Результат интеллектуальной деятельности: Способ получения рекомбинантного антимикробного пептида UBI18-35, рекомбинантная плазмидная ДНК pET31b-2хUBI18-35 и штамм-продуцент Escherichia coli BL21 Rosetta DE3 pLysS/ pET31b-2хUBI18-35 антимикробного пептида UBI18-35
Вид РИД
Изобретение
Изобретение относится к биотехнологии, генетической и белковой инженерии, конкретно - к получению рекомбинантного антимикробного пептида UBI18-35, способного проникать в мембрану микробных клеток, накапливаться в очаге гнойного воспаления и предназначенного для дифференциальной диагностики бактериального воспаления. Для получения рекомбинантного антимикробного пептида UBI18-35 конструируют рекомбинантную плазмидную ДНК рЕТ31b-2xUBI18-35 размером 5858 п.н., кодирующую рекомбинантный пептид UBI18-35. В состав плазмидной ДНК также входит промотор транскрипции Т7 РНК-полимеразы, участок связывания рибосомы; фрагмент ДНК плазмиды ген β-лактамазы, детерминирующий устойчивость клеток Escherichia coli к ампициллину, в качестве генетического маркера, ген кетостероидизомеразы. Полученной плазмидной ДНК трансформируют клетки штамма Escherichia coli BL21 Rosetta DE3 pLysS с получением штамма-продуцента рекомбинантного антимикробного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой. Для получения рекомбинантного пептида UBI18-35 проводят культивирование штамма-продуцента, клетки разрушают, выделяют тельца включения, тельца включения солюбилизируют в буфере, содержащем мочевину, белок слияния выделяют методом металл-аффинной хроматографии, проводят отделение рекомбинантного пептида от кетостероидизомеразы и шести гистидиновых остатков с использованием бромциана, и очищают рекомбинантный пептид методом гельфильтрационной хроматографии.
Получение векторных молекул с целью создания систем для диагностики и адресной терапии социально-значимых заболеваний крайне востребовано в современной биомедицине. В настоящее время пептиды (молекулы белковой природы размером от 3 до 50 а.к.о.) широко используются в диагностических целях. Они обеспечивают высокоспецифичное связывание с клетками-мишенями, но имеют меньшие размеры по сравнению с белками и низкую иммуногенность. Таким образом, пептиды являются перспективными объектами современной молекулярной медицины [Shadidi, М., 2003. Identification of novel carrier peptides for the specific delivery of therapeutics into cancer cells. FASEB J. 17, 256-258, Патент WO 9851794 (A1), 19.11.1998].
Антимикробные пептиды (АМП) были обнаружены в различных организмах; существуют многочисленные примеры АМП, найденных в бактериях (низин, педиоцин РА-1 и писциколин 126), растениях (дефензин и снакин-1) и в животных (магаинин, дефензин и протегрин). АМП также обнаружены в одноклеточных организмах и грибах [Ingham, А.В., 2007. Recombinant production of antimicrobial peptides in heterologous microbial systems. Biotechnol Appl Biochem. 47, 1-9].
Существуют две основные генетические группы антимикробных пептидов млекопитающих: кателицидины и дефензины [Selsted, М.Е., 2005. Mammalian defensins in the antimicrobial immune response. Nat Immunol. 6(6), 551-7]. Все они вырабатываются и функционируют как часть иммунной защиты организма.
Ключевым преимуществом антимикробных пептидов является высокая специфичность и способность связываться только с живыми бактериями [Akhtar, М.S., 2008. An imaging analysis of (99m)Tc-UBI (29-41) uptake in S. aureus infected thighs of rabbits on ciprofloxacin treatment. Eur J Nucl Med Mol Imaging 35, 1056-1064]. Благодаря способности пептидов встраиваться в мембрану микробной клетки, достигается высокоселективное накопление пептидов в очаге гнойного воспаления, тогда как накопления в стерильном воспалении не происходит [Amblard, M., 2006. Methods and protocols of modern solid phase Peptide synthesis. Molecular biotechnology 33, 239-254].
Первый антимикробный пептид убиквицидин (UBI), относящийся к классу дефензинов, был выделен из макрофагов мыши, позже идентичный пептид был обнаружен в эпителиальных клетках дыхательных путей человека. Меченные 99mTc производные (фрагменты) антимикробного пептида убиквицидина, в частности, UBI29-41, на первой стадии клинических испытаний продемонстрировал 100% общей чувствительности, 80 % специфичности и 94,4% точности среди пациентов с инфекциями мягких тканей и остеомиелитом, при этом оптимальное время формирования изображения составило 30 минут после внутривенного введения радиофармпрепарата [Akhtar, M.S., 2005. Antimicrobial Peptide 99mTc-Ubiquicidin 29-41 as Human Infection-Imaging Agent: Clinical Trial. J. Nucl. Med. 46 (4), 567-573]. Исследование специфичности детектирования стерильного воспаления и бактериальной инфекции с использованием 99mTc-меченных синтетических антимикробных производных убиквицидина (UBI 29–41, 18–35, 31–38), синтетического hLf 1–11, нейтрофильных пептидов человека (дефензинов), по сравнению с применением 99mTc-ципрофлоксацина (Infecton©), показало эффективное аккумулирование антимикробных пептидов в очаге инфекции, что указывает на преимущественное связывание этих пептидов не с активированными лейкоцитами, а с живыми микроорганизмами. При этом 99mTc-ципрофлоксацин аккумулировался как в очагах стерильного воспаления, так и бактериальной инфекции [Welling, M., 2001. 99mTc-labeled antimicrobial peptides for detection of bacterial and Candida albicans infections. J. Nucl. Med. 42, 788-794].
Группа пептидов, полученных из убиквицидина, представляет большой интерес не только для дифференциальной диагностики микробного воспаления, но также для мониторинга эффективности антимикробной терапии. Показана эффективность использования радиоактивно меченых пептидов - производных UBI, у пациентов с остеомиелитом, септическим артритом, бациллемией, медиастинитом после операции на сердце, инфекцией суставов после протезирования, вызванных различными видами патогенных микроорганизмов [Sasser, T.A., 2013. Bacterial infection probes and imaging strategies in clinical nuclear medicine and preclinical molecular imaging. Curr Top Med Chem. 13, 479-487].
Наиболее перспективными пептидами производными убиквицидина считаются фрагменты UBI29-41 и UBI18-35, которые позволяют эффективно визуализировать очаг инфекции при отсутствии неспецифического накопления [Welling, M., 2001. 99mTc-labeled antimicrobial peptides for detection of bacterial and Candida albicans infections. J. Nucl. Med. 42, 788-794]. Пептид UBI18-35 демонстрирует более высокую селективность связывания с микробными клетками Staphylococcus aureus и Candida albicans в очаге воспаления, поэтому его применение по сравнению с UBI29-41 является более перспективным. Но, несмотря на это, пептид UBI18-35 применяется реже, что обусловлено большей сложностью и стоимостью твердофазного синтеза пептида длинной 18 а.к.о. по сравнению с UBI29-41, имеющим длину 12 а.к.о. [Welling, M. M., 2000. Technetium-99m labelled antimicrobial peptides discriminate between bacterial infections and sterile inflammations. Eur J Nucl Med. 27, 292-301]. Антимикробные пептиды могут быть выделены из природных источников (Патент WO2017176041 (A1), 12.10.2017; патент WO9851794 (A1), 19.11.1998). Наиболее широко для получения пептидов используется метод химического синтеза (Патент RU 2624020 , 20.03.2016).
Пептид убиквицидин может быть получен методом твердофазного синтеза или с использованием трансгенных животных (Патент WO 1998054314 A1 Antimicrobial peptides derived from ubiquicidine 3.12.1998).
Процесс трансгенеза животных включает выделение зародышевых клеток (зиготы, эмбриональных стволовых клеток, клеток раннего эмбриона), перенос в них чужеродного фрагмента ДНК одним из стандартных способов (эндоцитоз, электропорация, микроинъекция, бомбардировка микрочастицами, нагруженными рекомбинантной ДНК, инфецирование рекомбинантными вирусами). Далее генетически модифицированные зародышевые клетки животного переносят в матку приемной матери для получения трансгенного организма. Далее пептид или его фрагменты выделяют из тканей или биологических жидкостей организма, например молока. К недостатку метода можно отнести высокую трудоемкость, длительность, высокую стоимость процедуры трансгенеза с последующим отбор (скрининг) трансгенных организмов, а также сравнительно низкие выходы пептида [Maksimenko O.G., 2013. Use of Transgenic Animals in Biotechnology: Prospects and Problems Acta Naturae, 5(1), 33-46].
Для получения синтетического пептида наиболее широко в настоящее время используется твердофазный синтез. Твердофазный синтез начинается со связывания карбоксильной группы защищенной аминокислоты пептида с соответствующей смолой, обычно через сложноэфирную или амидную связь. Проводят защиту групп боковых цепей аминокислот с помощью различных химических реагентов. Далее следуют повторяющиеся циклы наращивания аминокислотной последовательности, включающие: деблокирование аминогруппы промежуточно-образующейся N-терминально защищенной пептидной смолы, активация карбоксильной группы следующей за ней NH2-защищенной аминокислоты, сочетание аминокислот с формированием пептидной связи. В качестве сочетающего реагента применяют различные используемые в синтезе пептидов активирующие реагенты, например, карбодиимиды. Активированное аминокислотное производное обычно используется в 1,5-4-х кратном избытке. В случаях если происходит неполное сочетание, реакцию повторяют. После получения полноразмерной последовательности пептида проводят финальное деблокирование, снятие защиты с боковых групп аминокислот и отщепление готового пептида от смолы, например, с помощью трифторуксусной кислоты. Пептидный синтез широко используется для структурно-функциональных исследований, для получения аналогов биологически активных пептидов, в том числе циклических производных с заданными свойствами (например, с пролонгированным, усиленным или избирательным действием), а также аналогов с остатками небелковых аминокислот, для изучения антигенных свойств фрагментов белков и получения специфичных к отдельным участкам полипептидных цепей антител, используемых в структурно-функциональном анализе и в создании диагностических систем и вакцин [Овчинников, Ю.А., 1987. Биоорганическая химия. Просвещение. Москва]. Однако, получение необходимого количества пептида, особенно гидрофобного и/или с длинной последовательности более 15 а.к.о. методом твердофазного синтеза имеет ряд ограничений [Amblard, M., 2006. Methods and protocols of modern solid phase Peptide synthesis. Mol Biotechnol. 33, 239-254; Guzmán, F.S., 2007. Peptide synthesis: chemical or enzymatic. Electronic Journal of Biotechnology. 10, 279-314], а именно: возникновение ошибочных последовательностей, пропуск аминокислот из-за неполноты протекания стадий деблокирования и аминообразования; самоассоциация пептидных цепей за счет формирования водородных связей, вызывающая экранирование N-концевой аминогруппы; а также высокая стоимость синтеза. Дополнительным негативным фактором химического синтеза является применение в работе токсичных растворителей и реагентов. Таким образом, получение антимикробных пептидов в достаточных количествах для клинического применения методом твердофазного синтеза является высоко затратным.
Для масштабирования процесса наработки пептида с целью создания диагностических систем выгодной альтернативой является микробный синтез с использованием технологии рекомбинантных ДНК.
Экспрессия гетерологичных белков в бактериальной системе в настоящее время представляется наиболее простым и недорогим способом получения больших количеств белка. Однако при экспрессии антимикробных пептидов возникают некоторые трудности, связанные с риском протеолитической деградации пептида и его токсичностью по отношению к клетке-хозяину [Zorko, M., 2010. Production of Recombinant Antimicrobial Peptides in Bacteria // Antimicrobial Peptides. Methods in Molecular Biology. 618, 61-76]. Для преодоления этих проблем экспрессия АМП осуществляется в составе с белками-партнерами. Таким образом, за последнее десятилетие рекомбинантным путем был получен ряд АМП с различными белками-партнерами, например, морицин (moricin), который экспрессировали в составе белка-слияния с мальтозо-связывающим белком [Hara, S., 1996. Production in Escherichia coli of moricin, a novel type antibacterial peptide from the silkworm. Bombyx mori. Biochem. Biophys. Res. Commun . 220, 664-669], цекропин X (cecropin X), экспрессированный совместно с фактором некроза опухоли (TNFα) [Shen, Y., 2007. High-level expression of cecropin X in Escherichia coli. Int. J. Mol. Sci. 8, 478-491], человеческий β-дефензин 2 (human beta-defensin 2) в составе тиоредоксина (Trx) [Xu, Z.N., 2006. High-level expression of a soluble functional antimicrobial peptide, human beta-defensin 2, in Escherichia coli. Biotechnol. Progress. 22, 382-386], лактоферрицин, в составе белка-слияния с анионным пептидом [Kim, H. K., 2006. Expression of the cationic antimicrobial peptide lactoferricin fused with the anionic peptide in Escherichia coli. Appl. Microbiol. Biotechnol. 72, 330-338] и пр.
Поскольку антимикробный пептид UBI18-35 проявляет токсичность в отношении клеток E. coli, была выбрана стратегия экспрессии UBI18-35, в составе белка-слияния с кетостероидизомеразой (KSI).
Аминокислотная последовательность фермента KSI была заимствована из бактерии Pseudomonas testosteroni и в дальнейшем модифицирована - пять остатков аспарагиновой кислоты были заменены остатками глутаминовой кислоты - это обеспечило устойчивость белка к кислотному гидролизу, при этом не лишая его способности эффективно индуцировать образование телец включения [Патент US 7829311 B2; 14.07.2008]. Благодаря этому свойству получен ряд различных пептидов в составе белка слияния с KSI, в том числе противоопухолевые пептиды p53pAnt и PNC27 [Rodríguez, V., 2014. Design and implementation of a high yield production system for recombinant expression of peptides. Microb. Cell Fact. 13:65, 1-10], трансмембранный домен нейротрофин рецептора р75 [Li, Q., 2012. Expression and purification of the p75 neurotrophin receptor transmembrane domain using a ketosteroid isomerase tag. Microb. Cell Fact. 11:45, 1-8], пептидные рецепторы аденозина [Britton, Z.T., 2012. An expression and purification system for the biosynthesis of adenosine receptor peptides for biophysical and structural characterization. Protein Expr. Purif. 84(2), 224-35].
За счет гетерогенной экспрессии в составе единого белка с кетостероидизомеразой (KSI) получены два антимикробных пептида: дермцидин (dermcidin) [Cipakova, I., 2006. Expression and purification of human antimicrobial peptide, dermcidin, in Escherichia coli. Protein Expr. Purif. 45, 269-274] и лактоферрицин [Park T-J., 2009. Cloning, expression, isotope labeling, purification, and characterization of bovine antimicrobial peptide, lactophoricin in Escherichia coli // Protein Expr. Purif. 65(1), 23-29].
Известен плазмидный вектор pET31b+, который содержит ген KSI. С использованием вектора pET31b+ получены рекомбинантный пептид rP11-4 (QQRFEWEFEQQ), предназначенный для использования в тканевой инженерии [Riley, J.M., 2009. Bioproduction and characterization of a pH responsive self-assembling peptide. Biotechnol Bioeng. 103, 241-251], и четыре пептида - антагониста αvβ6 интегрина: A20fmdv2 (NAVPNLRGDLQVLAQKVART), A20lap2 (GFTTGRRGDLATIHGLNRPF), DBD1 (EKCPNLRGDLQVLAQKVCRT), TGFb3 (NEDDHQRGDLGRLKKQKDHH), предназначенные для диагностики и целевой доставки к опухолям поджелудочной железы и толстой кишки [Wagstaff, J.L., 2010. Production of recombinant isotopically labelled peptide by fusion to an insoluble partner protein: generation of integrin αvβ6 binding peptides for NMR. Mol Biosyst. 6, 2380-2385].
Технической задачей настоящего изобретения является получение рекомбинантного векторного антимикробного пептида UBI18-35, способного встраиваться в мембрану микробных клеток, предназначенного для дифференциальной диагностики бактериального воспаления. В Российской Федерации и за рубежом рекомбинантные аналоги векторного антимикробного пептида UBI18-35, способного встраиваться в мембрану микробных клеток, а также способы получения пептида UBI18-35 с использованием технологии рекомбинантных ДНК и микробных штаммов-продуцентов, отсутствуют. Применение данного способа синтеза является более приемлемым и экономически выгодным для получения векторного антимикробного пептида UBI18-35 в препаративных количествах.
Поставленная техническая задача достигается конструированием плазмиды pET31b- 2xUBI18-35 путем встраивания в плазмидный вектор pET31b+ фрагмента ДНК, кодирующего две молекулы рекомбинантного антимикробного пептида UBI18-35 разделенного последовательностью, кодирующей метионин, получением штамма-продуцента, за счет трансформации полученной плазмидой клеток Escherichia coli BL21 Rosetta DE3 pLysS, обеспечивающего синтез двух молекул рекомбинантного антимикробного пептида UBI18-35 разделенных остатком метионина в составе белка-слияния с кетостероидизомеразой (KSI) и остатком из шести гистидинов, выделением белка-слияния методом аффинной хроматографии, последующего разделения пептидов между собой, от белка-слияния и остатков из 6 гистидинов с использованием бромциана, очистки пептида UBI18-35 методом гельфильтрационной хроматографии.
Сущность изобретения заключается в следующем.
Генно-инженерными методами [Маниатис, Т.Э., 1984. Молекулярное клонирование. Мир. Москва] получают плазмиду pET31b-2xUBI18-35, содержащую последовательность, кодирующую две молекулы рекомбинантного антимикробного пептида UBI18-35, полученную методом отжига двух синтетических олигонуклеотидных последовательностей c последующей амплификацией методом полимеразной цепной реакции (ПЦР).
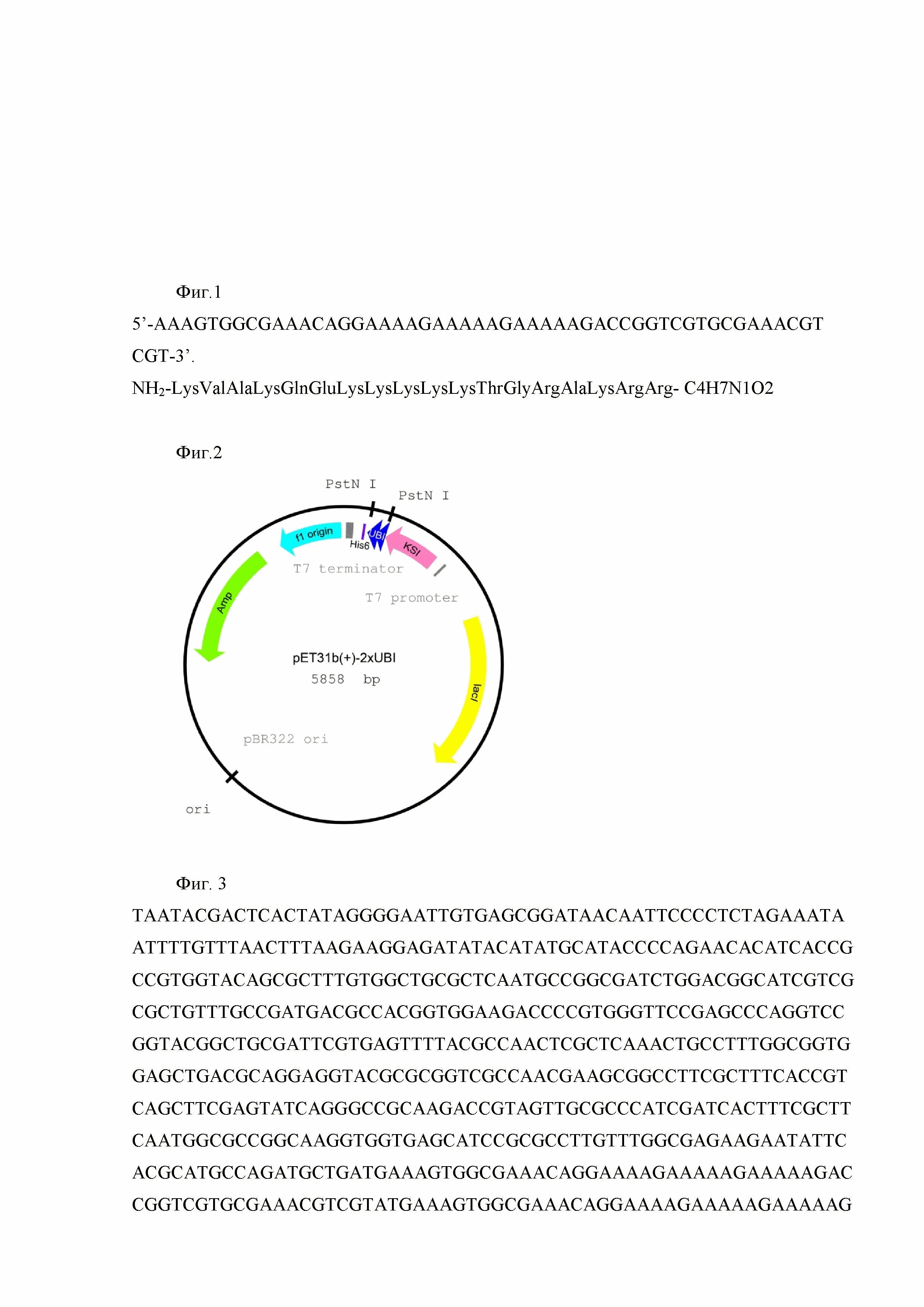
Клетки E.coli BL21 Rosetta DE3 pLysS, содержащие ген РНК-полимеразы фага Т7 под индуцибельным lacUV5 промотором, трансформируют сконструированной плазмидой pET31b-2xUBI18-35 и выращивают в течение ночи. Ночную культуру (1/100) засевают в свежую среду LB с добавлением ампициллина (50 мкг/мл). Синтез РНК-полимеразы индуцируют добавлением изопропилтиогалактазида (ИПТГ) в концентрации 0,1-0,5 мМ в тот момент, когда культура достигает среднелогарифмической фазы роста. Индуцированные клетки растят 4 часа при 28°С, после чего собирают центрифугированием при 5000 g. Индуцированные клетки E.coli BL21 Rosetta DE3 pLysS / pET31b-2xUBI18-35 используют для очистки антимикробного пептида UBI18-35, экспрессированного в составе белка слияния с кетостероидизомеразой и остатком из шести гистидинов. Для получения антимикробного пептида UBI18-35 выделенный химерный белок обрабатывают бромцианом. В результате получают 2 молекулы рекомбинантного антимикробного пептида UBI18-35, имеющего следующую аминокислотную последовательность: NH2-KVAKQEKKKKKTGRAKRR-C4H7N1O2 и молекулярную массу 2,252 кДа, кодируемого нуклеотидной последовательностью 5'- AAAGTGGCGAAACAGGAAAAGAAAAAGAAAAAGACCGGTCGTGCGAAACGTCGT-3' (Фиг. 1).
Учитывая вырожденность генетического кода, известно, что заданная аминокислотная последовательность KVAKQEKKKKKTGRAKRR может быть закодирована различными сочетаниями нуклеотидов. В тоже время, нуклеотидная последовательность кодирующая пептид играет решающую роль для успешной экспрессии и трансляции чужеродного пептида в клетке организма-продуцента, в связи с тем, что может содержать сочетания нуклеотидов распознаваемые клеткой, например, как сайты для действия ферментов гидролиза, рестрикции, модификации, остановки трансляции и проч. В связи с этим дизайн кодирующей пептид нуклеотидной последовательности является критичным для успешной экспрессии продукта и получения рекомбинантного пептида.
Исходным генетическим материалом для конструирования рекомбинантной плазмиды pET31b-2xUBI18-35 являются:
а) фрагмент плазмиды pET31b+ (Novagen), обработанный рестриктазой AlwN I (Pst I), обеспечивающий встраивание фрагмента ДНК, кодирующего две молекулы антимикробного пептида UBI18-35 и его экспрессию в составе белка-слияния с кетостероидизомеразой и остатком из шести гистидинов под контролем позднего промотора Т7 ДНК-полимеразы;
б) фрагмент ДНК, кодирующий две молекулы антимикробного пептида UBI18-35, полученный методом полимеразной цепной реакции с использованием синтетических олигонуклеотидов.
Полученная в результате плазмида pET31b-2xUBI18-35 (Фиг. 2) характеризуется следующими признаками:
- имеет размер 5858 п.н.;
- кодирует рекомбинантный антимикробный пептид UBI18-35 в составе белка слияния с кетостероидизомеразой и остатком из 6 гистидинов.
- состоит из следующих элементов:
1) фрагмент ДНК, размером 111 п.о., кодирующий 2 молекулы антимикробного пептида UBI18-35, разделенных остатком метионина.
2) фрагмент плазмиды pET31b+, обеспечивающий эффективную транскрипцию ДНК, кодирующей две молекулы антимикробного пептид UBI18-35, и их экспрессию.
- содержит:
а) сайт инициации репликации плазмиды pBR322;
б) промотор бактериофага Т7;
в) генетические маркеры: AMPr - ген β-лактамазы определяющий устойчивость E. coli к ампициллину, и lacI - репрессор лактозного оперона E. coli;
г) гены, кодирующие стартовый кодон, кетостероидизомеразу, последовательность из шести гистидиновых остатков, стоп-кадон;
д) сайт связывания с рибосомой;
е) уникальные сайты узнавания рестрикционной эндонуклеазой PstNI (прототип AlwNI);
Таким образом, впервые получена плазмидная ДНК, обеспечивающая продукцию в клетках E. coli рекомбинантного антимикробного пептида UBI18-35, способного встраиваться в мембрану микробных клеток, предназначенного для дифференциальной диагностики бактериального воспаления.
Предлагаемый штамм-продуцент E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 характеризуется следующими признаками:
Морфологические признаки. Клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаризованной LB-среде - колонии круглые, гладкие, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или LB-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки. Клетки растут при температуре от 4°C до 40°C при оптимуме рН от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к пенициллиновым антибиотикам (до 500 мкг/мл).
Штамм-продуцент E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 отличается от штамма-реципиента E. coli BL21 Rosetta DE3 pLysS только наличием рекомбинантной плазмидной ДНК pET31b-2xUBI18-35, которая придает ему устойчивость к пенициллиновым антибиотикам.
Штамм-продуцент получают путем трансформации компетентных клеток E.coli BL21 Rosetta DE3 pLysS соответствующей рекомбинантной плазмидной ДНК.
Клетки E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 являются продуцентом антимикробного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой. При индукции изопропилтио-D-галактозидом, а также и без индукции происходит биосинтез UBI18-35 в составе белка-слияния с кетостероидизомеразой, который накапливается в нерастворимой форме в тельцах включения в клетках в количестве более 10% суммарного белка бактерий.
Пример 1. Конструирование плазмиды pET31b-2xUBI18-35.
Для получения гена UBI18-35, содержащего удвоенную последовательность, кодирующую UBI18-35 разделенную между собой и ограниченную с обоих концов остатками метионина, используют два синтетических олигонуклеотидных праймера длинной по 81 н.о.:
Прямой праймер
5'-tcaccagatgctgatgaaagtggcgaaacaggaaaagaaaaagaaaaagaccggtcgtgcgaaacgtcgtatgaaagtggc-3'
Обратный праймер
5'-gacacagcatctgcatacgacgtttcgcacgaccggtctttttctttttcttttcctgtttcgccactttcatacgacgtt-3'.
Последовательности обеспечивающих наличие в амплифицированном фрагменте сайтов рестрикции PstN1 выделены подчеркиванием, области перекрывания праймеров выделены двойным подчеркиванием, кодоны метионина выделены жирным шрифтом.
Готовят реакционную смесь содержащую: по 0,4 мкМ каждого праймера, смесь dNTP по 0,2 мМ каждого, 0,5 е.а. Encyclo полимеразы в 1× Encyclo ПЦР буфере (Евроген, Россия).
Реакцию проводят в амплификаторе (ThermoCycler, Applied Biosystems) по следующей программе: начальная денатурация 95°С - 1 мин и далее 15 циклов - денатурация 95°С - 15 сек, отжиг 60°С - 3 мин, элонгация 72°С - 30 сек, финальная элонгация 72°С - 1 мин.
Продукт ПЦР разделяют в 1,2 % агарозном геле с использованием 1× ТАЕ буфера с добавлением красителя бромистого этидия. Электрофорез проводят при постоянном электрическом токе 200 мА, напряжении 150 Вольт, мощности 1 Вт/см2.
Выделяют из геля фрагмент массой 143 п.н., соответствующий нуклеотидной последовательности:
5'- TCACCAGATGCTGATGAAAGTGGCGAAACAGGAAAAGAAAAAGAAAAAGACCGGTCGTG CGAAACGTCGTATGAAAGTGGCGAAACAGGAAAAGAAAAAGAAAAAGACCGGTCGTGCG AAACGTCGTATGCAGATGCTGTGTC-3'
с использованием набора Cleanup Standard (Евроген, Россия).
Выделенный фрагмент и плазмидный вектор pET31b+ в отдельных пробирках гидролизуют рестриктазой PstN1 (Сибэнзим, Россия). Рестрикционную смесь разделяют в 1,2 % агарозном геле и выделяют из геля линеаризованный в результате гидролиза плазмидный вектор и фрагмент массой 126 п.н., соответствующий удвоенной нуклеотидной последовательности кодирующей 2 молекулы антимикробного пептида UBI18-35, разделенных между собой и ограниченных с обоих концов остатками метионина.
Выделенный линеаризованный плазмидный вектор pET31b+ дефосфорилируют с использованием термостабильной щелочной фосфатазы (СибЭнзим, Россия). Очищают линеаризованный вектор с использованием набора Cleanup Standard (Евроген, Россия), смешивают с выделенным фрагментом в соотношении вектор : вставка равным 1:10 и лигируют с использованием Т4 ДНК-лигазы (СибЭнзим, Россия). Полученной лигазной смесью трансформируют клетки E.coli XL blue. Трансформированные колонии отбирают на селективной агаризованной среде LB с добавлением ампициллина (50 мг/мл) и выделяют плазмиду, используя набор MiniPrep (Евроген, Россия). Успешность клонирования фрагмента подтверждают методом ПЦР с праймерами на Т7 промоторную и Т7 терминирующую области плазмиды. Подтверждение соответствия клонированной последовательности ожидаемой проводят секвенированием на автоматическом секвенаторе (ABI 3130XL, Applied Biosystems) (Фиг. 3).
Пример 2. Получение штамма-продуцента продуцент E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35
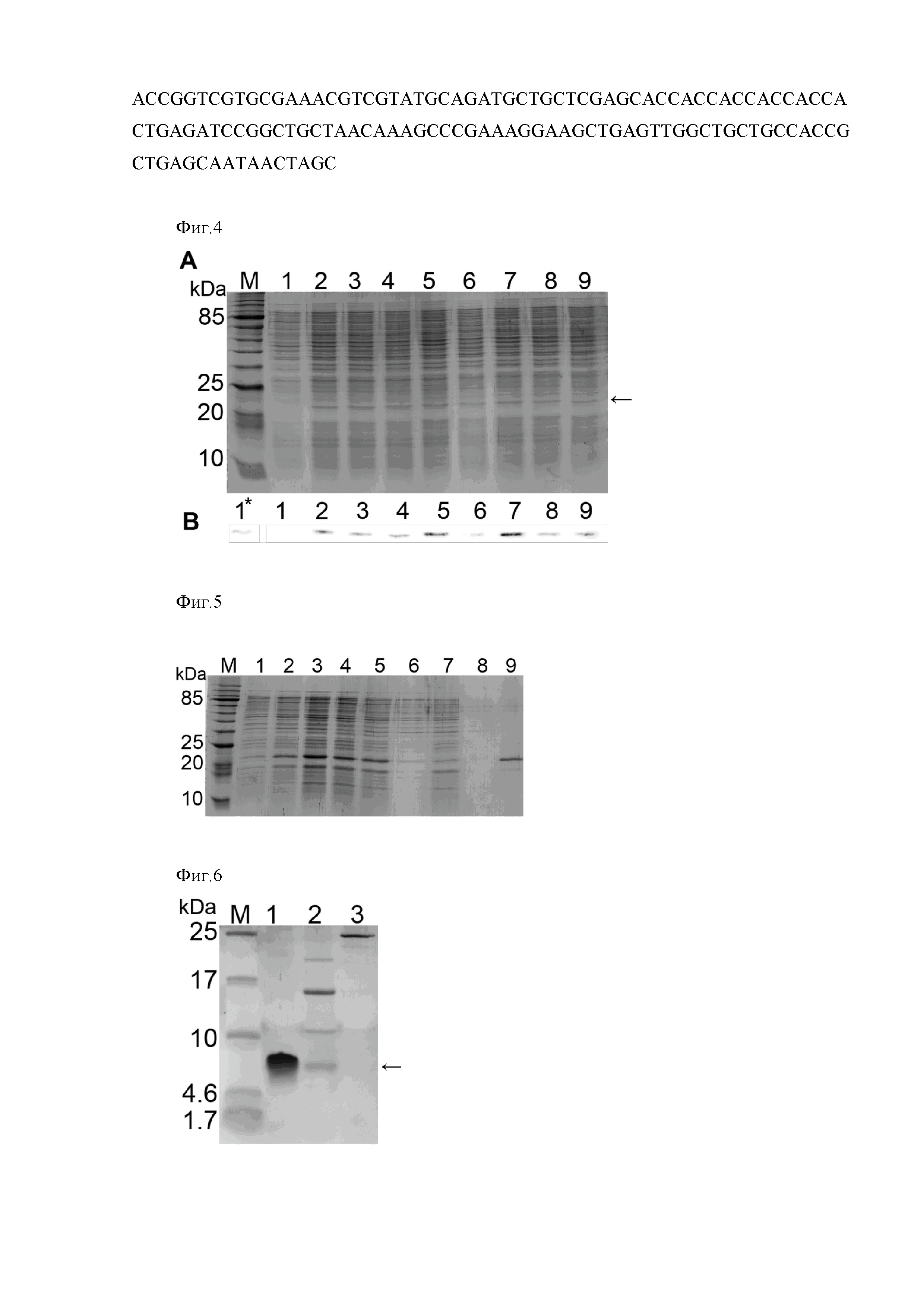
Клетки E.coli BL21 Rosetta DE3 pLysS, несущие ген РНК-полимеразы фага Т7 под индуцибельным lacUV5 промотором, трансформируют [Маниатис, Т.Э., 1984. Молекулярное клонирование. Мир. Москва] сконструированной плазмидой pET31b-2xUBI18-35. Клетки E.coli BL21 Rosetta DE3 pLysS, трансформированные плазмидой pET31b-2xUBI18-35, выращивают в течение ночи. Ночную культуру (1/100) засевают в свежую среду LB с добавлением ампициллина (50 мкг/мл). Синтез РНК-полимеразы индуцируют добавлением ИПТГ в концентрации 0,1-1 мМ, предпочтительно 0,5 мМ, в тот момент, когда культура достигает среднелогарифмической фазы роста. Индуцированные клетки растят в течение 4 часов при температуре 22-37°С, предпочтительно 28°С, после чего собирают центрифугированием при 5000 g и анализируют методом электрофореза по Лэммли [Laemmli U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 227, 680-685] в 16% SDS-полиакриламидном геле (ПААГ) для подтверждения успешности индукции синтеза UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом (Фиг. 4).
Результаты данного анализа, показывают наличие в индуцированной культуре клеток E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 дополнительного белка с молекулярной массой около 20 кДа (Фиг. 4, дорожки 2-9), который отсутствует в контрольном лизате клеток E.coli BL21 Rosetta DE3 pLysS/ pET31b (Фиг. 4, дорожка 1).
Пример 3. Очистка рекомбинантного антимикробного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом из клеток E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 методом металл-аффинной хроматографии.
Клетки E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 выращивают в течение ночи. Ночную культуру (1/100) засевают в свежую среду LB с добавлением ампициллина (50 мкг/мл), экспрессию пептида индуцируют добавлением ИПТГ до конечной концентрации 0,1-0,5 мМ в тот момент, когда культура достигает среднелогарифмической фазы роста. Индуцированные клетки растят в течение 4-16 часов при 28°С, после чего собирают центрифугированием при 5000 g 4°С в течение 20 минут. Осадок замораживают -80°С для повышения эффективности лизиса клеток. Осадок растворяют в лизирующем буфере (50 mM Tris, , 100 mM NaCl, pH 8.0.) из расчета 5 мл буфера на 1 г осадка и добавляют лизоцим до конечной концентрации 1 мг/мл. Клетки ресуспендируют и обрабатывают ультразвуком, импульсами по 30 секунд с перерывом 30 сек во льду, 5 циклов. Инкубируют во льду 30 минут, центрифугируют 20 мин 10000 g при 4°С. Осадок растворяют в промывочном буфере 40 mM Tris, 500 mM NaCl, 15 mM имидазол, 8 M мочевина, pH 8.0 из расчета 10 мл на 1 г осадка и центрифугируют 20 мин 10000 g при 4°С. Отбирают супернатант, содержащий рекомбинантный белок и очищают с помощью аффинной хроматографии на Ni-NTA агарозе (Qiagen, США), согласно инструкции производителя. На хроматографическую колонку, упакованную 5 мл Ni-NTA агарозы и уравновешенную буфером 40 mM Tris, 500 mM NaCl, 15 mM имидазол, 8 M мочевина, pH 8.0, наносят 10 мл супернатанта со скоростью потока 1 мл/мин. Для удаления неспецифически сорбирующихся белков E.coli проводят предварительную элюцию 40 мл буфера, содержащего 40 mM Tris, 500 mM NaCl, 15 mM имидазол, 8 M мочевина, pH 8.0. Пептид в составе белка-слияния элюируют 5 мл буфера, содержащего 40 mM Tris, 500 mM NaCl, 300 mM имидазол, 8 M мочевина, pH 8.0.
Полученные белковые фракции, содержащие рекомбинантный UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом, диализируют против дистиллированной воды (две смены по 24 ч при комнатной температуре), анализируют электрофорезом в ПААГ с SDS по Лэммли и лиофилизируют в течение суток (Freezone Labconco). На Фиг. 5 приведены результаты электрофоретического анализа очищенного методом металл-аффиннной хроматографии пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и последовательностью из шести гистидиновых аминокислотных остатков.
Пример 4. Отделение рекомбинантного антимикробного пептида UBI18-35 от белка партнера кетостероидизомеразы и 6-гистидинового тага
Водный раствор белка-слияния KSI-2xUBI18-35 полученный после диализа, предварительно лиофилизируют. Сухой белок растворяют в минимальном количестве 75 % муравьиной кислоты и добавляют 100 молярный (в расчете на количество содержащихся в белке остатков метионина) избыток бромциана. Реакцию проводят при комнатной температуре в течение 12-24 часов в темноте. Реакцию прекращают добавлением воды в 10-кратном избытке по отношению к исходному объему. Затем пробу диализируют против 1000 кратного избытка дистиллированной воды дважды в течение 24 часов, замораживают при минус 20°С и лиофилизируют. Эффективность отщепления пептида от белка-слияния оценивают методом SDS-ПААГ-трицин [Schägger H., 2006. Tricine-SDS-PAGE. Nat Protoc. 1, 16-22] электрофореза (Фиг. 6).
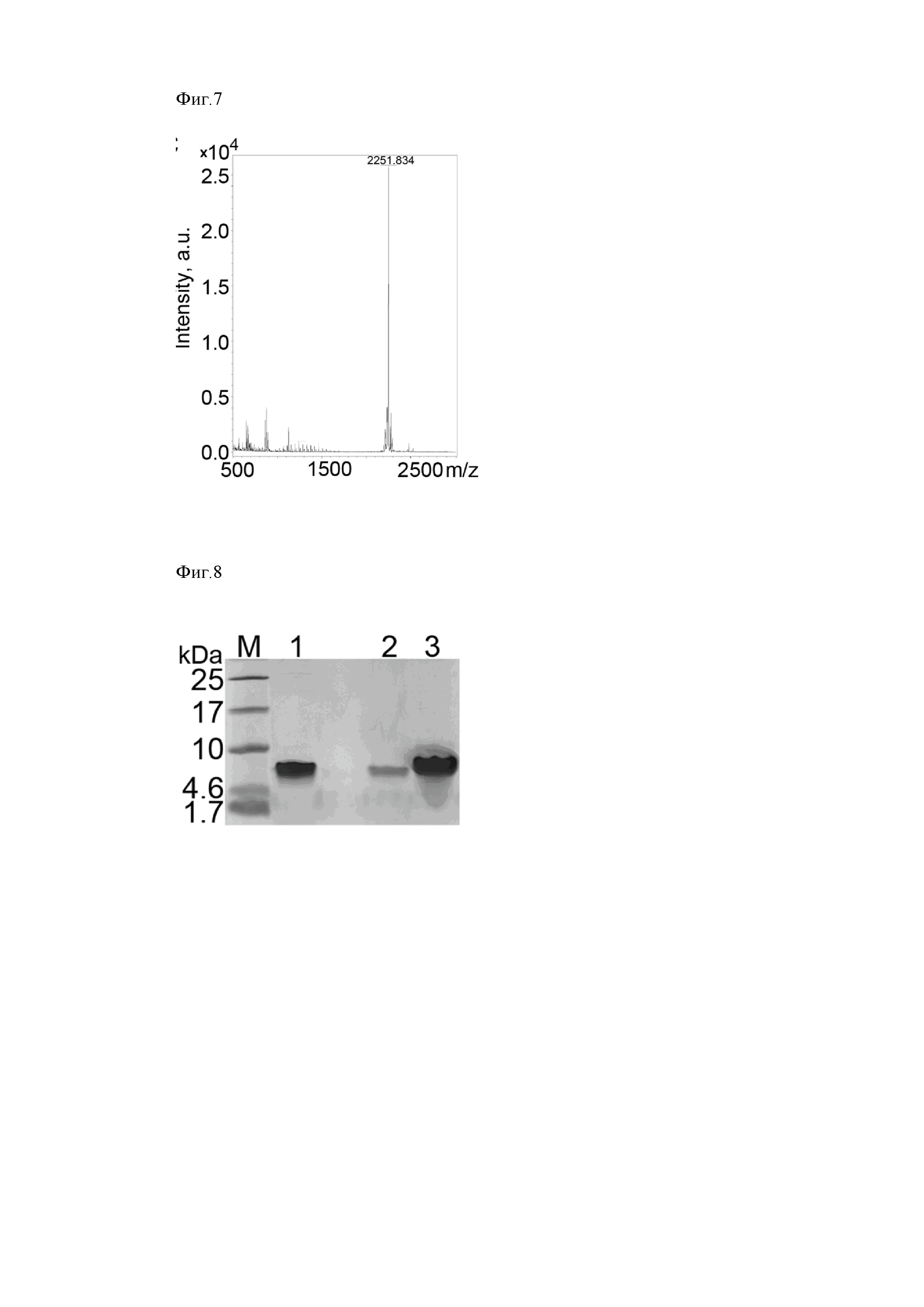
Соответствие аминокислотной последовательности рекомбинантного пептида UBI18-35 заявленной подтверждают методом MALDI-TOF-MS спектрометрии (Фиг. 7). Для проведения масс-спектрометрического анализа образца пептида в качестве матрицы используют 4-гидроксикоричную кислоту. Предварительно высушенную пробу растворяют в 0,1 % растворе трифторуксусной кислоты (ТФУ) в воде, смешивают с рабочим раствором матрицы, растворенной в 0,1 % растворе ТФУ в ацетонитриле, и наносят 2 мкл на подложку из нержавеющей стали и высушивают на воздухе. Масс-спектр получают на приборе MALDI-TOF Brucker MicroFlex mass spectrometer (Bruker, Germany).
На МАЛДИ масс-спектре наблюдается пик целевого пептида массой 2251,8 Да, что соответствует массе пептида UBI18-35 c гомосерин-лактоном на С-конце пептида, образующегося в результате реакции отщепления пептида от белка-слияния по остатку метионина под действием бромциана аминокислотной последовательностью (NH2-KVAKQEKKKKKTGRAKRR-C4H7N1O2) (Фиг. 7).
Пример 5. Получение препарата рекомбинантного антимикробного пептида UBI18-35 из клеток E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35.
Проводят очистку белка по методу, описанному в примере 3. Далее проводят отделение пептида от кетостероидизомеразы и последовательности из 6 гистидиновых аминокислотных остатков по методу, описанному в примере 4.
Очистку пептида UBI18-35 от прочих примесей белковой природы проводят методом гель-фильтрации на колонке с диаметром 30 мм и длиной 145 мм упакованной Bio-Gel P 2. Для этого колонку уравновешивают буфером 50 mM Tris HCl, 100 mM NaCl, 8 M мочевина (pH 8.0) и наносят смесь, содержащую пептид. Элюцию проводят в изократическом режиме при скорости потока 3 мл/мин буфером, содержащем 50 mM Tris HCl, 100 mM NaCl, 8 M мочевина (pH 8.0) в течение 25 минут с использованием хроматографической системы NGC™ Chromatography Systems (Bio-Rad). Фракцию, содержащую пептид, собирают и диализуют против воды используя Slide-A-Lyzer, 2000 MWCO (Pierce).
На Фиг. 8 приведены данные анализа методом трицин-ПААГ электрофореза фракции, собранной на 25-26 минутах хроматографирования. На электрофореграмме в пробе присутствует полоса, соответствующая пептиду UBI18-35, мигрирующего на уровне масс 7 кДа, совпадающая с миграцией синтетического пептида UBI18-35, используемого в качестве контроля.
Пример 6. Анализ способности рекомбинантного антимикробного пептида UBI18-35 полученного в E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 встраиваться в мембрану микробных клеток
Рекомбинантный антимикробный пептид UBI18-35 метят флуоресцентной меткой Cyanine5. Для этого 2 мг лиофилизированного рекомбинантного антимикробного пептида UBI18-35 растворяют в 450 мкл 0,1 M натрий-фосфатного буфера (pH 8,4) и смешивают с 5 мг Cyanine5-ester-NHS (Lumiprobe), растворенного в 50 мкл ДМСО, и инкубируют на ротаторе (Multi RS-60, BioSan) при 4 oC в течение ночи в темноте. После инкубации диализуют против натрий-фосфатного буфера, используя Slide-A-Lyzer, 2000 MWCO (Pierce).
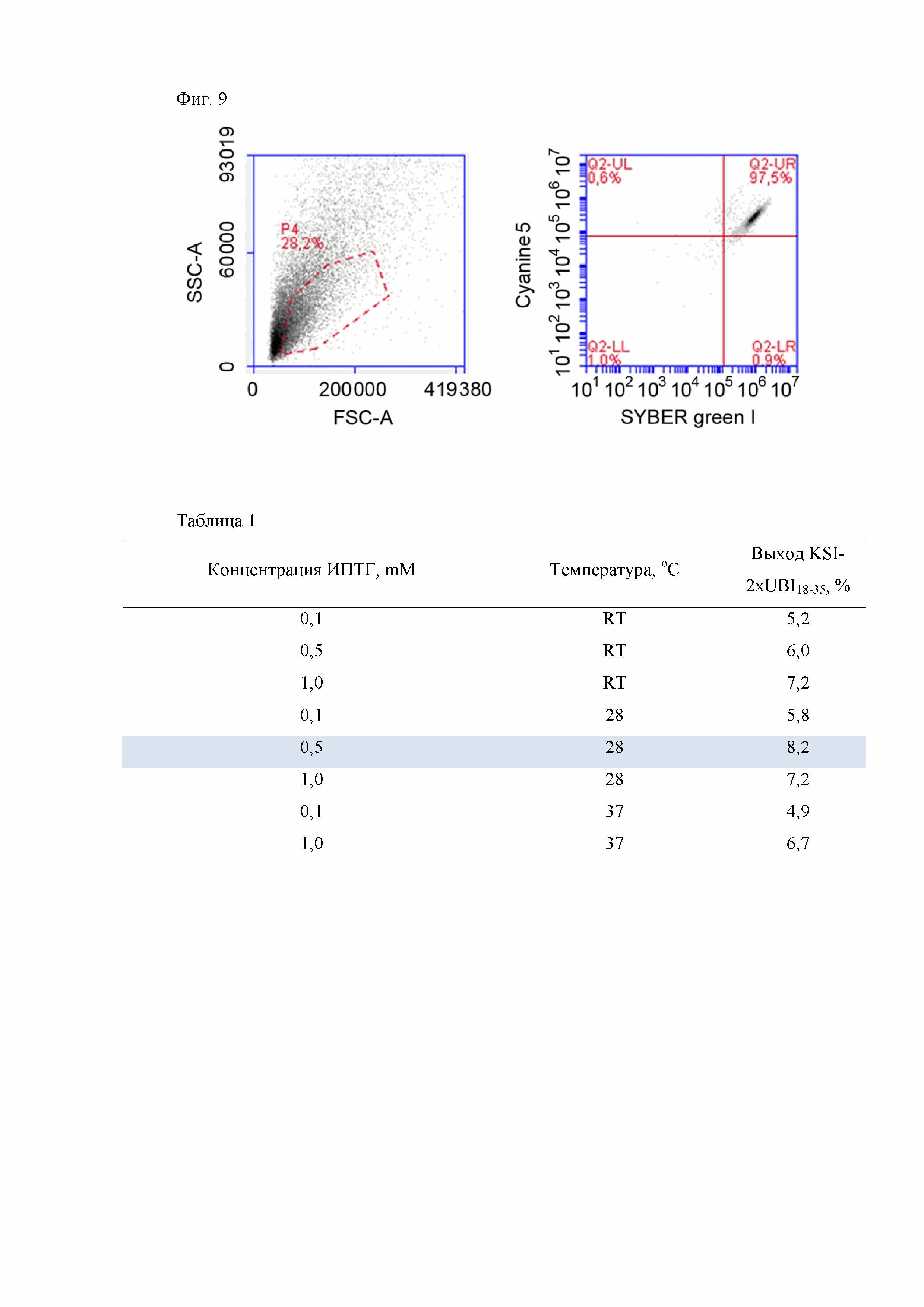
Для исследования связывание Cyanine5-UBI18-35 с микробными клетками 30 мкл Cyanine5-UBI18-35 (1 мМ) в натрий-фосфатном буфере (pH 7,4) добавляют к 420 мкл натрий-фосфатного буфера (pH 7.0), содержащего 0,01% (v/v) Tween-80, и смешивают с 50 мкл натрий-фосфатного буфера (pH 7,0) содержащего 106 микробных клеток Staphylococcus aureus. Смесь инкубируют на роторе при 4°C в течение 1 часа в темноте, центрифугируют при 3000 g 5 минут. Супернатант удаляют, дважды промывают осадок клеток, и растворяют в 200 мкл натрий-фосфатного буфера (pH 7.4). Клетки анализирую методом проточной цитомтерии (BD Accuri C6, BD Bioscience, CA). Дополнительно клетки окрашиваются с использованием флуоресцентного красителя Syber Green I. Эффективность связывания рекомбинантного антимикробного пептида UBI18-35 с клетками S. aureus по данным проточной цитофлуориметрии составляет 96,90±0,55% (Фиг. 9).
Обоснование режима экспрессии рекомбинантного антимикробного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом в клетках E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35
Технические параметры способа основаны на результатах анализа данных экспериментальных исследований (Таблица 1), которые показали, что:
- использование для индукции экспрессии ИПТГ в конечной концентрации 0,5 мМ обеспечивает оптимальное количество синтезируемого E.coli BL21 Rosetta DE3 pLysS / pET 31b-2xUBI18-35 рекомбинантного антимикробного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом.
- оптимальное количество рекомбинантного антимикробного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом нарабатывается при температуре культивирования E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 - 28°С. При температуре 22°С выход целевого продукта несколько ниже, тогда как увеличение температуры до 37°С приводит к значительному снижению выхода целевого продукта.
Таким образом, впервые получена плазмидная ДНК pET31b-2xUBI18-35, кодирующая аминокислотную последовательность рекомбинантного антимикробного пептида UBI18-35, в составе белка слияния с кетостероидизомеразой на N-конце пептида и последовательность из 6 гистидиновых остатков на С-конце, штамм-продуцент рекомбинантного пептида UBI18-35 E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35, а также описан метод получения рекомбинантного антимикробного пептида UBI18-35, способного встраиваться в мембрану микробных клеток и предназначенного для дифференциальной диагностики бактериального воспаления.
Изобретение иллюстрируется следующими чертежами:
Фиг. 1. Нуклеотидная последовательность и кодируемая ею аминокислотная последовательность антимикробного пептида UBI18-35.
Фиг. 2. Общая схема структурной организации плазмиды pET31b-2х UBI18-35. UBI - ген, кодирующий две молекулы антимикробного пептида UBI18-35, KSI–ген кетостероидизомеразы, Т7 - промотор фага Т7, AMP-ген устойчивости к ампициллину, 6xHis - ген, кодирующий 6 остатков гистидина, lac I-ген, кодирующий репрессор лактозного оперона E. coli;
Фиг. 3. Секвенирование кодирующего 2xUBI18-35 фрагмента в составе рекомбинантной плазмиды pET31b-2xUBI18-35. Сиквенс фрагмента, полученного в результате ПЦР с использованием в качестве матрицы рекомбинантной плазмиды pET31b-2xUBI18-35 и праймеров на Т7 промоторную и Т7 терминирующую области плазмиды (T7 forward: 5'-taatacgactcactataggg-3', T7 reverse: 5'-gctagttattgctcagcgg-3').
Фиг. 4. Оптимизация условий экспрессии пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и остатком из 6 гистидинов в клетках E.coli BL21 Rosetta DE3 pLysS / pET31b-2xUBI18-35. (А) Электрофоретический анализ в 16% SDS-ПААГ. М - набор белковых маркеров молекулярного веса (СибЭнзим), лизаты клеток E. coli, содержащих плазмиду pET31b-2xUBI18-35: до индукции экспрессии белка KSI-2xUBI18-35 в условиях культивирования в присутствие 1% глюкозы в среде (дорожка 1) и после индукции (дорожка 2) ИПТГ в конечной концентрации 0,1 мМ, культивирование при 22°С, (дорожка 3) -0,5 мМ ИПТГ, культивирование при 22°С, (дорожка 4) - 1 мМ ИПТГ, культивирование при 22°С, (дорожка 5) -0,1 мМ ИПТГ, культивирование при 28°С, (дорожка 6) -0,5 мМ ИПТГ, культивирование при 28°С, (дорожка 7) -1 мМ ИПТГ, культивирование при 28°С, (дорожка 8) -0,1 мМ ИПТГ, культивирование при 37°С, (дорожка 9) -0,5 мМ ИПТГ, культивирование при 37°С.
(Б) Вестерн-блот анализ лизата клеток E. coli, содержащих плазмиду pET31b-2xUBI18-35 до индукции экспрессии белка KSI-2xUBI18-35 без добавления в среду глюкозы (наблюдается слабая экспрессия белка) (дорожка 1'); (дорожки 1-9) - нумерация образцов в соответствие с SDS-ПААГ электрофореграммой.
Фиг. 5. Электрофореграмма в 16 % SDS-ПААГ белковых фракций и очищенного рекомбинантного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом. M - набор белковых маркеров молекулярного веса (СибЭнзим), (дорожка 1) - лизат клеток E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 до индукции белка KSI-2xUBI18-35, (дорожка 2) - лизат клеток E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35 после индукции белка KSI-2xUBI18-35 (дорожка 3) - общий клеточный лизат, (дорожка 4) - общий клеточный экстракт, (дорожка 5) - тельца включения KSI-2xUBI18-35, растворенные в буфере, содержащем 40 мМ трис-HCl, 500 мМ NaCl, 15мМ имидазол, 8М мочевину, pH 8.0, (дорожка 6) - осадок, после растворения телец включения KSI-2xUBI18-35, (дорожки 7, 8) - фракции, не взаимодействующие с хроматографическим сорбентом, (дорожки 9) - фракция, элюируемая буфером (40 мМ трис-HCl, 500 мМ NaCl, 300 мМ имидазол, 8 М мочевина, pH 8.0), содержащая антимикробный пептид UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом.
Фиг. 6. Анализ эффективности отщепления пептида UBI18-35, от белка-слияния KSI и 6-гистидинового тага с использованием бромциана. Электрофореграмма в 16,5 % трицин-ПААГ. M - набор низкомолекулярных белковых маркеров (Spectra Scientific Spectra Multicolor Low Range Protein Ladder, Thermo Scientific), (дорожка 1) - синтетический пептид UBI18-35 (контроль), (дорожка 2) - белковые фракции, содержащие целевой рекомбинантный пептид UBI18-35, (дорожка 3) - белок слияния KSI-2xUBI18-35 до обработки бромцианом.
Фиг. 7. МАЛДИ масс-спектр (А) рекомбинантного пептида UBI18-35 и (Б) пептида UBI18-35, полученного методом твердофазного синтеза.
Фиг. 8. Анализ хроматографических фракций, содержащих очищенный рекомбинантный пептид UBI18-35. Электрофореграмма в 16,5 % трицин-ПААГ. M - набор низкомолекулярных белковых маркеров (Spectra Scientific Spectra Multicolor Low Range Protein Ladder, Thermo Scientific), (дорожка 1) - синтетический пептид UBI18-35 (контроль), (дорожка 3) - фракция, собранная с 25 по 26 минуты хроматографирования методом гель-фильтрации на колонке упакованной Bio-Gel P 2 (рекомбинантный пептид UBI18-35).
Фиг. 9 Анализ связывания рекомбинантного антимикробного пептида UBI18-35, меченого Cyanine5, с микробными клетками S. aureus методом проточной цитометрии (клетки окрашены с использованием SYBER Green I).
ПРИЛОЖЕНИЕ
Фиг. 1. Нуклеотидная последовательность и кодируемая ею аминокислотная последовательность антимикробного пептида UBI18-35
Фиг. 2. Общая схема структурной организации плазмиды pET31b-2xUBI18-35
Фиг. 3. Секвенирование кодирующего 2xUBI18-35 фрагмента в составе рекомбинантной плазмиды pET31b-2xUBI18-35.
Фиг. 4. Оптимизация условий экспрессии пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом в клетках E.coli BL21 Rosetta DE3 pLysS / pET31b-2xUBI18-35.
Фиг. 5. Электрофореграмма в 16 % SDS-ПААГ белковых фракций и очищенного рекомбинантного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом.
Фиг. 6. Анализ эффективности отщепления пептида UBI18-35, от белка-слияния KSI-2xUBI18-35 с использованием бромциана.
Фиг. 7. МАЛДИ масс-спектр (вверху) рекомбинантного и синтетического пептидов UBI18-35
Фиг. 8. Анализ хроматографической фракции, содержащей очищенный рекомбинантный пептид UBI18-35.
Фиг. 9. Анализ связывания рекомбинатного антимикробного пептида UBI18-35, меченого Cyanine5, с микробными клетками S. aureus методом проточной цитометрии (клетки окрашены с использованием SYBER Green I).
Таблица 1. Обоснование режима экспрессии рекомбинантного антимикробного пептида UBI18-35 в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом в клетках E.coli BL21 Rosetta DE3 pLysS/ pET31b-2xUBI18-35
Способ дооперационного компьютерного инверсионного моделирования ацетабулярного компонента при диспластическом коксартрозе
Средство, обладающее противоописторхозной активностью
Средство для профилактики заражения описторхозом
Способ диагностики когнитивной дисфункции у пациентов с сахарным диабетом 1 типа методом протонной магнитно-резонансной спектроскопии
Способ прогнозирования течения саркоидоза органов дыхания по морфологическому строению гранулемы
Способ комбинированного лечения тяжелой степени акне
Способ сохранения жизнеспособности конечностей при глубоких отморожениях
Способ хирургического лечения ахалазии кардии iii-iv стадии, осложненной s-, l-образной деформацией пищевода
Способ билиопанкреатического шунтирования
Способ выделения и сохранения тотальной микробиоты кишечника человека для трансплантации
Способ иммобилизации биомолекул на поверхности магнитоуправляемых наночастиц железа покрытых углеродной оболочкой
Способ генерации антиген-специфических цитотоксических клеток с противоопухолевой активностью при раке молочной железы
Рекомбинантная плазмидная днк pet31b-phlip, обеспечивающая синтез рекомбинантного ph-зависимого векторного пептида phlip, способ получения рекомбинантного пептида и рекомбинантный ph-зависимый векторный пептид для диагностики и целевой доставки к зонам локального ацидоза
Средство для профилактики заражения описторхозом
4-[(пурин-6-ил)аминополиметиленкарбонил]-производные 3,4-дигидро-3-метил-7,8-дифтор-2h-[1,4]бензоксазина, обладающие противоопухолевой активностью

