Lin-HSC) с левым глазом (L, контрольный глаз, обработанный Lin+HSC) в моменты времени (после инъекции) 1 месяц (1M), 2 месяца (2M) и 6 месяцев (6M); каждая линия в конкретном графике сравнивает глаза отдельной мыши.
На фиг.15 изображены изменения в сосудистой сети и нервных клетках сетчатки мышей rd1/rd1 (C3H/HeJ, левые секции) или мышей дикого типа (C57BL/6, правые секции). Показана сосудистая сеть промежуточных (верхние панели) или глубоких (средние панели) плексиформных слоев в тотальных препаратах сетчатки (красный: коллаген IV, зеленый: CD31) и срезах (красный: DAPI, зеленый: CD31, нижние секции) тех же препаратов сетчатки (P: день после родов). (GCL: слой ганглионарных клеток, INL: внутренний нуклеарный слой, ONL: внешний нуклеарный слой).
На фиг.16 показано, что введение Lin-HSC останавливает дегенерацию нервных клеток у мышей rd1/rd1. (A, B и C) сосудистая сеть сетчатки в промежуточных (Int.) или глубоких плексиформных слоях и срезах глаза, обработанного Lin-HSC (правые секции), и противоположного глаза, обработанного контрольными клетками (CD31-) (левые секции), в моменты P30 (A), P60 (B) и P180 (C). (D) Средняя общая длина сосудистой сети (+ или - стандартная ошибка среднего значения) в сетчатках, обработанных Lin-HSC или контрольными клетками (CD31"), в моменты P30 (слева, n=10), P60 (посередине, n=10) и P180 (справа, n=6). Данные, полученные для промежуточного (Int.) и глубокого плексиформного слоев, показаны раздельно (ось Y: относительная длина сосудистой сети). (E) Среднее число клеточных ядер в ONL сетчаток, обработанных контрольными клетками (CD31-) или Lin-HSC, в моменты P30 (слева, n=10), P60 (посередине, n=10) или P180 (справа, n=6) (ось Y: относительное число клеточных ядер в ONL). (F) Линейная корреляция между длиной сосудистой сети (ось X) и числом клеточных ядер в ONL (ось Y) сетчаток, обработанных Lin-HSC или контрольными клетками, в моменты P30 (слева), P60 (посередине) и P180 (справа).
На фиг.17 показано, что функция сетчатки восстанавливается под воздействием Lin-HSC. Электроретинографические (ERG) кривые используют для измерения функции сетчаток, обработанных Lin-HSC или контрольными клетками (CD31"). (A и B) Типичные случаи восстановленных и не восстановленных сетчаток через 2 месяца после введения. Показаны срезы сетчатки правого глаза, обработанного Lin-HSC (A), и левого глаза того же животного, обработанного контрольными клетками CD31- (B) (зеленый: сосудистая сеть, окрашенная CD31, красный: ядра, окрашенные DAPI). (C) Результаты ERG животного, используемого для получения данных, представленных в секциях (A) и (B).
На фиг.18 показано, что популяция человеческих клеток костного мозга может восстановить дегенерирующую сетчатку у мыши rd1 (A-C). Восстановление также наблюдается у другой модели дегенерации сетчатки, rd10 (D-K). (A) Человеческие Lin-HSC (hLin-HSC), меченные зеленым красителем, могут дифференцироваться в клетки сосудов сетчатки после введения в стекловидное тело мышей C3SnSmn.CB17-Prkdc SCID. (B и C) Сосудистая сеть сетчатки (левые секции; вверху: промежуточный плексиформный слой, внизу: глубокий плексиформный слой) и нервные клетки (правая секция) в глазу, обработанном hLin-HSC (B), или в противоположном контрольном глазу (C) через 1,5 месяца после инъекции. (D-K) Восстановление мышей rd10 под воздействием Lin-HSC (введенных на P6). Типичные сетчатки показаны в моменты P21 (D: Lin-HSC, H: контрольные клетки), P30 (E: Lin-HSC, I: контрольные клетки), P60 (F: Lin-HSC, J: контрольные клетки) и P105 (G: Lin-HSC, K: контрольные клетки) (для каждого момента времени обработанные и контрольные глаза принадлежат одному животному). Сосудистую сеть сетчатки (на верхнем изображении в каждой секции представлен промежуточный плексиформный слой; на среднем изображении в каждой секции представлен глубокий плексиформный слой) окрашивают CD31 (зеленый) и коллагеном IV (красный). На нижнем изображении в каждой секции показан поперечный срез, полученный из той же сетчатки (красный: DAPI, зеленый: CD31).
На фиг.19 показано, что кристаллин αA подвергается повышающей регуляции в восстановленных клетках внешнего нуклеарного слоя после обработки Lin-HSC, но не в противоположных глазах, обработанных контрольными клетками. Левая панель: IgG контроль в восстановленной сетчатке, средняя панель: кристаллин αA в восстановленной сетчатке, правая панель: кристаллин αA в невосстановленной сетчатке.
На фиг.20 приведены таблицы генов, которые подвергаются повышающей регуляции в сетчатках мыши, обработанных Lin-HSC по настоящему изобретению. (A) Гены, экспрессия которых в сетчатках мыши повышается в 3 раза после обработки мышиными Lin-HSC. (B) Гены кристаллина, которые подвергаются повышающей регуляции в сетчатках мыши, обработанных мышиными Lin-HSC. (C) Гены, экспрессия которых в сетчатках мыши повышается в 2 раза после обработки человеческими Lin-HSC. (D) Гены нейротрофических факторов или факторов роста, экспрессия которых подвергается повышающей регуляции в сетчатках мыши, обработанных человеческими Lin-HSC.
Фиг.21 иллюстрирует распределение поверхностных антигенов CD31 и интегрина α6 на CD133-положительных (DC133+) и CD133-отрицательных (CD133-) человеческих популяциях Lin-HSC. В левых секциях приведены диаграммы рассеяния, полученные методом проточной цитометрии. В центральных и правых секциях приведены гистограммы, демонстрирующие уровень экспрессии специфического антитела на клеточной популяции. Ось Y соответствует числу событий, а ось X соответствует интенсивности сигнала. Если закрашенная гистограмма сдвинута вправо от незакрашенной (контрольной) гистограммы, это означает, что флуоресцентный сигнал имеет повышенную интенсивность, а уровень экспрессии антитела выше фонового.
Фиг.22 иллюстрирует постнатальное развитие сетчатки у мышей дикого типа C57/B16, выращиваемых в атмосфере с нормальным содержанием кислорода (нормоксия) в период P0-P30 после родов.
Фиг.23 иллюстрирует индуцированную кислородом модель ретинопатии у мышей C57/B16, выращиваемых в атмосфере с высоким содержанием кислорода (гипероксия; 75% кислорода) в период P7-P12 с последующим периодом нормоксии в течение P12-P17.
Фиг.24 демонстрирует восстановление сосудов после обработки популяциями Lin-HSC у модели индуцированной кислородом ретинопатии (OIR).
На фиг.25 показаны восстановленные фоторецепторы внешнего нуклеарного слоя (ONL) мышей rd1 после введения Lin-HSC в стекловидное тело, которые преимущественно представляют собой колбочки. Небольшой процент фоторецепторов сетчатки мышей дикого типа (верхняя секция) представляет собой колбочки, что подтверждается экспрессией красного/зеленого опсина колбочек (A), тогда как большая часть клеток ONL является положительной по специфичному для палочек родопсину (B). После обработки неиммунной сывороткой наблюдается аутофлуоресценция сосудистой сети сетчатки (C), однако нуклеарные слои совсем не окрашиваются опсинами, специфичными для палочек или колбочек. Сетчатки мышей rd/rd (нижние секции) имеют уменьшенный внутренний нуклеарный слой и почти полностью атрофированный ONL, причем оба указанных слоя являются отрицательными по опсину колбочек (D) или палочек (секция G). Контрольные глаза, обработанные CD31-HSC, идентичны необработанным сетчаткам rd/rd и не окрашиваются опсином колбочек (E) или палочек (H). Противоположные глаза, обработанные Lin-HSC, имеют значительно уменьшенный, но явно присутствующий ONL, который преимущественно состоит из колбочек, что доказывается положительной иммунореактивностью по отношению к красному/зеленому опсину колбочек (F). Также присутствует небольшое число палочек (I).
На фиг.26 показаны диаграммы рассеяния, полученные методом проточной цитометрии для популяций стволовых клеток с положительной или отрицательной линией дифференцировки (верхняя левая и нижняя левая диаграммы, соответственно), которые демонстрируют процент клеток, экспрессирующих антиген CD44 (окрашены красным); а также диаграммы, полученные для CD31-отрицательных и CD31-положительных клеточных популяций (верхняя правая и нижняя правая диаграммы, соответственно), которые демонстрируют процент клеток, экспрессирующих антиген CD44 (окрашены красным).
На фиг.27 показаны диаграммы рассеяния, полученные методом проточной цитометрии для клеточных популяций отрицательной линии дифференцировки, которые экспрессируют антиген CD44 на значительном уровне (левый ряд диаграмм), а также для субпопуляций клеток костного мозга, которые не экспрессируют значительных количеств антигена CD44 (правый ряд диаграмм), на этих диаграммах показано относительное процентное содержание клеток, экспрессирующих другие поверхностные антигены.
На фиг.28 показаны микрофотографические изображения сетчатки мыши, обработанной путем введения в стекловидное тело клетками из популяции MLBM данного изобретения (левая секция), по сравнению с сетчаткой мыши, обработанной путем введения в стекловидное тело клетками CD44L0.
На фиг.29 показаны микрофотографические изображения сетчаток глаз, обработанных клетками из популяции MLBM (CD44HI) и клетками CD44LD.
На фиг.30 приведены столбчатые диаграммы, демонстрирующие благоприятные эффекты клеточной популяции MLBM, заключающиеся в уменьшении патогенного ангиогенеза и стимуляции полезной физиологической реваскуляризации мышиных сетчаток в индуцированной кислородом модели ретинопатии или ретинопатии недоношенных. На верхнем графике сравнивается преретинальный участок пучка новых сосудов в контрольной сетчатке (первый столбик), сетчатке, обработанной клетками CD44LD (средний столбик), и в сетчатках, обработанных клетками из популяции MLBM (правый столбик). На нижнем графике сравнивается участок разрушения сосудов в контрольной сетчатке (первый столбик), сетчатке, обработанной клетками CD44LD (средний столбик), и в сетчатках, обработанных клетками из популяции MLBM (правый столбик).
На фиг.31 приведено микрофотографическое изображение, демонстрирующее, что только клетки из популяции MLBM внедряются в сосудистую сеть сетчатки, причем данные клетки экспрессируют эндотелиальный фактор роста сосудов (VEGF), что подтверждает зеленое окрашивание клеток в нижней части изображения.
На фиг.32 приведены микрофотографические изображения, демонстрирующие, что клетки из популяции CD11b+ MLBM данного изобретения селективно направлены на сосудистую сеть сетчатки.
На фиг.33 приведены микрофотографические изображения, демонстрирующие, что клетки костного мозга CD44- CD11b- не обладают избирательностью в отношении сосудистой сети сетчатки.
На фиг.34 показана аминокислотная последовательность фрагмента T2 TrpRS (SEQ ID NO: 3) и его варианта T2-TrpRS-GD (SEQ ID NO: 4).
На фиг.35 показана аминокислотная последовательность мини-TrpRS (SEQ ID NO: 5).
На фиг.36 показана аминокислотная последовательность T1-TrpRS (SEQ ID NO: 6).
На фиг.37 показано нормальное развитие сосудов сетчатки мыши, модель индуцированной кислородом ретинопатии (OIR), и эффект восстановления после трансплантации внутрь стекловидного тела клеток костного мозга Lin-. Мышь рождается с сетчаткой, в значительной степени лишенной сосудов, как показано на 2 день после родов (P2) (секция a, тотальный препарат сетчатки), причем сосуды находятся в поверхностном слое сетчатки, занимая одну плоскость, как показано в секции b. В секциях b,d и f приведены изображения, полученные в результате 3-мерного воспроизведения конфокальных z-серий наборов данных анфас, повернутых на 90 градусов. В течение первой недели после рождения поверхностные сосуды сетчатки растут в радиальном направлении от диска зрительного нерва, почти достигая периферии на день P10 (c). Затем развивается глубокая сосудистая сеть сетчатки в результате ветвления поверхностного слоя, которое происходит в течение второй недели (d). Наконец, образуется третье сплетение сосудов между первыми двумя и примерно к P30 формируется зрелая сосудистая сеть сетчатки (e,f). В секции g приведены результаты, полученные на P10, которые показывают, что гипоксия вызывает у модели OIR разрушение центральных сосудов. В секции h показано, что после возвращения к среде с нормальным содержанием кислорода на P12, в центральной сетчатке начинается реваскуляризация и на границе между васкуляризированным (периферическим) и лишенным сосудов (центральным) слоями сетчатки формируются характерные преретинальные пучки новых сосудов. Данные пучки интенсивно окрашиваются изолектином. В секциях I-n показано, что гематопоэтические клетки-предшественники Lin- стимулируют восстановление сосудов у модели OIR. Клетки Lin-, введенные в стекловидное тело перед воздействием атмосферы с высоким содержанием кислорода, значительно ускоряют реваскуляризацию в центральной сетчатке по сравнению с противоположным глазом, обработанным средой на P17. В то время как в сетчатках, обработанных средой, наблюдается уменьшение поверхностной сосудистой сети (I) и полное отсутствие глубокой сосудистой сети (k, m), в противоположном глазу, обработанном клетками Lin-, наблюдается относительно нормальная сосудистая сеть сетчатки (J) с тремя сплетениями (k, m). В секции o показано, что на P17, в глазах OER, обработанных клетками Lin-, реваскуляризация полностью завершается гораздо чаще, чем в необработанных глазах, или в глазах, обработанных средой. Сосуды визуализируют путем перфузии сердца флуоресцеином-декстраном, секции a-f,i,j, и лектином GS, секции g,h,k-n. Ядра в секциях k-n метят DAPI.
На фиг.38 показано, что клетки Lin- ускоряют реваскуляризацию сетчатки и уменьшают формирование преретинального пучка новых сосудов в OIR. В секциях a-d показан метод компьютерного анализа изображения, используемый для вычисления площади разрушения сосудов сетчатки, а также формирование преретинального пучка новых сосудов (красный) в тотальных препаратах сетчатки глаз OIR на 17 день после родов. В секции e показаны сетчатки, обработанные клетками Lin- перед воздействием среды с высоким содержанием кислорода, в которых наблюдается почти 6-кратное уменьшение области разрушения по сравнению с необработанными контролями, и приблизительно 5-кратное уменьшение по сравнению с глазами, обработанными только средой. В секции f показано, что обработка клетками Lin- значительно уменьшает двухмерную площадь пучков новых сосудов по сравнению с необработанными глазами и глазами, обработанными средой. В секции g показано, что трансплантация клеток Lin- эффективно уменьшает площадь разрушения не только при введении перед воздействием среды с высоким содержанием кислорода, но и при введении в период P9-P12 в процессе воздействия среды с высоким содержанием кислорода и сразу после возвращения к нормальному содержанию кислорода. (На графиках представлены средние значения ± SEM; *p < 0,001).
На фиг.39 показано, что при обработке клетками костного мозга долговременные токсические эффекты являются небольшими или отсутствуют. Сетчатки, анализируемые на 5 или 6 месяцы после обработки клетками Lin-, имеют нормальную сосудистую сеть, а нейронный слой сетчатки выглядит гистологически неизмененным на поперечных срезах (a-f, необработанные сетчатки по сравнению с сетчатками, обработанными клетками Lin-, через 6 месяцев после трансплантации). Опухоли отсутствуют и единственными аномалиями являются "розетки", встречающиеся иногда в нейронном слое сетчатки, которые также можно видеть в контрольных необработанных глазах (g,h).
На фиг.40 показано, что клетки CD44HI преобладают в популяции Lin- и эффективно стимулируют восстановление сосудов в модели OIR. В секции a показано, что костный мозг содержит фракции CD44HI и CD44LD, причем популяция Lin- обогащена клетками CD44HI по сравнению с контрольными CD-клетками. На вкладках показаны светорассеивающие свойства клеток CD44HI, типичные для моноцитов и гранулоцитов, в то время как светорассеивающие свойства клеток CD44LD характерны для лимфоцитов. В секции b показаны типичные сетчатки глаз, обработанных клетками костного мозга CD44LD и CD44HI, на день P17, перед воздействием кислорода. В нижних панелях приведены значения площадей разрушения и реваскуляризации на день P17, используемые для получения данных, показанных в секции c. В секции c показано, что клетки CD44HI уменьшают разрушение сосудов и преретинальную реваскуляризацию с эффективностью, подобной наблюдающейся при обработке глаз клетками Lin-. Области разрушения сосудов (*) и преретинальной реваскуляризации (**) значительно ниже в глазах, обработанных CD44HI и Lin-, чем в необработанных глазах, или глазах, обработанных средой (во всех случаях p<105). Площадь разрушения в глазах, обработанных клетками Lin-, также снижена по сравнению с глазами, обработанными CD44HI (p=0,03), но в гораздо меньшей степени. Площади преретинальной реваскуляризации в глазах, обработанных Lin- и CD44HI, практически не различаются (p=0,25).
На фиг.41 показано, что субпопуляция CD44HI экспрессирует миелоидные маркеры. В секции a для дополнительной характеристики популяций CD44 используют двухцветную проточную цитометрию. Все клетки метят антителом против CD44 и одновременно разными указанными антителами. Популяция CD44HI интенсивно взаимодействует с антителами против CD11a, CD11b и Ly6GC. Фракции клеток CD44hi являются положительными по CD14, F4/80, cKit и CD115. Большинство данных антигенов присутствует на клетках миелоидной линии дифференцировки. Клетки CD44LD интенсивно взаимодействуют с антителами против Ter119 и CD45R B220, которые являются маркерами эритробластов и B-клеток, соответственно.
На фиг.42 показано, что клетки CD44HI локализуются в сетчатке около сосудов. С помощью конфокальной визуализации создают ряд изображений в z-измерении, которые затем преобразуют в трехмерные (3D). В секции a иллюстрируется CD31-меченный сосудистый эндотелий и экспрессия GFP после введения клеток костного мозга. Клетки костного мозга локализуются около сосудов. С помощью трехмерных изображений визуализируют просвет сосудов и относительное положение GFP+ клеток костного мозга. Номера, приведенные в секции (b), соответствуют положениям поперечных срезов, указанным в секции (a). Сигнал GFP детектируют вне просвета во всех случаях, за исключением секции b, среза №3, который проходит через клетку и характеризуется интенсивной флуоресценцией, где выражено проступание сигнала.
На фиг.43 показан анализ in situ клеток костного мозга CD44HI, введенных в модель OIR. Мечение контрольной необработанной сетчатки показывает присутствие эндогенных периваскулярных клеток F4/80+ (a-c). Вводимые клетки CD44HI направлены на сосудистую сеть сетчатки и имеют локализацию, морфологию и экспрессию F4/80, подобные наблюдающимся в эндогенных клетках (d-i). Трансплантированные периваскулярные клетки костного мозга не экспрессируют CD44, в то время как клетки, не связанные с сосудистой сетью сетчатки, сохраняют экспрессию CD44 (j-o).
На фиг.44 приведены результаты анализа экспрессии с использованием микрочипов, свидетельствующие о высоком уровне экспрессии миелоид-ассоциированных генов в популяции CD44HI, в то время как клетки CD44LD экспрессируют гены, ассоциированные с лимфоидными клетками. Для сравнения профилей экспрессии генов в указанных популяциях клеток костного мозга используют микрочипы AFFYMETRIX®. Уровни экспрессии анализируемых генов различаются, как минимум, в 5 раз. В популяции CD44HI наблюдается значительно более высокий уровень экспрессии CD44, чем в клетках CD44LD.
На фиг.45 показано, что клетки CD44HI могут дифференцироваться в клетки, имеющие микроглиальные характеристики. В секциях A и B показано, что вводимые клетки CD44HI экспрессируют CD11b и F4/80 и подобны эндогенной микроглии в отношении морфологии и периваскулярной локализации. В секции C приведено 3-мерное изображение периваскулярной локализации вводимых клеток CD44HI. В секции D с большим увеличением показана морфология вводимых клеток CD44HI.
На фиг.46 показано, что клетки CD44HI можно выделить путем отрицательной селекции. В секции A показано, что после истощения костного мозга методом MACS с использованием антител, специфичных к CD45R/B220, TER119 и CD3e, можно получить популяцию клеток, которая более чем на 90 процентов состоит из клеток CD44HI. В секции B показано, что отрицательная фракция (популяция CD44HI) практически не содержит клетки CD45R/B220, TER119 и CD3e. В секции C показано, что полученные методом отрицательной селекции клетки CD44HI сохраняют специфичность к сетчатке и способность к дифференцировке.
На фиг.47 показано, что экспрессия HIF-1 имеет большое значение для клеток-предшественников, поскольку он опосредует восстановление у модели OIR. В секциях (a и b) показаны типичные окрашенные лектином GS тотальные образцы сетчатки глаз, обработанных клетками CD44HI, полученными из миелоид-специфичных мышиных клеток с нокаутом гена HIF-1α (a), или клетками CD44HI, полученными из мышей дикого типа (b). В секциях (c и d) приведены изображения тех же сетчаток, что и в секциях a и b, с указанием размеров площадей разрушения сосудов (светлые участки) и пучков новых сосудов (темные участки). В секции (e) приведены скомпилированные данные, свидетельствующие о значительном уменьшении восстановительной активности в глазах, обработанных клетками CD44HI, полученными из клеток с нокаутом HIF-1α. *p≤0,0003, **p=0,024, n=15, результаты выражены в виде средних значений ± SEM. Для статистических вычислений используют парные глаза.
На фиг.48 показано, что при сравнении штаммов C57BL/6J и BALB/cByJ обнаружено измеримое различие в числе микроглиальных клеток CD11b+ в процессе ишемической фазы модели OIR. В секции (a) приведены изображения тотальных препаратов сетчатки, показывающие, что оба штамма имеют примерно одинаковую степень разрушения сосудов в центральной области сетчатки на 12 день после родов (P12); на P17 в сосудистой сети сетчаток наблюдаются сильные различия, причем в сетчатках C57BL/6J присутствует большое число пучков новых сосудов и небольшая степень реваскуляризации центральной области сетчатки. И наоборот, в сетчатках BALB/cByJ к этому моменту наблюдается низкая степень или отсутствие преретинальной реваскуляризации и практически полное восстановление сосудов. В секции (b) показано, что после 48 часов ишемии сетчатки C57BL/6J (слева) содержат меньше микроглиальных клеток CD11b+, чем сетчатки BALB/cByJ (справа). На всех изображениях диск оптического нерва располагается внизу справа. В секции (c) приводятся данные по количеству микроглиальных клеток CD11b+ в зависимости от времени для двух штаммов, которые указывают на то, что сетчатки C57BL/6J содержат меньше микроглиальных клеток после возвращения к нормальному содержанию кислорода на P12 (0 часов ишемии), и на P14 (48 часов ишемии) в два раза меньше микроглиальных клеток, чем в сетчатках мышей BALB/cByJ. p ≤ 0,02 для BALB/cByJ по сравнению с C57BL/6J во все моменты времени, n=8-11. В секции (d) показано, что уменьшение количества микроглиальных клеток у C57BL/6J индуцирует сильное уменьшение существующей сети микрососудов в процессе развития сетчатки. Введение липосом с клодронатом на P5 приводит к значительному уменьшению микроглиальных клеток CD11b+ и исчезновению капилляров на P8. Приведенные изображения получены для таких же участков сетчатки с диском зрительного нерва в нижней правой части. В секции (e) показано, что введение липосом с клодронатом в C57BL/6J на P2 вызывает значительное уменьшение микроглиальных клеток CD11b+ и сильное подавление и разрушение сосудистой сети сетчатки на P6 по сравнению с контрольным противоположным глазом, обработанным липосомами, содержащими PBS; на вкладке показано, что меченные препараты липосом (красные) специфически поглощаются микроглиальными клетками CD11b+ (зеленые), но не клетками сосудов, меченными лектином GS (синие), или другими клетками CD11b-.
На фиг.49 приведены графические результаты FACS-анализа клеток, выделенных из костного мозга человека, позволяющие определить число клеточных маркеров, экспрессированных на клетках.
На фиг.50 приведены графические результаты FACS-анализа клеток CD44W, выделенных из костного мозга человека.
На фиг.51 приведены графические результаты FACS-анализа клеток, выделенных из костного мозга человека, позволяющие идентифицировать клеточные маркеры CD11a, CD11b, CD33 и CD114.
На фиг.52 приведены диаграммы, демонстрирующие площадь пучков и площадь разрушения в сетчатках мышей, обработанных человеческими и мышиными клетками CD44W, по сравнению с контрольными сетчатками, обработанными PBS.
На фиг.53 приведены графические результаты FACS-анализа клеток, выделенных из периферической крови человека, позволяющие идентифицировать ряд клеточных маркеров.
На фиг.54 приведены диаграммы, демонстрирующие площадь пучков и площадь разрушения в сетчатках мышей, обработанных клетками CD44HI, полученными путем положительной селекции с использованием CD14 и CD33, и клетками CD14-, по сравнению с сетчатками, обработанными PBS (контроль).
На фиг.55 приведены графические результаты FACS-анализа клеток, выделенных из периферической крови человека.
На фиг.56 приведены диаграммы, демонстрирующие площадь пучков и площадь разрушения в сетчатках мышей, обработанных разными клеточными популяциями данного изобретения, по сравнению с контрольными сетчатками, обработанными PBS.
На фиг.57 показана дифференцировка мононуклеарных клеток пуповинной крови человека в эндотелиальные клетки.
На фиг.58 показано иммунное окрашивание мононуклеарных клеток пуповинной крови человека антителом против KDR (VEGFR2).
На фиг.59 показано иммунное окрашивание мононуклеарных клеток пуповинной крови человека антителом против PECAM (CD31).
На фиг.60 приведены диаграммы, демонстрирующие площадь пучков и площадь разрушения в сетчатках мышей, обработанных миелоподобными клетками данного изобретения, полученными из пуповины человека, по сравнению с сетчатками, обработанными только PBS.
На фиг.61 приведены диаграммы, демонстрирующие площадь пучков и площадь разрушения в сетчатках мышей, обработанных миелоподобными клетками данного изобретения, полученными из пуповины человека, по сравнению с сетчатками, обработанными только PBS.
На фиг.62 показаны микрофотографии свежих мононуклеарных клеток пуповинной крови, идентифицированных с помощью лентивирусной экспрессии eGFP.
На фиг.63 показаны (вверху) результаты анализа FACS и (внизу) диаграммы, демонстрирующие площадь пучков и площадь разрушения в сетчатках мышей, обработанных миелоподобными клетками пуповинной крови человека, выделенными путем положительной селекции с использованием CD14, по сравнению с сетчатками, обработанными только PBS или клетками CD14-.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ
Костный мозг, периферическая кровь и кровь пуповины содержат субпопуляцию миелоподобных клеток, которая экспрессирует антиген CD44 (т.е., рецептор гиалуроновой кислоты) и CD11b (интегрин αM). Миелоподобную популяцию клеток костного мозга, обогащенную CD44- и CD11b-экспрессирующими клетками, можно выделить из костного мозга путем обработки клеток костного мозга антителом против антигена CD44 (анти-CD44) и/или антителом против антигена CD11b (анти-CD11b) с последующим отбором клеток, способных к иммунному взаимодействию с антителом. Затем антитело можно отделить от клеток с помощью хорошо известных в данной области способов. Отбор клеток можно осуществить, например, с помощью проточной цитометрии с использованием антител, связанных с гранулами, или нанесенных на гранулы, и затем фильтрации или других способов разделения, хорошо известных в данной области. Большая часть отобранных клеток относится к отрицательной линии дифференцировки и экспрессирует как антиген CD44, так и антиген CD11b, независимо от того, какое антитело использовалось при выделении.
Помимо костного мозга популяции миелоподобных клеток, экспрессирующие CD44 и CD11b, также можно выделить из периферической крови и пуповинной крови. Предпочтительно популяцию миелоподобных клеток выделяют из костного мозга человека, периферической крови человека или пуповинной крови человека.
Костный мозг содержит стволовые клетки. Стволовые клетки обычно идентифицируют по распределению антигенов на поверхности клеток (подробное описание можно найти в докладе, подготовленном Национальными институтами здравоохранения, Управление по научной политике, в июне 2001 г: Stem Cells: Scientific Progress and Future Directions, приложение E: Stem Cell Markers, который включен в данное описание в качестве ссылки во всей полноте). Приблизительно 75% гематопоэтических стволовых клеток отрицательной линии дифференцировки, выделенных из костного мозга, также являются CD44-положительными. В предпочтительном воплощении основная часть клеток из популяции MLBM представляет гематопоэтические стволовые клетки отрицательной линии дифференцировки (т.е., CD44+Lin-HSC).
Настоящее изобретение предлагает способ уменьшения сосудистой и нейронной дегенерации сетчатки млекопитающего, страдающего глазным заболеванием. Выделенную популяцию миелоподобных клеток по данному изобретению вводят в сетчатку млекопитающего, предпочтительно в стекловидное тело. Клетки вводят в количестве, достаточном для уменьшения сосудистой и/или нейронной дегенерации сетчатки. Предпочтительно, выделенная популяция миелоподобных клеток является аутологичной для млекопитающего, подлежащего лечению. Предпочтительно, клетки из популяции миелоподобных клеток вводят в физиологически приемлемой среде, такой как физиологический раствор, забуференный фосфатом (PBS).
Предпочтительный способ включает выделение популяции миелоподобных клеток из костного мозга млекопитающего, подлежащего лечению, и затем введение клеток млекопитающему в количестве, достаточном для уменьшения сосудистой и/или нейронной дегенерации сетчатки. Клетки можно получить от млекопитающего, страдающего дегенеративным заболеванием глаза, предпочтительно на ранней стадии заболевания глаза, или от здорового млекопитающего, имеющего предрасположенность к дегенеративным заболеваниям глаз (т.е., генетическую предрасположенность). В последнем случае выделенную популяцию миелоподобных клеток можно хранить после выделения, и ее можно вводить профилактически на ранних стадиях развивающегося глазного заболевания. Предпочтительно больная сетчатка содержит активированные астроциты, на которые направлены клетки из популяции миелоподобных клеток. Соответственно, если имеет место ассоциированный глиоз, полезно провести раннюю обработку глаза. Альтернативно, чтобы стимулировать локальную пролиферацию активированных астроцитов, перед введением аутологичной популяции миелоподобных клеток сетчатку можно обработать лазером.
Гематопоэтические стволовые клетки представляют собой стволовые клетки, способные развиваться в клетки крови разных типов, например B-клетки, T-клетки, гранулоциты, тромбоциты и эритроциты. Поверхностные антигены линии дифференцировки представляют собой группу клеточных поверхностных белков, которые являются маркерами линий зрелых клеток крови и включают CD2, CD3, CD11, CD11a, Mac-1 (CD11b:CD18), CD14, CD16, CD19, CD24, CD33, CD36, CD38, CD45, CD45RA, мышиный Ly-6G, мышиный TER-119, CD56, CD64, CD68, CD86 (B7.2), CD66b, человеческий лейкоцитарный антиген DR (HLA-DR) и CD235a (гликофорин A). Гематопоэтические стволовые клетки, которые не экспрессируют значительных уровней данных антигенов, обычно относят к отрицательной линии дифференцировки (Lin-). Человеческие гематопоэтические стволовые клетки, как правило, дополнительно экспрессируют такие поверхностные антигены, как CD31, CD34, CD117 (c-kit) и/или CD133. Мышиные гематопоэтические стволовые клетки, как правило, дополнительно экспрессируют такие поверхностные антигены, как CD34, CD117 (c-kit), Thy-1 и/или Sca-1.
Указанные миелоподобные клетки по данному изобретению CD44+ CD11b способны внедряться в развивающуюся сосудистую сеть и затем дифференцироваться с образованием сосудистых эндотелиальных клеток.
В данном описании и в прилагающейся формуле изобретения фраза "взрослый" в применении к костному мозгу и клеткам костного мозга, относится к костному мозгу, выделенному после родов, т.е., из молодых и взрослых особей, но не из эмбрионов. Соответственно, термин "взрослое млекопитающее" относится как к молодым (после рождения), так и к половозрелым млекопитающим, но не к эмбрионам или внутриутробным субъектам.
Выделенные популяции миелоподобных клеток по настоящему изобретению селективно направлены на астроциты и способны внедряться в сосудистую сеть сетчатки после введения в стекловидное тело глаза млекопитающего, такого как мышь или человек, из которого были выделены клетки. Выделенные популяции миелоподобных клеток по настоящему изобретению включают клетки, способные внедряться в сосудистую сеть сетчатки, которые можно использовать для лечения заболеваний сетчатки, связанных с нарушением образования новых сосудов и с дегенерацией сосудов, а также для восстановления повреждения сосудов сетчатки. Популяция миелоподобных клеток по настоящему изобретению также стимулирует восстановление нейронов сетчатки и повышающую регуляцию антиапоптотических генов. Кроме того, популяцию миелоподобных клеток по данному изобретению можно использовать для лечения дефектов сетчатки глаз новорожденных млекопитающих, таких как млекопитающие, страдающие индуцированной кислородом ретинопатией, или ретинопатии недоношенности.
Обнаружено, что клетки костного мозга, которые не экспрессируют CD44 (клетки CD44LD), обычно экспрессируют один или несколько из следующих маркеров: Ter119, CD45RB220 и CD3e. Используя данный факт, можно выделить миелоподобные клетки по настоящему изобретению CD44HI с помощью способа, включающего отрицательную селекцию по клеточному маркеру. Данный способ включает контактирование совокупности клеток костного мозга, клеток периферической крови или клеток пуповинной крови с антителами, специфичными к Ter119, CD45RB220 и CD3e, удаление из совокупности клеток костного мозга клеток, способных к иммунному взаимодействию с антителами против Ter119, CD45RB220 и CD3e, и извлечение миелоподобных клеток костного мозга, которые не экспрессируют Ter119, CD45RB220 и CD3e. С помощью данного способа можно выделить популяцию миелоподобных клеток, в которых более 90 процентов составляют клетки, экспрессирующие CD44.
Настоящее изобретение также предлагает способ лечения глазных заболеваний млекопитающих, включающий выделение из костного мозга млекопитающего популяции миелоподобных клеток и введение клеток из данной популяции в стекловидное тело глаза млекопитающего в количестве, достаточном для подавления заболевания. Способ по настоящему изобретению можно использовать для лечения глазных заболеваний, таких как дегенеративные заболевания сетчатки, дегенеративные заболевания сосудов сетчатки, ишемические ретинопатиии, сосудистые кровотечения, протекание сосудов и хориоидиопатии у новорожденных, молодых или половозрелых млекопитающих. Примеры таких заболеваний включают возрастную дегенерацию желтого пятна (ARMD), диабетическую ретинопатию (DR), предполагаемый глазной гистоплазмоз (POHS), ретинопатию недоношенных (ROP), серповидноклеточную анемию и пигментную дегенерацию сетчатки, а также повреждения сетчатки.
Число клеток из популяции миелоподобных клеток, вводимых в глаз, должно быть достаточным для подавления болезненного состояния глаза. Например, количество вводимых клеток должно быть эффективным для восстановления повреждения сетчатки глаза, стабилизации сосудистой сети сетчатки, созревания сосудистой сети сетчатки и предотвращения или устранения протекания сосуда и кровотечения из сосуда.
Клетки из популяции миелоподобных клеток по настоящему изобретению можно трансфицировать терапевтически полезными генами, такими как гены, кодирующие антиангиогенные белки для применения в клеточной генной терапии глаза, и гены, кодирующие нейротрофические агенты для усиления эффектов восстановления нейронов.
Трансфицированные клетки могут содержать любой ген, терапевтически пригодный для лечения нарушений сетчатки. В одном предпочтительном воплощении трансфицированные клетки из популяции миелоподобных клеток по настоящему изобретению содержат ген, функционально кодирующий антиангиогенный пептид, в том числе белки, или белковые фрагменты, такие как TrpRS или их антиангиогенные (т.е., ангиостатические) фрагменты, например фрагменты TrpRS, обозначаемые T2-TrpRS (SEQ ID NO: 3, фиг.34), T2-TrpRS-GD (SEQ ID NO: 4, фиг.34), которые представляют собой предпочтительные ангиостатические пептиды, а также мини-TrpRS (SEQ ID NO: 5, фиг.35) и T1-TrpRS (SEQ ID NO: 6, фиг.36). Трансфицированные клетки из популяции миелоподобных клеток, кодирующие антиангиогенный пептид по настоящему изобретению можно использовать для лечения заболеваний сетчатки, связанных с аномальным развитием сосудов, таких как диабетическая ретинопатия и подобные заболевания. Предпочтительно клетки из популяции миелоподобных клеток представляют собой клетки человека.
В другом предпочтительном воплощении трансфицированные клетки из клеточной популяции MLBM по настоящему изобретению содержат ген, функционально кодирующий нейротрофический агент, такой как фактор роста, нейротрофин-3, нейротрофин-4, нейротрофин-5, нейротрофический фактор мерцательного эпителия, нейротрофический фактор пигментированного эпителия сетчатки, инсулиноподобный фактор роста, нейротрофический фактор глиальной клеточной линии, нейротрофический фактор мозга и т.п. Такие нейротрофические клетки из популяции миелоподобных клеток можно использовать для стимуляции восстановления нейронов при нейронных дегенеративных заболеваниях сетчатки, таких как глаукома и пигментная дегенерация сетчатки, при лечении повреждений нервов сетчатки и т.п. Описано применение нейротрофического фактора мерцательного эпителия для лечения пигментной дегенерации сетчатки (см. Kirby et al., 2001, Mol Ther. 3(2):241-8; Farrar et al., 2002, EMBO Journal 21:857-864). По имеющимся данным, нейротрофический фактор мозга модулирует гены, связанные с ростом в поврежденных ганглиях сетчатки (см. Foumier, et al., 1997, J. Neurosci. Res. 47:561-572). Описано, что нейротрофический фактор, продуцирующийся линией глиальных клеток, замедляет дегенерацию фоторецепторов при пигментной дегенерации сетчатки (см. McGee et al., 2001, Mol Ther. 4(6):622-9).
Настоящее изобретение также предлагает способы лечения глазных ангиогенных заболеваний путем введения трансфицированных клеток из популяции миелоподобных клеток по настоящему изобретению в стекловидное тело глаза. Такие трансфицированные клетки из популяции миелоподобных клеток включают клетки из популяции MLBM, трансфицированные терапевтически полезным геном, таким как ген, кодирующий антиангиогенный или нейротрофический генный продукт. Предпочтительно трансфицированные клетки из популяции миелоподобных клеток представляют собой клетки человека.
Предпочтительно, по меньшей мере, приблизительно 1×105 клеток из популяции MLBM или трансфицированных клеток из популяции миелоподобных клеток вводят в стекловидное тело глаза млекопитающего, страдающего дегенеративным заболеванием сетчатки. Необходимое число вводимых клеток может зависеть от тяжести дегенерации сетчатки, возраста млекопитающего и других факторов, хорошо известных рядовым специалистам в области лечения заболеваний сетчатки. Клетки из популяции миелоподобных клеток можно вводить в один прием, или в виде нескольких доз в течение периода времени, определенного врачом, отвечающим за лечение.
Популяции миелоподобных клеток по настоящему изобретению можно использовать для лечения повреждений сетчатки и дефектов сетчатки, участвующих в разрыве или деградации сосудистой сети сетчатки или нейронной дегенерации сетчатки. Популяции человеческих миелоподобных клеток также можно использовать для получения линии генетически идентичных клеток, т.е., клонов, пригодных для применения в регенеративном или восстановительном лечении сосудистой сети сетчатки, а также для лечения или уменьшения нейронной дегенерации сетчатки. Кроме того, популяции миелоподобных клеток по настоящему изобретению можно использовать в качестве исследовательских инструментов при изучении развития сосудов сетчатки и для доставки генов к некоторым клеткам-мишеням, таким как астроциты.
Развитие сосудов сетчатки мышей
Модель глазного ангиогенеза. Глаз мыши представляет собой признанную модель для изучения развития сосудов сетчатки млекопитающих, например развития сосудов сетчатки человека. В процессе развития сосудистой сети сетчатки мыши в условиях ишемии кровеносные сосуды развиваются в тесной связи с астроцитами. Эти глиальные элементы мигрируют у зародыша человека третьего триместра развития или новорожденного грызуна в сетчатку из диска зрительного нерва вдоль слоя ганглионарных клеток и распространяются в радиальном направлении. При развитии сосудистой сети сетчатки мыши эндотелиальные клетки используют эту уже сформированную астроцитарную матрицу для установления характера сосудистой сети сетчатки (см. фиг.1 (a и b)). На фиг.1 (a и b) приведены схематические диаграммы развития сетчатки мыши. В секции (a) изображено развитие первичного сплетения (темные линии в верхней левой части диаграммы), наложенного на астроцитарную матрицу (светлые линии), а в секции (b) показана вторая фаза формирования сосудов сетчатки. На фиг.1 GCL обозначает ганглионарный клеточный слой; IPL обозначает внутренний плексиформный слой; INL обозначает внутренний нуклеарный слой; OPL обозначает внешний плексиформный слой; ONL обозначает внешний нуклеарный слой; RPE обозначает пигментный эпителий сетчатки; ON обозначает зрительный нерв; и P обозначает периферию.
В момент рождения сосудистая сеть сетчатки фактически отсутствует. На 14 день после родов (P14) развиваются комплексные первичные (поверхностные) и вторичные (глубокие) слои сосудов сетчатки и появляется зрение. Вначале спицеобразные перипапиллярные сосуды растут радиально на существующей астроцитарной сети в направлении периферии, а затем между ними постепенно образуются связи в результате формирования капиллярного сплетения. Данные сосуды растут в виде монослоя в нервном волокне до P10 включительно (фиг.1(a)). В период P7-P8 первичное сплетение начинает давать боковые ветви, которые проникают в сетчатку до внешнего плексиформного слоя, где они образуют вторичный или глубокий плексиформный слой сетчатки. На P21 вся сеть подвергается экстенсивному ремоделированию и образуется третий, или промежуточный, плексиформный слой на внутренней поверхности внутреннего нуклеарного (фиг.1(b)).
Модель ангиогенеза в сетчатке новорожденной мыши используют для исследования роли HSC в процессе глазного ангиогенеза по нескольким причинам. В данной соответствующей физиологическим условиям модели перед появлением эндогенных кровеносных сосудов существует большая астроцитарная матрица, что позволяет оценить роль межклеточного взаимодействия в процессе формирования новых сосудов. Кроме того, известно, что последовательный и повторяющийся процесс формирования сетчатки у новорожденных проходит в условиях гипоксии, подобно многим заболеваниям сетчатки, в развитии которых, как известно, играет роль ишемия.
Обогащение эндотелиальных клеток-предшественников (EPC) из костного мозга
Хотя экспрессия клеточных поверхностных маркеров популяцией EPC, присутствующей в препаратах HSC, хорошо охарактеризована, еще не определены маркеры, позволяющие однозначно идентифицировать EPC. Чтобы получить препарат, обогащенный EPC, из мононуклеарных клеток костного мозга мышей удаляют клетки, положительные по маркерам гематопоэтической линии дифференцировки (Lin+), т.е., B-лимфоциты (CD45), T-лимфоциты (CD3), гранулоциты (Ly-6G), моноциты (CD11) и эритроциты (TER-11. Для дополнительного обогащения EPC используют антиген Sca-1. При сравнении результатов, полученных после введения в стекловидное тело одинакового числа клеток Lin-Sca-1+ или клеток Lin-, различие среди двух групп не обнаружено. В действительности, при введении только клеток Lin-Sca-1+ наблюдается гораздо более интенсивное внедрение в развивающиеся кровеносные сосуды.
Популяции Lin-HSC обогащают EPC, используя функциональные анализы. Кроме того, популяции Lin+HSC значительно отличаются от популяций Lin-HSC в функциональном отношении. Также анализируют эпитопы, обычно используемые для идентификации EPC каждой фракции (по результатам ранее опубликованных аналитических исследований in vitro). Хотя все указанные маркеры связаны не только с фракцией Lin-, их количество в Lin-HSC примерно на 70-1800% выше, чем во фракции Lin+HSC (фиг.1(c)). На фиг.1, секция (c) приведены результаты анализа методом проточной цитометрии разделенных клеток костного мозга Lin+HSC и Lin-HSC. В верхнем ряду секции с показана гистограмма распределения немеченных антителом гематопоэтических стволовых клеток. R1 обозначает область измеримого селектируемого положительного PE-окрашивания; R2 обозначает GFP-положительную область. В среднем ряду показаны гистограммы для Lin-HSC, а в нижнем ряду показаны гистограммы для Lin+HSC. Клетки C57B/6 метят PE-конъюгированными антителами против Sca-1, c-kit, Flk-1/KDR, CD31. Данные Tie-2 получают с использованием мышей Tie-2-GFP. Проценты, приведенные в углах гистограмм указывают долю меченных клеток во всей популяции Lin- или Lin+HSC. Интересно, что признанные маркеры EPC, такие как Flk-1/KDR, Tie-2 и Sca-1, экспрессируются на низком уровне и, следовательно, не используются для дальнейшего фракционирования.
Lin-HSC можно выделить путем (a) извлечения костного мозга из взрослого млекопитающего; (b) выделения совокупности моноцитов из костного мозга; (c) мечения моноцитов набором биотин-конъюгированных антител против одного или нескольких линиеспецифических поверхностных антигенов, предпочтительно, линиеспецифических поверхностных антигенов, выбранных из группы, состоящей из CD2, CD3, CD4, CD11, CD11a, Mac-1, CD14, CD16, CD19, CD24, CD33, CD36, CD38, CD45, Ly-6G (мышиный), TER-119 (мышиный), CD45RA, CD56, CD64, CD68, CD86 (B7.2), CD66b, человеческий лейкоцитарный антиген DR (HLA-DR) и CD235a (гликофорин A); (d) удаления моноцитов, положительных по одному или нескольким линиеспецифическим поверхностным антигенам, из совокупности моноцитов; и (e) выделения популяции гематопоэтических стволовых клеток отрицательной линии дифференцировки.
Если Lin-HSC выделяют из костного мозга взрослого человека, предпочтительно, моноциты метят набором биотин-конъюгированных антител против линиеспецифических поверхностных антигенов CD2, CD3, CD4, CD11a, Mac-1, CD14, CD16, CD19, CD33, CD38, CD45RA, CD64, CD68, CD86 (B7.2) и CD235a. Если Lin-HSC выделяют из костного мозга взрослой мыши, предпочтительно, моноциты метят набором биотин-конъюгированных антител против линиеспецифических поверхностных антигенов CD3, CD11, CD45, Ly-6G и TER-119.
Введенные в стекловидное тело клетки HSC Lin - содержат EPC, направленные на астроциты и способные внедряться в развивающуюся сосудистую сеть сетчатки
Чтобы определить, действительно ли введенные в стекловидное тело Lin-HSC могут быть направлены на конкретные типы клеток сетчатки, использовать астроцитарную матрицу и участвовать в ангиогенезе сетчатки, приблизительно 105 клеток из композиции Lin-HSC по настоящему изобретению или клеток Lin+HSC (контроль, приблизительно 105 клеток), выделенных из костного мозга взрослых мышей (GFP или LacZ трансгенных), вводят в глаза мышей на 2 день после родов (P2). Через четыре дня после инъекции (P6) многие клетки из композиции Lin-HSC по настоящему изобретению, полученной из GFP или LacZ трансгенных мышей, прилипают к сетчатке и имеют характерный удлиненный вид эндотелиальных клеток (фиг.2 (a)). Фиг.2 иллюстрирует приживление Lin-HSC на развивающейся мышиной сетчатке. Как показано на фиг.2, секция (a), через четыре дня после введения (P6) в стекловидное тело клетки eGFP+ Lin-HSC присоединяются к сетчатке и дифференцируются на сетчатке.
Во многих областях сетчатки GFP-экспрессирующие клетки располагаются в соответствии с находящимися под ними астроцитами и напоминают кровеносные сосуды. Данные флуоресцентные клетки находятся впереди эндогенной развивающейся сосудистой сети (фиг.2 (b)). И наоборот, только небольшое число Lin+HSC (фиг.2 (c)) или мезентериальных эндотелиальных клеток взрослых мышей (фиг.2 (d)) присоединяется к поверхности сетчатки. Чтобы определить, действительно ли клетки из введенной популяции Lin-HSC могут присоединяться к сетчаткам с развитыми сосудами, композицию Lin-HSC вводят в глаза взрослых субъектов. Интересно, что не наблюдается присоединение клеток к сетчатке или внедрение их в развитые нормальные кровеносные сосуды сетчатки (фиг.2 (e)). Это свидетельствует о том, что композиции Lin-HSC по настоящему изобретению не разрушают нормальной развитой сосудистой сети и не инициируют аномальной васкуляризации в нормальных развитых сетчатках.
Чтобы определить взаимосвязь между введенными композициями Lin-HSC по настоящему изобретению и астроцитами сетчатки, используют трансгенную мышь, которая экспрессирует глиальный фибриллярный кислый белок (GFAP, маркер астроцитов) и управляемый промотором зеленый флуоресцентный белок (GFP). Анализ сетчаток указанных GFAP-GFP трансгенных мышей, обработанных Lin-HSC из трансгенных мышей eGFP, показывает, что наблюдается совместная локализация введенных eGFP EPC и эндогенных астроцитов (фиг.2 (f-h), стрелки). Наблюдающиеся процессы eGFP+Lin-HSC проходят в соответствии с нижележащей сетью астроцитов (стрелки, фиг.2 (g)). Анализ таких глаз показывает, что введенные меченные клетки только присоединяются к астроцитам; на день P6, когда периферия сетчатки еще не имеет эндогенных сосудов, наблюдается прилипание введенных клеток к астроцитам в еще не васкуляризованных участках сетчаток мышей. Неожиданным является то, что введенные меченные клетки можно обнаружить в глубоких слоях сетчатки, точно в участках, где впоследствии развиваются сосуды сетчатки (фиг.2 (I), стрелки).
Чтобы определить, действительно ли введенные Lin-HSC стабильно внедряются в развивающуюся сосудистую сеть сетчатки, сосуды сетчатки впоследствии анализируют в нескольких временных точках. Уже на P9 (семь дней после инъекции) Lin-HSC внедряются в структуры CD31+ (фиг.2 (j)). На P16 (14 дней после инъекции) клетки уже экстенсивно внедряются в сосудоподобные структуры сетчатки (фиг.2 (k)). Если перед умерщвлением животных внутрь сосудов ввести родамин-декстран (чтобы идентифицировать функциональные кровеносные сосуды сетчатки), можно обнаружить, что большая часть клеток Lin-HSC связана с проходимыми сосудами (фиг.2 (l)). Наблюдаются два типа распределения меченных клеток: (1) в соответствии с одним типом меченные клетки распределены вдоль сосудов между немечеными эндотелиальными клетками; и (2) в соответствии с другим типом распределения сосуды состоят только из меченных клеток. Введенные клетки также внедряются в сосуды глубокого сосудистого сплетения (фиг.2 (m)). В то время как единичное включение EPC, полученных из Lin-HSC, в новообразованную сосудистую сеть описано ранее, данные, свидетельствующие о том, что сосудистая сеть полностью состоит из данных клеток, опубликованы впервые. Указанные результаты демонстрируют, что клетки полученной из костного мозга популяции Lin-HSC, введенные в стекловидное тело, могут эффективно внедряться в любой слой формирующегося сосудистого сплетения сетчатки.
Гистологический анализ не относящихся к сетчатке тканей (например, мозга, печени, сердца, легких, костного мозга), проведенный не более чем через 5 или 10 дней после введения в стекловидное тело, не показывает присутствия каких-либо GFP-положительных клеток. Данный факт указывает на то, что субпопуляция клеток из фракции Lin-HSC селективно направлена на астроциты сетчатки и способна стабильно внедряться в развивающуюся сосудистую сеть сетчатки. Поскольку эти клетки обладают многими характерными чертами эндотелиальных клеток (ассоциируются с астроцитами сетчатки, имеют удлиненную морфологию, стабильно внедряются в проходимые сосуды и отсутствуют во внесосудистых участках), они представляют EPC, присутствующие в популяции Lin-HSC. Астроциты-мишени относятся к типу, наблюдающемуся при многих гипоксических ретинопатиях. Хорошо известно, что глиальные клетки являются выступающим компонентом новообразованных ветвей пучков сосудов, наблюдающихся при DR и других формах повреждений сетчатки. В условиях реактивного глиоза и индуцированной ишемией реваскуляризации активированные астроциты пролиферируют, продуцируют цитокины и осуществляют повышающую регуляцию GFAP, подобно тому, как это происходит в процессе формирования сосудистой матрицы неонатальной сетчатки многих видов млекопитающих, в том числе людей.
Популяции Lin-HSC направлены на активированные астроциты в глазах взрослых мышей, как и в глазах новорожденных, клетки Lin-HSC вводят в глаза взрослых с сетчаткой, поврежденной путем фотокоагуляции (фиг.3 (a)) или кончиком иглы (фиг.3 (b)). В обоих моделях популяция клеток с рельефным окрашиванием GFAP наблюдается только вокруг участка повреждения (фиг.3 (a и b)). После введения клетки композиций Lin-HSC локализуются в участке повреждения и остаются специфически ассоциированными с GFAP-положительными астроцитами (фиг.3 (a и b)). Также обнаружено, что присутствующие в данных участках клетки Lin-HSC мигрируют в более глубокий слой сетчатки, причем уровень миграции подобен наблюдающемуся в процессе неонатального формирования глубокой сосудистой сети сетчатки. Неповрежденные фрагменты сетчатки не содержат клеток Lin-HSC, идентичных наблюдаемым после введения Lin-HSC в нормальные неповрежденные сетчатки взрослых особей (фиг.2 (e)). Полученные результаты показывают, что композиции Lin-HSC могут быть селективно направлены на активированные глиальные клетки, присутствующие в сетчатках взрослых, пораженных глиозом, а также в сетчатках новорожденных, подвергающихся васкуляризации.
Введенные в стекловидное тело Lin - HSC могут восстанавливать и стабилизировать дегенерирующую сосудистую сеть
Поскольку введенные в стекловидное тело композиции Lin-HSC направлены на астроциты и способны внедряться в нормальную сосудистую сеть сетчатки, данные клетки также стабилизируют дегенерирующую сосудистую сеть при ишемических или дегенеративных заболеваниях сетчатки, ассоциированных с глиозом и дегенерацией сосудов. Мышь rd/rd представляет собой модель дегенерации сетчатки, в которой через месяц после рождения происходит полная дегенерация фоторецепторов и сосудистых слоев сетчатки. У таких мышей сосудистая сеть сетчатки развивается нормально до P16, после чего происходит регрессия глубоких сосудистых сплетений; у большинства мышей глубокие и промежуточные сплетения почти полностью дегенерируют на P30.
Чтобы определить, действительно ли HSC могут восстанавливать регрессирующие сосуды, Lin+ или Lin-HSC (из мышей Balb/c) вводят в стекловидное тело мышей rd/rd на P6. На P33 сосуды самого глубокого слоя сетчатки мышей, обработанных клетками Lin+, почти полностью отсутствуют (фиг.4 (a и b)). И наоборот, большая часть сетчаток, обработанных Lin-HSC, на P33 имеет почти нормальную сосудистую сеть с тремя параллельными, хорошо сформированными сосудистыми слоями (фиг.4 (a и d)). Количественный анализ данного эффекта показывает, что средняя длина сосудов глубокого сосудистого сплетения в глазах мышей rd/rd, обработанных Lin-, почти в три раза больше, чем в необработанных глазах, или глазах, обработанных Lin+ (фиг.4 (e)). Неожиданным является то, что введение композиции Lin-HSC, полученной из костного мозга взрослой мыши rd/rd (FVB/N), также приводит к восстановлению дегенерирующей сосудистой сети сетчатки новорожденной мыши rd/rd (фиг.4 (f)). Дегенерация сосудистой сети в глазах мышей rd/rd наблюдается уже через 2-3 недели после рождения. Введение Lin-HSC только на P15 также приводит к частичной стабилизации дегенерирующей сосудистой сети у мышей rd/rd в течение, по меньшей мере, одного месяца (фиг.4 (g и h)).
Композиция Lin-HSC, введенная более молодым (например, на P2) мышам rd/rd, также внедряется в развивающуюся поверхностную сосудистую сеть. Обнаружено, что на P11 данные клетки мигрируют на уровень глубокого сосудистого сплетения и образуют картину, идентичную наблюдающейся во внешнем сосудистом слое сетчатки мышей дикого типа (фиг.5 (a)). Чтобы более точно описать схему, посредством которой клетки из введенных композиций Lin-HSC внедряются в дегенерирующую сосудистую сеть сетчатки мышей rd/rd и стабилизируют ее, композицию Lin-HSC, полученную из мышей Balb/c, вводят в глаза мышей Tie-2-GFP FVB. Мыши FVB имеют генотип rd/rd и, поскольку они экспрессируют гибридный белок Tie-2-GFP, все эндогенные кровеносные сосуды являются флуоресцентными.
Если немеченые клетки из композиции Lin-HSC вводят в глаза новорожденных Tie-2-GFP с последующим внедрением данных клеток в развивающуюся сосудистую сеть, в эндогенных немеченых сосудах Tie-2-GFP должны присутствовать немеченые пробелы, которые соответствуют внедрению введенных немеченых Lin-HSC. Последующее окрашивание другим сосудистым маркером (например, CD-31) позволяет получить изображение целого сосуда и определить, действительно ли неэндогенные эндотелиальные клетки являются частью сосудистой сети. Через два месяца после инъекции CD31-положительные, Tie-2-GFP-отрицательные сосуды наблюдаются в сетчатках глаз, обработанных композицией Lin-HSC (фиг.5 (b)). Интересным является тот факт, что большая часть восстановленных сосудов содержит Tie-2-GFP-положительные клетки (фиг.5 (c)). Распределение перицитов, определяемое с помощью окрашивания актином гладкой мускулатуры, не изменяется после введения Lin-HSC, независимо от того, происходит ли восстановление сосудов (фиг.5 (d)). Полученные результаты отчетливо демонстрируют, что после введения в стекловидное тело клетки Lin-HSC мигрируют в сетчатку, участвуют в формировании нормальных кровеносных сосудов сетчатки и стабилизируют эндогенную дегенерирующую сосудистую сеть у генетически дефектной мыши.
Ингибирование ангиогенеза сетчатки трансфицированными клетками Lin - HSC
Заболевания сосудов сетчатки чаще обусловлены аномальной пролиферацией сосудов, чем дегенерацией. Трансгенные клетки, направленные на астроциты, можно использовать для доставки антиангиогенного белка и ингибирования ангиогенеза. Клетки из композиций Lin-HSC трансфицируют геном T2-триптофанил-тРНК-синтетазы (T2-TrpRS). T2-TrpRS представляет собой фрагмент TrpRS размером 43 кДа, способный эффективно ингибировать ангиогенез сетчатки (фиг.6 (a)). На P12 сетчатки глаз, обработанных на P2 композицией Lin-HSC, трансфицированных контрольной плазмидой (ген T2-TrpRS отсутствует), имеют нормальные первичные (фиг.6 (c)) и вторичные (фиг.6 (d)) сосудистые сплетения. Если на P2 в глаза вводят композицию Lin-HSC по настоящему изобретению, трансфицированную T2-TrpRS, анализ, проведенный через 10 дней, показывает, что первичная сеть имеет значительные нарушения (фиг.6 (e)), а формирование глубокой сосудистой сети сетчатки почти полностью подавляется (фиг.6 (T)). Несколько сосудов, присутствующих в таких глазах, значительно разрежены большими зазорами между сосудами. Степень ингибирования под действием T2-TrpRS-секретирующих Lin-HSC приведена в таблице 1.
T2-TrpRS продуцируется и секретируется клетками композиции Lin-HSC in vitro, и после введения указанных трансфицированных клеток в стекловидное тело в сетчатке можно обнаружить фрагмент T2-TrpRS размером 30 кДа (фиг.6 (b)). Данный фрагмент размером 30 кДа присутствует только в сетчатках, обработанных трансфицированными Lin-HSC, такое уменьшение кажущейся молекулярной массы по сравнению с рекомбинантным или синтезированным in vitro белком может быть обусловлено процессингом или деградацией T2-TrpRS in vivo. Полученные результаты показывают, что композиции Lin-HSC, направленные на активированные астроциты, можно использовать для доставки функционально активных генов, таких как гены, экспрессирующие ангиостатические молекулы, в сосудистую сеть сетчатки. Хотя существует возможность, что наблюдаемый ангиостатический эффект обусловлен клеточной активностью, это является маловероятным, поскольку глаза, обработанные идентичными, но не трансфицированными T2 композициями Lin-HSC, имеют нормальную сосудистую сеть сетчатки.
|
Введенные в стекловидное тело популяции Lin-HSC локализуются на астроцитах сетчатки, внедряются в сосуды и могут использоваться для лечения многих заболеваний сетчатки. Хотя большая часть клеток, введенных композиций HSC, прилипает к астроцитарной матрице, небольшое число мигрирует вглубь сетчатки, перемещаясь к участкам, где впоследствии развивается глубокая сосудистая сеть. Однако хотя GFAP-положительные астроциты не наблюдаются в данной области до 42 дня после рождения, это не исключает вероятности того, что GFAP-отрицательные глиальные клетки уже присутствуют, обеспечивая сигнал для локализации Lin-HSC. Проведенные ранее исследования показывают, что многие заболевания связаны с реактивным глиозом. В частности, при DR глиальные клетки и их внеклеточный матрикс связаны с патологическим ангиогенезом.
Поскольку клетки введенных композиций Lin-HSC специфически присоединяются к GFAP-экспрессирующим глиальным клеткам, независимо от типа повреждения, композиции Lin-HSC по настоящему изобретению можно использовать для направления на преангиогенные повреждения сетчатки. Например, при ишемических ретинопаптиях, таких как диабет, реваскуляризация протекает в ответ на гипоксию. Путем направления композиций Lin-HSC к участкам патологической реваскуляризации развитие новой сосудистой сети можно стабилизировать путем предотвращения нарушений в новообразованной сети, таких как кровоизлияние или отек (случаи утраты зрения, связанные с DR), и можно ослабить гипоксию, которая изначально стимулирует реваскуляризацию. Аномальные кровеносные сосуды могут восстанавливаться до нормального состояния. Кроме того, ангиостатические белки, такие как T2-TrpRS можно доставлять к участкам патологического ангиогенеза путем применения трансфицированных композиций Lin-HSC и индуцированной лазером активации астроцитов. Поскольку лазерную фотокоагуляцию часто используют в клинической офтальмологии, данный способ часто применяется для лечения многих заболеваний сетчатки. Хотя такие способы, основанные на использовании клеток, разрабатываются для противораковой терапии, их применение для лечения болезней глаз имеет больше преимуществ, поскольку внутриглазное введение делает возможной доставку большого числа клеток непосредственно к участку заболевания.
Нейротрофическое и сосудотрофическое восстановление под действием Lin - HSC
С помощью MACS Lin-HSC выделяют из костного мозга мышей с повышенной экспрессией зеленого флуоресцентного белка (eGFP), C3H (rd/rd), FVB (rd/rd), как описано выше. Выделенные из указанных мышей Lin-HSC, содержащие EPC, вводят на P6 в стекловидное тело глаз мышей C3H или FVB. Сетчатки собирают в разные моменты времени после введения (1 месяц, 2 месяца и 6 месяцев) после инъекции. Сосудистую сеть анализируют методом сканирующей лазерной конфокальной микроскопии после окрашивания антителами против CD31 и путем гистологии сетчатки после окрашивания ядер DAPI. Чтобы идентифицировать гены, потенциально участвующие в данном процессе, с помощью микрочипов анализируют мРНК сетчаток в разные моменты времени.
На P21 в глазах мышей rd/rd наблюдается полная дегенерация как чувствительной иннервации, так и сосудистой сети сетчатки. Глаза мышей rd/rd, обработанные Lin-HSC на P6, сохраняют нормальную сосудистую сеть сетчатки вплоть до 6 месяцев; во всех временных точках (1M, 2M и 6M) состояние как глубоких, так и промежуточных слоев, значительно лучше, чем у контролей (см. фиг.12). Кроме того, авторы обнаружили, что сетчатки, обработанные Lin-HSC также являются более толстыми (1M; в 1,2 раза, 2M; в 1,3 раза, 6M; в 1,4 раза) и содержат больше клеток во внешнем нуклеарном слое (1M; в 2,2 раза, 2M; в 3,7 раза, 6M; в 5,7 раз) по сравнению с контрольными глазами, обработанными Lin+HSC. Крупномасштабный геномный анализ "восстановленных" (например, под действием Lin-HSC) сетчаток по сравнению с контрольными (не обработанными или обработанными клетками, отличными от Lin-) сетчатками rd/rd показывает, что имеет место значительная повышающая регуляция генов, кодирующих sHSP (маленькие белки теплового шока) и специфические факторы роста, связанные с восстановлением сосудов и нейронов, в том числе генов, кодирующих белки, перечисленные на фиг.20, секции A и B.
Полученные из костного мозга популяции Lin-HSC обеспечивают значительную и воспроизводимую защиту нормальной сосудистой сети и сильно увеличивают фоторецепторные и другие нейронные слои у мышей rd/rd. Данный нейротрофический восстановительный эффект коррелирует со значительной повышающей регуляцией маленьких белков теплового шока и факторов роста и может служить основой для разработки терапевтических способов лечения неизлечимых в настоящее время дегенеративных заболеваний сетчатки.
Сетчатки мышей rd1/rd1 с полной дегенерацией сосудов и нейронов
Нормальное развитие сосудов и нейронов сетчатки новорожденных мышей хорошо охарактеризовано и аналогично изменениям, наблюдающимся в третьем триместре человеческих зародышей (Dorrell et al., 2002, Invest. Ophthalmol. Vis. ScL 43:3500-3510). Мыши, гомозиготные по гену rd1, имеют много общих характеристик с дегенерацией человеческой сетчатки (Frasson et al., 1999, Nat. Med. 5: 1183-1187), у них происходит быстрая утрата фоторецепторов (PR), сопровождающаяся тяжелой сосудистой атрофией, в результате мутации гена, кодирующего фосфодиэстеразу цГМФ PR (Bowes et al., 1990, Nature 347:677-680). Для анализа сосудистой сети в процессе развития сетчатки и ее последующей дегенерации используют антитела против коллагена IV (CIV), белка внеклеточного матрикса (ECM) зрелой сосудистой сети и CD31 (PECAM-I), маркера эндотелиальных клеток (фиг.15). Сетчатки rd1/rd1 (C3H/HeJ) развиваются нормально примерно до 8 дня после родов (P), а затем начинается дегенерация внешнего нуклеарного слоя (ONL), содержащего фоторецепторы. ONL быстро дегенерирует и клетки погибают в результате апоптоза, так что на P20 остается только один нуклеарный слой. Двойное окрашивание тотальных препаратов сетчатки антителами против CIV и CD31 позволяет обнаружить характерные особенности дегенерации сосудов у мышей rd1/rd1, подобные описанным другими авторами (Blanks et al., 1986, J. Comp. Neurol. 254:543-553). Первичный и глубокий сосудистые слои сетчатки развиваются нормально до P12, после чего происходит быстрая утрата эндотелиальных клеток, как показывает отсутствие окрашивания CD31. Нормальное распределение CD31-положительных эндотелиальных клеток наблюдается до P12, но затем данные клетки быстро исчезают. Интересно, что CIV-положительное окрашивание присутствует во всех анализируемых моментах времени, это позволяет предположить, что сосуды и связанный с ними ECM образуются нормально, однако после P13 остается только матрикс, а CD31-положительные клетки отсутствуют (фиг.15, средние панели). Промежуточное сосудистое сплетение тоже дегенерирует после P21, однако в этом случае дегенерация прогрессирует медленнее, чем в глубоком сплетении (фиг.15, верхняя секция). Для сравнения с мышью rd1/rd1 показаны сосудистые и нейронные слои сетчатки нормальной мыши (правые секции, фиг.15).
Нейропротективное действие клеток костного мозга на мышей rd1/rd1
Введенные в стекловидное тело Lin-HSC внедряются в эндогенную сосудистую сеть сетчатки во всех трех сосудистых сплетениях и предотвращают дегенерацию сосудов. Интересно, что введенные клетки фактически никогда не присутствуют во внешнем нуклеарном слое. Данные клетки либо внедряются в формирующиеся сосуды сетчатки, либо находятся в непосредственной близости от таких сосудов. Мышиные Lin-HSC (из C3H/HeJ) вводят в стекловидное тело глаз мышей C3H/HeJ (rd1/rd1) на P6, непосредственно перед началом дегенерации. На P30 глаза, обработанные контрольными клетками (CD31-), имеют типичный фенотип rd1/rd1, т.е., наблюдается почти полная дегенерация глубокого сосудистого сплетения и в каждой анализируемой сетчатке присутствует ONL. Глаза, обработанные Lin-HSC, сохраняют нормальные промежуточные и глубокие сосудистые сплетения. Неожиданным является то, что значительно больше клеток наблюдается во внутреннем нуклеарном слое (INL) и ONL глаз, обработанных Lin-HSC, чем в глазах, обработанных контрольными клетками (фиг.16 (A)). Данный восстанавливающий эффект Lin-HSC можно наблюдать через 2 месяца (фиг.16 (B)) и вплоть до 6 месяцев после инъекции (фиг.16 (C)). Различия в сосудистой сети промежуточного и глубокого сплетений глаз, обработанных Lin-HSC, а также INL и ONL, содержащих нейронные клетки, являются значительными во все анализируемые моменты времени, при сравнении восстановленных и не восстановленных глаз (фиг.16 (B и C)). Количественное определение данного эффекта проводят путем измерения общей длины сосудистой сети (фиг.16 (D)) и определения числа DAPI-положительных клеточных ядер, присутствующих в ONL (фиг.16 (E)). Данные, полученные для всех временных точек, обрабатывают с помощью простого линейнорегрессионного анализа.
Статистически значимую корреляцию наблюдают между восстановлением сосудов и восстановлением нейронов (например, толщиной ONL) на P30 (p < 0,024) и P60 (p < 0,034) в глазах, обработанных Lin-HSC (фиг.16 (F)). Корреляция остается высокой, хотя статистически не значимой (p < 0,14), на P180 при сравнении сетчаток, обработанных Lin-HSC, с сетчатками, обработанными контрольными клетками (фиг.16 (F)). И наоборот, в сетчатках, обработанных контрольными клетками, не наблюдается значительной корреляции между сохранением сосудистой сети и ONL ни в одной временной точке (фиг.16 (F)). Полученные результаты показывают, что введение в стекловидное тело Lin-HSC приводит к восстановлению сосудов и нейронов сетчатки у мышей rd1/rd1. Вводимые клетки не наблюдаются в ONL или каком-либо месте, кроме как внутри кровеносных сосудов, или в непосредственной близости от них.
Функциональное восстановление сетчаток rd/rd, обработанных Lin - HSC
Электроретинограммы (ERG) получают на мышах через 2 месяца после введения контрольных клеток или мышиных Lin-HSC (фиг.17). Чтобы подтвердить наличие сосудистого и нейронного восстановления, после получения электроретинограмм проводят иммуногистохимический и микроскопический анализ каждого глаза. Типичные диаграммы ERG обработанных, восстановленных и контрольных, невосстановленных глаз, показывают, что после цифрового вычитания (обработанные глаза минус необработанные) продуцируется отчетливо детектируемый сигнал с амплитудой порядка 8-10 микровольт (фиг.17). Отчетливо видно, что сигналы, полученные от обоих глаз, сильно отклоняются от нормы. Однако соответствующие и детектируемые ERG получают от глаз, обработанных Lin-HSC. Во всех случаях ERG от контрольных глаз не детектируется. Хотя амплитуды сигналов в восстановленных глазах значительно ниже нормы, сигналы соответственно наблюдаются всегда, когда имеет место гистологическое восстановление, и имеют порядок величины, полученной в других, генетических исследованиях восстановления. В целом полученные результаты демонстрируют некоторую степень функционального восстановления в глазах, обработанных Lin-HSC.
К восстанавливаемым типам клеток сетчатки rd/rd преимущественно относятся колбочки
Восстановленные и невосстановленные сетчатки анализируют иммуногистохимическими методами с использованием антител, специфичных к опсину палочек или колбочек. Опсин палочек или колбочек анализируют в тех же глазах, для которых были получены диаграммы ERG, представленные на фиг.17. В сетчатках мышей дикого типа колбочки составляют менее 5% от присутствующих фоторецепторов (Soucy et al., 1998, Neuron 21: 481-493) и характер иммуногистохимического окрашивания с использованием красного/зеленого опсина колбочек, как показано на фиг.25 (A), или родопсина палочек, как показано на фиг.25 (B), согласуется с указанной долей колбочек. Если сетчатки дикого типа окрашивают преиммунным IgG, в участках чувствительной иннервации сетчаток не наблюдается окрашивание, отличающееся от аутофлуоресценции кровеносных сосудов (фиг.25 (C)). Через два месяца после рождения сетчатки необработанных мышей rd/rd имеют практически атрофированный внешний нуклеарный слой, который не окрашивается антителами против красно-зеленого опсина колбочек (фиг.25 (D)) или родопсина (фиг.25 (G)). В глазах, обработанных контрольными, CD31-, HSC, также отсутствует положительное окрашивание, свидетельствующее о присутствии опсина колбочек (фиг.25 (E))) или палочек (фиг.25 (H)). И наоборот, противоположные глаза, обработанные Lin-HSC, содержат приблизительно от 3 до 8 рядов ядер в сохраненном внешнем нуклеарном слое; большая часть данных клеток имеет положительное окрашивание по опсину колбочек (фиг.25 (F)) и примерно 1-3% положительного окрашивания по опсину палочек (фиг.25 (I)). Удивительно, что данное состояние почти противоположно исходному состоянию нормальной сетчатки мыши, в котором преобладают палочки. Полученные результаты показывают, что введение Lin-HSC сохраняет колбочки длительное время, в течение которого они дегенерируют в отсутствие данных клеток.
Lin - HSC, полученные из человеческого костного мозга (hBM), также восстанавливают дегенерирующие сетчатки
Lin-HSC, выделенные из костного мозга человека, ведут себя подобно мышиным Lin-HSC. Костный мозг собирают у доноров-людей и удаляют Lin+HSC, получая популяцию человеческих Lin-HSC (hLin-HSC). Полученные клетки метят ex vivo флюоресцентным красителем и вводят в глаза мышей C3SnSmn.CB17-Prkdc SCID. Введенные hLin-HSC мигрируют и направляются к участкам ангиогенеза сетчатки посредством механизма, идентичного наблюдаемому при введении мышиных Lin-HSC (фиг.18 (A)). Помимо направленности на сосуды человеческие Lin-HSC также оказывают интенсивное восстановительное действие на сосудистые и нейронные клеточные слои мышей rd1/rd1 (фиг.18 (B и C)). Данное наблюдение подтверждает присутствие в человеческом костном мозге клеток, направленных на сосудистую сеть сетчатки и способных предотвращать дегенерацию сетчатки.
Lin - HSC оказывают сосудо- и нейротрофическое действие на мышей rd10/rd10
Хотя мышь rd1/rd1 представляет собой наиболее широко используемую и лучше всего охарактеризованную модель дегенерации сетчатки (Chang et al. 2002, Vision Res. 42:517-525), данная дегенерация протекает очень быстро и в этом отношении отличается от обычной, более медленной дегенерации, наблюдаемой при заболевании человека. У данного штамма дегенерация фоторецепторных клеток начинается примерно на P8, когда сосудистая сеть сетчатки еще быстро растет (фиг.15). Последующая дегенерация глубокой сосудистой сети сетчатки происходит даже несмотря на то, что промежуточное сплетение еще формируется и, следовательно, сетчатки мышей rd1/rd1 никогда полностью не развиваются, в отличие от сетчаток большинства людей с данным заболеванием. Мышиную модель rd10, которая имеет более медленную дегенерацию и больше напоминает дегенеративное состояние человеческой сетчатки, используют для исследования Lin-HSC-опосредованного восстановления сосудов. У мыши rd10 дегенерация фоторецепторных клеток начинается примерно на P21, а дегенерация сосудов начинается вскоре после этого.
Поскольку нормальное нейросенсорное развитие сетчатки в основном завершается к дню P21, дегенерация начинается после завершения дифференциации сетчатки и с данной точки зрения является более сходной с дегенерацией человеческой сетчатки, чем дегенерация мышиной модели rd1/rd1. hLin-HSC или контрольные клетки, полученные из мышей rd10, вводят в глаза на P6 и затем сетчатки анализируют в разные моменты времени. На P21 сетчатки глаз, обработанных Lin-HSC и контрольными клетками, имеют нормальный вид с полным развитием всех сосудистых слоев и нормальным развитием INL и ONL (фиг.18 (D и H)). Примерно на P21 начинается дегенерация сетчатки, которая прогрессирует с возрастом. На P30 в сетчатках, обработанных контрольными клетками, наблюдается тяжелая сосудистая и нейронная дегенерация (фиг.18 (I)), тогда как сетчатки, обработанные Lin-HSC, сохраняют почти нормальные сосудистые слои и фоторецепторные клетки (фиг.18 (E)). Различие между восстановленными и невосстановленными глазами выражено сильнее в более поздние моменты времени (сравните фиг.18 (F и G) и 18 (J и K)). В глазах, обработанных контрольными клетками, прогрессирование сосудистой дегенерации отчетливо обнаруживается с помощью иммуногистохимического окрашивания антителами против CD31 и коллагена IV (фиг.18 (I-K)). Глаза, обработанные контрольными клетками, являются почти полностью отрицательными по CD31, но сохраняются коллаген IV-положительные сосудистые "треки", свидетельствуя о том, что скорее происходит восстановление сосудов, чем их неполное формирование. И наоборот, глаза, обработанные Lin-HSC, имеют и CD31- и коллаген IV-положительные сосуды, очень похожие на сосуды нормальных глаз дикого типа (сравните фиг.18 (F и I)).
Анализ экспрессии генов в сетчатках мышей rd/rd после обработки Lin - HSC
Чтобы идентифицировать предполагаемые медиаторы нейротрофического восстановления, проводят крупномасштабный геномный анализ (с использованием микрочипов) восстановленных и невосстановленных сетчаток. Экспрессию генов в сетчатках мышей rd1/rd1, обработанных Lin-HSC, сравнивают с экспрессией генов в необработанных сетчатках, а также в сетчатках, обработанных контрольными клетками (CD31-). Все сравнения проводят с тройными повторами. Считают, что гены присутствуют, если уровни их экспрессии превышают уровни фона во всех трех повторах, по меньшей мере, в 2 раза. Гены, экспрессия которых в Lin-HSC-защищенных сетчатках в 3 раза выше, чем в сетчатках, обработанных контрольными клетками, и в необработанных сетчатках мышей rd/rd, приведены на фиг.20, секция A и B. Коэффициент вариации (COV) для уровней экспрессированных генов рассчитывают путем деления стандартного отклонения на средний уровень экспрессии каждой реплицированной кРНК. Кроме того, рассчитывают корреляцию уровней экспрессии и шумовых отклонений путем сопоставления среднего значения и стандартного отклонения (SD). Корреляцию уровня экспрессии гена и стандартного отклонения рассчитывают для каждого гена, что позволяет определить фоновые уровни и порог достоверного уровня экспрессии. В целом полученные данные попадают в приемлемые пределы (Tu et al., 2002, Proc. Natl. Acad. Sci. USA 99: 14031-14036). Гены, которые отдельно обсуждаются ниже, имеют уровни экспрессии, превышающие указанные критические уровни экспрессии. Также определяют значения двустороннего критерия Стьюдента для рассматриваемых генов. Во всех случаях значения p являются приемлемыми (около или ниже 0,05) и демонстрируют, что существует подобие среди реплик и могут присутствовать значительные различия среди разных тестируемых групп. Многие гены с существенно повышенным уровнем экспрессии, в том числе MAD и Ying Yang-1 (YY-I) (Austen et al., 1997, Curr. Top. Microbiol. Immunol. 224: 123-130.), кодируют белки, участвующие в защите клеток от апоптоза. Ряд генов кристаллина, обладающих гомологией последовательности и функциями, подобными функциям известных белков теплового шока, участвующих в защите клеток от стресса, также подвергаются повышающей регуляции после обработки Lin-HSC. Иммуногистохимический анализ показывает, что экспрессия α-кристаллина локализована в ONL (фиг.19). На фиг.19 показано, что повышающая регуляция кристаллина αA наблюдается в клетках восстановленного внешнего нуклеарного слоя после обработки Lin-HSC, но не в противоположных глазах, обработанных контрольными клетками. В левой секции показано окрашивание IgG (контроль) в восстановленной сетчатке. В средней секции показан кристаллин αA в восстановленной сетчатке. В правой секции показан кристаллин αA в невосстановленной сетчатке.
Матричную РНК из сетчаток мышей rd1/rd1, восстановленных под действием человеческих Lin-HSC, гибридизуют со специфическим человеческим микрочипом Affymetrix U133A. В результате анализа, проведенного в жестких условиях, обнаружен ряд генов, экспрессирующих специфичную для человека мРНК на уровне, превышающем фоновый, который значительно выше в сетчатках, восстановленных под действием Lin-HSC, чем в сетчатках, восстановленных под действием мышиного Lin-HSC, и в невосстановленных сетчатках, обработанных человеческими контрольными клетками (фиг.20, секция C). С помощью биоинформационного метода с использованием микрочипов обнаружены ген CD6, молекулы клеточной адгезии, экспрессирующейся на поверхности недифференцированных и новодифференцированных гематопоэтических стволовых клеток CD34+, и ген интерферона альфа 13, также экспрессирующийся гематопоэтическими стволовыми клетками, подтверждающие результаты анализа. Кроме того, в образцах мышиной сетчатки, восстановленной под действием человеческого Lin-HSC, несколько факторов роста и нейротрофических факторов экспрессируются на уровне выше фонового (фиг.20, секция D).
Маркеры, связанные с линией дифференцировки гематопоэтических клеток, используют для отрицательной селекции популяции клеток Lin-HSC костного мозга, содержащих EPC. Хотя субпопуляция полученных из костного мозга Lin-HSC, которые могут служить в качестве EPC, не содержит традиционных маркеров клеточной поверхности, поведение данных клеток при развитии или повреждении сосудистой сети сетчатки полностью отличается от поведения Lin+ или популяций взрослых эндотелиальных клеток. Данные клетки селективно направлены на участки ангиогенеза сетчатки и участвуют в формировании проходимых кровеносных сосудов. Наследственные дегенеративные заболевания сетчаток зачастую связаны с утратой сосудистой сети сетчатки. Эффективное лечение таких заболеваний требует восстановления функции, а также сохранения сложной архитектуры ткани. Хотя в нескольких последних исследованиях изучается применение клеточной доставки трофических факторов или самих стволовых клеток, иногда необходимо использовать некоторые их сочетания. Например, применение терапии на основе факторов роста для лечения дегенеративных заболеваний сетчатки приводит к нерегулируемому повышенному росту кровеносных сосудов и, как следствие, к тяжелому нарушению нормальной архитектуры ткани сетчатки. Применение нервных или ретинальных стволовых клеток для лечения дегенеративных заболеваний сетчатки может способствовать восстановлению функции нейронов, однако для поддержания функциональной целостности сетчатки также необходима функциональная сосудистая сеть. Внедрение клеток из популяции Lin-HSC в сосуды сетчатки мышей rd/rd стабилизирует дегенеративную сосудистую сеть, не нарушая структуры сетчатки. Такой восстанавливающий эффект также наблюдается при введении клеток мышам rd/rd на P15. Поскольку дегенерация сосудов у мышей rd/rd начинается на P16, данное наблюдение расширяет терапевтическое окно для эффективной обработки Lin-HSC. В глазах, обработанных клетками Lin-HSC, сохраняются ретинальные нейроны и фоторецепторы и поддерживается зрительная функция.
Lin-HSC, полученные из костного мозга взрослых, оказывают сильное сосудо- и нейротрофическое действие при введении в стекловидное тело мышей с дегенеративным заболеванием сетчатки. Такой восстанавливающий эффект длится до 6 месяцев после обработки и является наиболее выраженным при введении Lin-HSC до полной дегенерации сетчатки (в течение 16 дней после рождения у мышей, у которых полная дегенерация сетчатки развивается к 30 дню после родов). Данное восстановление наблюдается у двух мышиных моделей дегенерации сетчатки и, в особенности, оно может осуществляться под действием HSC, полученных из костного мозга взрослых, если реципиент представляет собой иммунодефицитного грызуна с дегенерацией сетчатки (например, мышь SCID), или если донор представляет собой мышь с дегенерацией сетчатки. Хотя в некоторых последних исследованиях описано частичное фенотипическое восстановление у мышей или собак с дегенерацией сетчатки после вирусного спасения гена с помощью гена дикого типа (Ali, et al., 2000, Nat Genet 25:306-310; Takahashi et al., 1999, J. Virol. 73:7812-7816; Acland et al., 2001, Nat. Genet. 28:92-95.), настоящее изобретение впервые раскрывает клеточный терапевтический эффект, достигаемый путем восстановления сосудов. Таким образом, возможное применение такого способа для лечения группы заболеваний (например, пигментной дегенерации сетчатки), ассоциированных более чем со 100 известными мутациями, является более практичным, чем создание индивидуальных способов генной терапии для лечения каждой известной мутации.
Точная молекулярная основа эффекта нейротрофического восстановления остается неизвестной, однако он наблюдается только при наличии сопутствующих стабилизации/восстановления сосудов. Присутствие введенных стволовых клеток самих по себе не является достаточным условием для генерирования нейротрофического восстановления, и полное отсутствие во внешнем нуклеарном слое нейронов, образовавшихся из стволовых клеток, исключает возможность того, что введенные клетки будут трансформироваться в фоторецепторы. Результаты анализа экспрессии генов с использованием микрочипов демонстрируют значительную повышающую регуляцию генов, заведомо связанных с антиапоптотическими эффектами. Поскольку гибель нейронов, наблюдающаяся при дегенерации сетчатки, в основном происходит в результате апоптоза, такая защита может иметь большое терапевтическое значение для продления жизни фоторецепторов и других нейронов, играющих важную роль в поддержании зрительной функции при данных заболеваниях. C-myc представляет собой фактор транскрипции, который участвует в апоптозе путем повышающей регуляции разных нижестоящих апоптоз-индуцирующих факторов. У мышей rd/rd экспрессия C-myc в 4,5 раза выше, чем у мышей дикого типа, что указывает на возможность его участия в дегенерации фоторецепторов, наблюдающейся у мышей rd1/rd1. Известно, что гены Mad1 и YY-1, подвергающиеся интенсивной повышающей регуляции в Lin-HSC-защищенных сетчатках (фиг.20, секция A), подавляют активность c-myc, ингибируя c-myc-индуцированный апоптоз. Также показано, что сверхэкспрессия Mad1 подавляет Fas-индуцированную активацию каспазы-8, другого важного компонента апоптотического пути. Повышающая регуляция двух указанных молекул может играть роль в защите сетчатки от сосудистой и нейронной дегенерации путем предотвращения инициации апоптоза, который обычно приводит к дегенерации у мышей rd/rd.
Другой ряд генов, которые подвергаются интенсивной повышающей регуляции в Lin-HSC-защищенных сетчатках, включает членов семейства кристаллина (фиг.20, секция B). Подобно белкам теплового шока и другим стресс-индуцированным белкам кристаллины могут активироваться в результате стресса сетчатки и обеспечивать защиту от апоптоза. Аномально низкая экспрессия кристаллина αA коррелирует с утратой фоторецепторов у крысиной модели дистрофии сетчатки, а недавно проведенный протеомный анализ сетчатки мыши rd/rd демонстрирует индукцию повышающей регуляции кристаллина в ответ на дегенерацию сетчатки. На основе результатов анализа с использованием микрочипов, проведенного авторами настоящего изобретения на EPC-восстановленных сетчатках мышей rd/rd, можно сделать вывод, что повышающая регуляция кристаллинов играет ключевую роль в EPC-опосредованной нейропротекции сетчатки.
Такие гены, как c-myc, Mad1, Yx-1 и гены кристаллинов, вероятно являются нижестоящими медиаторами нейронного восстановления. Нейротрофические агенты могут регулировать экспрессию антиапоптотических генов, хотя анализ с использованием микрочипов, проведенный авторами настоящего изобретения на сетчатках, восстановленных мышиными стволовыми клетками, не показывает индукцию известных нейротрофических факторов на повышенных уровнях. С другой стороны, анализ восстановления, опосредованного стволовыми клетками костного мозга человека, с использованием специфичных для человека чипов, демонстрирует небольшое, но значимое увеличение экспрессии генов нескольких факторов роста.
Подвергающиеся повышающей регуляции гены включают несколько членов семейства факторов роста фибробластов и отоферлин. Мутации гена отоферлина связаны с генетическими нарушениями, приводящими к глухоте вследствие слуховой невропатии. Возможно, что продукция отоферлина введенными Lin-HSC также вносит вклад в предотвращение ретинальной невропатии. Исторически давно известно, что сосудистые изменения, наблюдающиеся у пациентов и животных с дегенерацией сетчатки, являются вторичными по отношению к пониженной метаболической потребности в гибели фоторецепторов. Результаты, полученные в настоящем изобретении, показывают, что, по меньшей мере, у мышей с наследственной дегенерацией сетчатки сохранение нормальной сосудистой сети может также помочь сохранить компоненты внешнего нуклеарного слоя. Последние публикации подтверждают концепцию, что тканеспецифичная сосудистая сеть имеет трофические эффекты, превышающие эффекты просто предоставления "питания" сосудами. Например, после индукции эндотелиальные клетки печени могут продуцировать, посредством активации VEGFR1, факторы роста, играющие важную роль в регенерации и поддержании гепатоцитов при поражении печени (LeCouter et al. 2003, Science 299:890- 893).
Описано, что подобные характерные взаимодействия между эндотелиальными клетками сосудов и соседними клетками паренхимы печени участвуют в органогенезе печени задолго до формирования функциональных кровеносных сосудов. Эндогенная ретинальная сосудистая сеть у субъектов с дегенерацией сетчатки может не вносить большой вклад в восстановление, однако если данную сосудистую сеть усилить эндогенными предшественниками, полученными из популяций гематопоэтических стволовых клеток костного мозга, это может сделать сосудистую сеть более устойчивой к дегенерации и в то же время обеспечить выживание нейронов и сосудистых клеток сетчатки. У людей с дегенерацией сетчатки задержка наступления полной дегенерации может привести к продлению способности видеть на несколько лет. У животных, обработанных Lin-HSC, наблюдается значительное сохранение ERG, которое может быть достаточным условием для поддержания зрения.
В клинической практике широко известно, что значительное уменьшение числа фоторецепторов и других нейронов может происходить при сохранении зрительной функции. Однако после преодоления критического порога происходит потеря зрения. Поскольку почти все наследственные заболевания сетчатки человека характеризуются ранним, но медленным началом, субъекта с дегенерацией сетчатки можно диагностировать и лечить путем введения в стекловидное тело трансплантата аутологичных стволовых клеток костного мозга по данному изобретению, с целью замедления дегенерации сетчатки и сопутствующей потери зрения. Для повышения направленности и внедрения стволовых клеток по данному изобретению желательно присутствие активированных астроцитов (Otani et al., 2002, Nat. Med. 8: 1004-1010); этого можно достичь путем ранней обработки, когда присутствует ассоциированный глиоз, или путем стимуляции локальной пролиферации активированных астроцитов с помощью лазера. Чтобы повысить восстанавливающий эффект, перед внутриглазным введением стволовые клеток можно трансфицировать ex vivo одним или несколькими нейротрофическими веществами. Данный подход можно использовать для лечения других нейронных дегенеративных нарушений зрения, таких как глаукома, которые характеризуются дегенерацией ганглионарных клеток сетчатки.
Популяции Lin-HSC из костного мозга взрослых содержат популяцию EPC, которая посредством направленности на активированные астроциты и внедрения в развитую матрицу может стимулировать ангиогенез, не нарушая структуру сетчатки. Lin-HSC, также обеспечивают длительный эффект нейротрофического восстановления в глазах с дегенерацией сетчатки. Кроме того, генетически модифицированные аутологичные композиции Lin-HSC, содержащие EPC, после трансплантации в ишемические или имеющие аномальную васкуляризацию глаза могут стабильно внедряться в новые сосуды и нейронные слои и непрерывно доставлять терапевтические молекулы в определенный участок в течение длительных периодов времени. Такая локальная доставка генов, которые экспрессируют фармакологические средства в физиологически значимых дозах, является новой концепцией лечения неизлечимых в настоящее время глазных заболеваний.
Фоторецепторы нормальной сетчатки мышей, например, преимущественно представляют собой палочки, однако после восстановления под действием Lin-HSC данного изобретения внешний нуклеарный слой содержит в основном колбочки. Большинство случаев наследственной дегенерации сетчатки человека обусловлены, в первую очередь, палочкоспецифичными дефектами, а утрата колбочек предположительно является вторичной по отношению к дисфункции палочек и, по всей вероятности, связана с уменьшением уровня некоторых трофических факторов, экспрессируемых в палочках.
ПРИМЕРЫ
Пример 1. Выделение и обогащение клеток; Получение популяций A и B мышиных Lin - HSC
Общий способ. Все анализы in vivo проводят в соответствии с Руководством Национального института здравоохранения по содержанию и применению лабораторных животных, а все процедуры, используемые в данных анализах, утверждены Комитетом по содержанию и применению животных научно-исследовательского института имени Скриппса (TSRI, La Jolla, CA). Клетки костного мозга извлекают из взрослых мышей B6.129S7-Gtrosa26, Tie-2GFP, ACTbEGFP, FVB/NJ (мыши rd/rd) или Balb/cBYJ (The Jackson Laboratory, ME).
Затем моноциты мышей разделяют в градиенте плотности полисахарозы HISTOPAQUE® (Sigma, St. Louis, MO), метят конъюгированными с биотином линиеспецифичными антителами (CD45, CD3, Ly-6G, CD11, TER-119, Pharmingen, San Diego, CA) и отбирают фракцию Lin-. Клетки положительной линии дифференцировки (Lin+) отделяют и удаляют из Lin-HSC, используя устройство для разделения в магнитном поле (сортер AUTOMACSTM, Miltenyi Biotech, Auburn, CA). Полученную популяцию Lin-HSC, содержащую предшественники эндотелиальных клеток, дополнительно характеризуют с помощью проточного цитометра FACSTM Calibur (Becton Dickinson, Franklin Lakes, NJ) с использованием следующих антител: PE-конъюгированный-Sca-1, c-kit, KDR и CD31 (Pharmingen, San Diego, CA). Клетки костного мозга Tie-2-GFP используют для характеристики Tie-2.
Чтобы собрать эндотелиальные клетки взрослых мышей, мезентериальную ткань хирургически удаляют из мыши ACTbEGFP, гидролизуют путем обрабатки коллагеназой (Worthington, Lakewood, NJ) и затем фильтруют через фильтр 45 мкм. Фильтрат собирают и инкубируют со средой для выращивания эндотелиальных клеток (Clonetics, San Diego, CA). Характеристики эндотелиальных клеток подтверждают путем наблюдения морфологического окрашивания mAb против CD31 (Pharmingen) и культуры анализируют на формирование трубчатых структур в матриксе MATRIGELTM (Beckton Dickinson, Franklin Lakes, NJ).
Популяция A мышиных Lin - HSC. Клетки костного мозга выделяют из мышей ACTbEGFP с помощью описанного выше общего способа. Клетки Lin-HSC анализируют методом проточной цитометрии FACS на антигенные маркеры клеточной поверхности CD31, c-kit, Sca-1, Flk-1 и Tie-2. Результаты показаны на фиг.1 (c). Примерно 81% Lin-HSC содержат маркер CD31, примерно 70,5% Lin-HSC содержат маркер c-kit, примерно 4% Lin-HSC содержат маркер Sca-1, примерно 2,2% Lin-HSC содержат маркер Flk-1 и примерно 0,91% клеток Lin-HSC содержат маркер Tie-2. И наоборот, Lin+HSC, выделенные из указанных клеток костного мозга, имеют значительно отличающийся профиль клеточных маркеров (т.е., CD31: 37,4%; c-kit: 20%; Sca-1: 2,8%; Flk-: 0,05%).
Популяция В мышиных Lin - HSC. Клетки костного мозга выделяют из мышей Balb/C, ACTbEGFP и C3H с помощью описанного выше общего способа. Клетки Lin-HSC анализируют на маркеры клеточной поверхности (Sca-1, Flk-1/KDR, c-kit (CD117), CD34, CD31 и разные интегрины: αl, α2, α3, α4, α5, α6, αL, αM αV, αX, α11b, β1, β2, β3, β4, β5 и β7). Результаты приведены в таблице 2.
|
Пример 2. Введение клеток в стекловидное тело мышиной модели
В веке мыши делают надрез тонким лезвием, чтобы обнажить глазное яблоко на P2 - P6. Популяцию А отрицательной линии дифференцировки HSC по настоящему изобретению (приблизительно 105 клеток примерно в 0,5-1 мкл среды для культивирования клеток) затем вводят в стекловидное тело с помощью шприца с иглой 33 калибра (Hamilton, Reno, NV).
Пример 3. Трансфекция EPC
Мышиные Lin-HSC (популяция A) трансфицируют ДНК, кодирующей фрагмент T2 TrpRS и содержащей маркер His6 (SEQ ID NO: 1, фиг.7), с использованием реагента для трансфекции FuGENETM6 (Roche, Indianapolis, IN) в соответствии с инструкцией производителя. Клетки Lin-HSC (примерно 106 клеток на мл) суспендируют в среде OPTI-MEM® (Invitrogen, Carlsbad, CA), содержащей фактор стволовых клеток (PeproTech, Rocky Hill, NJ). Затем добавляют смесь ДНК (примерно 1 мкг) и реагента FuGENE (примерно 3 мкл) и инкубируют приблизительно при 37°C в течение примерно 18 часов. После инкубации клетки промывают и собирают. Степень трансфекции в данной системе составляет приблизительно 17% по данным анализа FACS. Продукцию T2-TrpRS подтверждают методом вестерн-блоттинга. Аминокислотная последовательность His6-меченного T2-TrpRS показана в SEQ ID NO: 2, фиг.8.
Пример 4. Иммуногистохимия и конфокальный анализ
Из мышиных сетчаток, собранных в разные моменты времени, получают либо тотальные препараты, либо срезы после замораживания. Чтобы получить тотальные препараты, сетчатки фиксируют 4% параформальдегидом и затем блокируют 50% фетальной бычьей сывороткой (FBS) и 20% нормальной козлиной сывороткой в течение одного часа при комнатной температуре. Сетчатки обрабатывают первичными антителами и детектируют с использованием вторичных антител. В качестве первичных антител используют антитела против коллагена IV (Chemicon, Temecula, CA), против β-gal (Promega, Madison, WI), против GFAP (Dako Cytomation, Carpenteria, CA), против α-актина гладкой мускулатуры (α-SMA, Dako Cytomation). В качестве вторичных антител используют антитела, конъюгированные с флуоресцентными маркерами Alexa 488 или 594 (Molecular Probes, Eugene, OR). Изображения получают с использованием конфокального микроскопа MRC 1024 (Bio-Rad, Hercules, CA). С помощью программного обеспечения LASERSHARP® (Bio-Rad) создают трехмерные изображения, позволяющие анализировать три разных слоя развития сосудов в тотальном препарате сетчатки. Для получения 3-мерных изображений используют разницу в пиксельной интенсивности GFP у мышей с повышенным уровнем GFP (eGFP) и у мышей GFAP/wtGFP, определяемую методом конфокальной микроскопии.
Пример 5. Количественный анализ ангиогенеза в сетчатке мышей in vivo
Чтобы провести анализ T2-TrpRS, первичное и глубокое сплетения воссоздают на основе трехмерных изображений мышиных сетчаток. Первичное сплетение делят на две категории: нормальное развитие или остановленное развитие сосудов. Категории ингибирования развития глубоких сосудов интерпретируют на основе процента ингибирования развития сосудов, используя следующие критерии: полное ингибирование образования глубокого сплетения обозначают "полное", нормальное развитие сосудов (включающее в себя ингибирование менее 25%) обозначают "нормальное" и остальное обозначают "частичное." Чтобы получить данные по восстановлению для мышей rd/rd, с помощью линз 10× фиксируют четыре отдельных участка глубокого сплетения в каждом тотальном препарате сетчатки. Общую длину сосудистой сети рассчитывают для каждого изображения и суммируют, после чего сравнивают результаты, полученные в разных группах. Чтобы получить достоверную информацию, Lin-HSC вводят в один глаз, а Lin+HSC в другой глаз той же мыши. Необработанные контрольные сетчатки берут у мышей того же помета.
Пример 6. Мышиные модели повреждения сетчатки взрослых особей
Модели повреждения получают либо с помощью диодного лазера (150 мВ, 1 секунда, 50 мм), либо путем механического прокалывания мышиной сетчатки иглой 27 калибра. Через пять дней после повреждения в стекловидное тело вводят клетки. Глаза собирают у мышей еще через пять дней.
Пример 7. Нейротрофическое восстановление регенерации сетчатки
Полученные из костного мозга взрослых мышей гематопоэтические стволовые клетки отрицательной линии дифференцировки (Lin-HSC) оказывают сосудотрофическое и нейротрофическое восстанавливающее действие на мышиную модель дегенерации сетчатки. В стекловидное тело правых глаз мышей возрастом 10 дней вводят примерно 0,5 микролитров, содержащих приблизительно 105 Lin-HSC по настоящему изобретению, и затем через два месяца глаза анализируют на присутствие сосудистой сети сетчатки и количество ядер в нейронном слое. В качестве контроля в левые глаза тех же мышей вводят примерно такое же количество Lin+HSC и анализируют подобным способом. Как показано на фиг.9, в глазах, обработанных Lin-HSC, сосудистая сеть сетчатки выглядит почти нормальной, внутренний нуклеарный слой является почти нормальным, а внешний нуклеарный слой (ONL) имеет примерно от 3 до 4 слоев ядер. И наоборот, в противоположном глазу, обработанном Lin-HSC, средний сосудистый слой сетчатки в значительной степени атрофирован, внешний сосудистый слой сетчатки атрофирован полностью; внутренний нуклеарный слой в значительной степени атрофирован, а внешний нуклеарный слой атрофирован полностью. Это наглядно иллюстрируется у мыши 3 и мыши 5. У мыши 1 отсутствует эффект восстановления, и такое отсутствие наблюдается примерно у 15% обработанных мышей.
Если зрительную функцию анализируют с помощью электроретинограмм (ERG), возобновление положительных ERG наблюдается как при сосудистом, так и при нейронном восстановлении (мыши 3 и 5). В отсутствие сосудистого или нейронного восстановления положительные ERG не наблюдаются (мышь 1). Данную корреляцию между сосудо- и нейротрофическим восстановлением глаз мышей rd/rd под действием Lin-HSC по настоящему изобретению иллюстрирует полученный с использованием регрессионного анализа график, приведенный на фиг.10. Корреляция между нейронным (ось y) и сосудистым (ось x) восстановлением наблюдается в промежуточном (r=0,45) и глубоком сосудистом слое (r=0,67).
На фиг.11 показано отсутствие какой-либо статистически значимой корреляции между сосудистым и нейронным восстановлением под действием Lin+HSC. Результаты количественного анализа восстановления сосудов представлены на фиг.12. Приведенные на фиг.12 результаты, полученные для мышей через 1 месяц (1M), 2 месяца (2M) и 6 месяцев (6M) после введения, демонстрируют, что длина сосудов в глазах, обработанных Lin-HSC по настоящему изобретению (темные столбики), значительно выше, чем в необработанных глазах тех же мышей (светлые столбики), особенно, через 1 и 2 месяца после введения. Эффект нейротрофического восстановления количественно определяют путем подсчета ядер во внутренних и внешних нуклеарных слоях примерно через два месяца после введения Lin-HSC или Lin+HSC. Результаты представлены на фиг.13 и 14.
Пример 8. Популяция человеческих Lin - HSC
Клетки костного мозга выделяют из здоровых взрослых людей-добровольцев с помощью описанного выше общего способа. Затем моноциты разделяют в градиенте плотности полисахарозы, используя HISTOPAQUE® (Sigma, St. Louis, MO). Популяцию Lin-HSC выделяют из мононуклеарных клеток костного мозга с помощью системы для разделения в магнитном поле (сортер AUTOMACSTM, Miltenyi Biotech, Auburn, CA), используя следующие конъюгированные с биотином линиеспецифичные антитела: CD2, CD3, CD4, CD11a, Mac-1, CD14, CD16, CD19, CD33, CD38, CD45RA, CD64, CD68, CD86, CD235a (Pharmingen).
Человеческую популяцию Lin-HSC дополнительно разделяют на две субпопуляции в зависимости от экспрессии CD133. Клетки метят конъюгированными с биотином антителами против CD133 и разделяют на CD133-положительные и CD133-отрицательные субпопуляции.
Пример 9. Введение человеческих и мышиных клеток в стекловидные тела мышиных моделей дегенерации сетчатки
В качестве моделей дегенерации сетчатки используют штаммы мышей C3H/HeJ, C3SnSmn.CB17-Prkdc SCID и rd10. Мыши C3H/HeJ и C3SnSmn.CB17-Prkdc SCID (The Jackson Laboratory, Maine) являются гомозиготными по мутации дегенерации сетчатки 1 (rd1), которая обуславливает раннее развитие тяжелой дегенерации сетчатки. Данная мутация расположена в экзоне 7 гена Pde6b, кодирующего β-субъединицу фосфодиэстеразы цГМФ фоторецепторов-палочек. Мутация данного гена обнаружена у пациентов-людей с аутосомной рецессивной пигментной дегенерацией сетчатки (RP). Мыши C3SnSmn.CB17-Prkdc SCID, также являющиеся гомозиготными по спонтанной мутации тяжелого комбинированного иммунодефицита (Prkdc SCID), используются в экспериментах по переносу человеческих клеток. Дегенерация сетчатки у мышей rd10 вызывается мутацией в экзоне 13 гена Pde6b gene. Данные мыши также представляют собой клинически подходящую модель RP с более поздним развитием и более мягкой дегенерацией сетчатки, чем у rd1/rd1). Все анализы проводят в соответствии с Руководством Национального института здравоохранения по содержанию и применению лабораторных животных, а все процедуры, используемые в данных анализах, утверждены Комитетом по содержанию и применению животных научно-исследовательского института имени Скриппса.
В веке мыши делают надрез тонким лезвием, чтобы обнажить глазное яблоко P2 - P6. Мышиную популяцию А или человеческую популяцию С клеток HSC отрицательной линии дифференцировки настоящего изобретения (приблизительно 105 клеток примерно в 0,5-1 мкл среды для культивирования клеток) затем вводят в стекловидное тело глаз мышей с помощью шприца с иглой 33 калибра (Hamilton, Reno, NV). Чтобы визуализировать вводимые человеческие клетки, перед введением их метят красителем (зеленый краситель для мечения клеток CMFDA, Molecular Probes).
Сетчатки собирают в разные моменты времени, фиксируют 4% параформальдегидом (PFA) и метанолом и затем блокируют 50% FBS/20% NGS в течение одного часа при комнатной температуре. Чтобы окрасить сосудистую сеть, сетчатки инкубируют с антителами против CD31 (Pharmingen) и коллагена IV (Chemicon) и затем со вторичными антителами, конъюгированными с Alexa 488 или 594 (Molecular Probes, Eugene, Oregon). Сетчатки укладывают на плоскости, делая четыре радиальных расслабляющих надреза и получают тотальный препарат. Изображения сосудистой сети в промежуточном и глубоком плексиформных слоях сетчатки (см. Dorrell et al., 2002 Invest Ophthalmol. Vis. ScL 43:3500-3510) получают с помощью конфокального микроскопа Radiance MP2100 и программного обеспечения LASERSHARP® (Biorad, Hercules, California). Чтобы провести количественный анализ сосудистой сети, произвольно выбирают четыре независимых участка (900 мкм × 900 мкм) из средней части промежуточного или глубокого сосудистого слоя и измеряют общую длину сосудистой сети с помощью аналитической программы LASERPIX® (Biorad). Общую длину указанных четырех участков одного плексиформного слоя используют для следующего анализа.
Чтобы получить криостатные срезы, закрепленные на плоскости сетчатки подвергают повторному погружению. Сетчатки помещают в 4% PFA и держат в течение ночи, после чего инкубируют с 20% сахарозой. Сетчатки погружают в вещество с оптимальной температурой резания (OCT: Tissue-Tek; Sakura FineTech, Torrance, CA). Криостатные срезы (10 мкм) снова гидратируют в PBS, содержащем краситель для ядер DAPI (Sigma-Aldrich, St. Louis, Missouri). Изображения трех разных участков (шириной 280 мкм, объективная выборка), содержащих DAPI-меченные ядра, в одном срезе, который содержит диск зрительного нерва и целую периферическую сетчатку, получают с помощью конфокального микроскопа. Число ядер, расположенных в ONL трех независимых участков одного среза, подсчитывают, суммируют и используют для последующего анализа. Взаимозависимость длины сосудистой сети глубокого сплетения и числа клеточных ядер в ONL определяют с помощью простого линейно-регрессионного анализа.
После адаптации к темноте в течение ночи мышей анестезируют путем внутрибрюшинного введения 15 мкг/г кетамина и 7 мкг/г ксилазина. Электроретинограммы (ERG) регистрируют на поверхности роговицы каждого глаза после расширения зрачка (1% атропин сульфат), используя золотой петлевой электрод для роговицы наряду с электродом сравнения во рту и заземляющим электродом на хвосте. Импульсы получают с помощью фотостимулятора Grass (PS33 Plus, Grass Instruments, Quincy, MA), прикрепленного к внешней стороне купола Ganzfeld с высокой отражательной способностью. Регистрируют ответы палочек на коротковолновые (Реттен 47A; λmax=470 нм) вспышки света в интервале интенсивностей до максимально допустимой для фотостимулятора (0,668 кд·с/м2). Сигналы ответов усиливают (усилитель CP511 AC, Grass Instruments), преобразуют в цифровую форму (PCI-1200, National Instruments, Austin, TX) и анализируют с помощью компьютера. Каждая мышь служит собственным внутренним контролем, поскольку ERG регистрируют как на обработанных, так и на необработанных глазах. При слабых сигналах для получения средних значений используют до 100 разверток. Усредненные ответы от необработанных глаз после перевода в цифровую форму вычитают из ответов обработанных глаз и полученную разность сигналов используют для характеристики функционального восстановления.
С использованием микрочипов анализируют экспрессию Lin-HSC-таргетированных генов сетчатки. Мышам rd/rd на P6 вводят либо Lin-, либо CD31- HSC. Сетчатки данных мышей анализируют через 40 дней после введения в среде, не содержащей РНКаз (в данный момент времени происходит заметное восстановление сосудистой сети сетчатки и фоторецепторного слоя). Одну четверть каждой сетчатки анализируют как тотальный препарат, чтобы подтвердить достижение таргетирования на нормальные HSC, а также защиты сосудов и нейронов. РНК из сетчаток с успешным введением очищают, используя фенол/хлороформный метод выделения РНК TRIzol (Life Technologies, Rockville, MD). РНК гибридизуют с чипами Affymetrix Mu74Av2 и анализируют экспрессию генов, используя программное обеспечение GENESPRING® (SiliconGenetics, Redwood City, CA). Очищенные человеческие или мышиные HSC вводят в стекловидное тело мышей на P6. На P45 сетчатки анатомируют и объединяют в группы 1) обработанные человеческими HSC, восстановленные мышиные сетчатки, 2) обработанные человеческими HSC, невосстановленные мышиные сетчатки, и 3) обработанные мышиными HSC, восстановленные мышиные сетчатки, после чего РНК очищают и гибридизуют со специфичными для человека чипами U133A Affymetrix. Для идентификации генов уровень экспрессии которых превышает фоновый и является повышенным в сетчатках, восстановленных под действием человеческих HSC, используют программное обеспечение GENESPRING®. Затем профили экспрессии пар зондов анализируют отдельно для каждого из данных генов и сравнивают с результатами экспериментов, проводимых с использованием нормального человеческого микрочипа U133A и dChip, чтобы определить специфическую гибридизацию с человеческими последовательностями и устранить ошибочно положительные результаты, обусловленные межвидовой гибридизацией.
На фиг.21 показаны результаты проточной цитометрии, сравнивающие экспрессию поверхностных антигенов CD31 и интегрина альфа 6 на CD133-положительных (CD133+) и CD133-отрицательных (CD133-) популяциях человеческих Lin-HSC по настоящему изобретению. В левых секциях показаны диаграммы разброса для результатов проточной цитометрии. В центральных и правых секциях приведены гистограммы, демонстрирующие уровень экспрессии конкретного антитела на клеточной популяции. На оси Y откладывают число событий, а на оси X откладывают интенсивность сигнала. Обведенные гистограммы, полученные для контрольных антител изотипа IgG, отражают уровень неспецифического фонового окрашивания. Закрашенные гистограммы демонстрируют уровень экспрессии специфического антитела на клеточной популяции. Если закрашенная гистограмма сдвинута вправо по отношению к обведенной (контрольной) гистограмме, это означает, что произошло увеличение флуоресцентного сигнала, и что уровень экспрессии антитела выше фонового. Сравнение положения пиков закрашенных гистограмм двух клеточных популяций позволяет определить разницу в экспрессии белка на клетках данных популяций. Например, уровень экспрессии CD31 как в клетках CD133+, так и в клетках CD133- данного изобретения выше фонового; однако в популяции CD133+ больше клеток, экспрессирующих пониженные уровни CD31, чем в популяции CD133-. Из полученных результатов видно, что две популяции экспрессируют разные уровни CD31, и что интегрин альфа 6 экспрессируется в основном на клетках из популяции Lin- и, следовательно, может служить маркером клеток, выполняющих функцию сосудо- и нейротрофического восстановления.
Если в стекловидные тела глаз новорожденных мышей SCID вводят CD133-положительную и CD133-отрицательную субпопуляции Lin-HSC, наибольшая степень внедрения в развивающуюся сосудистую сеть наблюдается в CD133-отрицательной субпопуляции, которая экспрессирует оба поверхностных антигена CD31 и интегрин α6 (см. фиг.21, внизу). CD133-положительная субпопуляция, которая не экспрессирует CD31 или интегрин α6 (фиг.21, вверху), направлена на участки периферической обусловленной ишемией реваскуляризации, но не при введении в глаза, в которых протекает ангиогенез.
Восстановленные и невосстановленные сетчатки анализируют методом иммуногистохимии с использованием антител, специфичных к опсину палочек или колбочек. Анализ опсина палочек или колбочек проводят в глазах, для которых получали ERG, приведенные на фиг.17. В сетчатках мышей дикого типа колбочки составляют менее 5% от присутствующих фоторецепторов (Soucy et al. 1998, Neuron 21: 481-493) и результаты иммуногистохимического окрашивания с использованием красного/зеленого опсина колбочек, как показано на фиг.25 (A), или родопсина палочек, как показано на фиг.25 (B), подтверждают указанное содержание клеток колбочек. Антитела, специфичные к родопсину палочек (rho4D2), получают от Dr. Robert Molday, Университет Британской Колумбии, и используют в соответствии с описанной ранее методикой (Hicks et al., 1986, Exp. Eye Res. 42: 55-71). Кроличьи антитела, специфичные к красному/зеленому опсину колбочек, получают от Chemicon (AB5405) и используют в соответствии с инструкциями производителя.
Пример 10. Введение мышиных клеток в стекловидное тело мышиных моделей дегенерации сетчатки, индуцированной кислородом
Чтобы получить модель индуцированной кислородом дегенерации сетчатки (OIR), новорожденных мышей дикого типа C57B16 подвергают воздействию повышенного содержания кислорода (75% кислорода) в период от P7 до P12 после рождения. На фиг.22 показано нормальное постнатальное развитие сосудов у мышей C57B16 от P0 до P30. На P0 наблюдается только прорастание поверхностных сосудов в районе диска зрительного нерва. В течение нескольких следующих дней первичная поверхностная сеть распространяется в направлении к периферии, достигая отдаленных участков на день P10. Между P7 и P12 развивается вторичное (глубокое) сплетение. На P17 присутствует обширная поверхностная и глубокая сеть сосудов (фиг.22, вкладки). В последующие дни наряду с развитием третичного (промежуточного) слоя сосудов происходит ремоделирование и на P21 достигается образование взрослой структуры.
И наоборот, у описанной здесь модели OIR после воздействия 75% кислорода на P7-P12 происходит сильное нарушение нормальной последовательности событий (фиг.23). Популяции Lin-HSC взрослых мышей данного изобретения вводят на P3 в стекловидное тело глаза мыши, которую затем подвергают OIR, в другой глаз в качестве контроля вводят PBS или CD31-отрицательные клетки. На фиг.24 показано, что популяции Lin-HSC могут предотвращать дегенеративное действие высоких уровней кислорода на развивающуюся мышиную сетчатку. В обработанных глазах на Р17 наблюдается полностью развитая поверхностная и глубокая сосудистая сеть сетчатки, тогда как в контрольных глазах присутствуют большие участки, лишенные сосудов, и фактически отсутствуют глубокие сосуды (фиг.24). Анализируют примерно 100 глаз мышиных моделей OIR. Нормальную васкуляризацию наблюдают в 58% глаз, обработанных популяциями Lin-HSC, по сравнению с 12% контрольных глаз, обработанных клетками CD31-, и 3% контрольных глаз, обработанных PBS.
Пример 11. Выделение миелоподобных клеток костного мозга из костного мозга мыши путем селекции по CD44
Клетки костного мозга выделяют из взрослых мышей (The Jackson Laboratory, ME). Целый костный мозг обрабатывают мышиным антителом против CD44, после чего CD44-экспрессирующие клетки выделяют с помощью проточной цитометрии. Клетки отделяют от антитела и хранят в буферном растворе до последующего применения. Также выделяют популяцию клеток, которая не экспрессирует CD44 на значительном уровне (CD44LDBM).
Пример 12. Выделение миелоподобных клеток костного мозга из мышиного костного мозга путем селекции по CD44
Клетки костного мозга также получают в результате положительной селекции с использованием антитела против CD11b вместо CD44, как описано в примере 11. Выделяют популяцию миелоподобных клеток костного мозга, которая характеризуется как CD44HI и CD11b+ и обладает такими же параметрами активности, как и популяция CD44HI, выделенная по способу примера 11 с использованием CD44. Также выделяют популяцию CD44LD CD11b+, которая является неактивной.
Пример 13. Характеристика клеточных популяций MLBM
Хотя роль CD44 в данном контексте не ясна, возможно, данный рецептор опосредует выживание клеток, миграцию и/или дифференциацию клеток в обогащенной гиалуроновой кислотой среде стекловидного тела после введения клеток в глаз. В нефракционированном костном мозге мыши присутствуют разные популяции клеток CD44HI (т.е., MLBM) и CD44LD. Клеточная популяция MLBM составляет 76% популяции Lin-, используемой в предыдущих примерах, и приблизительно только 37% и 4%, соответственно, популяций клеток костного мозга Lin+ и CD31-/CD34-/CD11b-, экспрессирующих CD44 (фиг.26). Соответственно, существует превосходная корреляция между экспрессией CD44 и сосудотрофической и нейротрофической активностью трех указанных популяций, т.е. клетки Lin- являются наиболее эффективными, тогда как клетки CD31-/CD34-/CD11b-, соответственно, обладают наименьшей эффективностью. С помощью ряда линиеспецифичных антител определяют, что большая часть клеток CD44HI имеет строго миелоидные характеристики (фиг.27). Подобным образом, почти все клетки костного мозга CD44HI также характеризуются как CD11b+ (фиг.27).
MLBM, полученные в результате положительной селекции с использованием антитела против CD11b по способу примера 12 (CD44HI CD11b+), обладают такими же характеристиками активности, как и MLBM, выделенные в результате селекции с использованием антитела против CD44 в экспериментах по сосудистому таргетированию.
Характеристика полученных в примере 12 клеточной популяции MLBM и клеток CD44LD CD11b+ по антигенам клеточной поверхности приведена ниже в таблице 3. В таблице 3 увеличение числа символов плюс (+) соответствует относительно повышенной экспрессии антигена. Символ минус (-) означает, что экспрессия не детектируется.
|
|
Пример 14. Сосудотрофический и нейротрофический эффекты клеточной популяции MLBM
Клеточная популяция MLBM, полученная по способу примера 11, сохраняет свойства клеток Lin- в отношении направленности на сосуды и сосудо- и нейротрофического эффектов, тогда как клетки CD44LDBM обладают низкой активностью, или являются неактивными. Активность, связанную с направленностью на сосуды, демонстрируют путем введения клеток из популяции GFP+ MLBM в стекловидное тело мышей на 7 день после рождения (P7) с последующим анализом сетчаток на P14. После мечения кровеносных сосудов изолектином GS наблюдают направленность клеток GFP+ на сосудистую сеть сетчатки и их околососудистую локализацию, без очевидного внедрения. Данные события часто происходят при применении MLBM, но редко встречаются или отсутствуют в глазах, обработанных CD44LDBM (фиг.28).
Сосудо- и нейротрофическую активность клеточной популяции MLBM, полученной по способу примера 11, анализируют, используя мышиную модель дегенерации сетчатки, как описано выше для Lin-HSC. У мыши rd1/rd1 наблюдаются характерные признаки дегенеративного заболевания сетчатки, в том числе гибель фоторецепторов и атрофия глубокой сосудистой сети сетчатки. Как описано выше, клетки костного мозга Lin-HSC сохраняют глубокую сосудистую сеть сетчатки и частично восстанавливают фоторецепторы. Клеточная популяция MLBM по настоящему изобретению выполняет такую же функцию (фиг.29).
Модель индуцированной кислородом ретинопатии имеет признаки ретинопатии недоношенных. Патология, связанная с данной моделью, значительно уменьшается после обработки глаз клетками из популяции MLBM. Действие, оказываемое клетками популяции MLBM на данную модель, подобно действию, наблюдаемому при применении описанных выше Lin-HSC. В глазах, обработанных клетками популяции MLBM, наблюдается значительное уменьшение двух параметров, используемых для количественного определения степени патологии у данной модели: площади разрушения сосудов и площади пучков новообразованных сосудов. И наоборот, в глазах, обработанных клетками CD44LDBM, не наблюдается улучшение по сравнению с контрольными глазами, обработанными средой (фиг.30).
Помимо направленности на сосудистую сеть сетчатки клетки популяции MLBM дифференцируются в макрофагоподобные (F4/80+) клетки, проникают в сетчатку и занимают положение прямо напротив пигментного эпителия сетчатки (RPE). Такая локализация облегчает осуществление наблюдаемых эффектов восстановления сосудов и фоторецепторов клетками популяции MLBM. Далее, после того, как клетки популяции MLBM займут положение вблизи RPE, они начинают продуцировать фактор роста эндотелия сосудов (VEGF), как показано путем введения клеток популяции MLBM, полученных из мыши VEGF-GFP, в которой зеленый флуоресцентный белок (GFP) экспрессируется при активации гена VEGF (фиг.31). Таким образом, клетки популяции MLBM находятся в VEGF "активированном" состоянии. После введения клетки из популяции MLBM пополняют свои ряды эндогенными клетками того же типа, поскольку в районе RPE наблюдаются как GFP+ (введенные), так и GFP- (эндогенные) клетки. Такая локализация наблюдается у мышей дикого типа при нормальном развитии сосудов сетчатки, в восстановленных сетчатках мышей rd1/rd1 и у модели индуцированной кислородом ретинопатии.
Подобные характеристики сосудистого нацеливания получены и для клеточной популяции MLBM примера 12. На фиг.32 показано, что на P20 клетки CD44HI CD11b+ примера 12 (зеленые), введенные на P2 с помощью способа, подобного используемому в примере 11 для популяции с высоким содержанием CD44, специфически направлены на сосудистую сеть (красная). На фиг.33 показано, что CD44LD CD11b- примера 12 не обладают специфической направленностью на сосудистую сеть.
Клеточная популяция MLBM по настоящему изобретению обеспечивает эффективное и универсальное лечение глазных заболеваний. Клетки можно легко выделить из аутологичного костного мозга, минимизируя таким образом возможную иммуногенность, часто наблюдающуюся в способах лечения, основанных на применении клеток. Кроме того, клеточную популяцию MLBM по данному изобретению можно трансфицировать с целью доставки функциональных генов в сетчатку.
Пример 15. Дополнительная характеристика субпопуляций клеток костного мозга
Как описано в предыдущих примерах, все эксперименты проводят в соответствии с Руководством Национального института здравоохранения по содержанию и применению лабораторных животных, а все экспериментальные процедуры утверждены Комитетом по содержанию и применению животных TSRI. OIR индуцируют у мышей C57B16 в соответствии с описанным выше способом. На 7 день после родов детенышей и их матерей переводят с комнатной атмосферы на среду, содержащую 75% кислорода, и держат в ней 5 дней, после чего возвращают к комнатной атмосфере. Уровни кислорода регистрируют с помощью анализатора кислорода, утвержденного FDA (AX-300, Teledyne Analytical Instruments, CA, USA). В данных условиях в процессе воздействия повышенного содержания кислорода в центральной сетчатке образуются обширные участки с пониженным числом сосудов, а после возвращения к нормальному содержанию кислорода наблюдается аномальная преретинальная реваскуляризация, достигающая пика примерно на P17 с последующим рассасыванием (фиг.37, секции g-I, фиг.2, секции a, c).
Получение клеток: Выделение клеток костного мозга мышей, в основном, проводят следующим образом: клетки костного мозга получают из бедренной кости и большеберцовой кости мышей actGFP и обрабатывают двумя разными способами. В первом способе мононуклеарные клетки разделяют в градиенте плотности, используя FICO/LITE LM® (Atlanta Biologicals, Norcross, Georgia), и метят конъюгированными с биотином линиеспецифичными антителами (CD45R/B220, CD3e, Ly-6G/C, CD11b, Ter119, Pharmingen, San Diego, CA). Затем клетки инкубируют со стрептавидином или антибиотиновыми магнитными гранулами и сортируют, используя систему для клеточного сортинга MACS (Miltenyi Biotech, Auburn, CA), с получением популяций Lin-HSC. Во втором способе целый костный мозг инкубируют с антителом против CD44, конъюгированным с флуоресцентной меткой. Затем, используя метод клеточного сортинга с активацией флуоресценции (FACS), выделяют клетки CD44HI (т.е., клеточную популяцию MLBM, в которой большая часть клеток экспрессирует CD44) и клетки CD44LD (т.е., клеточную популяцию, в которой меньшая часть клеток экспрессирует CD44).
Характеристика клеток костного мозга: Дополнительный анализ клеточных субпопуляций, полученных с помощью вышеуказанных способов, проводят с использованием двух процедур: (1) двухцветной проточной цитометрии в сочетании с антителами против поверхностных маркеров линии дифференцировки и клеток-предшественников, включающих в себя CD11a, CD11b, Ly6G/C, CD43, F4/80, CD14, cKit, CD34, α6 интегрин и CD115 (поставляемыми Pharmingen, San Diego, CA); и (2) анализа экспрессии генов с использованием чипов AFFYMETRIX® Mu430 (Affymetrix, Santa Clara, CA) с помощью стандартных способов, известных в данной области. Для анализа экспрессии генов используют программное обеспечение GENESPRING® (Agilent Technologies, Palo Alto, CA).
Введение в стекловидное тело: В веке мыши делают надрез путем осторожного рассечения, чтобы обнажить глазное яблоко на P2-P7 (перед воздействием повышенного содержания кислорода). В стекловидное тело одного глаза каждого животного вводят примерно 150000-250000 клеток костного мозга в 0,5 мкл среды (PBS, содержащий 0,5% БСА и 2 мМ EDTA) с помощью шприца Hamilton и иглы 33 калибра (Hamilton, Reno, Nevada). В противоположный контрольный глаз вводят примерно такое же число контрольных клеток или только среду, в некоторых случаях введение совсем не производят, чтобы наблюдать естественное развитие заболевания. В экспериментальных подгруппах трансплантацию клеток осуществляют в более позднем возрасте, между P9 и P12.
Окрашивание сосудистой сети сетчатки: На P17 сетчатки собирают и визуализируют сосудистую сеть, чтобы определить местоположение и охарактеризовать введенные клетки. В некоторых случаях, чтобы визуализировать проходимые сосуды, перед препарированием сетчаток животных анестезируют и в сердце вводят меченный флуоресцеином высокомолекулярный декстран (FTTC Dextran, Sigma). В других случаях для окрашивания кровеносных сосудов и GFP-экспрессирующих клеток используют иммуногистологические методы. Сетчатки фиксируют в 4% растворе перфторуксусной кислоты (PFA) и метаноле, после чего блокируют 20% FBS/20% NGS в течение одного часа при комнатной температуре. Затем сетчатки инкубируют с изолектином GS-TB4, конъюгированным с ALEXA® 594, чтобы идентифицировать сосуды (Molecular Probes, Eugene, Oregon). Сетчатки укладывают на плоскости, делая четыре радиальных расслабляющих надреза, с получением тотальных препаратов, или погружают в OCT и делают поперечные криостатные срезы сетчатки, которые перед креплением подвергают контрастному окрашиванию с использованием DAPI.
Чтобы охарактеризовать трансплантированные клетки, в подгруппах глаз с помощью иммуногистологических методов идентифицируют следующие клеточные маркеры: F4/80 (Caltag, Burlingame, CA), CD44, CD31 (Pharmingen, San Diego, CA) и NG2 (Chemicon, Temecula, CA). Во всех сетчатках наблюдается тройное окрашивание: лектином, антителами против GFP и одного из вышеуказанных маркеров.
Визуализация и анализ изображения: Изображения сосудистой сети сетчатки получают с использованием сканирующего лазерного конфокального микроскопа RADIANCE® 2100MP (Biorad, Hercules, CA). Количественный анализ разрушения сосудов и реваскуляризации проводят следующим образом: Площадь разрушения сосудов измеряют путем тщательного определения контура бессосудистых зон в центральных участках сетчаток, окрашенных лектином GS, и вычисления общей площади с помощью программного обеспечения PHOTOSHOP® (Adobe) или VOLOCITY® (Improvision, Lexington MA). Подобным образом рассчитывают площадь преретинальной реваскуляризации ("пучки"), используя конфокальные изображения, сфокусированные на преретинальной плоскости и выбирая пучки на основе пиксельных интенсивностей (в пучках маркеры более яркие, чем в нормальной сосудистой сети). Затем выбранные участки суммируют, получая общую площадь реваскуляризации. Статистическое сравнение разных экспериментальных групп проводят с использованием критерия Стьюдента.
Трехмерные изображения сосудистой сети сетчатки и периваскулярных клеток костного мозга получают путем сбора z-серий конфокальных изображений и придания им объемности с помощью программного обеспечения VOLOCITY®. В результате на поперечном срезе можно увидеть сосуды сетчатки и определить положение трансплантированных клеток костного мозга относительно просвета сосуда.
Развитие сосудов сетчатки и мышиная модель индуцированной кислородом ретинопатии. Нормальное развитие сосудов сетчатки у новорожденных мышей, выращиваемых в условиях нормального содержания кислорода, показано на фиг.37, секции a-f. На 2 день после родов (P2) наблюдаются только развивающиеся поверхностные сосуды, занимающие одну плоскость вокруг оптического диска (фиг.37, секции a, b). В течение следующей недели первичная поверхностная сеть распространяется в направлении периферии, достигая отдаленных участков примерно на P12 (фиг.37, секция c). В интервале P7-P12 развивается вторичное (глубокое) сплетение (фиг.37, секция d). К концу первого месяца в полностью васкуляризованной сетчатке происходит ремоделирование (фиг.37, секция e) наряду с развитием третичного (промежуточного) слоя сосудов и завершается формирование взрослой структуры (фиг.37, секция f).
И наоборот, у модели OIR воздействие среды, содержащей 75% кислорода, в период P7-P12 приводит к тяжелому нарушению нормальной последовательности событий: происходит значительная регрессия сформировавшейся поверхностной сети сосудов в центральной сетчатке, особенно, вдоль артерий (фиг.37, секция g (P10) и секции h, i (P17)), а развитие глубокого сплетения сильно замедляется (фиг.37, секции k,m, поперечные срезы сетчатки на P17). После возвращения к условиям с нормальным содержанием кислорода на P12 снова начинается рост сосудов, но в аномальном режиме. По существу, теперь это условия с относительно пониженным содержанием кислорода для сетчатки со значительной степенью гиповаскуляризации. На P17 на периферии можно идентифицировать некоторые глубокие сосуды, однако аномальные преретинальные новообразованные пучки сосудов, ассоциированные с просачиванием внутрисосудистого красителя, можно видеть в средней части периферии, на границе между гиповаскуляризованной центральной сетчаткой и периферией с более высокой степенью васкуляризации (фиг.37, секция h). В течение последующих дней в бессосудистых областях поверхностные и глубокие сосуды развиваются медленно, однако присутствуют пучки новообразованных сосудов, выступающие над внутренней ограничительной мембраной (ILM) сетчатки в стекловидное тело, зачастую наблюдающиеся вплоть до P21 или даже позже. В период P25-P30 происходит ремоделирование сосудистой сети сетчатки и она становится похожа на нормальную сосудистую сеть данного возраста.
Введение предшественников гематопоэтических клеток перед воздействием повышенного содержания кислорода стимулирует восстановление сосудов сетчатки после индуцированного кислородом разрушения сосудов. Введение Lin-HSC данного изобретения на P2-P7 сильно изменяет способность сосудистой сети сетчатки к восстановлению после воздействия повышенного содержания кислорода (фиг.37, секции j,l,n,o, и фиг.38, секции b,d,e,f,g). Введение только среды не вызывает такие изменения. Более чем в 50% случаев на P17 в глазах, обработанных Lin-HSC, наблюдается полностью развитая поверхностная и глубокая сосудистая сеть сетчатки, тогда как в противоположных глазах, обработанных средой, присутствуют обширные бессосудистые участки и практически отсутствуют глубокие сосуды (фиг.37, секции l,n, по сравнению с секциями h,i,k,m, и с секцией o). В некоторых случаях, особенно, если повреждение противоположного контрольного глаза является очень тяжелым, восстановление глаз, обработанных клетками Lin-, не завершается на P17, однако в подавляющем большинстве случаев состояние данных глаз значительно лучше. Такое сравнение глаз одного животного дополнительно подтверждает эффективность Lin-HSC, фактически компенсирующих действие большинства других генетических факторов и факторов окружающей среды.
Разрушению сосудов в данной модели уделяется недостаточно внимания, поскольку большая часть исследований анализирует только формирование преретинальных пучков нововобразованных сосудов в ряде срезов сетчатки. Разрушение сосудов и образование пучков можно исследовать в одной сетчатке путем анализа цифрового изображения, полученного с помощью конфокального микроскопа (см., например, фиг.38, секции a-d). P17 выбирают как основную временную точку для анализа, поскольку в данный момент времени формирование пучков зачастую является максимальным, хотя в контрольных глазах еще присутствует значительное разрушение сосудов. Новый метод объединенного анализа позволяет установить существенную разницу между обработанными и контрольными глазами. Разрушение сосудов, измеренное на P17 в сетчатках, обработанных Lin-, значительно уменьшается (площадь разрушения уменьшается более чем на 75%) по сравнению с глазами, обработанными только средой, или с не обработанными глазами (фиг.38, секция e). В данном отношении различие между глазами, обработанными средой, и необработанными глазами не наблюдается. Подобным образом, в глазах, обработанных клетками Lin-, наблюдается примерно 70% уменьшение области пучков новых сосудов по сравнению с глазами, обработанными средой, и более чем 80% уменьшение по сравнению с необработанными контрольными глазами (фиг.38, секция f). Таким образом, обработка глаз Lin-HSC оказывает сильное влияние на два основных типа сосудистого повреждения и параметры восстановления мышиной модели OIR, т.е. одновременно уменьшает формирование пучков новообразованных сосудов и ускоряет "физиологическую" внутрисетчаточную реваскуляризацию.
Ускоренное восстановление также наблюдается, если обработку проводят в процессе гипероксии и при возвращении к нормоксии, однако эффект уменьшается. Эксперименты, описанные до настоящего времени, включают в себя инъекции, проводимые на дни P2-P7, перед воздействием гипероксии. Чтобы определить, действительно ли клетки Lin- могут также влиять на восстановление сосудов при более позднем введении, в течение фазы гипероксии цикла и при возвращении к нормоксии, введение осуществляют на P9, P11 или P12, после чего сетчатки анализируют в разные моменты времени. Результаты, приведенные на фиг.38, секция g, демонстрируют, что введение Lin-HSC приводит к эффективному ускорению восстановления сосудов и уменьшению области разрушения при введении в процессе гипероксии и на P12. Однако эффект немного ослабляется, указывая на то, что максимальная эффективность достигается, если обработку проводят перед воздействием среды с высоким содержанием кислорода.
После обработки предшественниками гематопоэтических клеток Lin- структура и функция сетчатки сохраняются в течение длительного времени. Также исследуются долговременные эффекты и возможные побочные эффекты Lin-HSC. Для этой цели 12 сетчаток выделяют из мышей возрастом 3-6 месяцев после введения клеток Lin- и воздействия повышенного содержания кислорода в соответствии с известной моделью (фиг.39). Во всех случаях опухоли отсутствуют и сохраняется гистология нервной ткани сетчатки. Единственной значительной аномальностью является встречающееся иногда образование "розеток" в сетчатке, которые также присутствуют в контрольных глазах (фиг.39, секции g,h). Сосудистая сеть сетчатки глаз, обработанных Lin-HSC, имеет нормальный вид, и очевидные отличия от контрольных необработанных сетчаток не обнаружены (фиг.39, секции a-f).
Также исследуют длительность существования трансплантированных клеток. Клетки GFP+ присутствуют только в небольшом проценте глаз (10%), свидетельствуя о том, что основная часть введенных клеток не выживает дольше нескольких месяцев. Выжившие клетки зачастую располагаются в непосредственной близости от сосудистой сети сетчатки. Электроретинограммы, полученные в период от 17 дней до 6 месяцев после трансплантации, показывают, что функция сетчатки глаз, обработанных Lin-HSC, не отличается от функции сетчатки нормальных контрольных глаз такого же возраста, не имеющих OIR. Чтобы определить, могут ли трансплантированные клетки существовать в глазах или распространяться системно, селезенки и/или печени, выделенные из 15 мышей, анализируют на присутствие клеток GFP+ в период приблизительно от 7 до 10 дней после введения. Вне глаз такие клетки не обнаружены.
Подтверждение активного клеточного типа: Популяцию Lin - обогащают клетками CD44 HI. Чтобы лучше понять механизмы, участвующие в данных процессах, и упростить процедуру селекции клеток, предпринимают попытку идентифицировать один маркер, который можно было бы использовать для выделения активных HSC из костного мозга. Анализируют широкий ряд кандидатов-маркеров клеток-предшественников костного мозга, выбранных на основе таких характеристик, как участие в миграции и дифференциации клеток. С помощью проточной цитометрии данные маркеры подвергают скринингу, сравнивая их экспрессию в активных клетках Lin- и в клетках BM, которые, как показано ранее, являются неактивными в ряде экспериментальных систем. Обнаружено, что CD44 по-разному экспрессируется в двух указанных популяциях: содержание клеток CD44HI в популяции Lin- (76%) значительно выше, чем в контрольной популяции клеток BM (4%) (фиг.40, секция a). Показано, что CD44, представляющий собой клеточный поверхностный рецептор гиалуроновой кислоты, участвует в регуляции нескольких клеточных функций, которые могут играть важную роль в опосредовании эффекта восстановления и включают в себя выживание, миграцию и дифференциацию. Распределение клеток CD44HI, а именно их высокое содержание в активной клеточной популяции и низкое содержание в контрольных клетках с пониженной активностью, указывает на то, что CD44 при необходимости может служить эффективным показателем активности.
Например, клетки CD44HI стимулируют восстановление сосудов у модели OIR, а клетки CD44LD не стимулируют. Эффективность клеток CD44HI подтверждается их способностью обеспечивать восстановление сосудов у модели OIR. С помощью экспериментальной схемы, используемой при введении клеток Lin-, показано, что клетки CD44HI стимулируют восстановление сосудов сетчатки у данной модели с эффективностью, подобной эффективности клеток Lin- (фиг.40, секции b, c). И наоборот, клетки CD44LD не оказывают положительного влияния на восстановление. Следует отметить, что в сетчатках животных, обработанных CD44LD, введенные клетки зачастую отсутствуют или присутствуют в небольших количествах, свидетельствуя о том, что данные клетки обладают пониженной способностью выживать в стекловидном теле и/или мигрировать в сетчатку. Неизвестно, являются ли клетки CD44HI единственной активной субпопуляцией костного мозга, или одной из нескольких субпопуляций, обладающих указанной активностью.
Клетки CD44 HI предположительно экспрессируют гены и маркеры миелоидного происхождения. Дальнейшую характеристику популяции CD44HI проводят путем крупномасштабного анализа экспрессии и путем мечения антителами против маркеров, специфичных для Lin- и клеток-предшественников, с последующим анализом методом проточной цитометрии (фиг.41 и фиг.44). Оба метода показывают, что клетки CD44HI имеют профиль экспрессии, соответствующий миелоидному происхождению. На этих клетках наблюдается интенсивная экспрессия CD11a, CD11b и Ly6G/C на уровне белка, тогда как с помощью проточной цитометрии детектируют менее интенсивное положительное окрашивание по F4/80, CD14, cKit и CD115. Анализ экспрессии показывает, что некоторые миелоид-специфичные гены, включающие в себя CD204, CD114, CD33 и CD115, экспрессируются на высоком уровне по сравнению с клетками CD44LD (фиг.44). И наоборот, на уровне белка популяция CD44LD экспрессируют на значительном уровне Ter119 и CD45R B220, которые являются маркерами эритробластов/эритроцитов и B-клеток, соответственно. С помощью экспрессионного чипа обнаружено, что в CD44LD по сравнению с клетками CD44HI интенсивно экспрессируется ряд генов, ассоциированных с лимфоцитами, которые включают в себя CD19, CD79a и CD22 (фиг.44). Таким образом, анализ на транскрипционном и белковом уровне позволяет идентифицировать активную популяцию CD44HI как имеющую исходно миелоидное происхождение, в то время как неактивные клетки CD44LD в основном являются лимфоидными.
Анализ трансплантированных клеток in situ - доказательство дифференциации: После более четкого определения популяции активных клеток костного мозга исследуют судьбу данных клеток после введения. Для этого сетчатки моделей OIR, обработанные CD44HI, анализируют методом иммуногистохимии с использованием разных маркеров. Подавляющее большинство введенных клеток направляется к сосудистой сети сетчатки и занимает периваскулярную локализацию, зачастую образуя удлиненные структуры, тесно связанные с сосудами хозяина (фиг.42, секция a). С использованием антител против CD31 и NG2 данные маркеры не удается обнаружить на GFP-экспрессирующих периваскулярных клетках костного мозга, это позволяет предположить, что данные клетки не дифференцируются в эндотелиальные клетки или перициты, соответственно. Кроме того, трансплантированные клетки не участвуют в формировании какой-либо части просвета сосуда (фиг.42, секция b), следовательно, вряд ли данные клетки дифференцируются в эндотелиальные клетки. И наоборот, многие, но не все периваскулярные клетки GFP+ в глазах, обработанных CD44HI, несут макрофагальный/микроглиальный маркер F4/80 (фиг.43, секции d-I). Такие введенные клетки F4/80+ имеют внешний вид, очень напоминающий эндогенные периваскулярные клетки, которые также содержат F4/80 (фиг.43, секции a-c), позволяя предположить, что трансплантированные клетки приобретают характерные особенности нативных клеток модели OIR.
Одно из возможных преимуществ клеточной терапии, особенно по сравнению с традиционным фармацевтическим способом лечения, заключается в способности клеток отвечать на локальные стимулы и подвергаться модификации при изменениях окружающей среды. Обнаружено, что на P17 (через 10 дней после введения) в трансплантированных клетках, направленных на сосудистую сеть сетчатки и занимающих периваскулярную локализацию, наблюдается снижение экспрессии CD44 до недетектируемых уровней (фиг.43, секции j-o). Однако в клетках, не связанных с сосудистой сетью, сохраняется экспрессия CD44. Таким образом, в субпопуляциях имплантированных клеток, которые изначально были отобраны методом FACS на основе высокой экспрессии CD44, наблюдается понижающая регуляция данного рецептора in-vivo, которая коррелирует с расположением клеток в сетчатке. Это позволяет предположить, что вводимые клетки фактически подвергаются селективным изменениям (дифференциации) в среде глаза.
Описанные выше результаты свидетельствуют о том, что клеточную терапию можно использовать для лечения ROP и других ишемических ретинопатий. Результаты, полученные на мышиной модели, показывают, что данный способ позволяет эффективно уменьшить сосудистую патологию, связанную с воздействием высокого содержания кислорода, и характеризуется низкой токсичностью или отсутствием токсичности. Преимущество клеточной терапии по сравнению с однофакторной терапией может заключаться в способности клеток адаптироваться к изменениям окружающей среды и отвечать на них. Развитие от однофакторных способов лечения до применения сочетаний лекарственных средств и вмешательств с целью отбора и доставки модифицированных адаптируемых клеток, которые могут контролировать и осуществлять сложную последовательность ответов при взаимодействии с тканью хозяина, является многообещающей новой концепцией. В данном контексте настоящее изобретение предлагает "смену парадигмы" в подходе к лечению ишемических ретинопатий/васкулопатий, т.е., оно делает упор на заживлении и стабилизации вместо подавления и разрушения.
Выделенные популяции миелоподобных клеток по данному изобретению, направленные на сосудистую сеть сетчатки, могут использоваться для доставки ангиостатических средств и оказывать сосудо- и нейротрофическое действие на модели дегенерации сетчатки. В настоящем исследовании конкретные субпопуляции клеточных популяций MLBM с высокой эффективностью ускоряют восстановление OIR. Интересно, что активные клетки экспрессируют маркеры, позволяющие предположить, что они имеют миелоидное происхождение и могут подвергаться дифференциации и модификации после трансплантации.
Клеточная терапия, стимулирующая васкуляризацию, впервые была использована в области кардиологии с целью восстановления пораженных инфарктом артерий. Значительное количество данных указывают на то, что некоторые клетки костного мозга эффективно улучшают кровоснабжение и сердечную функцию. Однако пока не ясно, какие типы клеток отвечают за наблюдаемые эффекты. Большое число исследований, изучающих потенциальную роль полученных из костного мозга предшественников эндотелиальных клеток (EPC), позволяют сделать вывод, что данные клетки присутствуют в новых или коллатеральных сосудах, однако небольшое число внедренных клеток, указанных в некоторых из этих исследований, вызывают вопросы относительно их значения. Кроме того, гетерогенные популяции костного мозга, такие как мононуклеарные клетки или нефракционированные клетки, которые содержат очень небольшое число стволовых клеток и/или EPC, также могут значительно повышать развитие коллатеральных сосудов, свидетельствуя о возможности присутствия других механизмов помимо непосредственного внедрения в сосуды. Без связи с какой-либо теорией, данные клетки могут играть поддерживающую, паракринную роль, в рамках которой факторы, секретируемые данными клетками, оптимизируют состояние сосудистой сети хозяина. Показано, что многие субпопуляции костного мозга являются источником ангиогенных факторов, а моноцитарные клетки, как известно, секретируют ряд таких факторов. Таким образом, существует вероятность того, что клетки костного мозга участвуют в паракринной регуляции, дополняя роль EPC в формировании коллатеральных сосудов и взаимодействуя с иммунной системой хозяина.
Хотя точные механизмы, участвующие в данной системе, пока не ясны, значительный прогресс достигнут в отношении понимания природы функциональных клеток костного мозга. Идентификация активной миелоидной популяции костного мозга, представленной клетками по настоящему изобретению, позволяет сделать некоторые предположения относительно механизма действия. Миелоидные клетки, особенно моноциты и макрофаги, обладают способностью влиять на рост кровеносных сосудов посредством секреции ангиогенных факторов роста. Кроме того, показано, что макрофаги являются более устойчивыми к гипоксии, чем клетки других типов, и отвечают на низкое содержание кислорода секрецией ангиогенных факторов. Следовательно, введение миелоидных предшественников в ишемические сетчатки предоставляет клетки, способные противостоять гипоксическим условиям и стимулировать восстановление сосудов в паракринной манере. Присутствие полученных из хозяина периваскулярных клеток F4/80+ в сетчатке OIR позволяет предположить, что данный тип клеток играет роль в данном процессе, и что доставка большого пула таких клеток (или их предшественников) путем непосредственной трансплантации в глаз может усиливать данный эффект. Данный сценарий разъясняет парадоксальное наблюдение, сделанное при проведении настоящих исследований, что введение клеточных популяций по настоящему изобретению стимулирует реваскуляризацию сетчатки и в то же время подавляет преретинальную реваскуляризацию. Хотя механизм этого явления полностью не известен, возможно, ускоренная "физиологическая" реваскуляризация может снижать гипоксию, которой подвергается сетчатка, и, как следствие, уменьшать стимулированное ишемией формирование пучков новообразованных сосудов.
Идея поддержки роста сосудов миелоподобными клетками может быть связана с более ранней работой, проведенной с использованием мышиных моделей дегенерации сетчатки rd1 и rd10. Введенные миелоидные предшественники могут поддерживать глубокую сосудистую сеть сетчатки посредством секретируемых факторов и предотвращать дегенерацию сосудов, наблюдаемую у данных моделей. Показано, что некоторые секретируемые макрофагами ангиогенные факторы, такие как bFGF, также могут обладать нейротрофической активностью. Таким образом, наблюдаемое уменьшение гибели фоторецепторов у мышей rd после введения клеточных популяций по настоящему изобретению может опосредоваться паракринным механизмом, в котором нейротрофические факторы продуцируются трансплантированными миелоидными клетками костного мозга. В подтверждение данного механизма настоящее исследование демонстрирует, что выделенные клеточные популяции MLBM данного изобретения могут восстанавливать сосуды и нейроны у модели rd с эффективностью, наблюдаемой при введении выделенных клеток MLBM.
Чтобы осуществить клиническое лечение ROP, клетки эмбриональной пуповинной крови собирают при рождении недоношенного ребенка высокого риска, затем клетки сортируют, обогащая их конкретной субпопуляцией, которая опосредует эффект восстановления, и затем полученные аутологичные клетки-предшественники можно ввести в глаз младенца.
Одно из существующих в настоящее время ограничений для применения клеточной терапии обусловлено тем, что во многих случаях точные молекулярные механизмы действия еще не ясны, и на самом деле у разных моделей эти механизмы могут быть разными. Однако в действительности это, т.е. способность отвечать на изменение условий и стимулов разными путями и широким набором сигналов, может являться самым большим преимуществом клеточных терапий. Такое разнообразие ответов может наблюдаться не только среди разных экспериментальных систем и задач, но и с течением времени в одной системе. Другими словами, такие клетки могут секретировать в один момент времени одни факторы, а впоследствии другие факторы, и, в конечном счете, если потребность в них уменьшается, секреция всех факторов может прекратиться. Существующая в настоящее время терапия на основе химических лекарственных средств не может достичь такого эффекта, основанного на том, что клетки функционируют по принципу обратной связи. Изменение клеточных маркеров в трансплантированных клетках in vivo, наблюдаемое в настоящем изобретении, подтверждает данную концепцию.
Пример 16. Клетки MLBM дифференцируются в клетки с микроглиальными характеристиками
Анализ сетчаток после введения клеток CD44HI показывает, что популяция CD44HI клеток костного мозга после введения в глаз дифференцируется в микроглию. Микроглия представляет собой резидентную миелоидную популяцию сетчатки и экспрессирует характеристические маркеры, включающие в себя CD11b и F4/80. Данные клетки также отличаются разветвленной (с ответвлениями) морфологией и имеют периваскулярную локализацию. Локализацию, морфологию и экспрессию поверхностных маркеров клеток CD44HI анализируют в разные моменты времени после введения в глаза. Обнаружено, что введенные клетки CD44HI GFP+ имеют все указанные характеристики эндогенной микроглии сетчатки (фиг.45). В секциях A и B на фиг.45 показано, что введенные клетки CD44HI экспрессируют CD11b и F4/80 и имеют такую же морфологию и периваскулярную локализацию, как и эндогенная микроглия. В секции C приведен анализ 3-мерных изображений, демонстрирующий, что введенные клетки CD44HI локализуются в периваскулярной области. В секции D показано изображение морфологии введенных клеток CD44HI с высоким увеличением.
Пример 17. Выделение клеток MLBM путем отрицательной селекции
Для экспериментальных и клинических применений желательно вводить клетки, которые не содержат связанных с поверхностью средств селекции, таких как антитела и/или магнитные гранулы. Одним из способов достижения этой цели является выделение клеток CD44HI с использованием отрицательной селекции. В результате характеристики профилей экспрессии поверхностных маркеров клеточных популяций CD44HI и CD44LD, описанных в данном документе, было обнаружено, что клетки CD44LD интенсивно экспрессируют Ter119 и CD45RB220, маркеры эритроидных клеток и B-клеток, соответственно. Антитела против данных маркеров, а также против T-клеточного маркера CD3e, эффективно метят популяцию CD44LD и позволяют удалить ее путем разделения в магнитном поле или методом FACS, оставляя "нетронутые" клетки CD44HI в качестве продукта. Клетки, полученные путем разделения методом FACS с использованием данной стратегии, имеют типичные функциональные характеристики клеточных популяций MLBM по настоящему изобретению (фиг.46).
На фиг.46, секция A, показано, что истощение мышиного костного мозга методом MACS с использованием антител, специфичных к CD45R/B220, Ter119 и CD3e, позволяет получить популяцию клеток, более 90 процентов которой составляют клетки CD44HI. В секции B показано, что отрицательная фракция (популяция CD44HI) практически не содержит клеток CD45R/B220, Ter119 и CD3e. В секции C показано, что полученные методом отрицательной селекции клетки CD44HI сохраняют способность к таргетированию и дифференциации.
Пример 18. Экспрессия индуцируемого гипоксией фактора 1α (HIF-1α) имеет большое значение для восстановления индуцированного гипоксией повреждения сосудов
HIF-1α является хорошо изученным модулятором клеточного ответа на низкое содержание кислорода и регулятором экспрессии ангиогенных генов, регулирующих транскрипцию ряда генов, которые потенциально участвуют в восстановлении сосудов и включают в себя VEGF, IGF, TGF-α и другие. Направленные делеции фактора транскрипции HIF-1α у мышей C57BL/6J получают путем вмешательства в события, предшествующие экспрессии cre, управляемой промотором лизозима M (lysMcre), что обеспечивает специфическую делецию фактора в миелоидной линии дифференцировки. Авторы настоящего изобретения выделяют миелоидные предшественники CD44HI из костного мозга указанных мышей, как описано в данном документе, и анализируют их способность стимулировать восстановление сосудов у модели OIR по сравнению с клетками CD44HI мышей дикого типа, при трансплантации на P7. Важно, что различия в экспрессии поверхностных маркеров или в светорассеивающих свойствах миелоид-специфичного штамма с нокаутом по HIF-1α и штамма дикого типа не обнаружены. Однако клетки, дефицитные по HIF-1α, не способны стимулировать восстановление, тогда как клетки дикого типа значительно ускоряют восстановление сосудов (фиг.47). Этот факт также указывает на то, что миелоидные клетки составляют активную популяцию, опосредующую восстановление сосудов, поскольку подавление экспрессии гена HIF-1α в данной модели нокаута является специфичным для клеток миелоидной линии. Все другие клетки костного мозга предположительно сохраняют нормальную активность.
Пример 19. Микроглия участвует в реваскуляризации сетчатки
Микроглия сетчатки исторически рассматривается как иммунокомпетентные клетки, которые отвечают на воспаление и инфекцию, осуществляя фагоцитоз осколков, образованных в результате ремоделирования при нормальном развитии или в процессе дегенеративного заболевания. Роль микроглии в стимулировании васкуляризации сетчатки окончательно не ясна. Настоящие исследования демонстрируют, клетки костного мозга взрослых, экспрессирующие поверхностные маркеры миелоидных предшественников, могут дифференцироваться в микроглию и обеспечивать ускоренное восстановление сосудистой сети после гипоксического повреждения. Данный процесс является HIF-1α-зависимым и описывает новую роль миелоидных предшественников в модуляции ангиогенеза.
Исходя из того, что трансплантированные миелоидные клетки-предшественники костного мозга дифференцируются в микроглию у модели OIR и стимулируют реваскуляризацию и восстановление центральной сетчатки, можно сделать вывод, что микроглия играет важную роль в стимуляции и поддержании васкуляризации сетчатки при нормальном развитии. Для получения модели OIR чаще всего используют мышиный штамм C57BL/6J, поскольку у него стабильно наблюдается значительное разрушение сосудов и формирование преретинальных сосудистых пучков. И наоборот, у штамма BALB/cByJ в тех же условиях не наблюдается образование преретинальных пучков на детектируемом уровне и после возвращения к нормоксии очень быстро происходит реваскуляризация центральных областей разрушения сосудов, образовавшихся в период гипероксии (фиг.48, секция a). Поскольку причина таких различий не ясна, у обоих штаммов анализируют микроглию сетчатки. Обнаружено, что у мышей BALB/cByJ микроглия присутствует в участках разрушения сосудов центральной сетчатки после 48 часов ишемии/гипоксии в более высоком количестве, чем у мышей C57BL/6J (фиг.48, секции b, c). Данный результат показывает, что повышенное содержание эндогенной микроглии сетчатки в фазе восстановления частично защищает штамм BALB/cByJ. Без связи с какой-либо теорией, миелоидные предшественники костного мозга, трансплантированные мышам C57BL/6J, могут заменить и/или пополнить эндогенную микроглию, присутствующую в данном штамме в пониженном количестве, обеспечивая эффект восстановления, подобный наблюдающемуся у необработанных мышей BALB/cByJ.
Чтобы дополнительно охарактеризовать механизмы, участвующие в восстановлении сосудов у модели OIR, исследуют роль микроглии в нормальном развитии сосудов сетчатки. Для этого изменяют число микроглиальных клеток, используя липосомы, нагруженные клодронатом, которые селективно поглощаются фагоцитирующими клетками, такими как макрофаги и микроглия, и индуцируют у них апоптоз. Специфичность микроглиального поглощения в данных экспериментах демонстрируют с использованием меченных липосом, которые через четыре дня после введения локализуются исключительно совместно с микроглией CD11b+ (фиг.48, секция e, вкладка). В клетках сосудах или других клетках CD11b- меченное вещество не обнаружено. Введение в стекловидное тело нагруженных клодронатом липосом на P5 приводит к значительному уменьшению микроглиальных клеток и сильному разрушению сосудистой сети сетчатки на P8. Наблюдаемое исчезновение крупных капилляров анатомически коррелирует с участками опосредованной клодронатом гибели микроглиальных клеток, указывая на то, что микроглия необходима для развития и/или поддержания новых сосудов (фиг.48, секция d).
Подобным образом, введение нагруженных клодронатом липосом на P2 приводит к 28% уменьшению области васкуляризованной сетчатки (n=6) на P6 по сравнению с противоположными глазами, обработанными контрольными липосомами, нагруженными PBS (фиг.48, секция e). Вследствие очевидного различия в плотности сосудистых сетей, также сравнивают общую площадь сосудов, которая уменьшается на 41% после обработки липосомами с клодронатом. Полученные результаты показывают, что микроглия играет важную роль в поддержании незрелых сосудов, а также в росте новых сосудов в процессе развития сетчатки.
Пример 20. Человеческие клетки MLBM CD44 HI
Данные, полученные для популяции миелоподобных клеток костного мозга, также можно распространить на человеческие клетки.
Популяцию миелоидных клеток выделяют из костного мозга человека. Выделенные клетки представляют собой клетки, экспрессирующие CD44HI, которые также экспрессируют CD11a, CD11b, CD11c, CD14, CD33 и CD46 (фиг.49 и 51). Человеческие клетки CD44HI имеют физические свойства, такие как светорассеяние при проточной цитометрии, подобные свойствам мышиных клеток CD44HI (фиг.50). Например, клетки экспрессируют и CD44, и CD11b. Человеческая популяция CD44HI отличается от мышиной тем, что часть человеческих клеток CD44HI составляют лимфоциты, тогда как клетки CD44HI мышиного костного мозга являются исключительно миелоидными. Чтобы получить популяцию человеческих клеток, максимально подобные описанным ранее мышиным клеткам, популяцию CD44HI выделяют из костного мозга человека, истощенного по лимфоцитам. Полученные клетки костного мозга человека эффективно уменьшают патологию сосудов у мышиной модели OIR (фиг.52).
Затем определяют, присутствуют ли в периферической крови активные клетки. Вначале из периферической крови выделяют моноциты, используя антитело против CD14. Данные клетки выбирают, поскольку основным компонентом клеток CD44HI костного мозга являются моноциты. Миелоидные клетки выделяют из костного мозга с помощью антитела против. Анализ человеческой периферической крови на антигены, экспрессирующиеся совместно с CD44, проводят после лизиса красных кровяных клеток (rbc) (фиг.53). Обе клеточные популяции CD44HI, полученная путем положительной селекции по CD14, и полученная путем положительной селекции по CD33, являются эффективными в отношении мышиной модели OIR (фиг.54). Данные клетки также экспрессируют CD11b.
Используют способ селекции на основе разных физических свойств клеток. Для отбора конкретных клеточных популяций можно использовать проточную цитометрию, которая позволяет измерять размер клеток и зернистость клеток. В данном случае, после лизиса красных кровяных клеток хлоридом аммония, моноциты и гранулоциты отбирают только по светорассеянию, не применяя никаких средств селекции (т.е., антител). С помощью данного способа из периферической крови можно получить популяцию клеток, подобных CD44HI (которая содержит в качестве основных компонентов моноциты и гранулоциты) (фиг.55). Обнаружено, что данные клетки уменьшают разрушение сосудов и реваскуляризацию у мышиной модели OIR. Моноциты и гранулоциты, выделенные по отдельности, также обладают эффективностью в отношении модели also OIR (фиг.56).
Пример 21. Клетки пуповинной крови человека CD14 +
Пуповинная кровь человека является богатым источником гематопоэтических колониеобразующих клеток или стволовых клеток (HCFC или HSC). Некоторые клинические исследования демонстрируют, что кровь из пуповины и плаценты содержит количества стволовых клеток, достаточные для восстановления пораженного излучением костного мозга.
Предшественники эндотелиальных клеток (EPC), полученные из костного мозга, играют важную роль в реваскуляризации ишемических тканей и в реэндотелизации поврежденных кровеносных сосудов.
Авторы настоящего изобретения обнаружили, что гематопоэтические клетки-предшественники пуповинной крови человека содержат одну или несколько популяций, способных внедряться в сосудистую сеть сетчатки в участках ишемии и/или дегенерации и участвовать в восстановлении функциональных кровеносных сосудов. Большая часть клеток экспрессирует CD44 и примерно 97% клеток экспрессирует CD11b. Данные клетки, а также их эндотелиальное потомство, выращенное in vitro, вводят в стекловидное тело иммунодефицитных мышей SCID, страдающих от индуцированной кислородом ишемии. Мононуклеарные клетки пуповинной крови человека получают методом центрифугирования в градиенте плотности Ficoll. Неотселектированные мононуклеарные клетки пуповинной крови человека выращивают на культуре, покрытой фибронектином.
После культивирования in vitro в течение четырех дней появляются колонии с морфологией эндотелиальных клеток (EC). Колония эндотелиальных клеток-предшественников представляет собой совокупность тонких плоских клеток, выделяющуюся из центрального кластера клеток круглой формы.
На 7 день EPC, культивируемые в контрольной среде, имеют морфологию колоний эндотелиальных клеток, клетки которых имеют тонкую плоскую веретенообразную форму и выделяются из кластера круглых/полигональных клеток. Удлиненные эндотелиальные клетки образуют либо рассеянные колонии, либо плотно упакованные кипы. На 13 день культивируемые EPC подобным образом дифференцируются в EC (фиг.57).
Чтобы подтвердить эндотелиальный профиль, проводят непрямое иммуноокрашивание, используя антитела против рецептора 2 (VEGFR2) фактора роста эндотелия сосудов (VEGF) и CD31. Ядра окрашивают 4'-6-диамидино-2-фенилиндол-2HCl (фиг.58, 59).
В стекловидное тело мышиной модели OIR вводят свежеполученные клетки пуповинной крови или клетки пуповинной крови человека после дифференцировки в культуре в течение семи дней. После введения свежеполученных мононуклеарных клеток пуповинной крови (CBMN) или клеток, дифференцированных в культуре в течение семи дней, анализируют участки разрушения сосудов (желтые) и пучки новообразованных сосудов по сравнению с обработкой PBS (фиг.60 и 61). Свежие клетки CBMN идентифицируют с использованием лентивирусной экспрессии eGFP (фиг.62)
В мононуклеарных клетках пуповинной крови человека идентифицируют популяцию моноцитов, экспрессирующих CD14. Свежие CD14-положительные клетки, выделенные методом MACS (чистота 98%), вводят в стекловидное тело мышиной модели OIR. Авторы настоящего изобретения анализируют участки разрушения сосудов и образование пучков. Как показано на фиг.63, введенные клетки очевидно индуцируют восстановление сосудов по сравнению с PBS.
Многочисленные вариации и модификации описанных выше воплощений можно осуществить без отступления от сущности и объема новых признаков данного изобретения. Предполагается, или подразумевается, что описанные здесь конкретные воплощения никак не ограничиваются.

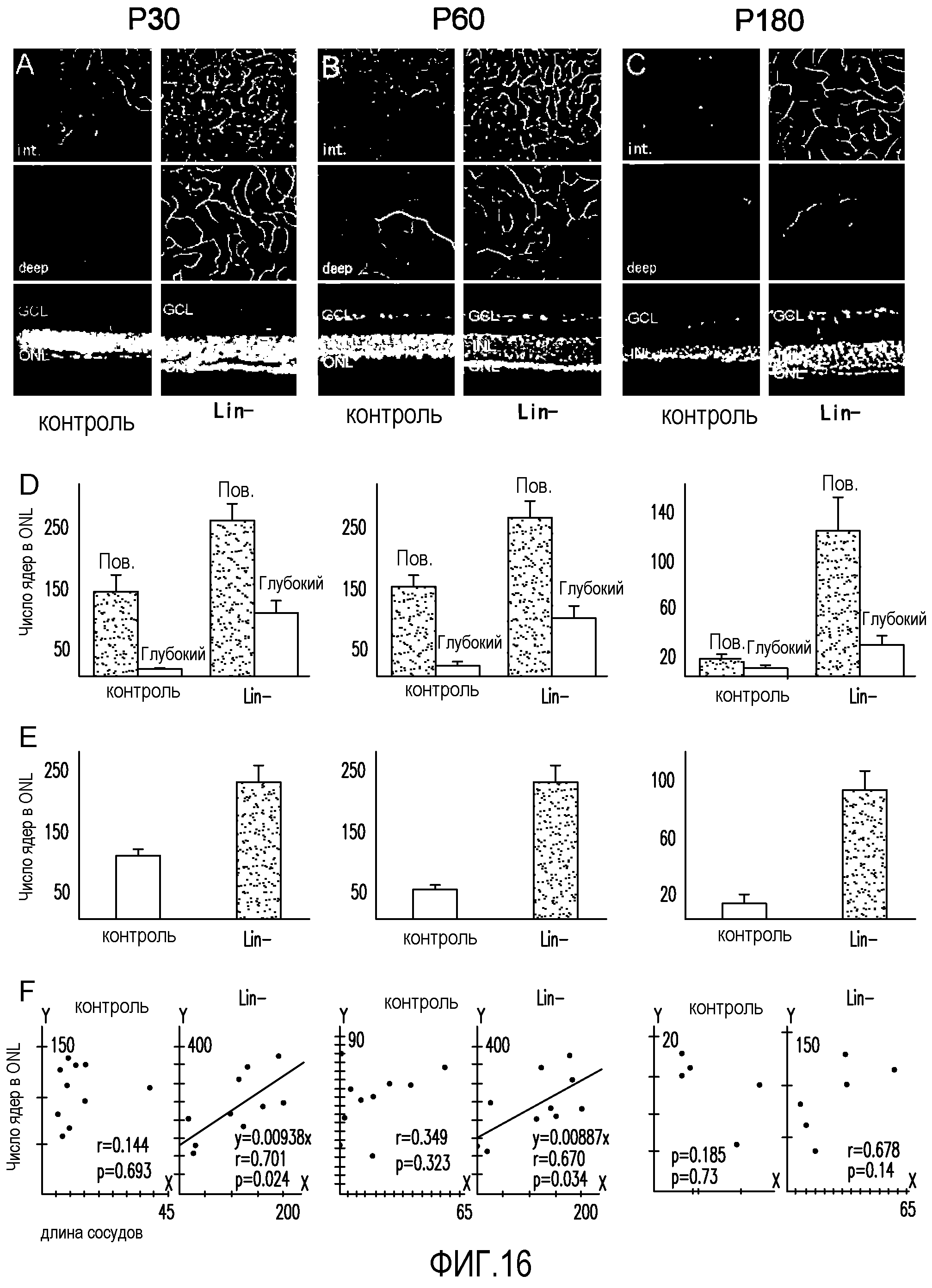

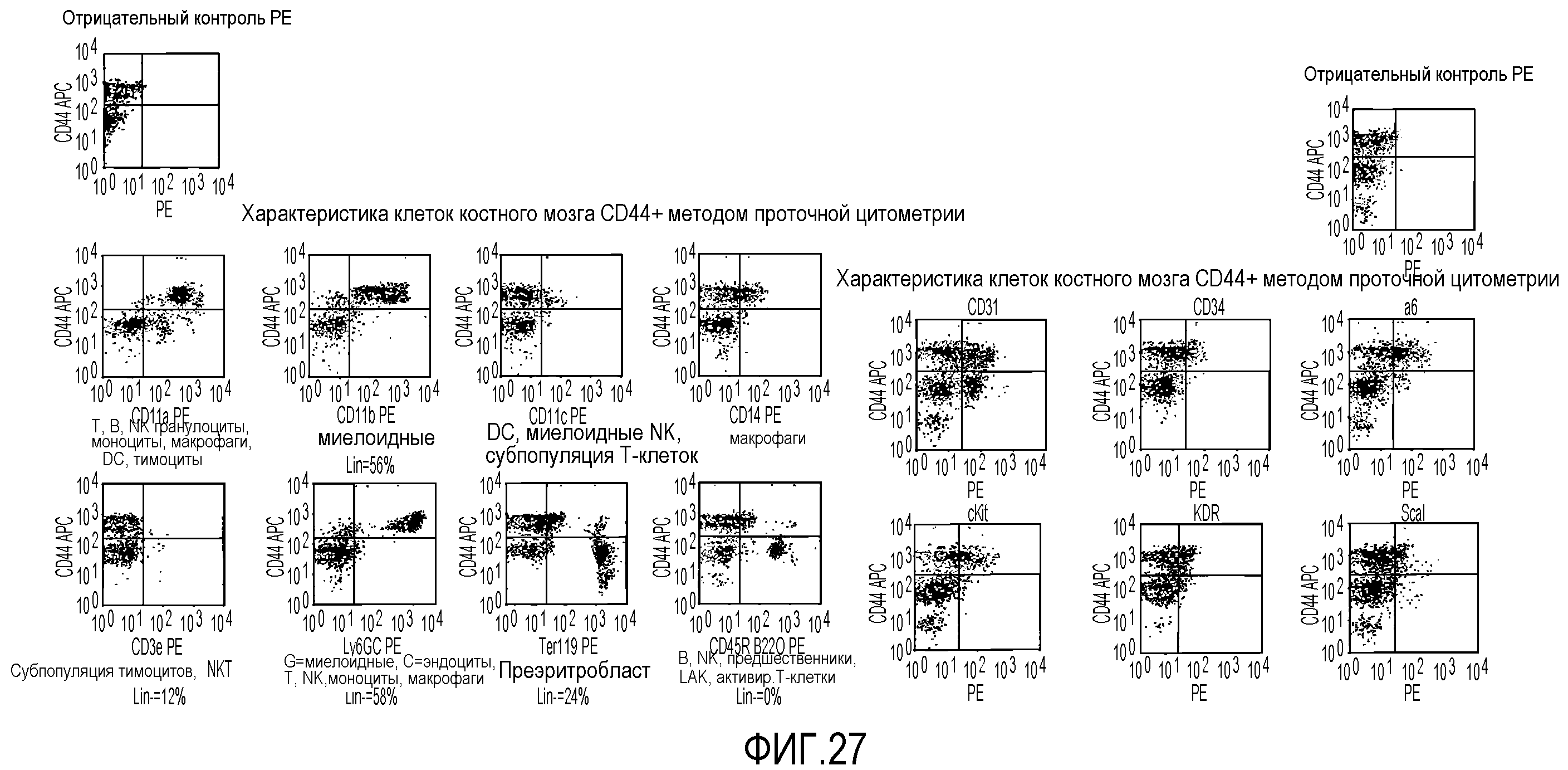



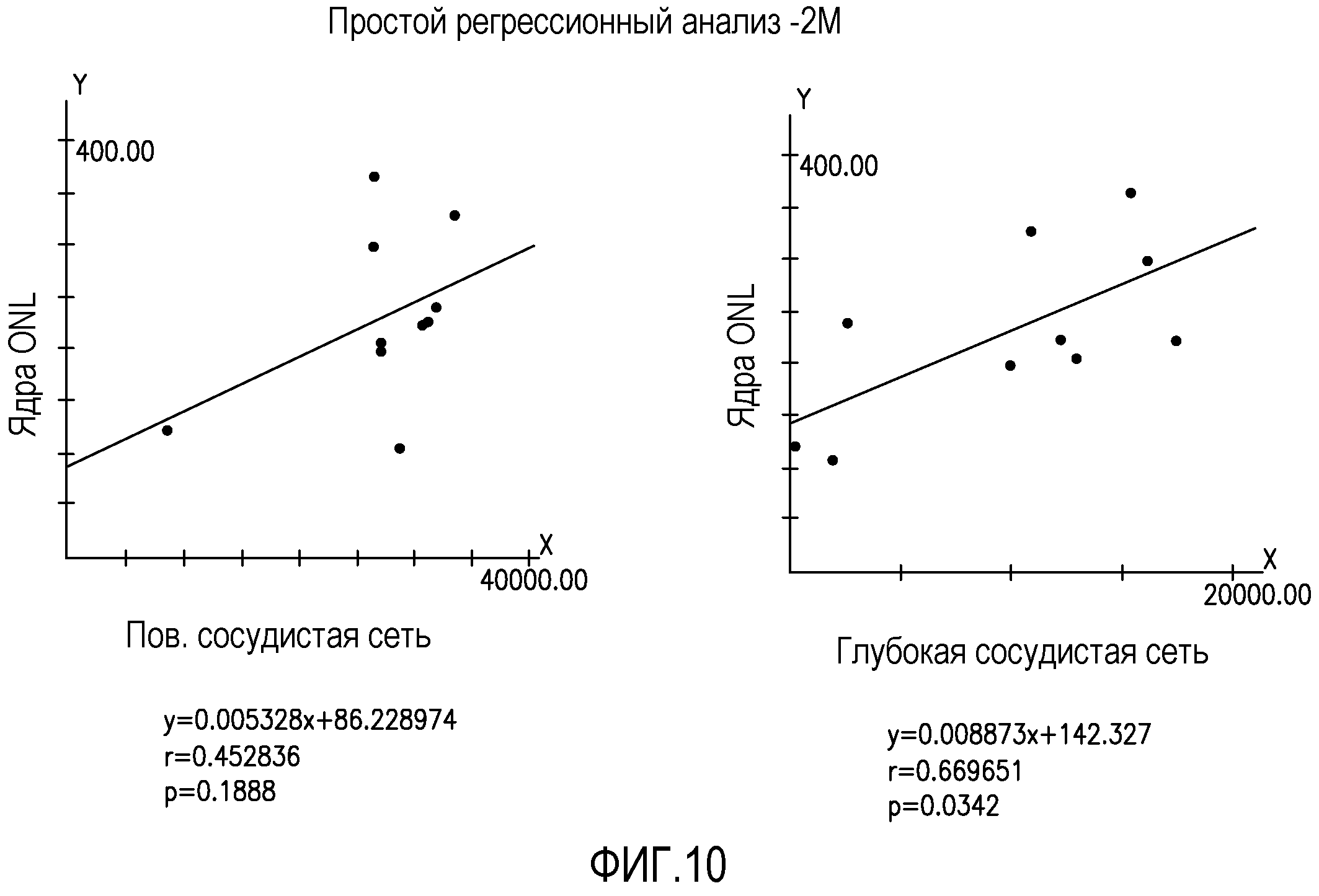
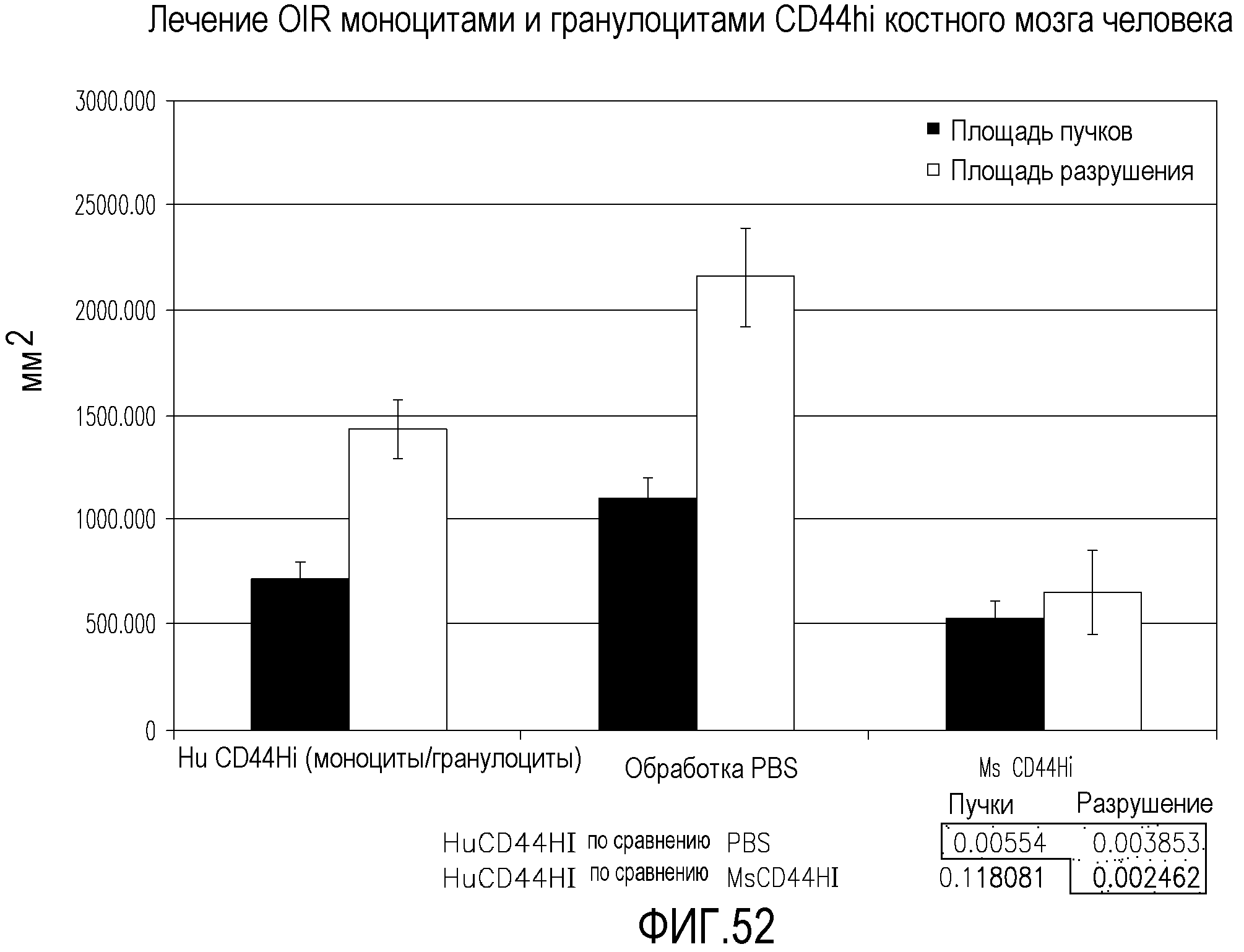

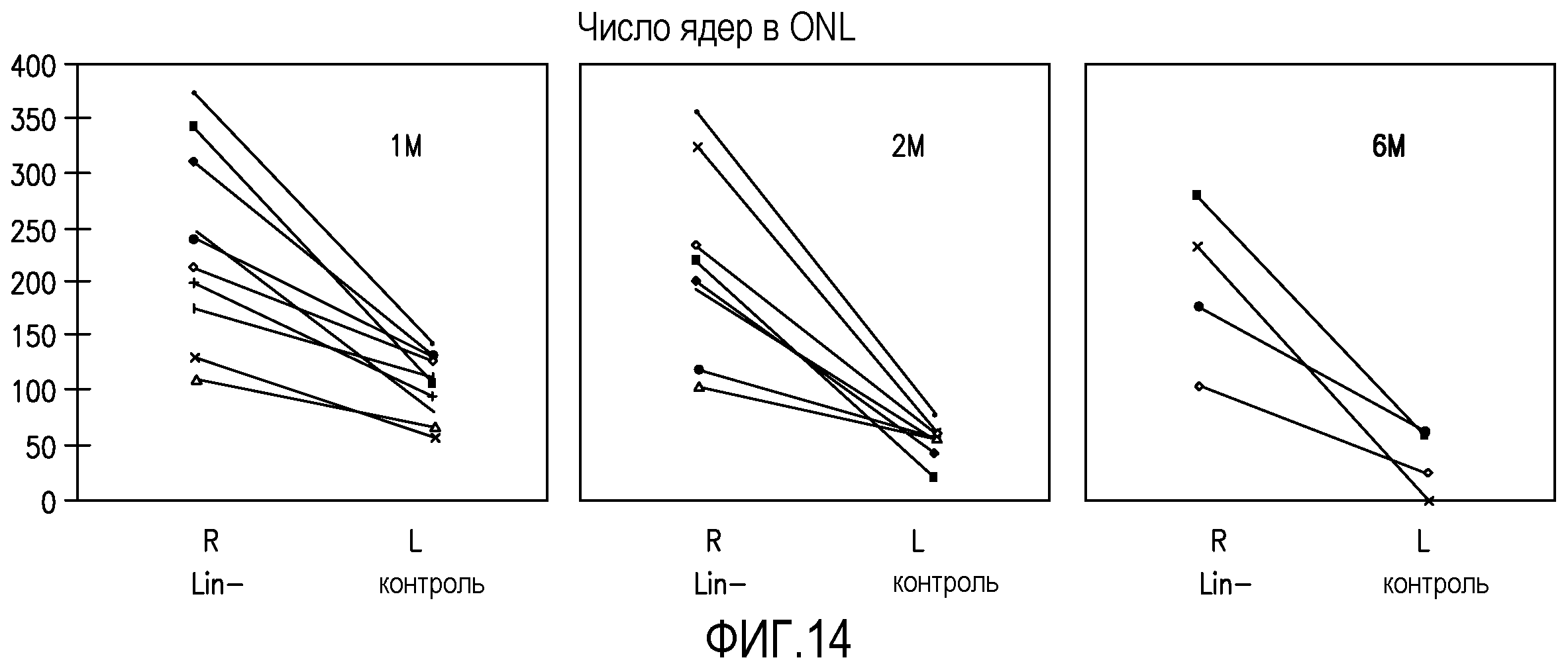

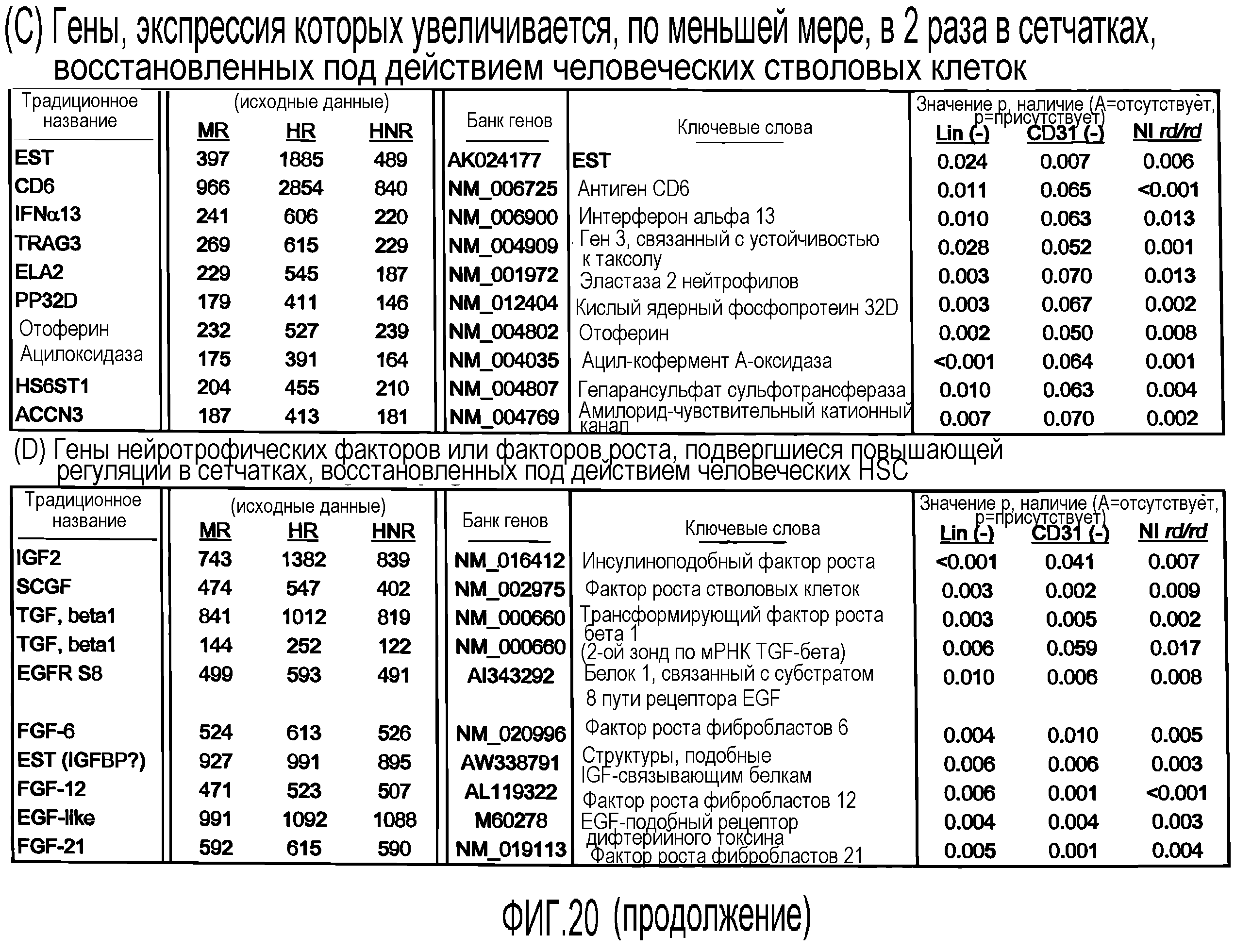

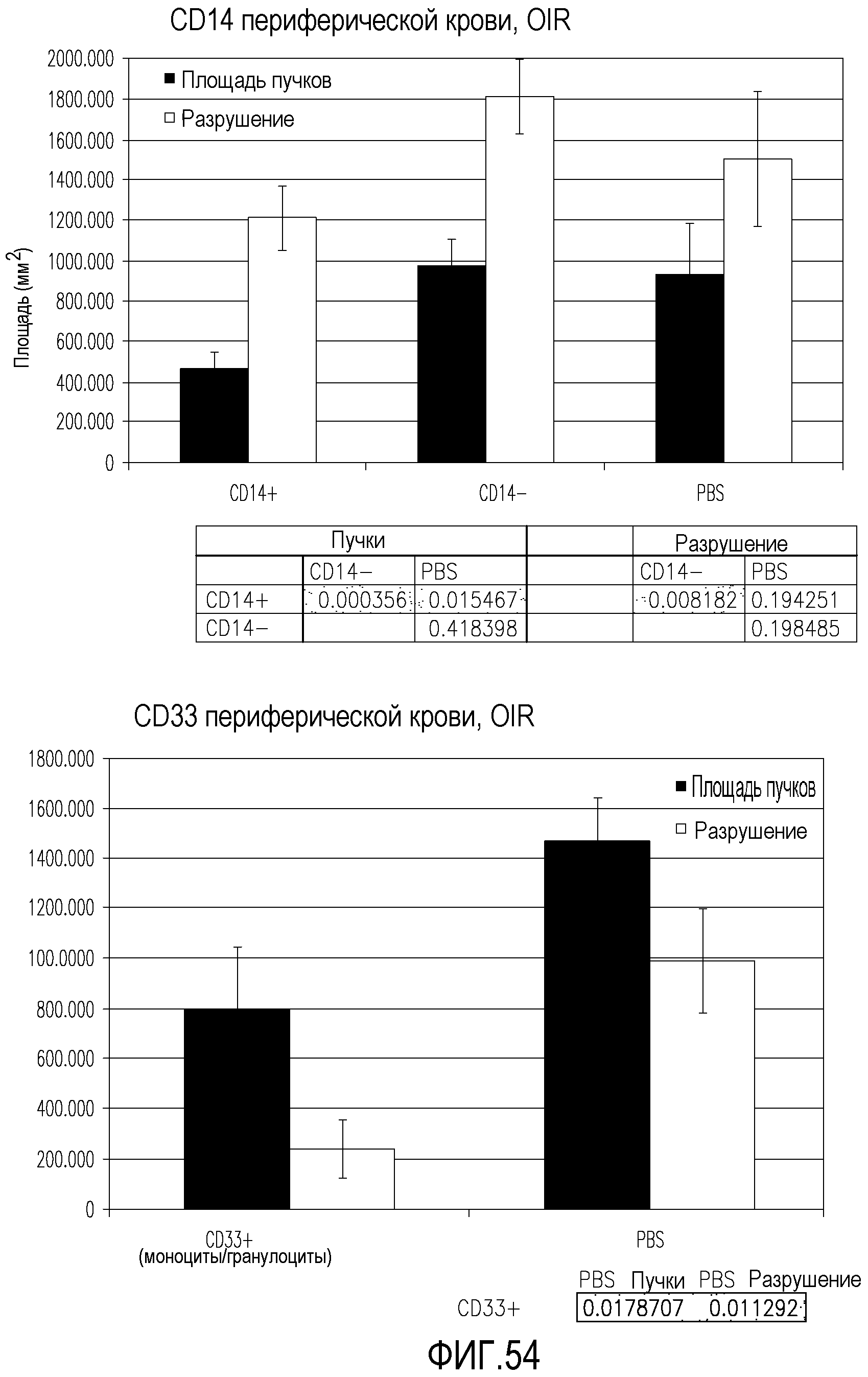
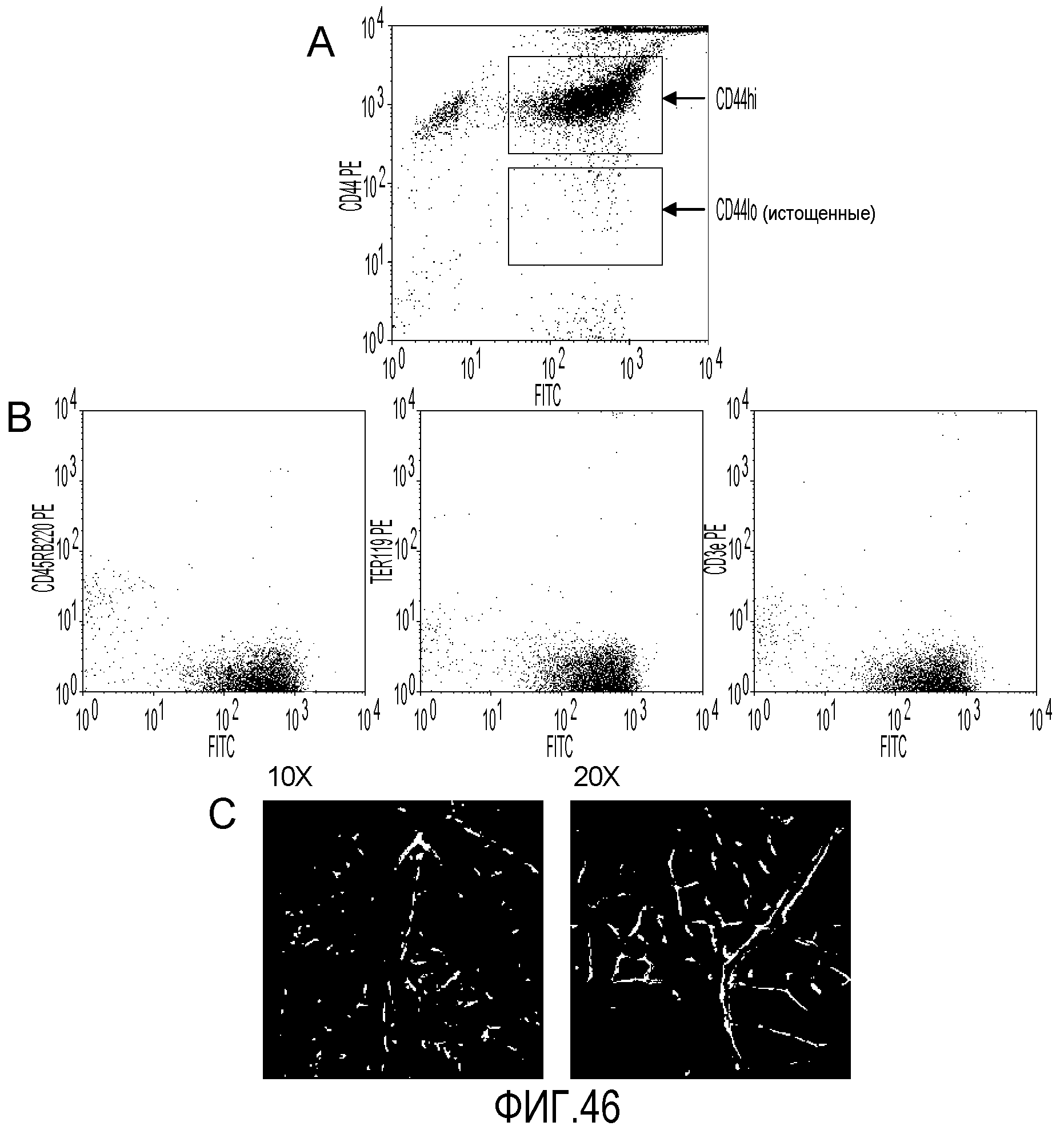
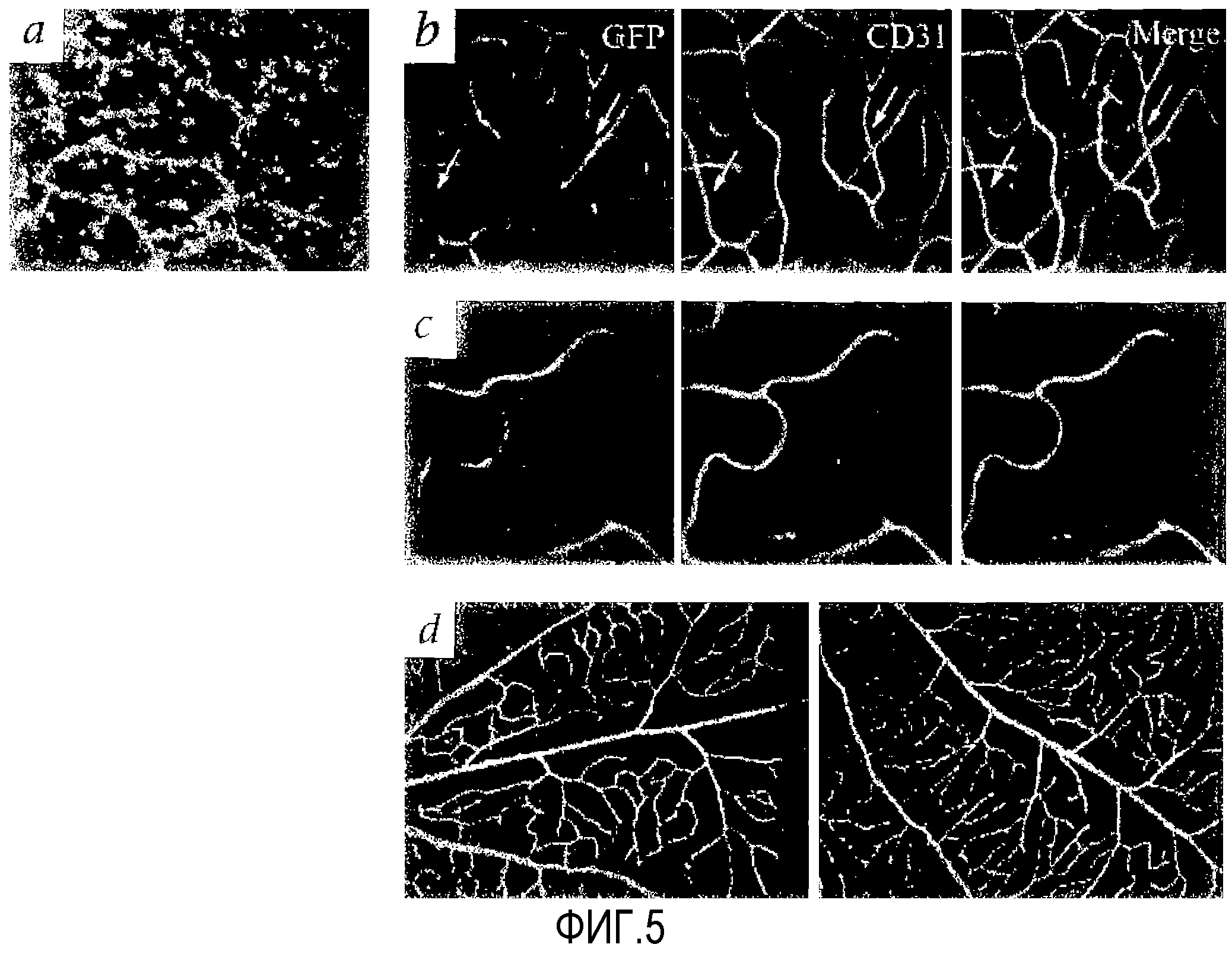
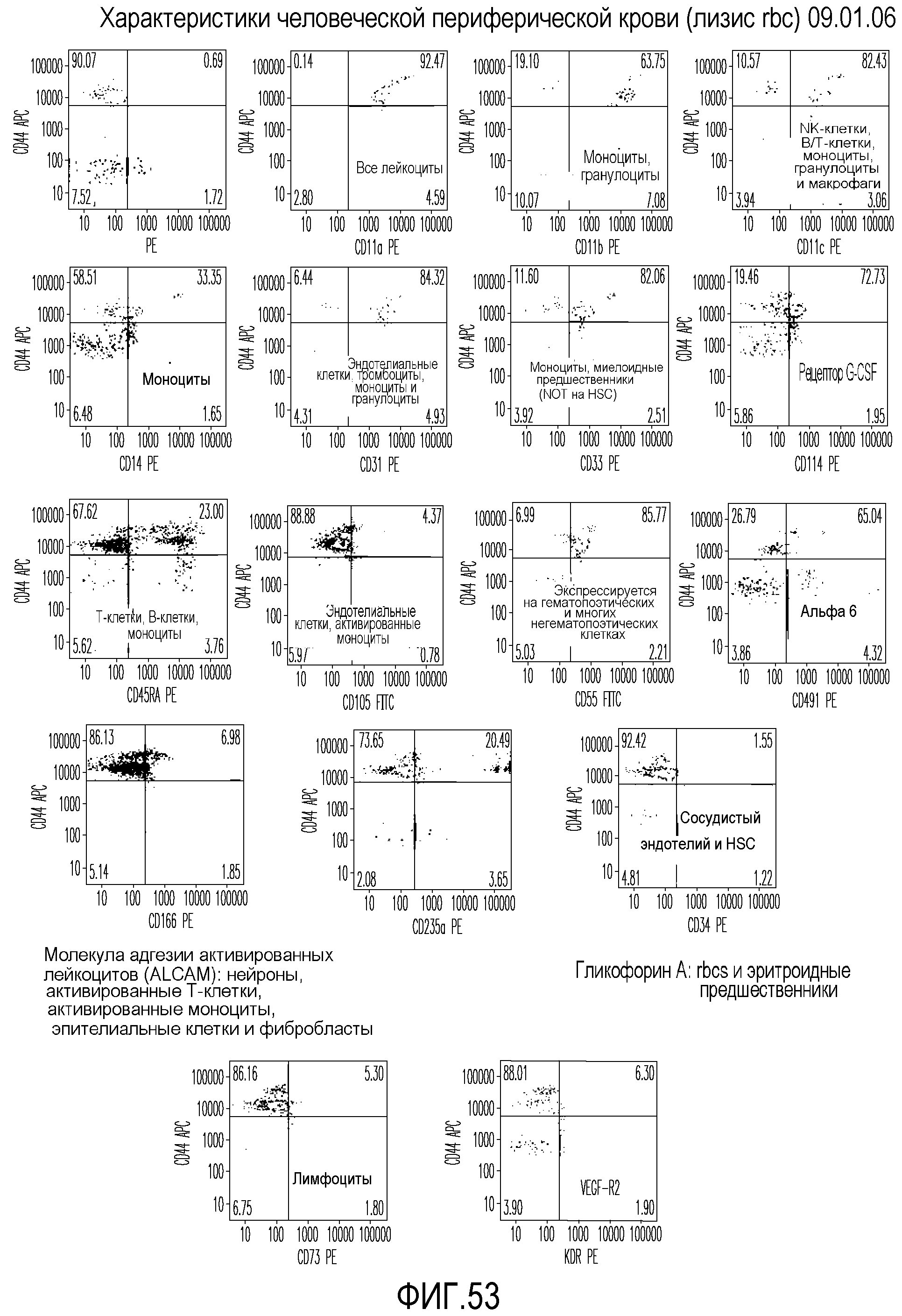

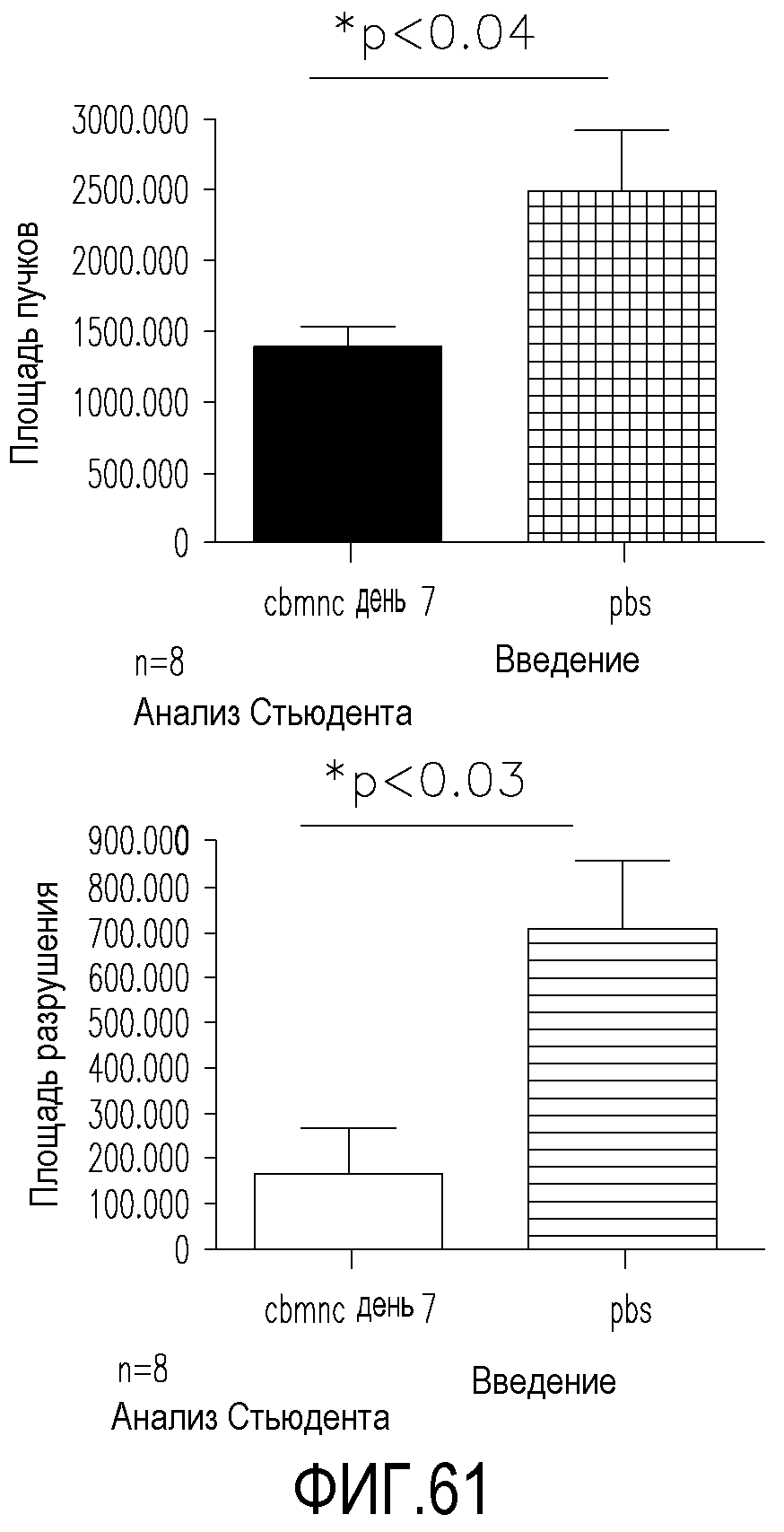
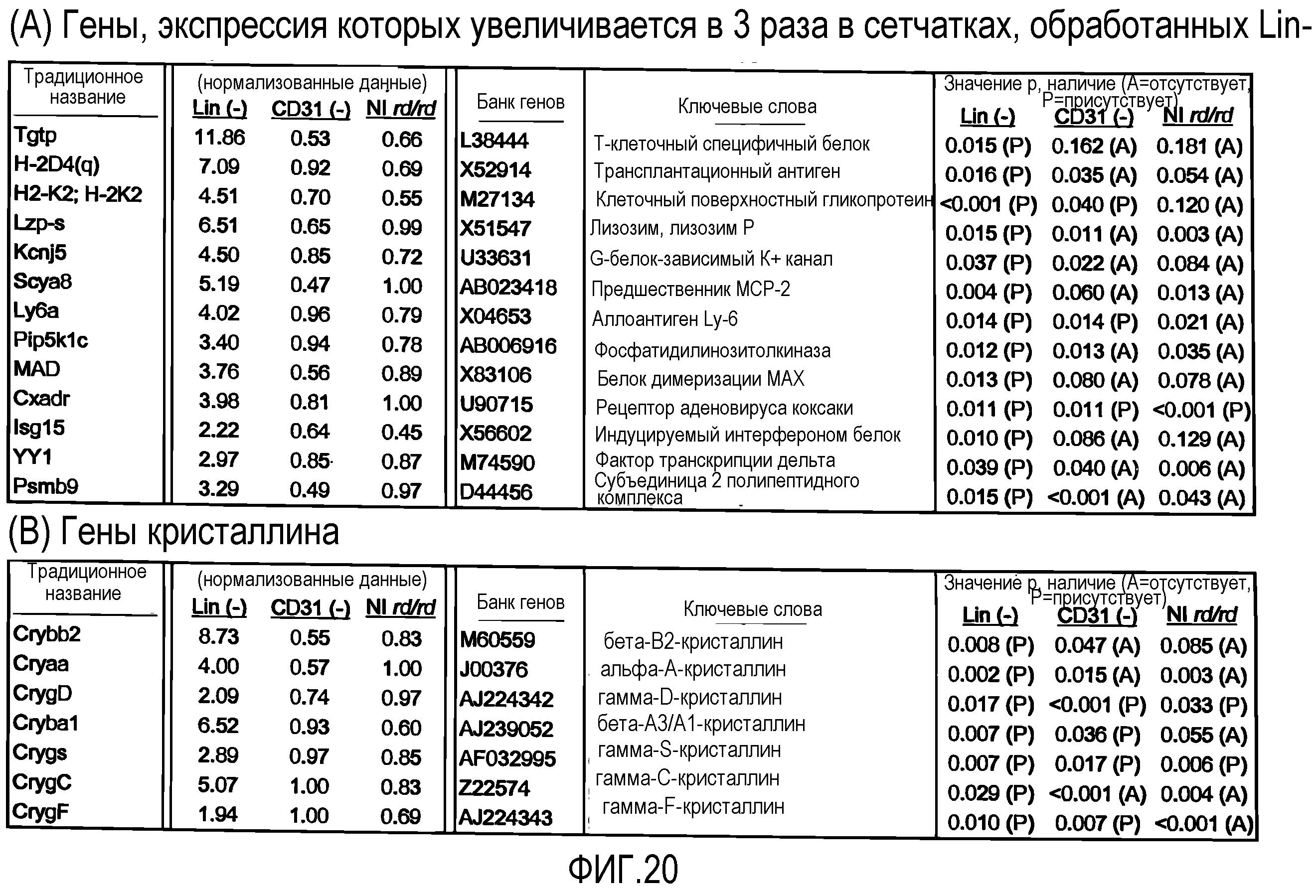


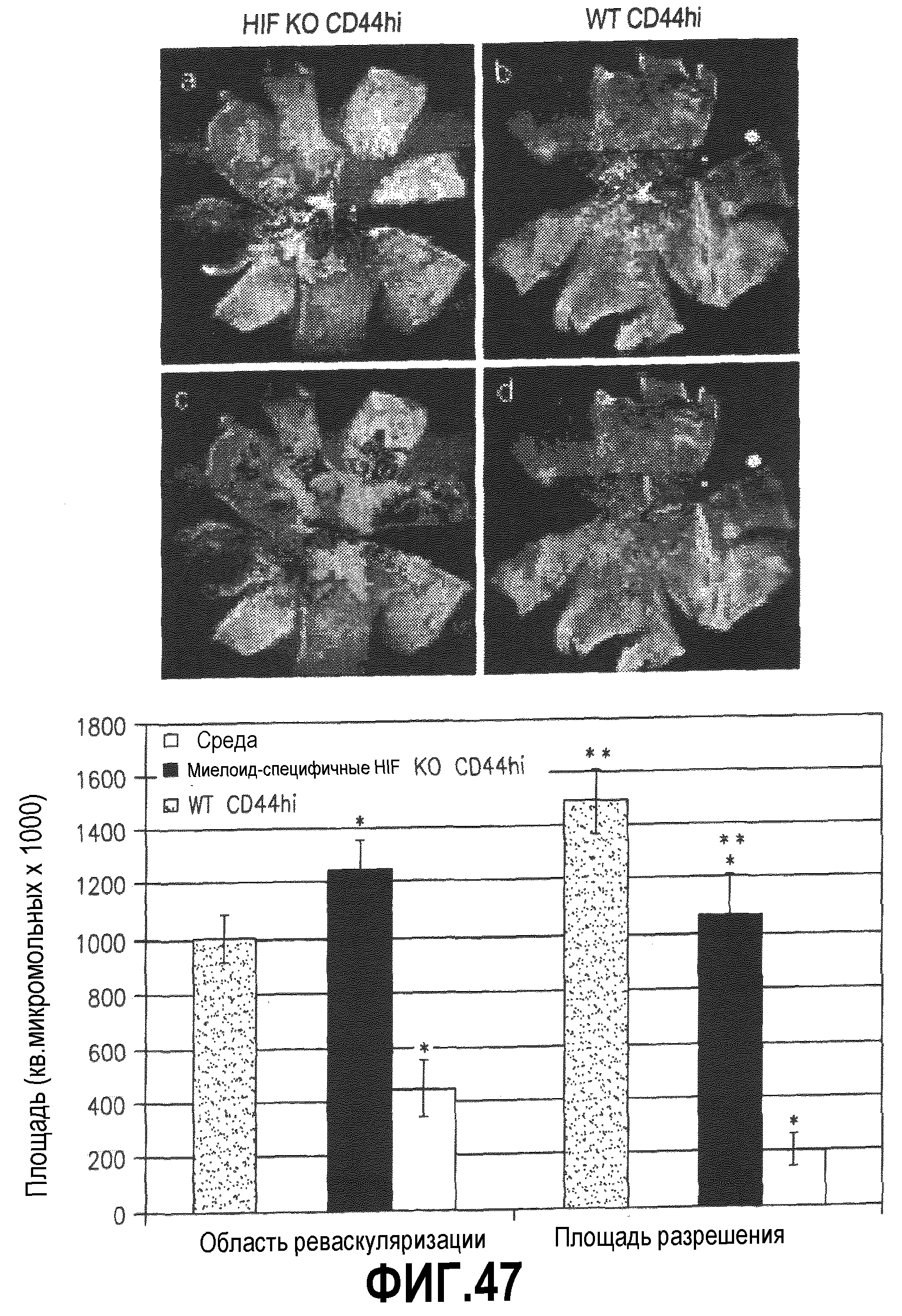
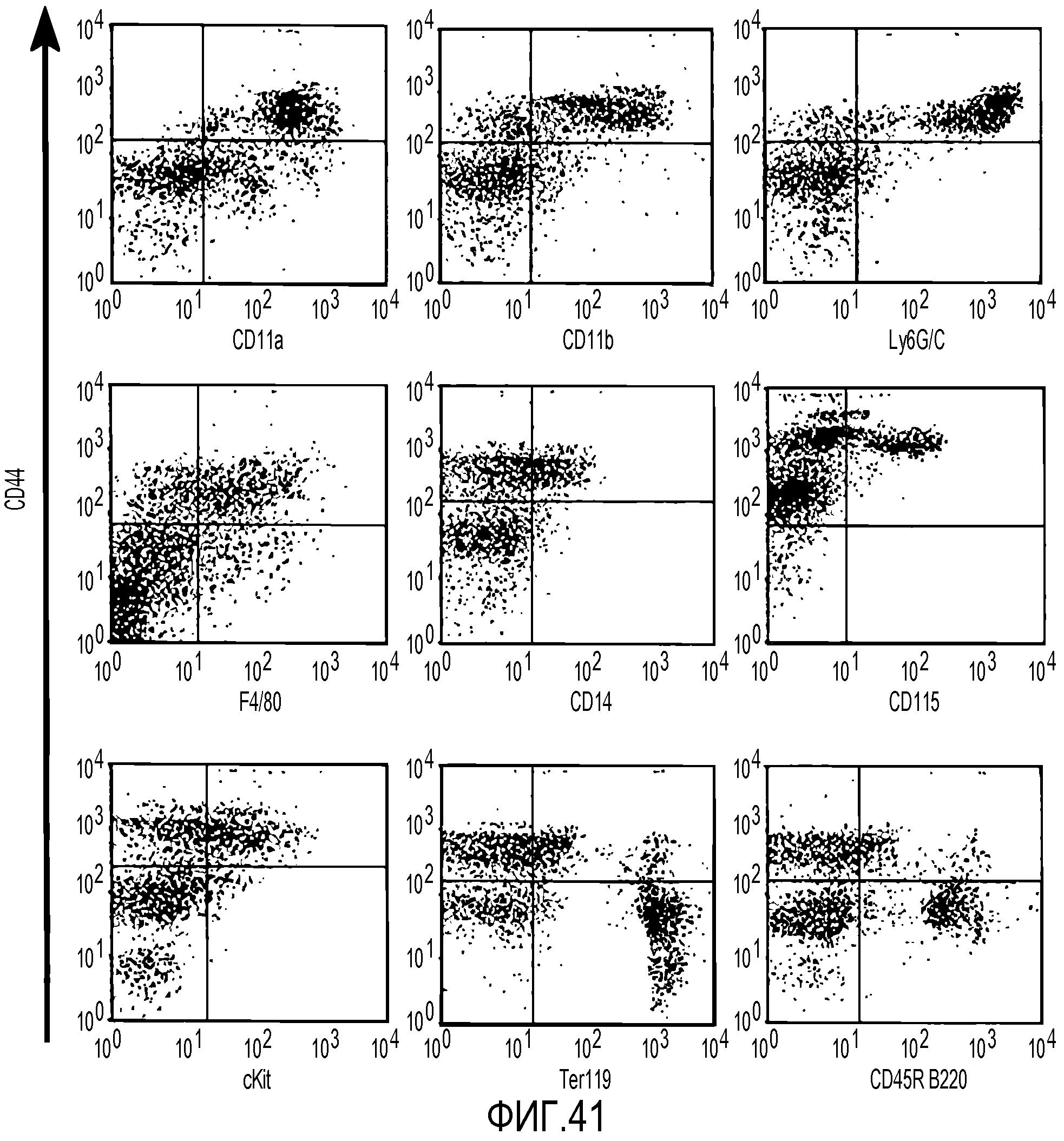
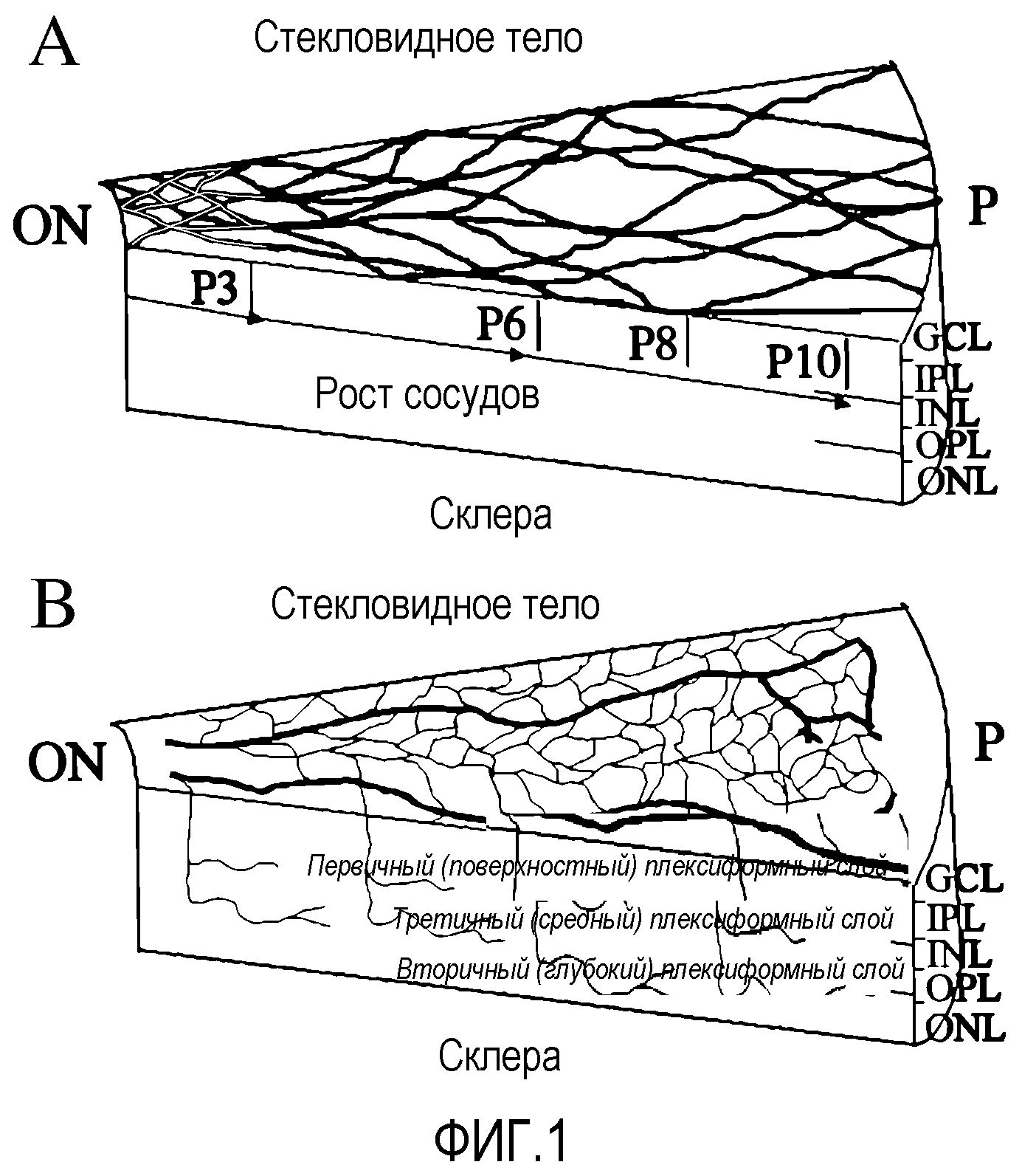


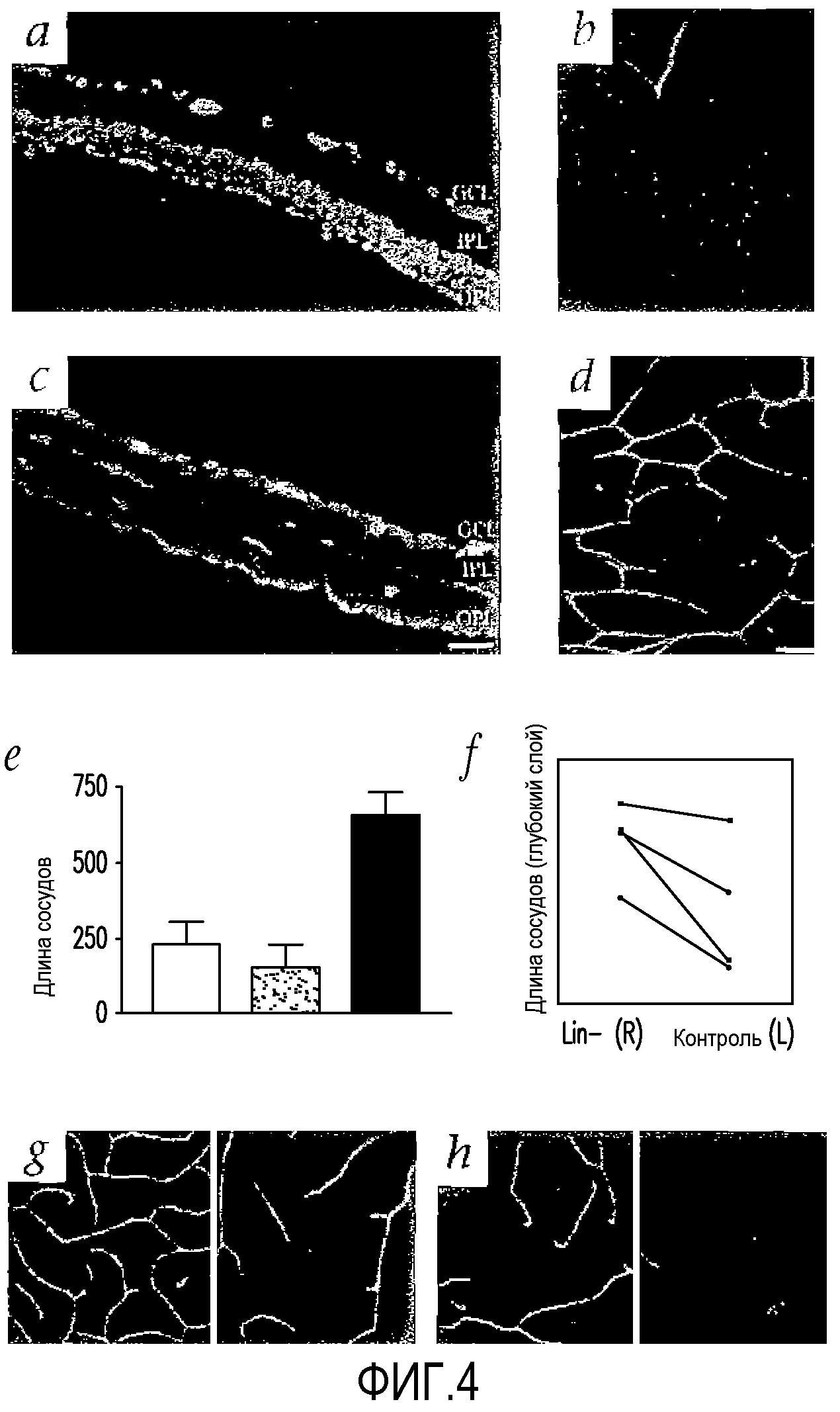
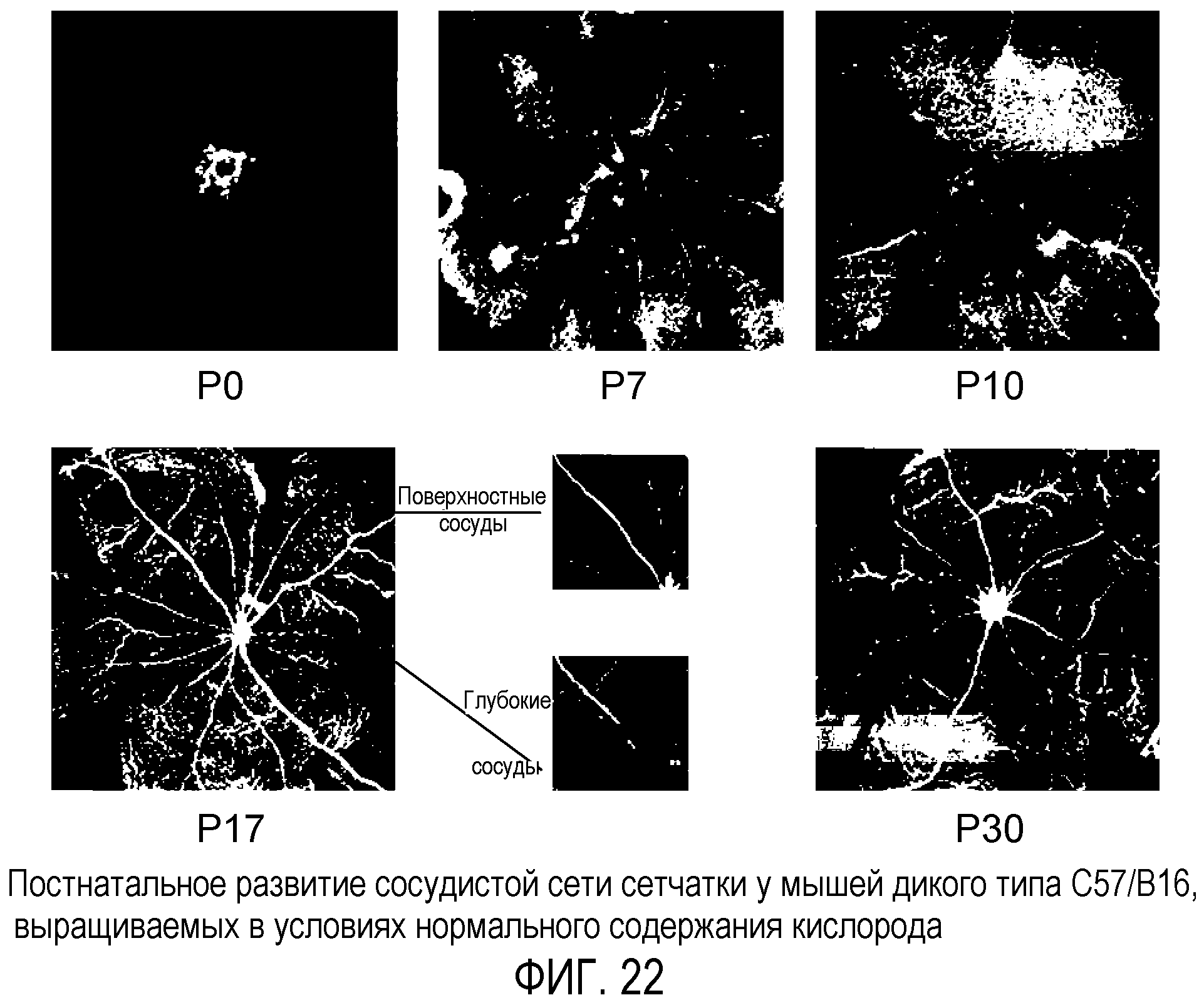
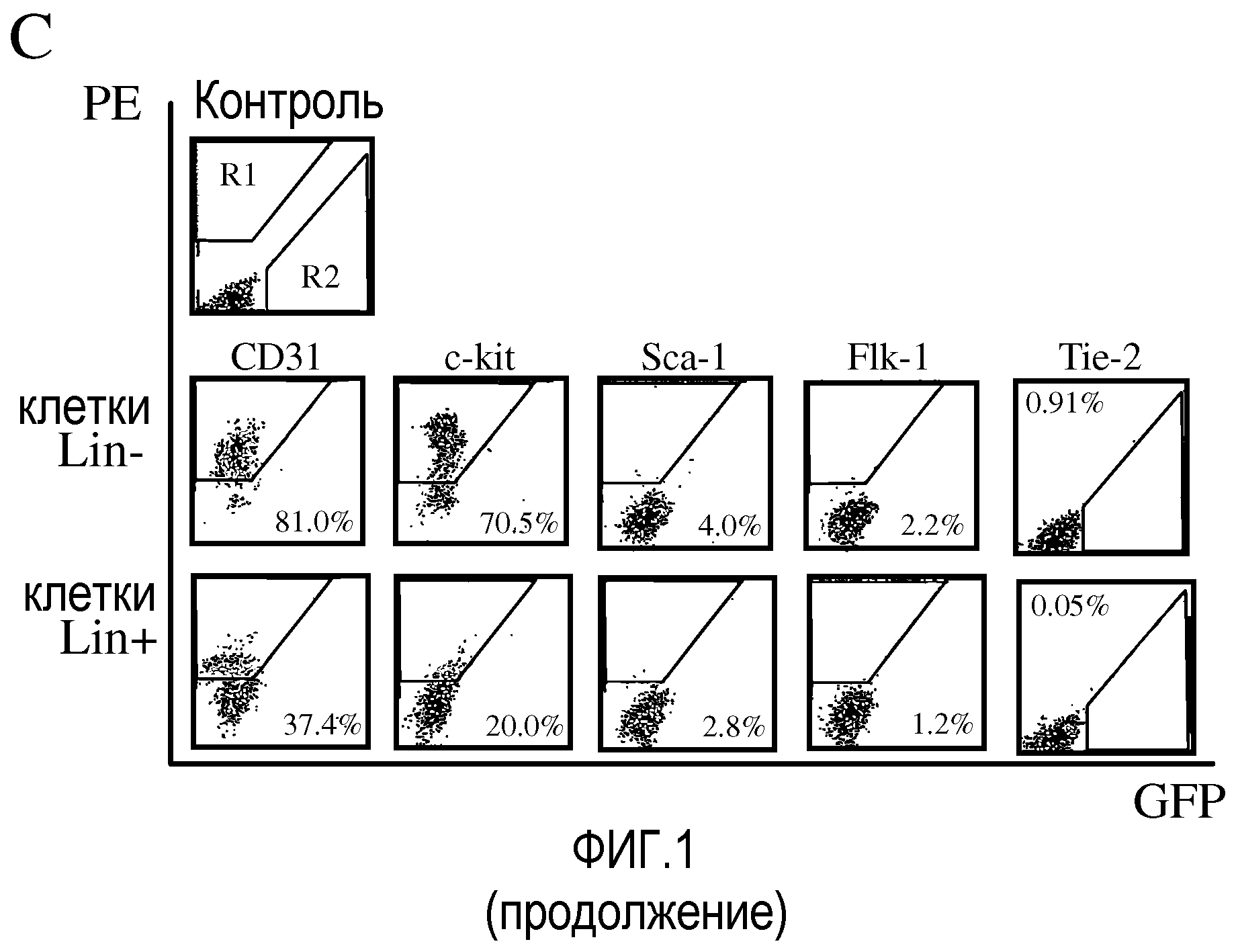
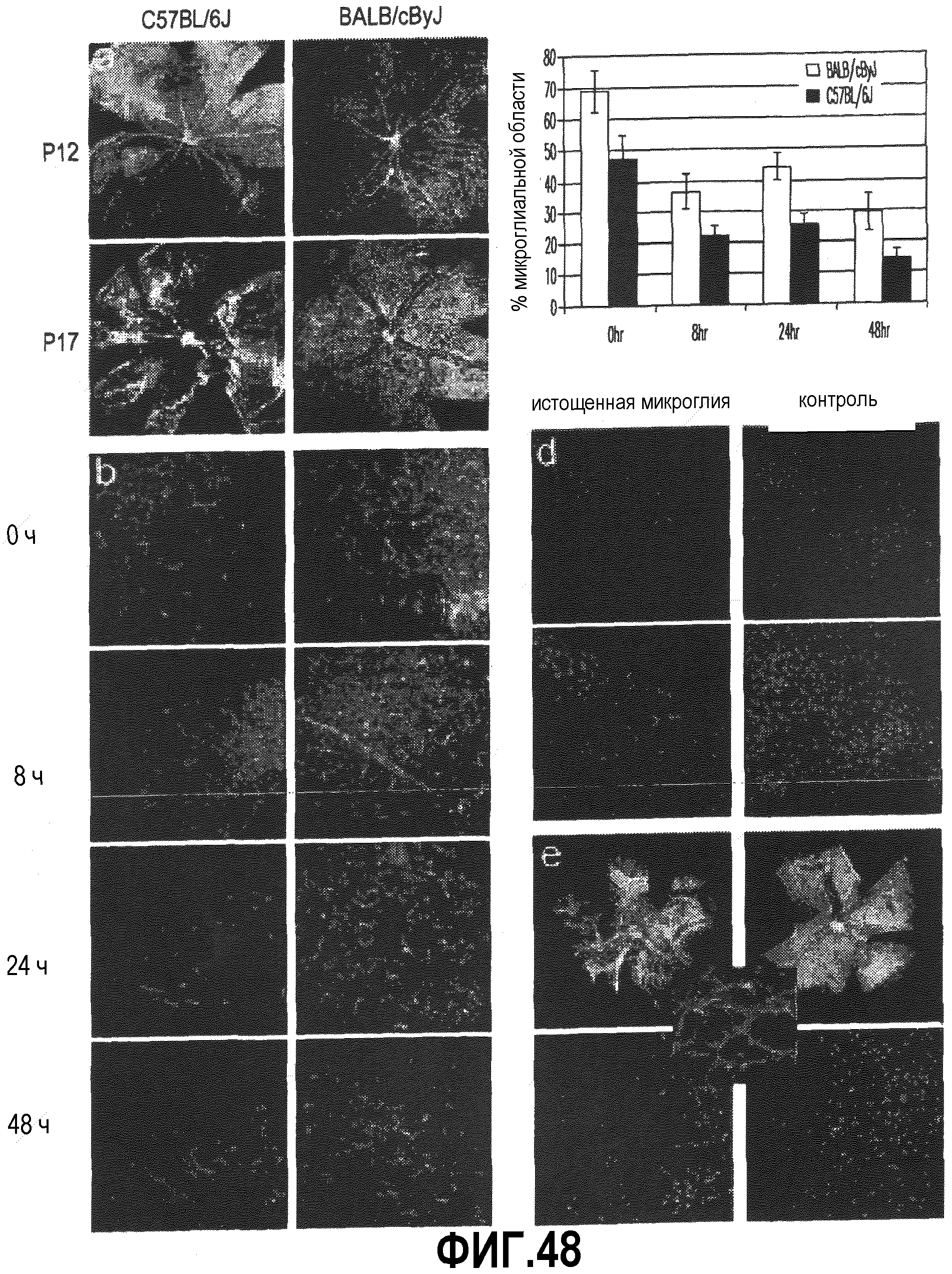
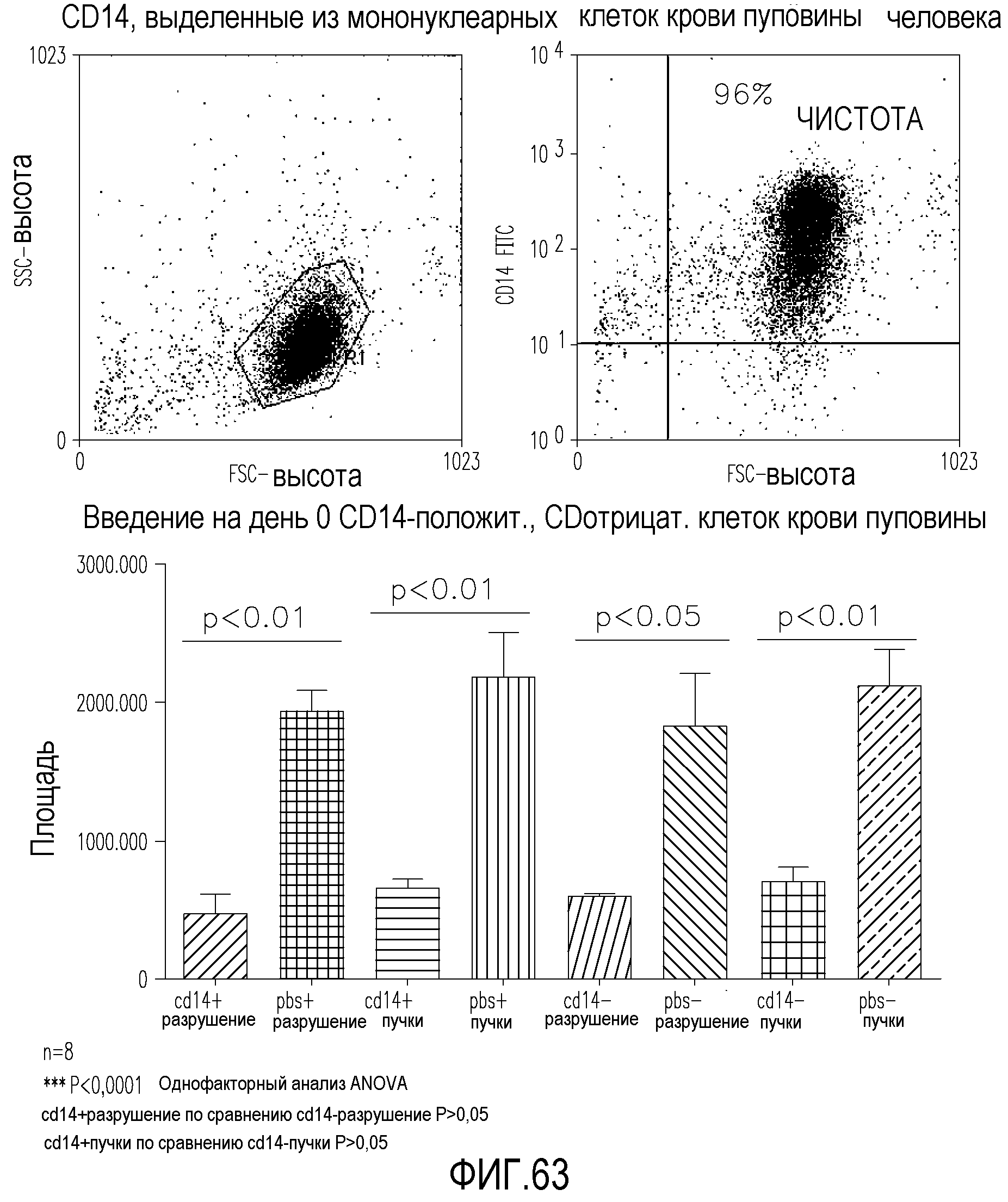

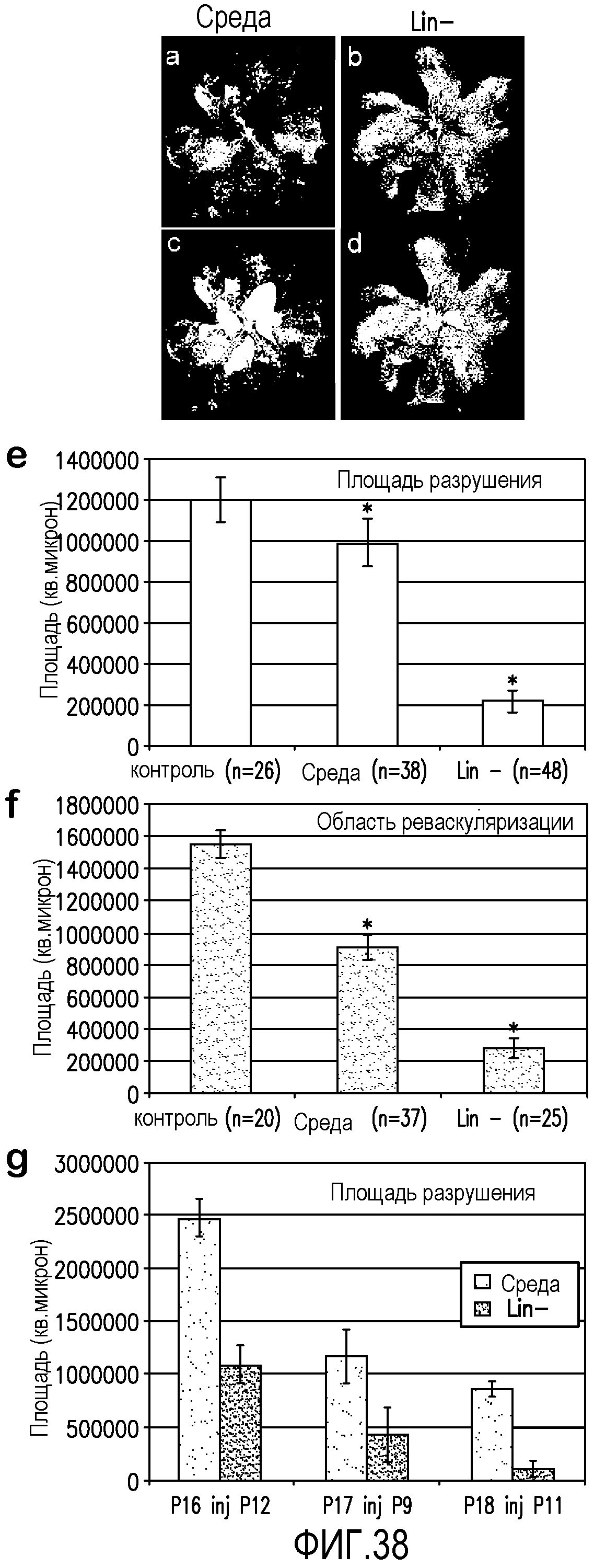

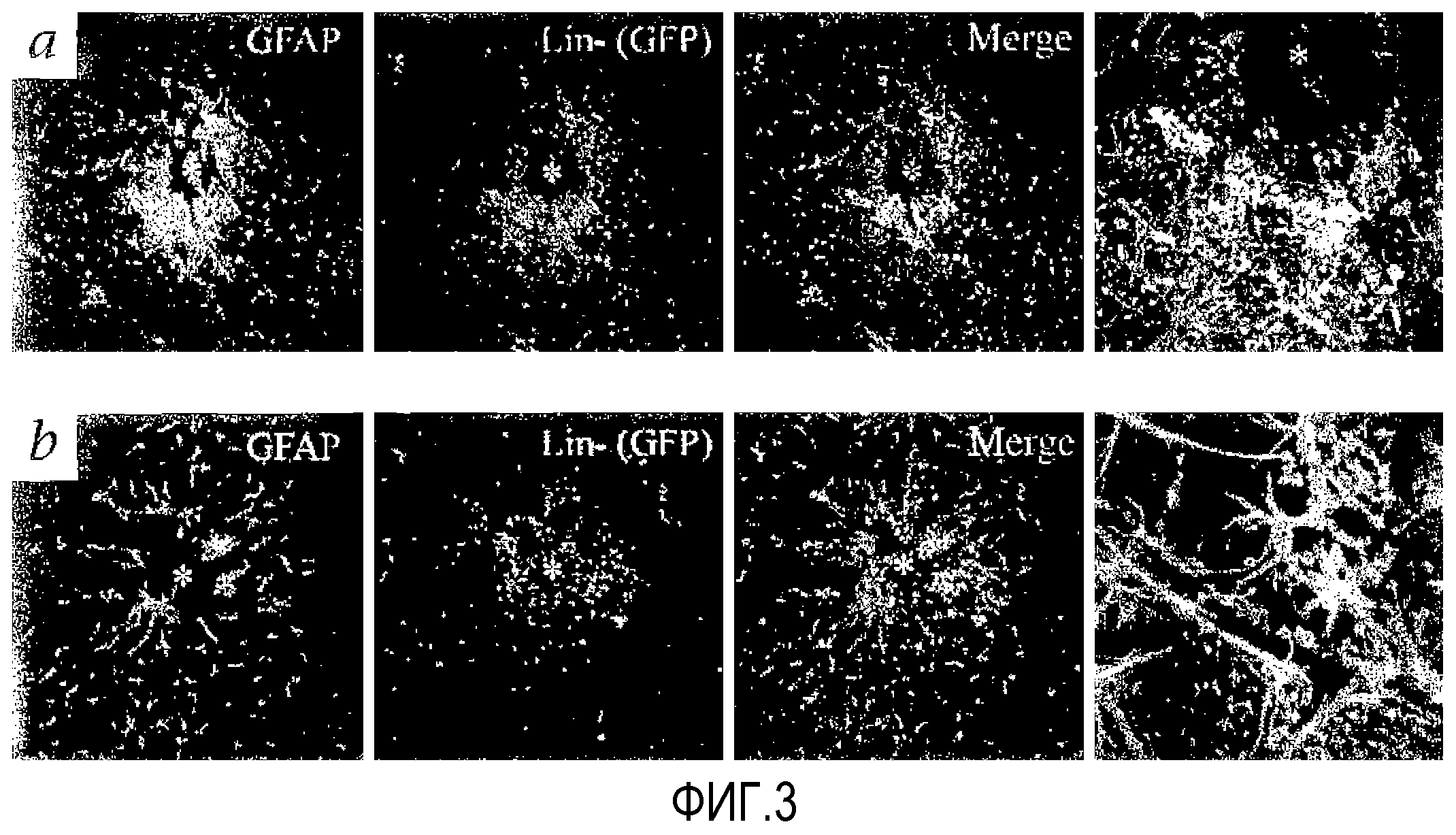

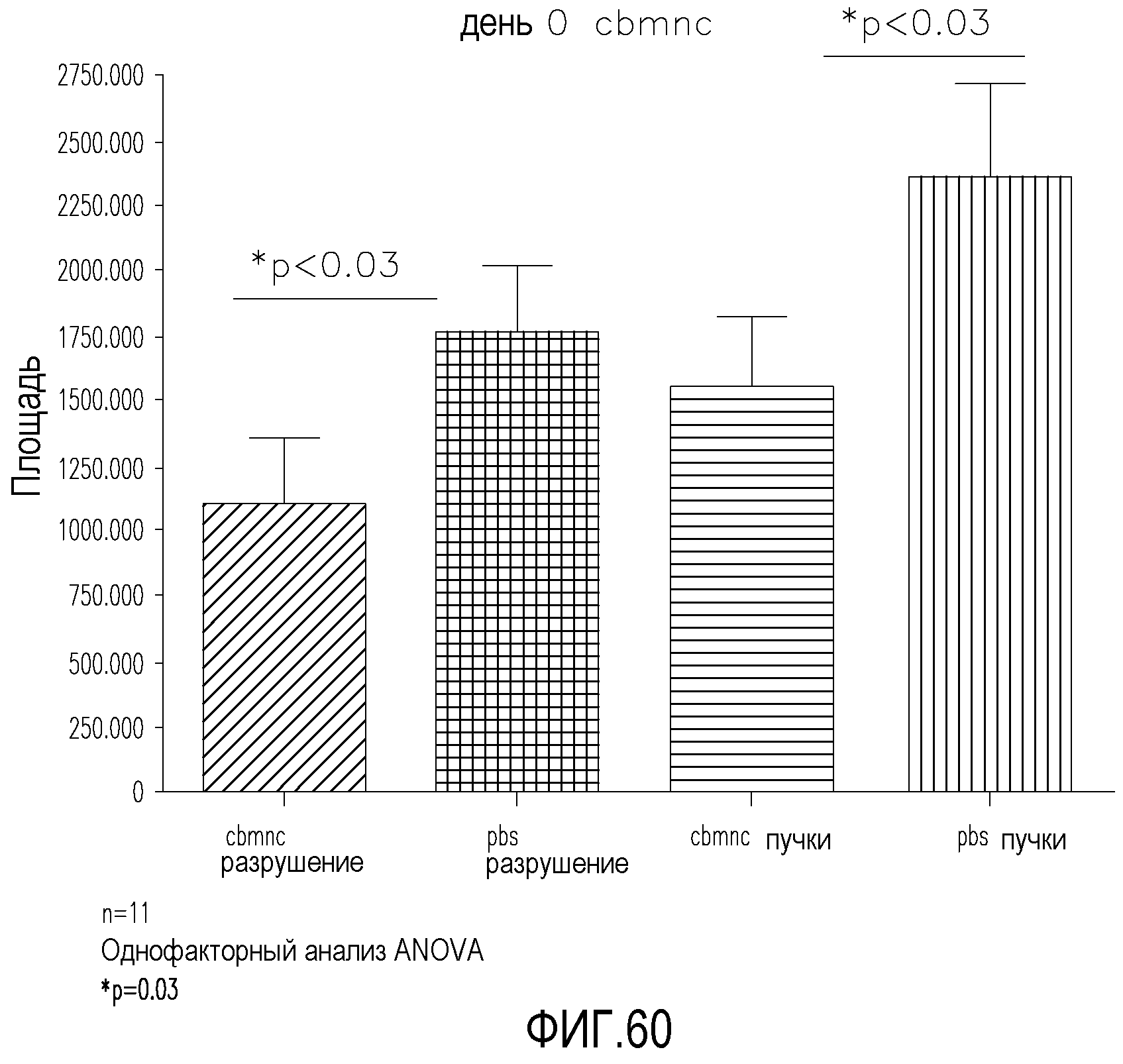
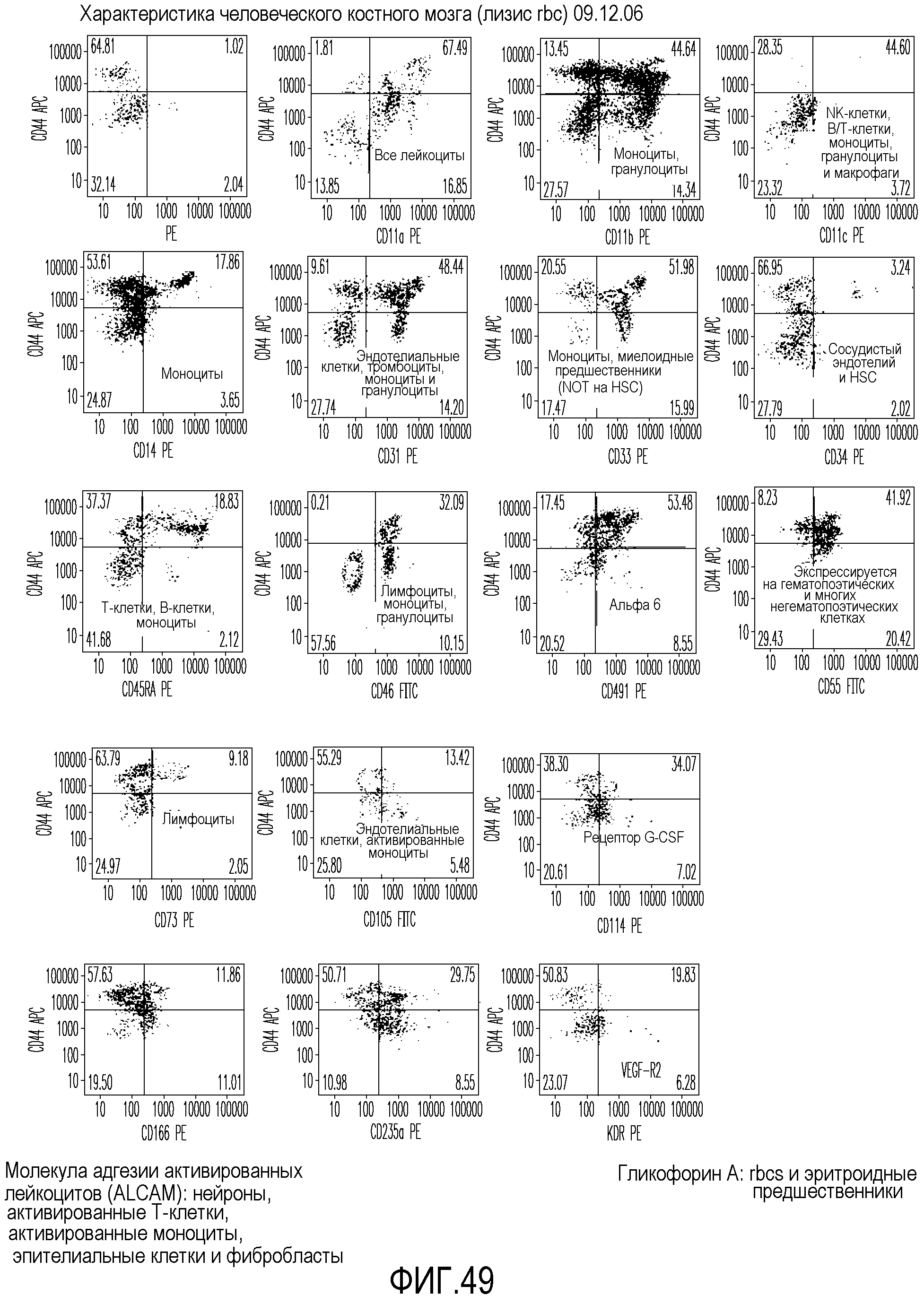
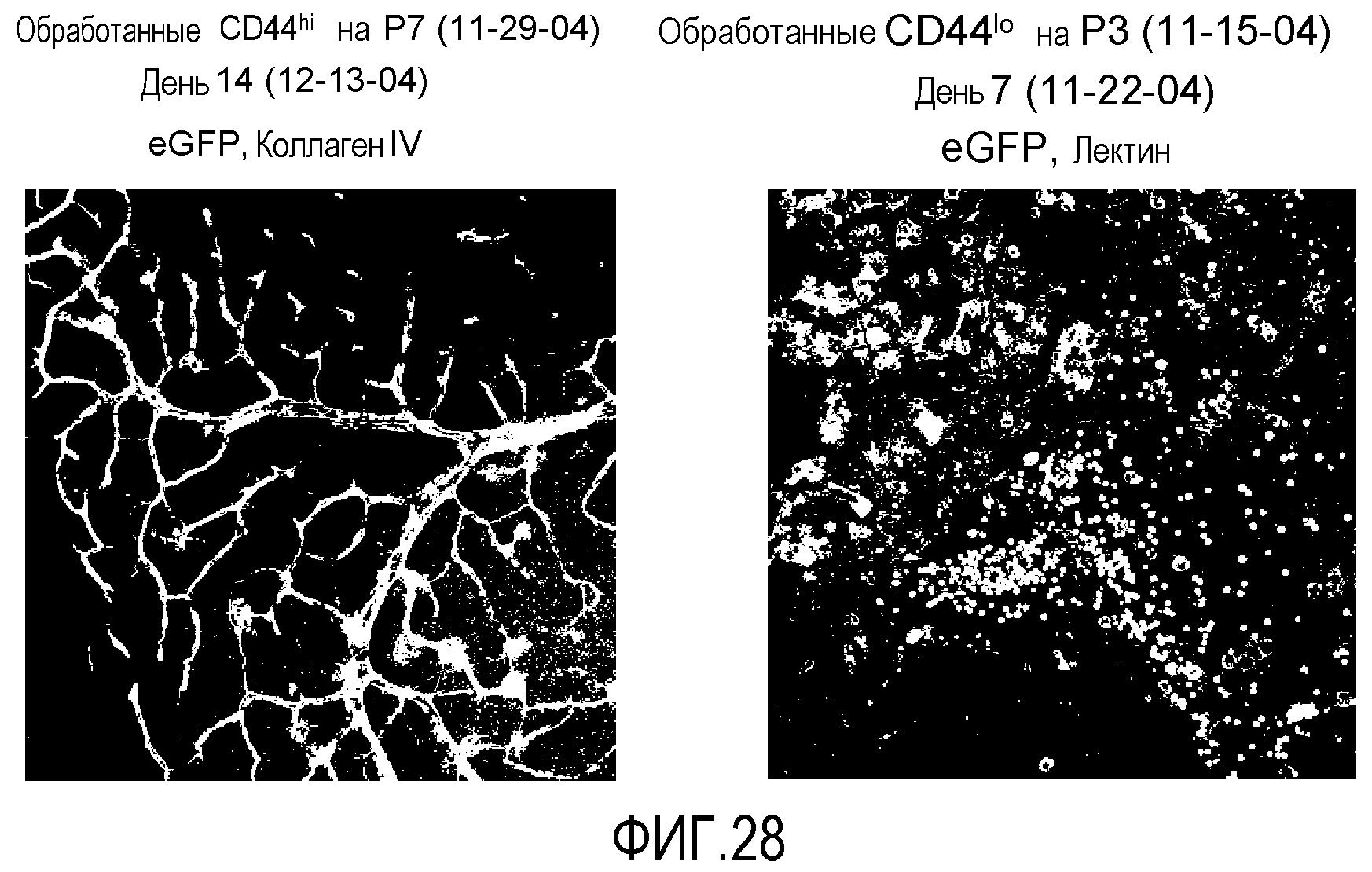
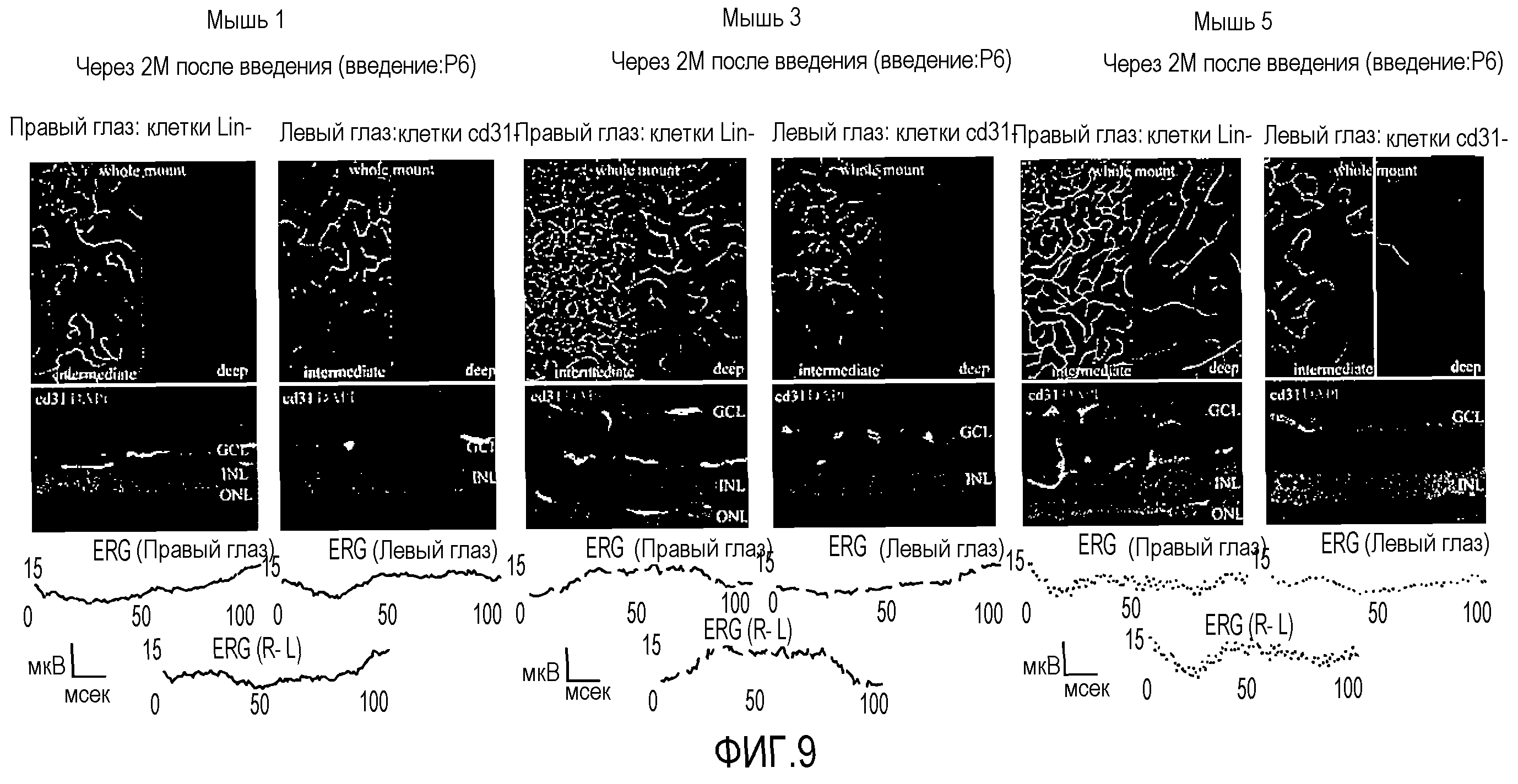
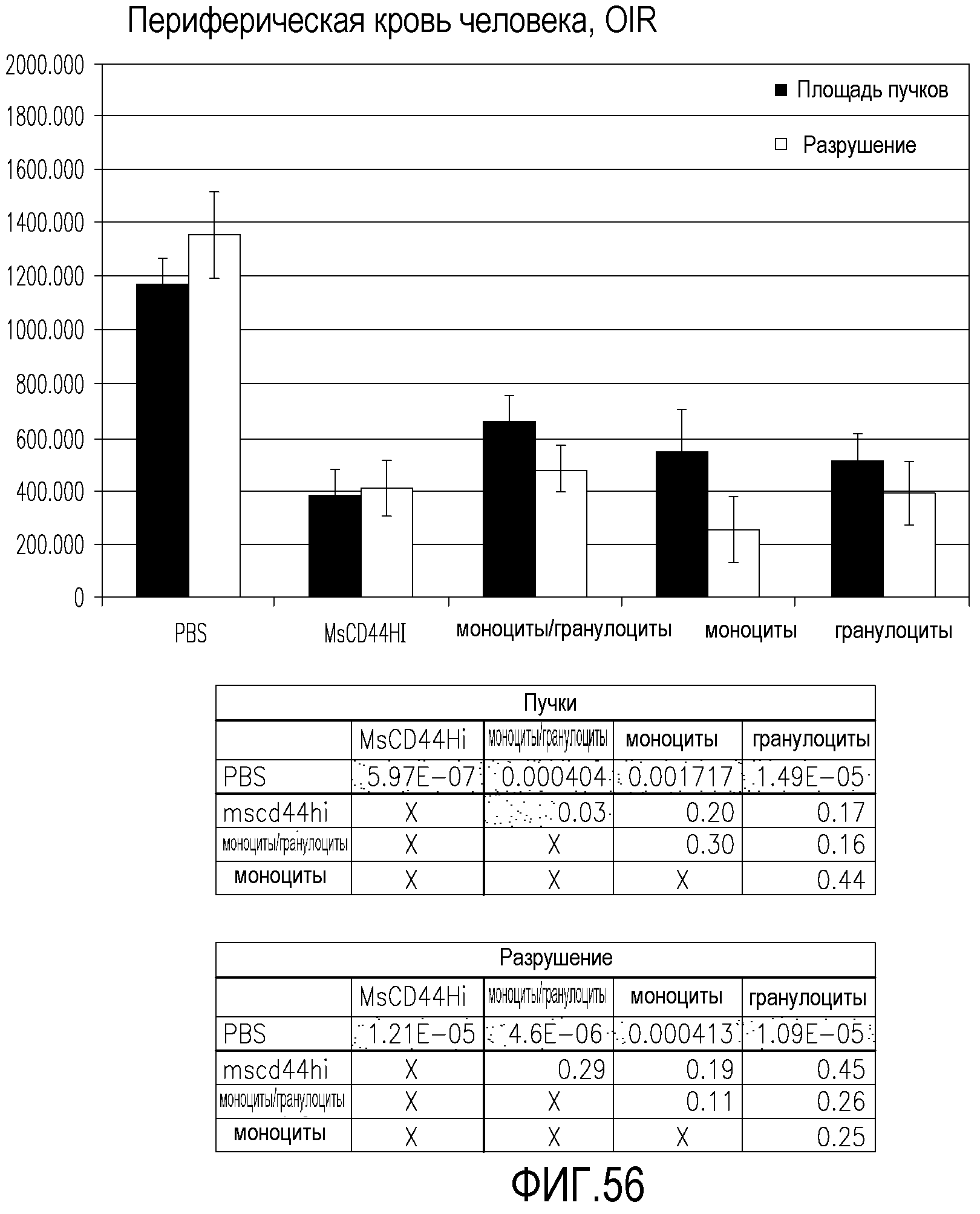

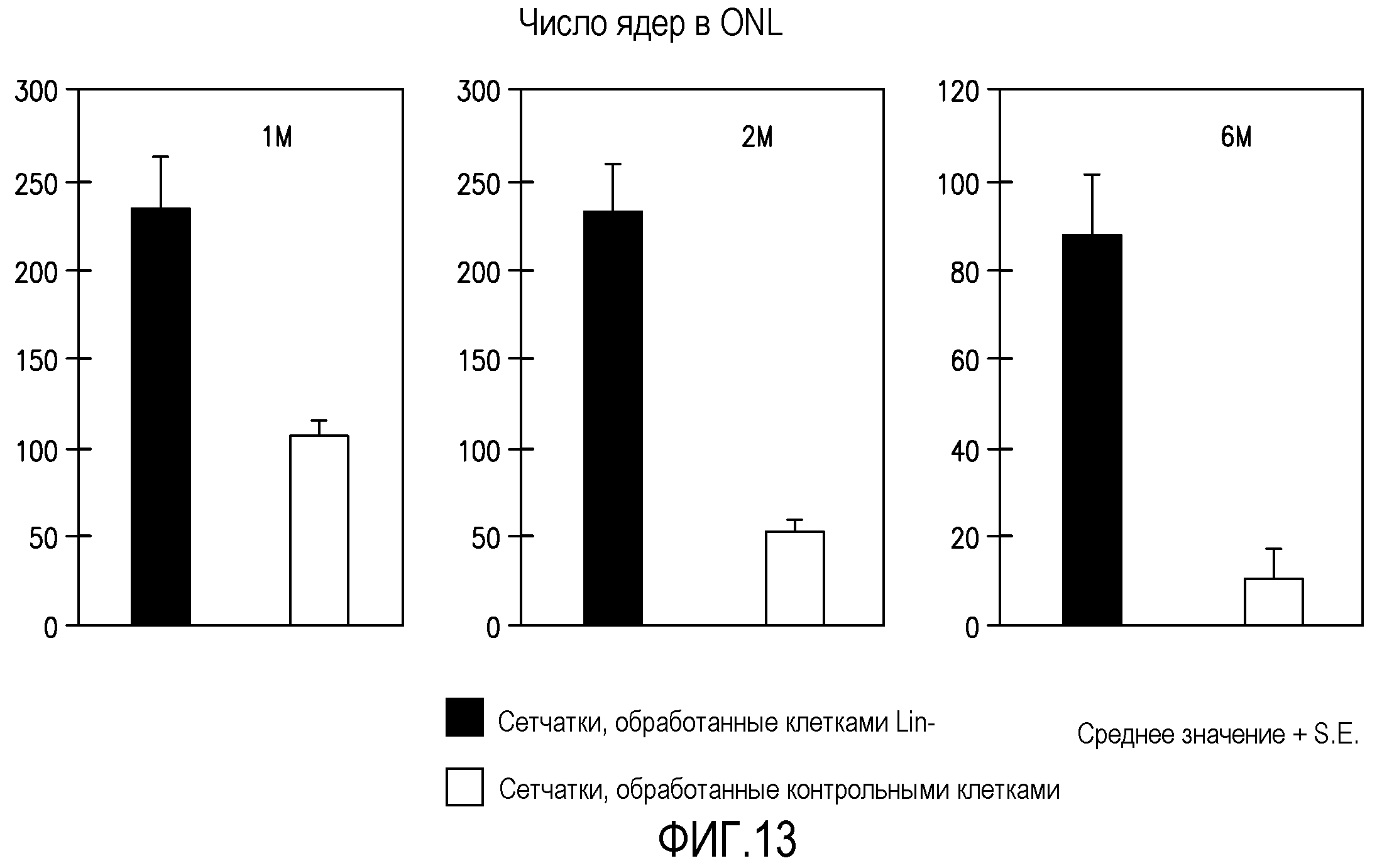
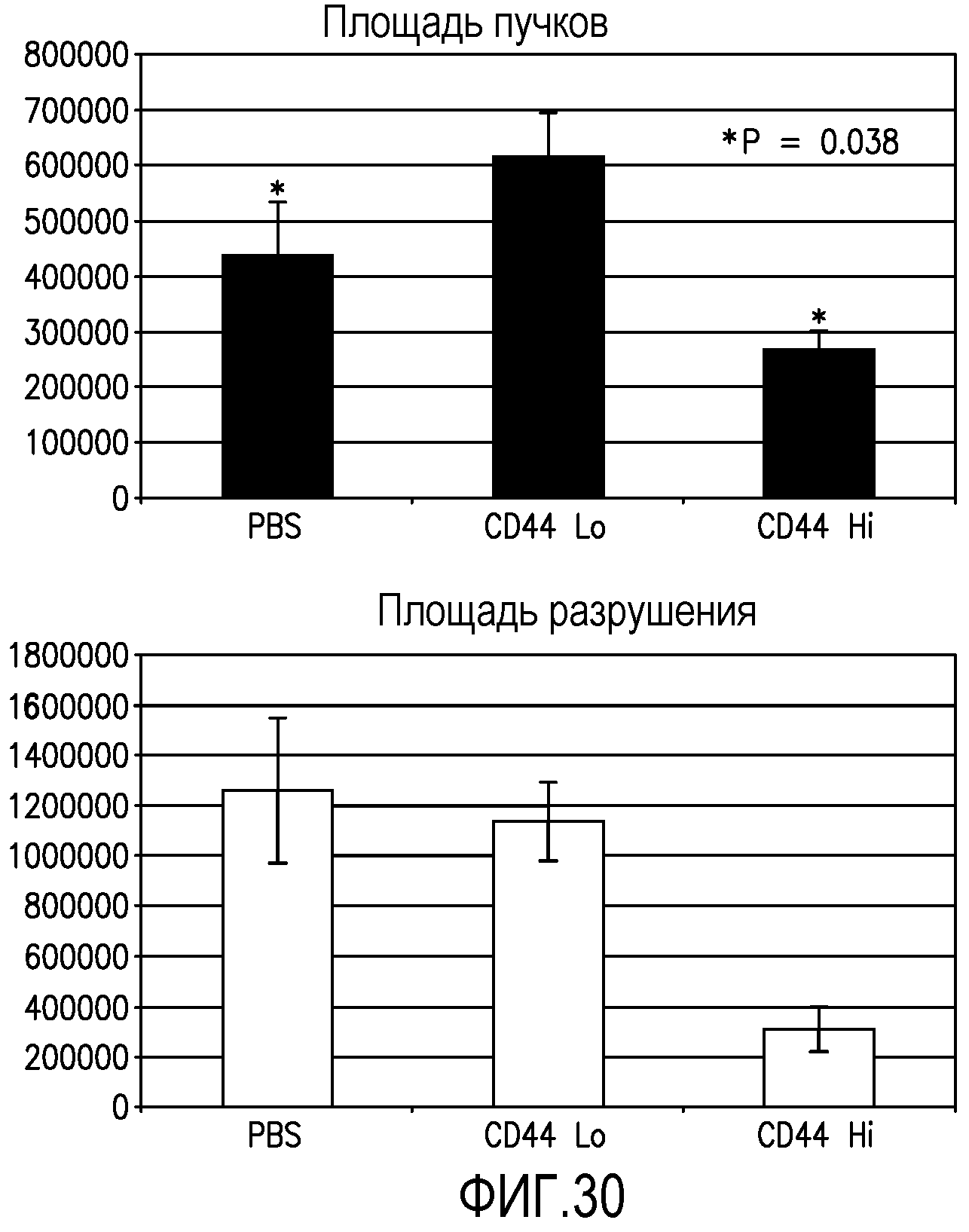


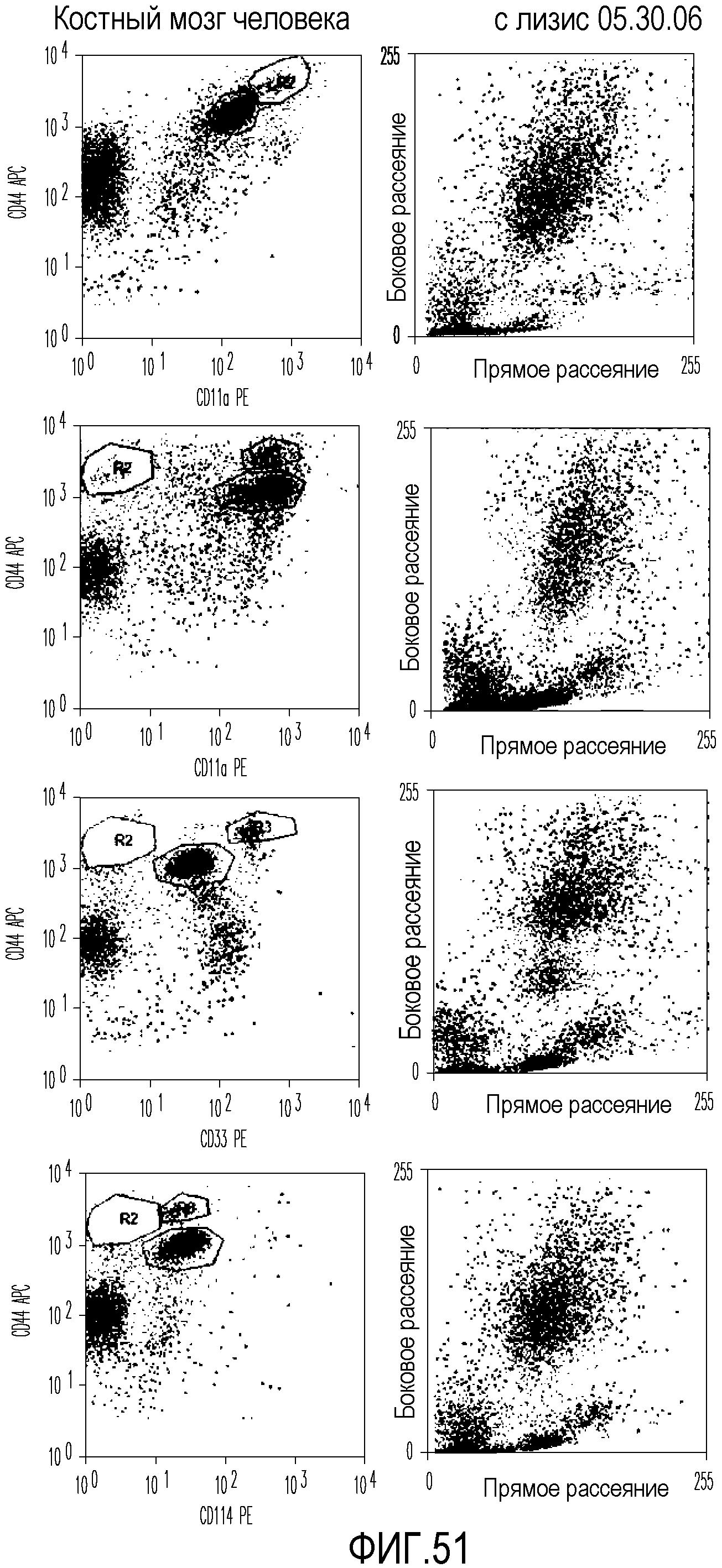

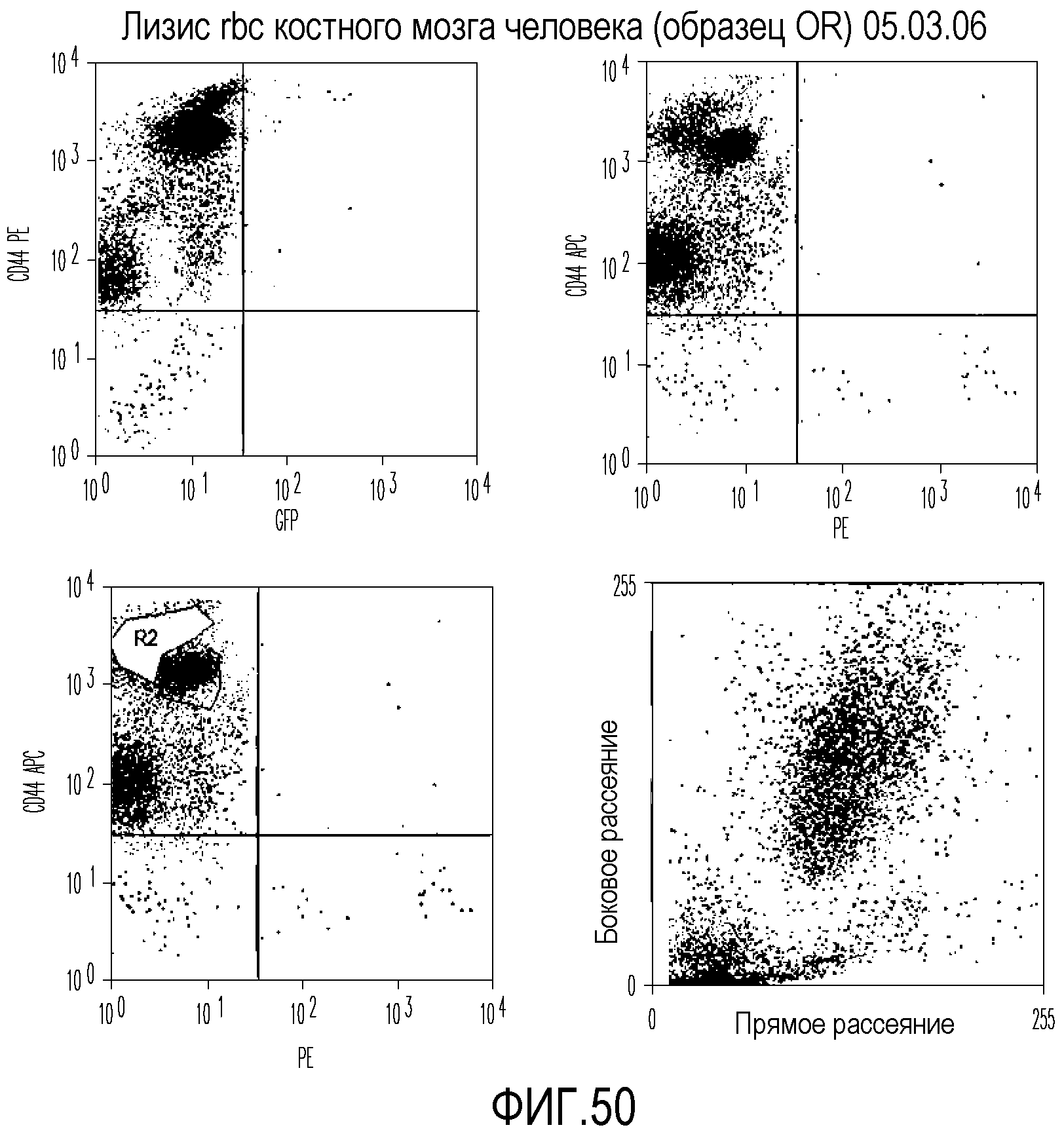

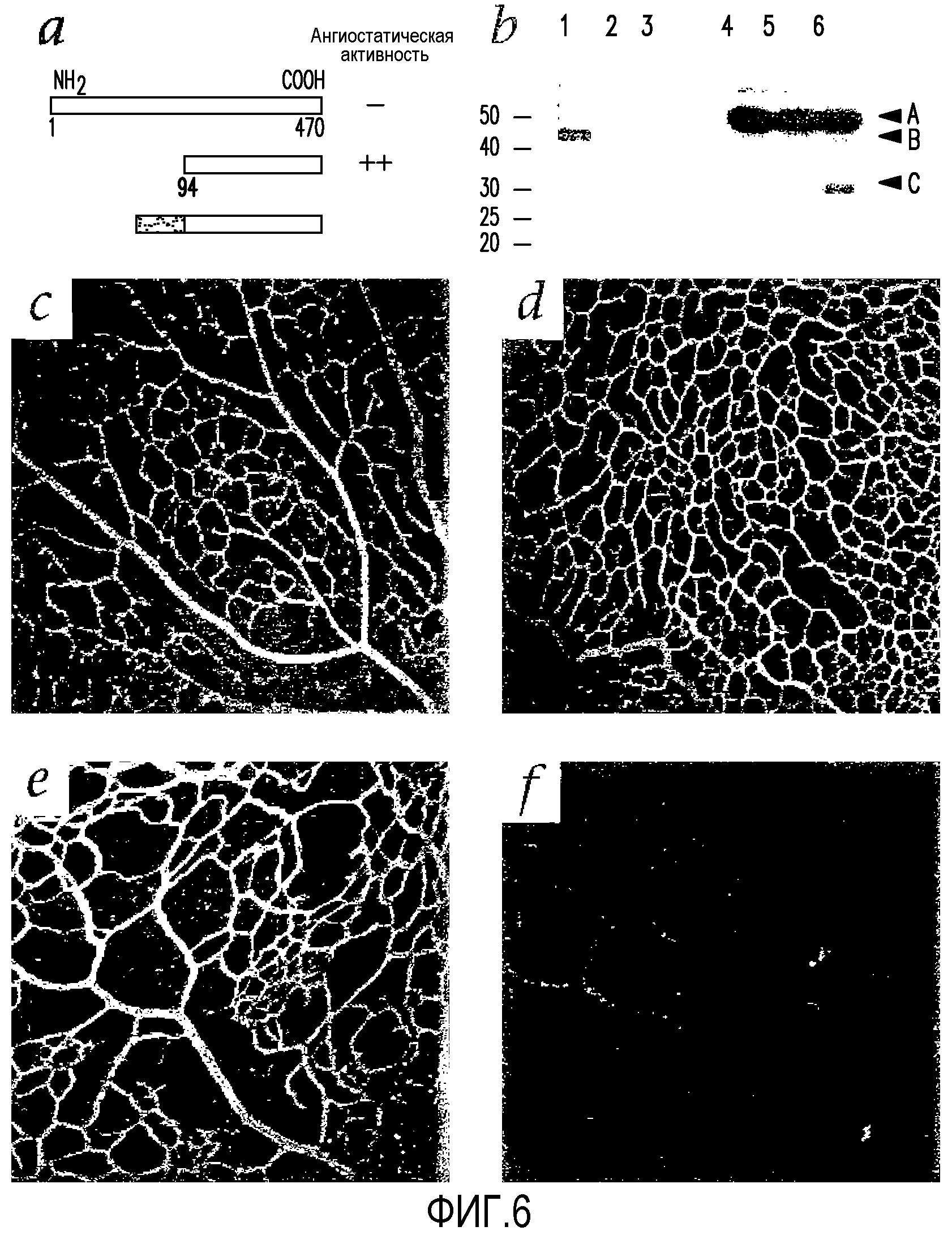

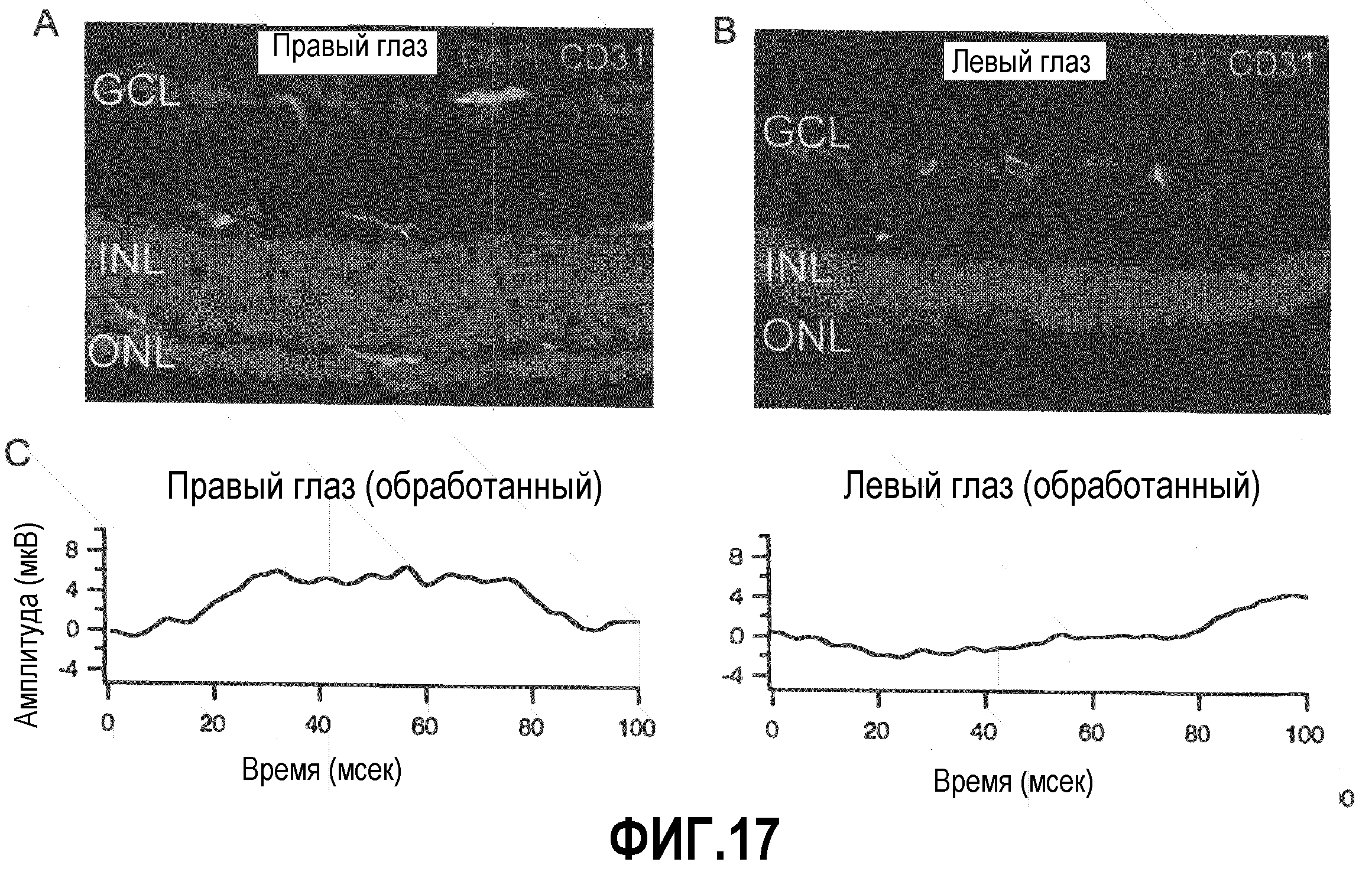

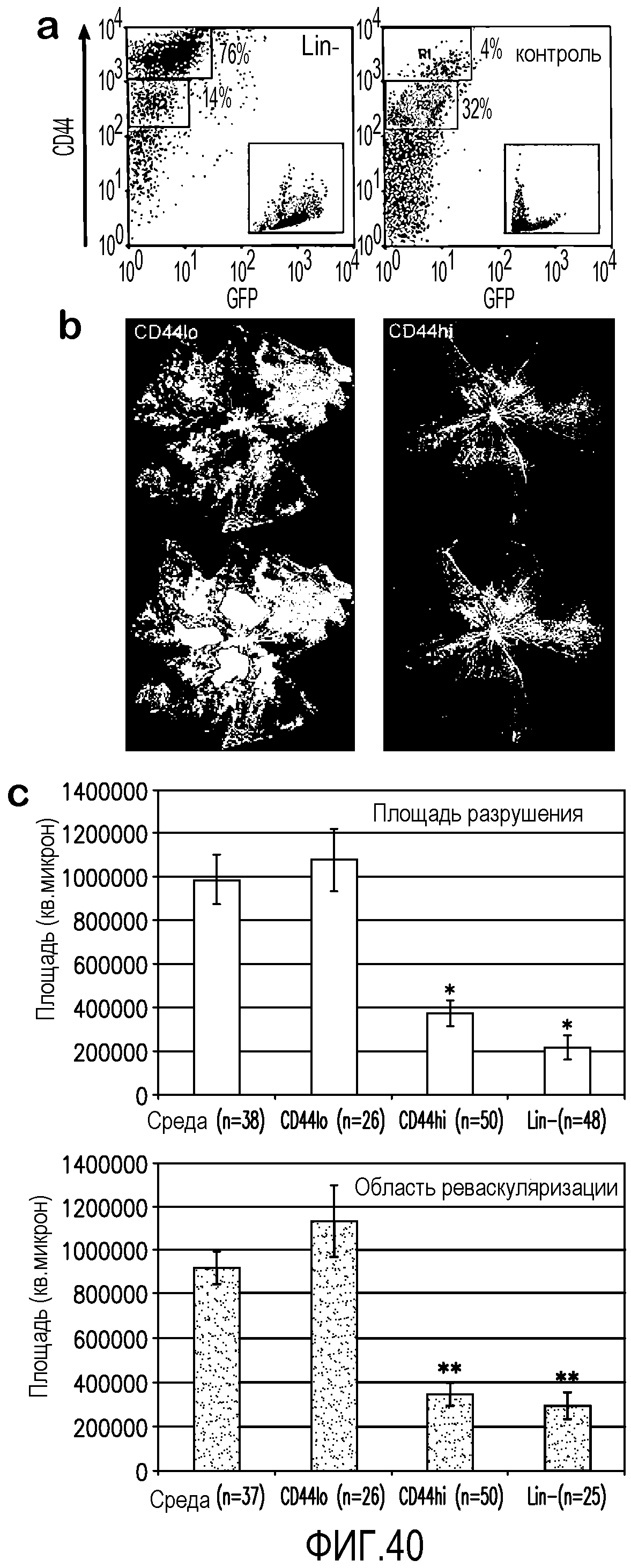
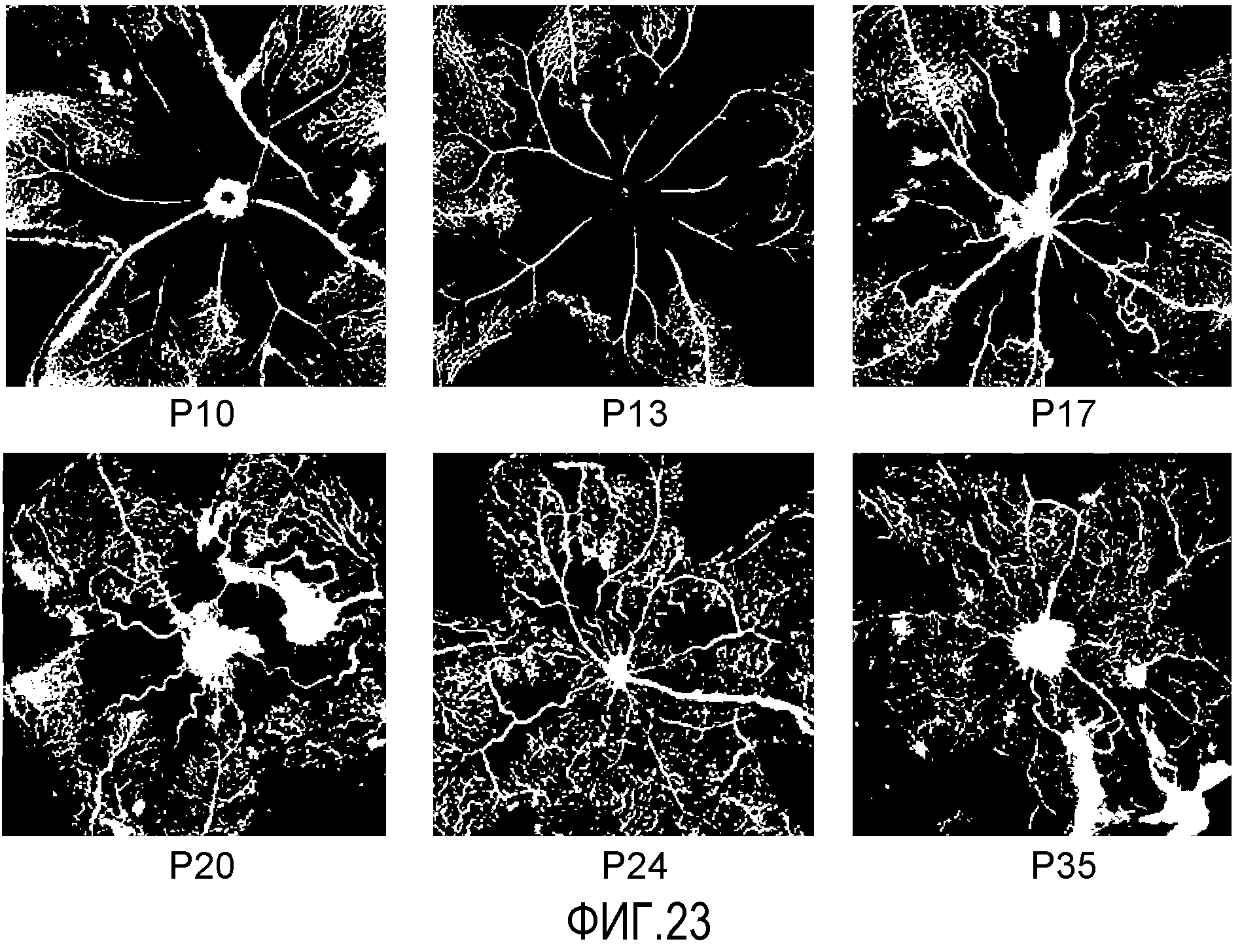

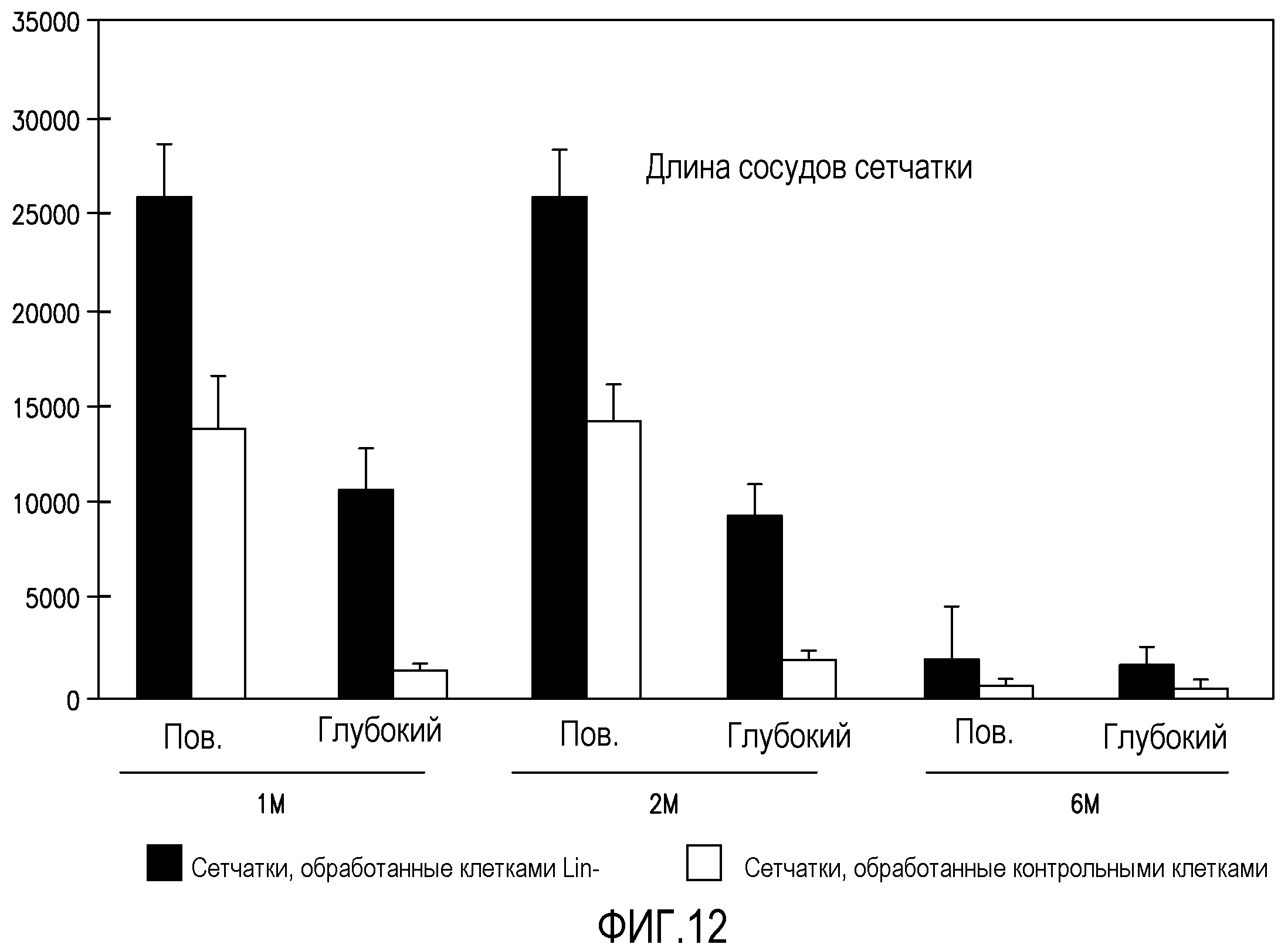
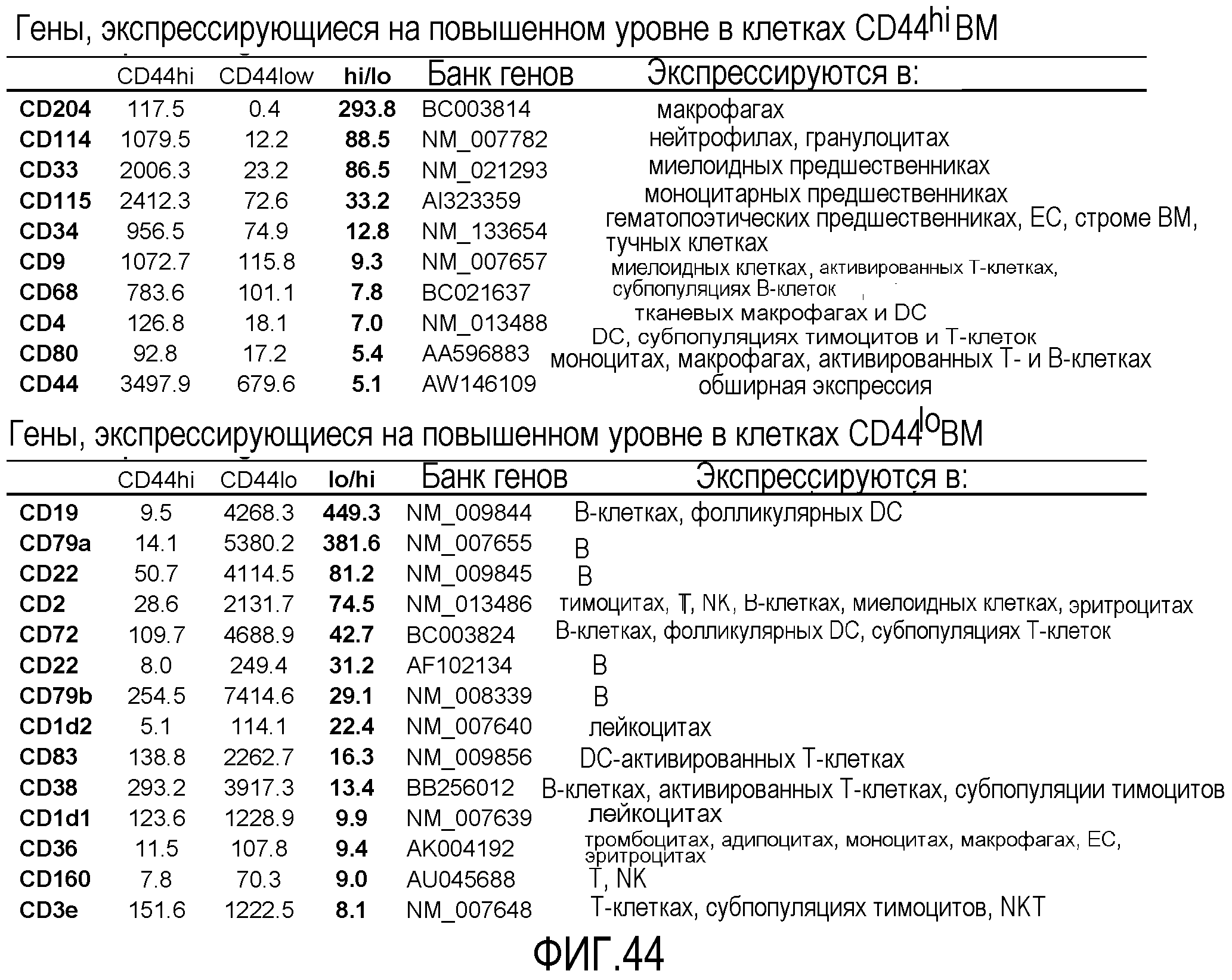

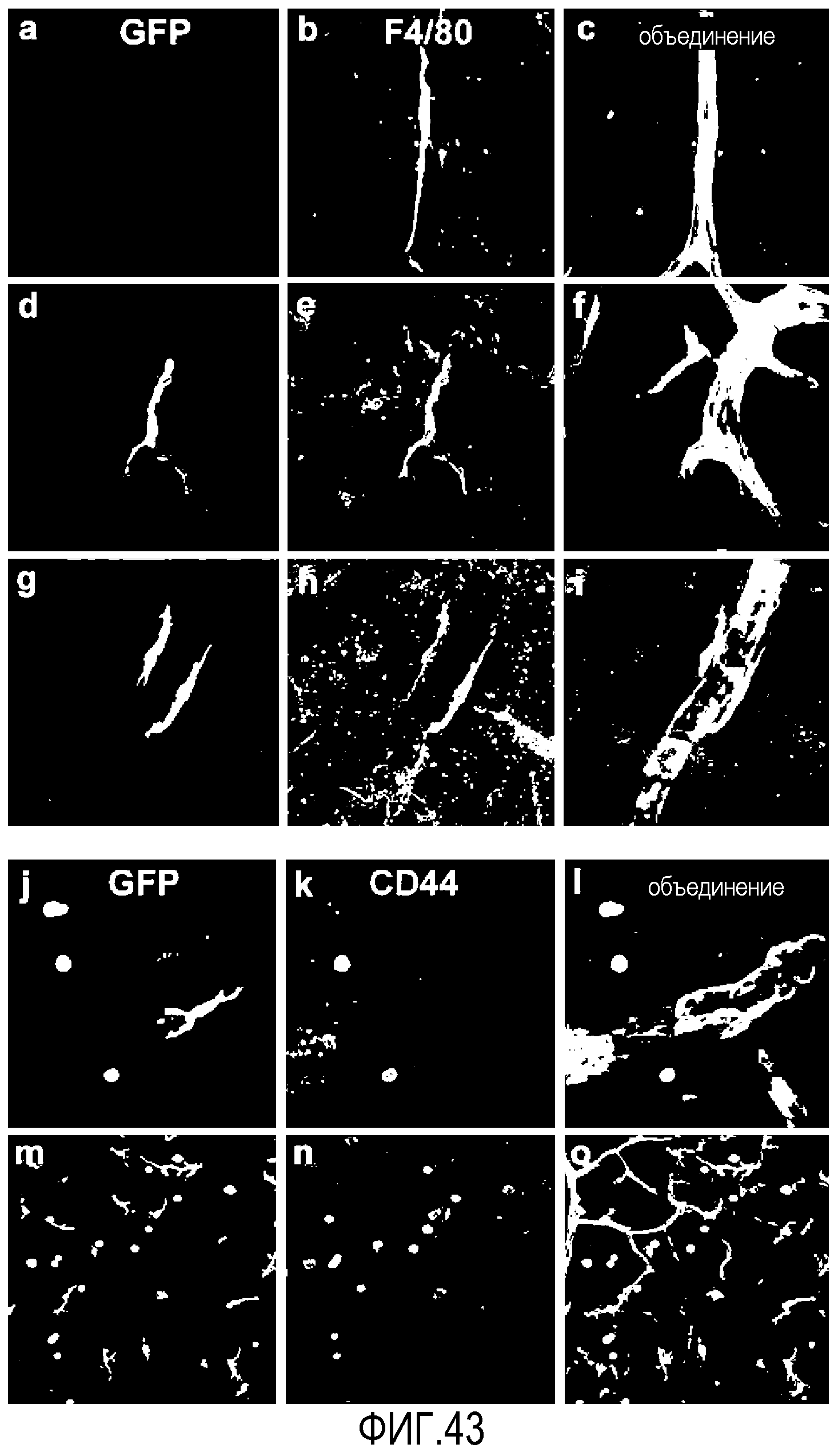
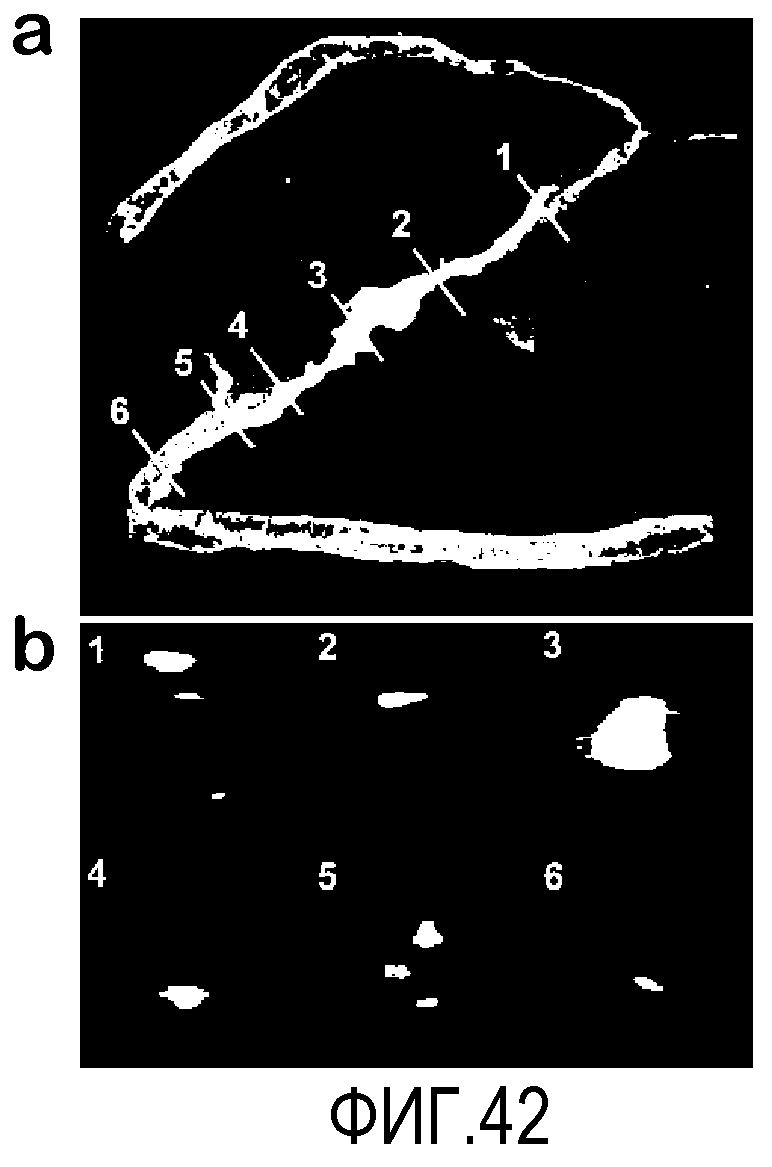



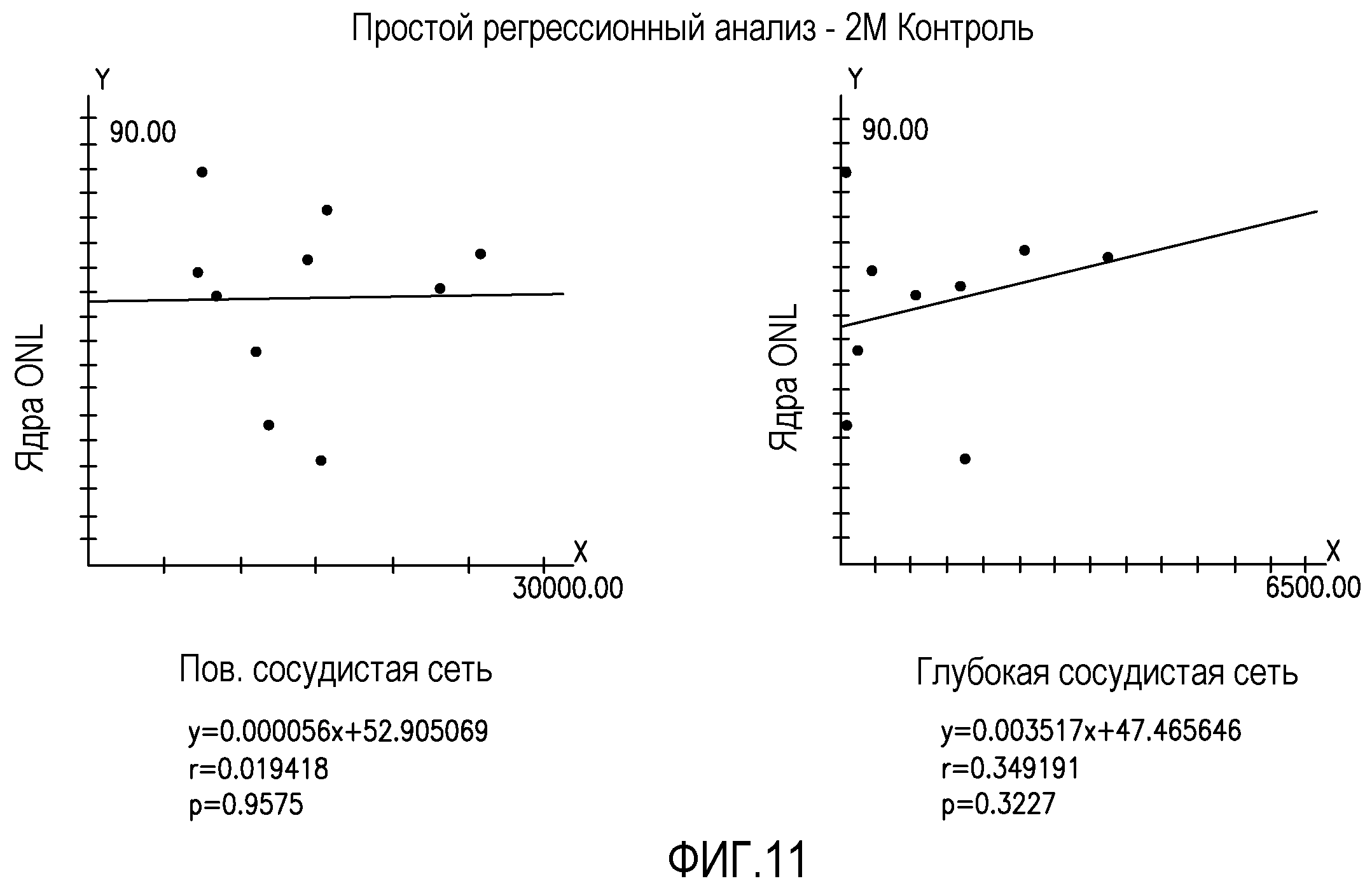
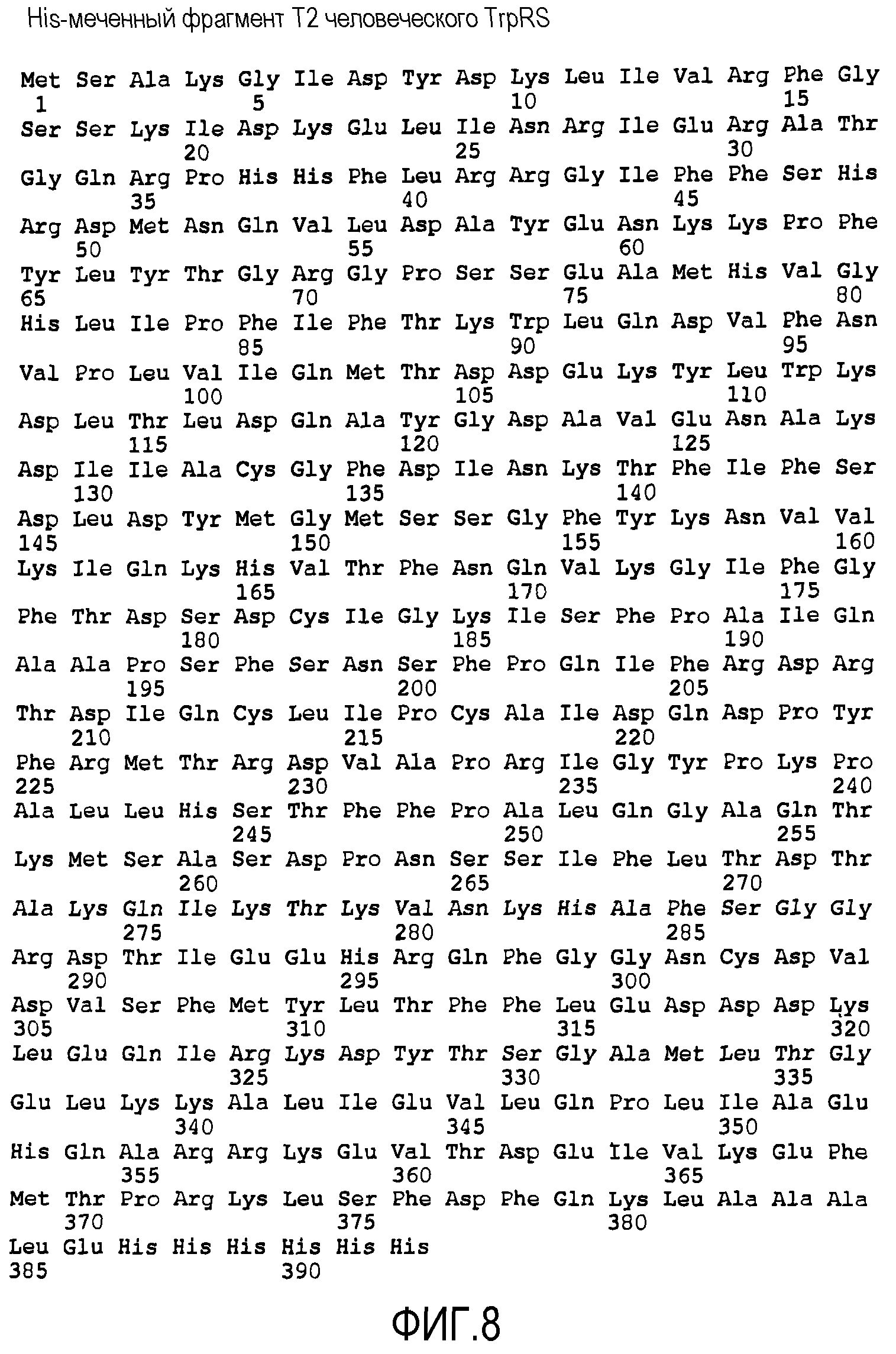
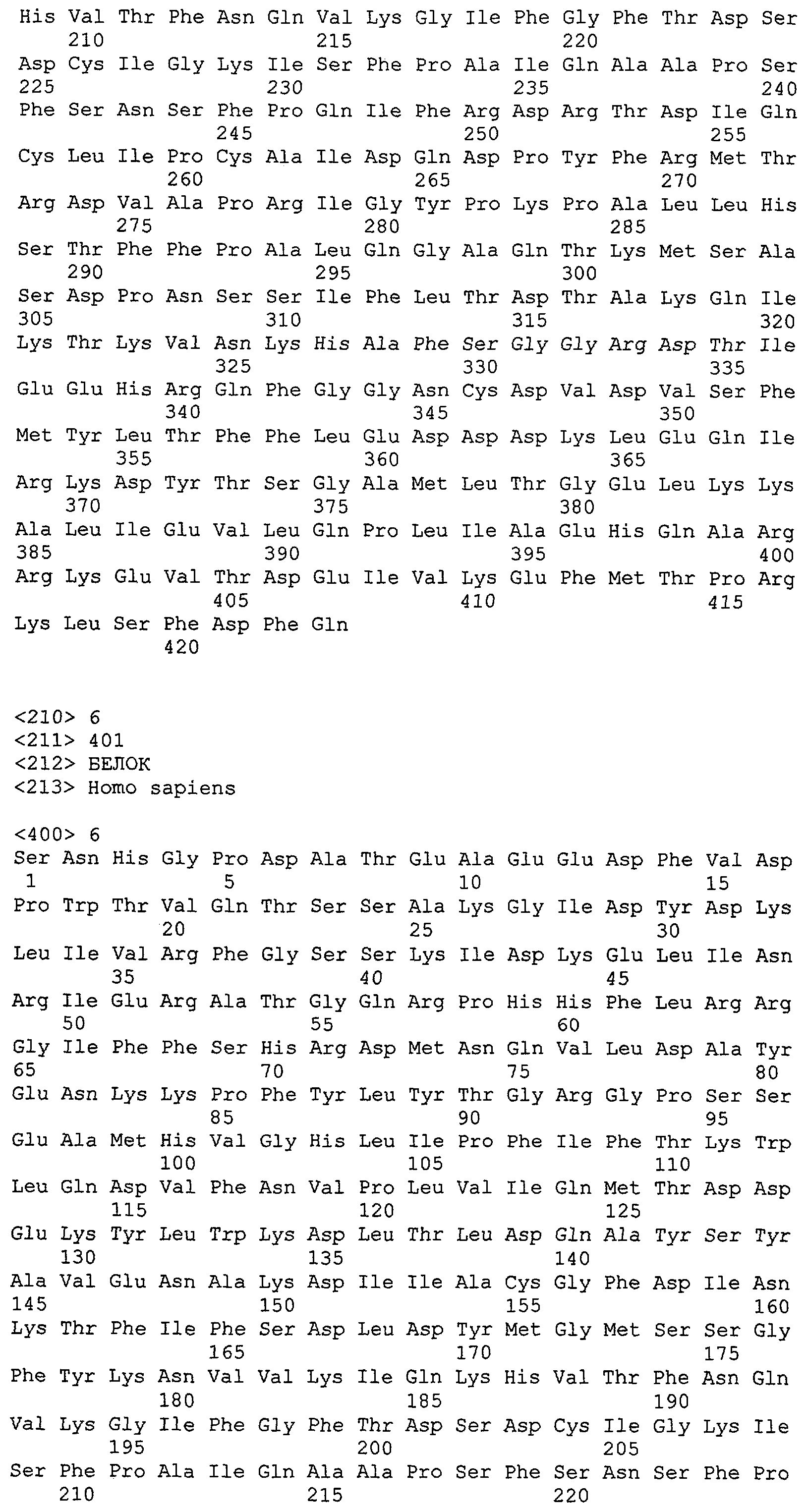

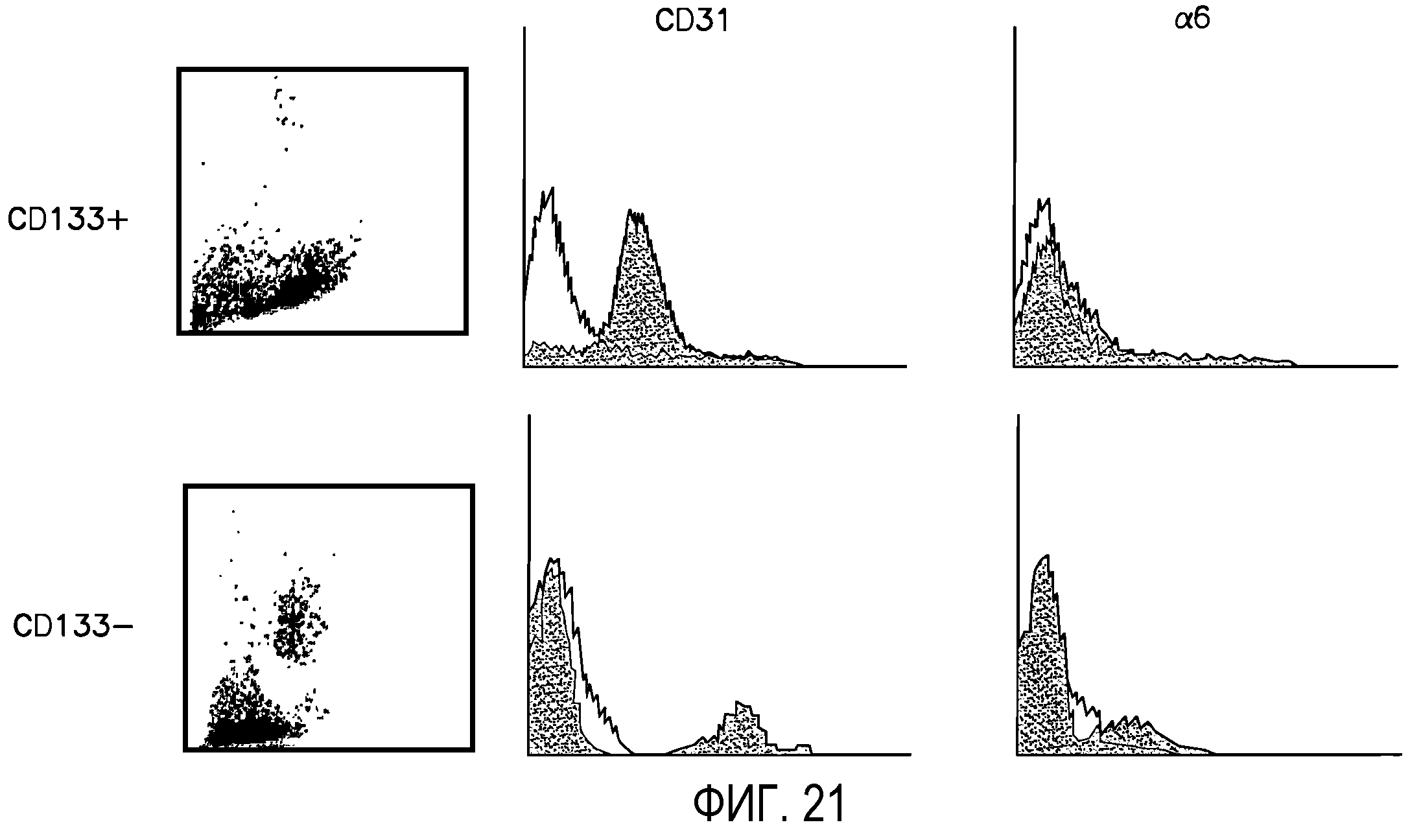
Основанная на наночастицах, нацеленная на опухоли доставка лекарственных средств
Днк-вакцина против пролиферирующих эндотелиальных клеток и способы ее применения
Гематопоэтические стволовые клетки и способы лечения неоваскулярных заболеваний глаз с их помощью
Гематопоэтические стволовые клетки и способы лечения неоваскулярных заболеваний глаз с их помощью
Способы и композиции для модуляции ангиогенеза с использованием тирозинкиназы src
Основанная на наночастицах, нацеленная на опухоли доставка лекарственных средств
Гематопоэтические стволовые клетки и способы лечения неоваскулярных заболеваний глаз с их помощью
Гематопоэтические стволовые клетки и способы лечения неоваскулярных заболеваний глаз с их помощью

