Результат интеллектуальной деятельности: Синтетическая ДНК, кодирующая интерлейкин-7 человека, содержащий ее экспрессионный вектор (варианты), штамм-продуцент интерлейкина-7 человека и способ получения интерлейкина-7 человека
Вид РИД
Изобретение
Изобретение относится к биотехнологии, в частности к технологии получения рекомбинантного интерлейкина-7 человека.
Интерлейкин-7 (ИЛ-7) представляет собой цитокин, стимулирующий пролиферацию В- и Т-лимфоцитов и их предшественников, а также пролиферацию и моторную активность лимфангиобластов. Утрата организмом гена ИЛ-7 (нокаут) вызывает опустошение тимуса, тотальную лимфопению, тяжелый иммунодефицит, снижение интенсивности (в 7-10 раз) ответа иммунокомпетентных клеток на действие стимуляторов. При этом полностью блокируется процесс формирования гамма/дельта Т-лимфоцитов, снижается генерация альфа/бета Т-клеток. При избытке ИЛ-7 уровень В- и Т-лимфоцитов в крови повышен. Также установлена физиологическая роль ИЛ-7 в функционировании лимфатической системы [ru.wikipedia.org/wiki/Интерлейкин_7].
Предполагается [Кетлинский С.А., Симбирцев А.С. Цитокины. С.-Пб., 2008], что ИЛ-7 можно будет успешно использовать для лечения ряда патологических состояний, связанных с низким содержанием В- и Т-лимфоцитов, в частности:
- для восстановления содержания Т-клеток, главным образом CD4+, в сочетании с высокоактивной антиретровирусной терапией ВИЧ-инфицированных;
- для лечения лимфопении, связанной с применением цитостатиков и радиационной терапии при лечении онкологических заболеваний;
- для терапии онкологических заболеваний в сочетании с локальной гипертермией;
- для регенерации Т-клеток после пересадки гематопоэтических стволовых клеток;
- для лечения лимфовенозной недостаточности.
Молекула интерлейкина-7 представляет собой гликозилированную полипептидную цепь длиной 152 аминокислотных остатка. В молекуле присутствуют 6 остатков цистеина, образующих три дисульфидных мостика, а также три сайта N-гликозилирования и несколько потенциальных сайтов О-гликозилирования. Секрецию синтезированного белка направляет сигнальная последовательность, содержащая 25 аминокислотных остатков [ru.wikipedia.org/wiki/Интерлейкин_7; Кетлинский С.А., Симбирцев А.С. Цитокины. С.-Пб., 2008].
Получение ИЛ-7 из природных источников в значимых количествах не представляется возможным, поскольку его концентрация в сыворотке не превышает десятков пикограмм на миллилитр. Поэтому для получения ИЛ-7 используются рекомбинантные клетки бактерий, дрожжей, высших эукариот, трансформированные соответствующими векторами, экспрессирующими ИЛ-7 человека. Однако в клетках бактерий синтезируется негликозилированная форма ИЛ-7, отличающаяся нестабильностью при введении in vivo по сравнению с нативной (гликозилированной) формой. ИЛ-7, синтезированный в клетках дрожжей, содержит углеводные цепи, отличающиеся от углеводных цепей нативного белка, что приводит к повышению его иммуногенности [RU 2562169, 2014].
Наиболее перспективными продуцентами ИЛ-7 человека для использования в качестве лекарственного средства могут служить рекомбинантные культивируемые клетки высших эукариот, в частности клетки ВНK, COS, СНО, HEK-293, в которых обеспечиваются синтез, гликозилирование, фолдирование, отщепление сигнальной последовательности и секрецию в среду культивирования белков, идентичных нативным белкам человека. В частности, описан синтез ИЛ-7 человека в ряде культивируемых клеток млекопитающих (WO 2004018681, WO 8903884). В результате были получены клетки HEK-293, COS, BHK, синтезировавшие ИЛ-7 человека при культивировании в течение 10 дней. Идентичность полученных продуктов интерлейкину-7 человека была установлена, однако не приведены какие-либо данные о продуктивности полученных клеток-продуцентов.
Наиболее близкой к используемой технологии получения ИЛ-7 является технология, описанная в патенте WO 2004018681, где описывается получение ИЛ-7 продуцированием клеток яичников китайского хомячка (СНО), HEK-293, клеток человека или бактерии. В частности, приведена структура ДНК, используемые экспрессионные вектора и т.п. Очистка проводится многостадийной хроматографией.
Недостатком данной технологии является относительно невысокий выход ИЛ-7.
Наиболее близким в отношении штамма продуцента является ранее полученный авторами штамм-продуцент ИЛ-7 CHO-IL7/13, полученный трансфекцией клеток CHOdhfr плазмидой pIPvES-DHFR/IL7, содержащей ген интерлейкина-7 человека, с удельной продукцией 6,5 пг на клетку в сутки и волюметрической продуктивностью до 11 мкг/мл (RU 2562169, 2014). Подобный уровень волюметрической продукции является недостаточным для экономически целесообразного производства данного рекомбинантного белка.
Известна технология выделения и очистки рекомбинантного ИЛ-7, произведенного в клетках Е. coli HMS174 (DE3) pLysS под контролем промотора Т7. Полученные нерастворимые тельца были отделены от клеточных остатков фильтрацией с дальнейшей гомогенизацией с 6 М гуанидин гидрохлоридом. Денатурированный ИЛ-7 очищали гель-хроматографией с использованием Prep-Grade Pharmacia Superdex 200, а затем жидкостной хроматографией низкого давления, используя комбинацию гидрофобного взаимодействия, катионообменной и гель-хроматографии [Ouellette Т. et al. Production and purification of refolded recombinant human IL-7 from inclusion bodies // Protein Expr Purif. 2003, 30(2): 156-66].
Недостатком способа является низкий выход целевого продукта, а также получение негликозилированного ИЛ-7.
Известна технология экспрессирования активного рекомбинантного белка IL-7 человека в дрожжах с использованием системы экспрессии Pichia pastoris. ИЛ-7 выделяли из ферментационной надосадочной жидкости, получая 35 мг/л при 95%-ной чистоте с использованием общей SP Sepharose FF катионообменной хроматографии [Luo Y. et al. Expression, purification, and functional characterization of recombinant human interleukin-7 // Protein Expr Purif. 2009, 63(1): 1-4)].
Недостатком способа является получение ИЛ-7, который содержит не встречающиеся у млекопитающих углеводные остатки.
В случае экспрессии ИЛ-7 в клетках млекопитающих присутствие в составе белка сиаловых кислот существенно изменяет изоэлектрическую точку рекомбинантного белка и делает невозможным использование хроматографических способов очистки, разработанных для негликозилированного ИЛ-7, и требует разработки новой технологии выделения и очистки ИЛ-7.
Задачей, стоявшей перед авторами, являлась разработка технологии получения рекомбинантного ИЛ-7 человека с более высоким выходом целевого продукта.
Технической задачей являлись создание эффективных экспрессионных векторов, получение трансфецированных данными векторами клеток яичников китайского хомячка, отбор наиболее продуктивного по данному признаку клона клеток с последующим повышением его продуктивности, разработка оптимальной технологии выделения и очистки рекомбинантного интерлейкина-7.
Технический результат достигался
- созданием синтетической ДНК, кодирующей интерлейкин-7 человека, содержащей ген интерлейкина-7 человека с кодонным составом, оптимизированным для экспрессии в клетках яичника китайского хомячка, и имеющей последовательность Seq ID No. 1;
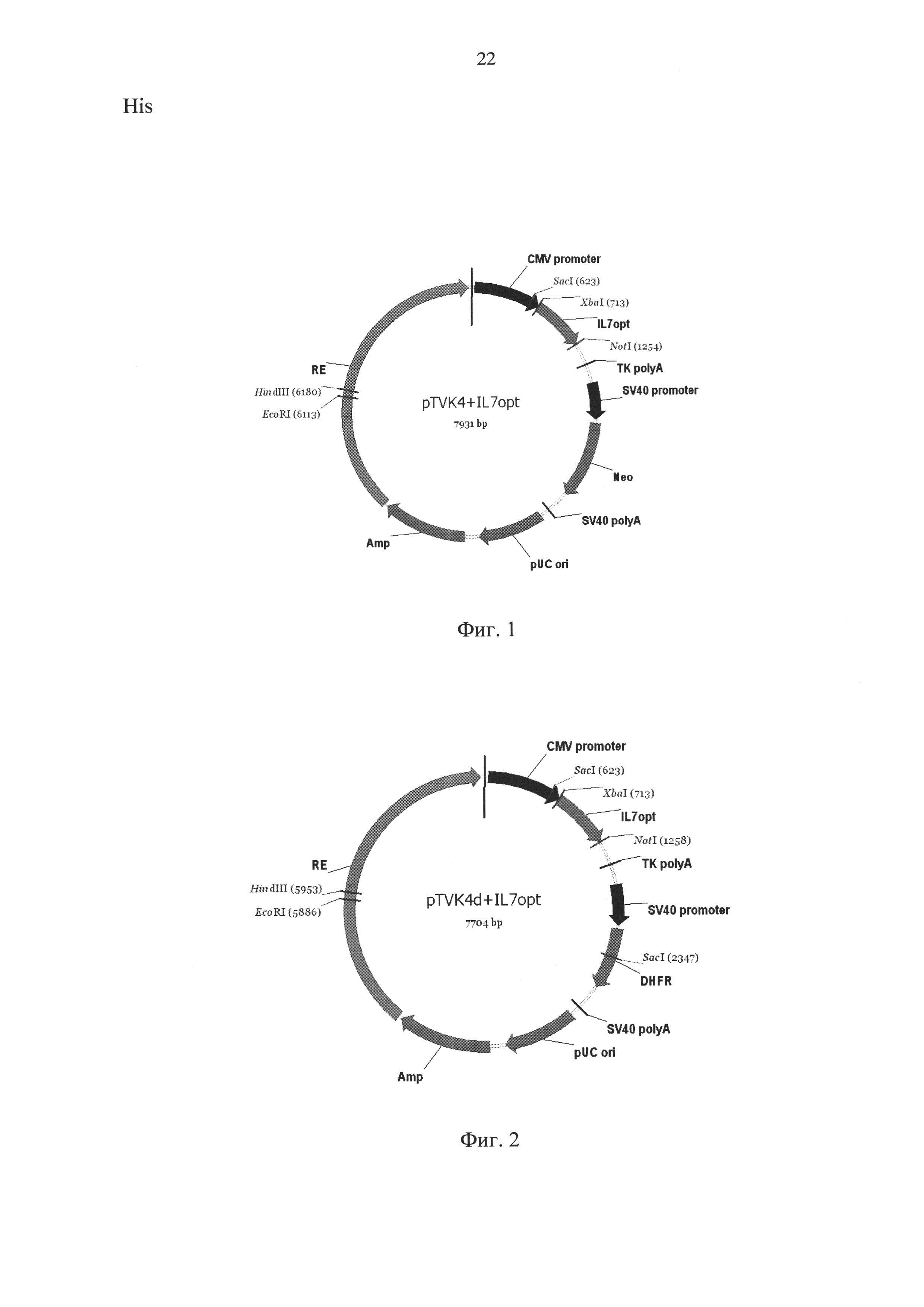
- конструированием плазмидного вектора экспрессии pTVK4/IL7, схема которого приведена на фиг. 1, содержащего вышеуказанную ДНК и в качестве селективного маркера ген устойчивости к неомицину/G418;
- конструированием плазмидного вектора экспрессии pTVK4d/IL7, схема которого приведена на фиг. 2, содержащего конструирование плазмидного вектора экспрессии в качестве гена селекции дигидрофолатредуктазу;
- получением штамма клеток яичников китайского хомячка - продуцента рекомбинантного интерлейкина-7 человека, депонированного в коллекции РККК под номером РККК(П) 770Д;
- созданием технологии получения ИЛ-7, включающего двухстадийный способ очистки рекомбинантного ИЛ-7 из культуральной жидкости, путем гидрофобной хроматографии на каскаде из двух колонок с сорбентами разной степени гидрофобности- фенил-сефарозой и бутил-сефарозой и стадию ионообменной хроматографии на катеоните SP-сефарозе.
Заявляемая последовательность была синтезирована с помощью автоматизированного химического синтеза и вставлена по сайтам рестрикции XbaI и NoiI в два экспрессионных вектора, pTVK4 и pTVK4d, содержащие, помимо экспрессионных элементов, указанных на фиг. 3, специализированный регуляторный элемент UCOE (Williams S. et al. CpG-island fragments from the HNRPA2B1/CBX3 genomic locus reduce silencing and enhance transgene expression from the hCMV promoter/enhancer in mammalian cells. BMC Biotechnology 2005, 5:17), обозначенный на Фиг. 1 и 2, как RE, при этом pTVK4 несет ген устойчивости к неомицину, a pTVK4d - ген дигидрофолатредуктазы. В результате были получены две экспрессионные плазмиды, pTVK4AL7 и pTVK4d/IL7,
Штамм клеток яичников китайского хомячка CHO-IL7 16/16 - продуцент рекомбинантного интерлейкина-7 человека был помещен в Специализированную коллекцию культур клеток позвоночных Российской коллекции клеточных культур 25.06.2015 под №РККК(П) 770Д.
Штамм характеризуется следующими свойствами:
Родословная штамма: родительская клеточная линия - CHOdhfr-. Котрансфекция плазмидами pTVK4d/IL7 и pTVK4/IL7, отбор стабильного высокопродуктивного клона на среде без гипоксантина/тимидина с 150 мкг/мл G-418 и постепенным увеличением концентрации метотрексата.
Число пассажей к моменту паспортизации и депонирования - 15. Стандартные условия выращивания: среда CDM4CHO (HyClone) без гипоксантина/тимидина с 6 мМ глутамина, 20 нМ метотрексата, 150 мкг/мл G-418, 37°С, 5% CO2.
Культуральные свойства: суспензионное культивирование в пробирках, колбах или 6-луночных платах на орбитальном шейкере с частотой вращения 130-180 об/мин. Посевная доза 0,3-0,5×106/мл, пересев каждые 3-4 суток. Время удвоения 25-27 часов.
Данные по видовой принадлежности: Cricetulus griseus (ПЦР-анализ с видоспецифичными праймерами).
Продуцирует рекомбинантный гликозилированный интерлейкин-7 человека (оценка с помощью иммуноферментного анализа, вестерн-блота, биологического теста на тимоцитах мыши)
Рекомбинантный интерлейкин-7 человека секретируется в культуральную среду в количестве около 10 пг на клетку в сутки. Стабильность культивирования - не менее 20 пассажей.
Способ криоконсервирования: 45% свежей среды CDM4CHO, 45% кондиционной среды, 10% DMSO, 3-5×106 клеток/мл, заморозка до -70°С со скоростью 1°С/мин, далее помещение в жидкий азот, жизнеспособность после размораживания и отмывки от криоконсерванта 90% (с трипановым синим).
Способ получения ИЛ-7 включает технологию выделения и очистки интерлейкина-7 человека из супернатанта штамма CHO-IL7 16/16, основанную на проведении двух хроматографических стадий. На первой стадии используется гидрофобная хроматография с применением каскада из двух колонок с сорбентами разной степени гидрофобности - фенил-сефарозой и бутил-сефарозой. При этом первая колонка с фенил-сефарозой выполняет функцию избирательного удаления присутствующего в культуральной среде неионного детергента, который необходим для уменьшения механического повреждения клеток при суспензионном культивировании, но в то же время осложняет проведение различных процедур очистки, включая хроматографию. Кроме того, на фенил-сефарозе адосбируется большинство близких по гидрофобности к ИЛ-7 примесных белков. Подобный подход позволяет обеспечить элюцию ИЛ-7 со второй колонки простым изменением состава элюента с высоким выходом и степенью очистки.
Использование на второй стадии хроматографии катионита SP-сефарозы позволяет достичь высокой степени очистки от примесей, в том числе от эндотоксина, белков и ДНК штамма-продуцента, что необходимо для получения фармацевтических субстанций. Преимущество предложенного метода заключается, помимо небольшого числа стадий, также в его технологичности, поскольку на стадиях отсутствует необходимость смены буферов вне колонок.
Сущность изобретения иллюстрируется следующими последовательностями и чертежами.
В приложении приведен список последовательностей Seq ID No 1 - ДНК, кодирующей интерлейкин-7 человека
Фиг. 1 - карта экспрессионной плазмиды pTVK4/IL7.
CMV promoter - промотор предранних белков цитомегаловируса
IL7opt - оптимизированный ген интерлейкина-7 человека
TK polyA - сайт полиаденилирования РНК из гена тимидинкиназы
HSV
SV40 promoter - промотор ранних белков вируса SV40
Neo - ген устойчивости к неомицину
SV40 polyA - сайт полиаденилирования ранних белков вируса SV40
pUC ori - точка начала репликации
Amp - ген устойчивости к ампициллину
RE - регуляторный элемент, усиливающий экспрессию целевого гена
Фиг. 2 - карта экспрессионной плазмиды pTVK4d/IL7.
CMV promoter - промотор предранних белков цитомегаловируса
IL7opt - оптимизированный ген интерлейкина-7 человека
TK polyA - сайт полиаденилирования РНК из гена тимидинкиназы
HSV
SV40 promoter - промотор ранних белков вируса SV40
DHFR - ген дигидрофолатредуктазы
SV40 polyA - сайт полиаденилирования ранних белков вируса SV40
pUC ori - точка начала репликации
Amp - ген устойчивости к ампициллину
RE - регуляторный элемент, усиливающий экспрессию целевого гена
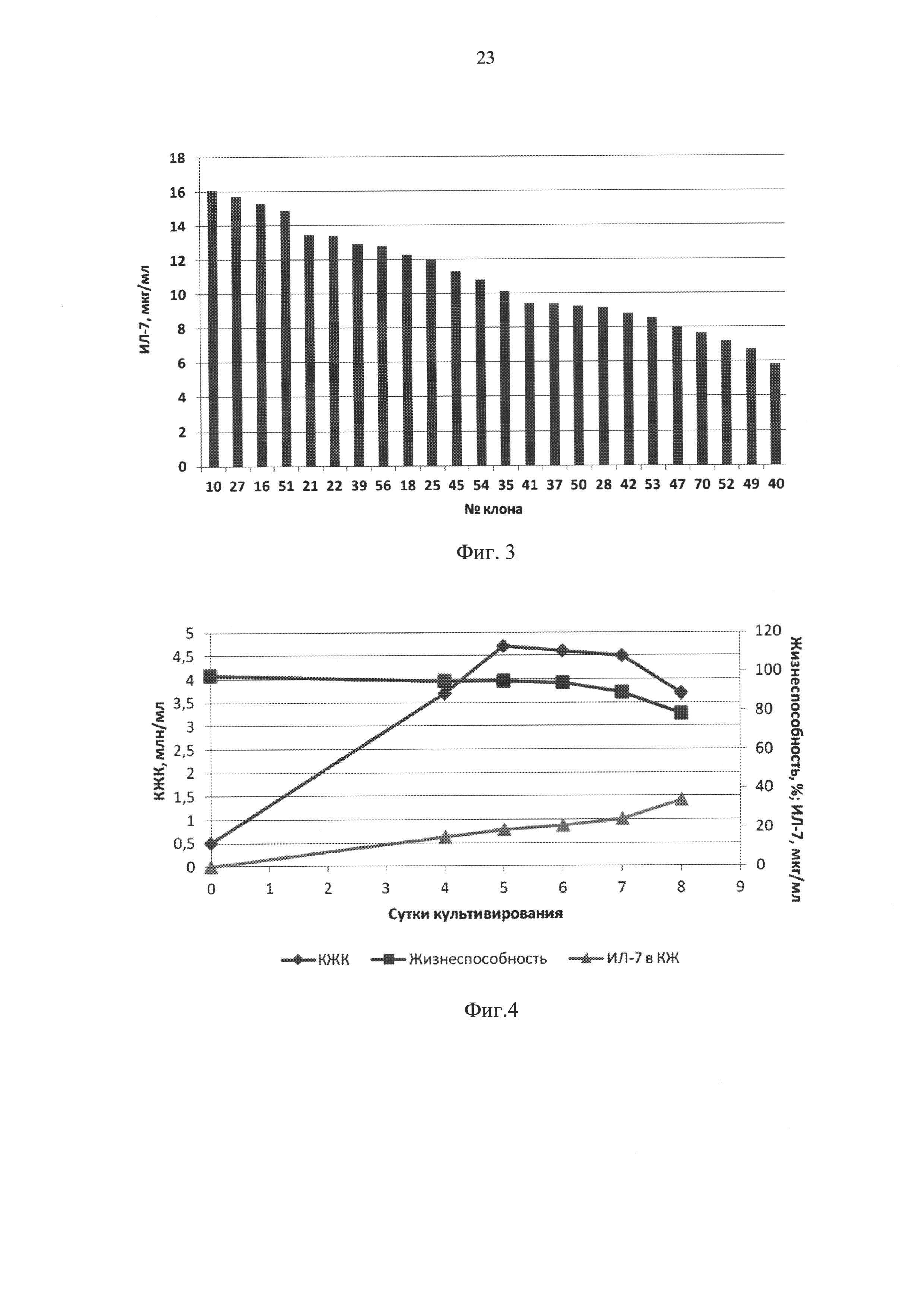
Фиг. 3 - концентрация ИЛ-7 в супернатантах через 4 суток культивирования 24 кандидатных клонов в 6-луночной плате по данным твердофазного ИФА.
Фиг. 4 - динамика роста клеток штамма-продуцента CHO-IL7 16/16, их жизнеспособности и концентрации ИЛ-7 в культуральной жидкости при культивировании при 37°С, где КЖК - концентрация жизнеспособных клеток.
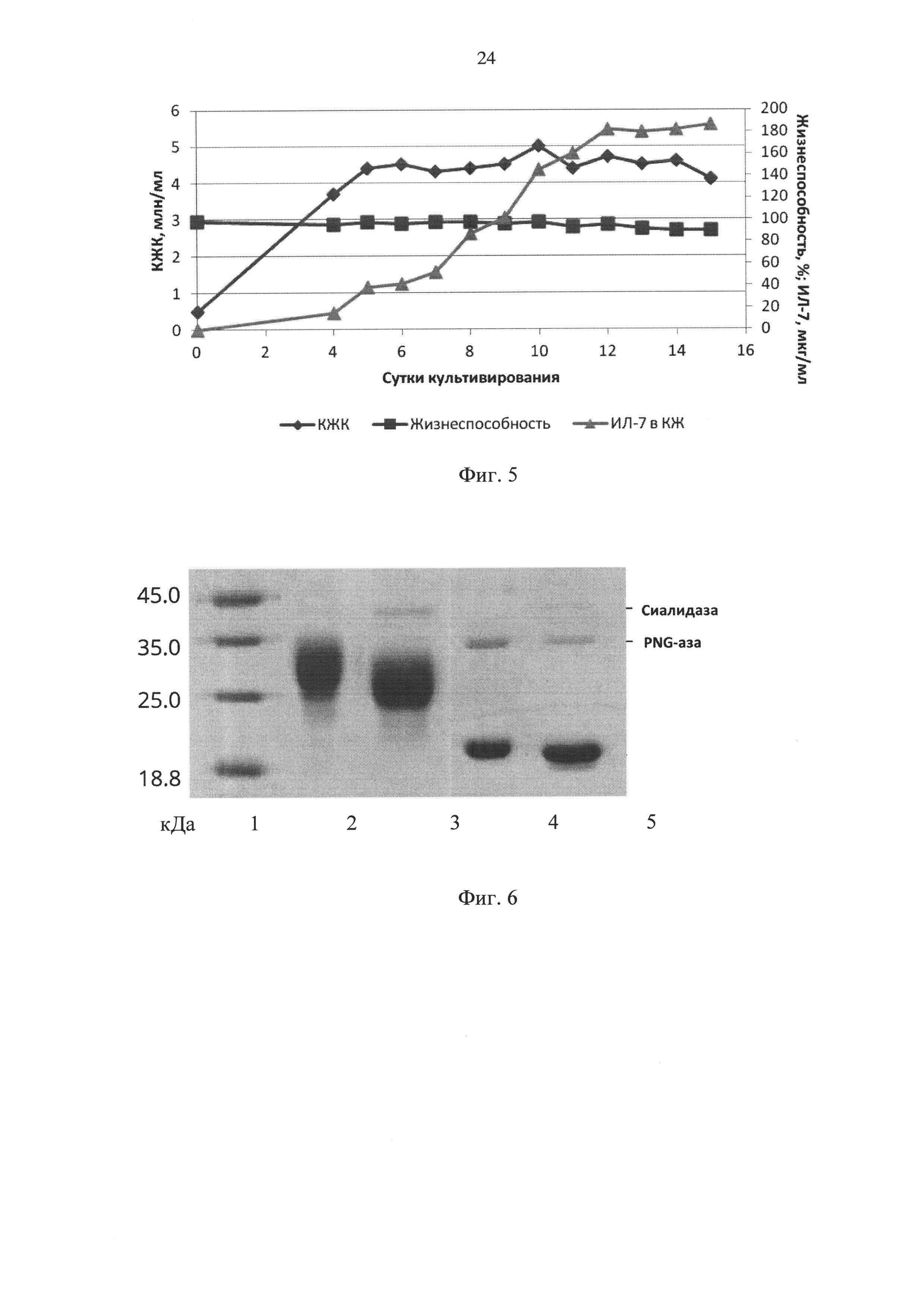
Фиг. 5 - динамика роста клеток штамма-продуцента CHO-IL7 16/16, их жизнеспособности и концентрации ИЛ-7 в культуральной жидкости при культивировании при 32°С, начиная с 4 суток, где КЖК - концентрация жизнеспособных клеток.
Фиг. 6 - электрофорез рекомбинантного ИЛ-7, очищенного из культуральной жидкости штамма-продуцента CHO-IL7 16/16, после различных вариантов дегликозилирования.
Дорожки:
1 - маркер молекулярных весов (кДа)
2 - необработанный образец ИЛ-7
3 - ИЛ-7, обработанный сиалидазой
4 - ИЛ-7, обработанный PNG-азой
5 - ИЛ-7, обработанный смесью ферментов для дегликозилирования.
Справа подписаны полосы, соответствующие ферментам.
Фиг. 7 - определение биологической активности ИЛ-7, очищенного из культуральной жидкости штамма-продуцента CHO-IL7 16/16.
Показана пролиферация тимоцитов мыши на фоне различных разведений международного стандарта ИЛ-7 (удельная активность 105 единиц активности/мкг) или двух образцов рекомбинантного ИЛ-7, выделенных из культуральной жидкости штамма-продуцента CHO-IL7 16/16, на фоне 20 Ед/мл ИЛ-1 человека. Учет интенсивности пролиферации с помощью ХТТ-теста, ось ординат - оптическая плотность при длине волны 450 нм.
Промышленная применимость изобретения поясняется следующими примерами.
Пример 1. Дизайн нуклеотидной последовательности гена, кодирующего интерлейкин-7 человека, осуществляли с учетом частоты использования кодонов в геноме китайского хомячка. Полученная последовательность, представленная в Seq ID No 1, кодирует последовательность интерлейкина-7 человека, включая сигнальный пептид. Данная последовательность была синтезирована с помощью автоматизированного химического синтеза и вставлена по сайтам рестрикции XbaI и NotI в два экспрессионных вектора, pTVK4 и pTVK4d, содержащие, помимо экспрессионных элементов, указанных на фиг. 3, специализированный регуляторный элемент UCOE (Williams S. et al. CpG-island fragments from the HNRPA2B1/CBX3 genomic locus reduce silencing and enhance transgene expression from the hCMV promoter/enhancer in mammalian cells. BMC Biotechnology 2005, 5:17), обозначенный на Фиг. 1 и 2, как RE, при этом pTVK4 несет ген устойчивости к неомицину, a pTVK4d - ген дигидрофолатредуктазы. В результате были получены две экспрессионные плазмиды, pTVK4/IL7 и pTVK4d/IL7, соответственно, карты которых представлены на Фиг. 1 и 2. Правильность синтеза гена и сборки плазмид были подтверждены сиквенированием.
Пример 2. Получение высокопродуктивного штамма культивируемых клеток - продуцента ИЛ-7 человека.
Клетки линии яичников китайского хомячка, дефицитные по гену дигидрофолатредуктазы (CHOdhfr-), адаптированные к суспензионному росту в бессывороточной среде, культивировали в питательной среде CDM4CHO («HyClone», США) с добавлением 6 мM аланилглутамина, 0,1 мМ гипоксантина, 0,016 мМ тимидина и 0,1% Pluronic F-68 («Sigma», США). Постоянную трансфекцию клеток смесью плазмидных ДНК векторов pTVK4/IL7 и pTVK4d/IL7 (1:1) проводили при помощи реагента для трансфекции «Freestyle» («Life Technologies», США). Через 48 часов после трансфекции клетки переводили на селективную среду, не содержащую гипоксантина и тимидина с добавлением 150 мкг/мл G-418 («Life Technologies»), и далее пересевали каждые 72 часа. После перевода на селективную среду в течение первых четырех пассажей жизнеспособность трансфецированных клеток снизилась до 10%, затем стала повышаться и на седьмом пассаже составила 87%. Полученный пул клеток переводили на среду, содержащую 150 мкг/мл G-418 и 5 нМ, 10 нМ или 100 нМ метотрексата (МТХ). На фоне 5 нМ МТХ снижения жизнеспособности клеток практически не происходило, при использовании 100 нМ МТХ в течение трех пассажей произошло снижение жизнеспособности клеток <5% и при дальнейшем пассировании роста клеток получено не было. При использовании МТХ в концентрации 10 нМ наблюдалось снижение жизнеспособности до 40% к третьему пассажу и затем ее восстановление до 85% к седьмому пассажу. Эта культура была клонирована методом серийных разведений с целью отбора индивидуальных клонов. Было получено 486 единичных клонов, концентрацию ИЛ-7 в супернатантах этих клонов оценивали с помощью иммуноферментного анализа (ИФА). На основании полученных результатов было отобрано 70 клонов, в супернатантах которых концентрация ИЛ-7 была выше 5 мкг/мл. Клетки этих клонов были пересеяны в 24-луночную плату. Через трое суток из лунок платы были отобраны супернатанты и проведен второй анализ концентрации ИЛ-7 в супернатантах, на основании которого было выбрано 24 лучших по продукции белка клона. Клетки этих клонов были пересеяны в 6-луночную плату в плотности 0,2 млн/мл. Через четверо суток культивирования с помощью ИФА повторно оценивали концентрацию ИЛ-7 в супернатантах. Результаты представлены на Фиг. 3.
На основании полученных данных было отобрано 8 клонов, имеющих продукцию ИЛ-7 на 4-е сутки от 12 мкг/мл до 16 мкг/мл. Культивирование этих клонов продолжили на шейкере в микробиореакторах типа TubeSpin 50 в объеме 5 мл. После адаптации клеток к росту на шейкере оценивали их динамику роста и продукции, а также удельную продуктивность клеток выбранных клонов. Полученные данные представлены в таблице №1.
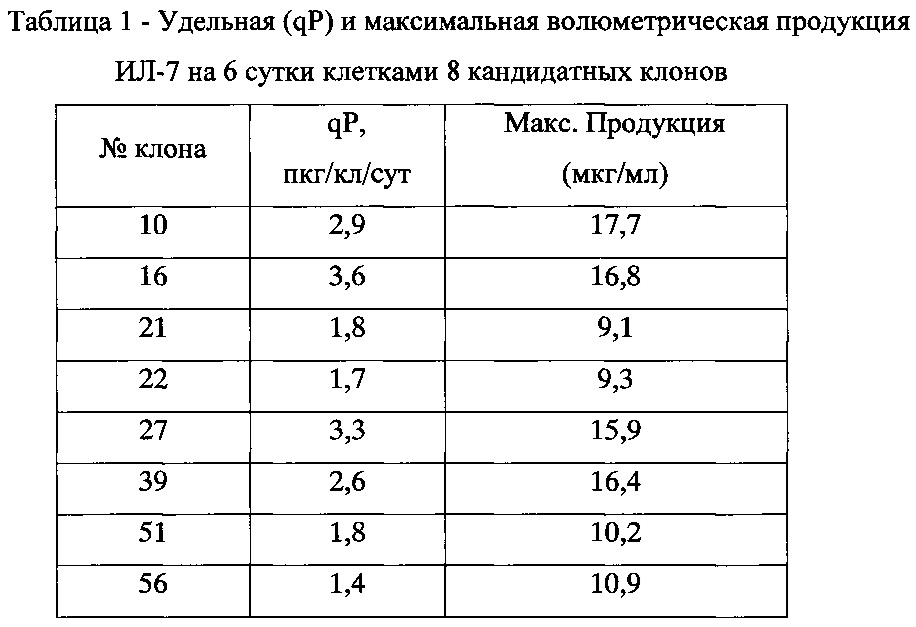
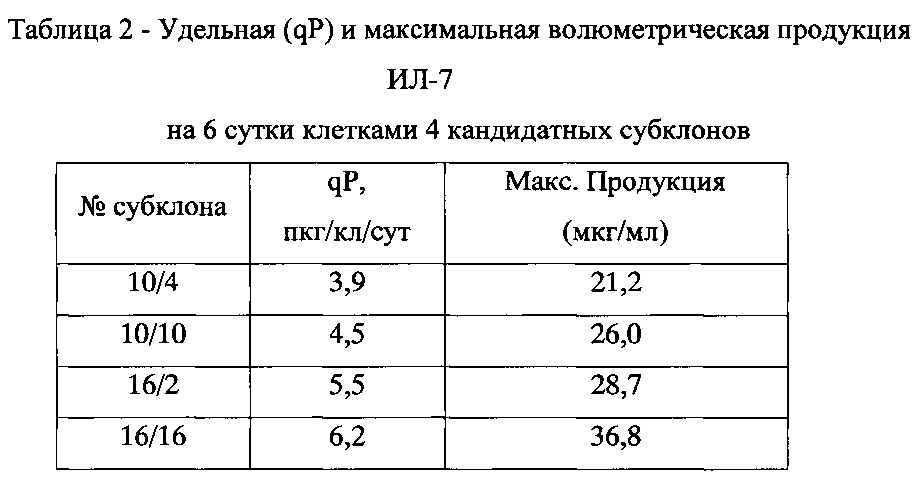
Учитывая полученные данные, для дальнейшей работы были выбраны клоны №10, показавший максимальную волюметрическую продуктивность, и №16, имеющий наибольшую удельную продуктивность. Клетки клонов культивировали в среде, содержащей 150 мкг/мл G-418 и 20 нМ МТХ. Как и при адаптации к росту при 10 нМ МТХ, вначале наблюдалось снижение жизнеспособности культур, через 6-7 пассажей интенсивность роста восстановилась, после чего клетки обоих клонов были повторно клонированы методом лимитирующих разведений. Было получено 180 субклонов из клона №10 и 166 субклонов из клона №16. Путем последовательных скринингов концентрации ИЛ-7 в супернатантах, как описано выше, было отобрано по 2 субклона, полученных от клонов №10 и №16, с максимальной продуктивностью. После анализа продукции ИЛ-7 клетками 4 кандидатных субклонов в микробиореакторах типа TubeSpin 50 было проведено окончательное сравнение продуктивности (таблица 2). Наиболее высокопродуктивным субклоном оказался субклон 16/16. Культура клеток этого клона была масштабирована для создания мастер-банка и депонирована с авторским наименованием CHO-IL7 16/16 в Специализированную коллекцию культур клеток позвоночных Российской коллекции клеточных культур под номером РККК(П) 770Д.
Пример 3. Оптимизация условий культивирования штамма-продуцента СНО-IL7 16/16.
Для подбора оптимальных условий культивирования клеток штамма-продуцента CHO-IL7 16/16, обеспечивающих накопление максимального уровня рекомбинантного ИЛ-7 в культуральной жидкости (КЖ), тестировали питательную добавку с химически определенным составом EX-CELL CD Hydrolysate Fusion (Sigma, США), которую вносили в питательную среду в количестве 5% от объема в первые сутки культивирования. Клетки культивировали в минибиореакторах типа TubeSpin-50 при непрерывном перемешивании при 37°С. Стартовая плотность составляла 0,5×106/мл. Ежедневно, начиная с 4 суток, определяли концентрацию жизнеспособных клеток (КЖК), жизнеспособность по исключению трипанового синего, отбирали образцы КЖ для определения концентрации ИЛ-7 в динамике, которое выполнялось с помощью твердофазного ИФА.
Как видно из результатов, графически представленных на Фиг. 4, клетки сохраняли жизнеспособность выше 80% в течение 7 суток, за это время концентрация ИЛ-7 в КЖ достигала 35 мкг/мл. С 5 по 7 сутки КЖК составляла около 4,5×106/мл.
Для создания более благоприятных условий для ферментации была испытана стратегия культивирования, при которой, начиная с 4 суток, температуру в инкубаторе снижали до 32°С. В остальном условия были аналогичны описанным выше. Этот подход позволил радикально улучшить результаты, удлинив время сохранения высокой жизнеспособности до 15 суток, при этом максимальная КЖК составила 5×106/мл, а уровень жизнеспособных клеток около 4,5×106/мл сохранялся с 5 до 14 суток (Фиг. 5). За это время, как видно из Фиг. 5, концентрация ИЛ-7 в КЖ к 12 суткам достигла 180 мкг/мл и далее не менялась.
Пример 4. Выделение рекомбинантного интерлейкина-7 из культуральной жидкости штамма-продуцента CHO-IL7 16/16
Штамм-продуцент CHO-IL7 16/16 культивировали в среде CDM4CHO с 5% питательной добавки EX-CELL CD Hydrolysate Fusion по схеме со снижением температуры до 32°С на 4 сутки, как описано в примере 3, в колбах Эрленмейера с рабочим объемом 150 мл. На 12 сутки от начала культивирования к 1 л культуральной жидкости добавляли ингибитор сериновых протеаз фенилметилсульфонил фторид и азид натрия до конечных концентраций 1 мМ и 0,02%, соответственно, после чего осветляли путем центрифугирования в центрифуге JE-6 (Beckman) в роторе JA-10 при 10000 об/мин 30 мин при температуре 4°С.
К супернатанту добавляли 143 г порошка сульфата аммония (NH4)2SO4 (конечная концентрация 1 М) и наносили раствор на хроматографическую колонку (XK26x20), заполненную 50 мл сорбента Phenyl-sepharose FF (GE), последовательно соединенную с аналогичной колонкой, содержащей 70 мл сорбента Fractogel TSK Butyl-650(S) (Tosoh). Колонки предварительно уравновешивали буфером, содержащим 1М (NH4)2SO4, 50 мМ однозамещенного фосфата натрия, рН 6,5. Скорость потока устанавливали равной 10 мл/мин и после нанесения раствора, содержащего целевой белок, пропускали 250 мл буфера для уравновешивания. Адсорбированный белок элюировали с колонки с сорбентом Fractogel TSK Butyl-650(S) буфером, содержащим 50 мМ однозамещенного фосфата натрия, рН 6,5 под контролем УФ-монитора при длине волны 280 нм. Отбирали фракцию, соответствующую пику поглощения на мониторе, объем который составлял около 50 мл. Контроль над выходом целевого белка подтверждали, используя набор для иммуноферментного определения ИЛ-7 (R&D Systems). Выход ИЛ-7 после произведенных манипуляций составлял около 90% по отношению к исходному содержанию после культивирования.
Для проведения дальнейшей процедуры хроматографической очистки 50 мл раствора, элюированные с колонки с сорбентом Fractogel TSK Butyl-650(S), содержащие ИЛ-7, разбавляли дистиллированной водой, устанавливая проводимость раствора равной 4 mS. Вторую стадию хроматографии проводили на колонке XK26x20, содержащей сорбент SP-Sepharose FF (GE). Колонку уравновешивали 30 мМ Na-фосфатным буфером, рН 6,5. Раствор ИЛ-7 (4 mS) наносили на колонку со скоростью потока 20 мл/мин, после чего пропускали еще 100 мл уравновешивающего буферного раствора. Абсорбированный белок элюировали с колонки, используя раствор 30 мМ фосфатного буфера, содержащего 0,2М хлорида натрия, под контролем УФ монитора при длине волны 280 нм при скорости потока 5 мл/мин. Фракцию, содержащую ИЛ-7, в соответствии с однопиковым профилем элюции собирали в объеме 20 мл.
Согласно данным ИФА выход ИЛ-7 составлял 70% от исходного содержания. Степень чистоты, согласно данным обращенно-фазной ВЭЖХ составляла 95-97%. Содержание эндотоксина было менее 20 ЕЭ/мг белка (определение с помощью LAL-теста), примесь ДНК штамма-продуцента - менее 1 нг/мг (определение с помощью набора СНО Host Cell DNA на основе ПНР в реальном времени, Cygnus Technologies, США), примесь белка штамма-продуцента - менее 100 нг/мг (определение с помощью ИФА-набора СНО Host Cell Protein, Cygnus Technologies, США).
Пример 5. Анализ гликозилирования и специфической активности белка, продуцируемого клетками штамма CHO-IL7 16/16.
Полученный, как описано в примере 4, рекомбинантный ИЛ-7 человека обрабатывали ферментами для селективного дегликозилирования: нейраминидазой, отщепляющей концевые сиаловые кислоты, PNG-азой, отщепляющей N-гликаны, или смесью ферментов для тотального дегликозилирования. Обработку ферментами проводили по инструкции изготовителя (NEB, США). Затем образцы анализировали с помощью денатурирующего гель-электрофореза с додецилсульфатом натрия.
Результат, представленный на Фиг. 6, показывает наличие широкой белковой полосы с кажущейся молекулярной массой 25-33 кДа у необработанного ферментами белка. После обработки сиалидазой полоса остается широкой, но ее кажущаяся масса снижается до 24-30 кДа. Обработка PNG-азой приводит к визуализации узкой полосы с массой около 19 кДа. При обработке смесью ферментов для полного дегликозилирования кажущаяся масса рекомбинантного ИЛ-7 снижается еще на 0,5 кДа. В целом, полученные данные указывают на наличие в составе молекул рекомбинантного ИЛ-7, выделенных из культуральной жидкости штамма CHO-IL7 16/16, N- и О-гликанов, что характерно для гликопротеинов, секретируемых клетками млекопитающих. При этом в составе N-гликанов присутствует значительное количество сиаловых кислот: исходя из разницы молекулярных масс десиалированного и интактного ИЛ-7 в 1-3 кДа и массы одной сиаловой кислоты в 0,2 кДа, их количество может колебаться от 5 до 15 на одну молекулу ИЛ-7.
Присутствие значительного количества сиаловых кислот также должно существенно изменять изоэлектрическую точку белка. Для оценки изоэлектрической точки белка, полученного по примеру 4, было проведено капиллярное изоэлектрическое фокусирование с использованием системы капиллярного электрофореза «Капель-105М» и набора реагентов для изофокусирования в капилляре производства BD (США). Результаты показали присутствие в препарате белка пяти групп изоформ с изоэлектрическими точками от 4,85 до 6,25. При этом теоретическая изоэлектрическая точка рекомбинантного ИЛ-7, рассчитанная на основе первичной аминокислотной последовательности, составляет 8,72. Это также подтверждает факт обильного гликозилирования, позволяя оценить количество сиаловых кислот в составе одной молекулы рекомбинантного ИЛ-7 от 6 до 11.
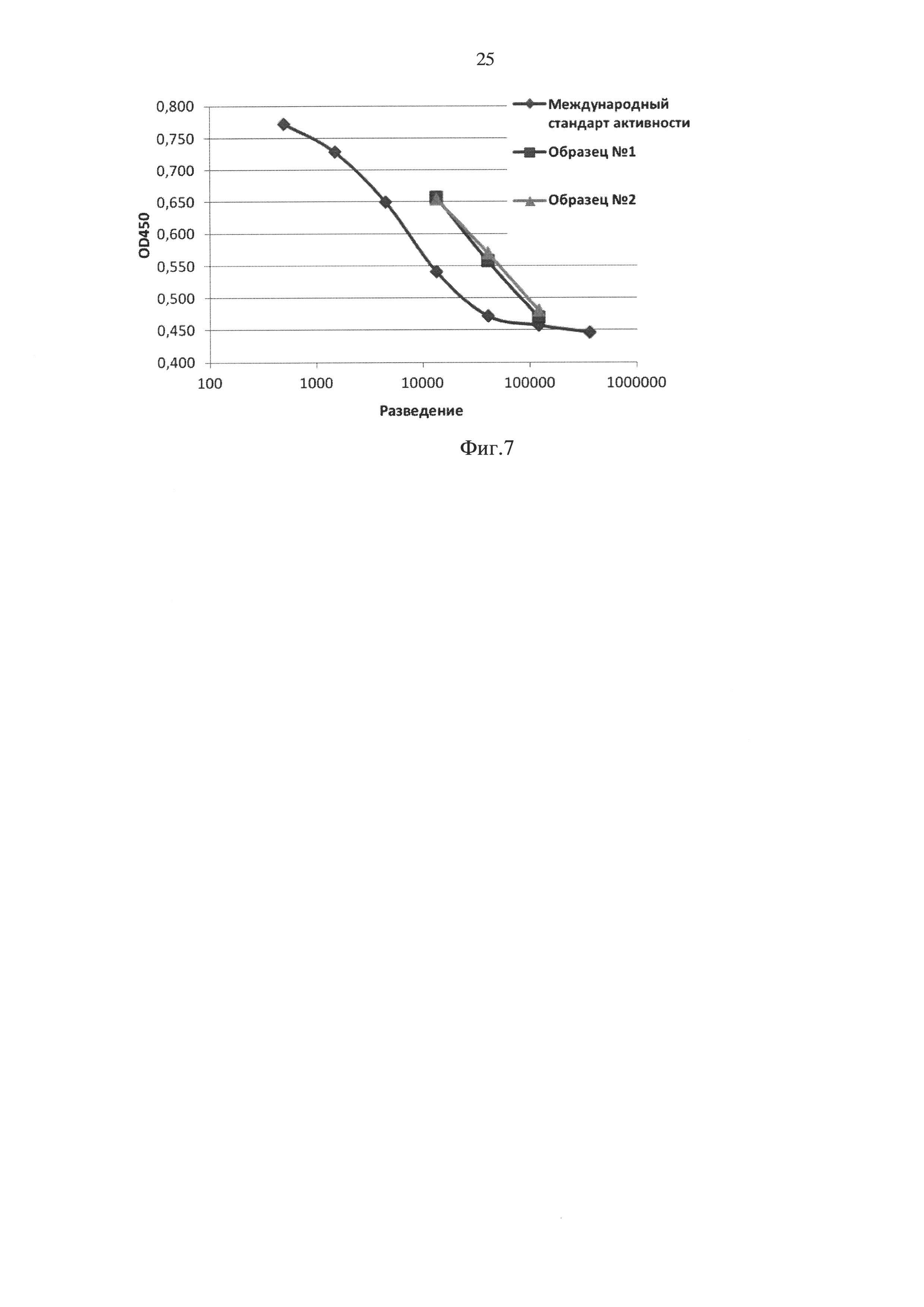
Определение биологической активности полученного по примеру 4 рекомбинантного ИЛ-7 человека выполняли на первичной культуре тимоцитов мыши, по способности данного цитокина стимулировать пролиферацию тимоцитов на фоне субоптимальных концентраций интерлейкина-1 [Herbelin A., Machavoine F., Schneider Е. et al. IL-7 is requisite for IL-1-induced thymocyte proliferation. Involvement of IL-7 in the synergistic effects of granulocyte-macrophage colony-stimulating factor or tumor necrosis factor with IL-1 // J. Immunology - 1992. - V.148. - №1. - P. 99-105].
Тимоциты выделяли из селезенок мышей-гибридов F1 (C57BL/6×CBA) асептически, дважды отмывали в стерильном забуференном физиологическом растворе и ресуспендировали в среде RPMI-1640 (Sigma) с 2,0 мМ L-глутамина, 50 мкМ 2-меркаптоэтанола и 80 мг/л гентамицина (полная среда). Тимоциты инкубировали в 96-луночном плоскодонном планшете в 4-х параллелях по 1,5×106 клеток на лунку в присутствии различных концентраций рекомбинантного ИЛ-7 и международного стандарта активности ИЛ-7 (WHO Reference Reagent Interleukin-7, NIBSC code: 90/530) на фоне 20 Ед/мл ИЛ-1 человека. Инкубацию проводили в полной среде с 2% телячьей эмбриональной сыворотки в течение 72 ч в СО2-инкубаторе при температуре 37°С. За 5 ч до окончания инкубации в культуры вносили ХТТ (Life Technologies) в конечной концентрации 0,33 мг/мл и феназин метосульфат (Sigma) в конечной концентрации 2 мкг/мл. Через 3-5 часов после внесения оптическую плотность в лунках определяли на планшетном счетчике Victor (Perkin-Elmer) при длине волны 450 нм. Для стандартного образца строили калибровочный график зависимости OD450 от разведения. Далее по этому калибровочному графику рассчитывали активность исследуемых образцов, с учетом их разведений.
Результат измерения активности в двух независимо полученных образцах рекомбинантного ИЛ-7 представлен на фиг. 7. Расчетная удельная активность ИЛ-7, полученного по примеру 4, составила для двух независимо полученных образцов 3,3×108 Ед/мг и 3,5×108 Ед/мг, соответственно, что более чем в 3 раза превышает удельную активность международного стандарта ИЛ-7, полученного из клеток Е. coli.
Из полученных результатов следует, что продуцируемый штаммом-продуцентом CHO-IL7 16/16 рекомбинантный ИЛ-7, очищенный по примеру 4, представляет собой обильно гликозилированный белок с высокой биологической активностью.


Устройство для отведения воды из деформационного шва (варианты)
Штамм клеток cho-se-9/4 - продуцент химерного антитела против эритропоэтина человека и химерное антитело, продуцируемое данным штаммом
Моноклональное антиидиотипическое антитело аи-g1, обладающее антигенными свойствами морфина
Моноклональное антитело к бтш70
Рекомбинантный белок для иммунизации против холеры
Способ получения различных видов морфологии поверхности карбида кремния
Способ контроля качества аммиачной тепловой трубы
Антитела против белка р17 вич-1 субтипа а
Устройство для отведения воды из деформационного шва (варианты)
Антитела против анафилатоксина c5a человека
Вакцина против герпеса

