Результат интеллектуальной деятельности: СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ L-МЕТИОНИНА ИЗ О-ФОСФО-L-ГОМОСЕРИНА И МЕТАНТИОЛА
Вид РИД
Изобретение
Настоящее изобретение относится к способу получения L-метионина, в котором O-фосфо-L-гомосерин и метантиол ферментативно превращают в L-метионин и Н3РО4. Такого превращения достигают путем воздействия фермента O-фосфо-L-гомосерин (OHPS)-зависимой метионинсинтазы. Настоящее изобретение также относится к O-фосфо-L-гомосерин-зависимым метионинсинтазам, т.е. белкам, которые способны ферментативно превращать O-фосфо-L-гомосерин и метантиол в L-метионин и Н3РО4. Настоящее изобретение также относится к микроорганизмам, генетически модифицированным таким образом, чтобы производить L-метионин из О-фосфо-L-гомосерина и метантиола.
Описанные ферменты и процессы также могут быть с успехом использованы для синтеза производных метионина, таких как S-аденозилметионин, глутатион, цистеин, S-аденозилгомоцистеин и метилтиоаденозин. Настоящее изобретение также относится к методам скрининга ферментов, катализирующих превращение O-фосфо-L-гомосерина и метантиола в L-метионин и H3PO4.
L-метионин является незаменимой аминокислотой, образующейся в ходе метаболизма, при превращении L-аспартата, путем активации, в L-аспартил фосфат с последующим восстановлением до полуальдегида L-аспартата и L-гомосерина. У бактерий, также как и у грибов, L-гомосерин подвергается О-ацетилированию с образованием или сукцинила, или ацетильного эфира, который, сам по себе, подвергается конденсации серы, или непосредственно с сульфидом (SH2) с образованием L-гомоцистеина, или опосредованно с L-цистеином с образованием L-цистатионина и его дальнейшей конверсией в гомоцистеин. Метилирование гомоцистеина происходит с помощью метилтетрагидрофолата, с образованием L-метионина.
В растениях, L-гомосериновый эфир, используемый в качестве предшественника цистатионина, представляет собой О-фосфо-L-roMocepHH. Цистатионин затем превращается в гомоцистеин под действием цистатионин бета-лиазы. Метилирование гомоцистеина затем приводит к образованию метионина. O-фосфо-L-гомосерин также образуется в ходе метаболизма в клетках бактерий, растений, грибов и млекопитающих, как прямой предшественник L-треонина, при участии пиридоксальфосфата треонин-синтазы (КФ (код фермента) 4.2.3.1). У растений существует жесткое регулирование потока углерода в метионин и треонин в O-фосфо-L-гомосерин точке перехода (Amir et al., TRENDS Plant Science 7 (2002), 153). В литературе (смотри, например, Kreft et al., Plant. Physiol. 104 (1994), 1215; Ravanel et al., Arch. Biochem. Biophys. 316 (1995), 572) имеются сведения о том, что и L-цистеин, и сульфид (SH2) могут быть конденсированы растительной цистатионин-гамма-синтазой с O-фосфо-L-гомосерином в L-цистатионин и L-гомоцистеин, с одновременным высвобождением фосфата. Сообщалось также, что выбор субстрата тиола очень ограничен, помимо L-цистеина, лишь немногие субстраты являются приемлемыми.
Хотя метионин является незаменимой аминокислотой, он не синтезируется de novo у животных, которые должны получать извне метионин или метионин-содержащие белки, или относящиеся к ним серосодержащие соединения. В частности, метионин имеет важное значение для кормления в птицеводстве и животноводстве, он не содержится в достаточном количестве в используемых растительных кормах. Таким образом, для эффективного фермерства необходим дополнительный источник метионина. На сегодняшний день, большинство, если не весь метионин, используемый для кормления животных, поступает из нефтехимии. Одним из ограничений производства метионина является стоимость энергии, затрачиваемой на восстановление серы. Таким образом, существует потребность в альтернативных способах получения этой аминокислоты, предпочтительно, использующих возобновляемые ресурсы, которые позволили бы использовать микроорганизмы, синтез метионина в которых сопоставим с производством метионина в промышленном масштабе.
Настоящее изобретение направлено на удовлетворение этой потребности путем создания способа получения L-метионина, в котором О-фосфо-L-гомосерин и метантиол ферментативно превращают в Ьнметионин и Н3РО4. В таком способе источником серы является метантиол. В этом соединении сера представлена в виде сульфида, уже восстановленная. Кроме того, использование O-фосфо-L-гомосерина в качестве предшественника, экономит, по меньшей мере, два атома углерода на молекулу синтезированного метионина по сравнению со стандартным метаболическим путем, который основан на использовании ацетилированных или сукцинилированных производных гомосерина. В целом этот путь дает гораздо лучшие результаты в синтезе метионина и его производных.
Таким образом, настоящее изобретение относится к способу производства L-метионина, в котором L-метионин ферментативно получают в согласно следующей схеме реакции:
O-фосфо-L-гомосерин + CH3-SH <=> L-метионин + Н3РО4
В настоящее время отсутствуют данные о существовании в природе белка, обладающего способностью превращать O-фосфо-L-гомосерин и метантиол в L-метионин. Авторы настоящего изобретения рассмотрели возможность получения L-метионина экономически эффективным способом из О-фосфо-L-гомосерина и метантиола, и, с этой целью, разработали белки, которые не встречаются в природе, и которые способны превращать О-фосфо-L-гомосерин и метантиол в L-метионин. Как видно из приведенных примеров, авторы настоящего изобретения разработали систему, которая позволяет создавать ферменты, обладающие способностью превращать О-фосфо-L-гомосерин и метантиол в L-метионин и Н3РО4. Кроме того, авторы изобретения успешно применили эту систему для создания новых вариантов ферментов, полученных из существующих ферментов, обладающих способностью превращать О-фосфо-L-гомосерин и метантиол в L-метионин и H3PO4. Для этой цели они отталкивались от существующих ферментов, не обладающих способностью превращать О-фосфо-L-гомосерин и метантиол в L-метионин и Н3РО4, получали мутанты из таких существующих ферментов, и отбирали ферменты, демонстрирующие способность превращать О-фосфо-L-гомосерин и метантиол в L-метионин и Н3РО4. Таким образом, авторы изобретения выявили новую ферментативную активность, не описанную ранее, и предложили надежные и воспроизводимые способы получения соответствующих ферментов. Соответствующие ферменты в контексте настоящего изобретения называют O-фосфо-L-гомосерин-зависимыми метионинсинтазами.
Таким образом, настоящее изобретение, в частности, относится к способу производства L-метионина, в котором O-фосфо-L-гомосерин и метантиол ферментативно превращают в L-метионин и H3PO4 согласно следующей схеме реакции:
O-фосфо-L-гомосерин + CH3-SH <=> L-метионин + H3PO4
где ферментативное превращение осуществляют путем использования O-фосфо-L-гомосерин-зависимой метионинсинтазы.
В принципе, любая O-фосфо-L-гомосерин-зависимая метионинсинтаза, т.е. любой белок, обладающий способностью превращать О-фосфо-L-гомосерин и метантиол в L-метионин и H3PO4) может быть использован в способе по изобретению. Настоящее изобретение впервые описывает белки, обладающие этой способностью, и предлагает способы получения дополнительных белков, обладающих этой способностью. В частности, настоящее изобретение показывает, что, взяв за основу, растительную цистатионин-гамма-синтазу (КФ 2.5.1.48), которая в природе не обладает способностью превращать О-фосфо-L-гомосерин и метантиол в L-метионин, можно получить, в результате мутации и отбора, ее варианты, обладающие этой способностью.
О-фосфо-L-гомосерин-зависимые метионинсинтазы будут описаны ниже, в контексте белка по изобретению, и любая описанная О-фосфо-L-гомосерин-зависимая метионинсинтаза, может быть использована в способе по изобретению.
Как видно из приведенных примеров, авторы изобретения успешно создали несколько различных белков, которые демонстрируют способность превращать О-фосфо-L-гомосерин и метантиол в L-метионин и H3PO4. Последовательности этих белков показаны в SEQ ID NO: от 6 до 29. Эти белки получены, путем мутации и отбора в скрининговой системе, как описано в примерах, из растительной цистатионин гамма-синтазы (КФ 2.5.1.48), которая, в природе, не способна превращать О-фосфо-L-гомосерин и метантиол в L-метионин.
Кроме того, отталкиваясь от последовательностей, представленных в SEQ ID NO: от 6 до 29, возможно предложить дополнительные белки, обладающие активностью по превращению О-фосфо-L-гомосерина и метантиола в L-метионин и Н3РО4. То есть, например, возможно дальнейшее увеличение аффинности белков к субстрату О-фосфо-L-гомосерина и/или к субстрату метантиола, или улучшение других свойств белка, как описано ниже. Таким образом, в предпочтительном варианте осуществления способа по изобретению, ферментативного превращения О-фосфо-L-гомосерина и метантиола в L-метионин и H3PO4 достигают путем использования белка, выбранного из группы, состоящей из:
(a) белка, содержащего любую из аминокислотных последовательностей SEQ ID NO: от 6 до 29;
(b) белка, имеющего последовательность, идентичную по меньшей мере на 60% любой из SEQ ID NO: от 6 до 29, и обладающего ферментативной активностью превращения О-фосфо-L-гомосерина и метантиола в L-метионин и Н3РО4.
Ферментативная активность по превращению О-фосфо-L-гомосерина и метантиола в L-метионин и Н3РО4 может, например, быть оценена с помощью анализа, как описано в приведенных примерах. Для этой цели можно, например, использовать штамм S. cerevisiae, который имеет ауксотрофный фенотип по метионину. Примерами такого штамма являются штаммы, в которых ферменты гомосерин трансацетилаза и гомоцистеинсинтаза удалены или инактивированы. Предпочтительно, оба фермента удалены или инактивированы. Как показано на Фигуре 1, S. cerevisiae, зависит от обязательного использования О-ацетилгомосерина для синтеза гомоцистеина, который затем превращается в метионин. Фермент гомосерин трансацетилаза, ответственный за синтез О-ацетилгомосерина, кодируется геном МЕТ2. После инактивации МЕТ2, гомосерин не может больше превращаться в О-ацетилгомосерин, и весь поток гомосерина перенаправляется в сторону фосфогомосерина. Поскольку реакция, катализируемая МЕТ2, является единственным источником О-ацетилгомосерина в дрожжах, инактивация гена МЕТ2 приводит к появлению дрожжевых штаммов, имеющих строгий ауксотрофный фенотип по метионину. Тем не менее, гомоцистеин (последний предшественник метионина) может быть получен или из цистеина (через транссульфурационный путь), или из переработки S-аденозилметионина. Для того чтобы быть уверенными, что метионин не может быть синтезирован вообще, ген МЕТ6, кодирующий гомоцистеин метилтрансферазу, отвечающую за синтез метионина из гомоцистеина и метилтетрагидрофолата, также удаляют.
Таким образом, в анализе для проверки способности белка превращать О-фосфо-L-гомосерин и метантиол в L-метионин и H3PO4, может быть, предпочтительно, использован штамм S. cerevisiae, в котором гены МЕТ2 и/или МЕТ6, предпочтительно оба гена, удалены или инактивированы. Штамм с одновременно инактивированными met2Δ met6Δ (двойной мутант), не может расти в отсутствие метионина и, в частности, не может расти в присутствии метантиола в качестве источника серы. Такой штамм уже не в состоянии синтезировать О-ацетил-гомосерин, но производит О-фосфо-гомосерин. Такой штамм дрожжей затем может быть трансформирован молекулой нуклеиновой кислоты, кодирующей белок, который будет протестирован на способность превращать О-фосфо-L-гомосерин и метантиол в L-метионин и Н3РО4. Штамм выращивали в/на среде, содержащей метантиол в качестве единственного источника серы; способность расти на такой среде свидетельствует о том, что экспрессированный белок в состоянии превращать O-фосфо-L-гомосерин (OHPS) и метантиол в L-метионин; еще более предпочтительным является использование штамма, в котором ген, кодирующий треонинсинтазу, также инактивирован, например, путем делетирования или разрушения. В met2Δ met6Δ мутантных штаммах, OPHS синтезируется в реакции, катализируемой гомосерин-киназой, кодируемой геном THR1, но он не может эффективно накапливаться из-за его активного превращения в треонин с помощью треонин-синтазы, кодируемой геном THR4. Такие тройные met2Δ met6Δ thr4Δ мутантные штаммы, таким образом, позволяют детектировать очень низкую активность OPHS-зависимой метионинсинтазы, и были использованы для первого скрининга клеток с целью выделения новых белков, обладающих OPHS-зависимой метионинсинтазной активностью.
Эта ферментативная активность в дальнейшем может быть подтверждена с помощью in vitro анализа, в котором О-фосфо-L-гомосерин и метантиол инкубируют in vitro в подходящих условиях с бесклеточным экстрактом из штамма дрожжей, экспрессирующего белок, который будет протестирован, или с (частично) очищенным белком, который будет протестирован, и в котором производство метионина детектируется с помощью жидкостной хроматографии с тандемной масс-спектрометрией (ЖХ/МС/МС) с использованием С3 метионина в качестве внутреннего контроля (Ravanel et al. Archives of Biochemistry and Biophysics 316 (1995), 572-584).
О-фосфо-L-гомосерин, используемый в таком in vitro анализе в качестве субстрата, может быть получен, например, способом, показанным на Фигуре 2 (Barclay et al., J. Chem. Soc, Chem. Com (1994) 815-816).
Как упоминалось выше, примерами белков, обладающих ферментативной активностью по превращению O-фосфо-L-гомосерина и метантиола в L-метионин и Н3РО4, являются белки, имеющие аминокислотную последовательность, в соответствии с любой из последовательностей SEQ ID NO: от 6 до 29. Таким образом, в одном предпочтительном варианте осуществления способа по изобретению используют белок, содержащий аминокислотную последовательность, в соответствии с любой из последовательностей SEQ ID NO: от 6 до 29. Тем не менее, конечно, также можно использовать варианты этих белков, то есть белки с аминокислотной последовательностью, имеющей высокую степень идентичности аминокислотной последовательности, в соответствии с любой из SEQ ID NO: от 6 до 29, и обладающие ферментативной активностью по превращению О-фосфо-L-гомосерина и метантиола в L-метионин и Н3РО4. Идентичность последовательности составляет по меньшей мере 60%, предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или, по меньшей мере 95%, и наиболее предпочтительно по меньшей мере 96%, 97%, 98% или 99% любой из последовательностей, в соответствии с SEQ ID NO: от 6 до 29. Предпочтительно, степень идентичности определяют путем сравнения соответствующей последовательности с аминокислотной последовательностью, в соответствии с любой из последовательностей SEQ ID NO: от 6 до 29. Когда сравниваемые последовательности не имеют одной и той же длины, степень идентичности, предпочтительно, относится или к проценту аминокислотных остатков в более короткой последовательности, идентичных аминокислотным остаткам в более длинной последовательности, или к проценту аминокислотных остатков в более длинной последовательности, идентичных аминокислотным остаткам в более короткой последовательности. Степень идентичности последовательностей может быть определена методами, хорошо известными в данной области техники, с использованием подходящих, предпочтительно, компьютерных алгоритмов, таких как CLUSTAL. При использовании Clustal-анализа для того, чтобы определить, является ли конкретная последовательность идентичной, например, на 80% эталонной последовательности, могут быть использованы настройки по умолчанию, или, предпочтительно, следующие настройки: Matrix: blosum 30; Open gap penalty: 10.0; Extend gap penalty: 0.05; Delay divergent: 40; Gap separation distance: 8 для сравнения аминокислотных последовательностей. Для сравнений нуклеотидных последовательностей, значение "Extend gap penalty" устанавливают предпочтительно равным 5,0.
Предпочтительно, степень идентичности рассчитывают по всей длине последовательности. Кроме того, если в контексте настоящего изобретения используют термин "гомология", этот термин, предпочтительно, означает "идентичность последовательности".
Способ по изобретению также позволяет получать другие серосодержащие соединения, являющиеся производными L-метионина. Примерами таких соединений являются S-аденозилметионин, глутатион, цистеин, S-аденозилгомоцистеин, метилтиоаденозин и 2-оксо-4-метилтиобутаноат.
Таким образом, настоящее изобретение также относится к способу получения S-аденозилметионина, который включает в себя способ получения L-метионина по изобретению, описанный выше, и, в котором L-метионин дополнительно преобразуют в S-аденозилметионин в соответствии со следующей реакцией:
Метионин + АТФ => S-аденозилметионин + РР + Ρ
Эта ферментативная реакция известна в данной области техники, и ферменты, катализирующие эту реакцию, известны в данной области техники.
Эти ферменты называются S-аденозилметионин синтазы (КФ 2.5.1.6). Примерами соответствующих ферментов являются SAM1 и SAM2 в дрожжах. Таким образом, в случае, когда такой способ осуществляется в организме, такой организм, предпочтительно, сверхэкспрессирует соответствующий фермент (ы), который (е) способен превращать L-метионин в S-аденозилметионин.
Также, может быть полезной дальнейшая модификация такого организма, чтобы предотвратить поток S-аденозилметионина в другие метаболические пути. В дрожжах, может, например, быть полезной, инактивация активности аденозин-киназы (КФ 2.7.1.20, кодируемой геном дрожжей AD01) для того, чтобы уменьшить поток S-аденозилметионина в S-аденозилгомоцистеин.
Кроме того, настоящее изобретение также относится к способу получения цистеина, который включает способ получения L-метионина по изобретению, описанный выше, и в котором L-метионин далее превращают в цистеин. Превращение L-метионина в цистеин известно из предшествующего уровня техники, и может быть достигнуто за счет средств и методов, известных специалистам в данной области. Например, L-метионин может быть сначала превращен в S-аденозилметионин, который затем превращают в S-аденозилгомоцистеин в соответствии со следующей реакцией:
S-аденозилметионин + метил-акцептор => S-аденозилгомоцистеин + метилированный акцептор
Эта реакция катализируется S-аденозилметионин-зависимой метилтрансферазой.
S-аденозилгомоцистеин может быть далее превращен в L-гомоцистеин в соответствии со следующей реакцией:
S-аденозилгомоцистеин <=> L-гомоцистеин + аденозин
Эта реакция катализируется S-аденозилгомоцистеин гидролазой, КФ 3.3.1.1. Впоследствии L-гомоцистеин может быть превращен в L-цистатионин в соответствии со следующей реакцией:
L-гомоцистеин + L-серин <=> L-цистатионин + H2O
Эта реакция катализируется цистатионин-бета-синтазой (КФ 4.2.1.22). И, наконец, L-цистатионин превращают в цистеин в соответствии со следующей реакцией:
L-цистатионин + Н2О <=> L-цистеин + NH3 + оксобутаноат
Эта реакция катализируется цистатионин-гамма-лиазой (КФ 4.4.1 0.1).
Если такой способ осуществляется в дрожжах, предпочтительно, сверхэкспрессированы следующие гены: SAM1, SAM2, SAH1, STR4 и STR1. Кроме того, ген STR2, кодирующий у дрожжей цистатионин гамма-синтазу, должен быть делетирован для того чтобы снизить обратный синтез, из цистеина в гомоцистеин. Кроме того, такие дрожжи, преимущественно, также содержат мутацию МЕТ6, ликвидирующую метилтетрагидрофолат-зависимую метионинсинтазу, а также делецию гена DUG2, который участвует в основном пути деградации глутатиона.
Кроме того, настоящее изобретение также относится к способу получения глутатиона, который включает способ получения L-метионина по изобретению, описанный выше, и в котором L-метионин дополнительно превращают в глутатион. Превращение L-метионина в глутатион известно в данной области техники и может быть осуществлено с помощью средств и методов, известных специалистам в данной области. Например, L-метионин может быть сначала превращен в S-аденозилметионин, который дополнительно превращают в цистеин, как описано выше, и полученный таким образом цистеин, дополнительно превращают в Glu-Cys (гамма-L-глутамил-L-цистеин) в соответствии со следующей реакцией:
АТФ + L-глутамат + L-цистеин <=> гамма-L-глутамил-L-цистеин + АДФ + Pi
Эта реакция катализируется глутамат-цистеинлигазой (КФ 6.3.2.2).
Полученный таким образом Glu-Cys затем превращают в глутатион в соответствии со следующей реакцией:
АТФ + гамма-L-глутамил-L-цистеин + глицин = глутатион + АДФ + Pi
Эта реакция катализируется глутатионсинтазой (КФ 6.3.2.3). Если такой способ осуществляют в дрожжах, предпочтительно, чтобы дрожжи были сконструированы так, как это описано выше в связи со способом производства цистеина, и такие дрожжи также должны сверхэкспрессировать гены GSH1 и GSH2, участвующие в превращении цистеина в глутатион. В наиболее предпочтительном варианте ген GSH1 экспрессирует устойчивый фермент по типу обратной связи.
Кроме того, настоящее изобретение также относится к способу получения 2-оксо-4-метилтиобутаноата, который включает способ получения L-метионина по изобретению, описанный выше, и в котором L-метионин дополнительно превращают в 2-оксо-4-метилтиобутаноат в соответствии со следующей реакцией:
Метионин + 2-оксо кислота => 2-оксо-4-метилтиобутаноат + L-аминокислота
Эта ферментативная реакция известна в данной области техники, и ферменты, катализирующие эту реакцию, известны в данной области техники. Эти ферменты называются метионинтрансаминазы (КФ 2.6.1.88). Примерами соответствующих ферментов являются AR08, ВАТ1, ВАТ2 в дрожжах. Таким образом, в случае, когда такой способ осуществляется в организме, такой организм, предпочтительно, сверхэкспрессирует соответствующий фермент (ы), который (ые) способен превращать L-метионин в 2-оксо-4-метилтиобутаноат.
Также может быть полезной дальнейшая модификация такого организма, чтобы предотвратить поток 2-оксо-4-метилтиобутаноата в другие метаболические пути. В дрожжах, например, может быть полезной, инактивация активности фенилпируват декарбоксилазы (КФ 4.1.1.43, кодируемой геном ARO10 в дрожжах) или активности пируватдекарбоксилазы (КФ 4.1.1.1 кодируемой генами PDC1, PDC5 и PDC6 в дрожжах) для того, чтобы уменьшить поток 2-оксо-4-метилтиобутаноата в 3-(метилтио)пропионовый альдегид.
Способ по настоящему изобретению может быть осуществлен in vitro или in vivo. Под реакцией in vitro понимают реакцию, в которой используют не клетки, а бесклеточную систему. Таким образом, термин "in vitro", преимущественно, означает в бесклеточной системе. Термин "in vitro", в одном варианте осуществления, означает в присутствии изолированного фермента. В одном варианте осуществления фермент, используемый в способе, используют в очищенном виде.
Для проведения процесса in vitro субстраты для реакции и фермента инкубируют в условиях (буфер, температура и т.д.), позволяющих ферменту быть активным, и иметь место ферментативному превращению. Реакцию проводят в течение времени, достаточного для получения L-метионина. Продукция L-метионина может быть измерена способами, известными в данной области техники.
Фермент может быть в любой подходящей форме, позволяющей ферментативной реакции иметь место. Он может быть очищенным, частично очищенным, в виде сырых клеточных экстрактов или в виде частично очищенных клеточных экстрактов. Также возможно, что фермент иммобилизован на подходящем носителе.
В другом варианте осуществления, способ по изобретению осуществляют в культуре, в присутствии организма, предпочтительно, микроорганизма, производящего белок, обладающий способностью превращать О-фосфо-L-гомосерин и метантиол в L-метионин и H3PO4. Белок представляет собой белок, описанный здесь.
Организм, используемый в таком способе, предпочтительно, представляет собой клетку-хозяина в соответствии с настоящим изобретением, как описано здесь.
Настоящее изобретение также относится к белку, обладающему способностью ферментативно превращать О-фосфо-L-гомосерин и метантиол в L-метионин. В контексте настоящего изобретения, такие ферменты называют OHPS-зависимыми метионинсинтазами. Как упоминалось выше, до настоящего времени нет никаких сведений о существовании в природе белков, обладающих способностью превращать О-фосфо-L-гомосерин и метантиол в L-метионин, и, настоящее изобретение впервые предлагает белки, позволяющие получать L-метионин в соответствии со способом по изобретению, как описано выше.
В предпочтительном варианте осуществления, белок по изобретению, способный ферментативно превращать О-фосфо-L-гомосерин и метантиол в L-метионин, получают из цистатионин-гамма-синтазы (КФ 2.5.1.48) в результате мутации. Такой мутацией может быть замена одного или нескольких аминокислотных остатков и/или удаление одного или нескольких аминокислотных остатков и/или добавление одного или нескольких аминокислотных остатков в аминокислотной последовательности цистатионин гамма-синтазы (КФ 2.5.1.48).
Цистатионин-гамма-синтазы известны и описаны в различных организмах. Например, для растений известны более чем 350 последовательностей цистатионин-гамма-синтазы, в частности, для A. thaliana, Nicotiana tabacum, Triticum aestivum, Solanum lycopersicum, Lemna paucicostata, Solanum tuberosum, Spinacia olerácea, Astragalus racemosus, Astragalus bisulcatus, Astragalus sinicus and Neptunia amplexicaulis.
Цистатионин-гамма-синтазы также известны для бактерий и грибов. Для бактерий описаны более 22,000 последовательностей; примерами последовательностей бактерий и грибов являются последовательности из Saccharomyces cerevisiae, Neurospora crassa, Salmonella erterica, Escherichia coli, Agrobacterium tumefaciens, Alcaligenes faecalis, Aneurinibacillus aneurinilyticus, Bacillus pumius, Bacillus subtilis, Corynebacterium glutamicum, Helicobacter pylori, Lysinibacillus sphaericus, Mycobacterium tuberculosis, Pectobacterium carotovorum, Pseudomonas dacunhae, Pseudomonas putida, Streptomyces phaeochromogenes.
Способы получения белков, обладающих активностью по превращению О-фосфо-L-гомосерина и метантиола в L-метионин и Н3РО4 описаны в приведенных примерах и более подробно будут описаны ниже. В принципе, любая цистатионин-гамма-синтаза (КФ 2.5.1.48) может быть использована в качестве исходного материала для получения белка, обладающего активностью по превращению O-фосфо-L-гомосерина и метантиола в L-метионин и Н3РО4. Предпочтительно, белок, обладающий активностью по превращению O-фосфо-L-гомосерина и метантиола в L-метионин и H3PO4, полученный из цистатионин-гамма-синтазы, (КФ 2.5.1.48) имеет последовательность, идентичную по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, еще более предпочтительно по меньшей мере на 90%, и наиболее предпочтительно по меньшей мере на 95%, аминокислотной последовательности природной цистатионин-гамма-синтазы.
В одном предпочтительном варианте осуществления, цистатионин-гамма-синтаза (КФ 2.5.1.48), из которой происходит OHPS-зависимая метионинсинтаза, представляет собой растительную цистатионин-гамма-синтазу (КФ 2.5.1.48), предпочтительно цистатионин-гамма-синтазу (КФ 2.5.1.48) CGS1 из Arabidopsis thaliana, наиболее предпочтительно, цистатионин гамма-синтазу (КФ 2.5.1.48), имеющую аминокислотную последовательность SEQ ID NO: 1. В еще более предпочтительном варианте OHPS-зависимую метионинсинтазу, используемую в способе по изобретению, получают из последовательности SEQ ID NO: 2. Эта последовательность соответствует последовательности SEQ ID NO: 1, за исключением того, что остаток глицина в положении 84 заменен на остаток серина. Эта замена, то есть mto мутация, снимает репрессию трансляции, оказываемую S-аденозилметионином на CGS1 (Onoue et al., Journal of Biological Chemistry 286 (2011), 14903-14911). В наиболее предпочтительном варианте осуществления, OHPS-зависимую метионинсинтазу, используемую в способе по изобретению, получают из последовательности SEQ ID NO: 3. Эта последовательность соответствует последовательности SEQ ID NO: 2 за исключением того, N-концевая хлоропластная сигнальная последовательность (аминокислотные остатки от 1 до 57) была удалена, и остаток метионина добавлен на N-конце.
Примерами OHPS-зависимых метионинсинтаз, которые могут быть использованы в способе по настоящему изобретению, являются
(i) OHPS-зависимые метионинсинтазы, полученные из цистатионин-гамма-синтазы, имеющей аминокислотную последовательность SEQ ID NO: 3, путем замены или делеции по меньшей мере одного аминокислотного остатка в SEQ ID NO: 3, выбранного из группы, состоящей из:
(a) пролина 10;
(b) аспарагина 11;
(c) глутамина 15;
(d) изолейцина 27;
(e) аланина 30;
(f) лейцина 45;
(g) серина 47;
(h) валина 60;
(i) аланина 68;
(j) фенилаланина 150;
(k) треонина 178;
(l) аспартата 183;
(m) изолейцина 185;
(n) треонина 220;
(о) метионина 232;
(p) валина 245;
(q) аланина 257;
(r) аспарагина 259;
(s) фенилаланина 261;
(t) фенилаланина 275;
(u) изолейцина 287;
(v) гистидина 289;
(w) тирозина 324;
(х) глицина 326;
(у) пролина 356;
(2) треонина 371;
(аа) валина 396;
(bb) пролина 405;
(сс) аспартата 431;
(dd) изолейцина 436;
(ее) изолейцина 457;
(ff) аспартата 459;
(gg) пролина 470;
(hh) глутамата 472;
(ii) аланина 506;
(jj) изолейцина 507;
или
(ii) OHPS-зависимые метионинсинтазы, полученные из цистатионин-гамма-синтазы, аминокислотная последовательность которой, по меньшей мере, на 60% идентична аминокислотной последовательности SEQ ID NO: 3, путем замены или делеции по меньшей мере одной аминокислоты, соответствующей любой из перечисленных выше, от (а) до (jj), в SEQ ID NO: 3.
Предпочтительно, последовательность идентичнна по меньшей мере на 70%, еще более предпочтительно по меньшей мере на 80% и, наиболее предпочтительно по меньшей мере на 90%.
Термин "замена" означает, что аминокислоту, находящуюся в указанном положении, замещают другим аминокислотным остатком. В контексте настоящего изобретения, фраза "замещенный другим аминокислотным остатком" означает, что соответствующие аминокислотные остатки в указанном положении могут быть замещены любыми другими возможными аминокислотными остатками, предпочтительно, аминокислотными остатками, выбранными из группы, состоящей из аланина, аргинина, аспарагина, аспарагиновой кислоты, цистеина, глутамина, глутаминовой кислоты, глицина, гистидина, изолейцина, лейцина, лизина, метионина, фенилаланина, пролина, серина, треонина, триптофана, тирозина и валина. Предпочтительные замены для определенных положений указаны ниже. Аминокислотные остатки, расположенные в позиции, соответствующей позиции, выбранной из группы, включающей позиции от (а) до (jj), перечисленные выше, в аминокислотной последовательности SEQ ID NO: 3, могут быть идентифицированы специалистом способами, известными в данной области техники. Например, такие аминокислотные остатки могут быть идентифицированы путем сравнения изучаемой последовательности с последовательностью SEQ ID NO: 3, и путем идентификации позиций, соответствующих указанным выше позициям в SEQ ID NO: 3. Сравнение может быть проведено методами и способами, известными специалисту в данной области, например, с использованием такого известного компьютерного алгоритма, как алгоритм Липмана-Пирсона (Science 227 (1985), 1435) или алгоритма CLUSTAL. Предпочтительно, чтобы в таком сравнении, максимальная гомология приходилась на консервативные аминокислотные остатки, присутствующие в аминокислотных последовательностях.
При сравнении аминокислотных последовательностей таким способом, независимо от того, есть ли в аминокислотных последовательностях вставки или делеции, позиции соответствующих аминокислотных остатков могут быть определены в заданной последовательности.
По одному из вариантов осуществления, OHPS-зависимые метионинсинтазы по настоящему изобретению имеют аминокислотную последовательность, в которой
(i) аминокислотный остаток в положении 10 в аминокислотной последовательности SEQ ID NO: 3 или в положении, соответствующем этой позиции, замещен на лейцин; и/или
(ii) аминокислотный остаток в положении 11 в аминокислотной последовательности SEQ ID NO: 3 или в положении, соответствующем этой позиции, замещен на аспартат; и/или
(iii) аминокислотный остаток в положении позиции 15 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на аргинин; и/или
(iv) аминокислотный остаток в положении 27 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на серии; и/или
(v) аминокислотный остаток в положении 30 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на треонин; и/или
(vi) аминокислотный остаток в положении 45 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на серии; и/или
(vii) аминокислотный остаток в положении 47 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на треонин; и/или
(viii) аминокислотный остаток в положении 60 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на аспартат; и или
(ix) аминокислотный остаток в положении 68 в аминокислотной последовательности SEQ ID NO: 3, в положении, соответствующем этой позиции, замещен на треонин; и/или
(х) аминокислотный остаток в положении 150 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на лейцин; и/или
(xi) аминокислотный остаток в положении 178 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на изолейцин; и/или
(xii) аминокислотный остаток в положении 183 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на глутамат; и/или
(xiii) аминокислотный остаток в положении 185 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на валин; и/или
(xiv) аминокислотный остаток в положении 220 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на серии; и/или
(xv) аминокислотный остаток в положении 232 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на лейцин; и/или
(xvi) аминокислотный остаток в положении 245 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на аланин; и/или
(xvii) аминокислотный остаток в положении 257 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на треонин; и/или
(xviii) аминокислотный остаток в положении 259 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на аспартат или серии; и/или
(xiх) аминокислотный остаток в положении 261 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на серии; и/или
(xx) аминокислотный остаток в положении 275 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на лейцин; и/или
(xxi) аминокислотный остаток в положении 287 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на валин или фенилаланин; и/или
(xxii) аминокислотный остаток в положении 289 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на тирозин или аргинин; и/или
(xxiii) аминокислотный остаток в положении 324 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен фенилаланином; и/или
(xxiv) аминокислотный остаток в положении 326 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на серии; и/или
(xxv) аминокислотный остаток в положении 356 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на треонин; и/или
(xxvi) аминокислотный остаток в положении 371 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на аланин; и/или
(xxvii) аминокислотный остаток в положении 396 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на аланин; и/или
(xxviii) аминокислотный остаток в положении 405 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на серин; и/или
(xxix) аминокислотный остаток в положении 431 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на глицин; и/или
(xxx) аминокислотный остаток в положении 436 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на треонин; и/или
(xxxi) аминокислотный остаток в положении 457 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на лейцин; и/или
(xxxii) аминокислотный остаток в положении 459 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на аспарагин; и/или
(xxxiii) аминокислотный остаток в положении 470 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на серии; и/или
(xxxiv) аминокислотный остаток в положении 472 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на глицин; и/или
(xxxv) аминокислотный остаток в положении 506 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на глицин; и/или
(xxxvi) аминокислотный остаток в положении 507 в аминокислотной последовательности SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещен на валин.
Настоящее изобретение также относится к вариантам, описанным выше от (i) до (xxxvi), в которых аминокислотный остаток, указанный как замещающий аминокислотный остаток в положении SEQ ID NO: 3, не является конкретным аминокислотным остатком, но представляет собой аминокислотный остаток, консервативный по отношению к указанному замещающему аминокислотному остатку.
Оценить, является ли аминокислота консервативной по отношению к другой аминокислоте, можно методами, известными в данной области техники. Одной из возможностей является использование матрицы РАМ 250; альтернативно, могут быть использованы матрицы семейства Blosum.
В одном варианте осуществления, настоящее изобретение относится к OHPS-зависимой метионинсинтазе, имеющей аминокислотную последовательность SEQ ID NO: 3, или аминокислотную последовательность, идентичную, по меньшей мере, на 60% последовательности SEQ ID NO: 3, где аминокислотные остатки в положении 356 в SEQ ID NO: 3, или в положении, соответствующем этой позиции, замещены другим аминокислотным остатком. В предпочтительном варианте осуществления настоящее изобретение относится к такому белку, в котором, по меньшей мере, один аминокислотный остаток, замещен в положении, выбранном из группы, состоящей из позиций 10, 11, 15, 27, 28, 30, 32, 45, 47, 60, 68, 104, 150, 178, 183, 185, 220, 232, 245, 257, 259, 261, 275, 287, 289, 324, 326, 371, 396, 405, 431, 436, 457, 459, 470, 472, 506 и 507, предпочтительно, выбранном из группы, состоящей из позиций 10, 11, 15, 30, 32, 45, 47, 68, 104, 150, 178, 183, 185, 220, 232, 245, 257, 259, 261, 275, 287, 289, 326, 371, 396, 405, 431, 436, 459, 470, 472, 506 и 507, еще более предпочтительно, выбранном из группы, состоящей из позиций 275 и 396.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции являются следующие:
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 10, 27, 60, 324 и 457.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 32, 287, 289 и 356.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 10, 232, 245, 259, 356, 431 и 436.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 11, 15, 30, 45, 47, 68, 178, 356, 371 и 459.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 32 и 356.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 32, 60, 324 и 457.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 32, 287, 289 и 356.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 32, 232, 245, 259, 356, 431 и 436.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 32, 45, 47, 68, 178, 356, 371 и 459.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 232, 245, 259, 356, 431 и 436.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 178, 356, 371 и 459.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 150, 257, 259, 261, 275, 289, 356 и 506.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 185, 356 и 405.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 275, 356, 396 и 472.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 275, 326, 356 и 396.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 220, 275, 356 и 396.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 83, 275, 356, 396 и 507.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 275, 287, 356, 396 и 507.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 275, 356, 396 и 470.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 275, 356 и 507.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 275, 356 и 396.
В одном варианте осуществления, позициями, в которых происходят замены и/или делеции, являются следующие: позиции 275, 287 и 356.
Предпочтительными заменами в этих позициях являются те, которые указаны выше.
OHPS-зависимые метионинсинтазы, которые могут быть использованы в способе по изобретению, представляют собой ферменты, которые могут быть получены из любой из описанных выше OHPS-зависимых метионинсинтаз путем делеции одной или нескольких N-концевых аминокислот, соответствующих аминокислотам от 1 до 103 аминокислотной последовательности SEQ ID NO: 3. Как показано на примерах укороченные формы вышеописанных OHPS-зависимых метионинсинтаз, например, укороченные формы, в которых, по сравнению с SEQ ID NO: 3, 31 аминокислотный остаток или 103 аминокислотных остатка удалены на N-конце, также все еще обладают эффективной OHPS-зависимой метионинсинтазной активностью.
В предпочтительном варианте OHPS-зависимая метионинсинтаза по изобретению также обладает ферментативной активностью по превращению О-фосфо-L-гомосерина и сульфида в L-гомоцистеин + Н3РО4. Предпочтительно, сульфид представляет собой металл, такой как сульфид Na2S. В этом случае превращение происходит в соответствии со следующей схемой реакции:
О-фосфо-L-гомосерин + Na2S <=> L-гомоцистеин + Н3РО4
Активность по превращению О-фосфо-L-гомосерина (OPHS) и сульфида металла в L-гомоцистеин и H3PO4, может быть выявлена с помощью анализа, который является в основном таким же, как описано выше в связи с детекцией активности по превращению О-фосфо-L-гомосерина (OPHS) и метантиола в L-метионин и Н3РО4. В этом случае, используют met2Δ met25Δ двойной мутантный штамм дрожжей и его выращивают на среде с Na2S в качестве единственного источника сульфата. В самом деле, МЕТ25 кодирует единственную, известную в дрожжах, гомоцистеин-синтазную активность, катализирующую конденсацию О-ацетил-L-гомосерина с сульфидом. Чувствительность анализа можно легко повысить, используя штаммы S. cerevisiae, сконструированные таким образом, чтобы, в дополнение к двойным мутациям met2 met25, они включали в себя, мутацию thr4, приводящую к накоплению O-фосфо-L-гомосерина.
Эта ферментативная активность также может быть дополнительно подтверждена с помощью анализа in vitro, в котором O-фосфо-L-гомосерин и Na2S инкубируют in vitro в подходящих условиях с бесклеточным экстрактом из штамма дрожжей, экспрессирующего белок, который будет протестирован, или с (частично) очищенным белком, который будет протестирован, и в котором производство гомоцистеина определяют с помощью колориметрического метода (Becker et al., Journal of Biochemical Chemistry 244 (1969), 2418). Настоящее изобретение также относится к молекулам нуклеиновых кислот, кодирующих белок по изобретению. Молекулой нуклеиновой кислоты может быть молекула ДНК или РНК, предпочтительно, молекула ДНК.
Настоящее изобретение также относится к вектору, содержащему молекулу нуклеиновой кислоты по изобретению. В предпочтительном варианте осуществления, вектор представляет собой вектор, обеспечивающий экспрессию нуклеиновой кислоты по изобретению так, чтобы осуществлялась продукция белка по изобретению. Таким образом, в предпочтительном варианте осуществления изобретения, нуклеиновая кислота по изобретению, функционально связана с последовательностями, контролирующими экспрессию, которые обеспечивают возможность экспрессии в требуемой системе клетки-хозяина или в клетке-хозяине. Термин "функционально связанный", так как он использован в настоящем описании, относится к связи между одной или более последовательностями, контролирующими экспрессию, и кодирующей областью в молекуле нуклеиновой кислоты, которая будет экспрессирована, таким образом, что экспрессия достигается в условиях, совместимых с последовательностью, контролирующей экспрессию.
Экспрессия включает транскрипцию гетерологичной последовательности ДНК, предпочтительно, в транслируемую мРНК. Регуляторные элементы, контролирующие экспрессию в клетках растений, животных, грибов, а также бактерий хорошо известны специалистам в данной области техники. Они включают промоторы, энхансеры, сигналы терминации, сигнальные последовательности и тому подобное. Примеры, вместе с подробными пояснениями относительно векторов, приведены ниже.
Промоторы для использования вместе с молекулой нуклеиновой кислоты могут быть гомологичными или гетерологичными по своему происхождению и/или по отношению к гену, который будет экспрессирован. Подходящими промоторами являются, например, промоторы, контролирующие конститутивную экспрессию. Тем не менее, промоторы, активирующиеся только в ответ на внешние воздействия, также могут быть использованы. В данном контексте могут быть использованы искусственные и/или химически индуцируемые промоторы.
Вектор по настоящему изобретению может быть введен в (микро)организм с тем, чтобы быть экспрессированным и привести к продуцированию белка, способного превращать O-фосфо-L-гомосерин и метантиол в L-метионин.
Обзор различных экспрессионных систем приведен, например, в Methods in Enzymology 153 (1987), 385-516, in Bitter et al. (Methods in Enzymology 153 (1987), 516-544) и in Sawers et al. (Applied Microbiology and Biotechnology 46 (1996), 1-9), Billman-Jacobe (Current Opinion in Biotechnology 7 (1996), 500-4), Hockney (Trends in Biotechnology 12 (1994), 456-463), Griffiths et al. (Methods in Molecular Biology 75 (1997), 427-440). Обзор экспрессионных систем дрожжей, например, сделан Hensing et al. (Antonie van Leuwenhoek 67 (1995), 261-279), Bussineau et al. (Developments in Biological Standardization 83 (1994), 13-19), Gellissen et al. (Antonie van Leuwenhoek 62 (1992), 79-93, Fleer (Current Opinion in Biotechnology 3 (1992), 486-496), Vedvick (Current Opinion in Biotechnology 2 (1991), 742-745) и Buckholz (Bio/Technology 9 (1991), 1067-1072).
Экспрессионные векторы были широко описаны в литературе. Как правило, они содержат не только ген селективного маркера и ориджин репликации (точку начала репликации), обеспечивающие репликацию в выбранном хозяине, но и бактериальный или вирусный промотор, и, в большинстве случаев, сигнал терминации транскрипции. Между промотором и сигналом терминации транскрипции существует, как правило, по меньшей мере, один сайт рестрикции или полилинкер, обеспечивающий возможность вставки кодирующей последовательности ДНК. В качестве промоторной последовательности может быть использована последовательность ДНК, контролирующая в природе транскрипцию соответствующего гена, если она активна в выбранном организме-хозяине. Тем не менее, эта последовательность может также быть заменена на другие промоторные последовательности. Можно использовать конститутивные промоторы, контролирующие конститутивную экспрессию гена, а также индуцируемые промоторы, предусматривающие планомерный контроль экспрессии гена. Бактериальные и вирусные промоторные последовательности, обладающие этими свойствами, подробно описаны в литературе. Последовательности, регулирующие экспрессию в микроорганизмах (например, E.coli,: S. cerevisiae) достаточно подробно описаны в литературе. Промоторами, обеспечивающими наиболее высокую экспрессию расположенной ниже последовательности, являются, например, промотор Т7 (Studier et al., Methods in Enzymology 185 (1990), 60-89), lacUV5, trp, trp-lacllV5 (DeBoer et al., В Rodriguez and Chamberlin (Eds), Promoters, Structure and Function; Praeger, NY, (1982), 462-481; DeBoer et al., Proc Natl Acad Sci USA (1983), 21-25), Ip1, rae (Boros et al., Gene 42 (1986), 97-100). Индуцируемые промоторы, предпочтительно, используют для синтеза полипептидов. Эти промоторы часто обеспечивают более высокий выход полипептидов, чем конститутивные промоторы. Для того чтобы получить оптимальное количество полипептида, часто используют двухэтапный процесс. Сначала клетки-хозяева культивируют в оптимальных условиях до сравнительно высокой клеточной плотности. На втором этапе, индуцируют транскрипцию в зависимости от типа используемого промотора.
В этом отношении, наиболее подходящим является tac промотор, который может быть индуцирован лактозой или IPTG (изопропил - бета-D-тиогалактопиранозид) (DeBoer et al., Proc. Natl. Acad. Sci. USA 80 (1983), 21-25). Сигналы терминации транскрипции также описаны в литературе.
Трансформация клетки-хозяина полинуклеотидом или вектором по изобретению может быть осуществлена стандартными методами, как описано, например, в Sambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USA; Methods in Yeast Genetics, A Laboratory Course Manual, Cold Spring Harbor Laboratory Press, 1990. Клетку-хозяина культивируют в питательной среде, соответствующей требованиям для конкретной клетки-хозяина, в частности, в отношении величины рН, температуры, концентрации соли, аэрации, антибиотиков, витаминов, микроэлементов и т.д.
Настоящее изобретение также относится к клетке-хозяину, которая содержит/трансформирована молекулой нуклеиновой кислоты или вектором по изобретению. В предпочтительном варианте такая клетка-хозяин экспрессирует белок по изобретению и способна превращать O-фосфо-L-гомосерин и метантиол в L-метионин. В принципе, клеткой-хозяином может быть любая клетка-хозяин, например, клетка животного, растения, гриба, бактерии. В предпочтительном варианте осуществления, клетка-хозяин представляет собой клетку бактерии или гриба. В более предпочтительном варианте осуществления, клетка-хозяин представляет собой клетку бактерии, например, из рода Escherichia, Corynebacterium, Clostridium, Bacillus или Acinetobacter, наиболее предпочтительными из этих видов являются Е. coli, Corynebacterium glutamicum, Bacillus subtilis или Acinetobacter villandi.
В другом предпочтительном варианте клетка-хозяин представляет собой клетку гриба, например, из рода Saccharomyces, Candida, Ashbya, Kluyveromyces, Pichia, Yarrowia, Zygosaccharomyces, Aspergillus, Debaryomyces или Torulopsis, более предпочтительно, из рода: S.cerevisiae, Saccharomyces maximus, Candida Maltosa, Ashbya gossypii, Kluveromyces Lactis, Pichia pastoris Pichia stipitis, Yarrowia lipolitica, Aspergillus Niger, Aspergillus nidullans, Debaryomyces hansenii или Torulopsis utilis. В более предпочтительном варианте осуществления клетка-хозяин представляет собой дрожжевую клетку.
Как указано выше, настоящее изобретение также относится к способу получения молекулы нуклеиновой кислоты, кодирующей белок, способный превращать О-фосфо-L-гомосерин и метантиол в L-метионин. Такой способ включает следующие стадии:
(i) осуществление мутации в молекуле нуклеиновой кислоты, кодирующей цистатионин-гамма-синтазу (КФ 2.5.1.48) таким образом, чтобы получить мутантные цистатионин-гамма-синтазы;
(ii) экспрессию мутантных молекул нуклеиновой кислоты, полученных на стадии (i) в клетке-хозяине в условиях культивирования, позволяющих отбирать молекулы нуклеиновых кислот, кодирующие белок, способный ферментативно превращать О-фосфо-L-гомосерин и метантиол в L-метионин;
(iii) идентификацию тех клеток-хозяев, которые экспрессируют белок, способный ферментативно превращать О-фосфо-L-гомосерин и метантиол в L-метионин; и
(iv) получение из клетки-хозяина, идентифицированной на стадии (iii), молекулы нуклеиновой кислоты, кодирующей мутированную цистатионин-гамма-синтазу, способную ферментативно превращать О-фосфо-L-гомосерин и метантиол в L-метионин.
Настоящее изобретение также относится к способу получения белка, способного ферментативно превращать О-фосфо-L-гомосерин и метантиол в L-метионин, причем указанный способ включает в себя стадию экспрессии молекулы нуклеиновой кислоты, полученной в соответствии с описанным выше способом по изобретению, и выделение кодированного белка.
Термин "цистатионин-гамма-синтаза (КФ 2.5.1.48)" относится к любому ферменту, имеющему ферментативную активность цистатионин-гамма-синтазы. Цистатионин-гамма-синтаза представляет собой фермент, который катализирует следующую реакцию
O4-сукцинил-L-гомосерин + L-цистеин <=> L-цистатионин + сукцинат
Эта активность может быть измерена с помощью методов, известных в данной области техники (Ravanel, Biochem. J. 331 (1998), 639-648).
В соответствии со стадией (i) описанного выше способа по изобретению, мутированный вариант молекулы нуклеиновой кислоты, кодирующей цистатионин-гамма-синтазу, сделан таким образом, чтобы получить мутантную цистатионин-гамма-синтазу. Методы мутагенеза молекул нуклеиновых кислот, хорошо известны специалистам в данной области техники. Таким образом, можно вставить различные типы мутаций в молекулы нуклеиновых кислот методами, обычно используемыми в молекулярной биологии (смотри, например, Sambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USA), что приведет к синтезу цистатионин-гамма-синтазы с модифицированной аминокислотной последовательностью по сравнению с исходной последовательностью. В принципе, возможен любой тип мутации на уровне молекулы нуклеиновой кислоты, например, делеции, вставки и/или замены. В предпочтительном варианте мутация, осуществленная в нуклеиновой кислоте, приводит к замене аминокислоты на уровне аминокислотных последовательностей.
Можно осуществить только одну мутацию в молекуле нуклеиновой кислоты, но также, конечно, можно осуществить одну или несколько мутаций в молекуле нуклеиновой кислоты, ведущих к одной или более мутаций на уровне аминокислот. В принципе, не существует верхнего предела по количеству мутаций. Тем не менее, предпочтительно, чтобы мутантная молекула нуклеиновой кислоты имела последовательность, идентичную по меньшей мере на 60%, более предпочтительно по меньшей мере на 70%, еще более предпочтительно по меньшей мере на 80%, еще более предпочтительно по меньшей мере на 90%, и наиболее предпочтительно по меньшей мере на 95% исходной последовательности, кодирующей цистатионин-гамма-синтазу, используемую на стадии (i) описанного выше способа.
В предпочтительном варианте, мутантная молекула нуклеиновой кислоты, полученная на стадии (i), кодирует мутантный вариант цистатионин-гамма-синтазы, который имеет не более 20 изменений в аминокислотных остатках по сравнению с цистатионин-гамма-синтазой, кодируемой исходной нуклеиновой кислотой, более предпочтительно не более 15 изменений, еще более предпочтительно не более 10 изменений и наиболее предпочтительно не более 5 изменений.
Термин "молекула нуклеиновой кислоты, кодирующая цистатионин гамма-синтазу (КФ 2.5.1.48)" на стадии (i) способа по изобретению, относится к молекуле нуклеиновой кислоты, кодирующей белок, обладающий ферментативной активностью цистатионин-гамма-синтазы, как указано выше. В принципе, любая молекула нуклеиновой кислоты, кодирующая такой фермент, может быть использована в способе по изобретению в качестве исходного материала. Цистатионин-гамма-синтазы были подробно описаны выше, также как и их распространение в природе. Любая нуклеиновая кислота, кодирующая любую из этих цистатионин-гамма-синтаз, может быть использована на стадии (i) способа по изобретению. В предпочтительном варианте осуществления, молекула нуклеиновой кислоты, используемая на стадии (i) представляет собой молекулу нуклеиновой кислоты, кодирующую растительную цистатионин-гамма-синтазу. Еще более предпочтительно, цистатионин-гамма-синтаза происходит из A. thaliana, предпочтительно нуклеотидная последовательность, кодирующая цистатионин-гамма-синтазу A. thaliana, такая каквЕО. ID NO: 1.
Нуклеотидная последовательность молекулы нуклеиновой кислоты, используемой на стадии (i) может быть адаптирована для использования в клетке-хозяине, которую используют на стадии (ii). Например, можно изменить частоту встречаемости кодонов с тем, чтобы в большей степени соответствовать частоте встречаемости кодонов трансляционной машины используемых клеток-хозяев. Таким образом, молекула нуклеиновой кислоты может быть модифицирована таким образом, чтобы она имела оптимальную частоту встречаемости кодонов по отношению к клетке-хозяину, используемой на стадии (ii).
Кроме того, могут быть необходимы или желательны другие модификации, чтобы адаптировать некоторые молекулы нуклеиновых кислот, такие как молекула нуклеиновой кислоты, кодирующая растительную цистатионин-гамма-синтазу, к способу по изобретению. Например, растительные цистатионин-гамма-синтазы, как правило, содержат на N-конце хлоропластную сигнальную последовательность, которая направляет белок в хлоропласты. В зависимости от того, какую клетку-хозяина используют на стадии (и), может быть необходимо или желательно удаление этой сигнальной последовательности. Если, например, на стадии (i) используют дрожжевые клетки, то желательно удалить эту хлоропластную сигнальную последовательность. Кроме того, растительная цистатионин-гамма-синтаза может содержать регуляторную область (также известную как mto область), которая обеспечивает регуляцию трансляции, опосредованную увеличением внутриклеточного пула S-аденозилметионина (SAM). SAM способен связываться с образующимся пептидом цистатионин-гамма-синтазы во время трансляции в кармане, образованной этой mto областью и частью рибосомы, что приводит к остановке трансляции. Может быть желательным избежать этого регулирования при применении способа по изобретению. Таким образом, в предпочтительном варианте, на стадии (i) используют последовательность, кодирующую растительную цистатионин-гамма-синтазу, модифицированную таким образом, чтобы избежать регулирования SAM. Такие мутации были описаны в предшествующем уровне техники, например, в Onoue et al. (Journal of Biological Chemistry 286 (2011), 14903-14911).
Дополнительно, также может быть возможным использование в качестве исходной молекулы нуклеиновой кислоты, последовательности, которая приводит к более стабильной версии цистатионин-гамма-синтазы. Известно, что идентичность второй N-концевой аминокислоты, определяет стабильность всего белка (Varshavsky, Annual Review of Biochemistry 81 (2012), 167-176).
Таким образом, целесообразно, избегать дестабилизирующих остатков (в частности, Туг, Gin, Leu, Phe, Asp, Lys, Arg) и предпочесть стабилизирующие остатки, такие как Val или Ser (Varshavsky, Annual review of Biochemistry 81 (2012), 167-176).
Экспрессия мутантной молекулы нуклеиновой кислоты согласно стадии (ii) способа может быть достигнута способами, известными специалисту в данной области техники. Экспрессионные системы также уже были описаны выше в связи с векторами и клетками-хозяевами по изобретению. На стадии (ii) способа по изобретению используют клетку-хозяина для экспрессии мутантной цистатионин-гамма-синтазы, что позволяет отбирать молекулы нуклеиновых кислот, кодирующие белок, способный ферментативно превращать О-фосфо-L-гомосерин и метантиол в L-метионин. В этом контексте, например, можно использовать клетки-хозяева, которые могут выжить только при выращивании на среде, содержащей метантиол, в качестве единственного источника серы, если они способны превращать О-фосфо-L-гомосерин и метантиол в L-метионин. Одним из примеров таких клеток-хозяев являются клетки Saccharomyces cerevisiae, которые были генетически модифицированы, так, чтобы быть, по меньшей мере, лишенными функции met2. Ген МЕТ2 кодирует гомосеринтрансацетилазу, катализирующую превращение гомосерина и ацетил-КоА в О-ацетил гомосерин и КоА. Инактивация гена МЕТ2 приводит к появлению дрожжевых штаммов, имеющих ауксотрофный фенотип по метионину. Предпочтительно, чтобы в штамме дрожжей, с инактивированным геном МЕТ2, ген met6 также был инактивирован. Ген МЕТ6 кодирует гомоцистеинметилтрансферазу, превращающую гомоцистеин и метилтетрагидрофолат в метионин. Поскольку S. cerevisiae, не имеет иных способов синтеза незаменимой аминокислоты метионина, кроме как ее синтез из гомоцистеина, штамм S. cerevisiae, лишенный функции МЕТ2 (и, возможно, также функции МЕТ6) может выжить только, если он способен производить метионин другими способами.
Условия культивирования на стадии (ii) подобраны таким образом, что клетка-хозяин может выжить, только если она способна превращать О-фосфо-L-гомосерин и метантиол в L-метионин. Это может быть достигнуто путем добавления метантиола в качестве единственного источника серы в культуральную среду.
Еще более предпочтительно, чтобы в дополнение к генам МЕТ2 и МЕТ6 был также инактивирован ген THR4. Такая инактивация нацелена на повышение внутриклеточного пула OPHS. В этом случае трансформированные клетки-хозяева выращивают в среде, содержащей и треонин и метантиол вместо метионина. Если используется такая клетка-хозяин, то возможно выделить мутанты, обладающие очень слабой активностью OPHS-зависимой метионинсинтазы.
Клетки-хозяева, идентифицированные на стадии (iii) способа по изобретению, в дальнейшем используют для получения молекул нуклеиновых кислот, которые кодируют мутированную цистатионин-гамма-синтазу, способную ферментативно превращать O-фосфо-L-гомосерин и метантиол в L-метионин. Это может быть сделано методами, известными специалистам в данной области техники.
Выделенная молекула нуклеиновой кислоты может быть затем использована для экспресии соответствующего фермента, или она может быть введена в другие клетки-хозяева.
В предпочтительном варианте осуществления, способ получения молекулы нуклеиновой кислоты, кодирующей белок, способный превращать О-фосфо-L-гомосерин и метантиол в L-метионин, включает несколько раундов скрининга, то есть мутаций и выделения соответствующих мутантов, обладающих необходимой активностью.
В этом контексте, предпочтительно, чтобы в первом раунде скрининга использовали клетку-хозяина, в которой гены МЕТ2, МЕТ6 и THR4 инактивированы. Как описано выше, с использованием такой клетки-хозяина, можно отобрать мутанты, обладающие очень слабой активностью OPHS-зависимой метионинсинтазы. Отобранные мутанты подвергают дальнейшему раунду мутагенеза и впоследствии отбирают по OPHS-зависимой метионинсинтазной активности в клетке-хозяине, в которой инактивированы только гены МЕТ2 и МЕТ6, а ген thr4 активен. Такой отбор является более жестким, так как часть OPHS используется треонинсинтазой. В этом случае клетки-хозяева, используемые для отбора клонов с желательными ферментативными свойствами, выращивают в среде, содержащей метантиол в качестве источника серы.
Второй раунд отбора может быть выполнен итеративно и давление отбора может быть увеличено за счет уменьшения количества метантиола, добавленного к среде, и отбора трансформированных штаммов, демонтстрирующих быстрый рост.
Фигура 1 показывает упрощенную схему пути биосинтеза метионина в S.cerevisiae (взято из: Thomas D. and Y. Surdin-Kerjan; Microbiological and Molecular Reviews 61 (1997); Metabolism of sulfur amino-acids in Saccharomyces cerevisiae, page 503-532).
Фигура 2 показывает химический путь синтеза О-фосфо-L-гомосерина.
Фигура 3 схематически показывает укороченые формы CGS1, используемые в качестве исходного материала для создания мутантных версий фермента.
Фигура 4 показывает рост штаммов YA246-4A, экспрессирующих или AD242, или AD328 CGS1 вариант, или осуществляющих контроль вектора pAL06 в среде А, содержащей 0,05 мМ метионина.
Фигура 5 показывает рост штаммов на основе YA246-4A, экспрессирующих CGS1-4 (A), CGS1-5 (В) и CGS1-1 (С) мутантные семейства, соответственно, в среде А, содержащей 0,1 мМ метантиола. CGS1-4 (G84S) и отрицательные контроли с pAL06 пустым вектором показаны на всех графиках.
Фигура 6 показывает рост штаммов на основе YA247-5A, экспрессирующих CGS1-4 семейство мутантов AD242, AD328, MUT24 и MUT27 в среде А, содержащей 1 мМ (А) или 0,1 мМ (В) метантиола. CGS1-4 (G84S) и отрицательные контроли с pAL06 показаны на всех графиках.
Фигура 7 показывает сравнение идентифицированных мутантов CGS.
Описание последовательностей:
SEQ ID NO: 1 аминокислотная последовательность цистатионин-гамма-синтазы (КФ 2.5.1.48) CGS1 из Arabidopsis thaliana (ААС25687.1).
SEQ ID NO: 2 аминокислотная последовательность цистатионин-гамма-синтазы (КФ 2.5.1.48) CGS1 из Arabidopsis thaliana (ААС25687.1), в которой глицин 84 заменен на серии. Эту последовательность также называют CGS1 G84S или CGS1-0.
SEQ ID NO: 3 аминокислотная последовательность цистатионин-гамма-синтазы (КФ 2.5.1.48) CGS1w3 Arabidopsis thaliana (ААС25687,1), в которой глицин 84 заменен на серии, в которой удалены 57 N-концевых аминокислот и добавлен остаток метионина на N-конце. Эту последовательность также называют CGS1 1-4G84S.
SEQ ID NO: 4 укороченная форма аминокислотной последовательности цистатионин-гамма-синтазы (КФ 2.5.1.48) CGS1 из Arabidopsis thaliana (ААС25687.1), в которой 88 N-концевых аминокислот удалены, и остаток метионина добавлен на N-конце. Эту последовательность также называют CGS1 1-5G84S.
SEQ ID NO: 5 укороченная форма аминокислотной последовательности цистатионин-гамма-синтазы (КФ 2.5.1.48) CGS1 из Arabidopsis thaliana (ААС25687.1), в которой 160 N-концевых аминокислот удалены, и остаток метионина добавлен на N-конце. Эту последовательность также называют CGS1 1-1 G84S.
SEQ ID NO: 6 последовательность мутанта MUT02.
SEQ ID NO: 7 последовательность мутанта MUT04.
SEQ ID NO: 8 последовательность мутанта MUT13.
SEQ ID NO: 9 последовательность мутанта MUT18.
SEQ ID NO: 10 последовательность мутанта MUT19.
SEQ ID NO: 11 последовательность мутанта AD309.
SEQ ID NO: 12 последовательность мутанта AD310.
SEQ ID NO: 13 последовательность мутанта AD311.
SEQ ID NO: 14 последовательность мутанта AD312.
SEQ ID NO: 15 последовательность мутанта AD313.
SEQ ID NO: 16 последовательность мутанта AD242.
SEQ ID NO: 17 последовательность мутанта AD328.
SEQ ID NO: 18 последовательность мутанта AD329.
SEQ ID NO: 19 последовательность мутанта MUT24.
SEQ ID NO: 20 последовательность мутанта MUT27.
SEQ ID NO: 21 последовательность мутанта MUT67.
SEQ ID NO: 22 последовательность мутанта MUT68.
SEQ ID NO: 23 последовательность мутанта MUT70.
SEQ ID NO: 24 последовательность мутанта MUT71.
SEQ ID NO: 25 последовательность мутанта MUT72.
SEQ ID NO: 26 последовательность мутанта MUT74.
SEQ ID NO: 27 последовательность мутанта MUT75.
SEQ ID NO: 28 последовательность мутанта MUT78.
SEQ ID NO: 29 последовательность мутанта MUT79.
Содержание документов, приведенных в данной заявке, включено посредством ссылки во всей своей полноте.
Другие аспекты и преимущества настоящего изобретения будут описаны в следующих примерах, которые приведены с целью иллюстрации, а не в качестве ограничения.
Примеры
Пример 1: Получение укороченных форм CGS1, использованных в качестве исходного материала для получения мутантов
Ген CGS1-4 является синтетическим геном. Синтез проводили с помощью MWG Eurofins оперона (Ebersberg) с использованием алгоритма GENEius с целью оптимизации частоты встречаемости кодонов для улучшения экспрессии гена в S. cerevisiae. Таблица частоты встречаемости кодонов S.cerevisiae была взята из базы данных частоты встречаемости кодонов Kazusa (http://www.kazusa.or.jp/codon). Укороченные формы CGS1-5 и CGS1-1 были подготовлены методом ПЦР с использованием CGS1-4 в качестве матрицы, с прямыми олигонуклеотидами GTACCGCTCGAGATGGTTGCTGGTAAGTGGTCTAACAATC для CGS1-5 и GTACCGCTCGAGATGTCTGTTCAATTGACCGATTCTAAG для CGS1-1. Для обеих форм CGS1-5 и CGS1-1, был использован одинаковый обратный олигонуклеотид AGTACGGGATCCTCAAATGGCTTCCA. Описанные укороченные модифицированные формы CGS1 схематически показаны на Фигуре 3.
Пример 2: Получение мутантов укороченных форм CGS1
Укороченные формы генов CGS1 были клонированы в плазмиду pAL06, дрожжевой репликативный вектор получен из вектора pRS316 (Sikorski RS & Hieter П., Genetics 1989, 122: 19-27). Плазмида pAL06 обеспечивает возможность экспрессии клонированного гена под контролем сильного дрожжевого промотора TEFL. CGS1 библиотеки, были генерированы путем гипермутагенной ПЦР с разными концентрациями дезоксинуклеотидтрифосфатов (dNTP) с использованием протокола, описанного Vartanian JP et al., (Nucleic Acids Res. 1996 24: 2627-2631).
Оба различия в концентрациях: [дТТФ] больше [дЦТФ] и [дГТФ] больше [дАТФ] были использованы в мутагенных ПЦР. Либо [дТТФ]/[дЦТФ]=[дГТФ]/[дАТФ]=1000 мкМ/200 мкМ или [дТТФ]/[дЦТФ]=[дГТФ]/[дАТФ]=1000 мкМ/150 мкМ были использованы в присутствии 0,5 мМ MnCl2.
Пример 3: Скрининг мутантов, которые могут производить L-метионин из О-фосфо-L-гомосерина и метантиола
Штаммы дрожжей YA247-5A (МАТ-α, ade2, his3, Ieu2, met2::loxP, met6::HIS3, trp1, ura3) или YA246-4A (МАТ-α, ade2, his3, Ieu2, met2::loxP, met6::HIS3, thr4::loxP, trp1, ura3) были трансформированы CGS1 библиотеками в минимальной среде, содержащей метионин, но не содержащей урацил, чтобы отобрать маркер плазмиды. После выращивания клеток в течение 48 часов клетки собирали, промывали и ресуспендировали в минимальной жидкой среде с метилмеркаптаном в качестве источника серы. Через 7 дней, культуры собирали и разводили в той же жидкой среде, до значения оптической плотности OD(оптическая плотность)600 равного приблизительно 0,2. Проводили три последовательных цикла разведения.
Плазмиды, присутствующие в полученных в результате растущих дрожжевых клетках, были затем экстрагированы, амплифицированы в E.coli, и полученная ДНК использована для повторной трансформации штаммов YA247-5А (МАТ-α, ade2, his3, Ieu2, met2::loxP, met6::HIS3, trp1, ura3) или YA246-4A (MAT-α, ade2, his3, Ieu2, met2::loxP, met6::HIS3, thr4::loxP, trp1, ura3). Затем трансформированные дрожжи были отобраны на твердой минимальной среде, содержащей метионин, но не содержащей урацил. Способность каждой колонии расти с метилмеркаптаном в качестве источника серы затем оценивали при выращивании отдельных клонов в жидкой среде, содержащей метантиол в качестве источника серы. В каждом цикле, плазмиды, содержащиеся в дрожжевых колониях, демонстрирующих лучшие темпы роста в присутствии метантиола, экстрагировали и секвенировали. Отобранными мутантами был трансформирован штамм YA247-5A для in vitro анализа, и мутанты были использованы в качестве исходных материалов для новой гипермутагенной ПЦР.
С использованием описанной выше процедуры скрининга (отбора) было выявлено большое количество мутантов CSG1, которые были способны расти на метантиоле в качестве единственного источника серы, и которые, следовательно, как предполагают, имеют возможность конвертировать О-фосфо-L-гомосерин и метантиол в L-метионин. Были определены последовательности этих мутантов, и мутанты суммированы в следующей таблице 1А, где позиции мутаций указаны со ссылкой на последовательность SEQ ID NO: 3 (CGS 1-4). Мутанты также приведены на Фигурах 7A-D, в которых они сравниваются.
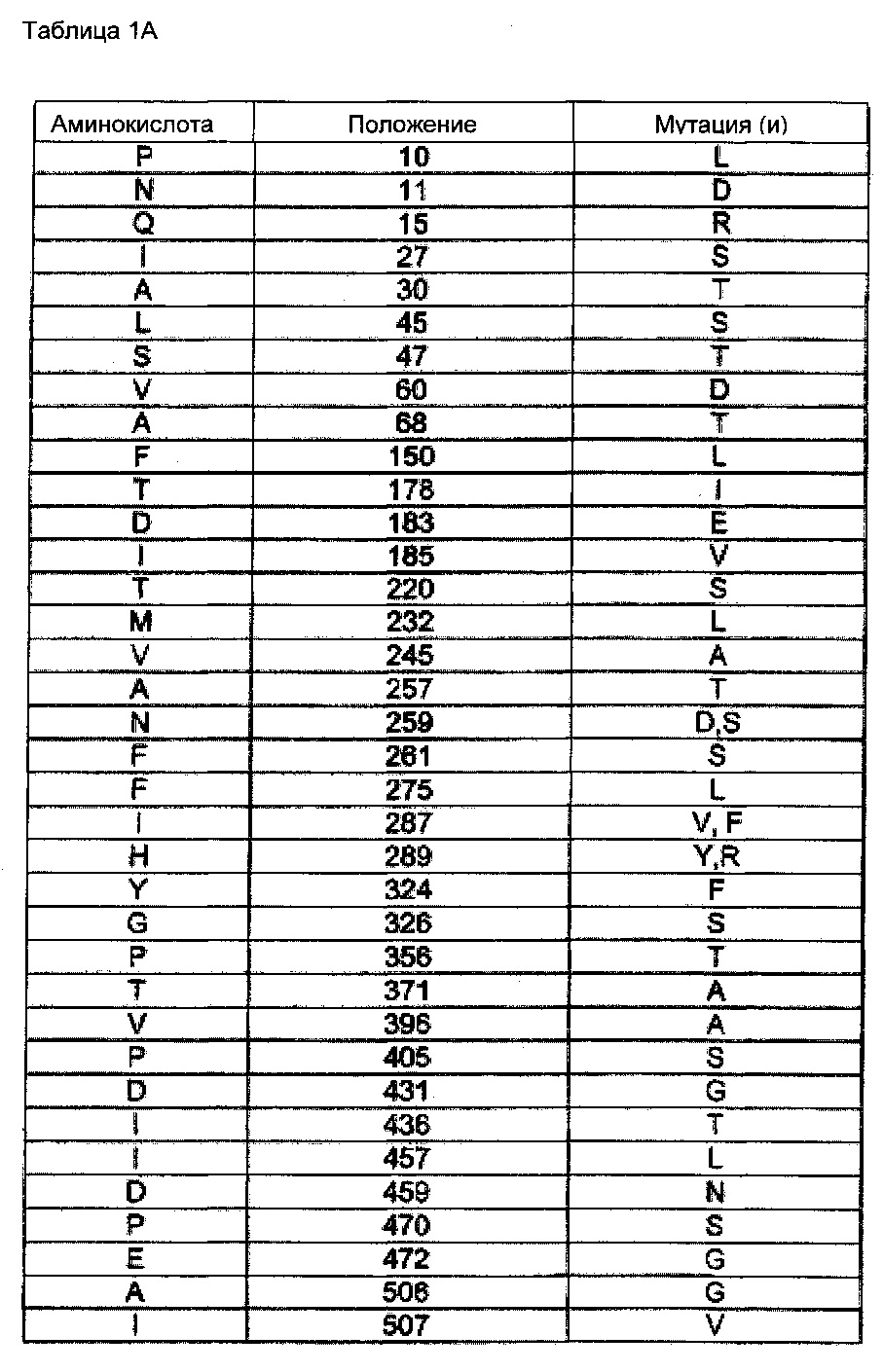
Следующая таблица 1В перечисляет все проанализированные мутанты и указывает положения найденных мутаций по отношению к полной длине последовательности SEQ ID NO: 1 (CGS1). Положение по отношению к соответствующей исходной последовательности указано в скобках.
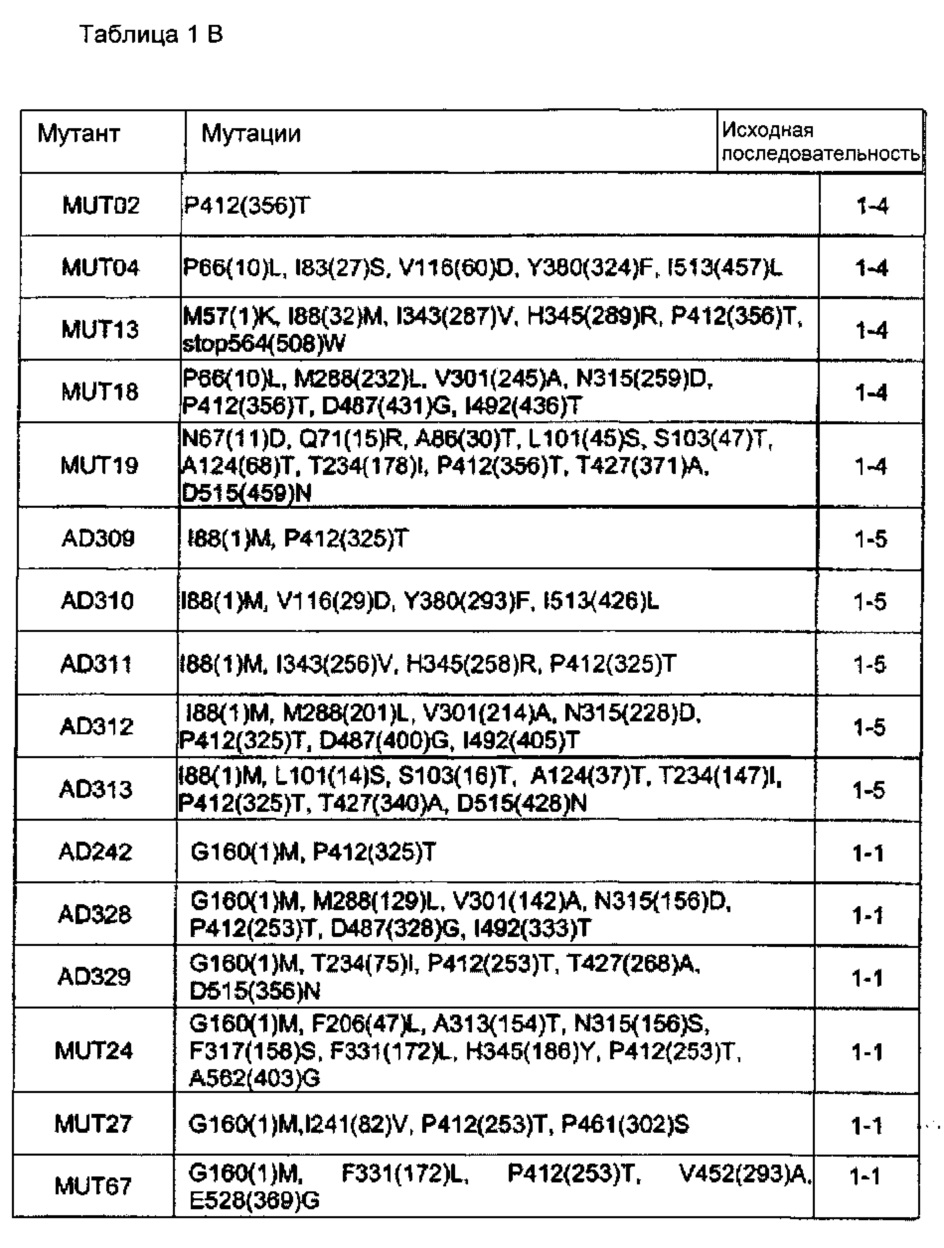

Исходная последовательность 1-4 соответствует SEQ ID NO: 3.
Исходная последовательность 1-5 соответствует SEQ ID NO: 4.
Исходная последовательность 1-1 соответствует SEQ ID NO: 5.
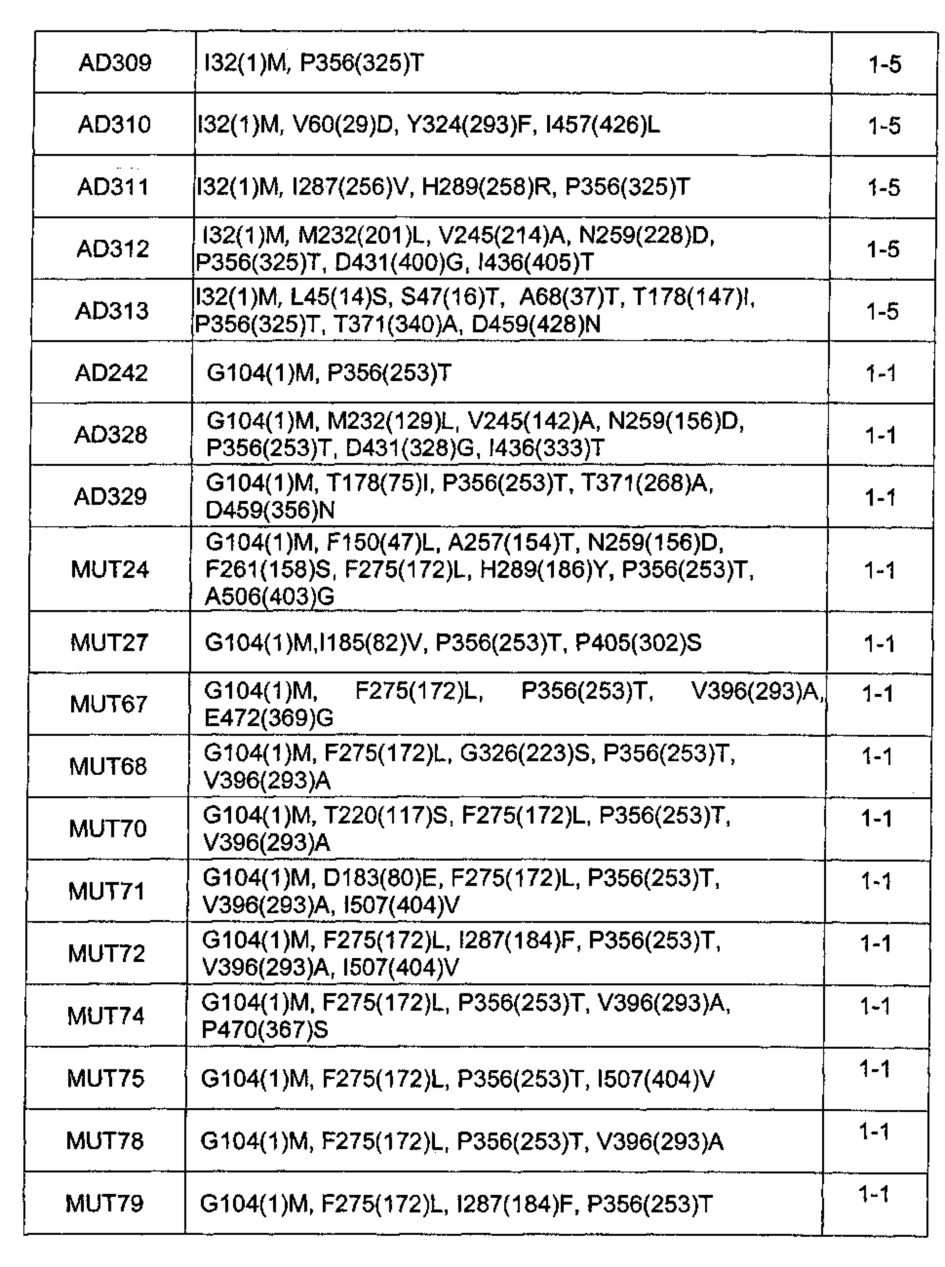
В следующей таблице 1С перечислены все проанализированные мутанты, и указаны положения обнаруженных мутаций по отношению к последовательности SEQ ID NO: 3 (CGS1-4). Положение по отношению к соответствующей исходной последовательности указано в скобках.

Пример 4:
Тест на ферментативную активность у дрожжей Saccharomyces cerevisiae Экспрессия любого из описанных выше мутантов в штаммах Saccharomyces cerevisiae YA247-5A (ΜΑΤ-α, ade2, his3, Ieu2, met2::loxP, met6::HIS3, trp1, ura3) и YA246-4A (ΜΑΤ-α, ade2, his3, Ieu2, met2::loxP, met6::HIS3, thr4::loxP, trp1, ura3) избавляет штаммы от их ауксотрофии по метионину.
Другими словами, при экспрессии любого из указанных выше мутантов, дрожжевые штаммы YA247-5A и YA246-4A, дефектные по синтезу метионина, растут на минимальной среде с добавлением аденина, гистидина, лейцина, триптофана и урацила.
Экспериментальный дизайн теста:
Мутантные нуклеотидные последовательности были индивидуально клонированы в репликативную плазмиду pAL06 (производную от pRS316) ниже промотора транскрипции (pTEF1) и выше терминатора транскрипции (tADH1).
Полученные таким образом двенадцать плазмид, индивидуально трансформировали в дрожжевые штаммы YA247-5A и YA246-4A. Трансформанты выращивали в среде A (Difco™ азотистое основание дрожжей 6,7%, глюкоза 2%, аденин 0,3 мМ, лейцин 0,75 мМ, гистидин 1,3 мМ, триптофан 0,1 мМ), дополненной 0,2 мМ метионином.
Каждый из трансформантов затем засевали (OD590=0,015) в среду А, содержащую 0,5 мМ метионина, или среду А, содержащую 0,1 или 1 мМ метантиола.
Рост контролировали последующим измерением оптической плотности при 590 нм (OD590). Соответствующий рост каждого клона в двух средах сравнивали с ростом отрицательных контролей YA246-4A или YA247-5A, трансформированных пустым вектором pAL06. В среде А с метионином, все проверенные штаммы имеют время генерации около 4 часов. Например, рост штамма YA246-4A, экспрессирующего мутанты CGS 1-4 (G84S), AD242, AD328, и контрольный вектор pAL06 показаны на Фигуре 4.
Рост в контексте накопления фосфогомосерина:
Рост штаммов на основе YA246-4A, которые накапливают фосфогомосерин и экспрессируют CGS1 белковые мутанты в среде А, содержащей 0,1 мМ метантиол, показан на Фигуре 5.
Во всех случаях, никакого роста не наблюдалось в контроле CGS1-4 (G84S) и для штаммов с плазмидой pAL06.
Для семейства CGS1-4 (Фигура 5А), время генерации составляет от 13 часов для MUT19 до 42 часов для MUT02. Для семейства CGS1-5 (Фигура 5В), время генерации составляет от 9.5 часов для AD312 и AD313 до 12.5 часов для AD310. Для семейства CGS1-1 (Фигура 5С), время генерации составляет около 9.5 часов для AD242, AD328 и AD329.
Рост в контексте ненакопления фосфогомосерина:
Рост штаммов на основе YA247-5A, которые не накапливают фосфогомосерин и экспрессируют CGS1 белковые мутанты в среде А, содержащей 1 мМ метантиол или 0,1 мМ метантиол, показан на Фигурах 6А и 6В соответственно.
С 1 мМ метантиолом не наблюдалось никакого роста для CGS1-1 и для отрицательного контроля, трансформированного пустым вектором pAL06. Время генерации составляет, проблизительно, 21 ч для AD242 или AD328, 10,5 часов для MUT27 и 8 часов для MUT24. С 0,1 мм метантиолом рост наблюдается только с мутантами MUT24 и MUT27. Время генерации составляет соответственно 19 ч и 40 ч.
Пример 5: In vitro активность OPHS-зависимой метионинсинтазы
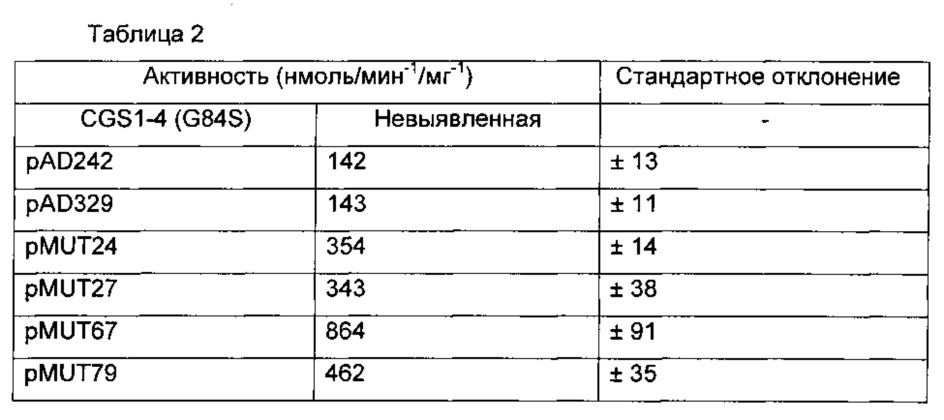
In vitro активность фермента, экспрессирующегося в дрожжах, была протестирована в неочищенном лизате дрожжей.
Вслед за проверкой активности осуществлялся мониторинг синтеза метионина в лизате, в присутствии O-фосфо-L-гомосерина (OPHS) и метантиола (CH3SNa). Лизаты клеток дрожжей YA246-5A, YA246-5A, несущих пустую плазмиду pAL06, YA246-5A, экспрессирующего CGS1-4 и YA246-5A, экспрессирующиего мутанты AD246, AD239, MUT24, MUT27, MUT67 или MUT79 сравнили.
Экспериментальная процедура:
Подготовка лизата
Дрожжевые клетки сначала выращивали в полной среде. Эту первую культуру высевали в 100 мл среды A (OD590=0,3), которую инкубировали при 28°С при перемешивании в течение 16 часов.
Общее количество белка определяли с использованием анализа по Брэдфорду.
Чтобы начать реакцию, от 0,03 до 0,06 мг общего белка инкубировали при 37°С в 100 мМ Трис рН 8,0, 0,2 мМ пиридоксаль фосфата, 5 мМ CH3SNa и 25 мМ OPHS в общем объеме 100 мкл в течение 15 минут.10 мкл аликвоты реакционной смеси отбирали через 15 и 60 минут и реакцию останавливали добавлением 90 мкл хлорной кислоты. Количество метионина в этих аликвотах было определено с помощью ЖХ/МС (жидкостная хроматография с масс-спектрометрией) с использованием 13CMet (метионин) в качестве внутреннего стандарта. Количество образованного метионина нормализовали количеством белка, используемого в исследовании.
Полученные результаты приведены в таблице 2:
Пример 6: In vitro анализ для измерения гомоцистеинсинтазной активности
Гомоцистеинсинтазную активность ферментов, экспрессированных в дрожжах, измеряли путем in vitro анализа с использованием неочищенного лизата дрожжей. Вслед за проверкой активности осуществлялся мониторинг синтеза метионина в лизате в присутствии O-фосфо-L-гомосерина (OPHS) и метантиола (CH3SNa). Лизаты из клеток дрожжей YA246-5A, YA246-5A, несущие пустую плазмиду pAL06, YA246-5A, экспрессирующий CGS1-4 и YA246-5A, экспрессирующий мутанты AD242, AD328, MUT24, MUT27, MUT67 или MUT79 сравнивали.
Экспериментальная процедура: Получение лизата
Дрожжевые клетки сначала выращивали в полной среде. Эту первую культуру высевали в 100 мл среды A (OD590=0.3), которую инкубировали при 28°С при перемешивании в течение 16 часов. Общее количество белка определяли с использованием анализа по Брэдфорду. Количество образованного гомоцистеина определяли колориметрическим методом.
Чтобы начать реакцию, от 0,03 до 0,06 мг общего белка инкубировали в 0,1 М Трис рН 8, 0,2 мМ пиридоксаль фосфате, 10 мМ Na2S, 12,5 мм OPHS в 100 мкл. Инкубировали 15 минут при 30°С.
Добавляли 500 мкл 1% NaNO2 (растворенного в 0,4Н H2SO) инкубировали в течение 5 минут.
Добавляли 100 мкл NH4SO3NH2 и инкубировали в течение 2 минут.
Добавляли 750 мкл 1 объема HgCl2.
Добавляли 4 объема сульфаниламида.
Добавляли 2 объема N-(1-нафтил)этилендиамин дигидрохлорида.
Инкубировали в течение 15 минут.
Измеряли оптическую плотность при длине волны 450 нм (возрастает пропорционально количеству гомоцистеина, количество определяли путем сравнения с результатами, полученными с известным диапазоном гомоцистеина). Результаты приведены в таблице 3.
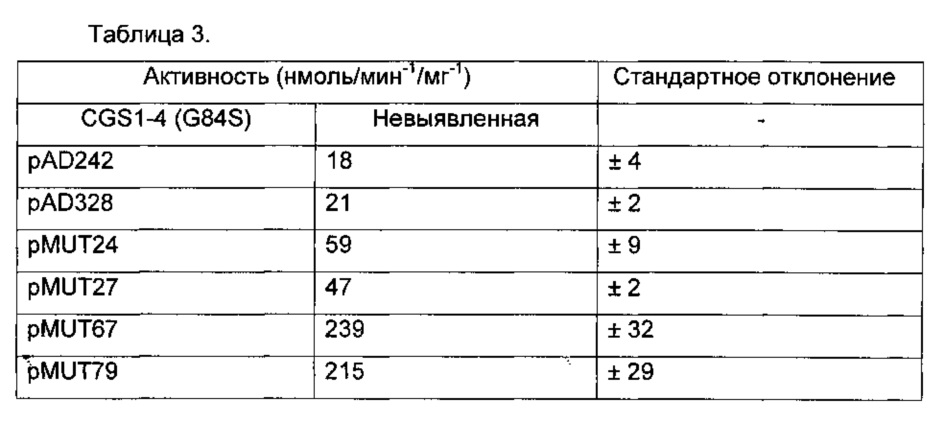

















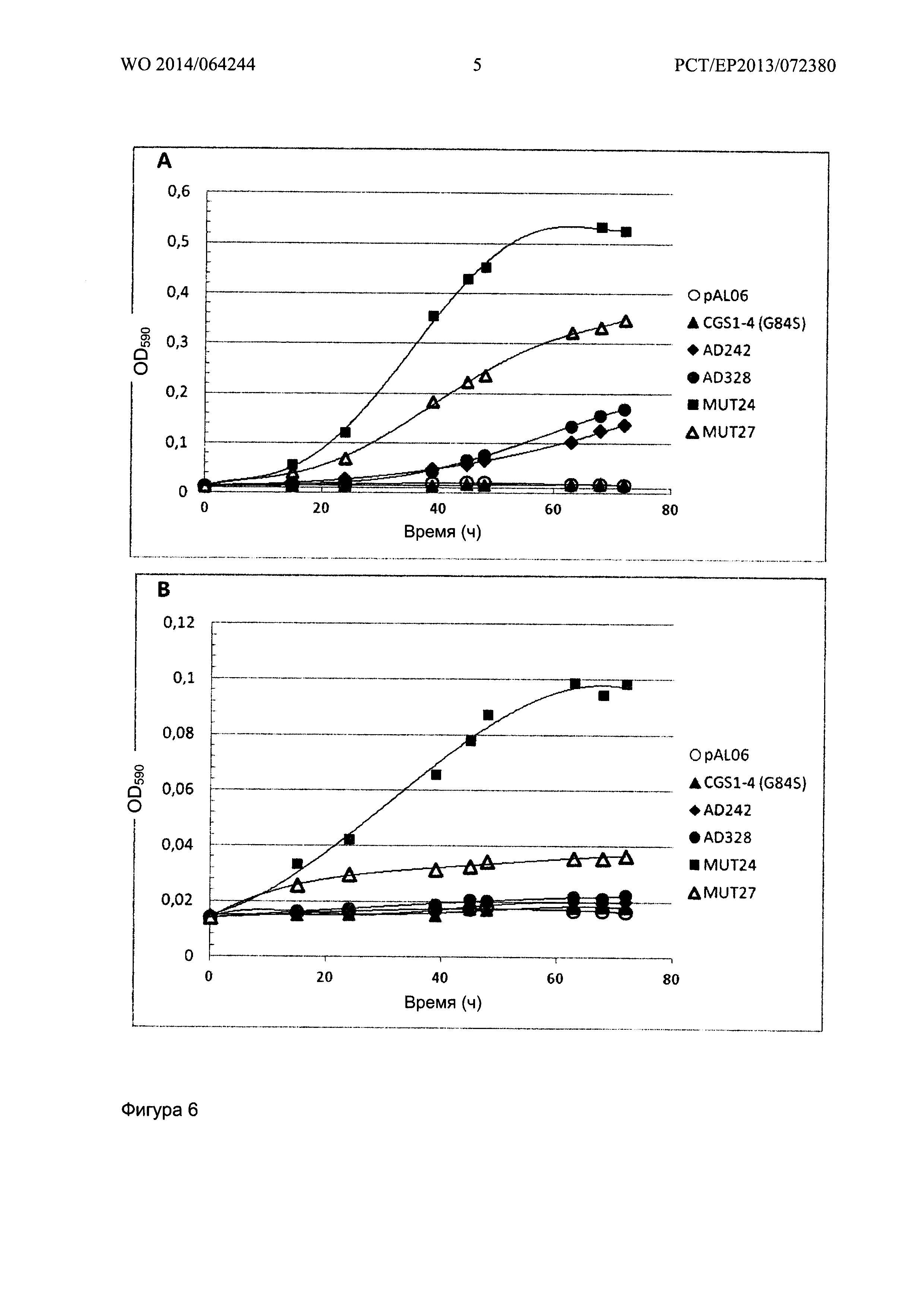







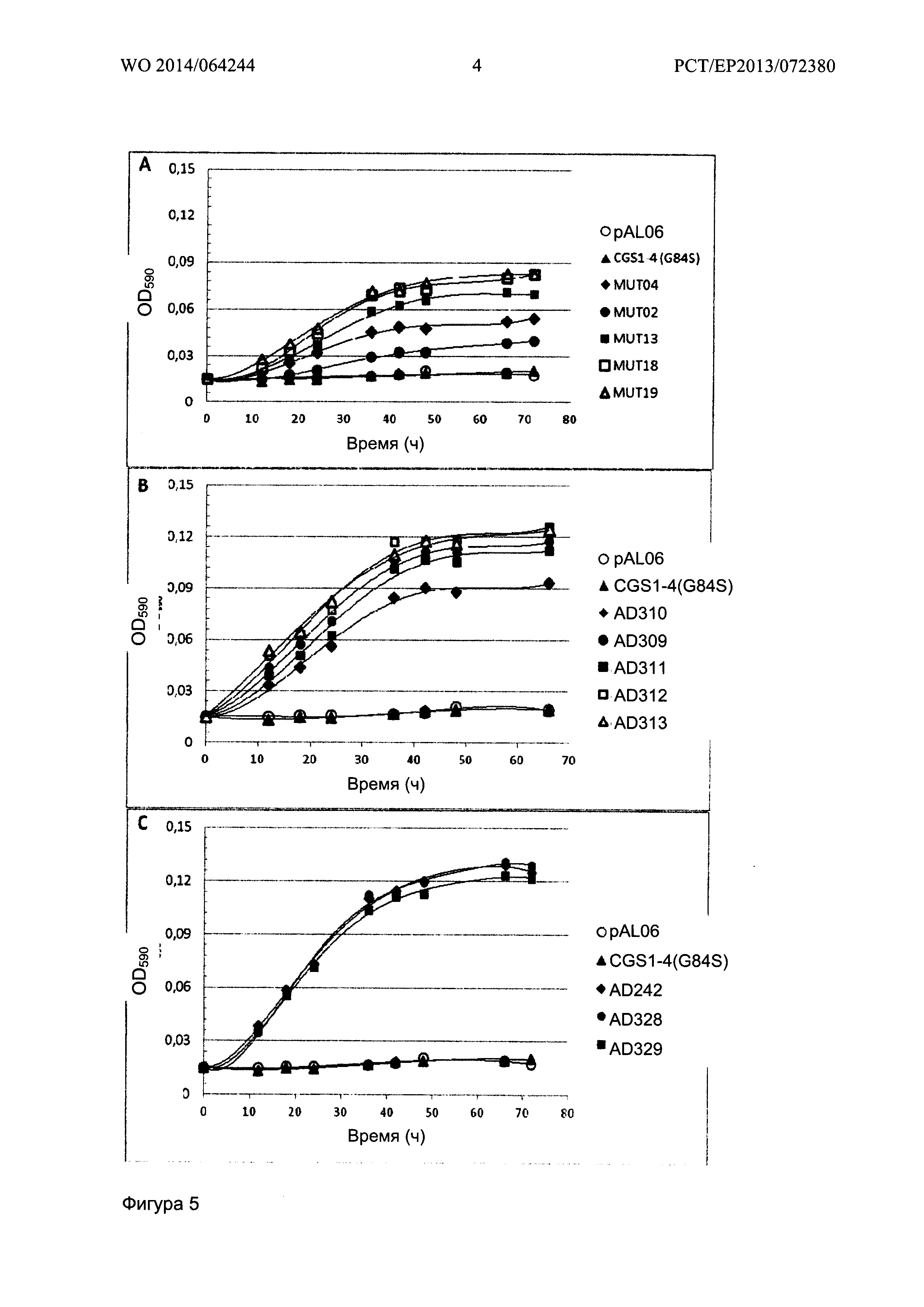









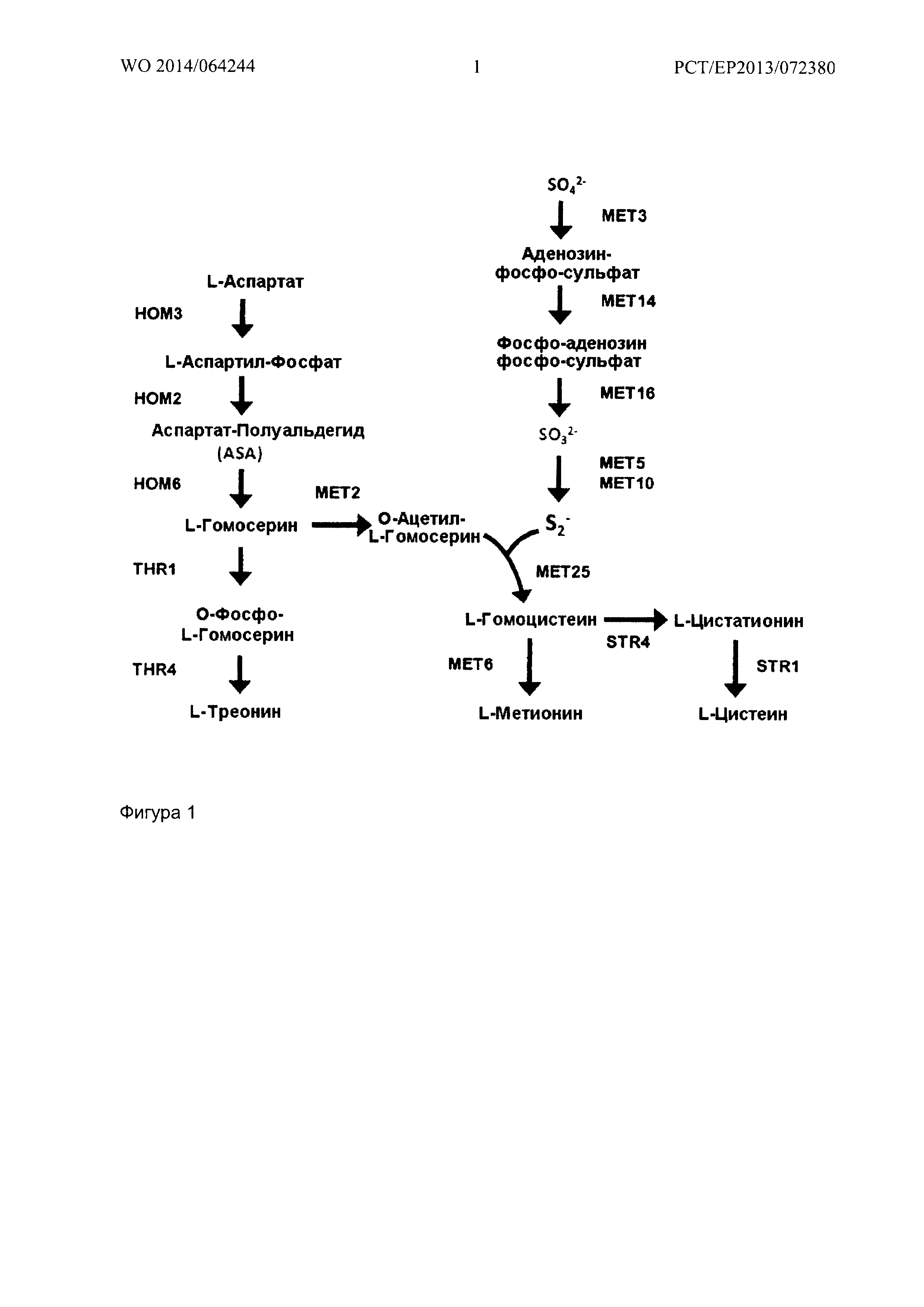



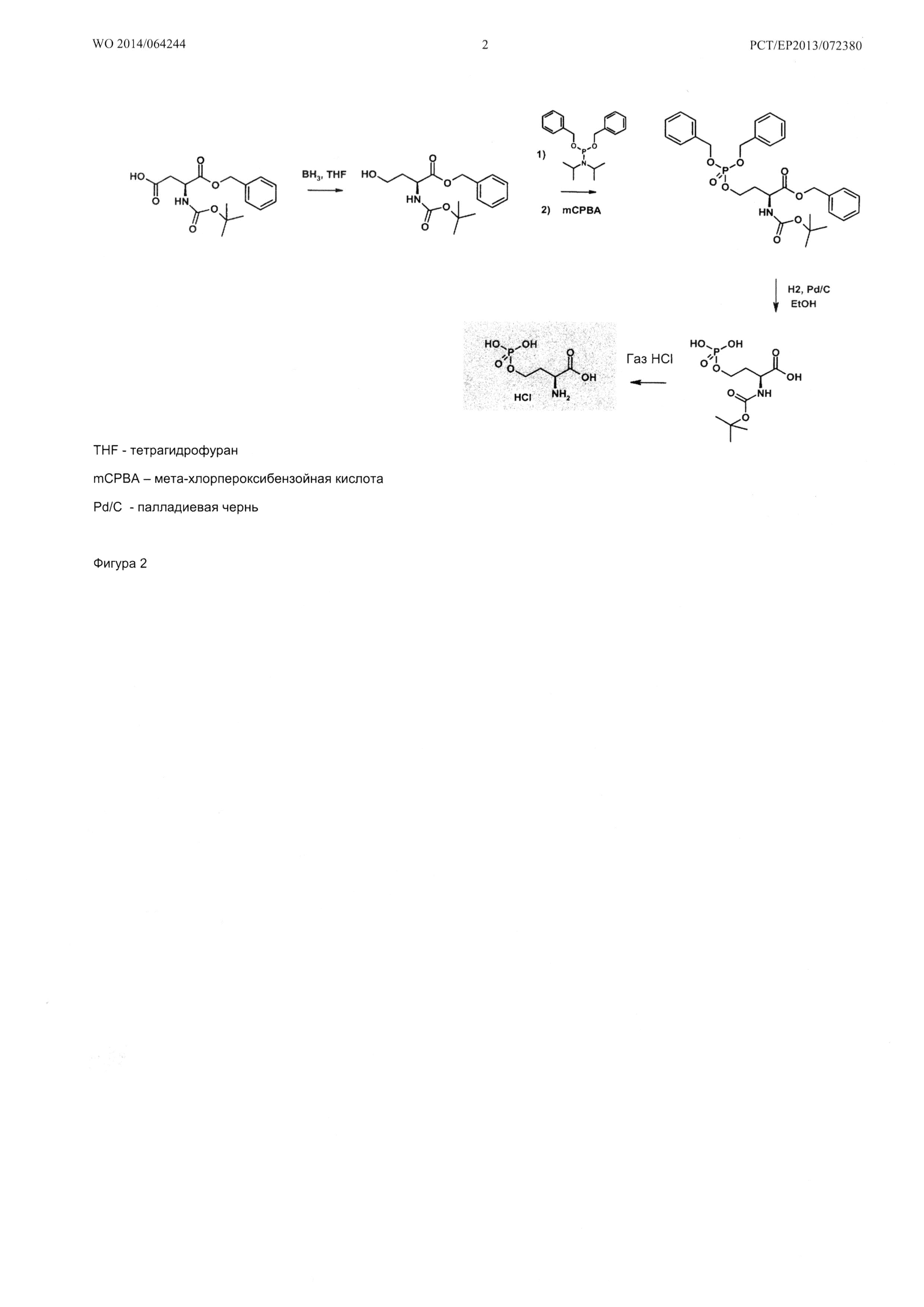
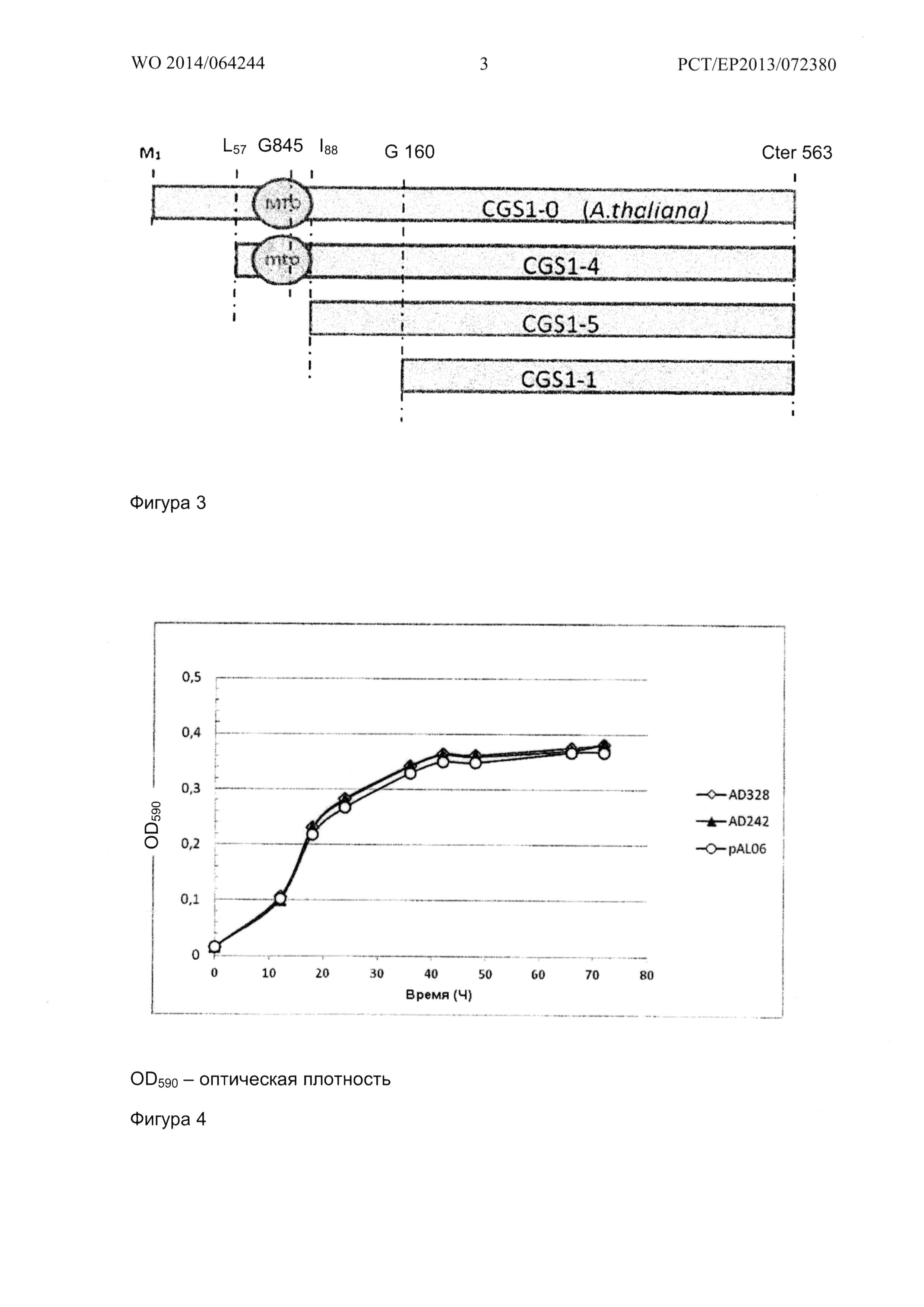
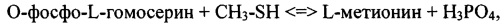

Термостабильная композиция для животных, содержащая смесь ферментов
Способ получения акролеина из глицерола или глицерина
Способ получения аминокислоты из 2-аминобутиролактона
Способ получения 2-гидроксибутиролактона
Способ получения 2,4-дигидроксимасляной кислоты
Способ получения акролеина из глицерина
Способ получения 2,4-дигидроксибутирата
Способ получения олефина посредством каталитической конверсии по меньшей мере одного спирта
Способ получения метионина
Штамм penicillium funiculosum, продуцирующиий комплекс ферментов - целлюлазу, эндо-1,4-β-ксиланазу, целлобиогидролазу, β-глюкозидазу, эндо-1,3(4)- β-глюканазу, ферулоил-эстеразу, жидкая кормовая добавка и сухой корм для сельскохозяйственных животных
Способ получения ацетоина

