Результат интеллектуальной деятельности: НОВЫЙ РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM CANESCENS PEP3 И ПОЛУЧЕНИЕ НА ЕГО ОСНОВЕ КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА ПРОТЕАЗ ЭНДО- И ЭКЗОДЕЙСТВИЯ
Вид РИД
Изобретение
Изобретение относится к области биотехнологии и может быть использовано в пищевой и кормовой промышленности для получения комплексного препарата грибных протеаз эндо- и экзодействия (пенициллопепсина, пролилспецифичной эндопептидазы и лейцинаминопептидазы), с целью дальнейшего применения для получения белковых гидролизатов из растительного сырья.
Благодаря высокой активности кислой аспартатной протеазы - пенициллопепсина, обладающей широкой субстратной специфичностью и способностью к ограниченному гидролизу высокомолекулярных белковых субстратов, способности лейцинаминопептидазы к удалению N-концевых гидрофобных аминокислот, придающих пептидам горький вкус, а также способности пролилспецифичной эндопептидазы эффективно гидролизовать пептидные связи, соединяющие остатки пролина, изобретение может использоваться для получения белковых гидролизатов без горечи, с повышенными органолептическими свойствами и улучшенной усвояемостью.
Грибные протеазы, благодаря уникальной специфичности действия, обеспечивающей получение продуктов с заданными свойствами, нашли широкое применение в пищевой промышленности.
Основной проблемой при получении белковых продуктов является горечь белковых гидролизатов, обусловленная присутствием остатков гидрофобных аминокислот и пролина в пептидах. Пролин - циклическая аминокислота, играет ключевую физиологическую роль и защищает пептиды от ферментативного гидролиза, что может вызывать такие заболевания, как депрессия, болезнь Паркинсона и целиакия [1].
Для устранения горечи при получении белковых гидролизатов используют протеазы с определенной специфичностью действия. К таким протеазам относятся пепсинподобные аспартатные протеазы, лейцинаминопептидазы (LAP) и пролилспецифичные эндопептидазы.
Пепсинподобные аспартатные протеазы характеризуются широкой субстратной специфичностью и способностью расщеплять пептидные связи между громоздкими гидрофобными остатками [2, 3]. В результате гидролиза белковых субстратов пепсином образуются, преимущественно, мелкие пептиды от 3 до 30 остатков. Несмотря на широкую специфичность, пепсины дают воспроизводимый выход продуктов, т.е. при одинаковых условиях гидролиза белка образуются одинаковые пептиды [4-7]. Наиболее хорошо известными грибными аспартатными протеазами являются аспергиллопепсин и пенициллопепсин [8].
Гомологичный пенициллопепсин (КФ 3.4.23.20), синтезируемый грибами рода Penicillium, обладает широкой субстратной специфичностью, предпочтительно катализирует гидролиз гидрофобных остатков в положении Р1 и Р1', обладает молокосвертывающей способностью [9, 10] применяется в пищевой промышленности для получения пептидов, не образующих горечь [11].
Лейцинаминопептидаза (КФ 3.4.11.1) - экзоаминопептидаза, катализирует расщепление полипептидов, начиная с N-конца с образованием свободных аминокислот, фермент не строго специфичен по отношению к лейцину, но "предпочитает" расположенные на N-конце аминокислоты с разветвленной алифатической цепью [12].
Способность грибных аминопептидаз высвобождать отдельные N-концевые аминокислоты, также как и их термостабильность, делает их перспективными для контроля степени гидролиза и развития вкуса различных белковых субстратов [13, 14]. Так как интенсивность горечи пептида пропорциональна количеству гидрофобных аминокислот на N-конце, LAP используют для удаления единичных или пары гидрофобных аминокислот с N-конца полипептидной цепи при снятии горечи белковых гидролизатов [15-17].
Пепсинподобные аспартатные протеазы и LAP, как правило, являются основными компонентами протеолитического комплекса мицелиальных грибов [18-21].
Пролилспецифичная эндопептидаза (КФ 3.4.21.26) - сериновая эндопептидаза, гидролизующая пролин и, в меньшей степени, аланин в олигопептидах. Фермент гидролизует пептиды с карбоксильной стороны остатков пролина [22].
Пролилспецифичная эндопептидаза - единственный фермент эндодействия, способный эффективно гидролизовать пептидные связи, соединяющие остатки пролина, в богатых пролином белках (глютен, казеин, коллаген и желатин) [1].
В мировой практике основными продуцентами комплексных ферментных препаратов грибных эндо- и экзо-пептидаз являются штаммы рода Aspergillus. Например ФП Flavourzyme 500L (Novozymes, США) получают на основе штамма A. oryzae, ФП Corolase PN-L (АВ enzymes, Финляндия) - на основе штамма A. sojae. Штамм A. oryzae также является продуцентом для получения комплексных препаратов Fungal Protease 500000 и Fungal Protease Concentrate (Genencor International, США), а также Fungal Protease 100 и Protease FNP (Enzeco, США) [23, 24].
Мицелиальные грибы Aspergillus oryzae и Aspergillus sojae являются близкими родственниками. На сегодняшний день в грибах были идентифицированы следующие эндопептидазы: две нейтральные эндопептидазы (NP1 и NP2) [25, 26], щелочная эндопептидаза (ALP) [25, 26] и аспартатная эндопротеаза аспергиллопепсин I (РерО) [27]. Экзопептидазы A. oryzae представлены лейцинаминопептидазами [25, 26], карбоксипептидазами [25], дипептидилпептидазой IV (dppIV) и дипептидилпептидазой V (dppV) [26]. Дипептидилпептидаза IV высвобождает Х-Pro остатки с N-конца и может иметь особое значение для деградации пшеничного глютена, однако коммерческие препараты на основе штаммов A. oryzae, в состав которых входят пролил дипептидилпептидазы, характеризуются высокой стоимостью, что делает их применение нецелесообразным [25,26]. Высокой эффективностью при гидролизе пшеничного глютена характеризуются пролилспецифичные эндопептидазы, не входящие в состав большинства коммерческих комплексных препаратов грибных протеаз [1, 24,26].
Пролилспецифичные эндопептидазы синтезируют Xanthomonas sp., Aeromonas hydrophilic, Pseudomonas sp. KU-22, Sphingomonas capsulate, Lactobacillus helveticus, Halobacterium halobium S9, Myxococcus xanthus, Aspergillus oryzae, A. niger [1, 28, 29].
Однако F. meningosepticum, Xanthomonas sp., A. hydrophila, Pseudomonas sp.KU-22, Myxococcus xanthus относятся к патогенным бактериям, и применение пролилспецифичной эндопептидазы, полученной на их основе, в пищевой промышленности не допускается [1].
В пивоваренной промышленности для стабилизации пива широко применяется ФП Brewers Clarex (DSM, США), содержащий пролилспецифичную эндопептидазу A.niger [30]. Однако ФП Brewers Clarex характеризуется низкой активностью пролилспецифичной протеазы (≥5 PPU/g) при высокой стоимости препарата.
В России единственными продуцентами комплекса грибных эндо- и экзопептидаз являются штаммы A. oryzae 12 и A. oryzae 107, полученные на основе A. oryzae 387 (ВКПМ F-683) путем многоступенчатой селекции с использованием эффективных методов мутагенеза [31, 32]. Широкому внедрению производства ФП на основе указанных штаммов препятствует недостаточный уровень протеолитической активности продуцентов. Наиболее близким по технической сущности к предлагаемому изобретению является мутантный штамм A. oryzae 107 (ВКПМ F-929), являющийся продуцентом комплекса кислых и слабокислых протеаз [32].
При глубинном культивировании штамма A. oryzae 107 на средах на основе ячменной муки (3%), пшеничных отрубей (3%) и KH2PO4 (1,5%), в течение 40 ч в зависимости от температуры культивирования 30 и 32°C, активность внеклеточных протеаз, определенная модифицированным методом Ансона с использованием в качестве субстрата бычьего гемоглобина (при рН 4,7), составляет 20,8 -24,8 ед. ПС/мл.
Техническая задача, на решение которой направлено данное изобретение, состоит в получении высокоактивного рекомбинантного штамма мицелиального гриба - продуцента комплекса грибных протеаз эндо- и экзодействия для применения в процессах преработки зернового сырья.
Технический результат от предлагаемого изобретения состоит в использовании нового рекомбинантного штамма и высокоактивного ферментного препарата пенициллопепсина, лейцинаминопептидазы и пролилспецифичной эндопептидазы, полученного на его основе, в процессах переработки растительного белок-содержащего сырья в белковые гидролизаты без горечи, с повышенными органолептическими свойствами и улучшенной усвояемостью для различных отраслей АПК.
Сущность объекта изобретения - новый рекомбинантный штамм гриба Penicillium canescens РЕР3 - продуцент комплекса ферментов: пенициллопепсина, лейцинаминопептидазы и пролилспецифичной пептитидазы, и комплексный ферментный препарат протеаз эндо- и экзодействия на основе P. canescens РЕР3.
Изобретение обеспечивает получение высокоактивного комплексного ФП грибных протеаз эндо- и экзодействия в результате глубинного культивирования нового рекомбинантного штамма P. canescens РЕР3 на ферментационных средах на основе соевой шелухи и кукурузного экстракта.
Рекомбинантный штамм P. canescens РЕР3 создан на основе ауксотрофного штамма P. canescens RN3-11-7 NiaD-, являющегося мутантом штамма P. canescens ВКПМ F-178 [33], путем трансформации экспрессионными плазмидами:
- pPrXyl-PepA, содержащей гомологичный ген pepА, кодирующий пенициллопепсин P. canescens;
- pPrAbf-Lap1, несущей ген lap1, кодтрующий лейцинаминопептидазу А. oryzae;
- pPrXyl-Epo, содержащей ген epo1, кодирующий пролилспецифичную пептидазу A. niger.
Заявляемый штамм Penicillium canescens РЕР3 депонирован во Всероссийской коллекции микроорганизмов (ВКМ) Института биохимии и физиологии микроорганизмов им. Г.К. Скрябина РАН под №ВКМ F-4662D.
Штамм Penicillium canescens РЕР3 (ВКМ F-4662D) характеризуется следующими признаками:
Культурально-морфологические и физиолого-биохимические особенности штамма
Температурный оптимум роста - 28-30°C, рН 4,5-6,0. Аэроб.
Среда Чапека с дрожжевым автолизатом (CYA), 26°C. Диаметр колонии 40-45 мм, отчетливо радиально складчатая, средней плотности, ростовая зона плотная, шириной 1,5-2,0 мм. Мицелий белый, конидиальная зона голубовато-серая, конидиогенез обильный. Экссудат и растворимый пигмент отсутствуют. Обратная сторона палевая, буроватая до темно-коричневой.
Среда Мальц-агар (МА), 26°C. Диаметр колонии 30-35 мм, отчетливо радиально складчатая, средней плотности, ростовая зона плотная, шириной 1,5-2,0 мм. Мицелий белый, пушистый, конидиальная зона голубовато-серая, конидиогенез средний. Экссудат и растворимый пигмент отсутствуют. Обратная сторона палевая, в центре слегка оранжевая.
Среда Чапека с глицерином (GN25), 26°C. Диаметр колонии 16-20 мм, отчетливо радиально складчатая, средней плотности, ростовая зона плотная, шириной 1,5-2,0 мм. Мицелий белый, прижатый, конидиальная зона голубовато-серая, конидиогенез слабый. Экссудат и растворимый пигмент отсутствуют. Обратная сторона светлая.
Конидиеносцы двухъярусные, терминальные, фуркатные, шероховатые, длиной более 150 мкм, шириной 3-4 мкм. Метулы шероховатые, 10-18×2,5-3,5 мкм, фиалиды ампуллиформные, 7-8×2,2-2,5 мкм, конидии округлые, иногда овальные, гладкие, 2,2-3,0×2,0-3,0 мкм.
Данный вид мицелиального гриба относится к IV группе патогенности в соответствии с санитарно-эпидемиологическими правилами «Безопасность работы с микроорганизмами III-IV групп патогенности (опасности) и возбудителями паразитарных болезней» (СП 1.3.2322-08), утвержденными Постановлением Главного государственного санитарного врача РФ №4 от 28 января 2008 г.
Полученный рекомбинантный штамм P. canescens РЕР3 отличается от исходного штамма P. canescens RN3-11-7 NiaD- по морфологическим признакам - более интенсивному спороношению при росте на среде РМ и повышенной способностью при глубинном культивировании на жидких средах к биосинтезу пенициллопесина, пролилспецифичной пептидазы и лейцинаминопептидазы.
Штамм может храниться в лиофилизированном состоянии в течение нескольких лет и/или на скошенном агаре со средой SM при +4°C при обязательных пересевах не реже одного раза в течение 5-6 мес.
Среда SM (% масс): глюкоза - 1, KH2PO4 - 1, дрожжевой экстракт - 0,5, агар-агар - 2.
Заявляемый штамм способен синтезировать пенициллопепсин с активностью 50-60 ед/мл, лейцинаминопептидазу с активностью 10-15 ед/мл и пролилспецифичную пептидазу с активностью 0,5-0,6 ед/мл при выращивании в качалочных колбах объемом 750 мл со 100 мл ферментационной среды на качалке (240 об/мин) при 30°C в течение 144 ч. Состав ферментационной среды (% масс): соевая шелуха - 4,5, кукурузный экстракт - 5,0, KН2РО4 - 2,5, рН естественный.
Штамм P.canescens РЕР3 и ферментный препарат, полученный на его основе, будут содержать комплекс протеолитических ферментов эндо- и экзодействия, активный в широком диапазоне рН 3,5-7,0. Преимуществом ферментного препарата РЕР3 будет являться возможность применения в пищевой промышленности в процессах, происходящих при кислых значениях рН в меньших дозировках, чем препараты с нейтральным оптимумом действия. Например, в хлебопечении для повышения качества продукции за счет быстрого расщепления белка на крупные фрагменты при рН 3,5-5,5. В производстве спирта на стадии брожения сусла для улучшения и ускорения процесса сбраживания. В пивоварении для гидролиза белковых веществ при затирании при рН 4,5-5,0 для увеличения выхода экстрактивных веществ в сусло, интенсификации процесса дрожжегенерации и для предотвращения коллоидного помутнения пива, за счет гидролиза белковой составляющей коллоидных частиц, что сокращает расходы за счет исключения этапа холодной стабилизации. Благодаря специфичности действия протеаз препарат также будет гидролизовать глиадины глютена, обладающие антипитательными свойствами.
Возможность использования изобретения иллюстрируется примерами, которые не ограничивают объем и сущность притязаний, связанных с ними.
Пример 1. Получение целевой плазмиды рРеР заключалось в клонировании гена пенициллопепсина из P.canescens (pepA) в универсальный вектор, обеспечивающий экспрессию целевого гена под контролем промотора xylA из P.canescens
Ген рерА получают методом инверсного ПЦР. На первом этапе находят две консервативные области в выравнивании генов, отличающихся наибольшей степенью гомологии с pepА из P.canescens. Затем проводят рестрикцию геномной ДНК P.canescens с использованием эндонуклеазы HindIII, полученный набор фрагментов лигируют. Далее амплифицируют пробу ДНК с использованием инверсных праймеров, комплементарных к консервативным областям:

где I - инозин (универсальный нуклеотид)
Пробу ДНК, размером 1500 п.о. секвенируют по обеим цепям, получая последовательность 5' и 3' концевых участков гена pepА из P.canescens.
Полинуклеотидная последовательность гена рерА и транслируемый полипептид представлены на фиг. 1.
Пример 2. Получение штамма P.canescens РЕР3 ВКМ F-4662D.
Для создания экспрессионных плазмид ПЦР-методом амплифицируют гены целевых ферментов.
Используя в качестве матрицы геномную ДНК P.canescens, амплифицируют полинуклеотидную последовательность размером 1326 п.о., соответствующую целевому гену пенициллопепсина (фиг. 1). Для этого синтезируют следующие праймеры:

Используя в качестве матрицы геномную ДНК A.niger, амплифицируют полинуклеотидную последовательность размером 1578 п.о., соответствующую целевому гену пролилспецифичной пептидазы (AN: АМ270168).
Для этого синтезируют следующие праймеры:
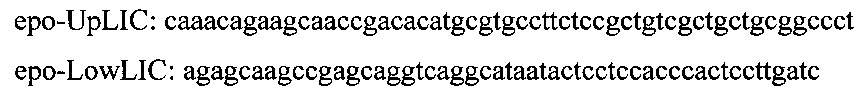
Используя в качестве матрицы геномную ДНК A. oryzae, ПЦР-методом амплифицируют полинуклеотидную последовательность размером 1134 п.о., соответствующую целевому гену лейцинаминопептидазы (AN: ХМ_001825693). Для этого синтезируют следующие праймеры:

Фрагменты ДНК, соответствующие гену пенициллопесина (рерА) и гену пролилспецифичной пептидазы (еро), клонируют в векторы, содержащие полинуклеотидные последовательности, соответствующие промоторной и терминаторной областям гена ксиланазы А P.canescens. Для этого используют метод независимого лигирования, описанный в [34]. Таким образом получают экспрессионные плазмиды PrXyl-PepA и pPrXyl_Epo (фиг. 2 и фиг. 3).
Фрагмент ДНК, соответствующий гену лейцинаминопептидазы (lap1), клонируют в вектор, содержащий полинуклеотидные последовательности, соответствующие промоторной и терминаторной областям гена арабинофуранозидазы P.canescens, с использованием метода независимого лигирования. Таким образом получают экспрессионную плазмиду pPrAbf_Lap1 (фиг. 4).
Трансформацию реципиентного штамма P.canescens RN3-11-7 проводят полученными экспрессионными плазмидами pPrXyl-PepA (5 мкг ДНК), pPrXyl-Epo (5 мкг ДНК), pPrAbf-Lap1 (5 мкг ДНК) совместно с трансформационной плазмидой pSTA10 (1 мкг ДНК), несущей селективный niaD признак.
Трансформанты отбирают на средах с 10 мМ нитрата натрия. Из 20 полученных трансформантов методом PCR отобран 1 клон, содержащий все три целевых гена.
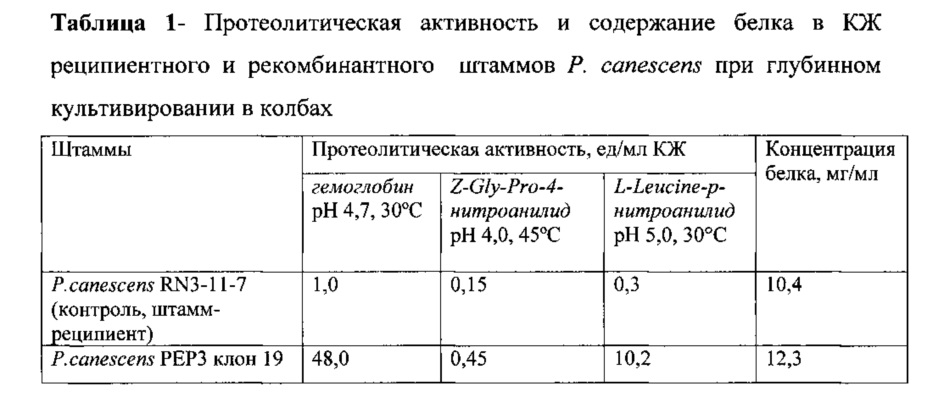
Проверка протеолитических активностей: пенициллопепсина (рН 4,7, 30°C, субстрат гемоглобин), пролилспецифичной пептидазы (рН 4,0, 45°C, субстрат - Z-Gly-Pro-4-нитроанилид) и лейцинаминопептидазы (рН 5,0, 30°C, субстрат - L-Leucine-p-нитроанилид) при глубинном культивировании отобранного трансформанта и реципиентного штамма в колбах (табл. 1) показывает увеличение всех трех активностей у полученного трансформанта.
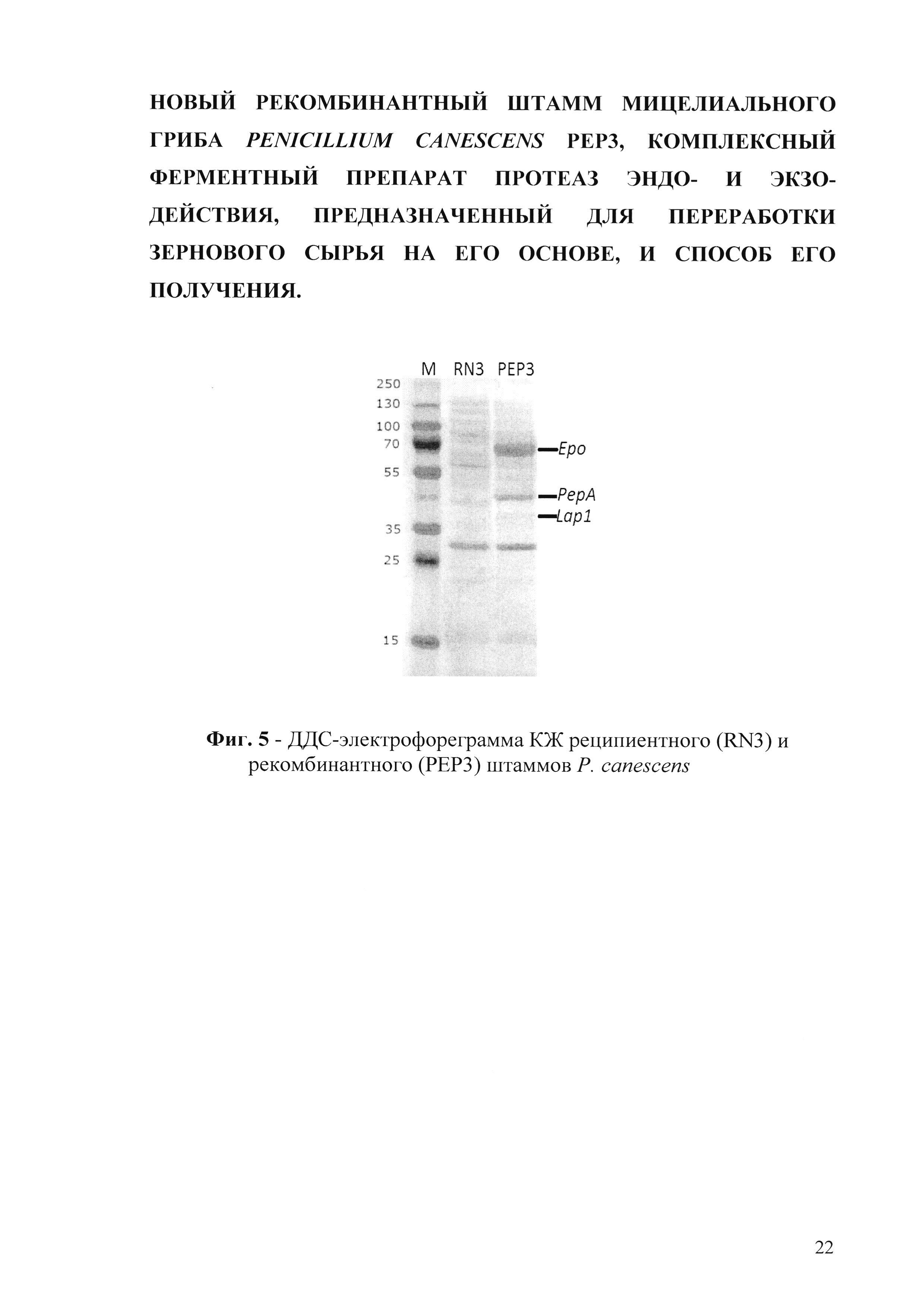
Результаты ДДС-электрофоретического анализа КЖ штаммов Р. canescens подтверждают наличие в рекомбинантном штамме белков с молекулярными массами 66 кДА, что соответствует ожидаемой массе пролилспецифичной пептидазы, 43 кДА, что соответствует массе пенициллопепсина и 37 кДА, что соответствует ожидаемой массе лейцинаминопептидазы (фиг. 5). Таким образом получают штамм Р. canescens РЕР3.
Пример 2. Культивирование штамма P. canescens РЕР3 ВКМ F-4662D в лабораторном ферментере объемом 3 л, оснащенном системой автоматического регулирования рН, pO2 и температуры, а также барботерами для подачи воздуха в аппарат и двухъярусной мешалкой. Получение инокулята и культивирование проводят на ферментационной среде следующего состава, %: соевая шелуха - 4,5, кукурузный экстракт - 5, KН2РО4 - 2,5. В качестве пеногасителя перед стерилизацией вносят 0,1% Лапрола. Ферментеры засевают 10% вегетативного посевного материала, выращенного в колбах при 28-30°C в течение 24 ч. Температура культивирования - 28±0,2°C; рН естественный (4,0-4,2 в начале ферментации, 6,0-6,1 в конце ферментации). Через каждые 24 ч отбирают пробы КЖ и после удаления биомассы определяют активности целевых ферментов и концентрацию растворимого белка.
В культуральной жидкости на 144 ч роста максимальная активность пенициллопепсина составляет 60 ед/мл, активность пролилспецифичной пептидазы - 0,55 ед/мл, активность лейцинаминопептидазы - 15 ед/мл, содержание растворимого белка в культуральной жидкости - 16 мг/мл.
Пример 3. Из культуральной жидкости штамма, наработанной в соответствии с примером 2, получают концентрированный комплексный ферментный препарат методом лиофильного-высушивания ультраконцентрата (табл. 2).
Эффективность полученного ФП оценивают при гидролизе богатых пролином растительных белков (пшеничной муки и пшеничного глютена) в сравнении с ФП Flavourzyme 500L (Novozymes, США). Гидролиз субстратов проводят в пластиковых микропробирках объемом 2 мл при 40°C при постоянном перемешивании. Концентрация субстрата составляет 250 мг/мл, ФП вносят в дозировке 0,5% к массе субстрата (сырья). Концентрацию субстрата выбирают с учетом параметров реальных технологических процессов в пищевой промышленности (пивоварение, хлебопечение и пр.). Через 3 ч гидролиза полученные ферментолизаты инкубируют при 85°C в течение 5 мин для инактивации протеаз и центрифугируют при 10750 g в течение 5 мин. В супернатантах определяют степень гидролиза белка как отношение концентрации низкомолекулярных белковых продуктов к содержанию общего растворимого белка (табл. 3), используя метод Хартри-Лоури [35]. Концентрацию низкомолекулярных белковых продуктов определяют, предварительно осадив высокомолекулярные белковые соединения 20% трихлоруксусной кислотой.
Осадок, полученный при действии ФП на глютен, используют для качественной оценки гидролиза методом ДДС-электрофореза (Фиг. 6).
Применение ФП РЕР3, полученного из КЖ нового рекомбинантного штамма P.canescens РЕР3, при обработке растительного сырья в течение 3 ч обеспечивает степень гидролиза белков пшеницы - 83%, а глютена - 53%, позволяя увеличить степень гидролиза на 4-3% по сравнению с применением коммерческого ФП Flavourzyme 500L.
Результаты ДДС-электрофореза также свидетельствуют о высокой эффективности нового ФП при устранении наиболее трудно гидролизуемых фракций глютена - глиадинов, обладающих антипитательными свойствами при высокой концентрации субстрата.
Используемые источники литературы
1. Kang С., Yu X., Xu Y. Gene cloning and enzymatic characterization of an endoprotease Endo- Pro-Aspergillus niger. J Ind Microbiol Biotechnol. 2013. Vol. 40. P. 855-864.
2. Fruton J.S. The specificity and mechanism of pepsin action. Enzymol. Relat. Areas. Mol. Biol., 1970, V. 33, P. 401-443.
3. Ryle A. The porcine pepsinand pepsinogens. Methods Enzymol, 1970, V. 19, P. 316-336.
4. Zhang Z. and Smith D.L. Determination of amide hydrogen exchange by mass spectrometry: a new tool for protein structure elucidation. Protein Sci., 1993, V. 2, P. 522-531.
5. Voynick I.M. and Fruton J.S. The comparative specificity of acid proteinases. Proceedings of the National Academy of Sciences, 1971, V. 68, №2, P. 257-259.
6. Palashoff M.H. Determining the specificity of pepsin for proteolyticdigestion. Chemistry Master's Theses, 2008, P. 1
7. Davies D.R. The structure and function of the aspartic proteinases. Annu. Rev. Biophys. Biophys. Chern, 1990, V. 19, P. 189-215.
8. Monoda M., Capocciaa S., Le chennea В., Zaugga C., Holdomb M., Jousson O. Secreted proteases from pathogenic fungi. Int. J. Med. Microbiol., 2002, V. 292, P. 405-419.
9. Enzyme Database - BRENDA
URL: http://www.brenda-enzymes.org/enzyme.php?ecno=3.4.23.20
10. Davidson R., Gertler A. and Hofmann T. Aspergillus oryzae acid proteinase. Purification and properties, and formation of 7r-chymotrypsin. Biochem. J., 1975, V. 147, P. 45-53
11. Source Book of Enzymes. John S. White, Dorothy C. White. Published by Taylor Francis Inc, United States, 1997, 1328 P.
12. Enzyme Database - BRENDA
URL: http://brenda-enzymes.org/enzyme.php?ecno=3.4.11.1
13. Nampoothiri K.M., Nagy V., Kovacs K., Szakacs G. and Pandey A. L-leucine aminopeptidase production by filamentous Aspergillus fungi. Letters in Applied Microbiology, 2005, V. 41, P. 498-504.
14. Toldra F., Aristoy A.C., Flores M. Contribution of muscle amino peptidase to flavor development in dry cured ham. Food Res., 2000, Int. 33, P. 181-185.
15. Ney K.H., Retzlaff G. The shelf life of foods and beverage. In: Charalambous G (ed.) Proceedings of the 4th International Flavor Conference. Amsterdam: Elsevier, 1985, P. 54.
16. Rao M.B., Tanksale A.M., Ghatge M.S., Deshpande V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol, and Mol. Biol. Rev, 1998, V. 62(3), P. 597-63573-550.
17. Clemente A. Enzymatic protein hydrolysates in human nutrition. Trends Food Sci Technol., 2000, V. 11, P. 254-262.
18. Sumantha A., Larroche C., Pandey A. Microbiology and Industrial Biotechnology of Food-Grade Proteases: A Perspective. Food Technol. Biotechnol. 2006. V. 44. №2. P. 211-220.
19. Nampoothiri K.M., Nagy V., Kovacs K., Szakacs G. and Pandey A. L-leucine aminopeptidase production by filamentous Aspergillus fungi. Letters in Applied Microbiology. 2005. V. 41. P. 498-504.
20. Ward O.P., Rao M.B., Kulkarni A. Proteases, production. Applied Microbiology: Industrial. 2009. P. 495-511.
21. Shie-Jea Lin, Li-Lin Chen, Chiou-Yen Wen and Wen-Shen Chu. Extracellular leucine aminopeptidase produced by Aspergillus oryzae LL1 and LL2. African Journal of Microbiology Research. 2010. V. 4 (3). P. 158-168.
22. Enzyme Database - BRENDA
URL: http://brenda-enzymes.org/enzyme.php?ecno=3.4.21.26
23. Patent US 20080299607 A1. Enzymatic Process To Produce Highly Functional Soy Protein from Crude Soy Material. 2008.
24. Enzyme Development Corporation
URL: http://www.enzymedevelopment.com/enzymes/proteases/
25. Doumas A., Van Den Broek P., Affolter M., Monod M. Characterization of the Prolyl Dipeptidyl Peptidase Gene (dppIV) from the Koji Mold Aspergillus oryzae. Appl. Environ. Microbiol. 1998. V. 64 (12). P. 4809-4815.
26. Merz M., Eisele Т., Berends P., Appel D., Rabe S., Blank I., Stressler Т., Fischer L. Flavourzyme, an Enzyme Preparation with Industrial Relevance: Automated Nine-Step Purification and Partial Characterization of Eight Enzymes. J Agric Food Chem. 2015. V.63(23). P. 5682-93.
doi: 10.1021/acs.jafc.5b01665.
27. Tsujita, Y., and A. Endo. Purification and characterization of the two molecular forms of Aspergillus oryzae acid protease. Biochim. Biophys. Acta. 1976. V. 445 (1). P. 194-204
28. Patent EP 0522428 A1. Prolyl endopeptidase and production thereof. 1993.
29. Patent US 7323327 B2. Genes encoding novel proteolytic enzymes. 2008.
30. Patent EP 2402425 A1. Improved brewing process. 2012.
31. Патент РФ. 2008. №2315097.
32. Патент РФ. 2008. №2315098.
33. Чулкин A.M., Вавилова Е.А., Беневоленский С.В.
Транскрипционный регулятор углеродной катаболитной репрессии CreA мицелиального гриба Penicillium canescens. Молекулярная биология, 2010, Т. 44(4), с. 1-11.
34. Aslanidis С., de Jong P.J. Ligation-independent cloning of PCR products (LIC-PCR). Nucleic Acid Res. 1990. V. 18. №20. P. 6069-6074.
35. Hartree E.F. Determination of protein: A modification of the lowry method that gives a linear photometric response. Analytical Biochemistry J. 1972. V. 48 (2). P. 422-427 doi: 10.1016/0003-2697(72)90094-2.







Рекомбинантный фрагмент днк, кодирующий альфа-фетопротеин (афп) человека, содержащий "медленные" кодоны, кодирующие лейцин, экспрессионная плазмида, содержащая указанный фрагмент, клетка saccharomyces cerevisiae, трансформированная указанной плазмидой, и способ получения рекомбинантного афп человека
Участок связывания антигена (fab), в том числе гуманизированный fab, против ботулинического нейротоксина с (варианты), способ получения fab с использованием дрожжей, способ и набор для детекции ботулинического нейротоксина с
Способ получения гидролизата из шротов и жмыхов масличных культур
Штаммы-продуценты ферментов и способ получения мультиэнзимной композиции протеолитического и гемицеллюлолитического действия, предназначенной для применения в кормопроизводстве
Рекомбинантный штамм мицелиального гриба penicillium canescens cl14, продуцирующий компонент целллюлосомы clostridium thermocellum, и способ его культивирования
Рекомбинантный штамм мицелиального гриба penicillium verruculosum ( варианты) и способ получения ферментного препарата с его использованием (варианты)
Штамм мицелиального гриба trichoderma longibrachiatum tw-14-220 - продуцент целлюлаз, бета - глюканаз и ксиланаз для кормопроизводства и способ получения кормового комплексного ферментного препарата
Рекомбинантный фрагмент днк, кодирующий альфа-фетопротеин (афп) человека, содержащий "медленные" кодоны, кодирующие лейцин, экспрессионная плазмида, содержащая указанный фрагмент, клетка saccharomyces cerevisiae, трансформированная указанной плазмидой, и способ получения рекомбинантного афп человека
Участок связывания антигена (fab), в том числе гуманизированный fab, против ботулинического нейротоксина с (варианты), способ получения fab с использованием дрожжей, способ и набор для детекции ботулинического нейротоксина с
Способ получения гидролизата из шротов и жмыхов масличных культур
Штаммы-продуценты ферментов и способ получения мультиэнзимной композиции протеолитического и гемицеллюлолитического действия, предназначенной для применения в кормопроизводстве
Рекомбинантный штамм мицелиального гриба penicillium canescens cl14, продуцирующий компонент целллюлосомы clostridium thermocellum, и способ его культивирования
Рекомбинантный штамм мицелиального гриба penicillium verruculosum ( варианты) и способ получения ферментного препарата с его использованием (варианты)
Генетическая конструкция для обеспечения экспрессии комплекса ферментов эндоглюканаз и ксиланаз в клетках гриба penicillium verruculosum и способ получения комплексных ферментных препаратов на его основе, предназначенных для кормопроизводства
Штамм мицелиального гриба trichoderma longibrachiatum tw-14-220 - продуцент целлюлаз, бета - глюканаз и ксиланаз для кормопроизводства и способ получения кормового комплексного ферментного препарата
Гуманизированные антигенсвязывающие фрагменты (fab) против вируса бешенства, изолированный фрагмент днк, кодирующий fab против вируса бешенства, клетка дрожжей, трансформированная фрагментом днк, и способ получения fab против вируса бешенства с использованием дрожжей
Рекомбинантный штамм мицелиального гриба aspergillus awamori - продуцент комплекса ферментов глюкоамилазы и ксиланазы

