Результат интеллектуальной деятельности: 3β-(БЕНЗИЛОКСИ)-17α-МЕТИЛ-ПРЕГН-5-ЕН-20-ОН ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ КОГНИТИВНЫХ РАССТРОЙСТВ
Вид РИД
Изобретение
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗРОБРЕТЕНИЕ
Настоящее изобретение относится к области психических расстройств, и, в частности, к области когнитивных расстройств. Изобретение относится к конкретному производному прегненолона, "3β-(бензилокси)-17α-метил-прегн-5-ен-20-ону" (3Bn17MeP), которое не может быть метаболизировано в активные метаболиты прегненолона, и к его применению для лечения когнитивных расстройств.
УРОВЕНЬ ТЕХНИКИ
Когнитивные расстройства определяют как расстройства, приводящие к нарушению когнитивной функции человека, что делает независимую жизнь в обществе затруднительной или невозможной без лечения.
Когнитивные расстройства относятся к нарушениям в любых сферах когнитивного функционирования, включая обучение и память (например, кратковременную и долговременную память, память усвоения, консолидации, вспоминания и распознавания), исполнительные функции (например, рабочую память, когнитивную гибкость, принятие решений), комплексное внимание, язык, перцептивно-моторные способности или социальное познание. Когнитивные функции необходимы для деятельности людей в целом. Следовательно, когнитивные расстройства влияют на личные, образовательные или профессиональные социальные достижения, что в конечном итоге приводит к потере способности к самостоятельному передвижению и к самообслуживанию.
Когнитивные расстройства могут быть вызваны очень различными по своей природе заболеваниями и могут проявляться на любом этапе жизни. Например, когнитивные расстройства могут быть следствием генетического врожденного дефекта, физического повреждения головного мозга, нейродегенеративного заболевания или снижения когнитивных способностей, связанного со старением.
Согласно пятому изданию Руководства по диагностике и статистическому учету психических расстройств (Fifth Edition of the Diagnostic and Statistical Manual of Mental Disorders (DSM-5™)), когнитивные расстройства включают в себя две категории заболеваний, которые отличаются параметрами развития и продолжительностью жизни: нарушения неврологического развития и нейрокогнитивные расстройства.
Нарушения неврологического развития представляют собой группу психических нарушений, которые проявляются в раннем периоде развития (то есть до поступления в начальную школу). Как правило, нарушения неврологического развития могут быть вызваны врожденными генетическими дефектами (например, синдромом Дауна, синдромом ломкой Х-хромосомы), аномалиями развития (например, недостаточным развитием мозга), материнскими заболеваниями (например, плацентарным заболеванием) или перинатальными факторами окружающей среды (например, воздействием алкоголя на плод).
Нарушения неврологического развития характеризуются умственной отсталостью, адаптационными и коммуникативными нарушениями. Последствия для лиц с нарушениями неврологического развития варьируются в зависимости от тяжести когнитивных нарушений, но всегда препятствуют личной, общественной и образовательной деятельности. Например, при синдроме Дауна, который является наиболее частой генетической причиной умственной отсталости, когнитивные нарушения в основном проявляются снижением памяти и исполнительных функций (Grieco et al., 2015). Происходит задержка во времени восприятия языка и письменного текста, и умственное развитие у взрослых не превышает уровня развития 8 летнего ребенка с коэффициентом умственного развития <70. Следовательно, люди с синдромом Дауна являются в высокой степени зависимыми, нуждающимися, например, в специальной и индивидуальной образовательной поддержке и подвергающимися высокому уровню безработицы (например, 50% в США). Более половины людей с синдромом Дауна характеризуются неадаптированным поведением и/или затруднениями в установлении социальных отношений (Grieco et al., 2015; Kumin and Schoenbrodt, 2016).
Нарушения неврологического развития широко распространены. Самой по себе умственной отсталостью страдает 1% населения, при этом синдромом Дауна составляет 0,1% случаев (DSM-5 ™). Однако в настоящее время отсутствует лекарственное средство, одобренное для лечения когнитивных нарушений, вызванных нарушениями неврологического развития.
Нейрокогнитивные расстройства представляют собой группу психических расстройств, характеризующихся ключевыми когнитивными нарушениями, который представляет собой снижение когнитивного уровня, достигнутого ранее. Этиология нейрокогнитивных расстройств включает нейродегенеративные расстройства (например, болезнь Альцгеймера, лобно-височную лобарную дегенерацию, болезнь Паркинсона), сосудистые заболевания, травматические повреждения головного мозга, злоупотребление психоактивными веществами/ лекарственными препаратами, инфекцию, вызванную вирусом иммунодефицита человека (ВИЧ), или прионную инфекцию.
В зависимости от степени воздействия на способность человека с нейрокогнитивными расстройствами управлять самим собой, нейрокогнитивные расстройства могут подразделять на легкие формы (также называемые умеренными когнитивными нарушениями) или основные формы (также называемые деменцией).
Основные нейрокогнитивные расстройства приводят к неспособности выполнять без посторонней помощи повседневные действия, такие как оплата счетов или соблюдение режима приема лекарств. Лица с легкими нейрокогнитивными нарушениями остаются независимыми, но за счет больших затрат усилий и использования компенсационных стратегий (DSM-5TM).
Нейрокогнитивные расстройства вызываются многими заболеваниями, поэтому их распространенность среди населения мира трудно оценить. Однако среди пожилого населения, основные нейрокогнитивные расстройства поражают до 2% людей в возрасте 65 лет и до 30% людей в возрасте 85 лет. Распространенность легких нейрокогнитивных расстройств достигает 10% в возрасте 65 лет и 25% в возрасте старше 70 лет (DSM-5TM). Рост частоты возникновения нейрокогнитивных расстройств взаимосвязан с процессом постоянно продолжающегося старения населения мира. Таким образом, создание методов лечения, позволяющих улучшать когнитивные функции, является чрезвычайно актуальной задачей.
В настоящее время одобрено применение фармакологических препаратов с целью ограничения или отсрочки снижения когнитивных способностей, связанного с болезнями Альцгеймера и Паркинсона. Эти фармакологические препараты представляют собой ингибиторы ацетилхолинэстеразы (например, донепезил, ривастигмин, галантамин) и ингибитор глутаматергических рецепторов N-метил-D-аспартата (NMDA) (мемантин). Недавно в разных странах были проведены исследования по их эффективности и переносимости у пациентов (Haute Autorité de Santé, 2016; National Institute for Health and Care Excellence, 2011; Breton et al., 2015). На основании этих исследований был сделан вывод, что эти лекарственные препараты не представляют особого терапевтического интереса по следующим причинам:
- Отсутствие долговременного эффекта по ограничению или отсрочки снижения когнитивных способностей.
- Отсутствие умеренного кратковременного эффекта по ограничению или отсрочки снижения когнитивных способностей.
- Неэффективность в отношении улучшения познавательных способностей.
- Существенный риск возникновения неблагоприятных эффектов; сообщалось о желудочно-кишечных, сердечно-сосудистых и психоневрологических осложнениях. Частота возникновения этих неблагоприятных эффектов является высокой, в результате чего в 30% случаев лечению прекращают (Haute Autorité de Santé, 2016).
Кроме того, проводились испытания соединения обоих этих фармакологических классов на пациентах с нарушениями неврологического развития, но они не дали положительных терапевтических результатов, например, при синдроме Дауна и синдроме ломкой Х-хромосомы. (Hanney et al., 2012; Kishnani et al., 2010; NCT01120626; NCT00584948, https://clinicaltrials.gov/).
Вследствие этого, для лечения когнитивных расстройств необходимо создание эффективных и безопасных фармакологических препаратов.
Было показано, что применение соединений (ортостерических антагонистов), блокирующих активность рецептора CB1 посредством ингибирования его ортостерического сайта связывания, сайта, в котором связываются эндоканнабиноиды для активации рецептора, облегчает когнитивные нарушения при синдроме ломкой Х-хромосомы (Busquets-Garcia et al., 2013). Одно из этих соединений, римонабант, было выпущено на рынок под торговой маркой Acomplia®. К сожалению, доступные ортостерические антагонисты, такие как римонабант, подавляют полностью активность рецептора, а также могут действовать как обратные агонисты рецептора CB1, то есть они не только ингибируют активацию CB1, но также индуцируют противоположные сигнальные ответы рецептора. Из-за этого обратного агонистического действия и полного ингибирования активности рецептора, доступные методы, основанные на введении ортостерических антагонистов CB1, также характеризуются рядом серьезных неблагоприятных эффектов. Из-за этих неблагоприятных эффектов, серийное производство препарата Acomplia® было приостановлено, а разработка других методов ингибирования ортостерического сайта CB1 было прекращено.
Ортостерические антагонисты CB1 и, в частности, препарат Acomplia®, обладают приведенными ниже известными неблагоприятными эффектами, которые делают их непригодными для лечения когнитивных расстройств.
1. Они снижают потребление пищи, что указывает на общее нарушение системы поощрения головного мозга.
2. Они вызывают тревожное поведение у животных и ощущение тревогу у людей.
3. Они вызывают связанное с депрессией поведение у животных и депрессивный синдром у людей.
4. Они вызывают судороги и общие поведенческие и клинические нарушения при проведении исследований по безопасности и токсикологии лекарственных средств.
5. Они обладают гепатотоксическим действием.
6. Они обладают неспецифическим действием, ингибируя, например, вызываемую морфином анальгезию в результате прямого взаимодействия с опиоидными рецепторами (Seely et al., 2012).
Очевидно, что эти эффекты делают невозможным применение антагонистов CB1 в качестве метода лечения когнитивных расстройств, в частности потому, что когнитивными расстройствами страдают группы молодых и пожилых людей, которые являются более уязвимыми и/или ослабленными.
Недавно было обнаружено, что при гиперактивации рецептора CB1, концентрация стероидного гормона прегненолона увеличивается (3000%) в головном мозге. Затем прегненолон связывается со специфическим сайтом рецептора CB1, отличным от сайта, связываемого агонистами CB1, и действует как специфический ингибитор эндогенной передачи сигналов рецептора CB1 (eCB1-SSi). Таким образом, прегненолон селективно ингибирует CB1-индуцированную активацию пути MAPK (митоген-активированная протеинкиназа), а не CB1-индуцированное ингибирование аденилатциклазы. Несмотря на это ограниченное молекулярное действие, прегненолон подавляет большинство поведенческих эффектов, вызванных гиперактивацией рецептора CB1 у грызунов (Vallée et al., 2014).
К сожалению, прегненолон нельзя применять в качестве медикаментозного лечения, потому что он малодоступен, имеет очень короткий период полувыведения и по ходу метаболизма превращается в активные стероиды.
В патентном документе WO2012/160006 в качестве производного прегненолона раскрыт 3β-(бензилокси)-17α-метил-прегн-5-ен-20-он, который не метаболизируется в стероиды, и его применение для ингибирования рецептора CB1.
Разработка метода лечения когнитивных расстройств, основанного на специфическом ингибировании активации сигнального пути рецептора CB1, который может применяться на людях, представляет несколько новых возможностей. Следовательно, такое соединение должно одновременно обладать всеми приведенными ниже характеристики.
1. Оно должно улучшать показатели в различных сферах когнитивного функционирования, учитывая вариабельность когнитивных симптомов у пациентов с когнитивными расстройствами.
2. Оно должно демонстрировать эффективность в экспериментальных моделях, которые приблизительно моделируют когнитивные процессы человека, для того чтобы достигать лучшего переноса результатов моделирования применительно к человеку.
3. Оно должен улучшать когнитивную деятельность при ряде заболеваний/состояний с учетом неоднородности этиологий когнитивных расстройств.
4. Оно не должно иметь неблагоприятных эффектов, свойственных ингибиторам CB1. В частности, оно не должно вызывать: a) снижения потребления пищи; b) усугубления поведения, связанного с тревогой и депрессией; c. судорог и клинических признаков нарушения центральной нервной системы.
5. Оно не должно модифицировать связывание других рецепторов, во избежание нецелевых эффектов или модификации активности других терапевтических препаратов, как, например, это происходит в случае римонабанта.
6. Оно не должно оказывать неспецифических влияний на поведение, включающих, но этим не ограничивая, седативный эффект, возбудимость, измененное спонтанное поведение, которые могут препятствовать проявлению его терапевтических эффектов.
7. Оно должно иметь приемлемый запас безопасности. Общепризнано, что максимальная доза, не вызывающая неблагоприятного эффекта, должна быть как минимум в 10 раз выше терапевтической дозы.
Ни для одного из ранее описанных специфических ингибиторов активации сигнального пути и других антагонистов рецепторов CB1 не были описаны все упомянутые выше ключевые характеристики. Насколько известно авторам изобретения, ни одно соединение, применяемое для лечения когнитивных расстройств или, в более общем смысле, поведенческих расстройств, не обладает всеми этими характеристиками. Так, например, три основных класса психоактивных лекарственных средств, анксиолитики, антидепрессанты и нейролептики, а также одобренные лекарственные средства для лечения когнитивных расстройств, связанных с болезнью Альцгеймера, вызывают неблагоприятные поведенческие эффекты в диапазоне их терапевтических доз. Например: а) анксиолитические препараты вызывают сонливость, снижают концентрацию внимания и ухудшают память; b) антидепрессанты вызывают возбудимость, бессонницу и снижение либидо; c) нейролептики вызывают гормональные нарушения, седативный эффект, дискинезию и непроизвольные движения. d) ингибиторы ацетилхолинэстеразы и мемантин вызывают неблагоприятные эффекты со стороны желудочно-кишечного тракта, сердечно-сосудистой системы и нервнопсихической системы.
Следовательно, соединение, обладающее всеми характеристиками, описанными в пунктах с 1 по 7, могло бы стать важной инновацией в области лекарственных средств, предназначенных для лечения когнитивных расстройств, а также и важной инновацией в области психиатрии в целом.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится в целом к конкретному производному прегненолона и к его применению для лечения когнитивного расстройства.
Более конкретно, изобретение относится к соединению формулы (I)
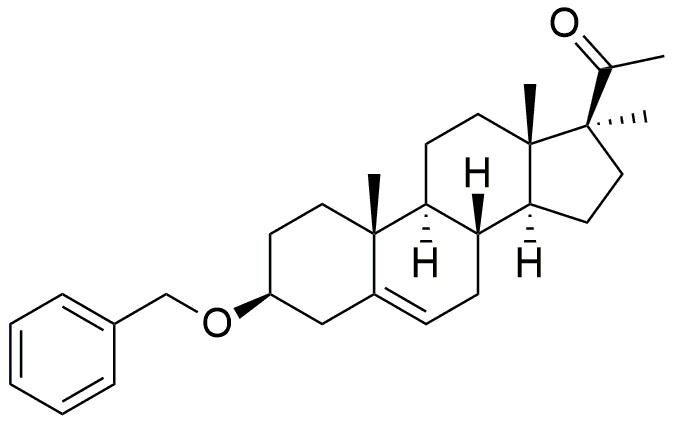
Формула (I)
и его применению при лечении когнитивных расстройств.
Согласно изобретению, лечение когнитивных расстройств с помощью соединения формулы (I) включает:
- Полное или частичное купирование нарушения, вызванного когнитивным расстройством, в том числе улучшение когнитивной функции.
- Предотвращение дальнейшего снижения когнитивных способностей.
- Отсрочка или замедление снижения когнитивных способностей или дальнейшей утраты когнитивных способностей.
Настоящее изобретение наделено следующими отличительными признаками.
1. Оно улучшает когнитивное функционирование в различных сферах когнитивного процесса (например, декларативная/социальная память, распознавание, исполнительные функции). Поэтому оно сможет соответствовать критериям эффективности практически для всех видов когнитивных расстройств.
2. Оно улучшает результаты выполнения когнитивных тестов, которые успешно применяются для обнаружения когнитивных нарушений как у людей (в виртуальном радиальном лабиринте), так и у мышей (в радиальном лабиринте) с использованием очень похожих методик.
3. Оно улучшает когнитивную деятельность в различных экспериментальных моделях заболеваний/состояний (например, синдрома Дауна, синдрома ломкой X-хромосомы, когнитивных расстройств, связанных со старением). Поэтому оно сможет соответствовать критериям эффективности для когнитивных расстройств какого угодно происхождения.
4. Оно не имеет неблагоприятных эффектов, свойственных ингибиторам CB1. Оно не вызывает: a) снижения потребления пищи и массы тела; b) усугубления поведения, связанного с тревогой и депрессией как при нормальных, так и при патологических состояниях; c) нейротоксичности, судорог и клинических признаков нарушения центральной нервной системы. d) гепатотоксичности и генотоксичности.
5. Оно не модифицирует связывание широкого набора рецепторов (85), в том числе опиоидных рецепторов.
6. Оно не оказывает неспецифических влияний на поведение, включающих, но этим не ограничивая, седативный эффект, возбудимость, измененное спонтанное поведение, которые могут препятствовать проявлению его терапевтических эффектов.
7. Оно имеет очень большой запас безопасности (>3500).
Это изобретение также относится к способу лечения описанного в изобретении когнитивного расстройства путем введения субъекту, нуждающемуся в этом, эффективного количества соединения формулы I. В некоторых примерах, субъект, нуждающийся в проведении терапии, страдает от синдрома ломкой X-хромосомы, синдрома Дауна или от связанного с возрастом снижения когнитивных способностей.
Это изобретение также относится к фармацевтической композиции для лечения описанного в изобретении когнитивного расстройства, включающей, в качестве активного ингредиента, соединение формулы I. В некоторых примерах, фармацевтическая композиция предназначена для лечения субъекта с синдромом ломкой X-хромосомы, синдромом Дауна или со связанным с возрастом снижением когнитивных способностей.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
ФИГУРА 1. Cхема синтеза соединения формулы (I) (3Bn17MeP) методом А.
ФИГУРА 2. Cхема синтеза соединения формулы (I) (3Bn17MeP) методом B.
ФИГУРА 3. Cхема синтеза соединения формулы (I) (3Bn17MeP) методом C.
ФИГУРА 4. Cхема синтеза соединения формулы (I) (3Bn17MeP) методом D.
ФИГУРА 5. In vitro воздействия 3Bn17MeP на ингибирование клеточного дыхания, на увеличение степени фосфорилирования митоген-активируемых киназ (p-MAPK) и на снижение содержания циклического аденозинмонофосфата (cAMP), вызванное стимуляцией рецепторов CB1 Δ9-тетрагидроканнабинолом (THC).
ФИГУРА 5A. Воздействие 3Bn17MeP (1; 2,5; 5; 50 и 100 нМ) на индуцированное THC (1 мкМ) ингибирование клеточного дыхания в клетках линии HEK293, трансфицированных рецептором CB1 человека дикого типа. Данные выражены в процентах воздействия THC (пунктирная линия при 0% представляет воздействие носителя THC; пунктирная линия при 100% представляет воздействие THC в отсутствие 3Bn17MeP). 3Bn17MeP дозозависимо блокирует ингибирование клеточного дыхания, вызванное THC, со значительными эффектами от 2,5 нМ. р <0,001, THC по сравнению с носителем THC в отсутствие 3Bn17MeP, критерий Тьюки. p <0,001, 3Bn17MeP (2,5; 5; 50 и 100 нМ) по сравнению с его носителем в присутствии THC, критерий Тьюки.
ФИГУРА 5B. Воздействие 3Bn17MeP (100 нМ) на индуцированное THC (1 мкМ) ингибирование клеточного дыхания в клетках линии HEK293, трансфицированных рецептором CB1 человека дикого типа (HEK293-hCB1-WT) или мутантным рецептором CB1 человека (HEK293-hCB1- Mut), в котором сайт связывания прегненолона был сделан недействующим (hCB1p.E1.49G; Vallée et al., 2014). 3Bn17MeP ингибирует вызванное THC (1 мкМ) снижение клеточного дыхания в клетках линии HEK293, трансфицированных hCB1-WT (белая полоса), тогда как 3Bn17MeP не ингибирует этот эффект THC в клетках линии HEK293, трансфицированных с помощью hCB1-Mut. p <0,01, hCB1-WT по сравнению с hCB1-Mut в присутствии 3Bn17MeP и THC, критерий Стьюдента для одной выборки).
ФИГУРА 5C. Воздействие 3Bn17MeP (0,1; 0,3; 1; 3 и 9 мкМ) на вызванное THC (10 мкМ) увеличение степени фосфорилирования MAPK (p-Erk1/2MAPK). 3Bn17MeP ингибирует вызванное THC увеличение степени фосфорилирования MAPK.
ФИГУРА 5D. Воздействие 3Bn17MeP на вызванное THC снижение уровней cAMP в клетках линии CHO, стабильно трансфицированных с помощью hCB1. 3Bn17MeP не изменял снижение cAMP, вызванное THC. NT, без обработки с помощью THC.
ФИГУРА 6. Воздействие 3Bn17MeP на нарушение распознавания объектов в доклинической модели синдрома Дауна.
ФИГУРА 6A. 3Bn17MeP восстанавливает кратковременное последовательное распознавание объектов у мышей линии Ts65Dn после его длительного перорального введения (0,6 мкг/мл) с питьевой водой. **р <0,01; ***p <0,001, 1-е представление объекта против 2-го представления того же объекта после задержки в 5 мин, и ##p <0,01; ###p <0,001, после задержки в 35 мин (тест Фишера).
ФИГУРА 6B. 3Bn17MeP восстанавливает долговременное распознавание объектов у мышей линии Ts65Dn после его повторных пероральных введений в кукурузном масле (15 мкг/кг; два раза в день). ***p <0,001, мыши дикого типа (WT) по сравнению с мышами линии Ts65Dn, которым вводили носитель, используемый для 3Bn17MeP, (0 мкг/кг) и ###p <0,001, носитель 3Bn17MeP (0 мкг/кг) по сравнению с 3Bn17MeP (15 мкг/кг) у мышей линии Ts65Dn (критерий Тьюки). Нет значимой разницы между мышами дикого типа (WT) и мышами линии Ts65Dn, которым вводили 3Bn17MeP (15 мкг/кг, критерий Тьюки).
ФИГУРА 7. Воздействие 3Bn17MeP на нарушение декларативной/реляционной памяти в доклинической модели синдрома Дауна. Длительное пероральное введение 3Bn17MeP с питьевой водой (0,6 мкг/мл) мышам линии Ts65Dn восстанавливает характеристики их памяти (измеряемые процентом правильных ответов) до уровня мышей дикого типа (WT). Пунктирная линия показывает уровень случайности. *p <0,05, мыши дикого типа (WT) по сравнению с мышами линии Ts65Dn, которым вводили носитель для 3Bn17MeP (0 мкг/мл; критерий Стьюдента для одной выборки). #p <0,05, носитель для 3Bn17MeP (0 мкг/мл) против 3Bn17MeP (0,6 мкг/мл) у мышей линии Ts65Dn (критерий Стьюдента для одной выборки). Отсутствие значимого эффекта между мышами дикого типа WT и мышами линии Ts65Dn, которым вводили 3Bn17MeP (0,6 мкг/мл; критерий Стьюдента для одной выборки).
ФИГУРА 8. Воздействие 3Bn17MeP на нарушение рабочей памяти в доклинической модели синдрома Дауна. Длительное пероральное введение 3Bn17MeP с питьевой водой (0,06 мкг/мл) мышам линии Ts65Dn восстанавливает характеристики их памяти (измеряемые процентом правильных ответов) до уровня мышей дикого типа (WT). ***p <0,001, мыши дикого типа (WT) по сравнению с мышами линии Ts65Dn, которым вводили носитель для 3Bn17MeP (носитель; критерий Тьюки). ##p <0,01, носитель против 3Bn17MeP (0,06 мкг/мл) у мышей линии Ts65Dn (критерий Тьюки). Отсутствие значимого эффекта между мышами дикого типа WT и мышами линии Ts65Dn, которым вводили 3Bn17MeP (0,06 мкг/мл; критерий Тьюки).
ФИГУРА 9. Воздействие 3Bn17MeP на нарушение распознавания объектов в доклинической модели синдрома ломкой Х-хромосомы. Повторные пероральные введения 3Bn17MeP в кукурузном масле (15 мкг/кг; два раза в сутки) корректируют снижение индекса различения у мышей линии fmr1-KO относительно мышей дикого типа (WT). **p <0,01, мыши дикого типа (WT) по сравнению с мышами линии fmr1-KO, которым вводили носитель для 3Bn17MeP (0 мкг/кг; критерий Тьюки). #p <0,05, носитель 3Bn17MeP (0 мкг/кг) по сравнению с 3Bn17MeP (15 мкг/кг) у мышей линии fmr1-KO (критерий Тьюки). Отсутствие значимого эффекта между мышами дикого типа WT и мышами линии fmr1-KO, которым вводили 3Bn17MeP (15 мкг/кг; критерий Тьюки).
ФИГУРА 10. Воздействие 3Bn17MeP на нарушение декларативной/реляционной памяти в доклинической модели возрастных когнитивных нарушений. Длительное пероральное введение 3Bn17MeP с питьевой водой (0,6 мкг/мл) улучшает характеристики памяти у пожилых мышей (измеряемые процентом правильных ответов). Пунктирная линия показывает уровень случайности. *p<0,05, носитель 3Bn17MeP (0 мкг/мл) при сравнении с 3Bn17MeP (0,6 мкг/мл; критерий Стьюдента для одной выборки).
ФИГУРА 11. Воздействия повторных введения 3Bn17MeP и римонабанта на снижение потребления пищи и массы тела у мышей дикого типа.
ФИГУРА 11A. Воздействия однократное введение 3Bn17MeP (0; 0.05; 5; 15 и 30 мг/кг; перорально в кукурузном масле) на суммарное снижение потребления стандартного корма, измеряемого через 3 и 13 часов после выключения света. 3Bn17MeP не оказывает воздействия на снижение потребления пищи.
ФИГУРА 11B. Воздействия однократного введения римонабанта (0; 10 мг/кг; интраперитонеально) на суммарное снижение потребления стандартного корма, измеряемое через 3 и 13 часов после выключения света. Римонабант снижал потребление пищи. **p<0,01; римонабант по сравнению с носителем (0 мг/кг; двухфакторный дисперсионный анализ, основной эффект лечения).
ФИГУРА 11C. Воздействия повторных (39 дней, один раз в сутки) введений 3Bn17MeP (0, 0,05; 5; 15 и 30 мг/кг, перорально в кукурузном масле) на массу тела мышей, которых кормили стандартным кормом. 3Bn17MeP не оказывает воздействия на массу тела.
ФИГУРА 11D. Воздействия повторных (39 дней, один раз в сутки) введений римонабанта (0 и 10 мг/кг, интраперитонеально) на массу тела мышей, которых кормили стандартным кормом. Римонабант снижал массу тела.
ФИГУРА 12. Воздействия 3Bn17MeP и римонабанта на поведение, связанное с тревогой и депрессией, у мышей дикого типа.
ФИГУРА 12A. Воздействия однократного введения 3Bn17MeP в кукурузном масле (30 мг/кг) или носителя (0 мг/кг) на поведение, подобное состоянию тревоги, измеряемое процентом затраченного времени и посещений в открытых рукавах приподнятого крестообразного лабиринта. 3Bn17MeP не оказывает воздействия на связанное с тревогой поведение.
ФИГУРА 12B. Воздействия однократного интраперитонеального введения римонабанта (10 мг/кг) или носителя (0 мг/кг) на поведение, подобное состоянию тревоги, измеряемое процентом затраченного времени и посещений в открытых рукавах приподнятого крестообразного лабиринта. Римонабант снижает затраченное время и число посещений в открытые рукава. ***p<0,001; **p<0,01; римонабант по сравнению с носителем (0 мг/кг) (критерий Стьюдента для одной выборки).
ФИГУРА 12C. Воздействия повторных (28 дней, один раз в сутки) введений 3Bn17MeP (0, 0,05; 5; 15 и 30 мг/кг, перорально в кукурузном масле) на поведение, связанное с депрессией, измеряемые в тесте на предпочтение сахарозы. 3Bn17MeP не оказывает воздействия на потребление сахарозы.
ФИГУРА 12D. Воздействия повторных (28 дней, один раз в сутки) введений римонабанта (0 и 10 мг/кг; интраперитонеально) на связанное с депрессией поведение, измеряемые в тесте на предпочтение сахарозы. Римонабант снижает потребление сахарозы. *p<0,05, римонабант по сравнению с носителем (0 мг/кг; критерий Стьюдента для одной выборки).
ФИГУРА 13. Оценка нейротоксичности и генотоксичности 3Bn17MeP и римонабанта in vitro.
ФИГУРА 13A. Воздействия 3Bn17MeP, римонабанта (0; 10; 30 и 100 мкM) и референсного положительного контроля стауроспорина (100 нM) на токсичность в отношении первичных культур кортикальных нейронов крысы. 3Bn17MeP не оказывает токсического действия в этих условиях, тогда как римонабант проявляет дозозависимое токсическое действие в этих условиях. ***p<0,001; *p<0,05, римонабант по сравнению с носителем (0 мкM) (критерий Холма-Сидака).
ФИГУРА 13B. Воздействие 3Bn17MeP (0; 0,1; 0,3; 1; 3; 10; 30 и 100 мкM), римонабанта (0; 0,1; 0,3; 1 и 100 мкM) и референсного положительного контроля этопозида (3 мкM) на фосфорилирование гистона H2AX в клетках линии HeLa. 3Bn17MeP не воздействует на фосфорилированные H2AX клетки, в то время как римонабант повышает число фосфорилированных H2AX клеток при 100 мкM. ***p<0,001, римонабант по сравнению с носителем (0 мкM) (критерий Холма-Сидака).
ФИГУРА 14. Оценка гепатотоксичности 3Bn17MeP и римонабанта in vitro.
ФИГУРА 14A. Воздействие 3Bn17MeP, римонабанта (0; 0,1; 0,3; 1; 3; 10; 30 и 100 мкM) и референсного положительного контроля ацетаминофена (APAP, 50 мM) на выживаемость первичных культур гепатоцитов крыс. 3Bn17MeP не проявляет гепатотоксического действия, в то время как римонабант увеличивает гибель гепатоцитов при концентрациях от 3 мкM. ***p<0,001, римонабант по сравнению с носителем (0 мкM) (критерий Холма-Сидака).
ФИГУРА 14B. Воздействие 3Bn17MeP, римонабанта (0; 0,1; 0,3; 1; 3; 10; 30 и 100 мкM) и референсного положительного контроля циклоспорина A (10 мкM) на ингибирование желчных канальцев в первичных культурах гепатоцитов крыс. 3Bn17MeP не оказывает воздействие на желчные канальцы, в то время как римонабант снижает число канальцев при концентрациях от 1 мкM. *** p<0,001; **p<0,01, римонабант по сравнению с носителем (0 мкM) (однофакторный дисперсионный анализ с последующим апостериорным тестом Бонферрони).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение в целом относится к соединению формулы (I):
Формула (I)
для применения при лечении когнитивного расстройства.
Соединение формулы (I) корректирует когнитивные нарушения, наблюдаемые при когнитивных расстройствах. В случае всех моделей когнитивных функциях и заболеваний, которые были подвергнуты оценке, введение 3β-(бензилокси)-17α-метил-прегн-5-ен-20-она (3Bn17MeP) полностью восстанавливает когнитивные характеристики до уровней здоровых людей, взятых в качестве контроля.
3Bn17MeP проявляет эффективность в нескольких сферах когнитивного процесса, включающих долговременную память, распознавание и исполнительные функции. Настоящее изобретение позволяет достигать положительного эффекта вследствие широкого спектра воздействий 3Bn17MeP на когнитивные функции. Следовательно, 3Bn17MeP, по-видимому, является универсальным усилителем когнитивных функций.
Соединение по настоящему изобретению, 3Bn17MeP, не блокирует всю активность рецептора-мишени, а только часть его активности. Специфическим рецептором-мишенью соединения по настоящему изобретению является рецептор CB1.
Считается, что соединение формулы (I) воспроизводит эффекты недавно открытого механизма головного мозга, который обеспечивает эндогенную отрицательную обратную связь, регулируя чрезмерную активацию рецептора CB1 (Vallée et al., 2014). Важно отметить, что этот регуляторный механизм запускается только тогда, когда CB1 чрезмерно активирован, что, вероятно, имеет место при когнитивном расстройстве, а не тогда, когда активация рецептора находится в более физиологическом диапазоне.
Этим может объяснить, почему соединение по настоящему изобретению является чрезвычайно эффективным для коррекции когнитивных расстройств и, следовательно, является эффективным для лечения когнитивных расстройств. Кроме того, вследствие его механизма действия, специфически направленного на активацию сигнального пути, соединение по настоящему изобретению не влияет на поведение как таковое у здоровых субъектов, у которых активация сигнального пути CB1 находится в физиологическом диапазоне.
Механизм действия соединения по настоящему изобретению в этом отношении сильно отличается от механизма действия ортостерических антагонистов CB1, которые, в результате блокирования связывание эндогенных и экзогенных агонистов рецептора CB1, вызывают полное ингибирование всей активности CB1 и нарушают поведение как таковое. Кроме того, не существует физиологического механизма действия антагонистов, то есть, насколько известно авторам изобретения, не существует эндогенных соединений, которые, подобно антагонистам, блокируют связывание агонистов CB1 с рецептором. В силу искусственной природы этого механизма действия, эти антагонисты, помимо коррекции чрезмерной активации рецептора-мишени, обычно снижают его активность ниже базальных уровней, нарушая физиологию и вызывая побочные эффекты.
Различие в механизме действия между соединением по настоящему изобретению и ортостерическими антагонистами, такими как римонабант, объясняет, почему оба препарата способны корректировать когнитивные расстройства, но не обладают другими общими для них воздействиями на поведение.
Следовательно, соединение по настоящему изобретению не является ортостерическим антагонистом CB1 и, как таковое, не блокирует все клеточные эффекты, вызванные активацией рецептора CB1, как это происходит в случае применения ортостерического антагониста CB1, римонабанта.
Еще более предпочтительным, в более широком смысле, является то, что при применении соединения по настоящему изобретению не обнаруживаются проявления неблагоприятных эффектов.
Отсутствие нежелательного эффекта и, во многих случаях, отсутствие воздействия соединения по настоящему изобретению на здоровых людей контрольной группы, вероятно, обусловлено специфической структурой и механизмом действия соединения по настоящему изобретению.
Авторы изобретения доказывают, что соединение по настоящему изобретению обладает сопутствующими характеристиками, которые делают его уникальным и идеальным средством для лечения когнитивных расстройств. Эти характеристики включают, но этим не ограничивая, перечисленные ниже характеристики.
1. Соединение по настоящему изобретению обладает уникальным механизмом действия преодоления чрезмерной активации рецептора CB1, адаптированным к эндогенному механизму головного мозга, но при этом, как представляется, оно не оказывает воздействия на базальную активность рецептора. Нарушение базальной активности целевой системы часто является причиной некоторых неблагоприятных эффектов, вызываемых антагонистами. Кроме того, соединение по настоящему изобретению действует очень селективно и не взаимодействует ни с одним из 85 протестированных рецепторов. Нецелевые эффекты часто являются причиной некоторых неблагоприятных эффектов новых химических структурных элементов. Таким образом, клеточная активность рецептора CB1 ингибируется избирательно и специфичным для активация сигнального пути способом.
2. Соединение по настоящему изобретению характеризуется очень высокую эффективностью in vivo при коррекции широкого спектра когнитивных нарушений, включая нарушения долгосрочной памяти, распознавания и исполнительных функций.
3. Соединение по настоящему изобретению демонстрирует эффективность in vivo в моделях различных когнитивных расстройств, включая синдром Дауна, синдром ломкой Х-хромосомы и возрастное снижение когнитивных способностей.
4. Соединение по настоящему изобретению не вызывает неблагоприятных эффектов, свойственных ортостерическим антагонистам CB1, включающих, но этим не ограничивая, a) снижение потребления пищи; b) рост случаев поведения, связанного с тревогой и депрессией; c) нейротоксичность, индуцирование клонических судорог и клинических признаков, связанных с поражением центральной нервной системы; d) гепатотоксичность и генотоксичность.
5. Соединение по настоящему изобретению не оказывает неспецифических воздействий на поведение даже в дозах, которые в тысячи раз превышают эффективную дозу для улучшения когнитивных функций. Эти эффекты включают, но этим не ограничиваются, изменения спонтанного поведения, седативный эффект, возбудимость.
6. Соединение по настоящему изобретению не проявляет неблагоприятных или токсических эффектов in vitro и in vivo в дозах, которые в тысячи раз превышают эффективную дозу для улучшения когнитивных функций. Следовательно, соединение по настоящему изобретению имеет высокий индекс безопасности (> 3500).
Согласно настоящему изобретению, "когнитивные расстройства" можно воспринимать как умственные расстройства, когнитивные или интеллектуальные нарушения, когнитивные или умственные нарушения, умственную отсталость или снижение когнитивных способностей, и они включают все заболевания/состояния, которые могут быть связаны с когнитивными расстройствами.
Когнитивная деятельность или когнитивная функция может быть определена как все формы умственной деятельности, посвященной обработке информации. Они включают в себя элементарные функции, такие как восприятие и двигательные навыки, а также процессы высокого порядка, такие как внимание, обучение и память, суждение или исполнительные функции. Нарушение когнитивной деятельности связано с нарушениями в любых когнитивных сферах, включающие:
- Комплексное внимание: устойчивое внимание, разделенное внимание, избирательное внимание и скорость обработки. Например, отсутствие концентрации комплексного внимания может приводить к трудностям в сохранении информации в уме в среде с отвлекающими факторами или к ошибкам при выполнении рутинных задач.
- Исполнительные функции: планирование, принятие решений, рабочая память, реакция на обратную связь/исправление ошибок, избавление от привычек/торможение и психическая гибкость. Например, недостаточность исполнительных функций может приводить к трудностям в выполнении многостадийных проектов, в планировании повседневной деятельности или в последующих социальных общениях.
- Обучение и память: непосредственная память и оперативная память, включая свободное узнавание или узнавание с подсказкой, долгосрочную и очень долговременную память и скрытное обучение. Например, последствия нарушения обучения и памяти могут проявляться в потере понимания при оплате счетов или повторении одних и тех же фактов в разговоре.
- Речь: выразительная речь и восприятие речи. Например, нарушение речи может быть связано с использованием общих, а не конкретных терминов, что приводит к грамматическим ошибкам. Тяжелые формы нарушения речи приводят к эхолалии или мутизму.
- Перцептивно-моторные способности: рисование, праксис (обучение запоминанию моторных последовательностей) и гнозис (например, распознавание лиц и цветов): перцепционно-моторные дефициты могут приводить к трудностям в исполнении ранее известных действий, таких как использование инструментов или вождение автомобиля.
- Общественное признание: способность распознавать эмоции, выражаемые другими людьми, и учитывать мысли и намерения, выражаемые другими людьми в необычных ситуациях. Нарушение общественного признания может приводить, например, к снижению эмпатии, к явно выраженному поведению или речи, выходящих за рамки общепринятого социального диапазона (например, неподходящая одежда), или к снижению способности поддержания разговоров.
В данном случае, когнитивное расстройство может относится к патологическому состоянию, при котором когнитивные нарушения превышают стандартный когнитивный уровень индивидуумов соответствующей возрастной группы. Кроме того, когнитивные расстройства могут рассматриваться как снижения когнитивных способностей, которые ассоциируются со старением здоровых людей (называемых возрастными снижениями когнитивных способностей).
Для иллюстрации основных когнитивных расстройств, авторы изобретения используют критерии Руководства по диагностике и статистическому учету психических расстройств (DSM-5™, которые относятся к нарушению неврологического развития и нейрокогнитивным расстройствам. Однако это руководство не является единственным описанием когнитивных расстройств, и оно включает аналогичные расстройства, описанные в других руководствах по диагностике, в том числе, но этим не ограничивая, в Классификации психических расстройств и расстройств поведения МКБ-10 и, в более общем смысле, все расстройства, в первую очередь связанные с нарушением когнитивных способностей.
В связи с этим, когнитивные расстройства включают в себя нарушения неврологического развития и нейрокогнитивные расстройства.
Нарушения неврологического развития относятся к расстройствам, которые проявляются на ранней стадии развития, часто до того, как ребенок начинает посещать начальную школу. Они характеризуются аномалиями развития, приводящими к нарушениям концептуального, практического и социального функционирования. Следовательно, индивидуум не в состоянии соответствовать возрастным стандартам интеллектуального развития, академической успеваемости, личной независимости и/или участия в общественной жизни. Диапазон нарушений развития варьируется от очень специфических пределов обучения или исполнительных функций до глобальных нарушений социальных навыков или умственных способностей.
Нарушения неврологического развития могут быть связаны с конкретной причиной (например, известным генетическим заболеванием или фактором окружающей среды) или могут быть точно не установленными, если их этиология неизвестна. Примеры конкретных причин включают генетические нарушения, такие как, но этим не ограничивая, синдром Дауна, синдром ломкой Х-хромосомы, туберозный склероз, синдром Ретта, синдром Уильяма, расщепление позвоночника. Медицинские состояния включают патологии, такие как эпилепсия, нарушения обмена веществ; аномалии развития (например, пороки развития головного мозга), материнские заболевания (например, плацентарное заболевание) или перинатальные факторы окружающей среды (например, воздействие алкоголя, токсинов или тератогенов на плод).
Нарушения неврологического развития включают семь категорий нарушений, которые определяют как указано ниже.
1. Умственная отсталость характеризуются недостаточностью общих умственных способностей, таких как рассуждение, решение проблем, планирование, абстрактное мышление, суждение, академическое обучение и обучение на собственном опыте. Общая задержка развития диагностируется у индивидуумов, которые не могут быть подвергнуты систематической оценке умственной деятельности, в том числе у детей, которые слишком малы для участия в стандартизированном тестировании.
2. Коммуникативные расстройства включают недостаточность владения языком (то есть использования общепринятой системы символов для общения), речи (то есть выражения мыслей артикулированными звуками), а также вербального и невербального общения. В эту категорию входят языковые расстройства, расстройства речевых звуков, расстройство беглости речи, возникшее в детстве (заикание), социальное (прагматическое) коммуникативное расстройство и коммуникативное расстройство с неустановленной причиной.
3. Расстройство аутистического спектра характеризуется постоянной недостаточностью социального общения и взаимодействия, ограниченными интересами и повторяющимся поведением. Расстройство аутистического спектра также называют аутистическим расстройством, синдромом Аспергера или первазивным расстройством развития.
4. Синдром дефицита внимания с гиперактивностью (ADHD) связан с выраженной невнимательностью и/или гиперактивностью и импульсивностью, которые не согласуются с ожидаемым уровнем развития и напрямую влияют на социальную и академическую/профессиональную деятельность. Эти симптомы не являются единственным проявлением оппозиционного поведения, неповиновения, враждебности или непонимания задач или инструкций.
5. Специфическое нарушение способности к обучению характеризуется определенной недостаточностью способности эффективно и точно воспринимать или обрабатывать информацию. Это связано с постоянными трудностями в обучении и использовании академических навыков в одной или более областях, таких как чтение (например, дислексия), письменное выражение (например, дисграфия) и математика (например, дискалькулия).
6. Двигательные расстройства, связанные с нервным развитием, характеризуются аномальными непроизвольными или неконтролируемыми движениями тела. Эти расстройства могут вызывать отсутствие запланированных движений или чрезмерное количество непроизвольных движений. Более конкретно, двигательные расстройства могут представлять собой расстройство координации, связанное с нервным развитием (недостаточность приобретения и реализации скоординированных двигательных навыков), расстройство стереотипных движений (повторяющееся, кажущееся управляемым и явно бесцельное моторное поведение) и тиковые расстройства (стереотипные моторные движения или внезапные, быстрые, повторяющееся и неритмичное вокализации). Продолжительность, предполагаемая этиология и клинические проявления определяют конкретное диагностированное тиковое расстройство (например, расстройство Туретта).
7. Другие нарушения неврологического развития относят к тем случаям, когда симптомы являются характерными для расстройства психического развития, но не соответствуют полным критериям для любого из нарушений в диагностическом классе нарушений неврологического развития.
Следует отметить, что нарушения неврологического развития часто проявляются совместно; например, люди с расстройством аутистического спектра часто имеют умственную отсталость, а многие дети с синдромом дефицита внимания с гиперактивностью (ADHD) также страдают специфическим нарушением способности к обучению.
Нарушения неврологического развития должны диагностироваться врачом-клиницистом на основании диагностических критериев DSM-5™ или любого другого средства диагностики психического здоровья. Тесты, используемые для оценки характера и степени нарушений, могут быть выбраны среди тестов, оценивающих умственную деятельность и/или адаптивное поведение. Соответствующие тесты следует выбирать в соответствии с возрастом пациента, а также с характером и тяжестью предполагаемых нарушений. Ниже приведены неполный перечень примеров шкал для оценки общих или специфических функций, которые проверены и одобрены, в том числе и для пациентов детского возраста.
Шкалы оценки общего когнитивного здоровья (Hessl et al., 2016; Esbensen et al., 2017):
- Батареи тестов CANTAB
- Когнитивная батарея NIH Toolbox для оценки умственной отсталости
Шкалы для оценки независимого функционирования и адаптивного поведения (Berry-Kravis et al., 2013; Esbensen et al., 2017):
- Система оценки адаптивной поведения - третье издание (ABAS-III) или более последнее
- Опросник по оценке отклоняющегося от нормы поведения (ABC)
- Шкалы адаптивного поведения Вайнленда (VABS)
- Шкалы оценок Коннорса
Шкалы оценки качества жизни пациента (Varni et al., 2001; Burckhardt and Anderson, 2003):
- Опросник по качеству жизни для педиатрических пациентов (PedsQL)
- Шкала оценки качества жизни (QOLS)
Тесты для оценки конкретных функций
- Исполнительные функции: Опросник для бальной оценки поведения на основе опросников по оценке исполнительных функций (BRIEF), Висконсинский тест сортировки карточек (WCST), Виртуальный радиальный лабиринт (методика оценки рабочей памяти) (Marighetto et al., 2012)
- Обучение и память: Виртуальный радиальный лабиринт (методика декларативной/реляционной памяти) (Sellami et al., 2017)
- Социальное познание: Шкала социальной ответственности
- Язык: Бостонский тест на именования (BNT), Токен тест
- Качество сна: Питтсбургский индекс качества сна (PSQI)
Нейрокогнитивные расстройства относятся к психическим расстройствам, характеризующимся первичной клинической недостаточностью когнитивной функции. Эта недостаточность должны быть приобретенный, а не обусловленной развитием, представляя собой снижение от ранее достигнутого когнитивного уровня. Когнитивная недостаточность, связанная с нейрокогнитивными расстройствами, должна влиять на повседневную деятельность человека и может приводить к потере способности человека управлять самим собой. Нейрокогнитивные расстройства могут влиять на одну или более когнитивных областей, среди которых комплексное внимание, исполнительные функции, обучение и память, язык, перцептивные моторные/зрительно-пространственные функции и социальное познание.
Нейрокогнитивные расстройства могут быть вызваны различными патологиями, включая, но этим не ограничивая, нейродегенеративные заболевания (например, болезнь Альцгеймера, лобно-височную лобарную дегенерацию, болезнь телец Леви, болезнь Паркинсона или болезнь Хантингтона), сосудистые заболевания, травматическое повреждение головного мозга, употребление психоактивных веществ/лекарственных веществ, вирус иммунодефицита человека (ВИЧ), прионную болезнь.
Нейрокогнитивные расстройства подразделяются на следующие категории.
1. Делирий: относится к нарушениям познавательной деятельности, которые развиваются в течение короткого периода времени (часов). Делирий является прямым следствием другого медицинского состояния, интоксикации или отмены психоактивных веществ (например, злоупотребления наркотиками, лекарственными препаратами, токсическими веществами). Делирий может длиться от нескольких часов до нескольких месяцев.
2. Основные и легкие нейрокогнитивные расстройства: относятся к снижению когнитивных способностей (в одной или более когнитивных областях), которое мешает быть самостоятельным в повседневной жизни. Основные нейрокогнитивные расстройства (также называемые деменцией) характеризуются значительной потерей человеком способности управлять самим собой. Как правило, людям требуется, как минимум, помощь для выполнения сложных задач, таких как прием лекарственных препаратов или оплата счетов. Легкие нейрокогнитивные расстройства (также называемые легкими когнитивными нарушениями) характеризуются более слабым снижением когнитивных способностей, чем основные нейрокогнитивные расстройства. Легкие нейрокогнитивные расстройства не препятствуют способности человека управлять самим собой, но выполнение сложных повседневных задач требует больших усилий или компенсационных стратегий.
Более того, когда у пациента проявляются симптомы когнитивной недостаточности, которые не могут быть обусловлены точными причинами, может быть диагностировано неуточненное легкое или основное нейрокогнитивное расстройство.
Диагностика нейрокогнитивных расстройств должна выполняться врачом с использованием субъективных и/или объективных оценок когнитивной функции. Субъективная когнитивная оценка заключается в получении информации от пациента и/или от его опекунов об изменениях когнитивных функций, которые проявляются в повседневной деятельности (например, распоряжение финансовыми средствами и прием лекарственных препаратов).
Объективная оценка должна проводиться с использованием одного или более стандартизированных тестов, направленных на оценку когнитивные функции. Доступно несколько шкал (Sheehan, 2012). Некоторые из них ориентированы на проведение опроса по повседневной деятельности, такого как Опрос функциональной активности, или по качеству жизни, такого Шкала оценки качества жизни (QOLS). Другие ориентированы на оценку конкретных когнитивных функций или глобальное когнитивное функционирование. Неисчерпывающими примерами тестов на глобальную когнитивную функцию являются Краткая шкала оценки психического статуса, Ускоренный тест для оценки психического статуса, Монреальская шкала оценки когнитивных функций, тесты "Мини-Ког" (Mini-Cog), серия тестов CANTAB, Повторяемая батарея тестов по оценке нейропсихологического состояния (RBANS) и Когнитивные батарея NIH Toolbox для оценки умственной отсталости.
Кроме того, методики виртуального радиального лабиринта, разработанные A. Marighetto и его коллегами, позволяют обнаруживать скрытую недостаточность рабочей памяти и декларативной/реляционной памяти, связанную с нормальным и патологическим старением (Etchamendy et al., 2012; Marighetto et al., 2012; Sellami et al., 2017). Эти методики радиального лабиринта имеют соответствующие эквиваленты для грызунов, что позволяет улучшить перенос данных исследований по потенциальным терапевтическим средствам для лечения когнитивных расстройств от экспериментальных животных на людей.
Следовательно, настоящее изобретение относится к соединению формулы (I):
Формула (I)
для применения для профилактики или лечения когнитивного расстройства, выбранного из нарушений неврологического развития, нейрокогнитивных расстройств и возрастного снижения когнитивных способностей. Введение субъекту эффективного количества соединения формулы (I) снижает, отменяет, стабилизирует (то есть предотвращает или ограничивает ухудшение) когнитивное расстройство или нарушение. В результате, у субъекта, в отношении которого применяли эффективную схему лечения, обнаруживается улучшение после объективного клинического обследования или по одному или нескольким параметрам нейропсихологических тестов или по шкалам качества жизни, соответствующим патологии, возрасту, языку и культуре субъекта. В качестве варианта, применение эффективной схемы лечения субъекта с помощью соединения формулы (I) позволяет сохранять когнитивную функцию, в силу чего это предотвращает или замедляет ухудшение после первого проявления или постановки диагноза когнитивного расстройства по сравнению с ожидаемым ухудшением при отсутствии лечения.
Термины "3β-бензилокси-17α—метил-прегн-5-ен-20-он" или "C29H40O2" или "3Bn17MeP" или "1-((3S,8R,9S,10R,13S,14S,17S)-3-(бензилокси)-10,13,17-триметил-2,3,4,7,8,9,10, 11,12,13,14,15,16,17-тетрадекагидро-1H-циклопента[a]фенантрен-17-ил)этан-1-он" обозначают производное прегненолона по настоящему изобретению, которое имеет следующую формулу (I):
Формула (I)
Способ получения
Настоящее изобретение также относится к способу получения соединения по изобретению. Соединение формулы (I) может быть получено различными способами синтеза, как это показано ниже.
Во-первых, соединение 3β-бензилокси-17α-метил-прегн-5-ен-20-он может быть получено в 2 стадии из прегненолона (метод A).
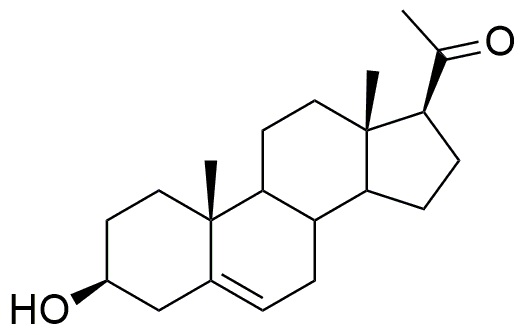
Прегненолон
- Введение метильной группы в положение C17 осуществляется в результате реакции между промежуточным соединением енолацетатом, образующимся по реакции прегнолона и уксусного ангидрида, и реактивом Гриньяра с получением соединения формулы (II).
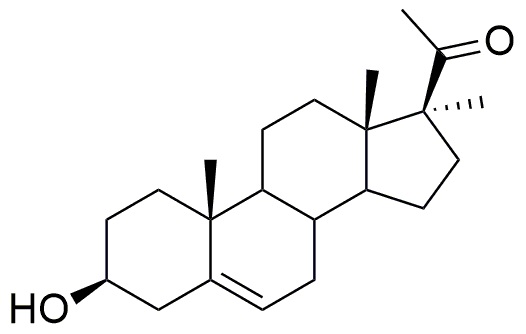
Формула (II)
Затем гидроксильную группу на C3 подвергают бензилированию с помощью реагента 2-бензилокси-1-метилпиридиния трифлата с получением 3β-бензилокси-17α—метил-прегн-5-ен-20-он формулы (I).
Формула (I)
Как видно на фигурах 1-4 и как описано ниже, могут быть использованы и другие методы получения 3β-бензилокси-17α—метил-прегн-5-ен-20-она.
Например, последнюю стадию алкилирования гидроксильной группы в положении C3 можно проводить в основных условиях (NaH, t-BuOK или другие основания) в присутствии бензилхлорида или бензилбромида (метод B). Также можно проводить алкилирование в кислой среде с использованием трихлорацетимидата в присутствии каталитического количества органической кислоты.
В другом методе (методе C) сначала может быть проведена защита функции прегненолон-кетона через образование ацеталя с получением соединения формулы (III).
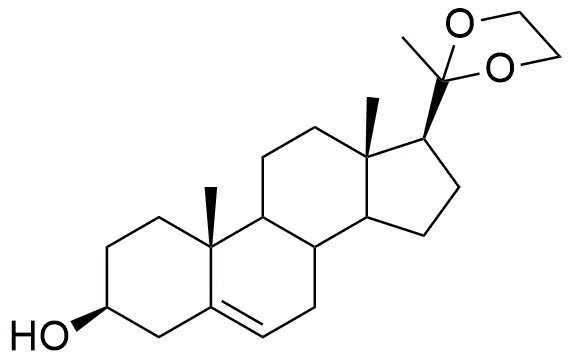
Формула (III)
Затем свободный спирт алкилируют с помощью бензильной группы (в кислых или основных условиях или через 2-бензилокси-1-метилпиридиния трифлат, как описано выше) с получением соединения формулы (IV).
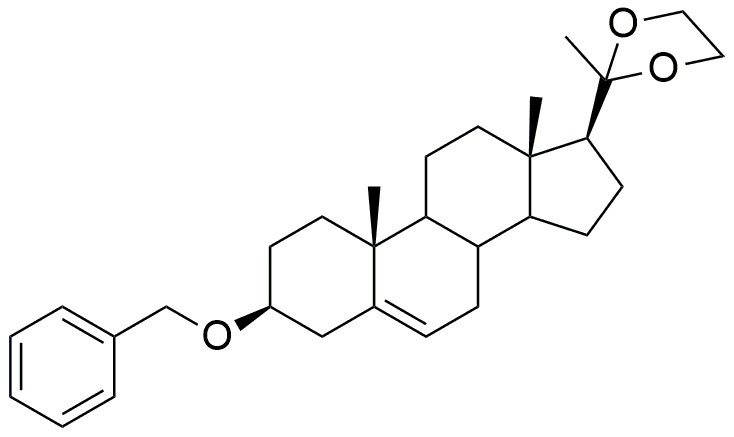
Формула (IV)
Ацеталь подвергают гидролизу в кислой среде с получением соединения формулы (V).
Формула (V)
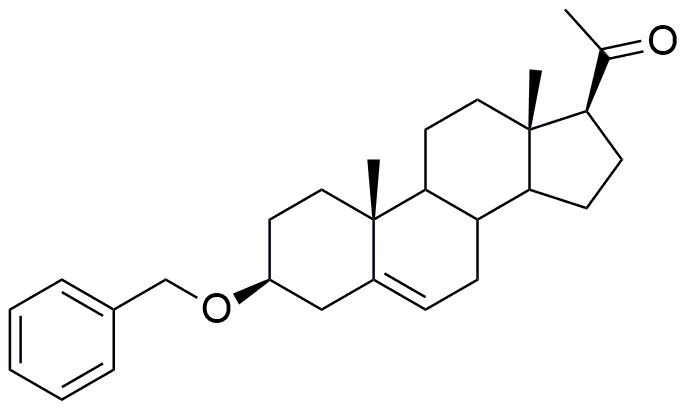
Прямое метилирование с использованием метилйодида в присутствии гидрида натрия дает 3β-бензилокси-17α—метил-прегн-5-ен-20-он формулы (I).
Формула (I)
Другим вариантом является метод C без осуществления защиты прегненолонкетоновой группы (метод D).
Настоящее изобретение также относится к фармацевтической композиции, содержащей соединение формулы (I):
Формула (I)
и, по меньшей мере, один фармацевтически приемлемый эксципиент.
Форма фармацевтической композиции, способ введения, дозировка и режим дозирования, естественно, зависят от подлежащего лечению состояния, тяжести заболевания, возраста, массы тела и пола пациента и других подобных параметров.
Несмотря на то, что соединение по настоящему изобретению может быть введено в чистом виде, предпочтительно приготавливать его в форме фармацевтической композиции в соответствии со стандартной фармацевтической практикой. Соответственно, в изобретение также предлагается фармацевтическая композиция, включающая соединение формулы (I) в смеси, по меньшей мере, с одним фармацевтически приемлемым эксципиентом.
Кроме того, в настоящем изобретении предлагается способ приготовления фармацевтической композиции, который включает смешивание соединения формулы (I) вместе, по меньшей мере, с одним фармацевтически приемлемым эксципиентом.
Фармацевтическая композиция обычно содержит, по меньшей мере, один фармацевтически приемлемый эксципиент. Приемлемые эксципиенты для терапевтического применения хорошо известны в фармацевтике и описаны, например, в руководстве Remington’s Pharmaceutical Sciences, 21st Edition 2011. Выбор фармацевтического эксципиента может быть осуществлен с учетом предполагаемого способа введения и стандартной фармацевтической практики. Эксципиент должен быть приемлемым в том смысле, что он не должен оказывать вредного воздействия на реципиента. По меньшей мере, один фармацевтически приемлемый эксципиент может представлять собой, например, связующее вещество, разбавитель, носитель, смазывающее вещество, разрыхлитель, смачивающее средство, диспергирующее средство, суспендирующее средство и другие подобные эксципиенты.
Способы введения (доставки) указанного выше соединения включают, но этим не ограничиваются, пероральный (например, в виде таблетки, капсулы или в виде раствора для приема внутрь), местный, мукозальный (например, в виде назального спрея или аэрозоля для ингаляции), назальный, гастроинтестинальный, интраспинальный, интраперитонаельный, внутримышечный, внутривенный, внутриматочный, внутриглазной, интрадермальный, интракраниальный, интратрахеальный, интравагинальный, интрацеребровентрикулярный, интрацеребральный, подкожный, офтальмологический (включая интравитреальный или внутрикамерный), трансдермальный, трансдермальный, ректальный, буккальный, эпидуральный, сублингвальный.
Предпочтительные способы введения включают пероральный, мукозальный, парентеральный и сублингвальный.
Например, соединение можно вводить перорально в форме таблеток, таблеток с покрытием, пилюль, капсул, мягких желатиновых капсул, пероральных порошков, гранул, вагинальных суппозиториев, эликсиров, растворов или суспензий, которые могут содержать ароматизаторы или окрашивающие вещества, для немедленного, отсроченного, модифицированного, пролонгированного, импульсного высвобождения или контролируемого высвобождения.
Таблетки могут содержать эксципиенты, такие как микрокристаллическая целлюлоза, лактоза, цитрат натрия, карбонат кальция, двухосновный фосфат кальция и глицин, разрыхлитель, такой как крахмал (предпочтительно, кукурузный, картофельный или тапиоковый крахмал), гликолят крахмала натрия, кроскармеллоза натрия и некоторые сложные силикаты, связующее вещество, такое как поливинилпирролидон, гидроксипропилметилцеллюлоза (HPMC), гидроксипропилцеллюлоза (HPC), сахароза, желатин и гуммиарабик, смазывающее вещество, такое как стеарат магния, стеариновая кислота, бегенат глицерина. Твердые композиции подобного типа могут также использоваться в качестве наполнителей в твердых желатиновых капсулах. Предпочтительные эксципиенты в этом отношении включают лактозу, сахарозу, сорбит, маннит, картофельный крахмал, кукурузный крахмал, амилопектин, производные целлюлозы или желатин. Твердые желатиновые капсулы могут содержать гранулы соединения по изобретению.
Могут быть приготовлены мягкие желатиновые капсулы, при этом капсулы включают соединение по изобретению, растительное масло, воск, жир или другой подходящий носитель для мягких желатиновых капсул. В качестве примера, приемлемым носителем может быть масляный носитель, такой как растительное масло с длинноцепочечным триглицеридом (например, кукурузное масло).
Диспергируемые порошки и гранулы, подходящие для приготовления водной суспензии путем добавления воды, могут содержать активный ингредиент в смеси с диспергирующими средствами, смачивающими средствами и суспендирующими средствами и одним или более консервантами, дополнительными эксципиентами, например, также может присутствовать подсластители, ароматизаторы и окрашивающие вещества. Эти композиции могут быть сохранены путем добавления антиоксиданта, такого как аскорбиновая кислота.
Жидкие лекарственные формы для перорального введения могут включать фармацевтически приемлемые растворы, эмульсии, суспензии, сиропы и эликсиры, включающие инертные разбавители, обычно используемые в данной области, такие как вода или масляный носитель. Жидкая лекарственная форма может быть представлена в виде сухого продукта для разведения водой или другой подходящей средой перед применением. Такие композиции могут также включать адъюванты, такие как смачивающие средства, эмульгаторы и суспендирующие средства, комплексообразующие вещества, такие как 2-гидроксипропил-бета-циклодекстрин, сульфобутиловый эфир-бета-циклодекстрин, а также подсластители, ароматизаторы, отдушки, окрашивающие вещества или красители с такими разбавителями как вода, этанол, пропиленгликоль и глицерин, а также их комбинации. Эти композиции могут быть сохранены добавлением антиоксиданта, такого как бутилированный гидроксианизол или альфа-токоферол.
Тонкоизмельченный порошок соединений по изобретению может быть приготовлен, например, путем микронизации или методами, известными в данной области. Соединения по настоящему изобретению могут быть измельчены с использованием известных методов измельчения, таких как мокрое измельчение, для получения размера частиц, подходящего для формования таблеток и для других типов композиций.
В случае парентерального введения соединение по настоящему изобретению, примеры такого введения включают один или более из следующих способов ведения: внутривенный, интраартериальный, интраперитонеальный, интратекальный, интравентрикулярный, интрауретральный, интрастернальный, внутригрудинный интракраниальный, внутримышечный или подкожный способ введения лекарственного средства; и/или путем использования инфузии.
Соединение по изобретению может быть введено парентерально в форме легкодоступной композиции или композиции типа депо.
Фармацевтические композиции для парентерального введения легкодоступного состава могут быть в форме стерильного водного или масляного раствора или суспензии для инъекций в нетоксичном парентерально приемлемом разбавителе или растворителе, и могут содержать вспомогательные вещества, такие как суспендирующие, стабилизирующие, диспергирующие, смачивающие вещества, и/или комплексообразующие агенты, такие как циклодекстрин, например, 2-гидроксипропил-бета-циклодекстрин, сульфобутиловый эфир-бета-циклодекстрин.
Композиция типа депо для парентерального введения может быть приготовлена обычными методами с фармацевтически приемлемым эксципиентом, включающим, но этим не ограничивая, биосовместимые и биоразлагаемые полимеры (например, поли-β-капролактон, полиэтиленоксид, полигликолевую кислоту, поли [(молочная кислота) -со (гликолевая кислота) ...)], поли (молочная кислота) ...), небиоразлагаемые полимеры (например, сополимер этилена и винилацетата, полиуретан, полиэфирамид, поливинилхлорид .. .), водные и неводные среды (например, воду, кунжутное масло, хлопковое масло, соевое масло, касторовое масло, миндальное масло, масляные эфиры, этиловый спирт или фракционированные растительные масла, пропиленгликоль, диметилсульфоксид (DMSO), тетрагидрофуран (THF), 2-пирролидон, N-метилпирролидинон, N-винилпирролидинон ...).
В качестве варианта, активный ингредиент может быть в сухой форме, такой как порошок, кристаллическое или лиофилизированное твердое вещество, для смешивания с подходящей средой. Приготовление подходящих парентеральных композиций в стерильных условиях легко осуществляется стандартными фармацевтическими методами, хорошо известными специалистам в данной области.
Как было указано, соединение по настоящему изобретению может быть введено интраназально или путем ингаляции, и его удобно вводить в форме сухого порошка с помощью порошкового ингалятора или распыляемого аэрозоля из контейнера под давлением, с помощью насоса, спрея или небулайзера с использованием подходящего пропеллента, например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана (например, фирмы Ineos Fluor), диоксида углерода или другого подходящего газа. В случае аэрозоля под давлением, стандартная доза может быть отмерена путем использования клапана для доставки отмеренного количества. Контейнер под давлением, насос, спрей или небулайзер могут содержать раствор или суспензию активного соединения. Капсулы и картриджи (сделанные, например, из желатина) для использования в ингаляторе или инсуффляторе могут быть приготовлены таким образом, чтобы они содержали порошковую смесь соединения и подходящую порошковую основу, такую как лактоза или крахмал. Для композиций, подходящих и/или адаптированных для ингаляционного введения, предпочтительно, чтобы соединение или соль формулы (I) находились в форме с уменьшенным размером частиц, и, более предпочтительно, чтобы форму с уменьшенным размером частиц получали, или ее можно было бы получать путем микронизации. Предпочтительный уменьшенный размер частиц (например, микронизированных частиц) соединения, соли или сольвата определяется значением D50 от приблизительно 0,5 до примерно 50 микрон (например, при измерении с помощью лазерной дифракции).
В качестве варианта, соединение по настоящему изобретению может быть введено в форме суппозитория или пессария, или оно может быть нанесено местно в форме геля, гидрогеля, лосьона, раствора, крема, мази или присыпки. Соединение по настоящему изобретению может быть также введено дермально или трансдермально, например, с использованием трансдермального пластыря. Соединения по изобретению могут быть также введены ингаляционно или ректально. Они могут быть также введены окулярно. Для офтальмологического применения, соединения могут быть приготовлены в виде микронизированных суспензий в изотоническом стерильном физиологическом растворе с корректированной величиной pH или, предпочтительно, в виде растворов в изотоническом стерильном физиологическом растворе с корректированной величиной pH, необязательно, в комбинации с консервантом, таким как хлорид бензилалкония. В качестве варианта они могут быть приготовлены в мази, такой как вазелин.
Для местного нанесения на кожу, средство по настоящему изобретению может быть приготовлено в виде подходящей мази, содержащей активное соединение, суспендированное или растворенное, например, в смеси с одним или более из следующих компонентов: минеральное масло, жидкий вазелин, белый вазелин, пропиленгликоль, сополимер полиоксиэтилена и полиоксипропилена, эмульгированный воск и вода. В качестве варианта, соединение может быть приготовлено в виде подходящего лосьона или крема, суспендированного или растворенного, например, в смеси одного или нескольких из следующих компонентов: минеральное масло, моностеарат сорбитана, полиэтиленгликоль, жидкий парафин, полисорбат 60, воск цетиловых эфиров, цетеариловый спирт, 2-октилдодеканол, бензиловый спирт и вода.
Как правило, фактическую дозу, которая будет наиболее подходящей для индивидуального субъекта, будет определять лечащий врач. Конкретный уровень дозы и частота дозирования для любого конкретного человека могут варьировать и будут зависеть от ряда факторов, включая активность конкретного применяемого соединения и продолжительность действия этого соединения, возраст, массу тела, общее состояние здоровья, пол, режим питания, способ и время введения, скорость выведения, комбинацию лекарств, тяжесть конкретного состояния и индивидуума, подвергаемого терапии.
Вышеуказанное соединение может быть введено субъекту для его применения при лечении когнитивных расстройств в любой дозе, подходящей для достижения терапевтического эффекта.
При пероральном и парентеральном введении человеку, суточная доза лекарственного средства может быть введена в форме разовой дозы или в форме разделенных доз.
Предлагаемый диапазон доз соединения по настоящему изобретению для введения человеку (с массой тела приблизительно 70 кг) составляет, в том числе, но не ограничивая, от 1 мкг до 1000 мг, более типично, от 1 мкг до 500 мг, более типично, от 1 мкг до 100 мг, более типично, от 1 мкг до 50 мг, более типично, от 1 мкг до 10 мг, более типично, от 1 мкг до 5 мг, более типично, от 1 мкг до 1 мг, более типично, от 1 мкг до 600 мкг, более типично, от 1 мкг до 200 мкг , более типично, от 1 мкг до 100 мкг, более типично, от 1 мкг до 60 мкг, более типично, от 10 мкг до 1000 мг, более типично, от 10 мкг до 500 мг, более типично, от 10 мкг до 100 мг, более типично, от 10 мкг до 50 мг, более типично, от 10 мкг до 10 мг, более типично, от 10 мкг до 5 мг, более типично, от 10 мкг до 1 мг, более типично, от 10 мкг до 600 мкг, более типично, от 10 мкг до 200 мкг, более типично, от 10 мкг до 100 мкг, более типично, от 20 мкг до 1000 мг, более типично, от 20 мкг до 600 мг, более типично, от 20 мкг до 200 мг, более типично, от 20 мкг до 60 мг, более типично, от 20 мкг до 20 мг, более типично, от 20 мкг до 6 мг, более типично, от 20 мкг до 2 мг, более типично, от 20 мкг до 600 мкг, более типично, от 20 мкг до 200 мкг активного ингредиента в разовой дозе, в расчете на массу свободной кислоты. Разовая доза может быть введена, например, от 1 до 4 раз в сутки. Доза будет зависеть от способа введения. Следует иметь в виду, что может потребоваться проведение обычной корректировки дозирования в зависимости от возраста и массы тела пациента, а также от тяжести состояния. Дозировка также будет зависеть от способа введения. Окончательно, точную доза и способ введения будет определять лечащий врач или ветеринар.
Термины "подходящая доза", "эффективное количество" соединения по изобретению относятся к эффективному количеству, достаточному для предотвращения, облегчения, устранения, контролирования, лечения или ингибирования когнитивного расстройства. Предполагается, что термин "контролирование" обозначает все процессы, которые могут представлять собой замедление, прерывание, купирование или остановку прогрессирования заболеваний и состояний, описанных в изобретении, но не обязательно указывает на полное устранение всех симптомов заболевания и состояния. Используемые для введения дозы могут быть адаптированы в зависимости от различных параметров, в частности, в зависимости от используемого способа введения, соответствующей патологии или, в качестве варианта, от желаемой продолжительности лечения. Естественно, что форма фармацевтической композиции, способ введения, дозировка и режим дозирования, как правило, зависят от подвергаемого лечению состояния, тяжести заболевания, возраста, массы тела и пола субъекта, и других подобных параметров. Предполагается, что предлагаемые ниже эффективные дозы не огранивают изобретение и представляют собой предпочтительные диапазоны доз. Однако предпочтительная доза может быть адаптирована применительно к индивидуальному субъекту, и для любого специалиста в данной области известно, как определить предпочтительную дозу без проведения излишних экспериментов.
Изобретение также относится к способу лечения когнитивных расстройств у субъекта, нуждающегося в этом, включающему введение эффективного количества формулы соединения (I):
Формула (I)
указанному пациенту.
Все раскрытые выше варианты осуществления включены в этот аспект.
В другом аспекте настоящее изобретение относится к применению соединения формулы (I):
Формула (I)
для лечения когнитивных расстройств.
Все раскрытые выше варианты осуществления включены в этот аспект.
В дополнительном варианте осуществления настоящее изобретение относится к применению соединения формулы (I):
Формула (I)
для производства фармацевтического препарата для лечения когнитивных расстройств.
Все раскрытые выше варианты осуществления включены в этот аспект.
ПРИМЕРЫ
Пример 1: Синтез 3Bn17MeP
3β-бензилокси-17α—метил-прегн-5-ен-20-он (3Bn17MeP) представляет собой химическое соединение, содержащее 7 хиральных центров 3S, 8S, 9S, 10R, 13S, 14S, 17S, изображенных в формуле (I):
Формула (I)
Стереохимическая конфигурация в этих центрах идентична стереохимической конфигурации исходного материала прегненолона.
Этот пример представляет собой способ получения 3β-бензилокси-17α-метил-прегн-5-ен-20-она (3Bn17MeP) методом А, приведенном на фигуре 1.
Синтез промежуточного соединения енолацетата
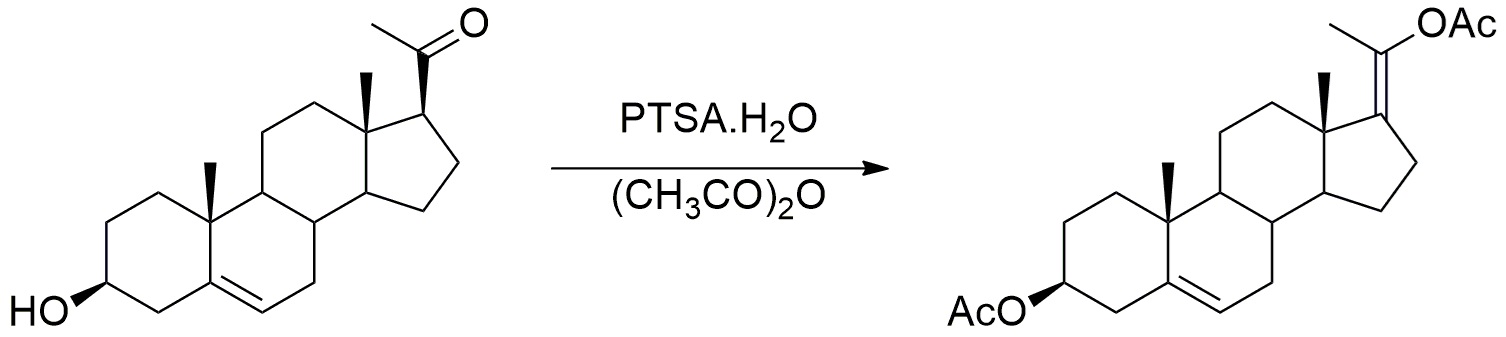
Моногидрат п-толуолсульфоновой кислоты (1,12 г, 5,9 ммоль, 0,93 экв) добавляли к раствору прегненолона (2 г, 6,3 ммоль, 1 экв) в уксусном ангидриде (230 мл). Реакционную среду перемешивали в течение 5 часов при кипячении с обратным холодильником, и медленно отгоняли уксусный ангидрид. После охлаждения до 20°C, реакционную среду выливали в измельченный лед, и затем смесь экстрагировали диэтиловым эфиром. Органический слой промывали насыщенным водным раствором Na2CO3, сушили над Na2SO4, затем испаряли при пониженном давлении. Остаток очищали хроматографией на силикагеле (элюент: циклогексан/AcOEt от 100/0 до 90/10) с получением енолацетата прегнолона (2,2 г; 85%) в виде белого твердого вещества.
Синтез 17α-метил-прегненолона
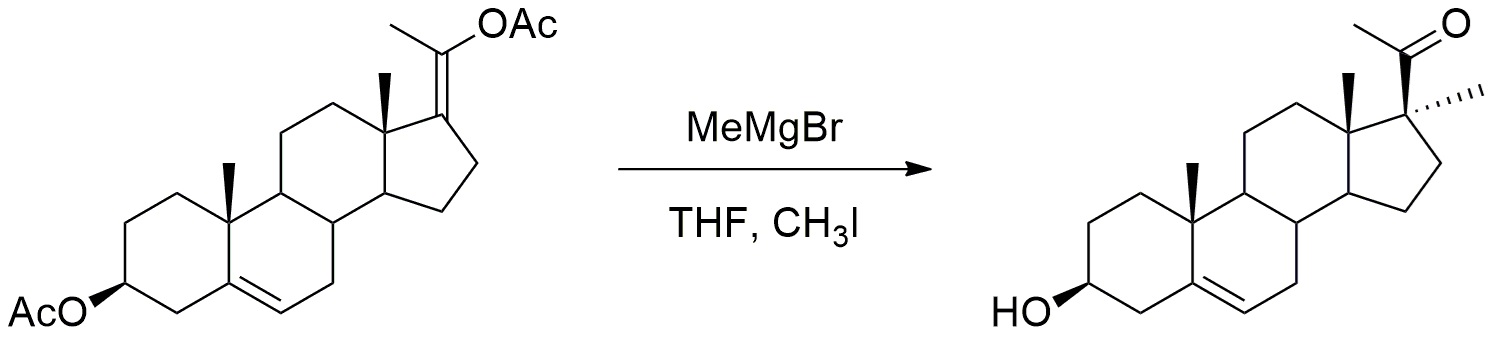
Как показано ниже, MeMgBr2 (3M в Et2O, 25 мл, 75 ммоль, 10 экв) добавляли к раствору енолацетата прегнолона (3 г, 7,5 ммоль, 1 экв) в безводном тетрагидрофуране (65 мл). Реакционную среду перемешивали в течение 1 часа при кипячении с обратным холодильником, затем давали остыть до 20°C. Добавляли CH3I (4,6 мл, 75 ммоль, 10 экв), и реакционную среду перемешивали при кипячении с обратным холодильником. Добавление CH3I повторяли каждые 45 минут до 40 эквивалентов. После охлаждения до 20°C, добавляли водный раствор NH4Cl, и смесь экстрагировали этилацетатом. Органический слой промывали солевым раствором, сушили над Na2SO4, затем испаряли при пониженном давлении. Остаток очищали хроматографией на силикагеле (элюент: циклогексан/AcOEt 75/25) с получением 17α-метил-прегнолона (~ 600 мг; 25%) в виде белого твердого вещества.
Синтез 3β-(бензилокси)-17α-метил-прегн-5-ен-20-она
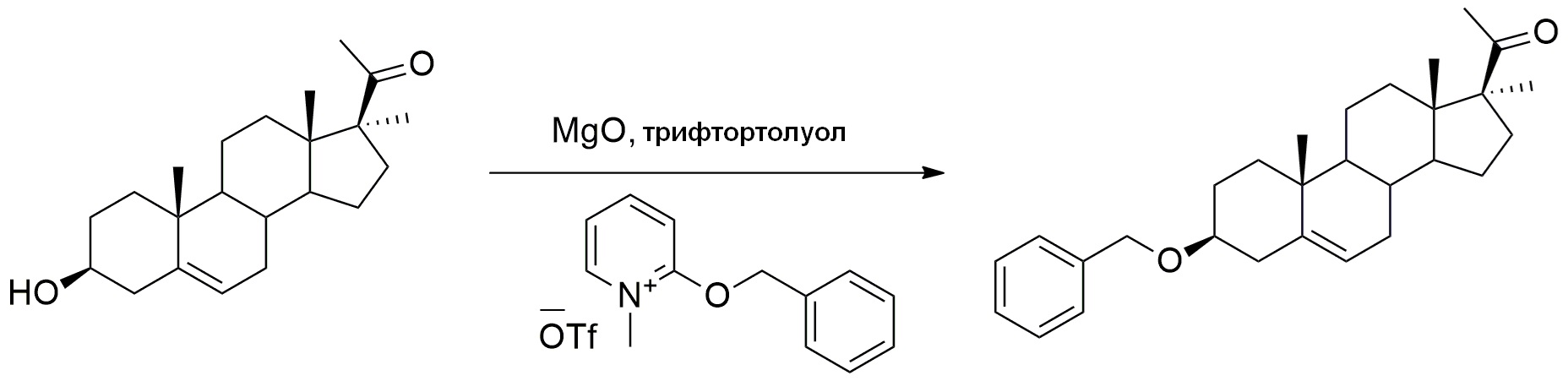
MgO (100 мг, 2,42 ммоль, 2 экв) и 2-бензилокси-1-метилпиридиния трифлат (850 мг, 2,42 ммоль, 2,0 экв) добавляли к раствору 17α-метил-прегненолона (400 мг, 1,21 ммоль, 1 экв) в трифтортолуоле (10 мл). Реакционную среду перемешивали в течение одной ночи при 85°C, затем фильтровали через целит и испаряли при пониженном давлении. Остаток очищали хроматографией на силикагеле (элюент: циклогексан/AcOEt 95/5), затем кристаллизовали в ацетоне с получением 3β-(бензилокси)-17α-метил-прегн-5-ен-20-она (0,28 г, 36%) в виде белого твердого вещества.
Пример 2. Специфическое ингибирование активности рецептора CB1 с помощью 3Bn17MeP
Было получено представление о воздействии соединения по настоящему изобретению, 3Bn17MeP, на клеточную активность в результате исследования его способности ингибировать несколько эффектов, вызванных стимуляцией CB1. В частности, были исследованы способность 3Bn17MeP (i) подавлять ингибирование клеточного дыхания, (ii) ингибировать увеличение степени фосфорилирования Erk1/2MAPK (p-Erk1/2MAPK, регулируемых внеклеточными сигналами киназ из семейства митоген-активируемых протеинкиназ) и (iii) предотвращать ингибирование индуцированной циклическим аденозинмонофосфатом (cAMP) стимуляции рецептора CB1 с помощью Δ9-тетрагидроканнабинола (THC). Были исследованы клеточное дыхание, p-Erk1/2MAPK и cAMP, поскольку эти клеточные ответы определяют специфическое ингибирование активации сигнального пути рецептора CB1 прегнолоном (Vallée et al., 2014). То есть прегнолон блокирует индуцированное THC ингибирование клеточного дыхания, ингибирует индуцированное THC увеличение p-Erk1/2MAPK, но не влияет на индуцированное THC ингибирование cAMP. В противоположность этому, ортостерические антагонисты CB1, такие как римонабант, ингибируют оба ответа.
Селективность воздействия 3Bn17MeP на рецептор CB1 изучали путем анализа связывания этого соединения с 85 рецепторами.
Материалы и методы
Влияние 3Bn17MeP на ингибирование клеточного дыхания, индуцированное THC.
Цель этих исследований заключалась в оценке влияние 3Bn17MeP на индуцированное THC (1 мкМ) ингибирование клеточного дыхания в клетках HEK-293, временно трансфицированных рецепторами CB1 человека (hCB1). Клетки HEK-293 были выбраны в связи с тем, что они не экспрессируют эндогенные рецепторы CB1, они могут быть легко трансфицированы и их ранее использовали в экспериментах по изучению активности рецептора CB1 in vitro (Shore et al., 2014) и в экспериментах, в которых было показано, что прегнолон способен ингибировать индуцированное THC снижение клеточного дыхания (Vallée et al., 2014).
В первом исследовании (фигура 5A), клетки HEK-293 временно трансфицировали плазмидой, экспрессирующей hCB1 дикого типа (HEK-293-hCB1-WT). Сначала клетки обрабатывали с помощью 3Bn17MeP (0, 1, 2,5, 5, 50 и 100 нМ, растворенного в 0,01% ацетонитриле). После 15 минут инкубации, добавляли в чашки для культивирования THC (0, 1 мкМ, растворенный в EtOH, 0,0034%) на 30 минут.
Во втором исследовании (фигура 5B), клетки HEK-293 временно трансфицировали либо плазмидой, экспрессирующей hCB1 дикого типа (HEK-293-hCB1-WT), либо мутантной плазмидой, экспрессирующей hCB1p.E1.49G (HEK-293-hCB1-Mut). В этом мутанте глутаминовая кислота в положении 1.49 была заменена глицином. Было показано, что эта мутация сохраняет воздействие THC, но подавляет воздействие прегненолона на клеточную активность, вызванную стимуляцией рецептора CB1 (Vallée et al., 2014). И действительно, глутамат в положении 1.49 играет важную роль для связывания прегненолона с рецептором CB1. Сначала клетки обрабатывали с помощью 3Bn17MeP (100 нМ растворяли в 0,01% ацетонитриле). Через 15 минут инкубации, в чашки для культивирования добавляли THC (0, 1 мкМ, растворенный в EtOH, 0,0034%) на 30 минут.
Клеточное дыхание измеряли на калиброванном респирометре Oxygraph, снабженном электродом Кларка. Для измерения клеточного дыхания использовали скорость потребления кислорода (ОС). Воздействие THC на скорость ОС в отсутствии и в присутствии 3Bn17MeP выражали в процентах от исходного уровня ОС для клетки, обработанной носителем 3Bn17MeP и носителем THC из того же эксперимента.
Воздействие 3Bn17MeP на увеличение p-Erk1/2MAPK, индуцированное THC
Цель этого исследования заключалась в оценке воздействия 3Bn17MeP на увеличение фосфорилирования p-Erk1/2MAPK, индуцированного путем введения THC в клетки линии STHdhQ7/Q7 (линия клеток, полученных из полосатого тела от нокаутной трансгенной мыши, содержащей гомозиготные локусы Хантингтина с гуманизированным экзоном 1 с 7 полиглутаминовыми повторами). Клетки STHdhQ7/Q7 были выбраны в связи с тем, что они стабильно экспрессируют высокие уровни эндогенных рецепторов CB1. Эти клетки ранее использовали в экспериментах по изучению активности рецепторов CB1 in vitro после стимуляции эндоканнабиноидами (Laprairie et al., 2014), и в условиях данного исследования они позволяют наиболее надежно анализировать CB1-индуцированное увеличение фосфорилирования MAPK.
Влияние 3Bn17MeP в 5 дозах (0,1, 0,3, 1, 3 и 9 мкМ, растворенных в DMSO 0,9%) испытывали при воздействиях 10 мкМ THC (растворенного в DMSO 0,05%) на фосфорилирование MAPK. Клетки предварительно обрабатывали с помощью 3Bn17MeP или носителя за 30 минут перед обработкой либо с помощью THC, либо с помощью носителя, в течение 30 минут).
Фосфорилирование MAPK (белков p-Erk1/2MAPK) измеряли с помощью наборов AlphaLISA SureFire Ultra с использованием нефосфорилированных белков Erk1/2MAPK в качестве контролей загрузки. Число импульсов p-Erk1/2MAPK нормализовали путем расчета процента увеличения числа импульсов p-Erk1/2MAPK, индуцированного THC, или носителем в отсутствие и в присутствии 3Bn17MeP.
Воздействие 3Bn17MeP на ингибирование cAMP, индуцированное THC
Цель этого исследования заключалась в оценке воздействия 3Bn17MeP на снижение cAMP, индуцированное введением THC, в клетках яичника китайского хомячка (СНО), стабильно экспрессирующих человеческий рецептор CB1 CHO-hCB1.
В этих экспериментах были выбраны клетки CHO-hCB1 в связи с тем, что они не экспрессируют рецептор CB1 эндогенно, и их ранее использовали в экспериментах по изучению воздействия агонистов CB1 (Rinaldi-Carmona et al., 1996), в том числе THC, на cAMP и P-MAPK и в экспериментах, в которых было показано, что прегненолон способен ингибировать THC-индуцированные эффекты на P-MAPK, но не на cAMP (Vallée et al., 2014).
Воздействие 3Bn17MeP (1 нМ, 10 нМ, 100 нМ и 1 мкМ, растворенного в DMF (N, N-диметилформамиде), 0,01%, фигура 5C) было испытано при сравнении с функциональной зависимостью "доза-ответная реакция" THC (0,3, 1, 3, 10, 30, 100 и 300 нМ, растворенный в этаноле, 0,0063%).
Клетки CHO-CB1-C2 подвергали обработке путем одновременного добавления THC и тестируемого соединения в течение 45 минут. Для поддержания базального уровня cAMP при всех условиях испытания также одновременно добавляли форсколин (2,5 мкМ).
После завершения обработки, клетки лизировали для проведения количественного определения cAMP. Все измерения в одном эксперименте выполняли в трех экземплярах. Количественное определение cAMP проводили методом конкурентного флуоресцентного иммуноанализа. Данные выражали в % дельта-флуоресценции (дельта F), которую рассчитывали следующим образом: дельта F% = (флуоресценция образца - флуоресценция отрицательного контроля) / флуоресценция отрицательного контроля.
Селективность связывания 3Bn17MeP in vitro.
Исследовали специфичность воздействия 3Bn17MeP на рецептор CB1 путем анализа воздействия этого соединения на связывание других 85 рецепторов. Эту серию анализов проводили с целью оценки специфичности связывания 3Bn17MeP, сравнивая ее с профилем прегненолона.
Были проведены испытания потенциальной способности 3Bn17MeP и прегнолона при концентрации 10 мкМ предотвращать связывание лигандов 85 рецепторов (CEREP с высокой пропускной способностью профилирования+4 стероидных рецептора+каннабиноидный рецептор типа 2). CEREP с высокой пропускной способностью профилирования состоит из обширной коллекции 80 трансмембранных и растворимых рецепторов, ионных каналов и рецепторов, связанных с G-белком. Он был специально разработан для предоставления информации для определения приоритетности наиболее перспективных соединений в процессе отбора на основе скрининга от соединений с требуемым типом активности до основного соединения (hit-to-lead). Для того чтобы лучше адаптировать анализ к 3Bn17MeP, в дополнение к спектру стероидных рецепторов, уже содержащихся в CEREP с высокой пропускной способностью профилирования, добавляли рецепторы андрогенов, эстрогенов, прогестерона и PXR, а затем рецепторы каннабиноидов типа 2.
Результаты
3Bn17MeP ингибирует снижение клеточного дыхания, индуцированное THC, в HEK-293-hCB1-WT (фигура 5A). При ID100 3Bn17MeP (50 нМ), воздействие THC полностью прекращается. В отличие от этого, в HEK-293-hCB1-Mut THC-индуцированное снижение клеточного дыхания невосприимчиво к действию 3Bn17MeP (100 нМ, фигура 5B), что указывает на то, что 3Bn17MeP ингибирует эффект THC в результате взаимодействия с рецепторами CB1.
В клетках STHdhQ7/Q7, стабильно экспрессирующих эндогенные рецепторы CB1, 3Bn17MeP дозозависимо снижает THC-индуцированное фосфорилирование MAPK (фигура 5C) при величине ED50 приблизительно 1 мкМ.
В отличие от этого, 3Bn17MeP не модифицирует ингибирование cAMP, индуцированное THC, при любой из доз, испытанных на клетках СНО, стабильно экспрессирующих рецепторы CB1 человека (фигура 5D).
3Bn17MeP (10 мкМ) не модифицирует связывание какого-либо из 85 рецепторов, которые были протестированы in vitro с использованием CEREP с высокой пропускной способностью профилирования, включая основные стероидные рецепторы, рецепторы PXR и рецепторы CB1 и CB2 (таблица 1). В этом отношении 3Bn17MeP является более селективным, чем прегненолон (10 мкМ), который предотвращает (> 80%) связывание рецепторов глюкокортикоидов, андрогенов и прогестерона и в меньшей степени (> 40%) связывание центральных и периферических рецепторов бензодиазепина.
Таблица 1. Сравнение связывания 3Bn17MeP и прегненолона с 85 рецепторами
|
Воздействия 3Bn17MeP in vitro суммированы в таблице ниже.
Таблица 2. Оценка in vitro механизма действия 3Bn17MeP
|
NA=не анализировали, потому что не было обнаружено никакого эффекта
Заключение
Следовательно, 3Bn17MeP in vitro действует как специфический ингибитор активация сигнального пути CB1 (CB1-SSi). Вследствие этого, 3Bn17MeP ингибирует снижение клеточного дыхания, вызванное THC в клетках HEK-293, трансфицированных рецепторами CB1 человека. 3Bn17MeP оказывает ингибирующее действие на стимуляцию рецептора CB1 посредством связывания сайта, который отличается от сайта связывания THC. 3Bn17MeP ингибирует усиление фосфорилирования MAPK, индуцированное THC в клетках STHdhQ7/Q7, эндогенно экспрессирующих рецепторы CB1, но не влияет на снижение уровней cAMP, индуцированное THC в клетках CHO, трансфицированных рецепторами CB1 человека.
Кроме того, 3Bn17MeP in vitro является более селективным, чем его эндогенный аналог прегненолон. И действительно, прегненолон (10 мкМ) предотвращает связывание (> 80%) рецепторов прогестерона, глюкокортикоидов и андрогенов, а также связывание бензодиазепиновых рецепторов (> 40%). 3Bn17MeP (10 мкМ) не изменяет связывание этих рецепторов или каких-либо других рецепторов (всего 85), которые исследовали с использованием CEREP с высокой пропускной способностью профилирования.
Пример 3. Эффективность 3Bn17MeP на доклинических моделях когнитивных расстройств.
Доклиническая оценка эффективности 3Bn17MeP при когнитивных расстройствах может проводиться на связанных с когнитивными расстройствами доклинических моделях, в том числе на доступных экспериментальных моделях нарушений неврологического развития (например, синдрома Дауна и синдрома ломкой Х-хромосомы), нейрокогнитивных расстройств (например, болезни Альцгеймера, болезни Паркинсона) и возрастного снижение когнитивных способностей на животных.
Экспериментальные модели на мышах синдрома Дауна, синдрома ломкой Х-хромосомы и возрастного снижение когнитивных способностей были выбраны потому, что:
- синдром Дауна является наиболее распространенным генетическим врожденным дефектом у человека и характеризуется выраженной умственной отсталостью (Grieco et al., 2015).
- синдром ломкой Х-хромосомы является основной моногенной причиной умственной отсталости и аутизма (Hunter et al., 2014).
- линии мышей Ts65Dn и fmr1-KO, экспериментальные модели на мышах синдрома Дауна и синдрома ломкой Х-хромосомы, соответственно, являются наиболее широко используемыми доклиническими моделями нарушения неврологического развития (Chang et al., 2013).
- старение является самым высоким фактором риска, связанный с нейрокогнитивными расстройствами и, в частности, с нейродегенеративными заболеваниями, такими как болезнь Альцгеймера (DSM-5tm). Следовательно, возрастные когнитивные нарушения часто являются предпосылкой патологического снижения когнитивных функций.
Исследовали положительное воздействие от введения 3Bn17MeP на три когнитивные функции, память распознавания, декларативную/реляционную память и рабочую память.
- Распознавание относится к типу извлечения памяти, происходящему, когда ранее испытанное событие совпадает с сохраненной памятью об этом событии.
- Декларативная/реляционная память: у человека декларативная память представляет собой тип долговременной памяти, относящейся к интенциональному вспоминанию информации, предыдущего опыта и концепций. Одним свойством декларативной памяти является способность устанавливать взаимосвязь между отличающимися в пространстве и/или во времени элементами совокупности всей познаваемой реальности (Sellami et al., 2017). В действительности, декларативная/реляционная память является гибкой, позволяющей вспоминать и манипулировать полученными раздельно двумя информационными составляющими, для того чтобы делать выводы или принимать решение.
- Рабочая память представляет собой тип исполнительной функции, позволяющей временно хранить и обрабатывать информацию. Это необходимо для выполнения повседневных действий, таких как поддержание беседы, рассуждение, понимание прочитанного.
Эти процессы являются кардинальными для нормального когнитивного функционирования, и их нарушения, вероятно, являются наиболее ограничивающими дееспособность когнитивными симптомами. Например, для овладения языком необходимо распознавать и запоминать слова, а также подвергать анализу их значение, для того чтобы понимать самому и быть понятым другими. Более того, нарушения этих процессов обычно выявляются при заболеваниях, вызывающих когнитивные расстройства, и воспроизводятся в экспериментальных доклинических моделях этих заболеваний на мышах, включая мышей лини Ts65Dn, мышей лини fmr1-KO и престарелых мышей.
Так, например, были проведены следующие исследования для оценки эффективности применения 3Bn17MeP при когнитивных расстройствах:
а) нарушение распознавания объектов, связанное с синдромом Дауна;
b) нарушение декларативной/реляционной памяти, связанное с синдромом Дауна;
c) нарушение рабочей памяти, связанное с синдромом Дауна;
d) нарушение распознавания объектов, связанное с синдромом ломкой Х-хромосомы;
е) нарушение долговременной реляционной/декларативной памяти, связанное со старением
В следующих доклинических исследованиях, 3Bn17MeP вводили перорально в двух разных формах, которые можно было безопасно применять на людях.
3Bn17MeP может быть солюбилизирован в кукурузном масле, растительном масле с длинноцепочечным триглицеридом. Эту липидную композицию вводили перорально через желудочный зонд в жидком виде.
3Bn17MeP можно растворять в водопроводной воде, содержащей 0,3% 2-гидроксипропил-β-циклодекстрина (HP-β-CD). Этот раствор вводили перорально мышам в виде ежедневного питья ad libitum в стеклянной бутылке. Водопроводную воду, содержащую 0,3% HP-β-CD, использовали в качестве носителя для контроля. В процессе введения 3Bn17MeP бутылки взвешивали 3 раза в неделю для контроля потребление 3Bn17MeP. Объем раствора, потребляемого ежедневно одной мышью, составлял приблизительно 4 мл. Например, концентрация 0,6 мкг/мл 3Bn17MeP в питьевой воде соответствует приблизительно дозе 80 мкг/кг для мыши с массой тела 30 г.
Доклинические модели, используемые для оценки эффективности 3Bn17MeP, описываются следующим образом:
- Доклиническая модель синдрома Дауна: мыши линии Ts65Dn характеризуются трипликацией хромосомы 16, которая содержит большинство генов, ортологичных хромосоме 21 человека (Gardiner, 2015). Мыши линии Ts65Dn хорошо воспроизводят когнитивные нарушения, наблюдаемые при синдроме Дауна, и являются наиболее изученной доклинической моделью синдрома Дауна (Gardiner, 2015). Экспериментальных мышей выводят путем скрещивания самок TS65Dn (B6EiC3Sn.BLiA-Ts(1716)65Dn/DnJ; Jackson Laboratory; Stock No: 5252) с самцами B6EiC3Sn.BLiAF1 (Jackson Laboratory; Stock No: 3647). Трансгенные мыши (Ts65Dn) и их однопометники дикого типа (WT) происходят из смешанного генетического фона DBA/2J x C57Bl/6J x C3H/HeJ.
- Доклиническая модель синдрома ломкой Х-хромосомы: мыши линии fmr1-KO характеризуются целенаправленной делецией экзона 5 последовательности гена fmr1 (Kazdoba et al., 2014). Эта делеция приводит к сайленсингу гена fmr1 и, соответственно, к подавлению кодируемого белка, белка умственной отсталости хрупкой хромосомы Х (FMRP). Основная роль FMRP заключается в регулировании локального синтеза белка в нейронах путем ограничения трансляции мРНК. Потеря FMRP приводит к нарушению созревания нейронов, что связано с сенсомоторными, поведенческими и когнитивными нарушениями (Kazdoba et al., 2014). В частности, у мышей линии fmr1-KO проявляется нарушение долговременной памяти, включая нарушение распознавания объектов (Busquets-Garcia et al., 2013). Экспериментальных мышей разводят в результате скрещивания мышей fmr1-KO (линии FVB.129P2-Pde6b+ Tyrc-ch Fmr1tm1Cgr/J; The Jackson Laboratory, Stock No: 4624) с мышами дикого типа (WT; FVB.129P2-Pde6b+ Tyrc-ch/AntJ; The Jackson Laboratory, Stock No: 4828). Трансгенные мыши и их однопометники дикого типа происходят из генетического фона FVB.
- Доклиническая модель возрастного снижения когнитивных способностей: известно, что у мышей C57Bl/6J в возрасте от 19 до 22 месяцев проявляются когнитивные расстройства при сравнении с молодыми взрослыми мышами C57Bl/6J в возрасте 3-4 месяцев, выведенными в тех же условиях (Sellami et al. др., 2017).
Пример 3a. Эффективность применения 3Bn17MeP при нарушении распознавания объектов, связанном с синдромом Дауна.
У мышей тест на распознавание объектов, который существует в нескольких вариантах, основан на спонтанном предпочтении новизны. Соответственно, мыши исследуют в течение более длительного времени новые объекты (Ennaceur, 2010) и демонстрируют "узнавание" объекта, с которым они уже сталкивались, исследуя его в течение более короткого времени. Память об одном конкретном объекте может быть оценена после короткой отсрочки по времени (например, от нескольких минут до одного часа) или после длительной отсрочки (например, через 24 часа).
Цель этих исследований заключается в оценке влияние введения 3Bn17MeP на нарушение распознавания объектов у мышей линии Ts65Dn в доклинической модели синдрома Дауна.
В первом исследовании (фигура 6A), проводили тест на краткосрочное распознавание объектов с использованием последовательной процедуры, в которой объекты предъявляли по одному. Узнавание ранее встреченного объекта, соответственно, проявлялась в сокращении времени исследования этого объекта. Эта процедура была выбрана в связи с тем, что она наиболее близка к задачам распознавания объектов, используемым при тестировании человека (например, к Кембриджской автоматизированной батареи нейробиологических тестов (Cantab Pattern Recognition Memory)), и при применении которой было показано, что у субъектов с синдромом Дауна обнаруживаются нарушения в распознавании объектов. В этой задаче, объекты/шаблоны представляли последовательно, и распознавание оценивали после отсрочки в несколько минут (Edgin et al., 2012).
Во втором исследовании (фигура 6B), проводили тестирование на долгосрочное распознавание объектов. Хотя нарушение долговременной памяти не является одним из основных нарушений, наблюдаемых у пациентов с синдромом Дауна, этот тест использовали для демонстрации воздействия ортостерических антагонистов CB1 на когнитивные функции. По этой причине, было важно проанализировать влияние 3Bn17MeP на решение этой конкретной задачи.
Материалы и методы
Влияние 3Bn17MeP на краткосрочное последовательное распознавание объектов в доклинической модели синдрома Дауна
Мышам предоставляли возможность исследовать 3 различных объекта (A, B, C), представленных один за другим и в следующих одной из двух возможных последовательностей: A-C-C-B-A и A-B-C-C-A. В открытом поле, мышей подвергали воздействию объекта А в течение 5 мин, а затем снова помещали в их домашнюю клетку. После отсрочки в течение 5 минут, мышей подвергали воздействию объекта B или C в течение 5 минут, а затем помещали обратно их в домашнюю клетку и так далее. Следовательно, задержка в результате их удержания в клетке между двумя представлениями объекта C составляла 5 минут, тогда как задержка между двумя представлениями объекта A составляла 35 минут. Время, потраченное на изучение объекта A или C во время первой презентации, сравнивали со временем исследования того же объекта во второй презентации. Этот параметр использовали для оценки характеристик распознавания объектов.
3Bn17MeP (0,6 мкг/мл в 0,3% HP-β-CD) или носитель (0,3% HP-β-CD) вводили перорально ad libitum в качестве ежедневного питья. Самцам и самкам мышей Ts65Dn и WT (в возрасте от 3 до 4 месяцев; n=7-11 на группу) предоставляли свободный доступ к носителю или раствору 3Bn17MeP. Перед процедурой распознавания объектов, мышей подвергали тестированию в радиальном лабиринте. Распознавание объектов проводили через 2 месяца введения 3Bn17MeP или носителя.
Влияние 3Bn17MeP на долгосрочное распознавание объектов в доклинической модели синдрома Дауна
Долговременную память у мышей можно оценить с помощью теста на распознавание объекта, в котором память об одном конкретном объекте оценивают через 24 часа после знакомства с ним. Мышам давали возможность исследовать 2 идентичных объекта в L-образном лабиринте. На следующий день проводили тест на распознавание объектов путем замены одного объекта на новый объект. В соответствии со спонтанным предпочтением новизны, мыши исследуют новые объекты в течение более длительного времени (Ennaceur, 2010). Отношение времени, потраченного на исследование нового объекта, к времени, потраченному на исследование знакомых объектов, используют в качестве индекса различения между знакомым объектом и новым объектом. Следовательно, этот параметр используют для оценки характеристик распознавания объектов.
Эффект повторного перорального введения 3Bn17MeP в дозе 15 мкг/кг в кукурузном масле или носителя (кукурузное масло; 2 мл/кг) изучали на самцах мышей Ts65Dn и WT в возрасте от 2 до 4 месяцев (n=7-8 на группу). 3Bn17MeP вводили дважды в сутки (в 9:00 и 17:00) в течение 7 дней подряд. Тест распознавания объектов проводился на 7-й день, через 3 часа после 13-го по счету введения 3Bn17MeP или носителя.
Результаты
В протоколе краткосрочного последовательного распознавания объектов (фигура 6A), мыши дикого типа были способны распознавать уже встречавшиеся им объекты, о чем свидетельствовало сокращение времени исследования между первым и вторым представлением одного и того же объекта, и это сокращение времени обнаруживалось как после 5-минутной задержки, так и после 35-минутной задержки, между презентациями объектов. В отличие от этого, мыши Ts65Dn, получавшие носитель, не демонстрировали значительного уменьшения времени исследования между последовательными представлениями объектов, независимо от времени задержки, что указывало на наличие нарушений краткосрочного распознавания объектов у мышей Ts65Dn. Мыши линии Ts65Dn, которым вводили 3Bn17MeP (0,6 мкг/мл), тратили значительно меньше времени на исследование знакомого объекта с задержками во времени через 5 и 35 минут, обнаруживая характеристики, аналогичные характеристикам мышей дикого типа.
В протоколе долговременного распознавания объектов (фигура 6B), мыши Ts65Dn, которым вводили носитель из кукурузного масла, демонстрирует выраженное нарушение распознавания объектов по сравнению с мышами WT (фигура 6B). Введение 3Bn17MeP (15 мкг/кг) увеличивало индекс различения у мышей Ts65Dn относительно мышей WT.
Таким образом, введение 3Bn17MeP полностью восстанавливало способность распознавания объектов у мышей в этой доклинической модели синдрома Дауна.
Пример 3b. Эффективность воздействия 3Bn17MeP на нарушение декларативной/реляционной памяти, связанное с синдромом Дауна.
Цель данного исследования заключается в оценке влияния введения 3Bn17MeP на нарушение декларативной/реляционной памяти у мышей линии Ts65Dn, в модели синдрома Дауна. Проводили описанное далее тестирование декларативной/реляционной памяти с использованием процедуры радиального лабиринта, которую уже успешно применяли для выявления нарушений как у пациентов с синдромом Дауна (в виртуальном радиальном лабиринте), так и у мышей линии Ts65Dn (в радиальном лабиринте) с использованием очень похожего протокола.
Материалы и методы
Выполнение задания на реляционную память проводили в радиальных лабиринтах с 8 рукавами. Диаметр центральной площадки составлял 30 см, длина рукавов составляла 55 см, ширина составляла 10 см, и высота от пола составляла 100 см. Каждый рукав был снабжен выдвижной дверцей на входе и системой подачи гранулированного корма на его конце. Каждый лабиринт помещали в комнату с визуальными ориентирами на стенах, для того чтобы обеспечить выделение цели по положению в пространстве. Внутри лабиринта ориентиры отсутствовали. Каждый лабиринт был полностью автоматизирован путем использования видео-телевизионного сопровождения целей (IMETRONIC, Pessac, France). Это позволяло автоматически управлять дверцами рукавов (открывание/закрывание) и доставкой корма в зависимости от местоположения мыши.
Мышей с ограниченным питанием (85% их исходной массы тела на протяжении всего исследования) сначала подвергали 3 сеансам ознакомления с лабиринтами. Во время каждого сеанса ознакомления, мышь свободно исследовала лабиринт до тех пор, пока она не посещала каждый из 8 рукавов, или пока не проходило 30 минут. В начале каждого сеанса ознакомления, мышь помещали на центральную платформу со всеми закрытыми руками. Через одну минуту, во все рукава помещали приманку в виде пищевого вознаграждения (шарик из сухих макаронных изделий, Panzani "Pâtes Perles", из 100% твердой пшеницы, приблизительно 10 мг каждый) и открывали одновременно вход во все рукава. Рукав считался посещенным, как только мышь достигала кормушки в конце рукава. Дверь посещенного рукава закрывали после того, как мышь возвращалась в центр лабиринта.
Мышей подвергали одному ежедневному сеансу ознакомления. В конце второго сеанса, если мышь не смогла посетить все рукава в течение отведенного времени или собрать пищевые вознаграждения из четырех последних посещенных рукавов, ее отправляли на дополнительный сеанс в тот же день, и еще на следующий день, если не достигалось это условие. Если после третьего дня ознакомления мышь все еще не могла исследовать все рукава или собрать пищевые вознаграждения из четырех последних посещенных рукавов, ее исключали из экспериментов.
Обучение начинали через 24 часа после последнего сеанса ознакомления. Стадия обучения заключалась в изучении положения гранул корма в лабиринте. Положение пищевого вознаграждения не менялось на протяжении всей процедуры. Использовали шесть рукавов из восьми (3 пары (A, B или C) смежных рукавов). Один рукав каждой пары содержал на конце пищевое вознаграждение, в другом рукаве каждой пары пищевое вознаграждение отсутствовало.
В начале каждого ежедневного сеанса, мышь помещали в центр лабиринта без доступа к рукавам. Затем двери первой пары рукавов открывали, обеспечивая доступ к обоим рукавам пары. После того, как мышь посещала один рукав и возвращалась на центральную платформу, дверь закрывали, и животное удерживали на центральной платформе. Две двери снова открывали, обеспечивая сразу же после этого доступ к паре рукавов (интервал между испытаниями 0 секунд). После того, как животное посещало один из рукавов и возвращалась на центральную платформу, двери снова закрыли перед следующим испытанием. В течение каждого сеанса, мышей подвергали 20 повторным и последовательным презентациям 3 пар рукавов, поочередно.
На каждом сеансе измеряли количество правильных ответов. Считалось, что мышь освоила тест, если она достигла критерия обучения. Этот критерий выражался, по меньшей мере, (i) 75% правильных ответов (для трех пар) за два последовательных сеанса и (ii) 62% правильных ответов для каждой из 3 пар за два последовательных сеанса.
Проведение теста на гибкость обучения начинали на следующий день после достижения критерия обучения. Но в действительности, мыши могли подвергаться тесту на гибкость обучения в разные дни.
В этой задаче на гибкость обучения, формировали новую пару рукавов путем рекомбинации 2 рукавов из двух разных пар, с которыми мыши ознакомились на этапе приобретения информации. Положение корма в лабиринте не изменяли, но изменяли способ представления рукавов. И действительно, вместо пар A и B, мышам представляли так называемую пару AB. Пара АВ состояла из комбинации двух соседних рукавов пар А и В (то есть один рукав с пищевым вознаграждением, а другой рукав без пищевого вознаграждения). Эта пара представляла собой контрольное испытание теста на гибкость обучения. Зависимая переменная, позволяющая измерять характеристику гибкости мышления (то есть декларативную/реляционную память), представляет собой процент правильных ответов при презентации пары AB.
3Bn17MeP (0,6 мкг/мл в 0,3% HP-β-CD) или носитель (0,3% HP-β-CD) вводили перорально ad libitum в качестве ежедневного питья. Самцам и самкам мышей Ts65Dn и WT (в возрасте от 3 до 4 месяцев; n=7 на группу) обеспечивали свободный доступ к носителю или раствору 3Bn17MeP (0,6 мкг/мл) за 7 дней до начала проведения процедуры радиального лабиринта и до конца исследования.
После окончания проведения исследования поведения мышей, их умерщвляли, собирали образцы крови и головного мозга. Количественно определяли содержание 3Bn17MeP в плазме и в головном мозге методом жидкостной хроматографии с тандемной масс-спектрометрией (LC/MS-MS).
Результаты
В тесте на гибкость обучения, мыши Ts65Dn, которым вводили носитель, характеризовались наличием нарушений по сравнению с мышами дикого типа. Точность выбора у мышей Ts65Dn, которым вводили носитель, не превышала уровня случайности (фигура 7). Введение 3Bn17MeP (0,6 мкг/мл) повышало точность выбора у мышей Ts65Dn, которая достигала уровня для мышей WT. Таким образом, в этой доклинической модели синдрома Дауна, применение 3Bn17MeP позволяет полностью восстанавливать гибкость обучения, причем эта функция лежит в основе скоррегированных способностей декларативной/реляционной памяти.
3Bn17MeP способен проникать как в головной мозг, так и в плазму мышей Ts65Dn. Средние концентрации 3Bn17MeP в плазме и в головном мозге составляли, соответственно, 1,33 нг/мл и 5,91 нг/г, что дает величину соотношения головной мозг/плазма, равную 4,4.
Пример 3c. Эффективность воздействия 3Bn17MeP при расстройстве рабочей памяти, связанным с синдромом Дауна.
Цель данного исследования заключается в оценке эффективности введения 3Bn17MeP при расстройстве рабочей памяти у мышей Ts65Dn в модели синдрома Дауна. Проводили описанное далее тестирование рабочей памяти с использованием процедуры радиального лабиринта, которую уже успешно применяли для выявления нарушений как у пациентов с синдромом Дауна (в виртуальном радиальном лабиринте), так и у мышей линии Ts65Dn (в радиальном лабиринте) с использованием очень похожего протокола.
Материалы и методы
Выполнение задания на рабочую память проводили в радиальных лабиринтах с 8 рукавами, описанных в примере 3а. Процедуру ограничения приема пищи и ознакомления с лабиринтами проводили так же, как описано в примере 3а. Проведение теста на рабочую память начинали через 24 часа после последнего сеанса ознакомления. Каждой мыши обеспечивали проведение 15 ежедневных сессий обучения (5 блоков по 3 дня), каждая из которых состояла из 23 испытаний. Во время проведения теста на рабочую память использовали шесть рукавов из восьми (3 пары (A, B или C) смежных рукавов). В начале каждого ежедневного сеанса, мышь помещали в центр лабиринта без доступа к рукавам. Затем двери первой пары рукавов открывали, обеспечивая доступ к обоим рукавам пары. После того, как мышь посещала один рукав и возвращалась на центральную платформу, дверь закрывали, и животное удерживали на центральной платформе в течение 10 секунд (интервал между испытаниями, ITI), затем две двери снова открывали, обеспечивая сразу же после этого доступ к паре рукавов. После того, как животное посещало один из рукавов и возвращалось на центральную платформу, двери снова закрыли на 10 секунд перед следующим испытанием. В течение каждого сеанса, мышей подвергали многократным и последовательным представлениям трех пар рукавов, поочередно. При первом представлении каждой пары рукавов, пищевое вознаграждение присутствовало в обеих рукавах. При последующих представлениях пар рукавов, пищевое вознаграждение присутствовало только в рукаве, противоположном рукаву, который был посещен ранее, независимо от правильности предыдущего выбора ответа. Зависимая переменная, позволяющая измерять характеристику рабочей памяти, представляет собой процент правильных ответов в каждом блоке из трех сеансов.
3Bn17MeP (0,06 мкг/мл в 0,3% HP-β-CD) или носитель (0,3% HP-β-CD) вводили перорально ad libitum в качестве ежедневного питья. Самцам и самкам мышей Ts65Dn и WT (в возрасте от 3 до 4 месяцев; n=24-26 на группу) обеспечивали свободный доступ к носителю или раствору 3Bn17MeP (0,06 мкг/мл) за 7 дней до начала проведения процедуры радиального лабиринта и до конца исследования.
Результаты
Процент правильных ответов был сильно снижен у мышей Ts65Dn, которым вводили носитель, по сравнению с мышами WT (фигура 8). Введение 3Bn17MeP (0,06 мкг/мл) увеличивало количество правильных ответов у мышей Ts65Dn по сравнению с мышами, которым вводили носитель, до уровня количества правильных ответов у мышей WT в конце процедуры тестирования рабочей памяти. Таким образом, 3Bn17MeP корректирует нарушение рабочей памяти в этой доклинической модели синдрома Дауна.
Пример 3d. Эффективность применения 3Bn17MeP при нарушении распознавания объектов, связанном с синдромом ломкой Х-хромосомы.
Цель данного исследования заключается в оценке воздействия введения 3Bn17MeP на нарушение распознавания объектов у мышей линии fmr1-KO в доклинической модели синдрома ломкой Х-хромосомы.
Материалы и методы
Процедуру распознавания объекта проводили, как описано в Примере 3а.
Воздействие повторных пероральных введений 3Bn17MeP в дозе 15 мкг/кг в кукурузном масле или носителя (кукурузного масла; 2 мл/кг) изучали на самцах мышей линий fmr1-KO и WT в возрасте от 2 до 4 месяцев (n=7-8 на группу). 3Bn17MeP вводили два раза в сутки (в 9:00 и 17:00) в течение 7 дней подряд. Тест на распознавание объектов проводили на 7-й день, через 3 часа после 13-го введения 3Bn17MeP или носителя.
Результаты
Мыши линии Fmr1-KO, которым вводили носитель из кукурузного масла, демонстрирует нарушение распознавания объектов по сравнению с мышами WT (фигура 9). Введение 3Bn17MeP (15 мкг/кг) увеличивало индекс различения мышей линии fmr1-KO относительно уровня мышей WT. Таким образом, введение 3Bn17MeP полностью восстанавливало способность распознавания объектов в этой доклинической модели Fragile X.
Пример 3e. Эффективность воздействия 3Bn17MeP на долговременное нарушение реляционной/декларативной памяти, связанное со старением.
Представленный далее когнитивный тест был успешно использован для обнаружения нарушений декларативной/реляционной памяти как у людей пожилого возраста (в виртуальном радиальном лабиринте), так и у пожилых мышей (в радиальном лабиринте) с использованием очень похожей процедуры (Sellami et al., 2017). И действительно, как пожилые люди, так и пожилые мыши, не справляются с выполнением заданий теста на гибкость обучения, который требует вспоминания и манипулирования двумя информационными составляющими, полученными раздельно в двух различных временных последовательностях.
Материалы и методы
Выполнение задания на реляционную память проводили в радиальных лабиринтах с 8 рукавами, описанных в примере 3b. Процедуру ограничения приема пищи и ознакомления с лабиринтами проводили так же, как описано в примере 3b. Проведение теста на реляционную память начинали через 24 часа после последнего сеанса ознакомления. Стадия ознакомления заключалась в изучении положения гранулированного корма в лабиринте. Положение пищевого вознаграждения не менялось на протяжении всей процедуры. Один рукав каждой пары содержал в своем конце пищевое вознаграждение, в другом рукаве каждой пары пищевое вознаграждение отсутствовало.
Обучение основано на процедуре "да - нет". Рукав представляли один за другим. Ожидается, что субъекты будут посещать рукав с пищевым вознаграждением быстрее, чем рукав без пищевого вознаграждения. В начале каждого ежедневного сеанса, мышь помещали в центр лабиринта без доступа к рукавам. Затем открывали дверь первого рукава. После того, как мышь посещала один рукав и возвращалась на центральную платформу (или после задержки в 90 секунд, если она не посещала рукав), дверь закрывали, и животное удерживали на центральной платформе. Затем дверь рукава тут же открывают (интервал между испытаниями 0 секунд). После того, как животное посетило рукав и вернулось на центральную платформу (или после задержки в 90 секунд, если оно не посетило рукав), дверь снова закрывали перед следующей попыткой. Каждое ежедневный сеанс состоял из 40 последовательных презентаций рукавов (то есть испытаний), организованных в две отдельные временные последовательности. В первой последовательности (20 испытаний), представляли поочередно четыре рука лабиринта (два с пищевым вознаграждением и два без пищевого вознаграждения). Затем проводили вторую последовательность (20 испытаний); четыре других рукава лабиринта представляли альтернативно (два с пищевым вознаграждением и два без пищевого вознаграждения). Рукава представляют в их соответствующей последовательности на протяжении всей фазы обучения.
В каждом испытании измеряли время запаздывания посещения рукава. Считалось, что мышь освоила тест, когда она достигала критерия обучения. Этот критерий характеризовался (i) по меньшей мере, 50% уменьшением времени для посещения рукавов с пищевым вознаграждением по сравнению с рукавами без пищевого вознаграждения в среднем за два последовательных сеанса и (ii) по меньшей мере, 30% уменьшением времени для посещения для посещения рукавов с пищевым вознаграждением в среднем за последний сеанс.
Проведение теста на гибкость обучения начинали на следующий день после достижения критерия обучения. Но в действительности, мыши могли подвергаться тесту на гибкость обучения в разные дни.
Гибкость обучения оценивали в следующих условиях. Виртуально разделяли восемь рукавов лабиринта на пары, каждая из которых состояла из двух смежных рукавов, один с пищевым вознаграждением, а другой без пищевого вознаграждения. Пары состояли из одного рукава, информацию о котором мышь усвоила в первой последовательности, и другого рукава, информацию о котором мышь усвоила во второй последовательности. Положение корма в лабиринте не изменяли, но изменяли способ представления рукавов. В начале проведения сеанса на гибкости обучения, мышь помещали в центр лабиринта без доступа к рукавам. Затем открывали двери первой пары рукавов, обеспечивая доступ к обоим рукавам пары. После того, как мышь посещала один рукав и возвращалась на центральную платформу, дверь закрывали, и животное удерживали на центральной платформе. Две двери снова открывали, обеспечивая сразу же после этого доступ к паре рукавов (интервал между испытаниями 0 секунд). После того, как животное посещало один из рукавов и возвращалась на центральную платформу, двери снова закрыли перед следующим испытанием. Мышей подвергалась 20 повторным и последовательным представлениям пар рукавов поочередно.
Для каждой пары измеряли процент правильных ответов. Точность выбора пар, состоящих из рукавов, которые были представлены в разных последовательностях, отражала характеристику реляционной памяти.
3Bn17MeP (0,6 мкг/мл в 0,3% HP-β-CD) или носитель (0,3% HP-β-CD) вводили перорально ad libitum в качестве ежедневного питья. Самцам мышей C57Bl/6J в возрасте от 19 до 22 месяцев (n=9-12 на группу) предоставляли свободный доступ к носителю или раствору 3Bn17MeP (0,6 мкг/мл) за 7 дней до начала проведения процедуры радиального лабиринта и до конца исследования.
После окончания проведения исследования поведения мышей, их умерщвляли, собирали образцы крови и головного мозга. Количественно определяли содержание 3Bn17MeP в плазме и в головном мозге методом жидкостной хроматографии с тандемной масс-спектрометрией (LC/MS-MS).
Результаты
В тесте на гибкость обучения, точность выбора у пожилых мышей, которым вводили носитель, не превышала уровня случайности (50%, фигура 10). Введение 3Bn17MeP (0,6 мкг/мл) значительно повышало точность выбора у пожилых мышей, достигая более 70% правильных ответов. Таким образом, применение 3Bn17MeP позволяет восстанавливать способность реляционной памяти у пожилых мышей.
3Bn17MeP способен проникать как в головной мозг, так и в плазму пожилых мышей. Средние концентрации 3Bn17MeP в плазме и мозге составляли, соответственно, 1,28 нг/мл и 6,75 нг/г, что дает величину соотношения головной мозг/плазма, равную 5,3.
Заключение
Соединение по настоящему изобретению демонстрирует in vivo очень высокую эффективность при коррекции широкого спектра когнитивных нарушений, включая нарушения долговременной декларативной/реляционной памяти, распознавания и исполнительных функций. 3Bn17MeP демонстрирует эффективность в нескольких моделях когнитивных расстройств, включая синдром Дауна, синдром ломкой Х-хромосомы и возрастное снижение когнитивных способностей. Следовательно, 3Bn17MeP обладает широким спектром воздействия на когнитивную функцию и, в силу этого, вероятно, является усилителем глобальных когнитивных функций.
Пример 4. 3Bn17MeP не оказывает побочных поведенческих эффектов, свойственных ортостерическим антагонистам CB1.
Проводили оценку фармакологического профиля и воздействий 3Bn17MeP на фенотипы, связанные с неблагоприятными эффектами ортостерического антагониста CB1 римонабанта. Ортостерические антагонисты CB1, такие как Acomplia®, были изъяты из обращения и применения вследствие их неблагоприятных эффектов. Следовательно, для того чтобы терапевтическое средство, ингибирующее CB1, применялось на практике при лечении людей, оно не должно иметь известных неблагоприятных эффектов, свойственных ортостерическим антагонистам CB1.
Известными неблагоприятными эффектами ортостерических антагонистов рецепторов CB1 и, в частности, римонабанта являются: 1) снижение потребления пищи и массы тела, что является признаком неспецифического воздействия на сигнальные пути механизмов вознаграждения; 2) индуцирование поведения, связанного с тревогой и депрессией.
А связи с этим, проводили сравнение воздействие 3Bn17MeP на снижение потребления пищи, массы тела и на поведение, связанное с тревогой и депрессией, у мышей с воздействием ортостерического антагониста CB1 римонабанта.
Известно, что римонабант вызывает изменения спонтанного поведения у грызунов, такие как царапание, гиперактивность с последующим седативным действием и судороги, при дозах, которые в 6 раз превышают терапевтическую дозу для лечения ожирения (Zavatti et al., 2011; EPAR discussion, EMEA 2006). В действительности, воздействие 3Bn17MeP на спонтанное поведение изучали на мышах.
Пример 4а. Отсутствие вызываемых 3Bn17MeP нежелательных эффектов на снижение потребления пищи и массы тела у мышей дикого типа при сравнении с римонабантом.
Целью этих экспериментов была оценка способности однократного введения 3Bn17MeP снижать потребление пищи у худых мышей ad libitum, получавших стандартную лабораторную пищу, и сравнение этих потенциальных эффектов с эффектами ортостерического антагониста CB1 римонабанта. Исследовали снижение потребления пищи и массы тела, поскольку снижение потребления пищи и массы тела является прототипным следствием воздействия ортостерических антагонистов CB1 как на мышей, так и на людей (Carai et al., 2006).
Материалы и методы
Воздействие однократного введения 3Bn17MeP на снижение потребления пищи у худых мышей, получавших стандартный корм ad libitum.
Спонтанное потребление стандартного корма мышами оценивали во время темной фазы цикла свет-темнота. Количество съеденной пищи измеряли при выключенном свете через 3 и 13 часов после выключения света.
Воздействие однократного введения 3Bn17MeP (0,05, 5, 15 и 30 мг/кг, перорально в кукурузном масле, 5 мл/кг) или носителя (кукурузного масла) на снижение потребления пищи сравнивали с воздействие римонабанта (10 мг/кг, интраперитонеально в 0,9% NaCl, содержащем 2% ДМСО и 2% Твин 80, 10 мл/кг) или носителя (2% ДМСО и 2% Твин 80 в 0,9% NaCl) самцам мышей C57BL/6J (n=7-8 на группа). 3Bn17MeP и римонабант вводили за 3 часа и за 30 минут до начала темной фазы, соответственно.
Воздействие повторяющихся введений 3Bn17MeP на массу тела худых мышей, получавших стандартный корм ad libitum
Воздействие повторяющихся введений самцам мышей C57BL/6J (n=7-8 на группу) 3Bn17MeP (0,05, 5, 15 и 30 мг/кг, перорально в кукурузном масле, 5 мл/кг) или носителя (кукурузного масла) на массу тела сравнивали с воздействием римонабанта (10 мг/кг), интраперитонеально в 0,9% NaCl, содержащем 2% ДМСО и 2% Tween80, 10 мл/кг) или носителя (2% DMSO и 2% Tween80 в 0,9% NaCl). 3Bn17MeP и римонабант вводили 1 раз в сутки в течение 40 дней. Мышей взвешивали перед введением лекарственного средства каждый день в течение первой недели лечения, а затем в течение четырех дней в неделю.
Результаты
3Bn17MeP (0,05, 5; 15 и 30 мг/кг) не влиял на снижение потребления пищи по сравнению с носителем (фигура 11A), в то время как римонабант (10 мг/кг) значительно снижал потребления пищи в течение первых 3 часов после выключения света (фигура 11B). В тех же дозах, 3Bn17MeP не модифицировал процесс изменения массы тела в течение 40 дней повторного введения по сравнению с носителем (фигура 11C). В противоположность этому, римонабант (10 мг/кг) снижал массу тела мышей за этот период (фигура 11D).
Пример 4b. Отсутствие нежелательных воздействий, индуцируемых 3Bn17MeP, на поведение, подобное тревоге и депрессии, у мышей дикого типа при сравнении с римонабантом.
Проводили исследование поведения, связанного с тревогой и депрессией, поскольку усиление состояния тревоги и депрессии является следствием повторного лечения ортостерическими антагонистами CB1 как у грызунов, так и у людей (Bellocchio et al., 2013; Moreira et al., 2009). Поведение, подобное тревожному состоянию, изучали в приподнятом крестообразном лабиринте (EPM), поскольку эта модель широко используется на грызунах для оценки предполагаемых анксиогенных или анксиолитических эффектов фармакологических соединений (Walf and Frye, 2007). Поведение, связанное с депрессией, изучали с помощью теста предпочтения сахарозы, который широко используется в качестве модели ангедонии, одного из основных симптомов депрессии (Overstreet, 2012).
Цель этих экспериментов заключалась в оценке воздействия повторного лечения с помощью 3Bn17MeP на поведение, связанное с депрессией и тревогой, у мышей. Эти воздействия сравнивали с воздействием антагониста ортостерических рецепторов CB1 римонабанта, который, как известно, вызывает поведение, подобное тревоге и депрессии, как у мышей, так и у людей.
Материалы и методы
Исследование тревожного поведения в приподнятом крестообразном лабиринте
Приподнятый крестообразный лабиринт был изготовлен из поливинилхлорида серого цвета и состоял из четырех приподнятых рукавов. Рукава (высотой 60 см, длиной 37 см, шириной 6 см каждый) были расположены крестообразно, при этом два противоположных рукава были окружены стенками, а два других рукава были открыты. Видеокамера, размещенная на вершине лабиринта и подключенная к системе слежения, позволяла автоматически оценивать передвижения мышей в лабиринте. Интенсивность освещения лабиринта устанавливали на уровне 45 люкс в открытых рукавах. Одну мышь помещали в центре приподнятого крестообразного лабиринта и давали ей возможность свободно исследовать весь лабиринт в течение 5 минут. Измеряли затраченное время на посещение и количество посещений открытых и закрытых рукавов. Снижение процента посещений и/или времени, проведенного в открытых рукавах, отражает повышение уровня тревожности.
Изучали воздействие разового перорального введения 3Bn17MeP, растворенного в кукурузном масле, в дозе 30 мг/кг, или носителя (кукурузного масла; 5 мл/кг) на двухмесячных самцов мышей C57Bl/6J (n=10) на группу) в тесте в приподнятом крестообразном лабиринте. Тест в приподнятом крестообразном лабиринте проводили через 3 часа после введения 3Bn17MeP. Римонабант растворяли в носителе, содержащем DMSO (2%), Tween80 (2%) в растворе NaCl 0,9% для инъекций. В рамках этого же исследования, исследовали воздействие интраперитонеальной инъекции римонабанта в дозе 10 мг/кг или только одного носителя (10 мл/кг) на двухмесячных самцов мышей C57Bl/6J (n=10 на группу) в тесте в приподнятом крестообразном лабиринте. Тест в приподнятом крестообразном лабиринте проводили через 30 минут после инъекции римонабанта.
Поведение, подобное депрессии, в тесте на предпочтение сахарозы
Тест на предпочтение сахарозы проводили в домашних клетках мышей. Раствор сахарозы, содержащий 2% сахарозы (D (+) сахарозы) в питьевой воде, разливали в пластиковые градуированные бутыли (объем каждой бутылки 45 мл). Идентичные бутыли заполняли питьевой водой. Всех мышей сначала приучали к бутылкам и к раствору сахарозы. Помещали одну бутылку с водой и одну бутылку с сахарозой в питательный бункер каждой домашней клетки. Для гарантии того, что животные не будут проявлять предпочтения в отношении стороны размещения питья, половину бутылок, содержащих каждый раствор, помещали с левой стороны питательного бункера, а другую половину бутылок размещали с правой стороны питательного бункера. Мыши имели доступ к растворам с начала темной фазы в период суточного пика потребления пищи и воды (с 19:00 до 8:30). Через семь дней проводили тест на предпочтение сахарозы. Растворы воды и сахарозы выставляли таким же образом, как это описано для периода обучения, за исключением того, что мыши имели доступ к растворам с 12 до 22 часов. В каждый момент времени бутылки взвешивали, при этом 1 г соответствовал 1 мл. Объем потребления рассчитывали путем вычитания конечной массы бутылки из исходной массы бутылки.
Изучали воздействия повторных пероральных введений 3Bn17MeP, растворенного в кукурузном масле, в дозах 0,05; 5; 15 или 30 мг/кг, или носителя (кукурузного масла, 5 мл/кг) на двухмесячных самцах мышей C57Bl/6J (n=7-8 на группу) в тесте на предпочтение сахарозы. Римонабант растворяли в носителе, содержащем диметилсульфоксид (DMSO, 2%), Tween80 (2%) в растворе NaCl 0,9% для инъекций. В рамках этого же исследования, изучали воздействие повторных интраперитонеальных инъекций римонабанта в дозе 10 мг/кг или только одного носителя (10 мл/кг) на двухмесячных самцов мышей C57Bl/6J (n=8 на группу) в тесте на предпочтения сахарозы. 3Bn17MeP и римонабант вводили 1 раз в сутки в течение 28 дней. Тест на предпочтения сахарозы проводили на 28-й день, растворы сахарозы и водные растворы выставляли через 3 часа после введения 3Bn17MeP и через 30 минут после введения римонабанта.
Результаты
3Bn17MeP, вводимый в разовой дозе 30 мг/кг, не влиял на процент затраченного времени и на процент посещений открытых рукавов приподнятого крестообразного лабиринта (фигура 12A). В противоположность этому, римонабант в дозе 10 мг/кг заметно сокращал затраченное время и количество посещений открытых рукавов (фигура 12B).
Длительное введение 3Bn17MeP в дозах до 30 мг/кг не влияло на потребление сахарозы ни при одной из испытанных доз (0,05; 5; 15; 30 мг/кг), тогда как римонабант (10 мг/кг) снижал потребление сахарозы наполовину при тех же самых условиях (фигуры 12C и D).
Эти результаты показывают, что повторные введения 3Bn17MeP не вызывают депрессивного или тревожного поведения у мышей. В отличие от этого, ортостерический антагонист рецептора CB1 римонабант (10 мг/кг) вызывает ангедонию в тесте на предпочтения сахарозы и усиливает тревожное поведение в тесте в приподнятом крестообразном лабиринте. Эти результаты согласуются с ранее проведенными исследованиями, в которых сообщалось об аналогичных эффектах римонабанта у грызунов при дозах всего 1 мг/кг (Beyer et al., 2010; Patel and Hillard, 2006).
Пример 4c. 3Bn17MeP не влияет на спонтанное поведение у мышей.
3Bn17MeP не оказывал заметного влияния на поведение как таковое у грызунов, как показал анализ видеоданных спонтанного поведения грызунов в домашней клетке в течение 24 часов после перорального введения 3Bn17MeP мышам в дозе 30 мг/кг.
Заключение
В таблице 3 приведено сравнение неблагоприятных эффектов при применении римонабанта с эффектами при применении 3Bn17MeP.
Таблица 3. 3Bn17MeP не проявляет неблагоприятных эффектов, свойственных римонабанту.
|
Эффекты 3Bn17MeP сильно отличаются от эффектов ортостерического антагониста CB1 римонабанта.
3Bn17MeP не имеет типичных неблагоприятных эффектов, свойственных римонабанту и другим ортостерическим антагонистам CB1, таких как снижение потребления пищи, рост поведения, связанного с тревогой и депрессией. Это отсутствие эффектов у 3Bn17MeP наблюдалось для всех самых высоких доз, используемых в каждом тесте. Более того, 3Bn17MeP не вызывает модификации спонтанного поведения у мышей. Эти дозы 3Bn17MeP были в несколько раз выше, чем доза, применяемая для индуцирования регресса когнитивных нарушений, во всех протестированных моделях когнитивного расстройства.
Пример 5. 3Bn17MeP не токсичен in vitro и демонстрирует благоприятный профиль безопасности in vivo.
Пример 5а. Токсичность 3Bn17MeP in vitro
Для изучения потенциальной токсичности 3Bn17MeP in vitro использовали четыре модели: 1) нейротоксичность в первичной культуре кортикальных нейронов крыс; 2) гепатотоксичность и функция желчевыводящих путей в сэндвич культуре первичных гепатоцитов крысы; 3) генотоксичность, измеряемая фосфорилированием гистона H2AX (γH2AX) в клетках HeLa. В этих анализах, эффект 3Bn17MeP сравнивался с эффектом римонабанта; 4) кардиотоксичность, измеряемая ингибированием тока калиевых каналов (hERG) в клетках hERG-CHO. 3Bn17MeP тестировали до 100 мкМ.
Материал и методы
Нейротоксичность
Цель этого исследования заключалась в анализе цитотоксических эффектов 3Bn17MeP и римонабанта в первичной культуре корковых нейронов. Культивированные первичные корковые нейронов из эмбрионов крыс E19 обрабатывали 3Bn17MeP, римонабантом (в обоих случаях при концентрациях 0, 10, 30 или 100 мкМ в 0,1% NMP) или стауроспорином (100 нМ, используемого в качестве референсного соединения) и растворимым флуоресцентным маркером цитолиза. Затем за клетками наблюдали в режиме замедленной съемки в течение 72 часов. Затем клетки пермеабилизировали. Эта процедура позволяла выражать цитолиз в зависимости от времени в процентах от общего количества клеток на лунку.
Генотоксичность
Цель этого исследования заключалась в измерении фосфорилирования гистона H2AX (γH2AX), которое является клеточным ответом на повреждение ДНК, приводящее к разрывам двухцепочечной ДНК. Этот эксперимент проводили на клетках HeLa, обработанных 3Bn17MeP (0,1, 0,3, 1, 3, 10, 30 и 100 мкМ в NMP, 0,1%) или римонабантом (0,1, 0,3, 1 или 100 мкМ в NMP, 0,1%), в течение 24 часов. В качестве положительного контроля генотоксических эффектов добавляли этопозид в концентрации 3 мкМ. Проводили иммунофлуоресцентный анализ обработанных клеток специфическим антителом против фосфорилированного гистона gH2AX. Ядра окрашивали флуоресцентным средством для интеркалирования в ДНК. Окрашенные клетки визуализировали и анализировали на устройстве для визуализации BD Pathway 855 Imager®.
Гепатотоксичность и функция желчевыводящих путей.
Цель этого исследования заключалась в анализе цитотоксических эффектов 3Bn17MeP в первичной культуре гепатоцитов крыс. Процент подвергнутых цитолизу гепатоцитов в зависимости от времени определяли в режиме замедленной съемки изображений, полученных с помощью использования флуоресцентного маркера цитолиза. Количество желчных протоков после 48 часов лечения определяли с использованием флуоресцентного аналога соли желчной кислоты.
Выделяли первичные гепатоциты крыс линии Wistar в возрасте 10-12 недель с использованием двухэтапного метода перфузии коллагеназы. Через 24 часа клетки покрывали слоем матригеля для культивирования сэндвич-культуры. Через 24 часа клетки обрабатывали 3Bn17MeP или римонабантом (0, 0,1, 0,3, 1, 3, 10, 30 и 100 мкМ в NMP, 0,1%) и флуоресцентным маркером цитолиза, а затем постоянно наблюдали в режиме замедленной съемки изображений, полученных с помощью флуоресцентного и фазового контрастных веществ, в течение 48 часов. Затем клетки окрашивали с использованием флуоресцентного аналога соли желчных кислот для измерения состояния сети желчных канальцев и активности помпы, экспортирующей желчные кислоты. В качестве положительного контроля гепатотоксичности и потери желчных канальцев добавляли, соответственно, ацетаминофен (30 мМ) и циклоспорин A (1 мкМ).
Кардиотоксичность
Цель этого исследования заключалась в измерении воздействия 3Bn17MeP на ток калиевых каналов (hERG), который имеет решающее значение при реполяризации потенциалов действия кардиомиоцитов. Этот эксперимент проводили на клетках hERG-CHO-K1, обработанных 3Bn17MeP (0,1, 1 и 10 мкМ в DMSO, 1%) в течение 5 мин. В качестве положительного контроля кардиотоксических эффектов добавляли E-4031 в концентрации 0,3 мкМ. Ток hERG регистрировали автоматизированным методом локальной фиксации потенциала на целой клетке. Степень ингибирования (%) получали путем измерения разницы между амплитудами следового тока, индуцированного протоколом стимуляции (+20 мВ, 2 секунды и -40 мВ, 1 секунда), применяемой до и после инкубации лекарственного средства.
Результаты
3Bn17MeP не показал токсического эффекта ни при одной из испытанных доз и ни в одном из проведенных тестов на нейротоксичность (фигура 13A), генотоксичность (фигура 13B), гепатотоксичность (фигура 14A), функции желчевыводящих путей (фигура 14B) и кардиотоксичность.
В отличие от этого, римонабант проявлял значительную нейротоксичность при концентрации от 10 мкМ и индуцировал 80% гибели нервных клеток при концентрации от 30 мкМ (фигура 13A). Римонабант проявлял генотоксичность, поскольку он вызывал повреждение ДНК при концентрации 100 мкМ (фигура 13B). Римонабант проявлял гепатотоксический эффект, поскольку при концентрации от 10 мкМ он вызывал 100% гибель гепатоцитов и подавлял функцию желчных каналов (фигура 14A-B). Для 3Bn17MeP, максимальная испытанная доза (100 мкМ) в 2000 раз превышает ID100 3Bn17MeP для ингибирования THC-индуцированного снижения клеточного дыхания (50 нМ) в клетках HEK-293, трансфицированных рецепторами CB1 человека (фигура 5A).
Пример 5b. Безопасность 3Bn17MeP in vivo
Цель этого исследования (CERB, Франция) заключалась в оценке in vivo эффекта 3Bn17MeP у крыс Sprague-Dawley после перорального дозирования 3Bn17MeP один раз в сутки в течение 7 дней.
Материал и методы.
Крысам (5 самцам, 5 самкам) вводили перорально фиксированную дозу 3Bn17MeP (20 мг/кг в 10 мл/кг кукурузного масла) один раз в сутки в течение 7 дней.
Регистрацию случаев заболеваемости/смертности проводили два раза в сутки. Клинические наблюдения проводили перед первым дозированием, а затем ежедневно. Функциональные и нейроповеденческие тесты проводили перед первым введением дозы и в последний день лечения. Массу тела регистрировали в день-1, день-3, день-5 и день-7. Потребление пищи измеряли еженедельно.
Образцы крови для определения гематологических параметров и для клинического биохимического анализа собирали в день проведения некропсии (день-8).
Образцы крови для анализа концентрации лекарственного средства в плазме собирали после последней обработки, через 5 и 24 часа после введения дозы. Количественное определение проводили методом жидкостной хроматографии с тандемной масс-спектрометрией (LC/MS-MS).
Животных умерщвляли на день-8. Отобранные органы взвешивали, фиксировали, консервировали при некропсии и проводили гистопатологические исследования.
Результаты.
Единственные незначительные изменения были замечены в результатах анализа гематологических параметров и биохимического анализа крови. В случае гематологических параметров, наблюдалась тенденция к снижению количества лейкоцитов у мужчин и женщин и небольшая тенденция к снижению количества эритроцитов у мужчин. В случае биохимического анализа крови, наблюдалась тенденция к снижению общего уровня белка и повышению уровня триглицеридов. У женщин наблюдалась тенденция к снижению уровня триглицеридов. Но эти изменения не были связаны с какими-либо гистопатологическими изменениями.
На 7-й день, через 5 часов после последней обработки, средняя концентрация 3Bn17MeP в плазме составила 4652,4 нг/мл. Через 24 часа после лечения, средняя концентрация в плазме снижалась до 973,8 нг/мл. Эти результаты показали, что происходит всасывание 3Bn17MeP, и что через 24 часа после последнего введения он полностью не выводится из организма.
Для сравнения, средняя концентрация 3Bn17MeP в плазме, измеренная у мышей после одного месяца введения 3Bn17MeP в питьевой воде, при концентрации, позволяющей достигать регрессии при нарушениях долговременной памяти, связанных со старением (пример 7) и с синдромом Дауна (пример 4), составляла 1,3 нг/мл. Соотношение между этой концентрацией и концентрацией 3Bn17MeP в плазме, измеренное через 5 часов после последнего введения 3Bn17MeP в этом исследовании токсичности (4652,4 нг/мл), показало наличие высокой величины окна безопасности, которая составляла > 3500.
Заключение.
При применяемых условиях проведения экспериментов, 3Bn17MeP, вводимый перорально в течение 7 дней подряд, не вызывал никаких признаков токсичности при дозе 20 мг/кг/сутки у крыс линии Sprague-Dawley. Следовательно, после проведения повторного лечения разовой дозой 3Bn17MeP, можно считать, что максимальная переносимая доза (MTD) в случае 3Bn17MeP составляет величину выше 20 мг/кг/сутки.
Соотношение между концентрацией 3Bn17MeP в плазме, измеренной при проведении теста на безопасность и эксперимента по оценке эффективности, показывает наличие высокой величины для окна безопасности, которая составляла > 3500.
СПИСОК ЦИТИРУЕМЫХ ИСТОЧНИКОВ ИНФОРМАЦИИ
Arnone, M., Maruani, J., Chaperon, F., Thiébot, M.H., Poncelet, M., Soubrié, P., and Le Fur, G. (1997). Selective inhibition of sucrose and ethanol intake by SR 141716, an antagonist of central cannabinoid (CB1) receptors. Psychopharmacology (Berl.) 132, 104-106.
Bellocchio, L., Soria-Gómez, E., Quarta, C., Metna-Laurent, M., Cardinal, P., Binder, E., Cannich, A., Delamarre, A., Häring, M., Martín-Fontecha, M., et al. (2013). Activation of the sympathetic nervous system mediates hypophagic and anxiety-like effects of CB₁ receptor blockade. Proc. Natl. Acad. Sci. U.S.A. 110, 4786-4791.
Beringer, P. (2011). Remington: the science and practice of pharmacy. (Philadelphia; London: Lippincott Williams & Wilkins).
Berry-Kravis, E., Hessl, D., Abbeduto, L., Reiss, A.L., Beckel-Mitchener, A., and Urv, T.K. (2013). Outcome Measures for Clinical Trials in Fragile X Syndrome: Journal of Developmental & Behavioral Pediatrics 34, 508-522.
Beyer, C.E., Dwyer, J.M., Piesla, M.J., Platt, B.J., Shen, R., Rahman, Z., Chan, K., Manners, M.T., Samad, T.A., Kennedy, J.D., et al. (2010). Depression-like phenotype following chronic CB1 receptor antagonism. Neurobiol. Dis. 39, 148-155.
Breton, M.-C., Turgeon, M., Tremblay, É., Gosselin, C., Institut national d’excellence en santé et en services sociaux (Québec), and Bibliothèque numérique canadienne (Firme) (2015). Traitement pharmacologique de la maladie d’Alzheimer et des maladies apparentées: rapport d’évaluation des technologies de la santé.
Burckhardt, C.S., and Anderson, K.L. (2003). The Quality of Life Scale (QOLS): reliability, validity, and utilization. Health Qual Life Outcomes 1, 60.
Busquets-Garcia, A., Gomis-González, M., Guegan, T., Agustín-Pavón, C., Pastor, A., Mato, S., Pérez-Samartín, A., Matute, C., de la Torre, R., Dierssen, M., et al. (2013). Targeting the endocannabinoid system in the treatment of fragile X syndrome. Nature Medicine 19, 603-607.
Carai, M.A.M., Colombo, G., Maccioni, P., and Gessa, G.L. (2006). Efficacy of rimonabant and other cannabinoid CB1 receptor antagonists in reducing food intake and body weight: preclinical and clinical data. CNS Drug Rev 12, 91-99.
Chang, K.T., Ro, H., Wang, W., and Min, K.-T. (2013). Meeting at the crossroads: common mechanisms in Fragile X and Down syndrome. Trends Neurosci. 36, 685-694.
Edgin, J.O., Mason, G.M., Spanò, G., Fernández, A., and Nadel, L. (2012). Human and mouse model cognitive phenotypes in Down syndrome: implications for assessment. Prog. Brain Res. 197, 123-151.
Ennaceur, A. (2010). One-trial object recognition in rats and mice: methodological and theoretical issues. Behav. Brain Res. 215, 244-254.
Esbensen, A.J., Hooper, S.R., Fidler, D., Hartley, S.L., Edgin, J., d’Ardhuy, X.L., Capone, G., Conners, F.A., Mervis, C.B., Abbeduto, L., et al. (2017). Outcome Measures for Clinical Trials in Down Syndrome. American Journal on Intellectual and Developmental Disabilities 122, 247-281.
Etchamendy, N., Konishi, K., Pike, G.B., Marighetto, A., and Bohbot, V.D. (2012). Evidence for a virtual human analog of a rodent relational memory task: A study of aging and fMRI in young adults. Hippocampus 22, 869-880.
Gardiner, K.J. (2015). Pharmacological approaches to improving cognitive function in Down syndrome: current status and considerations. Drug Des Devel Ther 9, 103-125.
Grieco, J., Pulsifer, M., Seligsohn, K., Skotko, B., and Schwartz, A. (2015). Down syndrome: Cognitive and behavioral functioning across the lifespan. American Journal of Medical Genetics Part C: Seminars in Medical Genetics 169, 135-149.
Hanney, M., Prasher, V., Williams, N., Jones, E.L., Aarsland, D., Corbett, A., Lawrence, D., Yu, L.-M., Tyrer, S., Francis, P.T., et al. (2012). Memantine for dementia in adults older than 40 years with Down’s syndrome (MEADOWS): a randomised, double-blind, placebo-controlled trial. The Lancet 379, 528-536.
Hessl, D., Sansone, S.M., Berry-Kravis, E., Riley, K., Widaman, K.F., Abbeduto, L., Schneider, A., Coleman, J., Oaklander, D., Rhodes, K.C., et al. (2016). The NIH Toolbox Cognitive Battery for intellectual disabilities: three preliminary studies and future directions. Journal of Neurodevelopmental Disorders 8.
Hunter, J., Rivero-Arias, O., Angelov, A., Kim, E., Fotheringham, I., and Leal, J. (2014). Epidemiology of fragile X syndrome: a systematic review and meta-analysis. Am. J. Med. Genet. A 164A, 1648-1658.
Kazdoba, T.M., Leach, P.T., Silverman, J.L., and Crawley, J.N. (2014). Modeling fragile X syndrome in the Fmr1 knockout mouse. Intractable Rare Dis Res 3, 118-133.
Kishnani, P.S., Heller, J.H., Spiridigliozzi, G.A., Lott, I., Escobar, L., Richardson, S., Zhang, R., and McRae, T. (2010). Donepezil for treatment of cognitive dysfunction in children with Down syndrome aged 10-17. Am. J. Med. Genet. A 152A, 3028-3035.
Kumin, L., and Schoenbrodt, L. (2016). Employment in Adults with Down Syndrome in the United States: Results from a National Survey. Journal of Applied Research in Intellectual Disabilities 29, 330-345.
Laprairie, R.B., Bagher, A.M., Kelly, M.E.M., Dupré, D.J., and Denovan-Wright, E.M. (2014). Type 1 cannabinoid receptor ligands display functional selectivity in a cell culture model of striatal medium spiny projection neurons. J. Biol. Chem. 289, 24845-24862.
Marighetto, A., Brayda-Bruno, L., and Etchamendy, N. (2012). Studying the impact of aging on memory systems: contribution of two behavioral models in the mouse. Curr Top Behav Neurosci 10, 67-89.
Moreira, F.A., Grieb, M., and Lutz, B. (2009). Central side-effects of therapies based on CB1 cannabinoid receptor agonists and antagonists: focus on anxiety and depression. Best Practice & Research Clinical Endocrinology & Metabolism 23, 133-144.
Overstreet, D.H. (2012). Modeling depression in animal models. Methods Mol. Biol. 829, 125-144.
Patel, S., and Hillard, C.J. (2006). Pharmacological evaluation of cannabinoid receptor ligands in a mouse model of anxiety: further evidence for an anxiolytic role for endogenous cannabinoid signaling. J. Pharmacol. Exp. Ther. 318, 304-311.
Rinaldi-Carmona, M., Calandra, B., Shire, D., Bouaboula, M., Oustric, D., Barth, F., Casellas, P., Ferrara, P., and Le Fur, G. (1996). Characterization of two cloned human CB1 cannabinoid receptor isoforms. J. Pharmacol. Exp. Ther. 278, 871-878.
Seely, K.A., Brents, L.K., Franks, L.N., Rajasekaran, M., Zimmerman, S.M., Fantegrossi, W.E., and Prather, P.L. (2012). AM-251 and rimonabant act as direct antagonists at mu-opioid receptors: Implications for opioid/cannabinoid interaction studies. Neuropharmacology 63, 905-915.
Sellami, A., Al Abed, A.S., Brayda-Bruno, L., Etchamendy, N., Valério, S., Oulé, M., Pantaléon, L., Lamothe, V., Potier, M., Bernard, K., et al. (2017). Temporal binding function of dorsal CA1 is critical for declarative memory formation. Proc. Natl. Acad. Sci. U.S.A. 114, 10262-10267.
Sheehan, B. (2012). Assessment scales in dementia. Ther Adv Neurol Disord 5, 349-358.
Shore, D.M., Baillie, G.L., Hurst, D.H., Navas, F., Seltzman, H.H., Marcu, J.P., Abood, M.E., Ross, R.A., and Reggio, P.H. (2014). Allosteric Modulation of a Cannabinoid G Protein-coupled Receptor: BINDING SITE ELUCIDATION AND RELATIONSHIP TO G PROTEIN SIGNALING. Journal of Biological Chemistry 289, 5828-5845.
Vallée, M., Vitiello, S., Bellocchio, L., Hébert-Chatelain, E., Monlezun, S., Martin-Garcia, E., Kasanetz, F., Baillie, G.L., Panin, F., Cathala, A., et al. (2014). Pregnenolone can protect the brain from cannabis intoxication. Science 343, 94-98.
Varni, J.W., Seid, M., and Kurtin, P.S. (2001). PedsQL 4.0: reliability and validity of the Pediatric Quality of Life Inventory version 4.0 generic core scales in healthy and patient populations. Med Care 39, 800-812.
Walf, A.A., and Frye, C.A. (2007). The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat Protoc 2, 322-328.
Zavatti, M., Carnevale, G., Benelli, A., and Zanoli, P. (2011). Effects of the cannabinoid antagonist SR 141716 on sexual and motor behaviour in receptive female rats. Clin. Exp. Pharmacol. Physiol. 38, 771-775.
Diagnostic and statistical manual of mental disorders: DSM-5 (2013, Washington, D.C: American Psychiatric Association).
Rapport d’évaluation des médicaments indiqués dans le traitement symptomatique de la maladie d’ALZHEIMER (2016). Commission de la Transparence. Haute Autorité de Santé.
Donepezil, galantamine, rivastigmine and memantine for the treatment of Alzheimer's disease (2011). Technology appraisal guidance. National Institute for Health and Care Excellence.
EPAR discussion. https://www.ema.europa.eu/documents/-scientific-discussion/acomplia-epar-scientific-discussion_en.pdf.
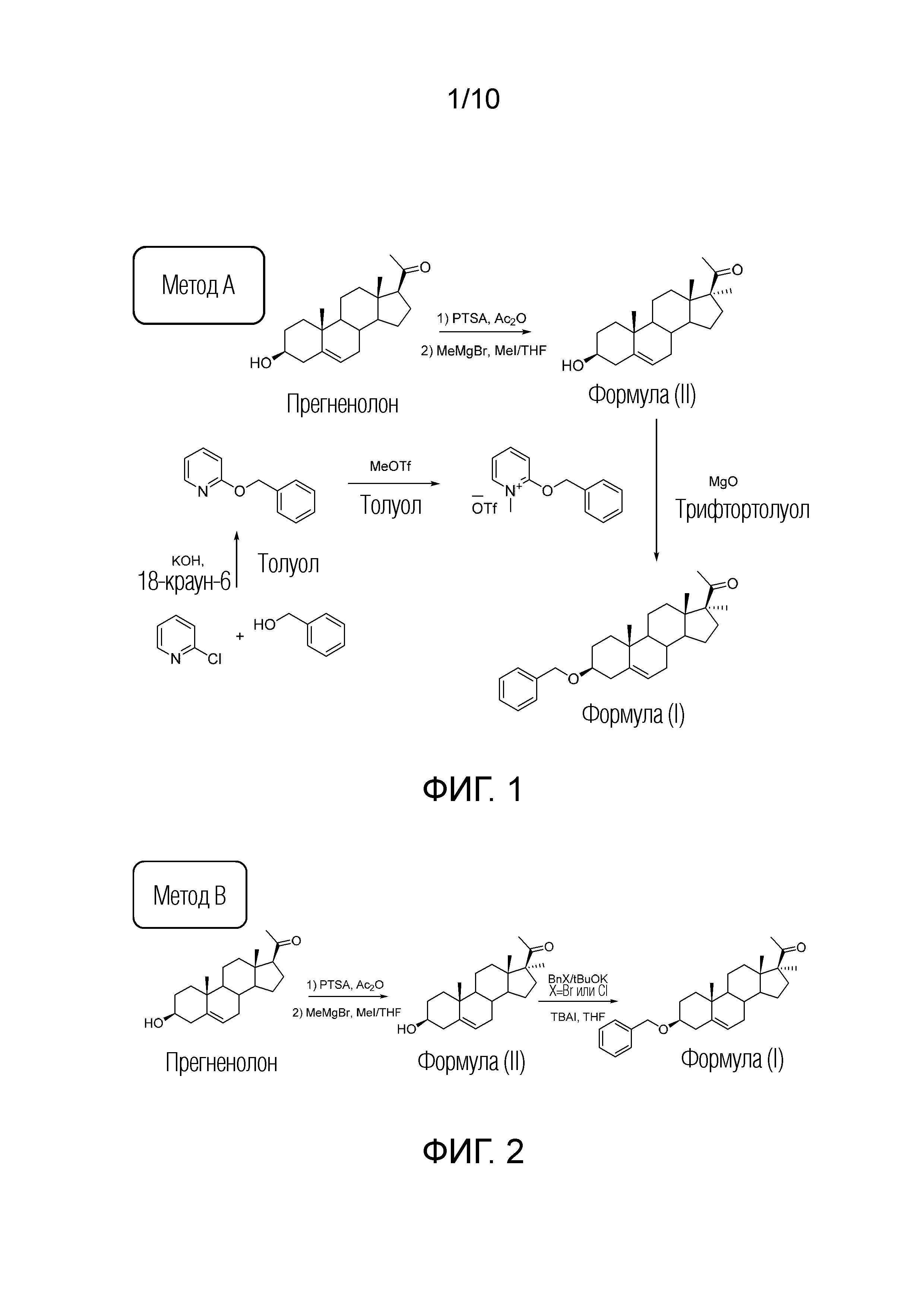
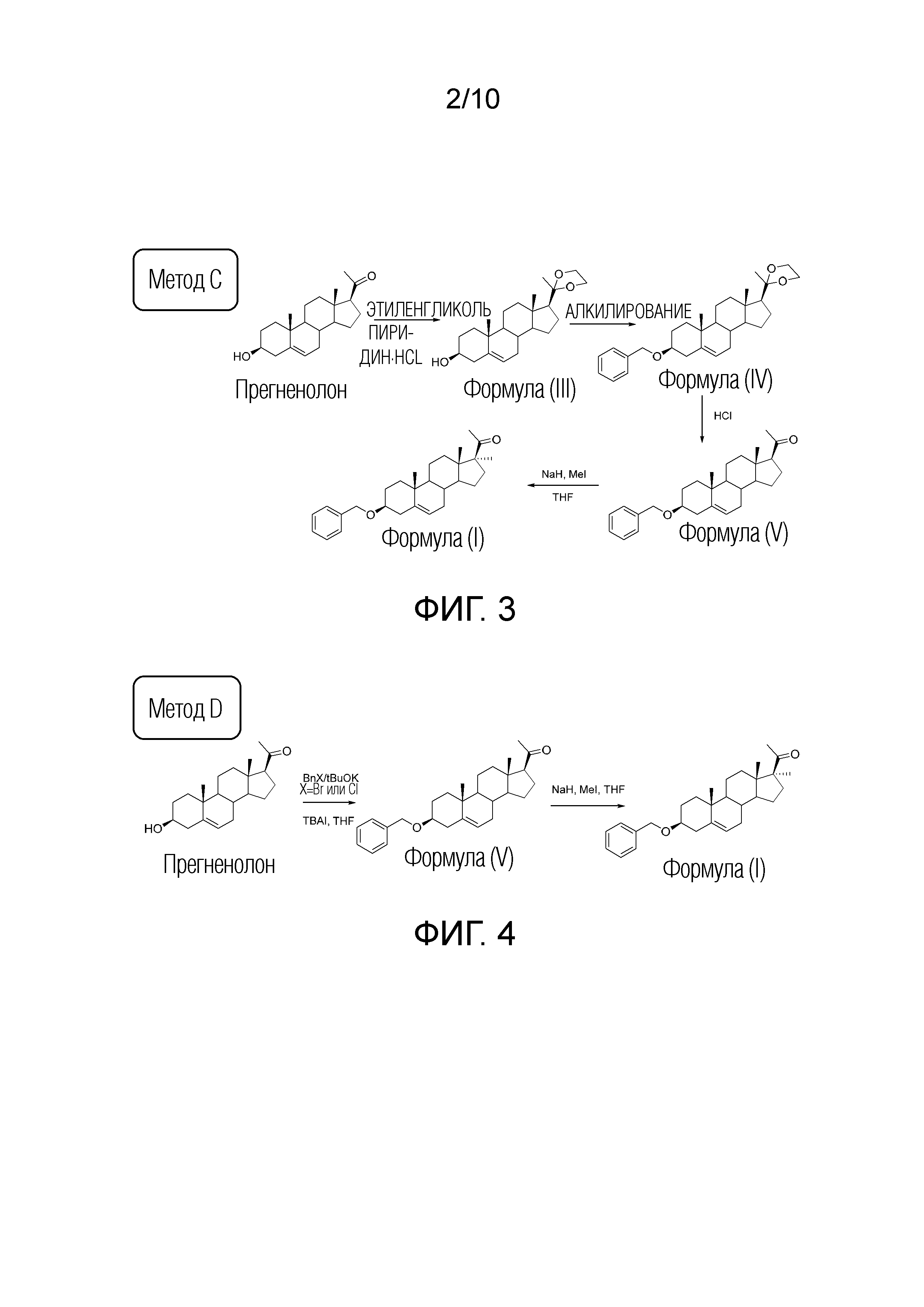
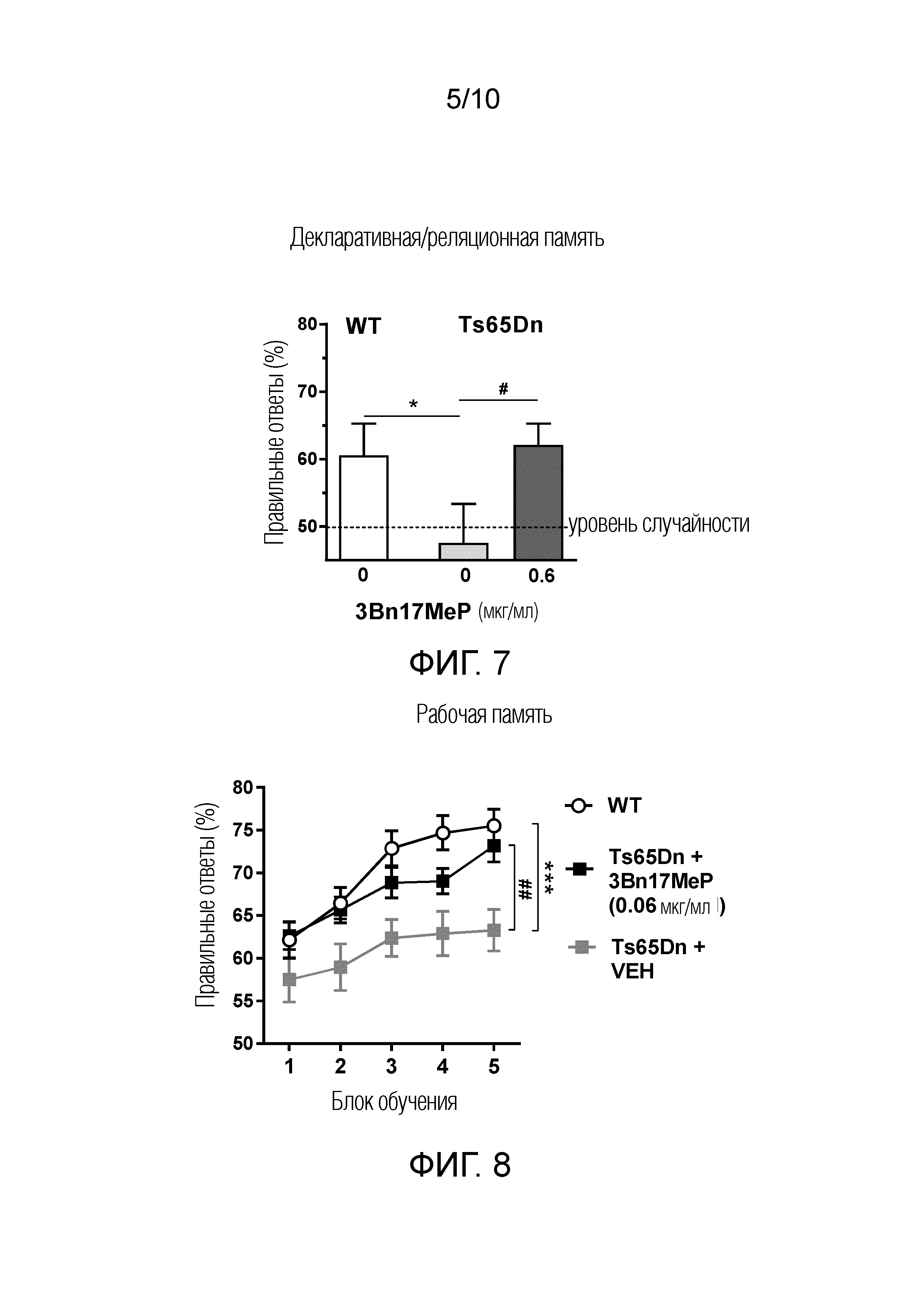

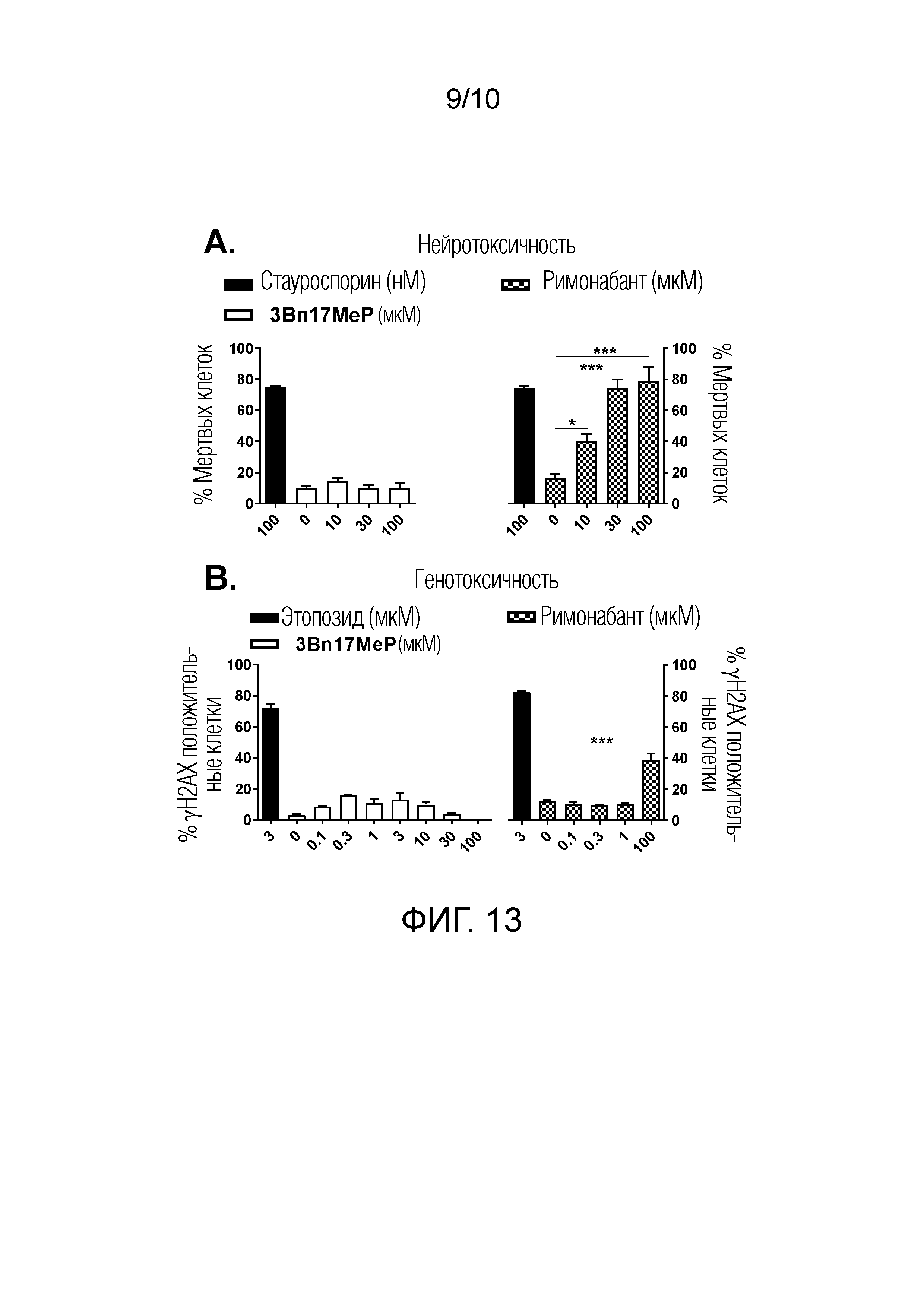
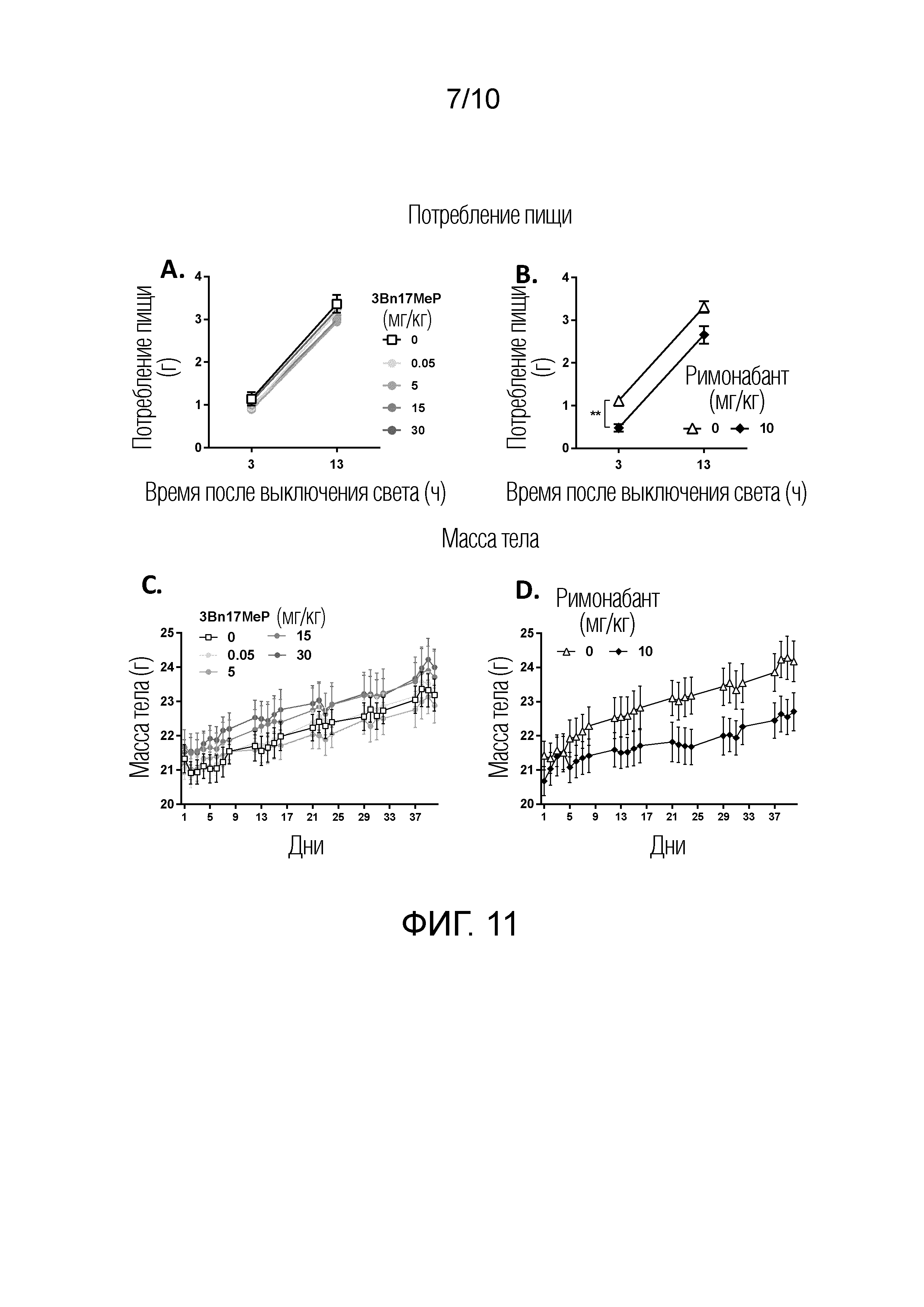
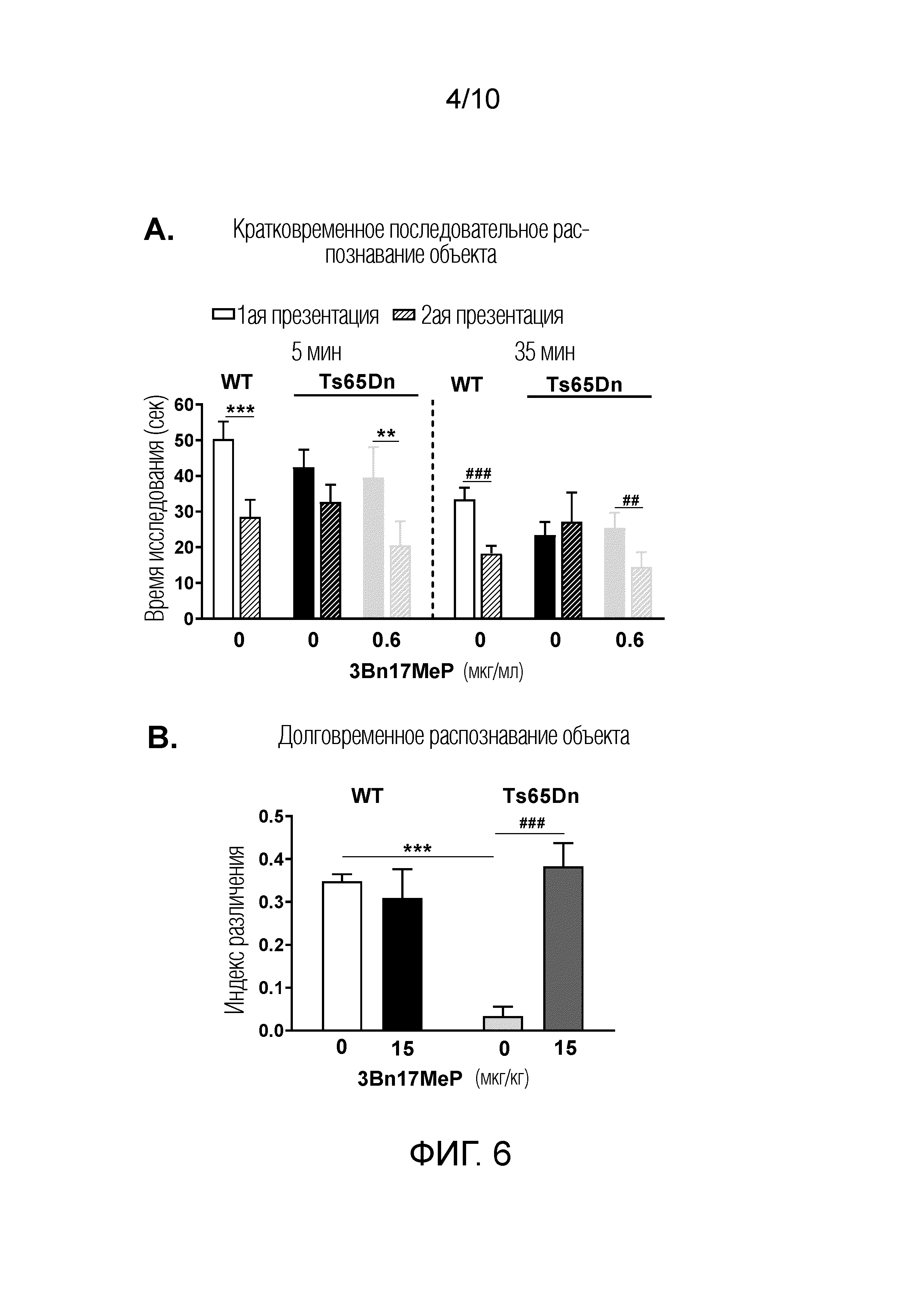
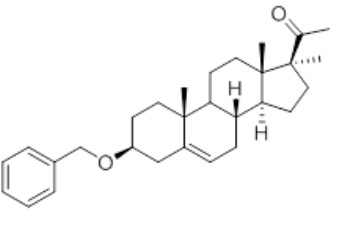
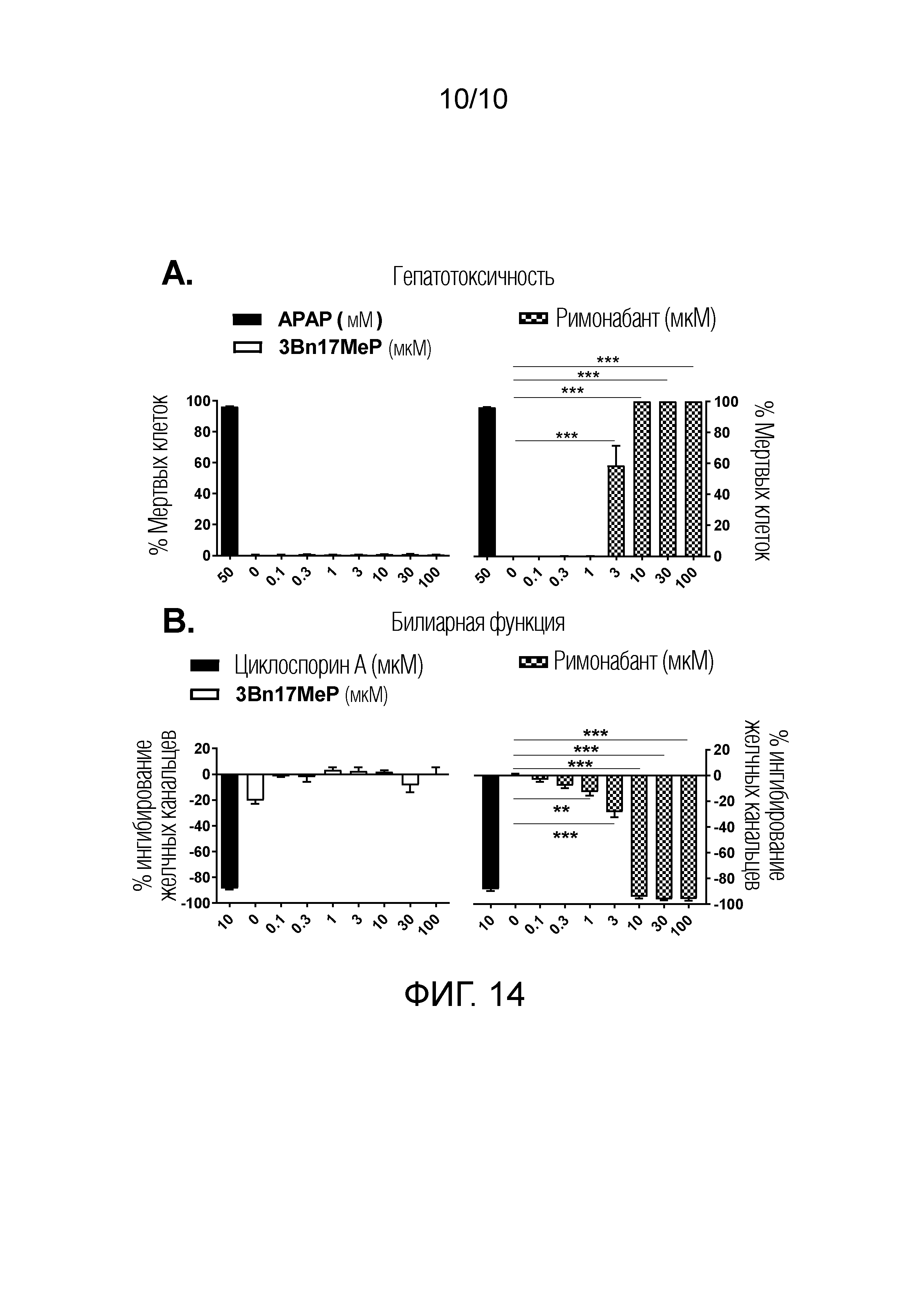
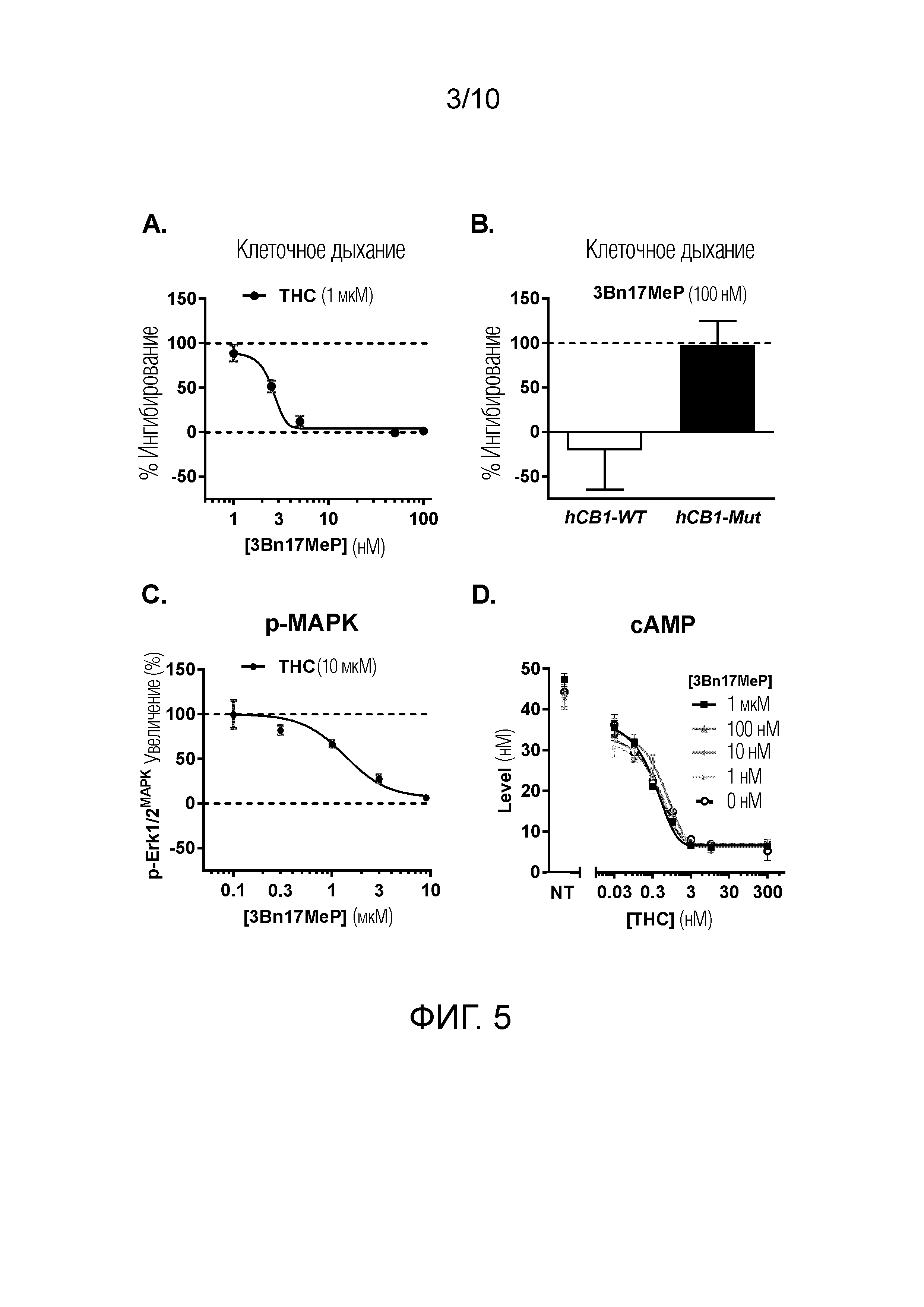
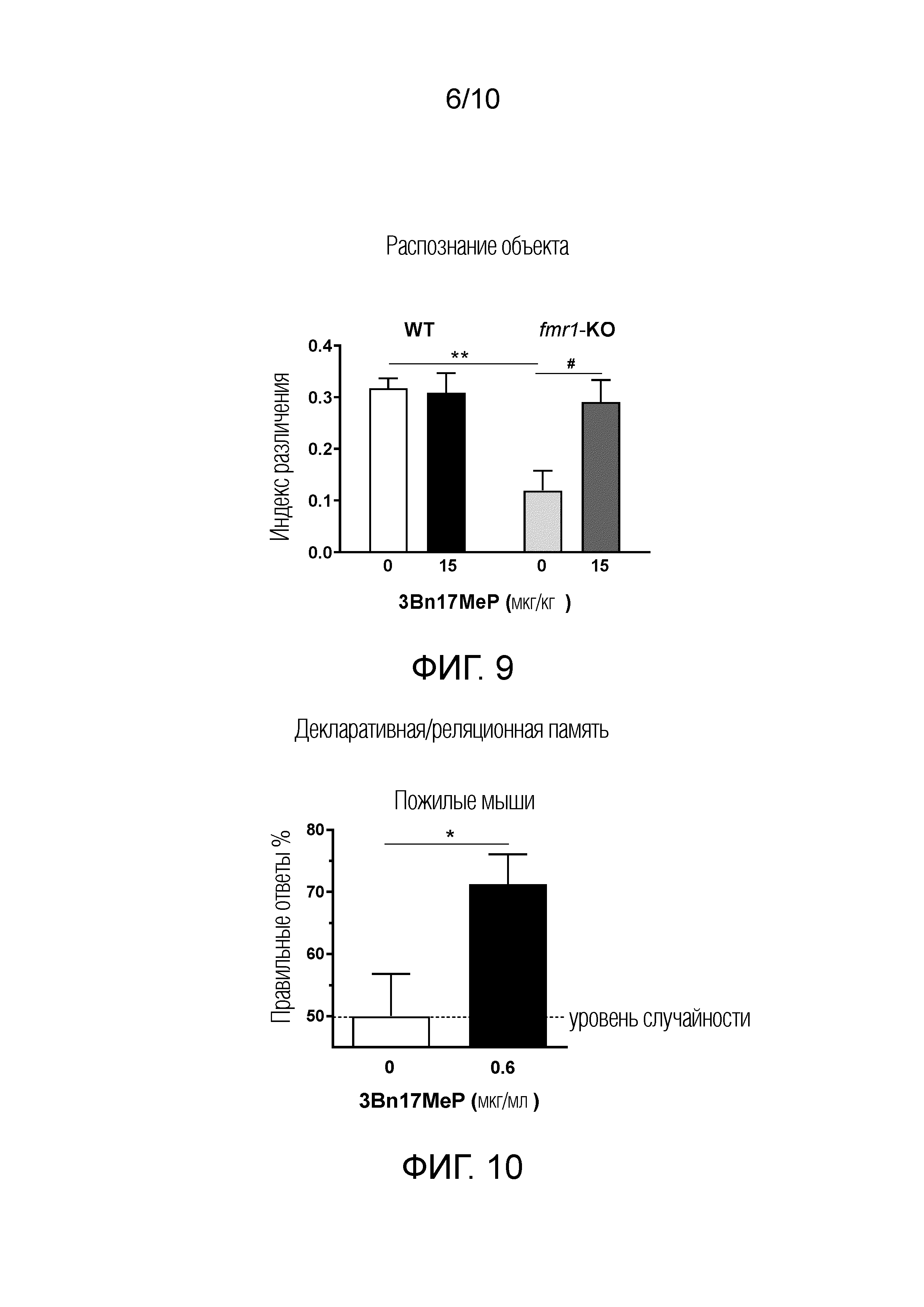
Антагонисты рецептора св1
Защитный материал от воздействия среды для тугоплавкой подложки, содержащей кремний
Производные прегненолона
Селективные ингибиторные пептиды nox-1 и их применение
Печать адгезивного шаблона на необрастающей подложке

