Результат интеллектуальной деятельности: ЛИНИЯ КЛЕТОК PIGAS, СОДЕРЖАЩАЯ СТАБИЛЬНО ИНТЕГРИРОВАННЫЙ В ГЕНОМ САЙТ СВЯЗЫВАНИЯ ТРАНСКРИПЦИОННОГО ФАКТОРА STAT-1
Вид РИД
Изобретение
Область техники настоящего изобретения
Изобретение относится к области биотехнологии, молекулярной биологии, фармакологии, клеточной биологии и генной инженерии, в частности, к получению линии генномодифицированных клеток меланомы человека, используемых для. определения биоактивности IFN-γ.
Настоящее изобретение может быть использовано для определения биологической активности IFN-γ, а также для идентификации молекул различной природы (химических соединений, пептидов и полипептидов) и тестирования активности различных фармацевтических препаратов, демонстрирующих интерфероно-подобную активность или же способных модулировать активность IFN-γ-активированного сигнального пути.
Предшествующий уровень техники настоящего изобретения
В ответ на воздействие патогенов различной природы клетки иммунной системы синтезируют целый спектр цитокинов, модулирующих иммунный ответ и обеспечивающих защитные функции организма. Среди цитокинов важное место занимают представители интерферонового семейства. Интерфероны представляют собой эволюционно древний компонент врожденного и адаптивного иммунного ответа. Они оказывают противовирусное и антибактериальное действие, участвуют в противоопухолевом иммунном ответе и в развитии атопического дерматита [1]. Нарушение в продукции интерферонов детектируется в ряде заболеваний воспалительной и аутоиммунной природы [2].
Все интерфероны делятся на 3 основных типа (тип I, тип II и тип III), каждый из которых узнает свой рецептор на поверхности клетки [3]. Группа интерферонов типа I самая многочисленная. Она включает в себя 13 разновидностей: IFN-α, IFN-β, IFN-ε, IFN-κ, IFN-ω, IFN-δ и IFN-τ. Единственным представителем интерферонов типа II является IFN-γ. К интерферонам типа III относят несколько различных подтипов IFN-A (IFN-λ1, IFN-λ1, IFN-λ3).
IFN-γ входит в группу цитокинов с плейотропным действием, играющих важную роль в иммунном ответе. Ген IFN-γ человека локализуется в 12-ой хромосоме, содержит 4 экзона и кодирует полипептид из 166 аминокислотных остатков. Первоначально считалось, что продукция IFN-γ свойственна только узкому спектру, клеток иммунной системы, среди которых натуральные киллеры (NK), CD4+ Th1-лимфоциты и цитотоксические CD8+ Т-лимфоциты. Позднее стало очевидно, что В-лимфоциты, NKT-клетки и профессиональные антиген-презентирующие клетки (АРС) (макрофаги, дендритные клетки) также способны к синтезу данного цитокина. IFN-γ-продуцирующая способность у различных типов иммунных клеток разная и тесно связана с особенностями устройства гена, кодирующего этот цитокин [4].
Нарушения в IFN-γ-зависимой системе имеют драматические последствия для организма [5]. Мыши, дефектные по гену IFN-γ, оказываются гиперчувствительны к бактериальным, паразитическим и вирусным инфекциям, вызываемым вирусом коровьей оспы, мышиным вирусом энцефаломиелита, паразитическими протистами Leishmania major, Toxoplasma gondii, бактерией Listeria monocytogenes и некоторыми слабовирулетными микобактериями. У людей среди клинических проявлений, связанных с нарушением в IFN-γ-системе, отмечают сильную восприимчивость к слабовирулентным микобактериям, включая осложнения при вакцинации БЦЖ вплоть до летального исхода. Помимо повышенной восприимчивости к вирусным и бактриальным инфекциям, для детей с дефектной IFN-γ-системой характерна пониженная мобильности нейтрофилов и активность NK-клеток, что связано с ключевой ролью IFN-γ в регуляции воспалительной реакцияии и иммунорегуляции. Кроме того, IFN-γ оказывается важным фактором и при ряде аутоиммунных патологий, включая системную красную волчанку, множественный склероз и инсулин-зависимый сахарный диабет.
IFN-γ используется как важный терапевтический агент. Он оказывает противовирусное, антипаразитическое, микробицидное, иммуномодулирующее и антипролиферативное действия. Влиянию со стороны данного цитокина подвержены компоненты как врожденного, так и адаптивного иммунитета. IFN-γ стимулирует активацию макрофагов и NK-клеток [6]. Кроме того, в присутствии IFN-γ происходит увеличение эффективности презентации процессированного опухолевого антигена в составе комплекса МНС I класса-пептид-β2-микроглобулин Т-клетками. IFN-γ может препятствовать развитию опухоли, оказывая антипролиферативное и проапоптотическое влияние на трансформированные клетки [7], [8]. Активация IFN-γ/STAT-1 сигнального каскада ингибирует опухоль-ассоциированный ангиогенез [9]. С другой стороны, IFN-γ способен индуцировать ускользание опухоли от иммунного надзора [10]-[12].
IFN-γ синтезируется в виде предшественника, содержащего сигнальный пептид размером в 23 аминокислотных остатка. В процессе созревания IFN-γ претерпевает посттрансляционные модификации, включая С-концевой процессинг и N-гликозилирование. Посттрансляционные модификации IFN-γ важны для его биологической активности. В частности, гликозилирование по Asn25 обеспечивает более эффективную димеризацию и секрецию белка [13].
Биологически активная форма IFN-γ представляет собой нековалентный антипараллельный гомодимер, состоящий из гликопротеинов с молекулярной массой массой 20-25 kDa.
Существует несколько способов получения IFN-γ в препаративных количествах. Одним из первых был разработан метод выделения IFN-γ из компонентов человеческой крови. Данный метод имеет ряд существенных недостатков, препятствующих его удачному технологическому внедрению. В частности, выделение нативных цитокинов экономически невыгодно из-за низкого выхода продукта (менее 1 мг/л донорской крови), а также возможности контаминации препатарата различными вирусами, передающихся через кровь и ее компоненты [14]. Технологии получения рекомбинантных белков позволяют избежать описанных трудностей путем эффективного синтеза требуемых белков в прокариотических и эукариотических экспрессионных системах. На настоящий момент на фармацевтическом рынке, преобладает рекомбинантный IFN-γ.
Гетерологическая экспрессия IFN-γ в эукариотических клетках, особенно в клетках млекопитающих, позволяется получить зрелый гликопротеин, прошедший через этапы посттрансляционной модификации, что должно гарантировать уровень его. биологической активности сопоставимый с нативным белком. Однако высокая стоимость компонентов питательных сред для эукариот и необходимость дорогостоящих стадий очистки рекомбинантного IFN-γ, делают коммерчески невыгодным производство данного белка путем экспрессии в клетках млекопитающих [15].
Кроме того, ряд исследований указывает на то, что вид используемой эукариотической системы влияет на качество получаемого рекомбинантного белка. В частности, с использованием различных техник, основанных на масс-спектрометрическом анализе, показано, что рекомбинантный IFN-γ имеет разную, степень и сайты N-гликозилирования, а также структуру С-концевой части молекулы в зависимости от того, получен он в линии клеток яичников китайских хомячков СНО (Chinese hamster ovary cells), в клетках молочных желез трансгенных мышей или в линии клеток насекомых Spodoptera frugiperda (Sf9), инфицированных бакуловирусом. [16]. Демонстрируется также, что IFN-γ, полученный в клетках линии СНО, представляет собой гетерогенную популяцию, включающую в себя молекулы с различных уровнем и характером посттрансляционных модификаций [17]. Как уже отмечалось выше, посттрансляционные модификации, такие как гликозилирование, могут сказываться на биоактивности, способности к связыванию рецептора, чувствительности к протеолизу, иммуногенности, а также стабильности терапевтического рекомбинантного белка in vivo. Рекомбинантный IFN-γ человека, полученный в клетках насекомых, отличается от нативного IFN-γ человека по своим фармакокинетическим свойствам. Время выведения такого IFN-γ из кровотока кролика значительно меньше, чем нативного белка [18]. Для рекомбинантного IFN-γ, полученного в клеках мелкопитающих, характерна модификая структурно важной С-концевой части молекулы, что может существенно снизить его биоактивность [16].
Существуют методы экспрессии и очистки функционально активного рекомбинантного IFN-γ в клетках одноклеточных эукариот, например в штаммах дрожжей Pichia pastoris. Серьезным недостатком данного подхода является крайне сложные условия ферментации этого типа дрожжей, необходимость строго поддерживать концентрацию индуктора, в частности метанола, в процессе биосинтеза [19].
Еще одним способом наработки рекомбинантного IFN-γ человека является его микробиологический синтез в прокариотических системах. Штаммы-продуценты E.coli широко используются для получения биологически активного рекомбинантного IFN-γ человека в количестве, достаточном как для проведения структурно-функциональных исследований, так и для использования в качестве терапевтичекого агента.
Следует отметить, что используемые на настоящий момент способы выделения и очистки рекомбинантного IFN-γ включают в себя процедуры по ренатурации белка из телец включения. Процесс рефолдинга достаточно сложный и сопряжен с опасностью потери биологической активности выделяемого белка [20]. Вне зависимости от метода, использованного для получения IFN-γ, функциональная активность конечного выделенного и очищенного продукта должна быть проверена и подтверждена.
Кроме того, периодическая проверка функциональной активности требуется и во время хранения IFN-γ. В процессе хранения очищенный рекомбинантный IFN-γ человека может подвергнуться модификациям, способным сильно снизить (вплоть до полного элиминирования) его биологическую активность [21]. В бактериальных клетках отсутствует система посттрансляционной модификации белков, в частности система энзиматического гликозилирования. Однако показано, что рекомбинантный IFN-γ, полученный путем микробиологического синтеза в штаммах E.coli, может, содержать углеводные остатки [22]. В нормальных физиологичеких условиях в клетках E.coli протекает процесс гликирования, заключающийся в неэнзиматической неконтролируемой ковалентной модификации белков углеводными остатками (часто глюкозой). Более того, после ковалентного присоединения остатков глюкозы белок, может претерпевать дальнейшие изменения и в отсутствии Сахаров, что приводит к образованию конечных продуктов глубокого гликирования (advanced glycation end products (AGEs)) [23]. Гликирование рекомбинантного IFN-γ человека отражается на его стабильности. При хранении гликированная фракция очищенного IFN-γ· подвергается протеолизу и ковалентной димеризации, что приводит к потере противовирусной активности препарата [21].
Для оценки удельной биологической активности IFN-γ как природного происхождения, так и полученного методами генной инженерии, используют тест по супрессии цитопатического действия вируса на клеточную культуру. Наиболее часто используются следующие сочетания клетка/вирус: клетки бычьих почек Madin-Darby (MDBK), лимфоидные клетки человека Л-41, перевиваемые клетки почки эмбриона свиньи СПЭВ/вирус везикулярного стоматита (VSV); клетки линии Vero, Нер2С, L929, Л-41/вирус энцефаломиокардита мышей (EMCV); клетки фибробластов человека/вирус леса Семлики, вирус Синдбис или иные [24]. Метод достаточно трудоемкий и требует от исследователя повышенной осторожности в виду работы с вирусными частицами. На настоящий момент тест по супрессии цитопатического действия вируса на клеточную культуру является единственным доступным методом определения биологической активности IFN-γ. Учитывая востребованность процедуры проверки функциональной активности IFN-γ как для лабораторной практики, так и для фармацевтической отрасли, существует необходимость в разработке и создании новых подходов и тестовых систем для решения данной задачи.
Один из возможных подходов к созданию тест-системы для определения биоактивности IFN-γ основывается на способности функционального IFN-γ запускать внутриклеточный сигнальный каскад с участием транскрипционного фактора STAT-1 (Signal Transducer and Activator of Transcription 1).
Обработка клеток функциональным IFN-γ приводит к активации транскрипционного фактора STAT-1. IFN-γ связывается с рецепторным комплексом IFNGR (IFNGR1/IFNGR2) на поверхности клеток. Цитоплазматические домены IFNGR ассоциированы с двумя нерецепторными тирозиновыми киназами JАК1 и JAK2. Образование компекса IFN-γ/IFNGR1/IFNGR2 индуцирует JАК1/JАК2-зависимое фосфорилирование цитоплазматических доменов рецептора, что создает условия для связывания, фосфорилирования и активации транскрипционного фактора STAT-1. Активный фосфорилированный фактор STAT-1 димеризуется и транслоцируется в ядро, где активирует транскрипцию генов, промоторные области которых содержат последовательность GAS (IFN-γ-activated site) [5].
Существуют способы количественной оценки активности транскрипционных факторов. Эти способы основаны на использовании репортерных экспрессионных конструкций, представляющих собой генетические элементы, состоящие из репортерного гена, минимального промотора и сайта связывания для транскрипционного фактора, чья активность детектируется. В качестве репортерного гена может быть использован любой ген, продукт которого легко поддается, количественному измерению и не влияет на физиологию клетки. Под минимальным промотором понимается промотор, который сам по себе не способен направлять экспрессию репортерного гена и требует добавления перед ним специальных регуляторных элементов, таких как сайты связывания для транскрипционного фактора. Сайты связывания, в свою очередь, представляют из себя нуклеотидные последовательности, которые располагаются в непосредственной близости от минимального промотора, распознаются и связываются специфическими транскрипционными факторами, что способствует активации транскрипции репортерного гена с минимального промотора. Репортерные экспрессионные конструкции вводятся в клетки путем трансфекции. В том случае, если в клетке присутствует соответствующий активный транскрипционный фактор, наблюдается экспрессия репортерного гена.
Раскрытие настоящего изобретения
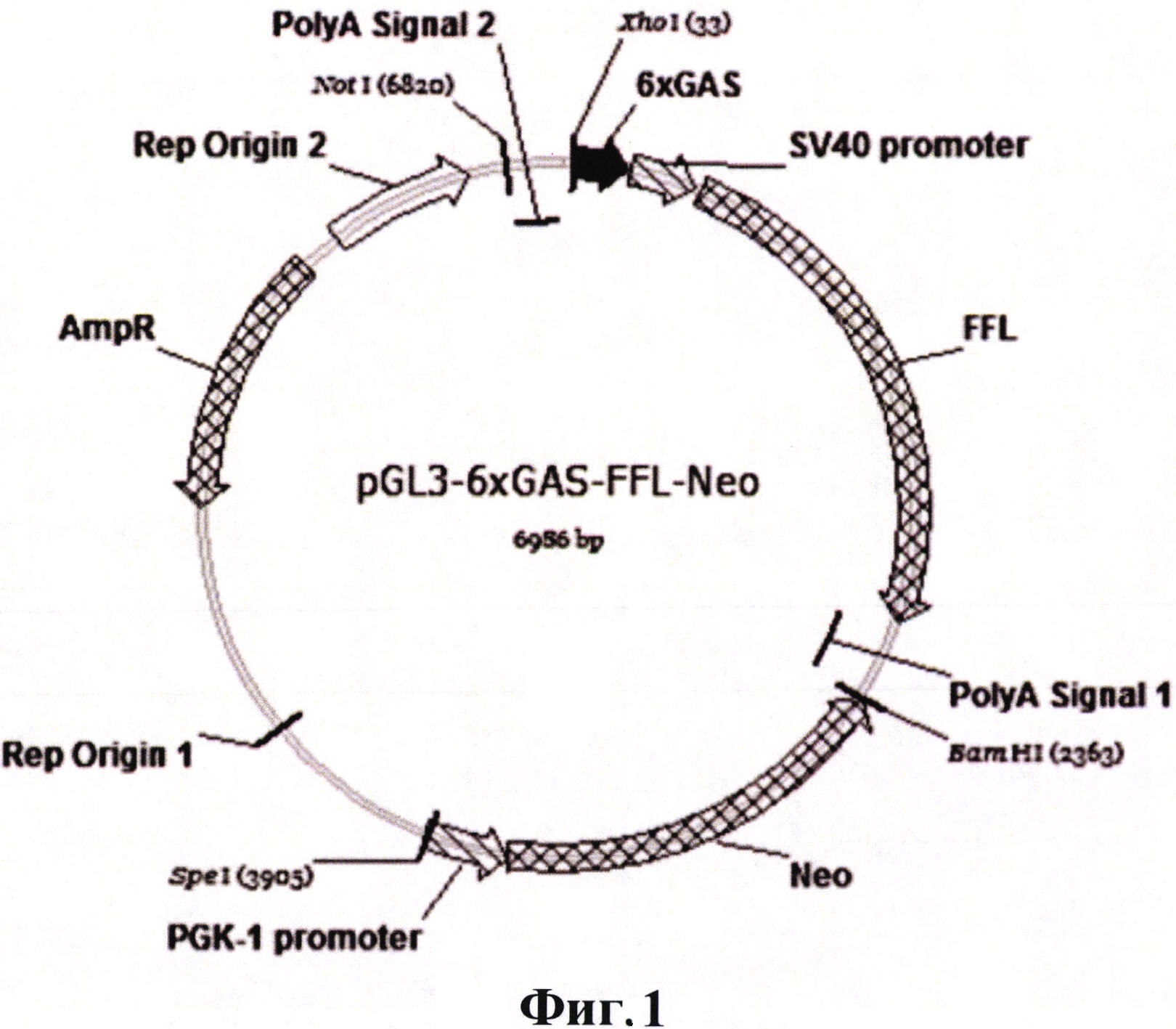
Сущностью настоящего изобретения является создание новой линии клеток PiGAS, содержащей стабильно интегрированную в геном репортерную конструкцию 6xGAS-Luc//Neo, позволяющую детектировать биоактивность IFN-γ. Способность клеток линии PiGAS выступать в качестве модельной системы для детекции активности интерферонов достигается при помощи генной модификации клеток линии меланомы человека MelP плазмидой pGAS(x6)-Luc//Neo, содержащей репортерную кассету 6xGAS-Luc, встроенную в вектор, предназначенный для экспрессии в клетках млекопитающих, из шести тандемно повторяющихся сайтов связывания GAS (IFN-γ-activated site) транскрипционного фактора STAT-1, минимального промотора SV40 и гена люциферазы светлячка Photinus pyralis (Luc). Помимо репортерной кассеты 6xGAS-Luc плазмида pGAS(x6)-Luc//Neo содержит ген селективного маркера неомицин-фосфотрансферазы (Neo), что позволяет выращивать клетки линии PiGAS на селективной среде, содержащей антибиотик G418.
Технический результат заключается в получении клеток линии PiGAS, стабильно трансфицированных плазмидой pGAS(x6)-Luc//Neo, предназначенной для определения биоактивности IFN-γ человека.
Целью настоящего изобретения является расширение коллекции уникальных клеточных штаммов, которые можно использовать для детекции биологической активности как рекомбинантных, так и эндогенных белков, в частности IFN-γ. Эта задача является особенно актуальной для современной биотехнологии и молекулярной биологии. Интерфероны преимущественно получают методом микробиологического синтеза в различных искусственно сконструированных штаммах P. Pastoris, Ps. putida и Ε.coli. В результате отсутствия посттрансляционных модификаций, свойственных природным эндогенным интерферонам, а также особенностям процедуры очистки рекомбинантных белков полученные препараты нуждаются в проверке их биологичекой активности. При хранении (особенно в водных растворах) интерфероны подвергаются модификации и химическому разложению за счет протекания процессов протеолиза, окисления, дисульфидного обмена и прочих, что также сказывается на их биологической активности. На настоящий момент биологическую активность, препаратов интерферона определяют в основном с помощью тест-систем, в основе которых лежит способность интерферонов подавлять цитопатическое действие вируса (например, вируса везикулярного стоматита) в культуре клеток. Метод достаточно трудоемкий и имеет ряд недостатков в виду необходимости использования вирусных частиц, что в совокупности диктует необходимость разработки новых методов определения биоактивности интерферонов.
Поставленная задача решается тем, что получена новая линия клеток PiGAS, несущая стабильно интегрированную в геном репортерную экспрессионную конструкцию, представляющую собой последовательно расположенные 6 повторов сайта связывания GAS (IFN-γ-activated site) транскрипционного фактора STAT-1, минимальный промотор SV40 и репортерный ген люциферазы светлячка Photinus pyralis (Luc), встроенные в вектор, предназначенный для экспрессии в клетках млекопитающих. Линия PiGAS также содержит интегрированный в геном ген неомицин фосфотрансферазы II (Neo) под контролем конститутивного протомора PGK-1, что обеспечивает возможность выращивания клеток в ростовой среде в присутствии селективного антибиотика G418.
Полученная клеточная линия PiGAS может быть использована в качестве тест-системы для определения биоактивности IFN-γ. Она позволяет достоверно детектировать биоактивный IFN-γ в концентрации не менее 0,05 нг/мл.
Полученная клеточная линия PiGAS обладает стабильными культуральными и морфологическими свойствами и депонирована во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под регистрационным номером Н-161.
Краткое описание чертежей
Фигура 1 демонстрирует карту полученной плазмиды pGAS(x6)-Luc//Neo, содержащей репортерную экспрессионную кассету для детекции биоактивности IFN-γ, состоящую их гена люциферазы светлячка (Luc) под контролем промотора SV40, которому предшествуют 6 повторов сайта связывания GAS (IFN-γ-activated site) транскрипционного фактора STAT-1 (SEQ ID NO: 1), а также селективный маркерный ген неомицин фосфотрансферазы II (Neo) под контролем конститутивного промотора PGK-1 (SEQ ID NO: 2), обеспечивающий устойчивость эукариотических клеток к антибиотику генетицину (G418).
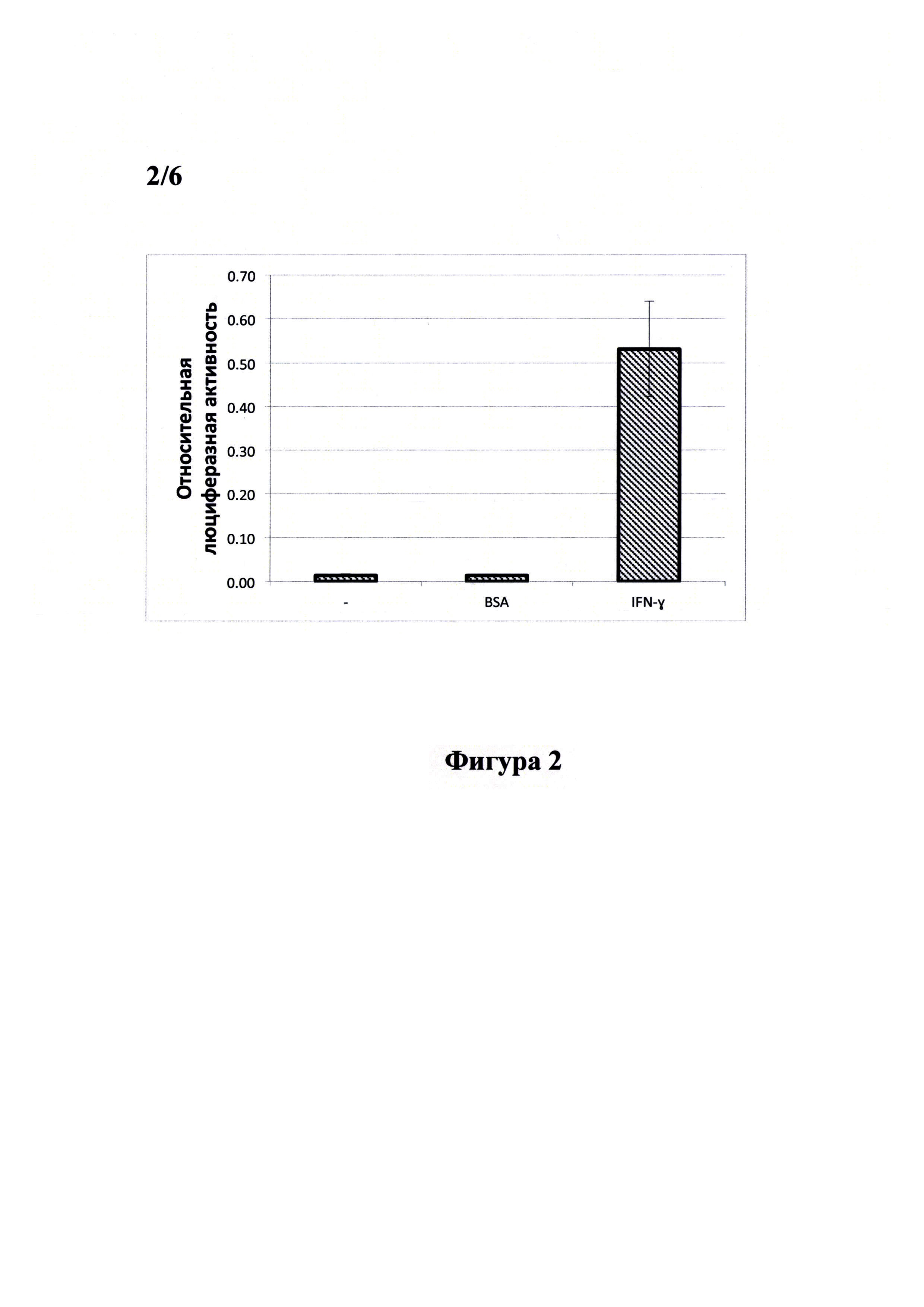
Фигура 2 демонстрирует результат двойного люциферазного теста, проведенного с лизатами клеток линии меланомы человека MelP, стимулированных IFN-γ. Клетки линии MelP высаживали в лунки 24-луночного планшета по 120000. клеток в лунку, выращивали до достижения 60-80% монослоя, после чего трансфицировали смесью плазмид pGAS(x6)-Luc//Neo и pRL-CMV (Promega, США) при помощи набора для липофекции эукариотических клеток Unifectin-56 (UnifectGroup, Россия) согласно рекомендациям производителя. Спустя 4 часа проводили замену среды в лунках на свежую культуральную среду, содержащую 10 нг/мл рекомбинантного IFN-γ или 100 нг/мл бычьего сыворочного альбумина и инкубировали клетки в течение 24 часов. Далее клетки лизировали и проводили двойной люциферазный тест согласно рекомендациям производителя (Promega, США). Все измерения проводили в четырех повторах. Результаты представлены в виде гистограммы. Под относительной люциферазной активностью понимается отношение активности люциферазы светлячка Photinus pyralis (Luc) к активности люциферазы Renilla.
Стимуляция клеток рекомбинантным IFN-γ человека вызывает увеличение сигнала относительной активности люциферазы в 37 раз по сравнению с нестимулированными клетками.
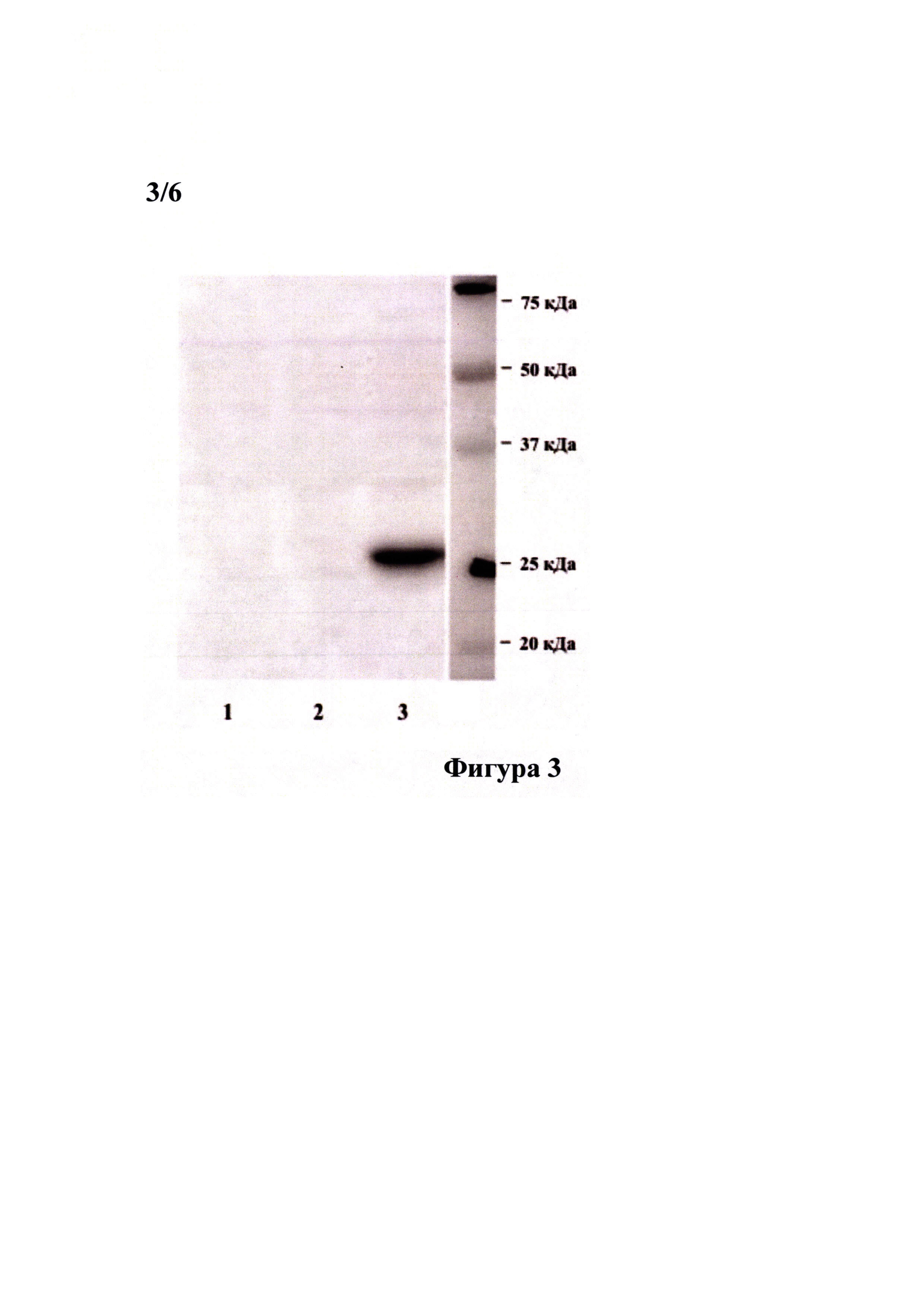
Фигура 3 демонстрирует результат анализа лизата клеток линии меланомы человека MelP, трансфицированных плазмидами pGAS(x6)-Luc//Neo и pGAS(x6)-Luc, на присутствие белка NPTII (неомицин фосфотрансферазы И) методом вестерн-блот анализа со специфическими антителами. Плазмида pGAS(x6)-Luc отличается от плазмиды pGAS(x6)-Luc//Neo отсутствием селективного маркерного гена неомицин фосфотрансферазы II (Neo) под контролем конститутивного промотора PGK-1. Клетки линии MelP высаживали в лунки 6-луночного планшета по 500000 клеток в лунку, выращивали до достижения 60-80% монослоя, после чего трансфицировали соответствующими плазмидами при помощи набора для липофекции эукариотических клеток Unifectin-56 (UnifectGroup, Россия) согласно рекомендациям производителя. Через 24 часа клетки лизировали в буфере Лэмли (BioRad) и использовали для анализа. Все образцы разделяли в 12,5% ДСН-ПААГ в редуцирующих условиях, после чего проводили вестерн-блот анализ с использованием антител к неомицин фосфотрансферазе II (Millipore, США). Для оценки молекулярной массы белков использовали маркер молекулярной массы Prescision Plus Protein™ Standard (Bio-Rad, США).
По результатам вестерн-блот анализа в образце, соответствующем лизату клеток линии MelP, трансфицированных плазмидой pGAS(x6)-Luc//Neo, детектируется белок весом 29 кДа, отсутствующий в нетрансфицированных клетках или клетках, трансфицированных плазмдой pGAS(x6)-Luc. Размер детектируемого белка соответствует размеру неомицин фосфотрансферазы II. 1 - нетрансфицированные клетки линии меланомы человека MelP, 2 - клетки линии меланомы человека MelP, трансфицированные плазмидой pGAS(x6)-Luc, 3 - клетки линии меланомы человека MelP, трансфицированные плазмидой pGAS(x6)-Luc//Neo.
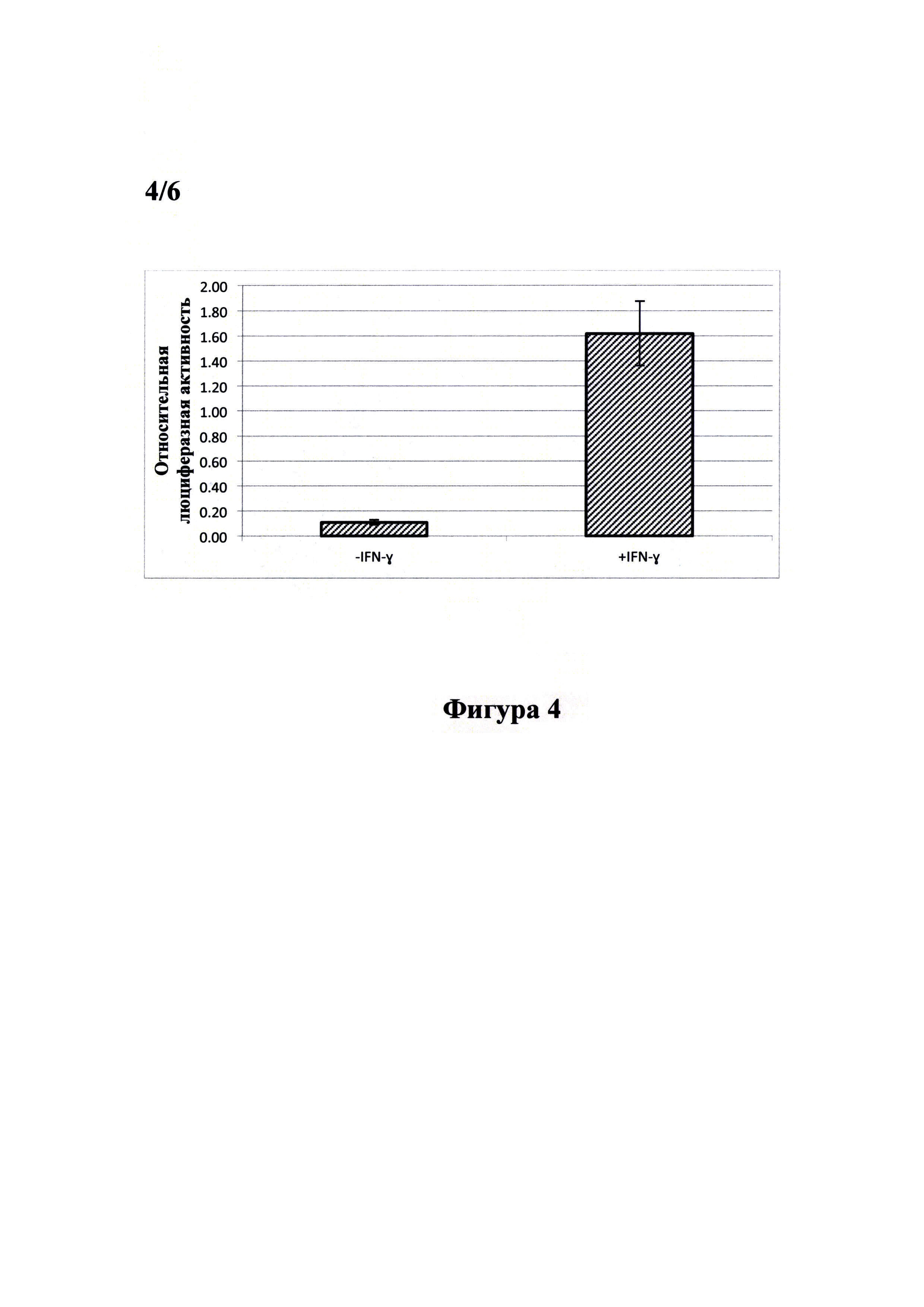
Фигура 4 демонстрирует результат анализа активности STAT-1-зависимого репортера в клеточной линии PiGAS, содержащей стабильно интегрированную в геном экспрессионную кассету 6xGAS-Luc//Neo. Клетки линии PiGAS высаживали в лунки. 24-луночного планшета по 120000 клеток в лунку. Спустя 4-6 часов, убедившись, что клетки полностью прикрепились ко дну планшета и восстановили морфологию, проводили замену среды в лунках на свежую культуральную среду, содержащую 10 нг/мл рекомбинантного IFN-γ человека и инкубировали клетки в течение 24 часов. Далее получали клеточные лизаты, в которых измеряли активность люциферазы при помощи люциферазного теста (Promega, США) и концентрацию общего белка при помощи бицинхониновой кислоты (Sigma-Aldrich, США). Для каждого лизата проводили нормировку значения активности люциферазы на концентрацию общего белка. Все измерения проводили в четырех повторах. Результаты представлены в виде гистограммы. Под относительной люциферазной активностью понимается отношение активности люциферазы светлячка Photinus pyralis (Luc) к концентрации общего белка.
Стимуляция клеток рекомбинантным IFN-γ человека вызывает увеличение сигнала относительной активности люциферазы в 15 раз по сравнению с нестимулированными клетками.
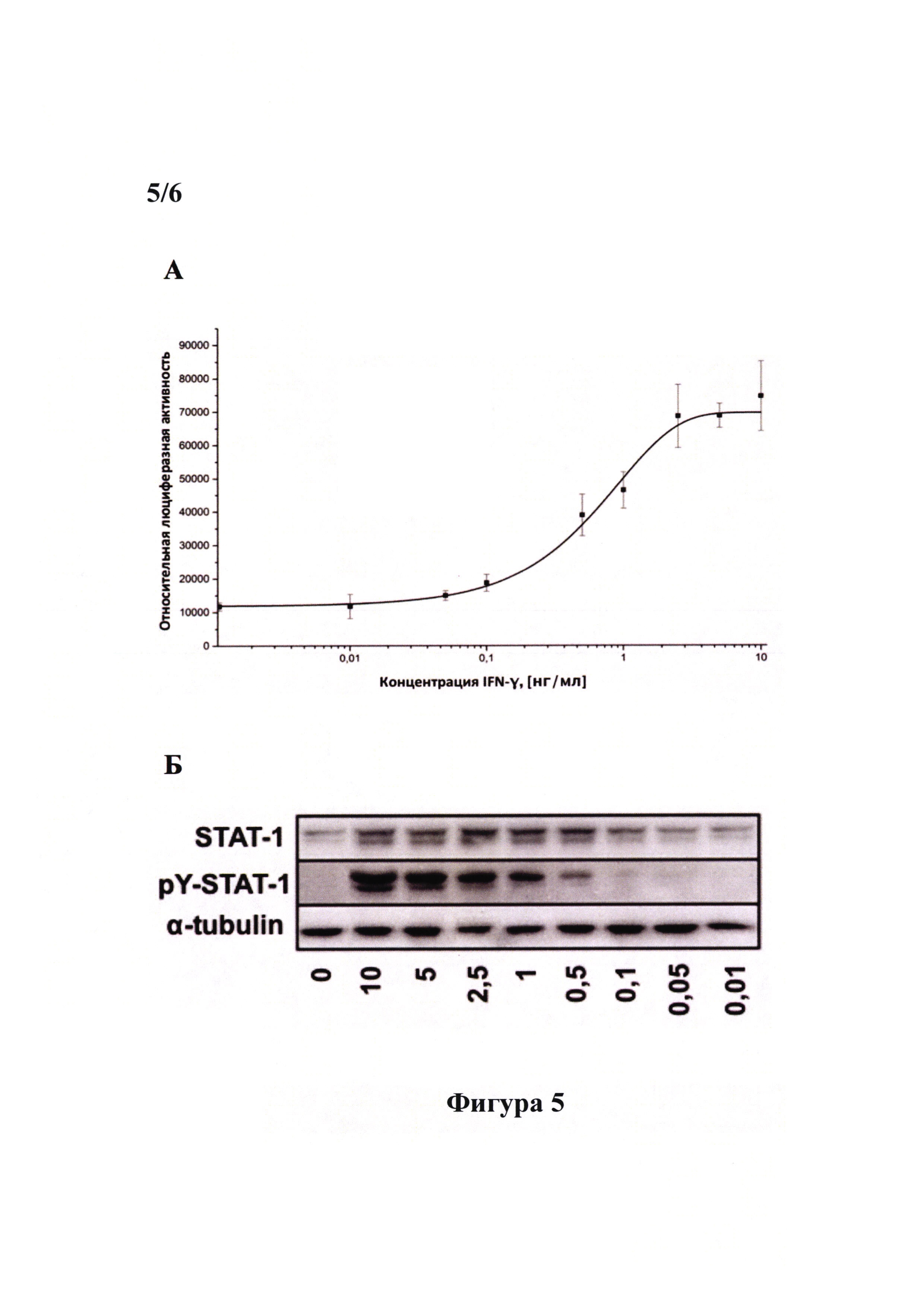
Фигура 5 демонстрирует зависимость уровня активности люциферазы (А) и уровень фосфорилирования транскрипционного фактора STAT-1 (Б) в репортерной клеточной линии PiGAS, содержащей стабильно интегрированную в геном экспрессионную кассету 6xGAS-Luc//Neo, от концентрации рекомбинантного IFN-γ человека. Клетки линии PiGAS высаживали в лунки 96-луночного планшета по 30000 клеток в лунку. Спустя 16 часа проводили замену среды в лунках на свежую культуральную среду, содержащую рекомбинантный IFN-γ человека в концентрации от 0,01 нг/мл до 10 нг/мл и инкубировали клетки в течение 24 часов. Далее получали клеточные лизаты. (А) В клеточных лизатах измеряли активность люциферазы при помощи люциферазного теста (Promega, США) и концентрацию общего белка при помощи бицинхониновой кислоты (Sigma-Aldrich, США). Для каждого лизата проводили нормировку значения активности люциферазы на концентрацию общего белка. Все измерения проводили в четырех повторах. Результаты представляли в виде графика зависимости относительной люциферазной активности от концентации рекомбинантного IFN-γ человека. Под относительной люциферазной активностью понимается отношение активности люциферазы светлячка Photinus pyralis (Luc) к концентрации общего белка. Репортерная клеточная линия PiGAS достоверно детектирует биоактивный IFN-γ в концентрации не менее 0,05 нг/мл. (Б) В клеточных лизатах также проводили детекцию фосфорилированной формы транскрипционного фактора STAT-1 (pY-STAT-1) и тотального STAT-1 при помощи вестерн-блот анализа с использованием специфических антител (Cell signaling, США). pY-STAT-1 детектируется в лизатах клеток линии PiGAS, стимулированных IFN-γ в концентрации не ниже 0,1 нг/мл.
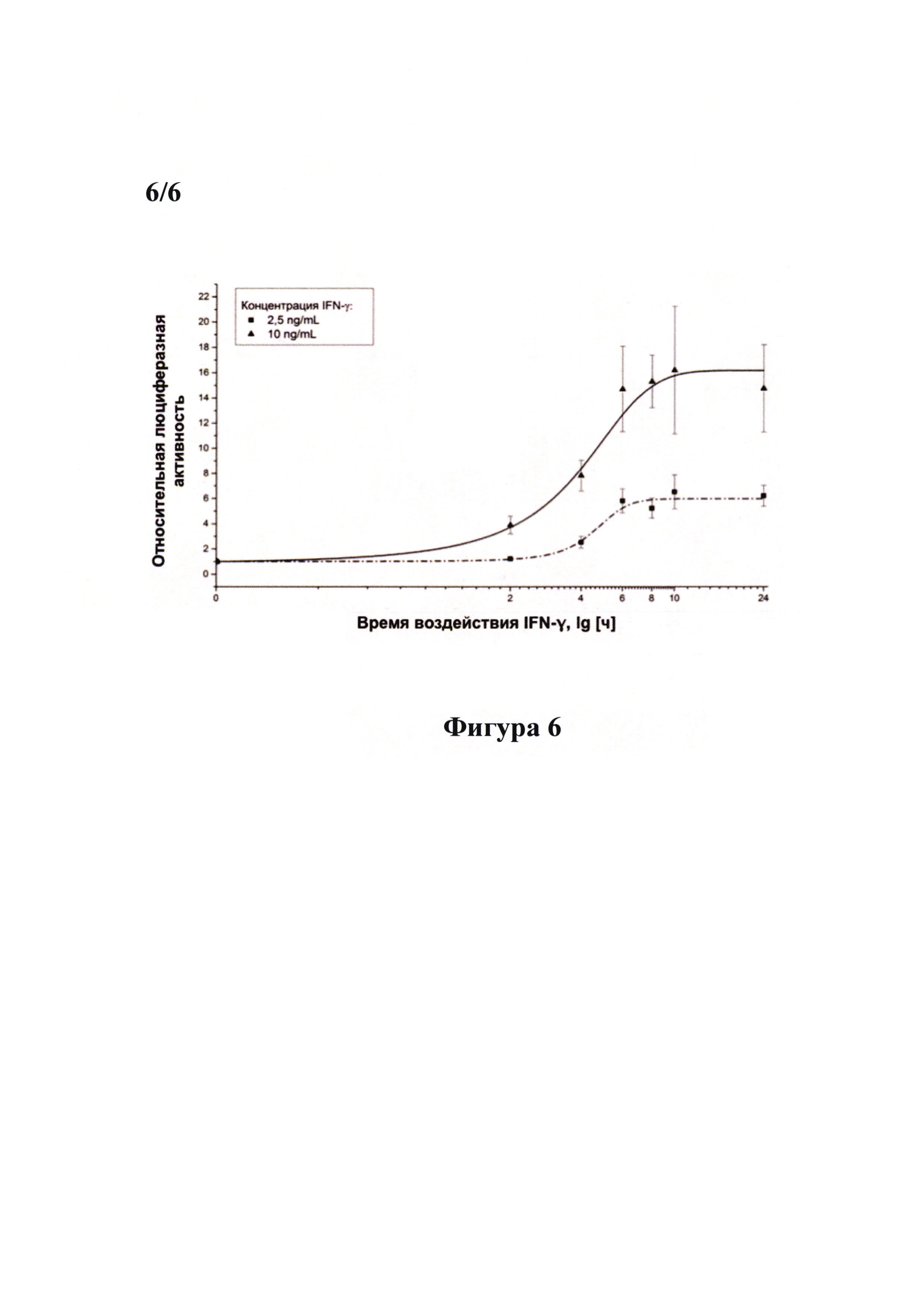
Фигура 6 демонстрирует зависимость уровня активности люциферазы в репортерной линии PiGAS, содержащей стабильно интегрированную в геном экспрессионную кассету 6xGAS-Luc//Neo, от времени стимуляции клеток рекомбинантным IFN-γ человека. Клетки линии PiGAS засевали в лунки 96-луночного планшета по 30000 клеток в лунку. Спустя 16 часов проводили замену среды в лунках на свежую культуральную среду, содержащую рекомбинантный IFN-γ человека в концентрации 2,5 нг/мл (прерывистая линия) или 10 нг/мл (сплошная линия), инкубировали клетки в течение 2, 4, 6, 8, 10 или 24 часов, после чего получали клеточные лизаты. Для каждого временного значения в качестве контроля использовали клетки линии PiGAS, нестимулированные IFN-γ. В клеточных лизатах измеряли активность люциферазы при помощи люциферазного теста (Promega, США) и концентрацию общего белка при помощи бицинхониновой кислоты (Sigma-Aldrich, США). Для каждого лизата проводили нормировку значения активности люциферазы на концентрацию общего белка. Все измерения проводили в четырех повторах. Результаты представляли в виде отношения нормированной активности люциферазы в лизатах клеток, стимулированных IFN-γ, к нормированной активности люциферазы в лизатах клеток, нестимулированных IFN-γ. В лизатах клеточной линии PiGAS достоверно детектируется увеличение активности люциферазы (по сравнению с нестимулированными клетками) спустя 4 часа после стимуляции клеток биоактивным IFN-γ в концентрациях 2,5 нг/мл и 10 нг/мл.
Примеры осуществления настоящего изобретения
Пример 1. Получение генетической репортерной экспрессионной кассеты для детекции уровня активности IFN-γ /STAT-1-зависимого сигнального каскада, встроенной в вектор, предназначенный для экспрессии в клетках млекопитающих (плазмида pGAS(x6)-Luc//Neo)
Генетическая репортерная экспрессионная кассета 6xGAS-Luc//Neo состоит из. нескольких отдельных структурных блоков, несущих различное функциональное значение. Она включает в себя 1) репортерный ген люциферазы светлячка Photinus pyralis (Luc) под контролем минимального промотора SV40, которому предшествуют 6 повторов сайта связывания GAS (IFN-γ-activated site) транскрипционного фактора. STAT-1 (SEQ ID NO: 1); и 2) селективный маркерный ген неомицин фосфотрансферазы II (Neo) под контролем конститутивного промотора PGK-1 (SEQ ID NO: 2). Репортерный экспрессионный элемент 6xGAS-Luc//Neo обеспечивает возможность детекции активности ΙΡΝ-γ/STAT-1-зависимого сигнального каскада, а наличие гена Neo дает возможность выращивать клетки, содержащие данную плазмиду, на среде с антибиотиком генетицином (G418).
Фрагмент ДНК, включающий в себя элемент экспрессионной репортерной кассеты 6xGAS-Luc//Neo, был получен с использованием традиционных генно-инженерных методов и подходов.
Нуклеотидная последовательность для связывания транскрипционного фактора STAT-1 под названием GAS (IFN-γ-activated site) была взята из промоторной области гена гуанилат-связывающего белка GBP (guanylate-binding protein) [25]. Олигонуклеотиды (5'- TCGAGAGTTTCATATTACTCTAAATCG -3' и 5'-TCGACGATTTAGAGTAATATGAAACTC -3'), которые при отжиге друг на друга дают ДНК-фрагмент, содержащий мономерную последовательность GAS, фланкированную сайтами рестрикции XhoI и SalI, были подвергнуты самолигированию с последующей обработкой полученного мультимерного продукта рестриктазой Salí, ДНК-полимеразой Pfu и рестриктазой XhoI. Полученная смесь мультимеров разного размера была подвергнута электрофорезу в агарозном геле. ДНК-фрагмент размером 162 п. о., соответствующий тандемным повторам из шести последовательностей GAS, был выделен из геля и клонирован в вектор pBluescript-SK(-) (Stratagene, США) по сайтам XhoI и SmaI. Далее фрагмент ДНК 6xGAS (170 п. о.) был вырезаны из плазмиды pBluescript-SK(-) по сайтам XhoI и ВаmHI и клонирован в вектор pGL3promoter (Promega, США) по сайтам XhoI и BglII.
В полученную плазмиду pGAS(x6)-Luc по сайтам SalI и ВаmHI был также вставлен фрагмент, кодирующиий ген неомицин фосфотрансферазы II (Neo) под контролем конститутивного промотора PGK-1, предварительно вырезанный из плазмиды pΡΝΤ по сайтам XhoI и ВаmHI. Ген Neo кодирует селективный маркер - ген устойчивости к антибиотику генитицину (G418), позволяющий проводить селекцию в клетках млекопитающих при получении клонов клеток, содержащих стабильно интегрированную в геном репортерную конструкцию 6xGAS-Luc//Neo.
Пример 2. Проверка функциональной активности полученной генетической конструкции pGAS(x6)-Luc//Neo, содержащей ген люциферазы светлячка Photinus pyralis (Luc) под контролем STAT-1-зависимого промотора и ген неомицин фосфотрансферазы II (Neo) под контролем конститутивного промотора PGK-1.
Проверка функциональной активности плазмиды pGAS(x6)-Luc//Neo осуществлялась в два этапа с использование клеточной линии меланомы человека MelP. На первом была доказана функциональная активность экспрессинной кассеты 6xGAS-Luc, состоящей из 6 повторов сайта связывания GAS (IFN-γ-activated site) транскрипционного фактора STAT-1, минимального промотора SV40 и репортерного гена люциферазы светлячка Photinuspyralis (Luc).
Для этого клетки линии MelP трансфицировали смесью опытной (pGAS(x6)-Luc//Neo) и контрольной (pRL-CMV (Promega, США)) плазмид. Контрольная плазмида кодирует ген люциферазы Renilla под контролем конститутивного промотора и служит для оценки жизнеспособности клеток, эффективности трансфекции и используется для нормировки сигнала от люфицеразы светлячка (Luc). Трансфекцию проводили при помощи набора для липофекции эукариотических клеток Unifectin-56 (UnifectGroup, Россия) согласно рекомендациям производителя. Трансфицированные клетки обрабатывали рекомбинантным белком IFN-γ человека в концентрации 10 нг/мл или бычьим сыворочным альбумином в концентрации 100 нг/ мл (в качестве отрицательного контроля) в течение 24 часов. В присутствии IFN-γ происходит фосфорилирование и активация транскрипционного фактора STAT-1, в результате чего фосфо-STAT-1 формирует гомодимеры, транслоцируется в ядро и связывается с нуклеотидной последовательностью GAS в промоторной области репортерного гена люциферазы светлячка (Luc), тем самым запуская экспрессию этого гена с последующей наработкой люциферазы. Активность люциферазы в клеточных лизатах анализировали при помощи двойного люциферазного теста согласно рекомендациям производителя (Promega, США). Результаты представляли в виде отношения активности люциферазы светлячка Photinus pyralis (Luc) к активности люциферазы Renilla. По результатам данного теста было установлено, что в присуствии IFN-γ детектируется тридцатисемикратное увеличение уровня активности люциферазы, что подтверждает функциональную активность экспрессинной кассеты 6xGAS-Luc.
На втором этапе оценки активности плазмиды pGAS(x6)-Luc//Neo была доказана функциональность экспрессионного элемента, кодирующего ген неомицин фосфотрансферазы II (Neo) под контролем конститутивного протомора PGK-1. Клетки линии MelP трансфицировали плазмидой pGAS(x6)-Luc//Neo, а также плазмидой pGAS(x6)-Luc при помощи набора для липофекции эукариотических клеток Unifectin-56 (UnifectGroup, Россия) согласно рекомендациям производителя. Плазмида pGAS(x6)-Luc была использована в качестве отрицательного контроля, поскольку в ней отсутствует экспрессионный элемент PGK-1-Neo. Трансфицированные клетки культивировали в ростовой среде (среда RPMI-1640 с 10% эмбриональной бычьей сыворотки, 2 мМ L-Глутамина, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина) в течение 24 часов, после чего получали клеточные лизаты и осуществляли анализ наличия в них неомицин фосфотрансферазы II при помощи вестерн-блот анализа со специфическими антителами (Millipore, США). Вестерн-блот анализ показал наличие белка с молекулярной массой около 29 кДа, что соответствует молекулярной массе к неомицин фосфотрансферазы II.
Пример 3. Получение новой клеточной линии PiGAS, содержащей стабильно интегрированную в геном генетическую конструкцию 6xGAS-Luc//Neo
Для получения клонов клеток со стабильно интегрированной в геном репортерной кассетой 6xGAS-Luc//Neo использовали клетки линии меланомы человека MelP, трансфицированные плазмидой pGAS(x6)-Luc//Neo.
Трансфекцию клеток осуществляли при помощи набора для липофекции эукариотических клеток Unifectin-56 (UnifectGroup, Россия) согласно рекомендациям производителя. Селекцию клонов проводили в ростовой среде (среда RPMI-1640 с 10% эмбриональной бычьей сыворотки, 2 мМ L-Глутамина, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина), содержащей 800 мкг/мл антибиотика G418. После формирования резистентными клетками колоний последние переносили в 96-луночный планшет. Те клоны, которые покрывали дно лунки 96-луночного планшета на 50-80%, переносили в лунки 24-луночного планшета, где культивировали до состояния, когда клон клеток достигал монослоя, после чего переносили в лунки 6-луночного планшета..
После формирования в лунках 6-луночного планшета монослоя из клона клеток линии меланомы человека MelP, трансфицированных плазмидой pGAS(x6)-Luc//Neo, проводили анализ способности клона продуцировать люциферазу светлячка в ответ на обработку IFN-γ. Для этого клетки каждого клона высаживали в 8 лунок 96-луночного планшета из расчета 30000 клеток в лунку. Спустя 4 часа, убедившись, что клетки прикрепились к дну лунок и восстановили распластанную морфологию, меняли культуральную среду в 4 лунках на свежую ростовую среду, а в других 4 лунках на свежую ростовую среду с добавлением 10 нг/мл IFN-γ. После чего культивировали клетки в течение 24 часов. По окончании культивирования ростовую среду удаляли, клетки промывали однократным раствором PBS, после чего лизировали. В клеточных лизатах определяли активность люциферазы светлячка Photinus pyralis при помощи люциферазного теста и концентрацию общего белка при помощи бицинхониновой кислоты (Sigma-Aldrich, США). Далее значения люциферазной активности нормировали на концентрацию общего белка и делали вывод о способности каждого из клонов отвечать на стимуляцию IFN-γ. По результатам проверки полученных клонов клеток линии меланомы человека MelP, трансфицированных плазмидой pGAS(x6)-Luc//Neo, был отобран клон, для которого было характерное оптимальное соотношение сигнал (нормированная активность люциферазы в клетках, стимулированных IFN-γ)/ шум (нормированная активность люциферазы в нестимулированных клетках). Клон получил название PiGAS. Клетки клона PiGAS выращивали в чашке Петри диаметром 6 см, после чего в чашке Петри 10 см.
Пример 4. Определение предела чувствительности клеточной линии PiGAS к IFN-γ
Для определения предела чувствительности клеточной линии PiGAS к IFN-γ клетки линии PiGAS высаживали в лунки 96-луночного планшета из расчета 30000 клеток в лунку и культивировали в течение 16 часов в ростовой среде (RPMI-1640 с 10% эмбриональной бычьей сыворотки, 2 мМ L-Глутамина, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина). Далее ростовую среду в лунках меняли на свежую с добавлением рекомбинантного IFN-γ в концентрации от 0,01 нг/мл до 10 нг/мл и продолжали культивирование в течение следующих 24 часов. По окончании культивирования ростовую среду удаляли, клетки промывали однократным раствором. PBS, после чего лизировали. В клеточных лизатах определяли активность люциферазы светлячка Photinus pyralis при помощи люциферазного теста и концентрацию общего белка при помощи бицинхониновой кислоты (Sigma-Aldrich, США). Значения люциферазной активности нормировали на концентрацию общего белка. Результаты, представляли в виде графика зависимости относительной люциферазной активности, отражающей отношение активности люциферазы светлячка Photinus pyralis (Luc) к концентрации общего белка, от концентации рекомбинантного IFN-γ человека.
Кроме того, в полученных лизатах также анализировали наличие фосфорилированной и нефосфорилированной форм STAT-1 при помощи вестерн-блот анализа с использованием специфических антител к STAT-1 (Cell Signaling, США) и фосфо-STAT-1 (Cell Signaling, США). Наличие фосфорилированной формы STAT-1 служит косвенным признаком активации IFN-γ/STAT-1-сигнального пути и указывает на биоактивность IFN-γ.
Клеточная линия PiGAS, содержащая стабильно интегрированную в геном репортерную экспрессионную конструкцию 6xGAS-Luc//Neo, достоверно детектирует биоактивный IFN-γ в концентрации не менее 0,05 нг/мл. Таким образом, клеточная линия PiGAS может быть использована для определения биоактивного IFN-γ.
Пример 5. Определение оптимального времени воздействия IFN-γ на клеточную линию PiGAS
Для определения оптимального времени воздействия биоактивного IFN-γ на клетки линии PiGAS, достаточного для индукции люциферазной активности репортерной кассеты, стабильно интегрированной в геном клеток PiGAS, клетки линии PiGAS высаживали в лунки 96-луночного планшета из расчета 30000 клеток в лунку и культивировали в течение 16 часов в ростовой среде (RPMI-1640 с 10% эмбриональной бычьей сыворотки, 2 мМ L-Глутамина, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина) до достижения 65-85% монослоя, после чего среду в лунках меняли на свежую с добавлением рекомбинантного IFN-γ человека в концентрации 2,5 нг/мл, либо 10 нг/мл. Стимуляцию клеток IFN-γ проводили в течение различных интервалов времени: 2, 4, 6, 8, 10 и 24 часа. При этом для каждого временного значения в качестве контроля использовали клетки линии PiGAS, нестимулированные IFN-γ. Далее ростовую среду удаляли, клетки промывали однократным раствором PBS, получали клеточные лизаты, в которых определяли активность люциферазы светлячка при помощи люциферазного теста и концентрацию общего белка при помощи бицинхониновой кислоты (Sigma-Aldrich, США). Значения люциферазной активности нормировали на концентрацию общего белка. Все измерения проводили в четырех повторах. Результаты представляли в виде графика зависимости относительной люциферазной активности, отражающей отношение нормированной активности люциферазы в лизатах клеток, стимулированных IFN-γ, к нормированной активности люциферазы в лизатах клеток, нестимулированных IFN-γ, от времени воздействия IFN-γ.
Для стимуляции клеток линии PiGAS биоактивным IFN-γ (в концентрации как 2,5 нг/мл, так и 10 нг/мл) минимальным эффективным временем воздействия является значение 4 часа. При инкубации клеток линии PiGAS с биоактивным IFN-γ в течение 6 часов значение детектируемой относительной люциферазной активности достигает максимума. Увеличение времени воздействия свыше 6 часов нецелесообразно, поскольку не приводит к дальнейшему росту детектируемой относительной люциферазной активности.
Родословная клеточной линии;
Линия клеток PiGAS получена из клеток линии меланомы человека MelP [26] путем трансфекции их плазмидой pGAS(x6)-Luc//Neo.
Трансфицированные клетки содержат стабильно интегрированную в геном репортерную конструкцию 6xGAS-Luc, состоящую из 6 повторов сайта связывания транскрипционного фактора STAT-1, минимального промотора SV-40 и гена люциферазы светлячка Photinus pyralis (Luc), а также ген устойчивости к селективному антибиотику G418 под контролем своего конститутивного промотора PGK-1.
Число пассажей:
К моменту составления паспорта клеточная линия прошла более 15 пассажей.
Стандартные условия культивирования:
Питательная среда RPMI-1640, содержащая 10% эмбирональной бычьей сыворотки, 2 мМ L-глутамина и смесь антибиотиков (пенициллин - 100 ед/мл, стрептомицин - 100 мкг/мл, G418 - 400 мкг/мл). Культивирование осуществляется при 37°С в атмосфере 5% СО2. Пассаж 1 раз в 3-4 суток. Кратность рассева 1:4-1:6, оптимальная плотность 15000-20000 клеток/см2. Метод снятия: 0,05% раствор трипсина.
Культуральные свойства:
Клетки имеют преимущественно адгезионный, преимущественно монослойный характер роста в стандартных условиях культивации.
Маркерные признаки:
Функциональный ответ в люциферазном тесте, основанном на продукции люциферазы светлячка Photinus pyralis в ответ на обработку клеток IFN-γ в концентрации не менее 0,05 нг/мл в течение не менее 18 часов или в концентрации 10 нг/мл в течение не менее 4 часов. Устойчивость к антибиотику G418 (800 мкг/мл).
Контаминация:
Бактерии и грибы в культуре не обнаружены при длительном наблюдении. Тест на микоплазму отрицателен.
Условия криоконсервации:
Среда для замораживания: 80% ростовая среда, 10% эмбриональная бычья сыворотка, 10% DMSO. Количество замораживаемых клеток: 1,0-2,0×106 клеток/мл, 1 мл клеточной суспензии в ампуле. Режим замораживания: жидкий азот, снижение температуры на 1°С в минуту до -70°С. Хранение в жидком азоте при температуре -196°С. Размораживание быстрое, при 37°С. Клетки разводят в 5 мл ростовой среды и осаждают центрифугированием, ресуспендируют в 5 мл той же среды и переносят в культуральный флакон с площадью роста 25 см2. Жизнеспособность клеток оценивают по включению трипанового синего. Жизнеспособность клеток после размораживания составляет не менее 70%.
Список процитированной литературы
[1] A. Bufe, К. Gehlhar, Ε. Grage-Griebenow, and M. Ernst, "Atopic phenotype in children is associated with decreased virus-induced interferon-alpha release.," Int. Arch. Allergy Immunol, vol. 127, no. 1, pp.82-8, Jan. 2002.
[2] F.-C. Lin and H.A. Young, "The talented interferon-gamma," Adv. Biosci. Biotechnol, vol. 04, no. 07, pp. 6-13, 2013.
[3] S. Pestka, "The Interferons: 50 Years after Their Discovery, There Is Much More to Learn," J. Biol. Chem., vol. 282, no. 28, pp. 20047-20051, 2007.
[4] J.R. Schoenborn and С.B. Wilson, "Regulation of interferon-gamma during innate and adaptive immune responses.," Adv. Immunol, vol. 96, pp. 41-101, Jan. 2007.
[5] K. Schroder, P.J. Hertzog, T. Ravasi, and D.A. Hume, "Interferon-gamma: an overview of signals, mechanisms and functions.," J. Leukoc. Biol, vol. 75, no. 2, pp. 163-89, Feb. 2004.
[6] X. Hu and L. B. Ivashkiv, "Cross-regulation of signaling pathways by interferon-gamma: implications for immune responses and autoimmune diseases.," Immunity, vol. 31, no. 4, pp. 539-50, Oct. 2009.
[7] J.F. Bromberg, C.M. Horvath, Z. Wen, R.D. Schreiber, and J.E. Darnell, "Transcriptionally active Stat1 is required for the antiproliferative effects of both interferon alpha and interferon gamma.," Proc. Natl. Acad. Sci. U.S.Α., vol. 93, no. 15, pp. 7673-8, Jul. 1996.
[8] Y.E. Chin, M. Kitagawa, K. Kuida, R.A. Flavell, and X.Y. Fu, "Activation of the STAT signaling pathway can cause expression of caspase 1 and apoptosis.," Mol. Cell. Biol, vol. 17, no. 9, pp. 5328-37, Sep.1997.
[9] T.E. Battle, R.A. Lynch, and D.A. Frank, "Signal Transducer and Activator of Transcription 1 Activation in Endothelial Cells Is a Negative Regulator of Angiogenesis," Cancer Res., vol. 66, no. 7, pp. 3649-3657, Apr. 2006.
[10] S. Nguyen, V. Beziat, N. Dhedin, M. Kuentz, J.P. Vernant, P. Debre, and V. Vieillard, "HLA-E upregulation on IFN-gamma-activated AML blasts impairs CD94/NKG2A-dependent NK cytolysis after haplo-mismatched hematopoietic SCT.," Bone Marrow Transplant., vol. 43, no. 9, pp.693-9, May 2009.
[11] L.  , M. Corvaisier, В. Charreau, A. Moreau, E. Godefroy, A. Moreau-Aubry, F. Jotereau, and N. Gervois, "Expression and release of HLA-E by melanoma cells and melanocytes: potential impact on the response of cytotoxic effector cells.," J. Immunol, vol. 177, no. 5, pp.3100-7, Sep.2006.
, M. Corvaisier, В. Charreau, A. Moreau, E. Godefroy, A. Moreau-Aubry, F. Jotereau, and N. Gervois, "Expression and release of HLA-E by melanoma cells and melanocytes: potential impact on the response of cytotoxic effector cells.," J. Immunol, vol. 177, no. 5, pp.3100-7, Sep.2006.
[12] A.E.  , A.D. Chernisheva, I.R. Zakeeva, A.B. Danilova, A.O. Danilov, V.M. Moiseenko, D. Geraghty, Ν. V. Gnuchev, G.P. Georgiev, Α. V Kibardin, and S. S. Larin, "[HLA-E molecule induction on the surface of tumor cells protects them from cytotoxic lymphocytes]," Vopr. Onkol, vol. 55, no. 2, pp.224-9, Jan. 2009.
, A.D. Chernisheva, I.R. Zakeeva, A.B. Danilova, A.O. Danilov, V.M. Moiseenko, D. Geraghty, Ν. V. Gnuchev, G.P. Georgiev, Α. V Kibardin, and S. S. Larin, "[HLA-E molecule induction on the surface of tumor cells protects them from cytotoxic lymphocytes]," Vopr. Onkol, vol. 55, no. 2, pp.224-9, Jan. 2009.
[13] T. Sareneva, J. Pirhonen, K. Cantell, N. Kalkkinen, and I. Julkunen, "Role of N-glycosylation in the synthesis, dimerization and secretion of human interferon-gamma.," Biochem. J., vol. 303 (Pt 3, pp.831-40, Nov. 1994.
[14] I.A. Braude, "Purification of natural human immune interferon induced by A-23187 and mezerein.," Methods Enzymol, vol. 119, pp. 193-9, Jan. 1986.
[15] Енсен Анна Дам (DK), Патент РФ 2296130 "Варианты полипептида гамма-интерферона", 2002.
[16] D.С. James, M.H. Goldman, M. Hoare, N. Jenkins, R.W. Oliver, B.N. Green, and R.B. Freedman, "Posttranslational processing of recombinant human interferon-gamma in animal expression systems.," Protein Set, vol. 5, no. 2, pp. 331-40, 1996.
[17] E.M. Curling, P.M. Hayter, A.J. Baines, A.T. Bull, K.Gull, P.G. Strange, and N. Jenkins, "Recombinant human interferon-gamma. Differences in glycosylation and proteolytic processing lead to heterogeneity in batch culture.," Biochem. J., vol. 272, no. 2, pp. 333-7, Dec. 1990.
[18] T. Sareneva, K. Cantell, L. Pyhâla, J. Pirhonen, and I. Julkunen, "Effect of carbohydrates on the pharmacokinetics of human interferon-gamma.," J. Interferon Res., vol. 13, no. 4, pp.267-9, Aug. 1993.
[19] D. Wang, H. Ren, J.-W. Xu, P.-D. Sun, and X.-D. Fang, Expression, purification and characterization of human interferon-γ in Pichia pastoris," Mol. Med. Rep., vol. 9, no.2, pp.715-719, Feb. 2014.
[20] А.И. Закабунин, Г.А. Барановская, Η.M. Пустошилова, В.Φ. Майстренко, Л.П. Гаврюченкова и О.А. Громова, Патент РФ 2132386 "Способ получения рекомбинантного интерферона-гамма человека", 1999.
[21] R. Mironova, T. Niwa, R. Dimitrova, M. Boyanova, and I. Ivanov, "Glycation and post-translational processing of human interferon-gamma expressed in Escherichia coli.," J. Biol. Chem., vol. 278, no. 51, pp. 51068-74, Dec. 2003.
[22] R. Mironova, T. Niwa, H. Hayashi, R. Dimitrova, and I. Ivanov, "Evidence for non-enzymatic glycosylation in Escherichia coli.," Mol. Microbiol, vol. 39, no. 4, pp. 1061-8, Feb. 2001.
[23] A.S. Eble, S.R. Thorpe, and J.W. Baynes, "Nonenzymatic glucosylation and glucose-dependent cross-linking of protein.," J. Biol. Chem., vol. 258, no. 15, pp. 9406-12, Aug. 1983.
[24] Министерство здравоохранения РФ, Общая фармакопейная статья "Биологические методы испытания препаратов интерферона с использованием культур клеток".
[25] Т. Decker, D.J. Lew, Y.S. Cheng, D.Ε. Levy, and J.Ε. Darnell, "Interactions of alpha- and gamma-interferon in the transcriptional regulation of the gene encoding a guanylate-binding protein.," EMBO J., vol. 8, no. 7, pp. 2009-14, Jul. 1989.
[26] О.С. Бурова, С.С. Ларин, Г.Η. Ворожцов, И.Н. Михайлова, А.Ю. Барышников, Л.Ф. Морозова, Г.П. Георгиев, Н.В. Гнучев, Л.В. Демидов, Т.Н. Палкина и А.М. Козлов, Патент РФ 2287575 "Клеточная линия меланомы человека mel p, используемая для получения противоопухолевых вакцин", 2005.
Линия клеток меланомы человека PiGAS для определения биологической активности IFN-γ с регистрационным номером ВКПМ Н-161, полученная путем трансфекции родительской линии клеток меланомы человека MelP плазмидой pGAS(x6)-Luc//Neo, кодирующей репортерную конструкцию 6xGAS-Luc, состоящую из последовательности SEQ ID NO: 1, включающую 6 повторов сайта связывания транскрипционного фактора STAT-1, минимального промотора SV-40 и гена люциферазы светлячка Photinus pyralis (Luc), а также последовательности SEQ ID NO: 2, включающую ген устойчивости Neo к селективному антибиотику G418 под контролем своего конститутивного промотора PGK-1.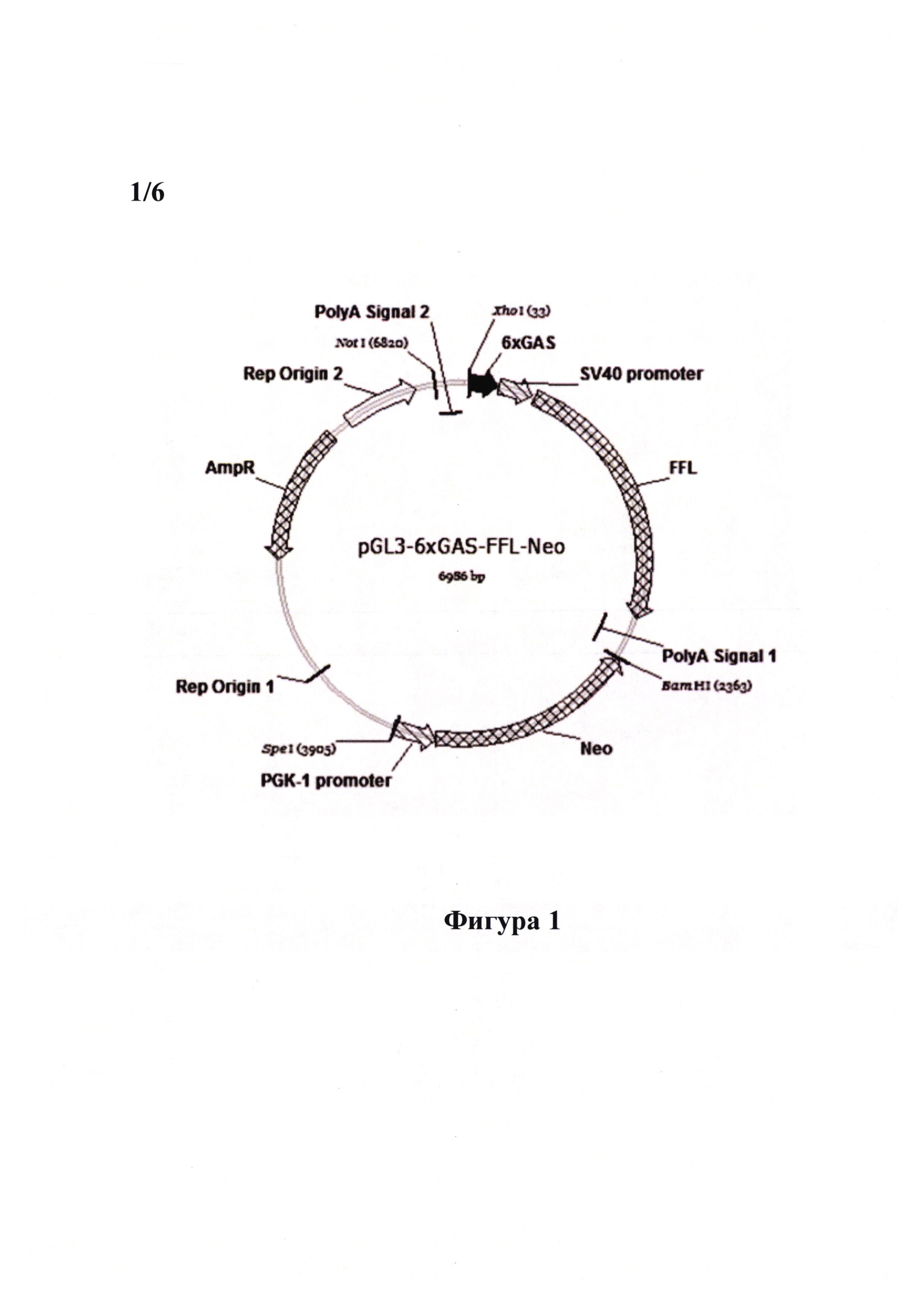


Многопрофильный промотор, экспрессирующий вектор и способ избирательного убийства раковых клеток с их использованием
Генетические конструкции ltf3, ltf5, ltf7, ltf10, ltf11 для получения рекомбинантного лактоферрина человека (варианты)
Линия клеток cho[v3d], секретирующих рекомбинантный фактор роста эндотелия сосудов (vegf) человека, изоформа а165, с экзогенным 3×ded-эпитопом
Способ разделения лактоферринов человека и козы с помощью дифференциальной иммуноаффинной хроматографии с использованием однодоменных мини-антител
Клеточная линия рака почки человека ibgvat r 6
Клеточная линия рака почки человека ibgvat r 27
Многопрофильный промотор, экспрессирующий вектор и способ избирательного убийства раковых клеток с их использованием
Генетические конструкции ltf3, ltf5, ltf7, ltf10, ltf11 для получения рекомбинантного лактоферрина человека (варианты)
Линия клеток cho[v3d], секретирующих рекомбинантный фактор роста эндотелия сосудов (vegf) человека, изоформа а165, с экзогенным 3×ded-эпитопом
Способ разделения лактоферринов человека и козы с помощью дифференциальной иммуноаффинной хроматографии с использованием однодоменных мини-антител
Клеточная линия рака почки человека ibgvat r 6
Клеточная линия рака почки человека ibgvat r 27
Противоопухолевый препарат
Клеточная линия меланомы человека mel ibr, используемая для получения противоопухолевых вакцин
Клеточная линия меланомы человека mel il, используемая для получения противоопухолевых вакцин
Клеточная линия меланомы человека mel p, используемая для получения противоопухолевых вакцин

