Результат интеллектуальной деятельности: Набор синтетических олигонуклеотидов для определения уровня экспрессии генов DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT сосны обыкновенной (pinus sylvestris l.) методом ПЦР в реальном времени
Вид РИД
Изобретение
Изобретение относится к области молекулярной биологии и лесной биотехнологии, в частности к наборам синтетических олигонуклеотидов, и может быть использовано для определения уровня экспрессии генов сосны обыкновенной (Pinus sylvestris L.) методом ПЦР в реальном времени.
Существенным недостатком традиционных методов получения сортов и форм древесных видов, таких как гибридизация и мутагенез, является длительность процесса, так как необходимо проводить продолжительную оценку полученных гибридных или мутантных потомств в специально созданных испытательных культурах. При этом объектом исследований являются только признаки и свойства, формируемые на базе созданного генотипа. Развитие молекулярно-генетических методов исследования позволяет существенно сократить длительность селекционного процесса и повысить его эффективность за счет прямого изучения экспрессии генов, связанных с формированием хозяйственно-важных признаков и свойств деревьев.
В настоящее время решение научных задач фундаментального и прикладного характера в области молекулярной биологии и лесной биотехнологии требует изучения уровня экспрессии генов. Одним из подходов изучения уровня экспрессии генов является определение количества продуктов работы генов - матричной РНК (мРНК) методом полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР). При этом фиксация результатов может быть выполнена по конечной точке с помощью электрофореза после использования классической ПЦР или с помощью флуоресцентной детекции при использовании ПЦР в реальном времени (ПЦР-РВ). Методы флуоресцентной детекции результатов ПЦР-РВ основаны на использовании меченых красителями зондов или на использовании интеркалирующих веществ (SYBR Green I/II и SYBR Gold). Существенным ограничением использования вышеуказанного подходя является отсутствие высокоспецифичных к определенным генам олигонуклеотидов, позволяющих определять наличие или отсутствие экспрессии целевого гена, а также давать количественную оценку уровня экспрессии методом ПЦР-РВ применительно к конкретным видам растений.
Из литературы известны последовательности праймеров, используемых для определения уровня экспрессии следующих генов сосны обыкновенной (Pinus sylvestris) с детекцией результатов ПЦР по конечной точке методом электрофореза:
1. Праймеры к генам LEA (late embryogenesis abundant protein) F- TCCGCAGAGGTTACAGACATCG, R- 5’CTATTTGCGC TCAGGAGTCGAA-3; P5CS2 (delta-1-pyrroline-5-carboxylate synthase F-GATCCCAAGAGGTCAGCA, R-GAATCCTGCTTGTGCTTATTCC; AbaH (abscisic acid and water-stress induced protein) F-AGGACAACGT TAATTCTGGCTC, R-AATCGGCCTTATAACCAGTGTCG; GapC1 (glyceraldehyde-3-phosphate dehydrogenase) F-ACGGTTTTGGTCGAATTG, R-CCCACGAGCTCGATATCAT [Voronova A., Jansons А.,  Expression of retrotransposon-like sequences in Scots pine (Pinus sylvestris) in response to heat stress. Environmental and Experimental Biology. 2011. 9: 121-127];
Expression of retrotransposon-like sequences in Scots pine (Pinus sylvestris) in response to heat stress. Environmental and Experimental Biology. 2011. 9: 121-127];
2. Праймеры к гену Peroxidase F - GCTCTAGCGGCTAAAGAGT и R - GAGGTCTGTGACGTTGAGA [Kovaleva V.A., Hrunyk N.I., Yusypovych Yu.M., Gout R.T. Expression of the peroxidase dene from Pinus Sylvestris L. in seedlings under abiotic stress. Науковий вісник НЛТУ України. 2016. 26(8): 87-95];
3. Праймеры к генам семейства Difensin PsDef1 F-GGGATGATGCAGGTTCAAGT, R-ACATTTTCTGCCAGCCACAT; PsDef2 F-TCCACTCAGTGCCCTTTTTC, R-ACCAGCCGAAAGTGCTACTG; PsDef3 F-AACCATTGGGATGATGGC, R-GCACTTTCGGCTGGTGAC; PsDef4 F-TGTGCTGCTCGTGTTAG, R-CGTTGGAAACCCTTCAGTA; PsDef5.1. F- TCTTGCTACTTGCAATACGT, R-GATACATGGTTTCTCGCAGA и гену 60S рибосомальнго белка F-CAAAGCTTGCAAAAAGCACA, R-TTCCCTTCCCCTTCTTGTCT [Шаловило Ю. I., Юсипович Ю. М., Ковальова В. А., Гут Р. Т. Вплив фітогормонів на експресію генів дефензинів сосни звичайної. Біологічні Студії. 2015. 9 (1): 15-24 (Shalovylo Y.I.,Yusypovych Y.M., Kovaleva V.A., R.T. Gout Shalovylo Y.I. The effect of phytohormones on expression of defensin gene in Scots pine. Studia Biologica. 2015. 9 (1): 15-24)];
4. Праймеры к генам Lipid-transfer protein F-ATGGCTGTGAAGAAGATG, R-TCAGTGAACCTTGGAACAG; 60S рибосомальнго белка F-CAAAGCTTGCAAAAAGCACA, R-TTCCCTTCCCCTTCTTGTCT [Груник Н. I., Ковальова В. А., Гут Р. Т. Особливості експресії гена ліпідтрансферного протеїну в органах сосни звичайної (Pinus sylvestris L.). Біологічні Студії. 2012. 6(2): 151-160 (Hrunyk N. I., Kovaleva V. A., Gout R. T. Patterns of expression of lipid-transfer protein gene in organs of scots pine (Pinus sylvestris L.). Studia Biologica. 2012. 6(2): 151-160)];
5. Праймеры к генам PsWUS/WOX5 (wuschel homeobox protein WOX) F- TCACAGCTCAAACGATACGG, R- CAGGGGCGTCCACTTCTAT; Actin F- CCAACAGGGAGAAAATGACG, R- CTGACGCCACATTTACCAAA [Nardmann J., Reisewitz P., Werr W. et al. Discrete Shoot and Root Stem Cell-Promoting WUS/WOX5 Functions Are an Evolutionary Innovation of Angiosperms. Mol. Biol. Evol. 2009. 26(8):1745-1755].
Из литературы также известны последовательности праймеров, используемые для определения уровня экспрессии методом ПЦР в реальном времени с использованием интеркалирующего красителя SYBR Green следующих генов сосны обыкновенной (Pinus sylvestris):
1. Праймеры к генам ADC (arginine decarboxylase) F-AGTCCGTGTGGCCTGTAATC, R-TGCACAGACACAACGTCAAA; SPDS (spermidine synthase) F-CCAACGTCCCATTAACCCTA, R-TGGCAAACAAAATGATGCTG; ACL5 (tSpm synthase) F-ACTGCTCACATTCCGTCCT, R-TTCGCCTTTGATTCTCTTGCT; DAO (diamine oxidase) F-AATGGGGAAGTTGGGAGTTC, R-CCCTCCTCAGTTTTCCAGTG; CAT (catalase) F-GGGAGGCAAACCTATGTGAA, R-TTGGTTGCATGACTGTGGTT; RAD51 (DNA repair gene) F-TATGGGGAATTTCGAACAGG, R-GTTCCCTCGGCATCAATAAA; KU80 (KU80-like protein) F-GCGGCTATGAGAATGTGGTT, R-AAAATCACCTGGAGCCATTC; ATG5 (autophagy-related genе) F-GGGTGAAGACAGCGTGAAAT, R-GCCATAGCTCTATTTGATCTG; ATG8 (autophagy-related gene) F-CCTGCTGATCTGACAGTTGGT, R-AGCAGCAGTTGGAGGTAGGT [Salo H. M., Sarjala T., Jokela A. et al. Moderate stress responses and specific changes in polyamine metabolism characterize Scots pine somatic embryogenesis. Tree Physiology. 2016. 36: 392-402];
2. Праймеры к генам CAT (catalase) F-GGGAGGCAAACCTATGTGAA, R-TTGGTTGCATGACTGTGGTT; βG (β-glucosidase) F-CAAATCTGTTTGTGCCGTTG, R-CTGACGAGCAATTCCCTGTT; RBR (retinoblastoma-related protein) F-ACAGGAAGCAACCTCAGTGC; R-TCCACTGTCTCATGCCCTAA (Vuosku J., Sutela S., Kestilä J. et al. Expression of catalase and retinoblastoma-related protein genes associates with cell death processes in Scots pine zygotic embryogenesis. BMC Plant Biology. 2015. 15:88];
3. Праймеры к генам ADC (arginine decarboxylase) F-AGTCCGTGTGGCCTGTAATC, R-TGCACAGACACAACGTCAAA; SPDS (spermidine synthase) F-CCAACGTCCCATTAACCCTA, R-TGGCAAACAAAATGATGCTG; ACL5 (tSpm synthase) F-ACTGCTCACATTCCGTCCT, R-TTCGCCTTTGATTCTCTTGCT; DAO (diamine oxidase) F-AATGGGGAAGTTGGGAGTTC, R-CCCTCCTCAGTTTTCCAGTG; PAO (flavoprotein-containing polyamine) F-CGAAATTGCAGAACCTCCAC, R-CGGCCACGAACTACTCATCT; SAMDC (S-adenosyl methionine decarboxylase) F-GCTTCGGCGAGGAAATATCTTA, R-TGTTTGCGGTCCAGTTG; SAMS (s-adenosyl methionine synthetase) F-ACTGCAAAGTGCTGGTT, R-ATGGGTCAGTGGCATAAG; LEA (late embryogenesis abundant protein) F-ACCCTCGCAGAGGTTACAGACA, R-TTGGCCTTCACTGACCCAGGA; CAT (catalase) F-GGGAGGCAAACCTATGTGAA, R-TTGGTTGCATGACTGTGGTT; ICE1 F-TTAGCTTGCTCTGCCCGAAA, R-TCACTTCCCAGTCCCAATGC; R2R3-MYB8 (R2R3-MYB transcription factor MYB8) F-CTTCCTGGAAAGATTTAATAGTGT, R-GGAGCCTGCAATACCCATA; PAL (phenylalanine ammonia-lyase) F-GAGGGAATTTCCAGGGCACA, R-GATCTCGGCCCCTTTCAGTC; psSTS (pinosylvin-forming stilbene synthase) F-ATGTTTCCGTACTCGCTCATAAC, R-ACAAGTTCAAGCGAATATGTGAA; UBQ (putative ubiquitin) F-GAAGGAGCAGTGGAGTCCTG, R-CAATTTCAGGGACGAGGA; TUBA (alpha tubulin) F-TGGCCGCATCTTCCTTGCCG, R-GATGGCCAGTGCCCAGCGA; [Muilu-Mäkelä R., Vuosku J., Saarinen M. et al. Coping with spring frost-effects on polyamine metabolism of Scots pine seedlings. iForest. 2017. 10: 227-236];
4. Праймеры к генам ADC (arginine decarboxylase) F-AGTCCGTGTGGCCTGTAATC, R-TGCACAGACACAACGTCAAA; AIH (agmatine iminohydrolase) F-TACCACATGCCTGCTGAATG, R-TCAGCAAAGACCGTTGACC; CPA (N-carbamoylputrescine amidohydrolase) F-TTCAGTCCAGGTGACACAGG, R-CTCTTGCTGCCTCTGGAAAC; SPDS (spermidine synthase) F-CCAACGTCCCATTAACCCTA, R-TGGCAAACAAAATGATGCTG; ACL5 (tSpm synthase) F-ACTGCTCACATTCCGTCCT, R-TTCGCCTTTGATTCTCTTGCT; SAMDC (S-adenosyl methionine decarboxylase) F-AAGGGGCAGCATTTCCACT, R-CTCCACCAGTGCTTCAAGGT; DAO (diamine oxidase) F-AATGGGGAAGTTGGGAGTTC, R-CCCTCCTCAGTTTTCCAGTG; PAO (flavoprotein-containing polyamine) F-CGAAATTGCAGAACCTCCAC, R-CGGCCACGAACTACTCATCT; CAT (catalase) F-GGGAGGCAAACCTATGTGAA, R-TTGGTTGCATGACTGTGGTT; MCA (metacaspase) F-CGGGAGAGGACATGCTAAAA, R-CTGGCCTAATTTTCCCAACA; TAT-D (TAT-D nuclease) F-TGGATGTTCCTTAAAGACAGTGG, R-TCTCACAGTATGGAGCGTCTG; E2F (E2F transcription factor) F-TGTGCTTGAGGGGATAGGAC, R-TTCAACCTCTCCAGGTCTCG; RBR (retinoblastoma-related protein) F-GGAATTGCAAAGATCCAACAA, R-ATCGCCTCGATTCAACAAAG [Vuosku J., Suorsa M., Ruottinen M. et al. Polyamine metabolism during exponential growth transition in Scots pine embryogenic cell culture. Tree Physiology. 2012. 32: 1274-1287];
5. Праймеры к генам Dirigent protein F- GTTAAGCACAAAAGTGAATCCAAG, R- TTCTACTTTTATGACATGAAGAACACC; Kunitz-type protease inhibitor F- AATGAATGAGAGACGCACCA, R- TCACAGCAACACAGTCGTTTT; Laccase F- CTGCTCAAAACATAAGCACAGC, R- GAACTTTCACTCTCGTGTCTTTCA; Transcription factor (NAC family) F- TGGAAAGCTACTGGGACAGAC, R-AGAGCCTTTTTCACCCCAAC; Cinnamoyl-CoA reductase F- GCTCACATTTTAGTTTATGAGACACG, R- CGGTGGAGGCTACATTCTG; Terpene synthase F- GCTCTCAGAATGTCGAAAGCA, R- GAAACCAGACACCAATGTTCC; Chalcone synthase F- CCGAATCCAAACAGAACTCC, R- CGCAGAATGGATGTTCGAC; β-glucosidase F- TTTCCGGCTGTGCAATTT, R-CTACGATGTGGGGATTCTCG; Serine-type protease inhibitor F- AGCAATTGCGAACACACACT, R- TGCACAAGGAATTGGAGAGA; Pectin methylesterase F- ATGGCTACGGCTTGAGAATG, R-GTCAATGGATTCGCTGCAA; Phospholipase A F- AGAGGCCCCAGTTCTAAGGT, R- CACAGGGAGCTTCGTTATGG; OPDA reductase F- TCCCGTCATAGACCACAATG, R-GTTCAGGACACTGATTAGGTGGT; Nitrate transporter F- AGTTGGAATTTCAGCGCAAC, R- ATCCCCATCACAGATCCTCTT; Transcription factor F- CCTGAGAGCCTCTACGTCAATAG, R- TGCTGTAAGTTCACGCCAAA; Oxidoreductase F- ACATCCATGGCCAATTTTTC, R- GTTGGCTAAACGAGGAGCAC; Invertase F- ACAGCCATCCGCATCATAA; R-GCACATGAAAACCTCCATCC; Hydrolase F- GAGATCGGGGATGAGGAGAT, R- AGGCTGGGATGACCAAATC; Cation/proton exchanger F- AGTTGGGTCCAATTGGGAAT, R- CAATACTGATGCGCTGATGG; TUBA F-GATCTGGTTCAGCCTGATGG, R-CCAGTCCGTACCTCGTCAAT; EF1-a F-CACCTTGGGAGTGAAGCAAATG, R-GGGAGTAGTGGCATCCATCTTG (Kovalchuk A., Raffaello T., Jaber E.et al. Activation of defence pathways in Scots pine barkafter feeding by pine weevil (Hylobius abietis). BMC Genomics. 2015. 16:352];
6. Праймеры к генам PsFTL2 (flowering locus T) F-AGCAGGCAGGTTAACAATGG, R-TTGGATCGCTTGGACTGGGA; GI (gigantea) F-GCAGATGGACTATGGAACC, R-GCTTTATGCCAATCTTGAAC; PRR1 (pseudo-response regulator) F-ACTCCAATACCAACAGTACCAA, R-ATATGTGAGGAAAGCTGATGC; ZTL (zeitlupe) F-ATGCAGCCTATGAATGACAC, R-CCAAATCCAGAATGAAAACATCG; Actin F-TGACATGGAGAAGATTTGGC, R-CATACATAGCAGGCACATTG; GAPDH (glyceraldehyde-3-phosphate dehydrogenase) F-CTGGTGTCTTCACCGACAAA, R-GGTGCTCATTAACCCCAACA [Avia K., Karkkainen K., Lagercrantz U. and Savolainen O. Association of FLOWERING LOCUS T/TERMINAL FLOWER1-like gene FTL2 expression with growth rhythm in Scots pine Pinus sylvestris). New Phytologist. 2014. 204: 159-170.];
7. Праймеры к гену PsDef1 (defensin 1) F-CAACGGGAAGTTGCGATT, R-TCAAGGGCAGGGTTTGTAAC (Jaber E., Xiao Ch., Asiegbu F. O. Comparative pathobiology of Heterobasidion annosum during challenge on Pinus sylvestris and Arabidopsis roots: an analysis of defensin gene expression in two pathosystems. Planta. 2014. 239:717-733];
8. Праймеры к генам PsHAP3A (LEC-type HAP3 gene) F-AGCTCGATCTTCGGTCTTCA, R-TGCCAACTTCGGACATCATA; PsVP1 (viviparous1) F-GTGCCCAATTTGGTTAATGG, R-GTGCCATCAAAGACAGCAGA [Uddenberg D., Valladares S., Abrahamsson M.et al. Embryogenic potential and expression of embryogenesis-related genes in conifers are affected by treatment with a histone deacetylase inhibitor. Planta. 2011. 234:527-539.]
9. Праймеры к генам Ef1-a (Elongation factor 1-α) F- ACTCTGGCAAGTCCACCACT, R- TCCTTCTCAAAACGCTCGAT; GH3 (GH3 homologue) F- ACGCTATCAGCTCACTGGTC, R-TTGGCTATGCGAAGAATCTC; iaa88 (transcription factor) F- GCAAATTCAAAGCCTAACGA, R- GCTGAGGAAAGCTCATGGTA; MtN21-like-a (noduline like) F- CTTTAAGCGGAGGGATGAAT, R-TTCCACGAAGAAAGCAAAAG; MtN21-like-b (noduline like) F- TTGGTCTCCTCGTCATCAAT, R- TCAGAAGAGAGCCGACAATC; 5NG4 (noduline like) F- CGCTGGATTCCATATCGTTT, R- AAGGGCCAATCAACATCAAG; Clv1-like (clavata like) F- TAATAATCTGAGCGGCAGCA, R- TTCGAGGAAACATCAAGCTG (Heller G., Lundén K., Finlay R. D. et al. Expression analysis of Clavata1-like and Nodulin21-likegenes from Pinus sylvestris during ectomycorrhiza formation. Mycorrhiza (2012) 22:271-277];
10. Праймеры к генам PsTPS1 (Terpene synthases 1) F- TACGGAATGCTAGACGAACTG, R-GCGGCCAGTTCAGACGAACTG; PsTPS2 (Terpene synthases 2) F- GGATCAGATTCTACCAGACG, R-GTTATGTCGTCCTTGACTCG; PsTPS3 (Terpene synthases 3) F-AGGGTATATGCAAGAGTCGC, R-GTATACACATTCCAGAGCTGC; Ubi (ubiquitin) F-GTTGATTTTTGCTGGCAA, R-CACCTCTCAGACGAAGTA [Köpke D., Schröder R., Fischer H. M. et al. Does egg deposition by herbivorous pine sawflies affect transcription of sesquiterpene synthases in pine? Planta. 2008. 228:427-438];
11. Праймеры к генам PER (peroxidase) F-GCAGGTACCTTCCATGATCG, R-ATTCTCGCCATTGCTGCT; AMP (antimicrobial peptide) F-TGCCCACGCAGTGAAATA, R-TGTTTGTGTTGGTAAGCTTGTTTT; Thau (thaumatin) F-TTGTTGCCACCATTCAGAGT, R-TGCTCAATTGCCAAGGTTC; metallothionein like protein F-GCACTTGCAGTCATTTTCCA, R-GGGATTCCAGATGGATGGTA; PR-10 (pathogenesis-related 10) F-TTTATTTTGCACGGTGAACG, R-TCATGTTGAGGAAAGTGGAGAA; MAP kinase F- GCCCTGCTCAAAATCAAAAT, R- TAGCAAGTCTGCATGGCATC [Sun H., Paulin L., Alatalo E., Asiegbu F. O. et al. Response of living tissues of Pinus sylvestris to the saprotrophic biocontrol fungus Phlebiopsis gigantea. Tree Physiology. 2011. 31: 438-451];
12. Праймеры к генам RAD51 F-TATGGGGAATTTCGAACAGG, R-GTTCCCTCGGCATCAATAAA; KU80 F-GAATGGCTCCAGGTGATTTT, R-AGGCGTTTATTTCCCTTGCT; LIG (DNA ligase) F-GCATTAGCCCTGTTCATCGT, R-CGCTTGGTCTGGATTCTTGT; MCA (metacaspase) F-CGGGAGAGGACATGCTAAAA, R-CTGGCCTAATTTTCCCAACA; TAT-D (Tat-D nuclease) (F-TGGATGTTCCTTAAAGACAGTGG, R-TCTCACAGTATGGAGCGTCTG; ACT (actin) F-GGACAGGTCATTACCGTTGG, R-GATACCCGCTGCTTCCATT; UBI (ubiquitin) F-GAAGGAGCAGTGGAGTCCTG, R-CAATTTCAGGGACGAGAGGA; GAPDH (glyceraldehyde-3- phosphate dehydrogenase) F- CTGGTGTCTTCACCGACAAA, R- GGTGCTCATTAACCCCAACA (Vuosku J., Sarjala T., Jokela et al. A. One tissue, two fates: different roles of megagametophyte cells during Scots pine embryogenesis. Journal of Experimental Botany. 2009. 60(4): 375-1386];
13. Праймеры к генам membrane intrinsic protein/Porine MIP1 F-GGCCAGAAAGTTGTCTCTGC, R-ATGAAGCCCTTCACAACACC; putative auxin induced F-TGGCTCCTAAGCCCACTAAT, R-CTCCACCCACTATGAACACG; anthocyanidine synthase F-GATGCGATAAAGGGAGTTGC, R-GGAGCACTTTTCTGCTTTT; disease resistance protein F-ACAATTGCCCGAAATTGAAG, R-ATTGCAACCCACCATTCTCT; unknown protein 2 F-AAAGGCATCCCCTGTTTCTT, R-GGACTAGCCCTGGGACTACC; unknown protein 3 F-ATTCATTGCGCTTTTCTGCT, R-CGCTTTGTGTTTAGCAGGTG; unknown protein 4 F-CGAAGCCATTGCTTTTCTTC, R-TCCTCATGGATCGAATCTCC; aldehyde dehydrogenase homolog F-CGTGGGGAATTCTCGTTTTA, R-TTGAGTGCAAGCCTAGCTGA; endoglucanase 1 F-AAGTCCGGATTCTTCAGTGC, R-AGAGTGCTGCAATGACTCCA; PR10/BetV1 F-AGCGAGCTGGACCTGTAACT, R-GGGCAAGGCTATCTTCCTTT; Clavata1 F-TAATAATCTGAGCGGCAGCA, R-TTCGAGGAAACATCAAGCTG; tau class glutathione S-transferase F-AATACATCGAGGAGGCATGG, R-TATGGGTCTTCGGGCATAAG; subtilisine like protease precursor F-CATGAACTATCCGACCATCG, R-GAGGCGGATCCTATGTTTGT; kanamycin F-GGACGGCGGCTTTGTTG, R-CTGCGTTGTCGGGAAGATG [Heller G., Adomas A., Li G. et al. Transcriptional analysis of Pinus sylvestris roots challenged with the ectomycorrhizal fungus Laccaria bicolor. BMC Plant Biology. 2008. 8:19];
14. Праймеры к генам ADC (arginine decarboxylase) F- AGAAATTGGGGATGCTGGAT, R- GCCATCACGATTGTATTCACC; ODC (ornithine decarboxylase) F- AAGCGGTGAAGCCATTAAAA, R- TTGCGTTGCAGACGTATTTC; Act (Actin) F- GCTTGCTTATGTAGCCCTTGA, R- GGTCTTGGCAATCCACATCT [Vuosku J., Jokela A., Läärä E.et al. Consistency of Polyamine Profiles and Expression of Arginine Decarboxylase in Mitosis during Zygotic Embryogenesis of Scots Pine. Plant Physiology. 2006. 142: 1027-1038].
Перечисленные олигонуклеотиды являются строго специфичными к указанным генам и не могут быть использованы при изучении экспрессии генов DHN1 (dehydrin 1), DHN7 (dehydrin 7), SGS3 (suppressor of gene silencing), CPK (calcium-dependent protein kinase), CYP450 (cytochrome P450), ERD3 (early responsive to dehydration 3), CCOAOMT1 (caffeoyl-CoA-O-methyltransferase 1), кроме генов LEA (late embryogenesis abundant protein) и ACT (actin). Для генов LEA и ACT в литературе не представлены последовательности олигонуклеотидов для изучения экспрессии с использованием флуоресценомеченных зондов. Однако, использование специфичных к генам флуоресцентномеченных зондов совместно со специфическими праймерами, позволяет существенно снизить вероятность неспецифичной амплификации ДНК за счет увеличение суммарной протяженности синтетических нуклеотидов для ПЦР, так как при использовании интеркалирующего красителя применяются только прямой и обратный праймеры, и не используется комплементарный к внутренней части амплифицированной ДНК зонд.
На основе анализа уровня техники была поставлена техническая задача, направленная на разработку набора синтетических олигонуклеотидов, состоящего из прямых праймеров, обратных праймеров и флуоресцентномеченных зондов специфичных к генам DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT сосны обыкновенной.
Технический результат заключается в повышении надежности оценки уровня экспрессии генов сосны обыкновенной (DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT), за счет использования комплементарных к внутренней части амплифицированной ДНК флуоресцентномеченных зондов, что приводит к увеличению суммарной протяженности синтетических нуклеотидов и снижению риска неспецифической амплификации.
Технический результат достигается тем, что набор синтетических олигонуклеотидов для определения уровня экспрессии генов DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT сосны обыкновенной (Pinus sylvestris L.) методом ПЦР в реальном времени, включающий следующие последовательности:
DHN1_F: GGAAGTGGAGAAGAAAGTAGGTT
DHN1_R: CCGGGGAGTTTCTCTTTGATTTTA
DHN1_Z: FAM-TAACAAGAAGGAAGGGGAAGGGAAAGAAGAAGAAG-BHQ1
DHN7_F: GGCAAAAAGAAGGAAGATGAGGG
DHN7_R: GGCAGCTTCTGTTTGATTTTATCAAC
DHN7_Z: FAM-TTCTTCCCCTCTTCACATCTACCACCCTCAGT-BHQ1
SGS3_F: ATCGGAGCTTGGAAGGGGA
SGS3_R: CTTGGTTTCCATTCCTCTGTTG
SGS3_Z: FAM-TAAGGAACGGGGATGGAAGGAGTCAGG-BHQ1
CPK_F: CTACAAAGCAGCATCAGCAGTG
CPK_R: CGATGGTCTTGCACGCGAAC
CPK_Z: FAM-TCGGGCGTGGGCAGTTCGGCGTA-BHQ1
CYP450_F: ATTTGGATTGGCTCGATTT
CYP450_R: GATGGTTTCCCGTGTAAT
CYP450_Z: FAM-TGCGGAGAAAATAATAGACGACCA-BHQ1
ERD3_F: GTTGATGGTCTTTTCAGTGCAG
ERD3_R: CTGCGGCTAGATTCTTGACA
ERD3_Z: FAM-TGTGTTGTTGGAAATGGACCGTATATTAAGACCTG-BHQ1
CCOAOMT1_F: TGCAGTATATATTGGAAACGAGC
CCOAOMT1_R: TTCTTGGCGTTAATGAGCTTC
CCOAOMT1_Z: FAM-TACTTCTGCCGATGAGGGTCAATTTCTGGG-BHQ1
LEA_F: TATGATCATGACCTACCAATTGGG
LEA_R: CGTAAGGCACTGTCACTGGAA
LEA_Z: FAM-TCTGGAACAATTGTAGATCCTGGGTCAGTGAAGG-BHQ1
ACT_F: TGAGCTTCGAGTTGCTCCAG
ACT_R: CCAGTTGTACGACCACTTGC
ACT_Z: FAM-TCCCAAGGCAAACAGAGAGAAGATGACTCAG-BHQ1
Изобретение иллюстрируют графические материалы.
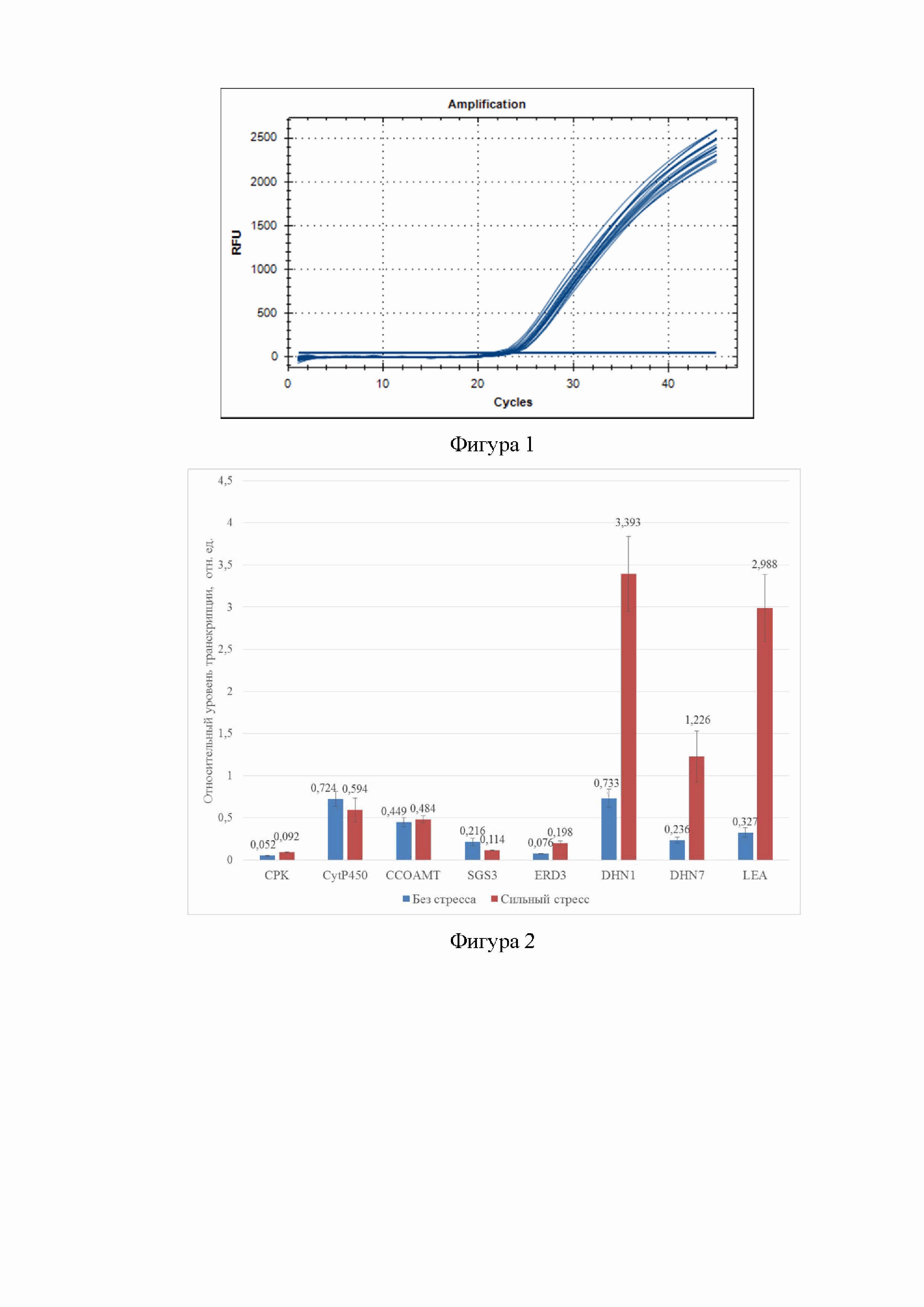
Фигура 1. Результаты ПЦР в режиме реального времени с применением специфических праймеров и зондов к гену CPK.
Фигура 2. Относительный уровень транскрипции генов CPK, CYP450, CCOAOMT1, SGS3, ERD3, DHN1, DHN7 и LEA в хвое сеянцев сосны обыкновенной при отсутствии стресса (2 часа после полива) и при сильном стрессе (10 дней после полива). Значения экспрессии гена рассчитывались относительно уровня экспрессии гена Actin (ACT).
Сущность изобретения.
Техническим результатом является создание набора синтетических олигонуклеотидов, позволяющего с высокой эффективностью и специфичностью определять уровень экспрессии генов DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT сосны обыкновенной методом ПЦР в режиме реального времени. Набор включает прямые праймеры (F), обратные праймеры (R) и зонды (Z) для каждого гена, имеющие следующие нуклеотидные последовательности:
1) ген DHN1(dehydrin 1):
DHN1_F: GGAAGTGGAGAAGAAAGTAGGTT
DHN1_R: CCGGGGAGTTTCTCTTTGATTTTA
DHN1_Z: FAM-TAACAAGAAGGAAGGGGAAGGGAAAGAAGAAGAAG-BHQ1
2) ген DHN7 (dehydrin 7):
DHN7_F: GGCAAAAAGAAGGAAGATGAGGG
DHN7_R: GGCAGCTTCTGTTTGATTTTATCAAC
DHN7_Z: FAM-TTCTTCCCCTCTTCACATCTACCACCCTCAGT-BHQ1
3) SGS3 (suppressor of gene silencing):
SGS3_F: ATCGGAGCTTGGAAGGGGA
SGS3_R: CTTGGTTTCCATTCCTCTGTTG
SGS3_Z: FAM-TAAGGAACGGGGATGGAAGGAGTCAGG-BHQ1
4) CPK (calcium-dependent protein kinase):
CPK_F: CTACAAAGCAGCATCAGCAGTG
CPK_R: CGATGGTCTTGCACGCGAAC
CPK_Z: FAM-TCGGGCGTGGGCAGTTCGGCGTA-BHQ1
5) CYP450 (cytochrome P450):
CYP450_F: ATTTGGATTGGCTCGATTT
CYP450_R: GATGGTTTCCCGTGTAAT
CYP450_Z: FAM-TGCGGAGAAAATAATAGACGACCA-BHQ1
6) ERD3 (early responsive to dehydration 3):
ERD3_F: GTTGATGGTCTTTTCAGTGCAG
ERD3_R: CTGCGGCTAGATTCTTGACA
ERD3_Z: FAM-TGTGTTGTTGGAAATGGACCGTATATTAAGACCTG-BHQ1
7) CCOAOMT1 (caffeoyl-CoA-O-methyltransferase 1):
CCOAOMT1_F: TGCAGTATATATTGGAAACGAGC
CCOAOMT1_R: TTCTTGGCGTTAATGAGCTTC
CCOAOMT1_Z: FAM-TACTTCTGCCGATGAGGGTCAATTTCTGGG-BHQ1
8) LEA (late embryogenesis abundant protein):
LEA_F: TATGATCATGACCTACCAATTGGG
LEA_R: CGTAAGGCACTGTCACTGGAA
LEA_Z: FAM-TCTGGAACAATTGTAGATCCTGGGTCAGTGAAGG-BHQ1
9) ACT (actin):
ACT_F: TGAGCTTCGAGTTGCTCCAG
ACT_R: CCAGTTGTACGACCACTTGC
ACT_Z: FAM-TCCCAAGGCAAACAGAGAGAAGATGACTCAG-BHQ1
Определение уровня экспрессии осуществляется известным способом за счет использования флуоресцентномеченного зонда, позволяющего проводить контроль динамики накопления продукта полимеразной цепной реакции в режиме реального времени. Флуоресцентномеченный зонд для ПЦР в режиме реального времени является синтетическим олигонуклеотидом, к которому присоединены молекула флуорофора (например, молекула карбоксифлуоресцеин (FAM)) и молекула гасителя флуоресценции (например, молекула Black Hole Quenchers (BHQ1)).
Созданный набор синтетических олигонуклеотидов позволяет эффективно и с высокой специфичностью проводить оценку уровня экспрессии генов DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT методом ПЦР в реальном времени. Высокая специфичность амплификации целевого участка ДНК обеспечивается увеличением суммарной протяженности комплементарных к целевому участку синтетических олигонуклеотидов до 61-82 пар оснований, по сравнению с детекцией результатов ПЦР на основе интеркалирующего красителя SYBR Green, когда применяются два праймера, общей протяженностью около 40-50 пар оснований.
Способ осуществления изобретения.
Созданный набор синтетических олгонуклеотидов применяется следующим образом.
1. Сбор образцов тканей сосны обыкновенной. В качестве исходного материала для изучения экспрессии генов могут служить хвоя, почки и корни.
2. Выделение суммарной РНК из растительной ткани с использованием одного из известных способов. Для выделения РНК могут быть использованы классические жидкофазные методы, включающие экстракцию нуклеиновых кислот с помощью лизирующего СТАВ-буфера и очистку хлороформом. Также для получения препаратов РНК могут быть использованы коммерческие наборы реагентов и колонок.
3. Получение комплементарной ДНК (кДНК) с помощью реакции обратной транскрипции с использованием универсального oligo(dT) праймера с применением стандартных протоколов. Реакцию обратной транскрипции проводят с применением коммерческих наборов, включающих препарат фермента обратной транскриптазы (например, обратная транскриптаза вируса лейкемии мышей, MMLV ревертаза).
4. Постановка ПЦР в режиме реального времени с использованием полученных препаратов кДНК и созданного набора синтетических олигонуклеотидов. ПЦР в режиме реального времени проводят с применением специализированного оборудования (термоциклера для ПЦР в реальном времени) в соответствие со следующим температурным режимом:
1) для олигонуклеотидов специфичных к генам SGS3, CPK, CYP450, CCOAOMT1 и ACT: предварительная денатурация при 95°С - 10 мин; 45 циклов: денатурация 95°С - 15 сек, отжиг 57°С - 30 сек, элонгация 72°С - 30 сек+детекция сигнала флуоресценции;
2) для олигонуклеотидов специфичных к генам DHN1, DHN7 и ERD3: предварительная денатурация при 95°С - 10 мин; 45 циклов: денатурация 95°С - 15 сек, отжиг 61(- 30 сек, элонгация 72°С - 30 сек+детекция сигнала флуоресценции.
5. Анализ результатов ПЦР в режиме реального времени и оценка изменения уровня экспрессии генов DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1 и ACT по сравнению с геном АСТ, относящийся к так называемым генам «домашнего хозяйства» («housekeeping genes») уровень экспрессии которых относительно постоянен. Для количественной оценки содержания матрицы в пробе при ПЦР-РВ используется показатель Ct - цикл полимеразной цепной реакции при котором уровень флуоресценции превысил пороговое значение.
Пример использования изобретения.
Изобретение использовано для изучения изменчивости уровня экспрессии генов DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1 и LEA у сеянцев сосны обыкновенной под воздействием водного стресса. В качестве референсного гена для расчета относительной экспрессии использовали ген Actin (АСТ). Сеянцы сосны обыкновенной с закрытой корневой системой подвергали воздействию засухи в течение длительного времени и через определенный временные отрезки брали образцы хвои для выделения суммарной РНК. Суммарная РНК выделена из хвои в соответствии с протоколом, предложенным М. Су и др. [Су М., Цзан В., Яо Н., Хуан М. Выделение высококачественной РНК из различных тканей Populus. Физиология растений. 2009. Т.56. №5. С.791-795.].
Для синтеза одноцепочной кДНК использован коммерческий набор «MMLV RT kit» (Евроген, Россия) и протокол компании-производителя:
1) для каждого образца РНК приготовили смесь №1 в пробирках для ПЦР, общий объем 9 мкл:
2 мкл - раствора РНК (1-2 мкг РНК)
1 мкл - oligo(dT) праймер
6 мкл - деионизированная вода
Инкубировали пробирки 2 мин при+70°C;
2) приготовили смесь №2 (указаны объемы для 1 образца РНК, общий объем 11 мкл):
4 мкл - 5X буфер для синтеза первой цепи кДНК (first-strand buffer)
2 мкл - DTT (20 мМ)
2 мкл - смесь dNTP (10 мМ)
1 мкл - MMLV ревертаза
2 мкл - деионизированная вода
3) добавили смесь №2 к смеси №1, перемешали и инкубировали 60 мин при 42°C.
4) для остановки реакции прогрели смесь 10 мин при 70°C, затем помещали на лед.
Для всех образцов кДНК в трехкратной повторности последовательно ставили ПЦР-РВ со специфическими олигонуклеотидами при следующих температурных режимах:
1) для олигонуклеотидов специфичных к генам SGS3, CPK, CYP450, CCOAOMT1 и ACT: предварительная денатурация при 95°С - 10 мин; 45 циклов: денатурация 95°С - 15 сек, отжиг 57°С - 30 сек, элонгация 72°С - 30 сек+детекция сигнала флуоресценции;
2) для олигонуклеотидов специфичных к генам DHN1, DHN7, ERD3 и LEA: предварительная денатурация при 95°С - 10 мин; 45 циклов: денатурация 95°С - 15 сек, отжиг 61(- 30 сек, элонгация 72°С - 30 сек+детекция сигнала флуоресценции.
Для проведения ПЦР-РВ использована готовая смесь для ПЦР «qPCRmix-HS» (Евроген, Россия) и термоциклер СFX96 Touch Thermal Cycler (Bio-Rad, США). Реакционная смесь для ПЦР-РВ, общим объемом 20 мкл, содержала: qPCRmix-HS - 4 мкл, кДНК - 2,0 мкл, прямой и обратный праймеры - по 0,3 мкл, флуоресцентный зонд (краситель FAM) - 0,3 мкл, деионизированная вода - 13,1 мкл.
После завершения реакции амплификации данные сохраняются в виде графиков, иллюстрирующих скорость накопления продуктов амплификации (фиг.1) и значений Ct (пороговый цикл), определенных в программе «CFX Manager». Значения Ct переносили в Microsoft Excel и выполняли математическую обработку данных. Относительный уровень транскрипции генов (R) вычисляли по формуле (1):
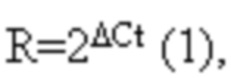 где
где
где ΔCt - разница значений пороговых циклов для референсного и целевого генов.
Уровень транскрипции генов выражался в относительных единицах (отн. ед.) (фиг.2).
Результаты. В результате изучения изменчивости экспрессии генов в хвое сеянцев сосны обыкновенной, выявлено 4,6-9,1 кратное увеличение уровня транскрипции генов DHN1, DHN7 и LEA -кратные под воздействием водного стресса.
Положительным эффектом предлагаемого изобретения является создание уникального набора синтетических олигонуклеотидов для изучения уровня экспрессии генов (DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT), обеспечивающего высокую надежность и снижение риска неспецифической амплификации ДНК во время полимеразной цепной реакции за счет использования флуоресцентномеченных зондов, что приводит к увеличения суммарной протяженности синтетических нуклеотидов.
Работа выполнена с использованием ресурсов ЦКП «Экология, биотехнологии и процессы получения экологически чистых энергоносителей» Поволжского государственного технологического университета, г. Йошкар-Ола при финансовой поддержке Министерства науки и высшего образования РФ (соглашение №075-15-2021-674 от 28.07.21).
Ручное устройство для образования лунок под посадку контейнеризированных сеянцев
Ручной бур для образования посадочных мест
Способ хранения желудей
Способ выработки окоренных сортиментов и рабочий орган для его осуществления
Транспортно-технологическая платформа со смещаемым центром тяжести
Захватно-срезающее устройство лесозаготовительной машины
Блок формирования раздражающих сигналов для сбора пчелиного яда
Прицепное устройство для образования посадочных лунок с фиксированным шагом посадки
Машина для комбинированной трелевки лесоматериалов
Судно-трансформер

